colonization of collagen scaffolds by adipocytes derived from mesenchymal stem cells of the common...
TRANSCRIPT

Biochemical and Biophysical Research Communications 411 (2011) 317–322
Contents lists available at ScienceDirect
Biochemical and Biophysical Research Communications
journal homepage: www.elsevier .com/locate /ybbrc
Colonization of collagen scaffolds by adipocytes derived from mesenchymalstem cells of the common marmoset monkey
Inga Bernemann a,⇑, Thomas Mueller b, Rainer Blasczyk b, Birgit Glasmacher a, Nicola Hofmann a
a Institute for Multiphase Processes, Leibniz Universität Hannover, Hannover, Germanyb Institute for Transfusion Medicine, Hannover Medical School, Hannover, Germany
a r t i c l e i n f o a b s t r a c t
Article history:Received 3 June 2011Available online 25 June 2011
Keywords:Common marmoset monkeyMSCsCollagen scaffoldsNon-human primateAdipogenic differentiation
0006-291X/$ - see front matter � 2011 Elsevier Inc. Adoi:10.1016/j.bbrc.2011.06.134
⇑ Corresponding author. Address: Institute for MUniversität Hannover, Callinstr. 36, D-30167 Hannove19389.
E-mail address: [email protected]
In regenerative medicine, human cell replacement therapy offers great potential, especially by cell typesdifferentiated from immunologically and ethically unproblematic mesenchymal stem cells (MSCs). Interms of an appropriate carrier material, collagen scaffolds with homogeneous pore size of 65 lm wereoptimal for cell seeding and cultivating. However, before clinical application and transplantation of MSC-derived cells in scaffolds, the safety and efficiency, but also possible interference in differentiation due tothe material must be preclinically tested. The common marmoset monkey (Callithrix jacchus) is a prefer-able non-human primate animal model for this aim due to its genetic and physiological similarities to thehuman.
Marmoset bone marrow-derived MSCs were successfully isolated, cultured and differentiated in sus-pension into adipogenic, osteogenic and chondrogenic lineages by defined factors. The differentiationcapability could be determined by FACS. Specific marker genes for all three cell types could be detectedby RT-PCR. Furthermore, MSCs seeded on collagen I scaffolds differentiated in adipogenic lineage showedafter 28 days of differentiation high cell viability and homogenous distribution on the material which wasvalidated by calcein AM and EthD staining. As proof of adipogenic cells, the intracellular lipid vesicles inthe cells were stained with Oil Red O. The generation of fat vacuoles was visibly extensive distinguishableand furthermore determined on the molecular level by expression of specific marker genes. The results ofthe study proved both the differential potential of marmoset MSCs in adipogenic, osteogenic and chon-drogenic lineages and the suitability of collagen scaffolds as carrier material undisturbing differentiationof primate mesenchymal stem cells.
� 2011 Elsevier Inc. All rights reserved.
1. Introduction
Regenerative medicine aims in the replacement of tissue due toloss or organ failure by implantation of an engineered biologicalsubstitute. This implant is either directly functional at the timeof implantation or matures to the desired functional tissue typeduring or after the integration. One particular challenge in tissueengineering poses the combination of appropriate cells on a suit-able biomaterial as a carrying structure. Besides the usage of stemcells or reprogrammed cells, the utilization of autologous cells, ob-tained relatively uninvasive from e.g. skin, bone marrow, blood orhair, is a feasible approach, due to the absence of allogenicity andimmune rejection. Although autologous somatic cells can be iso-lated, cultured, and expanded in vitro in large numbers and seededon diverse matrices [1] the outcome and survival of these cells re-
ll rights reserved.
ultiphase Processes, Leibnizr, Germany. Fax: +49 511 762
(I. Bernemann).
mains critical [2]. Furthermore, the transplantation into a humanpatient remains impossible due to ethical reasons. Before directclinical application of mesenchymal stem cells (MSCs) and cellseeded substitutes in cell and tissue replacement therapy, thesafety and feasibility must be preclinically tested. This should bepreferably carried out in a non-human primate animal model, sincemouse stem cells have proven to display great differences from thehuman [2].
The common marmoset (Callithrix jacchus) is such a model read-ily used in biomedical research due to its genetic and physiologicalsimilarities to the human [3,4]. Over the last years there has beenan increasing usage of marmoset as an alternative non-rodent spe-cies in preclinical safety evaluations on new pharmaceuticals [4–7]stem cell research [7–9] in, not only investigation of multiple scle-rosis research [10,11] in infectious and autoimmune diseases[12,13] but also in reproductive medicine, transplantation research[14] and immunology [15–17]. C. jacchus (cj) and homo sapiens arethought to have diverged from a common ancestor, a primitiveanthroid, which existed approximately 30 million years ago [18].Despite many years of evolution separation, the human DNA

318 I. Bernemann et al. / Biochemical and Biophysical Research Communications 411 (2011) 317–322
sequences are still similar enough to anneal to marmoset metapha-ses, which simplifies a lot of different standard molecular biologi-cal applications. Therefore, and due to the small size and easyhandling of the animals, the usage of marmoset cells in particularmarmoset MSCs in reconstructive medicine applications becomemore and more attractive.
The scaffolding material in cell-based tissue engineering deci-des the characteristics of the subsequent constructs. In this appli-cation, collagen is the most abundant protein in the human andanimal body, and thus an excellent and potential biomaterial forscaffolds for various applications in regenerative medicine. In bio-logical systems, it allows cell attachment, differentiation, organo-genesis, tissue regeneration and repair. Collagen scaffolds aremechanically stable with a high tensile strength and can be alteredinto different sizes and shapes with various physical and chemicalmodifications [1,19,20]. Collagen scaffolds are successful matricesfor adipose tissue engineering, because they are biocompatible,support tissue formation and demonstrate considerable stabilityto allow successful integration between the newly formed tissueand the surrounding host tissue [21]. The homogeneous pore struc-ture of the used scaffolds [1,19,20] may be decisively responsiblefor the attachment, the distribution and the differentiation of thecells in three dimensions.
Collagen type I scaffolds are also already well-accepted for hu-man implantation [1]. With a porosity of 98% and excellent degra-dability and biocompatibility properties, this material ispredestinated for functional human tissue engineering. Besidethe matrix itself, the choice of an appropriate cell type for engi-neered biological substitute is quite important.
Bone marrow-derived MSCs are the progenitors of multiple lin-eages, including bone, cartilage, muscle, and fat. They have mani-fold application and a high therapeutic potential. Thiscombination of marmoset MSCs and collagen I scaffolds is the firststep in clinical practice. The adipogenic differential potential of thecells in collagen matrix, especially behind the background of adi-pose tissue engineering, offers valuable clues to the applicabilityin human research.
In plastic and reconstructive surgery, autologous fat grafts areutilized as filling material for the reconstruction of soft tissue de-fects [22,23]. The current standard of care for soft tissue recon-struction and augmentation includes the utilization of syntheticand tissue transfer [24–27]. The need for secondary surgical proce-dures for autologous tissue harvest and the average of 40–60%reduction loss in graft volume over time are considered drawbacksof current autologous fat transplantation procedures [24,28,29].Hence, for adipose tissue engineering, the use of adult MSCs maybe advantageous over the use of differentiated adipocytes
Fig. 1. (A) Collagen scaffolds (bar = 3 mm). (B) The homogeneous and parallel pore stproliferation in the 3D matrix, SEM (bar = 100 lm). (C) Close-up visualizing of the pore
[18,30,31]. MSCs offer a potentially unlimited source of cells fortissue engineering application and the use of stem cells to produceadipose tissue has become more and more popular [32]. And, if thepatient’s own cells are used, biocompatibility complications can beeliminated [33].
Regarding adipose tissue engineering, we characterized the dif-ferentiation capability of cjMSCs into adipogenic lineage engraftedinto collagen scaffolds. To proof the functionality of cjMSC beforeseeding, we established multilineage differentiation of cjMSCs insuspension. We used isolated bone marrow-derived MSCs fromthe common marmoset monkey for the production of engineeredadipose tissue replacement which will allow implantations in anon-human primate animal model in vivo at a later point in time.
2. Materials and methods
All chemicals and reagents were obtained from Sigma–AldrichGmbH (Munich, Germany) unless otherwise specifically indicated.
2.1. Collagen scaffolds
Collagen scaffolds (Fig. 1A) were produced as described before[1,19,20,34] by directional solidification and subsequent freeze-drying by power-down technology. This process generated colla-gen scaffolds with a homogeneous pore structure and defined poresizes (Fig. 1B and C) [35]. Due to their biodegradable and biocom-patible behaviour collagen scaffolds are suitable 3D matrices for amultitude of cell types in tissue engineering [1,34]. Collagen type Iis also well-accepted for human implantation [1]. In this study weused collagen scaffolds with a height of 3 mm, a diameter of15 mm, an average pore size of 65 lm and a porosity of 98%.
2.2. Animal experiments
All experiments were in accordance the district government ofLower Saxonia, Hannover, and North-Rhine Westfaelia, Germany.Euthanasia of the animals and retrieval of the bone marrow wasapproved by the Institutional Animal Care. It should be highlightedthat all animals were initially killed for other studies or as veteri-nary measure, but not primarily for our experiments.
2.3. Isolation of marmoset bone marrow stem cells
The bone marrow of marmoset was isolated by rupture of thetibia and femur of each animal immediately after death was con-firmed by a veterinarian. The cavity was flushed with a hypoder-mic needle attached to a syringe. For preventing coagulation and
ructure of the collagen scaffolds allows optimized cell distribution, adhesion andstructure (bar = 50 lm).

I. Bernemann et al. / Biochemical and Biophysical Research Communications 411 (2011) 317–322 319
for cell singularization, a heparin PBS mix was utilized (5 IU/ml)and the bone marrow was separated by pipetting thoroughlyfor 3 min. After singularization, the cell suspension was trans-ferred into red cell lysis buffer (NH4Cl 0.15 M, KHCO3 10 mM,EDTA 0.1 mM) for 5 min and centrifuged at 200g for 10 min.The cell pellet was resuspended in the MSC culture medium con-sisting of Dulbecco’s Modified Eagle Medium (DMEM) (BiochromAG, Berlin, Germany), 15% FCS (Biochrom AG, Berlin, Germany),1% penicillin/streptomycin (Invitrogen GmbH, Karlsruhe, Ger-many), 10 lmol nystatin, 50 lM L-ascorbin acid-2-phosphateand plated on a cell culture dish.
2.4. Differentiation potential of marmoset MSCs
To demonstrate the general differentiation capability of mar-moset MSCs (cjMSCs) into adipogenic, osteogenic and chondro-genic lineage different supplements have to add to the culturemedium and the cells have been cultivated for several weeks. Allcells were visualized using Axiovert 200 microscope.
About 7.5 � 103 cells per cm2 were cultured for 3 weeks. Thenadipogenic differentiation was achieved by the addition of 1 lMdexamethasone, 0.2 mM indomethacin, 0.5 mM IBMX and 10 lg/ml insulin in DMEM with 10% FBS. After 3 days the medium wasreplaced by culture medium (DMEM, 10% FBS, insulin (20 lg/ml))for 2 days. This cultivation circle was repeated for four times. OilRed O dye was used to detect the generation of fat vacuoles. In or-der to do this, cells were washed with PBS and fixed for 20 min in4% paraformaldehyde, then washed in H2O twice, followed by ashort rinse in 50% ethanol. After incubation for 10 min in Oil RedO in acetone and 50% ethanol, removal of the staining dye and dif-ferentiation in 50% ethanol.
The osteogenic differentiation was induced by the addition ofosteogenic differentiation medium supplemented with 0.1 lMdexamethasone, 10 mM ß-glycerophosphate, 0.05 mM L-ascorbinacid-2-phosphate and 1% ITS (BD Biosciences, Heidelberg, Ger-many) in DMEM with 15% FBS. Von Kossa staining characterizedthe biological mineralization. The cells were washed in PBS andfixed with 10% phosphate-buffered paraformaldehyde, washedin PBS and distilled water. 1% silver nitrate solution was added.Then the dish was exposed to sunlight for 30 min, after whichthe plate was rinsed with water. Five percentage sodium thiosul-fate was added for 5 min, and then the plates were rinsed inwater.
For the chondrogenic differentiation, 2 � 105 cjMSCs were pel-leted by centrifugation at 500g for 5 min. The chondrogenic differ-entiation of the pellet micromass was induced by incubation for3 weeks in chondrogenic differentiation medium with 0.1 lMdexamethasone, 1 mM sodium pyruvate, 0.17 mM L-ascorbinacid-2-phosphate, 0.35 mM L-proline, 10 ng/ml transforminggrowth factor-beta 3 (RELIATech, Braunschweig, Germany) and1% ITS + Premix in DMEM with 15% FBS. Medium was changedevery 2–3days. After 3 weeks, the pellet was fixed with 10% para-formaldehyde, sectioned at 7 lm and stained with alcian blue(Merck, Darmstadt, Germany) as an indicator of sulfated glycos-aminoglycan [sGAG]-rich extracellular matrix and Hoechst 33342(Invitrogen, Karlsruhe, Germany), according to standard protocolsusing cryosections. The detection of extracellular matrix produc-tion was done by using an aggrecan marker (Acris Antibodies, Her-ford, Germany).
2.5. Flow cytometric analysis
The quantification of differentiation of cjMSCs was analysed in aFACSCalibur flow cytometer (BD Bio-Sciences, Heidelberg,Germany). Therefore, differentiated cells were suspended andsingularized. After 14 days of adipogenic differentiation, the cells
were fixated in 4% paraformaldehyde for 10 min. Then cells wereresuspended in 150 ll Nile Red (1 lg/ml) and incubated for60 min at RT. Partly the cells could not be pelletized. These cellswere stained separately according to the protocol and then ana-lysed. For analysis the osteogenic differentiated cells were stainedon day 14 with calcein in (12.5 lg/ml) over night at 4 �C. Then thecells were trypsinized and analyzed. The quantification of chon-drogensis was done after 3 weeks. Therefore, the cells of the pelletculture were singularized and suspended. The primary antibodyAggrecan (1:200) incubated for 2 h at 37 �C and Alexa Fluor 488rabbit anti mouse IgG secondary antibody (1:400) at 37 �C for1 h. After washing with PBS the cells were transferred into a FACStube and measured. Undifferentiated MSCs were used in all exper-iments as control.
2.6. Adipogenic differentiation in collagen scaffolds
The collagen scaffolds were seeded with 1 � 106 cells resolvedin 400 ll culture medium. On day 1 after seeding, the culture med-ium (DMEM with 10% FBS) was complemented with adipogenicsupplements. The control scaffolds were cultivated for 4 weeks inculture medium without differentiation supplements. The cell via-bility was determined using LIVE/DEAD Viabilty/Cytotoxicity Kit(Invitrogen, Karlsruhe, Germany). After 4 weeks of adipogenic dif-ferentiation, intracellular lipid vesicles were detected by Oil Red Ostaining.
2.7. Differentiation analysis by RT-PCR
The analysis of the general differentiation capability of MSCsand their adipogenic differentiation in collagen scaffolds was alsodetermined on a molecular level by RT-PCR. To analyze the expres-sion of typical marker genes for the respective differentiated celltypes, we selected for adipocytes PPAR-g, aP2 and CEBPa. For chon-drocyte development ACAN, ColA1 and CYR6.1 expression wasinvestigated. Osteoblast formation was displayed by TIMP2,MMP2 and RUNX2 expression. For quantification and control weutilized ß-actin and the ribosomal gene RPS29. For the RT-PCR weselect the following primer:
PPAR-g: 50-CTGTGAAGTTCAATGCACTG-30; 50-TGTTCCGTGACAATCTGTCTG-30
aP2: 50-GATGGCAATTGGAAGGTAGA-30; 50-AGAACTGAATACTGTCAGAG-30
CEBPa: 50-GTGGACAAGAACAGCAACGA-30; 50-GTCATTGTCACTGGTCAGCT-30
TIMP2: 50-CAAGATGCACATCACCCTCT-30; 50-TTCTTCTCTGTGACCCAGTC-30
MMP2: 50-GACACGCTGAAGAAGATGCA -30; 50-CTGCCTCTCCAT-CAT GGATTC-30
RUNX2: 50-AACTTCCTCTGCTCCGTGCT-30; 50-TGCGGTAGCATTTCTCAGCT-30
ColA1: 50-GAGACTACTGGATTGACCCC-30; 50-CCAGGTTGT-CATCTC CATAG-30
CYR6.1: 50-CTGTGGAACTGGTATCTCCA-30; 50-CTTCACACTCGAA-CATCCAG-30
ACAN: 50-GTGTTCCATTACAGAGCCATTT-30; 50-TCTCATTGGTGTCTCGGATG-30
ß-Actin: 50-CATTGTCACCAACTGGGAC-30; 50-CAGCCATGTATGTGGCCATC-30
RPS29: 50-GCAAGATGGGTCACCAGCAG-30; 50-CCAGTGTTTCCGT-CAGTACG-30
RNA of 5 � 105 cells respectively was isolated with the RNeasykit (Quiagen, Hilden, Germany) according to the manufacturer’sinstructions and normalized with a Nanodrop ND-1000 (PeqLab,Erlangen, Germany). 0.5 lg RNA was transcribed with a RT-kit(Applied Biosystems, Forster City, USA). The PCR-reaction mix
320 I. Bernemann et al. / Biochemical and Biophysical Research Communications 411 (2011) 317–322
contained 30 ll volume with 1 ll cDNA, 0.3 lM of each primer (TIBBiomol, Berlin, Germany), 1U Taq DNA polymerase (NatuTec,Frankfurt, Germany) and 333 lM DNTP mix under standard PCRconditions (initial denaturation 5 min 94 �C; 30 cycles 1 min at94 �C, annealing 1 min 62 �C, elongation 1 min 72 �C, final exten-sion 15 min 72 �C). PCR products were separated by electrophore-sis (1.5% agarose gel), and pictures were taken (Gel Doc UniversalHood II, BioRad, Munich, Germany).
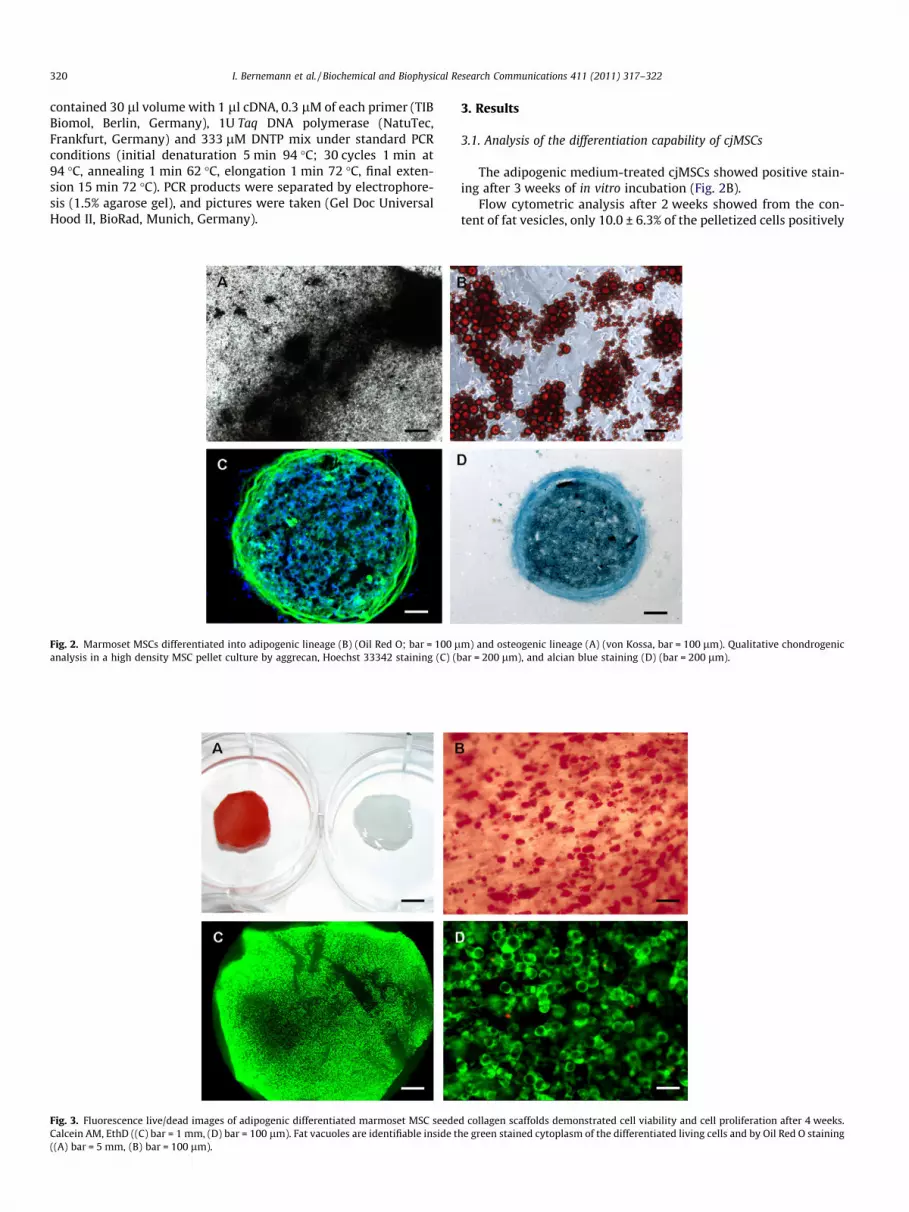
Fig. 2. Marmoset MSCs differentiated into adipogenic lineage (B) (Oil Red O; bar = 100 lanalysis in a high density MSC pellet culture by aggrecan, Hoechst 33342 staining (C) (b
Fig. 3. Fluorescence live/dead images of adipogenic differentiated marmoset MSC seedeCalcein AM, EthD ((C) bar = 1 mm, (D) bar = 100 lm). Fat vacuoles are identifiable inside t((A) bar = 5 mm, (B) bar = 100 lm).
3. Results
3.1. Analysis of the differentiation capability of cjMSCs
The adipogenic medium-treated cjMSCs showed positive stain-ing after 3 weeks of in vitro incubation (Fig. 2B).
Flow cytometric analysis after 2 weeks showed from the con-tent of fat vesicles, only 10.0 ± 6.3% of the pelletized cells positively
m) and osteogenic lineage (A) (von Kossa, bar = 100 lm). Qualitative chondrogenicar = 200 lm), and alcian blue staining (D) (bar = 200 lm).
d collagen scaffolds demonstrated cell viability and cell proliferation after 4 weeks.he green stained cytoplasm of the differentiated living cells and by Oil Red O staining

Fig. 4. RT-PCR analyse: detection of typical marker gene for adipogenic differen-tiation of cells in suspension on d15 and into scaffolds on d32 (A) and also forosteogenic (B) and chondrogenic differentiation (C) confirm the differentiationpotential of cjMSCs.
I. Bernemann et al. / Biochemical and Biophysical Research Communications 411 (2011) 317–322 321
identified as adipocytes, whereas 85.5% were positive in thesupernatant. Overall, 22.6 ± 4.7% of the cells could be successfullydifferentiated into adipocytes after 14 days. In general, the longerthe period of differentiation, the greater the quantity of lipid afat cell in culture can accumulate [36]. After 3 weeks of osteogenicdifferentiation, the generation of calcium deposits could distinctqualitative be proved by von Kossa staining (Fig. 2A). Flowcytometric analysis after 14 days showed that 11.34 ± 2.61% ofthe investigated cells positively identified as osteoblasts. The chon-drogenesis was qualitative and quantitative proven by histologicalinvestigations (Fig. 2C and D) and flow cytometric analysis. Theincrease of aggregan expression after 21 days indicates thechondrogenic differentiation (Fig. 2C). Additionally images ofalcian blue-stained cryosections verified qualitative chondrogenicdifferentiation potential of the cells (Fig. 2D) and proved thepresence of sulfated glycosaminoglycans (sGAG). Flow cytometricanalysis after 3 weeks showed that 9.46 ± 1.6% of the cells in thepellet culture positively identified as chondrocytes.
Specific marker genes for all three cell types (adipocytes, osteo-cytes and chondrocytes) could be detected by RT-PCR (Fig.4). Theseexperiments showed that isolated cjMSC can differentiate in a con-trolled manner to multiple lineages.
3.2. Analysis of the adipogenic differentiation potential of cjMSCs incollagen scaffolds
The viability after 4 weeks adipogenic differentiation was eval-uated and we observed a high cell proliferation and a high cellamount of viable cells and only a small quantity of dead cells(Fig. 3C and D). The cells adhered and proliferated three dimen-sionally into the scaffold. After 4 weeks there is a high density ofevenly distributed cells in and on the collagen scaffold. Fig. 3Cand D illustrates that the cells were grown multidimensional inthe scaffold. After 4 weeks the generation of fat vacuoles in colla-gen scaffolds was detected (Fig. 3A and B).
The histological investigation yielded that the cell distributionin collagen scaffold was dense and evenly distributed (Fig. 3A–D).Fat cells in all sections of the scaffold generate comprehensiveintracellular lipid vesicles. The macroscopic and microscopic re-sults proved the distinct adipogenic differentiation capability ofcjMSCs in collagen scaffolds.
Gene expression of adipogenic marker genes in differentiatedMSCs in scaffolds was demonstrated by RT-PCR. On day 0 and after4 weeks of adipogenic differentiation, the expression of this
specific marker genes was measured (Fig.4A). The increase after4 weeks is shown in Fig.4A. The results of the RT-PCR confirmedadipogenic differentiation capacity of cjMSCs in collagen scaffoldsand verified the results from the Oil Red O staining (Fig.3A and B).
4. Discussion
In the last years there has been tremendous progress in the fieldof regenerative medicine and stem cell research [37]. Not evennearly all cell types could be as proof of principle generated, butalso somatic cells reprogrammed into pluripotent lineages to over-come immunogenicity of allogenic embryonic stem cells [38].However, tissue functions are dependent on the arrangement, con-nection, and interaction of their composed cells and matrix ele-ments in 3D indicating that sole generation of certain cell typesin vitro display limited information about cell efficiency and func-tion in a transplant in vivo. To obtain more information about gen-erated biological tissue substitutes in terms of cell behaviour,differentiation potential and survival in human bodies we utilizeda non-human primate animal model to mimic their integration inhuman environment. Our animal model of the common marmosetfulfils a multitude of required conditions, like its direct translationto embryonic, autologous, and allogenic somatic stem cells andpotentially also to induced pluripotent cells (iPS).
In the past it has been demonstrated that collagen type I scaf-folds prepared from porcine collagen suspension is a suitable car-rier for stem cells and a compatible biodegradable material inthe field of tissue engineering [1,19,20,34,35,39]. It has been dem-onstrated that various cell types can attach, differentiate and pro-liferate in collagen scaffolds to form a specific tissue or organ,however this was never performed in a non-human primate modelwith human or primate MSCs.
Isolated marrow-derived mesenchymal stem cells of the com-mon marmoset monkey displayed a surprisingly excellent differ-entiation capability in adipogenic, osteogenic and chondrogeniclineage using defined protocols. In this study we established thedifferentiation of the cjMSCs and confirmed the applicability of di-verse functional and morphological assays for the marmosetspecies.
Furthermore, cjMSCs integrated well in collagen type I scaffoldswhere they differentiated excellently into adipogenic cells. Hence,for adipose tissue engineering research this combination is partic-ularly suitable. The existence of a large number of lipid vesicles andgene expression of several adipogenic markers could be demon-strated even after 4 weeks of cultivation, which is proof of functionand cell survival. The study also demonstrated that cjMSCs couldbe seeded and evenly distributed in collagen scaffolds. Even afteradipogenic differentiation, the cells in the scaffold showed high celldensity and cell viability.
In conclusion, in this study we have successfully establishedthe cultivation and differentiation of MSCs of a non-human pri-mate model into different somatic lineages, namely adipogenic,osteogenic and chondrogenic lineages. The generation of an adi-pose substitute by interaction of cjMSCs and collagen scaffoldscould also be observed and characterized. The non-human pri-mate MSCs attached and penetrated the collagen scaffolds sur-prisingly well and displayed an excellent adipogenicdifferentiation capability and survival. This study strongly sug-gests that the common marmoset monkey is a suitable modelfor soft tissue engineering in human regenerative medicine. Infurther studies our substitutes will be utilized in transplantationstudies as a preclinical approach. Thereby, the perspective of clin-ical application of adipose tissue engineering developments intohuman regenerative medicine increases and opens manifold med-ical application possibilities.

322 I. Bernemann et al. / Biochemical and Biophysical Research Communications 411 (2011) 317–322
Acknowledgments
We thank Resorba�, Germany, for the provision of the collagenand the Centre for Reproductive Medicine and Andrology Muenster(CeRA) for donation of the marmoset bone marrow. Thanks to A.Deiwick and C. Marx from IMP, Leibniz Universität Hannover, K.Egler from the MH Hannover for their outstanding technical sup-port and W. Hake for his excellent photographic work (Fig. 1A).This work is supported by funding from the Deutsche Forschungs-gemeinschaft (DFG, German research Foundation) for the Cluster ofExcellence REBIRTH (from Regenerative Biology to ReconstructiveTherapy) (EXC 62/1).
References
[1] M. Kuberka, I. Heschel, B. Glasmacher, G. Rau, Preparation of collagen scaffoldsand their applications in tissue engineering, Biomed. Tech. 47 (2002) 485–487.
[2] T. Cantz, G. Key, M. Bleidißel, A. Brenne, H.R. Schöler, Absence of Oct4expression in somatic tumor cell lines, Stem Cells 26 (2008) 692–697.
[3] Nature Publishing Group, The common marmoset: Biomedical supermodel,Nature 459 (2009) 7246.
[4] K. Mansfield, Marmoset models commonly used in biomedical research, Comp.Med. 53 (2003) 383–392.
[5] D. Smith, P. Trennery, D. Farningham, J. Klapwijk, The selection of marmosetmonkeys (Callithrix jacchus) in pharmaceutical toxicology, Lab. Anim. 35(2001) 117–130.
[6] U. Zühlke, G. Weinbauer, The common marmoset (Callithrix jacchus) as a modelin toxicology, Toxicol. Pathol. 31 (2003) 23–127.
[7] T. Mueller, G. Fleischmann, K. Eildermann, K. Maetz-Resing, P.A. Horn, E.Sasaki, R. Behr, A novel embryonic stem cell line derived from commonmarmoset monkey (Callithrix jacchus) exhibiting germ cell-like characteristics,Hum. Reprod. 24 (2009) 1359–1372.
[8] T. Mueller, T. Hupfeld, J. Roessler, M. Simoni, J. Gromoll, R. Behr, Molecularcloning and functional characterization of endogenous recombinant commonmarmoset monkey (Callithrix jacchus) follicle stimulating hormone, J. Med.Primatol. (2010), doi:10.1111/j.1600-0684.2010.00453.x [Epub ahead of print].
[9] E. Sasaki, K. Hanazawa, R. Kurita, A. Akatsuka, T. Yoshizaki, H. Ishii, Y. Tanioka,Y. Ohnishi, H. Suemizu, A. Sugawara, N. Tamaoki, K. Izawa, Y. Nakazaki, H.Hamada, H. Suemori, S. Asano, N. Nakatsuji, H. Okano, K. Tani, Establishment ofnovel embryonic stem cell lines derived from the common marmoset(Callithrix jacchus), Stem Cells 23 (2005) 1304–1313.
[10] B.A. t ‘Hart, L. Massacesi, Clinical, pathological, and immunologic aspects of themultiple sclerosis model in common marmosets (Callithrix jacchus), J.Neuropathol. Exp. Neurol. 68 (2009) 341–355.
[11] B.A. t’Hart, M. van Meurs, H.P. Brok, L. Massacesi, J. Bauer, L. Boon, R.E. Bontrop,J.D. Laman, A new primate model for multiple sclerosis in the commonmarmoset, Immunol. Today 21 (2000) 290–297.
[12] D.H. Crawford, G. Janossy, C.M. Hetherington, G.E. Francis, A.J. Edwards, A.V.Hoffbrand, H.G. Prentice, Immunological characterization of hemopoietic cellsin the common marmoset, rhesus monkey, and man. In search of a model forhuman marrow transplantation, Transplantation 31 (1981) 245–250.
[13] S. Potkay, Diseases of the Callitrichidae: a review, J. Med. Primatol. 21 (1992)189–236.
[14] C.M. Luetjens, J.B. Stukenborg, E. Nieschlag, M. Simoni, J. Wistuba, Completespermatogenesis in orthotopic but not in ectopic transplants of autologouslygrafted marmoset testicular tissue, Endocrinology 149 (2008) 1736–1747.
[15] A. Averdam, H. Kuhl, M. Sontag, T. Becker, A.L. Hughes, R. Reinhardt, L. Walter,Genomics and diversity of the common marmoset monkey NK complex, J.Immunol. 178 (2007) 7151–7161.
[16] M. Barros, V. Boere, J.P. Husto, C. Tomaz, Measuring fear and anxiety in themarmoset (Callithrix penicillata) with novel predator confrontation model:effects of diazepam, Behav. Brain Res. 108 (2000) 205–211.
[17] L. Boon, H.P. Brok, J. Bauer, A. Ortiz-Buijsse, M.M. Schellekens, S. Ramdien-Murli, E. Blezer, M. van Meurs, J. Ceuppens, M. de Boer, B.A. t’Hart, J.D. Laman,Prevention of experimental autoimmune encephalomyelitis in the commonmarmoset (Callithrix jacchus) using a chimeric antagonist monoclonal antibodyagainst human CD40 is associated with altered B-cell responses, J. Immunol.167 (2001) 2942–2949.
[18] J.K. Sherlock, D.K. Griffin, J.D. Delhanty, J.M. Parrington, Homologies betweenhuman and marmoset (Callithrix jacchus) chromosomes revealed bycomparative chromosome painting, Physiol. Genomics 33 (2) (1996) 214–219.
[19] H. Schoof, J. Apel, I. Heschel, G. Rau, Control of pore structure and size infreeze-dried collagen scaffolds, J. Biomed. Mater. Res. 58 (2002) 352–357.
[20] H. Schoof, L. Bruns, A. Fischer, I. Heschel, G. Rau, Dendritic ice morphology inunidirectionally solidified collagen scaffolds, J. Cryst. Growth 209 (2000) 122–129.
[21] M.N. Rahaman, J.J. Mao, Stem cell based composite tissue constructs forregenerative medicine, Biotechnol. Bioeng. 91 (3) (2005) 261–284.
[22] M.W. Strickberger, Evolution, third ed., Jones and Barclett Publishers, London,UK, 2000.
[23] A.J. Katz, R. Llull, M.H. Hedrick, J.W. Futrell, Emerging approaches to the tissueengineering of fat, Clin. Plast. Surg. 26 (1999) 587–603.
[24] A. Alhadlaq, M. Tang, J.J. Mao, Engineered adipose tissue from humanmesenchymal stem cells maintains predefined shape and dimension:implications in soft tissue augmentation and reconstruction, Tissue Eng. 11(3/4) (2005) 556–566.
[25] A. Atala, Future perspectives in reconstructive surgery using tissueengineering, Urol. Clin. North Am. 26 (1999) 157–165.
[26] K.J. Walgenbach, M. Voigt, A.W. Riabikhin, C. Andree, D.J. Schaefer, T.J. Galla, G.Bjorn, Tissue engineering in plastic reconstructive surgery, Anat. Rec. 263(2001) 372.
[27] E.S. Garfein, D.P. Orgill, J.J. Pribaz, Clinical applications of tissue engineeredconstructs, Clin. Plast. Surg. 30 (2003) 485.
[28] C.W.J. Patrick, Adipose tissue engineering: the future of breast and soft tissuereconstruction following tumor resection, Semin. Surg. Oncol. 19 (2000) 302–311.
[29] S.R. Coleman, Long-term survival of fat transplants, controlled demonstrations,Aesth. Plast. Surg. 19 (1995) 421.
[30] C.W. Patrick, Tissue engineering strategies for adipose tissue repair, Anat. Rec.263 (2001) 361–366.
[31] J. Smahel, Adipose tissue in plastic surgery, Ann. Plast. Surg. 16 (1986) 444–453.
[32] C.T. Gomillion, K.J.L. Burg, Stem cells and adipose tissue engineering,Biomaterials 27 (2006) 6052–6063.
[33] F.M. Gregoire, C.M. Smas, H. S Sul, Understanding adipocyte differentiation,Physiol. Rev. 78 (3) (1998) 783–809.
[34] D. von Heimburg, M. Kuberka, R. Rendchen, K. Hemmrich, G. Rau, N. Pallua,Preadipocyte-loaded collagen scaffolds with enlarged pore size for improvedsoft tissue engineering, Int. J. Artif. Organs 26 (2003) 1064–1076.
[35] M. Kuberka, D. von Heimburg, H. Schoof, I. Heschel, G. Rau, Magnification ofthe pore size in biodegradable collagen sponges, Int. J. Artif. Organs 25 (2002)67–73.
[36] Y.-H. Lee, S.-Y. Chen, R.J. Wiesner, Y.-F. Huang, Simple flow cytometric methodused to assess lipid accumulation in fat cells, J. Lipid Res. 45 (2004) 1162–1167.
[37] T. Vazin, W.J. Freed, Human embryonic stem cells: derivation, culture, anddifferentiation: a review, Restor. Neurol. Neurosci. 28 (2010) 589–603.
[38] H. Zaehres, J.B. Kim, H.R. Schöler, Induced pluripotent stem cells, MethodsEnzymol. 476 (2010) 309–325.
[39] M. Kuberka, I. Bernemann, Y. Marquardt, G. Rau, B. Glasmacher, Influence ofseeding methods on cellular growth in 3D collagen scaffolds for tissueengineering, Biomed. Tech. 49 (2) (2004) 634–635.