comparative genomic integration profiling of sleeping beauty transposons mobilized with high...
TRANSCRIPT

original article© The American Society of Gene & Cell Therapy
Molecular Therapy vol. 19 no. 8, 1499–1510 aug. 2011 1499
It has been previously shown that integrase-defective HIV-1-based gene vectors can serve, with moderate efficiency, as substrate for DNA transposition by a tran-siently expressed Sleeping Beauty (SB) transposase. Here, we describe the enhanced gene transfer properties of a HIV-1/SB hybrid vector that allows efficient DNA trans-position, facilitated by the hyperactive SB100X transpos-ase, from vector DNA intermediates in primary human cells. Potent transposase-dependent integration of genetic cargo carried by the hybrid HIV-1/SB vector (up to 160-fold above background) is reported in human cell lines as well as in primary human fibroblasts and kerati-nocytes. The efficiency of transgene integration in con-text of the newly developed hybrid vector is comparable with that of conventional lentiviral vectors (LVs). Integra-tion profiles of integrating HIV-1-derived vectors and SB transposons mobilized from LVs are investigated by deep sequencing of a large number of integration sites. A sig-nificant bias of lentiviral integrations in genes is reported, confirming that biological properties of the viral integra-tion machinery facilitate preferred insertion into actively transcribed genomic regions. In sharp contrast, lentivi-ral insertions catalyzed by the SB100X transposase are far more random with respect to genes. Based on these properties, HIV-1/SB vectors may become valuable tools for genetic engineering and therapeutic gene transfer.
Received 12 October 2010; accepted 22 February 2011; published online 5 April 2011. doi:10.1038/mt.2011.47
IntroductIonGenomic integration of transgenes plays a fundamental role in genetic engineering and therapeutic gene transfer. Virus-derived vectors are widely used due to the unique and intrinsic properties of viruses to transfer genetic cargo between cells. HIV-1-based self-inactivating lentiviral vectors (LVs) can efficiently insert their
genetic content into the genome of both proliferating and quies-cent cells, and have, therefore, attracted much attention as vehicles in gene therapy applications. However, a preference for integra-tion into transcription units has been demonstrated in various cell types.1–3 The lentiviral integrase is the primary determinant for viral integration,4 but several cellular proteins interact with the viral preintegration complex during integration.5 One of them, the transcriptional coactivator lens epithelium-derived growth factor p75 (LEDGF/p75), is an essential cofactor for integration.1,6 A bias toward transcriptionally active regions was not observed in LEDGF/p75-depleted cells, providing strong indications that LEDGF/p75 is a fundamental factor for the integration preference of HIV-1 and HIV-1-derived vectors.1,7
Integrase-defective lentiviral vectors (IDLVs) carrying mutated and inactive integrase proteins are prominent new tools for effi-cient and persistent gene transfer to nondividing cells.8–11 Hence, transgene expression from nonintegrating lentiviral DNA has been shown to reach therapeutic levels in neuronal cells,11 and efficient IDLV-derived gene expression has been reported in myotubes,8 liver,9 and human hematopoietic progenitor cells.10 In addition, IDLVs have been efficiently used as substrate for gene correction by homologous recombination in human cell lines and human embryonic stem cells.12,13 During lentiviral transduction, a fraction of the reverse-transcribed DNA is converted into circles with one or two long terminal repeats (LTRs) by homologous recombina-tion and nonhomologous end-joining, respectively. The biologi-cal function of the circular lentivirus-derived DNA is unknown, and the circles are traditionally considered dead-end products of reverse transcription that are eventually lost due to cell division.14 In proof-of-principle studies, we previously showed that circular DNA intermediates can act as substrates for alternative integration machineries added in trans to IDLV-transduced cells.15,16 We gen-erated an IDLV carrying the Flp recognition target (FRT) site, and demonstrated site-specific integration in a drug-selective approach by Flp recombination.15 To create a therapeutically more relevant hybrid vector, we inserted the Sleeping Beauty (SB) transposon
The first two authors contributed equally to this work.Correspondence: Jacob Giehm Mikkelsen, Department of Human Genetics, University of Aarhus, Wilh. Meyers Allé, Bldg. 1242, DK-8000 Aarhus C, Denmark. E-mail: [email protected]
Comparative Genomic Integration Profiling of Sleeping Beauty Transposons Mobilized With High Efficacy From Integrase-defective Lentiviral Vectors in Primary Human CellsBrian Moldt1, Csaba Miskey2,3, Nicklas Heine Staunstrup1, Andreas Gogol-Döring2, Rasmus O Bak1, Nynne Sharma1, Lajos Mátés2, Zsuzsanna Izsvák2,3, Wei Chen2, Zoltán Ivics2,3 and Jacob Giehm Mikkelsen1
1Department of Human Genetics, University of Aarhus, Aarhus, Denmark; 2Max Delbrück Center for Molecular Medicine, Berlin, Germany; 3University of Debrecen, Debrecen, Hungary

1500 www.moleculartherapy.org vol. 19 no. 8 aug. 2011
© The American Society of Gene & Cell TherapyIntegration of Lentiviral DNA by DNA Transposition
inverted repeats (IRs) into the HIV-1 vector in a context that allowed transposition from circular DNA substrates only.16 SB is a Tc1-like transposable element reconstructed from teleost fish.17 The integration pattern of SB seems to have low or no regional prefer-ence,18,19 and the element shows efficient gene insertion in numer-ous preclinical gene therapy applications.20–26 Nevertheless, the SB transposon is not actively transferred between cells, and the SB sys-tem, as a gene transfer tool, therefore relies on plasmid transfection for delivery. Lentiviral delivery of the transposon substrate and the novel hyperactive SB100X transposase27 resulted in a 11-fold transposase-directed increase in genomic insertion in HEK-293 cells obtained by mobilizing transposons carried on circular DNA intermediates.16 Mapping of 87 SB-mediated integrations demon-strated that the hybrid vector had adopted the random integration profile of SB with 30% of insertions mapped within genes, and therefore showed no bias toward transcription units.16 In a par-allel study, Vink and colleagues demonstrated transposition of a forward orientated SB transposon from nonintegrating LVs using an early generation hyperactive transposase, SB11.28 At best, this study showed only a twofold increase over background insertion when both transposon and transposase were delivered by IDLV transduction. Interestingly, however, 53% of the 161 insertion sites identified with this vector in HeLa cells were located within genes, whereas, in contrast, almost 77% of the integration sites mapped for conventional LVs were located within genes. Hence, accumulating findings support the idea that the integration profile of hybrid LVs can be diverted away from that of conventional LVs.29
In the present report, we further developed lentivirus-derived DNA as a platform for gene insertion by the SB transposase. We have improved the overall efficiency of integration resulting in high transgene insertion efficacies in both human cell lines and primary cells. Furthermore, we confirm detargeting of transcrip-tion units by the SB transposase by analyzing >1,000 chromosomal insertions, which represents a thus far unprecedented resolution of SB transposon insertion site mapping in the human genome. Our data show great promise for the use of HIV-1-based IDLV/SB hybrid vectors as vehicles in applications of gene transfer to primary cells.
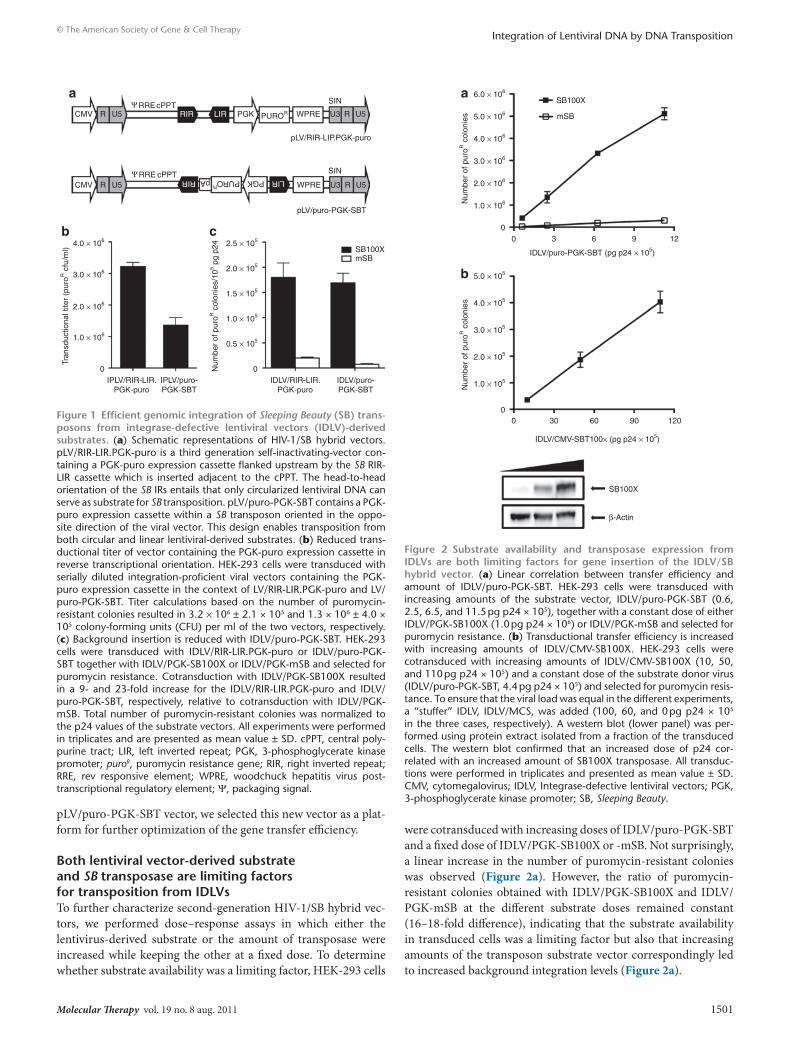
resultsSB transposition efficiency from lentiviral dnA substrates varies between hybrid vector designsWe and others have previously demonstrated that exogenous recombinases and transposases can access and integrate substrates delivered by IDLVs.15,16,28 Importantly, these novel hybrid vectors adopted the integration profiles of the transacting recombinase or transposase. However, the transfer efficacies of the hybrid vec-tors were significantly lower than the transfer efficiency obtained with conventional LVs. To improve the overall performance of the IDLV/SB hybrid vector approach, we set out to construct and evaluate a second-generation vector design based on trans-position directed by the SB100X transposase variant. In our first-generation vector, pLV/RIR-LIR.PGK-puro, the SB right and left inverted repeats (RIR and LIR, respectively) were introduced in a head-to-head orientation between the central polypurine tract and a puromycin resistance gene cassette (Figure 1a, top). In this context, the RIR and LIR defined the outer limits of a relatively
large 8.3-kb long SB transposon (containing the puroR gene as well as the HIV-1 cis elements) that relied on circularization of the viral vector prior to serving as a substrate for the SB transposase. The second-generation vector, pLV/puro-PGK-SBT, contains a fully functional SB transposon (with a puroR gene) without any prior need for circularization; hence both linear and circular lentiviral DNA can serve as substrate for SB transposition (Figure 1a, bot-tom). The orientation of the puroR gene cassette is directed oppo-site the transcriptional orientation of the vector genome to avoid premature termination of transcription in the packaging cells due to the poly(A) site in the transposon.
To compare the transduction efficiencies of the two vectors, integration-proficient lentiviral vectors (IPLV) were transferred in serial dilutions to HEK-293 cells. Transduction titers of 3.2 × 106 ± 2.1 × 105 colony-forming units (CFU)/ml and 1.3 × 106 ± 4.0 × 105 CFU/ml for IPLV/RIR-LIR.PGK-puro and IPLV/puro-PGK-SBT, respectively, were obtained (Figure 1b), indicating that transfer efficiency of the second-generation vector was reduced to about one third of the efficiency observed with the first-generation vec-tors. We did not seek to further explain this titer decline, but note that the second-generation vector encodes vector RNA with long complementary stretches of RNA which may potentially form sta-ble secondary RNA structures, and therefore influence processes like RNA packaging and reverse transcription. Based on previous work reporting high transfer efficiency of an IPLV vector contain-ing the transposon-embedded transgene cassette in the forward orientation,28 we also created a LV vector containing the SBT/PGK-puro transposon in the forward orientation. However, the transfer capacity of this vector was equal to the capacity measured with IPLV/puro-PGK-SBT (data not shown), and for the subse-quent experiments we therefore focused on the vector harboring the transposon in reverse orientation.
We next wanted to compare the two transposon designs as to how efficient the lentivirus-derived DNA could act as sub-strate for the SB transposase. Viral vector preparations carrying the inactive D64V integrase mutant were generated to abolish all active integrase-directed lentiviral insertions. HEK-293 cells were transduced with IDLV/RIR-LIR.PGK-puro or IDLV/puro-PGK-SBT together with a source of SB100X transposase (IDLV/PGK-SB100X) or an inactive SB transposase control (IDLV/PGK-mSB) both driven by the 3-phosphoglycerate kinase (PGK) promoter, demonstrating that the two HIV-1/SB designs had comparable capability to act as substrate for the SB transposase (1.8 × 105 ± 2.8 × 104 and 1.7 × 105 ± 1.9 × 104 puromycin-resistant colonies, normalized to p24 values, for IDLV/RIR-LIR.PGK-puro and IDLV/puro-PGK-SBT, respectively) (Figure 1c). Hence, despite an overall reduction in viral transfer, as evident from Figure 1b, the transductional titers after SB transposition were similar for two hybrid systems. Accordingly, in the absence of SB transpos-ase, IDLV/puro-PGK-SBT (7.4 × 103 ± 1.3 × 102) showed a notably lower level of background integration compared to IDLV/RIR-LIR.PGK-puro (2.8 × 104 ± 1.8 × 103) (Figure 1c). In summary, for IDLV/RIR-LIR.PGK-puro the number of puromycin-resistant colonies was increased ninefold over background in the presence of SB100X, whereas the transfer efficiency of IDLV/puro-PGK-SBT was increased 23-fold over background by SB100X-directed transposition. Due to the enhanced integration properties of the
Molecular Therapy vol. 19 no. 8 aug. 2011 1501
© The American Society of Gene & Cell TherapyIntegration of Lentiviral DNA by DNA Transposition
pLV/puro-PGK-SBT vector, we selected this new vector as a plat-form for further optimization of the gene transfer efficiency.
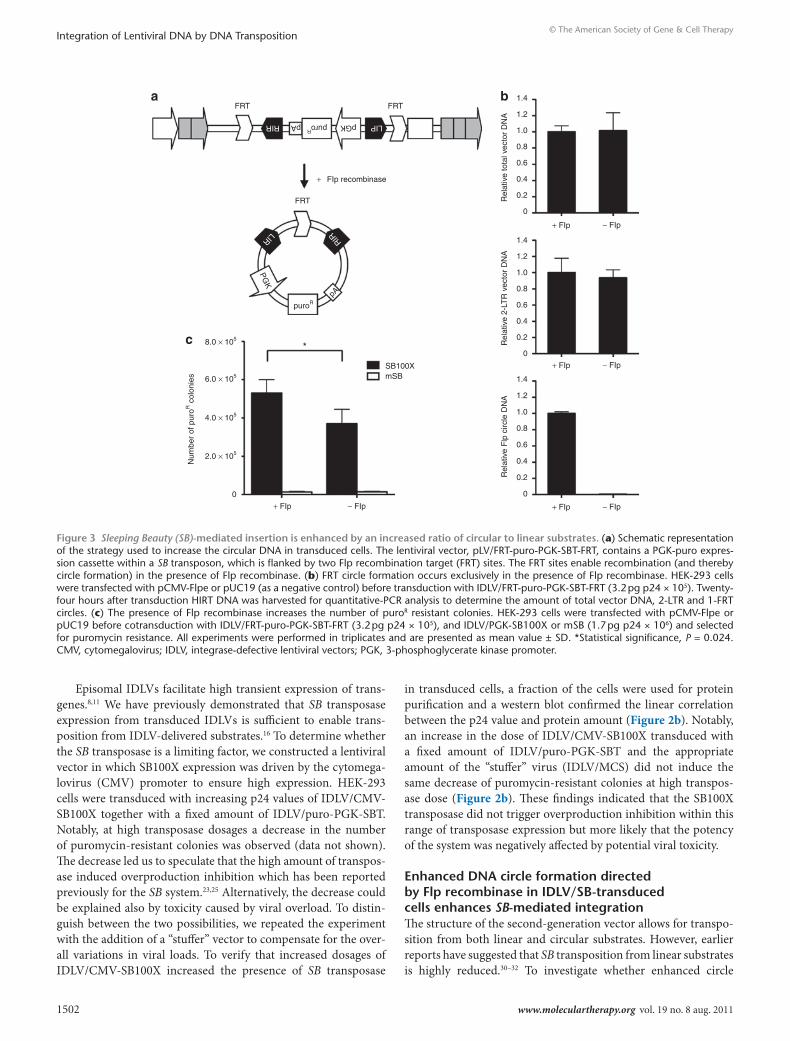
Both lentiviral vector-derived substrate and SB transposase are limiting factors for transposition from IdlVsTo further characterize second-generation HIV-1/SB hybrid vec-tors, we performed dose–response assays in which either the lentivirus-derived substrate or the amount of transposase were increased while keeping the other at a fixed dose. To determine whether substrate availability was a limiting factor, HEK-293 cells
were cotransduced with increasing doses of IDLV/puro-PGK-SBT and a fixed dose of IDLV/PGK-SB100X or -mSB. Not surprisingly, a linear increase in the number of puromycin-resistant colonies was observed (Figure 2a). However, the ratio of puromycin-resistant colonies obtained with IDLV/PGK-SB100X and IDLV/PGK-mSB at the different substrate doses remained constant (16–18-fold difference), indicating that the substrate availability in transduced cells was a limiting factor but also that increasing amounts of the transposon substrate vector correspondingly led to increased background integration levels (Figure 2a).
CMV R U5 RIR LIR PGK PUROR WPRE U3 U5R
SINRREcPPTΨ
CMV R U5
RIR LIRPGKpA PUROR WPRE U3 U5R
SINRRE cPPTΨ
pLV/RIR-LIP.PGK-puro
pLV/puro-PGK-SBT
a
4.0 × 106
3.0 × 106
2.0 × 106
1.0 × 106
0
2.5 × 105
2.0 × 105
1.5 × 105
1.0 × 105
0.5 × 105
0
Tran
sduc
tiona
l tite
r (p
uroR
cfu
/ml)
IPLV/RIR-LIR.PGK-puro
IPLV/puro-PGK-SBT
IDLV/RIR-LIR.PGK-puro
IDLV/puro-PGK-SBT
Num
ber
of p
uroR
col
onie
s/10
5 pg
p24
SB100XmSB
b c
Figure 1 efficient genomic integration of Sleeping Beauty (sB) trans-posons from integrase-defective lentiviral vectors (IdlV)-derived substrates. (a) Schematic representations of HIV-1/SB hybrid vectors. pLV/RIR-LIR.PGK-puro is a third generation self-inactivating-vector con-taining a PGK-puro expression cassette flanked upstream by the SB RIR-LIR cassette which is inserted adjacent to the cPPT. The head-to-head orientation of the SB IRs entails that only circularized lentiviral DNA can serve as substrate for SB transposition. pLV/puro-PGK-SBT contains a PGK-puro expression cassette within a SB transposon oriented in the oppo-site direction of the viral vector. This design enables transposition from both circular and linear lentiviral-derived substrates. (b) Reduced trans-ductional titer of vector containing the PGK-puro expression cassette in reverse transcriptional orientation. HEK-293 cells were transduced with serially diluted integration-proficient viral vectors containing the PGK-puro expression cassette in the context of LV/RIR-LIR.PGK-puro and LV/puro-PGK-SBT. Titer calculations based on the number of puromycin-resistant colonies resulted in 3.2 × 106 ± 2.1 × 105 and 1.3 × 106 ± 4.0 × 105 colony-forming units (CFU) per ml of the two vectors, respectively. (c) Background insertion is reduced with IDLV/puro-PGK-SBT. HEK-293 cells were transduced with IDLV/RIR-LIR.PGK-puro or IDLV/puro-PGK-SBT together with IDLV/PGK-SB100X or IDLV/PGK-mSB and selected for puromycin resistance. Cotransduction with IDLV/PGK-SB100X resulted in a 9- and 23-fold increase for the IDLV/RIR-LIR.PGK-puro and IDLV/puro-PGK-SBT, respectively, relative to cotransduction with IDLV/PGK-mSB. Total number of puromycin-resistant colonies was normalized to the p24 values of the substrate vectors. All experiments were performed in triplicates and are presented as mean value ± SD. cPPT, central poly-purine tract; LIR, left inverted repeat; PGK, 3-phosphoglycerate kinase promoter; puroR, puromycin resistance gene; RIR, right inverted repeat; RRE, rev responsive element; WPRE, woodchuck hepatitis virus post- transcriptional regulatory element; Ψ, packaging signal.
a 6.0 × 106
5.0 × 106
4.0 × 106
3.0 × 106
2.0 × 106
1.0 × 106
0
Num
ber
of p
uroR
col
onie
s
SB100X
mSB
0 3 6 9 12
IDLV/puro-PGK-SBT (pg p24 × 105)
5.0 × 105
4.0 × 105
3.0 × 105
2.0 × 105
1.0 × 105
0
Num
ber
of p
uroR
col
onie
s
b
IDLV/CMV-SBT100× (pg p24 × 105)
0 30 60 90 120
β-Actin
SB100X
Figure 2 substrate availability and transposase expression from IdlVs are both limiting factors for gene insertion of the IdlV/sB hybrid vector. (a) Linear correlation between transfer efficiency and amount of IDLV/puro-PGK-SBT. HEK-293 cells were transduced with increasing amounts of the substrate vector, IDLV/puro-PGK-SBT (0.6, 2.5, 6.5, and 11.5 pg p24 × 105), together with a constant dose of either IDLV/PGK-SB100X (1.0 pg p24 × 106) or IDLV/PGK-mSB and selected for puromycin resistance. (b) Transductional transfer efficiency is increased with increasing amounts of IDLV/CMV-SB100X. HEK-293 cells were cotransduced with increasing amounts of IDLV/CMV-SB100X (10, 50, and 110 pg p24 × 105) and a constant dose of the substrate donor virus (IDLV/puro-PGK-SBT, 4.4 pg p24 × 105) and selected for puromycin resis-tance. To ensure that the viral load was equal in the different experiments, a “stuffer” IDLV, IDLV/MCS, was added (100, 60, and 0 pg p24 × 105 in the three cases, respectively). A western blot (lower panel) was per-formed using protein extract isolated from a fraction of the transduced cells. The western blot confirmed that an increased dose of p24 cor-related with an increased amount of SB100X transposase. All transduc-tions were performed in triplicates and presented as mean value ± SD. CMV, cytomegalovirus; IDLV, Integrase-defective lentiviral vectors; PGK, 3-phosphoglycerate kinase promoter; SB, Sleeping Beauty.
1502 www.moleculartherapy.org vol. 19 no. 8 aug. 2011
© The American Society of Gene & Cell TherapyIntegration of Lentiviral DNA by DNA Transposition
Episomal IDLVs facilitate high transient expression of trans-genes.8,11 We have previously demonstrated that SB transposase expression from transduced IDLVs is sufficient to enable trans-position from IDLV-delivered substrates.16 To determine whether the SB transposase is a limiting factor, we constructed a lentiviral vector in which SB100X expression was driven by the cytomega-lovirus (CMV) promoter to ensure high expression. HEK-293 cells were transduced with increasing p24 values of IDLV/CMV-SB100X together with a fixed amount of IDLV/puro-PGK-SBT. Notably, at high transposase dosages a decrease in the number of puromycin-resistant colonies was observed (data not shown). The decrease led us to speculate that the high amount of transpos-ase induced overproduction inhibition which has been reported previously for the SB system.23,25 Alternatively, the decrease could be explained also by toxicity caused by viral overload. To distin-guish between the two possibilities, we repeated the experiment with the addition of a “stuffer” vector to compensate for the over-all variations in viral loads. To verify that increased dosages of IDLV/CMV-SB100X increased the presence of SB transposase
in transduced cells, a fraction of the cells were used for protein purification and a western blot confirmed the linear correlation between the p24 value and protein amount (Figure 2b). Notably, an increase in the dose of IDLV/CMV-SB100X transduced with a fixed amount of IDLV/puro-PGK-SBT and the appropriate amount of the “stuffer” virus (IDLV/MCS) did not induce the same decrease of puromycin-resistant colonies at high transpos-ase dose (Figure 2b). These findings indicated that the SB100X transposase did not trigger overproduction inhibition within this range of transposase expression but more likely that the potency of the system was negatively affected by potential viral toxicity.
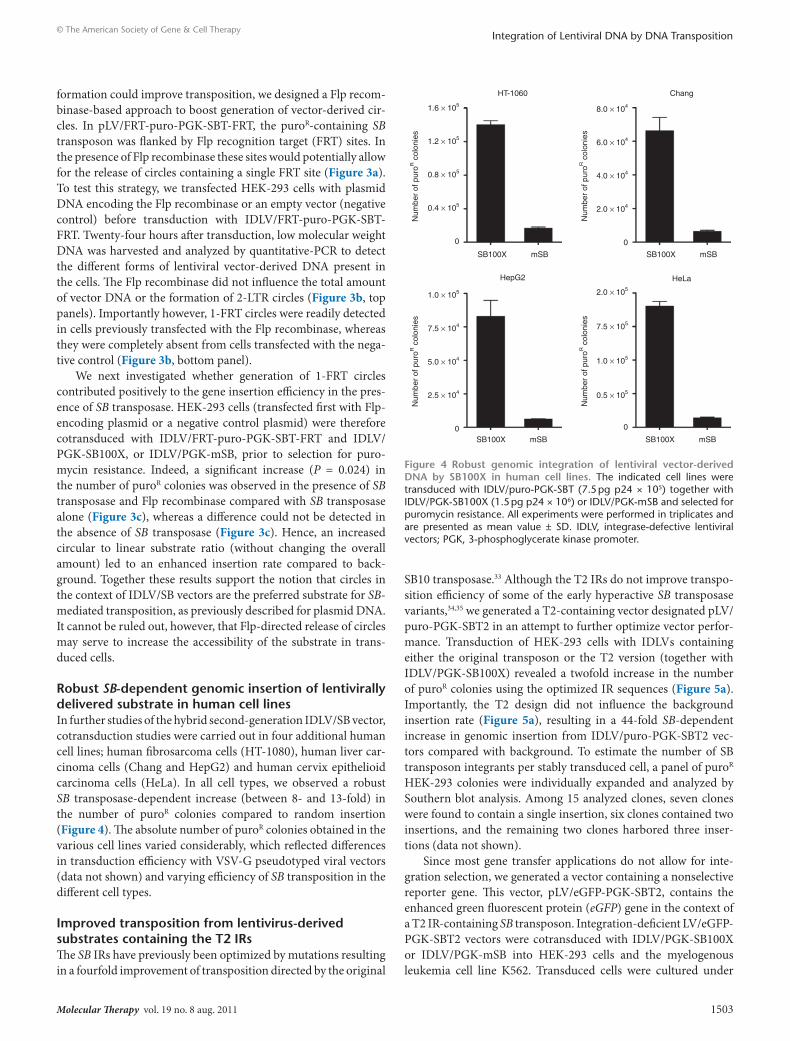
enhanced dnA circle formation directed by Flp recombinase in IdlV/sB-transduced cells enhances SB-mediated integrationThe structure of the second-generation vector allows for transpo-sition from both linear and circular substrates. However, earlier reports have suggested that SB transposition from linear substrates is highly reduced.30–32 To investigate whether enhanced circle
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0
+ FIp − FIp
Rel
ativ
e to
tal v
ecto
r D
NA
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0
+ FIp − FIp
Rel
ativ
e 2-
LTR
vec
tor
DN
A
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0
+ FIp − FIpR
elat
ive
Flp
circ
le D
NA
FRT
FRT
RIR
RIR
LIP
LIR
FRT
pA
pA
pGK
PG
K
puroR
puroR
FIp recombinase+
a
8.0 × 105
6.0 × 105
4.0 × 105
2.0 × 105
0
+ FIp − FIp
*
SB100XmSB
Num
ber
of p
uroR
col
onie
s
c
b
Figure 3 Sleeping Beauty (SB)-mediated insertion is enhanced by an increased ratio of circular to linear substrates. (a) Schematic representation of the strategy used to increase the circular DNA in transduced cells. The lentiviral vector, pLV/FRT-puro-PGK-SBT-FRT, contains a PGK-puro expres-sion cassette within a SB transposon, which is flanked by two Flp recombination target (FRT) sites. The FRT sites enable recombination (and thereby circle formation) in the presence of Flp recombinase. (b) FRT circle formation occurs exclusively in the presence of Flp recombinase. HEK-293 cells were transfected with pCMV-Flpe or pUC19 (as a negative control) before transduction with IDLV/FRT-puro-PGK-SBT-FRT (3.2 pg p24 × 105). Twenty-four hours after transduction HIRT DNA was harvested for quantitative-PCR analysis to determine the amount of total vector DNA, 2-LTR and 1-FRT circles. (c) The presence of Flp recombinase increases the number of puroR resistant colonies. HEK-293 cells were transfected with pCMV-Flpe or pUC19 before cotransduction with IDLV/FRT-puro-PGK-SBT-FRT (3.2 pg p24 × 105), and IDLV/PGK-SB100X or mSB (1.7 pg p24 × 106) and selected for puromycin resistance. All experiments were performed in triplicates and are presented as mean value ± SD. *Statistical significance, P = 0.024. CMV, cytomegalovirus; IDLV, integrase-defective lentiviral vectors; PGK, 3-phosphoglycerate kinase promoter.
Molecular Therapy vol. 19 no. 8 aug. 2011 1503
© The American Society of Gene & Cell TherapyIntegration of Lentiviral DNA by DNA Transposition
formation could improve transposition, we designed a Flp recom-binase-based approach to boost generation of vector-derived cir-cles. In pLV/FRT-puro-PGK-SBT-FRT, the puroR-containing SB transposon was flanked by Flp recognition target (FRT) sites. In the presence of Flp recombinase these sites would potentially allow for the release of circles containing a single FRT site (Figure 3a). To test this strategy, we transfected HEK-293 cells with plasmid DNA encoding the Flp recombinase or an empty vector (negative control) before transduction with IDLV/FRT-puro-PGK-SBT-FRT. Twenty-four hours after transduction, low molecular weight DNA was harvested and analyzed by quantitative-PCR to detect the different forms of lentiviral vector-derived DNA present in the cells. The Flp recombinase did not influence the total amount of vector DNA or the formation of 2-LTR circles (Figure 3b, top panels). Importantly however, 1-FRT circles were readily detected in cells previously transfected with the Flp recombinase, whereas they were completely absent from cells transfected with the nega-tive control (Figure 3b, bottom panel).
We next investigated whether generation of 1-FRT circles contributed positively to the gene insertion efficiency in the pres-ence of SB transposase. HEK-293 cells (transfected first with Flp-encoding plasmid or a negative control plasmid) were therefore cotransduced with IDLV/FRT-puro-PGK-SBT-FRT and IDLV/PGK-SB100X, or IDLV/PGK-mSB, prior to selection for puro-mycin resistance. Indeed, a significant increase (P = 0.024) in the number of puroR colonies was observed in the presence of SB transposase and Flp recombinase compared with SB transposase alone (Figure 3c), whereas a difference could not be detected in the absence of SB transposase (Figure 3c). Hence, an increased circular to linear substrate ratio (without changing the overall amount) led to an enhanced insertion rate compared to back-ground. Together these results support the notion that circles in the context of IDLV/SB vectors are the preferred substrate for SB-mediated transposition, as previously described for plasmid DNA. It cannot be ruled out, however, that Flp-directed release of circles may serve to increase the accessibility of the substrate in trans-duced cells.
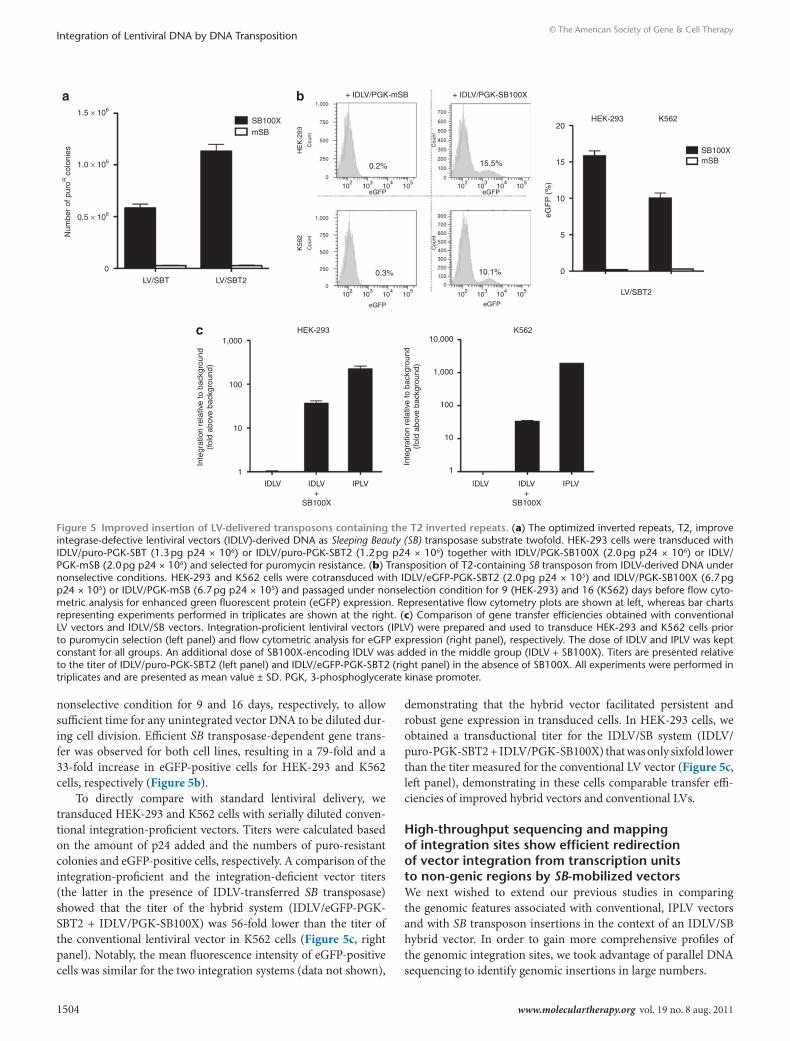
robust SB-dependent genomic insertion of lentivirally delivered substrate in human cell linesIn further studies of the hybrid second-generation IDLV/SB vector, cotransduction studies were carried out in four additional human cell lines; human fibrosarcoma cells (HT-1080), human liver car-cinoma cells (Chang and HepG2) and human cervix epithelioid carcinoma cells (HeLa). In all cell types, we observed a robust SB transposase-dependent increase (between 8- and 13-fold) in the number of puroR colonies compared to random insertion (Figure 4). The absolute number of puroR colonies obtained in the various cell lines varied considerably, which reflected differences in transduction efficiency with VSV-G pseudotyped viral vectors (data not shown) and varying efficiency of SB transposition in the different cell types.
Improved transposition from lentivirus-derived substrates containing the t2 IrsThe SB IRs have previously been optimized by mutations resulting in a fourfold improvement of transposition directed by the original
SB10 transposase.33 Although the T2 IRs do not improve transpo-sition efficiency of some of the early hyperactive SB transposase variants,34,35 we generated a T2-containing vector designated pLV/puro-PGK-SBT2 in an attempt to further optimize vector perfor-mance. Transduction of HEK-293 cells with IDLVs containing either the original transposon or the T2 version (together with IDLV/PGK-SB100X) revealed a twofold increase in the number of puroR colonies using the optimized IR sequences (Figure 5a). Importantly, the T2 design did not influence the background insertion rate (Figure 5a), resulting in a 44-fold SB-dependent increase in genomic insertion from IDLV/puro-PGK-SBT2 vec-tors compared with background. To estimate the number of SB transposon integrants per stably transduced cell, a panel of puroR HEK-293 colonies were individually expanded and analyzed by Southern blot analysis. Among 15 analyzed clones, seven clones were found to contain a single insertion, six clones contained two insertions, and the remaining two clones harbored three inser-tions (data not shown).
Since most gene transfer applications do not allow for inte-gration selection, we generated a vector containing a nonselective reporter gene. This vector, pLV/eGFP-PGK-SBT2, contains the enhanced green fluorescent protein (eGFP) gene in the context of a T2 IR-containing SB transposon. Integration-deficient LV/eGFP-PGK-SBT2 vectors were cotransduced with IDLV/PGK-SB100X or IDLV/PGK-mSB into HEK-293 cells and the myelogenous leukemia cell line K562. Transduced cells were cultured under
1.0 × 105
7.5 × 104
5.0 × 104
2.5 × 104
0
2.0 × 105
7.5 × 105
1.0 × 105
0.5 × 105
0
Num
ber
of p
uroR
col
onie
s
Num
ber
of p
uroR
col
onie
s
1.6 × 105
1.2 × 105
0.8 × 105
0.4 × 105
0
Num
ber
of p
uroR
col
onie
s
HT-1060
SB100X mSB
SB100X mSB SB100X mSB
8.0 × 104
6.0 × 104
4.0 × 104
2.0 × 104
0
Num
ber
of p
uroR
col
onie
s
Chang
SB100X mSB
HepG2 HeLa
Figure 4 robust genomic integration of lentiviral vector-derived dnA by sB100X in human cell lines. The indicated cell lines were transduced with IDLV/puro-PGK-SBT (7.5 pg p24 × 105) together with IDLV/PGK-SB100X (1.5 pg p24 × 106) or IDLV/PGK-mSB and selected for puromycin resistance. All experiments were performed in triplicates and are presented as mean value ± SD. IDLV, integrase-defective lentiviral vectors; PGK, 3-phosphoglycerate kinase promoter.
1504 www.moleculartherapy.org vol. 19 no. 8 aug. 2011
© The American Society of Gene & Cell TherapyIntegration of Lentiviral DNA by DNA Transposition
nonselective condition for 9 and 16 days, respectively, to allow sufficient time for any unintegrated vector DNA to be diluted dur-ing cell division. Efficient SB transposase-dependent gene trans-fer was observed for both cell lines, resulting in a 79-fold and a 33-fold increase in eGFP-positive cells for HEK-293 and K562 cells, respectively (Figure 5b).
To directly compare with standard lentiviral delivery, we transduced HEK-293 and K562 cells with serially diluted conven-tional integration-proficient vectors. Titers were calculated based on the amount of p24 added and the numbers of puro-resistant colonies and eGFP-positive cells, respectively. A comparison of the integration-proficient and the integration-deficient vector titers (the latter in the presence of IDLV-transferred SB transposase) showed that the titer of the hybrid system (IDLV/eGFP-PGK-SBT2 + IDLV/PGK-SB100X) was 56-fold lower than the titer of the conventional lentiviral vector in K562 cells (Figure 5c, right panel). Notably, the mean fluorescence intensity of eGFP-positive cells was similar for the two integration systems (data not shown),
demonstrating that the hybrid vector facilitated persistent and robust gene expression in transduced cells. In HEK-293 cells, we obtained a transductional titer for the IDLV/SB system (IDLV/puro-PGK-SBT2 + IDLV/PGK-SB100X) that was only sixfold lower than the titer measured for the conventional LV vector (Figure 5c, left panel), demonstrating in these cells comparable transfer effi-ciencies of improved hybrid vectors and conventional LVs.
High-throughput sequencing and mapping of integration sites show efficient redirection of vector integration from transcription units to non-genic regions by SB-mobilized vectorsWe next wished to extend our previous studies in comparing the genomic features associated with conventional, IPLV vectors and with SB transposon insertions in the context of an IDLV/SB hybrid vector. In order to gain more comprehensive profiles of the genomic integration sites, we took advantage of parallel DNA sequencing to identify genomic insertions in large numbers.
1.5 × 106
1.0 × 106
0.5 × 106
0
LV/SBT LV/SBT2
Num
ber
of p
uroR
col
onie
s
SB100XmSB
1,000
750
500
250
0
Cou
nt
Cou
nt
HE
K-2
93
1,000
750
500
250
0
Cou
nt
K56
210
210
310
410
5
eGFP
102
103
104
105
eGFP
102
103
104
105
eGFP10
210
310
410
5
eGFP
0.3% 10.1%
700
600
500
400
300
200
100
0
Cou
nt
700
900
600
500
400
300
200
100
0
0.2% 15.5%
+ IDLV/PGK-mSB + IDLV/PGK-SB100X
HEK-293 K56220
15
10
5
0
eGF
P (
%)
LV/SBT2
SB100XmSB
a b
HEK-293 K5621,000
100
10
1
IDLV IDLV+
SB100X
IPLV IDLV IDLV+
SB100X
IPLV
Inte
grat
ion
rela
tive
to b
ackg
roun
d(f
old
abov
e ba
ckgr
ound
)
10,000
1,000
100
10
1
Inte
grat
ion
rela
tive
to b
ackg
roun
d(f
old
abov
e ba
ckgr
ound
)
c
Figure 5 Improved insertion of lV-delivered transposons containing the t2 inverted repeats. (a) The optimized inverted repeats, T2, improve integrase-defective lentiviral vectors (IDLV)-derived DNA as Sleeping Beauty (SB) transposase substrate twofold. HEK-293 cells were transduced with IDLV/puro-PGK-SBT (1.3 pg p24 × 106) or IDLV/puro-PGK-SBT2 (1.2 pg p24 × 106) together with IDLV/PGK-SB100X (2.0 pg p24 × 106) or IDLV/PGK-mSB (2.0 pg p24 × 106) and selected for puromycin resistance. (b) Transposition of T2-containing SB transposon from IDLV-derived DNA under nonselective conditions. HEK-293 and K562 cells were cotransduced with IDLV/eGFP-PGK-SBT2 (2.0 pg p24 × 105) and IDLV/PGK-SB100X (6.7 pg p24 × 105) or IDLV/PGK-mSB (6.7 pg p24 × 105) and passaged under nonselection condition for 9 (HEK-293) and 16 (K562) days before flow cyto-metric analysis for enhanced green fluorescent protein (eGFP) expression. Representative flow cytometry plots are shown at left, whereas bar charts representing experiments performed in triplicates are shown at the right. (c) Comparison of gene transfer efficiencies obtained with conventional LV vectors and IDLV/SB vectors. Integration-proficient lentiviral vectors (IPLV) were prepared and used to transduce HEK-293 and K562 cells prior to puromycin selection (left panel) and flow cytometric analysis for eGFP expression (right panel), respectively. The dose of IDLV and IPLV was kept constant for all groups. An additional dose of SB100X-encoding IDLV was added in the middle group (IDLV + SB100X). Titers are presented relative to the titer of IDLV/puro-PGK-SBT2 (left panel) and IDLV/eGFP-PGK-SBT2 (right panel) in the absence of SB100X. All experiments were performed in triplicates and are presented as mean value ± SD. PGK, 3-phosphoglycerate kinase promoter.
Molecular Therapy vol. 19 no. 8 aug. 2011 1505
© The American Society of Gene & Cell TherapyIntegration of Lentiviral DNA by DNA Transposition
K562 cells were transduced with IDLV/puro-PGK-SBT2 together with IDLV/PGK-SB100X, or with the integration- proficient HIV vector (IPLV/puro-PGK-SBT2). To exclude possi-ble biases introduced by the nonrandom distribution of restriction enzyme recognition sites in the chromosomes, genomic DNA samples isolated from pooled puromycin-resistant cells were ran-domly fragmented by sonication. Sequences neighboring the inser-tion sites were amplified by linear amplification-mediated PCR36 using transposon IR-, or LTR-specific primers, and the resulting libraries were sequenced on an Illumina Genome Analyzer plat-form. Mapping the sequence reads on the human genome resulted in 1,283 unique IDLV/SB transposon integration sites and 1,165 uniquely mapped IPLV integration sites (Table 1 and Figure 6).
Random control sets of 10,000 loci were generated computation-ally, which were used as control datasets for the analysis.
Lentiviruses are known to prefer active transcription units for integration with a frequency of 70–80%. Previous studies showed no preference of SB for integration into genes in HeLa cells,37 and transposon insertions were very weakly associated with RefSeq genes in Huh-7 cells.19 In recent studies of transposition by the SB11 and SB100X hyperactive transposase versions from IDLV/SB hybrid vectors in HeLa cells, 53 and 30%, respectively, of the integrations were mapped within RefSeq genes.16,28 Analysis of our datasets revealed that 47% of the transposon insertions occurred within RefSeq genes, whereas 74% of lentiviral integrants were identified in genes (Table 1). Since the expected rate for random
chr1
chr2
chr3
chr4
chr5
chr6
chr7
chr8
chr9
chr10
chr11
chrX
chr22
chr21
chr20
chr19
chr18
chr17
chr16
chr15
chr14
chr13
chr12
LV
IDLV/SB
Figure 6 distribution of the lV and IdlV/sB integrations on human chromosomes. Mapped integrations of the datasets are depicted as triangles above (LV, blue) and under (IDLV/SB, red) the symbolic human chromosomes. The yellow chromosomal segments depict chromosomal regions that could not with certainty be assembled to the complete genome assembly. IDLV, integrase-defective lentiviral vectors.
table 1 comparison of integration profiles of conventional lentiviral vectors and IdlV/sB hybrid vectors in vector-transduced K562 cells
conventional lentiviral vector lenti-transposon hybrid vector
lV random seta (lV) IdlV/sB random seta (SB )
number of sites Frequency Frequency number of sites Frequency Frequency
RefSeq genes 863 0.74 0.41 610 0.48 0.40
RefSeq exons 47 0.040 0.025 25 0.019 0.018
RefSeq introns 817 0.70 0.39 588 0.46 0.38
RefSeq intergenic 302 0.26 0.59 673 0.52 0.60
2 kb upstream of RefSeq gene 9 0.0077 0.015 18 0.014 0.0094
2 kb downstream of RefSeq gene 21 0.018 0.018 27 0.021 0.011
±1 kb from CpG islands 14 0.012 0.025 22 0.017 0.012
Abbreviations: IDLV, Integrase-defective lentiviral vectors; SB, Sleeping Beauty.aRandom sets for the LV and IDLV/SB datasets were created as described in Materials and Methods section.
1506 www.moleculartherapy.org vol. 19 no. 8 aug. 2011
© The American Society of Gene & Cell TherapyIntegration of Lentiviral DNA by DNA Transposition
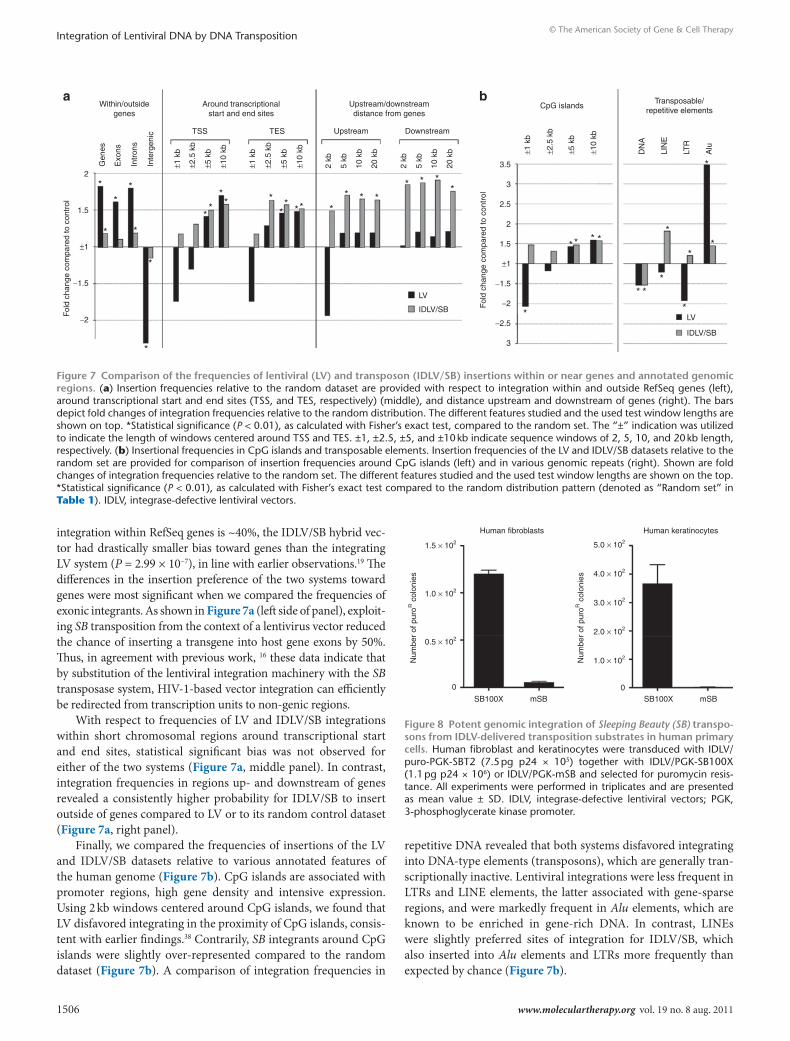
integration within RefSeq genes is ~40%, the IDLV/SB hybrid vec-tor had drastically smaller bias toward genes than the integrating LV system (P = 2.99 × 10−7), in line with earlier observations.19 The differences in the insertion preference of the two systems toward genes were most significant when we compared the frequencies of exonic integrants. As shown in Figure 7a (left side of panel), exploit-ing SB transposition from the context of a lentivirus vector reduced the chance of inserting a transgene into host gene exons by 50%. Thus, in agreement with previous work, 16 these data indicate that by substitution of the lentiviral integration machinery with the SB transposase system, HIV-1-based vector integration can efficiently be redirected from transcription units to non-genic regions.
With respect to frequencies of LV and IDLV/SB integrations within short chromosomal regions around transcriptional start and end sites, statistical significant bias was not observed for either of the two systems (Figure 7a, middle panel). In contrast, integration frequencies in regions up- and downstream of genes revealed a consistently higher probability for IDLV/SB to insert outside of genes compared to LV or to its random control dataset (Figure 7a, right panel).
Finally, we compared the frequencies of insertions of the LV and IDLV/SB datasets relative to various annotated features of the human genome (Figure 7b). CpG islands are associated with promoter regions, high gene density and intensive expression. Using 2 kb windows centered around CpG islands, we found that LV disfavored integrating in the proximity of CpG islands, consis-tent with earlier findings.38 Contrarily, SB integrants around CpG islands were slightly over-represented compared to the random dataset (Figure 7b). A comparison of integration frequencies in
repetitive DNA revealed that both systems disfavored integrating into DNA-type elements (transposons), which are generally tran-scriptionally inactive. Lentiviral integrations were less frequent in LTRs and LINE elements, the latter associated with gene-sparse regions, and were markedly frequent in Alu elements, which are known to be enriched in gene-rich DNA. In contrast, LINEs were slightly preferred sites of integration for IDLV/SB, which also inserted into Alu elements and LTRs more frequently than expected by chance (Figure 7b).
Gen
es
Exo
ns
Intr
ons
Inte
rgen
ic
Within/outsidegenes
Around transcriptionalstart and end sites
Upstream/downstreamdistance from genes
DownstreamUpstreamTSS TES
±1 k
b
±2.5
kb
±5 k
b
±10
kb
±1 k
b
±2.5
kb
±5 k
b
±10
kb
2 kb
5 kb
10 k
b
20 k
b
2 kb
5 kb
10 k
b
20 k
b
*
*
*
*
*
*
**
** *
**
** ** * *
* * **
*
LV
IDLV/SB
2
1.5
±1
−1.5
−2
Fol
d ch
ange
com
pare
d to
con
trol
3.5
3
2.5
2
1.5
±1
−1.5
−2
−2.5
3
±1 k
b
±2.5
kb
±5 k
b
±10
kb
DN
A
LIN
E
LTR
Alu
LV
IDLV/SB
Fol
d ch
ange
com
pare
d to
con
trol
*
** * *
* **
*
*
*
*
*
CpG islandsTransposable/
repetitive elements
a b
Figure 7 comparison of the frequencies of lentiviral (lV) and transposon (IdlV/sB) insertions within or near genes and annotated genomic regions. (a) Insertion frequencies relative to the random dataset are provided with respect to integration within and outside RefSeq genes (left), around transcriptional start and end sites (TSS, and TES, respectively) (middle), and distance upstream and downstream of genes (right). The bars depict fold changes of integration frequencies relative to the random distribution. The different features studied and the used test window lengths are shown on top. *Statistical significance (P < 0.01), as calculated with Fisher’s exact test, compared to the random set. The “±” indication was utilized to indicate the length of windows centered around TSS and TES. ±1, ±2.5, ±5, and ±10 kb indicate sequence windows of 2, 5, 10, and 20 kb length, respectively. (b) Insertional frequencies in CpG islands and transposable elements. Insertion frequencies of the LV and IDLV/SB datasets relative to the random set are provided for comparison of insertion frequencies around CpG islands (left) and in various genomic repeats (right). Shown are fold changes of integration frequencies relative to the random set. The different features studied and the used test window lengths are shown on the top. *Statistical significance (P < 0.01), as calculated with Fisher’s exact test compared to the random distribution pattern (denoted as “Random set” in Table 1). IDLV, integrase-defective lentiviral vectors.
Human fibroblasts Human keratinocytes
1.5 × 102 5.0 × 102
4.0 × 102
3.0 × 102
2.0 × 102
1.0 × 102
0
0.5 × 102
0
1.0 × 102
SB100X mSB SB100X mSB
Num
ber
of p
uroR
col
onie
s
Num
ber
of p
uroR
col
onie
s
Figure 8 Potent genomic integration of Sleeping Beauty (SB) transpo-sons from IdlV-delivered transposition substrates in human primary cells. Human fibroblast and keratinocytes were transduced with IDLV/puro-PGK-SBT2 (7.5 pg p24 × 105) together with IDLV/PGK-SB100X (1.1 pg p24 × 106) or IDLV/PGK-mSB and selected for puromycin resis-tance. All experiments were performed in triplicates and are presented as mean value ± SD. IDLV, integrase-defective lentiviral vectors; PGK, 3-phosphoglycerate kinase promoter.

Molecular Therapy vol. 19 no. 8 aug. 2011 1507
© The American Society of Gene & Cell TherapyIntegration of Lentiviral DNA by DNA Transposition
Potent gene insertion by IdlV/sB vectors in human primary cellsWe finally assessed the gene transfer efficacies of IDLV/SB hybrid vectors in human primary cells. Primary human fibroblasts and keratinocytes were cotransduced with IDLV/puro-PGK-SBT2 and IDLV/PGK-SB100X (or IDLV/PGK-mSB) and then, 1 day later, sub-jected to puromycin selection. The number of puromycin-resistant colonies obtained in human primary fibroblasts and keratinocytes were 23- and 160-folds higher, respectively, in the presence of SB transposase compared with background level observed in the pres-ence of mSB (Figure 8). In summary, our findings demonstrate that improved second-generation IDLV/SB vectors are efficiently redi-rected from transcription units to non-genic regions and possess a high transfer potential in human primary cells.
dIscussIonThe efficacy of gene insertion is a pivotal factor for successful long-term expression in dividing cells and tissues. Lentiviral vectors efficiently pass both the cellular- and nuclear membrane before integrating their genetic cargo into genomic DNA. The integra-tion profiles of lentiviruses and derived vectors have been exten-sively studied, and have raised some concerns due to a preference of insertion into transcriptional units.1–3
Several hybrid vector systems have tried to overcome the dis-advantages and combine the best features of existing vectors.39–44 Many of these new vector technologies were recently reviewed.45 In one of the more successful approaches, efficient genetic engi-neering was achieved by IDLV-based delivery of gene cassettes encoding zinc-finger nucleases.13 We previously described a HIV-1/SB hybrid vector that showed no bias toward integration in transcriptional units.16 This vector was reliant on viral DNA circle formation prior to SB transposition. In the present report, we explored a second-generation vector design that allows trans-position from both linear and circular viral DNA intermediates. Although the transfer efficiency of the second-generation vector was reduced relative to the first-generation design, an increased transposition activity resulted in a hybrid vector with improved transduction efficiency, as measured by an increased divergence of transduction titers obtained in the presence and absence of SB100X. During lentiviral transduction, episomal linear forms of the lentiviral vector genome are in excess and, hence, poten-tially accessible for the cellular transcriptional machinery.8,11 The availability of both linear and circular substrates during transfer of the second-generation hybrid vector may explain the improved performance of this vector. Alternatively, an increased transposition rate could reflect that the transposon carrying the transgene is shorter in the second-generation design relative to the original hybrid vector. Notably, a previous report describing a related vector design (with the transposon placed in the for-ward orientation) did not succeed in demonstrating vector effi-cacy using SB11 transposase,28 and to our knowledge, LVs have not previously been combined with other integration machiner-ies than SB.
The SB transposon has earlier been paired with both adeno-viral vectors and herpes simplex virus.39,40,44 However, the integra-tion preferences have not been determined for the adenovirus/SB or HSV/SB hybrid vectors as would be required for a direct
comparison with HIV-1/SB vectors. The adenovirus/SB hybrid vector showed effective delivery and integration in mouse liver. The function of this vector, however, relied on a DNA circulariza-tion step mediated by the Flp recombinase in transduced cells.39 Notably, circularization of the lentiviral vector genome occurs without the need of an extra recombinase and indeed circular transposition substrates may be the preferred donor of transpo-sons in the context of second-generation IDLV/SB hybrid vectors. In support of this notion, previous reports have described that cir-cular substrates are preferred by SB transposases.30–32 Moreover, we found that the IDLV/SB vector integration efficacy was enhanced by increasing the amount of DNA circles (by Flp recombination). However, the occurrence of transposition from linear substrates cannot be excluded at this stage. Moreover, it cannot be ruled out that Flp-directed release of circular substrates from the viral preintegration complex may serve to increase accessibility of the DNA transposon substrate.
Transposition of the SB T2 transposon from IDLV substrates increased the level of genomic integration 44-fold above back-ground in HEK-293 cells and at most 160-fold above background in primary human keratinocytes. Such potency could be obtained only with hyperactive SB100X transposase which is under certain experimental conditions 100 times more active than the original SB10 transposase.27 Hence, it appears that this transposase is par-ticularly potent when the source of transposon is limited or, alter-natively, when the amount of transposase is limited. Although we did not directly compare the activity of the original SB10 trans-posase with the hyperactive SB100X in the context of the IDLV/SB vector, our findings suggest that SB100X in the context of IDLV/SB vectors does not improve transposition with 100-fold relative to the original SB transposase, as has been shown in plasmid-based experiments. Hence, by comparing vectors integrated by the inactive mSB and mSB100X, we observed titer differences ranging from an eightfold improvement in HT-1080 cells to a 160-fold improvement in primary human keratinocytes. As opposed to plasmid transfection experiments carried out under optimized experimental conditions, we believe that the scarcity of both transposase and transposon sources after lentiviral delivery may confine the overall effect of SB100X in the IDLV/SB context. This notion is supported by the finding that both the lentiviral vector-derived substrate and the SB transposase are limiting factors for transposition from IDLVs. In the course of our studies, we also developed hybrid IDLV vectors containing the transgene cassette flanked by the recognition site for PhiC31 integrase.46 However, IDLV-directed expression of the PhiC31 integrase did not result in detectable levels of gene insertion from IDLV-derived substrates. At present, we favor the hypothesis that the ability of SB100X to direct transposition from a very scarce transposon source is para-mount to the functionality of the system.
Copy number analysis indicated that the majority of cells that were successfully transduced with the IDLV/SB vector carried only a single or two insertions of the vector, suggesting that insertion of the vector was accompanied by a minimal risk of genotoxicity. This is a potential advantageous feature in gene therapy applications. We performed a large-scale, genome-wide analysis using parallel DNA sequencing to assess the integration site selection properties of the improved IDLV/SB hybrid vector in K562 cells. The dataset,

1508 www.moleculartherapy.org vol. 19 no. 8 aug. 2011
© The American Society of Gene & Cell TherapyIntegration of Lentiviral DNA by DNA Transposition
considerably larger than any previously available, allowed us to apply a statistically robust study of the integration profiles and a variety of annotated genomic features associated with SB integra-tion sites. Most importantly, the transposon insertions launched from the hybrid IDLV/SB vector showed drastically smaller bias toward genes than the LV integrants. If the bias toward RefSeq genes is expressed as the ratio of the frequencies of genic inser-tions above random control, the factors obtained in this study for LV and IDLV/SB were 1.85 and 1.17, respectively, where factor 1 means no bias. Our results obtained with the IDLV/SB system very much resemble those of previous studies using plasmid-based SB vectors in human HeLa and Huh7 cells, where the bias ratios of SB integrants for RefSeq genes were 1.24 and 1.17, respectively.19,37 These results may indicate that launching SB transposons from LVs does not introduce a further bias into the integration profile of SB toward genes.
In summary, the bipartite hybrid vector described here pro-vides a novel integration tool almost as efficient as conventional LVs but with an altered genomic integration profile. Also, our system combining an optimized vector design with a hyperactive transposase appears to facilitate gene transfer and genomic inser-tion of the transgene with much higher efficiency than previously seen.16,28 By intuition, this vector system is safer than conventional LVs and less likely to cause genotoxicity, but so far this attractive feature is hypothetical and remains to be tested in an appropriate tumor-prone animal model. Safe and efficient vectors are crucial in therapeutic gene transfer. Future applications may incorporate technologies for targeted gene insertion into predetermined safe loci. The lentiviral preintegration complex appears to be highly sensitive to alteration47 and other strategies may be necessary to improve the targeting of LVs. The SB transposase has with some success been fused to the E2C zinc-finger DNA binding protein for targeted integration.48 The most efficient approach to date has been fusing the targeting protein with a transposase fragment. The fusion protein interacts with full length transposase and has resulted in targeted integration of almost 10% of all insertions.49 The strategy seems to be readily accessible and may be incorpo-rated into the IDLV/SB vector system for future testing. We believe that, based on its properties reported here, the newly developed hybrid vector system can become a useful tool in genetic engineer-ing, animal transgenesis and preclinical therapeutic gene transfer.
MAterIAls And MetHodsVector construction. The HIV-1-derived SB substrate vector pLV/puro-PGK-SBT was generated by inserting a SB transposon containing a puro-mycin resistance gene cassette (PCR-amplified from pT/PGK-puro)31 into the pLV/MCS (digested with MluI/AscI). To create the pLV/MCS, which is a HIV-1-based lentiviral vector containing five unique restriction sites between the central polypurine tract and the woodchuck hepatitis virus post-transcriptional regulatory element, we digested the pCCL.WPS.PGK-Puro.WHV15 with ClaI and XhoI, removing the PGK-puro cassette and inserted annealed oligonucleotides containing the multiple cloning site. pLV/RIR-LIR.PGK-puro and pLV/PGK-SB100X were previously described.16 The HIV-1/FRT/SB hybrid vector, pLV/FRT-puro-PGK-SBT-FRT, was constructed by inserting a SB transposon containing a puromycin resistance gene (PCR-amplified from pT/PGK-puro) flanked by FRT sites (included in the primer sequences) into MluI-/AscI-digested pLV/MCS. The T2 version of the HIV-1/SB hybrid vector, pLV/puro-PGK-SBT2, was
constructed by exchanging RIR (BamHI) and LIR (AscI and PacI) with PCR-amplified T2-RIR and T2-LIR (from pT2/SV40-neo).33 To generate the eGFP-expressing T2 hybrid vector, pLV/eGFP-PGK-SBT2, the PGK-puro fragment was removed from pLV/puro-PGK-SBT2 by ClaI/PacI digestion followed by insertion of a PGK-eGFP fragment amplified from pCCL.WPS.PGK.eGFP.WHS (a kind gift from Dr Aebischer, Swiss Federal Institute of Technology, EPFL, Lausanne, Switzerland). The lentiviral vec-tor expressing SB100X, pLV/CMV-SB100X, was created by exchanging the RSV promoter (ApaI/EcoRI) with the CMV promoter PCR-amplified from pEGFP.N1 (Clontech, Mountain View, CA). The lentiviral vector contain-ing an RSV-driven SB100X expression cassette was generated by amplify-ing the RSV promoter from pT/hAAT25 and the SB100X gene from pLV/PGK-SB100X. The two fragments were linked together by overlap PCR and inserted into the pLV/MCS plasmid digested with ApaI and XhoI.
Cell culture. HEK-293, 293T, HeLa, and HT-1080 were maintained in Dulbecco’s modified Eagle’s medium (Cambrex, Verviers, Belgium). K562, Chang, Hep2G, and human primary fibroblasts were maintained in RPMI (Cambrex) all supplemented with 10% fetal calf serum, 100 U/ml penicillin, 0.1 mg/ml streptomycin, and 265 mg/l l-glutamine. Human primary kera-tinocytes were grown in Epidermal Keratinocyte Medium (CELLnTEC, Bern, Switzerland) supplemented with 100 U/ml penicillin and 0.1 mg/ml streptomycin. When selection was applied a final concentrations of 1 µg/ml (cell lines) or 3 µg/ml (primary cells) puromycin (Sigma, St Louis, MI) were used. All cells were cultured at 37 °C and 5% (vol/vol) CO2.
Lentiviral production. Integration-deficient LVs pseudotyped with VSV-G were produced by CaPO4-transfection of 293T cells (seeded at 3 × 106 cells/dish in 10 cm dishes) with 3 μg pRSV-Rev, 3.75 μg pMD.2G (VSV-G), 13 μg pMDLg/pRREintD64V (or pMDLg/pRRE for production of integration-proficient vectors), and 13 μg lentiviral vector plasmid. The supernatant was harvested 2 days post-transfection and polybrene was added to a final concentration of 8 µg/ml prior to transfer to target cells.
Transduction of cell lines and primary cells. All cell lines (except K562) were seeded (1.5 × 106 cells/dish in 10 cm dishes) 1 day prior to transduc-tion. Human primary fibroblast and keratinocytes were seeded in six-well plates at a density 2 × 105 cells/well. One day post-transduction the cells were diluted and reseeded and 2 days later subjected to puromycin selec-tion. After 10 days of selection the resistant colonies were stained with methylene blue and counted. In experiments based on eGFP expression HEK-293 and K562 cells were seeded at 1.5 × 106 cells/dish in 10 cm dishes and 5 × 105 cells/well, respectively, in six-well plates 1 day prior to trans-duction. The cells were passaged for 9 (HEK-293) or 16 days (K562) before flow analysis for green fluorescence using a FACS Aria sorter and FACS DiVa 4.1 software (BD Biosciences, Franklin Lakes, NJ).
For titer determination of IPLVs HEK-293 cells were seeded in six-well dishes (2 × 105 cells/well) 1 day prior to transduction of serially diluted LVs. One day post-transduction puromycin selection was applied for 1 week before staining with methylene blue and counting. Viral titers were calculated by dividing the number of colonies with the dilution factor. All experiments were performed in triplicates.
Southern and western blot analysis. Southern blot analysis of puro-mycin-resistant clones of HEK-293 cells cotransduced with IDLV/puro-PGK-SBT2 and IDLV/PGK-SB100X was performed as follows. Fifteen micrograms of genomic DNA from each clone was digested overnight with AvrII (which cuts once inside the transposon vector) prior to gel electrophoresis and vacuum blotting. Random labeling of the puro probe (using a PCR-amplified labeling template) was carried out by using the Prime-It random primer labeling kit (Agilent Technologies, Santa Clara, CA) according to the manufacturer’s instructions. Hybridization was per-formed at 42 °C, whereas all washing procedures were done at 50 °C. For western blot analysis, protein was harvested from HEK-293 cells cotrans-duced with 4.4 × 105 pg p24 of IDLV/puro-PGK-SBT and 10–110 × 105

Molecular Therapy vol. 19 no. 8 aug. 2011 1509
© The American Society of Gene & Cell TherapyIntegration of Lentiviral DNA by DNA Transposition
IDLV/CMV-SB100X and 10–110 × 105 pg p24 IDLV/MCS to obtain equal viral load. Ten micrograms protein extract was separated by 12% sodium dodecyl sulfate/polyacrylamide gel electrophoresis and then electrotrans-ferred to a nitrocellulose membrane. Membranes were blocked with buf-fer containing 5% fat-free milk. After blocking and wash the membranes were incubated with polyclonal SB antigen (R&D Systems, Minneapolis, MN) for 2 hours. After a second wash the membranes were incubated with peroxidase-conjugated anti-rabbit secondary antibodies and chemilumi-nescence developed with ECL Plus Western Blotting Detection System detection kit (Amersham, Buckinghamshire, UK).
Quantitative-PCR and p24 measurement. Low molecular weight DNA was harvested by Hirt extraction 24 hours post-transduction. The relative amount of total HIV DNA, 2-LTR, and FRT circles was quantified by real time-PCR performed on an iCycler Thermal Cycler (Bio-Rad, Hercules, CA) using the DyNAmo HS SYBR Green qPCR Kit (Finnzymes, Espoo, Finland). Total HIV DNA, 2-LTR, and FRT circles were quantified by amplifying a part of the woodchuck hepatitis virus (WHV) post-transcrip-tional regulatory element present in the vector backbone, using primers spanning the LTR-LTR or FRT junction, respectively. Concentrations of p24 Gag were measured using a HIV-1 p24 ELISA kit (ZeptoMetrix, Buffalo, NY) following the manufacturer’s instructions.
Statistical analysis. Data from the Flp recombinase assay was analyzed for significance (P < 0.05) using the Mann–Whitney test.
Construction of integration site libraries. K562 cells were transduced with pLV/CMV-SB100X and IDLV/puro-PGK-SBT2 or with the IPLV, as described above. Genomic DNA from pools of puromycin-resistant colonies was extracted using the DNeasy kit (Qiagen, Valencia, CA). Three micro-gram of DNA was sonicated to small pieces ranging from 100 to 500 bp, with an average size of 250 bp using the Covaris S2 sonication device (see Supplementary Materials and Methods for settings). The sonicated DNA was ethanol-precipitated and 500 ng was used for linear amplification-mediated PCR.36 Briefly, biotinylated LTR-, and SB transposon IR-specific primers were used in 50 rounds of linear amplification to enrich DNA spe-cies containing vector-chromosomal DNA junctions. The single-stranded products were immobilized on streptavidin-coated magnetic beads using the Dynabeads kilobase BINDER kit (Invitrogen, Carlsbad, CA). Second strand synthesis was performed with random hexamer oligos (Roche, Basel, Switzerland) using Klenow fragment (New England Biolabs, Ipswich, MA). The free ends of the double-stranded DNA were blunt-ended and phospho-rylated using the End-It DNA End-Repair kit (Epicentre, Madison, WI). Klenow fragment exo- (New England Biolabs) and dATP were used to add a single “A” nucleotide to the 3′-ends. The captured DNA was then ligated to linkers with a “T” overhang. To avoid later amplification of the lentiviral sequences from the 3′-LTR, samples were digested with NarI, which has a recognition site just upstream of the 3′-LTR within the lentiviral vector. The NarI digest was performed also on the IDLV/SB samples to avoid the introduction of a possible bias. Next, the bead-bound DNA was subjected to nested PCRs using primers specific for the LTR or IR sequences and for the linker. During amplification, we used bar-coded primers so that we could pool different libraries in the subsequent steps. Finally, primers correspond-ing to Illumina adapter sequences were used to yield a directional library, in which sequences complementary to the Illumina genomic DNA sequences primer were located upstream to the transposon IRs or the lentiviral LTRs. Thus, the resulting libraries could be pooled and sequenced on a single flow cell lane on the Illumina Genome Analyzer platform with single end run settings. The Supplementary Materials and Methods provide specific details about the protocol, including primer and linker sequences as well as cycling conditions.
Computational analysis. We used a set of error-correcting barcodes for dis-tinguishing different data sets pooled into a single flow cell lane. Only reads starting exactly with the barcode (4 bp) followed by the adapter sequence
(24 bp for LV and 23 bp for SB) were used for the subsequent analysis. The remaining part of the reads containing putative genomic sequences had length 48/49 bp (replicate 1) or were cut after 50 bases (replicate 2). We mapped these sequences using Bowtie50 to the chromosomes of the human genome (NCBI built GRCh37/hg19, February 2009) excluding the Y-chromosome because K562 is a female cell line. We used only exact matches occurring uniquely in the reference, except for the analysis of repetitive regions for which we generated a second set of sites also containing reads with multiple exact matches. Reads mapped to exactly the same strand and genomic posi-tion were merged to a single site. To improve the data quality, we discarded all sites supported by <10 reads. In case of the IDLV/SB data set, we only accepted sites having the ubiquitous TA sequence adjacent to the integration site. The two technical replicates yielded very similar results and were there-fore pooled. For statistical analysis, we generated random control sets, each of 10,000 sites, as follows. We randomly selected 50-mers from the reference genome and applied Bowtie for mapping them back to the genome using the same settings as described above. We discarded all sites in the IDLV/SB control set that did not start at a TA sequence.
suPPleMentArY MAterIAlMaterials and Methods.
AcKnoWledGMentsThe authors thank Thomas Juhl Corydon and Uffe Birk Jensen (Department of Human Genetics, University of Aarhus, Denmark) for kindly providing human primary fibroblasts and keratinocytes, respec-tively. This work was made possible through the generous support by the Danish Medical Research Council, the Novo Nordisk Foundation, the Carlsberg Foundation, the Danish Cancer Society, Aage Bangs Foundation, the Augustinus Foundation, The Foundation of 17-12-1981, the Lundbeck Foundation, the Danish National Advanced Technology Foundation, the Bundesministerium für Bildung und Forschung (NGFNplus—ENGINE), and the EU (EU-FP6-STREP, contract number 018961). N.S., N.H.S, and B.M were funded by grants from the Novo Scholarship Programme in Biotechnology and Pharmaceutical Sciences, the Danish National Advanced Technology Foundation, and the Danish Cancer Society, respectively. The authors declared no con-flict of interest.
reFerences1. Ciuffi, A, Llano, M, Poeschla, E, Hoffmann, C, Leipzig, J, Shinn, P et al. (2005). A role
for LEDGF/p75 in targeting HIV DNA integration. Nat Med 11: 1287–1289.2. Schröder, AR, Shinn, P, Chen, H, Berry, C, Ecker, JR and Bushman, F (2002). HIV-1
integration in the human genome favors active genes and local hotspots. Cell 110: 521–529.
3. Mitchell, RS, Beitzel, BF, Schroder, AR, Shinn, P, Chen, H, Berry, CC et al. (2004). Retroviral DNA integration: ASLV, HIV, and MLV show distinct target site preferences. PLoS Biol 2: E234.
4. Lewinski, MK, Yamashita, M, Emerman, M, Ciuffi, A, Marshall, H, Crawford, G et al. (2006). Retroviral DNA integration: viral and cellular determinants of target-site selection. PLoS Pathog 2: e60.
5. Vandegraaff, N and Engelman, A (2007). Molecular mechanisms of HIV integration and therapeutic intervention. Expert Rev Mol Med 9: 1–19.
6. Llano, M, Saenz, DT, Meehan, A, Wongthida, P, Peretz, M, Walker, WH et al. (2006). An essential role for LEDGF/p75 in HIV integration. Science 314: 461–464.
7. Shun, MC, Raghavendra, NK, Vandegraaff, N, Daigle, JE, Hughes, S, Kellam, P et al. (2007). LEDGF/p75 functions downstream from preintegration complex formation to effect gene-specific HIV-1 integration. Genes Dev 21: 1767–1778.
8. Apolonia, L, Waddington, SN, Fernandes, C, Ward, NJ, Bouma, G, Blundell, MP et al. (2007). Stable gene transfer to muscle using non-integrating lentiviral vectors. Mol Ther 15: 1947–1954.
9. Bayer, M, Kantor, B, Cockrell, A, Ma, H, Zeithaml, B, Li, X et al. (2008). A large U3 deletion causes increased in vivo expression from a nonintegrating lentiviral vector. Mol Ther 16: 1968–1976.
10. Nightingale, SJ, Hollis, RP, Pepper, KA, Petersen, D, Yu, XJ, Yang, C et al. (2006). Transient gene expression by nonintegrating lentiviral vectors. Mol Ther 13: 1121–1132.
11. Yáñez-Muñoz, RJ, Balaggan, KS, MacNeil, A, Howe, SJ, Schmidt, M, Smith, AJ et al. (2006). Effective gene therapy with nonintegrating lentiviral vectors. Nat Med 12: 348–353.
12. Cornu, TI and Cathomen, T (2007). Targeted genome modifications using integrase-deficient lentiviral vectors. Mol Ther 15: 2107–2113.
13. Lombardo, A, Genovese, P, Beausejour, CM, Colleoni, S, Lee, YL, Kim, KA et al. (2007). Gene editing in human stem cells using zinc finger nucleases and integrase-defective lentiviral vector delivery. Nat Biotechnol 25: 1298–1306.

1510 www.moleculartherapy.org vol. 19 no. 8 aug. 2011
© The American Society of Gene & Cell TherapyIntegration of Lentiviral DNA by DNA Transposition
14. Farnet, CM and Haseltine, WA (1991). Circularization of human immunodeficiency virus type 1 DNA in vitro. J Virol 65: 6942–6952.
15. Moldt, B, Staunstrup, NH, Jakobsen, M, Yáñez-Muñoz, RJ and Mikkelsen, JG (2008). Genomic insertion of lentiviral DNA circles directed by the yeast Flp recombinase. BMC Biotechnol 8: 60.
16. Staunstrup, NH, Moldt, B, Mátés, L, Villesen, P, Jakobsen, M, Ivics, Z et al. (2009). Hybrid lentivirus-transposon vectors with a random integration profile in human cells. Mol Ther 17: 1205–1214.
17. Ivics, Z, Hackett, PB, Plasterk, RH and Izsvák, Z (1997). Molecular reconstruction of Sleeping Beauty, a Tc1-like transposon from fish, and its transposition in human cells. Cell 91: 501–510.
18. Liu, G, Geurts, AM, Yae, K, Srinivasan, AR, Fahrenkrug, SC, Largaespada, DA et al. (2005). Target-site preferences of Sleeping Beauty transposons. J Mol Biol 346: 161–173.
19. Yant, SR, Wu, X, Huang, Y, Garrison, B, Burgess, SM and Kay, MA (2005). High-resolution genome-wide mapping of transposon integration in mammals. Mol Cell Biol 25: 2085–2094.
20. Belur, LR, Frandsen, JL, Dupuy, AJ, Ingbar, DH, Largaespada, DA, Hackett, PB et al. (2003). Gene insertion and long-term expression in lung mediated by the Sleeping Beauty transposon system. Mol Ther 8: 501–507.
21. Chen, ZJ, Kren, BT, Wong, PY, Low, WC and Steer, CJ (2005). Sleeping Beauty-mediated down-regulation of huntingtin expression by RNA interference. Biochem Biophys Res Commun 329: 646–652.
22. Izsvák, Z and Ivics, Z (2004). Sleeping beauty transposition: biology and applications for molecular therapy. Mol Ther 9: 147–156.
23. Mikkelsen, JG, Yant, SR, Meuse, L, Huang, Z, Xu, H and Kay, MA (2003). Helper-Independent Sleeping Beauty transposon-transposase vectors for efficient nonviral gene delivery and persistent gene expression in vivo. Mol Ther 8: 654–665.
24. Ohlfest, JR, Demorest, ZL, Motooka, Y, Vengco, I, Oh, S, Chen, E et al. (2005). Combinatorial antiangiogenic gene therapy by nonviral gene transfer using the sleeping beauty transposon causes tumor regression and improves survival in mice bearing intracranial human glioblastoma. Mol Ther 12: 778–788.
25. Yant, SR, Meuse, L, Chiu, W, Ivics, Z, Izsvak, Z and Kay, MA (2000). Somatic integration and long-term transgene expression in normal and haemophilic mice using a DNA transposon system. Nat Genet 25: 35–41.
26. Liu, L, Mah, C and Fletcher, BS (2006). Sustained FVIII expression and phenotypic correction of hemophilia A in neonatal mice using an endothelial-targeted sleeping beauty transposon. Mol Ther 13: 1006–1015.
27. Mátés, L, Chuah, MK, Belay, E, Jerchow, B, Manoj, N, Acosta-Sanchez, A et al. (2009). Molecular evolution of a novel hyperactive Sleeping Beauty transposase enables robust stable gene transfer in vertebrates. Nat Genet 41: 753–761.
28. Vink, CA, Gaspar, HB, Gabriel, R, Schmidt, M, McIvor, RS, Thrasher, AJ et al. (2009). Sleeping beauty transposition from nonintegrating lentivirus. Mol Ther 17: 1197–1204.
29. Qasim, W, Vink, CA and Thrasher, AJ (2010). Hybrid lentiviral vectors. Mol Ther 18: 1263–1267.
30. Luo, G, Ivics, Z, Izsvák, Z and Bradley, A (1998). Chromosomal transposition of a Tc1/mariner-like element in mouse embryonic stem cells. Proc Natl Acad Sci USA 95: 10769–10773.
31. Yant, SR and Kay, MA (2003). Nonhomologous-end-joining factors regulate DNA repair fidelity during Sleeping Beauty element transposition in mammalian cells. Mol Cell Biol 23: 8505–8518.
32. Yant, SR, Park, J, Huang, Y, Mikkelsen, JG and Kay, MA (2004). Mutational analysis of the N-terminal DNA-binding domain of sleeping beauty transposase: critical
residues for DNA binding and hyperactivity in mammalian cells. Mol Cell Biol 24: 9239–9247.
33. Cui, Z, Geurts, AM, Liu, G, Kaufman, CD and Hackett, PB (2002). Structure-function analysis of the inverted terminal repeats of the sleeping beauty transposon. J Mol Biol 318: 1221–1235.
34. Geurts, AM, Yang, Y, Clark, KJ, Liu, G, Cui, Z, Dupuy, AJ et al. (2003). Gene transfer into genomes of human cells by the sleeping beauty transposon system. Mol Ther 8: 108–117.
35. Zayed, H, Izsvák, Z, Walisko, O and Ivics, Z (2004). Development of hyperactive sleeping beauty transposon vectors by mutational analysis. Mol Ther 9: 292–304.
36. Schmidt, M, Schwarzwaelder, K, Bartholomae, C, Zaoui, K, Ball, C, Pilz, I et al. (2007). High-resolution insertion-site analysis by linear amplification-mediated PCR (LAM-PCR). Nat Methods 4: 1051–1057.
37. Vigdal, TJ, Kaufman, CD, Izsvák, Z, Voytas, DF and Ivics, Z (2002). Common physical properties of DNA affecting target site selection of sleeping beauty and other Tc1/mariner transposable elements. J Mol Biol 323: 441–452.
38. Wang, GP, Ciuffi, A, Leipzig, J, Berry, CC and Bushman, FD (2007). HIV integration site selection: analysis by massively parallel pyrosequencing reveals association with epigenetic modifications. Genome Res 17: 1186–1194.
39. Yant, SR, Ehrhardt, A, Mikkelsen, JG, Meuse, L, Pham, T and Kay, MA (2002). Transposition from a gutless adeno-transposon vector stabilizes transgene expression in vivo. Nat Biotechnol 20: 999–1005.
40. Peterson, EB, Mastrangelo, MA, Federoff, HJ and Bowers, WJ (2007). Neuronal specificity of HSV/sleeping beauty amplicon transduction in utero is driven primarily by tropism and cell type composition. Mol Ther 15: 1848–1855.
41. Vargas, J Jr, Gusella, GL, Najfeld, V, Klotman, ME and Cara, A (2004). Novel integrase-defective lentiviral episomal vectors for gene transfer. Hum Gene Ther 15: 361–372.
42. Oehmig, A, Cortés, ML, Perry, KF, Sena-Esteves, M, Fraefel, C and Breakefield, XO (2007). Integration of active human beta-galactosidase gene (100 kb) into genome using HSV/AAV amplicon vector. Gene Ther 14: 1078–1091.
43. Kubo, S and Mitani, K (2003). A new hybrid system capable of efficient lentiviral vector production and stable gene transfer mediated by a single helper-dependent adenoviral vector. J Virol 77: 2964–2971.
44. Bowers, WJ, Mastrangelo, MA, Howard, DF, Southerland, HA, Maguire-Zeiss, KA and Federoff, HJ (2006). Neuronal precursor-restricted transduction via in utero CNS gene delivery of a novel bipartite HSV amplicon/transposase hybrid vector. Mol Ther 13: 580–588.
45. Müther, N, Noske, N and Ehrhardt, A (2009). Viral Hybrid Vectors for Somatic Integration - Are They the Better Solution? Viruses 1: 1295-1324.
46. Thyagarajan, B, Olivares, EC, Hollis, RP, Ginsburg, DS and Calos, MP (2001). Site-specific genomic integration in mammalian cells mediated by phage phiC31 integrase. Mol Cell Biol 21: 3926–3934.
47. Tan, W, Zhu, K, Segal, DJ, Barbas, CF 3rd and Chow, SA (2004). Fusion proteins consisting of human immunodeficiency virus type 1 integrase and the designed polydactyl zinc finger protein E2C direct integration of viral DNA into specific sites. J Virol 78: 1301–1313.
48. Yant, SR, Huang, Y, Akache, B and Kay, MA (2007). Site-directed transposon integration in human cells. Nucleic Acids Res 35: e50.
49. Ivics, Z, Katzer, A, Stüwe, EE, Fiedler, D, Knespel, S and Izsvák, Z (2007). Targeted Sleeping Beauty transposition in human cells. Mol Ther 15: 1137–1144.
50. Langmead, B, Trapnell, C, Pop, M and Salzberg, SL (2009). Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol 10: R25.