comorbidity between epilepsy and depression: role of hippocampal interleukin-1beta
TRANSCRIPT

Comorbidity between epilepsy and depression: role ofhippocampal interleukin-1β
Andrey M. Mazarati, MD, PhD1, Eduardo Pineda, PhD1, Don Shin, B.Sci1, Delia Tio, B.Sci2,Anna N. Taylor, PhD2, and Raman Sankar, MD, PhD1,31Department of Pediatrics, David Geffen School of Medicine at the University of California LosAngeles2Department of Neurobiology, David Geffen School of Medicine at the University of California LosAngeles3Department of Neurology, David Geffen School of Medicine at the University of California LosAngeles
AbstractDepression is a frequent comorbidity of temporal lobe epilepsy (TLE); however its mechanismsremain poorly understood and effective therapies are lacking. Augmentation of hippocampalinterleukin-1β (IL-1β) signaling may be a mechanistic factor of both TLE and clinical depression.We examined whether pharmacological blockade of hippocampal interleukin-1 receptor exertsantidepressant effects in an animal model of comorbidity between TLE and depression, whichdeveloped in Wistar rats following pilocarpine status epilepticus (SE). In post-SE animals,depression-like state was characterized by behavioral equivalents of anhedonia and despair;dysregulation of the hypothalamo-pituitary-adrenocortical axis; compromised raphe-hippocampalserotonergic transmission. Two-week long bilateral intrahippocampal infusion of humanrecombinant Interleukin-1 Receptor antagonist (IL-1ra) improved all of the examined depressiveimpairments, without modifying spontaneous seizure frequency and without affecting normalparameters in naïve rats. These findings implicate hippocampal IL-1β in epilepsy-associateddepression, and provide a rationale for the introduction of IL-1β blockers in the treatment ofdepression in TLE.
KeywordsTemporal lobe epilepsy; depression; comorbidity; brain inflammation; Interleukin-1β; hippocampus
IntroductionDepression represents one of the most common comorbidities of temporal lobe epilepsy (TLE),and has profound negative impact on the quality of life of TLE patients (Kanner, 2003;Kondziella et al., 2007). However, the causes and mechanisms of depression in TLE remain
© 2009 Elsevier Inc. All rights reservedCorresponding author: Andrey Mazarati, MD, PhD Dept. of Pediatrics, Neurology Division D. Geffen School of Medicine at UCLABOX 951752, 22-474 MDCC Los Angeles, CA 90095-1752 Phone: 1-310-206-5198 Fax: 1-310-825-5834 [email protected]'s Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resultingproof before it is published in its final citable form. Please note that during the production process errors may be discovered which couldaffect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptNeurobiol Dis. Author manuscript; available in PMC 2011 February 1.
Published in final edited form as:Neurobiol Dis. 2010 February ; 37(2): 461. doi:10.1016/j.nbd.2009.11.001.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

poorly understood, partly due to the lack of proper animal models. We previously reported thatrats which develop chronic epilepsy following pilocarpine status epilepticus (SE) exhibited setof interictal disorders congruent with depression. Specifically, the following depressiveimpairments were documented in post-SE animals: behavioral equivalents of anhedonia (i.e.loss of the ability to experience pleasure) and despair; dysregulation of hypothalamo-pituitary-adrenocortical (HPA) axis; compromised raphe-hippocampal serotonergic transmission(Mazarati et al., 2008; 2009a). As the pilocarpine model reproduces both epileptic anddepressive states, it may serve as a model of comorbidity between TLE and depression and assuch, can be used both for studying mechanisms of this condition and as a screening platformfor therapeutic interventions. Since depression is a multisymptomatic and a multifactorialdisorder, further studies are necessary to advance the validation of this model; however theimpairments established up-to-date allow initial experimental therapy studies.
One factor which may contribute to depression in TLE is hippocampal tissue inflammation,and particularly, enhanced interleukin-1β (IL-1β) signaling. Indeed, activation of hippocampalIL-1β and its receptor (IL-1R) have been established hallmarks of TLE both in clinical andexperimental settings, and have been implicated in mechanisms of epileptogenesis (Bartfai etal., 2007; Ravizza et al., 2008; Vezzani et al., 2002; 2008; Vezzani and Granata, 2005). At thesame time, IL-1β and other inflammatory cytokines may lead to depression conceivably viainducing perturbation in the HPA axis, as suggested by clinical observations, and confirmedby experimental studies (Capuron and Dantzer, 2003; Dunn et al., 2005; Leonard, 2006).Particularly, the dysregulation of the HPA axis (which represents a neuroendocrine hallmarkof depression; Kondziella et al., 2007; Swaab et al., 2005) can be induced by the directactivation of hippocampal IL-1β signaling (Melik Parsadaniantz et al., 1999).
Such connections between epilepsy and IL-1β on the one hand, and IL-1β and depression onthe other hand, prompted us to examine whether protracted pharmacological blockade ofhippocampal IL-1R exerts antidepressant effect in the post-SE model of comorbidity betweenepilepsy and depression.
MethodsSubjects
The experiments were performed in male Wistar rats (Charles River, Wilmington, MA), fiftydays old at the beginning of the study, in accordance with the policies of the National Institutesof Health.
Study design is outlined in Fig. 1. Detailed description of procedures is provided in earlierpublications (Mazarati et al., 2008;2009a).
Forced swim test (FST)FST allows examining state of despair, and is based on the innate ability of rats to adopt activestrategies in the inescapable stressful situation (Pucilowski and Overstreet, 1993). FSTconsisted of a single five-minute swimming session in the tank (height - 60 cm, diameter 30cm) filled with water at 22°–25°C. Swimming behavior was videotaped and analyzed offline.Cumulative immobility time (Fig. 2A) was calculated. The increased immobility time indicatesthe state of despair (Mazarati et al., 2008; Pucilowski and Overstreet, 1993).
Taste preference testThis test for anhedonia is based on the innate preference of rodents towards sweets (Pucilowskiet al., 1993). Taste preference was examined using saccharin solution consumption test(Mazarati et al., 2008; Pucilowski et al., 1993). The rat's cage was supplied with two 250 ml
Mazarati et al. Page 2
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

graduated bottles, one filled with water and another – with 0.1% saccharin solution. Tastepreference was expressed as percent of the volume of saccharin solution of a total volume offluid (saccharin plus water) consumed over 24 hours. The loss of preference for saccharin (i.e.consumption of statistically equal volumes of saccharin and water) is indicative of anhedonia(Mazarati et al., 2008; Pucilowski et al., 1993).
Plasma corticosterone (CORT) assayMeasurement of baseline plasma CORT level and dexamethasone/corticotropin releasinghormone (DEX/CRH) test were performed upon the completion of behavioral tests (Johnsonet al., 2006; Mazarati et al., 2009a; Pohorecky et al., 2004; Steimer et al., 2007). Between 8:00AM and 10:00 AM, 50–100 μl of blood was collected from the tail vein into the EDTA – coatedtubes. The animals were then injected into the tail vein with DEX (Sigma; 0.03 mg/kg). Sixhours later blood was collected again, and animals were injected into the tail vein with CRH(Sigma; 50 ng/kg); two blood samples were taken 30 and 60 min after CRH injection. CORTwas detected in 10 l plasma samples, using Immunochem™ Double Antibody Corticosterone125I RIA kit (MP Biomedicals, Orangeburg, NY). Dysregulation of the HPA axis in depressionconsists of the elevated baseline CORT level; failure of DEX to suppress CORT; exacerbatedand prolonged increase of CORT in response to CRH (the latter two impairments constitutepositive DEX/CRH test) (Johnson et al., 2006; Mazarati et al., 2008; 2009a; Pohorecky et al.,2004; Zobel et al., 2004).
Status epilepticus (SE)Two days after DEX/CRH test, animals received intraperitoneal injection of LiCl (130 mg/kg,Sigma, St. Louis, MO) and twenty four hours later - subcutaneous injection of pilocarpine (40mg/kg, Sigma). Behavioral seizures were monitored during SE. SE was characterized bycontinuous limbic seizures starting 10–15 min after pilocarpine injection. Three and eight hoursafter seizure onset, rats were injected intraperitoneally with diazepam (5 mg/kg) and phenytoin(50 mg/kg) in order to alleviate further seizures and to decrease mortality. In control animalspilocarpine was substituted with saline.
Monitoring of spontaneous seizuresDue to the complexity of the drug delivery system, monitoring of spontaneous seizures waslimited to video. Beginning from 8 weeks after SE, and until the end of experiments, animals'behavior was continuously recorded using digital camera. Seizures were analyzed off-line.Two seizure types were considered: focal seizures (motor arrest, facial twitches andmastication), and generalized clonic or clonic-tonic seizures (all body clonus, rearing or rearingand falling) (Mazarati et al., 2009a). Seizure monitoring served two purposes: first – for theselection of proper time points for the FST and CORT assays (as described below); second –for the evaluation of effects of treatment on seizure frequency. For the second purpose, thecumulative number of spontaneous seizures was calculated during two consecutive two-weekperiods: one immediately preceding the start of IL-1ra treatment, and another during the IL-1ratreatment. Data obtained for untreated post-SE rats as well as for rats treated with IL1-ra orsaline were analyzed for each of the groups by comparing seizure counts before treatmentswith those during treatments.
Treatment with Interleukin-1 receptor antagonist (IL-1ra)On order to achieve sustained blockade of hippocampal IL-1R, we used continuousintrahippocampal delivery of human recombinant IL-1ra (Amgen, Thousand Oaks, CA,Vezzani et al., 1999; Vezzani et al., 2000) by means of ALZET osmotic pumps. Earlier studiesproved that IL-1ra preserves its stability and physiological activity when used in conjunctionwith the ALZET pump at 37°C for as long as four weeks (Knedla et al., 2009; Sandberg et al.,
Mazarati et al. Page 3
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

1993); after two weeks of incubation in the ALZET pump at 37°C, IL-1ra retained nearly 100%of its initial activity (Knedla et al., 2009).
Ten weeks after SE, animals underwent surgical implantation of system for continuousintrahippocampal infusion of IL-1ra. The system (Durect Corporation, Cupertino, CA)consisted of the ALZET osmotic pump model 2002 (delivery rate 0.5 μl/hr, duration 14 days,volume 200 μl) connected to the guide cannula via a polypropylene catheter (Brain InfusionKit I). Osmotic pumps had been filled with the 140 μg of IL-1ra diluted in sterile saline, sothat animals received 10 μg of the drug per day; the dose was based on earlier reports (Vezzaniet al., 1999; 2000, 2002). The pumps then were activated and primed in sterile saline for 24hours at 37°C according to the manufacturer's recommendation. Afterwards, each animal wasimplanted with two systems to provide bilateral infusion. Under isoflurane anesthesia, guidecannulas were stereotaxically implanted bilaterally into the hippocampi (4.3 mm posterior and4 mm lateral from Bregma, 3.8 mm ventral from brain surface). Osmotic pumps were placedsubcutaneously between the shoulders. Control groups included untreated animals; animalsinfused with the heat-inactivated IL-1ra (exposed to 70°C water bath for 30 min; 140 μg in 0.2ml); and animals infused with saline (0.2 ml; both control treatments were administered usingsame delivery system as in experimental subjects). At the end of drug delivery (i.e. just beforefast cyclic voltammetry experiments described below), pumps were removed, and the residualvolume of content was aspirated and measured, in order to confirm the delivery of a test agent.The residual volume in the pumps was between 0 and 20 μl (i.e. did not exceed 10% of theoriginal volume).
Behavioral test and CORT assay during IL-1ra treatmentDuring the days 10–13 after the beginning of IL-1ra infusion, the animals were subjected toFST, taste preference test and plasma CORT assay. To avoid possible immediate effects ofseizures on FST, the test was only performed upon verification (by reviewing video recordings)that no seizures had developed for at least 6 hours prior to the test. For CORT assay, bloodwas only collected upon verification that animals had not developed seizures for at least 6 hoursprior (Mazarati et al., 2009a).
Fast cyclic voltammetry (FCV)FCV allows measuring strength of the raphe-hippocampal serotonergic transmission (Buninand Wightman, 1998; Jackson et al., 1995; Mazarati et al., 2008); it was preformed 14 daysafter the beginning of IL-1ra administration. Under urethane anesthesia (1.25 g/kg), animalswere implanted with a bipolar electrode into dorsal raphe (7.8 mm posterior from Bregma,midline, 6.5 mm ventral from brain surface) and with a nafion-coated carbon fiber electrode(World Precision Instruments, Sarasota, FL) into the hippocampus (4.3 mm posterior toBregma, 3 mm lateral from midline, 3.6 mm down from brain surface). The release of serotonin(5-HT) in the hippocampus was induced by electrical stimulation of raphe (bipolar square wavepulses, 100 Hz, 200 ms, 0.35 mA; Bunin and Wightman, 1998; Mazarati et al., 2008). Theamount of 5-HT released in response to the stimulation was measured by applying oxidizingcurrent to the carbon fiber electrode (scanned consecutively from 0.2 V to 1 V, −0.1 V and 0.2V, at a rate of 1000 V/s, Fig. 5B). Oxidative peaks were acquired before and after raphestimulation, using POT500 scanning potentiostat (World Precision Instruments) coupled withthe AcqKnowledge software (BIOPAC, Goleta, CA). The difference between peaks before andafter stimulation, known as faradaic current, reflects concentration of 5-HT oxidative productquinone and is directly proportionate to the amount of 5-HT (Wrona and Dryhurst, 1987).Compromised raphe-hippocampal serotonergic transmission (evident as the decrease of theamplitudes of faradaic currents) represents a biochemical hallmark of depression (Mann et al.,1989; Murphy et al., 1978). Since FCV is a non-survival procedure, it was done at one timepoint.
Mazarati et al. Page 4
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Data analysisData were analyzed using Prizm software (GraphPad, San Diego, CA). The study included thefollowing groups. Twenty four control animals (i.e. not subjected to SE), which subsequentlywere divided into IL-1ra-treated, saline-treated and untreated groups (8 animals per group).Twenty seven post-SE animals, also subsequently divided into IL-1ra-treated, saline-treatedand untreated groups (9 animals per group). Additionally, 6 post-SE rats were treated with theheat-inactivated IL-1ra. Statistical tests are described in respective figure legends.
ResultsEffects of IL-1ra on FST (Fig. 2B)
Treatment with IL-1ra had no effect on behavior in control animals as compared both withthemselves at the beginning of the study, and with saline-treated naïve rats. Untreated andsaline-treated post-SE animals, as well as those infused with the heat-inactivated IL1-ra,showed statistically significant increase in the immobility time during FST, as compared bothwith themselves prior to SE, and with respective controls.
Administration of IL-1ra to post-SE subjects resulted in significant shortening of theimmobility time, as compared with both untreated and saline-treated post-SE rats. However,immobility time remained significantly longer than in control subjects.
Effects of IL-1ra on taste preference (Fig. 3)Control and experimental animals prior to SE preferred saccharin solution over tap water withthe ratio of approximately 4:1. In control rats, IL-1ra did not affect preference towardssaccharin. Post-SE rats (untreated, treated with saline and with the heat-inactivated IL1-ra)consumed statistically equal amounts of saccharin and water. This redistribution betweensaccharin and water consumption was not accompanied by changes in the intake of total volumeof fluid (i.e. saccharin plus water; not shown). In post-SE animals, IL-1ra completely restoredpreference towards saccharin.
Since intrahippocampal infusion of heat-inactivated IL1-ra had not ameliorated behavioralsymptoms of depression, this control treatment was not included in further studies.
Effects of IL-1ra on the HPA axis (Fig. 4)In all control groups (i.e. IL-1ra, saline and untreated) baseline CORT level and the outcomeof DEX/CRH test were statistically similar during the first (i.e. at the beginning of the study)and the second assays (i.e. 12 weeks later). DEX decreased CORT level by 3.5 fold. CRHinduced the 4-fold rise in CORT at 30 min; the latter returned to the pre-CRH level at 60 min.
Both untreated and saline-treated post-SE animals showed interictal increase of baseline CORT(20% as compared with controls and themselves prior to SE). DEX failed to decreaseconcentration of CORT; after CRH injection, the rise of CORT was steeper (7-fold) and longer-lasting (present at 60 min) than in control rats. IL-1ra significantly decreased baseline CORTlevel in post-SE subjects as compared with saline-treated and untreated post-SE animals;CORT concentrations were statistically similar between IL-1ra – treated post-SE rats andcontrol subjects. IL-1ra partially restored the response to DEX and mitigated the exacerbatedresponse to CRH.
Effects of IL-1ra on 5-HT release from the hippocampus (Fig. 5)In controls, intrahippocampal infusion of IL-1ra did not alter amplitudes of faradaic currentsrecorded from the hippocampus after dorsal raphe stimulation. In untreated and saline-treated
Mazarati et al. Page 5
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

post-SE rats, the amplitude of faradaic currents was significantly lower than in controls, whichpointed towards compromised raphe-hippocampal serotonergic transmission. The latter waspartially restored by IL-1ra, although it was still significantly lower than in control subjects.
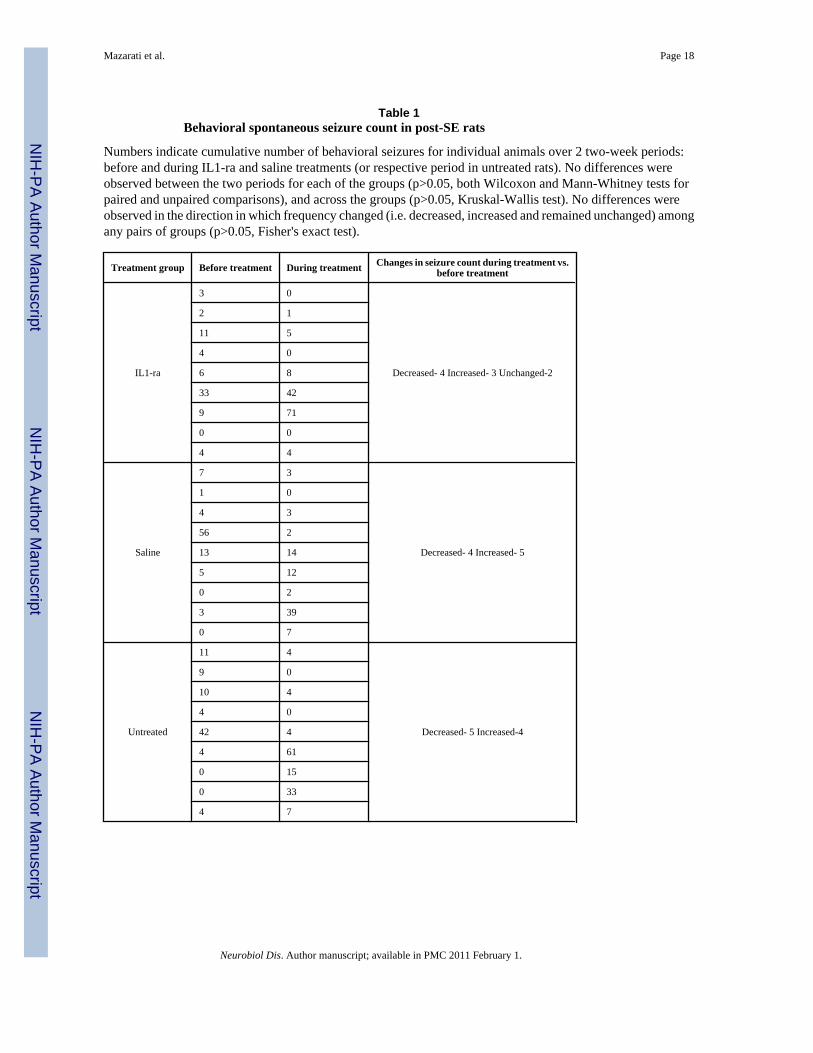
Effects of IL-1ra on seizures (Table 1)For all groups of post-SE animals (IL-1ra, saline and untreated) the number of seizuresdocumented before and during IL-1ra administration was statistically similar. All animalsexhibited both focal and generalized seizures; the latter type constituted at least 75% of alldocumented seizures. The distribution between the two seizure types was statistically similarbefore and during IL-1ra administration, as well as in untreated and saline-treated rats duringthe first and the second observation periods (data not shown). Furthermore, retrospectivecorrelation analysis did not reveal any statistical correlation between frequency of spontaneousseizures observed during the two weeks of treatment with IL-1ra and the immobility time inthe FST at the end of IL-1ra treatment (Spearmen coefficient of correlation r=−0.085, p>0.05).
Effects of different treatment regimensSince the administration of IL-1ra at 10 μg/day neither completely reversed all depressivesymptoms, nor reduced seizure frequency, we attempted to examine effects of the compoundat 30 μg/day (n=4). Two-three days after the onset of IL-1ra infusion, animals showed lack ofgrooming, motor exaltation (e.g. jumping in response to touch or moving the cage), andprogressive weight loss. For this reason, these studies were interrupted.
Another group of post-SE animals (n=5) was injected with IL-1ra acutely bilaterally into thehippocampus, using chronically implanted guide cannulas and Hamilton microsyringe, at 1μg (i.e. equivalent to the amount delivered over 2.5 hours in chronic experiments). The animalswere subjected to FST 2 weeks before and 30 min after IL-1ra injection. This treatment didnot improve behavioral deficit in the FST (not shown).
DiscussionProtracted blockade of hippocampal IL-1R attenuated behavioral, endocrine and biochemicalhallmarks of depression following SE. This effect did not appear to be a consequence of seizuremodification, since seizure frequency was neither altered by the treatment, nor statisticallycorrelated with the outcome of behavioral test under IL-1ra administration. IL-1ra improvedthe examined parameters of depression in epileptic rats selectively, without affecting responsesin naïve animals. Antidepressant action of IL-1ra was not due to an immediate blockade ofIL-1R, but apparently involved chronic downstream mechanisms, since acute drug injectionexerted no antidepressant effect.
The study confirmed our findings that post-SE animals exhibit interictal depression-likeimpairments (Mazarati et al., 2008; 2009a). Behavioral depressive deficits are not results ofnon-specific motor and sensory impairments induced by SE: despite the increased immobilitytime in the FST, and the loss of preference towards saccharin, SE impairs neither the animal'sability to swim, nor the normally present aversion to bitter quinine (Supplementary on-linematerial).
High incidence of depression among epilepsy patients, as well as reciprocal connectionbetween the two conditions has led to the hypothesis that depression and epilepsy share certainmechanisms (Kanner, 2003; Kondziella et al., 2007). We suggest that hippocampal IL-1β maybe a factor contributing to depression in TLE. Indeed, IL-1β signaling is enhanced in thehippocampus during the course of TLE, and may contribute to epileptogenesis (Bartfai et al.,2007; Ravizza et al., 2008; Vezzani and Granata, 2005). At the same time, brain inflammation,
Mazarati et al. Page 6
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

and particularly the augmentation of IL-1β signaling induced depression in experimentalsetting (Dunn et al., 2005). For example, inflammation induced both by lipopolysaccharideand by IL-1β was accompanied by behavioral symptoms of depression (Dunn and Swiergiel,2005; O'Connor et al., 2009; Yirmiya, 1996); lipopolysaccharide-induced depression wasreversed by IL-1ra (Konsman et al., 2008). Clinical observations also provide evidence for theassociation between inflammatory cytokines and depression (Capuron and Dantzer, 2003;Dunn et al., 2005; Leonard, 2006; van den Biggelaar et al., 2007).
Mechanisms through which the augmented hippocampal IL-1β signaling leads to depressionneed further investigation. The former has been proven to induce perturbations in the HPA axis(Melik Parsadaniantz et al., 1999), as seen under conditions of depression (Capuron andDantzer, 2003; Dunn et al., 2005; Leonard, 2006), and in post-SE animals (Mazarati et al.,2009a). In our studies, the dysregulation of the HPA axis following SE was mitigated by theblockade of hippocampal IL-1R. The observed effect of IL-1ra was consistent with previouslyreported increase of plasma CORT level by the IL-1β injected in the rat hippocampus (MelikParsadaniantz et al., 1999). As the DEX/CRH test reflects reduced inhibitory feedback controlwithin the HPA axis (Pariante and Miller, 2001; Watson et al., 2006), partial improvement ofthe response to DEX after intrahippocampal IL-1ra treatment was an apparent result of thenormalization of the afferent component of the HPA autoinhibitory loop (De Kloet and Reul,1987; Melik Parsadaniantz et al., 1999). Normalization of the response to CRH may beexplained based on the finding that the activation of hippocampal IL-1β stimulatedtranscription and processing of proopiomelanocortin (POMC, precursor of adrenocorticotropichormone, ACTH) in the anterior pituitary (Melik Parsadaniantz et al., 1999). It can be surmisedthat the sustained blockade of hippocampal IL-1β in post-SE subjects would mitigate theenhanced POMC transcription and cleavage; thus the administration of CRH to the IL-1ra –pretreated post-SE rats would target corticotropes in which ACTH production would have beennormalized, the latter being reflected in the mitigated rise in plasma CORT level.
Although the dysregulation of the HPA axis has been an accepted hallmark of clinicaldepression (Kondziella et al., 2007; Steimer et al., 2007; Watson et al., 2006; Zobel et al.,2004), exact mechanistic connection between the two are not known. One hypothesis links thedysregulation of the HPA axis to the deficiency of serotonergic transmission. Raphe-hippocampal serotonergic pathway plays important role in regulating mood and iscompromised in depression (Kondziella et al., 2007), as well as in the pilocarpine model(Mazarati et al., 2008). Among possible mechanisms involved in the raphe-hippocampalserotonergic deficiency in depression is the augmentation of mechanisms which inhibit 5-HTrelease. Specifically, depression is characterized by the upregulation of raphe 5-HT1Aautoreceptors (Boldrini et al., 2008; Stockmeier et al., 1998) – a component of short feedbackautoinhibitory loop involved in the control of 5-HT release (Aghajanian et al., 1990; Riad etal., 2000); thus the upregulation of raphe 5-HT1A receptors would result in the decrease of 5-HT release. The discussed mechanism may contribute to depression in TLE patients as well:recent study involving positron emission tomography using a selective 5-HT1A ligand showedincreased binding affinity of raphe 5-HT1A receptors in patients with TLE and concurrentdepression, and its positive correlation with the severity of clinical symptoms of depression(Lothe et al., 2008).
Enhanced level of circulating glucocorticoids may upregulate raphe 5-HT1A autoreceptors viaglucocorticoid receptor-mediated genomic mechanisms (Bellido et al., 2004; Man et al.,2002). The upregulation of raphe 5-HT1A autoreceptors and the involvement ofglucocorticoids in this process are yet to be shown directly in the pilocarpine model. However,our in-progress studies found that post-SE animals exhibited enhanced sensitivity to a selective5-HT1A blocker WAY100635 injected locally into raphe, as judged by effects of the compoundin the FST; furthermore, protracted pharmacological blockade of raphe glucocorticoid
Mazarati et al. Page 7
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

receptors by mifepristone improved behavioral symptoms of depression in post-SE rats withoutaffecting these parameters in naïve animals (Mazarati et al., 2009b; Supplementary on-linematerial) and reversed the enhanced sensitivity to the 5-HT1A ligand (personal unpublisheddata). These findings suggest that hypersensitization of raphe 5-HT1A receptors may be indeedoccurring in the pilocarpine model, and that increased exposure of raphe glucocorticoidreceptors to circulating CORT may contribute to the 5-HT1A hypersensitivity and depression.
Attenuation of depressive symptoms by IL-1ra was not an epiphenomenon of its anticonvulsanteffects. While this finding emphasizes the importance of IL-1β signaling as specific mechanismof depression associated with epilepsy, it may also be surprising, considering the discussedrole IL-1β in epileptogenesis. Several explanations can be offered on this matter. First, it shouldbe acknowledged that our experimental protocol may not be suitable for reliable examinationof antiepileptic therapies. In our laboratory the pilocarpine model is refined in such a way thatfrequency of recurrent seizures is relatively low due to aggressive anticonvulsant interventionsduring SE (Mazarati et al., 2008; 2009a) in order to afford examination of interictal phenomena,and as such it may prohibit adequate examination of antiepileptic treatments. Second, IL-1radid not affect severity of limbic seizures under intrahippocampal administration, although itwas anticonvulsant when administered into the lateral brain ventricle (Vezzani et al., 1999;2002), or systemically (Marchi et al., 2009); in such case, a broader exposure of brain to IL-1ramay be required for the drug to exert anticonvulsant effect. Third, it is plausible that a higherdose of IL-1ra would be antiepileptic; however this possibility could not be explored due tothe described side effects. On a related note, our finding that depression in post-SE animalscould not be completely abolished by IL-1ra, may be either due to the fact that the dose wasnot saturated to maximal therapeutic level, or that the exposure of hippocampal tissue to IL-1rawas limited to the sites of injection, or that other mechanisms (inflammatory and non-inflammatory alike) are involved in the described phenomenon. The latter explanations seemslikely: even considering inflammation only, other changes (e.g. proliferation of microglia,disruption of the blood-brain barrier; Ravizza et al., 2008; Vezzani and Granata, 2005) occurin TLE. Another inflammatory cytokine, tumor necrosis factor-α, has been directly implicatedin the enhanced neuronal excitability ensuing after a systemic inflammatory challenge resultingin decreased seizure threshold (Riazi et al 2008; Galic et al 2008).
The observed dissociation between antidepressant effects of IL-1ra and lack of its effects onseizure frequency is not surprising. Indeed, both clinical (Gilliam et al., 2007) and experimental(Mazarati et al., 2008) evidence suggest that the severity of depression in TLE does notnecessarily depend on frequency of seizures.
The importance of antidepressant action of IL-1ra is further emphasized by the fact that in thepilocarpine model, selective serotonin reuptake inhibitor (SSRI) fluoxetine was ineffective inmitigating depressive impairments (Mazarati et al., 2008). This finding on itself is notunexpected, as up to 1/3 of depression patients do not respond to SSRI (Barbui et al., 2002).In fact, our studies may offer an explanation as to the lack of antidepressant effects of fluoxetinein post-SE rats. A plausible mechanism through which SSRIs induce antidepressant effect isdesensitization of raphe 5-HT1A autoreceptors and subsequent strengthening of theraphehippocampal serotonergic transmission (Chaput et al., 1986; Le Poul et al., 1995). It isconceivable that the ongoing hippocampal inflammation and continuing positive drive of theHPA axis on raphe overrides fluoxetine-mediated desensitization of 5-HT1A autoreceptors. Insuch case, adjunctive therapy of IL-1R antagonists together with conventional antidepressantsmay be worth exploring.
In conclusion, the present study implicates hippocampal IL-1β in mechanisms of epilepsy-associated depression, and provides a rationale for the introduction of IL-1R antagonists in thetreatment of depression in TLE patients. Presently IL-1ra is used for the treatment of
Mazarati et al. Page 8
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

autoimmune diseases, such as rheumatoid arthritis (Kalliolias and Liossis, 2008), and its centralbioavailability in the healthy brain upon systemic administration is questionable. However,epilepsy is accompanied by the breakdown of the blood-brain barrier (BBB; Marcon et al.,2009; Ravizza et al., 2008; Rigau et al., 2007), which, in turn would likely render brain tissueaccessible to compounds, which normally do not permeate BBB. Indeed, experimental findingsshowed that peripherally administered IL-1ra exhibited neuroprotective effects in animalmodels of stroke and traumatic brain injury (Relton et al., 1996; Sanderson et al., 1999), thatis in conditions accompanied by the BBB leakage.
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgmentsThe work was supported by research grants MH079933 from the National Institutes of Health (A. Mazarati) and132081 from Epilepsy Foundation of America (A. Mazarati). IL-1ra was a gift from Amgen.
ReferencesAghajanian GK, Sprouse JS, Sheldon P, Rasmussen K. Electrophysiology of the central serotonin system:
receptor subtypes and transducer mechanisms. Ann N Y Acad Sci 1990;600:93–103. discussion 103.[PubMed: 2123618]
Barbui C, Hotopf M, Garattini S. Fluoxetine dose and outcome in antidepressant drug trials. Eur J ClinPharmacol 2002;58:379–386. [PubMed: 12242596]
Bartfai T, Sanchez-Alavez M, Andell-Jonsson S, Schultzberg M, Vezzani A, Danielsson E, Conti B.Interleukin-1 system in CNS stress: seizures, fever, and neurotrauma. Ann N Y Acad Sci2007;1113:173–177. [PubMed: 17656565]
Bellido I, Hansson AC, Gomez-Luque AJ, Andbjer B, Agnati LF, Fuxe K. Corticosterone stronglyincreases the affinity of dorsal raphe 5-HT1A receptors. Neuroreport 2004;15:1457–1459. [PubMed:15194873]
Boldrini M, Underwood MD, Mann JJ, Arango V. Serotonin-1A autoreceptor binding in the dorsal raphenucleus of depressed suicides. J Psychiatr Res 2008;42:433–442. [PubMed: 17574270]
Bunin MA, Wightman RM. Quantitative evaluation of 5-hydroxytryptamine (serotonin) neuronal releaseand uptake: an investigation of extrasynaptic transmission. J Neurosci 1998;18:4854–4860. [PubMed:9634551]
Capuron L, Dantzer R. Cytokines and depression: the need for a new paradigm. Brain Behav Immun2003;17(Suppl 1):S119–124. [PubMed: 12615197]
Chaput Y, de Montigny C, Blier P. Effects of a selective 5-HT reuptake blocker, citalopram, on thesensitivity of 5-HT autoreceptors: electrophysiological studies in the rat brain. Naunyn SchmiedebergsArch Pharmacol 1986;333:342–348. [PubMed: 3022157]
De Kloet ER, Reul JM. Feedback action and tonic influence of corticosteroids on brain function: a conceptarising from the heterogeneity of brain receptor systems. Psychoneuroendocrinology 1987;12:83–105.[PubMed: 3037584]
Dunn AJ, Swiergiel AH. Effects of interleukin-1 and endotoxin in the forced swim and tail suspensiontests in mice. Pharmacol Biochem Behav 2005;81:688–693. [PubMed: 15982728]
Dunn AJ, Swiergiel AH, de Beaurepaire R. Cytokines as mediators of depression: what can we learnfrom animal studies? Neurosci Biobehav Rev 2005;29:891–909. [PubMed: 15885777]
Galic MA, Riazi K, Heida JG, Mouihate A, Fournier NM, Spencer SJ, Kalynchuk LE, Teskey GC, PittmanQJ. Postnatal inflammation increases seizure susceptibility in adult rats. J Neurosci 2008;28:6904–6913. [PubMed: 18596165]
Gilliam FG, Maton BM, Martin RC, Sawrie SM, Faught RE, Hugg JW, Viikinsalo M, Kuzniecky RI.Hippocampal 1H-MRSI correlates with severity of depression symptoms in temporal lobe epilepsy.Neurology 2007;68:364–368. [PubMed: 17261683]
Mazarati et al. Page 9
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Jackson BP, Dietz SM, Wightman RM. Fast-scan cyclic voltammetry of 5-hydroxytryptamine. AnalChem 1995;67:1115–1120. [PubMed: 7717525]
Johnson SA, Fournier NM, Kalynchuk LE. Effect of different doses of corticosterone on depression-likebehavior and HPA axis responses to a novel stressor. Behav Brain Res 2006;168:280–288. [PubMed:16386319]
Kalliolias GD, Liossis SN. The future of the IL-1 receptor antagonist anakinra: from rheumatoid arthritisto adult-onset Still's disease and systemic-onset juvenile idiopathic arthritis. Expert Opin InvestigDrugs 2008;17:349–359.
Kanner AM. Depression in epilepsy: prevalence, clinical semiology, pathogenic mechanisms, andtreatment. Biol Psychiatry 2003;54:388–398. [PubMed: 12893113]
Knedla A, Riepl B, Lefevre S, Kistella S, Grifka J, Straub RH, Gay S, Scholmerich J, Muller-Ladner U,Neumann E. The therapeutic use of osmotic minipumps in the severe combined immunodeficiency(SCID) mouse model for rheumatoid arthritis. Ann Rheum Dis 2009;68:124–129. [PubMed:18339663]
Kondziella D, Alvestad S, Vaaler A, Sonnewald U. Which clinical and experimental data link temporallobe epilepsy with depression? J Neurochem 2007;103:2136–2152. [PubMed: 17887964]
Konsman JP, Veeneman J, Combe C, Poole S, Luheshi GN, Dantzer R. Central nervous action ofinterleukin-1 mediates activation of limbic structures and behavioural depression in response toperipheral administration of bacterial lipopolysaccharide. Eur J Neurosci 2008;28:2499–2510.[PubMed: 19087175]
Le Poul E, Laaris N, Doucet E, Laporte AM, Hamon M, Lanfumey L. Early desensitization of somato-dendritic 5-HT1A autoreceptors in rats treated with fluoxetine or paroxetine. Naunyn SchmiedebergsArch Pharmacol 1995;352:141–148. [PubMed: 7477436]
Leonard BE. HPA and immune axes in stress: involvement of the serotonergic system.Neuroimmunomodulation 2006;13:268–276. [PubMed: 17709948]
Lothe A, Didelot A, Hammers A, Costes N, Saoud M, Gilliam F, Ryvlin P. Comorbidity between temporallobe epilepsy and depression: a [18F]MPPF PET study. Brain 2008;131:2765–2782. [PubMed:18765418]
Man MS, Young AH, McAllister-Williams RH. Corticosterone modulation of somatodendritic 5-HT1Areceptor function in mice. J Psychopharmacol 2002;16:245–252. [PubMed: 12236633]
Mann JJ, Arango V, Marzuk PM, Theccanat S, Reis DJ. Evidence for the 5-HT hypothesis of suicide. Areview of post-mortem studies. Br J Psychiatry Suppl 1989:7–14. [PubMed: 2692642]
Marchi N, Fan Q, Ghosh C, Fazio V, Bertolini F, Betto G, Batra A, Carlton E, Najm I, Granata T, JanigroD. Antagonism of peripheral inflammation reduces the severity of status epilepticus. Neurobiol Dis2009;33:171–181. [PubMed: 19010416]
Marcon J, Gagliardi B, Balosso S, Maroso M, Noe F, Morin M, Lerner-Natoli M, Vezzani A, RavizzaT. Age-dependent vascular changes induced by status epilepticus in rat forebrain: implications forepileptogenesis. Neurobiol Dis 2009;34:121–132. [PubMed: 19320047]
Mazarati A, Siddarth P, Baldwin RA, Shin D, Caplan R, Sankar R. Depression after status epilepticus:behavioural and biochemical deficits and effects of fluoxetine. Brain 2008;131:2071–2083.[PubMed: 18559371]
Mazarati AM, Shin D, Kwon YS, Bragin A, Pineda E, Tio D, Taylor AN, Sankar R. Elevated plasmacorticosterone level and depressive behavior in experimental temporal lobe epilepsy. Neurobiol Dis2009a;34:457–461. [PubMed: 19285131]
Mazarati A, Shin D, Bragin A, Tio D, Taylor A, Sankar R. Hyperactivity of hypothalamo-pituitary-adrenocortical axis in a rat model of temporal lobe epilepsy: a mechanism of comorbidity betweenepilepsy and depression. Epilepsia 2009b;50(Suppl 10):68.
Melik Parsadaniantz S, Dauge V, Roques BP, Kerdelhue B. Acute intrahippocampal injection of humaninterleukin-1beta stimulates the anterior pituitary POMC transcription and increases plasma levelsof ACTH and corticosterone in the male rat. Neuroendocrinology 1999;69:77–87. [PubMed:9986920]
Murphy, DL.; Campbell, L.; Costa, JL. The brain serotonergic system in the affective disorders. In:Lipton, MA.; DiMascio, A.; Killam, KF., editors. Psychopharmacology: A generation of progress.Raven Press; New York: 1978. p. 1235-1247.
Mazarati et al. Page 10
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

O'Connor JC, Lawson MA, Andre C, Moreau M, Lestage J, Castanon N, Kelley KW, Dantzer R.Lipopolysaccharide-induced depressive-like behavior is mediated by indoleamine 2,3-dioxygenaseactivation in mice. Mol Psychiatry 2009;14:511–522. [PubMed: 18195714]
Pariante CM, Miller AH. Glucocorticoid receptors in major depression: relevance to pathophysiologyand treatment. Biol Psychiatry 2001;49:391–404. [PubMed: 11274650]
Pohorecky LA, Baumann MH, Benjamin D. Effects of chronic social stress on neuroendocrineresponsiveness to challenge with ethanol, dexamethasone and corticotropin-releasing hormone.Neuroendocrinology 2004;80:332–342. [PubMed: 15692218]
Pucilowski O, Overstreet DH. Effect of chronic antidepressant treatment on responses to apomorphinein selectively bred rat strains. Brain Res Bull 1993;32:471–475. [PubMed: 8221139]
Pucilowski O, Overstreet DH, Rezvani AH, Janowsky DS. Chronic mild stress-induced anhedonia:greater effect in a genetic rat model of depression. Physiol Behav 1993;54:1215–1220. [PubMed:8295967]
Ravizza T, Gagliardi B, Noe F, Boer K, Aronica E, Vezzani A. Innate and adaptive immunity duringepileptogenesis and spontaneous seizures: evidence from experimental models and human temporallobe epilepsy. Neurobiol Dis 2008;29:142–160. [PubMed: 17931873]
Relton JK, Martin D, Thompson RC, Russell DA. Peripheral administration of Interleukin-1 Receptorantagonist inhibits brain damage after focal cerebral ischemia in the rat. Exp Neurol 1996;138:206–213. [PubMed: 8620919]
Riad M, Garcia S, Watkins KC, Jodoin N, Doucet E, Langlois X, el Mestikawy S, Hamon M, DescarriesL. Somatodendritic localization of 5-HT1A and preterminal axonal localization of 5-HT1B serotoninreceptors in adult rat brain. J Comp Neurol 2000;417:181–194. [PubMed: 10660896]
Riazi K, Galic MA, Kuzmiski JB, Ho W, Sharkey KA, Pittman QJ. Microglial activation and TNFalphaproduction mediate altered CNS excitability following peripheral inflammation. Proc Natl Acad SciU S A 2008;105:17151–17156. [PubMed: 18955701]
Rigau V, Morin M, Rousset MC, de Bock F, Lebrun A, Coubes P, Picot MC, Baldy-Moulinier M,Bockaert J, Crespel A, Lerner-Natoli M. Angiogenesis is associated with blood-brain barrierpermeability in temporal lobe epilepsy. Brain 2007;130:1942–1956. [PubMed: 17533168]
Sandberg JO, Eizirik DL, Sandler S, Tracey DE, Andersson A. Treatment with an interleukin-1 receptorantagonist protein prolongs mouse islet allograft survival. Diabetes 1993;42:1845–1851. [PubMed:8243831]
Sanderson KL, Raghupathi R, Saatman KE, Martin D, Miller G, McIntosh TK. Interleukin-1 receptorantagonist attenuates regional neuronal cell death and cognitive dysfunction after experimental braininjury. J Cereb Blood Flow Metab 1999;19:1118–1125. [PubMed: 10532636]
Steimer T, Python A, Schulz PE, Aubry JM. Plasma corticosterone, dexamethasone (DEX) suppressionand DEX/CRH tests in a rat model of genetic vulnerability to depression. Psychoneuroendocrinology2007;32:575–579. [PubMed: 17512120]
Stockmeier CA, Shapiro LA, Dilley GE, Kolli TN, Friedman L, Rajkowska G. Increase in serotonin-1Aautoreceptors in the midbrain of suicide victims with major depression-postmortem evidence fordecreased serotonin activity. J Neurosci 1998;18:7394–7401. [PubMed: 9736659]
Swaab DF, Bao AM, Lucassen PJ. The stress system in the human brain in depression andneurodegeneration. Ageing Res Rev 2005;4:141–194. [PubMed: 15996533]
van den Biggelaar AH, Gussekloo J, de Craen AJ, Frolich M, Stek ML, van der Mast RC, WestendorpRG. Inflammation and interleukin-1 signaling network contribute to depressive symptoms but notcognitive decline in old age. Exp Gerontol 2007;42:693–701. [PubMed: 17350781]
Vezzani A, Balosso S, Ravizza T. The role of cytokines in the pathophysiology of epilepsy. Brain BehavImmun 2008;22:797–803. [PubMed: 18495419]
Vezzani A, Conti M, De Luigi A, Ravizza T, Moneta D, Marchesi F, De Simoni MG. Interleukin-1betaimmunoreactivity and microglia are enhanced in the rat hippocampus by focal kainate application:functional evidence for enhancement of electrographic seizures. J Neurosci 1999;19:5054–5065.[PubMed: 10366638]
Vezzani A, Granata T. Brain inflammation in epilepsy: experimental and clinical evidence. Epilepsia2005;46:1724–1743. [PubMed: 16302852]
Mazarati et al. Page 11
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Vezzani A, Moneta D, Conti M, Richichi C, Ravizza T, De Luigi A, De Simoni MG, Sperk G, Andell-Jonsson S, Lundkvist J, Iverfeldt K, Bartfai T. Powerful anticonvulsant action of IL-1 receptorantagonist on intracerebral injection and astrocytic overexpression in mice. Proc Natl Acad Sci U SA 2000;97:11534–11539. [PubMed: 11016948]
Vezzani A, Moneta D, Richichi C, Aliprandi M, Burrows SJ, Ravizza T, Perego C, De Simoni MG.Functional role of inflammatory cytokines and antiinflammatory molecules in seizures andepileptogenesis. Epilepsia 2002;43(Suppl 5):30–35. [PubMed: 12121291]
Watson S, Gallagher P, Smith MS, Ferrier IN, Young AH. The dex/CRH test--is it better than the DST?Psychoneuroendocrinology 2006;31:889–894. [PubMed: 16701957]
Wrona MZ, Dryhurst G. Oxidation chemistry of 5-hydroxytryptamine. 1. Mechanism and productsformed at micromolar concentrations. J Org Chem 1987;52:2817–2825.
Yirmiya R. Endotoxin produces a depressive-like episode in rats. Brain Res 1996;711:163–174.[PubMed: 8680860]
Zobel A, Wellmer J, Schulze-Rauschenbach S, Pfeiffer U, Schnell S, Elger C, Maier W. Impairment ofinhibitory control of the hypothalamic pituitary adrenocortical system in epilepsy. Eur ArchPsychiatry Clin Neurosci 2004;254:303–311. [PubMed: 15365705]
Mazarati et al. Page 12
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. Experimental designExplanations are in the Methods section.
Mazarati et al. Page 13
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
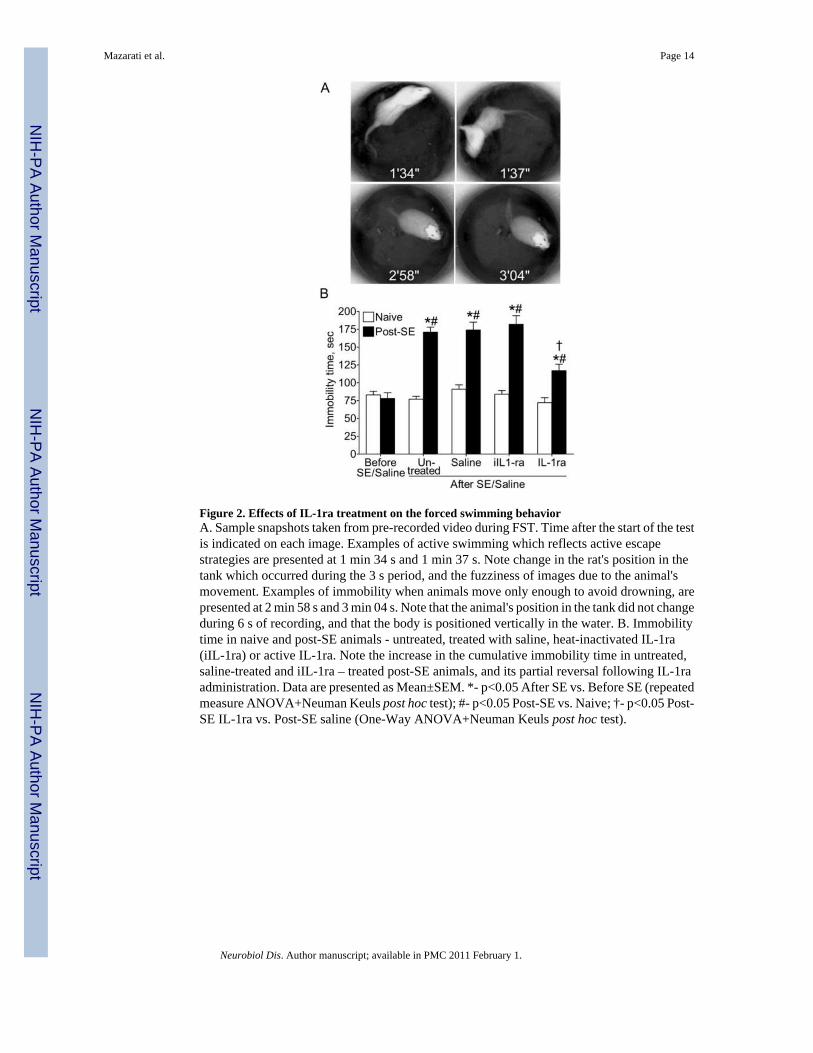
Figure 2. Effects of IL-1ra treatment on the forced swimming behaviorA. Sample snapshots taken from pre-recorded video during FST. Time after the start of the testis indicated on each image. Examples of active swimming which reflects active escapestrategies are presented at 1 min 34 s and 1 min 37 s. Note change in the rat's position in thetank which occurred during the 3 s period, and the fuzziness of images due to the animal'smovement. Examples of immobility when animals move only enough to avoid drowning, arepresented at 2 min 58 s and 3 min 04 s. Note that the animal's position in the tank did not changeduring 6 s of recording, and that the body is positioned vertically in the water. B. Immobilitytime in naive and post-SE animals - untreated, treated with saline, heat-inactivated IL-1ra(iIL-1ra) or active IL-1ra. Note the increase in the cumulative immobility time in untreated,saline-treated and iIL-1ra – treated post-SE animals, and its partial reversal following IL-1raadministration. Data are presented as Mean±SEM. *- p<0.05 After SE vs. Before SE (repeatedmeasure ANOVA+Neuman Keuls post hoc test); #- p<0.05 Post-SE vs. Naive; †- p<0.05 Post-SE IL-1ra vs. Post-SE saline (One-Way ANOVA+Neuman Keuls post hoc test).
Mazarati et al. Page 14
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
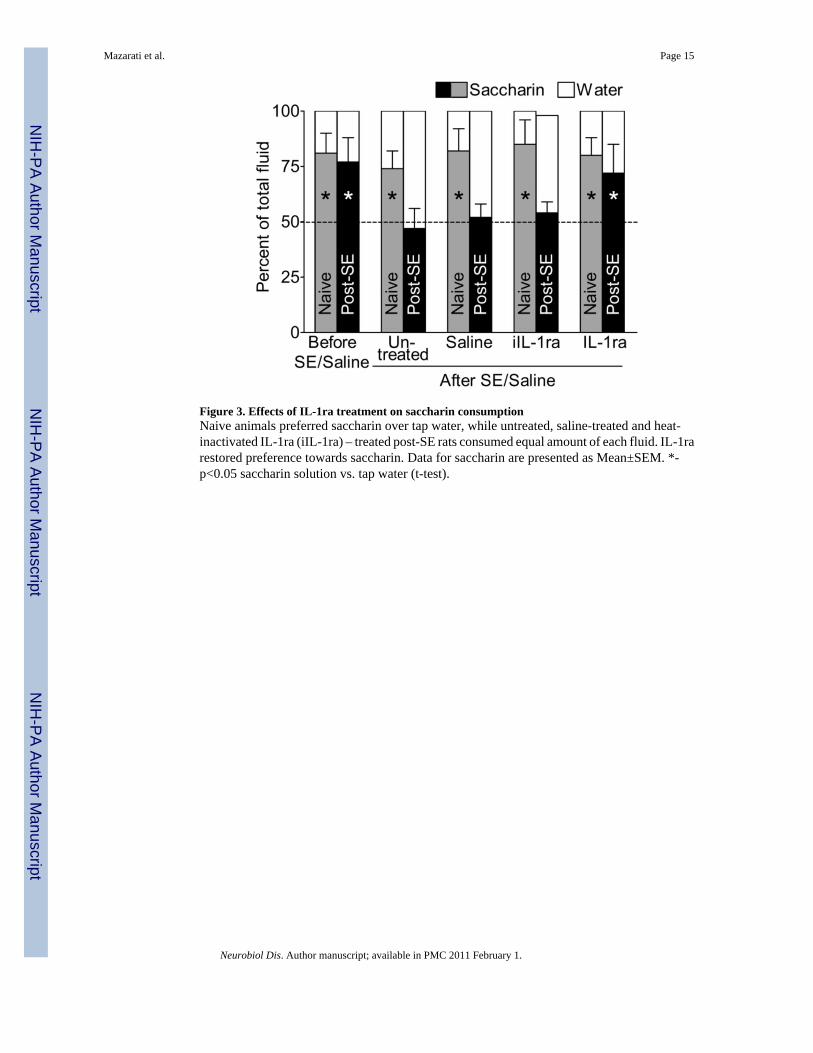
Figure 3. Effects of IL-1ra treatment on saccharin consumptionNaive animals preferred saccharin over tap water, while untreated, saline-treated and heat-inactivated IL-1ra (iIL-1ra) – treated post-SE rats consumed equal amount of each fluid. IL-1rarestored preference towards saccharin. Data for saccharin are presented as Mean±SEM. *-p<0.05 saccharin solution vs. tap water (t-test).
Mazarati et al. Page 15
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 4. Effects of IL-1ra treatment on baseline plasma CORT concentration and on DEX/CRHtestSince in all three groups of naïve animals all examined parameters were statistically similarfor the first and the second assays, in order to simplify the presentation of the findings, theoutcome of DEX/CRH test is presented only for the second assay (i.e. during IL-1ra treatment)and data in naive groups are combined for the three treatments (i.e. IL-1ra, saline, anduntreated). In saline-treated and untreated rats, SE led to the increase of baseline plasma CORTlevel; failure of DEX to decrease CORT concentration; and the exacerbated and longer-lastingresponse to CRH (since the responses in two groups were statistically close, the data arecombined for presentation purposes). IL-1ra decreased baseline plasma CORT level andpartially normalized the outcome of DEX/CRH test. Data are presented as Mean±SEM. Eventhough data for some groups were combined, statistical comparisons were done betweenrespective groups: #- p<0.05 Post-SE-Saline and Post-SE untreated vs. respective Naïvecontrols and at respective time points; †- p<0.05 post-SE IL-1ra vs. Naive IL-1ra; ‡- p<0.05post-SE IL-1ra vs. post-SE Saline and post-SE untreated (One-Way ANOVA + Neuman-Keulspost hoc test). *- p<0.05; **- p<0.01; ***- p<0.001 comparison within the same groups, fortwo adjacent time points between which the symbols are located (Repeated measure ANOVA+ Neuman-Keuls post hoc test).
Mazarati et al. Page 16
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
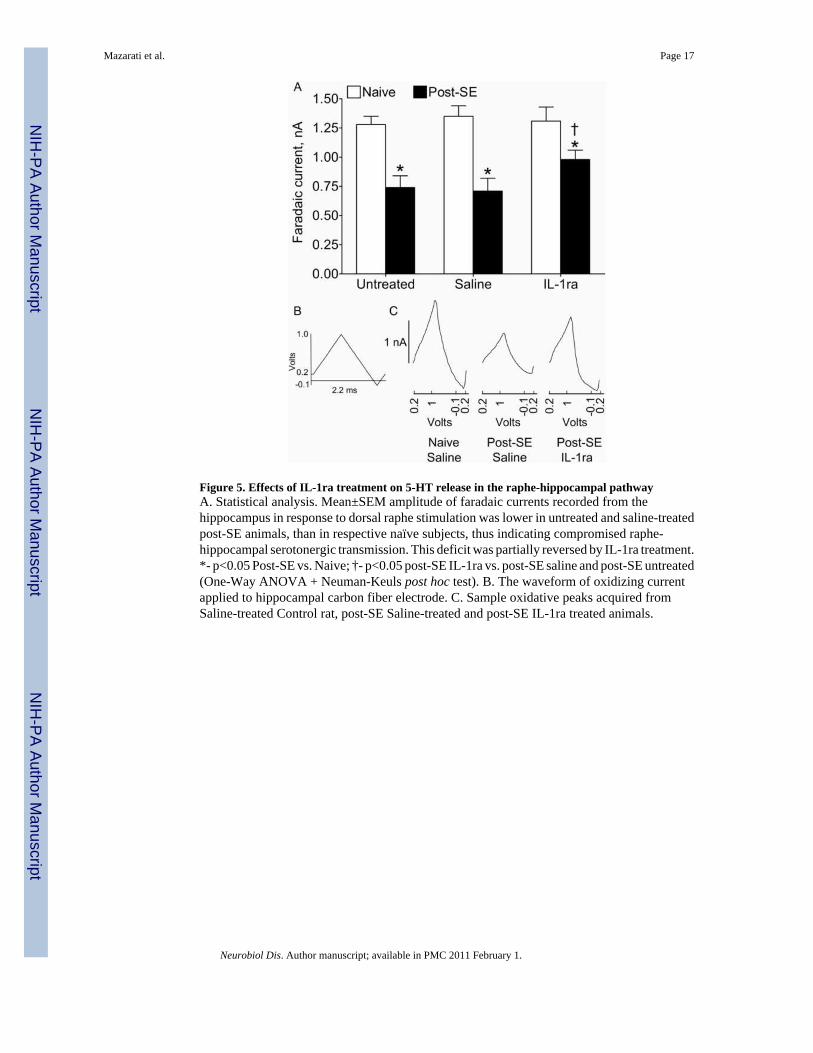
Figure 5. Effects of IL-1ra treatment on 5-HT release in the raphe-hippocampal pathwayA. Statistical analysis. Mean±SEM amplitude of faradaic currents recorded from thehippocampus in response to dorsal raphe stimulation was lower in untreated and saline-treatedpost-SE animals, than in respective naïve subjects, thus indicating compromised raphe-hippocampal serotonergic transmission. This deficit was partially reversed by IL-1ra treatment.*- p<0.05 Post-SE vs. Naive; †- p<0.05 post-SE IL-1ra vs. post-SE saline and post-SE untreated(One-Way ANOVA + Neuman-Keuls post hoc test). B. The waveform of oxidizing currentapplied to hippocampal carbon fiber electrode. C. Sample oxidative peaks acquired fromSaline-treated Control rat, post-SE Saline-treated and post-SE IL-1ra treated animals.
Mazarati et al. Page 17
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Mazarati et al. Page 18
Table 1Behavioral spontaneous seizure count in post-SE rats
Numbers indicate cumulative number of behavioral seizures for individual animals over 2 two-week periods:before and during IL1-ra and saline treatments (or respective period in untreated rats). No differences wereobserved between the two periods for each of the groups (p>0.05, both Wilcoxon and Mann-Whitney tests forpaired and unpaired comparisons), and across the groups (p>0.05, Kruskal-Wallis test). No differences wereobserved in the direction in which frequency changed (i.e. decreased, increased and remained unchanged) amongany pairs of groups (p>0.05, Fisher's exact test).
Treatment group Before treatment During treatment Changes in seizure count during treatment vs.before treatment
IL1-ra
3 0
Decreased- 4 Increased- 3 Unchanged-2
2 1
11 5
4 0
6 8
33 42
9 71
0 0
4 4
Saline
7 3
Decreased- 4 Increased- 5
1 0
4 3
56 2
13 14
5 12
0 2
3 39
0 7
Untreated
11 4
Decreased- 5 Increased-4
9 0
10 4
4 0
42 4
4 61
0 15
0 33
4 7
Neurobiol Dis. Author manuscript; available in PMC 2011 February 1.