deep-water agglutinated foraminiferal changes and stable isotope profiles across the...
TRANSCRIPT

Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
Deep-water agglutinated foraminiferal changes and stable isotopeprofiles across the Cretaceous–Paleogene boundary in the
Rotwandgraben section, Eastern Alps (Austria)
Danuta Peryt a,*, Roman Lahodynsky b, Tomasz Durakiewicz ca Institute of Paleobiology, Polish Academy of Sciences, 00-818 Warszawa, Poland
b Institut fur Geologie und Palaontologie, Universitat Innsbruck, Innrain 52, A-6020 Innsbruck, Austriac Institute of Physics, Maria Curie-Sklodowska University, Plac Marii Skłodowskiej-Curie 1, 20-031 Lublin, Poland
Received 8 March 1995; accepted 19 March 1997
Abstract
The studied interval extends from 2.5 m below to 1 m above the Cretaceous–Paleogene (K–P) boundary andencompasses the uppermost Abathomphalus mayaroensis, PO (subzones: POa — Guembelitria cretacea and POb —Globoconusa conusa), Pa or Parvularugoglobigerina eugubina and lowermost P1 (subzone: P1a or Subbotina pseudobul-loides) planktonic foraminiferal zones.
The agglutinated benthic foraminiferal assemblages of the uppermost Abathomphalus mayaroensis Zone aremoderately diversified and composed of mixed epifaunal and infaunal morphogroups. In the lower part of theGuembelitria cretacea Subzone benthic foraminifers are extremely rare and represented only by a few epifaunal species;in the upper part of this subzone they are still not abundant but assemblages are again moderately diversified. In theGloboconusa conusa Subzone deep-water agglutinated foraminifers are abundant, highly diversified and representedby mixed epifaunal and infaunal morphogroups.
Bulk rock stable isotope analysis shows a significant drop of both d13C and d18O values at the K–P boundary.It appears that a drastic decrease in abundance and diversity of deep-water agglutinated foraminifers as well as a
change of the community structure from mixed epifaunal and infaunal to one dominated by epifaunal morphogroupsare related to a reduction in marine primary productivity.
The instantaneous environmental stress and the collapse of primary productivity, are compatible with a large bolideimpact. © 1997 Elsevier Science B.V.
Keywords: Cretaceous–Paleogene boundary; agglutinated foraminifera; stable isotopes; paleoecology
1. Introduction events to a greater or lesser degree. While plank-tonic forms suffered almost complete extinction(only a few species survived) (Au-Liu and Olsson,One of the greatest mass extinctions took place1994; Longoria and Gamper, 1995), benthic formsat the Cretaceous–Paleogene (K–P) boundary.were not so severely affected by the biotic crisis,Foraminifera were affected by K–P boundaryat least in terms of extinction ( Keller, 1988; Zachoset al., 1989; Thomas, 1990; Kaiho, 1992; Widmark* Corresponding author. Tel.: 00-48-22-6978795;
e-mail: [email protected] and Malmgren, 1992; Kuhnt and Kaminski, 1993;
0031-0182/97/$17.00 © 1997 Elsevier Science B.V. All rights reserved.PII S0031-0182 ( 97 ) 00056-4
288 D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
Coccioni and Galeotti, 1994; Speijer and Van der depth of the basin, benthic foraminiferal assem-blages were to a lesser degree affected by the K–PZwaan, 1994).
It has been observed that the survival rate across boundary event(s) (e.g., Thomas, 1990).Causes of extinction and/or restructuring ofthe K–P boundary among various groups of
marine organisms depended upon the environment benthic foraminiferal assemblages across the K–Pboundary has been explained in different ways. Ininhabited (Hansen et al., 1987). With increasing
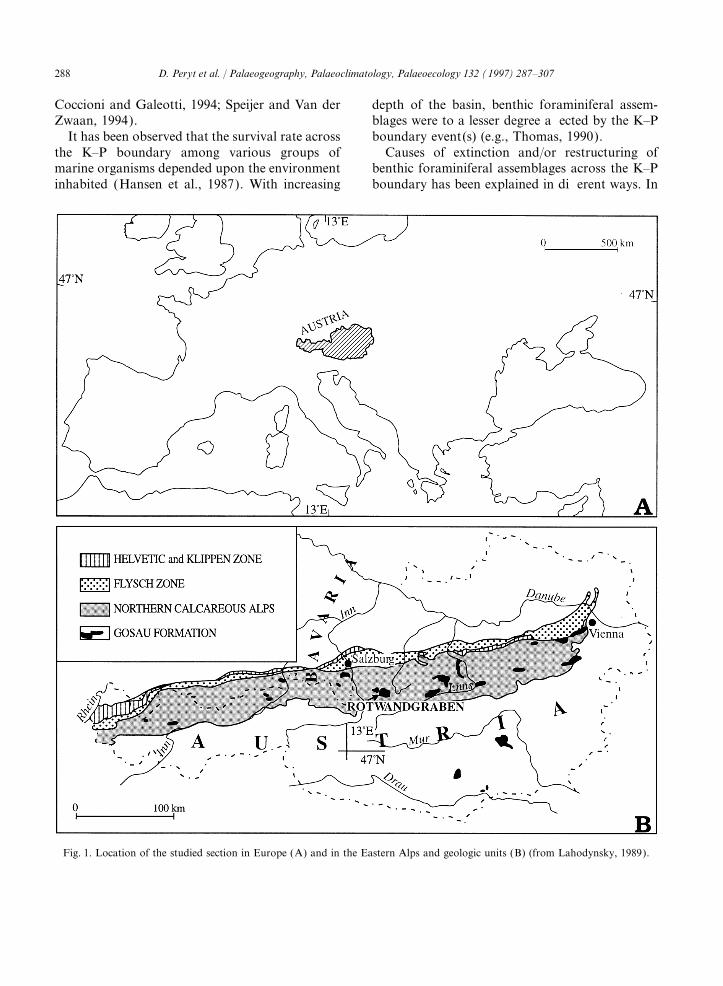
Fig. 1. Location of the studied section in Europe (A) and in the Eastern Alps and geologic units (B) (from Lahodynsky, 1989).

289D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
most of the hypotheses, causes of the K–P benthicforaminiferal turnover were directly or indirectlylinked with an extraterrestrial asteroid impact, e.g.a sudden temperature rise due to the impact of anextraterrestrial body into the ocean (Emiliani et al.,1981); a collapse of primary productivity as aresult of the instantaneous environmental stresswhich may be brought about by a bolide impact(Zachos et al., 1989; Thomas, 1990; Kaiho, 1992;Widmark and Malmgren, 1992; Kuhnt andKaminski, 1993); and a cooling event at the K–Pboundary, developing after the impact of an extra-terrestrial object (Speijer and Van der Zwaan,1994). Terrestrial causes were given by Keller(1988) to explain benthic foraminiferal turnoveracross the K–P boundary. Keller (1988) postulateda major drop in sea level and oxygen contentduring the earliest Paleocene to explain theobserved change in the benthic foraminiferalassemblages in the shallow-water El Kef section(Tunisia). Coccioni and Galeotti (1994) suggestedlow oxygen conditions to elucidate changes in themorphotypic structure of benthic foraminiferalassemblages in a middle bathyal environment atCaravaca (southeast Spain) at the K–P boundary.
The purpose of this paper is to trace the responseof deep-water agglutinated foraminifers assem-blages to the K–P transition event(s) in theRotwandgraben section and an attempt to explainthe relation between changes in foraminiferalassemblages and abiotic events at the K–Pboundary.
2. Material and methods
A total of 22 samples were micropaleontologi-cally and geochemically analyzed from thesection 2.5 m below to 1 m above the K–P bound-ary. The section was sampled at 20–50-cm intervalsbelow the K–P boundary and continuous samplingwas done at 1–2-cm intervals for the first 15 cmabove the boundary, then at 10-cm intervals
Fig. 2. Lithological column and stratigraphy across the K–Pboundary in the Rotwandgraben section (from Peryt et al.,1993).

290 D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
between 15 and 35 cm and at 20-cm intervals system, consisting of the Austro-Alpine, Flyschand Helvetic nappes.between 35 and 100 cm. Samples were processed
using glauber salt. An aliquot of about 200 speci- The Gosau basins were developed on a tectoni-cally mobile Triassic to Jurassic carbonate plat-mens from the >100-mm size fraction was used
for the faunal analyses. form that advanced northwards since the EarlyCretaceous and evolved into a submarine plateauThe taxonomy of deep-water agglutinated fora-
minifers (DWAF) adopted in this study follows with trench-like basins during the Late Cretaceous.These basins were loci of turbidite sedimentationthe systematics of Kaminski et al. (1988), Kuhnt
(1990), Geroch and Kaminski (1992), Kaminski during the Campanian to Paleocene (Hesse andButt, 1976; Butt, 1981; Preisinger et al., 1986).and Geroch (1993), and Kuhnt and Kaminski
(1993). Therefore, detailed taxonomic descriptions The K–P transition in the Rotwandgraben sec-tion is contained within the Zwieselalm Beds whichare not given here.
Stable carbon and oxygen determinations of range from Upper Maastrichtian to UpperPaleocene in the Gosau area. The Zwieselalm Bedsthese 22 samples and 3 additional ones were per-
formed on carbon dioxide with a modified MI-1305 consist of interbedded sandstones, siltstones, lime-stones, marly limestones and marls, which aremass spectrometer (Hałas, 1979; Hałas and
Skorzynski, 1980) with a triple-collector detection partly turbiditic and partly hemipelagic (Fig. 2).The sequence of Maastrichtian and Paleocenesystem. The gas was extracted from samples in a
vacuum line by decomposition of the samples with sediments in the Gosau Basin indicates sedimenta-tion in a deep-sea environment above the CCD100% phosphoric acid at 25°C (McCrea, 1950).
The results are expressed in per mil (‰) relative (Butt, 1981; Preisinger et al., 1986; Lahodynsky,1989). The preservation of foraminiferal tests into the PDB international standard.the Rotwandgraben section suggests depositionabove the CCD, but probably at and/or below theforaminiferal lysocline at a depth of about 2000 m3. Geological setting(Peryt et al., 1993).
K–P transitions containing a boundary clayand geochemical anomalies are exposed in theEastern Alps only within the flyschoid Upper 4. Biostratigraphy and deep-water agglutinated
foraminiferal assemblages (Plates I–IV )Campanian–Lower Eocene Upper Complex of theGosau Beds (Herm et al., 1981; Stradner et al.,1985, 1987; Preisinger et al., 1986; Lahodynsky, The following planktonic foraminiferal zones
have been distinguished in the studied section:1988, 1989; Stradner and Rogl, 1988; Peryt et al.,1993). Abathomphalus mayaroensis, PO with two sub-
zones: POa or Guembelitria cretacea and POb orThe studied section is located in the Gosau area(Fig. 1) in the Northern Calcareous Alps which is Globoconusa conusa, Pa or Parvularugoglobigerina
eugubina and P1 with P1a or Subbotina pseudobul-the southernmost of five major paleogeographiczones of the Eastern Alps — a complex nappe loides Subzone (Fig. 2) (Peryt et al., 1993).
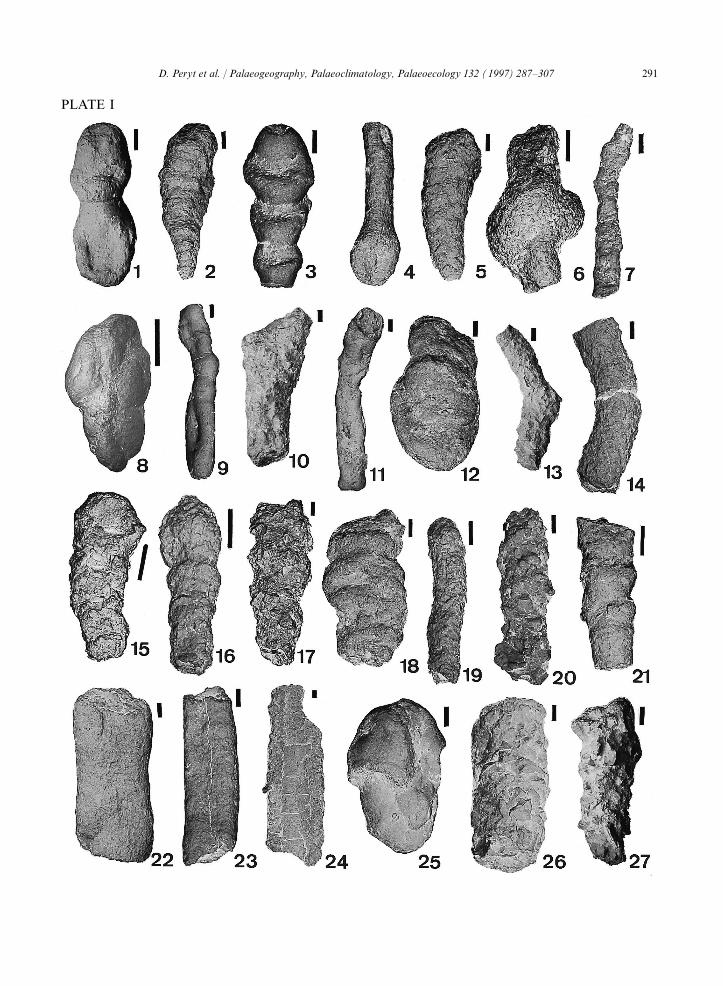
Plate I.
1=Hormosina velascoensis (Cushman), R21; 2=Pseudobolivina sp., R6; 3=Hormosina velascoensis (Cushman), R22; 4=Hyperamminaelongata Brady, R18; 5=Textularia sp., R2; 6=Glomospira irregularis (Grzybowski), R4; 7=Bathysiphon sp., R17; 8=Trochammi-noides contortus (Grzybowski), R18; 9=Kalamopsis grzybowskii (Grzybowski), R16; 10=Dendrophrya excelsa Grzybowski, R15;11=Bathysiphon sp., R19; 12=Paratrochamminoides sp., R2; 13=Rhabdammina sp., R8; 14=Rhabdammina sp., R13; 15=Subreophaxscalaris (Grzybowski), R20; 16=Reophax pilulifer Brady, R16; 17=Subreophax scalaris (Grzybowski), R20; 18=Subreophax guttifer(Brady), R1; 19=Rhabdammina cylindrica Glaessner, R16; 20=Rhizammina indivisa Brady, R16; 21=Bathysiphon sp., R6; 22=Bathysiphon gerochi Myatlyuk, R13; 23=Bathysiphon gerochi Myatlyuk, R16; 24=Dendrophrya robusta Grzybowski, R13; 25=Paratrochamminoides sp., R1; 26=Rhizammina indivisa Brady, R16; 27=Rhizammina indivisa Brady, R22. Scale bar=100 mm.
291D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
PLATE I

292 D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
PLATE II

293D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
More than fifty species of DWAF have been faunal and geochemical changes are related tothis interval.identified in the studied interval (Table 1; Fig. 3).
In the lowermost 3-cm-thick layer of sediment(samples 10–12) the abundance of DWAF species4.1. Abathomphalus mayaroensis Zonedecreases drastically. Only single specimens of afew species such as Ammodiscus cretaceus,DWAF assemblages from the uppermost part
of the planktonic foraminifer Abathomphalus may- Kalamopsis grzybowskii, Bathysiphon sp.,Glomospira irregularis, Recurvoides sp. andaroensis Zone are moderately diverse — the
number of species varies from 10 to 25 (Figs. 3 Marssonella oxycona were found (Fig. 3). Most ofthe other latest Maastrichtian species that disap-and 4). The assemblage is dominated by tubular
and branching forms of astrorhizids (Fig. 4), peared the K–P boundary in Rotwandgrabenrepresent mainly Lazarus species. Most of themi.e. Bathysiphon, Dendrophrya, Rhizammina,
Rhabdammina, Kalamopsis. Important contrib- return soon to the area, some others migrated toshallower environments.utors to the assemblages are also Trochammina,
Recurvoides, Dorothia, Marssonella, and Spiro- In the upper part of the subzone (samples 10–16)the DWAF density in the sediment is still very low,plectammina. Ammodiscus, Glomospira, Paratro-
chamminoides, Tritaxia, and Trochamminoides are but diversity increases. The number of speciesincreases to 12–17 (Figs. 3 and 4). Tubular andcommon.
Several DWAF species present in the uppermost branching forms of astrorhizids and trochamminidsconstitute 80–95% of the assemblage in this interval.Maastrichtian in the studied section have not been
recorded in the Lower Paleocene. These species The first Lazarus species to return are Trochamminasp., Saccammina grzybowskii, Reophax spp.,are: Psammosphaera scruposa, Trochamminoides
variolarius, Lituola lata, Gaudryina pyramidata, Eggerellina sp., Dendrophrya robusta, Rhizamminaindivisa. Several species also appear which were notCribrostomoides sp., Clavulinoides eggeri,
Subreophax guttifer, Textularia sp., Rzehakina epi- present in the uppermost Maastrichtian: Nothiaexcelsa, Glomospira gordialis, Subreophax scalaris,gona, Haplophragmoides sp., Dorothia bulletta.
However, at least some of the species from this Hyperammina elongata, Saccammina placenta,Trochammina globigeriniformis. These species eithergroup survived in other regions, e.g. Gaudryina
pyramidata, Clavulinoides eggeri, Psammosphaera were so rare in the pre-K–P boundary assemblagesthat they were not recorded in the studied samplesscruposa were recorded in the lowermost Paleocene
of the Biscay region (Kuhnt and Kaminski, 1993). or immigrated from other areas because of morehospitable environments.
4.2. PO zone — POa or Guembelitria cretaceaSubzone 4.3. PO zone — POb or Globoconusa conusa
SubzoneIn the studied section the interval is 10 cm thick
and consists of clay with a 1-cm-thick sandstone The interval is 50 cm thick and consists ofintercalated fine-grained sandstones and claysintercalation (Figs. 2 and 3). The zone begins just
above a 0.5–1-cm-thick rusty layer. The greatest (Figs. 2 and 3). When compared to the
Plate II.
1=Lituola lata (Grzybowski), R20; 2=Rzehakina fissistomata (Grzybowski), R18; 3=Rzehakina epigona (Rzehak), R20; 4=Trochamminoides subcoronatus (Grzybowski), R1; 5=Haplophragmoides walteri (Grzybowski), R20; 6=Hormosina gigantea Geroch,R22; 7=Saccamina placenta (Grzybowski), R22; 8=Hormosina ovulum (Grzybowski), R21; 9=Ammodiscus peruvianus Berry, R19;10=Glomospirella grzybowskii (Jurkiewicz), R17; 11=Trochamminoides variolarius (Grzybowski), R1; 12=Trochamminoides subcoro-natus (Grzybowski), R1; 13=Ammodiscus cretaceus (Reuss), R3; 14=Psammosphaera scruposa (Berthelin), R9; 15=Saccaminagrzybowskii (Schubert), R13. Scale bar=100 mm.

294 D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
PLATE III

295D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
Guembelitria cretacea Subzone, a considerable rounded trochospiral, plano-convex trochospiral,milioline and biconvex trochospiral morphotypes,increase in foraminiferal density in the sediment is
observed. The number of species in DWAF assem- while an infaunal mode of life prevails withinrounded planispiral, flattened ovoid, tapered andblages increases, and the next Lazarus taxa —
spiroplectamminids and rzehakinids — returned. cylindrical, spherical, flattened tapered morpho-types. However, this simple classification does notAstrorhizids, trochamminids and spiroplectam-
minids dominate the assemblages in this interval apply to mobile species which are living free onthe muddy sediment surface. Those may penetrate(Figs. 3 and 4).down into the unconsolidated mud in search forfood. Alternatively, they may move within the4.4. Pa or Parvularugoglobigerina eugubina Zone
and P1a or Subbotina pseudobulloides Subzone sediment because individual taxa may prefer sucha microhabitat which depends on the depth of theredox front (Jorissen, 1988; Barmawidjaja et al.,The interval is about 0.5 m thick and consists
of intercalated calcareous marlstones, clays and 1992).Therefore, the distribution of benthic foramini-sandstones (Figs. 2 and 3). The number of DWAF
species varies from 12 to 17. The taxonomic com- fers may vary to some degree in response tochanging environmental conditions and foodposition of DWAF assemblages increases by the
return of additional Lazarus species: Dorothia supply.The sea-floor organic flux is responsible forretusa, D. crassa, Spiroplectammina spectabilis
(Fig. 3). Several species also appear which are not both: food supply and oxygen concentration withinsediment pore waters. Infaunal species dominatepresent in the older sediments: Tritaxia sp.,
Dorothia sp., Arenobulimina sp., Arenobulimina in assemblages associated with relatively highorganic-carbon fluxes (Corliss and Chen, 1988),dorbignyi, Glomospira charoides, Spiroplectammina
israelskyi, Spiroplectammina sp., Hormosina gigan- and epifaunal ones in nutrient-poor environments(Gooday, 1983, 1994; Thomas, 1990).tea (Fig. 3). Astrorhizids and trochamminids
along with species of Spiroplectammina, Dorothia Jones and Charnock (1986) proposed a classifi-cation of agglutinated foraminifers based on theirand Marssonella dominate assemblages of this
interval (Fig. 4). life positions and feeding habits by distinguishingfour main morphogroups:
A, B and D — species with epifaunal lifeposition;5. Paleoecological response of DWAF to the K–P
boundary event A — primarily suspension feeders;B — passive and active herbivores, detrivores
and omnivores;Environmental requirements of Recent benthicforaminifers have been subject of many studies D — herbivores;
C — species with infaunal life position, mainlyand a broad classification of depth-related mor-photype associations has been proposed (Corliss, detrital/bacterial scavengers.
Table 2 shows how the agglutinated taxa1985; Corliss and Chen, 1988).An epifaunal mode of life is the most typical for recorded in the studied section are distributed
Plate III.
1=Spiroroplectammina israelskyi Frizzel, R22; 2=Spiroroplectammina dentata (Alth), R18; 3=Spiroroplectammina dentata (Alth),R18; 4=Spiroroplectammina sp., R18; 5=Spiroroplectammina israelskyi Frizzel, R19; 6=Dorothia bulletta (Carsey), R4; 7=Clavuli-noides eggeri (Cushman), R1; 8=Clavulinoides eggeri (Cushman), R1; 9=Tritaxia tricarinata (Reuss), R20; 10=Tritaxia sp., R20;11=Dorothia retusa (Cushman), R22; 12=Spiroplectammina spectabilis (Grzybowski), R20; 13=Dorothia sp., R20; 14=Dorothiabulletta (Carsey), R1; 15=Arenobulimina dorbignyi (Reuss), R20; 16=Dorothia crassa (Marsson), R5; 17=Arenobulimina dorbignyi(Reuss), R20; 18=Arenobulimina dorbignyi (Reuss), R20; 19=Paratrochamminoides sp., R7. Scale bar=100 mm.
296 D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
PLATE IV

297D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
among the various morphogroups established by Assemblage II ( lower part of the GuembelitriaJones and Charnock (1986). cretacea Subzone)
Four assemblages (I–IV ) can be recognized in In this interval benthic foraminifers are almostthe studied section related to changes in the relative completely absent. The few species found here areabundances of epifaunal and infaunal species epifaunal suspension-feeders (Kalamopsis grzybow-(Figs. 4 and 5). skii, Bathysiphon sp.) and deposit-feeders
(Ammodiscus cretaceus, A. peruvianus, Recurvoides5.1. Assemblage I (uppermost sp.), and the shallow infaunal detrital/bacterialAbathomphalus mayaroensis Zone) scavenger (Marssonella oxycona) (Figs. 4 and 5).
This assemblage reflects a collapse of the foodAssemblage I is composed of mixed epifaunal supply to the sea bottom.
and infaunal morphogroups, where epifaunal taxaconstitute 53–75% of the assemblage. The epifau-nal morphogroup is dominated by erect tubular 5.2. Assemblage III (upper part of theand branching taxa, primarily suspension-feeders Guembelitria cretacea Subzone)(Bathysiphon, Dendrophrya, Rhizammina,Rhabdammina, Kalamopsis), and trochospirally Assemblage III is composed almost entirely ofenrolled detritus feeders (Trochammina and epifaunal forms. They make up 80–95% of theRecurvoides). Within the epifaunal morphogroup, assemblage. The most common are suspension-tubular irregularly arranged taxa (Glomospira, feeding astrorhizids and trochamminids (Figs. 4Paratrochamminoides) and planispirally enrolled
and 5) with a most probably omnivorous feedingattached species (mainly Ammodiscus cretaceus)
habit.are also common; both groups are interpreted asThis assemblage indicates that very little food isactive deposit-feeders. Within the morphogroup
available on the sea bottom.with inferred infaunal mode of life Dorothia,Marssonella and Spiroplectammina are most abun-dant in the lower part (samples 1 and 2). Higher
5.3. Assemblage IV (Globoconusa conusa,in the zone Dorothia and Marssonella expand,Parvularugoglobigerina eugubina Zone andwhile Spiroplectammina decreases in abundanceSubbotina pseudobulloides Subzone)(Fig. 4). This group of species possesses tests with
calcareous cement and has represents a passiveEpifaunal forms make up 53–68% of the assem-detritus or bacterial scavenging feeding habit.
blage and are represented mainly by astrorhizidsAssemblage I is typical for deep-water marineand trochamminids. Within infaunal morpho-environments with normal primary productivitygroups, the spiroplectamminids dominate alongand a flux of organic detritus and calcium carbon-with Dorothia and Marssonella (Figs. 4 and 5). Aate that is sufficient to sustain infaunal bottom-significant presence in the assemblage of formsdwelling organisms.living within the sediment suggests a return toEpifaunal suspension-feeders have best adaptednormal pre-K–P boundary conditions at lowerto rapidly utilize food and they dominate the
assemblages. bathyal–abyssal depths.
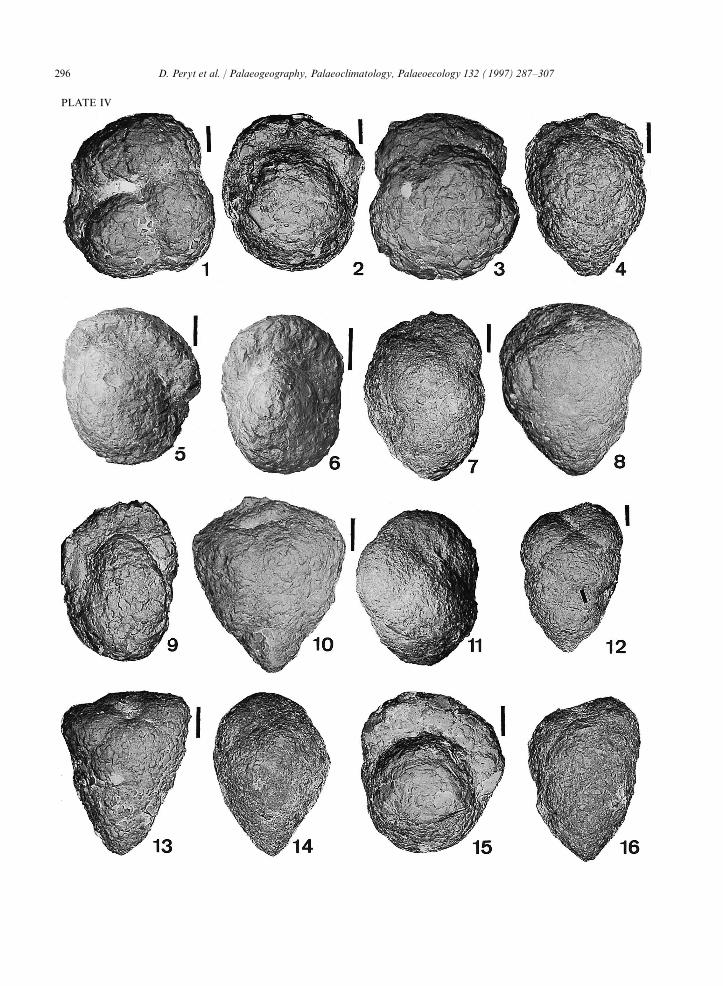
Plate IV.
1=Trochammina globigeriniformis Cushman, R15; 2=Haplophragmoides suborbicularis (Rzehak), R4; 3=Trochammina globigerini-formis Cushman, R16; 4=Arenobulimina dorbignyi (Reuss), R20; 5=Recurvoides sp., R18; 6=Recurvoides sp., R6; 7=Arenobuliminadorbignyi (Reuss), R20; 8=Arenobulimina dorbignyi (Reuss), R9; 9=Recurvoides sp., R18; 10=Marssonella oxycona (Reuss), R13;11=Arenobulimina sp., R20; 12=Arenobulimina dorbignyi (Reuss), R2; 13=Marssonella oxycona (Reuss), R15; 14=Arenobuliminadorbignyi (Reuss), R20; 15=Cribrostomoides sp., R4; 16=Arenobulimina dorbignyi (Reuss), R20. Scale bar=100 mm.

298 D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
Table 1Taxonomic citations of DWAF (in alphabetic order) recorded in the Rotwandgraben section
Ammodiscus cretaceus (Reuss) = Operculina cretacea ReussAmmodiscus peruvianus BerryArenobulimina dorbignyi (Reuss) = Bulimina d’orbignyi ReussArenobulimina sp.Bathysiphon gerochi MyatlyukBathysiphon sp.Clavulinoides eggeri (Cushman) = Pseudoclavulina eggeri CushmanCribrostomoides sp.Dendrophrya robusta GrzybowskiDorothia bulletta (Carsey) = Gaudryina bulletta CarseyDorothia crassa (Marsson) = Gaudryina crassa MarssonDorothia retusa (Cushman) = Gaudryina retusa CushmanDorothia sp.Eggerellina sp.Gaudryina pyramidata CushmanGlomospira charoides (Jones and Parker) = Trochammina squamata Jones and ParkerGlomospira gordialis (Jones and Parker) = Trochammina squamata Jones and Parker var. gordialis Jones and ParkerGlomospira irregularis (Grzybowski) = Ammodiscus irregularis GrzybowskiGlomospirella grzybowskii (Jurkiewicz) = Glomospira irregularis JurkiewiczHaplophragmoides suborbicularis (Rzehak) = Cyclammina suborbicularis RzehakHaplophragmoides walteri (Grzybowski) = Trochammina walteri GrzybowskiHaplophragmoides sp.Hormosina gigantea GerochHormosina ovulum (Grzybowski) = Reophax ovulum GrzybowskiHormosina velascoensis (Cushman) = Nodosinella velascoensis CushmanHyperammina elongata BradyKalamopsis grzybowskii (Grzybowski) = Hyperammina grzybowskii DylazankaLituola lata (Grzybowski) = Ammodiscus latus GrzybowskiMarssonella oxycona (Reuss) = Gaudryina oxycona ReussNothia excelsa (Grzybowski) = Dendrophrya excelsa GrzybowskiParatrochamminoides sp.Psammosphaera scruposa (Berthelin) = Haplophragmium scruposum BerthelinPseudobolivina sp.Recurvoides sp.Reophax pilulifer BradyRhabdammina cylindrica GlaessnerRhabdammina sp.Rhizammina indivisa BradyRzehakina fissistomata (Grzybowski) = Spiroloculina fissistomata GrzybowskiRzehakina epigona (Rzehak) = Silicina epigona RzehakSaccamina grzybowskii (Schubert) = Reophax grzybowskii SchubertSaccamina placenta (Grzybowski) = Reophax placenta GrzybowskiSpiroroplectammina dentata (Alth) = Textularia dentata AlthSpiroroplectammina israelskyi FrizzelSpiroplectammina spectabilis (Grzybowski) = Spiroplecta spectabilis GrzybowskiSpiroplectammina sp.Subreophax guttifer (Brady) = Reophax guttifer BradySubreophax scalaris (Grzybowski) = Reophax scalaris GrzybowskiTextularia sp.Tritaxia tricarinata (Reuss) = Tritaxia tricarinata ReussTritaxia sp.Trochammina globigeriniformis CushmanTrochammina sp.Trochamminoides contortus (Grzybowski) = Trochammina contorta GrzybowskiTrochamminoides subcoronatus (Grzybowski) = Trochammina subcoronata GrzybowskiTrochamminoides variolarius (Grzybowski) = Trochammina variolaria Grzybowski

299D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
Fig. 3. Distribution of deep-water agglutinated foraminifers in uppermost Maastrichtian and lowermost Paleocene deposits of theRotwandgraben section. Lithologies and other symbols are explained in Fig. 2.

300 D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
Fig. 4. Changes in DWAF assemblages across the K–P boundary at Rotwandgraben.A. Morphotypic assemblages.B. DWAF density (number of specimens per g of sediment).C. DWAF richness (number of species).D–G. Relative abundances of dominant agglutinated genera.H. Proportion of epifaunal and infaunal morphogroups within DWAF assemblages.

301D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
Table 2 because isotopic trends may be influenced to aList showing how the agglutinated taxa are interpreted to be large extent by relative amounts of different sedi-distributed among the various epifaunal and infaunal morpho- ment components and it is also difficult to assessgroups (classification after Jones and Charnock, 1986)
to what degree isotopic signals are original ordiagenetically altered.INFERRED MODE OF LIFE
According to Perch-Nielsen et al. (1982) isotopeEPIFAUNAL: profiles from the fine-fraction are similar, although
not identical to those from whole-rock samples.MORPHOGROUP A:
Because of the paucity of calcareous foramini-fers in the lowermost Paleocene and because ofRhabdammina spp.the very low carbonate content in this part of theDendrophrya robusta
Rhizammina indivisa section isotopic analyses were performed on whole-Kalamopsis grzybowskii rock samples only.Bathysiphon spp.Hyperammina elongata
6.1. Stable carbon isotopes
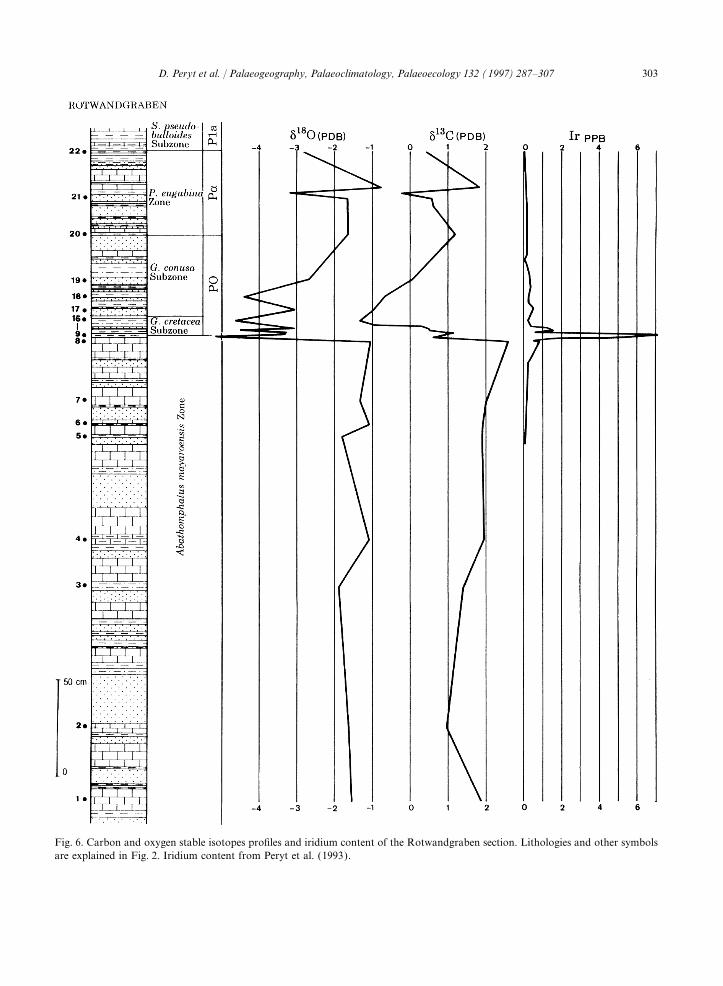
MORPHOGROUP B:The d13C values fluctuate significantly across the
B1: K–P boundary interval in the RotwandgrabenSaccamina grzybowskii section. The most characteristic feature for theSaccammina placenta K–P boundary d13C curve, the negative excursionPsammosphaera scruposa in the earliest Paleocene, is recorded here as wellNothia excelsa
(Fig. 6). The general shape of the d13C curve inthe Rotwandgraben section is very similar to theB2:
Ammodiscus cretaceus ones published by Romein and Smit (1981), Perch-Ammodiscus peruvianus Nielsen et al. (1982), and Smit (1990).Glomospirella grzybowskii The drop of d13C values shortly after the K–PGlomospira spp. boundary is interpreted as reflecting the globalTrochamminoides spp.
decrease in marine primary producivity.Paratrochamminoides spp.
B3: 6.2. Stable oxygen isotopesHaplophragmoides spp.Cribrostomoides sp.
Recurvoides sp.Trochammina spp.
INFAUNAL:
MORPHOGROUP C:
C1:Hormosina spp.Lituola lataPseudobolivina sp.Spiroplectammina spp.Textularia sp.Reophax spp.Subreophax spp.Tritaxia spp.Clavulinoides eggeriDorothia spp.Eggerellina sp.Arenobulimina spp.Marssonella oxyconaGaudryina pyramidata
C2:Rzehakina epigonaRzehakina fissistomata
In the upper part of the Abathomphalus mayaro-ensis Zone d18O values hardly vary except for anincreasing tendency upsection in the Zone. Very
6. Stable isotopes anomalies large negative shift of d18O of up to 4‰ wasmeasured at the K–P boundary, followed by an
A multitude of studies on stable isotopes across unstable signal in the lower PO Zone. In the higherthe Cretaceous–Paleogene boundary has been pub- part of the section d18O values tend to increase,lished (e.g., Romein and Smit, 1981; Hsu et al., with some pertubations in the middle part of the1982; Perch-Nielsen et al., 1982; Keller and Parvularugoglobigerina eugubina Zone (Fig. 6).Lindinger, 1989; Zachos et al., 1989; Barrera and The large amplitude of changes in d18O valuesKeller, 1990; Smit, 1990; Magaritz et al., 1992; reflects rather the effect of diagenesis than originalSchmitz et al., 1992). The published curves differ seawater temperature fluctuations.often significantly in shape.
Stable oxygen and carbon isotopic trends acrossthe K–P boundary intervals were reported from 7. Discussioncalcareous tests of benthic and planktonic foramin-ifers, fine-fraction and/or whole-rock samples. The Although our knowledge on foraminifers has
increased in the last fifteen years due to the greatwhole-rock data are thought to be least valuable

302 D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
Fig. 5. Idealized paleoenvironmental reconstruction of the habitat of deep-water agglutinated foraminifers on the sea bottom duringlatest Maastrichtian and earliest Paleocene in the Rotwandgraben section.
interest in K–P boundary events and mass extinc- the Early Paleocene and numerous taxa persistedacross the K–P boundary with little or no change.tions, agglutinated forms are still poorly under-
stood. Most of the papers dealing with benthic However, it has become evident that the extinctionrate is higher within benthic foraminiferal faunasforaminiferal assemblages focus on calcareous
species. in shallower seas than in deeper ones ( Keller,1988; Thomas, 1990; Kaiho, 1992; Speijer and VanPapers published in last few years ( Kuhnt, 1990;
Thomas, 1990; Kaiho, 1992; Widmark and der Zwaan, 1994).Deep-water benthic foraminiferal assemblagesMalmgren, 1992; Kuhnt and Kaminski, 1993;
Coccioni and Galeotti, 1994; Speijer and Van der in K–P sediments were studied by Kuhnt (1990),Thomas (1990), Kaiho (1992), Widmark andZwaan, 1994) confirm earlier opinions (e.g.,
Beckmann, 1960; Douglas and Woodruff, 1981; Malmgren (1992), Kuhnt and Kaminski (1993)and Coccioni and Galeotti (1994).Dailey, 1983) that the late Maastrichtian taxa of
deep-sea species differ only slightly from those in These authors performed morphotypic analyses
303D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
Fig. 6. Carbon and oxygen stable isotopes profiles and iridium content of the Rotwandgraben section. Lithologies and other symbolsare explained in Fig. 2. Iridium content from Peryt et al. (1993).

304 D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
in order to recognize the paleoecology of assem- with a coarsely agglutinated wall (Subreophax,blages. Thomas (1990) examined the proportions Ammobaculites, Recurvoides and Saccammina) andof foraminiferal species with infaunal and epifau- Group 3 by epifaunal suspension and detritus-nal morphotypes from the K–P boundary interval feeders, e.g. astrorhizids and ammodiscids. Thesein the Weddell Sea. She documented that the few three groups dominate assemblages in stratigraph-foraminiferal species which went extinct during the ically different parts of the K–P boundary interval:K–P event belonged to infaunal morphotypes and Group 3 along with Group 1 dominate latestthat epifaunal morphotypes increased in relative Maastrichtian DWAF assemblages; Group 2 is theabundance just after the boundary. This pattern main component of the assemblage in the sedi-of extinction and change in the structure of benthic ments directly above the boundary (PO Zone) andassemblages was interpreted as reflecting a strong Group 1 dominates assemblages from the marlydecrease in nutrient supply to the sea floor which upper portion of the K–P boundary clay (Pafavoured epifaunal species. Infaunal species which Zone). According to the authors the observeddepend on a relatively high food supply did not changes in the DWAF assemblages from uppersurvive the crisis (Thomas, 1990). A similar bathyal environment reflect a response of benthicpattern of changes within benthic foraminiferal biota to a collapse of the food web as a conse-assemblages across the K–P boundary interval was quence of drastically reduced primary productivitydocumented by Widmark and Malmgren (1992) in the earliest Paleocene.in the South Atlantic. The decrease in relative A comparison of DWAF assemblages from theabundance of infaunal morphotypes was interpre- K–P boundary interval from the Rotwandgrabented by them as a reflection of a decrease in primary section with those from the Basque region showsproductivity. a relatively high similarity with respect to both
In turn, Coccioni and Galeotti (1994) recorded taxonomic composition and changes in relativein the basal Danian at Caravaca (southeast Spain) abundances of dominant genera. Some differencesa sudden decrease of diversity within benthic fora-
are found as well. In the studied section,miniferal assemblages with an instantaneous
Ammobaculites is not recorded; Recurvoides,bloom of the infaunal detritus-feeders Bolivina andSaccammina, and Subreophax are present in theSpiroplectammina. A low-oxygen conditionDWAF assemblages to a lesser degree than in theevolved on the sea floor as a consequence ofBasque region. These differences are due toenhanced organic flux due to the mass mortalitydifferent depth of deposition of the two regions.in the upper water column and/or increased runoff
There is also a difference in morphotypic com-from land after the K–T event. This scenario wasposition of mainly Early Paleocene assemblages.given by Coccioni and Galeotti (1994) as the mainHowever, this difference may be more apparentfactor responsible for changes in morphotypicthan real. Epifaunal/shallow infaunal forms maystructure in benthic foraminiferal assemblages atbe included either with epifaunal morphotypes orCaravaca.with infaunal ones. Kuhnt and Kaminski (1993)Kuhnt (1990) and Kuhnt and Kaminski (1993)considered the PO Zone assemblage dominatedstudied agglutinated foraminiferal assemblages atby Subreophax, Ammobaculites, Recurvoides andthe K–P boundary interval from upper bathyalSaccammina as infaunal. Recurvoides andenvironments of the Umbrian Apennines (Italy)Saccammina comprise about 50% of thisand the Basque Basin (Spain). These authors dis-assemblage.covered a similar pattern of changes in DWAF
In our study the two genera are regarded asassemblages in both localities and distinguishedrepresenting an epifaunal life position, followingthree different faunal groups: Group 1 is domi-Jones and Charnock (1986) and Corliss and Chennated by calcareous agglutinating infauna and(1988). It seems very likely that these species whichis mainly represented by the generalived free in the topmost sediment layer after theSpiroplectammina and Dorothia; Group 2 is repre-
sented by non-calcareous infaunal morphotypes K–P boundary event migrated to the surface of

305D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
the sediment in search for food (e.g., Gooday, fluctuations of isotope values (e.g., Zachos et al.,1989; Schmitz et al., 1992) as well as CaCO31994).
Assemblages II and III from the Guembelitria content. K–P boundary levels are also charac-terized by an iridium anomaly (Alvarez et al.,cretacea Subzone are almost entirely composed of
astrorhizids and Trochammina (Fig. 4). This 1980).In the few centimeters of the lowermostclearly indicates that among agglutinated foramini-
fera the group least affected by the K–P boundary Paleocene sediments at Rotwandgraben a stableisotope anomaly (Fig. 6) as well as an iridiumcrisis in the Gosau region, were suspension-feeders
and species living on the surface of the sediment, anomaly (Peryt et al., 1993) have been recorded.The restriction of increased iridium values to ai.e. species with a predominantly epifaunal mode
of life. During the time of the collapse of the food very thin (<2 cm) layer in the Rotwandgrabensection indicates a rather short-lived source of theweb, these groups could survive on a very limited
food supply. Similar results for calcareous benthic anomaly.The iridium anomaly correlates with the begin-foraminifers were obtained by Thomas (1990) for
the Weddell Sea and Widmark and Malmgren ning of the d13C negative shift.As indicated by stable isotopic trends (e.g.,(1992) for the South Atlantic.
The following characteristics of the marine envi- Keller and Lindinger, 1989; Zachos et al., 1989;Schmitz et al., 1992), a gradual, long-term climaticronment may be suggested from the Gosau data:
(1) Assemblage I in which an epifaunal morpho- and oceanic instability does not seem to be a factorwhich was responsible for the sudden extinctiongroup predominates is typical for deep marine
basins which are usually somewhat oligotrophic. of many planktonic and benthic organisms(Zachos et al., 1989).On the other hand, significant numbers of the
infaunal genera Dorothia, Marssonella and A bolide impact suppressing sunlight longenough to reduce photosynthesis on a global scaleSpiroplectammina with calcareous cement indicate
that in the latest Maastrichtian the supply of food and to diminish primary productivity in the oceans(e.g., Alvarez et al., 1980; Smit, 1990) is consistentparticles and calcium carbonate was sufficient for
infaunal benthic life at depths. with the rapid change of morphotypic compositionin deep-water agglutinated foraminiferal assem-(2) Assemblage II — a paucity of foraminiferss
and low diversity suggest an instantaneous collapse blages and the coincidence of this change with aniridium anomaly. A drastic decrease in surface-of the food supply.
(3) Assemblage III is almost entirely composed water organic productivity resulted in the collapseof the nutrient supply for benthic organisms, theof epifaunal suspension-feeders (astrorhizids) and
omnivorous trochamminids, indicating the begin- extinction of several infaunal species of benthicforaminifers (Thomas, 1990) and the large changesning of restoration of marine primary surface
productivity. Limited food favours epifaunal in morphotype composition of benthic foramini-feral assemblages (Thomas, 1990; Kuhnt andmorphogroups.
(4) Assemblage IV is composed of epifaunal Kaminski, 1993).and infaunal morphogroups, with a similar contri-bution of infaunal groups with calcareouscement — Dorothia, Marssonella andSpiroplectammina. This suggests a return of pre- 8. ConclusionsK–P boundary conditions with sufficient supply offood particles and calcium carbonate for benthic (1) Significant faunal and geochemical changes
are observed in the Guembelitria cretacea Subzoneorganisms.K–P boundary levels in the marine record are of the Rotwandgraben section in Austria. Carbon
and oxygen stable isotopes as well as an iridiumassociated with a significant drop in d13C andd18O values (e.g., Perch-Nielsen et al., 1982; Zachos anomaly are recorded at the base of the Subzone.
(2) DWAF assemblages in the Guembelitriaet al., 1989), followed and preceded by smaller

306 D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
Butt, A., 1981. Depositional environments of the Upper Creta-cretacea Subzone are impoverished in abundanceceous rocks in the northern parts of the Eastern Alps. Cush-and diversity in comparison with under- and over-man Found. Foraminiferal Res., Spec. Publ. 20, 5–121.lying sediments. The assemblage from the lower
Coccioni, R., Galeotti, S., 1994. K–T boundary extinction: Geo-part of the subzone displays very low abundance logically instantaneous or gradual event? Evidence fromand diversity. The assemblage from the upper part deep-sea benthic foraminifera. Geology 22, 779–782.
Corliss, B.H., 1985. Microhabitats of benthic foraminiferais composed almost entirely of epifaunalwithin deep-sea sediments. Nature (London) 314, 435–438.morphogroups.
Corliss, B.H., Chen, C., 1988. Morphotype patterns of Norwe-(3) The low abundance of foraminifera in thegian Sea deep-sea benthic foraminifera and ecological impli-assemblages of the subzone as well as the change cations. Geology 16, 716–719.
in their morphotypic composition indicates a Dailey, D.H., 1983. Late Cretaceous and Paleocene benthic for-sudden breakdown of the food supply to benthic aminifers from Deep Sea Drilling Project Site 516, Rio
Grande Rise, Western South Atlantic Ocean. Init. Rep.organisms. The simultaneous occurrence of geo-DSDP 72, 757–782.chemical and iridium anomalies and the sudden
Douglas, R.G., Woodruff, F., 1981. Deep-sea benthic Foramini-extinction of planktonic foraminifers as well as thefera. In: Emiliani, C. (Ed.), The Oceanic Lithosphere. Thedrastic reorganization of DWAF assemblages at Sea, 7. Wiley–Interscience, New York, pp. 1233–1327.
the K–P boundary seem to be interrelated. Emiliani, C., Kraus, E.B., Shoemaker, E.M., 1981. Suddendeath at the end of the Mesozoic. Earth Planet. Sci. Lett.55, 317–334.
Geroch, S., Kaminski, M.A., 1992. The morphology, paleoecol-Acknowledgements ogy and systematics of Nothia excelsa (Grzybowski), a deep-
water agglutinated foraminifer. Ann. Soc. Geol. Pol. 62,The paper benefitted from reviews by Michael 255–265.
Gooday, A.J., 1983. Bathysiphon rusticus De Folin, 1886 andA. Kaminski (University College, London) and byBathysiphon folini, n. sp.: two large agglutinated foraminiferaan anonymous reviewer to whom the authors areabundant in abyssal N.E. Atlantic epibenthic sledge samples.greatly indebted. Thanks are due to E. Hara forJ. Foraminiferal Res. 13, 262–276.
processing samples, to G. Dziewinska for the Gooday, A.J., 1994. The biology of deep-sea foraminifera: Aphotos and to T. Dobroszycka for the drawings. review of some advances and their applications in paleocean-The Stefan Batory Foundation is gratefully ography. Palaios 9, 14–31.
Hałas, S., 1979. An automatic inlet system with pneumaticacknowledged for partial support of this researchchangeover valves for isotope ratio mass spectrometer.to D.P.J. Phys. E.: Sci. Instrum. 18, 417–420.
Hałas, S., Skorzynski, Z., 1980. An unexpensive device for digi-tal measurements of isotopic ratios. J. Phys. E.: Sci. Instrum.13, 346–349.References
Hansen, T., Ferrand, R., Montgomery, H.A., Billman, H.G.,Blechschmidt, G., 1987. Sedimentology and extinction pat-
Alvarez, L.W., Alvarez, W., Asaro, F., Michel, H.W., 1980.terns across the Cretaceous–Tertiary boundary interval inExtraterrestrial cause for the Cretaceous–Tertiary extinction.east Texas. Cretaceous Res. 8, 229–252.Science 208, 1095–1108.
Herm, D., v. Hillebrandt, A., Perch-Nielsen, K., 1981. DieAu-Liu, C.J., Olsson, R.K., 1994. On the origin of DanianKreide/Tertiargrenze im Lattengebirge (Nordliche Kalkal-normal perforate planktonic foraminifera from Hedbergella.pen) in mikropalaontologischer Sicht. Geol. Bavarica 82,J. Foraminiferal Res. 24, 61–74.319–344.Barmawidjaja, D.M., Jorissen, F.J., Puskaric, S., Van der
Hesse, R., Butt, A., 1976. Paleobathymetry of Cretaceous tur-Zwaan, G.J., 1992. Microhabitat selection by benthic fora-bidite basins of the East Alps relative to the calcite compen-minifera in the northern Adriatic Sea. J. Foraminiferal Res.sation level. J. Geol. 84, 505–533.22, 297–317.
Hsu, K.J., McKenzie, J.A., He, Q.X., 1982. Terminal Creta-Barrera, E., Keller, G., 1990. Stable isotope evidence for globalceous environmental and evolutionary changes. Geol. Soc.environmental changes and species survivorship across theAm., Spec. Pap. 190, 317–328.Cretaceous/Tertiary boundary. Paleoceanography 5,
Jones, R.W., Charnock, M.A., 1986. ‘‘Morphogroups’’ of867–890.agglutinating foraminifera — Their life positions and feedingBeckmann, J.P., 1960. Distribution of benthonic foraminiferahabits and potential applicability in (paleo)ecologicalat the Cretaceous–Tertiary boundary of Trinidad (West
Indies). Int. Geol. Cong., 21 Session, Norden, 5, 57–69. studies. Rev. Paleobiol. 4, 311–320.

307D. Peryt et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 132 (1997) 287–307
Jorissen, F.J., 1988. Benthic foraminifera from the Adriatic Sea; tion of calcareous nannoplankton at the Cretaceous–Tertiaryboundary. Geol. Soc. Am., Spec. Pap. 190, 353–371.principles of phenotypic variation. Utrecht Micropaleontol.
Bull. 37, 174 pp. Peryt, D., Lahodynsky, R., Rocchia, R., Boclet, D., 1993. TheCretaceous/Paleogene boundary and planktonic foramini-Kaiho, K., 1992. A low extinction rate of intermediate-water
benthic foraminifera at the Cretaceous/Tertiary boundary. fera in the Flyschgosau (Eastern Alps, Austria). Palaeo-geogr., Palaeoclimatol., Palaeoecol. 104, 239–252.Mar. Micropaleontol. 18, 229–259.
Kaminski, M.A., Geroch, S., 1993. A revision of foraminiferal Preisinger, A., Zobetz, E., Gratz, A., Lahodynsky, R., Becke,M., Mauritsch, H.J., Eder, G., Grass, F., Rogl, F., Stradner,species in the Grzybowski Collection. Grzybowski Found.,
Spec. Publ. 1, 239–323. H., Surenian, R., 1986. The Cretceous/Tertiary boundary inthe Gosau Basin, Austria. Nature (London) 322, 797–799.Kaminski, M.A., Gradstein, F.M., Berrgren, W.A., Geroch, S.,
Beckmann, J.P., 1988. Flysch-type agglutinated foramini- Romein, A.J.T., Smit, J., 1981. The Cretaceous/Tertiary bound-ary: Calcareous nannofossils and stable isotopes. Proc. K.feral assemblages from Trinidad: taxonomy, stratigraphy
and paleobathymetry. Abh. Geol. Bundesanst. 41, 155–227. Ned. Akad. Wet., Ser. B 84, 295–312.Schmitz, B., Keller, G., Stenwall, O., 1992. Stable isotope andKeller, G., 1988. Biotic turnover in benthic foraminifera across
the Cretaceous/Tertiary boundary at El Kef, Tunisia. Palaeo- foraminiferal changes across the Cretaceous–Tertiary bound-ary at Stevns Klint, Denmark: Arguments for long-term oce-geogr., Palaeoclimatol., Palaeoecol. 66, 153–172.
Keller, G., Lindinger, M., 1989. Stable isotope, TOC and anic instability before and after bolide-impact event.Palaeogeogr., Palaeoclimatol., Palaeoecol. 96, 233–260.CaCO3 record across the Cretaceous/Tertiary boundary at
El Kef, Tunisia. Palaeogeogr., Palaeoclimatol., Palaeoecol. Smit, J., 1990. Meteorite impact, extinctions and the Creta-ceous–Tertiary boundary. Geol. Mijnbouw 69, 187–204.73, 243–265.
Kuhnt, W., 1990. Agglutinated foraminifera of western Medi- Speijer, R.P., Van der Zwaan, G.J., 1994. Extinction and survi-vorship patterns in southern Tethyan benthic foraminiferalterranean Upper Cretaceous pelagic limestones (Umbrian
Apennines, Italy, and Betic Cordillera, Southern Spain). assemblages across the Cretaceous/Paleogene boundary. In:Speijer, R.P. (Ed.), Extinction and recovery patterns in ben-Micropaleontology 36, 297–330.
Kuhnt, W., Kaminski, A., 1993. Changes in the community thic foraminiferal paleocommunities across the Cretaceous/Paleogene and Paleocene/Eocene boundaries. Geol. Ultraiec-structure of deep water agglutinated foraminifers across the
K/T boundary in the Basque Basin (northern Spain). Rev. tina 124, 19–64.Stradner, H., Rogl, F., 1988. Microfauna and nannoflora of theEsp. Micropaleontol. 25, 57–92.
Lahodynsky, R., 1988. Lithostratigrapgy and sedimentology Knappengraben (Austria) across the Cretaceous/Tertiaryboundary. Ber. Geol. Bundesanst. 15, 25–26.across the Cretaceous/Tertiary boundary in the Flyschgosau
(Eastern Alps, Austria). Rev. Esp. Paleontol., No. Extraor- Stradner, H., Becke, M., Grass, F., Lahodynsky, R., Mauritsch,H.J., Preisinger, A., Rogl, F., Surenian, R., Zobetz, E., 1985.dinario, pp. 73–82.
Lahodynsky, R., 1989. The K/T boundary and ‘‘Yellow Clay’’ The Cretaceous–Tertiary boundary in the Gosau Formationof Austria. Terra Cognita 5, 247Layers in the Gosau Group, Northern Calcareous Alps,
Austria. In: Wiedmann, J. (Ed.), Proc. 3rd Int. Cretaceous Stradner, H., Eder, G., Grass, F., Lahodynsky, R., Mauritsch,H.J., Preisinger, A., Rogl, F., Surenian, R., Zeissl, W.,Symp., pp. 677–690.
Longoria, J.F., Gamper, M.A., 1995. Planktonic foraminiferal Zobetz, E., 1987. New K/T boundary sites in the GosauFormation of Austria. Terra Cognita 7, 212faunas across the Cretaceous–Tertiary succession of Mexico:
Implications for the Cretaceous–Tertiary boundary problem. Thomas, E., 1990. Late Cretaceous through Neogene deep-seabenthic foraminifers (Maud Rise, Weddell Sea, Antarctica).Geology 23, 329–332.
Magaritz, M., Benjamini, C., Keller, G., Moshkovitz, S., 1992. Proc. ODP, Sci. Results 113B, 751–794.Widmark, J.G.W., Malmgren, B., 1992. Benthic foraminiferalEarly diagenetic isotopic signal at the Cretaceous/Tertiary
boundary, Israel. Palaeogeogr., Palaeoclimatol., Palaeoecol. changes across the Cretaceous–Tertiary boundary in thedeep-sea: DSDP Sites 525, 527, and 465. J. Foraminiferal91, 291–304.
McCrea, J.M., 1950. On the isotopic geochemistry of carbon- Res. 22, 81–113.Zachos, J.C., Arthur, M.A., Dean, W.E., 1989. Geochemicalates and a paleotemperature scale. J. Chem. Phys. 18,
849–857. evidence for suppression of pelagic marine productivity atthe Cretaceous/Tertiary boundary. Nature (London) 337,Perch-Nielsen, K., McKenzie, J., He, Q.X., 1982. Biostratigra-
phy and isotope stratigraphy and the ‘‘catastrophic’’ extinc- 61–64.