compartmental origin of the striato-entopeduncular projection in the rat
TRANSCRIPT

THE .JOURNAL OF COMPARATIVE NEUROLOGY 331286-296 (1993)
Compartmental Origin of the Striato-Entopeduncular Projection
in the Rat
N. RAJAKUMAR, K. ELISEVICH, AND B.A. FLUMERFELT Departments of Anatomy and Clinical Neurological Sciences,
The University of Western Ontario, London, Ontario, Canada N6A 5C1
ABSTRACT The mammalian neostriatum is divisible into neurochemically and cytoarchitectonically
distinct striosome and matrix compartments. This compartmentalization is respected by many afferent and efferent projections of the striatum. The distribution of distinct types of neuroactive substances and receptors and the unique connections of the striosome and matrix suggest a functional segregation between these compartments. The present study examines the organization of efferent projections from each of the striatal compartments to the entopeduncu- lar nucleus (EPN), a major output center of the basal ganglia.
The fluorescent retrograde tracer fluorogold, or rhodamine-conjugated dextran, was injected into the lateral habenula or the ventrolateral nucleus of the thalamus of adult Wistar rats to identify the topographical organization of EPN-habenular and EPN-thalamic neurons. Fluorogold was then placed into the rostral or caudal parts of the EPN, identified from the previous experiment as areas containing predominantly EPN-habenular or EPN-thalamic neurons, respectively. Sections containing retrogradely labeled neurons in the neostriatum were simultaneously immunolabeled for calbindin-DZskD,, a calcium-binding protein found exclusively in the projection neurons of the matrix. The results indicate that the striatal projection to the EPN-habenular and EPN-thalamic parts of the EPN originates from striosome and matrix neurons, respectively. The duality of striatal outflow involving the EPN suggests a mechanism whereby the striosome is integrated into subcortical pathways that modulate the activity of the basal ganglia via the ascending serotoninergic projection from the dorsal raphe nucleus, whereas the matrix is involved in a loop that includes the thalamus and the cerebral cortex. o 1993 Wiley-Liss, Inc.
Key words: basal ganglia, thalamus, habenula, fluorogold, rhodamine dextran
The neostriatum, the major receptive component of the mammalian basal ganglia, receives massive projections from all areas of the neocortex (Kemp and Powell, '70; Kunzle, '75; Selemon and Goldman-Rakic, '85; Donoghue and Herkenham, '86; Gerfen, '89; Ragsdale and Graybiel, '901, monoaminergic centers of the midbrain (Anden et al., '64; Bentivoglio et al., '79; Van Der Kooy and Kuypers, '79; Parent et al., '83; Gerfen et al., '87; Feigenbaum and Graybiel, '89) and the intralaminar nuclei of the thalamus (Nauta et al., '74; Herkenham and Pert, '81; Jayaraman, '85; Dube et al., '88). The output of the striatum is mainly directed to the external and internal segments of the globus pallidus (Nauta and Mehler, '66; Loopuijt and Van Der Kooy, '85; Smith and Parent, '86; Beckstead and Cruz, '86; Jimenez-Castellanos and Graybiel, '89; Gimenez-Amaya and Graybiel, '90; Parent, '90) and the substantia nigra (Gerfen, '85; Smith and Parent, '86; Jimenez-Castellanos and Graybiel, '89; Desban et al., '89). The internal segment
of the globus pallidus or its homologue, the entopeduncular nucleus (EPN), forms an important output channel of the basal ganglia that directs projections mainly to the lateral habenula, the ventrolateral nucleus of the thalamus, and the pedunculopontine tegmental nucleus (Carter and Fibiger, '78; Larsen and McBride, '79; Nauta, '79; Van Der Kooy and Carter, '81; Vincent and Brown, '86). The rostral part of the EPN in the rat projects to the lateral habenula, which provides a large input to the dorsal raphe nucleus, an important source of the serotoninergic projection to the striatum (Kalen et al., '89). The caudal part of the EPN innervates the thalamus and the pontine tegmentum (Van Der Kooy and Carter, '81).
**
Accepted December 23, 1992. Address reprint requests to Dr. B.A. Flumerfelt, Department of Anatomy,
Health Sciences Centre, University of Western Ontario, London, Ontario, Canada N6A 5C1.
O 1993 WILEY-LISS, INC.

STRIATO-ENTOPEDUNCULAR PROJECTION 287
and can therefore be distinguished immunocytochemically from those of the striosomes (Gerfen et al., '85).
The differentiation of the striatal compartments and the segregation of their connections suggest that these compart- ments may be involved in different aspects of striatal function. The pattern of connections observed for the two compartments indicates that the striosome is integrated into subcortical pathways that modulate the activity of the basal ganglia, whereas the matrix is involved in a longer loop that incorporates the cerebral cortex, striatum, substan- tia nigra or EPN, and the thalamus. To examine this possibility, the striato-entopeduncular projection was stud- ied by fluorescent retrograde tracing techniques combined with immunocytochemical identification of markers spe- cific for striatal compartments. Initially, areas of the EPN, which project mainly to the lateral habenula or the ventro- lateral nucleus of the thalamus, were identified and then retrograde tracers were injected into these areas to label the striato-entopeduncular neurons. The distribution of retro- gradely labeled neurons in relation to the striatal compart- ments was determined by combining immunocytochemis- try for calbindin-D2gkD, or somatostatin in the same section. Part of the present results have been reported in abstract form (Rajakumar et al., '91).
The neostriatum was considered to be a homogeneous structure that receives information from diverse neocorti- cal areas. However, it is now well established that the entire neostriatum is divided into both cytoarchitectonically and neurochemically specific compartments (Olson et al., '72; Pert et al., '76; Graybiel and Ragsdale, '78; Herkenham and Pert, '81; Graybiel, '84; Gerfen et al., '85; Penny et al., '88). This compartmentalization was demonstrated by acetylcho- linesterase activity, which revealed that the neostriatum is divided into a cholinesterase-poor, three-dimensional laby- rinthine area referred to as the striosome and a surround- ing cholinesterase-rich matrix (Graybiel and Ragsdale, '78). It has since been shown that several neurotransmitters, modulators and receptors are also distributed differentially with respect to striosome and matrix compartments (Herkenham and Pert, '81; Gerfen, '84; Graybiel et al., '86; Besson et al., '88; Graybiel, '90). The striosome exhibits a denser immunoreactivity for substance P, enkephalin, dynorphin, and glutamic acid decarboxylase and contains a higher density of D1, M1 and mu-opiate receptor binding sites, whereas the matrix contains high levels of somato- statin and choline acetyltransferase immunoreactivity and D2 receptors (Herkenham and Pert, '81; Gerfen and Young, '88; Graybiel, '90).
The striosome-matrix compartmentalization is also re- spected by all major afferent inputs to the striatum. Al- though all cortical areas project to both compartments, the allocortex preferentially innervates the striosome, whereas the neocortex mainly projects to the matrix (Gerfen, '89, '92). The cortical input to the striosome arises mainly from the deep part of layer V while that to the matrix comes mainly from the superficial part of layer V and the supra- granular layers of the cortex (Gerfen, '89). The striosome receives its dopaminergic input from a chemically distinct group of neurons situated in the ventral tier of the substan- tia nigra pars compacta while the matrix is innervated by dopaminergic neurons of the dorsal tier of the substantia nigra pars compacta, the retrorubral area and the ventral tegmental area (Gerfen et al., '85, '87; Jimenez-Castellanos and Graybiel, '87). The thalamic input is directed mainly to the matrix, whereas the amygdala innervates both compart- ments (Herkenham and Pert, '81; Ragsdale and Graybiel, '88; Kita and Kitai, '90). The influence of the compartmen- tal distribution of afferents upon the striatum depends in part on the organization of striatal interneurons. The cholinergic and peptidergic interneurons of the striatum have their cell bodies distributed in both compartments with axons directed towards the matrix (Gerfen, '84; Ches- selet and Graybiel, '86; Graybiel et al., '86; Penny et al., '88).
The most common neurons of the striatum, the spiny projection neurons, are found in both compartments. Ger- fen ('84, '85) found in the rat that the projection neurons of the matrix innervate the substantia nigra pars reticulata, whereas the striosome neurons project to the substantia nigra pars compacta and islands of dopaminergic neurons in the pars reticulata. A similar segregation of striatal output has been reported in the cat (Jimenez-Castellanos and Graybiel, '89) and monkey (Gimenez-Amaya and Gray- biel, '90). Graybiel and coworkers also found a preferential output to the internal and external segments of the globus pallidus from the matrix (Jimenez-Castellanos and Gray- biel, '89; Gimenez-Amaya and Graybiel, '90). The projec- tion neurons of the matrix contain the calcium-binding protein, calbindin-DzgkD,, within their cell bodies and fibers
MATERIALS AND METHODS Retrograde tracing
Thirty adult female Wistar rats with body weights rang- ing from 280 to 310 g were used in this study. The rats were anaesthetized with an intraperitoneal injection of sodium pentobarbital (35 mgikg) and placed in a Kopf stereotaxic frame. Following surgical exposure, a fine glass micropi- pette (internal tip diameter 50-60 km) containing a 2% solution of fluorogold (Fluorochrome Inc., Englewood, CO) in 0.9% sodium chloride was lowered into the lateral habenula (n = 8) or the ventrolateral nucleus of the thala- mus (n = 8) according to the stereotaxic coordinates of Paxinos and Watson ('86). The tracer was injected by iontophoresis with a 5 pA positive current delivered in 7 second pulses every 14 seconds over 10 minutes from a constant current generator (Grass Instrument Co., Quincy, MA). Animals received routine postoperative care and analgesia (Talwin, s.c.). A day after placement of fluorogold into the ventrolateral nucleus of the thalamus or the lateral habenula, some animals also were injected stereotaxically with 0.5-1.0 pl of 5% rhodamine-conjugated dextran (Molec- ular Probes Inc., Eugene, OR) (Nance, '91) into the lateral habenula (n = 3) or the ventrolateral nucleus of the thala- mus (n = 3) on the ipsilateral side, respectively. Following a survival time of 9 to 10 days from the initial procedure, animals were perfused transcardially with a 0.1 M phos- phate buffer (PB) (pH 7.4) containing 4% paraformalde- hyde and the brain was dissected and immersed in 15% buffered (PB) sucrose a t 4°C. Alternate frozen transverse sections (40 krn) were mounted on chrome alum-gelatin coated slides, coverslipped with buffered (PB) 60% glycerine containing 0.1% p-phenalenediamine and examined under a fluorescence microscope. Selected areas were photographed with Kodak Ektachrome 400 or T-MAX 3200 film.
The areas of the EPN exclusively innervating the lateral habenula or the ventrolateral nucleus of the thalamus were carefully identified and their stereotaxic coordinates were determined. In another group of animals (n = l l ) , fluoro- gold was injected into the parts of the EPN identified as the

288 N. ICAJAKUMAR ET AL.
areas projecting either to the lateral habenula or to the ventrolateral nucleus of the thalamus (EPN-habenular neurons: AP 6.7, DV 2.5, L 2.9; EPN-thalamic neurons: AP 6.0, DV 2.2, L 2.7). The parameters of the tracer injection were optimized for the topographic separation of the sites of injection to ensure that there was no contamination of either injection site within the EPN. The 2% solution of fluorogold in 0.9% sodium chloride was injected by ionto- phoresis from a glass micropipette (internal tip diameter 30 p,m) with a 5 pA positive current delivered in 7 second pulses every 14 seconds for 6 minutes. This group of animals was perfused after an eight day survival period and processed in a manner similar to the previous group.
Immunocytochemistry Frozen sections (40 bm) through the EPN of normal rats
(n = 3) and every fourth transverse section through the striatum or the EPN from animals that received tracer injections were processed for ca1bindin-DzakD, immunoreac- tivity, which is present in cell bodies and processes of projection neurons of the striatal matrix. An indirect immunofluorescent technique was employed with a mono- clonal anti-calbindin-D28kDa immunoglobulin produced in a mouse hybridoma (Sigma Co., St. Louis, MO). The sections were incubated in 10% normal horse serum and 1% bovine serum albumin in PB (pH 7.4) for 45-60 minutes a t room temperature, rinsed in PB and then incubated in the primary antibody (1:200 dilution) in PB containing 1% normal horse serum and 0.1% bovine serum albumin for 36-40 hours at 4°C. They were then washed several times before being incubated in a solution containing biotinylated horse anti-mouse immunoglobulin (Vector, Burlingame, CA; 1:200) in PB for 45-60 minutes at room temperature. The sections were washed thoroughly in PB and incubated in fluorescein DCS conjugated to avidin (Vector; 1:400) in PB for 45-60 minutes. After several washes in PB, the sections were mounted and examined as described previ- ously for the tracing experiments. Control experiments including omission of the primary immunoglobulin and preadsorption of primary antibody with calcium-binding protein, calbindin-D2akD, (Sigma co.), were carried out to ensure the specificity of the immunocytochemical proce- dure.
Somatostatin immunoreactivity was examined in the striatum in cases where the entopeduncular nucleus was injected with fluorogold to confirm the pattern of compart- mentalization revealed by calbindin-DZskD, immunocyto- chemistry. The sections were incubated in a mixture of primary antibodies for calbindin-D28kD, and somatostatin. A well-characterized antibody (SS-320) directed against the somatostatin 28(1-12) peptide fragment (Benoit et al., '85) was used in combination with Texas Red-conjugated second- ary antibodies to identify the somatostatinergic fibers, which are seen mainly in the matrix.
RESULTS Entopeduncular nuclear efferents
Entopedunculo-habenularprojection. Iontophoretic ap- plication of fluorogold into the habenula (n = 8) resulted in a circumscribed dense unilateral deposition of tracer within the lateral habenular nucleus (Fig. 1). In four cases, the core of the injection site was confined to the lateral part of the lateral habenular nucleus, the area known to receive an EPN projection. The injection site was characterised by an intensely fluorescing granular neuropil surrounded by nu-
Fig. 1. A-C: Schematic reconstruction of serial sections through rostra1 (A), middle (B), and caudal (C) levels of the entopenduncular nucleus (EPN), showing the topographic distribution of entopedunculo- habenular (open circles) and entopedunculo-thalamic (filled circles) neurons following injection of fluorogold and rhodamine-conjugated dextran into the lateral habenula and the ventrolateral nucleus of the thalamus, respectively. Note the extent of the representative injection sites shown in black. CL, centrolateral nucleus of the thalamus; IC, internal capsule; LD, laterodorsal nucleus of the thalamus; LH, lateral hypothalamus; LHb, lateral hahenula; MD, mediodorsal nucleus of the thalamus; RT, reticulothalamic nucleus; SM, stria medullaris; VM, ventromedial nucleus of the thalamus; VPL, ventral posterolateral nucleus of the thalamus; VL, ventrolateral nucleus of the thalamus; ZI, zona incerta.
merous retrogradely labeled neurons. The retrograde label- ing was seen in the EPN and the adjacent lateral hypotha- lamic area of the ipsilateral side. Injection of rhodamine-

STRIATO-ENTOPEDUNCULAR PROJECTION 289
conjugated dextran, despite the large injection site in the lateral habenula, resulted in a comparable pattern of retrograde labeling within the EPN. However, the dextran also revealed the course of EPN-habenular fibers entering the ventrolateral aspect of the thalamus very clearly.
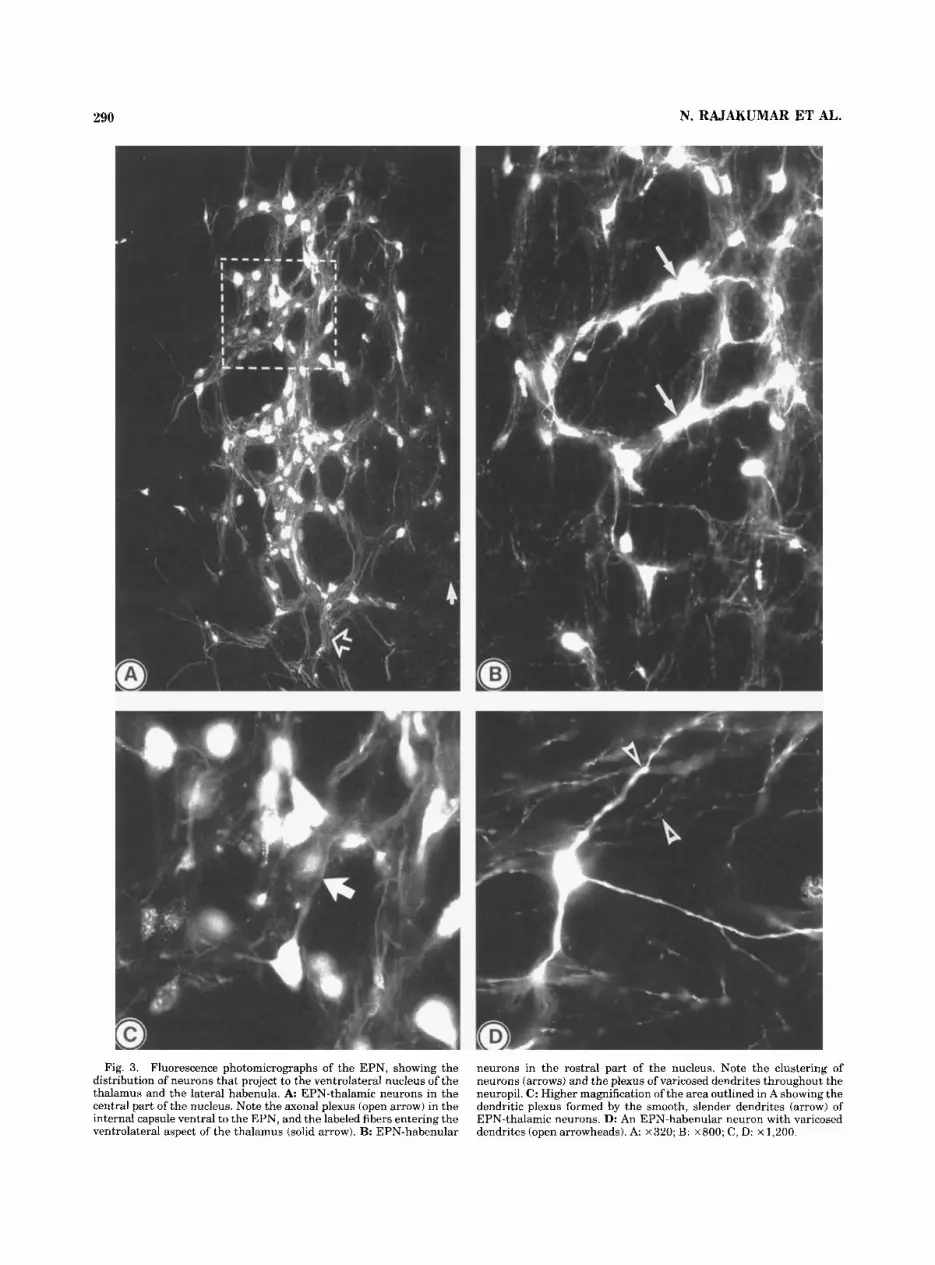
The labeled EPN neurons were of medium size, multipo- lar or fusiform in shape and were frequently found in clusters (Fig. 3B). Numerous retrogradely labeled neurons were seen clustered in the rostral third and the medial aspects of the middle third of the EPN and in the rostrolat- eral part of the lateral hypothalamic area. Injections involv- ing the entire area of the lateral habenular nucleus or only its lateral aspect resulted in a similar pattern of retrograde labeling within the EPN and the neighboring lateral hypo- thalamic area. The EPN-habenular neurons possessed long, slender, sparsely branched dendrites. The secondary and distal dendrites of all EPN-habenular neurons possessed a characteristic beaded appearance throughout their length (Fig. 3D).
Entopedunculo-thalamic projection. The fluorogold or the rhodamine-conjugated dextran injection sites in the thalamus involved the ventrolateral nucleus unilaterally without encroaching the other thalamic nuclei (Figs. 1,2B). In three animals, the injection site occupied mainly the rostral part of the ventrolateral nucleus while the remain- ing cases had injection sites mostly in the caudal parts of the nucleus. Labeled fibers emerged through the ventral and ventrolateral parts of the thalamus and passed dorsolat- erally into the internal capsule and formed an extensive plexus before entering the EPN through its ventral aspect (Fig. 3A).
Both rostral and caudal thalamic injections resulted in numerous retrogradely labeled neurons in the ipsilateral EPN and the adjacent hypothalamic area (Fig. 3A). None of
the cases showed labeling within the contralateral EPN. The labeled neurons of the EPN were of medium size and most of them were multipolar and possessed long, smooth dendrites that formed an extensive plexus throughout the nucleus (Fig. 3C). The retrogradely labeled neurons were seen mostly in the caudal two-thirds of the EPN (Fig. 1). The clustering was occasionally seen among the EPN- thalamic neurons. The rostral and caudal injections of tracer produced a slightly different pattern of retrograde labeling within the EPN, with rostral injections resulting in more rostral labeling than the caudal injections.
The injection of rhodamine- conjugated dextran produced larger injection sites involv- ing most of the lateral habenular nucleus or the ventrolat- eral nucleus of the thalamus. Numerous retrogradely labeled neurons were seen within the EPN. The dextran injections revealed the course of the axon bundles and the detailed arrangement of the dendritic arborization of EPN neurons. Both EPN-habenular and EPN-thalamic axons formed a prominent plexus in the internal capsule ventral to the EPN before entering through the ventral and ventrolateral aspects of the thalamus. The dendrites of EPN-thalamic neurons were extensively overlapped and extended a very long distance, particularly in the dorsoventral plane of the nucleus. However, they rarely extended into the territory of the EPN-habenular neurons. Injection of different tracers into the thalamus and habenula did not reveal any double- labeled neurons in the EPN, indicating a lack of collateral- ization within the EPN-thalamic and EPN-habenular sys- tems.
Immu- nocytochemical examination revealed moderately immuno- reactive fibers for ca1bindin-DzRkD, in the caudal two-thirds of the EPN, whereas the rostral third of the nucleus
Double labeling studies.
Calbindin-Dzako, Zmmunoreactivity in the EPN.
Fig. 2. A: Schematic diagram of the diencephalon, showing the position of a representative injection site in the ventrolateral nucleus of the thalamus. B: Fluorescence photomicrograph of the rhodamine- conjugated dextran injection site shown in A. C: Fluorescence photomicrograph illustrating the extent of a fluorogold injection site in the rostral EPN. Scale bars = 250 pn . See Figure 1 for abbreviations.
290 N. RAJAKUMAR ET AL.
Fig. 3. Fluorescence photomicrographs of the EPN, showing the distribution of neurons that project to the ventrolateral nucleus of the thalamus and the lateral habenula. A EPN-thalamic neurons in the central part of the nucleus. Note the axonal plexus (open arrow) in the internal capsule ventral to the EPN, and the labeled fibers entering the ventrolateral aspect of the thalamus (solid arrow). B: EPN-habenular
neurons in the rostra1 part of the nucleus. Note the clustering of neurons (arrows) and the plexus of varicosed dendrites throughout the neuropil. C: Higher magnification of the area outlined in A showing the dendritic plexus formed by the smooth, slender dendrites (arrow) of EPN-thalamic neurons. D: An EPN-habenular neuron with varicosed dendrites (open arrowheads). A ~ 3 2 0 ; B: ~ 8 0 0 ; C, D: x 1,200.

STRIATO-ENTOPEDUNCULAR PROJECTION 291
possessed very weak immunoreactivity. None of the EPN neurons showed immunoreactivity for calbindin-D28kDa. The combined immunocytochemical and tracing studies disclosed a close relationship between the EPN-thalamic neurons and the calbindin-Dz8kDa-containing fibers, whereas a similar association was not evident with the EPN- habenular neurons. The lateral hypothalamic area adjacent to the EPN and caudal parts of the globus pallidus con- tained a dense calbindin-Dz8kDa-positive fiber plexus and many immunoreactive cell bodies. However, none of the calbindin-containing hypothalamic or pallidal neurons were retrogradely labeled following injection of tracer into the EPN, suggesting that these are not responsible for the calbindin-immunoreactive fibers seen in the caudal EPN.
Striatal efferents to the EPN Striatal compartmentalization. Immunolabeling for cal-
bindin-DzskDa and somatostatin-28( 1-12) in the neostria- tum resulted in an identical compartmental pattern show- ing large areas containing calbindin-D28ma-immunoreactive cell bodies and somatostatin-positive fibers and small pock- ets of areas significantly low in these immunoreactive profiles. The areas of low and high immunoreactivity obtained with these antibodies correspond to the previously described striosome and matrix compartments, respec- tively.
The parameters adopted for the iontophoretic delivery of fluorogold into the EPN resulted in a small circumscribed deposition of tracer with the injection site not exceeding 200 pm in diameter. The core of the injection site consisted of bright fluorescing particles packed very densely within the neuropil (Fig. 2 0 . The area surrounding the core of the injection site was characterized by numerous retrogradely labeled neurons. However, the neuropil of the surrounding areas was devoid of any fluorescing particles, suggesting that the core is probably the effective site of uptake of the tracer. In all cases examined, the injection site was either confined to a small area in the rostral part of the EPN, which has been found to project to the habenula or to the caudal aspect of the nucleus, which projects to the thalamus.
Striosome projection to the EPN. Placement of tracer into the rostral part of the EPN (n = 6) resulted in numerous retrogradely labeled neurons in the neostriatum and the globus pallidus, ipsilaterally. The labeled striatal neurons were exclusively found within the striosomes as identified by the lack of ca1bindin-DzskDa immunoreactivity (Fig. 4A-F). The labeling was confined to the striosome compartments in the dorsal and lateral aspect of the striatum extending from the level of the genu of the corpus callosum to the anterior commissure. In a rostral to caudal progression through the neostriatum, the pattern of retro- grade labeling shifted from dorsolateral to ventromedial. Striosomes in more rostral, dorsal, and lateral regions were almost entirely filled with labeled neurons (Fig. 6A). How- ever, those in more caudal, ventral, and medial regions were only partially filled, suggesting that these striosome areas contain neurons projecting to more caudal parts of the EPN or to the pars compacta of the substantia nigra. Within striosomes, the retrogradely labeled neurons were often found in clusters of six to eight cells. The neurons were of medium size and multipolar and the labeling was limited to their cell bodies and proximal dendrites.
Following injection of tracer into the caudal third of the EPN (n = 51, the retrogradely labeled neurons were seen predominantly in
E P N injection sites.
Matrix projection to the EPN.
the calbindin-DzgkDa rich matrix compartment of the stria- turn, ipsilaterally (Figs. 5A, 6B). The matrix labeling was seen scattered in the lateral two-thirds of the striatum beginning at the level of the genu of the corpus callosum and continuing almost to the caudal end. The retrogradely labeled cells were more numerous towards the central part of the striatum. Clusters consisting of five to seven labeled cells were seen in many parts of the striatal matrix. Nevertheless, clustering was less evident in the matrix than in the striosomes. The retrogradely labeled matrix neurons appeared to be morphologically similar to those of the striosomes.
DISCUSSION The present study, employing fluorescent retrograde
tracing in combination with immunocytochemical character- ization of neuronal profiles, provides important informa- tion regarding the organization of neostriatal output sys- tems (Fig. 7) and the functional heterogeneity of striatal compartmentalization. The results indicate that: (1) the striosome neurons project to the entopedunculo-habenular neurons of the EPN and therefore are involved in a pathway influencing the serotoninergic neurons of the dorsal raphe nucleus through the lateral habenula; (2) the matrix neurons project to the entopedunculo-thalamic neu- rons of the EPN and thereby are associated with the pathway leading to the cerebral cortex through the ventro- lateral nucleus of the thalamus; and (3) the segregated connections of the striosome and the matrix compartments of the neostriatum and their involvement in functionally specific pathways confirm the functional heterogeneity proposed for the striatal compartments.
Methodological considerations In this study it was imperative to employ a retrograde
tracer with which the smallest possible injection site could be obtained, yet which possesses a superior sensitivity in order to yield a larger number of retrogradely labeled cells. The retrograde tracer fluorogold (Schmued and Fallon, '86) in combination with the injection parameters used for its delivery into the EPN, resulted in circumscribed injection sites not exceeding 200 pm in diameter and gave rise to numerous retrogradely labeled neurons found predomi- nantly in the striosome or matrix compartment of the striatum. With any tracing technique, retrograde labeling due to uptake and transport of tracer by fibers-of-passage is possible, especially in a center such as the EPN, which is penetrated by numerous fiber bundles. However, the dis- tinct patterns of retrograde labeling observed following injection into various parts of the rostral or caudal EPN indicate that in the present study, iontophoretic injection of fluorogold into the EPN did not provide significant retro- grade labeling by fibers-of-passage.
The second fluorescent tracer, rhodamine-conjugated dextran, provides excellent retrograde labeling following pressure injections, showing the entire dendritic arbor, soma, and axon of the labeled cells. Although the injection site involved a large area in the lateral habenula and the ventrolateral nucleus of the thalamus, the resulting retro- grade labeling was not significantly enhanced due to uptake of tracer by fibers-of-passage, as the pattern of labeling within the EPN was comparable to that following a similar fluorogold injection. Moreover, neither the habenula nor the ventrolateral nucleus of the thalamus contain any EPN efferent fibers directed to other target centers.

292 N. RAJAKUMAR ET AL.
Fig, 4. A, C, E: Fluorescence photomicrographs illustrating retro- gradely labeled neurons in the striosome compartment of the rostral (A, E) and middle (C) levels of the striatum following injection of fluorogold into the rostral EPN. B, D, F: Photomicrographs showing calbindin
immunoreactivity in the same sections seen in A, C, E, respectively. Note that the calbindin-poor striosomes (dashed lines) correspond to the areas containing retrogradely labeled neurons. Asterisks indicate identical fiber bundles in each pair. A-D. ~ 2 5 0 ; E, F: ~ 6 0 0 .

STRIATO-ENTOPEDUNCULAR PROJECTION 293
Fig. 5. A Fluorogold-labeled neurons in the central part of the striatum following injection of tracer into the caudal part of the EPN. B: Photomicrograph showing calbindin immunoreactivity in the same section shown in A. Note that the distribution of retrogradely labeled neurons corresponds to the calbindin-rich matrix. Asterisks indicate identical fiber bundles. A, B: x 250.
Fig. 6. A, B: Schematic drawings of the neostriatum at rostral levels illustrating the topographic distribution of retrogradely labeled neu- rons (black dots) following injection of fluorogold into the rostral (A) and caudal (B) parts of the EPN, respectively. Note that the striatal neurons that project to the rostral EPN are localized mainly within striosomes, whereas those that project to the caudal EPN are localized mostly within the matrix. The calbindin-poor dorsolateral area of the striatum (shown by broken lines) projects to both rostral and caudal parts of the EPN. A, anterior commissure; CC, corpus callosum; NA, nucleus accumbens.
Employing intracellular injections, Kawaguchi et al. ('90) found that striatal matrix neurons projecting to the substan- tia nigra provide axon collaterals to both the globus pallidus and the EPN. Since the extent of the terminal and pretermi- nal axonal field and the efficacy of tracer uptake are important factors in determining the limit of retrograde labeling, detectable retrograde labeling in the striatum following iontophoretic injection of fluorogold into the EPN has been considered an indication of a significant striatal projection to the EPN, either directly or via collateraliza- tion.
Efferent projections of the EPN The present study indicates that the rostral third and the
medial aspect of the middle third of the EPN provide a
5-HT 1 I- DORSAL RAPHE N.
t t I
S.N. COMPACTA
I 1 LATERAL HABENULA
- 1 t CAUDAL 2/3 OF EPN
i M I S.N. RETICULATA
CEREBRAL CORTEX
Fig. 7. Diagramatic representation of the efferent connections of striosome (Sj and matrix (M) compartments of the striatum. Note that the proposed striosome projection to the rostral EPN may control the serotoninergic input (5-HT) to the striatum from the dorsal raphe nucleus via the lateral habenula. The striosome also influences the dopaminergic input (DA) to the striatum through the substantia nigra pars compacta. The matrix directs its projections to the thalamus and to the cerebral cortex via the caudal EPN and the substantia nigra pars reticulata, thereby forming part of the cerebral cortex-basal ganglia- thalamus loop.
substantial input to the ipsilateral habenula, whereas a larger projection to the ipsilateral ventrolateral nucleus of the thalamus arises from the caudal two-thirds of the EPN. The study further demonstrates that the habenular and the thalamic projections arise from two separate populations of neurons, characteristically found in clusters. These observa- tions confirm previous reports on the organization of the efferent projections of the EPN in the rat (Herkenham and Nauta, "77; Van Der Kooy and Carter, '81). By means of the horseradish peroxidase tracing technique, Herkenham and Nauta ('77) found EPN-habenular neurons throughout the rostrocaudal extent of the EPN while Van Der Kooy and Carter ('81), employing the double labeling fluorescence

294
tracing method, observed EPN-habenular neurons in the rostral two-thirds of the EPN and EPN-thalamic neurons in the caudal third of the nucleus. Both these reports claim that the habenular projection is similar in size or larger than the thalamic projection. The present study demon- strates that EPN-thalamic neurons outnumber EPN- habenular neurons, which is compatible with the electro- physiological studies of Filion and Harnois ('78) and Larsen and Sutin ('78) who described a similar organization in the cat EPN. The discrepancy is partly due to the lower sensitivity of the techniques employed in the previous anatomical experiments. The two separate populations of neurons projecting to the lateral habenula and the thala- mus observed in the present study are in agreement with observations in the rat (Van Der Kooy and Carter, '811, cat (Filion and Harnois, '78; Larsen and Sutin, '78) and monkey (Parent and De Bellefeuille, '82). The present study also indicates a separation in the distribution of the dendritic fields of neurons projecting to the habenula and the thalamus within the EPN. The segregated nature of the EPN-habenular and the EPN-thalamic neurons and the preservation of this pattern across the species strongly indicate that these two efferent pathways of the EPN are involved in different aspects of basal ganglia function.
N. RAJAKUMAR ET AL.
Striatal compartmentalization The neostriatal compartments identified in the present
experiment employing somatostatin or calbindin-D28kna im- munoreactivity are identical to the histochemically distinct striosome and matrix compartments described previously by Graybiel and Ragsdale ('78). The ~albindin-&~k~, immu- noreactivity is localized within the cell bodies, fibers, and axon terminals of projection neurons of the matrix, thus enabling the identification of regions innervated by matrix neurons (Gerfen et al., '85).
In this study, the caudal two-thirds of the EPN showed calbindin-D28k~a-immunolabeled fibers, whereas the lateral hypothalamic area and the caudal part of the globus pallidus contained labeled neuronal cell bodies as well as fibers. Nevertheless, injection of fluorogold into the caudal EPN failed to show retrograde labeling in lateral hypotha- lamic neurons or calbindin-containing pallidal or midbrain neurons. Consequently, the immunoreactivity in the EPN is very likely contained within the striatal afferent fibers.
Striatal projection to the EPN The present study indicates that the projection to the
EPN arises from the entire rostrocaudal extent of the lateral two-thirds of the ipsilateral neostriatum. The EPN- habenular area in the rostral part of the EPN receives input from the striosomes in the dorsolateral region of the rostral two-thirds of the neostriatum, whereas the EPN-thalamic area in the middle third of the EPN receives input mainly from the matrix compartment in the caudal part of the neostriatum. The pattern of labeling observed in the pre- sent study suggests that independent rostrocaudally orga- nized topographical projections to the EPN arise from the striosome and matrix compartments of the neostriatum. Van Der Kooy and Carter ('81) employed the autoradio- graphic tracing method to show anterograde labeling in the rostral or caudal part of the rat EPN following injection of tracer into the rostral or caudal region of the neostriatum, respectively. Fink-Jensen and Mikkelsen ('89) found a similar dorsoventral topography in the striato-entopeduncu- lar projection by means of the horseradish peroxidase retrograde tracing method. However, no efforts were made
to characterize the origin of this projection in the rat in terms of striatal compartmentalization.
A preferential projection from the matrix to the globus pallidus has been described in the cat and monkey (Jimenez- Castellanos and Graybiel, '89; Gimenez-Amaya and Gray- biel, '90). In the first study, large wheatgerm agglutinin- conjugated horseradish peroxidase injections in the cat involved either the medial part of the caudal two-thirds or three-fourths, or the lateral part of the caudal three- fourths of the EPN (Jimenez-Castellanos and Graybiel, '89). Following injections of the same tracer into different parts of the internal or the external segments of the globus pallidus in the monkey, Gimenez-Amaya and Graybiel('90) concluded that the matrix is the principal source of projec- tion to the pallidum. However, they further stated that the striosomes embedded in the main fields of matrix labeling did contain variable numbers of labeled neurons. The present finding that the larger part of the EPN containing EPN-thalamic neurons receives a projection from the stria- tal matrix is in agreement with the reports on the cat and the monkey. The discrepancy observed in the striosome projection in the present study and that reported for the cat and the monkey may be partly due to interspecies variation. Nevertheless, employing a more sensitive tracer in combina- tion with smaller circumscribed injection sites would facili- tate the recognition of retrogradely labeled neurons in the striosome compartment, which comprises only 10-20% of the whole neostriatum.
Clustering of neurons Retrogradely labeled neurons appearing in clusters were
frequently seen in all projections examined in the present study including those of the striosome and matrix projec- tion neurons. The aggregation of neurons into small clus- ters has been described in Nissl preparations of the neostri- atum (Mensah, '77). A similar clustering of neurons was reported in the striatopallidal and striatonigral systems in the rat (Loopuijt and Van Der Kooy, '851, cat (Beckstead and Cruz, '86; Desban et al., '89) and monkey (Gimenez- Amaya and Graybiel, '90). Desban et al. ('89) and Gimenez- Amaya and Graybiel ('90) characterized striatal neuronal clusters labeled from the nigra in the cat and the pallidum in the monkey as being in the matrix compartment. Paskev- ich et al. ('91) described the frequent occurrence of discrete neuronal aggregates consisting of as many as sixty cells within larger neuron-dense areas morphologically resem- bling the striosomes in the rat neostriatum. These neurons were further characterised as having direct soma-somatic and soma-dendritic contacts with regions of "junctionlike" specialization. Levine and collaborators (Cepeda et al., '89; Walsh et al., '89) have shown dye-coupling between projec- tion neurons of the rat neostriatum but did not consider their compartmental location. No reports are available concerning the clustering of neurons in the EPN. The neuronal clustering observed in the neostriatum and the EPN in the present study may facilitate synchronization of those neurons that project to a particular target, or form part of a functional module such as the dominance columns described in the visual cortex (Wiesel et al., '74).
Functional considerations The present study has demonstrated that the EPN-
habenular neurons receive a projection from the striosome compartment of the neostriatum, whereas the EPN- thalamic neurons receive an input from the matrix (Fig. 7).
The striosome compartment in the rat has been shown to receive a prominent bilateral projection from the prelimbic

STRIATO-ENTOPEDUNCULAR PROJECTION 295
fully acknowledged. This investigation was supported by the Medical Research Council of Canada.
cortex (Donoghue and Herkenham, '861, which is known to have close connections with limbic centers, particularly the amygdala (Krettek and Price, '78; Kita and Kitai, '90). The amygdala also sends projections to the neostriatum and the nucleus accuinbens (Kita and Kitai, '90). Although amygda- loid fibers are distributed in both compartments, they terminate preferentially in the striosomes (Russchen et al., '85; Ragsdale and Graybiel, '88). Thus, the striosome compartment appears to be influenced mainly by an amyg- dala-centered limbic system, whereas the matrix is involved in the sensorimotor cortical system (Gerfen, '84, '89; Donoghue and Herkenham, '86).
The entopedunculo-thalamic projection, which utilizes GABA as its principal neurotransmitter substance, forms one of the major output pathways of the basal ganglia terminating in the ventrolateral-ventroanterior nuclear complex, a center having direct connections with the supple- mentary and premotor cortices. The striatal projection to the EPN-thalamic neurons of the EPN contains GABA, substance P, and dynorphin B as neuroactive substances (Graybiel, '90). According to the present study, this double inhibitory striatal output pathway originates from the matrix neurons, which are directly controlled by the cortico- striatal input from sensory, motor, and association areas.
The entopedunculo-habenular pathway, which utilizes GABA and somatostatin as its neurotransmitter sub- stances, terminates preferentially in the lateral aspect of the lateral habenula, an area known to receive projections from the prelimbic cortex (Greatex and Phillipson, '82; Vincent and Brown, '86). The lateral part of the lateral habenula provides a large projection to the dorsal and median raphe nuclei and the substantia nigra pars com- pacts (Herkenham and Nauta, '79). Serotoninergic input to the striatum arises mainly from the dorsal raphe nucleus (Steinbusch, '84) and is distributed preferentially in the matrix compartment (Lavoie et al., '88). By means of the in vivo microdialysis technique, Kalen et al. ('89) found that striatal serotonin release is regulated by the habenulo- raphe pathway.
Corticostriatal neurotransmission in the striatal matrix (Smith and Bolam, '90) and the mRNA contents of prepro- enkephalin, preprotachykinin, and glutamic acid decarbox- ylase in the striatal projection neurons are strongly influ- enced by the dopaminergic input from the substantia nigra pars compacta that receives projections from the striosome compartment (Bannon et al., '87; Morris et al., '88b; Vernier et al., '88). Furthermore, striatal prodynorphin mRNA is regulated by the serotoninergic raphe-striatal projection (Morris et al., '88a).
This study indicates that matrix neurons are influenced by the striosome compartment of the striatum through the mesencephalic dopaminergic and serotoninergic systems. This further suggests that in neurodegenerative diseases such as Parkinson's disease and schizophrenia, where both dopaminergic and serotoninergic systems are affected, (1) the involvement of the serotoninergic system may be secondary to the dopamine disturbance in the striosomes, and (2) the pathophysiological changes seen in both dopam- inergic and serotoninergic systems may be influenced by, or be secondary to, an alteration of striosome function.
ACKNOWLEDGMENTS The authors thank Dr. R. Benoit who provided the
somatostatin antibody. The skillful assistance of Mrs. J. Sholdice in preparing the photomicrographs is also grate-
LITERATURE CITED Anden, N.E., A. Carlsson, A. Dahlstrom, K. Fuxe, N.A. Hillarp, and K.
Larsson (1964) Demonstration and mapping out of nigroneostriatal dopamine neurons. Life Sci. 3523-530.
Bannon, M.J., P.J. Elliott, and E.B. Bunney (19871 Striatal tachykinin hiosynthesis: Regulation of mRNA and peptide levels by dopamine agonists and antagonists. Mol. Brain Res. 3131-37.
Beckstead, R.M., and C.J. Cruz (19861 Striatal axons to the globus pallidus, entopeduncular nucleus and substantia nigra come mainly from separate cell populations in the cat. Neuroscience 19:147-158.
Benoit, R., P. Bohlen, N. Ling, F. Esch, A. Baird, S.Y. Ying, W.B. Wehren- berg, R. Guillemin, J.H. Morrison, C. Bakhit, L. Koda, and F.E. Bloom (1985) Somatostatin 28(l-l2)-like peptides. In Y.C. Pate1 and G.S. Tannenbaum (eds): Somatostatin. New York: Plenum Press, pp. 99-107.
Bentivoglio, M., D. Van Der Kooy, and H.G.J.M. Kuypers (1979) The organization of the efferent projection of the substantia nigra in the rat. A retrograde fluorescent double labeling study. Brain Res. 174:l-17.
Besson, M.J., A.M. Graybiel, and M.A. Nastuk (1988) 13H1SCH 23390 binding to D1 dopamine receptors in the basal ganglia of the cat and primate delineation of striosome compartments and pallidal and nigral subdivisions. Neuroscience 26: 101-119.
Carter, D.A., and H.C. Fibiger (1978) The projections of the entopeduncuiar nucleus and globus pallidus in rat as demonstrated by autoradiography and horseradish peroxidase histochemistry. J. Comp. Neurol. 177r113- 124.
Chesselet, M.-F., and A.M. Graybiel (1986) Striatal neurons expressing somatostatin-like immunoreactivity: Evidence for a peptidergic interneu- ronal system in the cat. Neuroscience 17:547-571.
Cepeda, C., J.P. Walsh, C.D. Hull, S.G. Howard, N.A. Buchwald, and M.S. Levine (1989) Dye-coupling in the neostriatum of the rat: I. Modulation by dopamine-depleting lesions. Synapse 4:229-237
Desban, M., C. Gauchy, M.L. Kernel, M.J. Besson, and J. Glowinski (1989) Three-dimensional organization of the striosome compartment and patchy distribution of striatonigral projections in the matrix of the cat caudate nucleus. Neuroscience 29:55 1-566.
Donoghue, J.P., and M. Herkenham (1986) Neostriatal projections from individual cortical fields conform to histochemically distinct striatal compartments in the rat. Brain Res. 365:397-403.
Dube, L., A.D. Smith, and J.P. Bolam (1988) Identification of synaptic terminals of thahmic or cortical origin in contact with distinct medium- size spiny neurons in the rat neostriatum. J. Comp. Neurol. 2671455- 471.
Feigenbaum, L., and A.M. Graybiel (1989) Distinct nigrostriatal projection systems innervate striosomes and matrix in the primate striatum. Brain Res. 498:344-350.
Filion, M., and C. Harnois (1978) A comparison of projections of entopedun- cular neurons to the thalamus, the midbrain and the habenula in the cat. J. Comp. Neurol. 1811763-780.
Fink-Jensen, A., and J.D. Mikkelsen (1989) The striato-entopeduncular pathway in the rat. A retrograde transport study with wheatgerm agglutinin-horseradish peroxidase. Brain Res. 476: 194-198.
Gerfen, C.R. (1984) The neostriatal mosaic: Compartmentalization of corti- costriatal input and striatonigral output systems. Nature 311:461-464.
Gerfen, C.R. (1985) The neostriatal mosaic. I. Compartmental organization of projections from the striatum to the substantia nigra in the rat. J. Comp. Neurol. 236:154-176.
Gerfen, C.R. (1989) The neostriatal mosaic: Striatal patch-matrix organiza- tion is related to cortical lamination. Science 246:385-388.
Gerfen, C.R. (1992) The neostriatal mosaic: Multiple levels of compartmen- tal organization. TINS 15:135-139.
Gerfen, C.R., and W.S. Young 111 (1988) Distribution of striatonigral and striatopallidal peptidergic neurons in both patch and matrix compart- ments: An in situ hybridization histochemistry and fluorescent retro- grade tracing study. Brain Res. 460:161-167.
Gerfen, C.R., K.G. Baimbridge, and J.J. Miller (1985) The neostriatal mosaic: Compartmental distribution of calcium-binding protein and parvalbumin in the basal ganglia of the rat and monkey. Proc. Natl. Acad. Sci. USA 82:8780-8784.
Gerfcn, C.R., M. Herkenham, and J. Thibault (1987) The neostriatal mosaic: 11. Patch- and matrix-directed mesostriatal dopaminergic systems. J. Neurosci. 7:3915-3934.
Gimenez-Amaya, J.M., and A.M. Graybiel(1990) Compartmental origins of the striatopallidal projection in the primate. Neuroscience 341111-126.

296 N. RAJAKUMAR ET AL.
Nauta, W.J.H., and W.R. Mehler (1966) Projections of the lentiform nuclei in the monkey. Brain Res. 1:3-42.
Nauta, W.J.H., M.B. Pritz, and K.R.J. Lasek (1974) Merents to the rat caudoputamen studied with horseradish peroxidase. An evaluation of a retrograde neuroanatomical research method. Brain Res. 67.219-238.
Olson, L., A. Seiger, and K. Fuxe (1972) Heterogeneity ofstriatal and limbic dopamine innervation: Highly fluorescent islands in developing and adult rats. Brain Res. 44283-288.
Parent, A. (1990) Extrinsic connections of the basal ganglia. TINS 13254- 258.
Parent, A., and L. De Bellefeuille (1982) Organization of efferent projections from the internal segment of glohus pallidus in primate as revealed by fluorescence retrograde labeling method. Brain Res. 245:201-213.
Parent, A,, A. Mackey, and L. De Bellefeuille (1983) The subcortical afferents to caudate nucleus and putamen in primate: A fluorescence retrograde double labeling study. Neuroscience 10:1137-1150.
Paskevich, P.A., H.K. Evans, and V.B. Domesick (1991) Morphological assessment of neuronal aggregates in the striatum of the rat. J. Comp. Neurol. 305361-369.
Paxinos, G., and C. Watson (1986) The Rat Brain in Stereotaxic Coordinates. London: Academic Press.
Penny, G.R., C.J. Wilson, and S.T. Kitai (1988) Relationship of the axonal and dendritic geometry of spiny projection neurons to the compartmen- tal organization of the neostriatum. J. Comp. Neurol. 269:275-289.
Pert, C.B., M.J. Kuhar, and S.H. Snyder (1976) Opiate receptors: Autoradio- graphic localization in rat brain. Proc. Natl. Acad. Sci. USA 73:3729- 3733.
Ragsdale, C.W., and A.M. Graybiel (1988) Fibres from the hasolateral nucleus of the amygdala selectively innervate striosomes in the caudate nucleus of the rat. J. Comp. Neurol. 269.506-522.
Ragsdale, C.W., and A.M. Graybiel (1990) A simple ordering of neocortical areas established by the compartmental organization of their striatal projections. Proc. Natl. Acad. Sci. 87.6196-6199.
Rajakumar, N., K. Elisevich, and B.A. Flumerfelt (1991) Segregated efferent projections from neostriatal compartments to the entopeduncular nu- cleus in the rat. Proc. SOC. Neurosci. 17:1300 (Abstract).
Russchen, F.T., I. Bakst, D.G. Amaral, and -J.L, Price (1985) The amygdalos- triatal projections in the monkey. An anterograde tracing study. Brain Res. 329:241-257.
Schmued, L.C., and J.H. Fallon (1986) Fluoro-Gold: A new fluorescent retrograde axonal tracer with numerous unique properties. Brain Res. 377:147-154.
Selemon, L.D., and P.S. Goldman-Rakic (1985) Longitudinal topographyand interdigitation of corticostriatal projections in the Rhesus monkey. J. Neurosci. 5t776-794.
Smith, A.D., and J.P. Bolam (1990) The neural network of the basal ganglia as revealed by the study of synaptic connections of identified neurons. TINS I3:259-265.
Smith, Y., and A. Parent (1986) Differential connections of caudate nucleus and putamen in the squirrel monkey. Neuroscience 18:347-371.
Steinbusch, H.W.M. (1984) Serotonin-immunoreactive neurons and their projections in the CNS. In A. Bjorklund, T. Hokfelt, and M.J. Kuhar (eds): Handbook of Chemical Neuroanatomy, Vol. 3. Amsterdam: Elsevier.
Van Der Kooy, D., and D.A. Carter (1981) The organization of the efferent projections and striatal afferents of the entopeduncular nucleus and adjacent areas in the rat. Brain Res. 211:15-36.
Van Der Kooy, D., and H.G.J.M. Kuypers (1979) Fluorescent retrograde double labeling: Axonal branching in the ascending raphe and nigral projections. Science, Wash. 204:873-875.
Vernier, P., J.F. Julien, P. Rataboul, 0. Fourier, C. Feuerstein, and J. Mallet (1988) Similar time course changes in striatal levels of glutamic acid decarboxylase and proenkephalin mRNA following dopaminergic deafferentation in the rat. J. Neurochem. 51.1375-1380.
Vincent, S.R., and J.C. Brown (1986) Somatostatin immunoreactivity in the entopeduncular projection to the lateral habenula in the rat. Neurosci. Lett. 68:160-164.
Walsh, J.C., C. Cepeda, C. Hull, R. Fischer, M. Levine, and N.B. Buchwald (1989) Dye coupling in the neostriatum of the rat: 11. Decreased coupling between neurons during development. Synapse 4238-247.
Wiesel, T.N., D.H. Hubel, and D.M.K. Lam (1974) Autoradiographic demon- stration of ocular-dominance columns in the monkey striate cortex by means of transneuronal transport. Brain Res. 79273-279.
Graybiel, A.M. (1984) Correspondence between the dopamine islands and striosomes of the mammalian striatum. Neuroscience 13-1157-1187.
Graybiel, A.M. (1990) Neurotransmitters and neuromodulators in the basal ganglia. TINS 13344-254.
Graybiel, A.M., and C.W. Ragsdale (1978) Histochemically distinct compart- ments in the striatum of human, monkey, and cat demonstrated by acetylthiocholinesterase staining. Proc. Natl. Acad. Sci. 755723-5726.
Graybiel, A.M., R.W. Baughman, and F. Eckenstein (1986) Cholinergic neuropil of the striatum observes striosome boundaries. Nature 3233325- 627.
Great,rex, R.M., and O.T. Phillipson (1982) Demonstration of synaptic input from prefrontal cortex to the hahenula in the rat. Brain Res. 238.192- 197.
Herkenham, M., and W.J.H. Nauta (1977) Afferent connections of the habenular nuclei in the rat. A horseradish peroxidase study, with a note on the fibres-of-passage problem. J. Comp. Neurol. 173:123-146.
Herkenham, M., and W.J.H. Nauta (1979) Efferent connections of the habenular nuclei in the rat. J. Comp. Neurol. 187:19-48.
Herkenham, M., and C.B. Pert (1981) Mosaic distribution of opiate recep- tors, parafascicular projections and acetylcholinesterase in rat striatum. Nature 291:4 15-4 18.
Jayaraman, A. (1985) Organization of thalamic projections in the nucleus accumbcns and the caudate nucleus in cats and its relation with hippocampal and other subcortical afferents. J. Comp. Neurol. 231.396- 420.
Jimenez-Castellanos, J., and A.M. Graybiel (1987) Subdivisions of the dopamine-containing AS-Ag-AlO complex identified by their differential mesostriatal innervation of striosomes and extrastriosome matrix. Neu- roscience 23:223-242.
Jimenez-Castellanos, J., and A.M. Graybiel(1989) Compartmental origins of striatal efferent projections in the cat. Neuroscience 32297-321.
Kalen, P., R.E. Strecker, E. Rosengren, and A. Bjorklund (1989) Regulation of striatal serotonin release by the lateral hahenula-dorsal raphe path- way in the rat as demonstrated by in vivo microdialysis: Role of excitatory amino acids and GABA. Brain Res. 492187-202.
Kawaguchi, Y., C.J. Wilson, and P.C. Emson (1990) Projection subtypes of rat neostriatal matrix cells revealed by intracellular injection of hiocytin. J. Neurosci. 10:3421-3438.
Kemp, J.M., and T.P.S. Powell (1970) The cortico-striate projection in the monkey. Brain 93:525-546.
Kita, H., and S.T. Kitai (1990) Amygdaloid projections to the frontal cortex and the striatum in the rat. J. Comp. Neurol. 298:4049.
Krettek, J.E., and J.L. Price (1978) Amygdaloid projections to subcortical structures within the basal forebrain and brainstem in the rat and cat. J. Comp. Neurol. 178t225-254.
Kunzle, H. (1975) Bilateral projections from precentral motor cortex to the putamen and other parts of the basal ganglia. An autoradiographic study in macaca fascicularis. Brain Res. 88: 195-209.
Larsen, K.D., and R.L. McBride (1979) The organization of feline entopedun- cular nucleus projections: Anatomical studies. J. Comp. Neurol. 184.293- 308.
Larsen, K.D., and J. Sutin (1978) Output organization of the feline entopeduncular and subthalamic nuclei. Brain Res. 157:21-31.
Lavoie, B., Y. Smith, and A. Parent (1988) Distribution of serotonin immunoreactivity in primate basal ganglia. Proc. SOC. Neurosci. 14t719 (Abstract).
Loopuijt, L.D., and D. Van Der Kooy (1985) Organization of the striatum: Collateralization of its efferent axons. Brain Res. 3482%-99.
Mensah, P.L. (1977) The internal organization of the mouse caudate nucleus: Evidence for cell clustering and regional variation. Brain Res. 137:53-66.
Morris, B.J., S. Reimer, V. Hollt, and A. Herz (1988a) Regulation of striatal prodynorphin mRNA levels by the raphe-striatal pathway. Mol. Brain Res. 4.15-22.
Morris, B.J.: V. Hollt, and A. Herz (1988b) Dopaminergic regulation of striatal proenkephalin mRNA and prodynorphin mRNA: Contrasting effects of D1 and D2 antagonists. Neuroscience 255255632
Nance, D.M. (1991) Golgi-like labeling of neurons with fluorescent dextrans visualized with confocal scanning laser microscopy. Proc. SOC. Neurosci. I7:372 (Abstract).
Nauta, W.J.H. (1979) Projections of the pallidal complex: An autoradio- graphic study in the cat. Neuroscience 4.1853-1873.