comparison of salmonella enterica serovar bovismorbificans 2011 hummus outbreak strains with...
TRANSCRIPT

Accepted Manuscript
Comparison of Salmonella enterica serovar Bovismorbificans 2011 Hummus outbreakstrains with non-outbreak strains
Morris Blaylock, Reginald Blackwell, Sosina Merid, Scott Jackson, Michael Kotewicz,Gopal Gopinath, Sherry L. Ayers, Jason Abbott, Jonathan Sabo, Laura Ewing,Jayanthi Gangiredla, Solomon Gebru, Isha Patel, Brooke Jones, Kim Dudley, KarenJarvis, Darcy E. Hanes, Alpha A. Diallo, Junia Jean-Gilles Beaubrun, PhD, FDA/CFSAN/OARSA/DVA
PII: S0740-0020(14)00041-0
DOI: 10.1016/j.fm.2014.02.016
Reference: YFMIC 2114
To appear in: Food Microbiology
Received Date: 19 June 2013
Revised Date: 4 February 2014
Accepted Date: 18 February 2014
Please cite this article as: Blaylock, M., Blackwell, R., Merid, S., Jackson, S., Kotewicz, M., Gopinath,G., Ayers, S.L., Abbott, J., Sabo, J., Ewing, L., Gangiredla, J., Gebru, S., Patel, I., Jones, B., Dudley,K., Jarvis, K., Hanes, D.E., Diallo, A.A., Jean-Gilles Beaubrun, J., Comparison of Salmonella entericaserovar Bovismorbificans 2011 Hummus outbreak strains with non-outbreak strains, Food Microbiology(2014), doi: 10.1016/j.fm.2014.02.016.
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service toour customers we are providing this early version of the manuscript. The manuscript will undergocopyediting, typesetting, and review of the resulting proof before it is published in its final form. Pleasenote that during the production process errors may be discovered which could affect the content, and alllegal disclaimers that apply to the journal pertain.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
1
Title: Comparison of Salmonella enterica serovar Bovismorbificans 2011 Hummus 1
outbreak strains with non-outbreak strains. 2
3
Keywords: Salmonella, Bovismorbificans, hummus 4
5
Running Title: Molecular characterization of Salmonella enterica serovar Bovismorbificans 6
isolates. 7
8
Authors: Morris Blaylock1, Reginald Blackwell1, Sosina Merid1, Scott Jackson2, Michael 9
Kotewicz2, Gopal Gopinath2, Sherry L. Ayers3, Jason Abbott3, Jonathan Sabo3, Laura Ewing2, 10
Jayanthi Gangiredla2, Solomon Gebru2, Isha Patel2, Brooke Jones2, Kim Dudley2, Karen Jarvis2, 11
Darcy E. Hanes2, Alpha A. Diallo1, *Junia Jean-Gilles Beaubrun2. 12
13
*Corresponding author: Junia Jean-Gilles Beaubrun, PhD, FDA/CFSAN/OARSA/DVA, 8301 14
Muirkirk Rd, Rm 3404, Laurel MD 20708, 240-402-3578 (Lab), 301-785-4698 (cell), 301-210-15
5370 (Fax), [email protected]. 16
17
District of Columbia Public Health Laboratory, Department of Forensic Sciences, 401 E. Street 18
SW Washington, DC 200241, U.S. Food and Drug Administration, Laurel, MD 207082, U.S. 19
Food and Drug Administration, Center for Veterinary Medicine, Laurel MD 207083. 20

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
2
ABSTRACT 21
Eleven Salmonella enterica serovar Bovismorbificans isolates obtained from the U.S. District of 22
Columbia during a 2011 hummus-associated foodborne outbreak were compared to 12 non-23
outbreak isolates. All isolates from the outbreak demonstrated a single PFGE pattern that was 24
distinctly different from other isolates of S. Bovismorbificans as recorded in the PulseNet 25
Database. Results from molecular analyses of the hummus-associated S. Bovismorbificans 26
isolates indicate that the isolates from the outbreak were unique and have acquired an 80-90 kb 27
plasmid. The impact of this study is that the information gained will add and expand our 28
knowledge of diversity of the S. Bovismorbificans serovar. 29
INTRODUCTION 30
Outbreaks associated with a variety of Salmonella enterica serovars and food matrices 31
continue to increase each year, making identification, differentiation and characterization of the 32
various serotypes in foodborne outbreak a high priority for the field of food safety. Every year 33
approximately 40,000 cases of salmonellosis are reported in the United States. There are over 34
2500 different serovars of Salmonella enterica, and some of the most recent reported cases of 35
Salmonella enterica associated with foodborne outbreaks include: S. Typhimurium in ground 36
beef, S. Heidelberg in Kosher broiled chicken livers and ground turkey, S. Enteritidis in Turkish 37
pine nuts, eggs, alfalfa sprouts and spicy sprouts, S. Agona in fresh imported papayas, S. Hadar 38
in turkey burgers, S. Panama in Cantaloupe, S. Bareilly in spicy tuna, S. Braenderup in mangoes, 39
and S. Bredeney in Peanut Butter (20). 40
Human infections with S. enterica serovar Bovismorbificans are relatively infrequent in 41
the United States. Prior to 2011, the last major outbreak associated with S. Bovismorbificans 42

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
3
occurred in 2004 with 35 confirmed cases associated with the consumption of alfalfa sprouts 43
(20). From 1999 to 2009, only 758 illnesses associated with S. Bovismorbificans were reported, 44
compared to 1000 confirmed cases associated with S. Enteritidis in 1999 alone (Foodborne 45
Outbreak Online Database, CDC, www.cdc.gov/salmonella/outbreaks.html). The majority of 46
foodborne outbreaks associated with S. Bovismorbificans has occurred in Europe and have been 47
traced to pork products, lettuce and sprouts (18). In Finland in 1994, 201 cases were reported 48
from a large sprout-associated S. Bovismorbificans outbreak (16). In Sweden in 1994, there was 49
a large sprout-associated, S. Bovismorbificans outbreak (15) and a second nationwide outbreak 50
occurred in 2009 with 42 clinical isolates identified and was also associated with ready-to-eat 51
alfalfa sprouts. The sprouts samples were traced back to a domestic producer, but the seeds 52
originated in Italy (18). Over a 13 week period between November 2004 and March 2005, 525 53
cases of laboratory confirmed S. Bovismorbificans associated with raw pork were reported to 54
The Robert Koch Institute (18). 55
From August through November of 2011, sesame seed paste (tahini) and humus 56
containing a rare serotype of Salmonella caused illness in 23 people in 7 states, including the 57
District of Columbia (23). Reported cases were largely concentrated in the Mid-Atlantic region, 58
with eight in Washington, D.C., seven in Maryland, three in Virginia and one each in Delaware 59
and New Jersey. Three cases were also reported outside this region – one in California, one in 60
Michigan and one in New Hampshire (23). In this study, eleven S. Bovismorbificans isolates 61
from clinical and hummus food samples were obtained from the DC Public Health Laboratory 62
(DC_PHL) (Table 1). PFGE and conventional serotyping were used to identify the causal agent 63
as a unique strain of S. Bovismorbificans. Hummus has not previously been associated as a 64
source of Salmonella contamination. The DC_PHL and the FDA Office of Applied Research and 65

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
4
Safety Assessment (OARSA) collaborated to characterize the outbreak strains. In addition to the 66
original PFGE and the Kauffman White serotyping analyses, molecular serotyping using PCR, 67
Optical Mapping, Multiple-locus variable-number tandem-repeats analysis (MLVA), antibiotic 68
susceptibility testing, molecular fingerprinting using the Diversilab system, and plasmid analyses 69
were used to characterize these outbreak strains. 70
71
MATERIALS AND METHODS 72
Strains. A total of 23 isolates were examined in this study (Table 1). DC_PHL identified 11 S. 73
Bovismorbificans isolates from clinical and food samples (hummus) associated with the 2011 74
Washington DC outbreak. Two additional isolates associated with the outbreak were obtained 75
from the Michigan and Delaware Departments of Health, Division of Public Health laboratories. 76
Ten additional S. Bovismorbificans isolates, not associated with the outbreak, were also 77
examined; and these isolates were obtained from the FDA Center for Food Safety and Applied 78
Nutrition (CFSAN) and the Center for Veterinary Medicine (CVM) Salmonella culture 79
collections. Two outlier strains of Salmonella: S. Typhimurium and S. Newport were also 80
included in the studies (Table 1). 81
Molecular Serotyping (PCR Analysis). To evaluate the molecular serotyping PCR method’s 82
usefulness during an outbreak, all isolates used in this study were serotyped using the 83
conventional serotyping method and a PCR serotyping method described by Jean-Gilles 84
Beaubrun et al., (12,14). Genomic DNA was isolated using the NucliSENS EasyMag instrument 85
(BioMerieux, Inc. Hazelwood, MO) according to the manufacturer’s instructions. PCR products 86
were visualized using the Agilent 2100 Bio-analyzer (Agilent Technologies, Waldbronn, 87

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
5
Germany), and the DNA 1000 Reagents kit (Agilent Technologies) following the manufacturer’s 88
protocol. 89
Pulsed Field Gel Electrophoresis. All isolates of S. Bovismorbificans were analyzed for genetic 90
relatedness using pulsed-field gel electrophoresis (PFGE) with XbaI and BlnI according to the 91
U.S. CDC PulseNet protocol (21). Electrophoresis was performed with a CHEF-DR system 92
(Bio-Rad Laboratories, Hercules, California) using 1% SeaKem agarose in 0.5× Tris-borate-93
EDTA at 180 V. Running conditions consisted of one phase from 2.2 s to 63.8 s at a run time of 94
18 h. PFGE profiles were analyzed using Bionumerics software v3.5 (Applied Maths, Sint-95
Martens-Latem, Belgium) (3). 96
Antimicrobial Susceptibility Testing. All of the S. Bovismorbificans isolates were tested for 97
susceptibility to standardized panels of antimicrobial drugs by the broth microdilution method 98
(Sensititre® panel type: CMV1AGNF, Trek Diagnostic Systems, Westlake, OH) and the analysis 99
was conducted according to the rigorously standardized Clinical and Laboratory Standards 100
Institute (CLSI) protocols (6, 7, 8) ). CLSI approved interpretive criteria were used when 101
available; otherwise provisional National Antimicrobial Resistance Monitoring System 102
(NARMS) breakpoints were used (6, 7, 8). 103
Optical mapping. Genome mapping was conducted using Optical mapping. High molecular 104
weight DNA was prepared from bacterial colonies following the manufacturer’s procedures 105
(OpGen, Gaithersburg, MD). DNA immobilized on treated glass slides was digested with the 106
BamHI restriction enzyme, and stained with fluorescent dye. Contiguous immobilized 107
restriction fragments were sized and mapped across molecules ranging from 100,000 to 108
1,000,000 base pairs (bp). Images from a maximum of 50,000 molecules were collected and 109
assembled on the Argus mapping station, short DNA sequence contigs were assembled into a 110

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
6
complete circular chromosome. Optical maps were aligned and compared to reference 111
sequenced genomes (13). 112
Multiple-locus variable-number tandem-repeats analysis (MLVA). MLVA analysis was 113
conducted with the outbreak S. Bovismorbificans isolates using multiplex PCR and capillary 114
electrophoresis (QIAxcel System, Gaithersburg, MD). DNA templates were prepared from broth 115
cultures of LB grown at 37°C for 4 hrs. Cells were harvested and DNA was extracted using the 116
Wizard Genomic DNA Purification System (Promega, Madison, WI). Total DNA was 117
resuspended in 0.3 ml Tris-EDTA (Promega) buffer to give a final concentration 350 ng/ul to 118
550 ng/ul. S Enteritidis strain P125109 (Gene Bank AM933172.1) genome sequence was used to 119
search for variable number tandem repeats (VNTR) using the Tandem Repeat Finder (2, 19). 120
Five of the VNTR regions were selected and primers were designed using Primer3 (v. 0.4.0) 121
software (Primer3source forge.net) and designated as SE1, SE2 SE3, SE4 and SE5 (Table 2). 122
Two multiplex reactions for the five targeted loci (SE1-SE5) were used to generate MLVA 123
profiles with high diversity capacity using the QIAxcel bio-analyzer (QIAxcel System ). 124
PCR amplifications were performed using the Type-it Microsatellite PCR Kit (Qiagen 125
Inc., Balencia, CA) using a MJ Research PTC-200 Thermo Cycler (MJ Research, Ramsey, MN) 126
and 12.5 µl 2x Type-it Multiplex PCR Master Mix, 12.5 µl primer mix (2 µM of each primer), 6 127
µl RNase-free water, 5 µl 5x Q-Solution and 1.0 µl Template DNA. The PCR conditions were, 128
95 °C for 5 min; 35 cycles of 95°C for 30s, 60°C for 90s, 72°C for 30s; final extension at 68°C 129
for 10 min. PCR products were run on the QIAxcel System and the molecular sizes were 130
calculated using the Biocalculator software (eGene,Inc., Irvine CA) (19). 131
REP-PCR. The BioMerieux’s Diversilab® system is a Repetitive Extragenic Palindromic 132
(REP)-PCR based DNA fingerprinting analysis system (BioMerieux, Durham, NC). This method 133

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
7
was evaluated for its ability to differentiate or subtype the outbreak and non-outbreak isolates. 134
Cultures were grown for 18-24 h at 37±2°C on sheep Blood Agar (SBA). DNA was isolated 135
using Roche MagNa Pure Compact (Roche, Indianapolis, IN). Final DNA concentrations were 136
adjusted to 75 +/- 25 ng/µl. Data analysis was performed using BioMerieux’s Diversilab® 137
software. 138
Salmonella virulence plasmid analysis. The presence of the Salmonella virulence plasmid was 139
also assessed among the isolates. Plasmids were isolated using the Qiagen Plasmid Midi Kit 140
(Qiagen). Strains containing plasmid within the range of known Salmonella virulence plasmids 141
were evaluated for the presence of plasmid-associated virulence genes based on primers reported 142
by Carattoli et al., (4) and primers designed for this study that are specific to the origin of 143
replication genes such as qrmD, Orf4, Orf5, spvB and pef (Table 2) (4, 11). 144
145
RESULTS AND DISCUSSION 146
Molecular Serotyping: The Agilent Bio-analyzer 2100 provided a digital gel image depicting 147
the various sized amplicons which is used to generate the serotype-specific banding patterns 148
corresponding to each known serotype. Molecular serotyping was conducted and the results 149
confirmed that all 21 isolates used in this study are of the S. Bovismorbificans pattern BCEG as 150
reported by Jean-Gilles Beaubrun et al., (12) and confirming the results of the traditional 151
serotyping method. 152
PFGE: The PFGE banding pattern was identical for all S. Bovismorbificans isolates associated 153
with the hummus outbreak, and this pattern was distinguishable from the PFGE patterns obtained 154
from the non-outbreak isolates. There are five distinct clusters showing different banding 155

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
8
patterns (Figure 1). The S. Bovismorbificans isolates cultured directly from the hummus have a 156
PFGE pattern which is indistinguishable from the patterns of the outbreak clinical strains (Figure 157
1). As demonstrated in Figure 1 using XbaI, the hummus outbreak isolates contained several 158
fragments at approximately 880 kb, 523 kb, and 104-66 kb that are clearly distinguishable from 159
the fragments observed in the non-outbreak isolates (Figure 1). Concurrently, using BlnI, there is 160
a 336-291kb variable region in the outbreak isolates that is not observed in the non-outbreak 161
isolates. The results also show that SAL185 and SAL644 seem to be more closely related to 162
each other, but certainly distinguishable from the outbreak isolates. PFGE patterns demonstrate 163
that the outbreak isolates from DC_PHL distinctly cluster together compared to the other isolates 164
of S. Bovismorbificans. Isolates obtained from the Michigan State laboratory form two distinct 165
clusters (Figure 1). One isolate from Michigan (SAL 682), which was identified as an outbreak 166
isolate, matches the PFGE pattern obtained from the DC_PHL isolates. However, SAL676 did 167
not match the PFGE pattern although it was originally reported to be part of the outbreak. 168
Antimicrobial Susceptibility Testing: The antimicrobial drug (AMD) susceptibility assay 169
demonstrates that there is no resistance in the isolates and the results are represented in Figure 1. 170
From the 21 isolates tested, it is clear that 12 of the isolates are related to the same hummus 171
outbreak and 9 isolates are completely unrelated and all of the isolates of S. Bovismorbificans 172
were susceptible to the AMDs found in the antimicrobial panel. The presence of various 173
antibiotic resistant genes was examined using PCR in addition to the AMD test that was 174
conducted. For example the qrmD gene targeted on the plasmid analysis did not suggest the 175
presence of the quinolone antibiotic resistant genes. However, the consistent finding was that the 176
isolates from the hummus outbreak were certainly different as compared to the non-outbreak 177
isolates used in this study. 178

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
9
Optical Mapping: Optical mapping revealed that the outbreak strains possessed three unique 179
gene regions including 1) the evolutionary distances between outbreak and non-outbreak isolates, 180
2) the chromosome structure of the outbreak strain, which is usually compared to the most 181
similar sequenced Salmonella reference isolates from GenBank, 3) and lastly, a list of specific 182
chromosomal markers unique for the outbreak strain. Optical mapping of S. Bovismorbificans 183
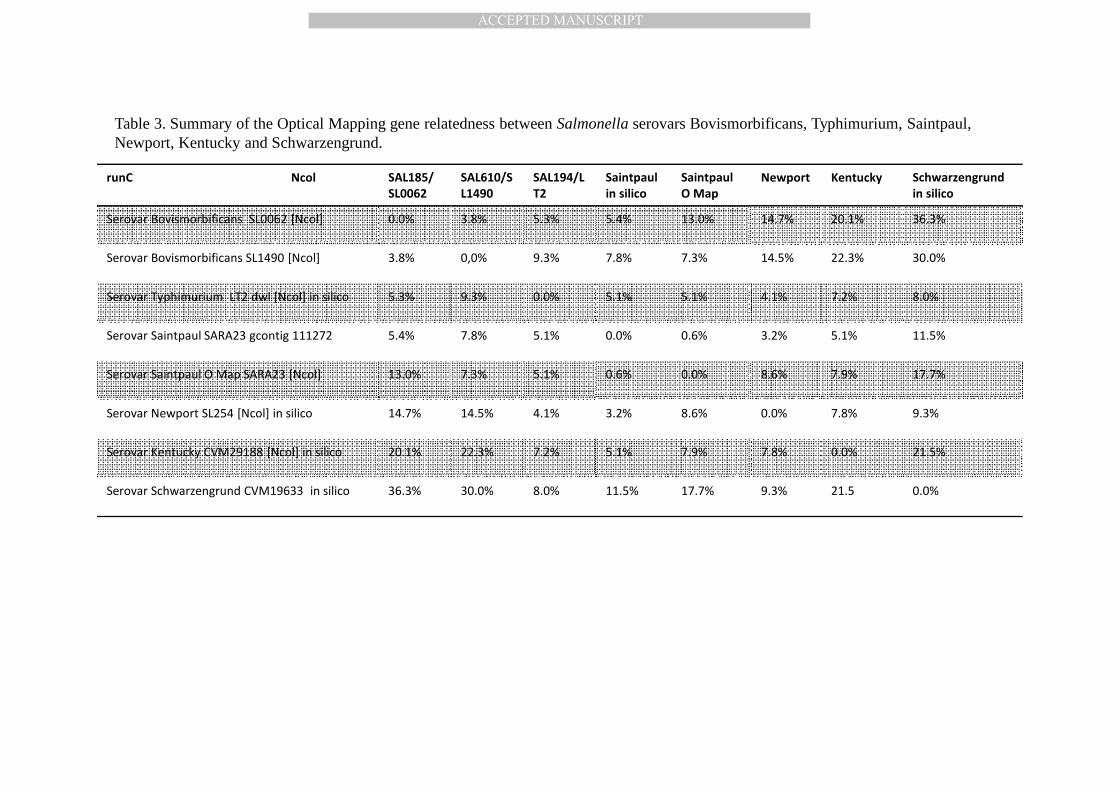
outbreak isolate SAL610 (SL1490) shows that it is most closely related to another non-outbreak 184
Bovismorbificans isolate SAL185 (SL0062) with a 3.8% difference (Table 3). The next closest 185
serovars are S. Typhimurium LT2 and S. Saintpaul SARA23, which were 5.3% and 5.4% 186
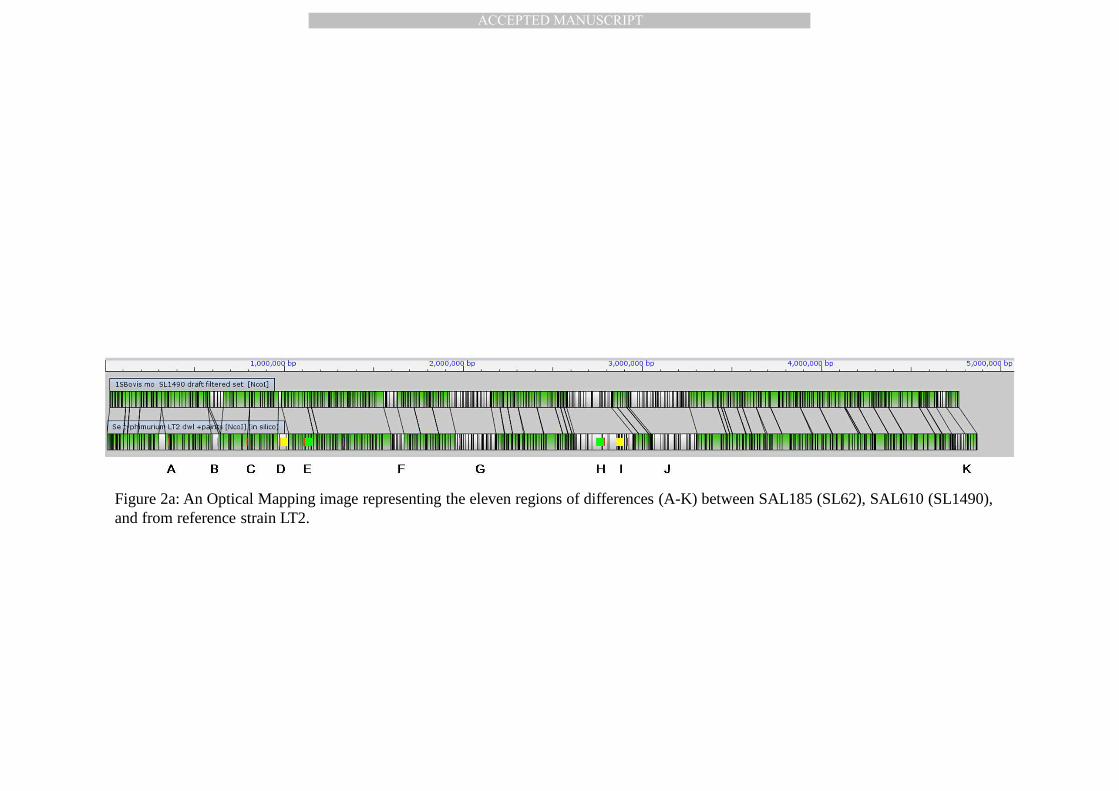
different. Optical mapping also revealed 11 major regions (A-K), excluding plasmids, that are 187
different relative to S. Typhimurium LT2 (4,857,372 kbp), and that the 4.740 Mbp chromosome 188
of S. Bovismorbificans is shorter by 117 kbp (Figure 2a and Table 3). OM results also showed 189
that the plasmid is unique to S. Bovismorbificans involved in the hummus associated outbreak. 190
Four genomic regions of interest , A, B, C, and D are simple insertion/deletion events; 191
region E is a variant Gifsy-2 prophage, and regions H – K are variable syntenic regions (VSRs). 192
Additionally genomic regions of interest H and I appear to be composites as they contained both 193
a VSR and variant prophages. VSRs are highly similar segments of the chromosome between 194
strains containing enough single nucleotide diversity to create and eliminate restriction sites 195
within sets of syntenic genes. A level of 1-3% nucleotide divergence creates enough change in 196
restriction fragment patterns that portions of otherwise aligned chromosomes appear as 197
unaligned sections of similar length. Upon closer examination, patterns of restriction site loss 198
and gain can be seen within the unaligned segments (Figure 2b). 199
200

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
10
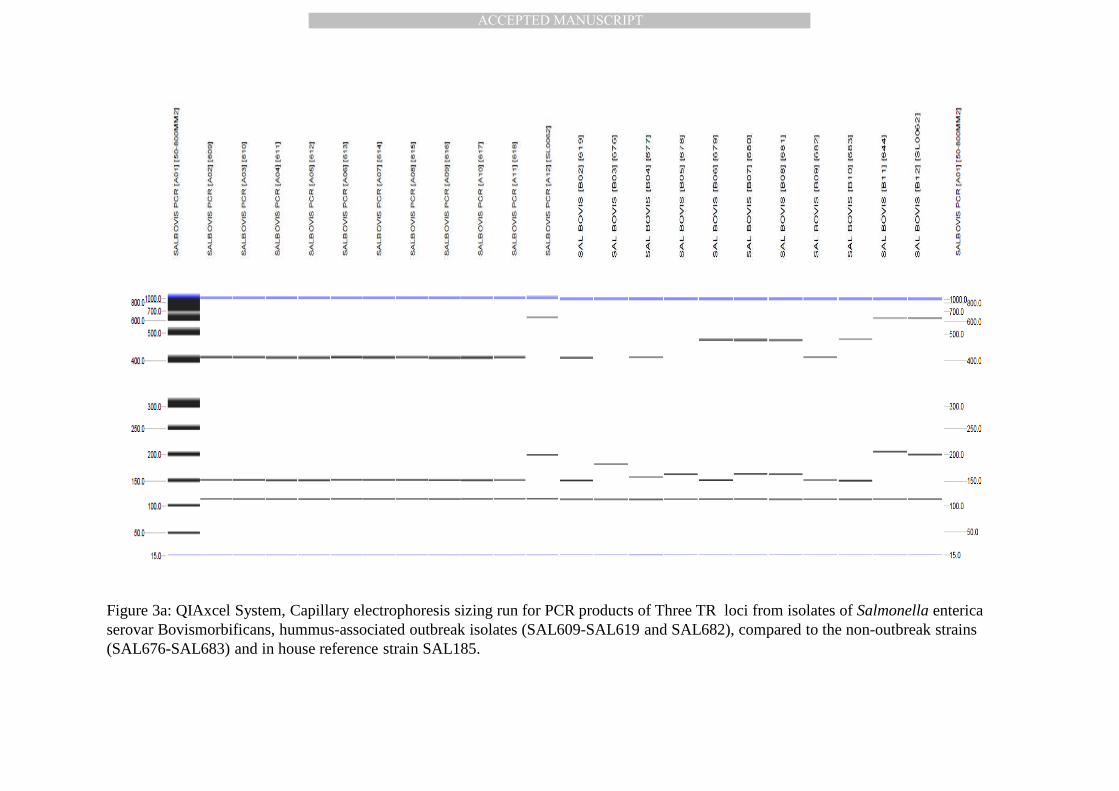
MLVA analysis. Multi-locus variable-number tandem repeat analysis (MLVA) consisted of 201
PCR-based amplification of five chromosomal loci and accurate sizing of the resulting sets of 202
PCR amplicons by electrophoresis. MLVA examines specific tandem repeats (TRs) found at 203
different genetic loci within the genome. The data from the MLVA analysis demonstrates clear 204
and distinguishable differences between the isolates from the hummus outbreak and the non-205
outbreak isolates (Figure 3a and 3b). Two multiplexes were used for the MLVA analysis of the 206
21 isolates of S. Bovismorbificans. The results show eight different MLVA banding patterns 207
amongst the isolates for the first multiplex reaction, although the isolates from the Hummus 208
outbreak cluster together with one consistent pattern (Figure 3a). The second multiplex 209
contained two targets, which seem to be present in 16 of the 21 isolates (Figures 3b). 210
REP-PCR: The results from the Diversilab®’s REP-PCR analysis show two main clusters, 211
which clearly separate the outbreaks isolates from the non-outbreak isolates (Figure 4). In 212
addition, the results from the REP-PCR analysis suggest that SAL185 and SAL644, which are in 213
house controls, clustered with the outbreak related isolates, which supports the hypothesis that 214
the repetitive region of SAL185 and SAL 644 may be more closely related to the hummus-215
associated outbreak isolates than some of the other S. Bovismorbificans isolates used in this 216
study. SAL185 and SAL644 contained a pattern of REP sequences that are ~90 (± 5) percent 217
similar to those of the hummus-associated outbreak isolates. Based on the grouping of the 23 218
isolates presented in Figures 1, 3a, 3b, and 4using PFGE, MLVA, and REP-PCR, the results 219
demonstrated that there were genetic differences between the isolates from the outbreak as 220
compared to the non-outbreak isolates. 221
Plasmid analysis. The size of the Salmonella virulence plasmid is serovar dependent, ranging 222
from ∼50 to 100 kb (1). Optical mapping suggest the presence of an 80 kb plasmid. The 223

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
11
plasmid was extracted and the origin of replication (ORi), repA gene was confirmed to indicate 224
the presence of an Inc FIIs plasmid in the S. Bovismorbificans isolates. Using the primer target 225
reported by Carattoli et al. (4) and primers designed for this study, the results shown in Table 4 226
confirm the presence of an Inc FIIs plasmid in the outbreak isolates. In addition, the PCR 227
product obtained suggests that the plasmids identified in the hummus outbreak isolates may be 228
closely related to plasmids found in S. Typhimurium. PCR products were obtained using the 229
primers targeted for pefA, spvB and Inc FIIs. 230
The plasmid in the outbreak isolates was confirmed using primer-specific targets as 231
demonstrated in Table 2. In a reports published in Genomes Announcements (11,12) the isolates 232
used in this study was whole genome sequenced and the results showed that outbreak isolates 233
contained two plasmids, one of which demonstrates similarity to the 93 Kb pSLT2 IncF-type 234
plasmid of S. Typhimurium and an Integration and Conjugative Element (ICE) similar to ICESb1 235
found in Salmonella bongori (17). The presence of recurrent and common plasmids in 236
epidemiologically unrelated Salmonella isolates of different serotypes suggests the successful 237
spread of these genetic determinants amongst Salmonella spp. This is to say that the hummus-238
associated strain may have acquired new genetic information through horizontal gene transfer 239
(HGT) from other Salmonella spp., thereby increasing their virulence. These findings are 240
significant since they suggest an uptake of a plasmid in the outbreak genome that may have 241
increased the probability of the pathogenicity of the hummus associated strains. The approach 242
used in this study using the Optical mapping and plasmid analysis could be applied to monitor 243
the circulation of plasmids within strains from different environments or to follow the horizontal 244
transmission of virulence genes among Enterobacteriaceae and track emerging pathogens. 245
246

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
12
CONCLUSION 247
Salmonella is one of the main causes of gastroenteritis in humans. S. Bovismorbificans have 248
been linked to foodborne outbreaks associated with alfalfa sprouts, homemade cheese, pasta 249
salad, striped bass, lettuce and pork (20). It has not been previously related to sesame paste 250
(tahini), which are used to make hummus. A report in MMWR and Food Safety News about the 251
District of Columbia outbreak investigation described the difficulties associated with the 252
hummus-associated outbreak in terms of identifying the probable cause or source of the 253
contamination (22, 23). In this study, S. Bovismorbificans isolates from the hummus-associated 254
outbreak were compared to non-outbreak related isolates using a polyphasic approach which 255
included PCR molecular serotyping, PFGE, AMD, optical mapping, MLVA, Diversilab®, and 256
plasmid analysis. The results showed the presence of plasmids in the hummus-associated isolates 257
that are not present in the other non-outbreak isolates. 258
The various molecular approaches used in this study support one another in determining 259
that the outbreak strains were unique unto themselves as compared to other non-outbreak S. 260
Bovismorbificans strains. In this multifaceted approach to investigate the uniqueness of the S. 261
Bovismorbificans isolates related to the hummus outbreak, PFGE analysis clustered the isolates 262
that were closely related, Optical mapping determined the presence of the plasmid unique to the 263
of outbreak isolates, and the other molecular approaches such as MLVA, and Diversilab also 264
show the uniqueness the outbreak isolates. Lastly, WGS results demonstrated the presence of 265
two plasmids, one of which possesses significant similarity to the S. Typhimurium 93 Kb pSLT2 266
IncF-type plasmid and an ICE similar to ICESb1 found in Salmonella bongori. 267

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
13
The expanded scientific knowledge though the use of this polyphasic molecular 268
approach to characterizing these pathogens has provided useful insights for both risk assessment 269
and methods development for food safety. The goal is to create faster method detection, 270
identification and characterization of Salmonellae in the event of an outbreak and thus improve 271
the safety of the food supply. Using Optical mapping and plasmid characterization may help 272
develop new approaches for epidemiology analysis. 273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
14
ACKNOWLEGMENTS 291
We acknowledge and thank everyone from the Department of Health from District of 292
Columbia Public Health Laboratory (DC_PHL) for providing the isolates used in this study, and 293
FDA scientists from the Center of Food Safety and Applied Nutrition (CFSAN), Office of 294
Applied Research and Safety Assessment (OARSA), Center for Veterinary Medicine (CVM), in 295
Laurel MD who were involved in the collaborative study. Also thanks to the Delaware and 296
Michigan Public Health for assisting with providing extra strains, Mark Mammel and Chris 297
Elkins at FDA for your support. 298
299
300
301
302
303
304
305
306
307
308
309
310

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
15
REFERENCES 311
1. Amavisit, P., D. Lightfoot, G. F. Browning, and P. F. Markham. 2003. Variation between 312
Pathogenic Serovars within Salmonella Pathogenicity Islands. J. Bacteriol. 185:3624-313
3635. 314
2. Benson, G. 1999. Tandem repeats finder: a program to analyze DNA sequences. Nucleic 315
Acids Res. 27:573–580. 316
3. BioNumerics. Keistraat 120, B-9830 Sint-Martens-Latem, Belgium. www.applied-317
maths.com. 318
4. Carattoli, A., A. Bertini, L. Villa, V. Falbo, K. L. Hopkins, E. J. Threlfall. 2005. 319
Identification of plasmids by PCR-based replicon typing. J. of Microbiological Meth. 63: 320
219-228. 321
5. Cavaco, L.M., H. Hasman, S. Xia, and F.M. Aarestrup. 2009. qnrD, a novel gene 322
conferring transferable quinolone resistance in Salmonella enterica Serovar Kentucky 323
and Bovismorbificans strains of Human Origin. Antimicrob. Agents Chemother. 53:603-324
608. 325
6. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial 326
Susceptibility Testing; Seventeenth Informational Supplement (M100-S17). 2007. 327
CLSI, Wayne, Pa. 328
7. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial 329
Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals; Informational 330
Supplement (M31-S1). 2004. CLSI, Wayne, Pa. 331

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
16
8. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial 332
Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals; Approved 333
Standard – Second Edition (M31-A2). 2002. CLSI, Wayne, Pa. 334
9. Gopal Gopinath, Junia Jean-Gilles Beaubrun, Chris Grim, Morris Blaylock, Reginald 335
Blackwell, Sosina Merid, Alpha Diallo, and Darcy Hanes. 2014. Whole Genome 336
Sequences of six Salmonella enterica Serovar Bovismorbificans Isolates Associated with 337
a 2011 Multistate Hummus Outbreak. Genome Announcements under review. 338
10. Gopal Gopinath, Junia Jean-Gilles Beaubrun, Chris Grim, and Darcy Hanes. 2014. Draft 339
Genome Sequences of nine Salmonella enterica Serovar Bovismorbificans isolates from 340
various sources. Genome Announcements under review. 341
11. Heithoff, D. M., W. R. Shimp, P. W. Lau, G. Badie, E. Y. Enioutina, R. A. Daynes, B. A. 342
Byrne, J. K. House, and M. J. Mahan. 2008. Human Salmonella Clinical isolates distinct 343
from those of animal origin. Appl. Environ. Microbiol. 74:1757-1766. 344
12. Jean-Gilles Beaubrun, J., C. Cheng, K. Chen, L. Ewing, H. Wang, M.C. Agpaoa, M. J. 345
Huang, E. Dickey, J.M. Du, D.M. Williams-Hill, B. Hamilton, S.A. Micallef, R.E. 346
Rosenberg-Goldstein, A. George, S.W. Joseph, A.R. Sapkota, A.P. Jacobson, B.D. Tall, 347
M.H. Kothary, and D.E. Hanes. 2012. The use of a PCR-Based Method for Identification 348
of Salmonella enterica serotypes Newport, Typhimurium, Javiana and Saint Paul from 349
various food matrices. Food Microbiology, 31: 199-209. 350
13. Kotewicz ML, Mammel MK, LeClerc JE, Cebula TA. 2008. Optical mapping and 454 351
sequencing of Escherichia coli O157:H7 isolates linked to the US 2006 spinach-352
associated outbreak. Microbiology 154:3518–3528. 353

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
17
14. Kim, S., J. G. Frye, J. Hu, P. J. Fedorka-Cray, R. Gautom, and D. S. Boyle. 2006. 354
Multiplex PCR-Based Method for Identification of Common Clinical Serotypes of 355
Salmonella enterica subsp. Enterica. J. Clin. Microbiol. 44:3608-3615. 356
15. Po¨nka¨, A., Y. Andersson, A. Siitonen, B. de Yong, M. Jahkola, O. Haikala, A. 357
Kuhmonen, and P. Pakkala. 1995. Salmonella in alfalfa sprouts. Lancet 345:462–463. 358
16. Puohiniemi, R., T. Heiskanen, and A. Siitonen. 1997. Molecular epidemiology of two 359
international sprout-borne Salmonella outbreaks. J. Clin. Microbiol. 35:2487-91. 360
17. Seth-Smith HM, Fookes MC, Okoro CK, Baker S, Harris SR, Scott P, Pickard D,Quail 361
MA, Churcher C, Sanders M, Harmse J, Dougan G, Parkhill J, Thomson NR.Structure, 362
diversity, and mobility of the Salmonella pathogenicity island 7 family of integrative and 363
conjugative elements within Enterobacteriaceae. J. Bacteriol. 2012 Mar;194(6):1494-504. 364
18. Rimhanen-Finne, R., R. T. Niskanen, T. Lienemann, T. Johansson, M. Sjo¨man, T. 365
Korhonen, S. Guedes, H. Kuronen, M. J. Virtanen, J. Ma¨ kinen, J. Jokinen, A. Siitonen 366
and M. Kuusi. 2011. Nationwide Outbreak of Salmonella Bovismorbificans Associated 367
with Sprouted Alfalfa Seeds in Finland, 2009. Zoonoses Public Health. 58: 589–596. 368
19. Rozen, S. and H. J. Skaletsky. 2000. Primer3 on the WWW for general users and for 369
biologist programmers. In: Krawetz S, Misener S (eds) Bioinformatics Methods and 370
Protocols: Methods in Molecular Biology. Humana Press, Totowa, NJ, pp 365-386 371
20. www.cdc.gov/salmonella/outbreaks.html 372
21. www.cdc.gov/pulsenet/protocols.html 373
22. www.cdc.gov/mmwr/preview/mmwrhtml/mm6146a3.htm. Multistate Outbreak of 374
Salmonella Serotype Bovismorbificans Infections Associated with Hummus 375
and Tahini — United States, 2011. 376

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
18
23. www.foodsafetynews.com/2012/11/government-reports-2011-salmonella-outbreak-377
linked-to-tahini. CDC: Salmonella from Tahini Sickened 23 Last Year. 378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
19
FIGURES 400
Figure 1: PFGE and AMD results of the Salmonella enterica serovar Bovismorbificans. 401
402
Figure 2a: An Optical Mapping image representing the eleven regions of differences (A-K) 403
between SAL185 (SL62), SAL610 (SL1490), and from reference strain LT2. 404
405
Figure 2b: An Optical Mapping image representing the variable syntenic regions of SAL185, 406
SAL609 and from reference strain LT2. 407
408
Figure 3a: QIAxcel System, Capillary electrophoresis sizing run for PCR products of Three TR 409
loci from isolates of Salmonella enterica serovar Bovismorbificans, hummus-associated outbreak 410
isolates (SAL609-SAL619 and SAL682), compared to the non-outbreak strains (SAL676-411
SAL683) and in house reference strain SAL185. 412
413
Figure 3b: QIAxcel System, Capillary electrophoresis sizing run for PCR products of two TR 414
loci from Isolates of Salmonella enterica serovar Bovismorbificans, hummus-associated 415
outbreak isolates (SAL609-SAL619 and SAL682), compared to the non-outbreak strains 416
(SAL676-SAL683) and in house reference strain SAL185. 417
418
Figure 4: A representation of the Rep-PCR results using Diversilab® software to demonstrate in 419
a dendogram, digital gel image the percent relatedness between hummus-associated outbreak 420
isolates (SAL609-SAL619 and SAL682), compared to the non-outbreak strains (SAL676-421
SAL683) and in house reference strain SAL185. 422

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
20
TABLES 423
Table 1. Summary of all the strains used in this study and the appropriate identification from 424
various laboratories. 425
426
Table 2. List of the primers used in this study for molecular characterization of Salmonella 427
serovar Bovismorbificans. 428
429
Table 3. . Summary of the Optical Mapping gene relatedness between Salmonella serovars 430
Bovismorbificans, Typhimurium, Saintpaul, Newport, Kentucky and Schwarzengrund. 431
432
Table 4. Results of the plasmids PCR analysis of Salmonella serovar Bovismorbificans from the 433
Hummus associated outbreak isolates compared to non-outbreak isolates. 434
435
436
437
438
439

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
ID DMB CVM DCPHL Source Serotypes Location Outbreak
SAL194 SARA2 LT2 Typhimurium
SAL609 SL1489 41673 11-399 Stool-RFBovismorbificans DC Hummus
SAL610 SL1490 41674 11-405 Stool-RFBovismorbificans DC Hummus
SAL611 SL1491 41675 11-440 Stool-RFBovismorbificans DC Hummus
SAL612 SL1492 41676 11-441 Stool-RFBovismorbificans DC Hummus
SAL613 SL1493 41677 11-466 Stool-RFBovismorbificans DC Hummus
SAL614 SL1494 41678 11-495 Stool-RFBovismorbificans DC Hummus
SAL615 SL1495 41679 11-510 Stool-RFBovismorbificans DC Hummus
SAL616 SL1496 41680 11-528 FoodBovismorbificans DC Hummus
SAL617 SL1497 41681 11-538 FoodBovismorbificans DC Hummus
SAL618 SL1498 41682 11-539 FoodBovismorbificans DC Hummus
SAL619 SL1499 41683 11-565 Stool-RFBovismorbificans DC Hummus
SAL676 SL1542 41838 12-180 Clinical Bovismorbificans DE Hummus
SAL677 SL1543 41839 12-198 Clinical Bovismorbificans MI N/A
SAL678 SL1544 41840 12-199 Clinical Bovismorbificans MI N/A
SAL679 SL1545 41841 12-200 Clinical Bovismorbificans MI N/A
SAL680 SL1546 41842 12-201 Clinical Bovismorbificans MI N/A
SAL681 SL1547 41843 12-202 Clinical Bovismorbificans MI N/A
SAL682 SL1548 41844 12-203 Clinical Bovismorbificans MI Hummus
SAL683 SL1549 41845 12-204 Clinical Bovismorbificans MI N/A
SAL185 SL0062 41672 Clinical Bovismorbificans MI N/A
SAL644 SL1541 17906 Human Bovismorbificans CVM N/A
SAL155 SL1541 Tomato Farm Newport VA N/A
Table 1. Summary of all the strains used in this study and the appropriate identification from various laboratories.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Table 2: List of the primers used in this study for molecular characterization of Salmonella serovar Bovismorbificans.
Primer Primer sequence Target Size bp
Forward Reverse
Plasmid 5’-TATTGCACTGGGTGTTCTGGGC-3’ 5’-TTGTAAGCCACTGCGAAAGACGC-3’ Pef A 473 This study
Plasmid 5’-AGCATCTCTGCCTCTCCCTTA-3’ 5’-CAGTATACCGCTGCCT-3’ spvB 1491 This study
Plasmid 5’-AGTTTTCACGAGATCAATTTACG-3’ 5’-CCTAAACTCTCAACAAGCTGAAG-3’ qnrD 578 This study
Plasmid5’-TCACACTCCTGCTGTCGGCGGAGC-3’
5’-TCCGTTGTCAGCAGCTTTTCAAGG-3’ orf5 454 This study
Plasmid 5’-TCAGTCAGCGAAAAGATCTCGAAC-3’ 5’-TGCTGCGAGATATCATGCGTCAGAG-3’ orf4 732 This study
Plasmid 5’-CTGTCGTAAGCTGATGGC-3’ 5’-CTCTGCCACAAACTTCAGC-3’ repA 270 Carattoli et. al.
MLVA 5’-AGGAAACACAGCCGCAAT-3’ 5’-AACCGGCTGTTCGATAACC-3’ SE1 151 This study
MLVA 5’-GCGCTAAACAAGCCGCTCAT-3’ 5’-TCGTTAACAACCTGCTGCTGT-3’ SE2 413 This study
MLVA 5’-ATGTTCTGGCGGACATGG-3’ 5’-GGCATCCTGATACGCTTTTG-3’ SE3 116 This study
MLVA 5’-TGGCTATTGGCGTTGAAAAT-3’ 5’-AAGGTAATAACAGAGTCATC-3’ SE4 488 This study
MLVA 5’-TCCTGCATACCAACGATCAC-3’ 5’-CTCTTGCGCCACCTCTTTAC-3’ SE5 112 This study
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Table 3. Summary of the Optical Mapping gene relatedness between Salmonella serovars Bovismorbificans, Typhimurium, Saintpaul, Newport, Kentucky and Schwarzengrund.
runC Ncol SAL185/
SL0062
SAL610/S
L1490
SAL194/L
T2
Saintpaul
in silico
Saintpaul
O Map
Newport Kentucky Schwarzengrund
in silico
Serovar Bovismorbificans SL0062 [Ncol] 0.0% 3.8% 5.3% 5.4% 13.0% 14.7% 20.1% 36.3%
Serovar Bovismorbificans SL1490 [Ncol] 3.8% 0,0% 9.3% 7.8% 7.3% 14.5% 22.3% 30.0%
Serovar Typhimurium LT2 dwl [Ncol] in silico 5.3% 9.3% 0.0% 5.1% 5.1% 4.1% 7.2% 8.0%
Serovar Saintpaul SARA23 gcontig 111272 5.4% 7.8% 5.1% 0.0% 0.6% 3.2% 5.1% 11.5%
Serovar Saintpaul O Map SARA23 [Ncol] 13.0% 7.3% 5.1% 0.6% 0.0% 8.6% 7.9% 17.7%
Serovar Newport SL254 [Ncol] in silico 14.7% 14.5% 4.1% 3.2% 8.6% 0.0% 7.8% 9.3%
Serovar Kentucky CVM29188 [Ncol] in silico 20.1% 22.3% 7.2% 5.1% 7.9% 7.8% 0.0% 21.5%
Serovar Schwarzengrund CVM19633 in silico 36.3% 30.0% 8.0% 11.5% 17.7% 9.3% 21.5 0.0%

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Lab ID Serotypes qnrDa Orf4a Orf5a pefAa
500 bp
spvBa
1300 bp
FIIsb
270 bp
*SAL194 Typhimurium - - - + + +
SAL609 Bovismorbificans - - - + + +
SAL610 Bovismorbificans - - - + + +
SAL611 Bovismorbificans - - - + + +
SAL612 Bovismorbificans - - - + + +
SAL613 Bovismorbificans - - - + + +
SAL614 Bovismorbificans - - - + + +
SAL615 Bovismorbificans - - - + + +
SAL616 Bovismorbificans - - - + + +
SAL617 Bovismorbificans - - - + + +
SAL618 Bovismorbificans - - - + + +
SAL619 Bovismorbificans - - - + + +
*SAL676 Bovismorbificans - - - + - -
*SAL677 Bovismorbificans - - - + - +
*SAL678 Bovismorbificans - - - + - -
*SAL679 Bovismorbificans - - - + - -
*SAL680 Bovismorbificans - - - + - -
*SAL681 Bovismorbificans - - - + - -
SAL682 Bovismorbificans - - - + - +
*SAL683 Bovismorbificans - - - + - -
*SAL185 Bovismorbificans - - - - - -
*SAL644 Bovismorbificans - - - - - -
Table 4. Results of the plasmids PCR analysis of Salmonella serovar Bovismorbificans from the Hummus associated outbreak
isolates compared to non-outbreak isolates.
* non-outbreak isolates, a is primers designed for this study, b Carattoli et al. (15)

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Figure 1: PFGE and MIC results of the Salmonella enterica serovar Bovismorbificans.
880 523 325-280
104-66
336-291
104-72
kb
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Figure 2a: An Optical Mapping image representing the eleven regions of differences (A-K) between SAL185 (SL62), SAL610 (SL1490), and from reference strain LT2.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Figure 2b: An Optical Mapping image representing the variable syntenic regions of SAL185 (SL62), SAL610 (SL1490) and from reference strain LT2.
#fragments locus1 locus 2 locus 3 locus 4
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Figure 3a: QIAxcel System, Capillary electrophoresis sizing run for PCR products of Three TR loci from isolates of Salmonella entericaserovar Bovismorbificans, hummus-associated outbreak isolates (SAL609-SAL619 and SAL682), compared to the non-outbreak strains (SAL676-SAL683) and in house reference strain SAL185.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Figure 3b: QIAxcel System, Capillary electrophoresis sizing run for PCR products of two TR loci from Isolates of Salmonella entericaserovar Bovismorbificans, hummus-associated outbreak isolates (SAL609-SAL619 and SAL682), compared to the non-outbreak strains (SAL676-SAL683) and in house reference strain SAL185.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Rep-PCR
Figure 4: A representation of the Rep-PCR results using Diversilab® software to demonstrate in a dendogram, digital gel image the percent relatedness between hummus-associated outbreak isolates (SAL609-SAL619 and SAL682), compared to the non-outbreak strains(SAL676-SAL683) and in house reference strain SAL185.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Research Highlights
• Comparing Bovismorbificans outbreak versus non outbreak isolates.
• Identification of a plasmid in the outbreak isolates.
• Using multiple molecular approaches for characterization of plasmids