degradation of p22 phox and inhibition of superoxide generation by ehrlichia chaffeensis in human...
TRANSCRIPT

Degradation of p22phox and inhibition of superoxidegeneration by Ehrlichia chaffeensis in humanmonocytes
Mingqun Lin and Yasuko Rikihisa*Department of Veterinary Biosciences, The Ohio StateUniversity, Columbus, OH 43210, USA.
Summary
Ehrlichia chaffeensis is an obligate intracellularbacterium which replicates in monocytes or macro-phages, the primary producers of reactive oxygenspecies (ROS). However, effects of ROS onE. chaffeensis infection and whether E. chaffeensismodulates ROS generation in host monocytes areunknown. Here, E. chaffeensis was shown to loseinfectivity upon exposure to O2
– or hydrogenperoxide. Upon incubation with human monocytes,E. chaffeensis neither induced O2
– generation byhuman monocytes, nor colocalized with nicotinamideadenine dinucleotide phosphate (NADPH) oxidasecomponents. Instead, it actively blocked O2
– genera-tion by monocytes stimulated with phorbol myristateacetate and caused the rapid degradation of p22phox, acomponent of NADPH oxidase. These effects werenot seen in neutrophil, which is another potent ROSgenerator, but a cell type that E. chaffeensis does notinfect. Trypsin pretreatment of monocytes preventedthe inhibition of O2
– generation by E. chaffeensis. Thedegradation of p22phox by E. chaffeensis was specificto subsets of monocytes with bound and/or intracel-lular bacteria, and the degradation could be reducedby heat treatment of the bacterium, lipopolysac-charide pretreatment of monocytes, or the incuba-tion with haemin. The degradation of p22phox byE. chaffeensis and its prevention by haemin or pro-tease inhibitors also occurred in isolated monocytemembrane fractions, indicating that host cytoplasmicsignalling is not required for these processes. Theamount of gp91phox was stable under all conditionsexamined in this study. These findings point to aunique survival mechanism of ROS-sensitive obligateintraleucocytic bacteria that involves the destabiliza-
tion of p22phox following the binding of bacteria to hostcell surface proteins.
Introduction
Ehrlichia chaffeensis, a Gram-negative obligate intracel-lular bacterium, replicates exclusively in the cytoplasm ofmonocytes/macrophages and causes an emerging tick-borne febrile illness called human monocytic ehrlichiosis(HME) (Paddock and Childs, 2003; Rikihisa, 2003). Giventhe importance of monocytes–macrophages in antimicro-bial defence, the elucidation of the host subversionmechanism by E. chaffeensis has become a central focusfor understanding pathogenesis of HME (Rikihisa, 2006).Among various antimicrobial defence mechanisms arereactive oxygen species (ROS) generated by thenicotinamide adenine dinucleotide phosphate (NADPH)oxidase, a multicomponent enzyme composed of amembrane-bound heterodimeric cytochrome b558 compo-nent (gp91phox and p22phox), three cytosolic subunits(p67phox, p47phox and p40phox) and a low-molecular-weightGTPase (Rac 1/2 or Rap1A) (Babior, 1999; Fang, 2004).When phagocytes are in a resting state, the NADPHoxidase is inactive by keeping its componentsdissociated. In response to invading pathogens or stimu-lating agents such as phorbol myristate acetate (PMA)and formyl-methionyl-leucyl-phenylalanine (fMLP), allcomponents of NADPH oxidase are rapidly assembledinto a holoenzyme to catalyse the production of superox-ide anion (O2
–) from oxygen. O2– serves as the starting
material for the production of powerful microbicidal ROS,including hydrogen peroxide (H2O2), oxidized halogens,hydroxyl radicals and singlet oxygen. As the effect of ROSon E. chaffeensis infection is still unknown, experimentsare warranted to define whether E. chaffeensis is suscep-tible to ROS.
Pathogenic microorganisms have evolved various strat-egies to counteract the ROS-induced killing. The strate-gies include evasion of ROS, suppression of ROSproduction, enzymatic detoxification and scavenging ofROS, iron sequestration, increased resistance to ROS bystress responses, and damage repair (Miller and Britigan,1997; Fang, 2004). In this regard, Anaplasma phagocyto-philum, an obligate intracellular bacterium of human
Received 11 August 2006; revised 9 September 2006; accepted 20September 2006. *For correspondence. E-mail [email protected];Tel. (+1) 614 292 9677; Fax (+1) 614 292 6473.
Cellular Microbiology (2007) 9(4), 861–874 doi:10.1111/j.1462-5822.2006.00835.xFirst published online 3 November 2006
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd

granulocytes and closely related to E. chaffeensis, pos-sesses a remarkable feature that sets it apart from manyother pathogenic bacteria (Carlyon and Fikrig, 2006;Rikihisa, 2006). Previous studies had shown thatA. phagocytophilum by itself does not induce NADPHoxidase activation, does not colocalize with NADPHoxidase components, and actively blocks O2
– generationin human or murine neutrophils or differentiated HL-60cells stimulated with Escherichia coli, PMA, fMLP orFc-OxyBurst immune complexes (Banerjee et al., 2000;Mott and Rikihisa, 2000; Mott et al., 2002; Wang et al.,2002; Carlyon et al., 2004; IJdo and Mueller, 2004;Borjesson et al., 2005). Furthermore, A. phagocytophilumlowers NADPH oxidase activity by several mechanisms,including reducing the amount of pre-existing p22phox
protein in neutrophils (Mott et al., 2002), downregulatingthe genes encoding Rac2 and gp91phox, and scavengingROS by superoxide dismutase (SOD) (Banerjee et al.,2000; Carlyon et al., 2002; 2004; IJdo and Mueller, 2004).
Given the importance of subversion of the ROS-mediated killing of obligate intraleucocytic bacteria, thepresent study examined the ROS susceptibility ofE. chaffeensis as well as A. phagocytophilum, influencesof E. chaffeensis on NADPH oxidase activation in humanmonocytes, and possible inhibitory mechanisms ofNADPH oxidase activation by E. chaffeensis.
Results
Ehrlichia chaffeensis and A. phagocytophilum are highlysusceptible to O2
– and H2O2
In order to defend against oxidative killing, many patho-gens employ ROS-detoxification enzymes to convertROS to less- or non-toxic species; these enzymes includeSOD, catalase/peroxidase and glutathione (GSH) in com-bination with GSH peroxidase and GSH reductase (Farrand Kogoma, 1991; Miller and Britigan, 1997; Fang,2004). Our analysis showed that these detoxificationenzymes, including [Cu, Zn]-SOD, Mn-SOD, catalases,peroxidases, GSH peroxidase/reductase and/or OxyR/SoxRS regulons are present in many facultative intracel-lular bacteria of monocytes–macrophages such asSalmonella, Brucella, Legionella and Mycobacterium spp.(Table S1). However, most genes known to confer resis-tance to oxidative stress were absent in the E. chaffeensisand A. phagocytophilum genomes (Dunning Hotoppet al., 2006). This feature is evolutionary conservedamong several genera in the order Rickettsiales, includingEhrlichia, Anaplasma, Neorickettsia, Wolbachia and Rick-ettsia (Table S1). Only ROS-resistance genes conservedin E. chaffeensis and A. phagocytophilum are sodB,which encodes cytosolic Fe-SOD, and genes that encodethe Ahp family proteins (AhpC), which might reduce
organic hydroperoxides (Seaver and Imlay, 2001; Ohashiet al., 2002; Dunning Hotopp et al., 2006). In addition,Herron et al. proposed a SOD-independent mechanism inwhich A. phagocytophilum might express an outermembrane-tethered cytochrome c oxidase that acceptselectrons from O2
–, thereby converting it to harmless O2
(Herron and Goodman, 2001). Given that considerablenumbers of genes in the completed genomes ofE. chaffeensis and A. phagocytophilum have no functionassigned (Dunning Hotopp et al., 2006), we cannot denythe possibility that these bacteria may have unusual ROSresistance.
We therefore examined the effects of O2– and H2O2 on
the infectivity of E. chaffeensis and A. phagocytophilum.For bacteria harbouring ROS detoxification enzymes,including E. coli, Brucella and Coxiella (Table S1), treat-ment with 10–100 mM H2O2 or paraquat (methyl viologen,1,1′-dimethyl-4,4′-bipyridinium dichloride), which can gen-erate both intracellular and extracellular O2
– (Hassan andFridovich, 1979; Hassett et al., 1987), has little effect onbacterial growth (Heinzen et al., 1992; Kim et al., 2000;Bhattacharya et al., 2004). The treatment of uninfectedTHP-1 or HL-60 cells with either 100 mM H2O2 or paraquatat 37°C for 30 min did not have noticeable effects on cellviability, morphology or growth, indicating that the hostcells were capable of scavenging this amount of ROS(data not shown). In contrast, pretreatment of host cell-free E. chaffeensis or A. phagocytophilum with 10 mMH2O2 or paraquat at 37°C for 30 min reduced their abilityto infect host cells by more than 70% relative to bacteriaincubated with medium alone (Fig. 1), indicating thatneither bacterial species could overcome this low level ofROS in vitro.
Ehrlichia chaffeensis blocks PMA-induced O2–
production by human peripheral blood mononuclearcells (PBMCs) and monocytes
Given the susceptibility of E. chaffeensis to O2– and H2O2,
we employed two methods to detect the production of O2–
by E. chaffeensis-infected human monocytes. Theluminol-dependent chemiluminescence (LDCL) assaymeasures total (intra- and extracellular) O2
– productionusing luminol, a small membrane-permeable luminogenicmolecule (Dahlgren et al., 1984), whereas the ferricyto-chrome c reduction assay measures extracellularlyreleased O2
–. LDCL assay showed that host cell-freeE. chaffeensis itself or THP-1 lysates control did notinduce O2
– generation by human peripheral blood mono-nuclear cells (PBMCs) up to 30 min of co-incubation(Fig. 2A, showing the result up to 15 min). Thus unlikemost pathogenic bacterial species studied (Underhill andOzinsky, 2002), the binding and internalization of
862 M. Lin and Y. Rikihisa
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Cellular Microbiology, 9, 861–874

E. chaffeensis to monocytes do not trigger ROSproduction.
However, in E. chaffeensis-infected hosts, ROS islikely generated once monocytes/macrophages are acti-vated. For example, Neorickettsia risticii, an obligateintracellular bacterium of horse monocytes/macrophagesand closely related to E. chaffeensis, does not induceROS in macrophages from naïve mice, but stimulatesthe production of ROS by macrophages from infection-induced immune mice (Williams et al., 1994). Therefore,we examined whether E. chaffeensis has ability toactively block ROS generation by monocytes with PMA,a well-studied direct stimulator for NADPH oxidase (Coxet al., 1985). Upon stimulation with PMA (0.5 mg ml-1),O2
– was produced by PBMCs; however, pre-incubationwith E. chaffeensis could actively block PMA-inducedO2
– release in a time-dependent manner (Fig. 2B): pre-
incubation of PBMCs with E. chaffeensis for 5 minblocked PMA-induced O2
– generation by 25 � 2% (n = 3;P < 0.05) and pre-incubation of PBMCs with E. chaffeen-sis for 30 min blocked O2
– generation by 64 � 8% (n = 4;P < 0.05).
Although LDCL assay has the advantage of not beingaffected by SOD or cytochrome c oxidases that arepresent in the host cells and the bacteria (Fridovich,1997), it has disadvantages of luminol radicals having aspontaneous reaction to reduce O2 to O2
– (although thiswas minimum as shown in Fig. 2A), and luminol beingexcited by other ROS in addition to O2
– (Miller and Fridov-ich, 1986). Therefore, we performed the ferricytochrome creduction assay that measures extracellular O2
–. Theresult was in agreement with the LDCL assay, showingsimilar inhibitory effects of PMA-induced O2
– generation inisolated human monocytes incubated with E. chaffeensisfor 30 min, then stimulated with PMA for 30 min (Fig. 2E).As E. chaffeensis was cultivated in THP-1 cells, unin-fected THP-1 cell lysates were used as a control through-out this study. Use of uninfected THP-1 cell lysates inplace of E. chaffeensis had no effect on PMA-inducedROS production by human monocytes (Fig. 2B and E).These results support that E. chaffeensis actively blocksproduction of both intracellular and extracellular O2
– inhuman monocytes.
The inhibition of PMA-induced O2– production by
E. chaffeensis is host cell-specific
We next tested whether E. chaffeensis prevents PMA-induced O2
– production by human neutrophils, a cell typethat E. chaffeensis does not infect. Neutrophils andPBMC were isolated from the same peripheral bloodspecimen and separately incubated with host cell-freeE. chaffeensis. Unlike in PBMCs, E. chaffeensis could notprevent PMA-induced O2
– production by human neutro-phils (Fig. 2C). Conversely, A. phagocytophilum did notblock PMA-induced O2
– production in PBMCs, a cell typethat A. phagocytophilum does not infect (Fig. 2E). Thehost cell-specific inhibition of ROS production byE. chaffeensis suggests the involvement of host cellsurface components. Therefore, we pretreated humanmonocytes with 0.25% trypsin for 15 min to removesurface-exposed proteins. Trypsin treatment alonedelayed and slightly inhibited PMA-induced O2
– produc-tion but did not prevent it, indicating that trypsin did notremove surface-exposed cytochrome b558; however, theremoval of other surface proteins from PBMCs reducedthe inhibition of PMA-induced O2
– generation byE. chaffeensis (Fig. 2D). This result suggests that inhibi-tion of O2
– production may require specific receptor–ligand interactions between the bacterium and its hostcell.
Fig. 1. Paraquat and H2O2 block the infection of E. chaffeensis andA. phagocytophilum in host cells in a dose-dependent manner. Hostcell-free E. chaffeensis (A) or A. phagocytophilum (B) wasincubated with paraquat or H2O2 (each at 10 mM, 0.1 mM or 1 mM),or a medium control at 37°C for 30 min. After washing,E. chaffeensis or A. phagocytophilum was added to infected hostTHP-1 or HL-60 cells respectively. The number of bacteria in thehost cells was determined at 3 days p.i. Data are presented asmeans and standard deviations of triplicate samples. The asteriskindicates significant difference compared with controls by Student’st-test (P < 0.01).
p22phox degradation in Ehrlichia-infected monocytes 863
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Cellular Microbiology, 9, 861–874

Ehrlichia chaffeensis reduces p22phox, but not gp91phox,in monocytes
To investigate the inhibitory mechanisms of O2– produc-
tion by E. chaffeensis in human monocytes, we first used
flow cytometry to examine the effect of E. chaffeensis onthe amount of plasma membrane-associated cytochromeb558, or its components p22phox and gp91phox. The mono-clonal antibody (mAb) 7D5 was used to detect cyto-chrome b558 on the surface of monocytes, because it
Fig. 2. E. chaffeensis blocks PMA-induced ROS production by human PBMCs and isolated monocytes. Human PBMCs were first incubatedwith luminol for 5 min, then THP-1 lysates or host cell-free E. chaffeensis (EC) were added (arrowhead) (A). Alternatively, human monocytesand neutrophils (PMN) isolated from the same blood specimen were pre-incubated with medium alone (Control), THP-1 lysate, host cell-freeE. chaffeensis (EC) (B, C, E), or A. phagocytophilum (AP) (E) for 5 or 30 min. In D, human PBMCs were pretreated with 0.25% trypsin ormedium control for 15 min prior to the incubation with E. chaffeensis (D). Luminol was added to the mixture and incubated for additional 5 min,then PMA was added (arrows) (B–D). Total ROS generation was continuously measured after luminol addition by LDCL (A–D). (E)SOD-inhibitable, extracellular release of O2
– was measured as DOD550 at 30 min after stimulation with PMA using the cytochrome c reductionassay. OD, optical density. Results shown are representative of three independent experiments in blood specimens derived from differenthealthy donors. The asterisk indicates significant difference compared with controls by Student’s t-test (P < 0.01).
864 M. Lin and Y. Rikihisa
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Cellular Microbiology, 9, 861–874

recognizes an epitope of gp91phox that is exposed on thesurface only when it is complexed with p22phox as a part ofcytochrome b558 (Nakamura et al., 1987; Burritt et al.,2001). The labelling of human monocytes with mAb 7D5showed increased population of monocytes with loweramount of plasma membrane-associated cytochrome b558
after infection with E. chaffeensis for 60 min than in unin-fected monocytes (Fig. 3A). The labelling of total gp91phox
or p22phox showed that the reduction in surface cyto-chrome b558 was due to increased population of mono-cytes with lower amounts of p22phox (Fig. 3B), butmonocytes population did not change with respect togp91phox labelling (Fig. 3C). THP-1 cell lysates used as thecontrol had no effect on the amount of membrane cyto-chrome b558, total p22phox or gp91phox protein (data notshown).
Using Western blot analysis, we further demonstratedthat the incubation of human monocytes withE. chaffeensis for 60 min reduced the amount of p22phox
protein by 59% � 5% (n = 5; P < 0.05) compared with thatin uninfected monocytes or monocytes incubated withuninfected THP-1 cell lysates (Fig. 4A and B). The amountof gp91phox relative to that of a-tubulin, which served as aloading control, was not affected by E. chaffeensis infec-tion (Fig. 4A and C). The amount of p22phox, but notgp91phox, was reduced in monocytes infected withE. chaffeensis with or without PMA stimulation (Fig. 4C).These results confirmed that the rapid decrease in theplasma-membrane cytochrome b558 in E. chaffeensis-infected monocytes was due to a reduction in the amountof p22phox, but not gp91phox. Furthermore, specific cleav-age products of approximately 17 and 15 kDa weredetected with an antibody (Ab) specific for the N-terminusof p22phox and a 20 kDa cleavage product was detectedwith an Ab specific for the C-terminus of p22phox (Fig. 4D),suggesting that the reduction in p22phox is caused bydegradation. The amount of p22phox was also reduced inmonocytes after incubation with E. chaffeensis for 30 min,although the amount reduced was less than those incu-bated for 60 min (data not shown).
Ehrlichia chaffeensis reduces p22phox in monocytemembrane fraction
To investigate whether intracellular signalling events arerequired for the reduction of p22phox protein caused byE. chaffeensis infection, we examined the effect ofE. chaffeensis on the isolated human monocyte mem-brane fraction. The incubation of E. chaffeensis with themonocyte membrane fraction for 30 min reducedmembrane-bound p22phox compared with the untreatedmembrane fraction or monocytes incubated with unin-fected THP-1 cell lysates (Fig. 4E). Furthermore, heat-treated E. chaffeensis did not reduce amounts of p22phox,
Fig. 3. E. chaffeensis infection reduces surface-exposedcytochrome b558 protein and total p22phox, but not gp91phox, in humanmonocytes. Human monocytes (5 ¥ 106 cells/group) in RPMImedium were incubated with THP-1 lysates (monocytes, black line)or host cell-free E. chaffeensis (EC, light grey) at 37°C for 1 h.Samples were then surface-labelled with a mouse IgG1 isotypecontrol (dark grey) or mAb 7D5 and then fixed in 1% PFA (A).Alternatively, cells were fixed in PFA, permeabilized with saponin,and labelled with control rabbit IgG (B, C, dark grey), rabbitanti-p22phox (B), or rabbit anti-gp91phox Abs (C). Samples wereanalysed by flow cytometry. Numbers in the brackets indicate themean fluorescence intensity of the gated areas positively labelledwith mAb 7D5, anti-p22phox or anti-gp91phox. Results shown arerepresentative of three independent experiments.
p22phox degradation in Ehrlichia-infected monocytes 865
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Cellular Microbiology, 9, 861–874
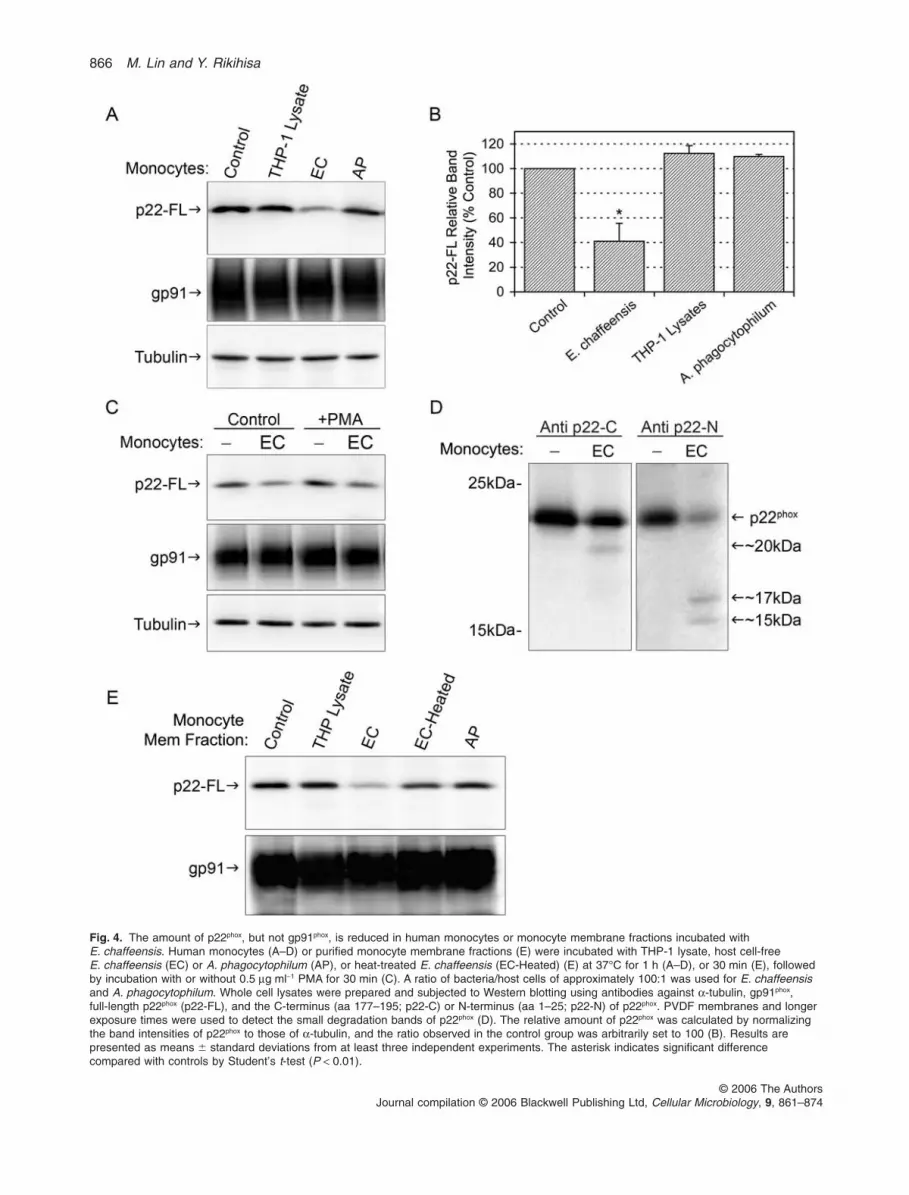
Fig. 4. The amount of p22phox, but not gp91phox, is reduced in human monocytes or monocyte membrane fractions incubated withE. chaffeensis. Human monocytes (A–D) or purified monocyte membrane fractions (E) were incubated with THP-1 lysate, host cell-freeE. chaffeensis (EC) or A. phagocytophilum (AP), or heat-treated E. chaffeensis (EC-Heated) (E) at 37°C for 1 h (A–D), or 30 min (E), followedby incubation with or without 0.5 mg ml-1 PMA for 30 min (C). A ratio of bacteria/host cells of approximately 100:1 was used for E. chaffeensisand A. phagocytophilum. Whole cell lysates were prepared and subjected to Western blotting using antibodies against a-tubulin, gp91phox,full-length p22phox (p22-FL), and the C-terminus (aa 177–195; p22-C) or N-terminus (aa 1–25; p22-N) of p22phox. PVDF membranes and longerexposure times were used to detect the small degradation bands of p22phox (D). The relative amount of p22phox was calculated by normalizingthe band intensities of p22phox to those of a-tubulin, and the ratio observed in the control group was arbitrarily set to 100 (B). Results arepresented as means � standard deviations from at least three independent experiments. The asterisk indicates significant differencecompared with controls by Student’s t-test (P < 0.01).
866 M. Lin and Y. Rikihisa
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Cellular Microbiology, 9, 861–874

suggesting that this effect requires heat-labile factors orproteins from E. chaffeensis (Fig. 4E). The incubation ofhuman monocytes or purified monocyte membrane frac-tions with host cell-free A. phagocytophilum did notreduce the amount of p22phox (Fig. 4A, B and E). Together,these results suggest that an interaction between thebacterium and specific host membrane proteins is neces-sary and sufficient to trigger the downregulation of p22phox
protein.
Lipopolysaccharide (LPS), haemin, or proteaseinhibitors interfere with the inhibition of ROS productionand the reduction of p22phox protein by E. chaffeensis
We previously showed that E. chaffeensis does notpossess genes encoding lipopolysaccharide (LPS)(lipid A) (Lin and Rikihisa, 2003), and the pretreatment ofhuman monocytes with E. coli LPS inhibits E. chaffeensisinfection in human monocytes (Lin and Rikihisa, 2004).LPS itself does not induce a considerable amount of O2
–
generation in neutrophils and monocyte/macrophages,but does prime them for enhanced O2
– generation uponsubsequent stimulation, possibly via enhancing NADPHoxidase holoenzyme assembly (Pabst and Johnston,1980; Guthrie et al., 1984; Cassatella et al., 1990; DeLeoet al., 1998). In agreement with these reports, incubationof PBMCs with E. coli LPS (1 mg ml-1) or addition ofE. chaffeensis to LPS-pretreated PBMCs for 30 min didnot produce measurable amount of O2
– (Fig. 5A), indicat-ing that E. chaffeensis did not have the ability to activateLPS-primed PBMCs. However, LPS-primed PBMCsgenerated more O2
– upon subsequent PMA stimulationthan non-primed PBMCs (Fig. 5B). Furthermore,E. chaffeensis was less effective at inhibiting PMA-induced O2
– production in LPS-primed PBMCs than innon-primed PBMCs (Fig. 5B).
To study whether there is a correlation in ROS pro-duction and the protein amount of p22phox, we used flowcytometry and Western blotting to examine the surfaceexpression of cytochrome b558 and total p22phox proteinamount. Results showed that LPS pretreatment ofhuman monocytes partially prevented both theE. chaffeensis-induced reduction in surface-exposedcytochrome b558 as measured with flow cytometry(Fig. 5C), and the reduction in total p22phox protein asdetected with Western blotting (Fig. 5D). The amount ofgp91phox remained unchanged in all groups (Fig. 5D),and the incubation of human monocytes with LPS alonehad little effect on the amount of surface-expressedcytochrome b558 (Fig. 5C), or the total amount of p22phox
and gp91phox protein (Fig. 5D). These data suggest thatthe inhibitory effects of E. chaffeensis in human mono-cytes can be counteracted by LPS-priming.
We further examined the factors that might be involvedin the downregulation of cytochrome b558 byE. chaffeensis. Cytochrome b558 contains one flavinadenine dinucleotide and two haem prosthetic groups(Nisimoto et al., 1995), and the haem groups play criticalroles in the formation and stability of the gp91phox/p22phox
heterodimer (Yu et al., 1997; DeLeo et al., 2000; Tailleet al., 2004). We therefore tested the effects of exogenoushaemin on the expression of plasma membrane-associated cytochrome b558 in monocytes infected withE. chaffeensis. Pre-incubation with haemin alone did notincrease the labelling of membrane-associated cyto-chrome b558 in human monocytes (Fig. 5C), or the amountof p22phox and gp91phox in monocytes or monocyte mem-brane fractions (Fig. 5D and E). However, haemin pre-treatment diminished the downregulation of membranecytochrome b558 in E. chaffeensis-infected monocytes asmeasured with flow cytometry (Fig. 5C), and the reductionof p22phox in monocytes and the monocyte membranefraction incubated with E. chaffeensis as detected withWestern blot analysis (Fig. 5D and E). The amount ofgp91phox remained unchanged in all treatment groups(Fig. 5D and E).
Previous studies showed that inhibition of haem biosyn-thesis causes p22phox degradation through the ubiquitina-tion and proteasome pathway (DeLeo et al., 2000).However, inhibitors of lysosome acidification or protea-somes have no influence on p22phox degradation inA. phagocytophilum-infected human neutrophils (Mottand Rikihisa, 2000). Similarly, Western blotting using anti-bodies against ubiquitin and p22phox indicated that p22phox
protein in monocytes is not ubiquitinated duringE. chaffeensis infection (data not shown); thus, p22phox ispresumably not degraded through the proteasomepathway that requires protein ubiquitination (Hoch-strasser, 1996). To determine whether the reduction ofp22phox by E. chaffeensis is due to degradation by non-proteasomal proteases, we pretreated human monocytesor monocyte membrane fractions with a cocktail of pro-tease inhibitors with broad specificity for aspartic, cysteineand serine proteases, as well as aminopeptidases. Suchtreatment during incubation with E. chaffeensis com-pletely abolished p22phox degradation (Fig. 5E). As wedetected the cleavage products of p22phox using antibod-ies against the N- and C-termini of p22phox (Fig. 4D), theseresults support that p22phox is cleaved by non-proteasomeproteases.
Reduction of p22phox protein and lack of colocalization ofp22/gp91phox with E. chaffeensis in individual infectedmonocytes
To examine the localization of p22phox and E. chaffeensisin individual human monocytes, we performed
p22phox degradation in Ehrlichia-infected monocytes 867
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Cellular Microbiology, 9, 861–874

laser-scanning confocal microscopy of doubleimmunofluorescence-labelled specimens using specificantibodies against E. chaffeensis and components ofcytochrome b558. We used a bacteria/host cell ratio of 10:1to produce partial infection; this enabled us to compare
individual infected and uninfected monocytes in thesame microscope field. The intensities of p22phox andgp91phox labelling were homogeneous among monocytespopulations in control groups not incubated withE. chaffeensis (Fig. 6A and B). However, the intensity of
868 M. Lin and Y. Rikihisa
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Cellular Microbiology, 9, 861–874

p22phox labelling in E. chaffeensis-infected monocyteswas strongly reduced at 60 min post infection (p.i.)(Fig. 6D–F), or to lesser extent after 30 min p.i. (data notshown), when compared with uninfected monocytes
within the same group incubated with E. chaffeensis. Incontrast, the labelling intensity of gp91phox did not differbetween uninfected and E. chaffeensis-infected individualmonocytes (Fig. 6G–I). Neither p22phox nor gp91phox colo-
Fig. 5. LPS, haemin, or protease inhibitors interfere with the inhibition of ROS production and the reduction of p22phox protein byE. chaffeensis.A. Human PBMCs were incubated with luminol for 5 min, and then LPS was added as indicated by open arrowhead. Alternatively, humanPBMCs were pretreated with LPS for 60 min, luminol was added and incubated for additional 5 min, then THP-1 cell lysates or E. chaffeensis(EC) were added (closed arrowhead). Total ROS generation was continuously measured after luminol addition.B. Human PBMCs were pre-incubated with LPS or medium alone for 60 min, and E. chaffeensis (EC) or THP-1 cell lysates was added andincubated for additional 30 min Luminol was added to the mixture and incubated for additional 5 min, then PMA was added (arrows). TotalROS generation was continuously measured after addition of luminol.C. After human monocytes were pretreated with medium control, LPS or haemin for 60 min, E. chaffeensis (EC) was added and incubated foradditional 60 min. The surface-exposed cytochrome b558 was analysed by flow cytometry using mAb 7D5 or control mouse IgG1. Numbers inthe parentheses indicate the mean fluorescence intensity of the gated areas positively labelled with mAb 7D5.D and E. Human monocytes (D) or human monocyte membrane fractions (E) were pretreated with LPS, haemin, or protease inhibitor cocktailset III (1:100 dilution) at 37°C for 30 min, then incubated with or without E. chaffeensis (EC) for 30 min in the presence of these compounds.Alternatively, E. chaffeensis was pretreated with DMSO control or haemin for 30 min, then incubated with monocyte membrane fractions foradditional 30 min (E). Unless indicated in the figure, a ratio of bacteria/host cell at 100:1 was used for E. chaffeensis. Samples were subjectedto Western blotting with antibodies against full-length p22phox (p22-FL), gp91phox or a-tubulin (monocytes only) (D, E). Results shown arerepresentative of at least three independent experiments in PBMCs or monocytes from different donors.
Fig. 6. LPS blocks p22phox reduction in infected monocytes and induces colocalization of p22phox with E. chaffeensis. Human monocytes werepretreated with or without LPS and then incubated with or without host cell-free E. chaffeensis (EC) at a bacteria/host cell ratio of 10:1 at 37°Cfor 60 min. Samples were either single-labelled with p22phox (A, C) or gp91phox Ab (B), or double-labelled with Abs against E. chaffeensis andp22phox (D–F and J–L) or gp91phox (B, G–I). Images were examined with a laser-scanning confocal microscope. At 1 h p.i. in groups not treatedwith LPS, most E. chaffeensis were located on the surface or in the peripheral cytoplasm and were not colocalized with p22phox (D–F, asteriskin F) or gp91phox (G–I, arrowhead in I). Furthermore, overall labelling of p22phox (D–F), but not that of gp91phox (G–I), was weaker in infectedmonocytes than in uninfected monocytes. In LPS-pretreated monocytes, more E. chaffeensis were found in the perinuclear cytoplasm, andabout 5–10% of E. chaffeensis were colocalized with p22phox (J–L, arrow). Bar: 5 mm.
p22phox degradation in Ehrlichia-infected monocytes 869
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Cellular Microbiology, 9, 861–874

calized with E. chaffeensis by confocal microscopy(Fig. 6D–I), suggesting that NADPH oxidase was notrecruited to E. chaffeensis-containing vacuoles. In agree-ment with the flow cytometry and Western blotting data(Fig. 5), the intensity of p22phox labelling was similar inuntreated or LPS-pretreated uninfected monocytes(Fig. 6A and C). However, the pretreatment of monocyteswith LPS not only partially prevented the reduction in theintensity of p22phox labelling by E. chaffeensis, but alsocaused the colocalization of p22phox with E. chaffeensis(p22phox was found in 5–10% of total E. chaffeensis-containing vacuoles by confocal microscopy) (Fig. 6J–L),suggesting the assembly of NADPH oxidase inE. chaffeensis-containing vacuoles.
Discussion
As an obligate intracellular bacterium that prolifer-ates exclusively in human monocytes/macrophages,E. chaffeensis must subvert the bactericidal abilities oftheir host cells. Given the extreme susceptibilities ofE. chaffeensis to O2
– and H2O2, its ROS-scavenging activ-ity appears insufficient to assure their survival in phago-cytes once host NADPH oxidase is activated. The currentstudy demonstrates that E. chaffeensis downregulatesNADPH oxidase activity: E. chaffeensis not only does notinduce O2
– production by itself, but also rapidly inhibitsPMA-stimulated O2
– production within 30 min of incuba-tion with its host human monocytes. Our result suggeststhat E. chaffeensis employs a similar strategy inA. phagocytophilum as reported by Mott et al. (Mott andRikihisa, 2000; Mott et al., 2002) to subvert ROS-mediated killing: the inhibition of host NADPH oxidaseactivity at the quite proximal site. The degradation ofp22phox occurred in isolated human monocyte membranefractions within 30 min incubation with E. chaffeensis,suggesting that cytoplasmic signalling events or tran-scriptional regulations in host cells are not required.The degradation was also observed in isolated humanneutrophil membrane fractions after incubation withA. phagocytophilum for 30 min (F. Kuribayashi, M. Lin andY. Rikihisa, unpubl. data). The present results alsosuggest that p22phox is likely degraded by protease(s)derived from either the host cell membrane orE. chaffeensis itself. Similarly, in the case of human neu-trophils, p22phox degradation by A. phagocytophilum is notdegraded via host lysosomes or proteasomes (Mott et al.,2002).
Lipopolysaccharide is known to enhance various micro-bicidal activities in monocytes/macrophages, includingphagocytosis, the production of a spectrum of pro-inflammatory cytokines, and NADPH oxidase holoenzymeassembly (DeLeo et al., 1998; 1999; Underhill andOzinsky, 2002). The treatment of human monocytes with
E. coli LPS overrides a number of immunosuppressivecytoplasmic signalling events induced by E. chaffeensis,and as a result inhibits E. chaffeensis infection in vitro (Linand Rikihisa, 2004). NADPH oxidase is known to be acti-vated and assembled on phagosomal membranes duringphagocytosis (DeLeo et al., 1999). Therefore, the colocal-ization of p22phox with intracellular E. chaffeensis inLPS-primed monocytes might be due to altered routeof internalization: phagocytosis instead of receptor-mediated endocytosis (Barnewall et al., 1997; Mott et al.,1999). Only part of E. chaffeensis population was colocal-ized with p22phox, consistent with our ROS assay resultthat LPS-priming cannot completely override anti-ROSability of E. chaffeensis (Fig. 5). It is also possible thatpre-incubation time of monocytes with LPS (1 h) prior toE. chaffeensis addition may not be sufficient for this effect.In addition, the current study showed that LPS preventsE. chaffeensis-induced p22phox degradation. These effectsof LPS may be one reason why E. chaffeensis andA. phagocytophilum lost their genes for LPS biosynthesis(Lin and Rikihisa, 2003). It is thus conceivable that mono-cytes primed in a similar manner in humans and animalsmay be less susceptible to E. chaffeensis infection.
The present data showed that haemin treatmentreduces the degradation of p22phox in human monocytesor the monocyte membrane fraction incubated withE. chaffeensis, suggesting that the haem prosthetic groupor iron might be involved in this degradation process.Haemin treatment also reduced the degradation of p22phox
in human neutrophils and neutrophil membrane fractionincubated with A. phagocytophilum (M. Lin and Y. Riki-hisa, unpubl. data), suggesting the involvement of ashared inhibitory mechanism. Haemin did not directlyreverse the degradation of p22phox through the expressionof additional cytochrome b558, because the surface or totalp22phox protein amount in the monocytes were notchanged by haemin preloading over a short incubationtime (30–60 min). Our results are in agreement with pre-vious reports on the critical role of haem in the productionof O2
– (Yu et al., 1997; DeLeo et al., 2000; Taille et al.,2004). Recently, an intracellular parasite Leishmaniapifanoi amastigote is shown to inhibit macrophage pro-duction of O2
– by inducing monocyte/macrophage haemdegradation and reducing host cytochrome b558 expres-sion, although detailed mechanism remains to be studied(Pham et al., 2005). Given the shared inhibitorymechanisms of ROS-mediated killing, the evolutionarilyconserved proteins between E. chaffeensis andA. phagocytophilum are currently under investigation ascandidates in haemin-reversible host p22phox degradation;analysis of factors from either bacterium could hold thekey for future elucidation of the molecular mechanisms ofROS inhibition in host leucocytes. In addition, futurestudies of host cell membrane-dependent p22phox degra-
870 M. Lin and Y. Rikihisa
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Cellular Microbiology, 9, 861–874

dation by E. chaffeensis and A. phagocytophilum mayshed light on the specific bacterial ligand and host cellreceptor involved in ROS inhibition.
Experimental procedures
Cultivation of E. chaffeensis, A. phagocytophilum andhost cells
Ehrlichia chaffeensis Arkansas strain and A. phagocytophilumHZ strain were cultivated in the human leukaemia cell linesTHP-1 and HL-60 respectively (Barnewall and Rikihisa, 1994;Rikihisa et al., 1997). Cells and bacteria were cultured in RPMI1640 medium supplemented with 10% fetal bovine serum (FBS,endotoxin level < 0.22 ng ml-1) and 2% L-glutamine. No anti-biotics were used throughout the study. The degree of bacterialinfection in host cells was assessed with Diff-Quik staining(Baxter Scientific Products, Obetz, OH) of cytocentrifugedpreparations.
Host cell-free E. chaffeensis or A. phagocytophilum were pre-pared by sonication of heavily infected cells (> 98% infectivity)with a W-380 ultrasonic processor (Heat Systems, Farmington,NY) as described (Lin et al., 2002). Unless otherwise indicated,host cell-free E. chaffeensis isolated from 2 ¥ 106 highly infectedTHP-1 cells was used for infecting 1 ¥ 106 human monocytes,which is equivalent to a ratio of approximate 100 bacteria per cell,which was shown to achieve 100% infection at 2 days p.i. (Linet al., 2002). THP-1 host cell lysates were prepared from thesame number of uninfected cells by the same procedure to serveas negative controls.
Preparation of human peripheral monocytes andneutrophils
Fresh buffy coat (~50 ml) was obtained from healthy volunteers.Following centrifugation at 250 g for 5 min, the plasma wasremoved, and ~10 ml of buffy coat was placed in a 50 ml centri-fuge tube containing 10 ml of Histopaque 1077 overlying 15 ml ofHistopaque 1119 (Sigma, St Louis, MO). After centrifugation at400 g for 30 min, PBMCs were collected from the interface over-lying Histopaque 1077, and neutrophils were collected from theinterface between the Histopaque 1077 and Histopaque 1119.Following hypotonic lysis of erythrocytes in 0.83% NH4Cl for5 min at room temperature, PBMCs or neutrophils were washedtwice and resuspended with Hanks’ balanced salt solution(HBSS) with Ca2+ and Mg2+ and without phenol red (Sigma)supplemented with 2 mg ml-1 dextrose (Abbott Laboratories,North Chicago, IL). Monocytes were separated from PBMCs byallowing them to adhere to 6 well culture plates at 37°C for 1 h.Purified cells contained > 95% neutrophils and > 90% mono-cytes, as assessed morphologically after Diff-Quik staining. Theconcentration of purified cells was determined by counting cells ina haemocytometer.
Isolation of monocyte membrane fraction
Membrane fractions from human peripheral blood monocyteswere prepared as described (Mankelow and Henderson, 2001).Briefly, monocytes in PBS (1 ¥ 108 cells ml-1) were disrupted by
sonication on ice four times for 10–15 s each with a W-380ultrasonic processor at setting 3, and centrifuged at 400 g for10 min at 4°C to remove unbroken cells. The supernatants werefurther centrifuged at 10 000 g for 10 min at 4°C to remove gran-ules and nuclei, and then centrifuged at 130 000 g for 40 min at4°C to obtain the membrane fractions.
Treatments of bacteria with paraquat or H2O2 andevaluation of infection
To determine the susceptibility of E. chaffeensis andA. phagocytophilum to endogenously generated or extracellularlysupplemented ROS, host cell-free bacteria were pretreated at37°C for 30 min with 10–100 mM paraquat (methyl viologen)(Sigma) or 10–100 mM H2O2 (Sigma). Treatment with RPMImedium at same condition served as a negative control. TreatedE. chaffeensis or A. phagocytophilum were washed, collectedand used to infect THP-1 or HL-60 cells respectively. The infectedcells were further cultured for 3 days, and the numbers of organ-isms were estimated in 100 cells each in triplicate culture wells aspreviously described (Barnewall and Rikihisa, 1994).
Measurement of the release of ROS or O2– by LDCL
and ferricytochrome c reduction assays
The release of ROS was measured by LDCL monitored with amodel 1251 luminometer (LKB Wallace, San Francisco, CA), withconstant shaking at 37°C as previously described (Mott andRikihisa, 2000). Briefly, PBMCs or neutrophils were suspendedin HBSS supplemented with 2 mg ml-1 dextrose (HBSSd) at5 ¥ 106 cells ml-1 in Clinicon cuvettes and incubated with orwithout host cell-free E. chaffeensis, A. phagocytophilum orTHP-1 lysate at 37°C for 5 min or 30 min, then PMA (0.5 mg ml-1
final concentration) was added. In some experiments, PBMCswere pretreated with 0.25% trypsin for 15 min in HBSS or1 mg ml-1 LPS from E. coli serotype 0111:B4 (Sigma) for 60 min.Trypsin was inactivated by adding 10% serum and removed bycentrifugation. Luminol (10 mM) was added once to the assaymixture 5 min prior to the addition of THP-1 lysate,E. chaffeensis, LPS or PMA, and LDCL was recorded every 10 sup to 15 or 30 min after the incubation with luminol.
The release of extracellular O2– was detected with a ferricyto-
chrome c reduction assay on the basis of SOD-inhibitable reduc-tion of ferricytochrome c as described (Mott and Rikihisa, 2000).Briefly, purified monocytes were suspended in RPMI medium at aconcentration of 5 ¥ 106 cells ml-1 in a 24 well Corning ultra-lowattachment plate (Fisher, Pittsburgh, PA), and incubated withfreshly prepared THP-1 lysate, host cell-free E. chaffeensis orA. phagocytophilum at a ratio of approximate 100 bacteria percell in triplicate wells for 30 min at 37°C in 5% CO2/95% air. Cellswere washed with HBSS to remove the medium and resus-pended in a 96 well plate containing 200 ml of reaction mixtureper well with 80 mM ferricytochrome c type III from horse heart(Sigma) with or without 50 mg ml-1 SOD from bovine erythrocytes(Sigma). Mixtures were then stimulated by adding PMA(0.5 mg ml-1, final concentration; Sigma) and incubated for 30 minat 37°C in 5% CO2/95% air. The optical density of the superna-tants at 550 nm (OD550) was measured in a microplate spectro-photometer (Molecular Devices, Sunnyvale, CA). The average ofthe OD550 from wells containing only ferricytochrome c was sub-
p22phox degradation in Ehrlichia-infected monocytes 871
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Cellular Microbiology, 9, 861–874

tracted from the individual OD550 of sample wells to calculate theSOD-inhibitable DOD550.
Treatment of human monocytes, neutrophils, membranefractions and host cell-free E. chaffeensis
Monocytes (2 ¥ 106 cells ml-1) in RPMI medium were pretreatedwith 1 mg ml-1 LPS, 10 mM iron-saturated haemin (Sigma), orprotease inhibitor cocktail at a 1:100 dilution for 1 h at 37°C andthen incubated with host cell-free E. chaffeensis at 37°C for 30 or60 min in the presence of these compounds. The Protease Inhibi-tor Cocktail Set III was EDTA-free and contained six proteaseinhibitors with broad specificity for the inhibition of aspartic, cys-teine and serine proteases and aminopeptidases: 100 mMAEBSF, 80 mM aprotinin, 5 mM bestatin, 1.5 mM E-64, 2 mMleupeptin and 1 mM pepstatin A (Calbiochem, San Diego, CA).Protease Inhibitor Cocktail Set II (Calbiochem) contained similarbroad spectrum protease inhibitors including 85 mM EDTA,20 mM AEBSF, 1.7 mM bestatin, 200 mM E-64 and 2 mM pep-statin A. THP-1 cell lysates, prepared similarly to the host cell-free bacterial lysates, served as negative controls. Alternatively,monocyte membrane fractions (equivalent to 1 ¥ 106 cells) orhost cell-free E. chaffeensis in PBS were incubated with haeminor protease inhibitor cocktail at 37°C for 30 min. Heat-treatedE. chaffeensis were prepared by boiling host cell-free bacteria ina water bath for 5 min. Membrane fractions and host cell-freebacteria were then co-incubated for additional 30 min in the pres-ence of these compounds. Samples were then subjected toimmunofluorescence labelling or Western blot analysis. Pretreat-ment of E. chaffeensis, monocytes, neutrophils or membranefraction with 1% DMSO (the solvent in the protease inhibitorcocktail) served as a negative control.
Flow cytometric analysis of plasma membrane-associated cytochrome b558 and total p22phox or gp91phox
Human monocytes were suspended at 2 ¥ 106 cells ml-1 in RPMImedium supplemented with 10% FBS and seeded in 6 wellplates. After pretreatment with LPS (1 mg ml-1) or haemin (10 mM)for 60 min at 37°C, host cell-free E. chaffeensis was added at aratio of approximate 100 bacteria per cell. After a 30 or 60 minincubation, the mixtures were centrifuged in 1.5 ml microcentri-fuge tubes and washed twice with PBS to remove unbound oruninternalized bacteria. Monocytes were labelled with 3 mg ofmAb 7D5 (a kind gift from Dr M. Nakamura) or a mouse IgG1
isotype control, followed by Alexa Fluor 488-conjugated goatanti-mouse secondary Ab, each in 200 ml PBS with 1% FBS;labelling was performed at 4°C for 60 min with rotation. Cellswere fixed in 500 ml of 1% paraformaldehyde (PFA, Sigma) inPBS with 1% FBS and analysed by flow cytometry on a BDFACSCalibur System (BD Biosciences, San Jose, CA). For label-ling of total p22phox or gp91phox, monocytes were first fixed with 2%PFA, permeabilized with 0.3% saponin in PBS, then incubatedwith a rabbit anti-full-length-p22phox Ab (p22-FL, Santa CruzBiotech, Santa Cruz, CA), a rabbit anti-gp91phox Ab (a kind giftfrom S. Tsunawaki), or normal rabbit IgG as a control, in PBScontaining 0.1% gelatin and 0.3% saponin. Samples were thenlabelled with an Alexa Fluor 488-conjugated goat anti-rabbit Aband analysed by flow cytometry.
Sample preparation and Western blot analysis
Following incubation with host cell-free E. chaffeensis orA. phagocytophilum, monocytes or neutrophils were washed andwhole cell lysates were prepared as described (Lin and Rikihisa,2004). The lysates or membrane fractions incubated with hostcell-free E. chaffeensis or A. phagocytophilum were mixed withequal volumes of 2¥ SDS sample buffer and boiled for 5 min.Samples were then subjected to Western blot analysis with rabbitpolyclonal antibodies against full-length p22phox or the N-/C-termini of p22phox (p22-N: polyclonal Ab against p22phox N-terminalamino acids 1–25, a gift from S. Imajoh-Ohmi; p22-C: polyclonalAb against p22phox C-terminal amino acids 177–195, a gift from S.Tsunawaki). An Ab against a-tubulin (Santa Cruz) was used as aloading control. To detect small p22phox cleavage fragments, a15% SDS-polyacrylamide gel and PVDF membrane (Schleicherand Schuell BioScience USA, Keene, NH) were used in theWestern blotting. Otherwise, 12% SDS-polyacrylamide gels andnitrocellulose membranes (Schleicher and Schuell BioScience)were used. A mAb against ubiquitin (Santa Cruz) was used todetect possible ubiquitinated proteins. Reacting bands were visu-alized with enhanced chemiluminescence by incubating themembrane with the LumiGLO™ chemiluminescent reagent(Amersham Biosciences). Images were then captured and den-sitometric analysis was performed using the LAS3000 imagedocumentation system (FUJIFILM Medical Systems USA, Stam-ford, CT). Band intensities were normalized against a-tubulin inthe corresponding samples.
Double immunofluorescence labelling andlaser-scanning confocal microscopy
Human monocytes were incubated with or without host cell-freeE. chaffeensis at 37°C for 30 min or 1 h. A ratio of approxi-mately 10 bacteria per cell was used so that the labelling ofp22phox in uninfected and infected monocytes could be observedwithin the same microscope field. Cells were washed with PBS,fixed in 2% PFA at room temperature for 15 min, and incubatedwith dog anti-E. chaffeensis and rabbit anti-p22phox or gp91phox
antibodies in PBS containing 0.1% gelatin and 0.3% saponin for1 h at room temperature. After washing with PBS, cells wereincubated with Alexa Fluor 555-conjugated goat anti-rabbit(Molecular Probes) and FITC-conjugated goat anti-dog second-ary antibodies (Jackson ImmunoResearch, West Grove, PA) for30 min. Cells were then washed, cytocentrifuged onto glassslides, and fluorescence images were analysed by an LSM 510laser-scanning confocal microscope (Carl Zeiss, Thornwood,NY).
Statistical analysis
Statistical analyses were performed by using by Student’s t-testand P < 0.05 was considered significant.
Acknowledgements
We thank Dr S. Tsunawaki (National Children’s MedicalResearch Center, Tokyo, Japan) for providing the Ab againstgp91phox and the C-terminus of p22phox; Dr S. Imajoh-Ohmi (Uni-versity of Tokyo, Tokyo, Japan) for the Ab against the N-terminus
872 M. Lin and Y. Rikihisa
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Cellular Microbiology, 9, 861–874

of p22phox; and Dr M. Nakamura (Nagasaki University, Nagasaki,Japan) for the mAb 7D5 against cytochrome b558. This work wassupported by Grant R01 AI30010 from the National Institutes ofHealth.
References
Babior, B.M. (1999) NADPH oxidase: an update. Blood 93:1464–1476.
Banerjee, R., Anguita, J., Roos, D., and Fikrig, E. (2000)Cutting edge: infection by the agent of human granulocyticehrlichiosis prevents the respiratory burst by down-regulating gp91phox. J Immunol 164: 3946–3949.
Barnewall, R.E., and Rikihisa, Y. (1994) Abrogation ofgamma interferon-induced inhibition of Ehrlichia chaffeen-sis infection in human monocytes with iron-transferrin.Infect Immun 62: 4804–4810.
Barnewall, R.E., Rikihisa, Y., and Lee, E.H. (1997) Ehrlichiachaffeensis inclusions are early endosomes which selec-tively accumulate transferrin receptor. Infect Immun 65:1455–1461.
Bhattacharya, J., GhoshDastidar, K., Chatterjee, A., Majee,M., and Majumder, A.L. (2004) Synechocystis Fe superox-ide dismutase gene confers oxidative stress tolerance toEscherichia coli. Biochem Biophys Res Commun 316:540–544.
Borjesson, D.L., Kobayashi, S.D., Whitney, A.R., Voyich, J.M.,Argue, C.M., and Deleo, F.R. (2005) Insights into pathogenimmune evasion mechanisms: Anaplasma phagocytophi-lum fails to induce an apoptosis differentiation program inhuman neutrophils. J Immunol 174: 6364–6372.
Burritt, J.B., DeLeo, F.R., McDonald, C.L., Prigge, J.R.,Dinauer, M.C., Nakamura, M., et al. (2001) Phage displayepitope mapping of human neutrophil flavocytochromeb558. Identification of two juxtaposed extracellulardomains. J Biol Chem 276: 2053–2061.
Carlyon, J.A., and Fikrig, E. (2006) Mechanisms of evasion ofneutrophil killing by Anaplasma phagocytophilum. CurrOpin Hematol 13: 28–33.
Carlyon, J.A., Chan, W.T., Galan, J., Roos, D., and Fikrig, E.(2002) Repression of rac2 mRNA expression by Ana-plasma phagocytophila is essential to the inhibition ofsuperoxide production and bacterial proliferation.J Immunol 169: 7009–7018.
Carlyon, J.A., Abdel-Latif, D., Pypaert, M., Lacy, P., andFikrig, E. (2004) Anaplasma phagocytophilum utilizes mul-tiple host evasion mechanisms to thwart NADPH oxidase-mediated killing during neutrophil infection. Infect Immun72: 4772–4783.
Cassatella, M.A., Bazzoni, F., Flynn, R.M., Dusi, S., Trinch-ieri, G., and Rossi, F. (1990) Molecular basis of interferon-gamma and lipopolysaccharide enhancement ofphagocyte respiratory burst capability. Studies on the geneexpression of several NADPH oxidase components. J BiolChem 265: 20241–20246.
Cox, J.A., Jeng, A.Y., Sharkey, N.A., Blumberg, P.M., andTauber, A.I. (1985) Activation of the human neutrophilnicotinamide adenine dinucleotide phosphate (NADPH)-oxidase by protein kinase C. J Clin Invest 76: 1932–1938.
Dahlgren, C., Briheim, G., and Stendahl, O. (1984) Measure-ment of luminol-dependent leukocyte chemiluminescenceoriginated from intracellular and extracellular events. InAnnals of Applied Bioluminescence Chemiluminescence:Proceedings. Kricka, L.J., Stanley, P.E., Thorpe, G.H.G.,and Whitehead, T.P. (eds). New York: Academic Press,pp. 335–338.
DeLeo, F.R., Renee, J., McCormick, S., Nakamura, M.,Apicella, M., Weiss, J.P., and Nauseef, W.M. (1998)Neutrophils exposed to bacterial lipopolysaccharideupregulate NADPH oxidase assembly. J Clin Invest 101:455–463.
DeLeo, F.R., Allen, L.A., Apicella, M., and Nauseef, W.M.(1999) NADPH oxidase activation and assembly duringphagocytosis. J Immunol 163: 6732–6740.
DeLeo, F.R., Burritt, J.B., Yu, L., Jesaitis, A.J., Dinauer, M.C.,and Nauseef, W.M. (2000) Processing and maturation offlavocytochrome b558 include incorporation of heme as aprerequisite for heterodimer assembly. J Biol Chem 275:13986–13993.
Dunning Hotopp, J.C., Lin, M., Madupu, R., Crabtree, J.,Angiuoli, S.V., Eisen, J., et al. (2006) Comparative genom-ics of emerging human ehrlichiosis agents. PLoS Genetics2: e21.
Fang, F.C. (2004) Antimicrobial reactive oxygen and nitrogenspecies: concepts and controversies. Nat Rev Microbiol 2:820–832.
Farr, S.B., and Kogoma, T. (1991) Oxidative stressresponses in Escherichia coli and Salmonella typhimurium.Microbiol Rev 55: 561–585.
Fridovich, I. (1997) Superoxide anion radical (O2-.), super-oxide dismutases, and related matters. J Biol Chem 272:18515–18517.
Guthrie, L.A., McPhail, L.C., Henson, P.M., and Johnston,R.B., Jr (1984) Priming of neutrophils for enhanced releaseof oxygen metabolites by bacterial lipopolysaccharide. Evi-dence for increased activity of the superoxide-producingenzyme. J Exp Med 160: 1656–1671.
Hassan, H.M., and Fridovich, I. (1979) Paraquat and Escheri-chia coli. Mechanism of production of extracellular super-oxide radical. J Biol Chem 254: 10846–10852.
Hassett, D.J., Britigan, B.E., Svendsen, T., Rosen, G.M., andCohen, M.S. (1987) Bacteria form intracellular free radicalsin response to paraquat and streptonigrin. Demonstrationof the potency of hydroxyl radical. J Biol Chem 262:13404–13408.
Heinzen, R.A., Frazier, M.E., and Mallavia, L.P. (1992)Coxiella burnetii superoxide dismutase gene: cloning,sequencing, and expression in Escherichia coli. InfectImmun 60: 3814–3823.
Herron, H.J., and Goodman, J.L. (2001) Evidence that thehuman granulocytic ehrlichiosis agent utilizes a noveloxygen radical detoxification system; cytochrome Cre-oxidation. In American Society for Rickettsiology – Bar-tonella as an Emerging Pathogen Group 2001 JointConference. Big Sky, MN, USA, Abstract �50.
Hochstrasser, M. (1996) Ubiquitin-dependent proteindegradation. Annu Rev Genet 30: 405–439.
IJdo, J., and Mueller, A.C. (2004) Neutrophil NADPH oxidaseis reduced at the Anaplasma phagocytophilumphagosome. Infect Immun 72: 5392–5401.
p22phox degradation in Ehrlichia-infected monocytes 873
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Cellular Microbiology, 9, 861–874

Kim, J.A., Sha, Z., and Mayfield, J.E. (2000) Regulationof Brucella abortus catalase. Infect Immun 68: 3861–3866.
Lin, M., and Rikihisa, Y. (2003) Ehrlichia chaffeensis andAnaplasma phagocytophilum lack genes for lipid A biosyn-thesis and incorporate cholesterol for their survival. InfectImmun 71: 5324–5331.
Lin, M., and Rikihisa, Y. (2004) Ehrlichia chaffeensis down-regulates surface Toll-like receptors 2/4, CD14 and tran-scription factors PU.1 and inhibits lipopolysaccharideactivation of NF-kB, ERK 1/2 and p38 MAPK in hostmonocytes. Cell Microbiol 6: 175–186.
Lin, M., Zhu, M.X., and Rikihisa, Y. (2002) Rapid activation ofprotein tyrosine kinase and phospholipase C-g2 andincrease in cytosolic free calcium are required by Ehrlichiachaffeensis for internalization and growth in THP-1 cells.Infect Immun 70: 889–898.
Mankelow, T.J., and Henderson, L.M. (2001) Inhibition of theneutrophil NADPH oxidase and associated H+ channel bydiethyl pyrocarbonate (DEPC), a histidine-modifying agent:evidence for at least two target sites. Biochem J 358:315–324.
Miller, E.K., and Fridovich, I. (1986) A demonstration thatO2- is a crucial intermediate in the high quantum yieldluminescence of luminol. J Free Radic Biol Med 2: 107–110.
Miller, R.A., and Britigan, B.E. (1997) Role of oxidants inmicrobial pathophysiology. Clin Microbiol Rev 10: 1–18.
Mott, J., and Rikihisa, Y. (2000) Human granulocytic ehrli-chiosis agent inhibits superoxide anion generation byhuman neutrophils. Infect Immun 68: 6697–6703.
Mott, J., Barnewall, R.E., and Rikihisa, Y. (1999) Humangranulocytic ehrlichiosis agent and Ehrlichia chaffeensisreside in different cytoplasmic compartments in HL-60cells. Infect Immun 67: 1368–1378.
Mott, J., Rikihisa, Y., and Tsunawaki, S. (2002) Effects ofAnaplasma phagocytophila on NADPH oxidase compo-nents in human neutrophils and HL-60 cells. Infect Immun70: 1359–1366.
Nakamura, M., Murakami, M., Koga, T., Tanaka, Y., andMinakami, S. (1987) Monoclonal antibody 7D5 raisedto cytochrome b558 of human neutrophils: immuno-cytochemical detection of the antigen in peripheralphagocytes of normal subjects, patients with chronicgranulomatous disease, and their carrier mothers. Blood69: 1404–1408.
Nisimoto, Y., Otsuka-Murakami, H., and Lambeth, D.J.(1995) Reconstitution of flavin-depleted neutrophil flavocy-tochrome b558 with 8-mercapto-FAD and characterizationof the flavin-reconstituted enzyme. J Biol Chem 270:16428–16434.
Ohashi, N., Zhi, N., Lin, Q., and Rikihisa, Y. (2002) Charac-terization and transcriptional analysis of gene clusters for atype IV secretion machinery in human granulocytic andmonocytic ehrlichiosis agents. Infect Immun 70: 2128–2138.
Pabst, M.J., and Johnston, R.B., Jr (1980) Increased produc-tion of superoxide anion by macrophages exposed in vitro
to muramyl dipeptide or lipopolysaccharide. J Exp Med151: 101–114.
Paddock, C.D., and Childs, J.E. (2003) Ehrlichia chaffeensis:a prototypical emerging pathogen. Clin Microbiol Rev 16:37–64.
Pham, N.-K., Mouriz, J., and Kima, P.E. (2005) Leishmaniapifanoi amastigotes avoid macrophage production ofsuperoxide by inducing heme degradation. Infect Immun73: 8322–8333.
Rikihisa, Y. (2003) Mechanisms to create a safe haven bymembers of the family Anaplasmataceae. Annals NY AcadSci 990: 548–555.
Rikihisa, Y. (2006) Ehrlichia subversion of host innateresponses. Curr Opin Microbiol 9: 95–101.
Rikihisa, Y., Zhi, N., Wormser, G.P., Wen, B., Horowitz, H.W.,and Hechemy, K.E. (1997) Ultrastructural and antigeniccharacterization of a granulocytic ehrlichiosis agent directlyisolated and stably cultivated from a patient in New Yorkstate. J Infect Dis 175: 210–213.
Seaver, L.C., and Imlay, J.A. (2001) Alkyl hydroperoxidereductase is the primary scavenger of endogenous hydro-gen peroxide in Escherichia coli. J Bacteriol 183: 7173–7181.
Taille, C., El-Benna, J., Lanone, S., Dang, M.C., Ogier-Denis,E., Aubier, M., and Boczkowski, J. (2004) Induction ofheme oxygenase-1 inhibits NAD(P)H oxidase activity bydown-regulating cytochrome b558 expression via thereduction of heme availability. J Biol Chem 279: 28681–28688.
Underhill, D.M., and Ozinsky, A. (2002) Phagocytosis ofmicrobes: complexity in action. Annu Rev Immunol 20:825–852.
Wang, T., Malawista, S.E., Pal, U., Grey, M., Meek, J.,Akkoyunlu, M., et al. (2002) Superoxide anion productionduring Anaplasma phagocytophila infection. J Infect Dis186: 274–280.
Williams, N.M., Cross, R.J., and Timoney, P.J. (1994) Res-piratory burst activity associated with phagocytosis ofEhrlichia risticii by mouse peritoneal macrophages. ResVet Sci 57: 194–199.
Yu, L., Zhen, L., and Dinauer, M.C. (1997) Biosynthesis of thephagocyte NADPH oxidase cytochrome b558. Role ofheme incorporation and heterodimer formation in matura-tion and stability of gp91phox and p22phox subunits. J BiolChem 272: 27288–27294.
Supplementary material
The following supplementary material is available for this articleonline:Table S1. Genes for defence against oxidative killing in the orderRickettsiales, other obligate intracellular bacteria, selected facul-tative intracellular bacteria of monocytes/macrophages, andE. coli.
This material is available as part of the online article from http://www.blackwell-synergy.com
874 M. Lin and Y. Rikihisa
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Cellular Microbiology, 9, 861–874