conservation of forest biodiversity and ecosystem properties in a pastoral landscape of the...
TRANSCRIPT

Conservation of forest biodiversity and ecosystem propertiesin a pastoral landscape of the Ecuadorian Andes
Chloe MacLaren • Hannah L. Buckley •
Roddy J. Hale
Received: 29 April 2013 / Accepted: 14 March 2014 / Published online: 1 April 2014
� Springer Science+Business Media Dordrecht 2014
Abstract High Andean cloud forests are home to a
diversity of unique wildlife and are important provid-
ers of ecosystem services to people in the Andean
regions. The extent of these cloud forests has been
widely reduced through conversion to pasture for
livestock, which threatens the forests’ ability to
support biodiversity and provide ecosystem services.
This paper explores whether impacts on woody plant
biodiversity and four ecosystem properties (woody
plant species richness, juvenile timber tree abundance,
soil organic matter content and soil moisture) from
converting forest to pasture can be mitigated if some
woody forest vegetation is maintained within pastures.
Woody vegetation in pastures was found to conserve
those woody plant species that are more tolerant to
exposure and grazing, but conservation of the high
montane cloud forest community required areas of
forest from which livestock were restricted. The
sampled sites clustered according to woody plant
species cover; these clusters represented a gradient
from pasture with patches of shrubs to mature forest.
Clusters differed in both woody plant species richness
and number of juvenile timber trees whereas soil
organic matter and soil moisture were observed to be
similar among all clusters. This suggests that the
different habitats may have some equivalent
ecosystem properties. We conclude that the presence
of woody vegetation in pastures may reduce some of
the impacts of converting forest to pasture, but should
not be considered a substitute for protecting large
areas of forest, which are essential for maintaining
woody plant species diversity in high Andean cloud
forest.
Keywords Conservation � Ecosystem
properties � Pasture � Forest �Woody vegetation �Andes
Introduction
Natural forests are fundamental to the conservation of
global biodiversity and ecosystem services (Pearce
2001; Lal 2004; Foley et al. 2007). Forests store
carbon (Lal 2004), regulate hydrology, climate and
soil nutrients (Reiners et al. 1994; Pearce 2001; Lal
2004; Foley et al. 2007; Ford et al. 2011), provide
timber, food and medicinal resources (Pearce 2001),
and provide habitat for a large number of species
(Gardner et al. 2009; Baez et al. 2010). However,
forested land is still in demand for agriculture, and
rapid deforestation continues worldwide, leading to
massive biodiversity losses and diminishing ecosys-
tem service provision (Pearce 2001; FAO 2010).
However, several authors are now proposing that these
declines can be mitigated by the use of agroecological
or ‘wildlife-friendly’ farming techniques, which aim
C. MacLaren � H. L. Buckley � R. J. Hale (&)
Department of Ecology, Lincoln University,
PO Box 85084, Lincoln 7647, Canterbury, New Zealand
e-mail: [email protected]
123
Agroforest Syst (2014) 88:369–381
DOI 10.1007/s10457-014-9690-9

to allow the persistence of native biodiversity in and
around the farming system (e.g., Scherr and McNeely
2008; Tscharntke et al. 2012).
One form of ‘wildlife-friendly’ farming is the
incorporation or retention of native trees into the
agricultural system to at least partially retain the
habitat attributes of natural forest (e.g., shading,
structural complexity) (Perfecto et al. 2005; Waldron
et al. 2012). In the Andes of Ecuador, it is common
practice to leave scattered remnant trees and forest
patches throughout highland pastoral landscapes, and
many farmers also allow shrubs and trees to
regenerate in their pastures (MacLaren, pers. obs).
This remnant and regenerating woody vegetation
may provide some habitat value for forest biodiver-
sity and conserve some forest ecosystem services
within pastoral landscapes. In this study, we inves-
tigated biodiversity and ecosystem service values of
woody vegetation in pastures of the upper Papallacta
valley, which lies on the eastern slopes of the
Ecuadorian Andes.
The native vegetation of the upper Papallacta valley
is high montane cloud forest, also referred to as ‘elfin
forest’ (Lauer and Rafiqpoor 2000), ‘montane ceja’
(Paniagua Zambrana et al. 2003), ‘high montane
evergreen forest’ (Salgado 2008) and ‘humid high
montane forest’ (Pillajo and Pillajo 2010). High
montane cloud forest is valued for its high biodiversity
(Baez et al. 2010), and is known to provide several
important ecosystem services. For the local people, the
forests are a source of income from tourism (native
charismatic species include the spectacled bear,
Tremarctos ornatus, and the mountain tapir, Tapirus
pinchaque), and a source of traditional resources such
as timber, foods, and medicines (Baez et al. 2010;
Pillajo and Pillajo 2010). These forests are also known
to be important in regulating local hydrology, partic-
ularly in terms of precipitation interception and
infiltration (Ataroff 2002).
Pastoral agriculture is the most widespread cause of
forest loss throughout the Andes (Jokisch and Lair
2002; Sarmiento 2002; Rodrıguez Morales 2009). In
the upper Papallacta valley the only extensive areas of
forest remain on the steep hillsides, and throughout
Ecuador, high montane cloud forest has been almost
entirely cleared from its range (Jokisch and Lair 2002;
Sarmiento 2002). When forest is converted to pasture,
local environmental conditions are altered by the
removal of the canopy cover and the loss of the
biogeochemical interactions between trees and soil
(Reiners et al. 1994; Holl 2006). This resulting open
pasture environment is likely to favour a different set
of plant species than those adapted to sheltered forest
conditions (Reiners et al. 1994). Any regenerating
vegetation in pasture areas is also subject to grazing
(Pettit et al. 1995; Posada et al. 2000), competition
with introduced pasture grasses (Sarmiento 1997), and
regular clearance by farmers, which may further
reduce the number of plant species that are capable
of survival.
The environment of adjacent forest can also be
affected by pasture, via edge effects. These manifest as
variety of changes to the physical environment of the
forest edges, such as increased light levels, and
increased variability in temperature and humidity
(Murcia 1995). Consequently, plant community com-
position is often found to vary between the edges and
interiors of habitat patches, with forest edges tending
to be dominated by a less diverse group of pioneer
species (Oosterhoorn and Kappelle 2000; Saunders
et al. 1991; Laurance et al. 2006; Tabarelli 2008;
Broadbent et al. 2008). Forest fragments that are small
relative to the extent of edge effects can have their
original community entirely displaced by ‘edge’
vegetation (Harrison and Bruna 1999; Tabarelli et al.
2008). Small forest fragments may also be less diverse
due to their size and isolation; smaller fragments
typically contain smaller populations, so there is a
higher probability of extinction through stochastic
processes (Collinge 2009). Isolation reduces the
number of seeds dispersing to a patch from other
patches, because the distance reduces the probability
of seeds arriving at the patch (Tscharntke et al. 2005;
Collinge 2009). Overall, increasing fragmentation of
forested landscapes tends to reduce biodiversity, and
drive the remaining plant communities toward pio-
neer-dominated communities (Tabarelli et al. 2008).
In this study, we aim to quantify the impacts of
agriculture on high montane cloud forest by investi-
gating how the woody plant community varies in
relation to agricultural intensity. Increased agricultural
intensity was represented by decreased vegetation
density (a surrogate for forest clearance) and increased
grazing intensity, as these are major impacts of
pastoral agriculture. Community composition was
also investigated in relation to altitude because this
is a well-known driver of compositional variation in
Andean vegetation (Young 1993; Lauer and Rafiqpoor
370 Agroforest Syst (2014) 88:369–381
123

2000; Salgado 2008). The specific questions addressed
in this study are: (1) how does the high montane forest
community differ between areas of low agricultural
intensity and high agricultural intensity, and (2) is
leaving woody vegetation in pastures an effective
method for conserving forest biodiversity within the
pastoral landscape?
The conversion of high montane cloud forest to
pasture is also likely to affect the ability of these
landscapes to provide ecosystem services. There is
evidence that climax forest communities provide a
greater supply and diversity of ecosystem services
than disturbed pioneer plant communities (Foley et al.
2007; Tabarelli et al. 2008) and that the focus on food
production in agricultural landscapes means other
ecosystem services, such as soil conditioning and
forest resource provision, are marginalised (Foley
et al. 2005). In this study we measured four ecosystem
properties, which are considered to represent the levels
of ecosystem service provision in different vegetation
types: species richness, which is linked to the overall
diversity and stability of ecosystem services
(Tscharntke et al. 2005; Isbell et al. 2011; Quijas
et al. 2012); soil organic matter content, which is a
reflection of soil nutrient status and carbon storage
(Lal 2009; Dominati et al. 2010); soil moisture
content, from which we gain an insight into the effects
of vegetation on local hydrology; and the abundance
of juvenile trees of timber species, which provides an
indication of the future timber supply. All ecosystem
properties were expected to be higher in forested areas
than in pasture areas, but retaining woody vegetation
in pastures could reduce this difference.
Methods
Data were collected between latitudes 0�22012500 and
0�23011000, and longitudes 78�9053500 and 78�11015000.The minimum altitude was 3,355 m.a.s.l, and the
maximum altitude was 3,752 m.a.s.l. All data were
collected from 83, 10 9 10 m (0.1 ha) quadrats
spread along the upper Papallacta valley in layout
aimed to be representative of the different vegetation
types and different levels of agricultural intensity.
Sites were placed at a minimum distance of 50 m apart
to reduce the effects of spatial autocorrelation.
The geographical coordinates and altitude (in
metres above sea level) of each site were recorded
using a Garmin eTrex 20 handheld GPS device.
Within each quadrat, percent canopy cover was
visually estimated from the centre point, and the
number of droppings from cattle, sheep, horses or
goats were counted. The area covered by forest within
a 100 m radius of the sample site was calculated using
ArcMap 10 (ESRI 2011), based on satellite imagery
from Google Earth (2012). The different vegetation
types (including forest, pasture, and pasture with
woody vegetation) were ground-truthed at each site
and at a variety of other points across the study area.
The abundance of every woody plant species
observed was recorded for each sample site. Only
plants taller than 30 cm in height were recorded, so
that only established individuals were included. Plant
species were identified using the illustrated guidebook
by Pillajo and Pillajo (2010), and plants not included
in the guidebook were later identified at the National
Herbarium of Ecuador. The number of juveniles of
potential timber tree species was also recorded at each
site. A juvenile tree was defined as being greater than
30 cm in height but less than 2.5 m in height or less
than 10 cm in breast-height diameter. Species were
designated as potential timber species if they regularly
grow large enough and have sufficiently durable wood
to be useful as a fencepost, or for larger constructions
(Pillajo and Pillajo 2010; pers. obs.). Twelve species
were identified as useful timber resources in the upper
Papallacta valley: Buddleja species, Escallonia myr-
tilloides, Grosvenoria rimbachii, Gynoxys spp., Hesp-
eromeles obtusifolia, Hesperomeles ferruginea,
Miconea bracteolata, Oreopanax ecuadorensis, Sara-
cha quitensis, Sessea crassifolia, and Vallea
stipularis.
At a subset of 40 sites, soil samples were taken from
the centre point and analysed at the Agrocalidad soil
laboratory in Tumbaco, Ecuador, for percent volu-
metric content of organic matter and percent gravi-
metric humidity.
Data analysis
Agglomerative cluster analysis (Bray-Curtis/Sørenson
distance), in conjunction with indicator species ana-
lysis, was undertaken in PC-ORD (McCune and
Mefford 2011) to group sites together based on their
community composition. Differences between each of
the clusters were assessed by calculating an inverse
Morisita-Horn Index, an index that is robust to high
Agroforest Syst (2014) 88:369–381 371
123

variability in plant abundances between sites (Magur-
ran 2004). Values of this index close to 0 indicate high
similarity and values close to 1 indicate low similarity.
Nonmetric multidimensional scaling (NMS) was
used to identify patterns in compositional variation
among sites (following Peck 2010 and Kent 2012).
Regression models were employed to determine which
environmental variables correlated with the changes in
plant community composition represented by each
axis of the NMS ordination. A generalised least
squares (GLS) regression approach was used because
this accounts for autocorrelation in the response
variable (Crawley 2007), which is often present in
ecological datasets where sample sites close together
can be more similar to each other than more distant
sites. One model was developed for each of the
significant NMS axes, with the response variable of
each model being the site scores of each axis from the
NMS ordination. The explanatory variables included
in the model were altitude, percent canopy cover,
livestock dropping density, and the proportion of area
within a 100 m radius of each site covered by forest.
All continuous explanatory variables were standard-
ised to a mean of zero and a standard deviation of one
before entering them into the model, so that the
regression coefficients were not affected by variables
of different scales.
Plots of the residuals and fitted values were used to
test the assumption of a linear relationship between the
explanatory variables and the response, and to check
for homoscedasticity in the response. Normal quan-
tile–quantile (QQ) plots were used to test the assump-
tion of normality in the residuals, and semivariograms
were used to check for the presence of spatial
autocorrelation in the residuals.
Analysis of variance was used to compare levels of
ecosystem properties among the community types
identified by the cluster analysis pairwise comparisons
were made using Tukey’s Honest Significant Differ-
ence in R (R Core Team 2012).
Results
Community types
A total of 65 plant species (Appendix) occurred in the
83 sample sites. Sites were assigned to nine clusters, as
using nine clusters resulted in the highest number of
significant indicator species associated with each
cluster. Four of these clusters contained three or fewer
members, resulting in an insufficient sample size to
determine any characteristic features of those clusters.
For this reason, these four clusters were excluded from
all further analyses.
The five common clusters were given descriptive
names based on some of their defining features: ‘high
altitude disturbed vegetation’, ‘low altitude disturbed
vegetation’, ‘mature forest’, ‘shrubby pasture’, and
‘regenerating pasture’ (Table 1). The ‘high altitude
disturbed’ cluster was found consistently in the upper
part of the valley (above 3,580 m.a.s.l.) and was
labelled ‘disturbed’ as member sites were typically
found in areas exposed to some agricultural pressure,
including pasture with woody vegetation, small forest
fragments, or forest sites near to the edge of forests.
Two points from this cluster were further into the
interior of the forest than the rest of the member sites.
These two sites were both beneath canopy gaps with
percent canopy covers of 50 and 60 %, which were
unusually low for forest sites in the Papallacta valley
study site. The ‘low altitude disturbed’ cluster was also
Table 1 Characteristics of each community type
Cluster Typically
occurs in
%
Canopy
cover
Altitude
(m.a.s.l.)
Mature forest Undisturbed
forest
interior
60–74 3,494–3,679
High altitude
disturbed
vegetation
Forest edges 15–51 3,574–3,634
Small forest
fragments
Pasture with
scattered
trees
Low altitude
disturbed
vegetation
Forest edges 30–66 3,414–3,510
Small forest
fragments
Pasture with
scattered
trees
Regenerating
pasture
Pasture with
scattered
trees
10–18 3,378–3,608
Open pasture
Shrubby pasture Open pasture 0 3,421–3,616
The interquartile range is given for % canopy cover and
altitude
372 Agroforest Syst (2014) 88:369–381
123

found in pasture with woody vegetation, small forest
fragments, or near to the edges of forests, but at lower
altitudes (generally below 3,520 m.a.s.l.). This cluster
contained several points that were inside forest areas;
however, all of these sites were accessible to cattle and
other livestock. The ‘mature forest’ group contained
sites that were located in interior forest locations and
these sites were protected from cattle and livestock
entry by small cliffs or impenetrable thickets of vines
and shrubs (MacLaren, pers. obs). The group labelled
‘shrubby pasture’ was made up of sites that were
essentially open pasture, with a variety of shrubs
present but little or no tree cover. The ‘regenerating
pasture’ cluster was named for its high abundance of
Baccharis latifolia (Table 2), a small tree species that
can regenerate easily in open areas (Pillajo and Pillajo
2010).
None of the community types shared more than
51 % similarity with any other cluster. The ‘mature
forest’, ‘high altitude disturbed’ and ‘low altitude
disturbed’ forests were most similar to one another,
with around 50 % similarity (Table 3). ‘Shrubby
pasture’ sites were most similar to ‘high altitude
disturbed’ sites and ‘regenerating pasture’ was most
similar to ‘low altitude disturbed’ sites, but the
similarity is weak in both cases (both have an inverse
Morisita-Horn index of 0.71, indicating only a 29 %
similarity between community types; Table 3). Sev-
eral species were found to occur significantly more
frequently and abundantly in certain clusters, and
these can be considered indicator species for those
clusters (Table 2). ‘Shrubby pasture’ had no signifi-
cant indicator species (Table 2), suggesting that this
cluster was more defined by a shared absence of
species than the presence of any particular species.
Patterns in community composition
The NMS ordination identified three relevant axes that
described the variation between the species assem-
blages of each plot (Fig. 1). Axis 1 was found to
explain approximately 30 % of the variance, while
both axis 2 and axis 3 explained approximately 20 %
each. Sites within the five common clusters are fairly
well separated in ordination space. Notably, ‘shrubby
pasture’ occurs at low values on axis 1 while ‘mature
forest’ occurs at high values, and ‘high altitude
disturbed’ sites occur at low values of axis 2 while
‘low altitude disturbed’ sites were present at high
values of axis 2. These trends were supported by the
regression models; all measures associated with
agricultural intensity including canopy cover, percent
surrounding forest cover, and number of animal
droppings, were significantly related to the first axis
of the NMS ordination (P \ 0.05; Table 4). Altitude
was significantly related to the second axis
(P \ 0.001; Table 4). No significant variables were
identified for the third axis (Table 4). None of the
three models (one for each axis) were found to violate
any model assumptions.
Table 2 Indicator species associated with each cluster
Cluster Species Growth form Indicator Value P value
Mature forest Calceolaria lamiifolia Climber 52.9 0.03
Munnozia jussieui Climber 51.7 0.02
Oreopanax ecuadorensis Tree 58.7 0.01
Monnina species Tree 45.2 0.04
High altitude disturbed vegetation Gynoxys (small-leafed morphotype) Tree 44.8 0.04
Miconia salicifolia Tree 62.6 0.00
Muehlenbeckia tamnifolia Climber 74.4 0.00
Ribes ecuadorensis Climber 66.1 0.00
Rubus species Climber 45.7 0.00
Salpichroa tristis Climber 46.5 0.01
Low altitude disturbed vegetation Jungia rugosa Climber/shrub 60.4 0.01
Solanum asperolatum Tree 75.2 0.00
Regenerating pasture Baccharis latifolia Tree 44.4 0.04
Shrubby pasture None – – –
Agroforest Syst (2014) 88:369–381 373
123

Table 3 Inverse Morisita-Horn index (beta-diversity) measures for each pairwise comparison of clusters
High altitude disturbed veg. Low altitude disturbed veg. Regenerating pasture Shrubby pasture
Mature forest 0.52 0.49 0.79 0.94
High altitude disturbed veg. 0.56 0.85 0.71
Low altitude disturbed veg. 0.71 0.81
Regenerating pasture 0.89
Values close to 1 indicate dissimilar community composition and abundance between cluster pairs
Fig. 1 The NMS ordination
plots showing community
composition in relation to
the NMS axes. Notable
trends include a shift from
the ‘shrubby pasture’
community type to ‘mature
forest’ along the first axis,
and from ‘high altitude
disturbed vegetation’ to
‘low altitude disturbed
vegetation’ along the second
axis. No particular trends are
obvious in relation to the
third axis, despite the
statistical significance of
this axis in the ordination
Table 4 Regression coefficients (‘Coeff.’) and P values (‘P’) for each explanatory variable in each axis model
Axis 1 Axis 2 Axis 3
Coeff. P Coeff. P Coeff. P
Altitude 0.05 0.250 -0.39 0.000 -0.08 0.146
Canopy cover 0.31 0.000 0.02 0.808 -0.03 0.657
Surrounding forest cover 0.17 0.015 0.08 0.234 -0.1 0.388
Number of droppings -0.15 0.005 0.02 0.741 -0.05 0.206
The three models are listed across the top of the table, and the explanatory variables are in the left-hand column. Positive coefficient
values indicate a positive relationship, negative values an inverse relationship with each NMS axis
374 Agroforest Syst (2014) 88:369–381
123
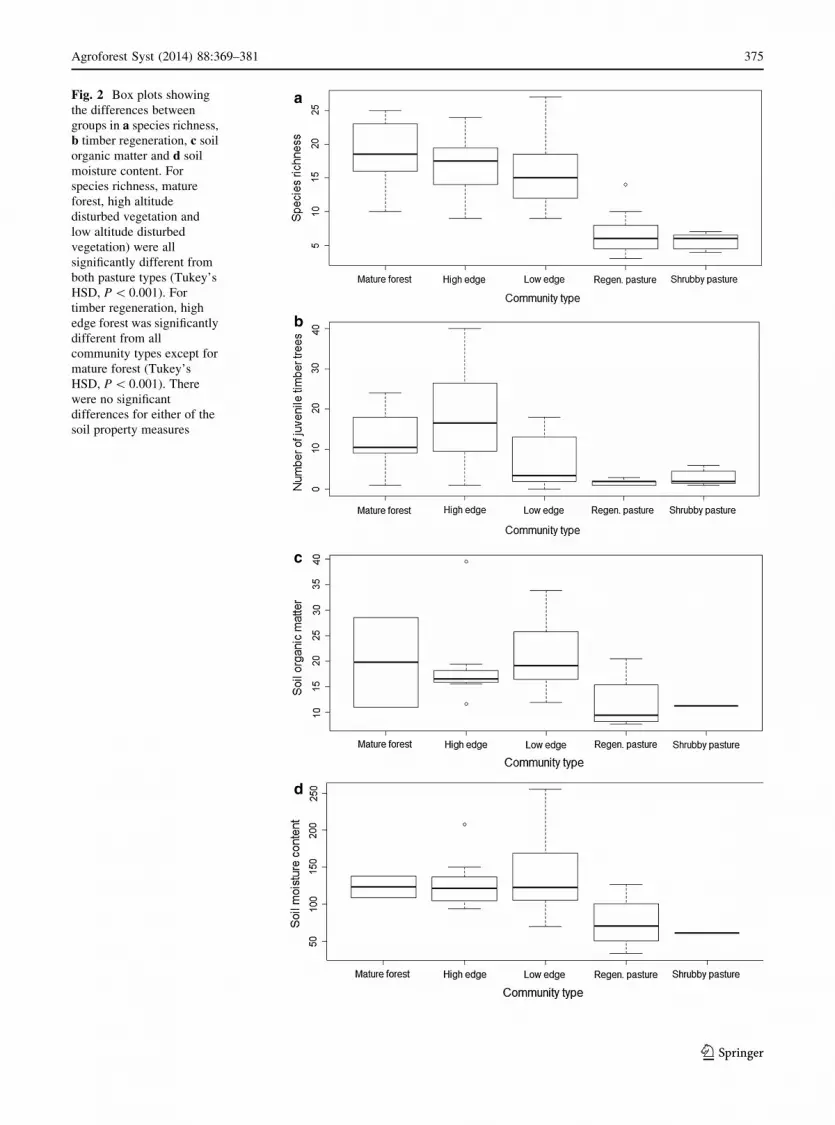
Fig. 2 Box plots showing
the differences between
groups in a species richness,
b timber regeneration, c soil
organic matter and d soil
moisture content. For
species richness, mature
forest, high altitude
disturbed vegetation and
low altitude disturbed
vegetation) were all
significantly different from
both pasture types (Tukey’s
HSD, P \ 0.001). For
timber regeneration, high
edge forest was significantly
different from all
community types except for
mature forest (Tukey’s
HSD, P \ 0.001). There
were no significant
differences for either of the
soil property measures
Agroforest Syst (2014) 88:369–381 375
123

Ecosystem properties
Significant differences in ecosystem properties among
community types were identified for species richness
(F(4,71) = 18.43, P \ 0.001) and timber regeneration
(F(4,71) = 11.55, P \ 0.001) (Fig. 2). Similar levels of
higher species richness were observed in mature forest,
high-altitude edge and low-altitude edge sites compared
to pasture-dominated sites (Fig. 2a). The three forested
cluster types tended to have higher and more variable
numbers of juvenile woody trees than the pasture-
dominated sites (Fig. 2b). There is some suggestion that
both soil ecosystem properties were lower in pasture
areas and higher in forested areas (Fig. 2c and d), but
these trends were not significant at the 5 % level.
Discussion
Five common and distinct plant community types
were identified in the upper Papallacta valley. Only
one occurred in undisturbed forest (‘mature forest’).
The other four community types were found in areas
influenced by agriculture, ranging from forest edges to
pasture areas with just a few woody shrubs. The
similarity of the plant communities to the mature
forest community decreased as canopy cover
decreased, grazing increased, and isolation from forest
increased. In addition, the number of juvenile trees and
woody plant species richness were higher in plant
communities that were more similar to the mature
forest community. These findings suggest that rem-
nant and regenerating vegetation is most effective for
woody plant biodiversity conservation when the
vegetation is present at high densities, when grazing
is kept to a minimum, and when large areas of forest
are conserved to act as source populations.
Grazing is known to reduce the presence of those
species that are most palatable to livestock and least
resilient to herbivory, whilst favouring an increase in
unpalatable and resilient species (Pettit et al. 1995;
Burns et al. 2011). Given that all member sites of the
mature forest cluster were known to be inaccessible to
cattle, while most disturbed vegetation sites were not, it
appears that grazing is a major edge effect that alters
forest communities within the upper Papallacta valley.
This is supported by the significance of livestock
dropping density, a proxy for grazing intensity, in
explaining vegetation composition (Table 3). Increased
light levels also likely play a role in determining
vegetation composition because canopy cover
explained significant variation, and the only two
disturbed vegetation sites that were not accessible to
cattle were found beneath canopy gaps. Most forest
communities have a suite of light-demanding species
that colonise treefall gaps and other open areas; these
also tend to be the species that survive on forest edges
and in forest fragments (Laurance et al. 2006).
The low levels of disturbance resulting from the edge
effects of grazing and increased light levels around
pastures caused a major change in community compo-
sition from mature forest to disturbed vegetation; neither
of the disturbed communities shared more than 51 %
similarity with mature forest (Table 3). However, the
disturbed plant communities were resilient to further
increases in agricultural pressure; both occurred
throughout areas of pasture with scattered trees, where
canopy cover was often substantially reduced and
grazing pressure often high, as well as in relatively
undisturbed forest edges (Table 1). This finding is
consistent with other studies of community responses to
agriculture, which show that undisturbed forest com-
munities tend to contain a group of highly forest-
dependent species, which disappear from the system
even at very low levels of agricultural intensity, while
other species of the same communities may be resistant
to relatively high agricultural intensity (Perfecto et al.
2005; Phalan et al. 2011; Pineda-Diez et al. 2012).
The amount of forest within a 100 m radius of a site
also affected the plant community structure, with sites
surrounded by greater amounts of forest being more
similar to the mature forest community (Table 3).
Forests provide a source of seeds, and can also be a
source of ecosystem functions such as pollination and
seed dispersal when it provides habitat for the
organisms that carry out these functions (Oosterhoorn
and Kappelle 2000; Jesus 2012). Therefore, these
source effects tend to increase the number and
diversity of species which establish in sites near to
forest in comparison with sites distant from forest
(Tscharntke et al. 2005; Collinge 2009) as found here.
Variation in relation to altitude was most obvious in
disturbed areas rather than in undisturbed forest. The
disturbed plant community in the upper Papallacta
valley is separated into two different community types
by altitude (the ‘low altitude disturbed’ type generally
occurring below 3520 m.a.s.l and the ‘high altitude
disturbed’ type generally occurring above 3580 m.a.s.l)
376 Agroforest Syst (2014) 88:369–381
123

while the mature forest community is found within both
altitudinal bands. This suggests that the creation of open
pasture environments generates more altitudinal varia-
tion than would otherwise be observed. The high
altitude disturbed vegetation was rich in plant species
generally associated with the treeline, or with the alpine
shrublands and grasslands (paramo) above the treeline.
These are more open environments than the forest, and
so plants from these high altitude habitats are likely to be
more suited to colonising edges than lower altitude
species. Without agriculture, the study area would be
entirely forested (it lies below the treeline at
3800 m.a.s.l.) and these open environment species
would not be able to establish, so the overall variation
would be limited to forest species. Such artificial
extensions of paramo species to lower altitudes is
thought to be widespread throughout the Andes as a
result of clearing forest for pasture (Sarmiento 2002).
The trends described so far indicate that forest
clearance and grazing in the upper Papallacta valley
lead to a change away from a mature cloud forest
community to an exposure-tolerant community. How-
ever, it may be possible for populations of some forest
species to survive in pasture under the right circum-
stances. In the regenerating pasture community, the
growth of pioneer plants may be aiding the regener-
ation of a wide range of other species, including forest
species such as O. ecuadorensis. The regenerating
pasture community is dominated by B. latifolia, a
small pioneer tree species that has previously been
found to promote the growth of seedlings of more
sensitive species (Posada et al. 2000). The branched
growth form of B. latifolia means that it protects the
area beneath its canopy from grazing and from
climatic exposure (Posada et al. 2000). Other plants
found in this community type with similar growth
forms (e.g., Berberis grandifolia and Barnadesia
arborea; MacLaren, pers. obs) may perform the same
function. This protection effect has been identified as
important for the regeneration of trees within wooded
pastures in Europe (Uytvanck et al. 2008), and could
be useful to promote the regeneration of forest trees
within pastoral areas of the upper Papallacta valley.
Community types and ecosystem properties
All ecosystem properties indicated higher ecosystem
service provision in mature forest and in both high and
low disturbed vegetation sites than in the pasture
community groups (Fig. 2). Notably, the high level of
timber regeneration occurring in the disturbed plant
communities, which included pasture with scattered
trees and small forest fragments, suggests that main-
taining trees and forest patches within the pastoral
landscape could be beneficial for protecting previ-
ously undisturbed forest from disturbance for timber
removal. The felling of forest trees for fenceposts is
known to be one of the greatest threats to forests
neighbouring agricultural areas of South America
(Murgueitio 2004). Post-and-wire fencing is widely
used in the upper Papallacta valley, so if trees are
completely cleared from pastures then farmers are
likely to seek timber in neighbouring forests.
The higher species richness found in forest frag-
ments and scattered tree communities compared with
open pasture communities may be beneficial to farmers.
Biodiversity is strongly linked to ecosystem service
provision (Quijas et al. 2010; Naeem et al. 2012), and
retaining biodiversity within pastoral areas may have
benefits for ecosystem services such as soil nutrients
(Murgueitio et al. 2011), beneficial insects (Bianchi
et al. 2006), and the productivity and nutritional value
of pasture herbs (Sanchez-Jardon et al. 2010). However,
further research is necessary because the relationship
between biodiversity and ecosystem services is known
to be complex and generally non-linear, with certain
species contributing more services than others
(Tscharntke et al. 2005; Bommarco et al. 2013). The
conservation of specific ecosystem services will require
a detailed knowledge of the relationship of those
services with different species and communities, and
with different land management scenarios.
Although the soil ecosystem properties suggested
that the mature forest and disturbed vegetation com-
munities provide greater organic matter and soil
moisture retention than open pasture communities,
this result was not significant. These ecosystem
properties would benefit from a more thorough study,
as it has been widely found in the literature that tree
cover can increase both soil organic matter and soil
moisture content in agricultural areas (Moreno 2008;
Jose 2009; Murgueitio et al. 2011).
Conclusions
The findings of this study indicate that agricultural
disturbance has had a large impact on the forest
Agroforest Syst (2014) 88:369–381 377
123

vegetation of the upper Papallacta valley and that
conservation of the mature cloud forest community
can only occur in a closed forest environment in
the absence of grazing. Protecting large forest
areas from further clearance should be a priority
for the conservation of high montane cloud forest
throughout the Ecuadorian Andes. It would also
help to encourage farmers to keep livestock out of
forested areas because this would eliminate the
edge effects caused by grazing (Burns et al. 2011).
If large forest areas cannot be protected, then
some of the loss of biodiversity caused by clearing
forest for pasture could be mitigated by retaining
scattered trees and forest fragments. These types
of vegetation are useful for the conservation of
plant species that are tolerant of higher light levels
and of grazing and may also be beneficial to the
farmers in terms of ecosystem services, notably as
a timber supply. In sum, scattered trees and forest
fragments in pastoral areas are a ‘wildlife-friendly’
solution for some species and some ecosystem
services, but the survival of the Andean cloud
forest will depend on the protection of large forest
areas.
Acknowledgments This study would have been impossible
without the advice and logistical assistance received from the
PRAA Project Team (Papallacta) of Care International, under
the auspices of the Ecuadorian Ministry of the Environment.
Thank you also to the communities of Tambo and Jamanco, on
whose properties this research was undertaken. Thanks to Pablo
Dominguez for help in the field, to Diana Fernandez of the
National Herbarium of Ecuador for help with identifying plants,
and to Brad Case for his assistance with spatial data and
calculations. Suggestions and advice from the Lincoln
University Spatial Ecology Group were invaluable during the
planning and analysis of this project. MacLaren was funded by a
Lincoln University postgraduate scholarship. The authors are
grateful to two anonymous reviewers, whose comments
improved this manuscript.
Appendix
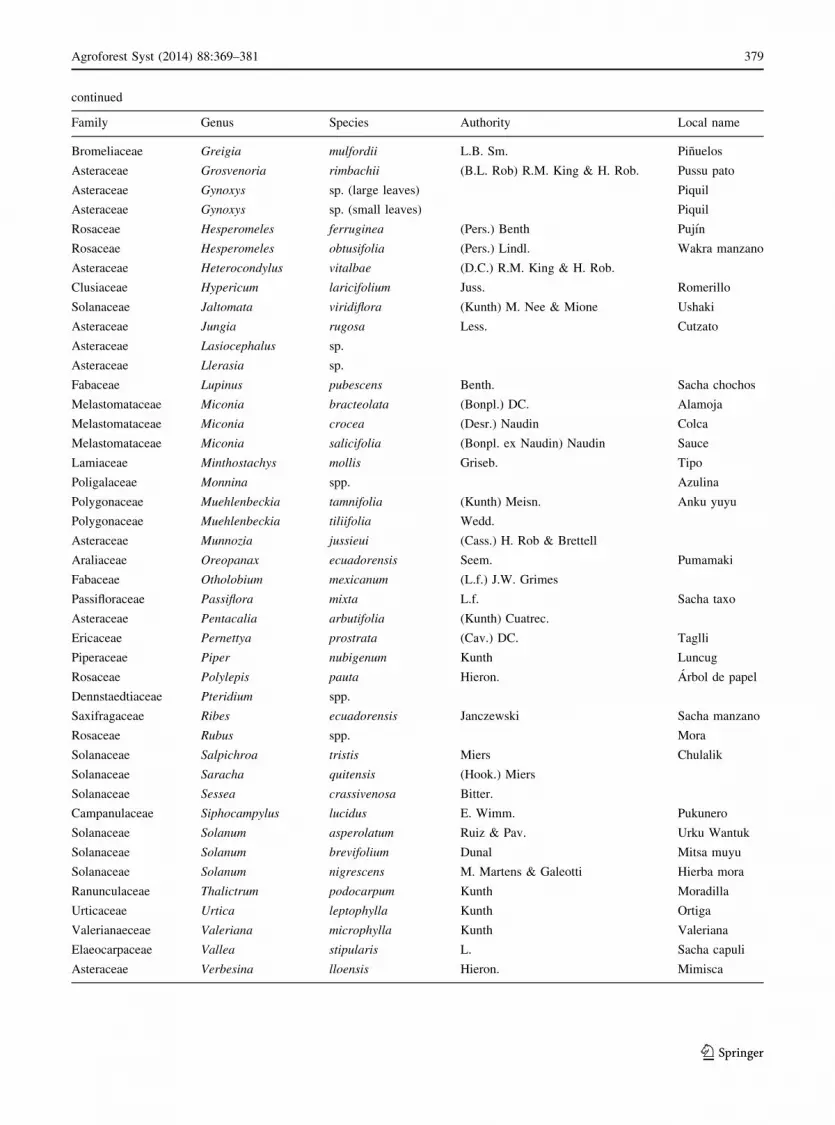
Family Genus Species Authority Local name
Rosaceae Acaena elongata L.
Asteraceae Baccharis buxifolia (Lam.) Pers.
Asteraceae Baccharis latifolia (Ruiz & Pav.) Pers. Chilco
Asteraceae Baccharis nitida (Ruiz & Pav.) Pers.
Asteraceae Baccharis prunifolia Kunth Chilco del cerro
Asteraceae Barnadesia arborea Kunth Alfilero
Scrophulariaceae Berberis grandiflora Turcz. Espino amarillo
Pterophyta Blechnum sp.
Melastomataceae Brachyotum ledifolium (Desr.) Triana Zarcilejo blanco
Scrophulariaceae Buddleja sp. Quijuar
Calceolariaceae Calceolaria lamiifolia Kunth Zapatitos
Campanulaceae Centropogon glabrifilis E. Wimm. Hierba de danta
Bambusoideae Chusquea scandens Kunth Bambu
Nyctaginaceae Colignonia ovalifolia Heimerl
Coriariaceae Coriaria ruscifolia L. Shanshi
Asteraceae Dendrophorbium lloense (Hieron ex Sodiro) C. Jeffrey Isca
Asteraceae Diplostephium floribundum (Benth.) Wedd.
Ericaceae Disterigma acuminatum (Kunth) Nied. Yurak muyu
Escalloniaceae Escallonia myrtilloides L.f. Chachaco
Onagraceae Fuchsia spp. Arete de monte
Asteraceae Gnaphalium elegans Kunth
378 Agroforest Syst (2014) 88:369–381
123
continued
Family Genus Species Authority Local name
Bromeliaceae Greigia mulfordii L.B. Sm. Pinuelos
Asteraceae Grosvenoria rimbachii (B.L. Rob) R.M. King & H. Rob. Pussu pato
Asteraceae Gynoxys sp. (large leaves) Piquil
Asteraceae Gynoxys sp. (small leaves) Piquil
Rosaceae Hesperomeles ferruginea (Pers.) Benth Pujın
Rosaceae Hesperomeles obtusifolia (Pers.) Lindl. Wakra manzano
Asteraceae Heterocondylus vitalbae (D.C.) R.M. King & H. Rob.
Clusiaceae Hypericum laricifolium Juss. Romerillo
Solanaceae Jaltomata viridiflora (Kunth) M. Nee & Mione Ushaki
Asteraceae Jungia rugosa Less. Cutzato
Asteraceae Lasiocephalus sp.
Asteraceae Llerasia sp.
Fabaceae Lupinus pubescens Benth. Sacha chochos
Melastomataceae Miconia bracteolata (Bonpl.) DC. Alamoja
Melastomataceae Miconia crocea (Desr.) Naudin Colca
Melastomataceae Miconia salicifolia (Bonpl. ex Naudin) Naudin Sauce
Lamiaceae Minthostachys mollis Griseb. Tipo
Poligalaceae Monnina spp. Azulina
Polygonaceae Muehlenbeckia tamnifolia (Kunth) Meisn. Anku yuyu
Polygonaceae Muehlenbeckia tiliifolia Wedd.
Asteraceae Munnozia jussieui (Cass.) H. Rob & Brettell
Araliaceae Oreopanax ecuadorensis Seem. Pumamaki
Fabaceae Otholobium mexicanum (L.f.) J.W. Grimes
Passifloraceae Passiflora mixta L.f. Sacha taxo
Asteraceae Pentacalia arbutifolia (Kunth) Cuatrec.
Ericaceae Pernettya prostrata (Cav.) DC. Taglli
Piperaceae Piper nubigenum Kunth Luncug
Rosaceae Polylepis pauta Hieron. Arbol de papel
Dennstaedtiaceae Pteridium spp.
Saxifragaceae Ribes ecuadorensis Janczewski Sacha manzano
Rosaceae Rubus spp. Mora
Solanaceae Salpichroa tristis Miers Chulalik
Solanaceae Saracha quitensis (Hook.) Miers
Solanaceae Sessea crassivenosa Bitter.
Campanulaceae Siphocampylus lucidus E. Wimm. Pukunero
Solanaceae Solanum asperolatum Ruiz & Pav. Urku Wantuk
Solanaceae Solanum brevifolium Dunal Mitsa muyu
Solanaceae Solanum nigrescens M. Martens & Galeotti Hierba mora
Ranunculaceae Thalictrum podocarpum Kunth Moradilla
Urticaceae Urtica leptophylla Kunth Ortiga
Valerianaeceae Valeriana microphylla Kunth Valeriana
Elaeocarpaceae Vallea stipularis L. Sacha capuli
Asteraceae Verbesina lloensis Hieron. Mimisca
Agroforest Syst (2014) 88:369–381 379
123

References
Ataroff M (2002) Precipitacion e intercepcion en los ecosist-
emas boscosos de los Andes venezolanos. Ecotropicos
15:195–202
Baez S, Ambrose K, Hofstede R (2010) Ecological and social
bases for the restoration of a High Andean cloud forest:
preliminary results and lessons from a case study in
northern Ecuador. In: Bruijnzeel LA, Scatena FN, Hamil-
ton LS (eds) Tropical montane cloud forests: science for
conservation and management. Cambridge University
Press, Cambridge, pp 628–643
Bianchi FJ, Booij CJ, Tscharntke T (2006) Sustainable pest
regulation in agricultural landscapes: a review on land-
scape composition, biodiversity and natural pest control.
Proc R Soc Biol Sci 273:1715–1727
Bommarco R, Kleijn D, Potts SG (2013) Ecological intensifi-
cation: harnessing ecosystem services for food security.
Trends Ecol Evol 28:230–238
Broadbent E, Asner G, Keller M, Knapp D, Oliveira P, Silva J
(2008) Forest fragmentation and edge effects from defor-
estation and selective logging in the Brazilian Amazon.
Biol Conserv 141:1745–1757
Burns BR, Floyd CG, Smale MC, Arnold GC (2011) Effects of
forest fragment management on vegetation condition and
maintenance of canopy composition in a New Zealand
pastoral landscape. Austral Ecol 36:153–166
Collinge SK (2009) Ecology of fragmented landscapes. The
John Hopkins University Press, Baltimore
Crawley MJ (2007) The R book. Wiley, Hoboken
Dominati E, Patterson M, Mackay A (2010) A framework for
classifying and quantifying the natural capital and eco-
system services of soils. Ecol Econ 69(9):1858–1868
ESRI (2011) ArcGIS desktop: release 10. Environmental Sys-
tems Research Institute, Redlands
Foley JA, Defries R, Asner GP, Barford C, Bonan G, Carpenter
SR, Chapin FS, Coe MT, Daily GC, Gibbs HK, Helkowski
JH, Holloway T, Howard EA, Kucharik CJ, Monfreda C,
Patz JA, Prentice IC, Ramankutty N, Snyder PK (2005)
Global consequences of land use. Science 309:570–574
Foley JA, Asner GP, Costa MH, Coe MT, DeFries R, Gibbs HK,
Howard EA, Olson S, Patz J, Ramankutty N (2007)
Amazonia revealed: forest degradation and loss of eco-
system goods and services in the Amazon Basin. Front Ecol
Environ 5:25–32
Food and Agriculture Organisation (FAO) (2010) Global forest
resources assessment. Forestry Department, Food and
Agriculture Organisation of the United Nations
Ford CR, Laseter SH, Swank WT, Vose JM (2011) Can forest
management be used to sustain water-based ecosystem
services in the face of climate change? Ecol Appl
21:2049–2067
Gardner TA, Barlow J, Chazdon R, Ewers RM, Harvey CA, Peres
CA, Sodhi NS (2009) Prospects for tropical forest biodi-
versity in a human-modified world. Ecol Lett 12:561–582
Google Earth (2012) ‘‘Papallacta’’, 0�22.125 to 78�11.150.
Google Earth Version 6. Accessed July 2012
Harrison S, Bruna E (1999) Habitat fragmentation and large-
scale conservation: what do we know for sure? Ecography
22:225–232
Holl KD (2006) Factors limiting tropical rain forest regeneration
in abandoned pasture: seed rain, seed germination, micro-
climate, and soil1. Biotropica 31:229–242
Isbell F, Calcagno V, Hector A, Connolly J, Harpole WS, Reich
PB, Scherer-Lorenzen M, Schmid B, Tilman D, van Ruij-
ven J, Weigelt A, Wilsey BJ, Zavaleta ES, Loreau M
(2011) High plant diversity is needed to maintain ecosys-
tem services. Nature 477:199–202
Jesus FM, Pivello VR, Meirelles ST, Franco GADC, Metzger JP
(2012) The importance of landscape structure for seed
dispersal in rain forest fragments. J Veg Sci 23:1126–1136
Jokisch BD, Lair BM (2002) One last stand? Forests and change
on Ecuador’s Eastern Cordillera. Geogr Rev 92:235–256
Jose S (2009) Agroforestry for ecosystem services and envi-
ronmental benefits: an overview. Agrofor Syst 76:1–10
Kent M (2012) Vegetation description and data analysis: a
practical approach, 2nd edn. Wiley, United Kingdom
Lal R (2004) Soil carbon sequestration to mitigate climate
change. Geoderma 123:1–22
Lal R (2009) Challenges and opportunities in soil organic matter
research. Eur J Soil Sci 60(2):158–169
Lauer W, Rafiqpoor MD (2000) Paramo de Papallacta: a phys-
iogeographical map 1:50,000 of the area around the Anti-
sana (Eastern Cordillera of Ecuador). Erdkunde 54:20–33
Laurance WF, Nascimento HEM, Laurance SG, Andrade AC,
Fearnside PM, Ribeiro JEL, Capretz RL (2006) Rain forest
fragmentation and the proliferation of successional trees.
Ecology 87:469–482
Magurran AE (2004) Measuring biological diversity. Blackwell
Science Ltd, Oxford
McCune B, Mefford MJ (2011) PC-ORD: multivariate analysis
of ecological data, version 6. MjM Software, Oregon
Moreno G (2008) Response of understorey forage to multiple
tree effects in Iberian dehesas. Agric Ecosyst Environ
123:239–244
Murcia C (1995) Edge effects in fragmented forests: implica-
tions for conservation. Trends Ecol Evol 10:58–62
Murgueitio E (2004) Silvopastoral systems in the Neotropics.
In: Mosquera-Losada MR, Rigueiro-Rodrıguez A, McAd-
am J (eds) Proceedings of an international congress on
silvopastoralism and sustainable management held in
Lugo, Spain, in April 2004. CAB International, United
Kingdom
Murgueitio E, Calle Z, Uribe F, Calle A, Solorio B (2011)
Native trees and shrubs for the productive rehabilitation of
tropical cattle ranching lands. For Ecol Manage 261:
1654–1663
Naeem S, Duffy JE, Zavaleta E (2012) The functions of bio-
logical diversity in an age of extinction. Science
336:1401–1406
Oosterhoorn M, Kappelle M (2000) Vegetation structure and
composition along an interior-edge-exterior gradient in a
Costa Rican montane cloud forest. For Ecol Manage
126:291–307
Paniagua Zambrana N, Maldonado Goyzueta C, Chumacero
Moscoso C (2003) Mapa de vegetacion de los alrededores
de la Estacion Biologica de Tunquini, Bolivia. Ecol Boliv
38:15–26
Pearce DW (2001) The economic value of forest ecosystems.
Ecosyst Health 7:284–296
380 Agroforest Syst (2014) 88:369–381
123

Peck JE (2010) Multivariate analysis for community ecologists:
step-by-step using PC-ORD. MjM Software, Oregon
Perfecto I, Vandermeer J, Mas A, Pinto LS (2005) Biodiversity,
yield, and shade coffee certification. Ecol Econ 54:435–446
Pettit NE, Froend RH, Ladd PG (1995) Grazing in remnant
woodland vegetation: changes in species composition and
life form groups. J Veg Sci 6:121–130
Phalan B, Onial M, Balmford A, Green RE (2011) Reconciling
food production and biodiversity conservation: land shar-
ing and land sparing compared. Science 333:1289–1291
Pillajo P, Pillajo M (2010) Plantas de Papallacta. Imprenta
Inkprima, Quito
Pineda-Diez de Bonilla E, Leon-Cortes JL, Rangel-Salazar JL
(2012) Diversity of bird feeding guilds in relation to habitat
heterogeneity and land-use cover in a human-modified
landscape in southern Mexico. J Trop Ecol 28:369–376
Posada JM, Aide TM, Cavelier J (2000) Cattle and weedy shrubs
as restoration tools of tropical montane rainforest. Restor
Ecol 8:370–379
Quijas S, Schmid B, Balvanera P (2010) Plant diversity
enhances provision of ecosystem services: a new synthesis.
Basic Appl Ecol 11(7):582–593
Quijas S, Jackson LE, Maass M, Schmid B, Raffaelli D, Bal-
vanera P (2012) Plant diversity and generation of ecosys-
tem services at the landscape scale: expert knowledge
assessment. J Appl Ecol 49:929–940
R Core Team (2012) R: a language and environment for sta-
tistical computing. R Foundation for Statistical Comput-
ing, Vienna
Reiners W, Bouwman A, Parsons W, Keller M (1994) Tropical
rain forest conversion to pasture: changes in vegetation and
soil properties. Ecol Appl 4:363–377
Rodrıguez Morales M, Chacın Moreno E, Ataroff Soler M
(2009) Transformacion del paisaje de selvas de montana en
la cuenca del rıo Capaz, Andes Venezolanos
Salgado S (2008) Composicion y diversidad de la flora en cuatro
localidades de la provincia del Carchi dentro del area de
intervencion del Proyecto Gisrena. In Boada C, Campana J
(eds) Composicion y diversidad de la flora y la fauna en
cuatro localidades en la provincia del Carchi. Un reporte de
las evaluaciones ecologicas rapidas. Ecociencia y GPC,
Quito, Ecuador
Sanchez-Jardon L, Acosta B, del Pozo A, Casado MA, Ovalle C,
Elizalde HF, Hepp C, de Miguel JM (2010) Grassland
productivity and diversity on a tree cover gradient in Not-
hofagus pumilio in NW Patagonia. Agric Ecosyst Environ
137:213–218
Sarmiento FO (1997) Arrested succession in pastures hinders
regeneration of Tropandean forests and shreds mountain
landscapes. Environ Conserv 24:14–23
Sarmiento FO (2002) Anthropogenic change in the landscapes
of highland Ecuador. Geogr Rev 92:213–234
Saunders DA, Hobbs RJ, Margules CR (1991) Biological con-
sequences of ecosystem fragmentation: a review. Conserv
Biol 5:18–32
Scherr SJ, McNeely JA (2008) Biodiversity conservation and
agricultural sustainability: towards a new paradigm of
‘ecoagriculture’ landscapes. Philos Trans R Soc Lond B
363:477–494
Tabarelli M, Lopes AV, Peres C (2008) Edge-effects drive
tropical forest fragments towards an early-successional
system. Biotropica 40:657–661
Tscharntke T, Klein AM, Kruess A, Steffan-Dewenter I, Thies C
(2005) Landscape perspectives on agricultural intensifica-
tion and biodiversity and ecosystem service management.
Ecol Lett 8:857–874
Tscharntke T, Clough Y, Wanger TC, Jackson L, Motzke I,
Perfecto I, Vandermeer J, Whitbread A (2012) Global food
security, biodiversity conservation and the future of agri-
cultural intensification. Biol Conserv 151:53–59
Uytvanck JV, Maes D, Vandenhaute D, Hoffmann M (2008)
Restoration of wood pasture on former agricultural land:
the importance of safe sites and time gaps before grazing
for tree seedlings. Biol Conserv 141:78–88
Waldron A, Justicia R, Smith L, Sanchez M (2012) Conserva-
tion through chocolate: a win–win for biodiversity and
farmers in Ecuador’s lowland tropics. Conserv Lett
5:213–221
Young KR (1993) Changes in Forest structure and regeneration
between two peruvian timberline margins. Arct Alp Res
25:167–174
Agroforest Syst (2014) 88:369–381 381
123