closely related parasitoids induce different pupation and foraging responses in drosophila larvae
TRANSCRIPT

Closely related parasitoids induce different pupation and foragingresponses in Drosophila larvae
Majeed Askari Seyahooei, Femmie J. L. Kraaijeveld-Smit, Ken Kraaijeveld,Jessica B. M. Crooijmans, Tom J. M. Van Dooren and Jacques J. M. van Alphen
M. Askari Seyahooei ([email protected]), F. J. L. Kraaijeveld-Smit, K. Kraaijeveld, J. B. M. Crooijmans, T. J. M. VanDooren and J. J. M. van Alphen, Inst. of Biology, Leiden Univ., PO Box 9516, NL�2300 RA Leiden, the Netherlands.
Few examples exist where parasites manipulate host behaviour not to increase their transmission rate, but their ownsurvival. Here we investigate fitness effects of parasitism by Asobara species in relation to the pupation behaviour of thehost, Drosophila melanogaster. We found that Asobara citri parasitized larvae pupate higher in rearing jars compared tounparasitized controls, while A. tabida pupated on or near the medium. No change in pupation site was found for threeother species. A follow-up experiment showed a non-random distribution of parasitized and unparasitized pupae over thedifferent jar parts. To test the adaptiveness of these findings, we performed pupal transfer experiments. Optimumpupation sites were found to be different between host individuals; wall individuals survived better than bottomindividuals, but bottom individuals did worse at the wall. Two parasitoid species that alter pupation site significantlyshowed high rates of diapause at their ‘preferred’ pupation site. For one of them, A. citri, pupation occurred at theoptimal site for highest survival (emergence plus diapause). From literature we know that pupation height and foragingactivity are genetically positively linked. Therefore, we implement a short assay for rover/sitter behavioural expression bymeasuring distance travelled during foraging after parasitism. For one out of three species, foraging activity was reduced,suggesting that this species suppresses gene expression in the for pathway and thereby reduces pupation height. Theparasitoid species used here, naturally inhabit widely different environments and our results are partly consistent with arole for ecology in shaping the direction of parasite-induced changes to host pupation behaviour. More parasitoids arefound on the wall of the rearing jar when they originate from dry climates, while parasitoids from wet climates pupate onthe humid bottom.
Parasite infection often induces changes in host behaviour(Combes 1991, 1998, Poluin 1998, Moore 2002). Thesechanges can result from resistance adaptations of the host,pathological side-effects of the infection or from the parasitemanipulating host behaviour in order to enhance its owntransmission rate. The latter is known as the ‘extendedphenotype’ (Dawkins 1982). Manipulation of host beha-viour by a parasite will be selected for when the normal hostbehaviour is suboptimal for parasite transmission. Manyexamples of such manipulation exist. Snails change habitatpreference when infected with a trematode parasite (Miuraet al. 2006). Birds infected with a vector-borne parasite havedecreased mobility, which increases predation risk andtherefore stimulates parasite dispersal (Holmstad et al.2006). Leptopilina boulardi, a hymenopteran parasitoid,has an increased tendency to lay eggs in already parasitizedhosts when infected with a virus, which promotes horizontaltransmission of the virus (Varaldi et al. 2003). Clearly,parasite-induced changes in host behaviour can haveprofound impacts on parasite fitness.
In addition to increasing their own transmission rate,parasites may also manipulate the behaviour of their host to
enhance their own survival directly. Some examples exist,especially in relation to host and parasitoids (parasites thatalways kill their host), a common threat to insects. They laytheir egg on (ecto) or inside (endo) the body of preimaginal(egg, larva or pupa) or adult stages of the host and completetheir development on or inside the host’s puparium or body(Godfray 1994, Quicke 1997). An example of hostbehavioural changes comes from the famous caterpillarsinfected with the ecto-parasitoid Cotesia glomerata whichdefends their parasitoid against natural enemies by behavingaggressively towards them and also via spinning a webaround the pupae (Brodeur and Vet 1994, Tanaka andOhsaki 2006). Similarly, the braconid parasitoid, Glypta-panteles sp. induces a behaviour in its host, Thyrinteinaleucocera to guard parasitoid pupae and this results inincreased parasitoid survival (Grosman et al. 2008).
In insects, one trait that has large consequences forsurvival, is pupation site selection behaviour. During theimmobile stage, insects are vulnerable to many dangers,including desiccation, fungal infection, predation/parasit-ism and super or hyperparasitism. Insects are thus understrong selection to optimize the timing and location of
Oikos 118: 1148�1157, 2009
doi: 10.1111/j.1600-0706.2009.17508.x,
# 2009 The Authors. Journal compilation # 2009 Oikos
Subject Editor: Tim Benton. Accepted 17 February 2009
1148

pupation in a way that minimizes such risks. Pupation siteselection occurs at the late larval stage and is affected bymoisture, temperature, density, phenotype/genotype, sex,species and the presence of predators or (super and/orhyper) parasitoids (Sameoto and Miller 1968, Barker 1971,Sokolowski and Hansell 1983, Sokolowski et al. 1986,Schnebel and Grossfield 1992, Pivnick 1993, Tanaka andOhsaki 2006, Riedl et al. 2007).
Parasitoids and hosts may often differ in optimalpupation strategy. For example, when the parasitoid spendsmore time in the puparium than the host (which they oftendo) it will be at risk of desiccation, predation andhyperparasitism by pupal parasitoids for longer and mayprefer a more humid and concealed pupation site. Further-more, differences in body size between parasitoid and hostresults in differences in surface/body mass ratio, which mayalso influence the optimal pupation site (Carton et al. 1986,Ellers 1998). Because it is the host that pupates, theparasitoid has to manipulate the host if it is to achieve itsown optimal pupation strategy. In spite of the considerableinterest in parasite-induced changes in host behaviour, littleis known of the effects of parasitism on pupation siteselection behaviour. One example that we do know of isthat of the Bertha armyworms, Mamestra configurate whichpupate in the leaf litter around the host plant whenparasitized by Microplitis mediator. Those not parasitizedall pupate on the host plant (Pivnick 1993). Although theseobservations are made, little experimental evidence ispresent to show that these behavioural changes actuallyincrease parasitoid survival.
Here we investigate whether the pupation behaviour ofDrosophila changes as a result of attack by Asobaraparasitoid wasps. Species in the genus Asobara cover awide range of ecological niches, including wet tropicalconditions, dry deserts and cool temperate conditions. Theyfurther differ markedly in generation time and hence in thetime spent in the host puparium. Thus, the different speciesof Asobara are expected to differ in the degree to which theyare in conflict with their host over pupation behaviour. Thisprovides an ideal system to test if parasitoids alter pupationbehaviour of their host in an adaptive manner.
We use five species of Asobara that span a range ofenvironments and that are all cultured on Drosophilamelanogaster. Two species are naturally adapted to dryclimates, while the other three originate from wet andhumid climatic conditions (Table 1). We hypothesize thatthe former are better adapted to lower humidity conditionsthan the latter. The conditions in our rearing jars show asteep gradient in humidity. Close to the medium at thebottom of the jar, conditions are much more moist than onthe glass higher up. Thus, if our expectation is correct and if
parasitoids are able to manipulate their host’s pupationbehaviour, we expect species from the dry areas to pupatehigher in the jars compared to larvae parasitized by waspsfrom wet areas. Therefore, we compare the pupationbehaviour of D. melanogaster larvae parasitized by differentspecies of Asobara to that of unparasitized larvae. Findingsuch differences, we then ask whether these might beadaptive. In order to test this, we performed a series oftranslocation experiments in which hosts pupae weretransferred from their selected (high or low in the jar) tothe unselected site. In the first experiment, we useunparasitized hosts only, to examine the adaptive signifi-cance of pupation site selection in individual larvae. This isimportant, since although we know that this behaviouraltrait has large consequences for survival (Sokolowski et al.1986, Rodriguez-del-Bosque and Smith 1996) there is noexperimental evidence to show that individually selectedsites are the sites which yield optimal survival. In the secondtranslocation experiment, we used parasitized hosts to assessthe adaptive value of parasite-induced alterations in pupa-tion behaviour. For parasitoid translocations we not onlyinvestigate the effect of translocation on emergence, but alsoon diapause and total survival (emergence�diapause).These translocations enable us to examine three possiblesources of variation which may address survival variance:
1) different effect of pupation sites0transferred fromwall to wall (WW)"bottom to bottom (BB).
2) variation of individuals which is caused by differencesbetween individuals in optimal pupation site0WW�WB and/or BB�BW.
3) pre-pupal stage determines the fitness0BB�BWand/or WW�WB.
Last, we try to touch upon the genetic mechanismunderlying parasitoid-induced changes in host pupationbehaviour. Foraging behaviour is regulated through theforaging pathway, which is largely controlled by the forgene. Alleles at the for gene induce two behaviouralphenotypes, known as ‘rovers’ and ‘sitters’. Rovers movemore during foraging and pupate higher compared to sitters(Graf and Sokolowski 1989, Sokolowski 1980). Thepleiotropic effects of foraging on pupation behavioursuggest that pupation behaviour is at least partly regulatedthrough the for pathway (Sokolowski and Hansell 1983).Thus parasitoids could achieve changes in pupation site byinfluencing the expression level of the for gene which resultsin rover/sitter phenotypic behaviour. As the first step to testthis hypothesis we did a short assay for rover/sitterphenotype behaviour by observing foraging behaviour ofparasitized versus unparasitized larvae.
Table 1. Origin, collection and rearing summary of the Asobara species used in this study. All of the parasitoid species have been reared onD. melanogaster since collection in our lab at 20918C and 16:8 h L:D.
Parasitoid Origin Climate at origin Collected
A. citri Africa, sub-Saharan Africa dry and hot Lamto, Cote, d’Ivoire 1995A. persimilis Australia Mediterranean hot and dry summer Sydney 1997A. tabida Holarctic, Europe and north America temperate and wet with cold winter Leiden 2006A. pleuralis southeast Asia, oriental tropic tropical wet forest Manado, Sulawesi Indonesia 2005A. japonica Japan Tokyo temperate and wet Tokyo 1995
1149

Material and methods
Strain of parasitoids and hosts
Parasitoids and hostWe used the five species of Asobara described in Table 1.The host used in all experiments was Drosophila melanoga-ster from a laboratory stock that has been maintained in ourlab since 1966 at 20918C and 16:8 h day:night regime.This stock is polymorphic for the rover and sitter alleles ofthe for gene.
Pupation height
To test whether different species of Asobara have differenteffects on host pupation behaviour, we set up an experimentusing the five species of Drosophila parasitoids (Table 1) anda control group (non-parasitized hosts). Each group con-sisted of 10�12 replicates. The experiment was performed insmall glass tubes (2.2�8.0 cm) containing a layer of agarmedium (3 ml vial�1) and a thin layer of yeast suspension(25 g dry yeast per 100 ml H2O) as food source (0.3 mlvial�1). To each tube we added 20 second instar host larvaeand two mated female wasps. The control vials were leftwithout parasitoids. Tubes were placed in a climate room at258C, 65% relative humidity and a 16:8 h light:darkregime. These condition resulted in very low parasitationrates for A. tabida and we repeated the experiment for thisspecies at 208C to provide optimum conditions for A. tabidaparasitism rate. Furthermore, vials were filled with foodmedium (recipe: /<http://www.utm.utoronto.ca/�w3forage//>), instead of agar medium. Wasps were removedfrom the treatment vials after 2 h. The vials were then left inthe climate room for five days, by which time all hosts hadpupated. We measured pupation height as the distancebetween the surface of the medium and the pupa (Soko-lowski and Hansell 1983). We also recorded whether thepupa was parasitized or not. Those that were not parasitizedwere discarded from the analyses.
Pupal translocation experiment
Translocation experiments were performed to test whetherthe choice of pupation site affected fitness of D. melano-gaster. Forty jars (3.5�8.5 cm) containing agar mediumand one ml yeast suspension (32 g yeast per 100 ml water)were prepared. To each we added � 100 2nd instar hostlarvae. When the larvae had pupated, 40 pupae werecollected from each jar: 20 from the medium on thebottom of the jar and 20 from the side of the jar. Thesepupae were then placed in new jars (10 pupae per jar)containing standard agar medium. Ten pupae that camefrom the side were placed back onto the side and ten pupaethat came from the side were placed on the medium. Tenpupae from the medium were placed onto the side and tenback on the medium. After emergence, the number of fliesper jar was scored. Per treatment group ten jars wereprepared and the experiment was performed twice. The jarswere kept at 208C.
A similar translocation experiment was performed forparasitized larvae. This time, 3 female parasitoids were
added to the jars containing � 100 2nd instar host larvae.All five parasitoid species were used. Just after pupation, perspecies, twenty jars were prepared for translocation side(bottom�bottom, bottom�wall, wall�wall, wall�bottom).Jars were placed at 258C, apart from the A. tabidaparasitized group, which was kept at 208C. After 4�5 weekswe scored the status for each translocated pupae: parasitizedor unparasitized, and emerged, dead or in diapauze.
Foraging path length
The pupation height experiment revealed significant differ-ences in pupation height of larvae parasitized by two out ofthe five species (A. tabida and A. citri; Results). To testwhether these differences could be caused by effects ofparasitism on expression of the for gene, as the first step tocheck the expression level of this gene we tested fordifferences in foraging path length by a short assay forrover/sitter behaviour, which is regulated by the samegenetic pathway. As a control, we also measured pathlengths of A. pleuralis parasitized larvae, where a change inpupation site was not significant in the pupation heightexperiment. The maximum individual variation in foragingbehaviour of D. melanogaster larvae occurs at the mid-thirdinstar larval stage (Sokolowski 1980, Sokolowski andHansell 1983). At this stage the larvae leave a clear trailon yeast which can be measured. This path length isroutinely used as a measure of foraging intensity (Soko-lowski 1980, Sokolowski and Hansell 1983). We measuredforaging path lengths of parasitized larvae and comparedthem to those of unparasitized control larvae. In eachexperiment 30 mid-2nd instar larvae were placed in a jar(3.5�8.5 cm) with a layer of agar and 0.3 ml of yeastsolution. To the groups in the parasitized treatment, weadded two mated female parasitoids for two hours. Five jarsper treatment (control vs parasitized) per parasitoid specieswere prepared for each experiment. Treatment and controljars of A. citri and A. pleuralis were kept in a climate roomat 258C, while A. tabida and its control jars were reared at208C. The immune reaction of parasitized larvae results inslightly slower growth rates and thus longer developmenttime compared to control larvae (Wertheim et al. 2005).Therefore we designed a time table in such a way that thecontrol jars for each treatment were started one day afterthose of the treatment larvae. This created mid-third instarlarvae of both parasitized and control groups on the sameday.
To measure foraging path length we used plastic petridishes (Ø 8.5 cm and 1.4 cm high) containing a layer ofagar on which we spread 1.5 ml of yeast solution (8.5 g per25 ml) (Sokolowski 1980). Path lengths of six larvae per jarwere measured individually by placing them in the centre ofthe petri dish and allowing them to forage for five minutes(Sokolowski 1980). The trail track was transferred onto atransparent sheet and its length was measured at a later time.For the parasitized group, larvae were dissected to determinewhether they were parasitized or not or had encapsulated theparasitoid larvae. Encapsulation by aggregation of haemo-cytes around the parasitoid eggs is a common strategy ininsects to escape parasitism (Nappi et al. 1975, Rizki andRizki 1984). Host larvae in the parasitized group that
1150

contained no wasp larvae or contained an encapsulatedparasitoid egg were excluded from the analyses.
Statistical analyses
Standard linear parametric models were unsuitable for theanalysis of pupation height, as this measure is boundedbelow (the medium) as well as above (the bottom of thestopper sealing the glass vial). We applied a Weibullparametric survival regression analysis in the free statisticalsoftware R 2.5.1 to our data (Ihaka and Gentleman 1996,Therneau and Grambsch 2000). Since data distributionswere similar in shape to Weiball. We included the randomjar effect as frailty in this model.
The results of the translocation experiments were analysedusing generalized linear models (GLM) with a quasibinomialdistribution in R. For host we estimated probabilities ofemergence. We then determined whether translocation sidecontributed to variation in the number of emerged flies bycomparing the full model to one from which translocationhad been dropped using a x2-test. We record the x2-valueand its associated p-value. Post-hoc t-tests from this modelwhere used to indicate which of the four translocation sides(BB, BW, WW, WB) differ from each other in emergencerate. The statistical analyses concerning translocation experi-ments involving parasitized pupae was similar to that of thehost experiment. However, three additional tests were addedper parasitoid species. In addition to emergence rate, weanalysed the number of parasites in diapause, the proportionof pupae surviving (emerged plus diapause) and the numberof flies versus parasitoids picked from either the bottom orthe wall at the start of the translocation experiment. Thelatter analyses was thus an independent repeat of thepupation height experiment.
Foraging pathlengths were analysed using generalizedlinear mixed effect models (LMEM, Pinheiro and Bates2000), in the R-package nlme. Treatment (parasitized orcontrol) was entered as a fixed effect while jar was entered asa random effect. The experiments for the three parasitoidspecies were not performed at the same time, and thereforeanalysed separately. In all three cases we searched for theminimum adequate model (Crawley 2005) by starting withthe maximal model and then dropping the main factor(treatment) if non-significant. Comparing the simpler tothe more complex model was done using a likelihood-ratiotest. We recorded these likelihood-ratios and their asso-ciated p-value.
Results
Pupation height
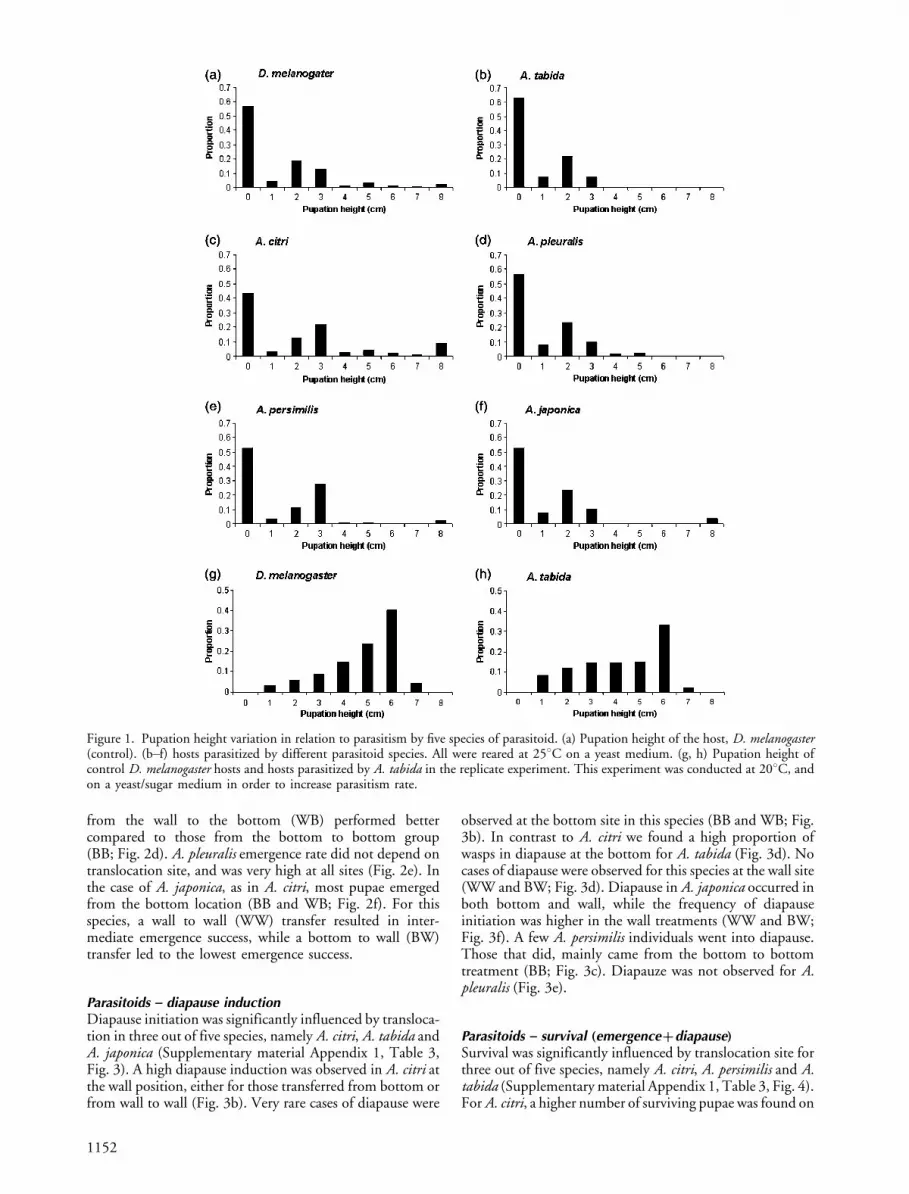
First we compared pupation heights of D. melanogasterlarvae parasitized by five different Asobara species and thoseof control larvae. Pupation height of host larvae differedsignificantly after parasitism by different species (likelihoodratio test, x2�5.16, DF�1, p�0.005) (Fig. 1a�f). Apost hoc z-test revealed that larvae parasitized by A. citripupated significantly higher than control larvae (z�2.52,p�0.01) and larvae parasitized by A. tabida had a non-significant tendency to pupate lower than control larvae
(z��1.67, p�0.09) Parameter estimates from this fullmodel also showed that the direction of pupation heightdifferent between parasitoid species. We observed somesimilarities in distribution pattern of pupation site. A. citriand A. persimilis showed a similar pattern, which differedfrom A. tabida, A. pleuralis and A. japonica. (Fig. 1a�f). Inthe replicate experiment of A. tabida, performed at a lowertemperature, parasitism induced a highly significant reduc-tion in pupation height (Fig. 1g�h, likelihood ratio test,x2�7.34, DF�1, p�0.006).
Pupal translocation experiments
Host � emergence (survival) successEmergence success was significantly influenced by translo-cation site in the case of unparasitized host pupae (Fig. 2aand 4a which are identical, DF�3, x2�69.6, p B0.001).Post-hoc t-tests (see complete overview of t- and p-valuesfor all species in Supplementary material Appendix 1)revealed that wall to wall and wall to bottom transfersresulted in equally high emergence success, which washigher than bottom to bottom or bottom to wall transfers.Bottom to wall transfers resulted in even lower emergencerates compared to its control, the bottom to bottomtransfers (Fig. 2a, 4a).
Parasitoids � number of flies versus waspsIn our pupation height experiment we found a significantincrease in pupation height for A. citri parasitized larvae anda reduction in A. tabida relative to control larvae.Furthermore, a similar distribution pattern to A. citri wasobserved in A. persimilis while distribution patterns ofA. pleuralis and A. japonica more resembled those ofA. tabida. Perhaps the effects of parasitoids on hostpupation behaviour are small, but consistently presentacross experiments. If this is the case, we would find adifferent number of flies versus parasitoids between the walland the bottom when picking pupae for the translocationexperiment. For A. citri and A. persimilis we expect to findmore parasitoids versus flies on the wall compared to thebottom, and vice versa for A. tabida, pleuralis and japonica.Table 2 shows that these expectations are met, except for A.citri, where the number of flies was indeed lower on the wallcompared to the bottom, but this difference was notsignificant. Perhaps this is due to the fact that in that caseparasitisation success was very high which results in very lownumbers of flies (Table 2).
Parasitoids � emergence successEmergence success was significantly influenced by translo-cation site for all but one species, namely A. pleuralis(Supplementary material Appendix 1, Table 3, Fig. 2). Theoptimal site to pupate at in terms of emergence successdiffered between parasitoid species. For A. citri most pupaeemerged from the bottom location, regardless of whetherthis was the original site of pupation (BB) or not (WB, Fig.2b). For A. persimilis the highest emergence rate wasobtained if bottom pupae were translocated to the wall(BW, Fig. 2c). A. tabida individuals emerged at the highestrate either when relocated back to the wall (WW) ortransferred from bottom to wall (BW). Those transferred
1151
from the wall to the bottom (WB) performed bettercompared to those from the bottom to bottom group(BB; Fig. 2d). A. pleuralis emergence rate did not depend ontranslocation site, and was very high at all sites (Fig. 2e). Inthe case of A. japonica, as in A. citri, most pupae emergedfrom the bottom location (BB and WB; Fig. 2f). For thisspecies, a wall to wall (WW) transfer resulted in inter-mediate emergence success, while a bottom to wall (BW)transfer led to the lowest emergence success.
Parasitoids � diapause inductionDiapause initiation was significantly influenced by transloca-tion in three out of five species, namely A. citri, A. tabida andA. japonica (Supplementary material Appendix 1, Table 3,Fig. 3). A high diapause induction was observed in A. citri atthe wall position, either for those transferred from bottom orfrom wall to wall (Fig. 3b). Very rare cases of diapause were
observed at the bottom site in this species (BB and WB; Fig.3b). In contrast to A. citri we found a high proportion ofwasps in diapause at the bottom for A. tabida (Fig. 3d). Nocases of diapause were observed for this species at the wall site(WW and BW; Fig. 3d). Diapause in A. japonica occurred inboth bottom and wall, while the frequency of diapauseinitiation was higher in the wall treatments (WW and BW;Fig. 3f). A few A. persimilis individuals went into diapause.Those that did, mainly came from the bottom to bottomtreatment (BB; Fig. 3c). Diapauze was not observed for A.pleuralis (Fig. 3e).
Parasitoids � survival (emergence�diapause)Survival was significantly influenced by translocation site forthree out of five species, namely A. citri, A. persimilis and A.tabida (Supplementary material Appendix 1, Table 3, Fig. 4).For A. citri, a higher number of surviving pupae was found on
Figure 1. Pupation height variation in relation to parasitism by five species of parasitoid. (a) Pupation height of the host, D. melanogaster(control). (b�f) hosts parasitized by different parasitoid species. All were reared at 258C on a yeast medium. (g, h) Pupation height ofcontrol D. melanogaster hosts and hosts parasitized by A. tabida in the replicate experiment. This experiment was conducted at 208C, andon a yeast/sugar medium in order to increase parasitism rate.
1152

the wall compared to the bottom (Fig. 4b). This is in contrastto the number emerging, which was highest at the bottom.This difference can be attributed to the fact that a largeproportion of the pupae that were transferred from eitherwall or bottom to wall went into diapause (Fig. 3b). Nosignificant differences in survival were observed forA. persimilis (Fig 4c). In contrast to A. citri, A. tabidaindividuals mostly went into diapause at the bottom (BB andWB). As a result, survival was similar for wall to wall, wall tobottom and bottom to wall transfers. Bottom to bottomtransfers still had significantly lower survival rates (Fig. 4d).For A. pleuralis emergence rate was high at all four sites, andno diapause events were observed, thus survival rate does not
differ from emergence rate (Fig. 2e and 4e are identical).Diapause in A. japonica mostly occurred for the walltreatments (WW and BW), where emergence was relativelylow compared to the bottom (Fig. 2f). This led to a very highsurvival rate for all four translocation treatments whicheliminated the transfer effects seen in emergence rate(Fig. 4f).
Foraging path length
In the short assay for rover/sitter phenotypic behaviour thedistance travelled while foraging did not differ betweenparasitized larvae and unparasitized larvae for A. citri and A.pleuralis (Fig. 5a�b: A. citri, likelihood-ratio test; x2�0.09,p�0.76; A. pleuralis, x2�0.04, p�0.84). Larvae para-sitized by A. tabida travelled less while foraging comparedto unparasitized larvae (Fig. 5c: x2�23.55, p B0.001).
Discussion
The pupation height experiment showed that attack by fivespecies of Asobara affected pupation height of D. melano-gaster larvae in different directions. Pupation heightchanged significantly after attack by two out of five speciesof Asobara. For one species, A. tabida, first only a trend wasobserved. Due to low parasitism rate at high temperatures,we were not able to firmly state that parasitism changespupation behaviour for this species. Therefore, for A. tabidawe repeated the experiment at 208C resulting in asignificant effect of parasitism on pupation height for thisspecies too. Although at 258C the effect of A. tabida wasnot significant, the direction of induced change remainedconsistent with the change observed at 208C (Fig. 1b, 1h).A. tabida significantly reduced pupation height whileA. citri increased pupation height. Further evidence forpupal site changes in response to parasitoid attack wasfound by collecting pupae from the wall and bottom at thestart of the translocation experiment. Relatively moreparasitoids compared to host pupae were found at thewall for A. persimilis, while for A. tabida, A. pleuralis andA. japonica, more parasitoids were found at the bottom.Pupal translocation experiments of the host showed thatoptimum pupation site differs between individuals. Overall,individuals pupating on the wall survived better than thosepupating on the bottom. However, bottom individuals didworse when translocated to the wall. Similar translocationexperiments were performed for parasitized larvae toinvestigate whether the change in pupation behaviour isadaptive for the parasitoid under our laboratory conditions.The results of the translocation experiments are summarizedin Table 4. Our results showed that diapause was induced atthe preferred site for both A. citri and A. tabida, the speciesthat showed significant change in pupation height. In termsof survival, the result obtained for A. citri was consistentwith an adaptive explanation (Table 4). Hosts attacked byA. citri tended to pupate on the wall and transferring pupaeto that location increased their survival rate. In contrast toour expectation, survival of A. tabida decreased at theselected pupation site. The analysis of foraging path lengthsshowed a significant reduction in foraging activity of larvae
Figure 2. The effect of translocation on the emergence rate for thehost and the five parasitoid species. BB�transferred from bottomto bottom, BW�transferred from bottom to wall, WB�transferred from wall to bottom and WW�transferred from wallto wall. Square symbols represent the median of all groups. Thebox represents 50% of all data points for that treatment(interquartile range). Error bars represent the minimum andmaximum observed pupation height, unless outliers are present.When outliers are present the error bar represents 1.5� theinterquartile range. To indicate the results of post-hoc t-tests toindicate which treatments differ within species we labelled eachbox with a letter. Boxes with identical letters do not differsignificantly from each other.
Table 2. The average number of fly pupae9SE collected from thewall or bottom at the start of the pupal translocation experiment(n�80 pupae per species). The p- and x2- values are the results ofGLM models that test whether the ratio of fly to wasp pupae differbetween wall and bottom.
Average no. of flies (out of 10) p-value (x2)
Wall Bottom
A. citri 0.2890.09 0.4390.13 0.41 (0.8)A. persimilis 0.2390.08 0.8390.15 B0.001 (15.3)A. tabida 7.0390.34 3.4590.29 B0.001 (104.8)A. pleuralis 1.3090.19 0.8090.15 0.04 (5.4)A. japonica 1.2890.30 0.5090.14 0.012 (15.3)
1153

parasitized by A. tabida. As both foraging path length andpupation height are to are large extent regulated by the forgene, this indicates that A. tabida may reduce the expressionof this gene.
Pupation site in unparasitized Drosophila larvae is knownto depend on moisture, temperature, density, phenotype/genotype (rover vs sitter) and species (Sameoto and Miller1968, Barker 1971, Sokolowski et al. 1986, Schnebel andGrossfield 1992). The decision of where to pupate seems tobe taken during the wandering stage which occurs a fewhours before pupation (Riedl et al. 2007). During this stage,larvae move away from the food source and return to thefood source many times, as if testing what the conditionsare like, before they choose a site to pupate. Although weknow that pupation site varies with both abiotic and bioticfactors, experimental evidence to show that pupal siteselection increases survival of the fruit fly is limited.Sokolowksi et al. (1986) showed that adult emergence inD. melanogaster of larvae that pupated on fruit decreased assoil water content increased. In contrast, those pupated onthe soil had lower adult emergence when soil water content
decreased. We observed an overall better survival forunparasitized hosts pupated on the wall. Perhaps this canbe explained by fast growing, fit, larvae being able to reachthe wall, filling up this space, while the weak, slow growinglarvae are forced (or not able to climb) to pupate at thebottom. Inconsistence with this explanation is the fact thatsurvival of sitter phenotypes is reduced when moved to arover phenotype pupation site. Thus our experiment partlysupports the hypothesis that one phenotype (rover) is fitterover another phenotype (sitter). However, the fact thatoptimal pupation sites differ between individuals within thesame population, also indicates a role for individualpupation site selection.
Finding a free enemy site for pupation could for both thehost, as well as the parasitoid, be a reason for pupation siteselection. Ohsaki and Sato (1994) showed that an oligo-phagous butterfly, Pieris rapae, chose a suboptimal food-resource host-plant for egg laying, which was less attractivefor parasitoids. This increased its survival but came at thecost of losing strengths in other traits. Furthermore,parasitoids face the treat of being parasitized by hyperpar-
Table 3. The effect of pupal translocation on emergence, survival and diapause rates.
Emergence success Survival success Diapause induction
x2 p-value x2 p-value x2 p-value
A. citri 81.5 B0.001 30.4 B0.001 259.0 B0.001A. persimilis 16.5 0.001 17.3 0.001 4.67 n.sA. tabida 74.4 B0.001 26.4 B0.001 53.2 B0.001A. pleuralis 2.7 n.s. 2.7 n.s. 0 .0 n.sA. japonica 40.3 B0.001 4.2 n.s. 49.5 B0.001
Figure 3. The effect of translocation on diapause rate for the hostin the five parasitoid species. BB�transferred from bottom tobottom, BW�transferred from bottom to wall, WB�transferredfrom wall to bottom and WW�transferred from wall to wall. SeeFig. 2 for box-plot details.
Figure 4. The effect of translocation on the survival rate(emergence plus diapause) for the host and the five parasitoidspecies. BB�transferred from bottom to bottom, BW�trans-ferred from bottom to wall, WB�transferred from wall to bottomand WW�transferred from wall to wall. See Fig. 2 for box-plotinformation.
1154

asitoids. To avoid this, they can manipulate their hostbehaviour, for example by leaving the food patch or nest, insearch for an enemy free space, in order to enhance its ownsurvival (Stamp 1981, Fritz 1982, Brodeur and McNeil1992, Grosman et al. 2008, Harvey et al. 2008).
Our results suggested that hosts pupate at higherlocations in the jar (where conditions are drier) whenparasitized by A. citri and A. persimilis. Both these speciesinhabit arid environments (Table 1), suggesting that theirsurvival is highest in dry conditions. Our translocationexperiments support this conclusion for A. citri, but foundno significant effect in A. persimilis. The three remainingspecies (A. japonica, A. pleuralis and A. tabida) are all foundin relatively humid environments. Hosts parasitized bythese species pupated on or near the medium in relativelymoist conditions. In the field, the optimal pupation siteshould minimize the risk of desiccation and microbialinfection (Alphen and Thunissen 1982). As mentionedbefore, another likely mechanism is that the behaviourchange may be to reduce hyper or superparasitism rate.Apart perhaps from desiccation, none of these risks werepresent in our experiments. Thus, it could be that our lab
stocks of these parasitoids have retained the ability tochange host pupation behaviour in the absence of the otherselective forces favouring these traits. To address this weneed more field experiments for each species at their site oforigin.
Unexpectedly, we found that pupation site affected theprobability of entering diapause in three out of five speciesof parasitoids. Such conditional diapause has not previouslybeen demonstrated in Drosophila parasitoids. Diapause ininsects can be induced by cold, heat and desiccation stress(Danks 1987, Denliger 2007) and is generally viewed as anadaptation to survive periods of unfavourable conditions(Masaki 2002). In A. japonica, diapause occurs at the wall,and this may be seen as a stress response to unfavourableconditions at this site. In contrast, in A. citri and A. tabida,diapause was most frequent at the site where the hoststended to pupate after parasitism (wall and bottom,respectively). Perhaps natural selection favours high ratesof diapause in these species, for example because successfulreproduction is unlikely after normal development time.Suitable humid conditions may be short-lived in the aridenvironments inhabited by A. citri, while the habitat ofA. tabida is strongly seasonal.
If diapause in our lab stocks is a genetic relic from theirnatural past, it is remarkable that it has been retained. Someof these species have been in culture in our lab for 13 years.During this time we certainly selected against diapausingindividuals. After a maximum of four weeks, when all non-diapausing individuals have emerged, rearing jars arediscarded, along with any diapausing individuals. Thegenetic basis of adult diapause induction is relatively well-understood in Drosophila melanogaster. An allele of the genetimeless known to induce diapause has recently spreadthrough D. melanogaster populations in Europe, wherediapause enhances winter survival (Tauber et al. 2007). Thisexample shows that diapause alleles can become fixed in apopulation rapidly.
A potential mechanism behind the change in hostpupation behaviour in response to parasitoid attack is aparasitoid-induced change in the expression of the for gene.This gene regulates foraging behaviour in the host,D. melanogaster (Sokolowski 1985). Rover phenotypesmove more during foraging and pupate further from thefood source than sitter phenotypes (Sokolowski and Hansell1983, Graf and Sokolowski 1989). A. citri could simply
Figure 5. Foraging path lengths (distance travelled) ofD. melanogaster larvae parasitized by three species of parasitoids.cont�unparasitized control larvae, par�parasitized larvae. SeeFig. 2 for boxplot information.
Table 4. Interpretation of the pupal translocation experiment involving parasitized larvae. The pupation manipulation column represent thedirection of change in response to parasitism as indicated by the pupation height and translocation experiments (for the latter: the ratio of fliesto wasps picked from the wall and bottom at the start of the experiment). In this column 2 represents strong evidence (supported by bothpupation and translocation experiment) and 1 represents weak evidence (support from either pupation height or translocation experimentonly). The consequence of this effect of parasitism for survival and diapause induction is shown as adaptive, maladaptive or neutral (noeffect). For this assessment we assume that being able to enter the diapause stage is a positive trait, enabling parasitoids to survive for a longerperiod. We indicate which of the following prediction are met: 1) there is a site difference, WW�BB or BB�WW, 2) individuals chose itsown optimal site, WW�WB and/or BB�BW, 3) pre-pupal stages determine success, BB�BW and/or WW�WB.
Originalcondition
Effect of parasitism onpupation site
Survival Diapause
Highest Consequence Prediction Highest Consequence Prediction
D. mel wet/cool � bottom and wall � 1, 2 and 3 not present � �A. citri dry /hot to wall (1) wall adaptive 1 wall adaptive 1A. persimilis dry/hot to wall (1) bottom and wall no effect 3 few events no effect �A. tabida wet/cool to bottom (2) wall maladaptive 1, 3 bottom adaptive 1A. pleuralis wet/hot to bottom (1) bottom and wall no effect � not present no effect �A. japonica wet/cool to bottom (1) bottom and wall no effect � wall maladaptive 1
1155

alter their host’s pupation site by stimulating for expression,while A. tabida could achieve pupation close to the foodsource by decreasing for expression. For A. tabida it isknown that alongside the egg, also venom is injected. Thisvenom induces a reduction in behavioural activity of thelarvae for the first 24 h after parasitism (Moreau et al.2002). For A. citri, such venom injection is not known tooccur (Prevost et al. 2005). Thus, perhaps this venomreduces activity in A. tabida right up until the time topupation. To test this hypothesis, we measured path lengthsof foraging D. melanogaster larvae that were parasitized byA. citri, A. tabida or A. pleuralis (as a control). Our resultsrevealed no correlation between pupation and foragingbehaviour by A. citri and A. pleuralis and hence for was notresponsible for the change in pupation behaviour observedin A. citri. Many genes besides for contribute to thedetermination of pupation site (Riedl et al. 2007). Ourresult indicates that the change in pupation behaviour afterattack by A. citri is regulated via genes other than foragingrelated genes. In contrast, we found a significant reductionin foraging path length of larvae parasitized by A. tabidacompared to unparasitized control larvae. Therefore, theresult for A. tabida is consistent with our hypothesis thatparasitoids can alter host behaviour by altering the expres-sion of the for gene. In order to confirm this observation, amore detailed study of host gene expression of the for geneand genes in this pathway in response to parasitism isunderway. Checking the expression level of the othercandidate genes for pupation height in the parasitizedlarvae by A. citri may provide evidence of other host geneexpression manipulation in this species.
In conclusion, we have shown that D. melanogaster larvaealter their pupation behaviour after attack by Asobaraparasitoids. The patterns are species-specific and may relateto the environmental conditions under which these specieslive in nature. While we show that individual pupation-siteselection by D. melanogaster larvae increases survival, wewere not able to show that parasitoid-induced changes wereconsistently adaptive. Last, the mechanistic processes under-lying the change in pupation behaviour appear to differbetween species. While we are currently still a long wayremoved from a coherent explanation for these patterns,their study will certainly enhance our understanding of theway in which parasites influence their host’s behaviours.
Acknowledgements � We would like to thank Prof. M. B.Sokolowski and Dr K. D. Williams from the Univ. of Torontoat Mississauga, Canada, for their support in terms of lab use andvaluable comments related to pupation height experiments.
References
Alphen, J. J. M. van and Thunissen, I. 1982. Host selection andsex allocation by Pachycrepoideus vindemiae Rondani(Pteromalidae) as a facultative hyperparasitoid of Asobaratabida Nees (Braconidae: Alysiinae) and Leptopilina heterotoma(Cynipidae: Eucoilidae). � Neth. J. Zool. 33: 497�514.
Barker, J. S. F. 1971. Ecological differences and competitiveinteraction between Drosophila melanogaster and Drosophilasimulans in small laboratory populations. � Oecologia 8:139�156.
Brodeur, J. and McNeil, J. N. 1992. Host behavior modificationby the endoparasitoid Aphidius nigripes � a strategy to reducehyperparasitism. � Ecol. Entomol. 17: 97�104.
Brodeur, J. and Vet, L. E. M. 1994. Usurpation of host behaviourby a parasitic wasp. � Anim. Behav. 48: 187�192.
Carton, Y. et al. 1986. The Drosophila parasitic wasps. � In:Ashburner, M. et al. (eds), The genetics and biology ofDrosophila. Vol. 3e. Academic Press, pp. 347�394.
Combes, C. 1991. Ethological aspects of parasite transmission.� Am. Nat. 138: 866�880.
Combes, C. 1998. Parasitism: the ecology and evolution ofintimate interactions. � Chicago Univ. Press.
Crawley, M. J. 2005. An introduction using R. � Wiley.Danks, H. V.1987. Insect dormancy: an ecological perspective.
� Biol. Survey of Canada (Terrestrial Arthropods), Ottawa.Dawkins, R. 1982. The extended phenotype: the long reach of the
gene. � Oxford Univ. Press.Denliger, D. L. 2007. Shutting down insect development:
diapause and cold storage. � Entomol. Res. 37: A15�A23.Ellers, J. 1998. life-history evolution in the parasitoid Asobara
tabida: on the tradeoff between reproduction and survival.PhD thesis. � Univ. of Leiden, the Netherlands.
Fritz, R. S. 1982. Selection for host modification by insectparasitoids. � Evolution 36: 283�288.
Godfray, H. C. J. 1994. Parasitoids behavioral and evolutionaryecology. � Princeton Univ. Press.
Graf, S. A. and Sokolowski, M. B. 1989. Rover/sitter Drosophilamelanogaster larval foraging polymorphism as a function oflarval development, food-patch quality, and starvation.� J. Insect Behav. 2: 301�313.
Grosman, A. H. et al. 2008. Parasitoid increases survival of itspupae by inducing hosts to fight predators. � PLoS One 3:e2276.
Harvey, J. A. et al. 2008. Do parasitized caterpillars protect theirparasitoids from hyperparasitoids? A test of the ‘usurpationhypothesis’. � Anim. Behav. 76: 701�708.
Holmstad, P. R. et al. 2006. Vector-borne parasites decrease hostmobility: a field test of freeze or flee behaviour of willowptarmigan. � Int. J. Parasitol. 36: 735�740.
Ihaka, R. and Gentleman, R. 1996. R: a language for data analysisand graphics. � J. Comput. Graph. Statist. 5: 299�314.
Masaki, S. 2002. Ecophysiological consequences of variability indiapause intensity. � Eur. J. Entomol. 99: 143�154.
Miura, O. et al. 2006. Parasites alter host phenotype and maycreate a new ecological niche for snail hosts. � Proc. R. Soc.Lond. B 273: 1323�1328.
Moore, J. 2002. Parasites and the behavior of animals. � OxfordUniv. Press.
Moreau, S. J. M. et al. 2002. Effects of parasitism by Asobaratabida (Hymenoptera: Braconidae) on the development,survival and activity of Drosophila melanogaster larvae.� J. Insect Physiol. 48: 337�347.
Nappi, A. J. et al. 1975. Parasite encapsulation in insects. � In:Maramorosch, K. and Shope, R. E. (eds), Invertebrateimmunity: mechanisms of invertebrate vector�parasite rela-tions. Academic Press, pp. 293�326.
Ohsaki, N. and Sato, Y. 1994. Food plant choice of Pierisbutterflies as a tradeoff between parasitoid avoidance andquality of plants. � Ecology 75: 59�68.
Pinheiro, J. C. and Bates, D. M. 2000. Mixed-effects models in Sand S-PLUS, statistics and computing series. � Springer.
Pivnick, K. A. 1993. Diapause initiation and pupation siteselection of the Braconid parasitoid Microplitis mediator(Haliday) � a case of manipulation of host behaviour.� Can. Entomol. 125: 825�830.
Poluin, R. 1998. Evolutionary ecology of parasites: from indivi-duals to communities. � Chapman and Hall.
1156

Prevost, G. et al. 2005. Asobara, braconid parasitoids of Drosophilalarvae: unusual strategies to avoid encapsulation without VLPs.� J. Insect Physiol. 51: 171�179.
Quicke, D. L. 1997. Parasitic wasps. � Chapman and Hall.Riedl, C. A. L. et al. 2007. Genetic and behavioral analysis of
natural variation in Drosophila melanogaster pupation position.� Fly 1: 23�32.
Rizki, R. M. and Rizki, T. M. 1984. Selective destruction of a hostblood cell type by a parasitoid wasp. � Proc. Natl Acad. Sci.USA 81: 6154�6158.
Rodriguez-del-Bosque, L. A. and Smith Jr., J. W. 1996. Rearingand biology of Lydella jalisco (Diptera: Tachinidae), a parasiteof Eoreuma loftini (Lepidoptera: Pyralidae) from Mexico.� Ann. Entomol. Soc. Am. 89: 88�95.
Sameoto, D. D. and Miller, R. S. 1968. Selection of pupation siteby Drosophila melanogaster and D. simulans. � Ecology 49:177�180.
Schnebel, E. M. and Grossfield, J. 1992. Temperature effects onpupation-height response in four Drosophila species grouptriads. � J. Insect Physiol. 38: 727�732.
Sokolowski, M. B. 1980. Foraging strategies of Drosophilamelanogaster: a chromosomal analysis. � Behav. Genet. 10:291�302.
Sokolowski, M. B. 1985. Genetics and ecology of Drosophilamelanogaster larval foraging and pupation behaviour.� J. Insect Physiol. 31: 857�864.
Sokolowski, M. B. and Hansell, R. I. C. 1983. Elucidating thebehavioral phenotype of Drosophila melanogaster larvae: corre-lations between larval foraging strategies and pupation height.� Behav. Genet. 13: 267�280.
Sokolowski, M. B. et al. 1986. Ecological genetics and behaviourof Drosophila melanogaster larvae in nature. � Anim. Behav. 34:403�408.
Stamp, N. E. 1981. Behavior of parasitized aposematic caterpillars� advantageous to the parasitoid or the host. � Am. Nat. 118:715�725.
Tanaka, S. and Ohsaki, N. 2006. Behavioral manipulation of hostcaterpillars by the primary parasitoid wasp Cotesia glomerata(L.) to construct defensive webs against hyperparasitism.� Ecol. Res. 21: 570�577.
Therneau, T. M. and Grambsch, P. M. 2000. Modelling survivaldata: extending the cox model. series: statistics for biology andhealth. � Springer.
Tauber, E. et al. 2007. Natural selection favors a newly derivedtimeless allele in Drosophila melanogaster. � Science 316:1895�1898.
Varaldi, J. et al. 2003. Infectious behavior in a parasitoid.� Science 302: 1930.
Wertheim, B. 2005. Genome-wide gene expression in response toparasitoid attack in Drosophila. � Genome Biol. 6: R94-1-20.
Supplementary material (available online as AppendixO17508 at/<www.oikos.ekol.lu.se/appendix/>). Appendix 1
1157