comparative analysis of mitochondrial genomes in bombina (anura; bombinatoridae)
TRANSCRIPT

Comparative Analysis of Mitochondrial Genomes in Bombina(Anura; Bombinatoridae)
Maciej Pabijan Æ Christina Spolsky ÆThomas Uzzell Æ Jacek M. Szymura
Received: 31 August 2007 / Accepted: 9 May 2008 / Published online: 12 August 2008
� Springer Science+Business Media, LLC 2008
Abstract The complete mitochondrial genomes of two
basal anurans, Bombina bombina and B. variegata (Anura;
Bombinatoridae), were sequenced. The gene order of their
mitochondrial DNA (mtDNA) is identical to that of
canonical vertebrate mtDNA. In contrast, we show that
there are structural differences in regulatory regions and
protein coding genes between the mtDNA of these two
closely related species. Corrected sequence divergence
between the mtDNA of B. bombina and B. variegata
amounts to 8.7% (2.3% divergence in amino acids).
Comparisons with two East Asian congeners show that the
control region contains two repeat regions, LV1 and LV2,
present in all species except for B. bombina, in which LV2
has been secondarily lost. The rRNAs and tRNAs are
characterized by low nucleotide divergence. The protein
coding genes are considerably more disparate, although
functional constraint is high but variable among genes, as
evidenced by dN/dS ratios. A mtDNA phylogeny
established the distribution of autapomorphic nonsynon-
omous substitutions in the mitogenomes of B. bombina and
B. variegata. Nine of 98 nonsynonomous substitutions led
to radical amino acid replacements that may alter mito-
chondrial protein function. Most radical substitutions were
found in ND2, ND4, or ND5, encoding mitochondrial
subunits of complex I of the electron transport system.
The extensive divergence between the mitogenomes of
B. bombina and B. variegata is discussed in terms of its
possible role in impeding gene flow in natural hybrid zones
between these two species.
Keywords mtDNA � Bombina � Divergence � Control
region � ETS complex I � Hybrid zone
Introduction
In eukaryotes, transmembrane proteins imbedded in the
inner membrane of the mitochondria are the sites of energy
conversion and cellular respiration. These proteins consist
of subunits of the electron transport system (ETS) and are
encoded by either nuclear DNA (nDNA) or mitochondrial
DNA (mtDNA). The products of both genomes and their
precise interactions are essential for aerobic metabolism. A
plethora of disorders in humans linked to sequence varia-
tion in mtDNA or mitochondrially expressed nDNA
illustrates the evolutionary constraint imposed on genes of
both genomes (Larsson and Clayton 1995; Schapira 2006).
Furthermore, specific mtDNA haplotypes have been shown
to influence individual fitness in humans, mice, Drosoph-
ila, and copepods (reviewed by Ballard and Whitlock 2004;
Gemmell et al. 2004; Ballard and Rand 2005). It has been
estimated that the ETS is comprised of approximately 90
different respiratory chain subunits, the majority of which
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00239-008-9123-3) contains supplementarymaterial, which is available to authorized users.
M. Pabijan (&) � J. M. Szymura
Department of Comparative Anatomy, Jagiellonian University,
Ingardena 6, 30-060 Krakow, Poland
e-mail: [email protected]
J. M. Szymura
e-mail: [email protected]
C. Spolsky � T. Uzzell
Center for Systematic Biology and Evolution, Academy
of Natural Sciences, 1900 Benjamin Franklin Parkway,
Philadelphia, PA 19103, USA
e-mail: [email protected]
T. Uzzell
e-mail: [email protected]
123
J Mol Evol (2008) 67:246–256
DOI 10.1007/s00239-008-9123-3

are transcribed in the nucleus, assembled in the cytoplasm,
and transported into the mitochondrion (Larsson and
Clayton 1995). The mitogenome encodes only 13 of these
protein subunits and 24 RNA components (12S and 16S
rRNA molecules, 22 tRNAs) necessary for mitochondrial
protein synthesis. Despite this small proportion, the high
mutation rate of mtDNA may drive the evolution of
mitochondrial-nuclear interactions by contributing ‘new’
amino acids for the scrutiny of natural selection (Wu et al.
2000; Schmidt et al. 2001; Willet and Burton 2004). These
results have prompted a reevaluation of the neutrality of
mtDNA and show the need for further research in this area.
The physical and functional contiguity of the nuclear
and mitochondrial proteins in the enzymatic complexes of
the ETS leads to coadaptation of the two genomes (Blier
et al. 2001; Ballard and Whitlock 2004; Ballard and Rand
2005). Moreover, interactions between nuclear-encoded
mtRNA polymerases and their promoters located in
mtDNA control regions have been shown to be species-
specific (Gaspari et al. 2004). The polymerases, and pos-
sibly additional transcription factors, may not bind to the
mtDNA as precisely if presented with a divergent, foreign
promoter (Burton et al. 2006). Hence, mitochondrial genes
cannot be exempt from the epistasis evolving between
geographically delimited sets of nuclear alleles. The dis-
association of these interactions in interpopulation or
interspecies hybrids may cause inferior mitochondrial
function (McKenzie et al. 2003; Ellison and Burton 2006;
Burton et al. 2006).
The hybridizing European fire-bellied and yellow-bel-
lied toads, Bombina bombina and B. variegata, have
evolved in different environments (Szymura 1993). The
lowland form, B. bombina, inhabits larger, permanent
ponds, while the mountain type, B. variegata, prefers
smaller transient ponds and puddles. The toads are
divergent at the molecular level, and each exhibits a suite
of phenotypic traits in morphology, life history, ecology,
and behavior, regarded as adaptations to their respective
environments (Szymura 1993). Hybrid zones between the
toads occur in areas of altitudinal transition along their
parapatric contact in central Europe (Szymura and Barton
1991; Yanchukov et al. 2006). Selection against hybrids
results from intrinsic hybrid dysfunction (Szymura and
Barton 1986; Kruuk et al. 1999). Despite extensive
hybridization, no introgression of mtDNA has taken place
outside of the narrow zones (Szymura et al. 2000; Yan-
chukov et al. 2006; Hofman et al. 2007; Hofman and
Szymura 2007). Clines in mtDNA are no wider, and are
sometimes narrower, than clines at multiple unlinked al-
lozyme loci and morphological traits (Hofman and
Szymura 2007). Postzygotic reproductive barriers between
B. bombina and B. variegata have thus been attributed to
substantial genetic divergence in the genomes of the toads
and the genetic incompatibility that ensues (Szymura
et al. 1985; Nurnberger et al. 2003; Spolsky et al. 2006),
with differences in mtDNA hypothesized to contribute to
the negative epistasis in recombined hybrids (Hofman and
Szymura 2007).
In this paper we describe the complete mtDNA
sequences of the two hybridizing European species, B.
bombina and B. variegata. We quantify divergence
between the two genomes and their functional components,
including the distinctive organization of their control
regions (Spolsky et al. 2006), and study the distribution of
substitutions across the mitogenomes. Using a phylogeny
based on four species of Bombina and five outgroup spe-
cies, we identify regions in the mtDNA of B. bombina and
B. variegata carrying nonsynonymous and radical amino
acid substitutions and, conversely, conservative nucleotide
and amino acid domains. Apart from augmenting our
understanding of postzygotic reproductive barriers in
Bombina, our results should be useful for elucidating the
mechanisms of molecular evolution of mtDNA at low
levels of divergence, and contribute to the growing number
of described mitogenomes of the most basal group of
anuran amphibians (archeobatrachians), whose mtDNA is
known to evolve particularly slowly (Igawa et al. 2008).
Materials and Methods
Laboratory Methods
A single individual of B. variegata from Radziszow near
Krakow, Poland (49�570 N, 19�480 E) and a B. bombina
from Tyniec, Poland (50�020 N, 19�480 E) were used as
sources of mtDNA. The mtDNAs were purified as by
Szymura et al. (1985, 2000). Aliquots of B. variegata
mtDNA were partially digested with either HindIII or XbaI
and subsequently cloned into compatible pUC18 sites
(Boehringer Mannheim). MtDNA was radioactively
labeled and used as a probe to screen for clones containing
mtDNA fragments. The fragments were sized to compare
with mitochondrial maps (Spolsky et al. 2006) and inserts
were sequenced with universal M13 (forward and reverse)
primers and additional walking primers. Bombina specific
primers amplifying overlapping fragments spanning the
entire mtDNA molecule on both strands were then
designed on the basis of the cloned B. variegata mtDNA
for both PCR and sequencing in B. bombina (PCR/
sequencing strategy and primers listed in Electronic Sup-
plementary Material, S1 and S2). Two sequencing
chemistries were used as appropriate for a Beckman CEQ
sequencer (B. variegata mtDNA clones) and Applied
Biosystem Analyzer (most B. bombina mtDNA
amplicons).
J Mol Evol (2008) 67:246–256 247
123
Molecular Analyses
The mtDNA sequences were edited and assembled using
SeqMan version 5.05. After alignment of the entire gen-
omes in ClustalW, the two rRNAs, 22 tRNAs, and 13
protein coding genes were identified by comparisons to the
mtDNA genomes of B. orientalis (AY585338 [San Mauro
et al. 2004a]) and B. fortinuptialis (AY458591 [Zhang
et al. 2005], incorrectly listed in the NCBI database as B.
bombina [cf. Spolsky et al. 2006]). Molecular evolutionary
analyses including base composition, sequence divergence,
numbers of transitions and transversions, and numbers of
synonymous and nonsynonymous substitutions were car-
ried out using MEGA v.3.1 (Kumar et al. 2004) and DnaSP
(Rozas et al. 2003). We also performed a sliding window
analysis of nucleotide divergence in the four Bombina
mtDNA genomes. We identified radical replacement
changes in the protein coding genes, i.e. the replacement of
one amino acid by another that belongs to a different class
defined by the properties of lateral chains (e.g., Doiron
et al. 2002). Because all mitochondrially encoded peptides
exhibit transmembrane domains, we used a mutation
matrix specifically designed for transmembrane proteins
for scoring (Jones et al. 1994). We then mapped the auta-
pomorphic and radical amino acid substitutions onto the
putative topologies of the mtDNA-encoded proteins by
applying the hidden Markov method for sequence feature
prediction implemented in PolyPhobius (Kall et al. 2005)
and TMHMM (Krogh et al. 2001). Because homologues
are likely to share the same secondary structures, PolyPh-
obius also uses existing information from homologous
sequences in protein databases.
Phylogenetic Analyses
Besides the Bombina genomes listed above, we also used the
closest living relatives of Bombina from which entire mito-
chondrial genomes are available, i.e., Discoglossus galganoi
(AY585339) and Alytes obstetricans pertinax (AY585337).
Mitochondrial genomes from three other representatives of
Archaeobatrachia (Frost et al. 2006; Roelants et al. 2007)
were used as more distant outgroups: Ascaphus truei
(AJ871087), Xenopus tropicalis (AY789013), and Pelobates
cultripes (AJ871086). Phylogenetic analyses were per-
formed on 12 mitochondrial H-strand protein-coding genes.
The L-strand-encoded ND6 gene was excluded because of
differences in base composition. The mitochondrial protein
coding sequences of these 12 genes were translated, con-
catenated to form a single chain, and aligned using ClustalW
as implemented in MEGA v.3.1.
Multiple substitutions at the same site confound phylo-
genetic inference and obscure evolutionary relationships.
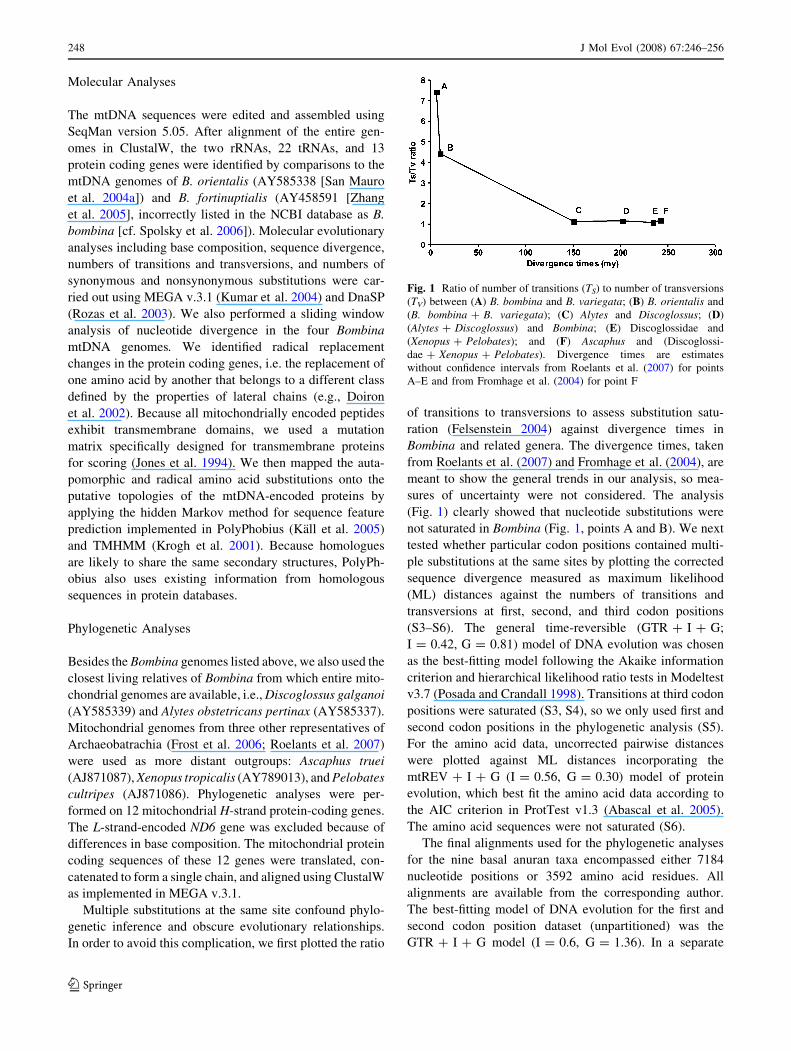
In order to avoid this complication, we first plotted the ratio
of transitions to transversions to assess substitution satu-
ration (Felsenstein 2004) against divergence times in
Bombina and related genera. The divergence times, taken
from Roelants et al. (2007) and Fromhage et al. (2004), are
meant to show the general trends in our analysis, so mea-
sures of uncertainty were not considered. The analysis
(Fig. 1) clearly showed that nucleotide substitutions were
not saturated in Bombina (Fig. 1, points A and B). We next
tested whether particular codon positions contained multi-
ple substitutions at the same sites by plotting the corrected
sequence divergence measured as maximum likelihood
(ML) distances against the numbers of transitions and
transversions at first, second, and third codon positions
(S3–S6). The general time-reversible (GTR + I + G;
I = 0.42, G = 0.81) model of DNA evolution was chosen
as the best-fitting model following the Akaike information
criterion and hierarchical likelihood ratio tests in Modeltest
v3.7 (Posada and Crandall 1998). Transitions at third codon
positions were saturated (S3, S4), so we only used first and
second codon positions in the phylogenetic analysis (S5).
For the amino acid data, uncorrected pairwise distances
were plotted against ML distances incorporating the
mtREV + I + G (I = 0.56, G = 0.30) model of protein
evolution, which best fit the amino acid data according to
the AIC criterion in ProtTest v1.3 (Abascal et al. 2005).
The amino acid sequences were not saturated (S6).
The final alignments used for the phylogenetic analyses
for the nine basal anuran taxa encompassed either 7184
nucleotide positions or 3592 amino acid residues. All
alignments are available from the corresponding author.
The best-fitting model of DNA evolution for the first and
second codon position dataset (unpartitioned) was the
GTR + I + G model (I = 0.6, G = 1.36). In a separate
Fig. 1 Ratio of number of transitions (TS) to number of transversions
(TV) between (A) B. bombina and B. variegata; (B) B. orientalis and
(B. bombina + B. variegata); (C) Alytes and Discoglossus; (D)
(Alytes + Discoglossus) and Bombina; (E) Discoglossidae and
(Xenopus + Pelobates); and (F) Ascaphus and (Discoglossi-
dae + Xenopus + Pelobates). Divergence times are estimates
without confidence intervals from Roelants et al. (2007) for points
A–E and from Fromhage et al. (2004) for point F
248 J Mol Evol (2008) 67:246–256
123
analysis, we partitioned the mtDNA genes into three
functional groups (protein coding genes, rRNAs, tRNAs)
and assigned models to each through Modeltest. For the
rRNAs and for the tRNAs, Modeltest suggested values of
GTR + G (G = 0.32) and GTR + I + G (I = 0.42,
G = 1.18), respectively. ML analyses of the unpartitioned
first and second codon position dataset and the amino acid
dataset were conducted in PHYML v2.4.4 (Guindon and
Gascuel 2003), starting with the BIONJ trees. Nonpara-
metric bootstrapping was used to test the reliabilities of the
ML trees (1000 pseudoreplicates). Bayesian inference (BI)
of the phylogeny for all three datasets was carried out using
MrBayes v3.04b (Huelsenbeck and Ronquist 2001) with
the previously specified models of sequence evolution; all
other priors were set to default values. One cold and three
heated chains were run for 5 million generations, with trees
sampled every 100 generations. Generations sampled
before the chains reached stationarity (20,000), as judged
by examining the log-likelihoods of the cold chains and
plots of the generation vs. log-likelihood values of the data,
were discarded as burn-in. At least two independent runs
were carried out for each dataset in the Bayesian analyses.
Results
Genome Organization and Nucleotide Composition
The complete mtDNA genomes have been deposited at the
GenBank database under accession numbers EU115993 (B.
bombina) and AY971143 (B. variegata). Within the genus,
the B. variegata mtDNA genome was the largest, at
18,551 bp, followed by B. fortinuptialis (17,575 bp), B.
orientalis (17,173 bp), and B. bombina (17,154 bp). The
differences in mtDNA genome size can be accounted for
by size variation in the control region. The mtDNA gen-
omes of Bombina exhibit the organization and gene content
of the canonical mtDNA genome of other vertebrates
(Table 1; Fig. 3) (Boore 1999). The gene arrangement and
start and stop codons in the Bombina mtDNA genomes are
identical in all studied species (Table 1). The putative
origin of L-strand replication (OL) in the Bombina mitog-
enomes was located within the WANCY tRNA cluster,
between the tRNAAsn and the tRNACys genes. The lengths
of the protein coding genes are also similar, with the
exception of the third codon in ND1 (ACT, threonine in B.
variegata), which is absent in B. bombina. The nucleotide
frequencies of the L-strands are very similar in all four
Bombina species (Table 1) and skewed against guanine
because of bias against this nucleotide at the second and
third codon positions. The transition-to-transversion ratio
was quite variable among genes, but always skewed toward
transitions (Table 2).
Control Region Structure
We compared the control regions in all four sequenced
Bombina mtDNAs (Fig. 2A). The size of the control region
varies nearly twofold in Bombina, consisting of 3072 bp in
B. variegata, 2373 bp in B. orientalis, 1990 bp in B. for-
tinuptialis, and 1675 bp in B. bombina (Fig. 2A). The B.
fortinuptialis control region is incomplete because of an
unsequenced stretch at the 30-end of the first tandem repeat
(Zhang et al. 2005). The latter, termed LV1 by Spolsky
et al. (2006), consists of 4–12 serial repeats 70–77 bp long
(Fig. 2B). A highly variable nucleotide sequence is found
downstream of LV1, with approximately the first 150 bp
representing incomplete repeat units. Three conserved
sequence blocks (CSB1–3) are localized downstream of
LV1. The region –180 bp relative to CSB1 is alignable in
all Bombina mitogenomes studied and encompasses a
poly(T) block, the pyrimidine-rich region, PP-1, of San
Table 1 Lengths, in base pairs, of structural features in the BombinamtDNA genomes and nucleotide compositions of mitochondrial L-strands
Feature Bbom Bvar Bori Bfor Start; stop
Total length 17,154 18,551 17,173 17,575
% A 30.0% 30.0% 30.0% 30.5%
% C 26.5% 26.1% 26.7% 25.4%
% G 15.8% 16.0% 15.5% 15.3%
% T 27.7% 27.9% 27.8% 28.8%
12S rRNA 933 933 933 929
16S rRNA 1599 1601 1599 1599
ATP6 683 683 683 683 ATG; TAA
ATP8 168 168 168 168 ATG; TAA
Cytb 1141 1141 1141 1141 ATG; T–
COI 1554 1554 1554 1554 GTG; TAA
COII 688 688 688 688 ATG; T–
COIII 784 784 784 784 ATG; T–
ND1 959 962 962 962 ATG; TAG
ND2 1045 1045 1045 1045 ATG; T–
ND3 343 343 343 343 ATG; T–
ND4 1378 1378 1378 1378 ATG; T–
ND4L 297 297 297 297 ATG; TAA
ND5 1809 1809 1809 1809 ATG; TAA
ND6 510 510 510 510 ATG; AGA
Control region 1675 3072 2373 1990
Intergenic DNA 42 41 42 37
Note: Initiation and termination codons (start; stop) are given in the
last column. Incomplete stop codons (T–) presumably become func-
tional by subsequent polyadenylation of the respective mRNAs. B.orientalis mtDNA sequence from San Mauro et al. (2004a;
AY585338). B. fortinuptialis mtDNA sequence from Zhang et al.
(2005; AY458591; incorrectly listed in the NCBI database as B.bombina [cf. Spolsky et al. 2006]). Bbom, B. bombina; Bvar, B.variegata; Bori, B.orientalis; Bfor, B. fortinuptialis
J Mol Evol (2008) 67:246–256 249
123
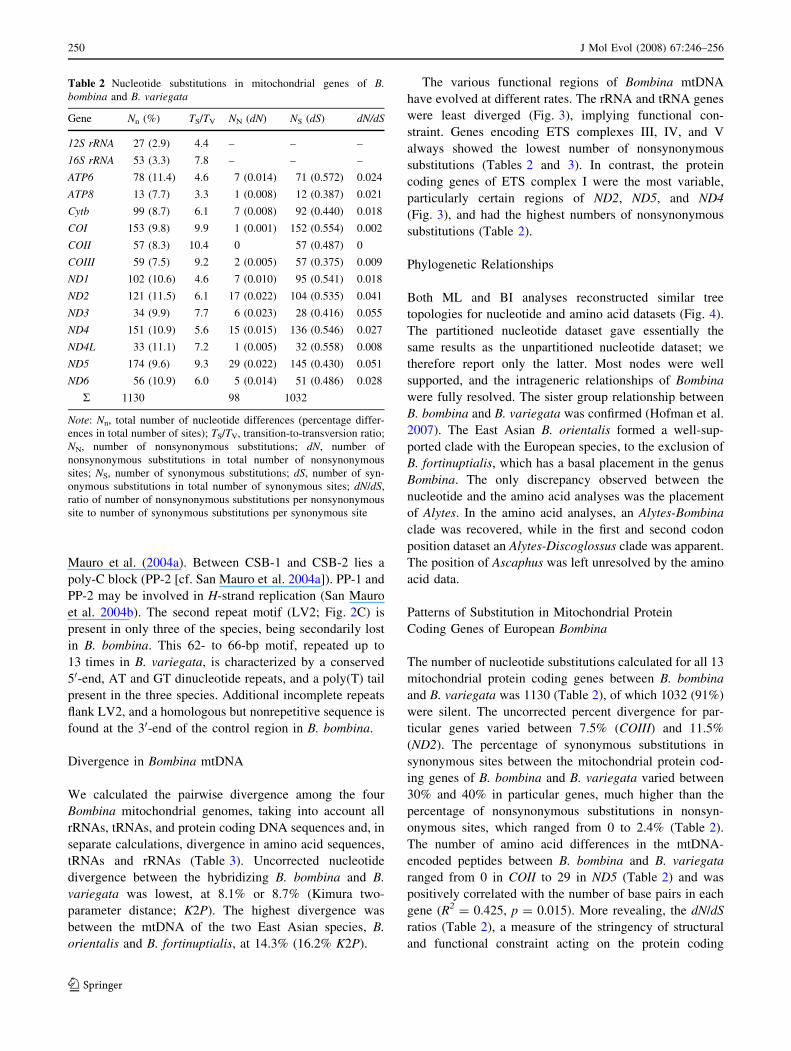
Mauro et al. (2004a). Between CSB-1 and CSB-2 lies a
poly-C block (PP-2 [cf. San Mauro et al. 2004a]). PP-1 and
PP-2 may be involved in H-strand replication (San Mauro
et al. 2004b). The second repeat motif (LV2; Fig. 2C) is
present in only three of the species, being secondarily lost
in B. bombina. This 62- to 66-bp motif, repeated up to
13 times in B. variegata, is characterized by a conserved
50-end, AT and GT dinucleotide repeats, and a poly(T) tail
present in the three species. Additional incomplete repeats
flank LV2, and a homologous but nonrepetitive sequence is
found at the 30-end of the control region in B. bombina.
Divergence in Bombina mtDNA
We calculated the pairwise divergence among the four
Bombina mitochondrial genomes, taking into account all
rRNAs, tRNAs, and protein coding DNA sequences and, in
separate calculations, divergence in amino acid sequences,
tRNAs and rRNAs (Table 3). Uncorrected nucleotide
divergence between the hybridizing B. bombina and B.
variegata was lowest, at 8.1% or 8.7% (Kimura two-
parameter distance; K2P). The highest divergence was
between the mtDNA of the two East Asian species, B.
orientalis and B. fortinuptialis, at 14.3% (16.2% K2P).
The various functional regions of Bombina mtDNA
have evolved at different rates. The rRNA and tRNA genes
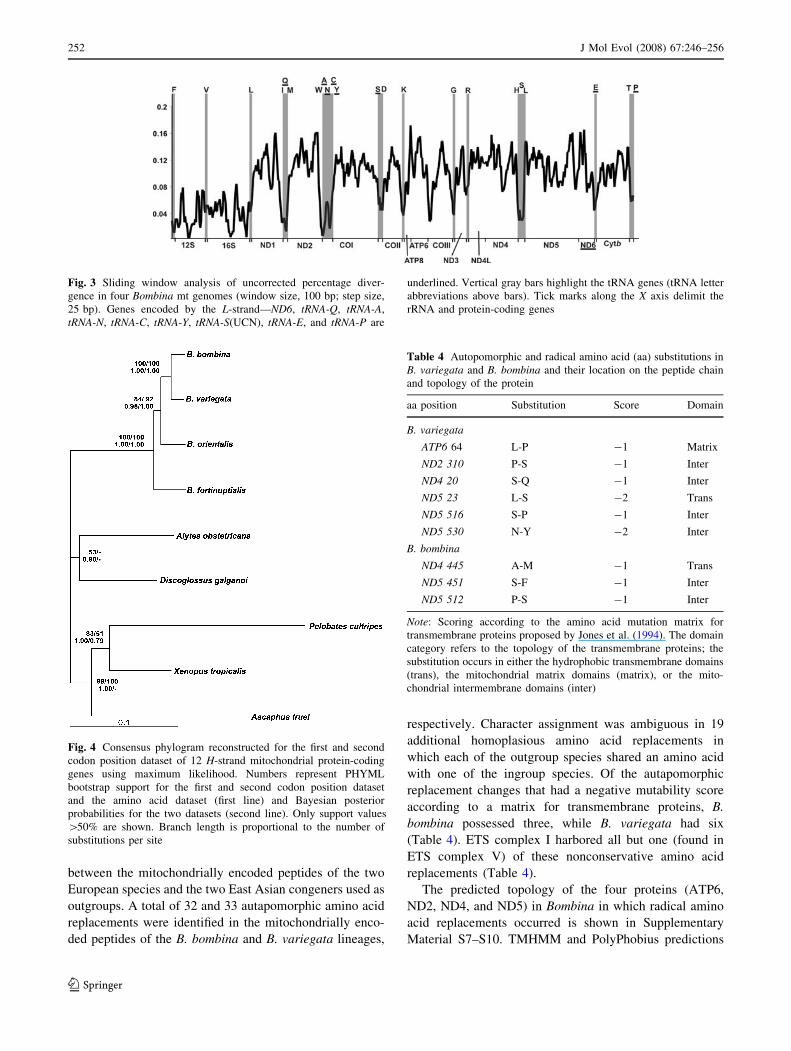
were least diverged (Fig. 3), implying functional con-
straint. Genes encoding ETS complexes III, IV, and V
always showed the lowest number of nonsynonymous
substitutions (Tables 2 and 3). In contrast, the protein
coding genes of ETS complex I were the most variable,
particularly certain regions of ND2, ND5, and ND4
(Fig. 3), and had the highest numbers of nonsynonymous
substitutions (Table 2).
Phylogenetic Relationships
Both ML and BI analyses reconstructed similar tree
topologies for nucleotide and amino acid datasets (Fig. 4).
The partitioned nucleotide dataset gave essentially the
same results as the unpartitioned nucleotide dataset; we
therefore report only the latter. Most nodes were well
supported, and the intrageneric relationships of Bombina
were fully resolved. The sister group relationship between
B. bombina and B. variegata was confirmed (Hofman et al.
2007). The East Asian B. orientalis formed a well-sup-
ported clade with the European species, to the exclusion of
B. fortinuptialis, which has a basal placement in the genus
Bombina. The only discrepancy observed between the
nucleotide and the amino acid analyses was the placement
of Alytes. In the amino acid analyses, an Alytes-Bombina
clade was recovered, while in the first and second codon
position dataset an Alytes-Discoglossus clade was apparent.
The position of Ascaphus was left unresolved by the amino
acid data.
Patterns of Substitution in Mitochondrial Protein
Coding Genes of European Bombina
The number of nucleotide substitutions calculated for all 13
mitochondrial protein coding genes between B. bombina
and B. variegata was 1130 (Table 2), of which 1032 (91%)
were silent. The uncorrected percent divergence for par-
ticular genes varied between 7.5% (COIII) and 11.5%
(ND2). The percentage of synonymous substitutions in
synonymous sites between the mitochondrial protein cod-
ing genes of B. bombina and B. variegata varied between
30% and 40% in particular genes, much higher than the
percentage of nonsynonymous substitutions in nonsyn-
onymous sites, which ranged from 0 to 2.4% (Table 2).
The number of amino acid differences in the mtDNA-
encoded peptides between B. bombina and B. variegata
ranged from 0 in COII to 29 in ND5 (Table 2) and was
positively correlated with the number of base pairs in each
gene (R2 = 0.425, p = 0.015). More revealing, the dN/dS
ratios (Table 2), a measure of the stringency of structural
and functional constraint acting on the protein coding
Table 2 Nucleotide substitutions in mitochondrial genes of B.bombina and B. variegata
Gene Nn (%) TS/TV NN (dN) NS (dS) dN/dS
12S rRNA 27 (2.9) 4.4 – – –
16S rRNA 53 (3.3) 7.8 – – –
ATP6 78 (11.4) 4.6 7 (0.014) 71 (0.572) 0.024
ATP8 13 (7.7) 3.3 1 (0.008) 12 (0.387) 0.021
Cytb 99 (8.7) 6.1 7 (0.008) 92 (0.440) 0.018
COI 153 (9.8) 9.9 1 (0.001) 152 (0.554) 0.002
COII 57 (8.3) 10.4 0 57 (0.487) 0
COIII 59 (7.5) 9.2 2 (0.005) 57 (0.375) 0.009
ND1 102 (10.6) 4.6 7 (0.010) 95 (0.541) 0.018
ND2 121 (11.5) 6.1 17 (0.022) 104 (0.535) 0.041
ND3 34 (9.9) 7.7 6 (0.023) 28 (0.416) 0.055
ND4 151 (10.9) 5.6 15 (0.015) 136 (0.546) 0.027
ND4L 33 (11.1) 7.2 1 (0.005) 32 (0.558) 0.008
ND5 174 (9.6) 9.3 29 (0.022) 145 (0.430) 0.051
ND6 56 (10.9) 6.0 5 (0.014) 51 (0.486) 0.028
R 1130 98 1032
Note: Nn, total number of nucleotide differences (percentage differ-
ences in total number of sites); TS/TV, transition-to-transversion ratio;
NN, number of nonsynonymous substitutions; dN, number of
nonsynonymous substitutions in total number of nonsynonymous
sites; NS, number of synonymous substitutions; dS, number of syn-
onymous substitutions in total number of synonymous sites; dN/dS,
ratio of number of nonsynonymous substitutions per nonsynonymous
site to number of synonymous substitutions per synonymous site
250 J Mol Evol (2008) 67:246–256
123
genes, shows that the strength of selection against nonsyn-
onymous change in the Bombina protein coding genes is
strong. According to this measure, the protein coding genes
can be arranged in the following order, from most conser-
vative to most variable: COII \ COI \ ND4L \\COIII\ Cytb \ ND1 \ ATP8 \ ATP6 \ ND4 \ ND6 \\ND2 \ ND5 \ ND3. Of the total of 98 nonsynonymous
substitutions between the mtDNA of B. bombina and B.
variegata, the majority (80) were found in ETS complex I of
the electron transport system (Table 3). Only seven
nonsynonymous substitutions were found in ETS complex
III, three in complex IV, and eight in complex V (Table 3).
Functional Significance of the Amino Acid
Substitutions
On the basis of the phylogenetic relationships among the
mtDNA genomes of the basal anurans (Fig. 4), we inferred
the derived state (either synapomorphic, autapomorphic, or
homoplasious) for all replacement substitutions observed
?
B. bombina
B. orientalis
B. fortinuptialis
5'
5'
5'
5'
3'
3'
3'
3'
B.variegataB.bombinaB.orientalisB.fortinuptialis
------------A TATTA TA-- AGGAC C T CTT TCC T AC T TTATATGC----A T--TA TG-- GGGGT C C CAT --- T TC -
CTATATATTT--A C--TA TAT- GGAC- C T CTT TCC T AT ------TATTTAGT TTT-- CATA G-AC- A C ATA TCC C AC T
TA ATG TA TAT TATGTATAAT GAGCATTCATCT T TCA GAATATCC ATA ATG TA TAT TATGTATAAT GAGCATTCATCT T TCA GAATATCC ATA ATG TA TAT TATGTATAAT GAGCATTCATCT T TCA GAATATCC ATA ATG TA TAT TATGTATAAT GAGCATTCATCT T TCA GAATATCC A
TAS
B.variegata
B.fortinuptialis
T T GG T CCAATATG GAAT TTA-- T A CATATGAC- A
AGCAGGAAC CC G CG ATA AA T TATA AT TAT TTTTT TAGCAGGAAC CC G CG ATA AA T TATA AT TAT TTTTT T
C A AA T CCATAAGG ATTC GGTG- T G GTG------ T T A GA C ATTTGTGA AC-C GGTG- C G GTGTAA--- TB.orientalis
AGCAGGAAC CC G CG ATA AA T TATA AT TAT TTTTT T
2VL1VL
(A)
(B)
(C)
Fig. 2 A comparison of control region structure in European and
East Asian Bombina species. (A) Large vertical bars represent
tandemly arranged repeat units within LV1 and LV2. Filled circles
represent conserved sequence blocks (CSB); small vertical bars
among the CSBs are pyrimidine-rich regions (PP-1 and PP-2). Length
is proportional to the number of nucleotides in each species. The
question mark in B. fortinuptialis marks an unsequenced fragment
after four repeats. (B) Alignment of the repeat unit LV1. Horizontal
bar denotes the putative termination associated sequence (TAS). (C)
Alignment of the repeat unit LV2. Conservative sites are in boldface
Table 3 Pairwise comparisons of divergence in functional units among Bombina mtDNA genomes
Total rRNA tRNA AA ETS complex I ETS complex III ETS complex IV ETS complex V
Bbom-Bvar 8.1 (8.7) 80 (3.1) 37 (2.4) 80.0 7.0 3.0 8.0 2.3
Bbom-Bori 12.4 (13.9) 152 (6.0) 62 (4.0) 146.8 16.0 10.0 16.5 4.4
Bbom-Bfor 13.8 (15.5) 160 (6.3) 116 (7.6) 215.6 18.0 20.0 16.0 5.7
Bvar-Bori 12.5 (14.0) 149 (5.9) 73 (4.8) 146.0 19.5 9.0 21.0 4.4
Bvar-Bfor 13.9 (15.7) 172 (6.8) 112 (7.3) 222.3 18.5 19.0 16.0 5.7
Bori-Bfor 14.3 (16.2) 178 (7.0) 111 (7.3) 216.0 18.5 23.0 25.0 6.2
Note: Bbom, B. bombina; Bvar, B. variegata; Bori, B.orientalis; Bfor, B. fortinuptialis. Total: the percentage of different nucleotides and Kimura
two-parameter corrected distances (K2P; percentages) in complete mtDNA genomes (excluding control regions). RNA: nucleotide differences in
mitochondrial RNA-encoding genes pooled for the 2 rRNAs and 22 tRNAs. Numbers in parentheses are uncorrected divergences given as
percentages. ETS complex: The numbers of nonsynonymous substitutions, calculated using Nei and Gojobori’s method (1986), for peptides in
subunits of the oxidative phosphorylation complex. ETS complex I: ND1–6, 4L. ETS complex III: Cytb. ETS complex IV: COI, COII, COIII.ETS complex V: ATP6, ATP8. AA: Poisson-corrected amino acid divergence for the concatenated mtDNA-encoded proteins, as a percentage
J Mol Evol (2008) 67:246–256 251
123
between the mitochondrially encoded peptides of the two
European species and the two East Asian congeners used as
outgroups. A total of 32 and 33 autapomorphic amino acid
replacements were identified in the mitochondrially enco-
ded peptides of the B. bombina and B. variegata lineages,
respectively. Character assignment was ambiguous in 19
additional homoplasious amino acid replacements in
which each of the outgroup species shared an amino acid
with one of the ingroup species. Of the autapomorphic
replacement changes that had a negative mutability score
according to a matrix for transmembrane proteins, B.
bombina possessed three, while B. variegata had six
(Table 4). ETS complex I harbored all but one (found in
ETS complex V) of these nonconservative amino acid
replacements (Table 4).
The predicted topology of the four proteins (ATP6,
ND2, ND4, and ND5) in Bombina in which radical amino
acid replacements occurred is shown in Supplementary
Material S7–S10. TMHMM and PolyPhobius predictions
Fig. 3 Sliding window analysis of uncorrected percentage diver-
gence in four Bombina mt genomes (window size, 100 bp; step size,
25 bp). Genes encoded by the L-strand—ND6, tRNA-Q, tRNA-A,
tRNA-N, tRNA-C, tRNA-Y, tRNA-S(UCN), tRNA-E, and tRNA-P are
underlined. Vertical gray bars highlight the tRNA genes (tRNA letter
abbreviations above bars). Tick marks along the X axis delimit the
rRNA and protein-coding genes
Fig. 4 Consensus phylogram reconstructed for the first and second
codon position dataset of 12 H-strand mitochondrial protein-coding
genes using maximum likelihood. Numbers represent PHYML
bootstrap support for the first and second codon position dataset
and the amino acid dataset (first line) and Bayesian posterior
probabilities for the two datasets (second line). Only support values
[50% are shown. Branch length is proportional to the number of
substitutions per site
Table 4 Autopomorphic and radical amino acid (aa) substitutions in
B. variegata and B. bombina and their location on the peptide chain
and topology of the protein
aa position Substitution Score Domain
B. variegata
ATP6 64 L-P -1 Matrix
ND2 310 P-S -1 Inter
ND4 20 S-Q -1 Inter
ND5 23 L-S -2 Trans
ND5 516 S-P -1 Inter
ND5 530 N-Y -2 Inter
B. bombina
ND4 445 A-M -1 Trans
ND5 451 S-F -1 Inter
ND5 512 P-S -1 Inter
Note: Scoring according to the amino acid mutation matrix for
transmembrane proteins proposed by Jones et al. (1994). The domain
category refers to the topology of the transmembrane proteins; the
substitution occurs in either the hydrophobic transmembrane domains
(trans), the mitochondrial matrix domains (matrix), or the mito-
chondrial intermembrane domains (inter)
252 J Mol Evol (2008) 67:246–256
123

were generally congruent, although the algorithm used in
the latter application consistently identified a higher num-
ber of transmembrane helices. In addition, the orientation
of the proteins was often contradictory. The locations of
the autapomorphic, radical amino acid substitutions in the
mitochondrial peptides of B. bombina and B. variegata are
given in Table 3 and Supplementary Material S7–S10.
The ATP6 subunit is part of the F0 segment of the ATP
synthase and takes part in the channeling of protons
through the inner membrane of the mitochondrion. The
functionally important amino acids of the ATP6 subunit,
including the polar residues comprising the proton channel
and residues in contact with nuclear subunits, are quite
conserved between humans and E. coli (Schon et al. 2001)
and Bombina (data not shown). The L-P substitution in B.
variegata, however, is clearly located outside of the
transmembrane helices and does not participate in nuclear-
mitochondrial interactions (Table 3, Supplementary Mate-
rial S7 [cf. Schon et al. 2001]), thus it probably does not
affect the functioning of the ATP synthase.
The remaining radical amino acid replacements
occurred in the ND proteins of ETS complex I. This
large, L-shaped enzyme is composed of 46 subunits, 7 of
which are hydrophobic central subunits encoded by
mtDNA (Brandt 2006). The molecular structure of this
complex remains unresolved, and how amino acid varia-
tion in specific subunits affects the functioning of the
complex is poorly known. The transmembrane domain
predictors applied to the Bombina complex I proteins
(Supplementary Material S8–S10) were often at odds as
to which amino acid residues constitute transmembrane
helices. Nonetheless, the domains in which the radical
replacement changes occurred were always well defined
and, thus, can be used as rough estimates of their loca-
tion. A single radical substitution, invoking a change from
a hydrophobic to a hydrophilic amino acid (L-S), was
detected in a transmembrane helix of ND5 in B. varieg-
ata. Other amino acid differences were located in
domains putatively outside of the transmembrane seg-
ments. Three involved proline, an amino acid with a
cyclic structure known to occur in the turns of polypep-
tide chains, and could therefore alter the secondary
structure of these surface domains. A total of three radical
substitutions occurred in a large (ca. 76 residues, positions
502–586; Supplementary Material S10) extramembrane
domain of the C-terminus of ND5.
Discussion
Despite the high degree of nucleotide divergence between
the Bombina mitochondrial genomes (up to 14% between
the two East Asian species) and their closest relatives (26%
between Discoglossus/Bombina and Alytes/Bombina), the
mitochondrial genomes of these amphibians retained
identical gene order and content and similar nucleotide
composition over the immense expanse of evolutionary
time since their divergence (during the late Triassic/early
Jurassic according to recent molecular estimates [Roelants
et al. 2007; San Mauro et al. 2004a]). The most divergent
mtDNA of the four species compared was that of B. for-
tinuptialis, which, together with B. maxima and B.
microdeladigitora, belongs to the group of large-bodied
Bombina species of Southeast Asia (Grobina). The ancient
split between the large-bodied and the small-bodied Bom-
bina species (B. bombina, B. variegata, and B. orientalis) is
supported by the grouping of B. orientalis with the Euro-
pean species in our mtDNA phylogeny, as well as by
karyotypic differences, i.e., the large-bodied species bear a
diploid chromosome complement of 2N = 28, while the
small-bodied species contain 2N = 24 (Szymura and Pas-
sakas-Szymczak 1988). B. orientalis, despite having a
distribution that is geographically proximate to the range of
B. fortinuptialis, is phylogenetically closer to the Western
Palearctic Bombina species than to its East Asian relatives.
Analysis by Yu et al. (2007) of DNA sequences from
several mitochondrial genes for Grobina and several
European and Asian Bombina species also supports this
grouping.
The average nucleotide divergence in mtDNA between
the two hybridizing species, B. bombina and B. variegata,
was 8.1 (8.7% K2P). This figure is congruent with earlier
estimates based on restriction enzymes (9.4% ± 1%
[Szymura et al. 1985], 6.0–8.1% [Szymura et al. 2000]). In
a different study, sequence divergence in Cytb haplotypes
between 62 individuals of both species amounted to
9.2% ± 0.2% K2P (Hofman and Szymura 2007), a value
slightly above the average for the entire mitogenome. The
nuclear sequence divergence between B. bombina and B.
variegata is lower than this and has been estimated as 0–
8% (Nurnberger et al. 2003) for several homologous,
putatively noncoding loci, 3.7% (3.8% K2P) for a partial
coding fragment of histone H3a (Frost et al. 2006; Gen-
Bank accession numbers DQ284275 and DQ284274 for B.
bombina and B. variegata, respectively), and 0.03%
(0.03% K2P) in exon 1 of the rhodopsin gene (Frost et al.
2006; DQ283920 and DQ283919 for B. bombina and B.
variegata, respectively). The divergence in 29 allozyme
loci between B. bombina and representatives of the major
subgroups of B. variegata, measured as Nei’s D distance,
ranged between 0.37 and 0.59 (Szymura 1993).
The Bombina control region contains highly conserved
regions, such as the CSBs and pyrimidine-rich regions, and
also length-variable regions (LV1 and LV2) comprised of
tandemly arranged repeat units. The conserved sequence
blocks (CSBs) and pyrimidine-rich stretches (PPs) possibly
J Mol Evol (2008) 67:246–256 253
123

play a role in transcription of mtDNA-encoded proteins
and H-strand replication in amphibians (San Mauro et al.
2004b). In Xenopus laevis, bidirectional promotors and
transcription factor binding sites have been localized
upstream of CSB2 and CSB3 (Antoshechkin and Bogen-
hagen 1995). These regions of high sequence conservation
are preceded by the first repeat region, LV1, consisting of
repeat units that are alignable in Bombina (Fig. 2B) and,
also, among archeobatrachians (San Mauro et al. 2004a),
suggesting homology. This sequence similarity indicates
functional constraint that may result from the presence of a
putative termination-associated sequence (TAS) in the LV1
repeat unit. TASs terminate H-strand synthesis and thus
produce the D-loop-containing form of mtDNA. The
presence of repeats that contain TAS sequences and pos-
sibly other regulatory elements downstream of LV1 raises
the possibility of a fine-tuning of mitochondrial metabo-
lism (through the regulation of mtDNA replication/
transcription) by the number of the repeat units in the
control regions, as suggested for lagomorph mtDNA (Ca-
sane et al. 1997). In contrast, the LV2 unit, tandemly
repeated at the 30 end of the Bombina control region, is
restricted to the genus Bombina and has been secondarily
lost in B. bombina. Apparently, the LV2 region in Bombina
is under less selective constraint than LV1, as evidenced by
the lack of sequence similarity to other archeobatrachian 30
repeat motifs (San Mauro et al. 2004a), its absence in B.
bombina, and no known regulatory function.
The varying dN/dS ratios observed among the protein
coding mitochondrial genes in Bombina suggest generally
high but, nonetheless, unequal levels of stringency of
structural and functional constraint. Overall, our results are
consistent with previous studies examining substitution
patterns among mitochondrial protein coding genes, in
which genes encoding proteins of ETS complexes III, IV,
and V were always much more conserved than those in
ETS complex I (e.g., Pesole et al. 1999; Doiron et al.
2002). Exceptional are perhaps the high dN/dS ratio in ND3
and relatively low ratios for ATP6 and ATP8 in Bombina.
Low ratios for ATP8 have also been documented in several
fish species (Roques et al. 2006 and references therein).
Closer inspection of the species-specific, nonsynony-
mous substitutions between the hybridizing European
Bombina species revealed that the majority can be
considered functionally neutral, yet 9 of the 98 nonsyn-
onymous substitutions between B. bombina and B.
variegata may have functional consequences. All but one
occur in ETS complex I, for which only a limited amount
of structural information is available. Moreover, the radical
replacement substitutions observed in ND4 and ND5
coincide with elevated KA/KS ratios. Both radical replace-
ments and high dN/dS ratios have been regarded as the
hallmarks of coadaptation between mitochondrial and
nuclear ETS subunits (Wu et al. 2000; Schmidt et al.
2001; Doiron et al. 2002) or evidence for the action of
positive selection (Mishmar et al. 2003; 2006). A possi-
ble alternative explanation is that the elevated rate of
amino acid substitution results from relaxed selective
constraint on sites at which the need for amino acid
conservation is lower (Elson et al 2004; Ingman and
Gyllensten 2007).
Mitochondrially encoded and nDNA-encoded proteins
build functional units constituting the mitochondrial ETS,
whereas species-specific promotor regions in mtDNA
probably coevolve with nDNA-encoded peptides. Muta-
tions in either genomic component influence the evolution
of the other, producing coadapted complexes (Blier et al.
2001). These associations are particularly prone to dis-
ruption in species hybrids (Sackton et al. 2003; Zeyl et al.
2005; Ellison and Burton 2008). Despite ongoing hybrid-
ization in spatially and temporally variable hybrid zones,
Hofman et al. (2007) have found no evidence for either
past or present mtDNA introgression between European
Bombina, which is consistent with the idea that later-gen-
eration hybrids are less fit because of cytonuclear
incompatibility leading to the disruption of mitochondrial
function (Burton et al. 2006). In this paper we have shown
that there are ample differences between the mtDNA
genomes of B. bombina and B. variegata that may have
functional consequences affecting mitochondrial oxidative
phosphorylation in hybrids. Further studies should incor-
porate experimental verification of our results through, e.g.,
in vitro assays of the efficiency of ETS complexes in
hybrid and nonhybrid individuals.
Acknowledgments The study was supported by grants from the
American Philosophical Society to J.M.S. and KBN Grant 2PO4C
087 29 to J.M.S. and M.P.
References
Abascal F, Zardoya R, Posada D (2005) ProtTest: selection of best-fit
models of protein evolution. Bioinformatics 12:2104–2105
Antoshechkin I, Bogenhagen DF (1995) Distinct roles for two purified
factors in transcription of Xenopus mitochondrial DNA. Mol
Cell Biol 15:7032–7042
Ballard JWO, Rand DM (2005) The population biology of mitochon-
drial DNA and its phylogenetic implications. Annu Rev Ecol
Evol 36:621–642
Ballard JWO, Whitlock MC (2004) The incomplete natural history of
mitochondria. Mol Ecol 13:729–744
Blier PU, Dufresne F, Burton R (2001) Natural selection and the
evolution of mtDNA-encoded peptides: insights from studies of
protein function and cytonuclear coadaptation. Trends Genet
17:400–406
Boore JL (1999) Animal mitochondrial genomes. Nucleic Acids Res
27:1767–1780
Brandt U (2006) Energy converting NADH: quinone oxidoreductase
(Complex I). Annu Rev Biochem 75:69–92
254 J Mol Evol (2008) 67:246–256
123

Burton RS, Ellison CK, Harrison JS (2006) The sorry state of F2
hybrids: consequences of rapid mitochondrial DNA evolution in
allopatric populations. Am Nat 168:S14–S24
Casane D, Dennebouy N, de Rochambeau H, Mounolou JC,
Monnerot M (1997) Nonneutral evolution of tandem repeats in
the mitochondrial DNA control region of Lagomorphs. Mol Biol
Evol 14:779–789
Doiron SL, Bernatchez L, Blier PU (2002) A comparative mitoge-
nomic analysis of the potential adaptive value of arctic charr
mtDNA introgression in brook charr populations (Salvelinusfontinalis Mitchill). Mol Biol Evol 19:1902–1909
Ellison CK, Burton RS (2006) Disruption of mitochondrial function
in interpopulation hybrids of Tigriopus californicus. Evolution
60:1382–1391
Ellison CK, Burton RS (2008) Interpopulation hybrid breakdown
maps to the mitochondrial genome. Evolution 62–63:631–638
Elson JL, Turnbull DM, Howell N (2004) Comparative genomics and
the evolution of human mitochondrial DNA: assessing the
effects of selection. Am J Hum Genet 74:229–238
Felsenstein J (2004) Inferring phylogenies. Sinauer Associates,
Sunderland, MA
Fromhage L, Vences M, Veith M (2004) Testing alternative
vicariance scenarios in Western Mediterranean discoglossid
frogs. Mol Phylogenet Evol 31:308–322
Frost DR, Grant T, Faivovich J, Bain RH, Haas A, Haddad CFB, De
Sa RO, Channing A, Wilkinson M, Donnelan SC, Raxworthy CJ,
Campbell JA, Blotto BL, Moler P, Drewes RC, Nussbaum RA,
Lynch JD, Green DM, Wheeler WC (2006) The amphibian tree
of life. Bull Am Mus Nat His 297:1–370
Gaspari M, Falkenberg M, Larsson N-G, Gustafsson CM (2004) The
mitochondrial RNA polymerase contributes critically to pro-
moter specificity in mammalian cells. EMBO J 23:4606–4614
Gemmell NJ, Metcalf VJ, Allendorf FW (2004) Mother’s curse: the
effect of mtDNA on individual fitness and population viability.
Trends Ecol Evol 19:238–244
Guindon S, Gascuel O (2003) PHYML—a simple, fast, and accurate
algorithm to estimate large phylogenies by maximum likelihood.
Syst Biol 52:696–704
Hofman S, Szymura JM (2007) Limited mtDNA introgression in a
Bombina hybrid zone. Biol J Linn Soc 91:295–306
Hofman S, Spolsky C, Uzzell T, Cogalniceanu D, Babik W, Szymura
JM (2007) Phylogeography of the fire-bellied toads, Bombina:
independent Pleistocene histories inferred from mitochondrial
genomes. Mol Ecol 16:2301–2316
Huelsenbeck JP, Ronquist F (2001) MrBayes: Bayesian inference of
phylogeny. Bioinformatics 17:754–755
Igawa T, Kurabayashi A, Usuki C, Fujii T, Sumida M (2008)
Complete mitochondrial genomes of three neobatrachian anu-
rans: a case study of divergence time estimation using different
data and calibration settings. Gene 407:116–129
Ingman M, Gyllensten U (2007) Rate variation between mitochon-
drial domains and adaptive evolution in humans. Hum Mol
Genet 16:2281–2287
Jones DT, Taylor WR, Thornton JM (1994) A mutation data matrix
for transmembrane proteins. FEBS Lett 339:269–275
Kall L, Krogh A, Sonnhammer ELL (2005) An HMM posterior
decoder for sequence feature prediction that includes homology
information. Bioinformatics 21:i251–i257
Krogh A, Larsson B, von Heijne G, Sonnhammer ELL (2001)
Predicting transmembrane protein topology with a hidden Markov
model: application to complete genomes. J Mol Biol 305:567–580
Kruuk LEB, Gilchrist JS, Barton NH (1999) Hybrid dysfunction in
fire-bellied toads (Bombina). Evolution 53:1611–1616
Kumar S, Tamura K, Nei M (2004) MEGA3: integrated software for
molecular evolutionary genetics analysis and sequence align-
ment. Brief Bioinform 5:150–163
Larsson NG, Clayton DA (1995) Molecular genetic aspects of human
mitochondrial disorders. Annu Rev Genet 29:151–178
McKenzie M, Chiotis M, Pinkert CA, Trounce IA (2003) Functional
respiratory chain analyses in murid xenomitochondrial cybrids
expose coevolutionary constraints of cytochrome b and nuclear
subunits of complex III. Mol Biol Evol 20:1117–1124
Mishmar D, Ruiz-Pesini E, Golik P, Macaulay V, Clark AG, Hosseini
S, Brandon M, Easley K, Chen E, Brown MD, Sukernik RI,
Olckers A, Wallace DC (2003) Natural selection shaped regional
mtDNA variation in humans. Proc Natl Acad Sci USA 100:
171–176
Mishmar D, Ruiz-Pesini E, Mondragon-Palomino M, Procaccio V,
Gaut B, Wallace DC (2006) Adaptive selection of mitochondrial
complex I subunits during primate radiation. Gene 378:11–18
Nei M, Gojobori T (1986) Simple methods for estimating the numbers
of synonymous and nonsynonymous nucleotide substitutions.
Mol Biol Evol 3:418–426
Nurnberger B, Hofman S, Forg-Brey G, Praetzel G, Maclean A,
Szymura JM, Abbott CM, Barton NH (2003) A linkage map for
the hybridising toads Bombina bombina and B. variegata (Anura:
Discoglossidae). Heredity 91:136–142
Pesole G, Gissi C, De Chirico A, Saccone C (1999) Nucleotide
substitution rate of mammalian mitochondrial genomes. J Mol
Evol 48:427–434
Posada D, Crandall KA (1998) Modeltest: testing the model of DNA
substitution. Bioinformatics 14:817–818
Roelants K, Gower DJ, Wilkinson M, Loader SP, Biju SD, Guillaume
K, Moriau L, Bossuyt F (2007) Global patterns of diversification
in the history of modern amphibians. Proc Natl Acad Sci USA
104:887–892
Roques S, Fox CJ, Villasana MI, Rico C (2006) The complete
mitochondrial genome of the whiting, Merlangius merlangus and
the haddock, Melanogrammus aeglefinus: a detailed genomic
comparison among closely related species of the Gadidae family.
Gene 383:12–23
Rozas J, Sanchez-DelBarrio JC, Messeguer X, Rozas R (2003)
DnaSP, DNA polymorphism analyses by the coalescent and
other methods. Bioinformatics 19:2496–2497
Sackton TB, Haney RA, Rand DM (2003) Cytonuclear coadaptation
in Drosophila: disruption of cytochrome c oxidase activity in
backcrossgenotypes. Evolution 57:2315–2325
San Mauro D, Garcıa-Parıs M, Zardoya R (2004a) Phylogenetic
relationships of discoglossid frogs (Amphibia: Anura: Disco-
glossidae) based on complete mitochondrial genomes and
nuclear genes. Gene 343:357–366
San Mauro D, Gower DJ, Oommen OV, Wilkinson M, Zardoya R
(2004b) Phylogeny of caecilian amphibians (Gymnophiona)
based on complete mitochondrial genomes and nuclear RAG1.
Mol Phylogenet Evol 33:413–427
Schapira AHV (2006) Mitochondrial disease. Lancet 368:70–82
Schmidt TR, Wu W, Goodman M, Grossman LI (2001) Evolution of
nuclear- and mitochondrial-encoded subunit interaction in cyto-
chrome c oxidase. Mol Biol Evol 18:563–569
Schon EA, Santra S, Pallotti F, Girvin ME (2001) Pathogenesis of
primary defects in mitochondrial ATP synthesis. Cell Dev Biol
12:441–448
Spolsky CM, Szymura JM, Uzzell T (2006) Mapping Bombinamitochondrial genomes: the conundrum of Carpathian Bombinavariegata (Anura: Discoglossidae). J Zool Syst Evol Res
44:100–104
Szymura JM (1993) Analysis of hybrid zones with Bombina. In:
Harrison R (ed) Hybrid zones and the evolutionary process.
Oxford University Press, New York
Szymura JM, Barton NH (1986) Genetic analysis of a hybrid zone
between fire-bellied toads, Bombina bombina and B. variegata,
near Cracow in southern Poland. Evolution 40:1141–1159
J Mol Evol (2008) 67:246–256 255
123

Szymura JM, Barton NH (1991) The genetic structure of the hybrid
zone between the fire-bellied toads Bombina bombina and B.variegata: comparisons between transects and between loci.
Evolution 45:237–261
Szymura JM, Passakas-Szymczak T (1988) A new chromosome
number for Bombina (Anura, Discoglossidae). Experientia
44:521–523
Szymura JM, Spolsky C, Uzzell T (1985) Concordant change in
mitochondrial and nuclear genes in a hybrid zone between two
frog species (genus Bombina). Experientia 41:1469–1470
Szymura JM, Uzzell T, Spolsky C (2000) Mitochondrial DNA
variation in the hybridizing fire-bellied toads, Bombina bombinaand B. variegata. Mol Ecol 9:891–899
Wu W, Schmidt TR, Goodman M, Grossman LI (2000) Molecular
evolution of cytochrome c oxidase subunit I in primates: is there
coevolution between mitochondrial and nuclear genomes? Mol
Phylogenet Evol 17:294–304
Willet CS, Burton RS (2004) Evolution of interacting proteins in the
mitochondrial electron transport system in a marine copepod.
Mol Biol Evol 21:443–453
Yanchukov AW, Hofman S, Szymura JM, Mezhzherin SV, Morozov-
Leonov SY, Barton NH, Nurnberger B (2006) Hybridization of
Bombina bombina and B. variegata (Anura, Discoglossidae) at a
sharp ecotone in western Ukraine: comparisons across transects
and over time. Evolution 60:583–600
Yu G, Yang J, Zhang M, Rao D (2007) Phylogenetic and systematic
study of the genus Bombina (Amphibia: Anura: Bombinatori-
dae): new insights from molecular data. J Herpetol 41:365–377
Zeyl C, Andreson B, Weninck E (2005) Nuclear-mitochondrial
epistasis for fitness in Saccharomyces cerevisiae. Evolution
59:910–914
Zhang P, Zhou H, Chen Y-Q, Kiu Y-F, Qu L-H (2005) Mitogenomic
perspectives on the origin and phylogeny of living amphibians.
Syst Biol 54:391–400
256 J Mol Evol (2008) 67:246–256
123