cognitive neuroscience of motor learning and motor control
TRANSCRIPT

J Phys Fitness Sports Med, 1(3): 369-380 (2012)
JPFSM: Review Article
Cognitive neuroscience of motor learning and motor control
Hiroaki Masaki1* and Werner Sommer2
1 Faculty of Sport Sciences, Waseda University, 2-579-15, Mikajima, Tokorozawa, Saitama 359-1192, Japan2 Institute for Psychology, Humboldt-University of Berlin, Rudower Chaussee 18, 12489 Berlin, Germany
Received: May 22, 2012 / Accepted: August 8, 2012
Abstract In this review, focus is given to the cognitive brain functions associated with motor learning and the control of learned motor behavior, as revealed by non-invasive studies in hu-mans. After providing a definition of motor control and learning, the tasks adopted in previous studies are first introduced, and some important findings about motor behavior and pertinent theoretical models are described. Relying mainly on findings from the event-related potential (ERP) technique, but also from neuroimaging, this review focuses on motor learning and motor control in skilled action, with an emphasis on movement preparation and performance monitor-ing.Keywords : event-related potentials, motor control, motor learning, movement preparation,
performance monitoring
Introduction
Living beings and man, in particular, continuously influence their environment and adapt to varying situ-ations. Adaptation is an active function and critical for survival (and reproduction), and often involves learning, traditionally defined as a relatively permanent change in a behavior potentiality that occurs as a result of reinforced practice1), and the flexible control over and execution of what has been learned. Among the important functions for interacting with the environment are movement and actions. There is strong evidence that the learning and control of such motor skills constitutes a specific domain of competence. Thus, changes in motor behavior (motor learning) usually require practice and often last longer than newly acquired declarative knowledge and memo-ries. For example, once an individual has learned to ride a bicycle, he or she usually maintains this motor skill over many years without further practice. In contrast, one is apt to quickly forget details of historical facts when the need to retain them subsides, for example, after an exam. Such observations suggest that motor skills and declara-tive knowledge are mediated by different neurocognitive mechanisms and brain systems. Indeed, learning of motor skills has been related to such neuroanatomical structures as the cerebellum, primary motor cortex (M1), or supple-mentary motor area (SMA), whereas the acquisition of declarative knowledge requires the medial temporal lobe and diencephalic structures2,3). In this review, focus is given to the cognitive brain functions associated with motor learning and the control
of learned motor behavior, as revealed by non-invasive studies in humans. After providing a definition of motor control and learning, the tasks adopted in previous studies are introduced, and some important findings about mo-tor behavior and pertinent theoretical models, such as the schema model by Schmidt (1975)4) are described. Relying mainly on findings obtained with the event-related po-tential (ERP) technique, but also with neuroimaging, the review focuses on motor learning and motor control in skilled actions, with an emphasis on movement prepara-tion and performance monitoring. A motor skill has been defined as the ability to carry out motor acts intentionally, in order to bring about prede-termined results5); motor skills are learned only through practice. Conversely, motor learning refers to the acqui-sition of skilled movements by practice, yielding long-lasting efficient movements. Therefore, an important point in motor learning is the permanence of altered be-havior potentiality and thus the strength of motor memory representations over time. This can be assessed in reten-tion tests after a suitable interval following an acquisi-tion phase. A further important point in motor learning is the generalizability of what has been learned to a new variation of the practiced skill or to a different situation or context6); generalizability can be assessed in transfer tests. Retention and transfer tests immediately following practice may be insufficient to establish motor learning. Often, immediate tests fail to reveal any learning effect even if learning has indeed occurred as revealed by a test on the next day or later. The interval between practice and retention or transfer tests is also called an offline period, during which consolidation of motor memories occurs7).
*Correspondence: [email protected]

370 JPFSM : Masaki H and Sommer W
Theories of motor control and learning
As a framework for the present review, some impor-tant concepts and models of motor control and learning will first be explained. One should distinguish between motor control of slow and rapid movements. When rela-tively slow movements are to be executed, one can use proprioceptive and visual feedback information in order to achieve the predetermined results by correcting any discrepancies between the attempted and achieved move-ment. The closed-loop theory8) emphasizes the importance of the perceptual trace, that is, the representation of an at-tempted movement. When there is a discrepancy between the perceptual trace and feedback information about an ongoing movement, an error signal will be sent to the mo-tor control system allowing for an appropriate correction of the ongoing movement. Closed-loop control occurs only when feedback information can be used, and there-fore does not operate for purely ballistic movements (see below). Examples of closed-loop movements are figure skaters making near perfect circles while keeping their balance or car drivers performing a turn, precisely adjust-ing steering directing and driving speed. In contrast to slow movements under closed-loop con-trol, ballistic movements are thought to be controlled by motor programs that are independent of feedback, which is referred to as open-loop control. Keele (1968) formally defined a motor program as “a set of muscle commands that are structured before a movement sequence begins, and that allows the entire sequence to be carried out un-influenced by peripheral feedback”9). Thus, performers of ballistic movements cannot use proprioceptive feedback to evaluate and modify their ongoing movement before it is completed. For example, hard punches in boxing and batting in baseball are primarily controlled by motor programs and cannot be altered during their execution by means of proprioceptive or visual feedback. Hence, learn-ing ballistic movements can only rely on the information returned after movement completion, whereas for non-ballistic skill acquisition closed-loop control can be used to evaluate success even during the movement.
Schema model of Schmidt
In order to initiate a movement according to the closed-loop theory8), one has to retrieve the intended action1 from previous experience (i.e., memory trace). This the-ory suggests that one must have stored representations of many different movements. However, this idea poses two
problems. First, if every conceivable motor experience was stored in memory, storage capacity would soon be exceeded (storage problem). Second, contrary to a predic-tion of the closed-loop theory, subjects can perform even novel skilled movements relatively well on the first occa-sion (novelty problem)11,12). To overcome the storage and novelty problems, Schmidt proposed the so-called schema theory4). According to this theory, only abstract forms of movements can be stored in memory (i.e., a schema or rule). In addition, we seem to use generalized motor programs (GMPs) that regulate similar kinds of movements, belonging to the same mo-tor class determined by the invariant characteristics of actions (e.g., throwing a ball and spiking a ball in vol-leyball share similar motor properties). A GMP contains an abstract representation about the temporal structure of events (i.e., phasing), the relative force, and the sequence of events that is needed to produce the action. Like a mathematical formula a schema represents the relationship between internal motor parameters and movement outcome. To determine this relationship, only a few experiences of performing the movement may be sufficient. Once an individual has constructed a recall schema, they can inversely determine specific parameters to be applied to the GMP. Thus they can easily succeed in throwing a ball at a target located at a distance that they have never aimed at before, by determining a specific force parameter from the recall schema and applying it to the GMP associated with the throwing movement. Ac-cording to the schema theory, the recall schema dominates in ballistic movements. For slower movements the recog-nition schema is more important. The recognition schema represents the relationship between sensory consequences and movement outcomes. By means of the recognition schema one’s own movement can be evaluated11). The schema theory and closed-loop theory make dif-ferent predictions for the optimal practice schedule of a novel movement. The closed-loop theory suggests that the repetition of the same movement is essential to learn a skill, strengthening the perceptual trace; this idea seems to fit an individual’s daily experience. However, the schema theory predicts better performance after practicing several different, but related motor tasks, and variations of a task, even with fewer trial numbers than repeatedly practic-ing the same task. In a test of these predictions, Shea and Kohl (1991) examined the learning effects for four prac-tice conditions in a target force production task (20 blocks consisting of five test trials: 150 N, Experiment 1)13). In the specific + space condition, the learners practiced only the test trials (150 N exertion, 100 test trials in total) with 16-s intervals between trials (intertrial interval: ITI). In the specific + specific condition, the learners performed three additional test trials during the ITI, resulting in 4-s intervals between trials (340 test trials in total). In the specific + variable condition, the learners exerted three trials during the ITI, but the target forces were 25 or 50
1 A movement can be defined as a brief, unitary activity of a muscle or body part, whereas an act or an action pat-tern is a complex sequential behavior such as putting on a jacket10). That is, an act is made up of a particular sequence of movements.

371JPFSM : Cognitive neuroscience of motor learning and motor control
these components are of special interest for motor control and motor learning. Scalp-recorded movement-related cortical potentials (MRCPs) start about 2 s prior to self-paced voluntary movements19,20) and include several different compo-nents: (1) a gradual and bilaterally symmetrical negativity (Bereitschaftspotential: BP), (2) a steeper asymmetrical negativity (negative slope: NS’), (3) a small pre-motion positivity (PMP), (4) a transient small negative deflection (motor potential: MP), and (5) a large positive deflection (reafferent potential: RAP)21,22). The negative slow wave, including the BP and NS’, are commonly referred to as the readiness potential (RP). As a post-movement com-ponent similar to MPs (pre-movement component), the N+5021) or fpMP (frontal peak of motor potential)23) was reported to be maximal over frontal regions contralateral to the responding limb. From the asymmetry of the BP as a function of the ef-fector prepared for movement, a popular measure for ef-fector-specific response preparation has been derived, the so-called lateralized readiness potential (LRP)24). When a task requires left- or right-hand movements, for example, a button press or dynamometer squeeze, the BP (i.e., NS’) is more negative over the primary motor cortex contra-lateral to the responding hand. The pure response-hand related asymmetry can be extracted by (1) calculating the difference between the contra- and ipsilateral EEG for each (required) hand and (2) by averaging these different waves. Negative deflections of the LRP reflect the activa-tion of the correct response hand in a given task, whereas positive deflections indicate the activation of the incorrect hand. Source localization and intracranial recordings indi-cate that the LRP is generated at least in part by the M124). This is also in line with the polarity inversion of the LRP when lateralized foot responses are required, because the representation of the foot muscles in M1 is located on the medial banks within the fissura interhemispherica, pro-jecting its negativity to the hemisphere ipsilateral to the activated foot25). This representation is specific to the M1 and not found in other motor areas. The BP is presumed to be part of a further ERP com-ponent, which is elicited when a warning stimulus or cue announces a second stimulus, which requires a response. Between the warning stimulus and the response signal, there is usually a negativity with a maximum at central re-cording sites, which has been termed the contingent nega-tive variation (CNV)26). When the inter-stimulus interval is long enough, a first negativity, following the warning signal, can be distinguished from a negativity preceding the response signal. The latter has been related to the BP even if such negativities can be observed also in situa-tions without response requirements. Both LRP and CNV have been shown to be useful in exploring motor control and motor programming, and are reviewed following.
N above or below the target force on test trials (i.e., 100, 125, 175, and 200 N). In the specific + alternative condi-tion, the learners performed a tracking task during the middle 12 s of the ITI. Poorer performance (i.e., larger errors) was found in the specific + variable condition than in the other conditions during the acquisition phase. How-ever, on a retention test on the next day, the specific + variable condition showed the best performance, whereas the specific + specific condition was worst (the two other conditions showed intermediate results). Similar results were also found in Experiment 213), and these results were consistent with their previous study14). This inversion of performance, between training and retention test, supports the predictions of the schema theory. The inversion of performance on retention tests or posi-tive transfer can also be explained by what is known as contextual interference, originally discovered in verbal learning studies15,16). Contextual interference is defined as “the effect on learning of the degree of functional interfer-ence found in a practice situation when several tasks must be learned and are practiced together (p. 244)”17). Accord-ing to contextual interference, an increased amount of inter-task interference will result in better performance on retention tests and therefore random practice should show better performance than blocked practice. For example, in learning the skills of tennis serve, volley, and backhand stroke, randomly-mixed practice of these movements should induce contextual variance, resulting in better learning than block-wise practice of each movement. Supporting this counterintuitive prediction of contextual interference, previous studies have demonstrated the ad-vantage of random practice in motor learning18).
Event-related potentials: tools of cognitive brain function
There are many methods for studying neural mecha-nisms of motor learning and motor control, such as functional magnetic resonance imaging (fMRI), positron emission tomography (PET), recordings of the electro-myogram (EMG), electroenphalogram (EEG), and event-related brain potentials (ERPs), transcranial magnetic stimulation (TMS), and the elicitation of reflexes. Be-cause of their relative ease and noninvasiveness, excellent time resolution, relative mobility, and inexpensiveness, ERPs are of special importance. Therefore, we will intro-duce this technique in some detail. ERPs are derived from EEG recordings by averaging selected time epochs synchronized to an event, for ex-ample, to a stimulus, muscle activation, or a button press. The average signal derived by this process consists of a complex waveform with positive and negative deflections during certain time intervals and with a specific voltage distribution across the head surface. Using such character-istics, so-called components can be defined; and in many cases they can be related to putative cognitive subpro-cesses that take place in specific brain systems. Some of

372 JPFSM : Masaki H and Sommer W
resonance imaging: fMRI) was found for both sequences; however, after three weeks of daily practice, the extent of activation in M1 associated with the trained sequence was larger than for the unpracticed sequence31). Although most cognitive neuroscientific studies of mo-tor learning used simple movement tasks (e.g., sequen-tial button presses or serial reaction time paradigms), an interesting study by Grafton and colleagues recorded PET during execution of a pursuit rotor task where par-ticipants tried to keep the tip of a stylus in contact with the target area on a rotating disk32). This study showed that motor execution itself is related to widely distributed networks in the cortex, basal ganglia, and cerebellum; whereas longitudinal increases in neuronal activity, rep-resenting learning, was found in the left motor cortex, left SMA, and pulvinar thalamus. Because Grafton et al. (1992) did not investigate the changes of these regions in a retention test, the true learning effects remain unclear. Interestingly, the authors failed to see any longitudinal changes in activity of the cerebellum in their pursuit ro-tor task. In their follow-up study where the pursuit rotor task was conducted on two consecutive days, they found that the activity of the putamen was still related to learn-ing on Day 233). Thus, the putamen seems to be involved in learning, which is consistent with other findings about this region29). Doyon and colleagues found similar results27) in highly skilled knitters by comparing brain activity (recorded by fMRI) during fully-automatized skillful stitching and a newly learned stitching. The authors found that the over-learned stitch induced activation of the parietal cortex, SMA, and putamen, but not the cerebellum; whereas the newly learned stitch activated the cerebellum as well as the putamen and globus pallidus. Differences between overlearned and learned stitching skills were found in activation of the basal ganglia and premotor regions as automaticity effect. Knitting is a MSL task and can be contrasted with other tasks such as target reaching and motor adaptation (e.g. joystick use). In these tasks, activ-ity in the putamen and motor areas was found during the early stages of learning, but activity in the cerebellum and parietal cortex was increased only during the late stages of learning27). To interpret these different findings about cerebellar activity, the model proposed by Doyon and Be-nali (2005)27) is helpful. They suggested that MSL relies on the cortico-striatal system, whereas motor adaptation relies on the cortico-cerebellar system (Fig. 1). Motor learning processes have been classified into three major stages8,34). (1) Early motor learning is also called the cognitive or verbal-motor stage, and is characterized by considerable improvement of a novel task, but incon-sistence in movement. (2) During the associative or mo-tor stage, less improvement is found, but movement, per se, becomes more consistent. (3) During the autonomous stage, learners automatically perform the task, and may even pay attention to other things during its execution.
Neural basis of motor learning
According to Doyon and Benali (2005)27), tasks in motor learning studies can be classified into two main catego-ries: (1) motor sequence learning (MSL) that is related to the acquisition of motor skills, and (2) motor adaptation that is related to the capacity to compensate for environ-mental changes. In addition, Willingham (1999) suggest-ed that four processes are involved in motor control28): (1) strategic processes associated with selecting environmen-tal goals, (2) perceptual-motor integration, (3) sequencing spatial targets, and (4) dynamic processes that translate the sequence of spatial targets into muscle activity. Mo-tor learning is necessary whenever one or more of these motor control processes needs improvement because the action goal cannot be achieved. Thus, when current per-ceptual-motor integration is altered – for example when wearing prism spectacles – a new relationship between vision and proprioception must be learned. Willingham (1999) suggested that learning the four motor control processes relies on different brain systems28). Thus, the dorsolateral frontal cortex appears to be involved in strategic process, posterior parietal cortex in perceptual-motor integration, and both the basal ganglia – especially the putamen and supplementary motor area (SMA) – are important for learning motor sequences, and interneurons in the spinal cord are responsible for dynamic movement control. For understanding the neural basis of motor learning, it is helpful to consider simple arbitrary associations between stimuli and responses. In sensorimotor learn-ing a categorical mapping between a sensory cue and an already-acquired skilled movement is made; therefore sensorimotor learning differs from acquiring a new motor skill29). In rhesus monkeys, activity of the premotor cor-tex changes during the acquisition of arbitrary sensory-motor associations, suggesting that the premotor cortex plays a role in the selection of movements on the basis of stimulus context30). Lalazar and Vaadia (2008) reviewed previous findings and emphasized the involvement of the hippocampus, the caudate, the globus pallidus, and the prefrontal cortex in learning the mapping between an ar-bitrary cue and a rewarded action during the early stages of learning. On the other hand, during the late stages of learning, the putamen showed efficient and selective fir-ing, representing retrieval of the association. Concerning more complex learning that alters kinematics, neuronal changes in the SMA relate to gains during early phases of motor learning, whereas changes in M1 seem to be in-volved in the later and slower phases of motor learning29). The activation of M1 in later phases of learning was also supported by other studies. Using two rapid sequences of finger movements with the non-dominant hand (a train-ing sequence and a mirror-reversed control sequence), Karni et al. (1995) found that in the first session (Day 1) a comparable extent of M1 activity (in functional magnetic
373JPFSM : Cognitive neuroscience of motor learning and motor control
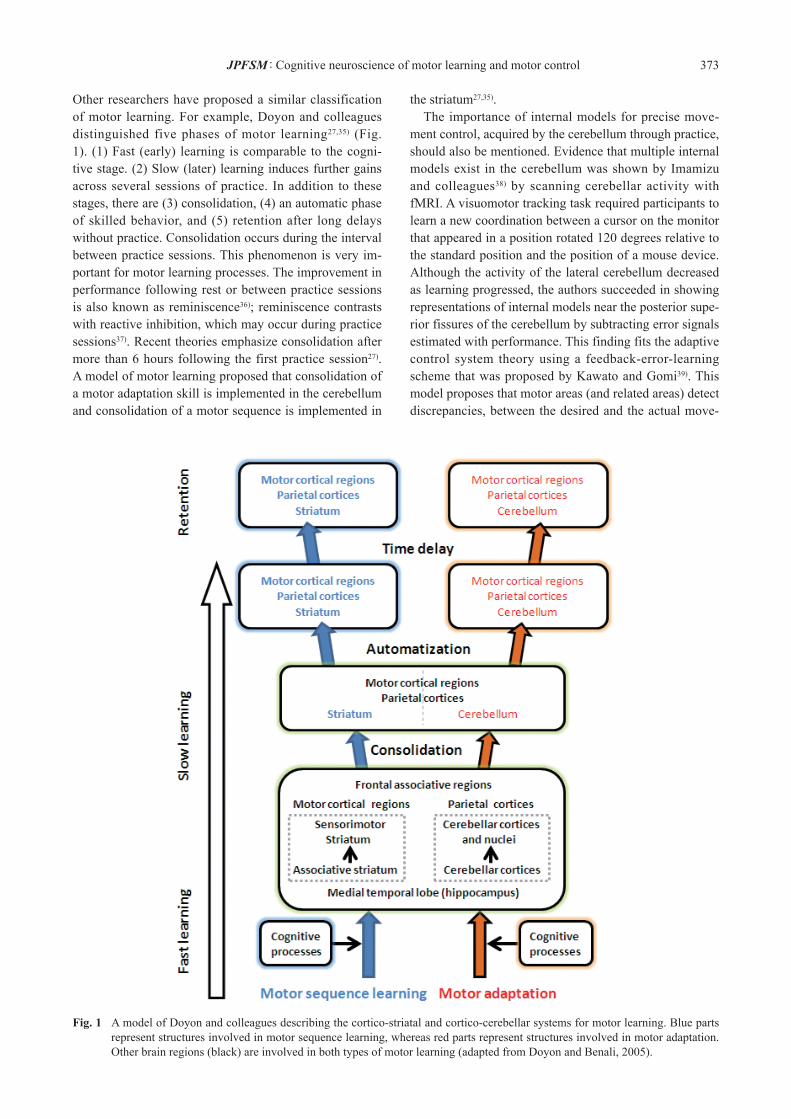
Other researchers have proposed a similar classification of motor learning. For example, Doyon and colleagues distinguished five phases of motor learning27,35) (Fig. 1). (1) Fast (early) learning is comparable to the cogni-tive stage. (2) Slow (later) learning induces further gains across several sessions of practice. In addition to these stages, there are (3) consolidation, (4) an automatic phase of skilled behavior, and (5) retention after long delays without practice. Consolidation occurs during the interval between practice sessions. This phenomenon is very im-portant for motor learning processes. The improvement in performance following rest or between practice sessions is also known as reminiscence36); reminiscence contrasts with reactive inhibition, which may occur during practice sessions37). Recent theories emphasize consolidation after more than 6 hours following the first practice session27). A model of motor learning proposed that consolidation of a motor adaptation skill is implemented in the cerebellum and consolidation of a motor sequence is implemented in
the striatum27,35). The importance of internal models for precise move-ment control, acquired by the cerebellum through practice, should also be mentioned. Evidence that multiple internal models exist in the cerebellum was shown by Imamizu and colleagues38) by scanning cerebellar activity with fMRI. A visuomotor tracking task required participants to learn a new coordination between a cursor on the monitor that appeared in a position rotated 120 degrees relative to the standard position and the position of a mouse device. Although the activity of the lateral cerebellum decreased as learning progressed, the authors succeeded in showing representations of internal models near the posterior supe-rior fissures of the cerebellum by subtracting error signals estimated with performance. This finding fits the adaptive control system theory using a feedback-error-learning scheme that was proposed by Kawato and Gomi39). This model proposes that motor areas (and related areas) detect discrepancies, between the desired and the actual move-
Fig. 1 A model of Doyon and colleagues describing the cortico-striatal and cortico-cerebellar systems for motor learning. Blue parts represent structures involved in motor sequence learning, whereas red parts represent structures involved in motor adaptation. Other brain regions (black) are involved in both types of motor learning (adapted from Doyon and Benali, 2005).

374 JPFSM : Masaki H and Sommer W
ment, as error signals that are conveyed to the cerebellum in order to regulate movement by feedback control. On the other hand, even without feedback, the model can produce near-perfect movements using an inverse model implemented in the cerebellum (feed-forward control). This system might be responsible for the underlying neural mechanisms of representations as proposed by the closed-loop and the schema theories.
ERP research on motor learning
So far, only a few ERP studies have tested motor learn-ing. Taylor (1978) found that the Bereitschaftspotential (BP) steadily increased only during the acquisition stage of a motor learning skill (i.e., six button presses in a spe-cific sequential pattern), but diminished over right-central (C4, ipsilateral side to responding hand) and midfrontal (Fz) regions after performance had reached asymptote40). This study adopted a sequential task where participants learned to chunk the response sequence, resulting in shorter response times. Based on other findings28,29), the increasing amplitude, during the early phase of learning, seemed to be due to increased activation of the SMA, a potential generator of the BP; and the diminishing am-plitude, after reaching performance asymptote, seemed to include less activation of both the SMA and M1. This study suggests that motor activity might be reduced after optimization of performance, although the result could also be explained by diminished attention demands after learning. Takasawa (1989) investigated how the BP alters as a function of practice in a target force production task41). Participants practiced exerting a target force (ranging 16 to 20 N for an easy task, and 17 to 19 N for a dif-ficult task) by isometric flexion of the index fingers (3 blocks consisting of 60 trials for each task on different days). The characteristics of this task are independent of visual information and only rely on feed-forward control of muscular contraction. Performance did not improve through practice, probably because the task was inher-ently difficult to learn. The BP did not systematically change in amplitude through practice, but it started earlier for the more difficult task. Following this study, Masaki (1997 unpublished PhD dissertation) tested the same force production task (target force range was 12 to 16 N) on 5 consecutive days. Although performance correctness increased, especially from Day 1 to Day 2, the amplitude of the BP did not change with practice. On the other hand, the BP started earlier as a function of practice, suggesting that motor programming implemented by the SMA needs time to be modified during the early stage of learning. In an fMRI study, Cross and colleagues investigated the neural underpinnings of contextual interference during learning of a set of three 4-element sequences with the left hand (right-handed participants), comparing blocked and random practice42). During training, the random prac-
tice group performed worse than the block group; but on the retention task, the random practice group performed better. Participants were allowed to rehearse the instructed sequence of button presses before they initiated the move-ment. During this study time the random group showed greater activity in the sensorimotor and premotor regions compared to the block group. These results suggest that reconstruction of the motor program, which should have been more in demand in the random group, relies on mo-tor related areas. This study, as well as the above-men-tioned ERP studies, indicates that motor preparation is a key process to investigating motor learning.
Motor control
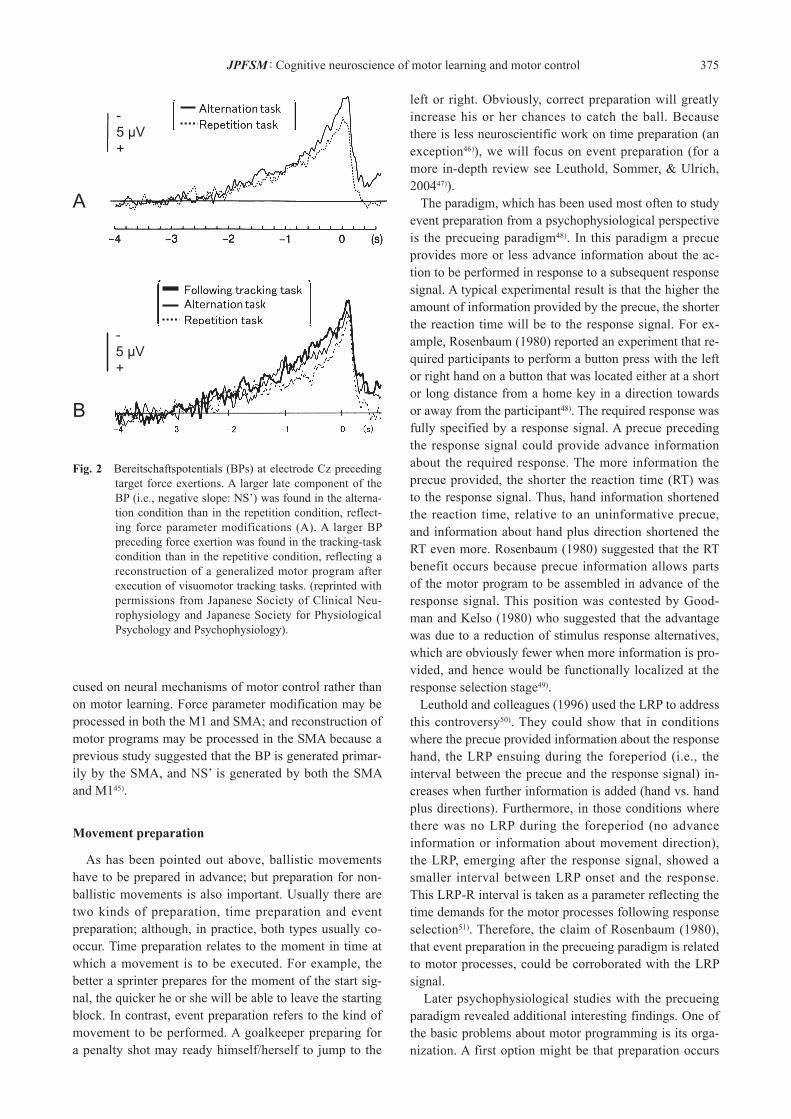
As mentioned above, it has been suggested that motor preparation is reflected in activity of the brain regions as-sociated with motor control and learning. Therefore, the BP has been investigated as an index of activity of those brain regions. Indeed, focusing on the BP, several studies have investigated brain activities associated with retrieval of the generalized motor program. In a target force pro-duction task, Masaki and colleagues compared a task rep-etition condition where participants repeatedly produced the same target force (i.e., 13 N) and an alternation con-dition where they alternatively produced three different target forces (i.e., 5, 13 and 21 N) in every trial43). When participants repeatedly exerted the same target force, they were considered to repeatedly apply the same force parameter to the motor program. Of course, they had to modify the force parameter at each trial using feedback information (knowledge of results), but the modifica-tion must have been less effortful than in the alternation condition, in which they had to apply a different force parameter to the motor program in every trial. Masaki et al. (1997) found a larger late component of the BP (i.e., NS’)21) in the alternation condition (in the 13 N target tri-als) than in the repetition condition (Fig. 2A). Following up this experiment, Masaki and colleagues investigated the effect of reconstructing motor programs on the BP44). The authors hypothesized that when differ-ent tasks implemented by distinct motor programs are performed alternatively, the motor programs for each task have to be reconstructed in every trial. Masaki et al. (1998) inserted visuomotor tracking tasks in between force pro-duction trials. They found a larger BP preceding force ex-ertion in the visuomotor tracking condition than in the re-petitive condition where only a force production task was executed repeatedly (Fig. 2B). In terms of morphology, the increased BP emerged earlier than in the force param-eter modification study, representing the reconstruction of motor programs in accordance with the temporal relation-ship between motor programming and response parameter supply. It should be noted that these studies counter-balanced the order of conditions across participants. Thus, they fo-
375JPFSM : Cognitive neuroscience of motor learning and motor control
cused on neural mechanisms of motor control rather than on motor learning. Force parameter modification may be processed in both the M1 and SMA; and reconstruction of motor programs may be processed in the SMA because a previous study suggested that the BP is generated primar-ily by the SMA, and NS’ is generated by both the SMA and M145).
Movement preparation
As has been pointed out above, ballistic movements have to be prepared in advance; but preparation for non-ballistic movements is also important. Usually there are two kinds of preparation, time preparation and event preparation; although, in practice, both types usually co-occur. Time preparation relates to the moment in time at which a movement is to be executed. For example, the better a sprinter prepares for the moment of the start sig-nal, the quicker he or she will be able to leave the starting block. In contrast, event preparation refers to the kind of movement to be performed. A goalkeeper preparing for a penalty shot may ready himself/herself to jump to the
left or right. Obviously, correct preparation will greatly increase his or her chances to catch the ball. Because there is less neuroscientific work on time preparation (an exception46)), we will focus on event preparation (for a more in-depth review see Leuthold, Sommer, & Ulrich, 200447)). The paradigm, which has been used most often to study event preparation from a psychophysiological perspective is the precueing paradigm48). In this paradigm a precue provides more or less advance information about the ac-tion to be performed in response to a subsequent response signal. A typical experimental result is that the higher the amount of information provided by the precue, the shorter the reaction time will be to the response signal. For ex-ample, Rosenbaum (1980) reported an experiment that re-quired participants to perform a button press with the left or right hand on a button that was located either at a short or long distance from a home key in a direction towards or away from the participant48). The required response was fully specified by a response signal. A precue preceding the response signal could provide advance information about the required response. The more information the precue provided, the shorter the reaction time (RT) was to the response signal. Thus, hand information shortened the reaction time, relative to an uninformative precue, and information about hand plus direction shortened the RT even more. Rosenbaum (1980) suggested that the RT benefit occurs because precue information allows parts of the motor program to be assembled in advance of the response signal. This position was contested by Good-man and Kelso (1980) who suggested that the advantage was due to a reduction of stimulus response alternatives, which are obviously fewer when more information is pro-vided, and hence would be functionally localized at the response selection stage49). Leuthold and colleagues (1996) used the LRP to address this controversy50). They could show that in conditions where the precue provided information about the response hand, the LRP ensuing during the foreperiod (i.e., the interval between the precue and the response signal) in-creases when further information is added (hand vs. hand plus directions). Furthermore, in those conditions where there was no LRP during the foreperiod (no advance information or information about movement direction), the LRP, emerging after the response signal, showed a smaller interval between LRP onset and the response. This LRP-R interval is taken as a parameter reflecting the time demands for the motor processes following response selection51). Therefore, the claim of Rosenbaum (1980), that event preparation in the precueing paradigm is related to motor processes, could be corroborated with the LRP signal. Later psychophysiological studies with the precueing paradigm revealed additional interesting findings. One of the basic problems about motor programming is its orga-nization. A first option might be that preparation occurs
A
B
-5 µV+
-5 µV+
Fig. 2 Bereitschaftspotentials (BPs) at electrode Cz preceding target force exertions. A larger late component of the BP (i.e., negative slope: NS’) was found in the alterna-tion condition than in the repetition condition, reflect-ing force parameter modifications (A). A larger BP preceding force exertion was found in the tracking-task condition than in the repetitive condition, reflecting a reconstruction of a generalized motor program after execution of visuomotor tracking tasks. (reprinted with permissions from Japanese Society of Clinical Neu-rophysiology and Japanese Society for Physiological Psychology and Psychophysiology).

376 JPFSM : Masaki H and Sommer W
in an abstract way, independent of the specific muscles to be specified. Since the LRP emerges at least in part from the M1, and possibly the premotor area24,52), one would not expect the LRP to increase when, in addition to the effector limb, concrete muscles can also be prepared. An increase in the LRP, with more preliminary information observed by Leuthold et al. (1996), already argues against this possibility and supports muscle-specific prepara-tion50). According to such a muscle-specific view of motor preparation, the LRP should increase further if additional parameters can be prepared. Muscle-specific preparation might come in several variants. Several muscle-specific parameters might be prepared in parallel. In this case the effects of additional pre-specified movement parameters would be additive. Alternatively, motor preparation might be organized hierarchically. In this case, a given param-eter might show an effect in the LRP only if certain other parameters are also set. Ulrich, Leuthold, and Sommer (1998) investigated these options in a precueing paradigm by requiring strong or weak flexion or extension move-ments with the left or right index finger53). As before, movement parameters, in addition to response hand, further increased the LRP, supporting muscle-specific preparation. It is important to note, however, this increase occurred only if the movement was fully specified by the precue. This finding supports a strictly hierarchical orga-nization of response programming at a level reflected in the LRP (M1). At a more central neural level, the findings were significant in indicating parallel programming: The CNV increased monotonically as a function of the number of pre-specified response parameters. Ulrich et al. (1998) interpreted these findings to indicate that there are two levels of motor programming, which are organized in dif-ferent ways. Centrally, there may be parallel organization of motor programs; whereas at the implementation level (peripheral level known as M1) of motor programming, it is organized in a hierarchical fashion. That peripheral-level organization may not always follow a strictly hierarchical principle was shown by Wild-Wall et al.54). These authors combined preliminary information about response hand, response finger (index, middle), and movement direction. At variance with the findings of Ulrich et al. (1998), hand plus finger increased the LRP above hand information, even when the move-ment was not fully specified – that is, when no informa-tion about movement direction was available. These find-ings indicate that the organization of peripheral response preparation may differ when effectors of the body, rather than functional parameters, are concerned. An important issue is whether the effects of the amount of precue information on LRP amplitude are data-limited or strategic. For example, participants might be taking advantage of the hand information more when it is ac-companied by other parameters just because they find preparation more rewarding. Therefore, Sangals, Sommer, and Leuthold (2002) attempted to encourage participants
to make maximum use of precue information by exerting time pressure or by blocking task conditions55). However, although these manipulations increased motor preparation in general, the effects of the amount of preliminary infor-mation remained unaltered, which strongly argues that strategies of the participants are not the basis of the previ-ous findings. Another interesting issue in the context of motor prepa-ration is its sensitivity to intervening events. Sangals and Sommer (2010) investigated this issue in three experi-ments where participants prepared to respond with the left or right hand, but also had to perform other tasks during the foreperiod (FP)56). At a central level (CNV), the inter-vening task seemed to cause the recruitment of additional processing resources enhancing CNV amplitudes. At the peripheral level (LRP), participants seemed to shy away from motor preparation when there was an increased like-lihood for an intervening task being required. In addition, there was evidence of a bottleneck-like postponement of response preparation. This study shows that motor prepa-ration not only requires due effort – it is also subject to interference. In order to make the best use of preparation, it should, therefore, be shielded from interfering stimuli or multitasking. That participants appeared to be conservative in their tendencies to prepare for a specific action was also shown by Scheibe et al.57) Here the precue did not convey infor-mation about a movement parameter, but about a response probability on either the left or right side, which varied from p = .50 over .75 to 1.00 for a response with a given response hand. An LRP developed only if the response hand was certain, but not when it was more unikely (p = .75). In contrast, the CNV parametrically increased with response probability. In a subsequent combined fMRI/ERP study, it was shown that these prior probability ef-fects on the CNV are related to the dorsolateral prefrontal cortex, the inferior frontal gyrus, and the inferior parietal lobule58). While the studies of motor preparation reviewed so far mainly used rather simple movements in artificial laboratory tasks, recently, the investigation of move-ment preparation was extended to more complex actions. Leuthold and Jentzsch (2009) showed that also for rapid aimed movements full precue information information results in larger late-CNV amplitudes than partial in-formation59). Whereas an older study had not found any effect from preparing various kinds of movements (e.g., single finger taps or different finger tap sequences) on the CNV60), Schröter and Leuthold (2009) showed the effect of sequence length on the CNV, with larger amplitudes for three-key responses compared to one-key responses61). Because the CNV is generated in the SMA and cingu-late motor area, the authors argued that sequential motor preparation is not limited to effector-specific areas. Larger CNV amplitudes in a more complex action, relative to a simple finger opposition condition, were also reported62).

377JPFSM : Cognitive neuroscience of motor learning and motor control
Recently Frömer and colleagues attempted to translate the precuing paradigm to a more naturalistic sport-related situation, namely “aimed throwing”63). The throwing situ-ation was simulated with a Nintendo® remote control (“Wii”), held and launched like a dart in a virtual real-ity scenario. All participants were trained to accurately “throw” the Wii at a target projected onto a screen. During the experimental session, participants stood upright while multi-channel EEG was recorded. The precue consisted of a target disc, which was either small or large, provid-ing a range of throwing difficulty. After the start of target presentations, participants had two seconds to aim their throw. After this interval, the bull’s eye of the target disc changed its color and then disappeared, encouraging im-mediate initiation of the ballistic throwing movement. A first important finding of this pilot study was that it is in-deed possible to record the EEG in such a situation where the participant is largely unrestrained. Another important finding was that the CNV, during the aiming period, was indeed larger for the more difficult condition (smaller tar-get), which likely required more programming effort. Fu-ture work can now focus not only on preparatory activity in trained participants, but also on neural activity during motor learning and on the role of feedback- and outcome-related neural activity during the learning process.
Performance monitoring
In skilled motor tasks learners develop the capability of evaluating their own movements, especially in those tasks that rely on open-loop control64). It is important to develop a perceptual trace or internal model to acquire a novel skill, because learners have to detect the discrep-ancy between representations of the required movement and the actual movement. This suggests that monitoring of performance is a critical process in acquiring a novel motor skill. Findings have accumulated, supporting the existence of performance monitoring in the brain, since the discovery of error (-related) negativity (Ne or ERN)65,66). The ERN is a negative-going deflection in the ERP that emerges with the commission of an erroneous response. It shows a fronto-central scalp distribution, reflecting neural activity of the anterior cingulate cortex (ACC)67), and is widely thought to be functionally related to performance moni-toring including error-detection65,66) and response-conflict detection68). Response-conflict emphasizes the crosstalk interference that occurs when two response activations overlap, during the parallel processing of incongruent stimuli, and elicits the ERN69). On the other hand, there is mounting evidence against the response-conflict ac-count70-73), Recently Anguera and colleagues tested the motor learning process in terms of performance monitoring in a visuomotor tracking task74). They found significant chang-es in performance with practice that were reflected in
diminishing errors from the early to late adaptation stage. In addition, a larger negativity in fronto-central regions was found during the early learning phase, as compared to a baseline or late learning phase, suggesting that more error monitoring is needed during the early stage. This study suggests the medial frontal negativity (ERN) to be a promising tool for motor learning research. It is also well known that feedback signals conveying a negative outcome elicit feedback-related negativity (FRN)75). FRN is thought to be similar to ERN in terms of scalp distribution, functional significance and neural generators. The reinforcement-learning theory of the FRN asserts that FRN represents a dopaminergic negative feedback reinforcement-learning signal generated when response outcomes are worse than expected76). Given that the neural mechanisms of motor learning are related to the basal ganglia, FRN-reflecting dopaminergic activity might be a good tool for motor learning studies. Indeed, even prior to the study of Miltner et al.75), Takasawa et al. (1990) investigated motivational processes during motor learning (target force production task) focusing on the FRN77). They found the FRN to increase when correct performance was followed by monetary reward. This study suggests the possibility of observing motivational aspects relying on basal ganglia, which is a critical brain system for motor learning. Another interesting ERP component for studies of motor learning is the N+50 (fpMP)21,23) seen in frontal regions contralateral to the responding limb. This component in-creases with the rate of force development78) and effort to perform a task79). Masaki et al. recorded the motor-elicited negativity that is thought to correspond to N+50, peaking at about 60 ms after force onset80). In a coincident-timing task, the peak force exerted by index finger flexion was to temporally coincide with the specific position of a visual stimulus rotating either slow or fast on a clock face. The motor-elicited negativity was larger in a condition where timing errors were relatively small compared to other conditions. This suggests that N+50 might partially rep-resent a monitoring or appraisal process of performance, differing from the ERN. In addition, it is possible that the larger motor-elicited negativity goes along with better performance. We should point out the difference between the ERN and N+50 in terms of function and generator source. Contrary to the ERN, which is considered to reflect error monitoring or response conflict monitoring, N+50 might reflect correctness monitoring or representations of the movement as well as kinematic factors80). Although it does not necessarily mean that N+50 exclusively represents better performance, further study is needed to examine if the negativity also can represent deviation errors (i.e., the degree of deviation from the target) that are observed in most motor-learning tasks. In addition, the ERN and N+50 do not seem to share neural generators. It is sug-gested that the ERN is generated by the rostral cingulate

378 JPFSM : Masaki H and Sommer W
motor area67), whereas, N+50 is thought to represent activ-ity of the sensorimotor cortex, suggesting Brodmann area 3 as its source45). Nevertheless, the generator of N+50 still remains somewhat unclear. N+50 also differs from the so-called correct-response negativity (CRN)81), because the CRN does not show an asymmetric distribution. As reviewed above, the internal models in cerebellum require error signals from motor and motor-related areas for the execution of desired movements39). Thus, it is pos-sible that the M1 functions as an error detection system of discrepancies between desired movements and actual movements. On the other hand, previous studies suggest that during late phases of motor learning, the M1 seems to be responsible for representations of trained motor skills31). Given that N+50 represents the activity of M1, as well as Brodmann area 3, it is also likely that the negativ-ity functions as a template of motor-skill representations for error detection.
Conclusions
This selective review has shown that cognitive neurosci-entific studies have contributed considerably to resolving controversial and unclear issues related to motor learn-ing and motor control. This does not only concern the elucidation of the underlying neuroanatomical systems but – more importantly – the mechanisms implemented in these systems. Therefore, imaging and ERP studies have already proven to be valuable tools for deepen-ing our understanding of motor learning and control. As these approaches are adapted to increasingly naturalistic situations, considerable progress can be expected in the understanding of motor learning and control, and in the application of this knowledge in sports science, motor re-habilitation, and educational fields.
Acknowledgments
This study was partly supported by a Grant-in-Aid for Scientific Research (C)24530925 from the Japan Society for the Promo-tion of Science to H. Masaki, and two Grants-in-Aid [the Global Center of Excellence Program (GCOE) and Kibankeisei (2010)] from the Ministry of Education, Culture, Sports, Science and Technology.
References
1) Kimble GA. 1961. Hilgard and Marquis’ Contitioning and Learning. Appleton-Century-Crofts, Inc, New York.
2) Squire LR, Wixted JT. 2011. The cognitive neuroscience of human memory since H.M. Annu Rev Neurosci 34: 259-288.
3) Squire LR. 1987. Memory and Brain. New York: Oxford Univ Press.
4) Schmidt RA. 1975. A schema theory of discrete motor skill learning. Psychol Rev 82: 225-260.
5) Whiting HA. 1975. Concepts in Skill Learning. Lepus Book, London.
6) Kantak SS, Winstein CJ. 2012. Learning-performance dis-tinction and memory processes for motor skills: a focused review and perspective. Behav Brain Res 228: 219-231.
7) Hallgató E, Győri-Dani D, Pekár J, Janacsek K, Nemeth D. 2012. The differential consolidation of perceptual and motor learning in skill acquisition. Cortex. In press.
8) Adams JA. 1971. A closed-loop theory of motor learning. J Mot Behav 3: 111-150.
9) Keele SW. 1968. Movement control in skilled motor perfor-mance. Psychol Bull 70: 387-403.
10) Breedlove SM, Watson NV, Rosenzweig MR. 2010. Biologi-cal Psychology: An Introduction to Behavioral, Cognitive, and Clinical Neuroscience. Sinauer Associates Inc., Sunder-land, MA, USA.
11) Schmidt RA, Wrisberg CA. 2000. Motor learning and perfor-mance. Human Kinetics, Champaign, Ill.
12) Schmidt RA, Lee TD. 1999. Motor control and learning. Hu-man Kinetics, Champaign.
13) Shea CH, Kohl RM. 1991. Composition of practice: influ-ence on the retention of motor skills. Res Q Exerc Sport 62: 187-195.
14) Shea CH, Kohl RM. 1990. Specificity and variability of prac-tice. Res Q Exerc Sport 61: 169-177.
15) Battig WF. 1966. Facilitation and interference. In: Acquisi-tion of skill (EA Bilodeau ed), New York: Academic Press: pp. 215-244.
16) Battig WF. 1972. Intratask interference as a source of facili-tation in transfer and retention. In: Topics in learning and performance (RF Thompson and JF Voss eds), New York: Academic Press pp. 131-159.
17) Magill RA, Hall KG. 1990. A review of the contextual in-terference effect in motor skill acquisition. Hum Mov Sci 9: 241-289.
18) Shea JB, Morgan RL. 1979. Contextual interference effects on the acquisition, retention, and transfer of a motor skill. J Exp Psychol Hum Learn 5: 179-187.
19) Kornhuber HH, Deecke L. 1965. Hirnpotentialänderungen bei Willkürbewegungen und passiven Bewegungen des Menschen: Bereitschaftspotential und reafferente Potentiale. Pflugers Arch 284: 1-17.
20) Vaughan HGJ, Costa LD, Gilden L, Schimmel H. 1965. Identification of sensory and motor components of cerebral activity in simple reactiontime tasks. Proc 73rd Conv Amer Psychol Assoc 1: 179-180.
21) Shibasaki H, Barrett G, Halliday E, Halliday AM. 1980. Com-ponents of the movement-related cortical potential and their scalp topography. Electroencephalogr Clin Neurophysiol 49: 213-226.
22) Deecke L, Grozinger B, Kornhuber HH. 1976. Voluntary fin-ger movement in man: Cerebral potentials and theory. Biol Cybern 23: 99-119.
23) Tarkka IM, Hallett M. 1991. Topography of scalp-recorded motor potentials in human finger movements. J Clin Neuro-physiol 8: 331-341.
24) Coles MGH. 1989. Modern mind-brain reading: Psycho-physiology, physiology, and cognition. Psychophysiology 26: 251-269.
25) Brunia CHM, Vingerhoets AJJM. 1980. CNV and EMG pre-ceding a plantar flexion of the foot. Biol Psychol 11: 181-191.
26) Walter WG, Cooper R, Aldridge VJ, McCallum WC, Winter AL. 1964. Contingent negative variation: An electric sign of

379JPFSM : Cognitive neuroscience of motor learning and motor control
Movement-Related Cortical Potentials: fMRI-Constrained EEG Dipole Source Analysis. NeuroImage 17: 161-173.
46) Müller-Gethmann H, Ulrich R, Rinkenauer G. 2003. Locus of the effect of temporal preparation: Evidence from the lateral-ized readiness potential. Psychophysiology 40: 597-611.
47) Leuthold H, Sommer W, Ulrich R. 2004. Preparing for action: Inferences from CNV and LRP. J Psychophysiol 18: 77-88.
48) Rosenbaum DA. 1980. Human movement initiation: Specifi-cation of arm, direction, and extent. J Exp Psychol Gen 109: 444-474.
49) Goodman D, Kelso JAS. 1980. Are movements prepared in parts? Not under compatible (naturalized) conditions. J Exp Psychol Gen 109: 475-495.
50) Leuthold H, Sommer W, Ulrich R. 1996. Partial advance information and response preparation: Inferences from the lateralized readiness potential. J Exp Psychol Gen 125: 307-323.
51) Masaki H, Wild-Wall N, Sangals J, Sommer W. 2004. The functional locus of the lateralized readiness potential. Psy-chophysiology 41: 220-230.
52) Leuthold H, Jentzsch I. 2001. Neural correlates of advance movement preparation: a dipole source analysis approach. Cog Brain Res 12: 207-224.
53) Ulrich R, Leuthold H, Sommer W. 1998. Motor programming of response force and movement direction. Psychophysiology 35: 721-728.
54) Wild-Wall N, Sangals J, Sommer W, Leuthold H. 2003. Are fingers special? Evidence about movement preparation from event-related brain potentials. Psychophysiology 40: 7-16.
55) Sangals J, Sommer W, Leuthold H. 2002. Influences of pre-sentation mode and time pressure on the utilisation of ad-vance information in response preparation. Acta Psychol (Amst) 109: 1-24.
56) Sangals J, Sommer W. 2010. The Impact of Intervening Tasks on Response Preparation. J Exp Psychol Hum Percept Per-form 36: 415-429.
57) Scheibe C, Schubert R, Sommer W, Heekeren HR. 2009. Electrophysiological evidence for the effect of prior probabil-ity on response preparation. Psychophysiology 46: 758-770.
58) Scheibe C, Ullsperger M, Sommer W, Heekeren HR. 2010. Effects of Parametrical and Trial-to-Trial Variation in Prior Probability Processing Revealed by Simultaneous Electro-encephalogram/Functional Magnetic Resonance Imaging. J Neurosci 30: 16709-16717.
59) Leuthold H, Jentzsch I. 2009. Planning of rapid aiming move-ments and the contingent negative variation: Are movement duration and extent specified independently? Psychophysiol-ogy 46: 539-550.
60) Prescott J. 1986. The effects of response parameters on CNV amplitude. Biol Psychol 22: 107-135.
61) Schröter H, Leuthold H. 2009. Motor programming of rapid finger sequences: Inferences from movement-related brain potentials. Psychophysiology 46: 388-401.
62) Kranczioch C, Mathews S, Dean P, Sterr A. 2010. Task com-plexity differentially affects executed and imagined move-ment preparation: evidence from movement-related poten-tials. PlosOne 5: e9284.
63) Frömer R, Hafner V, Sommer W. 2012. Aiming for the bull’s eye: Preparing for throwing investigated with event-related brain potentials. Psychophysiology 49: 335-344.
64) Schmidt RA, White JL. 1972. Evidence for an error detec-
sensorimotor association and expectancy in the human brain. Nature 203: 380-384.
27) Doyon J, Benali H. 2005. Reorganization and plasticity in the adult brain during learning of motor skills. Curr Opin Neu-robiol 15: 161-167.
28) Willingham DB. 1999. The Neural Basis of Motor-Skill Learning. Curr Dir Psychol Sci 8: 178-182.
29) Lalazar H, Vaadia E. 2008. Neural basis of sensorimotor learning: modifying internal models. Curr Opin Neurobiol 18: 573-581.
30) Mitz AR, Godschalk M, Wise SP. 1991. Learning-dependent neuronal activity in the premotor cortex: activity during the acquisition of conditional motor associations. J Neurosci 11: 1855-1872.
31) Karni A, Meyer G, Jezzard P, Adams MM, Turner R, Un-gerleider LG. 1995. Functional MRI evidence for adult mo-tor cortex plasticity during motor skill learning. Nature 377: 155-158.
32) Grafton ST, Mazziotta JC, Presty S, Friston KJ, Frackowiak RS, Phelps ME. 1992. Functional anatomy of human proce-dural learning determined with regional cerebral blood flow and PET. J Neurosci 12: 2542-2548.
33) Grafton ST, Roger MD, Woods P, Tyszka M. 1994. Function-al imaging of procedural motor learning: Relating cerebral blood flow with individual subject performance. Hum Brain Mapp 1: 221-234.
34) Fitts PM, Posner MI. 1967. Human Performance. Belmont, CA: Brooks/Cole.
35) Doyon J, Penhune V, Ungerleider LG. 2003. Distinct contri-bution of the cortico-striatal and cortico-cerebellar systems to motor skill learning. Neuropsychologia 41: 252-262.
36) Eysenck HJ, Thompson E. 1966. The effect of drive on per-formance and reminiscence in a complex tracing task. Br J Psychol 57: 99-106.
37) Kimble GA. 1949. An experimental test of a two-factor theo-ry of inhibition. J Exp Psychol 39: 15-23.
38) Imamizu H, Miyauchi S, Tamada T, Sasaki Y, Takino R, Pütz B, Yoshioka T, Kawato M. 2000. Human cerebellar activity reflecting an acquired internal model of a new tool. Nature 403: 192-195.
39) Kawato M, Gomi H. 1992. A computational model of four regions of the cerebellum based on feedback-error learning. Biol Cybern 68: 95-103.
40) Taylor MJ. 1978. Bereitschaftspontential during the acquisi-tion of a skilled motor task. Electroencephalogr Clin Neuro-physiol 45: 568-576.
41) Takasawa N. 1989. The effect of mental set on readiness po-tential. Jpn J Physiol Psychophysiol 7: 1-10 (in Japanese).
42) Cross ES, Schmitt PJ, Grafton ST. 2007. Neural substrates of contextual interference during motor learning support a model of active preparation. J Cogn Neurosci 19: 1854-1871.
43) Masaki H, Takasawa N, Yamazaki K. 1997. The effect of pa-rameter modification process during execution of a trigger pulling movement on readiness potential. Jpn J EEG EMG 25: 18-24 (in Japanese).
44) Masaki H, Takasawa N, Yamazaki K. 1998. The effect of mo-tor programming and the parameter-applying-processes on the readiness potential. Jpn J Physiol Psychophysiol 16: 85-91 (in Japanese).
45) Toma K, Matsuoka T, Immisch I, Mima T, Waldvogel D, Ko-shy B, Hanakawa T, Shill H, Hallett M. 2002. Generators of

380 JPFSM : Masaki H and Sommer W
tion mechanism in motor skills: A test of Adams’ closed-loop theory. J Mot Behav 4: 143-153.
65) Falkenstein M, Hohnsbein J, Hoormann J, Blanke L. 1991. Effects of crossmodal divided attention on late ERP compo-nents. II. Error processing in choice reaction tasks. Electroen-cephalogr Clin Neurophysiol 78: 447-455.
66) Gehring WJ, Goss B, Coles MGH, Meyer DE, Donchin E. 1993. A neural system for error detection and compensation. Psychol Sci 4: 385-390.
67) Ullsperger M, von Cramon D. 2003. Error monitoring using external feedback: specific roles of the habenular complex, the reward system, and the cingulate motor area revealed by functional magnetic resonance Imaging. J Neurosci 23: 4308-4314.
68) Carter CS, Braver TS, Barch DM, Botvinick MM, Noll D, Cohen JD. 1998. Anterior Cingulate Cortex, Error Detection, and the Online Monitoring of Performance. Science 280: 747-749.
69) Botvinick MM, Braver TS, Barch DM, Carter CS, Cohen JD. 2001. Conflict monitoring and cognitive control. Psychol Rev 108: 624-652.
70) Burle B, Roger C, Allain S, Vidal F, Hasbroucq T. 2008. Er-ror negativity does not reflect conflict: A reappraisal of con-flict monitoring and anterior cingulate cortex activity. J Cogn Neurosci 20: 1637-1655.
71) Carbonnell L, Falkenstein M. 2006. Does the error negativ-ity reflect the degree of response conflict? Brain Res 1095: 124-130.
72) Masaki H, Falkenstein M, Stürmer B, Pinkpank T, Sommer W. 2007. Does the error negativity reflect response conflict strength? Evidence from a Simon task. Psychophysiology 44:
579-585.73) Masaki H, Segalowitz S. 2004. Error negativity: A test of
the response conflict versus error detection hypotheses. In: Errors, conflicts, and the brain Current opinions on perfor-mance monitoring (M Ullsperger & M Falkenstein eds), pp 76-83, Leipzig: MPI of Cognitive Neuroscience.
74) Anguera JA, Seidler RD, Gehring WJ. 2009. Changes in Per-formance Monitoring During Sensorimotor Adaptation. J Neurophysiol 102: 1868-1879.
75) Miltner WHR, Braun CH, Coles MGH. 1997. Event-related brain potentials following incorrect feedback in a time-esti-mation task: Evidence for a “generic” neural system for error detection. J Cogn Neurosci 9: 788-798.
76) Holroyd CB, Coles MGH. 2002. The neural basis of human error processing: Reinforcement learning, dopamine, and the error-related negativity. Psychol Rev 109: 679-709.
77) Takasawa N, Takino R, Yamazaki K. 1990. Event-related po-tentials derived from feedback tones during motor learning. Jpn J Physiol Psychophysiol 8: 95-101 (in Japanese).
78) Slobounov SM, Ray WJ, Simon RF. 1998. Movement-related potentials accompanying unilateral finger movements with special reference to rate of force development. Psychophysi-ology 35: 537-548.
79) Slobounov S, Hallett M, Newell KM. 2004. Perceived effort in force production as reflected in motor-related cortical po-tentials. Clin Neurophysiol 115: 2391-2402.
80) Masaki H, Sommer W, Takasawa N, Yamazaki K. 2012. Neu-ral mechanisms of timing control in a coincident timing task. Exp Brain Res 218: 215-226.
81) Ford JM. 1999. Schizophrenia: The broken P300 and beyond. Psychophysiology 36: 667-682.