comparison of the properties of native and pentaammineruthenium(iii)-modified xylanase
TRANSCRIPT

IPST Technical Paper Series Number 605
Comparison of the Properties of Native and Pentaammineruthenium(!ll)-modified Xylanase
B.R. Evans, L.M. Lane, R. Margalit, G.M. Hathaway, A. Ragauskas, and J. Woodward
January 1996
Submitted toEnzyme and Microbiology Technology
Copyright® 1996by the Instituteof PaperScienceand Technology
For Members Only

COMPARISON OF THE PROPERTIES
OF NATIVE AND PENTAAMMINERUTHENIUM(III)-MODIFIED XYLANASE
Barbara R. Evans, _'¢ Lynette M. Lane, a Ruth Margalit, b Gary M. Hathaway, _Arthur Ragauskas, a and Jonathan Woodward _
a Oak Ridge National Laboratory _Oak Ridge, TN 37831-6194
b Jet Propulsion Laboratory 2Pasadena, CA 91109
° California Institute of TechnologyBeckman Institute
Pasadena, CA 91125
dlnstitute of Paper Science and TechnologyAtlanta, GA 30318
Corresponding author
'q'he submitted manuscript has been authored by acontractor of the U. S. Government under contract No.
DE-AC05-84OR21400. Accordingly, the U. S.
Government retains a nonexclusive, royalty-freelicense to publish or reproduce the published: form ofthis contribution, or allow others to do so, for U. S.Government purposes."
'Managed by Lockheed Martin Energy Systems, Inc., for the U.S. Department of Energyunder contract DE-AC05-84OR21400.
2Operating Division of California Institute of Technology under contract NAS7-918 withthe National Aeronautics and Space Administration.

Two xylanases, xynA of Bacillus pumilus and xyn II of Trichoderma reesei, were
purified, then modified by the attachment of pentaammineruthenium, resulting in the
generation of a xylanase with veratryl alcohol oxidase activity. Hydrolytic activity of
T. reesei xyn II on soluble xylans was unchanged by modification with
pentaammineruthenium; however, modification of B. pumilus xynA greatly reduced
xylan hydrolysis unless the active site of the xylanaSe was protected with xylose during
the modification. The presence of histidine, cysteine, or reduced glutathione during
xylan hydrolysis greatly increased the xylanase activity of the pentaammineruthenium-
modified B. pumilus xylanase. Glycine, glutamic acid, methionine, or oxidized
glutathione had no effect on xylanase activity.
keywords:
chemical modification/xytanase/mthenium/oxidase

Introduction
Ruthenium complexes generate hydrogen peroxide, superoxide, and hydroxyl
radicals by reaction with molecular oxygen. _'2Pentaammineruthenium can be attached to
proteins by coordination of the metal to a surface-accessible histidine residue of the protein
as a sixth ligand. Attachment of pentaamineruthenium(III) imparts unique redox properties
to myoglobin, azurin, and cytochrome c without perturbing their structure significantly. 3-5
Attachment of pentaammineruthenium to cellobiohydrolase t and II (CBH I and II), the
main cellulases of the fungus Trichoderma reesei, enables the enzymes to oxidize veratryl
alcohol, a substrate of lignin peroxidase, an oxidoreductase activity not present in the
native cellobiohydrolases. 64
Xylanases are used to decrease the mount of chemicals required in the bleaching
of wood pulp for paper manufacture. Improved efficacy of xylanase pretreatment would
further decrease the use of bleaching chemicals and improve the results from "totally
chlorine-free" bleaching. Hardwood xylans, such as birchwood xylan, are composed of
acetyl-4-O-methylglucuronoxylan with a degree of polymerization (DP) of about 200, and
with one in ten of the [_-D-xylopyranoside backbone units having a 1,2-1inked 4-0-
methylglucuronic acid moiety. The xylan found in cereals, such as oat spelt xylan, and
other grasses is arabino-4-methyl-glucuronoxylan with a DP of about 70 and with L-
arabinofuranosyl side chains. 9 The differences between the two types of xylan can affect
the bleaching of different wood pulps. _° The xylanase pretreatment aids the removal of

lignin from the wood pulp by hydrolyzing the xylan backbone of the hemicellulose to
which the lignin is covalently attached. If the xylanase were equipped with a redox group,
the bifunctional enzyme thus designed might be able to directly attack and depolymerize
the lignin in a manner similar to that employed by lignin peroxidases. In order to design
a bifunctional xylanase, we modified two xylanases, xynA from Bacillus pumilus and xyn
II from Trichoderma reesei, by attachment of pentaammineruthenium groups. Both of these
xylanases are small proteins with alkaline pi (molecular weight 22,000 and 21,000, and pis
9.3 and 9.0, respectively). They belong to the G family of xylanases, a group of
homologous small xylanases of alkaline pi that have been isolated from various fungi and
bacteria. 9'_1 The bacterial xylanase, xynA, possesses four histidine residues, at positions 11,
59, 60, and 160 from the amino terminus of the protein; it also has one cysteine residue at
position 82. The fungal xylanase xyn II has histidines at positions 22, 144, and 155 from
the amino terminus. The catalytically active residues of these xylanases had been reported
to be two glutamic acids, at positions 93 and 182 for xynA, 12 and at 86 and 177 for xyn
11.13 Histidine had not been reported to be a catalytically active residue in xylanases, 14q7
so no loss of hydrolytic activity due to attachment of pentaammineruthenium was
anticipated. The properties of the modified xylanases are reported here.
Materials and methods
Purification and modification of enzymes

Two xylanases, xynA of Bacillus pumilus and xyn II of Trichoderma reesei, were
purified from commercially available crude culture filtrates (Pulpzyme HB TM and SP431,
formerly Pulpzyme HA TM, generous gifts from NovoNordisk Biotech, Inc., Danbury, CT).
Purification stages of the proteins were analyzed by sodium dodecyl sulfate (SDS)
and isoelectric focusing polyacrylamide gels.
To purify the B. pumilus xylanase, 10-mi aliquots of culture filtrate were applied
to 5 x 50 cm columns of Biogel P-30 (BioRad) in 5 mM ammonium carbonate, pH 7.8.
Protein fractions from the P-30 chromatography were applied to 1.0 x 9 cm columns of
DEAE Sephadex in the same buffer. The purified xylanase appeared homogeneous on
SDS gels (Figure 1). Xylanase samples were lyophilized, and the dry protein was used for
the modification with pentaammineruthenium(II).
The Trichoderma culture filtrate was found to contain ali of the Trichoderma
cellulases, in addition to the two xylanases. The crude filtrate (30 ml) that had been
dialyzed overnight at 4°C agmst 5 mM ammonium carbonate, pH 7.8, was applied to a 50-
ml column of DEAE Sephadex in 5 mM ammonium carbonate, pH 7.8. At this pH,
xylanase II, with a pi of 9.0, should not bind to the ion-exchange matrix, whereas the
cellulase components should bind to the DEAE Sephadex. About 5-10% cellulase
(cellobiohydrolase II, endoglucanases I and II) was present in the eluted xylanase II
fraction....
Attachment of pentaammineruthenium to the xylanases was carried out as described
previously for CBH I. 6

Analytical methods
The presence of pentaammineruthenium attached to histidine was determined
spectrophotometrically by the appearance of an absorption peak at 360 nm and by the
appearance of a peak at 110 mV in square-wave voltammograms. _8 The ratio of Ru(NH3)s-
histidine to xylanase was calculated from the absorbance of the modified xylanase at 360
nmand the molar absorption coefficient of Ru(NH3)5-histidine (3000 M -_ cm-_), and from
the absorbance of the protein at 280 nm, using a molar absorption coefficient (56,000 M -_
cm -_) that was calculated from the tryptophan and tyrosine content of the protein. _2
Polyacrylamide gel electrophoresis was carried out with a Pharmacia Biotech
Phasff M system using Pharmacia Biotech precast IEF 3-9 and 10-15 gradient gels.
Pharmacia Biotech low-molecular-weight markers and IEF 3-10 markers were prepared and
used according to the manufacturer's instructions. Samples for SDS electrophoresis were
incubated with 2% SDS and 5% [3-mercaptoethanol for 5 min at 90°C before loading them
on the gels.
Assays of enzymatic activity
Oxidoreductase activity was assayed with the lignin peroxidase substrate veratryl
alcohol. 7'_9 Xylanase assays were carried out at 45°C in 50 mM sodium acetate, pH 6.0
(pH 5.0 for the Trichoderma xylanase) with 1% birchwood or oat spelt xylan and 0.010

mg/ml xylanase. Xylanase activity was assayed by measuring reducing sugar production
with the dinitrosalicylic acid reagent. 2° In determining the optimal pH for the B. pumilus
enzyme, 50 mM sodium phosphate buffer was used for the assays at pH 7.0, and 50 mM
Tris-HC1 buffer was used for the assays at pH 8.0, 8.5, or 9.0.
High-pressure liquid chromatography (HPLC) analysis
Purification of xylanase samples prior to tryptic digestion as well as fractionation
of tryptic peptides was carried out with a Perkin-Elmer Applied Biosystems Model 172
microbore liquid chromatography system. For reverse-phase chromatography, a 1.0 x 100
mm Nucleosil C18 column and a 0.5 x 150 mm Reliasil C18 column (Column Engineering,
Ontario, CA) were run using a gradient of 2% acetonitrile/0.05% trifluoroacetic acid to
90% acetonitrile/0.045% trifluoracetic acid, with a flow rate of 0.025 or 0.06 ml/min,
depending on column diameter. Whole xylanase was detected by monitoring at 280 nm;
tryptic peptides, by monitoring at 216 and 305 nm.
Tryptic digests
Samples (1.0 mg/ml) of native and modified xylanase were purified by C 18 reverse-.
phase HPLC before digestion with trypsin. Purified xylanase was dried and then
redissolved in 0.020 ml of 0.2 M ammonium bicarbonate, pH 8.5, to a final reaction

L
volume of 0.030 mi. Then 2 gl of a solution of 1 mg/ml modified trypsin (Promega) was
added, and digestion was allowed to proceed for 12 h at ambient temperature. Digestion
was terminated by the additionof 0.005 m! trifluoroacetic acid, followed by drying the
sample in vacuo.
Mass spectrometry and peptide sequencing
Analysis by mass spectrometry and peptide sequencing was carded out at the Protein
and Peptide Micro Analytical Facility, which is located ai the Beckman Institute, the
California Institute of Technology, Pasadena, California.
Matrix-assisted, time-of-flight (MALDI TOF) mass spectrometry was carried out
with a Lasertech II mass spectrometer (Perseptive Biosystems, Cambridge, MA), equipped
with a reflector, a 1.3-m flight tube, and a nitrogen laser. HPLC fractions (0.5 _1) were
spotted to the sample slides with sinapinic acid (10 mg/ml) and ec-cyano-4-hydroxycinnamic
acid. Acetylangiotensinogen (MW 1801.09) and bovine cytochrome c (MW 12,360.1)
were used as internal and external calibrams.
Peptides were identified by Edman degradation with a model 476 automated
microsequencer (Perkin Elmer/Applied Biosystems, Foster City, CA) and comparison with
the known sequence. The coupling buffer was N-methyl-piperidine, and the samples were
spotted to Porton peptide disks (Porton Instruments Division, Beckman Instruments,

Oxnard, CA).
Results
Purification of xylanases
The bacterial xylanase xynA was reportedly produced in a bacterial strain without
endogenous cellulases and was apparently homogenous, simplifying its purification. The
majority of the modification and pulping experiments were therefore carried out with this
xylanase. Contaminants from the culture media were removed by gel filtration on Biogel
P-30 in 5 mM ammonium carbonate, pH 7.8, followed by ion-exchange chromatography
on DEAE Sephadex in the same buffer (Figure 1).
In the case of the T. reesei xylanase xyn II, _3the culture filtrate obtained from
NovoNordisk contained the entire multicomponent cellulase system of the fimgus with xyn
II making up only about 10% of total protein. The relative amount of xylanase is
increased, but the cellulases are still present in large amounts, 21 necessitating purification
with multiple ion-exchange chromatography steps. To evaluate the suitability of this
enzyme, modification of this partially purified xylanase preparation was carried out. The
attachment of pemaarraninemthenium to xyn II did not adversely affect its xylanase activity
to the same degree as observed in the case of the bacterial xylanase described above. The
hydrolytic activity of pentaammineruthenium-modified xyn II towards birchwood and oat

l0
spelt xylans remained fairly close to that of the native control (modified, 162 and 168 _tmol
mg -_ min -_; native, 207 and 190 _tmol mg -_ min-1).
Properties of pentaammineruthenium(III)-modified xynA
The effect of modification with pentaammineruthenium upon the xylanase xynA
from Bacillus pumilus depended upon the modification conditions. Modification was
carried out for 2 or 4 h with a tenfold excess of aquopentaammineruthenium(II). The
properties of the xylanase after attachment of pentaamineruthenium were the same for the
2- and 4-h modification times, but attachment was found to be more reliable when a 4-h
modification time was used. A substantial loss of activity in hydrolysis of birchwood and
oat spelt xylans was observed for xynA modified with pentaammineruthenium. Loss of
activity was reduced when xynA was modified in the presence of 0.10 M xylose to protect
the active site of the enzyme (Table 1). Veratryl alcohol oxidase activity was higher for
xynA modified without xylose, and the number of pentaammineruthenium groups attached
to the xylanase, based on the absorption peak at 360 nm from the pentaammineruthenium-
histid'me, was 2 to 3, as opposed to 1 for enzyme protected with xylose.
The optimal pH for hydrolysis of birchwood xylan was not changed by the
attachment of pentaamminemthenium. For both native and modified xynA, maximum:.
hydrolysis was observed at pH 6.0 (Figure 2).

11
Effect of _ee amino acids on xylanase activity
When the inactivated xylanase was incubated with 50 mM histidine at pH 6.0,
activity towards xylan was partially restored. No restoration of activity occurred when the
modified xylanase was incubated with 50 mM glycine, implying that the imidazole group
of the histidine was required for restoration of activity (Figure 3). When the xylanase was
incubated with 200 mM histidme for 18 h at 4°C and then filtered on Sephadex G-25 to
remove excess histidine, the absorption peak at 360 nm from Ru(NH3)5-histidine
disappeared (data not shown). Since the absorption peak at 360 nm is characteristic of
pentaamminemthenium-histidine, its disappearance implies that the ruthenium complex was
displaced from the histidine on the xylanase by the excess free histidine. Further
investigations of the effects of various amino acids upon the xylanase activity of
inactivated, pentaammineruthenium-modified xynA are summarized in Table 2. Amino
acids were added directly to control and enzyme reaction mixtures containing 1%
birchwood xylan, and the net production of reducing sugar was determined for the reaction
containing enzyme. At the concentrations used in this study (5-50 mM), histidine doubled
the specific activity of the inactivated xylanase, but reduced glutathione and cysteine
increased it fourfold to fivefold. Oxidized glutathione, glutamic acid, glycine, and
methionine had no effect on the activity of the xylanase (Figure 3 and Table 2). Control
reactions in which xylan was omitted showed no product from the action of the modified
xylanase on cysteine, or on reduced or oxidized glutathione, which gave a reaction with the

12
dinitrosalicylic acid reducing sugar reagent.
Determination of site of attachment of pentaammineruthenium(lll)
To determine the site of attachment of the pentaammineruthenium on the mottled
xynA, Edman sequence determination and mass spectroscopy were carried out in tandem
on whole native and modified xynA and on tryptic peptides prepared from the two proteins.
Before further analysis by mass spectroscopy and tryptic digestion was carried out,
native and pentammineruthenium(I!I)-modified xynA were purified by reverse-phase HPLC
on Nucleosil C18. Mass spectroscopy of the HPLC-purified proteins gave a mass value for
native, unmodified xylanase of 22,577 + 25 amu and a mass value for
pentaammineruthenium(III)-modified xynA of 22,950 + 50 amu. The increase in mass of
370 amu observed for modified xylanase corresponds to that expected for the addition of
two pentaamineruthenium groups (mass 186) to the protein.
Native and pentaamminemthenium(III)-modified xynA were sequenced directly from
the amino terminus using Edman degradation. Peptide sequencing of the whole protein of
pentaammineruthenium(III)-modified xynA identified the first 20 amino acid residues and
determined that the ruthenium complex had not been attached to H11. The N-termin_
amino acid sequence was observed to differ from the published sequence of xynA from
Bacillus pumilus:
published sequence: RTITNNEMGNHSGYDYELWKD

13
observed sequence: ETIYDNRIGTHSGYDFELWKD
Another six differences between the published sequence and the observed sequence were
noted: N44 was changed to K, A159 was changed to E, R161 was missing, M173 was
changed to S, Q184 was changed to S, and S186 was changed to K. Overall, the percent
identity between the published sequence _7 and the sequence determined from the
NovoNordisk Pulpzyme HB TM xylanase was 93%. It appears that the NovoNordisk enzyme
may be isolated from a different strain of Bacillus pumilus than that used in the published
sequence determination.
The tryptic peptides generated from native and modified xylanase were compared
by analysis with HPLC and mass spectroscopy, a method that has been used to determine
the site of attachment of substrate analogs to exoglucanases and [3-glucosidases. 22 Tryptic
digests were carried out on native and modified xynA as described above. The tryptic
peptides were £ractionated by reverse-phase HPLC. The HPLC effluent was monitored at
216 nm to detect peptides and at 305 nm to detect the presenc e of pentaammineruthenium-
histidine (absorbance maximum is shifted to 305 nm at acid pH). When detection was
carried out at 216 nm, a peak present in the column profile of the tryptic digest of native
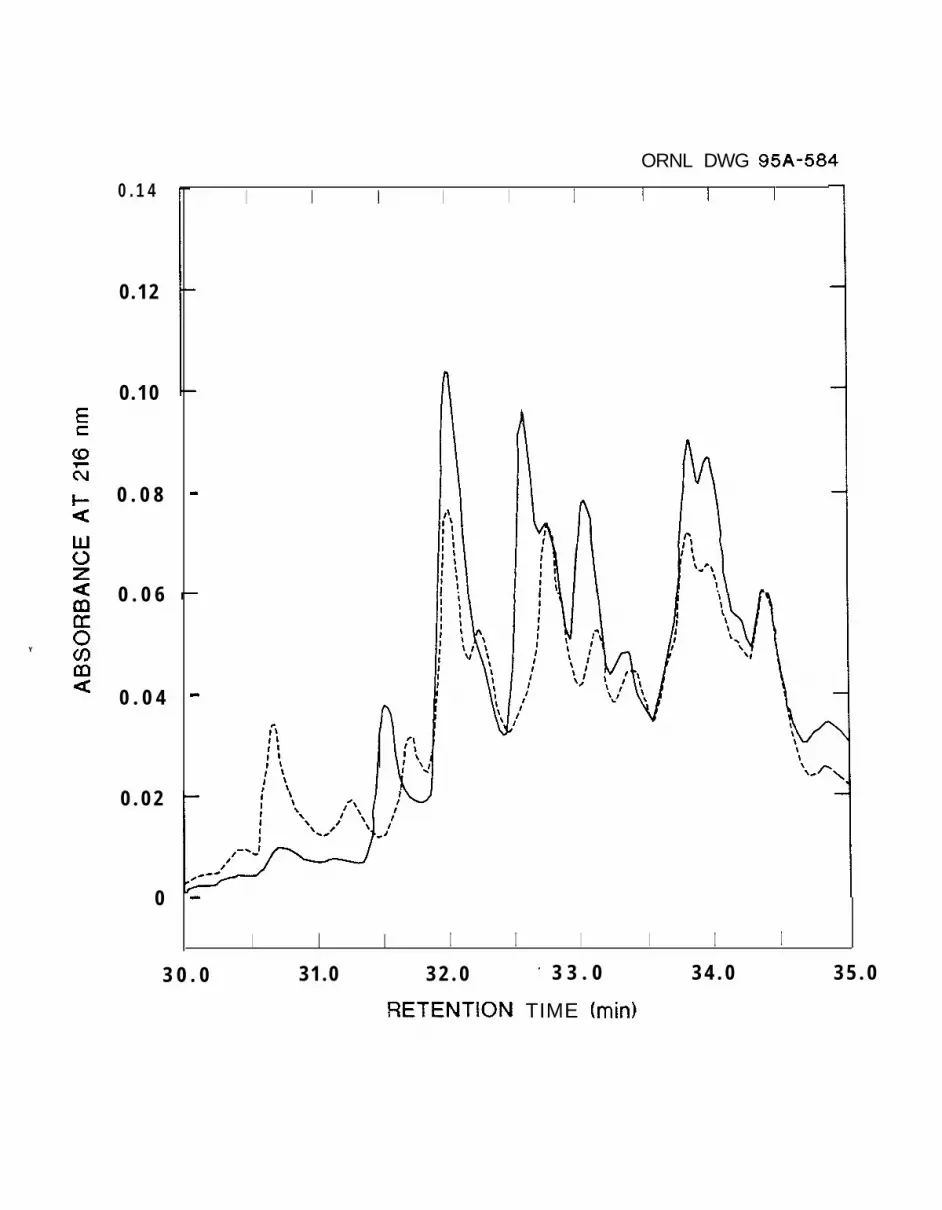
xylanase at 32.5 min was absent from the chromatogram of the pentaammineruthenium(III)-
modified xylanase (Figure 5). This peak, missing from the tryptic digest of the modified
xylanase, was found to contain several peptides, including one with a mass of 1279.5 amu.
The peptides present in the peak were sequenced, and the peptide of mass 1279.5 was
determined to correspond to residues 151-162 of the published sequence.

14
To establish that the peptide corresponding to 151-162 of the published sequence
was the site of attachment of pentaammineruthenium, the chromatogram of the tryptic
peptides from the modified xylanase was examined for the presence of a peptide
corresponding to the mass of the missing peptide plus one or two pentaammineruthenium
groups. When the chromatogram of the tryptic digest of modified xylanase was monitored
at 305 nm, several peaks were detected. One peak, which eluted at 31 min in the HPLC
chromatogram, had a high ratio of absorbance at 305 nm to absorbance at 216 nm,
indicating that it was likely to contain pentaammineruthenium-modified peptides. The
presence of a peptide with mass of 1650.8 was detected in the peak at 31 min from the
tryptic digest of the modified xylanase. The mass of this peptide (1279.5 amu)
corresponds to the mass of the native peptide consisting of residues 151-162, plus a mass
equivalent to two pentaammineruthenium groups. Sequence determination by Edman
degradation confirmed the identity of the peptide present in the peak eluting at 31 min as
corresponding to residues 151-162 of the xylanase'
TSGTVSVSEHFK
No tryptic peptides of mass 1650.8 were detected in the digest of the native xylanase. This
implies that the pentaammineruthenium is attached to the 151-162 peptide, which contains
one histidine at position 160. The location of the attached pentaammineruthenium thus
appears, by a process of elimination, to be the histidine at position 160, an amino acid
residue that is found to be conserved among the family G xylanases. _3 The second
pentaammineruthenium group could be attached to the glutamic acid residue adjacent to the

%
15
histidine on the same peptide.
Determination of kinetic parameters for native and pentaammineruthenium-modified xynA
To investigate the effect of attachment of pentaammuinemthenium to xynA, the
kinetic parameters of native and 4-h pentaammineruthenium-modified xynA were
determined for the hydrolysis of birchwood xylan (Figure 6 A and B). The Km values for
native and modified xynA were determined to be 6.44 and 3.76 mg/ml birchwood xylan,
respectively. For native and modified xynA, the Vma x values were 464 and 141 mM
reducing sugar min -_ (mg xylanase) -_, respectively. Loss of activity upon modification
would thus appear to result from the inability of the modified xylanase to carry out
catalysis, since the Vma x is decreased fourfold compared with the native enzyme. The
decrease in Km upon modification could be the result of a conformational change in the
active site caused by attachment of the pentaamminemthenkun groups.
Discussion
Initial studies have shown that oxidoreductase activity can be conferred upon
xylanases by attachment of pentaammineruthenium groups. The specific activity of the

16
modified xylanases towards the lignin monomeric substrate veratryl alcohol increases with
the number of pentaammineruthenium groups attached per molecule of xylanase.
The loss of xylanase activity observed due to modification of the B. pumilus
xylanase was unexpected, as histidine had not been reported to be a catalytically active
residue in xylanases. Two conserved glutamic acid residues had been reported to be
essential for catalysis of xylan hydrolysis in a number of xylanases. _4-_7 A comparison of
the amino acid sequences of the different xylanases indicates that three histidine residues
(positions 11, 59, and 60) are present in xynA from B. pumilus that are not conserved in
several other family G xylanases _'23 isolated from bacterial and fungal sources, including
T. reesei xyn II. The histidine at position 159 in the B. pumilus xynA is conserved among
ten bacterial and fungal xylanases, including the xylanases from B. pumilus, Bacillus
subtillus, and T. reesei. _3 Examination of the three-dimensional structure of the
xylanase 24'25reveals that His-159 is located inside the active site groove, while His-59 and
His-60 reside on a loop located below the active site. It is possible that this conserved
histidine may help carry out catalysis; thus, its modification results in a loss of hydrolytic
activity. Alternatively, attachment of the pentaammineruthenium group to a residue in the
active site cleft may obstruct access of the catalytic glutamic acid residues to the substrate.
The kinetic analysis of the reaction of native and pentaamminemthenium-modified xynA
with birchwood xylan appears to indicate that binding of the enzyme to the substrate is not
adversely affected and, in fact, would appear to be better for the modified xynA (an
approximately twofold decrease in KM). However, Vma x is decreased about fourfold for the

17
modified xynA, indicating a considerable decrease in the rate of the hydrolysis reaction.
Modification with pentaammineruthenium or other histidine-modifying reagents in
the presence or absence of xylose can be used as a probe of the role of histidines in the
active site of xylanases. A reversibly inactivated xylanase may be useful for applications
requiring strict limitation of the degree of xylan hydrolysis. A method using imidazole
instead of histidine, or a sulfhydryl reagent instead of cysteine, could be devised to increase
the cost-effectiveness of in situ reactivation of pentaammineruthenium-inactivated xylanase.
Treatment of wood pulps with xylanases to reduce the amount of chemicals required
for bleaching is a method that is currently being investigated and is of great interest to the
pulp and paper industries. 1°'26-29In one experiment, bleaching pretreatment of kraft pulps
using xylanases modified with pentaammineruthenium was shown to reduce the requirement
for chlorine dioxide to a greater degree than pretreatment with native xylanases (data not
shown). Further investigation and application of these preliminary results could, if
confirmed, result in improvement of the quality of paper while reducing the requirement
for bleaching chemicals, thus decreasing the amount ofhalogenated organic compounds that
would have to be removed from paper mill effluents. Penta_neruthenium and
pentammineruthenium-modified proteins have been reported to produce reactive oxygen
species and to carry out oxidation of organic compounds. '-3 Xylanases with attached redox
groups such as pentaammineruthenium could also be used to solubilize pulp residues and
depolymerize lignin in the treatment of effluent streams.

18
Acknowledgments
This research was sponsored by the Laboratory Directed Research and Development
Program at Oak Ridge National Laboratory, which is managed by Lockheed Martin Energy
SyStems, Inc., for the U.S. Department of Energy under contract number DE-AC05-
84OR21400. B. R. Evans is a postdoctoral research associate at Oak Ridge National
Laboratories through the Oak Ridge Institute for Science and Education at Oak Ridge
Associated Universities. The Jet Propulsion Laboratory is an operating division of the
California Institute of Technology under contract NAS7-918 with the National Aeronautics
and Space Administration.

19
References
1. Stanbury, D. M., Haas, O., and Taube, H. Reduction of oxygen by ruthenium(II)
ammines. Inorg. Chem. 1980, 19, 518-524.
2. Stanbury, D. M., Mulac, W. A., Sullivan, J. O., and Taube, H. Superoxide reactions
with (isonicotinamide)pentaammineruthenium(II) and (III). Inorg. Chem. 1980, 19,
3735-3740.
3. Margalit, R., Pecht, I., and Gray, H. B. Oxidation-reduction catalytic activity of a
pentaammineruthenium(III) derivative of sperm whale myoglobin, or.Am. Chem.
Soc. 1983, 105, 301-302.
4. Kosti6, N. M., Margalit, R., Che, C.-M., and Gray, H. B. Kinetics of long-distance
ruthenium-to-copper electron transfer in [pentaammineruthenium histi dine- 83] azurin.
J. Am. Chem. Soc. 1983, 105, 7765-7767.
5. Che, C.-M., Margalit, R., Chiang, H.-J., and Gray, H. B. Ruthenium-modified
proteins. Reactions of cis-[Ru(NH3)4(OH2)2]2+ with azurin, myoglobin, and
cytochrome c. Inorg. Chem. Acta 1987, 135, 33-35.

2O
6. Evans, B. R., Margalit, R., and Woodward, J. Attachment of
pentaammineruthenium(III) to Trichoderma reesei cellobiohydrolase I increases its
catalytic activity. Biochem. Biophys. Res. Comm. 1993, 195, 497-503.
7. Evans, B. R., Margalit, R., and Woodward, J. Veratryl alcohol oxidase activity of
a chemically modified cellulase protein. Arch. Biochem. Biophys. 1994, 312,
459 -466.
8. Evans, B. R., Margalit, R., and Woodward, J. Enhanced hydrolysis of soluble
substrates by a metallocellulase with veratryl alcohol oxidase activity. Appl.
Biochem. Biotechnol. 1995, 51/52, 225-239.
9. Coughlan, M. P., and Hazlewood, G. P. [3-1,4-D-Xylan-degrading enzyme systems:
Biochemistry, molecular biology and applications. Appl. Biochem. Biotechnol.
1993, 17, 259-289.
I0. Kantel'men, A., Hortling, B., Sundquist, J., Linko, M., and Viikari, L. Proposed
mechanism of the enzymatic bleaching of kraft pulps with xylanases. Holzforschung
1993, 47, 318-324...

23_
11. Wong, K. K. Y., Tan, L. U. L., and Saddler, J. N. Multiplicity of [3-1,4-xylanase
in microorganisms: Functions and applications. Eno'me Microbiol. Rev. 1988, 52,
305-317.
12. Ko, E. P., Akatsuka, H., Moriyama, H., Shinmyo, A., Hata, Y., Katsube, Y., Urabe,
I., and Okada, H. Site-directed mutagenesis at aspartate and glutamate residues of
xylanase from Bacillus pumilus. Biochem. J. 1992, 288, 117-121.
13. T6rr6nen, A., Mach, R. L., Messner, R., Gonzalez, R., Kalkkinen, N., Harkki, A.,
and Kubicek, C.P. The two major xylanases from Trichoderma reesei:
Characterization of both enzymes and genes. Bio/Technology 1992, 10, 1461-1465.
14. Marui, M., Nakanishi, K., and Yasui, T. Chemical modification of xylanases from
Streptomyces sp. Biosci. Biotech. Biochem. 1993, 57, 62-63.
15. Bray, M. R., and Clarke, A. J. Identification of a glutamate residue at the active
site of xylanase A from Schizophyllum commune. Eur. J. Biochem. 1994, 219,
821-827.

22
16. Paice, M. G., Bourbonnais, R., Desrochers, M., Jurasek, L., and Yaguchi, M. A
xylanase gene from Bacillus subtilis: Nucleotide sequence and comparison with B.
pumilus gene. Arch. Microbiol. 1986, 144, 201-202.
17. Fukusaki, E., Panbangred, W., Shinmyo, A., and Okada, H. The complete nucleotide
sequence of the xylanase (xynA) of Bacillus pumilus. FEBS Lett. 1984, 171,
197-201.
18. Margalit, R., Gray, H. B., Clarke, M. J., and Podbielski, L. Chemical and biological
properties of pentaammineruthenium-bleomycin complexes. Chem.-Biol.
Interactions 1986, 59, 231-245.
19. Tien, M., and Kirk, T. Kent. Lignin-degrading enzyme from Phanerochaete
chrysosporium' Purification, characterization, and catalytic properties of a unique
H202-requiring oxygenase. Proc. Natl. Acad. Sci. USA 1984, 81, 2280-2284.
20. Miller, G. L. Use of dinitrosalicylic acid reagent for determination of reducing
sugar. Anal. Chem. 1959, 31, 426-428.

23
21. Gamerith, G., Grotcher, R., Zellinger, S., Herzog, P., and Kubicek, C. P. Cellulase-
poor xylanases produced by Trichoderma reesei RUT C-30 on hemicellulose
substrates. Appl. Microbiol. Biotechnol. 1992, 38, 3! 5-322.
22. Tull, D., Miao, S., Withers, S. G., and Abersold, R. Identification of derivatized
peptides without radiolabels: Tandem mass spectrometric localization of the tagged
active-site nucleophiles of two cellulases and a [3-glucosidase. Analyt. Biochem.
1995, 224, 509-514.
23. T6rr6nen, A., Kubicek, C. P., and Henrissat, B. Amino acid sequence similarities
between low molecular weight endo-l,4-[3-xylanases and family H cellulases
revealed by clustering analysis. FEBS Lett. 1991, 321, 135-139.
24. Okada, H. Tertiary structure of xylanase and estimation of active sites by site-
directed mutagenesis. GBF Monogr. 1989, 12 (Adv. Prot. Des.), 81-86.
25. Arase, A., Tetsuya, Y., Urabe, I., Hata, Y., Katsube, Y., and Okada, H. Stabilization
of xylanase by random mutagenesis. FEBS Lett. 1982, 316, 123-127.

24
26. Ragauskas, A. J., Poll, K. M., and Cesternino, A. J. Effects of xylanase pretreatment
procedures on nonchlorine bleaching. Enzyme Microb. Technol. 1994, 16, 492-495.
27. Roberts, J. C., McCarthy, A. J., Flynn, N. J., and Broda, P. Modification of paper
properties by the pretreatment of pulp with $accharomonospora viridis xylanase.
Enzyme Microb. Technol. 1990, 12, 210-213.
28. Viikari, L., Kantelinen, A., R_itt6, M., and Sundquist, J. Enzymes in pulp and paper
processing. ACS Symp. Series 1991, 460 (Enzymes Biomass Convers.), 1991,
12-21.
29. Wong, K. K. Y., and Saddler, J. N. Trichoderma xylanases, their properties and
application. Crit. Rev. Biotechnol. 1992, 12, 413-435.

Table 1. Comparison of native and 4-h pentaammineruthenium-modified xylanase xynA
Specific activity a
Xylanase
Birchwood Oat spelt Veratrylpreparation
xylan xylan alcohol
Nativeenzyme 194+ 12 137+ 11 ND
(N= 4) (N= 5)
Trial1--Mod. 19.8 14.5 0.0115
Trial2--Mod. 17.9+ 0.7 14.0+ 13.1 0.0649
(N= 2) (N= 2)
Trial 3--Mod. (+ Xylose) 148+ 1.7 86.1 + 4.2 0.00938
(N=2) (N= 3)
,,
a Specific activities for catalytic activities on the indicated substrates in pmol (mg protein) -_mill -1.

Table 2. Effect of free amino acids on xylanase activity of pentaammineruthenium(III)-
modified xynA
Amino acid added Concentration Specific activitya
(mm)
None 0 18.7+2.9
Glycine 50 17.0
Glutamicacid 50 18.1
Histidine 50 37.1
Methionine 50 20.6
Cysteine 5.0 75.1
Cysteine 10 86.0+2.1
Glutathione(Red.) 10 104.1
Glutathione(Oxid.) 10 21.7
a Specific activity in _mol reducing sugar produced (mg xylanase)-_ min-_ after incubation
for 15 min at pH 6.0 and 45°C from the substrate birchwood xylan (1%).

Figure 1 SDS gel (10-15% gradient) of xynA preparations. Lanes 1 and 6, Pharmacia
LMW markers; Lane 2, Biogel P-30 filtered xynA (Pulpzyme HBTM); Lane 3, xynA after
Biogel P-30 filtration/DEAE Sephadex ion-exchange chromatography; Lane 4, xynA
modified with 50-fold excess pentaamminerathenium for 2 h; Lane 5, xynA modified with
50-fold excess pentaammineruthenium for 4 h. Protein samples were denatured by
incubation with 2.5% SDS and 5% [3-mercaptoethanol for 5 min at 90°C.
Figure 2 Effect of pH on hydrolysis of birchwood xylan by native and
pentaammineruthenium- modified xynA. The amount of reducing sugar produced by a
reaction mixture containing 0.020 mg/ml xylanase, 1% birchwood xylan, and 50 mM
buffer was measured after 15-min incubation at 45°C. Native xynA control = o; xynA
modified for 4 h with pentaammineruthenium = O.

Figure 3 Effect of free amino acids on xylanase activity of 4-h pentaammineruthenium-
modified xynA. Native xynA control = o. Pentaammineruthenium-modified xynA was
incubated with the following amino acids: no amino acids added = · ; with 10 mM
cysteine = V; with 10 mM reduced glutathione = *; with 10 mM oxidized glutathione =
[El; with 50 mM histidine = l; with 50 mM glutamic acid = zx;with 50 mM methionine
= _; with 50 mM glycine = 9.
Figure 4 Chromatograms of the HPLC profiles of tryptic peptides from digestion of native
(_) or pentaamminemthenium-modified (--) xylanase xynA.
Figure 5 A Michaelis Menten plot of kinetic data for native and pentaammineruthenium-
modified xynA hydrolysis of birchwood xylan. Native xynA = o; 4-h
pentaammineruthenium-modified xynA = O. B Double reciprocal plot of kinetic data for
native and pentaamminemthenium-modified xynA. Native xynA = o; 4-h
pentaamminemthenium-modified xynA = O.

abbreviated running title' Pentaammine-ruthenium-modified xylanase

.... _ ....... --"'· T ................. '_r _
MW (XlO-3)
94 '".-i:?'-:'. ' '-'-_ """ .,-..- ' .'
. _ .:. ..'..j - :o
42 :_''- .
3O
1 2 3 4 5 .6

" 0,,o" ' I I ........ I ..... I --(4)
I.,q::LC) -- o · --
('-'_ ·
-- ® -- O0._.JZ
O
-- o -- _,.T¢,)
_ ®
-- o ® LC)
0 ·
I , I, [ !0 LC) 0 LC) 0
(l uu) lvons ONlana3 l

OR
NL
DW
G95A
-644
20..............I
''""'1'
i!
II
I'
-
y,_
_''__
15
--
10/
-....
0..........I.,,
I.........
II,
II
.....I.....
-20
02
04
06
08
01
00
12
014
0
INC
UB
AT
ION
TIM
E(m
in)
ORNL DWG 95A-584
Y
0.14
0.12
0 .10
0 . 0 8
0 . 0 6
0 . 0 4
0 . 0 2
0
I I I I I I I I I
-
-
-
I k I I I I I I I
3 0 . 0 31 .0 3 2 . 0 - 3 3 . 0
f?ETENTtON TIME (mid
3 4 . 0 3 5 . 0

_oI O ·
-<rtn _-
c9
O · -- ,--z
O
O O _ CD
5
Z<I2.._J
0 cO >--X
0
..q-
cq
.
0 u_ 0 LC) O-
(L_u!w ¢qw) _¥ONS ONIOfiCl3_I

to _'i
tOI
0N'""
to
t_
z 0t_0
t.D
0'T'"--
O0 I1--"-I
0 0........
xJ !
0 0
0
e
0I
l I , ]_ [ _'[ i - · ·
_o c_ ,- o I
[-/5

