comparison of the mglu5 receptor positive allosteric modulator adx47273 and the mglu2/3 receptor...
TRANSCRIPT
Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/26823049
ComparisonofthemGlu5receptorpositiveallostericmodulatorADX47273andthemGlu2/3receptoragonistLY354740intestsforantipsychotic-likeactivity
ARTICLEinEUROPEANJOURNALOFPHARMACOLOGY·SEPTEMBER2009
ImpactFactor:2.53·DOI:10.1016/j.ejphar.2009.09.006·Source:PubMed
CITATIONS
43
READS
74
7AUTHORS,INCLUDING:
AndreasGravius
Merz
42PUBLICATIONS639CITATIONS
SEEPROFILE
SergioGreco
Merz
14PUBLICATIONS167CITATIONS
SEEPROFILE
LorenzoMorè
TheUniversityofWarwick
21PUBLICATIONS228CITATIONS
SEEPROFILE
WojciechDanysz
Merz
420PUBLICATIONS11,188CITATIONS
SEEPROFILE
Allin-textreferencesunderlinedinbluearelinkedtopublicationsonResearchGate,
lettingyouaccessandreadthemimmediately.
Availablefrom:LorenzoMorè
Retrievedon:03February2016

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Behavioural Pharmacology
Comparison of the mGlu5 receptor positive allosteric modulator ADX47273 and themGlu2/3 receptor agonist LY354740 in tests for antipsychotic-like activity
Chantal Schlumberger a, Małgorzata Pietraszek a, Andreas Gravius a, Kai-Uwe Klein b, Sergio Greco c,Lorenzo Morè a,1, Wojciech Danysz a,⁎a Dept. In vivo Pharmacology, Merz Pharmaceuticals GmbH, Eckenheimer Landstrasse 100, D-60318 Frankfurt am Main, Germanyb Dept. Non-clinical DMPK, Merz Pharmaceuticals GmbH, Eckenheimer Landstrasse 100, D-60318 Frankfurt am Main, Germanyc Dept. Biological Analytics, Merz Pharmaceuticals GmbH, Eckenheimer Landstrasse 100, D-60318 Frankfurt am Main, Germany
a b s t r a c ta r t i c l e i n f o
Article history:Received 7 February 2009Received in revised form 1 September 2009Accepted 10 September 2009Available online 16 September 2009
Keywords:Prepulse inhibitionLocomotor activitymGlu5 receptor positive allosteric modulatormGlu2/3 receptor agonistADX47273LY354740
Recently, it has been proposed that activation of either metabotropic glutamate receptors e.g. mGlu5 bypositive allosteric modulators or stimulation of mGluR2/3 receptors by agonists may offer new strategy inschizophrenia treatment. The aim of the present study was to compare the effect of mGlu5 receptor positiveallosteric modulator, ADX47273 (S-(4-Fluoro-phenyl)-{3-[3-(4-fluoro-phenyl)-[1,2,4]oxadiazol—5-yl]-pi-peridin-1-yl}-methanone), mGluR2/3 agonist, LY354740 ((1S,2S,5R,6S)-2-aminobicyclo[3.1.0]hexane-2,6-dicarboxylate monohydrate) and selected neuroleptics in animal models for positive schizophreniasymptoms. ADX47273 (3 and 10 mg/kg i.p.), the typical antipsychotic haloperidol (0.1 and 0.2 mg/kg i.p.),the atypical antipsychotics aripiprazole (1.25–5 mg/kg i.p.) and olanzapine (2.5 and 5 mg/kg i.p.) all reducedamphetamine-induced hyperlocomotion in Sprague–Dawley rats, unlike the mGlu2/3 receptor agonistLY354740 (1–10 mg/kg i.p.). Interestingly, haloperidol (0.1 and 0.2 mg/kg i.p.), aripiprazole (1.25–5 mg/kgi.p.) and olanzapine (1.25–5 mg/kg i.p.), but not ADX47273 (1–10 mg/kg i.p.), all reduced spontaneouslocomotion and rearings at doses effective against amphetamine-induced hyperlocomotion. This indicatesthat the effect of ADX47273 in combination with amphetamine may be specific, and also suggests a lack ofsedative side effects. Moreover, ADX47273 (30 mg/kg i.p.), haloperidol (0.1 and 0.2 mg/kg i.p.) andaripiprazole (5 and 10 mg/kg i.p.) reversed apomorphine (0.5 mg/kg s.c.)-induced deficits of prepulseinhibition, whereas neither LY354740 (1–10 mg/kg i.p.) nor olanzapine (1.25–5 mg/kg i.p.) produced thiseffect. Lack of effect of olanzapine was unexpected and at present no convincing explanation can beprovided. In conclusion, in selected rodent models for positive schizophrenia symptoms, ADX47273 showedbetter efficacy than LY354740.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
It is believed that the enhancement of dopaminergic transmissionin subcortical structures may lead to psychotic symptoms inschizophrenia (Meltzer and Stahl, 1976; Snyder et al., 1974), a theorywhich is supported by the efficacy of D2 receptor blockade by typicalneuroleptics (Seeman, 1987; Seeman, 1992).
Glutamatergic transmission may be also involved in the patho-mechanism of schizophrenia, and some NMDA receptor antagonistsinduce psychotic symptoms in healthy volunteers (Adler et al., 1998;Krystal et al., 1994; Luby et al., 1959). Drugs targeting theglutamatergic system, such as the mGlu receptor group II agonistLY404039 ((-)-(1R,4S,5S,6S)-4-amino-2-sulfonylbicyclo[3.1.0]hex-
ane-4,6-dicarboxylic acid), may provide therapeutic benefits forschizophrenic patients (Patil et al., 2007). Furthermore, LY404039shows fewer side effects e.g. prolactine release and weight gain thanolanzapine (Patil et al., 2007). Other mGlu receptor group II agonist,LY354740 ((1S,2S,5R,6S)-2-aminobicyclo[3.1.0]hexane-2,6-dicarbox-ylate monohydrate) was effective against ketamine-induced workingmemory impairment in healthy volunteers (Krystal et al., 2005) andphencyclidine-induced stereotypy, locomotion, and cortical gluta-mate efflux in rats (Moghaddam and Adams, 1998; Schlumbergeret al., 2009). Based on such animal data, it has been suggested thatLY354740's antipsychotic-like mechanism of action involves aninhibition of enhanced glutamate release in the prefrontal cortex.Such enhanced glutamate release resulting from disinhibition insubcortical regions is observed after PCP administration and believedalso to take place in schizophrenia (Moghaddam and Adams, 1998;Krystal et al., 2003; Conn et al., 2009). Preclinical experiments suggestthat also mGlu5 receptor positive allosteric modulators may havetherapeutic potential for this indication (Darrah et al., 2008; Kinney
European Journal of Pharmacology 623 (2009) 73–83
⁎ Corresponding author. Tel.: +49 69 1503 564; fax: +49 69 1503 795.E-mail address: [email protected] (W. Danysz).
1 Present address: School of Psychology, University of Nottingham, University Park,NG7 2RD Nottingham, UK.
0014-2999/$ – see front matter © 2009 Elsevier B.V. All rights reserved.doi:10.1016/j.ejphar.2009.09.006
Contents lists available at ScienceDirect
European Journal of Pharmacology
j ourna l homepage: www.e lsev ie r.com/ locate /e jphar

Author's personal copy
et al., 2005; Liu et al., 2008). The use of mGlu5 receptor positiveallosteric modulators for the treatment of schizophrenia is supportedby the fact that mGlu5 receptor knock-out mice (Kinney et al., 2003;Brody et al., 2004) have deficits in prepulse inhibition of the acousticstartle response which are alleviated by chronic clozapine adminis-tration (Gray et al., 2009). Similarly, blockade of the mGlu5 receptorsenhances impairment of prepulse inhibition produced by NMDAreceptor antagonists (Henry et al., 2002; Pietraszek et al., 2005).Prepulse inhibition impairment is observed both in schizophrenicpatients (Braff et al., 1978), and in animals, following dopaminomi-metics and NMDA receptor antagonists (Geyer et al., 2001; Swerdlowet al., 2008). Such deficit can be alleviated in animals by antipsychotics(Geyer et al., 2001; Swerdlow et al., 2008). Similarly, the mGlu5receptor positive modulator CDPPB (3-cyano-N-(1,3-diphenyl-1H-pyrazol-5-yl)benzamide) reduced amphetamine-induced deficits ofprepulse inhibition (Kinney et al., 2005; Lindsley et al., 2004). CDPPBaswell as anothermGlu5 receptor positivemodulator ADX47273 (S-(4-Fluoro-phenyl)-{3-[3-(4-fluoro-phenyl)-[1,2,4]oxadiazol—5-yl]-piperi-din-1-yl}-methanone) attenuated amphetamine-induced locomotoractivity, a standard preclinical test for assessment of antipsychotic-likeactivity (Kinney et al., 2005; Liu et al., 2008).
The aim of the present studywas therefore to compare the effect ofthe mGlu5 receptor positive allosteric modulator ADX47273 and themGlu2/3 receptor agonist LY354740 (representing a novel group ofantipsychotics lacking direct effects on the dopaminergic system)with selected neuroleptics in animal models relevant for the positivesymptoms of schizophrenia such as amphetamine-induced hyperlo-comotion and apomorphine-induced deficits of prepulse inhibition.
2. Material and methods
2.1. Animals
Male Sprague–Dawley rats obtained from Élevage Janvier (LeGenest Saint Isle, France) were used at a weight of 250–380 g. The ratswere allowed to acclimate for one week before the start of theexperiments, housed in groups of 4–5 rats per cage, and providedwithbedding, paper cloth and a red tunnel. Animals were kept in standardconditions (21±1 °C, 60±3%, water and chow ad libitum) under a12/12 h light–dark cycle (lights on at 7 a.m.). Experiments werecarried out between 9 a.m. and 6 p.m. The rats were acclimatised tothe experimental rooms for at least 30 min before all experiments.
All studies are approved by the Ethical Committee, Regierung-spraesidium Darmstadt, Hessen and were performed in accordancewith the recommendations and policies of the U.S. National Institutesof Health Guidelines for the Use of Animals.
2.2. Assessment of plasma and brain concentration of ADX47273
Rats were anaesthezised with a ketamine (90 mg/kg)/xylazine(10 mg/kg) mixture, and cannulated with silicone tubing via the rightjugular vein. Two days later, prior to the first blood sampling, animalswere connected to a counterbalanced system and tubing to performblood sampling in the freely moving rat. Baseline blood samples(200 µl) were taken immediately before application (predose)followed by the sampling 5, 10, 20, 40, 60, 80, 100 min, 2, 3, 4, 6, 8,12, 18, 24 and 28 h after application. Blood was collected inheparinised polypropylene tubes stored on ice and subsequentlycentrifuged at 645g for 10 min at 4 °C. The harvested plasma was keptat −20 °C until being assayed.
To 50 µl of rat plasma sample and calibration standard, 100 µlacetonitrile containing the internal standard (Griseofulvin, 300 ng/ml)was added. Samples and calibration standardswere vigorously shaken(10 s) and after 5 min at room temperature centrifuged for 10 min at6000×g and 20 °C. The particle free supernatant was diluted 1:1 withwater and an aliquot of the mixture was transferred to 200 µl sampler
vials and subsequently subjected to LC-MS/MS (high performanceliquid chromatography/mass spectrometry) with an injection volumeof 15 µl.
The HPLC pump flow rate was set to 300 µl/min and the test itemwas separated on a Gemini C6-Phenyl, 3 µm, 50×2.0 mm (Phenom-enex, Germany) analytical column with a pre-column (Gemini C6-Phenyl, 3 µm, 4×2.0 mm). Gradient elution (is used to accelerate theelution of strongly retained solutes by constantly changing thecomposition, and hence the polarity, of the mobile phase) with10 mM ammonium formate/0.1% formic acid as aqueous phase (A)and acetonitrile/0.1% formic acid as organic phase (B) was used for allitems: % B (t (min)), 5(0–0.2)–97(1.2–4.0)–5(4.2–6.0).
In a separate experiment plasma and brain concentrations werecompared. 30 min after i.p administration of ADX47273, rats weregiven an overdose of pentobarbital (150 mg/kg i.p.) and blood wastaken by cardiac puncture and the brains were removed as well. Bloodwas put in a vial with EDTA to avoid coagulation and was centrifugedafter 30 min (5000×g for 10 min). Plasma and brains were stored at−20 °C until analysis. The LC-MS/MS system used for analysis wasequipped with an Agilent 1100 HPLC coupled to an API 4000 Q Trapmass spectrometer with an electrospray ionization (ESI) source(Applied Biosystems, Darmstadt, Germany). The analytical columnwas a Symmetry C18, 5 µm, 50×2.1 mm (Waters, Eschborn,Germany). The mobile phase consisted of 60% water and 40%acetonitrile both containing 0.1% formic acid. The chromatographicrun was performed over 5 min and consisted of a gradient from 40%acetonitrile in water at the start, to a mobile phase with a compositionof 90% acetonitrile in water. For precipitation of proteins from theplasma, 100 µl of the internal standard (2-methyl-6-(phenylethynyl)pyridine hydrochloride, 10.2 ng/ml in acetonitrile) solution wereadded to a 20 µl sample and then mixed for 15 s using a vortex mixer.After centrifugation (10 min, 20,800g at 2 °C), 2 µl supernatant wastransferred to an HPLC vial with an insert and analyzed by LC-MS/MS.
For precipitation of brain protein, 4 ml aCSF (147 mM NaCl,2.7 mM KCl, 1.2 mM CaCl2, 0.85 mM MgCl2, pH 5.3) were added per1 g brain and homogenized using an Ultra Turrax. 660 µl of theinternal standard solution was then added to 330 µl of the sample andmixed for 15 s using a vortex mixer. After centrifugation (10 min,20,800g at 2 °C), the supernatant was transferred to an HPLC vial withan insert and analyzed by LC-MS/MS. The lowest limit of detection forADX47273 was 3 nM.
2.3. Locomotor activity
For the measurement of locomotor activity, Perspex® boxes (ENV-515-16, 43.2×43.2×30 cm) were placed in a noise-proof chamberequipped with a ventilator and a source of white light (5.6 W) at55 cm above a white floor (Med Associates Inc., St Albans, VT, USA).Four arrays of 16 infrared photo beams placed 3 cm above the floormeasured horizontal activity. Measurement of vertical activity wasassessed by two additional sets of 16 photo beams placed 15 cm abovethe floor. The output from the counters was integrated and analyzedon-line by a PC computer. Distance travelled and vertical movementswere assessed by further analysis as measures of locomotion andrearing, respectively. For interaction studies of antipsychotics orADX47273with amphetamine, animals were injectedwith ADX47273or antipsychotics and placed into the locomotor activity chambers for30 min before amphetamine was administered. For studies involvingthe interaction of LY354740 and amphetamine, animals were firsthabituated to the experimental chambers for 20 min and theninjected with LY354740. Amphetamine was administered 20 minafter LY354740. All measurements started directly after administra-tion of amphetamine and were continued for a total of 120 min. Dataare only shown from after the injection of amphetamine. For themeasurement of the effect of ADX47273 and neuroleptics onspontaneous locomotion and rearings, all compounds were injected
74 C. Schlumberger et al. / European Journal of Pharmacology 623 (2009) 73–83

Author's personal copy
30 min before start of measurement. A maximum of 8 rats fromdifferent groups were tested simultaneously in 8 open-field boxes in apseudorandom order.
2.4. Prepulse inhibition
Four ventilated, sound-proof startle chambers were used formeasurement of startle responses (Med Associates, St Albans, VT,USA), each containing acrylic animal holders (21 cm×9 cm) whichwere fastened via screws onto a piezoelectric force transducer. Forcetransducers were connected to an analogue-to-digital converter (MedAssociates, Model PHM-250B, St Albans, VT, USA) and connectedthrough an interface to an IBM-PC. Both non-startle-eliciting(prepulses) and startle-eliciting (pulse) noise bursts were generatedby a noise generator (Med Associates, Model PHM-255A, St Albans,VT, USA), whichwas connected to two loudspeakers (one for prepulseand pulse, a second one for white background noise (64 dB SPL)).Speakers were situated 7 cm from the animal holder in the back of thechamber. Each force transducer was calibrated prior to each testsession.
On the first day, animals were handled and placed into startlechambers for a 10 min period during which the 64 dB backgroundnoises was presented. On the next day, and two to three days beforethe proper test, animals were pretested in order to divide them intogroups with comparable prepulse inhibition levels and startleamplitude values.
The proper experiment started with a 5 min adaptation periodwith a 64 dB background white noise, which was continuedthroughout the session. Following the adaptation period, the startlesession began with 5 initial startle stimuli (118 dB SPL bursts of whitenoise, 40 ms duration) in order to partially habituate the animals tothe startle-eliciting stimulus. Rats exhibit a fast habituation to the firstfew presentations of startle stimuli, followed by stable reaction tonext presentations (Geyer et al., 1990). The first block of 5 startlestimuli was thus not included in the analysis. After the 5 initial stimuli,rats were presented with 84 trials which involved different types ofstimulus, followed by 5 final startle pulses (data not taken foranalysis). The 84 trials of different stimulus types included startlepulses (n=12), prepulses (68, 72 or 76 dB SPL noise burst pulses,20 ms duration; each n=8), prepulses in combination with a startlepulse (one type of prepulse was presented before a startle noise at aninterval of 120 ms from the prepulse's onset to pulse's onset, eachn=12) as well as no stimulus trails (only white background noisewas presented, n=12). The intertrial interval was varied between 7and 23 s, and the test lasted for 27 min.
LY354740 was injected i.p. 35 min prior to apomorphine, andADX47273 or antipsychotics were injected i.p. 20 min beforeapomorphine. Apomorphine was injected s.c. 10 min before thestart of measurements.
2.5. Data analysis and statistics
2.5.1. Locomotor activityLocomotion distances and rearing counts are expressed as mean±
S.E.M.. The time course of interactions between ADX47273, LY354740and the antipsychotics on amphetamine-induced hyperlocomotionwas analyzed via two-way ANOVA on repeated measurements,followed by Duncan's tests if significant. The influence of ADX47273,LY354740 and the antipsychotics on spontaneous horizontal locomo-tion, measured by recording the total distance travelled, was analyzedby one-way ANOVA followed by Duncan's test. The influence ofcompounds on spontaneous vertical locomotion (rearing) or amphet-amine-induced changes of vertical locomotion (rearing), measured byrecording the total rearing counts, was analyzed by one-way ANOVAfollowed by Duncan's if significant.
2.5.2. Prepulse inhibitionThe startle amplitude for each trial type presentation was
calculated as a mean value from 12 trials (for each trial, the maximumpeak to peak value was recorded in the 200 ms following the onset ofthe startle stimulus). Prepulse inhibition was calculated as thepercentage inhibition of the startle amplitude evoked by the pulsealone: %prepulse inhibition=100×((magnitude of the response inthe pulse-alone trial−magnitude of response in the prepulse+pulsetrial)/magnitude of the response in the pulse-alone trial). The meanstartle amplitude from the pulse-alone trial was analyzed by one-wayANOVA followed by Duncan's test if significant. The mean %prepulseinhibition was analyzed by two-way ANOVA on repeated measure-ments followed by Duncan's test if significant.
2.6. Drugs
ADX47273 (i.p.; synthesized by Merz Pharmaceuticals GmbH,Frankfurt am Main, Germany), was dissolved in 70% dimethylsulfoxide : 30% polyethylene glycol 400. LY354740 was dissolved inwater containing Tween 80 (10%). Amphetamine (i.p.; Sigma-Aldrich,Taufkirchen, Germany) was dissolved in saline, and apomorphine(0.5 mg/kg s.c.; TOCRIS, Bristol, UK)was dissolved in 0.1% ascorbic acidin water (the pH was adjusted to 6 with 1 N NaOH). Olanzapine (i.p.;Sequoia Research Products Ltd., Pangbourne, UK) was dissolved in asmall amount of 1 N HCl and diluted to the final concentration withwater. pHwas adjusted to 6with 1 NNaOH. Aripiprazole (i.p.; SequoiaResearch Products Ltd., Pangbourne, UK) was suspended in watercontaining Tween 80 (10%). Haloperidol (i.p.; Haldol-Janssen, Janssen-Cilag, Beerse, Belgium, ampoules of 5 mg/1 ml solution)was diluted tothe final concentration with physiological saline. All substances wereinjected in a volumeof 1 ml/kg except for aripiprazole,whichwas usedin a 2 ml/kg volume.
The injection times for LY354740 were chosen according to ourprevious behavioural experiments (Schlumberger et al., 2009) andprevious study (Johnson et al., 2002) showing that after p.o. adminis-tration, maximal plasma levels are reached rapidly (30 min) and staystable up to 4 h. Thus, it can be assumed that after i.p. administration itshould be even faster. Therefore, selected injection timeof 45 min seemsto correspond to high, nearly maximal plasma exposure.
3. Results
3.1. Concentration of ADX47273 in plasma and in the brain
ADX47273 (MW 369.37) reached high plasma levels already 5 minafter i.p. administration and then concentration decreased gradually(Fig. 1, for clarity only values for the first 4 h are shown). After this
Fig. 1. Plasma concentration of ADX47273 after i.p. administration of 10 mg/kg. Valuesare means±S.E.M of 4 animals (serial sampling in same subjects). See text for details ofpharmacokinetic parameters.
75C. Schlumberger et al. / European Journal of Pharmacology 623 (2009) 73–83
Author's personal copy
treatment Cmax (ng/ml) was 1171.1; tmax (h) was 0.1; t1/2z (h, terminalhalf-life, 2–18 h)was 3.4 and AUC0-∞ (ng⁎h/ml)was 1265.6. Dispositionphase half-life taking into account 0.33 to 1.67 h was 0.8 (h, 48 min).
In a separate experiment, plasma and brain homogenates levelswere compared at 30 min after i.p. administration i.e. time intervalused formost of the experiments. At 10 mg/kg, ADX47273had reached10.05 µM±2.71 µM (n=4) concentrations in brain homogenates and1.83 µM±0.54 µM (n=4) in plasma indicating clearly that this agentpenetrates to the brain and is even accumulated in this tissue.
3.2. Amphetamine-induced hyperlocomotion
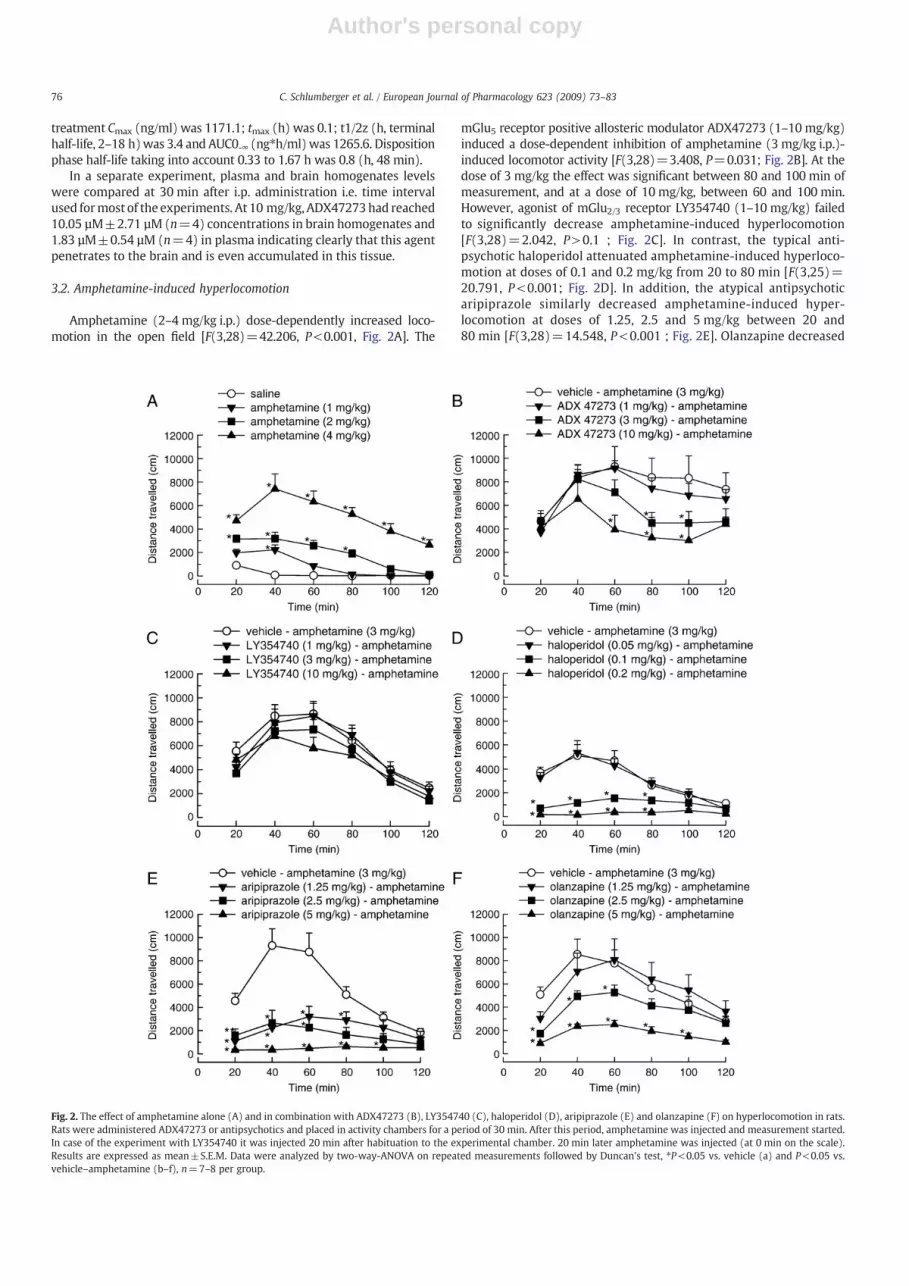
Amphetamine (2–4 mg/kg i.p.) dose-dependently increased loco-motion in the open field [F(3,28)=42.206, P<0.001, Fig. 2A]. The
mGlu5 receptor positive allosteric modulator ADX47273 (1–10 mg/kg)induced a dose-dependent inhibition of amphetamine (3 mg/kg i.p.)-induced locomotor activity [F(3,28)=3.408, P=0.031; Fig. 2B]. At thedose of 3 mg/kg the effect was significant between 80 and 100 min ofmeasurement, and at a dose of 10 mg/kg, between 60 and 100 min.However, agonist of mGlu2/3 receptor LY354740 (1–10 mg/kg) failedto significantly decrease amphetamine-induced hyperlocomotion[F(3,28)=2.042, P>0.1 ; Fig. 2C]. In contrast, the typical anti-psychotic haloperidol attenuated amphetamine-induced hyperloco-motion at doses of 0.1 and 0.2 mg/kg from 20 to 80 min [F(3,25)=20.791, P<0.001; Fig. 2D]. In addition, the atypical antipsychoticaripiprazole similarly decreased amphetamine-induced hyper-locomotion at doses of 1.25, 2.5 and 5 mg/kg between 20 and80 min [F(3,28)=14.548, P<0.001 ; Fig. 2E]. Olanzapine decreased
Fig. 2. The effect of amphetamine alone (A) and in combination with ADX47273 (B), LY354740 (C), haloperidol (D), aripiprazole (E) and olanzapine (F) on hyperlocomotion in rats.Rats were administered ADX47273 or antipsychotics and placed in activity chambers for a period of 30 min. After this period, amphetamine was injected and measurement started.In case of the experiment with LY354740 it was injected 20 min after habituation to the experimental chamber. 20 min later amphetamine was injected (at 0 min on the scale).Results are expressed as mean±S.E.M. Data were analyzed by two-way-ANOVA on repeated measurements followed by Duncan's test, ⁎P<0.05 vs. vehicle (a) and P<0.05 vs.vehicle–amphetamine (b–f), n=7–8 per group.
76 C. Schlumberger et al. / European Journal of Pharmacology 623 (2009) 73–83

Author's personal copy
amphetamine-induced hyperlocomotion from 20 to 60 min at2.5 mg/kg, whereas 5 mg/kg olanzapine almost abolished amphet-amine-induced hyperlocomotion from 20 to 100 min [F(3,28)=6.337, P=0.002; Fig. 2F].
3.3. Rearing behaviour during amphetamine-induced changes in rearingbehaviour
Amphetamine significantly increased the vertical activity (rearing)at doses 2 and 4 mg/kg [F(3,28)=12.579, P<0.001; Fig. 3A].The amphetamine-induced enhancement of rearings was changedneither by ADX47273 [F(3,28)=0.865, P>0.1; Fig. 3B] nor byLY354740 [F(3,26)=0.862, P>0.1; Fig. 3C] at any of the doses tested.However, the antipsychotics haloperidol [F(3,25)=11.420, P<0.001;Fig. 3D], aripiprazole [F(3,28)=5.828, P=0.003; Fig. 3E] andolanzapine [F(3,28)=10.115, P<0.001; Fig. 3F] dose-dependentlyalleviated amphetamine-induced rearings.
3.4. Spontaneous locomotor activity
In contrast to all tested antipsychotics, ADX47273 was void of anyeffect on spontaneous locomotion at all doses used [F(3,27)=1.827,P>0.1; Fig. 4A], and LY354740 has previously been shown by us tohave no effect on spontaneous locomotion in the same experimentalset-up (Schlumberger et al., 2009). Haloperidol decreased spontaneousexploration at the doses of 0.1 and 0.2 mg/kg [F(3,28)=24.233,P<0.001; Fig. 4B] and, at all doses tested, aripiprazole (1.25–5 mg/kg)strongly attenuated spontaneous locomotion, to approx. one third ofcontrol levels [F(3,28)=47.020, P<0.001; Fig. 4C]. Finally, olanzapine(1.25–5 mg/kg) dose-dependently alleviated and even abolishedspontaneous locomotion [F(3,28)=68.150, P<0.001; Fig. 4D].
3.5. Rearing behaviour during spontaneous locomotion
ADX47273 lacked any effect on rearing [F(3,27)=0.401, P>0.1;Fig. 5A] in contrast to all of the antipsychotics tested here. LY354740
Fig. 3. Effects of (A) amphetamine alone or in combination with ADX47273 (B), LY354740 (C), haloperidol (D), aripiprazole (E) and olanzapine (F) on total number of rearings. Ratswere administered ADX47273 or antipsychotics and placed in activity chambers for a period of 30 min. After this period, amphetamine was injected and measurement started.In case of the experiment with LY354740 it was injected 20 min after habituation to the experimental chamber. 20 min later amphetamine was injected. Results are expressed asmean±S.E.M. Data were analyzed by one-way-ANOVA followed by Duncan's test, ⁎P<0.05 vs. vehicle (A) and ⁎P<0.05 vs. vehicle–amphetamine (B–F), n=7–8 per group.
77C. Schlumberger et al. / European Journal of Pharmacology 623 (2009) 73–83

Author's personal copy
decreased rearing activity in the first 10 min of measurement, as hasbeen shown elsewhere (Schlumberger et al., 2009). Haloperidol dose-dependently decreased rearing behaviour at the doses of 0.1 and0.2 mg/kg [F(3,28)=15.989, P<0.001; Fig. 5B]. Moreover, aripipra-zole [1.25–5 mg/kg, F(3,28)=18.281, P<0.001; Fig. 5C] reduced, andolanzapine [F(3 ,28)=58.861, P<0.001; Fig. 5D] nearly abolishedrearing behaviour at all doses tested.
3.6. Effect of substances on apomorphine-induced deficit of prepulseinhibition and its changes to startle amplitude
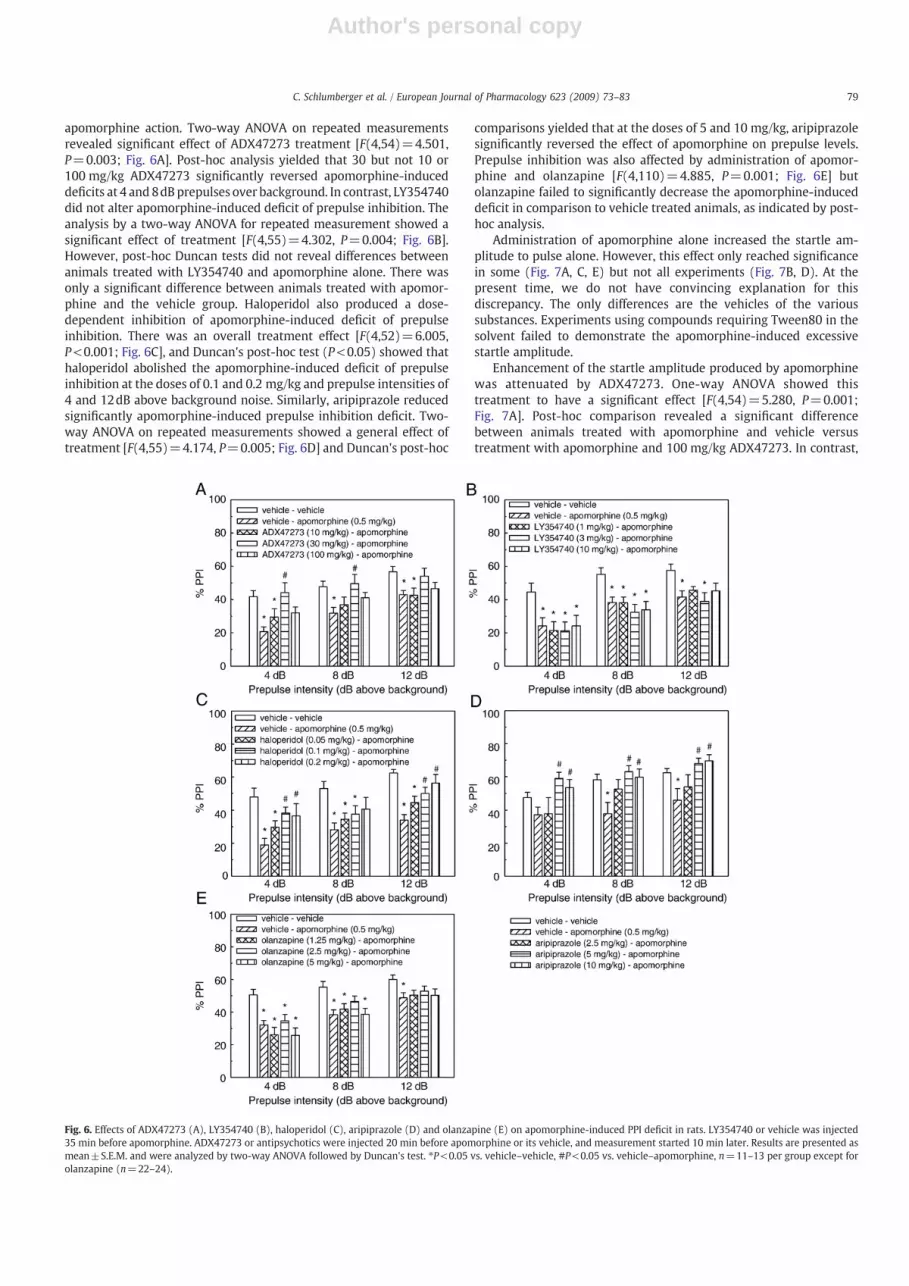
Apomorphine (0.5 mg/kg s.c.) induced a disruption of prepulseinhibition. ADX47273 (30 mg/kg) attenuated the apomorphine-induced impairment of prepulse inhibition, but the lowest and highesttested doses (10 and 100 mg/kg) failed to attenuate significantly
Fig. 4. Effects of ADX47273 (A), haloperidol (B), aripiprazole (C) and olanzapine (D) on spontaneous horizontal locomotion in rats. All substances were administered 30 min beforestart of measurement. Results are expressed as mean±S.E.M. and data were analyzed by one-way-ANOVA followed by Duncan's test, ⁎P<0.05 vs. vehicle, n=7–8 per group.
Fig. 5. Effects of ADX47273 (A), haloperidol (B), aripiprazole (C) and olanzapine (D) on vertical activity (rearings) in rats. All substances were administered 30 min before start ofmeasurement. Results are expressed as mean±S.E.M. and data were analyzed by one-way-ANOVA followed by Duncan's test, ⁎P<0.05 vs. vehicle, n=7–8 per group.
78 C. Schlumberger et al. / European Journal of Pharmacology 623 (2009) 73–83
Author's personal copy
apomorphine action. Two-way ANOVA on repeated measurementsrevealed significant effect of ADX47273 treatment [F(4,54)=4.501,P=0.003; Fig. 6A]. Post-hoc analysis yielded that 30 but not 10 or100 mg/kg ADX47273 significantly reversed apomorphine-induceddeficits at 4 and 8dB prepulses over background. In contrast, LY354740did not alter apomorphine-induced deficit of prepulse inhibition. Theanalysis by a two-way ANOVA for repeated measurement showed asignificant effect of treatment [F(4,55)=4.302, P=0.004; Fig. 6B].However, post-hoc Duncan tests did not reveal differences betweenanimals treated with LY354740 and apomorphine alone. There wasonly a significant difference between animals treated with apomor-phine and the vehicle group. Haloperidol also produced a dose-dependent inhibition of apomorphine-induced deficit of prepulseinhibition. There was an overall treatment effect [F(4,52)=6.005,P<0.001; Fig. 6C], and Duncan's post-hoc test (P<0.05) showed thathaloperidol abolished the apomorphine-induced deficit of prepulseinhibition at the doses of 0.1 and 0.2 mg/kg and prepulse intensities of4 and 12dB above background noise. Similarly, aripiprazole reducedsignificantly apomorphine-induced prepulse inhibition deficit. Two-way ANOVA on repeated measurements showed a general effect oftreatment [F(4,55)=4.174, P=0.005; Fig. 6D] and Duncan's post-hoc
comparisons yielded that at the doses of 5 and 10 mg/kg, aripiprazolesignificantly reversed the effect of apomorphine on prepulse levels.Prepulse inhibition was also affected by administration of apomor-phine and olanzapine [F(4,110)=4.885, P=0.001; Fig. 6E] butolanzapine failed to significantly decrease the apomorphine-induceddeficit in comparison to vehicle treated animals, as indicated by post-hoc analysis.
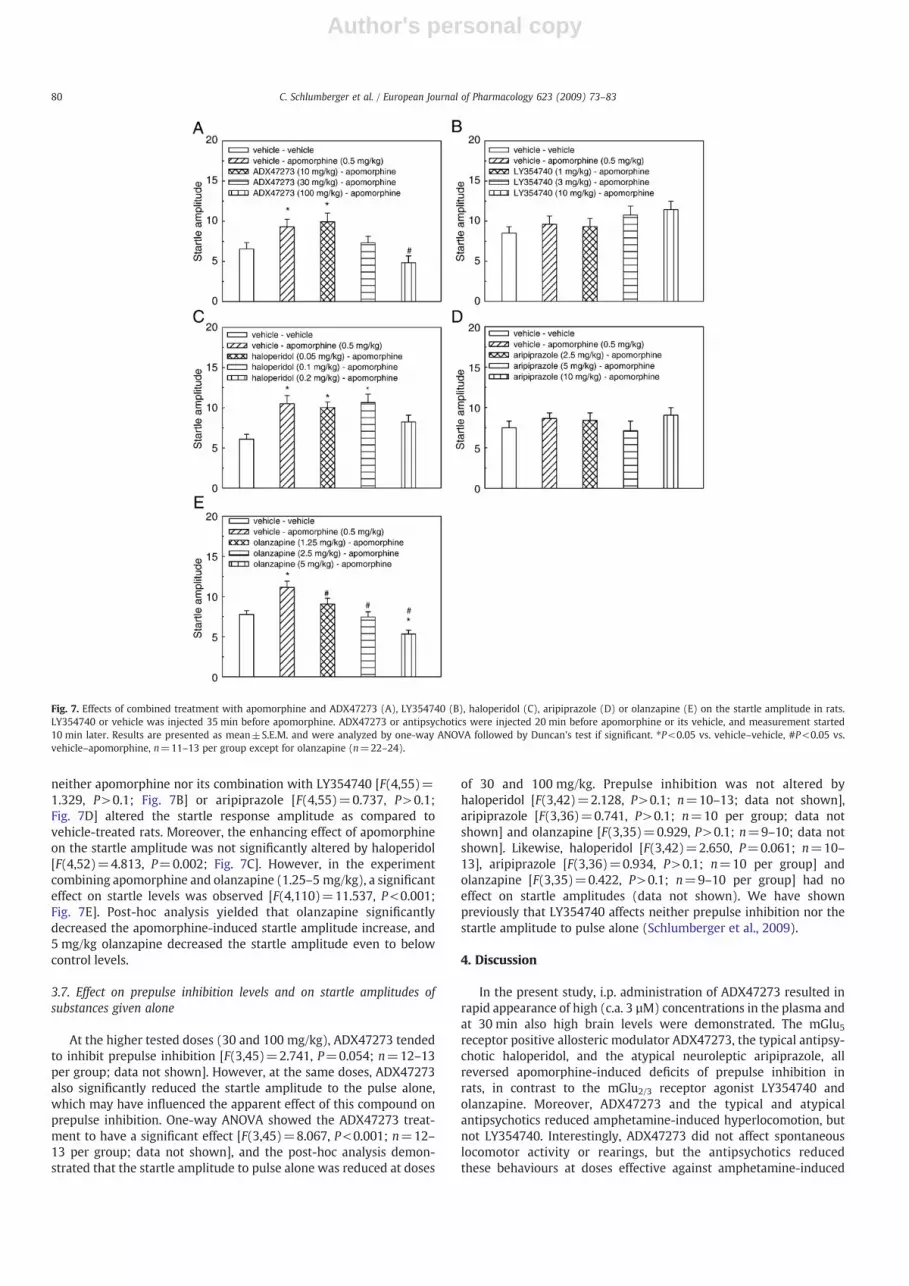
Administration of apomorphine alone increased the startle am-plitude to pulse alone. However, this effect only reached significancein some (Fig. 7A, C, E) but not all experiments (Fig. 7B, D). At thepresent time, we do not have convincing explanation for thisdiscrepancy. The only differences are the vehicles of the varioussubstances. Experiments using compounds requiring Tween80 in thesolvent failed to demonstrate the apomorphine-induced excessivestartle amplitude.
Enhancement of the startle amplitude produced by apomorphinewas attenuated by ADX47273. One-way ANOVA showed thistreatment to have a significant effect [F(4,54)=5.280, P=0.001;Fig. 7A]. Post-hoc comparison revealed a significant differencebetween animals treated with apomorphine and vehicle versustreatment with apomorphine and 100 mg/kg ADX47273. In contrast,
Fig. 6. Effects of ADX47273 (A), LY354740 (B), haloperidol (C), aripiprazole (D) and olanzapine (E) on apomorphine-induced PPI deficit in rats. LY354740 or vehicle was injected35 min before apomorphine. ADX47273 or antipsychotics were injected 20 min before apomorphine or its vehicle, and measurement started 10 min later. Results are presented asmean±S.E.M. and were analyzed by two-way ANOVA followed by Duncan's test. ⁎P<0.05 vs. vehicle–vehicle, #P<0.05 vs. vehicle–apomorphine, n=11–13 per group except forolanzapine (n=22–24).
79C. Schlumberger et al. / European Journal of Pharmacology 623 (2009) 73–83
Author's personal copy
neither apomorphine nor its combination with LY354740 [F(4,55)=1.329, P>0.1; Fig. 7B] or aripiprazole [F(4,55)=0.737, P>0.1;Fig. 7D] altered the startle response amplitude as compared tovehicle-treated rats. Moreover, the enhancing effect of apomorphineon the startle amplitude was not significantly altered by haloperidol[F(4,52)=4.813, P=0.002; Fig. 7C]. However, in the experimentcombining apomorphine and olanzapine (1.25–5 mg/kg), a significanteffect on startle levels was observed [F(4,110)=11.537, P<0.001;Fig. 7E]. Post-hoc analysis yielded that olanzapine significantlydecreased the apomorphine-induced startle amplitude increase, and5 mg/kg olanzapine decreased the startle amplitude even to belowcontrol levels.
3.7. Effect on prepulse inhibition levels and on startle amplitudes ofsubstances given alone
At the higher tested doses (30 and 100 mg/kg), ADX47273 tendedto inhibit prepulse inhibition [F(3,45)=2.741, P=0.054; n=12–13per group; data not shown]. However, at the same doses, ADX47273also significantly reduced the startle amplitude to the pulse alone,which may have influenced the apparent effect of this compound onprepulse inhibition. One-way ANOVA showed the ADX47273 treat-ment to have a significant effect [F(3,45)=8.067, P<0.001; n=12–13 per group; data not shown], and the post-hoc analysis demon-strated that the startle amplitude to pulse alone was reduced at doses
of 30 and 100 mg/kg. Prepulse inhibition was not altered byhaloperidol [F(3,42)=2.128, P>0.1; n=10–13; data not shown],aripiprazole [F(3,36)=0.741, P>0.1; n=10 per group; data notshown] and olanzapine [F(3,35)=0.929, P>0.1; n=9–10; data notshown]. Likewise, haloperidol [F(3,42)=2.650, P=0.061; n=10–13], aripiprazole [F(3,36)=0.934, P>0.1; n=10 per group] andolanzapine [F(3,35)=0.422, P>0.1; n=9–10 per group] had noeffect on startle amplitudes (data not shown). We have shownpreviously that LY354740 affects neither prepulse inhibition nor thestartle amplitude to pulse alone (Schlumberger et al., 2009).
4. Discussion
In the present study, i.p. administration of ADX47273 resulted inrapid appearance of high (c.a. 3 µM) concentrations in the plasma andat 30 min also high brain levels were demonstrated. The mGlu5receptor positive allosteric modulator ADX47273, the typical antipsy-chotic haloperidol, and the atypical neuroleptic aripiprazole, allreversed apomorphine-induced deficits of prepulse inhibition inrats, in contrast to the mGlu2/3 receptor agonist LY354740 andolanzapine. Moreover, ADX47273 and the typical and atypicalantipsychotics reduced amphetamine-induced hyperlocomotion, butnot LY354740. Interestingly, ADX47273 did not affect spontaneouslocomotor activity or rearings, but the antipsychotics reducedthese behaviours at doses effective against amphetamine-induced
Fig. 7. Effects of combined treatment with apomorphine and ADX47273 (A), LY354740 (B), haloperidol (C), aripiprazole (D) or olanzapine (E) on the startle amplitude in rats.LY354740 or vehicle was injected 35 min before apomorphine. ADX47273 or antipsychotics were injected 20 min before apomorphine or its vehicle, and measurement started10 min later. Results are presented as mean±S.E.M. and were analyzed by one-way ANOVA followed by Duncan's test if significant. ⁎P<0.05 vs. vehicle–vehicle, #P<0.05 vs.vehicle–apomorphine, n=11–13 per group except for olanzapine (n=22–24).
80 C. Schlumberger et al. / European Journal of Pharmacology 623 (2009) 73–83

Author's personal copy
hyperlocomotion. ADX47273 was not effective against amphetamine-induced increase in rearings.
Amphetamine administration dose-dependently induced hyperlo-comotion and increased rearings in rats, as reported in the literature(Arnt, 1995; Clark et al., 1991; Natesan et al., 2006). The typicalneuroleptic haloperidol, which blocks dopamine D2 receptors withhigh affinity, inhibited both amphetamine-induced hyperlocomotionand rearings at doses similar to previously reported (Cartmell et al.,1999; Maurel-Remy et al., 1995; Natesan et al., 2006). The present andprevious studies have also revealed an inhibitory effect of the atypicalneuroleptic olanzapine on hyperlocomotion elicited by amphetamine(Andersen and Pouzet 2001; Arnt, 1995). Apart from dopamine D2
receptors, olanzapine also blocks serotonin 5-HT2A receptors withhigh affinity (Arnt and Skarsfeldt, 1998; Horacek et al., 2006; Schotteet al., 1996). Although several studies have found that blockade ofdopamine D2 receptors is sufficient for inhibition of amphetamine-induced locomotion, antagonism of serotonin 5-HT2A receptor mayalso contribute to this effect (Arnt, 1995; Maurel-Remy et al., 1995).The recently registered atypical neuroleptic, aripiprazole, was shownto be a partial dopamine D2 receptor agonist, serotonin 5-HT2Areceptor antagonist and a serotonin 5-HT1A receptor partial agonist(Burris et al., 2002; McCreary et al., 2007; Newman-Tancredi et al.,2005). Inhibition of amphetamine-induced hyperlocomotion by thisdrug in our study is in agreement with previous reports (Natesanet al., 2006; Nordquist et al., 2008). In the present study, all of thetested neuroleptics inhibited hyperlocomotion evoked by amphet-amine at doses at which they also attenuated spontaneous locomotoractivity of rats, which is also in line with previous papers (Arnt, 1995;Nordquist et al., 2008).
In contrast to the neuroleptics, LY354740 did not decreaseamphetamine-induced locomotion and had no effect on rearings.The lack of effect of LY354740 on locomotion is in line with an earlierpublication (Cartmell et al., 1999). However, these authors observedinhibition of rearings evoked by amphetamine, but using s.c.application route as compared to i.p. administration used here andlower placement of photo beams (10 vs. 15 cm). It is noteworthy thatprevious studies (Cartmell et al., 1999; Moghaddam and Adams 1998;Schlumberger et al., 2009), have revealed that LY354740 attenuatedhyperlocomotion evoked by phencyclidine. The results of thosestudies indicate that LY354740 might be able to reverse the deficitproduced by dysfunction of glutamatergic but not dopaminergictransmission. Interestingly, other mGlu2/3 receptor agonists,LY379268 ((-)-2-oxa-4-aminobicyclo[3.1.0]hexane-4,6-dicarboxy-late) and LY404039, have been demonstrated to be effective againstboth phencyclidine and amphetamine-induced hyperlocomotion(Cartmell et al., 1999; Fell et al., 2008; Rorick-Kehn et al., 2007;Woolley et al., 2008). The reason for the different effects of LY354740and the other tested mGlu2/3 receptor agonists is not yet known.
Similarly to neuroleptics, the mGlu5 receptor positive allostericmodulator ADX47273 induced dose-dependent inhibition of amphet-amine-induced hyperlocomotion but not rearings, which indicatesthat mechanisms behind these behaviours might not be the same. Incontrast to neuroleptics, ADX47273 attenuated amphetamine-in-duced effect at doses at which it had no effect on spontaneouslocomotion. This suggests that mGlu5 receptor positive allostericmodulators may not induce sedation at therapeutic doses. The presentstudy is in agreement with a previous paper, which showedalleviation of amphetamine-induced locomotion in rats by anothermGlu5 receptor positive allosteric modulator CDPPB (Kinney et al.,2005). Moreover, ADX47273 has been demonstrated to attenuatehyperlocomotion evoked by direct (apomorphine) and indirect(amphetamine) dopamine receptor agonists as well as by theNMDA receptor antagonist phencyclidine in mice (Liu et al., 2008).However, we used a different species for amphetamine-inducedhyperlocomotion test, and thus these studies cannot be directlycompared due to the different effective doses. Previous studies have
found that stimulation of the mGlu5 receptor positively regulates thefunction of NMDA receptors in several brain regions (Attucci et al.,2001; Benquet et al., 2002; Doherty et al., 1997; Mannaioni et al.,2001; Pisani et al., 2001). This effect may contribute to the inhibitoryeffect of mGlu5 receptor positive allosteric modulators upon locomo-tor activity produced by NMDA receptor antagonists. For example, ithas been demonstrated that the group I mGlu receptor agonist (R,S)-3,5-dihydroxyphenylglycine (DHPG) and the mGlu5 receptor agonist(RS)-2-Chloro-5-hydroxyphenylglycine (CHPG) potentiate NMDA-evoked currents in the hippocampus, cerebral cortex and striatum(Attucci et al., 2001; Benquet et al., 2002; Doherty et al., 1997;Mannaioni et al., 2001; Pisani et al., 2001). The mGlu5 receptorpositive allosteric modulator CPPHA (N-{4-Chloro-2-[(1,3-dioxo-1,3-dihydro-2H-isoindol-2-yl)methyl]phenyl}-2-hy droxybenzamide),showed no effect when applied alone, but potentiated both DHPG-induced phosphorylation of the NR1 subunit of the NMDA receptor(serine at position 897) and the effect of DHPG on NMDA-evokedcurrents (Liu et al., 2006). ADX47273 has also been demonstrated toinhibit dopamine release in the nucleus accumbens, which can at leastpartially explain its inhibitory effect on amphetamine-inducedlocomotor activity (Liu et al., 2008). Moreover, in the striatum andnucleus accumbens, bothmGlu5 receptors and dopamine D2 receptorsare present on striatopallidal GABA neurons (Fuxe et al., 2008; Kerneret al., 1997; Tallaksen-Greene et al., 1998). In binding experiments, ithas been found that the mGlu5 receptor agonist, CHPG, produced asignificant decrease in the affinity of the high-affinity state ofdopamine D2 receptors for dopamine in membrane preparationsfrom rat striatum (Popoli et al., 2001). Antagonistic interactionsbetween dopamine D2 and mGlu5 receptors have also been proposedin nucleus accumbens (Diaz-Cabiale et al., 2002; Fuxe et al., 2008).Interestingly, it has been recently demonstrated that the mGlu5receptor positive allosteric modulator CDPPB attenuated amphet-amine-induced overactivation of pyramidal neurones in the orbito-frontal cortex, similarly to haloperidol (Homayoun and Moghaddam,2008). Furthermore, reversal of neuronal activity by both compoundswas associated with inhibition of amphetamine-induced stereotypy(Homayoun and Moghaddam, 2008).
In the present study, ADX47273 attenuated deficits of prepulseinhibition evoked by the dopamine D2/D1 receptor agonist apomor-phine. Similar effect was also produced by haloperidol, which is in linewith previous reports (Auclair et al., 2006; Mansbach et al., 1988).Administration of haloperidol alone neither affected prepulse inhibi-tion levels nor the startle amplitude to pulse alone. Aripiprazolerestored apomorphine-induced disruption of prepulse inhibition, ashas been described in literature, while the startle amplitude wasunchanged by any of the treatments (Auclair et al., 2006; Kohnomiet al., 2008; Nakai et al., 2008).
In the present study, olanzapine failed to reverse the deficits ofprepulse inhibition at doses at which it attenuated amphetamine-induced hyperlocomotion. Administration of apomorphine increasedthe startle amplitude, which was significantly reversed by all doses ofolanzapine and by ADX47273. However, at 30 and 100 mg/kg,ADX47273 also reduced the startle amplitude when administeredalone, which might conceal the effect of ADX47273 on apomorphine-induced enhancement of startle amplitude.
The nucleus accumbens has been proposed to be an important siteregarding the disruptive effects of dopaminominetics upon prepulseinhibition. Infusion of amphetamine or the dopamine D2 receptoragonist quinpirole into the nucleus accumbens disrupts prepulseinhibition in a dopamine D2 receptor-dependent manner (Swerdlowet al., 2007; Wan and Swerdlow, 1993). mGlu5 receptors, as well asmGlu2 and mGlu3 receptors, are localized in this structure (Fuxe et al.,2008; Kerner et al., 1997; Shigemoto et al., 1993; Spooren et al., 2003;Tallaksen-Greene et al., 1998). However, in the present study, themGlu2/3 receptor agonist LY354740 did not reverse deficits of prepulseinhibition evoked by apomorphine, in contrast to the mGlu5 receptor
81C. Schlumberger et al. / European Journal of Pharmacology 623 (2009) 73–83

Author's personal copy
positive allosteric modulator ADX47273. This is in line with previousstudies which showed that neither LY354740 nor other the mGlu2/3receptor agonist LY379268 were able to reverse deficits of prepulseinhibition evoked by apomorphine or amphetamine in rats (Galiciet al., 2005; Ossowska et al., 2000).
5. Conclusion
The present study shows that the used neuroleptics or potentialantipsychotics modulating glutamate receptors (LY354740,ADX47273) acting through diverse mechanisms have differentprofiles in animal models. The mGlu5 receptor positive allostericmodulator ADX47273 showed better efficacy than the mGlu2/3
receptor agonist LY354740 in rodent models for the positivesymptoms of schizophrenia. This is not unexpected, if we considerdifferent mechanisms of action i.e. the former probably produces itsantipsychiotic-like effects through indirect enhancement of NMDAreceptor function while the later likely acts through modulation ofglutamate release. In turn, therapeutic profile may differ as well,possibly providing added benefits to current therapies.
Moreover, since activation of the mGlu2/3 receptor has beenalready shown to be effective against the positive and negativesymptoms of schizophrenic patients (Patil et al., 2007), one could alsoexpect similar activity from mGlu5 receptor positive allostericmodulators. Therefore, mGlu5 receptor positive allosteric modulatorsmay also represent a promising treatment for schizophrenia.
References
Adler, C.M., Goldberg, T.E., Malhotra, A.K., Pickar, D., Breier, A., 1998. Effects of ketamineon thought disorder, working memory, and semantic memory in healthyvolunteers. Biol. Psychiatry 43, 811–816.
Andersen, M.P., Pouzet, B., 2001. Effects of acute versus chronic treatment with typicalor atypical antipsychotics on D-amphetamine-induced sensorimotor gating deficitsin rats. Psychopharmacology (Berl.) 156, 291–304.
Arnt, J., 1995. Differential effects of classical and newer antipsychotics on thehypermotility induced by two dose levels of D-amphetamine. Eur. J. Pharmacol.283, 55–62.
Arnt, J., Skarsfeldt, T., 1998. Do novel antipsychotics have similar pharmacologicalcharacteristics? A review of the evidence. Neuropsychopharmacology 18, 63–101.
Attucci, S., Carla, V., Mannaioni, G., Moroni, F., 2001. Activation of type 5 metabotropicglutamate receptors enhances NMDA responses in mice cortical wedges. Br. J.Pharmacol. 132, 799–806.
Auclair, A.L., Kleven, M.S., Besnard, J., Depoortere, R., Newman-Tancredi, A., 2006.Actions of novel antipsychotic agents on apomorphine-induced PPI disruption:influence of combined serotonin 5-HT1A receptor activation and dopamine D2receptor blockade. Neuropsychopharmacology 31, 1900–1909.
Benquet, P., Gee, C.E., Gerber, U., 2002. Two distinct signaling pathways upregulateNMDA receptor responses via two distinct metabotropic glutamate receptorsubtypes. J. Neurosci. 22, 9679–9686.
Braff, D., Stone, C., Callaway, E., Geyer, M., Glick, I., Bali, L., 1978. Prestimulus effects onhuman startle reflex in normals and schizophrenics. Psychophysiology 15,339–343.
Brody, S.A., Dulawa, S.C., Conquet, F., Geyer, M.A., 2004. Assessment of a prepulseinhibition deficit in a mutant mouse lacking mGlu5 receptors. Mol. Psychiatry 9,35–41.
Burris, K.D., Molski, T.F., Xu, C., Ryan, E., Tottori, K., Kikuchi, T., Yocca, F.D., Molinoff, P.B.,2002. Aripiprazole, a novel antipsychotic, is a high-affinity partial agonist at humandopamine D2 receptors. J. Pharmacol. Exp. Ther. 302, 381–389.
Cartmell, J., Monn, J.A., Schoepp, D.D., 1999. The metabotropic glutamate 2/3 receptoragonists LY354740 and LY379268 selectively attenuate phencyclidine versus D-amphetamine motor behaviors in rats. J. Pharmacol. Exp. Ther. 291, 161–170.
Clark, D., Furmidge, L.J., Petry, N., Tong, Z.Y., Ericsson, M., Johnson, D., 1991. Behaviouralprofile of partial D2 dopamine receptor agonists. 1. Atypical inhibition of D-amphetamine-induced locomotor hyperactivity and stereotypy. Psychopharma-cology (Berl.) 105, 381–392.
Conn, P.J., Lindsley, C.W., Jones, C.K., 2009. Activation of metabotropic glutamatereceptors as a novel approach for the treatment of schizophrenia. Trends Pharmacol.Sci. 30, 25–31.
Darrah, J.M., Stefani, M.R., Moghaddam, B., 2008. Interaction of N-methyl-D-aspartateand group 5 metabotropic glutamate receptors on behavioral flexibility using anovel operant set-shift paradigm. Behav. Pharmacol. 19, 225–234.
Diaz-Cabiale, Z., Vivo, M., Del Arco, A., O'Connor, W.T., Harte, M.K., Muller, C.E.,Martinez, E., Popoli, P., Fuxe, K., Ferre, S., 2002. Metabotropic glutamate mGlu5receptor-mediated modulation of the ventral striopallidal GABA pathway in rats.Interactions with adenosine A(2A) and dopamine D(2) receptors. Neurosci. Lett.324, 154–158.
Doherty, A.J., Palmer, M.J., Henley, J.M., Collingridge, G.L., Jane, D.E., 1997. (RS)-2-chloro-5-hydroxyphenylglycine (CHPG) activates mGlu5, but no mGlu1, receptorsexpressed in CHO cells and potentiates NMDA responses in the hippocampus.Neuropharmacology 36, 265–267.
Fell, M.J., Svensson, K.A., Johnson, B.G., Schoepp, D.D., 2008. Evidence for the role ofmetabotropic glutamate (mGlu)2 not mGlu3 receptors in the preclinical antipsy-chotic pharmacology of the mGlu2/3 receptor agonist (-)-(1R, 4S, 5S, 6S)-4-amino-2-sulfonylbicyclo[3.1.0]hexane-4, 6-dicarboxylic acid (LY404039). J. Pharmacol.Exp. Ther. 326, 209–217.
Fuxe, K., Marcellino, D., Rivera, A., Diaz-Cabiale, Z., Filip, M., Gago, B., Roberts, D.C.,Langel, U., Genedani, S., Ferraro, L., de la Calle, A., Narvaez, J., Tanganelli, S., Woods,A., Agnati, L.F., 2008. Receptor-receptor interactions within receptor mosaics.Impact on neuropsychopharmacology. Brain Res. Rev. 58, 415–452.
Galici, R., Echemendia, N.G., Rodriguez, A.L., Conn, P.J., 2005. A selective allostericpotentiator of metabotropic glutamate (mGlu) 2 receptors has effects similar to anorthosteric mGlu2/3 receptor agonist in mouse models predictive of antipsychoticactivity. J. Pharmacol. Exp. Ther. 315, 1181–1187.
Geyer, M.A., Swerdlow, N.R., Mansbach, R.S., Braff, D.L., 1990. Startle response models ofsensorimotor gating and habituation deficits in schizophrenia. Brain Res. Bull. 25,485–498.
Geyer, M.A., Krebs-Thomson, K., Braff, D.L., Swerdlow, N.R., 2001. Pharmacologicalstudies of prepulse inhibition models of sensorimotor gating deficits in schizo-phrenia: a decade in review. Psychopharmacology (Berl.) 156, 117–154.
Gray, L., van den Buuse, M., Scarr, E., Dean, B., Hannan, A.J., 2009. Clozapine reversesschizophrenia-related behaviours in the metabotropic glutamate receptor 5knockout mouse: association with N-methyl-D-aspartic acid receptor up-regula-tion. Int. J. Neuropsychopharmacol 12, 45–60.
Henry, S.A., Lehmann-Masten, V., Gasparini, F., Geyer, M.A., Markou, A., 2002. ThemGluR5 antagonist MPEP, but not the mGluR2/3 agonist LY314582, augments PCPeffects on prepulse inhibition and locomotor activity. Neuropharmacology 43,1199–1209.
Homayoun, H., Moghaddam, B., 2008. Orbitofrontal cortex neurons as a common targetfor classic and glutamatergic antipsychotic drugs. Proc. Natl. Acad. Sci. U. S. A. 105,18,041–18,046.
Horacek, J., Bubenikova-Valesova, V., Kopecek, M., Palenicek, T., Dockery, C., Mohr, P.,Hoschl, C., 2006. Mechanism of action of atypical antipsychotic drugs and theneurobiology of schizophrenia. CNS Drugs 20, 389–409.
Johnson, J.T., Mattiuz, E.L., Chay, S.H., Herman, J.L., Wheeler, W.J., Kassahun, K.,Swanson, S.P., Phillips, D.L., 2002. The disposition, metabolism, and pharmacoki-netics of a selective metabotropic glutamate receptor agonist in rats and dogs. DrugMetab. Dispos. 30, 27–33.
Kerner, J.A., Standaert, D.G., Penney Jr, J.B., Young, A.B., Landwehrmeyer, G.B., 1997.Expression of group one metabotropic glutamate receptor subunit mRNAs inneurochemically identified neurons in the rat neostriatum, neocortex, andhippocampus. Brain Res. Mol. Brain Res. 48, 259–269.
Kinney, G.G., Burno, M., Campbell, U.C., Hernandez, L.M., Rodriguez, D., Bristow, L.J.,Conn, P.J., 2003. Metabotropic glutamate subtype 5 receptors modulate locomotoractivity and sensorimotor gating in rodents. J. Pharmacol. Exp. Ther. 306, 116–123.
Kinney, G.G., O'Brien, J.A., Lemaire, W., Burno, M., Bickel, D.J., Clements, M.K., Chen, T.B.,Wisnoski, D.D., Lindsley, C.W., Tiller, P.R., Smith, S., Jacobson, M.A., Sur, C., Duggan,M.E., Pettibone, D.J., Conn, P.J., Williams Jr, D.L., 2005. A novel selective positiveallosteric modulator of metabotropic glutamate receptor subtype 5 has in vivoactivity and antipsychotic-like effects in rat behavioral models. J. Pharmacol. Exp.Ther. 313, 199–206.
Kohnomi, S., Suemaru, K., Kawasaki, H., Araki, H., 2008. Effect of aripiprazole on 5-HT2receptor-mediated wet-dog shake responses and disruption of prepulse inhibitionin rats. J. Pharmacol. Sci. 106, 645–650.
Krystal, J.H., Karper, L.P., Seibyl, J.P., Freeman, G.K., Delaney, R., Bremner, J.D., Heninger,G.R., Bowers, M.B., Charney, D.S., 1994. Subanesthetic effects of the noncompetitiveNMDA antagonist, ketamine, in humans — psychotomimetic, perceptual, cognitive,and neuroendocrine responses. Arch. Gen. Psychiatry 51, 199–214.
Krystal, J.H., D'Souza, D.C., Mathalon, D., Perry, E., Belger, A., Hoffman, R., 2003. NMDAreceptor antagonist effects, cortical glutamatergic function, and schizophrenia:toward a paradigm shift in medication development. Psychopharmacology (Berl.)169, 215–233.
Krystal, J.H., Abi-Saab, W., Perry, E., D'Souza, D.C., Liu, N., Gueorguieva, R., McDougall, L.,Hunsberger, T., Belger, A., Levine, L., Breier, A., 2005. Preliminary evidence ofattenuation of the disruptive effects of the NMDA glutamate receptor antagonist,ketamine, on working memory by pretreatment with the group II metabotropicglutamate receptor agonist, LY354740, in healthy human subjects. Psychopharma-cology (Berl.) 179 (1), 303–309.
Lindsley, C.W., Wisnoski, D.D., Leister, W.H., O'Brien, J.A., Lemaire, W., Williams Jr, D.L.,Burno, M., Sur, C., Kinney, G.G., Pettibone, D.J., Tiller, P.R., Smith, S., Duggan, M.E.,Hartman, G.D., Conn, P.J., Huff, J.R., 2004. Discovery of positive allosteric modulatorsfor the metabotropic glutamate receptor subtype 5 from a series of N-(1, 3-diphenyl-1H-pyrazol-5-yl)benzamides that potentiate receptor function in vivo.J. Med. Chem. 47, 5825–5828.
Liu, F., Zhang, G., Hornby, G., Vasylyev, D., Bowlby, M., Park, K., Gilbert, A., Marquis, K.,Andree, T.H., 2006. The effect of mGlu5 receptor positive allosteric modulators onsignaling molecules in brain slices. Eur. J. Pharmacol. 536, 262–268.
Liu, F., Grauer, S., Kelley, C., Navarra, R., Graf, R., Zhang, G., Atkinson, P.J., Popiolek, M.,Wantuch, C., Khawaja, X., Smith, D., Olsen, M., Kouranova, E., Lai, M., Pruthi, F.,Pulicicchio, C., Day, M., Gilbert, A., Pausch, M.H., Brandon, N.J., Beyer, C.E., Comery,T.A., Logue, S., Rosenzweig-Lipson, S., Marquis, K.L., 2008. ADX47273 [S-(4-fluoro-phenyl)-{3-[3-(4-fluoro-phenyl)-[1, 2, 4]-oxadiazol-5-yl]-piperidin-1-yl}-metha-none]: a novel metabotropic glutamate receptor 5-selective positive allosteric
82 C. Schlumberger et al. / European Journal of Pharmacology 623 (2009) 73–83

Author's personal copy
modulatorwithpreclinical antipsychotic-like andprocognitive activities. J. Pharmacol.Exp. Ther. 327, 827–839.
Luby, E.D., Cohen, R.C., Rosenbaum, B., Gottlieb, J.S., Kelly, R., 1959. Study of a newschizophrenomimetic drug: sernyl. Arch. Neurol. Psychiatr. 81, 363–369.
Mannaioni, G., Marino, M.J., Valenti, O., Traynelis, S.F., Conn, P.J., 2001. Metabotropicglutamate receptors 1 and 5 differentially regulate CA1 pyramidal cell function.J. Neurosci. 21, 5925–5934.
Mansbach, R.S., Geyer, M.A., Braff, D.L., 1988. Dopaminergic stimulation disruptssensorimotor gating in the rat. Psychopharmacology (Berl.) 94, 507–514.
Maurel-Remy, S., Bervoets, K., Millan, M.J., 1995. Blockade of phencyclidine-inducedhyperlocomotion by clozapine and MDL 100, 907 in rats reflects antagonism of 5-HT2A receptors. Eur. J. Pharmacol. 280, R9–R11.
McCreary, A.C., Glennon, J.C., Ashby Jr, C.R., Meltzer, H.Y., Li, Z., Reinders, J.H., Hesselink,M.B., Long, S.K., Herremans, A.H., van Stuivenberg, H., Feenstra, R.W., Kruse, C.G.,2007. SLV313 (1-(2, 3-dihydro-benzo[1, 4]dioxin-5-yl)-4-). Neuropsychopharma-cology 32, 78–94.
Meltzer, H.Y., Stahl, S.M., 1976. The dopamine hypothesis of schizophrenia: a review.Schizophr. Bull. 2, 19–76.
Moghaddam, B., Adams, B.W., 1998. Reversal of phencyclidine effects by a group IImetabotropic glutamate receptor agonist in rats. Science 281, 1349–1352.
Nakai, S., Hirose, T., Mori, T., Stark, A., Araki, H., Kikuchi, T., 2008. The effect of aripiprazoleonprepulse inhibition of the startle response innormal andhyperdopaminergic statesin rats. Int. J. Neurosci. 118, 39–57.
Natesan, S., Reckless, G.E., Nobrega, J.N., Fletcher, P.J., Kapur, S., 2006. Dissociationbetween in vivo occupancy and functional antagonism of dopamine D2 receptors:comparing aripiprazole to other antipsychotics in animal models. Neuropsycho-pharmacology 31, 1854–1863.
Newman-Tancredi, A., Assie, M.B., Leduc, N., Ormiere, A.M., Danty, N., Cosi, C., 2005.Novel antipsychotics activate recombinant human and native rat serotonin 5-HT1Areceptors: affinity, efficacy and potential implications for treatment of schizophre-nia. Int. J. Neuropsychopharmacol 8, 341–356.
Nordquist, R.E., Risterucci, C., Moreau, J.L., von Kienlin, M., Kunnecke, B., Maco, M.,Freichel, C., Riemer, C., Spooren, W., 2008. Effects of aripiprazole/OPC-14597 onmotor activity, pharmacological models of psychosis, and brain activity in rats.Neuropharmacology 54, 405–416.
Ossowska, K., Pietraszek, M., Wardas, J., Nowak, G., Zajaczkowski, W., Wolfarth, S., Pilc,A., 2000. The role of glutamate receptors in antipsychotic drug action. Amino Acids19, 87–94.
Patil, S.T., Zhang, L., Martenyi, F., Lowe, S.L., Jackson, K.A., Andreev, B.V., Avedisova, A.S.,Bardenstein, L.M., Gurovich, I.Y., Morozova, M.A., Mosolov, S.N., Neznanov, N.G.,Reznik, A.M., Smulevich, A.B., Tochilov, V.A., Johnson, B.G., Monn, J.A., Schoepp, D.D.,2007. Activation of mGlu2/3 receptors as a new approach to treat schizophrenia: arandomized phase 2 clinical trial. Nat. Med. 13, 1102–1107.
Pietraszek, M., Gravius, A., Schafer, D., Weil, T., Trifanova, D., Danysz, W., 2005. mGluR5,but not mGluR1, antagonist modifies MK-801-induced locomotor activity anddeficit of prepulse inhibition. Neuropharmacology 49, 73–85.
Pisani, A., Gubellini, P., Bonsi, P., Conquet, F., Picconi, B., Centonze, D., Bernardi, G.,Calabresi, P., 2001. Metabotropic glutamate receptor 5 mediates the potentiation ofN-methyl-D-aspartate responses in medium spiny striatal neurons. Neuroscience106, 579–587.
Popoli, P., Pezzola, A., Torvinen, M., Reggio, R., Pintor, A., Scarchilli, L., Fuxe, K., Ferre, S.,2001. The selective mGlu(5) receptor agonist CHPG inhibits quinpirole-inducedturning in 6-hydroxydopamine-lesioned rats and modulates the binding char-acteristics of dopamine D(2) receptors in the rat striatum: interactions withadenosine A(2a) receptors. Neuropsychopharmacology 25, 505–513.
Rorick-Kehn, L.M., Johnson, B.G., Knitowski, K.M., Salhoff, C.R., Witkin, J.M., Perry, K.W.,Griffey, K.I., Tizzano, J.P., Monn, J.A., McKinzie, D.L., Schoepp, D.D., 2007. In vivopharmacological characterization of the structurally novel, potent, selectivemGlu2/3 receptor agonist LY404039 in animal models of psychiatric disorders.Psychopharmacology (Berl.) 193, 121–136.
Schlumberger, C., Schafer, D., Barberi, C., More, L., Nagel, J., Pietraszek, M., Schmidt, W.J.,Danysz, W., 2009. Effects of a metabotropic glutamate receptor group II agonistLY354740 in animal models of positive schizophrenia symptoms and cognition.Behav. Pharmacol. 20, 56–66.
Schotte, A., Janssen, P.F., Gommeren, W., Luyten, W.H., Van Gompel, P., Lesage, A.S., DeLoore, K., Leysen, J.E., 1996. Risperidone compared with new and referenceantipsychotic drugs: in vitro and in vivo receptor binding. Psychopharmacology(Berl.) 124, 57–73.
Seeman, P., 1987. Dopamine receptors and the dopamine hypothesis of schizophrenia.Synapse 1, 133–152.
Seeman, P., 1992.Dopamine receptor sequences. Therapeutic levels of neuroleptics occupyD2 receptors, clozapine occupies D4. Neuropsychopharmacology 7, 261–284.
Shigemoto, R., Nomura, S., Ohishi, H., Sugihara, H., Nakanishi, S., Mizuno, N., 1993.Immunohistochemical localization of a metabotropic glutamate receptor, mGluR5,in the rat brain. Neurosci. Lett. 163, 53–57.
Snyder, S.H., Banerjee, S.P., Yamamura, H.I., Greenberg, D., 1974. Drugs, neurotrans-mitters, and schizophrenia. Science 184, 1243–1253.
Spooren, W., Ballard, T., Gasparini, F., Amalric, M., Mutel, V., Schreiber, R., 2003. Insightinto the function of group I and group II metabotropic glutamate (mGlu) receptors:behavioural characterization and implications for the treatment of CNS disorders.Behav. Pharmacol. 14, 257–277.
Swerdlow, N.R., Shoemaker, J.M., Bongiovanni, M.J., Neary, A.C., Tochen, L.S., SaintMarie, R.L., 2007. Strain differences in the disruption of prepulse inhibition ofstartle after systemic and intra-accumbens amphetamine administration. Pharma-col. Biochem. Behav. 87, 1–10.
Swerdlow, N.R., Weber, M., Qu, Y., Light, G.A., Braff, D.L., 2008. Realistic expectations ofprepulse inhibition in translational models for schizophrenia research. Psycho-pharmacology (Berl.) 199, 331–388.
Tallaksen-Greene, S.J., Kaatz, K.W., Romano, C., Albin, R.L., 1998. Localization ofmGluR1a-like immunoreactivity and mGluR5-like immunoreactivity in identifiedpopulations of striatal neurons. Brain Res. 780, 210–217.
Wan, F.J., Swerdlow, N.R., 1993. Intra-accumbens infusion of quinpirole impairssensorimotor gating of acoustic startle in rats. Psychopharmacology (Berl.) 113,103–109.
Woolley, M.L., Pemberton, D.J., Bate, S., Corti, C., Jones, D.N., 2008. The mGlu2 but notthe mGlu3 receptor mediates the actions of the mGluR2/3 agonist, LY379268, inmouse models predictive of antipsychotic activity. Psychopharmacology (Berl.)196, 431–440.
83C. Schlumberger et al. / European Journal of Pharmacology 623 (2009) 73–83