cladistic, phenetic and biogeographical analysis of the flightless dung beetle genus, gyronotus van...
TRANSCRIPT

Journal of Natural History 2001 35 1607ndash1625
Cladistic phenetic and biogeographical analysis of the macr ightless dungbeetle genus Gyronotus van Lansberge (Scarabaeidae Scarabaeinae)
in threatened eastern Afrotropical forests
ADRIAN L V DAVIS CLARKE H SCHOLTZ andJAMES DU G HARRISON
Department of Zoology amp Entomology University of Pretoria Pretoria
0002 South Africa e-mail adaviszoologyupacza
(Accepted 15 April 2000)
By virtue of their low vagility ightless insects are useful indicators of biogeo-graphical history Relationships of the ightless dung beetle genus Gyronotusare of particular interest due to its Gondwanaland ancestry distinctive relictdistribution along the south-eastern seaboard of Africa and its restriction toforests which are seriously threatened by exploitation Because of the limitednumber of diagnostic morphological characters it was necessary to code morpho-metric data in order to conduct distance and cladistic parsimony analysis ofinterspeci c relationships in Gyronotus There was a good correlation betweenrelationships indicated by the dendrogramscladograms and those determined byan examination of aedeagus character states both of which indicate a disjunctionbetween south and east African species and a broad separation between northernand southern South African species Comparison of the bilaterally asymmetricalaedeagi of Gyronotus with the symmetrical aedeagi of the sister genus AnachalcosHope suggests geographical polarization of character states from greater plesi-omorphy in east African Gyronotus to greater apomorphy in South Africanspecies particularly in the southernmost element in which the aedeagus showsextreme asymmetry Furthermore body shape follows a similar geographicalgradient in that the three Gyronotus species of tropical east Africa are signi cantlymore elongate than the three ovoid lowlandafromontane species of South AfricaAn examination of historical factors suggests that this spatially-restricted distribu-tion is the relict of a very old tropical lowland pattern In extant taxa thephylogenetic polarization is towards one of ve main centres of afrotropicalforest biodiversity in the geologically old Eastern Arc and the adjoining lowlandforest (Swahili centre of endemism) Survivors from old lineages may be onereason for such centres of high biodiversity
Keywords Afrotropical biogeographical cladistic dung ightless forestphenetic Scarabaeinae
Introduction
Eleven of the 20 extant genera of canthonine dung beetles in Africa are restrictedto the south-eastern seaboard between the Western Cape in South Africa and the
Journal of Natural HistoryISSN 0022-2933 printISSN 1464-5262 online copy 2001 Taylor amp Francis Ltd
httpwwwtandfcoukjournals
A L V Davis et al1608
East Usambara Mountains in north-eastern Tanzania (Scholtz and Howden 1987)where they are primarily found in forest habitats (Davis unpublished ) These 11genera may be regarded as relicts since they belong to a tribe with ancientGondwanaland a liations (HalŒter and Matthews 1966 HalŒter 1974 Cambefort1991) and are all species-poor This relictual distribution pattern is presumablyrelated to historical events particularly to the Miocene and Plio-Pleistocene upliftin east central Africa which separated the forests of eastern Africa from those inWest Africa (Lovett 1993 Burgess et al 1998) In East Africa survival of wetrainforest and its associated faunal relicts may be a consequence of the relativelystable surface temperatures (0ndash2 szlig C variation) in the Indian Ocean (Prell et al 1980)and continued coastal and orographic capture of high rainfall (Gillman 1949)during the Pleistocene cycles of cool dry and warm wet climate
The genus Gyronotus is one of the most species-rich of these eastern seaboardforest relicts It comprises six known species showing disjunct or parapatric distribu-tions (Scholtz and Howden 1987 Davis et al 1999) extending from the marginsof the bimodal rainfall region in the Eastern Cape South Africa (Davis 1993 1997)to the East Usambara Mountains which lie towards the northern end of the easternarc mountain system in north-eastern Tanzania All Gyronotus species are ightlessand are known to occur predominantly on ner-grained soils in lower altitude forestswith tropical or afromontane a nities Such ightless taxa with their low vagilityhave been shown to be most useful indicators of evolutionary and biogeographicalhistory (Bruhl 1997)
Both northern tropical and southern temperate (Palaeantarctic) Gondwanalandorigins have been hypothesized for afrotropical members of the tribe Canthonini(HalŒter 1974) However subsequent intra-continental dispersal vicariance eventsand extinctions have presumably shaped the modern relictual distribution patternsof the genera Phenetic cladistic and morphological analyses were conducted onGyronotus in order to examine their interspeci c relationships in comparison to thebiogeographical a nities of the species The results suggest a tropical generic a li-ation with a change in body shape from the lowland forest of East Africa to thelowland and low-lying afromontane forests of South Africa These geographicaldistribution patterns are discussed in an historical context
The ndings describe the relict distribution of an old dung beetle lineage ineastern Afrotropical forests In the present day these eastern forest habitats comprisean archipelago (Hamilton 1989 Low and Rebelo 1996 Burgess et al 1998) ofboth natural and anthropogenically-induced fragments which are seriouslythreatened by continuing exploitation Barnes (1990) predicted that these easternAfrotropical forests would be reduced by 95 by the year 2040 However recentanalyses suggest that the coastal portion between Somalia and Mozambique isalready reduced to a few thousand square kilometres of small fragments (Burgesset al 1998) Furthermore at least one of the species of Gyronotus dung beetles inthe present study (G glabrosus Scholtz and Howden) already deserves criticallyendangered (CR) status ( IUCN 1994) and may well be extinct (EX ) (Daviset al 1999)
Methods
Geographical distribution data for Gyronotus were derived from material heldby the University of Pretoria (now lodged in the Transvaal Museum Pretoria) andmaterial loaned by seven southern African and three European museums and
Cladistic and biogeographical analysis of Gyronotus 1609
institutions Specimens of four species were well-represented in these collections G
pumilus (Boheman) (1281 ) G carinatus Felsche (53 1 ) G Wmetarius Kolbe (41)and G mulanjensis Davis Scholtz and Harrison (33) However the institutionsheld only limited numbers of G dispar (Felsche) (7) and G glabrosus Scholtz andHowden (6) A seventh example of G glabrosus in the Howden Collection atCarleton University Ottawa Canada is the only other known museum specimenof this species During its known activity period in February 1998 unsuccessfulattempts were made to collect further material of this highly endemic species at itsone remaining known relatively undisturbed locality the afromontane Woodbushindigenous forest (23 szlig 52frac34 S 29 szlig 56frac34 E )
The morphology of Gyronotus showed relatively little variation and providedextremely few characters useful for cladistic analysis of interspeci c relationshipsIn such situations Chapill (1989) suggested that one alternative is to code morpho-metric data for cladistic analysis Since the use of phenetic characters for phylogeneticanalyses has been criticized (Wiley 1981) the results were compared with characterstates shown by the aedeagi These characters were the only morphological featureswhich showed diŒerences that could be interpreted with con dence as having strongphylogenetic signi cance
Twenty-seven morphometric characters used for distance or parsimony analysisof Gyronotus are listed in table 1 Measurements were made of all available specimensof G dispar (three males four females) and G glabrosus (four males two females)whereas ten specimens were measured for each of the other four species G WmetariusG pumilus G carinatus ( ve males ve females) and G mulanjensis (six malesfour females) The morphometric data matrix was treated in two diŒerent ways inorder to change the relative in uence of size and shape on the analyses Themeasurements (mm) were either left untransformed or were log10 transformed Bothtreatments were double standardized (Somers 1989) by subtracting each value fromthe row mean and each of these standardized values from the standardized columnmean For each treatment the data for each morphometric character was tested forsigni cant diŒerences between the six species using one-way ANOVA (table 1)Multiple range tests were conducted on each character using Tukeyrsquos HSD forunequal sample size (SpjotvollStoline Test ) On the basis of the results for thesemultiple range tests each character was coded for cladistic analysis using the diver-gence coding method of Thorpe (1984) This method provides a diŒerent code foreach signi cantly diŒerent character state and develops a species Ouml character statedistance matrix coded in categorical terms
Both distance and cladistic parsimony analyses were conducted on each of thetwo divergence matrices (table 2) using PHYLIP version 35 (Felsenstein 1993) andPAUP version 31 (SwoŒord 1993) Distance analyses were conducted using all 27characters in each matrix and species were linked by neighbour joining In contrastonly informative characters were used for cladistic parsimony analysis In the matrixderived from raw double-standardized data there were only 11 informativecharacters whereas in that derived from log10 double-standardized data there were14 informative characters In each analysis the character states were treated asunordered and the cladograms were centre-rooted
Each of the morphometric matrices was also subjected to principal componentsanalysis (PCA) As the rst component frequently correlates to size whereas follow-ing components correlate primarily to shape (Somers 1989 Fairbairn 1992) theordinate values for the rst two factors from each analysis were tested for correlation
A L V Davis et al1610
Table 1 List of morphometric characters for Gyronotus and the results of one-way ANOVAon interspeci c diŒerences between the characters either double-standardized rawmeasurements in mm or double-standardized log10 transformed measurements (seemethods)
F(5 47) F(5 47)(raw data)dagger ( log10 data)dagger
1 Distance between the tips of the clypeal teeth 17255 45972 Distance between the genal angles 6708 68373 Distance between the centre of the clypeal emargination and 10360 3422
a line drawn between the genal angles4 Distance between the centre of the clypeal emargination and 1384 1971
the posterior margin of the vertex5 Width between the anterior angles of the pronotal disc 4169 41236 Width between the lateral angles of the pronotal disc 3240 16667 Width between the posterior angles of the pronotal disc 493 20288 Length along the middle of the pronotal disc 4780 52659 Maximum depth of the pronotal disc between the dorsal 4695 3887
surface and the ventral surface of the profemora10 Depth of the pronotal disc between the dorsal surface and 1002 926
the pleural margin at the anterior end11 Depth of the pronotal disc between the dorsal surface and 1511 874
the pleural margin at the posterior end12 Distance from the centre of the posterior margin to a line 7113 2056
drawn between the posterior angles of the prothoraccic disc13 Length along the midline of the metasternum 665 105514 Distance between the left mesocoxa and left metacoxa 8648 43715 Distance between the right mesocoxa and the left metacoxa 1243 80316 Distance between the mesocoxae 1093 103517 Maximum depth from the elytra to the metasternum 3518 94618 Depth from the dorsal extremity of the left elytron to the 2037 889
pleural margin at the anterior end19 Maximum depth of the left elytron to the pleural margin 732 105820 Maximum height of the curvature along the lateral edge of 7638 1327
the left elytron above a line drawn between the anterior andposterior ends
21 Maximum width of the elytral pseudo-epipleurae 7702 280722 Maximum width of the right elytron Ouml 2 2496 388323 Width of the right elytron at the anterior end Ouml 2 1153 271824 Maximum length of the right elytron 5538 99225 Width of the pygidium along the basal ridge 1733 204226 Length of pygidium from the tip to the basal ridge 2326 227827 Length of the sternites along the midline of the abdomen 2709 1222
between the metasternum and the pygidium
daggerAll F numbers statistically signi cant plt0001
with both body length and the ratio of body length to maximum width of the elytraThese respective measurements were used as indices for body size and body shapeThe results provide some indication of the relative in uence of these two parameterson the four derived matrices used for distance and parsimony analyses
The relationships indicated by the cladograms are compared to characters shownby the aedeagi of Gyronotus those of Anachalcos Hope and that of Canthodimorpha
Davis Scholtz and Harrison which is a newly-discovered large-bodied monotypictropical genus from the northern coastline of Mozambique (Davis et al 1999)
Cladistic and biogeographical analysis of Gyronotus 1611
Tab
le2
Div
erge
nce
mat
rice
s(T
horp
e19
84)dagger
for
dist
ance
and
clad
isti
cpa
rsim
ony
anal
ysis
ofin
ter-
rela
tio
nshi
ps
betw
een
Gyro
notu
ssp
ecie
s
Ch
arac
ter
12
34
56
78
910
1112
1314
1516
1718
1920
2122
2324
2526
27
Der
ived
fro
mdo
uble
-sta
ndar
dize
dra
wph
enet
icm
easu
rem
ents
(mm
)C
har
in
form
ativ
eDaggeri
ii
ii
ii
ii
ii
G
pum
ilus
58
46
87
48
86
38
65
45
64
36
26
45
21
4G
ca
rinatu
s1
84
68
85
88
42
86
24
47
28
02
53
88
70
G
gla
bro
sus
18
46
87
58
84
28
63
44
71
21
42
39
97
4G
dis
par
a3
a7
20
40
09
80
5a
aa
08
6a
a1
30
37
9G
m
ula
nje
nsi
s5
34
42
57
32
37
26
44
46
78
66
a9
43
14
G
Wm
etari
us
80
41
23
53
44
84
16
43
48
36
66
84
57
9
Der
ived
fro
mdo
uble
-sta
ndar
dize
dlo
g 10tr
ansf
orm
edra
wm
easu
rem
ents
Ch
ar
info
rmat
iveDagger
ii
ii
ii
ii
ii
ii
ii
G
pum
ilus
57
46
75
38
88
43
68
76
64
47
34
35
32
5G
ca
rinatu
s1
75
77
a4
88
84
36
46
67
69
13
43
89
80
G
gla
bro
sus
17
87
74
38
85
34
65
66
71
32
30
28
98
6G
dis
par
97
96
75
a0
23
54
65
56
16
37
99
93
33
5G
m
ula
nje
nsi
s5
24
42
57
31
36
66
44
66
68
69
99
33
26
G
Wm
etari
us
90
00
01
33
33
8a
04
20
37
37
34
43
37
8
dagger Rec
oded
fro
ma
rang
eof
Otilde5
to5
toa
rang
efr
om
0to
10(1
0re
cod
edas
lsquoarsquo
wh
ich
isre
adas
one
unit
grea
ter
than
9in
the
dist
ance
anal
ysis
)Dagger A
llch
arac
ters
used
for
dist
ance
anal
ysis
on
lyin
form
ativ
ech
arac
ters
(i)
used
for
pars
imo
nyan
alys
is
A L V Davis et al1612
Although Anachalcos has been identi ed as the sister group (Scholtz and Howden1987) it is probably quite distant from Gyronotus which is both ightless and hasasymmetrical aedeagi However it is a useful comparison since the aedeagi ofAnachalcos are similar to those of most other afrotropical Canthonini in as muchas the parameres are symmetrical (Scholtz and Howden 1987) and lack the mem-branous terminal process on the left paramere which is characteristic of all Gyronotus
(Davis et al 1999)
Results
Geographical distribution
The six Gyronotus species showed either disjunct or parapatric distributions fromthe Eastern Cape in South Africa to north-eastern Tanzania ( gure 1) Two SouthAfrican species G pumilus (Eastern Cape southern KwaZulu-Natal ) and G
carinatus (northern KwaZulu-Natal ) showed a parapatric distribution pattern larg-ely restricted to coastal hills below 320m ( gure 2) The southern distribution limitsof G pumilus and the northern distribution limits of G carinatus coincide with thesouthern and northern limits of Walter and Liethrsquos (1964) temperatesubtropicalclimate type II (I )a ( gure 2) The respective northern and southern distribution
Fig 1 Geographical distribution of Gyronotus species from east to southern Africa(T Tanzania M Malawi SA South Africa) (S Swahili centre of endemism SMSwahili-Maputaland transition zone (Burgess et al 1998) Ma Maputaland centre ofendemism P Pondoland centre of endemism which occupies a narrow coastal zone(van Rensburg et al 1999))
Cladistic and biogeographical analysis of Gyronotus 1613
Fig 2 Geographical distribution of Gyronotus pumilus and G carinatus relative to altitudeand the subtropical coastal climate type II(I )a after Walter and Lieth (1964)
limits of G pumilus and G carinatus occur just north of Durban (29 szlig 51frac34 S 31 szlig 01frac34 E )at which latitude land greater than 1500 m in altitude swings away from the coastlineThe third southern African species was recorded only in a small localized region ofthe eastern escarpment of Northern Province in South Africa The combined distribu-tion of these species is sympatric with both coastal forest (particularly in the south)and the low-lying portion of forest classi ed as Afromontane (particularly in thenorth) (table 3) Distribution data for the east African species was geographicallymore limited ( gure 1) Most G Wmetarius were recorded from 215ndash850m in theEast Usambara Mountains of north-eastern Tanzania although a few specimenswere collected from the adjacent coastline The few G dispar were recorded fromsouth-eastern Tanzania around Lindi (10 szlig 00frac34 S 39 szlig 42frac34 E ) and along the RovumaRiver (11 szlig S 39 szlig E ) All G mulanjensis were recorded from 900ndash1000 m on MtMulanje (16 szlig 22frac34 S 35 szlig 07frac34 E ) in southern Malawi
Distance and cladistic analyses
The dendrograms and cladograms derived from the four diŒerent analyses areshown by gure 3 In essential details the inter-relationships among the species wereconsistent with their geographical distribution Each dendrogramcladogram showeda clear branching between east and southern African taxa and in each case there
A L V Davis et al1614
Tab
le3
Sum
mar
yof
know
nge
ogra
phic
aldi
stri
but
ion
and
habi
tat
asso
ciat
ion
sof
Gyro
notu
ssp
pbe
twee
nth
eE
aste
rnC
ape
Sout
hA
fric
aan
dno
rth
-eas
tern
Tan
zani
a
Len
gth
Geo
grap
hic
alra
nge
Alt
itu
din
alG
enu
san
dsp
ecie
s(m
m)dagger
(pat
chy
occu
rren
ce)
rang
e(m
)H
abit
atN
ote
s
Gyro
notu
sva
nL
ansb
erge
1874
South
Afr
ica
All
know
nre
cord
sin
fore
stG
pum
ilus
(Boh
eman
1857
)13
ndash15
Sout
h-ea
stco
asta
lhi
lls50
ndash450
For
est
469
of
128
info
rest
(gu
res
12
)G
ca
rinatu
sF
elsc
he19
1113
ndash15
Zul
ula
nd
coas
tal
hills
50ndash5
00F
ores
t5
7of
53in
fore
st(
gure
s1
2)
G
gla
bro
sus
Scho
ltz
and
Ho
wde
n19
8712
ndash12
5D
rak
ensb
erg
esca
rpm
ent
in78
0F
ores
t33
of
6in
fore
stN
ort
hern
Pro
vinc
e(
gure
s1
2)
Tro
pic
al
Afr
ica
Gm
ula
nje
nsi
sD
avis
Sch
olt
zan
dH
arri
son
118
ndash13
3M
tM
ulan
je
Mal
awi
(gu
re1
)90
0ndash10
00F
ores
tN
ot
reco
rded
F
ores
tan
d19
99op
enco
llect
ion
sm
ixed
DaggerG
Wm
etari
us
Ko
lbe
1894
125
ndash15
5M
ain
lyE
ast
Usa
mba
raM
ts50
ndash850
For
est
Fou
rtr
appe
dby
stre
amin
plu
sea
stco
ast
ofT
anza
nia
fore
stbu
tno
neou
tsid
e(
gure
1)
G
dis
par
(Fel
sche
1911
)16
ndash18
Sout
h-ea
stT
anza
nia
(gu
re1
)50
For
est
No
habi
tat
note
s
dagger Len
gth
wit
hhe
adde
exe
das
oppo
sed
tora
ised
asin
tabl
e5
Dagger C
L
Bel
lam
y(p
erso
nal
com
mun
icat
ion)
Cladistic and biogeographical analysis of Gyronotus 1615
Fig 3 Centre-rooted distance dendrograms and centre-rooted consensus parsimony clado-grams for inter-relationships between Gyronotus species (A B) Cladistic matrix derivedfrom double-standardized raw morphometric measurements by divergence coding(Thorpe 1984) (A) Distance dendrogram based on 27 characters (B) Parsimonycladogram based on 11 informative characters tree length 5 28 CI 5 0893 (C D)Cladistic matrix derived from double-standardized log10 transformed morphometricmeasurements by divergence coding (Thorpe 1984) (C ) Distance dendrogram basedon 27 characters (D) Parsimony cladogram based on 14 informative characters treelength 5 36 CI 5 0889 The numbers are the branch lengths from the nodes forunordered character states
was a similar resolution of relationships between South African species with thesouthernmost species G pumilus distanced from the two more northerly speciesHowever the results for the east African species indicated three diŒerent hypotheticalinter-relationships between species Of the three diŒerent patterns one each wasyielded from the matrices derived from double-standardized raw data whereas thethird was common to the dendrogram and cladogram yielded by the matrices derived
A L V Davis et al1616
from double-standardized log10 data These diŒerences may re ect the relativein uence of body size versus body shape according to the treatment of the datamatrices In the matrices derived from double-standardized raw data body size hasa stronger overall in uence than body shape (higher r2 and F numbers for regressionagainst Factor 1 which accounts for the greatest amount of variance in ordination(table 4) This emphasis is re ected by the closer linking of the two largest speciesG dispar and G Wmetarius Although both body size and body shape are de-emphasized in the matrices derived from double-standardized log10 data body shapehas the greater overall in uence (higher r2 and F numbers for regression againstboth Factors 1 and 2) This emphasis is re ected by the steps in common betweenthe elongate G dispar and G mulanjensis whereas the less elongate G Wmetarius isplaced on a separate branch ( gures 3C 3D)
Further phenetic analysis
As the cladograms were derived from morphometric data it is not surprisingthat the depicted interspeci c relationships are consistent with those indicated bythe three basic measurements of maximum length width and depth These measure-ments clearly separate the more elongate deeper east African species from the moreovoid atter southern African species (table 5) Ratios between the three dimen-sions indicate that width varies inversely with the other two dimensions whereaslength and depth vary proportionately to one another Thus the broadest species(G glabrosus) is also the attest and the most narrow species (G dispar) is alsothe deepest
Character states of aedeagi
F igure 4 illustrates the morphological diŒerences between the aedeagi ofGyronotus and those of large-bodied putative sister groups with primarily tropicaldistributions The parameres of tropical species in putative sister genera Anachalcos
procerus Gerstaecker and Canthodimorpha lawrencei Davis Scholtz and Harrisonare symmetrical and diverge at the tips Those of the east African Gyronotus alsodiverge at the tips but are asymmetrical due to a terminal process on the leftparamere The tips of the parameres of the southern African Gyronotus species aremore convergent and are also asymmetrical due to a terminal process projectingaway to the left of the left paramere
Discussion
Geographical polarization of characters
Comparison of Gyronotus aedeagi with those of the putative sister groupAnachalcos and that of Canthodimorpha suggests that bilateral symmetry anddivergence at the tips of the parameres are plesiomorphic character states ( gure 4)Although the aedeagi of the east African species of Gyronotus are characterized byasymmetry this is primarily due to the presence of a membranous terminal processwhich in ventral view projects to the right from the left paramere and occupies thespace between the divergent tips of the parameres ( gure 4) In contrast the SouthAfrican species are characterized by apomorphic convergence of the tips of theparameres ( gure 4mdashparameres slightly separated in the gure to facilitate drawing)In the two northernmost South African species asymmetry of the parameres ismainly due to the membranous terminal process which in ventral view projects
Cladistic and biogeographical analysis of Gyronotus 1617
Tab
le4
The
corr
elat
ion
ofG
yro
notu
sbo
dysi
ze(m
axim
umle
ngt
h)1
and
body
shap
e(r
atio
max
imu
mle
ngt
hm
axim
um
wid
thof
elyt
ra)2
wit
hor
din
ates
for
Fac
tors
1an
d2
deri
ved
fro
mpr
inci
pal
com
pon
ents
anal
ysis
ofm
orp
hom
etri
csda
tatr
eate
din
two
diŒ
eren
tw
ays
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
raw
mea
sure
men
tsra
wm
easu
rem
ents
log 10
mea
sure
men
tslo
g 10m
easu
rem
ents
(27
char
acte
rs)
(11
char
acte
rs)
(27
char
acte
rs)
(14
char
acte
rs)
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Pro
por
tion
ofov
eral
lvar
ianc
e(
)55
217
756
015
058
313
266
413
6R
2fo
rre
gres
sio
nof
body
size
on0
980
0002
092
000
20
060
230
080
24or
din
ates
1F
(15
1)fo
rre
gres
sio
nof
body
size
2870
60
0
0162
106
008
345
150
9
437
16
38
on
ordi
nat
es1
R2
for
regr
essi
on
ofbo
dysh
ape
on0
620
130
670
060
150
250
140
30or
din
ates
2F
(15
1)fo
rre
gres
sio
nof
body
shap
e83
55
7
55
103
79
3
358
63
173
1
873
22
26
on
ordi
nat
es2
p
lt0
05
p
lt0
02
p
lt0
001
A L V Davis et al1618
Tab
le5
Mor
pho
met
ric
mea
sure
men
tsfo
rsi
xsp
ecie
sof
Gyro
notu
s
Sout
hern
Afr
ican
spec
ies
Eas
tA
fric
ansp
ecie
sA
NO
VA
Mea
sure
men
tG
pum
ilus
G
cari
natu
sG
gla
bro
sus
G
dis
par
G
mula
nje
nsi
sG
Wm
etari
us
F(5
47)
Len
gth
(L)
147
2Ocirc0
64c
141
4Ocirc0
26cd
135
8Ocirc0
77d
206
4Ocirc0
41a
159
5Ocirc0
34b
162
0Ocirc0
26b
194
41
W
idth
(W)
905
Ocirc0
36b
900
Ocirc0
18b
944
Ocirc0
53b
107
4Ocirc0
42a
824
Ocirc0
22c
911
Ocirc0
46b
354
1
Dep
th(D
)6
02Ocirc
023
bd5
80Ocirc
021
cd5
57Ocirc
021
d8
24Ocirc
038
a6
09Ocirc
024
bc6
38Ocirc
027
b88
35
R
atio
sL
W0
62Ocirc
002
b0
64Ocirc
001
b0
70Ocirc
002
a0
52Ocirc
002
d0
52Ocirc
001
d0
56Ocirc
003
c87
16
L
D0
41Ocirc
001
a0
41Ocirc
001
a0
41Ocirc
002
a0
40Ocirc
002
ab0
38Ocirc
001
b0
39Ocirc
002
ab5
16
W
D0
67Ocirc
003
bc0
64Ocirc
002
c0
59Ocirc
002
d0
77Ocirc
004
ab0
74Ocirc
002
ab0
70Ocirc
003
b35
44
Sa
mp
lesi
ze10
106
710
10
Inea
chro
w
valu
esfo
llow
edby
adi
Œer
ent
lett
erdi
Œer
edsi
gni
cant
ly(T
ukey
rsquosH
SD
for
uneq
ual
nETH
plt
005
)
plt
000
1
Cladistic and biogeographical analysis of Gyronotus 1619
Fig 4 Distal and ventral views of canthonine aedeagi showing diŒerences in parameremorphology between (A) Anachalcos procerus Gerstaecker (distal ) (B)Canthodimorpha lawrencei Davis Scholtz and Harrison (ventral) (C ) east AfricanGyronotus van Lansberge (ventral ) (C1 G Wmetarius Kolbe C2 G mulanjensis DavisScholtz and Harrison C3 G dispar (Felsche)) and (D) South African Gyronotus(ventral ) (D1 G carinatus Felsche D2 G glabrosus Scholtz and Howden D3G pumilus (Boheman)) Scale bars 5 1 mm
away to the left of the left paramere However in the southernmost species theasymmetry is more extreme in that each paramere is radically diŒerent in shape(Davis et al 1999) The left paramere is more or less straight and bears the leftwardsaligned terminal process the right paramere is curved towards the mid-point andboth are quite unlike those of the other ve species in which each paramere issimilarly curved close to the distal end Thus there is clear biogeographical polariza-tion of the character states of the aedeagi with greater plesiomorphy in eastAfrican species
A L V Davis et al1620
Parallels between morphology biogeography and phenetic analysis of GyronotusThe interspeci c relationships indicated by the cladograms derived from phenetic
analysis largely corresponded to relationships suggested by morphological charactersof the aedeagi ( gure 4 Davis et al 1999) and these relationships were consistentwith the biogeographical distribution of the species along the south-eastern seaboardof Africa The clear separation of the elongate east African and more ovoid southernAfrican species is re ected by the diŒering position of the terminal process on theleft paramere of the aedeagi ( gure 4) The shape of this process in G Wmetarius
and in G mulanjensis diŒers only slightly in that the one is rounded and the otherangular Although it appears that these two species are very close the angularprocess of G dispar is su ciently similar that it is di cult to determine which ofthe cladograms if any re ects the true phylogenetic relationships between the eastAfrican species those strongly re ecting size and shape ( gures 3A 3B) or thosemore biased by shape ( gures 3C 3D) It is also possible that evolution of aedeagiand morphometric character states are divergent rather than parallel The relation-ships between the southern African species are resolved with greater consistencyThe shape of the parameres is similar in the two northerly species which diŒerprincipally only in the mono d shape of the outward projecting terminal process inG carinatus and the bi d shape of this process in G glabrosus ( gure 4) The moredistant phenetic separation of the southernmost species G pumilus is very stronglysupported by the characters of its aedeagus which diŒer radically from those of theother species However the outward projection of the terminal process indicates itscloser relationships to the other South African species than to the east Africanspecies
Biogeographical climatic and habitat associations of GyronotusSingle Gyronotus species are characteristic of several southern sub-centres of
endemism identi ed in mainland eastern coastal forests of Africa by Burgess et al(1998) These sub-centres occur in the East UsambaraTanga (G Wmetarius) Lindi(G dispar) and UdzungwaMulanje mountain regions (G mulanjensis) The coastalforests of southern Tanzania (Lindi subcentre) contain the most unique assemblageof endemic taxa along the east coast (Burgess et al 1998) and also harbour thelargest of the six known Gyronotus species (G dispar)
In terms of lowland versus afromontane forest association the biogeographicala nities of Gyronotus are dependent on the system used to classify oral zonationIn diŒerent regions altitudinal temperature zonation is modi ed by a combinationof factors including latitude rainfall regime aspect and the relative in uence ofmaritime versus continental climate (Hamilton 1989) Therefore instead of usinga single in exible classi cation system we have used three regional systems whichmake allowance for geographical diŒerences in the altitudinal occurrence of thesame temperature zones and their related ora These classi cation systems are thoseproposed by Lovett (1993) for the Eastern Arc Chapman and White (1970) forMalawi and Low and Rebelo (1996) for southern Africa Thus the East Africanspecies of Gyronotus may be described as elements of tropical lowland forests whichoccur below 1370 m in Malawi (Chapman and White 1970) and below 800m in theeastern arc (Lovett 1993) In the more southerly latitudes of South Africa mostlowland and highland forest is classi ed as afromontane (Low and Rebelo 1996)except for that on coastal sandveld of recent origin (Lawes 1990) AlthoughGyronotus have been recorded in coastal regions around East London (33 szlig 00frac34 S
Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

A L V Davis et al1608
East Usambara Mountains in north-eastern Tanzania (Scholtz and Howden 1987)where they are primarily found in forest habitats (Davis unpublished ) These 11genera may be regarded as relicts since they belong to a tribe with ancientGondwanaland a liations (HalŒter and Matthews 1966 HalŒter 1974 Cambefort1991) and are all species-poor This relictual distribution pattern is presumablyrelated to historical events particularly to the Miocene and Plio-Pleistocene upliftin east central Africa which separated the forests of eastern Africa from those inWest Africa (Lovett 1993 Burgess et al 1998) In East Africa survival of wetrainforest and its associated faunal relicts may be a consequence of the relativelystable surface temperatures (0ndash2 szlig C variation) in the Indian Ocean (Prell et al 1980)and continued coastal and orographic capture of high rainfall (Gillman 1949)during the Pleistocene cycles of cool dry and warm wet climate
The genus Gyronotus is one of the most species-rich of these eastern seaboardforest relicts It comprises six known species showing disjunct or parapatric distribu-tions (Scholtz and Howden 1987 Davis et al 1999) extending from the marginsof the bimodal rainfall region in the Eastern Cape South Africa (Davis 1993 1997)to the East Usambara Mountains which lie towards the northern end of the easternarc mountain system in north-eastern Tanzania All Gyronotus species are ightlessand are known to occur predominantly on ner-grained soils in lower altitude forestswith tropical or afromontane a nities Such ightless taxa with their low vagilityhave been shown to be most useful indicators of evolutionary and biogeographicalhistory (Bruhl 1997)
Both northern tropical and southern temperate (Palaeantarctic) Gondwanalandorigins have been hypothesized for afrotropical members of the tribe Canthonini(HalŒter 1974) However subsequent intra-continental dispersal vicariance eventsand extinctions have presumably shaped the modern relictual distribution patternsof the genera Phenetic cladistic and morphological analyses were conducted onGyronotus in order to examine their interspeci c relationships in comparison to thebiogeographical a nities of the species The results suggest a tropical generic a li-ation with a change in body shape from the lowland forest of East Africa to thelowland and low-lying afromontane forests of South Africa These geographicaldistribution patterns are discussed in an historical context
The ndings describe the relict distribution of an old dung beetle lineage ineastern Afrotropical forests In the present day these eastern forest habitats comprisean archipelago (Hamilton 1989 Low and Rebelo 1996 Burgess et al 1998) ofboth natural and anthropogenically-induced fragments which are seriouslythreatened by continuing exploitation Barnes (1990) predicted that these easternAfrotropical forests would be reduced by 95 by the year 2040 However recentanalyses suggest that the coastal portion between Somalia and Mozambique isalready reduced to a few thousand square kilometres of small fragments (Burgesset al 1998) Furthermore at least one of the species of Gyronotus dung beetles inthe present study (G glabrosus Scholtz and Howden) already deserves criticallyendangered (CR) status ( IUCN 1994) and may well be extinct (EX ) (Daviset al 1999)
Methods
Geographical distribution data for Gyronotus were derived from material heldby the University of Pretoria (now lodged in the Transvaal Museum Pretoria) andmaterial loaned by seven southern African and three European museums and
Cladistic and biogeographical analysis of Gyronotus 1609
institutions Specimens of four species were well-represented in these collections G
pumilus (Boheman) (1281 ) G carinatus Felsche (53 1 ) G Wmetarius Kolbe (41)and G mulanjensis Davis Scholtz and Harrison (33) However the institutionsheld only limited numbers of G dispar (Felsche) (7) and G glabrosus Scholtz andHowden (6) A seventh example of G glabrosus in the Howden Collection atCarleton University Ottawa Canada is the only other known museum specimenof this species During its known activity period in February 1998 unsuccessfulattempts were made to collect further material of this highly endemic species at itsone remaining known relatively undisturbed locality the afromontane Woodbushindigenous forest (23 szlig 52frac34 S 29 szlig 56frac34 E )
The morphology of Gyronotus showed relatively little variation and providedextremely few characters useful for cladistic analysis of interspeci c relationshipsIn such situations Chapill (1989) suggested that one alternative is to code morpho-metric data for cladistic analysis Since the use of phenetic characters for phylogeneticanalyses has been criticized (Wiley 1981) the results were compared with characterstates shown by the aedeagi These characters were the only morphological featureswhich showed diŒerences that could be interpreted with con dence as having strongphylogenetic signi cance
Twenty-seven morphometric characters used for distance or parsimony analysisof Gyronotus are listed in table 1 Measurements were made of all available specimensof G dispar (three males four females) and G glabrosus (four males two females)whereas ten specimens were measured for each of the other four species G WmetariusG pumilus G carinatus ( ve males ve females) and G mulanjensis (six malesfour females) The morphometric data matrix was treated in two diŒerent ways inorder to change the relative in uence of size and shape on the analyses Themeasurements (mm) were either left untransformed or were log10 transformed Bothtreatments were double standardized (Somers 1989) by subtracting each value fromthe row mean and each of these standardized values from the standardized columnmean For each treatment the data for each morphometric character was tested forsigni cant diŒerences between the six species using one-way ANOVA (table 1)Multiple range tests were conducted on each character using Tukeyrsquos HSD forunequal sample size (SpjotvollStoline Test ) On the basis of the results for thesemultiple range tests each character was coded for cladistic analysis using the diver-gence coding method of Thorpe (1984) This method provides a diŒerent code foreach signi cantly diŒerent character state and develops a species Ouml character statedistance matrix coded in categorical terms
Both distance and cladistic parsimony analyses were conducted on each of thetwo divergence matrices (table 2) using PHYLIP version 35 (Felsenstein 1993) andPAUP version 31 (SwoŒord 1993) Distance analyses were conducted using all 27characters in each matrix and species were linked by neighbour joining In contrastonly informative characters were used for cladistic parsimony analysis In the matrixderived from raw double-standardized data there were only 11 informativecharacters whereas in that derived from log10 double-standardized data there were14 informative characters In each analysis the character states were treated asunordered and the cladograms were centre-rooted
Each of the morphometric matrices was also subjected to principal componentsanalysis (PCA) As the rst component frequently correlates to size whereas follow-ing components correlate primarily to shape (Somers 1989 Fairbairn 1992) theordinate values for the rst two factors from each analysis were tested for correlation
A L V Davis et al1610
Table 1 List of morphometric characters for Gyronotus and the results of one-way ANOVAon interspeci c diŒerences between the characters either double-standardized rawmeasurements in mm or double-standardized log10 transformed measurements (seemethods)
F(5 47) F(5 47)(raw data)dagger ( log10 data)dagger
1 Distance between the tips of the clypeal teeth 17255 45972 Distance between the genal angles 6708 68373 Distance between the centre of the clypeal emargination and 10360 3422
a line drawn between the genal angles4 Distance between the centre of the clypeal emargination and 1384 1971
the posterior margin of the vertex5 Width between the anterior angles of the pronotal disc 4169 41236 Width between the lateral angles of the pronotal disc 3240 16667 Width between the posterior angles of the pronotal disc 493 20288 Length along the middle of the pronotal disc 4780 52659 Maximum depth of the pronotal disc between the dorsal 4695 3887
surface and the ventral surface of the profemora10 Depth of the pronotal disc between the dorsal surface and 1002 926
the pleural margin at the anterior end11 Depth of the pronotal disc between the dorsal surface and 1511 874
the pleural margin at the posterior end12 Distance from the centre of the posterior margin to a line 7113 2056
drawn between the posterior angles of the prothoraccic disc13 Length along the midline of the metasternum 665 105514 Distance between the left mesocoxa and left metacoxa 8648 43715 Distance between the right mesocoxa and the left metacoxa 1243 80316 Distance between the mesocoxae 1093 103517 Maximum depth from the elytra to the metasternum 3518 94618 Depth from the dorsal extremity of the left elytron to the 2037 889
pleural margin at the anterior end19 Maximum depth of the left elytron to the pleural margin 732 105820 Maximum height of the curvature along the lateral edge of 7638 1327
the left elytron above a line drawn between the anterior andposterior ends
21 Maximum width of the elytral pseudo-epipleurae 7702 280722 Maximum width of the right elytron Ouml 2 2496 388323 Width of the right elytron at the anterior end Ouml 2 1153 271824 Maximum length of the right elytron 5538 99225 Width of the pygidium along the basal ridge 1733 204226 Length of pygidium from the tip to the basal ridge 2326 227827 Length of the sternites along the midline of the abdomen 2709 1222
between the metasternum and the pygidium
daggerAll F numbers statistically signi cant plt0001
with both body length and the ratio of body length to maximum width of the elytraThese respective measurements were used as indices for body size and body shapeThe results provide some indication of the relative in uence of these two parameterson the four derived matrices used for distance and parsimony analyses
The relationships indicated by the cladograms are compared to characters shownby the aedeagi of Gyronotus those of Anachalcos Hope and that of Canthodimorpha
Davis Scholtz and Harrison which is a newly-discovered large-bodied monotypictropical genus from the northern coastline of Mozambique (Davis et al 1999)
Cladistic and biogeographical analysis of Gyronotus 1611
Tab
le2
Div
erge
nce
mat
rice
s(T
horp
e19
84)dagger
for
dist
ance
and
clad
isti
cpa
rsim
ony
anal
ysis
ofin
ter-
rela
tio
nshi
ps
betw
een
Gyro
notu
ssp
ecie
s
Ch
arac
ter
12
34
56
78
910
1112
1314
1516
1718
1920
2122
2324
2526
27
Der
ived
fro
mdo
uble
-sta
ndar
dize
dra
wph
enet
icm
easu
rem
ents
(mm
)C
har
in
form
ativ
eDaggeri
ii
ii
ii
ii
ii
G
pum
ilus
58
46
87
48
86
38
65
45
64
36
26
45
21
4G
ca
rinatu
s1
84
68
85
88
42
86
24
47
28
02
53
88
70
G
gla
bro
sus
18
46
87
58
84
28
63
44
71
21
42
39
97
4G
dis
par
a3
a7
20
40
09
80
5a
aa
08
6a
a1
30
37
9G
m
ula
nje
nsi
s5
34
42
57
32
37
26
44
46
78
66
a9
43
14
G
Wm
etari
us
80
41
23
53
44
84
16
43
48
36
66
84
57
9
Der
ived
fro
mdo
uble
-sta
ndar
dize
dlo
g 10tr
ansf
orm
edra
wm
easu
rem
ents
Ch
ar
info
rmat
iveDagger
ii
ii
ii
ii
ii
ii
ii
G
pum
ilus
57
46
75
38
88
43
68
76
64
47
34
35
32
5G
ca
rinatu
s1
75
77
a4
88
84
36
46
67
69
13
43
89
80
G
gla
bro
sus
17
87
74
38
85
34
65
66
71
32
30
28
98
6G
dis
par
97
96
75
a0
23
54
65
56
16
37
99
93
33
5G
m
ula
nje
nsi
s5
24
42
57
31
36
66
44
66
68
69
99
33
26
G
Wm
etari
us
90
00
01
33
33
8a
04
20
37
37
34
43
37
8
dagger Rec
oded
fro
ma
rang
eof
Otilde5
to5
toa
rang
efr
om
0to
10(1
0re
cod
edas
lsquoarsquo
wh
ich
isre
adas
one
unit
grea
ter
than
9in
the
dist
ance
anal
ysis
)Dagger A
llch
arac
ters
used
for
dist
ance
anal
ysis
on
lyin
form
ativ
ech
arac
ters
(i)
used
for
pars
imo
nyan
alys
is
A L V Davis et al1612
Although Anachalcos has been identi ed as the sister group (Scholtz and Howden1987) it is probably quite distant from Gyronotus which is both ightless and hasasymmetrical aedeagi However it is a useful comparison since the aedeagi ofAnachalcos are similar to those of most other afrotropical Canthonini in as muchas the parameres are symmetrical (Scholtz and Howden 1987) and lack the mem-branous terminal process on the left paramere which is characteristic of all Gyronotus
(Davis et al 1999)
Results
Geographical distribution
The six Gyronotus species showed either disjunct or parapatric distributions fromthe Eastern Cape in South Africa to north-eastern Tanzania ( gure 1) Two SouthAfrican species G pumilus (Eastern Cape southern KwaZulu-Natal ) and G
carinatus (northern KwaZulu-Natal ) showed a parapatric distribution pattern larg-ely restricted to coastal hills below 320m ( gure 2) The southern distribution limitsof G pumilus and the northern distribution limits of G carinatus coincide with thesouthern and northern limits of Walter and Liethrsquos (1964) temperatesubtropicalclimate type II (I )a ( gure 2) The respective northern and southern distribution
Fig 1 Geographical distribution of Gyronotus species from east to southern Africa(T Tanzania M Malawi SA South Africa) (S Swahili centre of endemism SMSwahili-Maputaland transition zone (Burgess et al 1998) Ma Maputaland centre ofendemism P Pondoland centre of endemism which occupies a narrow coastal zone(van Rensburg et al 1999))
Cladistic and biogeographical analysis of Gyronotus 1613
Fig 2 Geographical distribution of Gyronotus pumilus and G carinatus relative to altitudeand the subtropical coastal climate type II(I )a after Walter and Lieth (1964)
limits of G pumilus and G carinatus occur just north of Durban (29 szlig 51frac34 S 31 szlig 01frac34 E )at which latitude land greater than 1500 m in altitude swings away from the coastlineThe third southern African species was recorded only in a small localized region ofthe eastern escarpment of Northern Province in South Africa The combined distribu-tion of these species is sympatric with both coastal forest (particularly in the south)and the low-lying portion of forest classi ed as Afromontane (particularly in thenorth) (table 3) Distribution data for the east African species was geographicallymore limited ( gure 1) Most G Wmetarius were recorded from 215ndash850m in theEast Usambara Mountains of north-eastern Tanzania although a few specimenswere collected from the adjacent coastline The few G dispar were recorded fromsouth-eastern Tanzania around Lindi (10 szlig 00frac34 S 39 szlig 42frac34 E ) and along the RovumaRiver (11 szlig S 39 szlig E ) All G mulanjensis were recorded from 900ndash1000 m on MtMulanje (16 szlig 22frac34 S 35 szlig 07frac34 E ) in southern Malawi
Distance and cladistic analyses
The dendrograms and cladograms derived from the four diŒerent analyses areshown by gure 3 In essential details the inter-relationships among the species wereconsistent with their geographical distribution Each dendrogramcladogram showeda clear branching between east and southern African taxa and in each case there
A L V Davis et al1614
Tab
le3
Sum
mar
yof
know
nge
ogra
phic
aldi
stri
but
ion
and
habi
tat
asso
ciat
ion
sof
Gyro
notu
ssp
pbe
twee
nth
eE
aste
rnC
ape
Sout
hA
fric
aan
dno
rth
-eas
tern
Tan
zani
a
Len
gth
Geo
grap
hic
alra
nge
Alt
itu
din
alG
enu
san
dsp
ecie
s(m
m)dagger
(pat
chy
occu
rren
ce)
rang
e(m
)H
abit
atN
ote
s
Gyro
notu
sva
nL
ansb
erge
1874
South
Afr
ica
All
know
nre
cord
sin
fore
stG
pum
ilus
(Boh
eman
1857
)13
ndash15
Sout
h-ea
stco
asta
lhi
lls50
ndash450
For
est
469
of
128
info
rest
(gu
res
12
)G
ca
rinatu
sF
elsc
he19
1113
ndash15
Zul
ula
nd
coas
tal
hills
50ndash5
00F
ores
t5
7of
53in
fore
st(
gure
s1
2)
G
gla
bro
sus
Scho
ltz
and
Ho
wde
n19
8712
ndash12
5D
rak
ensb
erg
esca
rpm
ent
in78
0F
ores
t33
of
6in
fore
stN
ort
hern
Pro
vinc
e(
gure
s1
2)
Tro
pic
al
Afr
ica
Gm
ula
nje
nsi
sD
avis
Sch
olt
zan
dH
arri
son
118
ndash13
3M
tM
ulan
je
Mal
awi
(gu
re1
)90
0ndash10
00F
ores
tN
ot
reco
rded
F
ores
tan
d19
99op
enco
llect
ion
sm
ixed
DaggerG
Wm
etari
us
Ko
lbe
1894
125
ndash15
5M
ain
lyE
ast
Usa
mba
raM
ts50
ndash850
For
est
Fou
rtr
appe
dby
stre
amin
plu
sea
stco
ast
ofT
anza
nia
fore
stbu
tno
neou
tsid
e(
gure
1)
G
dis
par
(Fel
sche
1911
)16
ndash18
Sout
h-ea
stT
anza
nia
(gu
re1
)50
For
est
No
habi
tat
note
s
dagger Len
gth
wit
hhe
adde
exe
das
oppo
sed
tora
ised
asin
tabl
e5
Dagger C
L
Bel
lam
y(p
erso
nal
com
mun
icat
ion)
Cladistic and biogeographical analysis of Gyronotus 1615
Fig 3 Centre-rooted distance dendrograms and centre-rooted consensus parsimony clado-grams for inter-relationships between Gyronotus species (A B) Cladistic matrix derivedfrom double-standardized raw morphometric measurements by divergence coding(Thorpe 1984) (A) Distance dendrogram based on 27 characters (B) Parsimonycladogram based on 11 informative characters tree length 5 28 CI 5 0893 (C D)Cladistic matrix derived from double-standardized log10 transformed morphometricmeasurements by divergence coding (Thorpe 1984) (C ) Distance dendrogram basedon 27 characters (D) Parsimony cladogram based on 14 informative characters treelength 5 36 CI 5 0889 The numbers are the branch lengths from the nodes forunordered character states
was a similar resolution of relationships between South African species with thesouthernmost species G pumilus distanced from the two more northerly speciesHowever the results for the east African species indicated three diŒerent hypotheticalinter-relationships between species Of the three diŒerent patterns one each wasyielded from the matrices derived from double-standardized raw data whereas thethird was common to the dendrogram and cladogram yielded by the matrices derived
A L V Davis et al1616
from double-standardized log10 data These diŒerences may re ect the relativein uence of body size versus body shape according to the treatment of the datamatrices In the matrices derived from double-standardized raw data body size hasa stronger overall in uence than body shape (higher r2 and F numbers for regressionagainst Factor 1 which accounts for the greatest amount of variance in ordination(table 4) This emphasis is re ected by the closer linking of the two largest speciesG dispar and G Wmetarius Although both body size and body shape are de-emphasized in the matrices derived from double-standardized log10 data body shapehas the greater overall in uence (higher r2 and F numbers for regression againstboth Factors 1 and 2) This emphasis is re ected by the steps in common betweenthe elongate G dispar and G mulanjensis whereas the less elongate G Wmetarius isplaced on a separate branch ( gures 3C 3D)
Further phenetic analysis
As the cladograms were derived from morphometric data it is not surprisingthat the depicted interspeci c relationships are consistent with those indicated bythe three basic measurements of maximum length width and depth These measure-ments clearly separate the more elongate deeper east African species from the moreovoid atter southern African species (table 5) Ratios between the three dimen-sions indicate that width varies inversely with the other two dimensions whereaslength and depth vary proportionately to one another Thus the broadest species(G glabrosus) is also the attest and the most narrow species (G dispar) is alsothe deepest
Character states of aedeagi
F igure 4 illustrates the morphological diŒerences between the aedeagi ofGyronotus and those of large-bodied putative sister groups with primarily tropicaldistributions The parameres of tropical species in putative sister genera Anachalcos
procerus Gerstaecker and Canthodimorpha lawrencei Davis Scholtz and Harrisonare symmetrical and diverge at the tips Those of the east African Gyronotus alsodiverge at the tips but are asymmetrical due to a terminal process on the leftparamere The tips of the parameres of the southern African Gyronotus species aremore convergent and are also asymmetrical due to a terminal process projectingaway to the left of the left paramere
Discussion
Geographical polarization of characters
Comparison of Gyronotus aedeagi with those of the putative sister groupAnachalcos and that of Canthodimorpha suggests that bilateral symmetry anddivergence at the tips of the parameres are plesiomorphic character states ( gure 4)Although the aedeagi of the east African species of Gyronotus are characterized byasymmetry this is primarily due to the presence of a membranous terminal processwhich in ventral view projects to the right from the left paramere and occupies thespace between the divergent tips of the parameres ( gure 4) In contrast the SouthAfrican species are characterized by apomorphic convergence of the tips of theparameres ( gure 4mdashparameres slightly separated in the gure to facilitate drawing)In the two northernmost South African species asymmetry of the parameres ismainly due to the membranous terminal process which in ventral view projects
Cladistic and biogeographical analysis of Gyronotus 1617
Tab
le4
The
corr
elat
ion
ofG
yro
notu
sbo
dysi
ze(m
axim
umle
ngt
h)1
and
body
shap
e(r
atio
max
imu
mle
ngt
hm
axim
um
wid
thof
elyt
ra)2
wit
hor
din
ates
for
Fac
tors
1an
d2
deri
ved
fro
mpr
inci
pal
com
pon
ents
anal
ysis
ofm
orp
hom
etri
csda
tatr
eate
din
two
diŒ
eren
tw
ays
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
raw
mea
sure
men
tsra
wm
easu
rem
ents
log 10
mea
sure
men
tslo
g 10m
easu
rem
ents
(27
char
acte
rs)
(11
char
acte
rs)
(27
char
acte
rs)
(14
char
acte
rs)
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Pro
por
tion
ofov
eral
lvar
ianc
e(
)55
217
756
015
058
313
266
413
6R
2fo
rre
gres
sio
nof
body
size
on0
980
0002
092
000
20
060
230
080
24or
din
ates
1F
(15
1)fo
rre
gres
sio
nof
body
size
2870
60
0
0162
106
008
345
150
9
437
16
38
on
ordi
nat
es1
R2
for
regr
essi
on
ofbo
dysh
ape
on0
620
130
670
060
150
250
140
30or
din
ates
2F
(15
1)fo
rre
gres
sio
nof
body
shap
e83
55
7
55
103
79
3
358
63
173
1
873
22
26
on
ordi
nat
es2
p
lt0
05
p
lt0
02
p
lt0
001
A L V Davis et al1618
Tab
le5
Mor
pho
met
ric
mea
sure
men
tsfo
rsi
xsp
ecie
sof
Gyro
notu
s
Sout
hern
Afr
ican
spec
ies
Eas
tA
fric
ansp
ecie
sA
NO
VA
Mea
sure
men
tG
pum
ilus
G
cari
natu
sG
gla
bro
sus
G
dis
par
G
mula
nje
nsi
sG
Wm
etari
us
F(5
47)
Len
gth
(L)
147
2Ocirc0
64c
141
4Ocirc0
26cd
135
8Ocirc0
77d
206
4Ocirc0
41a
159
5Ocirc0
34b
162
0Ocirc0
26b
194
41
W
idth
(W)
905
Ocirc0
36b
900
Ocirc0
18b
944
Ocirc0
53b
107
4Ocirc0
42a
824
Ocirc0
22c
911
Ocirc0
46b
354
1
Dep
th(D
)6
02Ocirc
023
bd5
80Ocirc
021
cd5
57Ocirc
021
d8
24Ocirc
038
a6
09Ocirc
024
bc6
38Ocirc
027
b88
35
R
atio
sL
W0
62Ocirc
002
b0
64Ocirc
001
b0
70Ocirc
002
a0
52Ocirc
002
d0
52Ocirc
001
d0
56Ocirc
003
c87
16
L
D0
41Ocirc
001
a0
41Ocirc
001
a0
41Ocirc
002
a0
40Ocirc
002
ab0
38Ocirc
001
b0
39Ocirc
002
ab5
16
W
D0
67Ocirc
003
bc0
64Ocirc
002
c0
59Ocirc
002
d0
77Ocirc
004
ab0
74Ocirc
002
ab0
70Ocirc
003
b35
44
Sa
mp
lesi
ze10
106
710
10
Inea
chro
w
valu
esfo
llow
edby
adi
Œer
ent
lett
erdi
Œer
edsi
gni
cant
ly(T
ukey
rsquosH
SD
for
uneq
ual
nETH
plt
005
)
plt
000
1
Cladistic and biogeographical analysis of Gyronotus 1619
Fig 4 Distal and ventral views of canthonine aedeagi showing diŒerences in parameremorphology between (A) Anachalcos procerus Gerstaecker (distal ) (B)Canthodimorpha lawrencei Davis Scholtz and Harrison (ventral) (C ) east AfricanGyronotus van Lansberge (ventral ) (C1 G Wmetarius Kolbe C2 G mulanjensis DavisScholtz and Harrison C3 G dispar (Felsche)) and (D) South African Gyronotus(ventral ) (D1 G carinatus Felsche D2 G glabrosus Scholtz and Howden D3G pumilus (Boheman)) Scale bars 5 1 mm
away to the left of the left paramere However in the southernmost species theasymmetry is more extreme in that each paramere is radically diŒerent in shape(Davis et al 1999) The left paramere is more or less straight and bears the leftwardsaligned terminal process the right paramere is curved towards the mid-point andboth are quite unlike those of the other ve species in which each paramere issimilarly curved close to the distal end Thus there is clear biogeographical polariza-tion of the character states of the aedeagi with greater plesiomorphy in eastAfrican species
A L V Davis et al1620
Parallels between morphology biogeography and phenetic analysis of GyronotusThe interspeci c relationships indicated by the cladograms derived from phenetic
analysis largely corresponded to relationships suggested by morphological charactersof the aedeagi ( gure 4 Davis et al 1999) and these relationships were consistentwith the biogeographical distribution of the species along the south-eastern seaboardof Africa The clear separation of the elongate east African and more ovoid southernAfrican species is re ected by the diŒering position of the terminal process on theleft paramere of the aedeagi ( gure 4) The shape of this process in G Wmetarius
and in G mulanjensis diŒers only slightly in that the one is rounded and the otherangular Although it appears that these two species are very close the angularprocess of G dispar is su ciently similar that it is di cult to determine which ofthe cladograms if any re ects the true phylogenetic relationships between the eastAfrican species those strongly re ecting size and shape ( gures 3A 3B) or thosemore biased by shape ( gures 3C 3D) It is also possible that evolution of aedeagiand morphometric character states are divergent rather than parallel The relation-ships between the southern African species are resolved with greater consistencyThe shape of the parameres is similar in the two northerly species which diŒerprincipally only in the mono d shape of the outward projecting terminal process inG carinatus and the bi d shape of this process in G glabrosus ( gure 4) The moredistant phenetic separation of the southernmost species G pumilus is very stronglysupported by the characters of its aedeagus which diŒer radically from those of theother species However the outward projection of the terminal process indicates itscloser relationships to the other South African species than to the east Africanspecies
Biogeographical climatic and habitat associations of GyronotusSingle Gyronotus species are characteristic of several southern sub-centres of
endemism identi ed in mainland eastern coastal forests of Africa by Burgess et al(1998) These sub-centres occur in the East UsambaraTanga (G Wmetarius) Lindi(G dispar) and UdzungwaMulanje mountain regions (G mulanjensis) The coastalforests of southern Tanzania (Lindi subcentre) contain the most unique assemblageof endemic taxa along the east coast (Burgess et al 1998) and also harbour thelargest of the six known Gyronotus species (G dispar)
In terms of lowland versus afromontane forest association the biogeographicala nities of Gyronotus are dependent on the system used to classify oral zonationIn diŒerent regions altitudinal temperature zonation is modi ed by a combinationof factors including latitude rainfall regime aspect and the relative in uence ofmaritime versus continental climate (Hamilton 1989) Therefore instead of usinga single in exible classi cation system we have used three regional systems whichmake allowance for geographical diŒerences in the altitudinal occurrence of thesame temperature zones and their related ora These classi cation systems are thoseproposed by Lovett (1993) for the Eastern Arc Chapman and White (1970) forMalawi and Low and Rebelo (1996) for southern Africa Thus the East Africanspecies of Gyronotus may be described as elements of tropical lowland forests whichoccur below 1370 m in Malawi (Chapman and White 1970) and below 800m in theeastern arc (Lovett 1993) In the more southerly latitudes of South Africa mostlowland and highland forest is classi ed as afromontane (Low and Rebelo 1996)except for that on coastal sandveld of recent origin (Lawes 1990) AlthoughGyronotus have been recorded in coastal regions around East London (33 szlig 00frac34 S
Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

Cladistic and biogeographical analysis of Gyronotus 1609
institutions Specimens of four species were well-represented in these collections G
pumilus (Boheman) (1281 ) G carinatus Felsche (53 1 ) G Wmetarius Kolbe (41)and G mulanjensis Davis Scholtz and Harrison (33) However the institutionsheld only limited numbers of G dispar (Felsche) (7) and G glabrosus Scholtz andHowden (6) A seventh example of G glabrosus in the Howden Collection atCarleton University Ottawa Canada is the only other known museum specimenof this species During its known activity period in February 1998 unsuccessfulattempts were made to collect further material of this highly endemic species at itsone remaining known relatively undisturbed locality the afromontane Woodbushindigenous forest (23 szlig 52frac34 S 29 szlig 56frac34 E )
The morphology of Gyronotus showed relatively little variation and providedextremely few characters useful for cladistic analysis of interspeci c relationshipsIn such situations Chapill (1989) suggested that one alternative is to code morpho-metric data for cladistic analysis Since the use of phenetic characters for phylogeneticanalyses has been criticized (Wiley 1981) the results were compared with characterstates shown by the aedeagi These characters were the only morphological featureswhich showed diŒerences that could be interpreted with con dence as having strongphylogenetic signi cance
Twenty-seven morphometric characters used for distance or parsimony analysisof Gyronotus are listed in table 1 Measurements were made of all available specimensof G dispar (three males four females) and G glabrosus (four males two females)whereas ten specimens were measured for each of the other four species G WmetariusG pumilus G carinatus ( ve males ve females) and G mulanjensis (six malesfour females) The morphometric data matrix was treated in two diŒerent ways inorder to change the relative in uence of size and shape on the analyses Themeasurements (mm) were either left untransformed or were log10 transformed Bothtreatments were double standardized (Somers 1989) by subtracting each value fromthe row mean and each of these standardized values from the standardized columnmean For each treatment the data for each morphometric character was tested forsigni cant diŒerences between the six species using one-way ANOVA (table 1)Multiple range tests were conducted on each character using Tukeyrsquos HSD forunequal sample size (SpjotvollStoline Test ) On the basis of the results for thesemultiple range tests each character was coded for cladistic analysis using the diver-gence coding method of Thorpe (1984) This method provides a diŒerent code foreach signi cantly diŒerent character state and develops a species Ouml character statedistance matrix coded in categorical terms
Both distance and cladistic parsimony analyses were conducted on each of thetwo divergence matrices (table 2) using PHYLIP version 35 (Felsenstein 1993) andPAUP version 31 (SwoŒord 1993) Distance analyses were conducted using all 27characters in each matrix and species were linked by neighbour joining In contrastonly informative characters were used for cladistic parsimony analysis In the matrixderived from raw double-standardized data there were only 11 informativecharacters whereas in that derived from log10 double-standardized data there were14 informative characters In each analysis the character states were treated asunordered and the cladograms were centre-rooted
Each of the morphometric matrices was also subjected to principal componentsanalysis (PCA) As the rst component frequently correlates to size whereas follow-ing components correlate primarily to shape (Somers 1989 Fairbairn 1992) theordinate values for the rst two factors from each analysis were tested for correlation
A L V Davis et al1610
Table 1 List of morphometric characters for Gyronotus and the results of one-way ANOVAon interspeci c diŒerences between the characters either double-standardized rawmeasurements in mm or double-standardized log10 transformed measurements (seemethods)
F(5 47) F(5 47)(raw data)dagger ( log10 data)dagger
1 Distance between the tips of the clypeal teeth 17255 45972 Distance between the genal angles 6708 68373 Distance between the centre of the clypeal emargination and 10360 3422
a line drawn between the genal angles4 Distance between the centre of the clypeal emargination and 1384 1971
the posterior margin of the vertex5 Width between the anterior angles of the pronotal disc 4169 41236 Width between the lateral angles of the pronotal disc 3240 16667 Width between the posterior angles of the pronotal disc 493 20288 Length along the middle of the pronotal disc 4780 52659 Maximum depth of the pronotal disc between the dorsal 4695 3887
surface and the ventral surface of the profemora10 Depth of the pronotal disc between the dorsal surface and 1002 926
the pleural margin at the anterior end11 Depth of the pronotal disc between the dorsal surface and 1511 874
the pleural margin at the posterior end12 Distance from the centre of the posterior margin to a line 7113 2056
drawn between the posterior angles of the prothoraccic disc13 Length along the midline of the metasternum 665 105514 Distance between the left mesocoxa and left metacoxa 8648 43715 Distance between the right mesocoxa and the left metacoxa 1243 80316 Distance between the mesocoxae 1093 103517 Maximum depth from the elytra to the metasternum 3518 94618 Depth from the dorsal extremity of the left elytron to the 2037 889
pleural margin at the anterior end19 Maximum depth of the left elytron to the pleural margin 732 105820 Maximum height of the curvature along the lateral edge of 7638 1327
the left elytron above a line drawn between the anterior andposterior ends
21 Maximum width of the elytral pseudo-epipleurae 7702 280722 Maximum width of the right elytron Ouml 2 2496 388323 Width of the right elytron at the anterior end Ouml 2 1153 271824 Maximum length of the right elytron 5538 99225 Width of the pygidium along the basal ridge 1733 204226 Length of pygidium from the tip to the basal ridge 2326 227827 Length of the sternites along the midline of the abdomen 2709 1222
between the metasternum and the pygidium
daggerAll F numbers statistically signi cant plt0001
with both body length and the ratio of body length to maximum width of the elytraThese respective measurements were used as indices for body size and body shapeThe results provide some indication of the relative in uence of these two parameterson the four derived matrices used for distance and parsimony analyses
The relationships indicated by the cladograms are compared to characters shownby the aedeagi of Gyronotus those of Anachalcos Hope and that of Canthodimorpha
Davis Scholtz and Harrison which is a newly-discovered large-bodied monotypictropical genus from the northern coastline of Mozambique (Davis et al 1999)
Cladistic and biogeographical analysis of Gyronotus 1611
Tab
le2
Div
erge
nce
mat
rice
s(T
horp
e19
84)dagger
for
dist
ance
and
clad
isti
cpa
rsim
ony
anal
ysis
ofin
ter-
rela
tio
nshi
ps
betw
een
Gyro
notu
ssp
ecie
s
Ch
arac
ter
12
34
56
78
910
1112
1314
1516
1718
1920
2122
2324
2526
27
Der
ived
fro
mdo
uble
-sta
ndar
dize
dra
wph
enet
icm
easu
rem
ents
(mm
)C
har
in
form
ativ
eDaggeri
ii
ii
ii
ii
ii
G
pum
ilus
58
46
87
48
86
38
65
45
64
36
26
45
21
4G
ca
rinatu
s1
84
68
85
88
42
86
24
47
28
02
53
88
70
G
gla
bro
sus
18
46
87
58
84
28
63
44
71
21
42
39
97
4G
dis
par
a3
a7
20
40
09
80
5a
aa
08
6a
a1
30
37
9G
m
ula
nje
nsi
s5
34
42
57
32
37
26
44
46
78
66
a9
43
14
G
Wm
etari
us
80
41
23
53
44
84
16
43
48
36
66
84
57
9
Der
ived
fro
mdo
uble
-sta
ndar
dize
dlo
g 10tr
ansf
orm
edra
wm
easu
rem
ents
Ch
ar
info
rmat
iveDagger
ii
ii
ii
ii
ii
ii
ii
G
pum
ilus
57
46
75
38
88
43
68
76
64
47
34
35
32
5G
ca
rinatu
s1
75
77
a4
88
84
36
46
67
69
13
43
89
80
G
gla
bro
sus
17
87
74
38
85
34
65
66
71
32
30
28
98
6G
dis
par
97
96
75
a0
23
54
65
56
16
37
99
93
33
5G
m
ula
nje
nsi
s5
24
42
57
31
36
66
44
66
68
69
99
33
26
G
Wm
etari
us
90
00
01
33
33
8a
04
20
37
37
34
43
37
8
dagger Rec
oded
fro
ma
rang
eof
Otilde5
to5
toa
rang
efr
om
0to
10(1
0re
cod
edas
lsquoarsquo
wh
ich
isre
adas
one
unit
grea
ter
than
9in
the
dist
ance
anal
ysis
)Dagger A
llch
arac
ters
used
for
dist
ance
anal
ysis
on
lyin
form
ativ
ech
arac
ters
(i)
used
for
pars
imo
nyan
alys
is
A L V Davis et al1612
Although Anachalcos has been identi ed as the sister group (Scholtz and Howden1987) it is probably quite distant from Gyronotus which is both ightless and hasasymmetrical aedeagi However it is a useful comparison since the aedeagi ofAnachalcos are similar to those of most other afrotropical Canthonini in as muchas the parameres are symmetrical (Scholtz and Howden 1987) and lack the mem-branous terminal process on the left paramere which is characteristic of all Gyronotus
(Davis et al 1999)
Results
Geographical distribution
The six Gyronotus species showed either disjunct or parapatric distributions fromthe Eastern Cape in South Africa to north-eastern Tanzania ( gure 1) Two SouthAfrican species G pumilus (Eastern Cape southern KwaZulu-Natal ) and G
carinatus (northern KwaZulu-Natal ) showed a parapatric distribution pattern larg-ely restricted to coastal hills below 320m ( gure 2) The southern distribution limitsof G pumilus and the northern distribution limits of G carinatus coincide with thesouthern and northern limits of Walter and Liethrsquos (1964) temperatesubtropicalclimate type II (I )a ( gure 2) The respective northern and southern distribution
Fig 1 Geographical distribution of Gyronotus species from east to southern Africa(T Tanzania M Malawi SA South Africa) (S Swahili centre of endemism SMSwahili-Maputaland transition zone (Burgess et al 1998) Ma Maputaland centre ofendemism P Pondoland centre of endemism which occupies a narrow coastal zone(van Rensburg et al 1999))
Cladistic and biogeographical analysis of Gyronotus 1613
Fig 2 Geographical distribution of Gyronotus pumilus and G carinatus relative to altitudeand the subtropical coastal climate type II(I )a after Walter and Lieth (1964)
limits of G pumilus and G carinatus occur just north of Durban (29 szlig 51frac34 S 31 szlig 01frac34 E )at which latitude land greater than 1500 m in altitude swings away from the coastlineThe third southern African species was recorded only in a small localized region ofthe eastern escarpment of Northern Province in South Africa The combined distribu-tion of these species is sympatric with both coastal forest (particularly in the south)and the low-lying portion of forest classi ed as Afromontane (particularly in thenorth) (table 3) Distribution data for the east African species was geographicallymore limited ( gure 1) Most G Wmetarius were recorded from 215ndash850m in theEast Usambara Mountains of north-eastern Tanzania although a few specimenswere collected from the adjacent coastline The few G dispar were recorded fromsouth-eastern Tanzania around Lindi (10 szlig 00frac34 S 39 szlig 42frac34 E ) and along the RovumaRiver (11 szlig S 39 szlig E ) All G mulanjensis were recorded from 900ndash1000 m on MtMulanje (16 szlig 22frac34 S 35 szlig 07frac34 E ) in southern Malawi
Distance and cladistic analyses
The dendrograms and cladograms derived from the four diŒerent analyses areshown by gure 3 In essential details the inter-relationships among the species wereconsistent with their geographical distribution Each dendrogramcladogram showeda clear branching between east and southern African taxa and in each case there
A L V Davis et al1614
Tab
le3
Sum
mar
yof
know
nge
ogra
phic
aldi
stri
but
ion
and
habi
tat
asso
ciat
ion
sof
Gyro
notu
ssp
pbe
twee
nth
eE
aste
rnC
ape
Sout
hA
fric
aan
dno
rth
-eas
tern
Tan
zani
a
Len
gth
Geo
grap
hic
alra
nge
Alt
itu
din
alG
enu
san
dsp
ecie
s(m
m)dagger
(pat
chy
occu
rren
ce)
rang
e(m
)H
abit
atN
ote
s
Gyro
notu
sva
nL
ansb
erge
1874
South
Afr
ica
All
know
nre
cord
sin
fore
stG
pum
ilus
(Boh
eman
1857
)13
ndash15
Sout
h-ea
stco
asta
lhi
lls50
ndash450
For
est
469
of
128
info
rest
(gu
res
12
)G
ca
rinatu
sF
elsc
he19
1113
ndash15
Zul
ula
nd
coas
tal
hills
50ndash5
00F
ores
t5
7of
53in
fore
st(
gure
s1
2)
G
gla
bro
sus
Scho
ltz
and
Ho
wde
n19
8712
ndash12
5D
rak
ensb
erg
esca
rpm
ent
in78
0F
ores
t33
of
6in
fore
stN
ort
hern
Pro
vinc
e(
gure
s1
2)
Tro
pic
al
Afr
ica
Gm
ula
nje
nsi
sD
avis
Sch
olt
zan
dH
arri
son
118
ndash13
3M
tM
ulan
je
Mal
awi
(gu
re1
)90
0ndash10
00F
ores
tN
ot
reco
rded
F
ores
tan
d19
99op
enco
llect
ion
sm
ixed
DaggerG
Wm
etari
us
Ko
lbe
1894
125
ndash15
5M
ain
lyE
ast
Usa
mba
raM
ts50
ndash850
For
est
Fou
rtr
appe
dby
stre
amin
plu
sea
stco
ast
ofT
anza
nia
fore
stbu
tno
neou
tsid
e(
gure
1)
G
dis
par
(Fel
sche
1911
)16
ndash18
Sout
h-ea
stT
anza
nia
(gu
re1
)50
For
est
No
habi
tat
note
s
dagger Len
gth
wit
hhe
adde
exe
das
oppo
sed
tora
ised
asin
tabl
e5
Dagger C
L
Bel
lam
y(p
erso
nal
com
mun
icat
ion)
Cladistic and biogeographical analysis of Gyronotus 1615
Fig 3 Centre-rooted distance dendrograms and centre-rooted consensus parsimony clado-grams for inter-relationships between Gyronotus species (A B) Cladistic matrix derivedfrom double-standardized raw morphometric measurements by divergence coding(Thorpe 1984) (A) Distance dendrogram based on 27 characters (B) Parsimonycladogram based on 11 informative characters tree length 5 28 CI 5 0893 (C D)Cladistic matrix derived from double-standardized log10 transformed morphometricmeasurements by divergence coding (Thorpe 1984) (C ) Distance dendrogram basedon 27 characters (D) Parsimony cladogram based on 14 informative characters treelength 5 36 CI 5 0889 The numbers are the branch lengths from the nodes forunordered character states
was a similar resolution of relationships between South African species with thesouthernmost species G pumilus distanced from the two more northerly speciesHowever the results for the east African species indicated three diŒerent hypotheticalinter-relationships between species Of the three diŒerent patterns one each wasyielded from the matrices derived from double-standardized raw data whereas thethird was common to the dendrogram and cladogram yielded by the matrices derived
A L V Davis et al1616
from double-standardized log10 data These diŒerences may re ect the relativein uence of body size versus body shape according to the treatment of the datamatrices In the matrices derived from double-standardized raw data body size hasa stronger overall in uence than body shape (higher r2 and F numbers for regressionagainst Factor 1 which accounts for the greatest amount of variance in ordination(table 4) This emphasis is re ected by the closer linking of the two largest speciesG dispar and G Wmetarius Although both body size and body shape are de-emphasized in the matrices derived from double-standardized log10 data body shapehas the greater overall in uence (higher r2 and F numbers for regression againstboth Factors 1 and 2) This emphasis is re ected by the steps in common betweenthe elongate G dispar and G mulanjensis whereas the less elongate G Wmetarius isplaced on a separate branch ( gures 3C 3D)
Further phenetic analysis
As the cladograms were derived from morphometric data it is not surprisingthat the depicted interspeci c relationships are consistent with those indicated bythe three basic measurements of maximum length width and depth These measure-ments clearly separate the more elongate deeper east African species from the moreovoid atter southern African species (table 5) Ratios between the three dimen-sions indicate that width varies inversely with the other two dimensions whereaslength and depth vary proportionately to one another Thus the broadest species(G glabrosus) is also the attest and the most narrow species (G dispar) is alsothe deepest
Character states of aedeagi
F igure 4 illustrates the morphological diŒerences between the aedeagi ofGyronotus and those of large-bodied putative sister groups with primarily tropicaldistributions The parameres of tropical species in putative sister genera Anachalcos
procerus Gerstaecker and Canthodimorpha lawrencei Davis Scholtz and Harrisonare symmetrical and diverge at the tips Those of the east African Gyronotus alsodiverge at the tips but are asymmetrical due to a terminal process on the leftparamere The tips of the parameres of the southern African Gyronotus species aremore convergent and are also asymmetrical due to a terminal process projectingaway to the left of the left paramere
Discussion
Geographical polarization of characters
Comparison of Gyronotus aedeagi with those of the putative sister groupAnachalcos and that of Canthodimorpha suggests that bilateral symmetry anddivergence at the tips of the parameres are plesiomorphic character states ( gure 4)Although the aedeagi of the east African species of Gyronotus are characterized byasymmetry this is primarily due to the presence of a membranous terminal processwhich in ventral view projects to the right from the left paramere and occupies thespace between the divergent tips of the parameres ( gure 4) In contrast the SouthAfrican species are characterized by apomorphic convergence of the tips of theparameres ( gure 4mdashparameres slightly separated in the gure to facilitate drawing)In the two northernmost South African species asymmetry of the parameres ismainly due to the membranous terminal process which in ventral view projects
Cladistic and biogeographical analysis of Gyronotus 1617
Tab
le4
The
corr
elat
ion
ofG
yro
notu
sbo
dysi
ze(m
axim
umle
ngt
h)1
and
body
shap
e(r
atio
max
imu
mle
ngt
hm
axim
um
wid
thof
elyt
ra)2
wit
hor
din
ates
for
Fac
tors
1an
d2
deri
ved
fro
mpr
inci
pal
com
pon
ents
anal
ysis
ofm
orp
hom
etri
csda
tatr
eate
din
two
diŒ
eren
tw
ays
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
raw
mea
sure
men
tsra
wm
easu
rem
ents
log 10
mea
sure
men
tslo
g 10m
easu
rem
ents
(27
char
acte
rs)
(11
char
acte
rs)
(27
char
acte
rs)
(14
char
acte
rs)
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Pro
por
tion
ofov
eral
lvar
ianc
e(
)55
217
756
015
058
313
266
413
6R
2fo
rre
gres
sio
nof
body
size
on0
980
0002
092
000
20
060
230
080
24or
din
ates
1F
(15
1)fo
rre
gres
sio
nof
body
size
2870
60
0
0162
106
008
345
150
9
437
16
38
on
ordi
nat
es1
R2
for
regr
essi
on
ofbo
dysh
ape
on0
620
130
670
060
150
250
140
30or
din
ates
2F
(15
1)fo
rre
gres
sio
nof
body
shap
e83
55
7
55
103
79
3
358
63
173
1
873
22
26
on
ordi
nat
es2
p
lt0
05
p
lt0
02
p
lt0
001
A L V Davis et al1618
Tab
le5
Mor
pho
met
ric
mea
sure
men
tsfo
rsi
xsp
ecie
sof
Gyro
notu
s
Sout
hern
Afr
ican
spec
ies
Eas
tA
fric
ansp
ecie
sA
NO
VA
Mea
sure
men
tG
pum
ilus
G
cari
natu
sG
gla
bro
sus
G
dis
par
G
mula
nje
nsi
sG
Wm
etari
us
F(5
47)
Len
gth
(L)
147
2Ocirc0
64c
141
4Ocirc0
26cd
135
8Ocirc0
77d
206
4Ocirc0
41a
159
5Ocirc0
34b
162
0Ocirc0
26b
194
41
W
idth
(W)
905
Ocirc0
36b
900
Ocirc0
18b
944
Ocirc0
53b
107
4Ocirc0
42a
824
Ocirc0
22c
911
Ocirc0
46b
354
1
Dep
th(D
)6
02Ocirc
023
bd5
80Ocirc
021
cd5
57Ocirc
021
d8
24Ocirc
038
a6
09Ocirc
024
bc6
38Ocirc
027
b88
35
R
atio
sL
W0
62Ocirc
002
b0
64Ocirc
001
b0
70Ocirc
002
a0
52Ocirc
002
d0
52Ocirc
001
d0
56Ocirc
003
c87
16
L
D0
41Ocirc
001
a0
41Ocirc
001
a0
41Ocirc
002
a0
40Ocirc
002
ab0
38Ocirc
001
b0
39Ocirc
002
ab5
16
W
D0
67Ocirc
003
bc0
64Ocirc
002
c0
59Ocirc
002
d0
77Ocirc
004
ab0
74Ocirc
002
ab0
70Ocirc
003
b35
44
Sa
mp
lesi
ze10
106
710
10
Inea
chro
w
valu
esfo
llow
edby
adi
Œer
ent
lett
erdi
Œer
edsi
gni
cant
ly(T
ukey
rsquosH
SD
for
uneq
ual
nETH
plt
005
)
plt
000
1
Cladistic and biogeographical analysis of Gyronotus 1619
Fig 4 Distal and ventral views of canthonine aedeagi showing diŒerences in parameremorphology between (A) Anachalcos procerus Gerstaecker (distal ) (B)Canthodimorpha lawrencei Davis Scholtz and Harrison (ventral) (C ) east AfricanGyronotus van Lansberge (ventral ) (C1 G Wmetarius Kolbe C2 G mulanjensis DavisScholtz and Harrison C3 G dispar (Felsche)) and (D) South African Gyronotus(ventral ) (D1 G carinatus Felsche D2 G glabrosus Scholtz and Howden D3G pumilus (Boheman)) Scale bars 5 1 mm
away to the left of the left paramere However in the southernmost species theasymmetry is more extreme in that each paramere is radically diŒerent in shape(Davis et al 1999) The left paramere is more or less straight and bears the leftwardsaligned terminal process the right paramere is curved towards the mid-point andboth are quite unlike those of the other ve species in which each paramere issimilarly curved close to the distal end Thus there is clear biogeographical polariza-tion of the character states of the aedeagi with greater plesiomorphy in eastAfrican species
A L V Davis et al1620
Parallels between morphology biogeography and phenetic analysis of GyronotusThe interspeci c relationships indicated by the cladograms derived from phenetic
analysis largely corresponded to relationships suggested by morphological charactersof the aedeagi ( gure 4 Davis et al 1999) and these relationships were consistentwith the biogeographical distribution of the species along the south-eastern seaboardof Africa The clear separation of the elongate east African and more ovoid southernAfrican species is re ected by the diŒering position of the terminal process on theleft paramere of the aedeagi ( gure 4) The shape of this process in G Wmetarius
and in G mulanjensis diŒers only slightly in that the one is rounded and the otherangular Although it appears that these two species are very close the angularprocess of G dispar is su ciently similar that it is di cult to determine which ofthe cladograms if any re ects the true phylogenetic relationships between the eastAfrican species those strongly re ecting size and shape ( gures 3A 3B) or thosemore biased by shape ( gures 3C 3D) It is also possible that evolution of aedeagiand morphometric character states are divergent rather than parallel The relation-ships between the southern African species are resolved with greater consistencyThe shape of the parameres is similar in the two northerly species which diŒerprincipally only in the mono d shape of the outward projecting terminal process inG carinatus and the bi d shape of this process in G glabrosus ( gure 4) The moredistant phenetic separation of the southernmost species G pumilus is very stronglysupported by the characters of its aedeagus which diŒer radically from those of theother species However the outward projection of the terminal process indicates itscloser relationships to the other South African species than to the east Africanspecies
Biogeographical climatic and habitat associations of GyronotusSingle Gyronotus species are characteristic of several southern sub-centres of
endemism identi ed in mainland eastern coastal forests of Africa by Burgess et al(1998) These sub-centres occur in the East UsambaraTanga (G Wmetarius) Lindi(G dispar) and UdzungwaMulanje mountain regions (G mulanjensis) The coastalforests of southern Tanzania (Lindi subcentre) contain the most unique assemblageof endemic taxa along the east coast (Burgess et al 1998) and also harbour thelargest of the six known Gyronotus species (G dispar)
In terms of lowland versus afromontane forest association the biogeographicala nities of Gyronotus are dependent on the system used to classify oral zonationIn diŒerent regions altitudinal temperature zonation is modi ed by a combinationof factors including latitude rainfall regime aspect and the relative in uence ofmaritime versus continental climate (Hamilton 1989) Therefore instead of usinga single in exible classi cation system we have used three regional systems whichmake allowance for geographical diŒerences in the altitudinal occurrence of thesame temperature zones and their related ora These classi cation systems are thoseproposed by Lovett (1993) for the Eastern Arc Chapman and White (1970) forMalawi and Low and Rebelo (1996) for southern Africa Thus the East Africanspecies of Gyronotus may be described as elements of tropical lowland forests whichoccur below 1370 m in Malawi (Chapman and White 1970) and below 800m in theeastern arc (Lovett 1993) In the more southerly latitudes of South Africa mostlowland and highland forest is classi ed as afromontane (Low and Rebelo 1996)except for that on coastal sandveld of recent origin (Lawes 1990) AlthoughGyronotus have been recorded in coastal regions around East London (33 szlig 00frac34 S
Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

A L V Davis et al1610
Table 1 List of morphometric characters for Gyronotus and the results of one-way ANOVAon interspeci c diŒerences between the characters either double-standardized rawmeasurements in mm or double-standardized log10 transformed measurements (seemethods)
F(5 47) F(5 47)(raw data)dagger ( log10 data)dagger
1 Distance between the tips of the clypeal teeth 17255 45972 Distance between the genal angles 6708 68373 Distance between the centre of the clypeal emargination and 10360 3422
a line drawn between the genal angles4 Distance between the centre of the clypeal emargination and 1384 1971
the posterior margin of the vertex5 Width between the anterior angles of the pronotal disc 4169 41236 Width between the lateral angles of the pronotal disc 3240 16667 Width between the posterior angles of the pronotal disc 493 20288 Length along the middle of the pronotal disc 4780 52659 Maximum depth of the pronotal disc between the dorsal 4695 3887
surface and the ventral surface of the profemora10 Depth of the pronotal disc between the dorsal surface and 1002 926
the pleural margin at the anterior end11 Depth of the pronotal disc between the dorsal surface and 1511 874
the pleural margin at the posterior end12 Distance from the centre of the posterior margin to a line 7113 2056
drawn between the posterior angles of the prothoraccic disc13 Length along the midline of the metasternum 665 105514 Distance between the left mesocoxa and left metacoxa 8648 43715 Distance between the right mesocoxa and the left metacoxa 1243 80316 Distance between the mesocoxae 1093 103517 Maximum depth from the elytra to the metasternum 3518 94618 Depth from the dorsal extremity of the left elytron to the 2037 889
pleural margin at the anterior end19 Maximum depth of the left elytron to the pleural margin 732 105820 Maximum height of the curvature along the lateral edge of 7638 1327
the left elytron above a line drawn between the anterior andposterior ends
21 Maximum width of the elytral pseudo-epipleurae 7702 280722 Maximum width of the right elytron Ouml 2 2496 388323 Width of the right elytron at the anterior end Ouml 2 1153 271824 Maximum length of the right elytron 5538 99225 Width of the pygidium along the basal ridge 1733 204226 Length of pygidium from the tip to the basal ridge 2326 227827 Length of the sternites along the midline of the abdomen 2709 1222
between the metasternum and the pygidium
daggerAll F numbers statistically signi cant plt0001
with both body length and the ratio of body length to maximum width of the elytraThese respective measurements were used as indices for body size and body shapeThe results provide some indication of the relative in uence of these two parameterson the four derived matrices used for distance and parsimony analyses
The relationships indicated by the cladograms are compared to characters shownby the aedeagi of Gyronotus those of Anachalcos Hope and that of Canthodimorpha
Davis Scholtz and Harrison which is a newly-discovered large-bodied monotypictropical genus from the northern coastline of Mozambique (Davis et al 1999)
Cladistic and biogeographical analysis of Gyronotus 1611
Tab
le2
Div
erge
nce
mat
rice
s(T
horp
e19
84)dagger
for
dist
ance
and
clad
isti
cpa
rsim
ony
anal
ysis
ofin
ter-
rela
tio
nshi
ps
betw
een
Gyro
notu
ssp
ecie
s
Ch
arac
ter
12
34
56
78
910
1112
1314
1516
1718
1920
2122
2324
2526
27
Der
ived
fro
mdo
uble
-sta
ndar
dize
dra
wph
enet
icm
easu
rem
ents
(mm
)C
har
in
form
ativ
eDaggeri
ii
ii
ii
ii
ii
G
pum
ilus
58
46
87
48
86
38
65
45
64
36
26
45
21
4G
ca
rinatu
s1
84
68
85
88
42
86
24
47
28
02
53
88
70
G
gla
bro
sus
18
46
87
58
84
28
63
44
71
21
42
39
97
4G
dis
par
a3
a7
20
40
09
80
5a
aa
08
6a
a1
30
37
9G
m
ula
nje
nsi
s5
34
42
57
32
37
26
44
46
78
66
a9
43
14
G
Wm
etari
us
80
41
23
53
44
84
16
43
48
36
66
84
57
9
Der
ived
fro
mdo
uble
-sta
ndar
dize
dlo
g 10tr
ansf
orm
edra
wm
easu
rem
ents
Ch
ar
info
rmat
iveDagger
ii
ii
ii
ii
ii
ii
ii
G
pum
ilus
57
46
75
38
88
43
68
76
64
47
34
35
32
5G
ca
rinatu
s1
75
77
a4
88
84
36
46
67
69
13
43
89
80
G
gla
bro
sus
17
87
74
38
85
34
65
66
71
32
30
28
98
6G
dis
par
97
96
75
a0
23
54
65
56
16
37
99
93
33
5G
m
ula
nje
nsi
s5
24
42
57
31
36
66
44
66
68
69
99
33
26
G
Wm
etari
us
90
00
01
33
33
8a
04
20
37
37
34
43
37
8
dagger Rec
oded
fro
ma
rang
eof
Otilde5
to5
toa
rang
efr
om
0to
10(1
0re
cod
edas
lsquoarsquo
wh
ich
isre
adas
one
unit
grea
ter
than
9in
the
dist
ance
anal
ysis
)Dagger A
llch
arac
ters
used
for
dist
ance
anal
ysis
on
lyin
form
ativ
ech
arac
ters
(i)
used
for
pars
imo
nyan
alys
is
A L V Davis et al1612
Although Anachalcos has been identi ed as the sister group (Scholtz and Howden1987) it is probably quite distant from Gyronotus which is both ightless and hasasymmetrical aedeagi However it is a useful comparison since the aedeagi ofAnachalcos are similar to those of most other afrotropical Canthonini in as muchas the parameres are symmetrical (Scholtz and Howden 1987) and lack the mem-branous terminal process on the left paramere which is characteristic of all Gyronotus
(Davis et al 1999)
Results
Geographical distribution
The six Gyronotus species showed either disjunct or parapatric distributions fromthe Eastern Cape in South Africa to north-eastern Tanzania ( gure 1) Two SouthAfrican species G pumilus (Eastern Cape southern KwaZulu-Natal ) and G
carinatus (northern KwaZulu-Natal ) showed a parapatric distribution pattern larg-ely restricted to coastal hills below 320m ( gure 2) The southern distribution limitsof G pumilus and the northern distribution limits of G carinatus coincide with thesouthern and northern limits of Walter and Liethrsquos (1964) temperatesubtropicalclimate type II (I )a ( gure 2) The respective northern and southern distribution
Fig 1 Geographical distribution of Gyronotus species from east to southern Africa(T Tanzania M Malawi SA South Africa) (S Swahili centre of endemism SMSwahili-Maputaland transition zone (Burgess et al 1998) Ma Maputaland centre ofendemism P Pondoland centre of endemism which occupies a narrow coastal zone(van Rensburg et al 1999))
Cladistic and biogeographical analysis of Gyronotus 1613
Fig 2 Geographical distribution of Gyronotus pumilus and G carinatus relative to altitudeand the subtropical coastal climate type II(I )a after Walter and Lieth (1964)
limits of G pumilus and G carinatus occur just north of Durban (29 szlig 51frac34 S 31 szlig 01frac34 E )at which latitude land greater than 1500 m in altitude swings away from the coastlineThe third southern African species was recorded only in a small localized region ofthe eastern escarpment of Northern Province in South Africa The combined distribu-tion of these species is sympatric with both coastal forest (particularly in the south)and the low-lying portion of forest classi ed as Afromontane (particularly in thenorth) (table 3) Distribution data for the east African species was geographicallymore limited ( gure 1) Most G Wmetarius were recorded from 215ndash850m in theEast Usambara Mountains of north-eastern Tanzania although a few specimenswere collected from the adjacent coastline The few G dispar were recorded fromsouth-eastern Tanzania around Lindi (10 szlig 00frac34 S 39 szlig 42frac34 E ) and along the RovumaRiver (11 szlig S 39 szlig E ) All G mulanjensis were recorded from 900ndash1000 m on MtMulanje (16 szlig 22frac34 S 35 szlig 07frac34 E ) in southern Malawi
Distance and cladistic analyses
The dendrograms and cladograms derived from the four diŒerent analyses areshown by gure 3 In essential details the inter-relationships among the species wereconsistent with their geographical distribution Each dendrogramcladogram showeda clear branching between east and southern African taxa and in each case there
A L V Davis et al1614
Tab
le3
Sum
mar
yof
know
nge
ogra
phic
aldi
stri
but
ion
and
habi
tat
asso
ciat
ion
sof
Gyro
notu
ssp
pbe
twee
nth
eE
aste
rnC
ape
Sout
hA
fric
aan
dno
rth
-eas
tern
Tan
zani
a
Len
gth
Geo
grap
hic
alra
nge
Alt
itu
din
alG
enu
san
dsp
ecie
s(m
m)dagger
(pat
chy
occu
rren
ce)
rang
e(m
)H
abit
atN
ote
s
Gyro
notu
sva
nL
ansb
erge
1874
South
Afr
ica
All
know
nre
cord
sin
fore
stG
pum
ilus
(Boh
eman
1857
)13
ndash15
Sout
h-ea
stco
asta
lhi
lls50
ndash450
For
est
469
of
128
info
rest
(gu
res
12
)G
ca
rinatu
sF
elsc
he19
1113
ndash15
Zul
ula
nd
coas
tal
hills
50ndash5
00F
ores
t5
7of
53in
fore
st(
gure
s1
2)
G
gla
bro
sus
Scho
ltz
and
Ho
wde
n19
8712
ndash12
5D
rak
ensb
erg
esca
rpm
ent
in78
0F
ores
t33
of
6in
fore
stN
ort
hern
Pro
vinc
e(
gure
s1
2)
Tro
pic
al
Afr
ica
Gm
ula
nje
nsi
sD
avis
Sch
olt
zan
dH
arri
son
118
ndash13
3M
tM
ulan
je
Mal
awi
(gu
re1
)90
0ndash10
00F
ores
tN
ot
reco
rded
F
ores
tan
d19
99op
enco
llect
ion
sm
ixed
DaggerG
Wm
etari
us
Ko
lbe
1894
125
ndash15
5M
ain
lyE
ast
Usa
mba
raM
ts50
ndash850
For
est
Fou
rtr
appe
dby
stre
amin
plu
sea
stco
ast
ofT
anza
nia
fore
stbu
tno
neou
tsid
e(
gure
1)
G
dis
par
(Fel
sche
1911
)16
ndash18
Sout
h-ea
stT
anza
nia
(gu
re1
)50
For
est
No
habi
tat
note
s
dagger Len
gth
wit
hhe
adde
exe
das
oppo
sed
tora
ised
asin
tabl
e5
Dagger C
L
Bel
lam
y(p
erso
nal
com
mun
icat
ion)
Cladistic and biogeographical analysis of Gyronotus 1615
Fig 3 Centre-rooted distance dendrograms and centre-rooted consensus parsimony clado-grams for inter-relationships between Gyronotus species (A B) Cladistic matrix derivedfrom double-standardized raw morphometric measurements by divergence coding(Thorpe 1984) (A) Distance dendrogram based on 27 characters (B) Parsimonycladogram based on 11 informative characters tree length 5 28 CI 5 0893 (C D)Cladistic matrix derived from double-standardized log10 transformed morphometricmeasurements by divergence coding (Thorpe 1984) (C ) Distance dendrogram basedon 27 characters (D) Parsimony cladogram based on 14 informative characters treelength 5 36 CI 5 0889 The numbers are the branch lengths from the nodes forunordered character states
was a similar resolution of relationships between South African species with thesouthernmost species G pumilus distanced from the two more northerly speciesHowever the results for the east African species indicated three diŒerent hypotheticalinter-relationships between species Of the three diŒerent patterns one each wasyielded from the matrices derived from double-standardized raw data whereas thethird was common to the dendrogram and cladogram yielded by the matrices derived
A L V Davis et al1616
from double-standardized log10 data These diŒerences may re ect the relativein uence of body size versus body shape according to the treatment of the datamatrices In the matrices derived from double-standardized raw data body size hasa stronger overall in uence than body shape (higher r2 and F numbers for regressionagainst Factor 1 which accounts for the greatest amount of variance in ordination(table 4) This emphasis is re ected by the closer linking of the two largest speciesG dispar and G Wmetarius Although both body size and body shape are de-emphasized in the matrices derived from double-standardized log10 data body shapehas the greater overall in uence (higher r2 and F numbers for regression againstboth Factors 1 and 2) This emphasis is re ected by the steps in common betweenthe elongate G dispar and G mulanjensis whereas the less elongate G Wmetarius isplaced on a separate branch ( gures 3C 3D)
Further phenetic analysis
As the cladograms were derived from morphometric data it is not surprisingthat the depicted interspeci c relationships are consistent with those indicated bythe three basic measurements of maximum length width and depth These measure-ments clearly separate the more elongate deeper east African species from the moreovoid atter southern African species (table 5) Ratios between the three dimen-sions indicate that width varies inversely with the other two dimensions whereaslength and depth vary proportionately to one another Thus the broadest species(G glabrosus) is also the attest and the most narrow species (G dispar) is alsothe deepest
Character states of aedeagi
F igure 4 illustrates the morphological diŒerences between the aedeagi ofGyronotus and those of large-bodied putative sister groups with primarily tropicaldistributions The parameres of tropical species in putative sister genera Anachalcos
procerus Gerstaecker and Canthodimorpha lawrencei Davis Scholtz and Harrisonare symmetrical and diverge at the tips Those of the east African Gyronotus alsodiverge at the tips but are asymmetrical due to a terminal process on the leftparamere The tips of the parameres of the southern African Gyronotus species aremore convergent and are also asymmetrical due to a terminal process projectingaway to the left of the left paramere
Discussion
Geographical polarization of characters
Comparison of Gyronotus aedeagi with those of the putative sister groupAnachalcos and that of Canthodimorpha suggests that bilateral symmetry anddivergence at the tips of the parameres are plesiomorphic character states ( gure 4)Although the aedeagi of the east African species of Gyronotus are characterized byasymmetry this is primarily due to the presence of a membranous terminal processwhich in ventral view projects to the right from the left paramere and occupies thespace between the divergent tips of the parameres ( gure 4) In contrast the SouthAfrican species are characterized by apomorphic convergence of the tips of theparameres ( gure 4mdashparameres slightly separated in the gure to facilitate drawing)In the two northernmost South African species asymmetry of the parameres ismainly due to the membranous terminal process which in ventral view projects
Cladistic and biogeographical analysis of Gyronotus 1617
Tab
le4
The
corr
elat
ion
ofG
yro
notu
sbo
dysi
ze(m
axim
umle
ngt
h)1
and
body
shap
e(r
atio
max
imu
mle
ngt
hm
axim
um
wid
thof
elyt
ra)2
wit
hor
din
ates
for
Fac
tors
1an
d2
deri
ved
fro
mpr
inci
pal
com
pon
ents
anal
ysis
ofm
orp
hom
etri
csda
tatr
eate
din
two
diŒ
eren
tw
ays
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
raw
mea
sure
men
tsra
wm
easu
rem
ents
log 10
mea
sure
men
tslo
g 10m
easu
rem
ents
(27
char
acte
rs)
(11
char
acte
rs)
(27
char
acte
rs)
(14
char
acte
rs)
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Pro
por
tion
ofov
eral
lvar
ianc
e(
)55
217
756
015
058
313
266
413
6R
2fo
rre
gres
sio
nof
body
size
on0
980
0002
092
000
20
060
230
080
24or
din
ates
1F
(15
1)fo
rre
gres
sio
nof
body
size
2870
60
0
0162
106
008
345
150
9
437
16
38
on
ordi
nat
es1
R2
for
regr
essi
on
ofbo
dysh
ape
on0
620
130
670
060
150
250
140
30or
din
ates
2F
(15
1)fo
rre
gres
sio
nof
body
shap
e83
55
7
55
103
79
3
358
63
173
1
873
22
26
on
ordi
nat
es2
p
lt0
05
p
lt0
02
p
lt0
001
A L V Davis et al1618
Tab
le5
Mor
pho
met
ric
mea
sure
men
tsfo
rsi
xsp
ecie
sof
Gyro
notu
s
Sout
hern
Afr
ican
spec
ies
Eas
tA
fric
ansp
ecie
sA
NO
VA
Mea
sure
men
tG
pum
ilus
G
cari
natu
sG
gla
bro
sus
G
dis
par
G
mula
nje
nsi
sG
Wm
etari
us
F(5
47)
Len
gth
(L)
147
2Ocirc0
64c
141
4Ocirc0
26cd
135
8Ocirc0
77d
206
4Ocirc0
41a
159
5Ocirc0
34b
162
0Ocirc0
26b
194
41
W
idth
(W)
905
Ocirc0
36b
900
Ocirc0
18b
944
Ocirc0
53b
107
4Ocirc0
42a
824
Ocirc0
22c
911
Ocirc0
46b
354
1
Dep
th(D
)6
02Ocirc
023
bd5
80Ocirc
021
cd5
57Ocirc
021
d8
24Ocirc
038
a6
09Ocirc
024
bc6
38Ocirc
027
b88
35
R
atio
sL
W0
62Ocirc
002
b0
64Ocirc
001
b0
70Ocirc
002
a0
52Ocirc
002
d0
52Ocirc
001
d0
56Ocirc
003
c87
16
L
D0
41Ocirc
001
a0
41Ocirc
001
a0
41Ocirc
002
a0
40Ocirc
002
ab0
38Ocirc
001
b0
39Ocirc
002
ab5
16
W
D0
67Ocirc
003
bc0
64Ocirc
002
c0
59Ocirc
002
d0
77Ocirc
004
ab0
74Ocirc
002
ab0
70Ocirc
003
b35
44
Sa
mp
lesi
ze10
106
710
10
Inea
chro
w
valu
esfo
llow
edby
adi
Œer
ent
lett
erdi
Œer
edsi
gni
cant
ly(T
ukey
rsquosH
SD
for
uneq
ual
nETH
plt
005
)
plt
000
1
Cladistic and biogeographical analysis of Gyronotus 1619
Fig 4 Distal and ventral views of canthonine aedeagi showing diŒerences in parameremorphology between (A) Anachalcos procerus Gerstaecker (distal ) (B)Canthodimorpha lawrencei Davis Scholtz and Harrison (ventral) (C ) east AfricanGyronotus van Lansberge (ventral ) (C1 G Wmetarius Kolbe C2 G mulanjensis DavisScholtz and Harrison C3 G dispar (Felsche)) and (D) South African Gyronotus(ventral ) (D1 G carinatus Felsche D2 G glabrosus Scholtz and Howden D3G pumilus (Boheman)) Scale bars 5 1 mm
away to the left of the left paramere However in the southernmost species theasymmetry is more extreme in that each paramere is radically diŒerent in shape(Davis et al 1999) The left paramere is more or less straight and bears the leftwardsaligned terminal process the right paramere is curved towards the mid-point andboth are quite unlike those of the other ve species in which each paramere issimilarly curved close to the distal end Thus there is clear biogeographical polariza-tion of the character states of the aedeagi with greater plesiomorphy in eastAfrican species
A L V Davis et al1620
Parallels between morphology biogeography and phenetic analysis of GyronotusThe interspeci c relationships indicated by the cladograms derived from phenetic
analysis largely corresponded to relationships suggested by morphological charactersof the aedeagi ( gure 4 Davis et al 1999) and these relationships were consistentwith the biogeographical distribution of the species along the south-eastern seaboardof Africa The clear separation of the elongate east African and more ovoid southernAfrican species is re ected by the diŒering position of the terminal process on theleft paramere of the aedeagi ( gure 4) The shape of this process in G Wmetarius
and in G mulanjensis diŒers only slightly in that the one is rounded and the otherangular Although it appears that these two species are very close the angularprocess of G dispar is su ciently similar that it is di cult to determine which ofthe cladograms if any re ects the true phylogenetic relationships between the eastAfrican species those strongly re ecting size and shape ( gures 3A 3B) or thosemore biased by shape ( gures 3C 3D) It is also possible that evolution of aedeagiand morphometric character states are divergent rather than parallel The relation-ships between the southern African species are resolved with greater consistencyThe shape of the parameres is similar in the two northerly species which diŒerprincipally only in the mono d shape of the outward projecting terminal process inG carinatus and the bi d shape of this process in G glabrosus ( gure 4) The moredistant phenetic separation of the southernmost species G pumilus is very stronglysupported by the characters of its aedeagus which diŒer radically from those of theother species However the outward projection of the terminal process indicates itscloser relationships to the other South African species than to the east Africanspecies
Biogeographical climatic and habitat associations of GyronotusSingle Gyronotus species are characteristic of several southern sub-centres of
endemism identi ed in mainland eastern coastal forests of Africa by Burgess et al(1998) These sub-centres occur in the East UsambaraTanga (G Wmetarius) Lindi(G dispar) and UdzungwaMulanje mountain regions (G mulanjensis) The coastalforests of southern Tanzania (Lindi subcentre) contain the most unique assemblageof endemic taxa along the east coast (Burgess et al 1998) and also harbour thelargest of the six known Gyronotus species (G dispar)
In terms of lowland versus afromontane forest association the biogeographicala nities of Gyronotus are dependent on the system used to classify oral zonationIn diŒerent regions altitudinal temperature zonation is modi ed by a combinationof factors including latitude rainfall regime aspect and the relative in uence ofmaritime versus continental climate (Hamilton 1989) Therefore instead of usinga single in exible classi cation system we have used three regional systems whichmake allowance for geographical diŒerences in the altitudinal occurrence of thesame temperature zones and their related ora These classi cation systems are thoseproposed by Lovett (1993) for the Eastern Arc Chapman and White (1970) forMalawi and Low and Rebelo (1996) for southern Africa Thus the East Africanspecies of Gyronotus may be described as elements of tropical lowland forests whichoccur below 1370 m in Malawi (Chapman and White 1970) and below 800m in theeastern arc (Lovett 1993) In the more southerly latitudes of South Africa mostlowland and highland forest is classi ed as afromontane (Low and Rebelo 1996)except for that on coastal sandveld of recent origin (Lawes 1990) AlthoughGyronotus have been recorded in coastal regions around East London (33 szlig 00frac34 S
Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

Cladistic and biogeographical analysis of Gyronotus 1611
Tab
le2
Div
erge
nce
mat
rice
s(T
horp
e19
84)dagger
for
dist
ance
and
clad
isti
cpa
rsim
ony
anal
ysis
ofin
ter-
rela
tio
nshi
ps
betw
een
Gyro
notu
ssp
ecie
s
Ch
arac
ter
12
34
56
78
910
1112
1314
1516
1718
1920
2122
2324
2526
27
Der
ived
fro
mdo
uble
-sta
ndar
dize
dra
wph
enet
icm
easu
rem
ents
(mm
)C
har
in
form
ativ
eDaggeri
ii
ii
ii
ii
ii
G
pum
ilus
58
46
87
48
86
38
65
45
64
36
26
45
21
4G
ca
rinatu
s1
84
68
85
88
42
86
24
47
28
02
53
88
70
G
gla
bro
sus
18
46
87
58
84
28
63
44
71
21
42
39
97
4G
dis
par
a3
a7
20
40
09
80
5a
aa
08
6a
a1
30
37
9G
m
ula
nje
nsi
s5
34
42
57
32
37
26
44
46
78
66
a9
43
14
G
Wm
etari
us
80
41
23
53
44
84
16
43
48
36
66
84
57
9
Der
ived
fro
mdo
uble
-sta
ndar
dize
dlo
g 10tr
ansf
orm
edra
wm
easu
rem
ents
Ch
ar
info
rmat
iveDagger
ii
ii
ii
ii
ii
ii
ii
G
pum
ilus
57
46
75
38
88
43
68
76
64
47
34
35
32
5G
ca
rinatu
s1
75
77
a4
88
84
36
46
67
69
13
43
89
80
G
gla
bro
sus
17
87
74
38
85
34
65
66
71
32
30
28
98
6G
dis
par
97
96
75
a0
23
54
65
56
16
37
99
93
33
5G
m
ula
nje
nsi
s5
24
42
57
31
36
66
44
66
68
69
99
33
26
G
Wm
etari
us
90
00
01
33
33
8a
04
20
37
37
34
43
37
8
dagger Rec
oded
fro
ma
rang
eof
Otilde5
to5
toa
rang
efr
om
0to
10(1
0re
cod
edas
lsquoarsquo
wh
ich
isre
adas
one
unit
grea
ter
than
9in
the
dist
ance
anal
ysis
)Dagger A
llch
arac
ters
used
for
dist
ance
anal
ysis
on
lyin
form
ativ
ech
arac
ters
(i)
used
for
pars
imo
nyan
alys
is
A L V Davis et al1612
Although Anachalcos has been identi ed as the sister group (Scholtz and Howden1987) it is probably quite distant from Gyronotus which is both ightless and hasasymmetrical aedeagi However it is a useful comparison since the aedeagi ofAnachalcos are similar to those of most other afrotropical Canthonini in as muchas the parameres are symmetrical (Scholtz and Howden 1987) and lack the mem-branous terminal process on the left paramere which is characteristic of all Gyronotus
(Davis et al 1999)
Results
Geographical distribution
The six Gyronotus species showed either disjunct or parapatric distributions fromthe Eastern Cape in South Africa to north-eastern Tanzania ( gure 1) Two SouthAfrican species G pumilus (Eastern Cape southern KwaZulu-Natal ) and G
carinatus (northern KwaZulu-Natal ) showed a parapatric distribution pattern larg-ely restricted to coastal hills below 320m ( gure 2) The southern distribution limitsof G pumilus and the northern distribution limits of G carinatus coincide with thesouthern and northern limits of Walter and Liethrsquos (1964) temperatesubtropicalclimate type II (I )a ( gure 2) The respective northern and southern distribution
Fig 1 Geographical distribution of Gyronotus species from east to southern Africa(T Tanzania M Malawi SA South Africa) (S Swahili centre of endemism SMSwahili-Maputaland transition zone (Burgess et al 1998) Ma Maputaland centre ofendemism P Pondoland centre of endemism which occupies a narrow coastal zone(van Rensburg et al 1999))
Cladistic and biogeographical analysis of Gyronotus 1613
Fig 2 Geographical distribution of Gyronotus pumilus and G carinatus relative to altitudeand the subtropical coastal climate type II(I )a after Walter and Lieth (1964)
limits of G pumilus and G carinatus occur just north of Durban (29 szlig 51frac34 S 31 szlig 01frac34 E )at which latitude land greater than 1500 m in altitude swings away from the coastlineThe third southern African species was recorded only in a small localized region ofthe eastern escarpment of Northern Province in South Africa The combined distribu-tion of these species is sympatric with both coastal forest (particularly in the south)and the low-lying portion of forest classi ed as Afromontane (particularly in thenorth) (table 3) Distribution data for the east African species was geographicallymore limited ( gure 1) Most G Wmetarius were recorded from 215ndash850m in theEast Usambara Mountains of north-eastern Tanzania although a few specimenswere collected from the adjacent coastline The few G dispar were recorded fromsouth-eastern Tanzania around Lindi (10 szlig 00frac34 S 39 szlig 42frac34 E ) and along the RovumaRiver (11 szlig S 39 szlig E ) All G mulanjensis were recorded from 900ndash1000 m on MtMulanje (16 szlig 22frac34 S 35 szlig 07frac34 E ) in southern Malawi
Distance and cladistic analyses
The dendrograms and cladograms derived from the four diŒerent analyses areshown by gure 3 In essential details the inter-relationships among the species wereconsistent with their geographical distribution Each dendrogramcladogram showeda clear branching between east and southern African taxa and in each case there
A L V Davis et al1614
Tab
le3
Sum
mar
yof
know
nge
ogra
phic
aldi
stri
but
ion
and
habi
tat
asso
ciat
ion
sof
Gyro
notu
ssp
pbe
twee
nth
eE
aste
rnC
ape
Sout
hA
fric
aan
dno
rth
-eas
tern
Tan
zani
a
Len
gth
Geo
grap
hic
alra
nge
Alt
itu
din
alG
enu
san
dsp
ecie
s(m
m)dagger
(pat
chy
occu
rren
ce)
rang
e(m
)H
abit
atN
ote
s
Gyro
notu
sva
nL
ansb
erge
1874
South
Afr
ica
All
know
nre
cord
sin
fore
stG
pum
ilus
(Boh
eman
1857
)13
ndash15
Sout
h-ea
stco
asta
lhi
lls50
ndash450
For
est
469
of
128
info
rest
(gu
res
12
)G
ca
rinatu
sF
elsc
he19
1113
ndash15
Zul
ula
nd
coas
tal
hills
50ndash5
00F
ores
t5
7of
53in
fore
st(
gure
s1
2)
G
gla
bro
sus
Scho
ltz
and
Ho
wde
n19
8712
ndash12
5D
rak
ensb
erg
esca
rpm
ent
in78
0F
ores
t33
of
6in
fore
stN
ort
hern
Pro
vinc
e(
gure
s1
2)
Tro
pic
al
Afr
ica
Gm
ula
nje
nsi
sD
avis
Sch
olt
zan
dH
arri
son
118
ndash13
3M
tM
ulan
je
Mal
awi
(gu
re1
)90
0ndash10
00F
ores
tN
ot
reco
rded
F
ores
tan
d19
99op
enco
llect
ion
sm
ixed
DaggerG
Wm
etari
us
Ko
lbe
1894
125
ndash15
5M
ain
lyE
ast
Usa
mba
raM
ts50
ndash850
For
est
Fou
rtr
appe
dby
stre
amin
plu
sea
stco
ast
ofT
anza
nia
fore
stbu
tno
neou
tsid
e(
gure
1)
G
dis
par
(Fel
sche
1911
)16
ndash18
Sout
h-ea
stT
anza
nia
(gu
re1
)50
For
est
No
habi
tat
note
s
dagger Len
gth
wit
hhe
adde
exe
das
oppo
sed
tora
ised
asin
tabl
e5
Dagger C
L
Bel
lam
y(p
erso
nal
com
mun
icat
ion)
Cladistic and biogeographical analysis of Gyronotus 1615
Fig 3 Centre-rooted distance dendrograms and centre-rooted consensus parsimony clado-grams for inter-relationships between Gyronotus species (A B) Cladistic matrix derivedfrom double-standardized raw morphometric measurements by divergence coding(Thorpe 1984) (A) Distance dendrogram based on 27 characters (B) Parsimonycladogram based on 11 informative characters tree length 5 28 CI 5 0893 (C D)Cladistic matrix derived from double-standardized log10 transformed morphometricmeasurements by divergence coding (Thorpe 1984) (C ) Distance dendrogram basedon 27 characters (D) Parsimony cladogram based on 14 informative characters treelength 5 36 CI 5 0889 The numbers are the branch lengths from the nodes forunordered character states
was a similar resolution of relationships between South African species with thesouthernmost species G pumilus distanced from the two more northerly speciesHowever the results for the east African species indicated three diŒerent hypotheticalinter-relationships between species Of the three diŒerent patterns one each wasyielded from the matrices derived from double-standardized raw data whereas thethird was common to the dendrogram and cladogram yielded by the matrices derived
A L V Davis et al1616
from double-standardized log10 data These diŒerences may re ect the relativein uence of body size versus body shape according to the treatment of the datamatrices In the matrices derived from double-standardized raw data body size hasa stronger overall in uence than body shape (higher r2 and F numbers for regressionagainst Factor 1 which accounts for the greatest amount of variance in ordination(table 4) This emphasis is re ected by the closer linking of the two largest speciesG dispar and G Wmetarius Although both body size and body shape are de-emphasized in the matrices derived from double-standardized log10 data body shapehas the greater overall in uence (higher r2 and F numbers for regression againstboth Factors 1 and 2) This emphasis is re ected by the steps in common betweenthe elongate G dispar and G mulanjensis whereas the less elongate G Wmetarius isplaced on a separate branch ( gures 3C 3D)
Further phenetic analysis
As the cladograms were derived from morphometric data it is not surprisingthat the depicted interspeci c relationships are consistent with those indicated bythe three basic measurements of maximum length width and depth These measure-ments clearly separate the more elongate deeper east African species from the moreovoid atter southern African species (table 5) Ratios between the three dimen-sions indicate that width varies inversely with the other two dimensions whereaslength and depth vary proportionately to one another Thus the broadest species(G glabrosus) is also the attest and the most narrow species (G dispar) is alsothe deepest
Character states of aedeagi
F igure 4 illustrates the morphological diŒerences between the aedeagi ofGyronotus and those of large-bodied putative sister groups with primarily tropicaldistributions The parameres of tropical species in putative sister genera Anachalcos
procerus Gerstaecker and Canthodimorpha lawrencei Davis Scholtz and Harrisonare symmetrical and diverge at the tips Those of the east African Gyronotus alsodiverge at the tips but are asymmetrical due to a terminal process on the leftparamere The tips of the parameres of the southern African Gyronotus species aremore convergent and are also asymmetrical due to a terminal process projectingaway to the left of the left paramere
Discussion
Geographical polarization of characters
Comparison of Gyronotus aedeagi with those of the putative sister groupAnachalcos and that of Canthodimorpha suggests that bilateral symmetry anddivergence at the tips of the parameres are plesiomorphic character states ( gure 4)Although the aedeagi of the east African species of Gyronotus are characterized byasymmetry this is primarily due to the presence of a membranous terminal processwhich in ventral view projects to the right from the left paramere and occupies thespace between the divergent tips of the parameres ( gure 4) In contrast the SouthAfrican species are characterized by apomorphic convergence of the tips of theparameres ( gure 4mdashparameres slightly separated in the gure to facilitate drawing)In the two northernmost South African species asymmetry of the parameres ismainly due to the membranous terminal process which in ventral view projects
Cladistic and biogeographical analysis of Gyronotus 1617
Tab
le4
The
corr
elat
ion
ofG
yro
notu
sbo
dysi
ze(m
axim
umle
ngt
h)1
and
body
shap
e(r
atio
max
imu
mle
ngt
hm
axim
um
wid
thof
elyt
ra)2
wit
hor
din
ates
for
Fac
tors
1an
d2
deri
ved
fro
mpr
inci
pal
com
pon
ents
anal
ysis
ofm
orp
hom
etri
csda
tatr
eate
din
two
diŒ
eren
tw
ays
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
raw
mea
sure
men
tsra
wm
easu
rem
ents
log 10
mea
sure
men
tslo
g 10m
easu
rem
ents
(27
char
acte
rs)
(11
char
acte
rs)
(27
char
acte
rs)
(14
char
acte
rs)
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Pro
por
tion
ofov
eral
lvar
ianc
e(
)55
217
756
015
058
313
266
413
6R
2fo
rre
gres
sio
nof
body
size
on0
980
0002
092
000
20
060
230
080
24or
din
ates
1F
(15
1)fo
rre
gres
sio
nof
body
size
2870
60
0
0162
106
008
345
150
9
437
16
38
on
ordi
nat
es1
R2
for
regr
essi
on
ofbo
dysh
ape
on0
620
130
670
060
150
250
140
30or
din
ates
2F
(15
1)fo
rre
gres
sio
nof
body
shap
e83
55
7
55
103
79
3
358
63
173
1
873
22
26
on
ordi
nat
es2
p
lt0
05
p
lt0
02
p
lt0
001
A L V Davis et al1618
Tab
le5
Mor
pho
met
ric
mea
sure
men
tsfo
rsi
xsp
ecie
sof
Gyro
notu
s
Sout
hern
Afr
ican
spec
ies
Eas
tA
fric
ansp
ecie
sA
NO
VA
Mea
sure
men
tG
pum
ilus
G
cari
natu
sG
gla
bro
sus
G
dis
par
G
mula
nje
nsi
sG
Wm
etari
us
F(5
47)
Len
gth
(L)
147
2Ocirc0
64c
141
4Ocirc0
26cd
135
8Ocirc0
77d
206
4Ocirc0
41a
159
5Ocirc0
34b
162
0Ocirc0
26b
194
41
W
idth
(W)
905
Ocirc0
36b
900
Ocirc0
18b
944
Ocirc0
53b
107
4Ocirc0
42a
824
Ocirc0
22c
911
Ocirc0
46b
354
1
Dep
th(D
)6
02Ocirc
023
bd5
80Ocirc
021
cd5
57Ocirc
021
d8
24Ocirc
038
a6
09Ocirc
024
bc6
38Ocirc
027
b88
35
R
atio
sL
W0
62Ocirc
002
b0
64Ocirc
001
b0
70Ocirc
002
a0
52Ocirc
002
d0
52Ocirc
001
d0
56Ocirc
003
c87
16
L
D0
41Ocirc
001
a0
41Ocirc
001
a0
41Ocirc
002
a0
40Ocirc
002
ab0
38Ocirc
001
b0
39Ocirc
002
ab5
16
W
D0
67Ocirc
003
bc0
64Ocirc
002
c0
59Ocirc
002
d0
77Ocirc
004
ab0
74Ocirc
002
ab0
70Ocirc
003
b35
44
Sa
mp
lesi
ze10
106
710
10
Inea
chro
w
valu
esfo
llow
edby
adi
Œer
ent
lett
erdi
Œer
edsi
gni
cant
ly(T
ukey
rsquosH
SD
for
uneq
ual
nETH
plt
005
)
plt
000
1
Cladistic and biogeographical analysis of Gyronotus 1619
Fig 4 Distal and ventral views of canthonine aedeagi showing diŒerences in parameremorphology between (A) Anachalcos procerus Gerstaecker (distal ) (B)Canthodimorpha lawrencei Davis Scholtz and Harrison (ventral) (C ) east AfricanGyronotus van Lansberge (ventral ) (C1 G Wmetarius Kolbe C2 G mulanjensis DavisScholtz and Harrison C3 G dispar (Felsche)) and (D) South African Gyronotus(ventral ) (D1 G carinatus Felsche D2 G glabrosus Scholtz and Howden D3G pumilus (Boheman)) Scale bars 5 1 mm
away to the left of the left paramere However in the southernmost species theasymmetry is more extreme in that each paramere is radically diŒerent in shape(Davis et al 1999) The left paramere is more or less straight and bears the leftwardsaligned terminal process the right paramere is curved towards the mid-point andboth are quite unlike those of the other ve species in which each paramere issimilarly curved close to the distal end Thus there is clear biogeographical polariza-tion of the character states of the aedeagi with greater plesiomorphy in eastAfrican species
A L V Davis et al1620
Parallels between morphology biogeography and phenetic analysis of GyronotusThe interspeci c relationships indicated by the cladograms derived from phenetic
analysis largely corresponded to relationships suggested by morphological charactersof the aedeagi ( gure 4 Davis et al 1999) and these relationships were consistentwith the biogeographical distribution of the species along the south-eastern seaboardof Africa The clear separation of the elongate east African and more ovoid southernAfrican species is re ected by the diŒering position of the terminal process on theleft paramere of the aedeagi ( gure 4) The shape of this process in G Wmetarius
and in G mulanjensis diŒers only slightly in that the one is rounded and the otherangular Although it appears that these two species are very close the angularprocess of G dispar is su ciently similar that it is di cult to determine which ofthe cladograms if any re ects the true phylogenetic relationships between the eastAfrican species those strongly re ecting size and shape ( gures 3A 3B) or thosemore biased by shape ( gures 3C 3D) It is also possible that evolution of aedeagiand morphometric character states are divergent rather than parallel The relation-ships between the southern African species are resolved with greater consistencyThe shape of the parameres is similar in the two northerly species which diŒerprincipally only in the mono d shape of the outward projecting terminal process inG carinatus and the bi d shape of this process in G glabrosus ( gure 4) The moredistant phenetic separation of the southernmost species G pumilus is very stronglysupported by the characters of its aedeagus which diŒer radically from those of theother species However the outward projection of the terminal process indicates itscloser relationships to the other South African species than to the east Africanspecies
Biogeographical climatic and habitat associations of GyronotusSingle Gyronotus species are characteristic of several southern sub-centres of
endemism identi ed in mainland eastern coastal forests of Africa by Burgess et al(1998) These sub-centres occur in the East UsambaraTanga (G Wmetarius) Lindi(G dispar) and UdzungwaMulanje mountain regions (G mulanjensis) The coastalforests of southern Tanzania (Lindi subcentre) contain the most unique assemblageof endemic taxa along the east coast (Burgess et al 1998) and also harbour thelargest of the six known Gyronotus species (G dispar)
In terms of lowland versus afromontane forest association the biogeographicala nities of Gyronotus are dependent on the system used to classify oral zonationIn diŒerent regions altitudinal temperature zonation is modi ed by a combinationof factors including latitude rainfall regime aspect and the relative in uence ofmaritime versus continental climate (Hamilton 1989) Therefore instead of usinga single in exible classi cation system we have used three regional systems whichmake allowance for geographical diŒerences in the altitudinal occurrence of thesame temperature zones and their related ora These classi cation systems are thoseproposed by Lovett (1993) for the Eastern Arc Chapman and White (1970) forMalawi and Low and Rebelo (1996) for southern Africa Thus the East Africanspecies of Gyronotus may be described as elements of tropical lowland forests whichoccur below 1370 m in Malawi (Chapman and White 1970) and below 800m in theeastern arc (Lovett 1993) In the more southerly latitudes of South Africa mostlowland and highland forest is classi ed as afromontane (Low and Rebelo 1996)except for that on coastal sandveld of recent origin (Lawes 1990) AlthoughGyronotus have been recorded in coastal regions around East London (33 szlig 00frac34 S
Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

A L V Davis et al1612
Although Anachalcos has been identi ed as the sister group (Scholtz and Howden1987) it is probably quite distant from Gyronotus which is both ightless and hasasymmetrical aedeagi However it is a useful comparison since the aedeagi ofAnachalcos are similar to those of most other afrotropical Canthonini in as muchas the parameres are symmetrical (Scholtz and Howden 1987) and lack the mem-branous terminal process on the left paramere which is characteristic of all Gyronotus
(Davis et al 1999)
Results
Geographical distribution
The six Gyronotus species showed either disjunct or parapatric distributions fromthe Eastern Cape in South Africa to north-eastern Tanzania ( gure 1) Two SouthAfrican species G pumilus (Eastern Cape southern KwaZulu-Natal ) and G
carinatus (northern KwaZulu-Natal ) showed a parapatric distribution pattern larg-ely restricted to coastal hills below 320m ( gure 2) The southern distribution limitsof G pumilus and the northern distribution limits of G carinatus coincide with thesouthern and northern limits of Walter and Liethrsquos (1964) temperatesubtropicalclimate type II (I )a ( gure 2) The respective northern and southern distribution
Fig 1 Geographical distribution of Gyronotus species from east to southern Africa(T Tanzania M Malawi SA South Africa) (S Swahili centre of endemism SMSwahili-Maputaland transition zone (Burgess et al 1998) Ma Maputaland centre ofendemism P Pondoland centre of endemism which occupies a narrow coastal zone(van Rensburg et al 1999))
Cladistic and biogeographical analysis of Gyronotus 1613
Fig 2 Geographical distribution of Gyronotus pumilus and G carinatus relative to altitudeand the subtropical coastal climate type II(I )a after Walter and Lieth (1964)
limits of G pumilus and G carinatus occur just north of Durban (29 szlig 51frac34 S 31 szlig 01frac34 E )at which latitude land greater than 1500 m in altitude swings away from the coastlineThe third southern African species was recorded only in a small localized region ofthe eastern escarpment of Northern Province in South Africa The combined distribu-tion of these species is sympatric with both coastal forest (particularly in the south)and the low-lying portion of forest classi ed as Afromontane (particularly in thenorth) (table 3) Distribution data for the east African species was geographicallymore limited ( gure 1) Most G Wmetarius were recorded from 215ndash850m in theEast Usambara Mountains of north-eastern Tanzania although a few specimenswere collected from the adjacent coastline The few G dispar were recorded fromsouth-eastern Tanzania around Lindi (10 szlig 00frac34 S 39 szlig 42frac34 E ) and along the RovumaRiver (11 szlig S 39 szlig E ) All G mulanjensis were recorded from 900ndash1000 m on MtMulanje (16 szlig 22frac34 S 35 szlig 07frac34 E ) in southern Malawi
Distance and cladistic analyses
The dendrograms and cladograms derived from the four diŒerent analyses areshown by gure 3 In essential details the inter-relationships among the species wereconsistent with their geographical distribution Each dendrogramcladogram showeda clear branching between east and southern African taxa and in each case there
A L V Davis et al1614
Tab
le3
Sum
mar
yof
know
nge
ogra
phic
aldi
stri
but
ion
and
habi
tat
asso
ciat
ion
sof
Gyro
notu
ssp
pbe
twee
nth
eE
aste
rnC
ape
Sout
hA
fric
aan
dno
rth
-eas
tern
Tan
zani
a
Len
gth
Geo
grap
hic
alra
nge
Alt
itu
din
alG
enu
san
dsp
ecie
s(m
m)dagger
(pat
chy
occu
rren
ce)
rang
e(m
)H
abit
atN
ote
s
Gyro
notu
sva
nL
ansb
erge
1874
South
Afr
ica
All
know
nre
cord
sin
fore
stG
pum
ilus
(Boh
eman
1857
)13
ndash15
Sout
h-ea
stco
asta
lhi
lls50
ndash450
For
est
469
of
128
info
rest
(gu
res
12
)G
ca
rinatu
sF
elsc
he19
1113
ndash15
Zul
ula
nd
coas
tal
hills
50ndash5
00F
ores
t5
7of
53in
fore
st(
gure
s1
2)
G
gla
bro
sus
Scho
ltz
and
Ho
wde
n19
8712
ndash12
5D
rak
ensb
erg
esca
rpm
ent
in78
0F
ores
t33
of
6in
fore
stN
ort
hern
Pro
vinc
e(
gure
s1
2)
Tro
pic
al
Afr
ica
Gm
ula
nje
nsi
sD
avis
Sch
olt
zan
dH
arri
son
118
ndash13
3M
tM
ulan
je
Mal
awi
(gu
re1
)90
0ndash10
00F
ores
tN
ot
reco
rded
F
ores
tan
d19
99op
enco
llect
ion
sm
ixed
DaggerG
Wm
etari
us
Ko
lbe
1894
125
ndash15
5M
ain
lyE
ast
Usa
mba
raM
ts50
ndash850
For
est
Fou
rtr
appe
dby
stre
amin
plu
sea
stco
ast
ofT
anza
nia
fore
stbu
tno
neou
tsid
e(
gure
1)
G
dis
par
(Fel
sche
1911
)16
ndash18
Sout
h-ea
stT
anza
nia
(gu
re1
)50
For
est
No
habi
tat
note
s
dagger Len
gth
wit
hhe
adde
exe
das
oppo
sed
tora
ised
asin
tabl
e5
Dagger C
L
Bel
lam
y(p
erso
nal
com
mun
icat
ion)
Cladistic and biogeographical analysis of Gyronotus 1615
Fig 3 Centre-rooted distance dendrograms and centre-rooted consensus parsimony clado-grams for inter-relationships between Gyronotus species (A B) Cladistic matrix derivedfrom double-standardized raw morphometric measurements by divergence coding(Thorpe 1984) (A) Distance dendrogram based on 27 characters (B) Parsimonycladogram based on 11 informative characters tree length 5 28 CI 5 0893 (C D)Cladistic matrix derived from double-standardized log10 transformed morphometricmeasurements by divergence coding (Thorpe 1984) (C ) Distance dendrogram basedon 27 characters (D) Parsimony cladogram based on 14 informative characters treelength 5 36 CI 5 0889 The numbers are the branch lengths from the nodes forunordered character states
was a similar resolution of relationships between South African species with thesouthernmost species G pumilus distanced from the two more northerly speciesHowever the results for the east African species indicated three diŒerent hypotheticalinter-relationships between species Of the three diŒerent patterns one each wasyielded from the matrices derived from double-standardized raw data whereas thethird was common to the dendrogram and cladogram yielded by the matrices derived
A L V Davis et al1616
from double-standardized log10 data These diŒerences may re ect the relativein uence of body size versus body shape according to the treatment of the datamatrices In the matrices derived from double-standardized raw data body size hasa stronger overall in uence than body shape (higher r2 and F numbers for regressionagainst Factor 1 which accounts for the greatest amount of variance in ordination(table 4) This emphasis is re ected by the closer linking of the two largest speciesG dispar and G Wmetarius Although both body size and body shape are de-emphasized in the matrices derived from double-standardized log10 data body shapehas the greater overall in uence (higher r2 and F numbers for regression againstboth Factors 1 and 2) This emphasis is re ected by the steps in common betweenthe elongate G dispar and G mulanjensis whereas the less elongate G Wmetarius isplaced on a separate branch ( gures 3C 3D)
Further phenetic analysis
As the cladograms were derived from morphometric data it is not surprisingthat the depicted interspeci c relationships are consistent with those indicated bythe three basic measurements of maximum length width and depth These measure-ments clearly separate the more elongate deeper east African species from the moreovoid atter southern African species (table 5) Ratios between the three dimen-sions indicate that width varies inversely with the other two dimensions whereaslength and depth vary proportionately to one another Thus the broadest species(G glabrosus) is also the attest and the most narrow species (G dispar) is alsothe deepest
Character states of aedeagi
F igure 4 illustrates the morphological diŒerences between the aedeagi ofGyronotus and those of large-bodied putative sister groups with primarily tropicaldistributions The parameres of tropical species in putative sister genera Anachalcos
procerus Gerstaecker and Canthodimorpha lawrencei Davis Scholtz and Harrisonare symmetrical and diverge at the tips Those of the east African Gyronotus alsodiverge at the tips but are asymmetrical due to a terminal process on the leftparamere The tips of the parameres of the southern African Gyronotus species aremore convergent and are also asymmetrical due to a terminal process projectingaway to the left of the left paramere
Discussion
Geographical polarization of characters
Comparison of Gyronotus aedeagi with those of the putative sister groupAnachalcos and that of Canthodimorpha suggests that bilateral symmetry anddivergence at the tips of the parameres are plesiomorphic character states ( gure 4)Although the aedeagi of the east African species of Gyronotus are characterized byasymmetry this is primarily due to the presence of a membranous terminal processwhich in ventral view projects to the right from the left paramere and occupies thespace between the divergent tips of the parameres ( gure 4) In contrast the SouthAfrican species are characterized by apomorphic convergence of the tips of theparameres ( gure 4mdashparameres slightly separated in the gure to facilitate drawing)In the two northernmost South African species asymmetry of the parameres ismainly due to the membranous terminal process which in ventral view projects
Cladistic and biogeographical analysis of Gyronotus 1617
Tab
le4
The
corr
elat
ion
ofG
yro
notu
sbo
dysi
ze(m
axim
umle
ngt
h)1
and
body
shap
e(r
atio
max
imu
mle
ngt
hm
axim
um
wid
thof
elyt
ra)2
wit
hor
din
ates
for
Fac
tors
1an
d2
deri
ved
fro
mpr
inci
pal
com
pon
ents
anal
ysis
ofm
orp
hom
etri
csda
tatr
eate
din
two
diŒ
eren
tw
ays
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
raw
mea
sure
men
tsra
wm
easu
rem
ents
log 10
mea
sure
men
tslo
g 10m
easu
rem
ents
(27
char
acte
rs)
(11
char
acte
rs)
(27
char
acte
rs)
(14
char
acte
rs)
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Pro
por
tion
ofov
eral
lvar
ianc
e(
)55
217
756
015
058
313
266
413
6R
2fo
rre
gres
sio
nof
body
size
on0
980
0002
092
000
20
060
230
080
24or
din
ates
1F
(15
1)fo
rre
gres
sio
nof
body
size
2870
60
0
0162
106
008
345
150
9
437
16
38
on
ordi
nat
es1
R2
for
regr
essi
on
ofbo
dysh
ape
on0
620
130
670
060
150
250
140
30or
din
ates
2F
(15
1)fo
rre
gres
sio
nof
body
shap
e83
55
7
55
103
79
3
358
63
173
1
873
22
26
on
ordi
nat
es2
p
lt0
05
p
lt0
02
p
lt0
001
A L V Davis et al1618
Tab
le5
Mor
pho
met
ric
mea
sure
men
tsfo
rsi
xsp
ecie
sof
Gyro
notu
s
Sout
hern
Afr
ican
spec
ies
Eas
tA
fric
ansp
ecie
sA
NO
VA
Mea
sure
men
tG
pum
ilus
G
cari
natu
sG
gla
bro
sus
G
dis
par
G
mula
nje
nsi
sG
Wm
etari
us
F(5
47)
Len
gth
(L)
147
2Ocirc0
64c
141
4Ocirc0
26cd
135
8Ocirc0
77d
206
4Ocirc0
41a
159
5Ocirc0
34b
162
0Ocirc0
26b
194
41
W
idth
(W)
905
Ocirc0
36b
900
Ocirc0
18b
944
Ocirc0
53b
107
4Ocirc0
42a
824
Ocirc0
22c
911
Ocirc0
46b
354
1
Dep
th(D
)6
02Ocirc
023
bd5
80Ocirc
021
cd5
57Ocirc
021
d8
24Ocirc
038
a6
09Ocirc
024
bc6
38Ocirc
027
b88
35
R
atio
sL
W0
62Ocirc
002
b0
64Ocirc
001
b0
70Ocirc
002
a0
52Ocirc
002
d0
52Ocirc
001
d0
56Ocirc
003
c87
16
L
D0
41Ocirc
001
a0
41Ocirc
001
a0
41Ocirc
002
a0
40Ocirc
002
ab0
38Ocirc
001
b0
39Ocirc
002
ab5
16
W
D0
67Ocirc
003
bc0
64Ocirc
002
c0
59Ocirc
002
d0
77Ocirc
004
ab0
74Ocirc
002
ab0
70Ocirc
003
b35
44
Sa
mp
lesi
ze10
106
710
10
Inea
chro
w
valu
esfo
llow
edby
adi
Œer
ent
lett
erdi
Œer
edsi
gni
cant
ly(T
ukey
rsquosH
SD
for
uneq
ual
nETH
plt
005
)
plt
000
1
Cladistic and biogeographical analysis of Gyronotus 1619
Fig 4 Distal and ventral views of canthonine aedeagi showing diŒerences in parameremorphology between (A) Anachalcos procerus Gerstaecker (distal ) (B)Canthodimorpha lawrencei Davis Scholtz and Harrison (ventral) (C ) east AfricanGyronotus van Lansberge (ventral ) (C1 G Wmetarius Kolbe C2 G mulanjensis DavisScholtz and Harrison C3 G dispar (Felsche)) and (D) South African Gyronotus(ventral ) (D1 G carinatus Felsche D2 G glabrosus Scholtz and Howden D3G pumilus (Boheman)) Scale bars 5 1 mm
away to the left of the left paramere However in the southernmost species theasymmetry is more extreme in that each paramere is radically diŒerent in shape(Davis et al 1999) The left paramere is more or less straight and bears the leftwardsaligned terminal process the right paramere is curved towards the mid-point andboth are quite unlike those of the other ve species in which each paramere issimilarly curved close to the distal end Thus there is clear biogeographical polariza-tion of the character states of the aedeagi with greater plesiomorphy in eastAfrican species
A L V Davis et al1620
Parallels between morphology biogeography and phenetic analysis of GyronotusThe interspeci c relationships indicated by the cladograms derived from phenetic
analysis largely corresponded to relationships suggested by morphological charactersof the aedeagi ( gure 4 Davis et al 1999) and these relationships were consistentwith the biogeographical distribution of the species along the south-eastern seaboardof Africa The clear separation of the elongate east African and more ovoid southernAfrican species is re ected by the diŒering position of the terminal process on theleft paramere of the aedeagi ( gure 4) The shape of this process in G Wmetarius
and in G mulanjensis diŒers only slightly in that the one is rounded and the otherangular Although it appears that these two species are very close the angularprocess of G dispar is su ciently similar that it is di cult to determine which ofthe cladograms if any re ects the true phylogenetic relationships between the eastAfrican species those strongly re ecting size and shape ( gures 3A 3B) or thosemore biased by shape ( gures 3C 3D) It is also possible that evolution of aedeagiand morphometric character states are divergent rather than parallel The relation-ships between the southern African species are resolved with greater consistencyThe shape of the parameres is similar in the two northerly species which diŒerprincipally only in the mono d shape of the outward projecting terminal process inG carinatus and the bi d shape of this process in G glabrosus ( gure 4) The moredistant phenetic separation of the southernmost species G pumilus is very stronglysupported by the characters of its aedeagus which diŒer radically from those of theother species However the outward projection of the terminal process indicates itscloser relationships to the other South African species than to the east Africanspecies
Biogeographical climatic and habitat associations of GyronotusSingle Gyronotus species are characteristic of several southern sub-centres of
endemism identi ed in mainland eastern coastal forests of Africa by Burgess et al(1998) These sub-centres occur in the East UsambaraTanga (G Wmetarius) Lindi(G dispar) and UdzungwaMulanje mountain regions (G mulanjensis) The coastalforests of southern Tanzania (Lindi subcentre) contain the most unique assemblageof endemic taxa along the east coast (Burgess et al 1998) and also harbour thelargest of the six known Gyronotus species (G dispar)
In terms of lowland versus afromontane forest association the biogeographicala nities of Gyronotus are dependent on the system used to classify oral zonationIn diŒerent regions altitudinal temperature zonation is modi ed by a combinationof factors including latitude rainfall regime aspect and the relative in uence ofmaritime versus continental climate (Hamilton 1989) Therefore instead of usinga single in exible classi cation system we have used three regional systems whichmake allowance for geographical diŒerences in the altitudinal occurrence of thesame temperature zones and their related ora These classi cation systems are thoseproposed by Lovett (1993) for the Eastern Arc Chapman and White (1970) forMalawi and Low and Rebelo (1996) for southern Africa Thus the East Africanspecies of Gyronotus may be described as elements of tropical lowland forests whichoccur below 1370 m in Malawi (Chapman and White 1970) and below 800m in theeastern arc (Lovett 1993) In the more southerly latitudes of South Africa mostlowland and highland forest is classi ed as afromontane (Low and Rebelo 1996)except for that on coastal sandveld of recent origin (Lawes 1990) AlthoughGyronotus have been recorded in coastal regions around East London (33 szlig 00frac34 S
Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

Cladistic and biogeographical analysis of Gyronotus 1613
Fig 2 Geographical distribution of Gyronotus pumilus and G carinatus relative to altitudeand the subtropical coastal climate type II(I )a after Walter and Lieth (1964)
limits of G pumilus and G carinatus occur just north of Durban (29 szlig 51frac34 S 31 szlig 01frac34 E )at which latitude land greater than 1500 m in altitude swings away from the coastlineThe third southern African species was recorded only in a small localized region ofthe eastern escarpment of Northern Province in South Africa The combined distribu-tion of these species is sympatric with both coastal forest (particularly in the south)and the low-lying portion of forest classi ed as Afromontane (particularly in thenorth) (table 3) Distribution data for the east African species was geographicallymore limited ( gure 1) Most G Wmetarius were recorded from 215ndash850m in theEast Usambara Mountains of north-eastern Tanzania although a few specimenswere collected from the adjacent coastline The few G dispar were recorded fromsouth-eastern Tanzania around Lindi (10 szlig 00frac34 S 39 szlig 42frac34 E ) and along the RovumaRiver (11 szlig S 39 szlig E ) All G mulanjensis were recorded from 900ndash1000 m on MtMulanje (16 szlig 22frac34 S 35 szlig 07frac34 E ) in southern Malawi
Distance and cladistic analyses
The dendrograms and cladograms derived from the four diŒerent analyses areshown by gure 3 In essential details the inter-relationships among the species wereconsistent with their geographical distribution Each dendrogramcladogram showeda clear branching between east and southern African taxa and in each case there
A L V Davis et al1614
Tab
le3
Sum
mar
yof
know
nge
ogra
phic
aldi
stri
but
ion
and
habi
tat
asso
ciat
ion
sof
Gyro
notu
ssp
pbe
twee
nth
eE
aste
rnC
ape
Sout
hA
fric
aan
dno
rth
-eas
tern
Tan
zani
a
Len
gth
Geo
grap
hic
alra
nge
Alt
itu
din
alG
enu
san
dsp
ecie
s(m
m)dagger
(pat
chy
occu
rren
ce)
rang
e(m
)H
abit
atN
ote
s
Gyro
notu
sva
nL
ansb
erge
1874
South
Afr
ica
All
know
nre
cord
sin
fore
stG
pum
ilus
(Boh
eman
1857
)13
ndash15
Sout
h-ea
stco
asta
lhi
lls50
ndash450
For
est
469
of
128
info
rest
(gu
res
12
)G
ca
rinatu
sF
elsc
he19
1113
ndash15
Zul
ula
nd
coas
tal
hills
50ndash5
00F
ores
t5
7of
53in
fore
st(
gure
s1
2)
G
gla
bro
sus
Scho
ltz
and
Ho
wde
n19
8712
ndash12
5D
rak
ensb
erg
esca
rpm
ent
in78
0F
ores
t33
of
6in
fore
stN
ort
hern
Pro
vinc
e(
gure
s1
2)
Tro
pic
al
Afr
ica
Gm
ula
nje
nsi
sD
avis
Sch
olt
zan
dH
arri
son
118
ndash13
3M
tM
ulan
je
Mal
awi
(gu
re1
)90
0ndash10
00F
ores
tN
ot
reco
rded
F
ores
tan
d19
99op
enco
llect
ion
sm
ixed
DaggerG
Wm
etari
us
Ko
lbe
1894
125
ndash15
5M
ain
lyE
ast
Usa
mba
raM
ts50
ndash850
For
est
Fou
rtr
appe
dby
stre
amin
plu
sea
stco
ast
ofT
anza
nia
fore
stbu
tno
neou
tsid
e(
gure
1)
G
dis
par
(Fel
sche
1911
)16
ndash18
Sout
h-ea
stT
anza
nia
(gu
re1
)50
For
est
No
habi
tat
note
s
dagger Len
gth
wit
hhe
adde
exe
das
oppo
sed
tora
ised
asin
tabl
e5
Dagger C
L
Bel
lam
y(p
erso
nal
com
mun
icat
ion)
Cladistic and biogeographical analysis of Gyronotus 1615
Fig 3 Centre-rooted distance dendrograms and centre-rooted consensus parsimony clado-grams for inter-relationships between Gyronotus species (A B) Cladistic matrix derivedfrom double-standardized raw morphometric measurements by divergence coding(Thorpe 1984) (A) Distance dendrogram based on 27 characters (B) Parsimonycladogram based on 11 informative characters tree length 5 28 CI 5 0893 (C D)Cladistic matrix derived from double-standardized log10 transformed morphometricmeasurements by divergence coding (Thorpe 1984) (C ) Distance dendrogram basedon 27 characters (D) Parsimony cladogram based on 14 informative characters treelength 5 36 CI 5 0889 The numbers are the branch lengths from the nodes forunordered character states
was a similar resolution of relationships between South African species with thesouthernmost species G pumilus distanced from the two more northerly speciesHowever the results for the east African species indicated three diŒerent hypotheticalinter-relationships between species Of the three diŒerent patterns one each wasyielded from the matrices derived from double-standardized raw data whereas thethird was common to the dendrogram and cladogram yielded by the matrices derived
A L V Davis et al1616
from double-standardized log10 data These diŒerences may re ect the relativein uence of body size versus body shape according to the treatment of the datamatrices In the matrices derived from double-standardized raw data body size hasa stronger overall in uence than body shape (higher r2 and F numbers for regressionagainst Factor 1 which accounts for the greatest amount of variance in ordination(table 4) This emphasis is re ected by the closer linking of the two largest speciesG dispar and G Wmetarius Although both body size and body shape are de-emphasized in the matrices derived from double-standardized log10 data body shapehas the greater overall in uence (higher r2 and F numbers for regression againstboth Factors 1 and 2) This emphasis is re ected by the steps in common betweenthe elongate G dispar and G mulanjensis whereas the less elongate G Wmetarius isplaced on a separate branch ( gures 3C 3D)
Further phenetic analysis
As the cladograms were derived from morphometric data it is not surprisingthat the depicted interspeci c relationships are consistent with those indicated bythe three basic measurements of maximum length width and depth These measure-ments clearly separate the more elongate deeper east African species from the moreovoid atter southern African species (table 5) Ratios between the three dimen-sions indicate that width varies inversely with the other two dimensions whereaslength and depth vary proportionately to one another Thus the broadest species(G glabrosus) is also the attest and the most narrow species (G dispar) is alsothe deepest
Character states of aedeagi
F igure 4 illustrates the morphological diŒerences between the aedeagi ofGyronotus and those of large-bodied putative sister groups with primarily tropicaldistributions The parameres of tropical species in putative sister genera Anachalcos
procerus Gerstaecker and Canthodimorpha lawrencei Davis Scholtz and Harrisonare symmetrical and diverge at the tips Those of the east African Gyronotus alsodiverge at the tips but are asymmetrical due to a terminal process on the leftparamere The tips of the parameres of the southern African Gyronotus species aremore convergent and are also asymmetrical due to a terminal process projectingaway to the left of the left paramere
Discussion
Geographical polarization of characters
Comparison of Gyronotus aedeagi with those of the putative sister groupAnachalcos and that of Canthodimorpha suggests that bilateral symmetry anddivergence at the tips of the parameres are plesiomorphic character states ( gure 4)Although the aedeagi of the east African species of Gyronotus are characterized byasymmetry this is primarily due to the presence of a membranous terminal processwhich in ventral view projects to the right from the left paramere and occupies thespace between the divergent tips of the parameres ( gure 4) In contrast the SouthAfrican species are characterized by apomorphic convergence of the tips of theparameres ( gure 4mdashparameres slightly separated in the gure to facilitate drawing)In the two northernmost South African species asymmetry of the parameres ismainly due to the membranous terminal process which in ventral view projects
Cladistic and biogeographical analysis of Gyronotus 1617
Tab
le4
The
corr
elat
ion
ofG
yro
notu
sbo
dysi
ze(m
axim
umle
ngt
h)1
and
body
shap
e(r
atio
max
imu
mle
ngt
hm
axim
um
wid
thof
elyt
ra)2
wit
hor
din
ates
for
Fac
tors
1an
d2
deri
ved
fro
mpr
inci
pal
com
pon
ents
anal
ysis
ofm
orp
hom
etri
csda
tatr
eate
din
two
diŒ
eren
tw
ays
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
raw
mea
sure
men
tsra
wm
easu
rem
ents
log 10
mea
sure
men
tslo
g 10m
easu
rem
ents
(27
char
acte
rs)
(11
char
acte
rs)
(27
char
acte
rs)
(14
char
acte
rs)
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Pro
por
tion
ofov
eral
lvar
ianc
e(
)55
217
756
015
058
313
266
413
6R
2fo
rre
gres
sio
nof
body
size
on0
980
0002
092
000
20
060
230
080
24or
din
ates
1F
(15
1)fo
rre
gres
sio
nof
body
size
2870
60
0
0162
106
008
345
150
9
437
16
38
on
ordi
nat
es1
R2
for
regr
essi
on
ofbo
dysh
ape
on0
620
130
670
060
150
250
140
30or
din
ates
2F
(15
1)fo
rre
gres
sio
nof
body
shap
e83
55
7
55
103
79
3
358
63
173
1
873
22
26
on
ordi
nat
es2
p
lt0
05
p
lt0
02
p
lt0
001
A L V Davis et al1618
Tab
le5
Mor
pho
met
ric
mea
sure
men
tsfo
rsi
xsp
ecie
sof
Gyro
notu
s
Sout
hern
Afr
ican
spec
ies
Eas
tA
fric
ansp
ecie
sA
NO
VA
Mea
sure
men
tG
pum
ilus
G
cari
natu
sG
gla
bro
sus
G
dis
par
G
mula
nje
nsi
sG
Wm
etari
us
F(5
47)
Len
gth
(L)
147
2Ocirc0
64c
141
4Ocirc0
26cd
135
8Ocirc0
77d
206
4Ocirc0
41a
159
5Ocirc0
34b
162
0Ocirc0
26b
194
41
W
idth
(W)
905
Ocirc0
36b
900
Ocirc0
18b
944
Ocirc0
53b
107
4Ocirc0
42a
824
Ocirc0
22c
911
Ocirc0
46b
354
1
Dep
th(D
)6
02Ocirc
023
bd5
80Ocirc
021
cd5
57Ocirc
021
d8
24Ocirc
038
a6
09Ocirc
024
bc6
38Ocirc
027
b88
35
R
atio
sL
W0
62Ocirc
002
b0
64Ocirc
001
b0
70Ocirc
002
a0
52Ocirc
002
d0
52Ocirc
001
d0
56Ocirc
003
c87
16
L
D0
41Ocirc
001
a0
41Ocirc
001
a0
41Ocirc
002
a0
40Ocirc
002
ab0
38Ocirc
001
b0
39Ocirc
002
ab5
16
W
D0
67Ocirc
003
bc0
64Ocirc
002
c0
59Ocirc
002
d0
77Ocirc
004
ab0
74Ocirc
002
ab0
70Ocirc
003
b35
44
Sa
mp
lesi
ze10
106
710
10
Inea
chro
w
valu
esfo
llow
edby
adi
Œer
ent
lett
erdi
Œer
edsi
gni
cant
ly(T
ukey
rsquosH
SD
for
uneq
ual
nETH
plt
005
)
plt
000
1
Cladistic and biogeographical analysis of Gyronotus 1619
Fig 4 Distal and ventral views of canthonine aedeagi showing diŒerences in parameremorphology between (A) Anachalcos procerus Gerstaecker (distal ) (B)Canthodimorpha lawrencei Davis Scholtz and Harrison (ventral) (C ) east AfricanGyronotus van Lansberge (ventral ) (C1 G Wmetarius Kolbe C2 G mulanjensis DavisScholtz and Harrison C3 G dispar (Felsche)) and (D) South African Gyronotus(ventral ) (D1 G carinatus Felsche D2 G glabrosus Scholtz and Howden D3G pumilus (Boheman)) Scale bars 5 1 mm
away to the left of the left paramere However in the southernmost species theasymmetry is more extreme in that each paramere is radically diŒerent in shape(Davis et al 1999) The left paramere is more or less straight and bears the leftwardsaligned terminal process the right paramere is curved towards the mid-point andboth are quite unlike those of the other ve species in which each paramere issimilarly curved close to the distal end Thus there is clear biogeographical polariza-tion of the character states of the aedeagi with greater plesiomorphy in eastAfrican species
A L V Davis et al1620
Parallels between morphology biogeography and phenetic analysis of GyronotusThe interspeci c relationships indicated by the cladograms derived from phenetic
analysis largely corresponded to relationships suggested by morphological charactersof the aedeagi ( gure 4 Davis et al 1999) and these relationships were consistentwith the biogeographical distribution of the species along the south-eastern seaboardof Africa The clear separation of the elongate east African and more ovoid southernAfrican species is re ected by the diŒering position of the terminal process on theleft paramere of the aedeagi ( gure 4) The shape of this process in G Wmetarius
and in G mulanjensis diŒers only slightly in that the one is rounded and the otherangular Although it appears that these two species are very close the angularprocess of G dispar is su ciently similar that it is di cult to determine which ofthe cladograms if any re ects the true phylogenetic relationships between the eastAfrican species those strongly re ecting size and shape ( gures 3A 3B) or thosemore biased by shape ( gures 3C 3D) It is also possible that evolution of aedeagiand morphometric character states are divergent rather than parallel The relation-ships between the southern African species are resolved with greater consistencyThe shape of the parameres is similar in the two northerly species which diŒerprincipally only in the mono d shape of the outward projecting terminal process inG carinatus and the bi d shape of this process in G glabrosus ( gure 4) The moredistant phenetic separation of the southernmost species G pumilus is very stronglysupported by the characters of its aedeagus which diŒer radically from those of theother species However the outward projection of the terminal process indicates itscloser relationships to the other South African species than to the east Africanspecies
Biogeographical climatic and habitat associations of GyronotusSingle Gyronotus species are characteristic of several southern sub-centres of
endemism identi ed in mainland eastern coastal forests of Africa by Burgess et al(1998) These sub-centres occur in the East UsambaraTanga (G Wmetarius) Lindi(G dispar) and UdzungwaMulanje mountain regions (G mulanjensis) The coastalforests of southern Tanzania (Lindi subcentre) contain the most unique assemblageof endemic taxa along the east coast (Burgess et al 1998) and also harbour thelargest of the six known Gyronotus species (G dispar)
In terms of lowland versus afromontane forest association the biogeographicala nities of Gyronotus are dependent on the system used to classify oral zonationIn diŒerent regions altitudinal temperature zonation is modi ed by a combinationof factors including latitude rainfall regime aspect and the relative in uence ofmaritime versus continental climate (Hamilton 1989) Therefore instead of usinga single in exible classi cation system we have used three regional systems whichmake allowance for geographical diŒerences in the altitudinal occurrence of thesame temperature zones and their related ora These classi cation systems are thoseproposed by Lovett (1993) for the Eastern Arc Chapman and White (1970) forMalawi and Low and Rebelo (1996) for southern Africa Thus the East Africanspecies of Gyronotus may be described as elements of tropical lowland forests whichoccur below 1370 m in Malawi (Chapman and White 1970) and below 800m in theeastern arc (Lovett 1993) In the more southerly latitudes of South Africa mostlowland and highland forest is classi ed as afromontane (Low and Rebelo 1996)except for that on coastal sandveld of recent origin (Lawes 1990) AlthoughGyronotus have been recorded in coastal regions around East London (33 szlig 00frac34 S
Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)
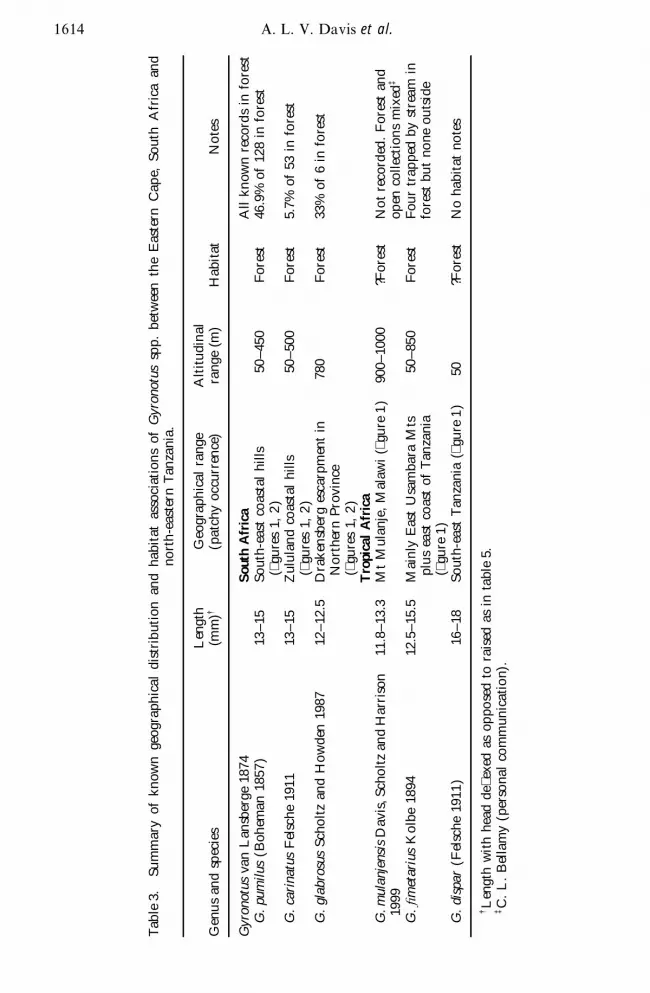
A L V Davis et al1614
Tab
le3
Sum
mar
yof
know
nge
ogra
phic
aldi
stri
but
ion
and
habi
tat
asso
ciat
ion
sof
Gyro
notu
ssp
pbe
twee
nth
eE
aste
rnC
ape
Sout
hA
fric
aan
dno
rth
-eas
tern
Tan
zani
a
Len
gth
Geo
grap
hic
alra
nge
Alt
itu
din
alG
enu
san
dsp
ecie
s(m
m)dagger
(pat
chy
occu
rren
ce)
rang
e(m
)H
abit
atN
ote
s
Gyro
notu
sva
nL
ansb
erge
1874
South
Afr
ica
All
know
nre
cord
sin
fore
stG
pum
ilus
(Boh
eman
1857
)13
ndash15
Sout
h-ea
stco
asta
lhi
lls50
ndash450
For
est
469
of
128
info
rest
(gu
res
12
)G
ca
rinatu
sF
elsc
he19
1113
ndash15
Zul
ula
nd
coas
tal
hills
50ndash5
00F
ores
t5
7of
53in
fore
st(
gure
s1
2)
G
gla
bro
sus
Scho
ltz
and
Ho
wde
n19
8712
ndash12
5D
rak
ensb
erg
esca
rpm
ent
in78
0F
ores
t33
of
6in
fore
stN
ort
hern
Pro
vinc
e(
gure
s1
2)
Tro
pic
al
Afr
ica
Gm
ula
nje
nsi
sD
avis
Sch
olt
zan
dH
arri
son
118
ndash13
3M
tM
ulan
je
Mal
awi
(gu
re1
)90
0ndash10
00F
ores
tN
ot
reco
rded
F
ores
tan
d19
99op
enco
llect
ion
sm
ixed
DaggerG
Wm
etari
us
Ko
lbe
1894
125
ndash15
5M
ain
lyE
ast
Usa
mba
raM
ts50
ndash850
For
est
Fou
rtr
appe
dby
stre
amin
plu
sea
stco
ast
ofT
anza
nia
fore
stbu
tno
neou
tsid
e(
gure
1)
G
dis
par
(Fel
sche
1911
)16
ndash18
Sout
h-ea
stT
anza
nia
(gu
re1
)50
For
est
No
habi
tat
note
s
dagger Len
gth
wit
hhe
adde
exe
das
oppo
sed
tora
ised
asin
tabl
e5
Dagger C
L
Bel
lam
y(p
erso
nal
com
mun
icat
ion)
Cladistic and biogeographical analysis of Gyronotus 1615
Fig 3 Centre-rooted distance dendrograms and centre-rooted consensus parsimony clado-grams for inter-relationships between Gyronotus species (A B) Cladistic matrix derivedfrom double-standardized raw morphometric measurements by divergence coding(Thorpe 1984) (A) Distance dendrogram based on 27 characters (B) Parsimonycladogram based on 11 informative characters tree length 5 28 CI 5 0893 (C D)Cladistic matrix derived from double-standardized log10 transformed morphometricmeasurements by divergence coding (Thorpe 1984) (C ) Distance dendrogram basedon 27 characters (D) Parsimony cladogram based on 14 informative characters treelength 5 36 CI 5 0889 The numbers are the branch lengths from the nodes forunordered character states
was a similar resolution of relationships between South African species with thesouthernmost species G pumilus distanced from the two more northerly speciesHowever the results for the east African species indicated three diŒerent hypotheticalinter-relationships between species Of the three diŒerent patterns one each wasyielded from the matrices derived from double-standardized raw data whereas thethird was common to the dendrogram and cladogram yielded by the matrices derived
A L V Davis et al1616
from double-standardized log10 data These diŒerences may re ect the relativein uence of body size versus body shape according to the treatment of the datamatrices In the matrices derived from double-standardized raw data body size hasa stronger overall in uence than body shape (higher r2 and F numbers for regressionagainst Factor 1 which accounts for the greatest amount of variance in ordination(table 4) This emphasis is re ected by the closer linking of the two largest speciesG dispar and G Wmetarius Although both body size and body shape are de-emphasized in the matrices derived from double-standardized log10 data body shapehas the greater overall in uence (higher r2 and F numbers for regression againstboth Factors 1 and 2) This emphasis is re ected by the steps in common betweenthe elongate G dispar and G mulanjensis whereas the less elongate G Wmetarius isplaced on a separate branch ( gures 3C 3D)
Further phenetic analysis
As the cladograms were derived from morphometric data it is not surprisingthat the depicted interspeci c relationships are consistent with those indicated bythe three basic measurements of maximum length width and depth These measure-ments clearly separate the more elongate deeper east African species from the moreovoid atter southern African species (table 5) Ratios between the three dimen-sions indicate that width varies inversely with the other two dimensions whereaslength and depth vary proportionately to one another Thus the broadest species(G glabrosus) is also the attest and the most narrow species (G dispar) is alsothe deepest
Character states of aedeagi
F igure 4 illustrates the morphological diŒerences between the aedeagi ofGyronotus and those of large-bodied putative sister groups with primarily tropicaldistributions The parameres of tropical species in putative sister genera Anachalcos
procerus Gerstaecker and Canthodimorpha lawrencei Davis Scholtz and Harrisonare symmetrical and diverge at the tips Those of the east African Gyronotus alsodiverge at the tips but are asymmetrical due to a terminal process on the leftparamere The tips of the parameres of the southern African Gyronotus species aremore convergent and are also asymmetrical due to a terminal process projectingaway to the left of the left paramere
Discussion
Geographical polarization of characters
Comparison of Gyronotus aedeagi with those of the putative sister groupAnachalcos and that of Canthodimorpha suggests that bilateral symmetry anddivergence at the tips of the parameres are plesiomorphic character states ( gure 4)Although the aedeagi of the east African species of Gyronotus are characterized byasymmetry this is primarily due to the presence of a membranous terminal processwhich in ventral view projects to the right from the left paramere and occupies thespace between the divergent tips of the parameres ( gure 4) In contrast the SouthAfrican species are characterized by apomorphic convergence of the tips of theparameres ( gure 4mdashparameres slightly separated in the gure to facilitate drawing)In the two northernmost South African species asymmetry of the parameres ismainly due to the membranous terminal process which in ventral view projects
Cladistic and biogeographical analysis of Gyronotus 1617
Tab
le4
The
corr
elat
ion
ofG
yro
notu
sbo
dysi
ze(m
axim
umle
ngt
h)1
and
body
shap
e(r
atio
max
imu
mle
ngt
hm
axim
um
wid
thof
elyt
ra)2
wit
hor
din
ates
for
Fac
tors
1an
d2
deri
ved
fro
mpr
inci
pal
com
pon
ents
anal
ysis
ofm
orp
hom
etri
csda
tatr
eate
din
two
diŒ
eren
tw
ays
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
raw
mea
sure
men
tsra
wm
easu
rem
ents
log 10
mea
sure
men
tslo
g 10m
easu
rem
ents
(27
char
acte
rs)
(11
char
acte
rs)
(27
char
acte
rs)
(14
char
acte
rs)
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Pro
por
tion
ofov
eral
lvar
ianc
e(
)55
217
756
015
058
313
266
413
6R
2fo
rre
gres
sio
nof
body
size
on0
980
0002
092
000
20
060
230
080
24or
din
ates
1F
(15
1)fo
rre
gres
sio
nof
body
size
2870
60
0
0162
106
008
345
150
9
437
16
38
on
ordi
nat
es1
R2
for
regr
essi
on
ofbo
dysh
ape
on0
620
130
670
060
150
250
140
30or
din
ates
2F
(15
1)fo
rre
gres
sio
nof
body
shap
e83
55
7
55
103
79
3
358
63
173
1
873
22
26
on
ordi
nat
es2
p
lt0
05
p
lt0
02
p
lt0
001
A L V Davis et al1618
Tab
le5
Mor
pho
met
ric
mea
sure
men
tsfo
rsi
xsp
ecie
sof
Gyro
notu
s
Sout
hern
Afr
ican
spec
ies
Eas
tA
fric
ansp
ecie
sA
NO
VA
Mea
sure
men
tG
pum
ilus
G
cari
natu
sG
gla
bro
sus
G
dis
par
G
mula
nje
nsi
sG
Wm
etari
us
F(5
47)
Len
gth
(L)
147
2Ocirc0
64c
141
4Ocirc0
26cd
135
8Ocirc0
77d
206
4Ocirc0
41a
159
5Ocirc0
34b
162
0Ocirc0
26b
194
41
W
idth
(W)
905
Ocirc0
36b
900
Ocirc0
18b
944
Ocirc0
53b
107
4Ocirc0
42a
824
Ocirc0
22c
911
Ocirc0
46b
354
1
Dep
th(D
)6
02Ocirc
023
bd5
80Ocirc
021
cd5
57Ocirc
021
d8
24Ocirc
038
a6
09Ocirc
024
bc6
38Ocirc
027
b88
35
R
atio
sL
W0
62Ocirc
002
b0
64Ocirc
001
b0
70Ocirc
002
a0
52Ocirc
002
d0
52Ocirc
001
d0
56Ocirc
003
c87
16
L
D0
41Ocirc
001
a0
41Ocirc
001
a0
41Ocirc
002
a0
40Ocirc
002
ab0
38Ocirc
001
b0
39Ocirc
002
ab5
16
W
D0
67Ocirc
003
bc0
64Ocirc
002
c0
59Ocirc
002
d0
77Ocirc
004
ab0
74Ocirc
002
ab0
70Ocirc
003
b35
44
Sa
mp
lesi
ze10
106
710
10
Inea
chro
w
valu
esfo
llow
edby
adi
Œer
ent
lett
erdi
Œer
edsi
gni
cant
ly(T
ukey
rsquosH
SD
for
uneq
ual
nETH
plt
005
)
plt
000
1
Cladistic and biogeographical analysis of Gyronotus 1619
Fig 4 Distal and ventral views of canthonine aedeagi showing diŒerences in parameremorphology between (A) Anachalcos procerus Gerstaecker (distal ) (B)Canthodimorpha lawrencei Davis Scholtz and Harrison (ventral) (C ) east AfricanGyronotus van Lansberge (ventral ) (C1 G Wmetarius Kolbe C2 G mulanjensis DavisScholtz and Harrison C3 G dispar (Felsche)) and (D) South African Gyronotus(ventral ) (D1 G carinatus Felsche D2 G glabrosus Scholtz and Howden D3G pumilus (Boheman)) Scale bars 5 1 mm
away to the left of the left paramere However in the southernmost species theasymmetry is more extreme in that each paramere is radically diŒerent in shape(Davis et al 1999) The left paramere is more or less straight and bears the leftwardsaligned terminal process the right paramere is curved towards the mid-point andboth are quite unlike those of the other ve species in which each paramere issimilarly curved close to the distal end Thus there is clear biogeographical polariza-tion of the character states of the aedeagi with greater plesiomorphy in eastAfrican species
A L V Davis et al1620
Parallels between morphology biogeography and phenetic analysis of GyronotusThe interspeci c relationships indicated by the cladograms derived from phenetic
analysis largely corresponded to relationships suggested by morphological charactersof the aedeagi ( gure 4 Davis et al 1999) and these relationships were consistentwith the biogeographical distribution of the species along the south-eastern seaboardof Africa The clear separation of the elongate east African and more ovoid southernAfrican species is re ected by the diŒering position of the terminal process on theleft paramere of the aedeagi ( gure 4) The shape of this process in G Wmetarius
and in G mulanjensis diŒers only slightly in that the one is rounded and the otherangular Although it appears that these two species are very close the angularprocess of G dispar is su ciently similar that it is di cult to determine which ofthe cladograms if any re ects the true phylogenetic relationships between the eastAfrican species those strongly re ecting size and shape ( gures 3A 3B) or thosemore biased by shape ( gures 3C 3D) It is also possible that evolution of aedeagiand morphometric character states are divergent rather than parallel The relation-ships between the southern African species are resolved with greater consistencyThe shape of the parameres is similar in the two northerly species which diŒerprincipally only in the mono d shape of the outward projecting terminal process inG carinatus and the bi d shape of this process in G glabrosus ( gure 4) The moredistant phenetic separation of the southernmost species G pumilus is very stronglysupported by the characters of its aedeagus which diŒer radically from those of theother species However the outward projection of the terminal process indicates itscloser relationships to the other South African species than to the east Africanspecies
Biogeographical climatic and habitat associations of GyronotusSingle Gyronotus species are characteristic of several southern sub-centres of
endemism identi ed in mainland eastern coastal forests of Africa by Burgess et al(1998) These sub-centres occur in the East UsambaraTanga (G Wmetarius) Lindi(G dispar) and UdzungwaMulanje mountain regions (G mulanjensis) The coastalforests of southern Tanzania (Lindi subcentre) contain the most unique assemblageof endemic taxa along the east coast (Burgess et al 1998) and also harbour thelargest of the six known Gyronotus species (G dispar)
In terms of lowland versus afromontane forest association the biogeographicala nities of Gyronotus are dependent on the system used to classify oral zonationIn diŒerent regions altitudinal temperature zonation is modi ed by a combinationof factors including latitude rainfall regime aspect and the relative in uence ofmaritime versus continental climate (Hamilton 1989) Therefore instead of usinga single in exible classi cation system we have used three regional systems whichmake allowance for geographical diŒerences in the altitudinal occurrence of thesame temperature zones and their related ora These classi cation systems are thoseproposed by Lovett (1993) for the Eastern Arc Chapman and White (1970) forMalawi and Low and Rebelo (1996) for southern Africa Thus the East Africanspecies of Gyronotus may be described as elements of tropical lowland forests whichoccur below 1370 m in Malawi (Chapman and White 1970) and below 800m in theeastern arc (Lovett 1993) In the more southerly latitudes of South Africa mostlowland and highland forest is classi ed as afromontane (Low and Rebelo 1996)except for that on coastal sandveld of recent origin (Lawes 1990) AlthoughGyronotus have been recorded in coastal regions around East London (33 szlig 00frac34 S
Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

Cladistic and biogeographical analysis of Gyronotus 1615
Fig 3 Centre-rooted distance dendrograms and centre-rooted consensus parsimony clado-grams for inter-relationships between Gyronotus species (A B) Cladistic matrix derivedfrom double-standardized raw morphometric measurements by divergence coding(Thorpe 1984) (A) Distance dendrogram based on 27 characters (B) Parsimonycladogram based on 11 informative characters tree length 5 28 CI 5 0893 (C D)Cladistic matrix derived from double-standardized log10 transformed morphometricmeasurements by divergence coding (Thorpe 1984) (C ) Distance dendrogram basedon 27 characters (D) Parsimony cladogram based on 14 informative characters treelength 5 36 CI 5 0889 The numbers are the branch lengths from the nodes forunordered character states
was a similar resolution of relationships between South African species with thesouthernmost species G pumilus distanced from the two more northerly speciesHowever the results for the east African species indicated three diŒerent hypotheticalinter-relationships between species Of the three diŒerent patterns one each wasyielded from the matrices derived from double-standardized raw data whereas thethird was common to the dendrogram and cladogram yielded by the matrices derived
A L V Davis et al1616
from double-standardized log10 data These diŒerences may re ect the relativein uence of body size versus body shape according to the treatment of the datamatrices In the matrices derived from double-standardized raw data body size hasa stronger overall in uence than body shape (higher r2 and F numbers for regressionagainst Factor 1 which accounts for the greatest amount of variance in ordination(table 4) This emphasis is re ected by the closer linking of the two largest speciesG dispar and G Wmetarius Although both body size and body shape are de-emphasized in the matrices derived from double-standardized log10 data body shapehas the greater overall in uence (higher r2 and F numbers for regression againstboth Factors 1 and 2) This emphasis is re ected by the steps in common betweenthe elongate G dispar and G mulanjensis whereas the less elongate G Wmetarius isplaced on a separate branch ( gures 3C 3D)
Further phenetic analysis
As the cladograms were derived from morphometric data it is not surprisingthat the depicted interspeci c relationships are consistent with those indicated bythe three basic measurements of maximum length width and depth These measure-ments clearly separate the more elongate deeper east African species from the moreovoid atter southern African species (table 5) Ratios between the three dimen-sions indicate that width varies inversely with the other two dimensions whereaslength and depth vary proportionately to one another Thus the broadest species(G glabrosus) is also the attest and the most narrow species (G dispar) is alsothe deepest
Character states of aedeagi
F igure 4 illustrates the morphological diŒerences between the aedeagi ofGyronotus and those of large-bodied putative sister groups with primarily tropicaldistributions The parameres of tropical species in putative sister genera Anachalcos
procerus Gerstaecker and Canthodimorpha lawrencei Davis Scholtz and Harrisonare symmetrical and diverge at the tips Those of the east African Gyronotus alsodiverge at the tips but are asymmetrical due to a terminal process on the leftparamere The tips of the parameres of the southern African Gyronotus species aremore convergent and are also asymmetrical due to a terminal process projectingaway to the left of the left paramere
Discussion
Geographical polarization of characters
Comparison of Gyronotus aedeagi with those of the putative sister groupAnachalcos and that of Canthodimorpha suggests that bilateral symmetry anddivergence at the tips of the parameres are plesiomorphic character states ( gure 4)Although the aedeagi of the east African species of Gyronotus are characterized byasymmetry this is primarily due to the presence of a membranous terminal processwhich in ventral view projects to the right from the left paramere and occupies thespace between the divergent tips of the parameres ( gure 4) In contrast the SouthAfrican species are characterized by apomorphic convergence of the tips of theparameres ( gure 4mdashparameres slightly separated in the gure to facilitate drawing)In the two northernmost South African species asymmetry of the parameres ismainly due to the membranous terminal process which in ventral view projects
Cladistic and biogeographical analysis of Gyronotus 1617
Tab
le4
The
corr
elat
ion
ofG
yro
notu
sbo
dysi
ze(m
axim
umle
ngt
h)1
and
body
shap
e(r
atio
max
imu
mle
ngt
hm
axim
um
wid
thof
elyt
ra)2
wit
hor
din
ates
for
Fac
tors
1an
d2
deri
ved
fro
mpr
inci
pal
com
pon
ents
anal
ysis
ofm
orp
hom
etri
csda
tatr
eate
din
two
diŒ
eren
tw
ays
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
raw
mea
sure
men
tsra
wm
easu
rem
ents
log 10
mea
sure
men
tslo
g 10m
easu
rem
ents
(27
char
acte
rs)
(11
char
acte
rs)
(27
char
acte
rs)
(14
char
acte
rs)
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Pro
por
tion
ofov
eral
lvar
ianc
e(
)55
217
756
015
058
313
266
413
6R
2fo
rre
gres
sio
nof
body
size
on0
980
0002
092
000
20
060
230
080
24or
din
ates
1F
(15
1)fo
rre
gres
sio
nof
body
size
2870
60
0
0162
106
008
345
150
9
437
16
38
on
ordi
nat
es1
R2
for
regr
essi
on
ofbo
dysh
ape
on0
620
130
670
060
150
250
140
30or
din
ates
2F
(15
1)fo
rre
gres
sio
nof
body
shap
e83
55
7
55
103
79
3
358
63
173
1
873
22
26
on
ordi
nat
es2
p
lt0
05
p
lt0
02
p
lt0
001
A L V Davis et al1618
Tab
le5
Mor
pho
met
ric
mea
sure
men
tsfo
rsi
xsp
ecie
sof
Gyro
notu
s
Sout
hern
Afr
ican
spec
ies
Eas
tA
fric
ansp
ecie
sA
NO
VA
Mea
sure
men
tG
pum
ilus
G
cari
natu
sG
gla
bro
sus
G
dis
par
G
mula
nje
nsi
sG
Wm
etari
us
F(5
47)
Len
gth
(L)
147
2Ocirc0
64c
141
4Ocirc0
26cd
135
8Ocirc0
77d
206
4Ocirc0
41a
159
5Ocirc0
34b
162
0Ocirc0
26b
194
41
W
idth
(W)
905
Ocirc0
36b
900
Ocirc0
18b
944
Ocirc0
53b
107
4Ocirc0
42a
824
Ocirc0
22c
911
Ocirc0
46b
354
1
Dep
th(D
)6
02Ocirc
023
bd5
80Ocirc
021
cd5
57Ocirc
021
d8
24Ocirc
038
a6
09Ocirc
024
bc6
38Ocirc
027
b88
35
R
atio
sL
W0
62Ocirc
002
b0
64Ocirc
001
b0
70Ocirc
002
a0
52Ocirc
002
d0
52Ocirc
001
d0
56Ocirc
003
c87
16
L
D0
41Ocirc
001
a0
41Ocirc
001
a0
41Ocirc
002
a0
40Ocirc
002
ab0
38Ocirc
001
b0
39Ocirc
002
ab5
16
W
D0
67Ocirc
003
bc0
64Ocirc
002
c0
59Ocirc
002
d0
77Ocirc
004
ab0
74Ocirc
002
ab0
70Ocirc
003
b35
44
Sa
mp
lesi
ze10
106
710
10
Inea
chro
w
valu
esfo
llow
edby
adi
Œer
ent
lett
erdi
Œer
edsi
gni
cant
ly(T
ukey
rsquosH
SD
for
uneq
ual
nETH
plt
005
)
plt
000
1
Cladistic and biogeographical analysis of Gyronotus 1619
Fig 4 Distal and ventral views of canthonine aedeagi showing diŒerences in parameremorphology between (A) Anachalcos procerus Gerstaecker (distal ) (B)Canthodimorpha lawrencei Davis Scholtz and Harrison (ventral) (C ) east AfricanGyronotus van Lansberge (ventral ) (C1 G Wmetarius Kolbe C2 G mulanjensis DavisScholtz and Harrison C3 G dispar (Felsche)) and (D) South African Gyronotus(ventral ) (D1 G carinatus Felsche D2 G glabrosus Scholtz and Howden D3G pumilus (Boheman)) Scale bars 5 1 mm
away to the left of the left paramere However in the southernmost species theasymmetry is more extreme in that each paramere is radically diŒerent in shape(Davis et al 1999) The left paramere is more or less straight and bears the leftwardsaligned terminal process the right paramere is curved towards the mid-point andboth are quite unlike those of the other ve species in which each paramere issimilarly curved close to the distal end Thus there is clear biogeographical polariza-tion of the character states of the aedeagi with greater plesiomorphy in eastAfrican species
A L V Davis et al1620
Parallels between morphology biogeography and phenetic analysis of GyronotusThe interspeci c relationships indicated by the cladograms derived from phenetic
analysis largely corresponded to relationships suggested by morphological charactersof the aedeagi ( gure 4 Davis et al 1999) and these relationships were consistentwith the biogeographical distribution of the species along the south-eastern seaboardof Africa The clear separation of the elongate east African and more ovoid southernAfrican species is re ected by the diŒering position of the terminal process on theleft paramere of the aedeagi ( gure 4) The shape of this process in G Wmetarius
and in G mulanjensis diŒers only slightly in that the one is rounded and the otherangular Although it appears that these two species are very close the angularprocess of G dispar is su ciently similar that it is di cult to determine which ofthe cladograms if any re ects the true phylogenetic relationships between the eastAfrican species those strongly re ecting size and shape ( gures 3A 3B) or thosemore biased by shape ( gures 3C 3D) It is also possible that evolution of aedeagiand morphometric character states are divergent rather than parallel The relation-ships between the southern African species are resolved with greater consistencyThe shape of the parameres is similar in the two northerly species which diŒerprincipally only in the mono d shape of the outward projecting terminal process inG carinatus and the bi d shape of this process in G glabrosus ( gure 4) The moredistant phenetic separation of the southernmost species G pumilus is very stronglysupported by the characters of its aedeagus which diŒer radically from those of theother species However the outward projection of the terminal process indicates itscloser relationships to the other South African species than to the east Africanspecies
Biogeographical climatic and habitat associations of GyronotusSingle Gyronotus species are characteristic of several southern sub-centres of
endemism identi ed in mainland eastern coastal forests of Africa by Burgess et al(1998) These sub-centres occur in the East UsambaraTanga (G Wmetarius) Lindi(G dispar) and UdzungwaMulanje mountain regions (G mulanjensis) The coastalforests of southern Tanzania (Lindi subcentre) contain the most unique assemblageof endemic taxa along the east coast (Burgess et al 1998) and also harbour thelargest of the six known Gyronotus species (G dispar)
In terms of lowland versus afromontane forest association the biogeographicala nities of Gyronotus are dependent on the system used to classify oral zonationIn diŒerent regions altitudinal temperature zonation is modi ed by a combinationof factors including latitude rainfall regime aspect and the relative in uence ofmaritime versus continental climate (Hamilton 1989) Therefore instead of usinga single in exible classi cation system we have used three regional systems whichmake allowance for geographical diŒerences in the altitudinal occurrence of thesame temperature zones and their related ora These classi cation systems are thoseproposed by Lovett (1993) for the Eastern Arc Chapman and White (1970) forMalawi and Low and Rebelo (1996) for southern Africa Thus the East Africanspecies of Gyronotus may be described as elements of tropical lowland forests whichoccur below 1370 m in Malawi (Chapman and White 1970) and below 800m in theeastern arc (Lovett 1993) In the more southerly latitudes of South Africa mostlowland and highland forest is classi ed as afromontane (Low and Rebelo 1996)except for that on coastal sandveld of recent origin (Lawes 1990) AlthoughGyronotus have been recorded in coastal regions around East London (33 szlig 00frac34 S
Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

A L V Davis et al1616
from double-standardized log10 data These diŒerences may re ect the relativein uence of body size versus body shape according to the treatment of the datamatrices In the matrices derived from double-standardized raw data body size hasa stronger overall in uence than body shape (higher r2 and F numbers for regressionagainst Factor 1 which accounts for the greatest amount of variance in ordination(table 4) This emphasis is re ected by the closer linking of the two largest speciesG dispar and G Wmetarius Although both body size and body shape are de-emphasized in the matrices derived from double-standardized log10 data body shapehas the greater overall in uence (higher r2 and F numbers for regression againstboth Factors 1 and 2) This emphasis is re ected by the steps in common betweenthe elongate G dispar and G mulanjensis whereas the less elongate G Wmetarius isplaced on a separate branch ( gures 3C 3D)
Further phenetic analysis
As the cladograms were derived from morphometric data it is not surprisingthat the depicted interspeci c relationships are consistent with those indicated bythe three basic measurements of maximum length width and depth These measure-ments clearly separate the more elongate deeper east African species from the moreovoid atter southern African species (table 5) Ratios between the three dimen-sions indicate that width varies inversely with the other two dimensions whereaslength and depth vary proportionately to one another Thus the broadest species(G glabrosus) is also the attest and the most narrow species (G dispar) is alsothe deepest
Character states of aedeagi
F igure 4 illustrates the morphological diŒerences between the aedeagi ofGyronotus and those of large-bodied putative sister groups with primarily tropicaldistributions The parameres of tropical species in putative sister genera Anachalcos
procerus Gerstaecker and Canthodimorpha lawrencei Davis Scholtz and Harrisonare symmetrical and diverge at the tips Those of the east African Gyronotus alsodiverge at the tips but are asymmetrical due to a terminal process on the leftparamere The tips of the parameres of the southern African Gyronotus species aremore convergent and are also asymmetrical due to a terminal process projectingaway to the left of the left paramere
Discussion
Geographical polarization of characters
Comparison of Gyronotus aedeagi with those of the putative sister groupAnachalcos and that of Canthodimorpha suggests that bilateral symmetry anddivergence at the tips of the parameres are plesiomorphic character states ( gure 4)Although the aedeagi of the east African species of Gyronotus are characterized byasymmetry this is primarily due to the presence of a membranous terminal processwhich in ventral view projects to the right from the left paramere and occupies thespace between the divergent tips of the parameres ( gure 4) In contrast the SouthAfrican species are characterized by apomorphic convergence of the tips of theparameres ( gure 4mdashparameres slightly separated in the gure to facilitate drawing)In the two northernmost South African species asymmetry of the parameres ismainly due to the membranous terminal process which in ventral view projects
Cladistic and biogeographical analysis of Gyronotus 1617
Tab
le4
The
corr
elat
ion
ofG
yro
notu
sbo
dysi
ze(m
axim
umle
ngt
h)1
and
body
shap
e(r
atio
max
imu
mle
ngt
hm
axim
um
wid
thof
elyt
ra)2
wit
hor
din
ates
for
Fac
tors
1an
d2
deri
ved
fro
mpr
inci
pal
com
pon
ents
anal
ysis
ofm
orp
hom
etri
csda
tatr
eate
din
two
diŒ
eren
tw
ays
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
raw
mea
sure
men
tsra
wm
easu
rem
ents
log 10
mea
sure
men
tslo
g 10m
easu
rem
ents
(27
char
acte
rs)
(11
char
acte
rs)
(27
char
acte
rs)
(14
char
acte
rs)
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Pro
por
tion
ofov
eral
lvar
ianc
e(
)55
217
756
015
058
313
266
413
6R
2fo
rre
gres
sio
nof
body
size
on0
980
0002
092
000
20
060
230
080
24or
din
ates
1F
(15
1)fo
rre
gres
sio
nof
body
size
2870
60
0
0162
106
008
345
150
9
437
16
38
on
ordi
nat
es1
R2
for
regr
essi
on
ofbo
dysh
ape
on0
620
130
670
060
150
250
140
30or
din
ates
2F
(15
1)fo
rre
gres
sio
nof
body
shap
e83
55
7
55
103
79
3
358
63
173
1
873
22
26
on
ordi
nat
es2
p
lt0
05
p
lt0
02
p
lt0
001
A L V Davis et al1618
Tab
le5
Mor
pho
met
ric
mea
sure
men
tsfo
rsi
xsp
ecie
sof
Gyro
notu
s
Sout
hern
Afr
ican
spec
ies
Eas
tA
fric
ansp
ecie
sA
NO
VA
Mea
sure
men
tG
pum
ilus
G
cari
natu
sG
gla
bro
sus
G
dis
par
G
mula
nje
nsi
sG
Wm
etari
us
F(5
47)
Len
gth
(L)
147
2Ocirc0
64c
141
4Ocirc0
26cd
135
8Ocirc0
77d
206
4Ocirc0
41a
159
5Ocirc0
34b
162
0Ocirc0
26b
194
41
W
idth
(W)
905
Ocirc0
36b
900
Ocirc0
18b
944
Ocirc0
53b
107
4Ocirc0
42a
824
Ocirc0
22c
911
Ocirc0
46b
354
1
Dep
th(D
)6
02Ocirc
023
bd5
80Ocirc
021
cd5
57Ocirc
021
d8
24Ocirc
038
a6
09Ocirc
024
bc6
38Ocirc
027
b88
35
R
atio
sL
W0
62Ocirc
002
b0
64Ocirc
001
b0
70Ocirc
002
a0
52Ocirc
002
d0
52Ocirc
001
d0
56Ocirc
003
c87
16
L
D0
41Ocirc
001
a0
41Ocirc
001
a0
41Ocirc
002
a0
40Ocirc
002
ab0
38Ocirc
001
b0
39Ocirc
002
ab5
16
W
D0
67Ocirc
003
bc0
64Ocirc
002
c0
59Ocirc
002
d0
77Ocirc
004
ab0
74Ocirc
002
ab0
70Ocirc
003
b35
44
Sa
mp
lesi
ze10
106
710
10
Inea
chro
w
valu
esfo
llow
edby
adi
Œer
ent
lett
erdi
Œer
edsi
gni
cant
ly(T
ukey
rsquosH
SD
for
uneq
ual
nETH
plt
005
)
plt
000
1
Cladistic and biogeographical analysis of Gyronotus 1619
Fig 4 Distal and ventral views of canthonine aedeagi showing diŒerences in parameremorphology between (A) Anachalcos procerus Gerstaecker (distal ) (B)Canthodimorpha lawrencei Davis Scholtz and Harrison (ventral) (C ) east AfricanGyronotus van Lansberge (ventral ) (C1 G Wmetarius Kolbe C2 G mulanjensis DavisScholtz and Harrison C3 G dispar (Felsche)) and (D) South African Gyronotus(ventral ) (D1 G carinatus Felsche D2 G glabrosus Scholtz and Howden D3G pumilus (Boheman)) Scale bars 5 1 mm
away to the left of the left paramere However in the southernmost species theasymmetry is more extreme in that each paramere is radically diŒerent in shape(Davis et al 1999) The left paramere is more or less straight and bears the leftwardsaligned terminal process the right paramere is curved towards the mid-point andboth are quite unlike those of the other ve species in which each paramere issimilarly curved close to the distal end Thus there is clear biogeographical polariza-tion of the character states of the aedeagi with greater plesiomorphy in eastAfrican species
A L V Davis et al1620
Parallels between morphology biogeography and phenetic analysis of GyronotusThe interspeci c relationships indicated by the cladograms derived from phenetic
analysis largely corresponded to relationships suggested by morphological charactersof the aedeagi ( gure 4 Davis et al 1999) and these relationships were consistentwith the biogeographical distribution of the species along the south-eastern seaboardof Africa The clear separation of the elongate east African and more ovoid southernAfrican species is re ected by the diŒering position of the terminal process on theleft paramere of the aedeagi ( gure 4) The shape of this process in G Wmetarius
and in G mulanjensis diŒers only slightly in that the one is rounded and the otherangular Although it appears that these two species are very close the angularprocess of G dispar is su ciently similar that it is di cult to determine which ofthe cladograms if any re ects the true phylogenetic relationships between the eastAfrican species those strongly re ecting size and shape ( gures 3A 3B) or thosemore biased by shape ( gures 3C 3D) It is also possible that evolution of aedeagiand morphometric character states are divergent rather than parallel The relation-ships between the southern African species are resolved with greater consistencyThe shape of the parameres is similar in the two northerly species which diŒerprincipally only in the mono d shape of the outward projecting terminal process inG carinatus and the bi d shape of this process in G glabrosus ( gure 4) The moredistant phenetic separation of the southernmost species G pumilus is very stronglysupported by the characters of its aedeagus which diŒer radically from those of theother species However the outward projection of the terminal process indicates itscloser relationships to the other South African species than to the east Africanspecies
Biogeographical climatic and habitat associations of GyronotusSingle Gyronotus species are characteristic of several southern sub-centres of
endemism identi ed in mainland eastern coastal forests of Africa by Burgess et al(1998) These sub-centres occur in the East UsambaraTanga (G Wmetarius) Lindi(G dispar) and UdzungwaMulanje mountain regions (G mulanjensis) The coastalforests of southern Tanzania (Lindi subcentre) contain the most unique assemblageof endemic taxa along the east coast (Burgess et al 1998) and also harbour thelargest of the six known Gyronotus species (G dispar)
In terms of lowland versus afromontane forest association the biogeographicala nities of Gyronotus are dependent on the system used to classify oral zonationIn diŒerent regions altitudinal temperature zonation is modi ed by a combinationof factors including latitude rainfall regime aspect and the relative in uence ofmaritime versus continental climate (Hamilton 1989) Therefore instead of usinga single in exible classi cation system we have used three regional systems whichmake allowance for geographical diŒerences in the altitudinal occurrence of thesame temperature zones and their related ora These classi cation systems are thoseproposed by Lovett (1993) for the Eastern Arc Chapman and White (1970) forMalawi and Low and Rebelo (1996) for southern Africa Thus the East Africanspecies of Gyronotus may be described as elements of tropical lowland forests whichoccur below 1370 m in Malawi (Chapman and White 1970) and below 800m in theeastern arc (Lovett 1993) In the more southerly latitudes of South Africa mostlowland and highland forest is classi ed as afromontane (Low and Rebelo 1996)except for that on coastal sandveld of recent origin (Lawes 1990) AlthoughGyronotus have been recorded in coastal regions around East London (33 szlig 00frac34 S
Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

Cladistic and biogeographical analysis of Gyronotus 1617
Tab
le4
The
corr
elat
ion
ofG
yro
notu
sbo
dysi
ze(m
axim
umle
ngt
h)1
and
body
shap
e(r
atio
max
imu
mle
ngt
hm
axim
um
wid
thof
elyt
ra)2
wit
hor
din
ates
for
Fac
tors
1an
d2
deri
ved
fro
mpr
inci
pal
com
pon
ents
anal
ysis
ofm
orp
hom
etri
csda
tatr
eate
din
two
diŒ
eren
tw
ays
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
Do
uble
-sta
ndar
dize
dD
oub
le-s
tand
ardi
zed
raw
mea
sure
men
tsra
wm
easu
rem
ents
log 10
mea
sure
men
tslo
g 10m
easu
rem
ents
(27
char
acte
rs)
(11
char
acte
rs)
(27
char
acte
rs)
(14
char
acte
rs)
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Fac
tor
1F
acto
r2
Pro
por
tion
ofov
eral
lvar
ianc
e(
)55
217
756
015
058
313
266
413
6R
2fo
rre
gres
sio
nof
body
size
on0
980
0002
092
000
20
060
230
080
24or
din
ates
1F
(15
1)fo
rre
gres
sio
nof
body
size
2870
60
0
0162
106
008
345
150
9
437
16
38
on
ordi
nat
es1
R2
for
regr
essi
on
ofbo
dysh
ape
on0
620
130
670
060
150
250
140
30or
din
ates
2F
(15
1)fo
rre
gres
sio
nof
body
shap
e83
55
7
55
103
79
3
358
63
173
1
873
22
26
on
ordi
nat
es2
p
lt0
05
p
lt0
02
p
lt0
001
A L V Davis et al1618
Tab
le5
Mor
pho
met
ric
mea
sure
men
tsfo
rsi
xsp
ecie
sof
Gyro
notu
s
Sout
hern
Afr
ican
spec
ies
Eas
tA
fric
ansp
ecie
sA
NO
VA
Mea
sure
men
tG
pum
ilus
G
cari
natu
sG
gla
bro
sus
G
dis
par
G
mula
nje
nsi
sG
Wm
etari
us
F(5
47)
Len
gth
(L)
147
2Ocirc0
64c
141
4Ocirc0
26cd
135
8Ocirc0
77d
206
4Ocirc0
41a
159
5Ocirc0
34b
162
0Ocirc0
26b
194
41
W
idth
(W)
905
Ocirc0
36b
900
Ocirc0
18b
944
Ocirc0
53b
107
4Ocirc0
42a
824
Ocirc0
22c
911
Ocirc0
46b
354
1
Dep
th(D
)6
02Ocirc
023
bd5
80Ocirc
021
cd5
57Ocirc
021
d8
24Ocirc
038
a6
09Ocirc
024
bc6
38Ocirc
027
b88
35
R
atio
sL
W0
62Ocirc
002
b0
64Ocirc
001
b0
70Ocirc
002
a0
52Ocirc
002
d0
52Ocirc
001
d0
56Ocirc
003
c87
16
L
D0
41Ocirc
001
a0
41Ocirc
001
a0
41Ocirc
002
a0
40Ocirc
002
ab0
38Ocirc
001
b0
39Ocirc
002
ab5
16
W
D0
67Ocirc
003
bc0
64Ocirc
002
c0
59Ocirc
002
d0
77Ocirc
004
ab0
74Ocirc
002
ab0
70Ocirc
003
b35
44
Sa
mp
lesi
ze10
106
710
10
Inea
chro
w
valu
esfo
llow
edby
adi
Œer
ent
lett
erdi
Œer
edsi
gni
cant
ly(T
ukey
rsquosH
SD
for
uneq
ual
nETH
plt
005
)
plt
000
1
Cladistic and biogeographical analysis of Gyronotus 1619
Fig 4 Distal and ventral views of canthonine aedeagi showing diŒerences in parameremorphology between (A) Anachalcos procerus Gerstaecker (distal ) (B)Canthodimorpha lawrencei Davis Scholtz and Harrison (ventral) (C ) east AfricanGyronotus van Lansberge (ventral ) (C1 G Wmetarius Kolbe C2 G mulanjensis DavisScholtz and Harrison C3 G dispar (Felsche)) and (D) South African Gyronotus(ventral ) (D1 G carinatus Felsche D2 G glabrosus Scholtz and Howden D3G pumilus (Boheman)) Scale bars 5 1 mm
away to the left of the left paramere However in the southernmost species theasymmetry is more extreme in that each paramere is radically diŒerent in shape(Davis et al 1999) The left paramere is more or less straight and bears the leftwardsaligned terminal process the right paramere is curved towards the mid-point andboth are quite unlike those of the other ve species in which each paramere issimilarly curved close to the distal end Thus there is clear biogeographical polariza-tion of the character states of the aedeagi with greater plesiomorphy in eastAfrican species
A L V Davis et al1620
Parallels between morphology biogeography and phenetic analysis of GyronotusThe interspeci c relationships indicated by the cladograms derived from phenetic
analysis largely corresponded to relationships suggested by morphological charactersof the aedeagi ( gure 4 Davis et al 1999) and these relationships were consistentwith the biogeographical distribution of the species along the south-eastern seaboardof Africa The clear separation of the elongate east African and more ovoid southernAfrican species is re ected by the diŒering position of the terminal process on theleft paramere of the aedeagi ( gure 4) The shape of this process in G Wmetarius
and in G mulanjensis diŒers only slightly in that the one is rounded and the otherangular Although it appears that these two species are very close the angularprocess of G dispar is su ciently similar that it is di cult to determine which ofthe cladograms if any re ects the true phylogenetic relationships between the eastAfrican species those strongly re ecting size and shape ( gures 3A 3B) or thosemore biased by shape ( gures 3C 3D) It is also possible that evolution of aedeagiand morphometric character states are divergent rather than parallel The relation-ships between the southern African species are resolved with greater consistencyThe shape of the parameres is similar in the two northerly species which diŒerprincipally only in the mono d shape of the outward projecting terminal process inG carinatus and the bi d shape of this process in G glabrosus ( gure 4) The moredistant phenetic separation of the southernmost species G pumilus is very stronglysupported by the characters of its aedeagus which diŒer radically from those of theother species However the outward projection of the terminal process indicates itscloser relationships to the other South African species than to the east Africanspecies
Biogeographical climatic and habitat associations of GyronotusSingle Gyronotus species are characteristic of several southern sub-centres of
endemism identi ed in mainland eastern coastal forests of Africa by Burgess et al(1998) These sub-centres occur in the East UsambaraTanga (G Wmetarius) Lindi(G dispar) and UdzungwaMulanje mountain regions (G mulanjensis) The coastalforests of southern Tanzania (Lindi subcentre) contain the most unique assemblageof endemic taxa along the east coast (Burgess et al 1998) and also harbour thelargest of the six known Gyronotus species (G dispar)
In terms of lowland versus afromontane forest association the biogeographicala nities of Gyronotus are dependent on the system used to classify oral zonationIn diŒerent regions altitudinal temperature zonation is modi ed by a combinationof factors including latitude rainfall regime aspect and the relative in uence ofmaritime versus continental climate (Hamilton 1989) Therefore instead of usinga single in exible classi cation system we have used three regional systems whichmake allowance for geographical diŒerences in the altitudinal occurrence of thesame temperature zones and their related ora These classi cation systems are thoseproposed by Lovett (1993) for the Eastern Arc Chapman and White (1970) forMalawi and Low and Rebelo (1996) for southern Africa Thus the East Africanspecies of Gyronotus may be described as elements of tropical lowland forests whichoccur below 1370 m in Malawi (Chapman and White 1970) and below 800m in theeastern arc (Lovett 1993) In the more southerly latitudes of South Africa mostlowland and highland forest is classi ed as afromontane (Low and Rebelo 1996)except for that on coastal sandveld of recent origin (Lawes 1990) AlthoughGyronotus have been recorded in coastal regions around East London (33 szlig 00frac34 S
Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)
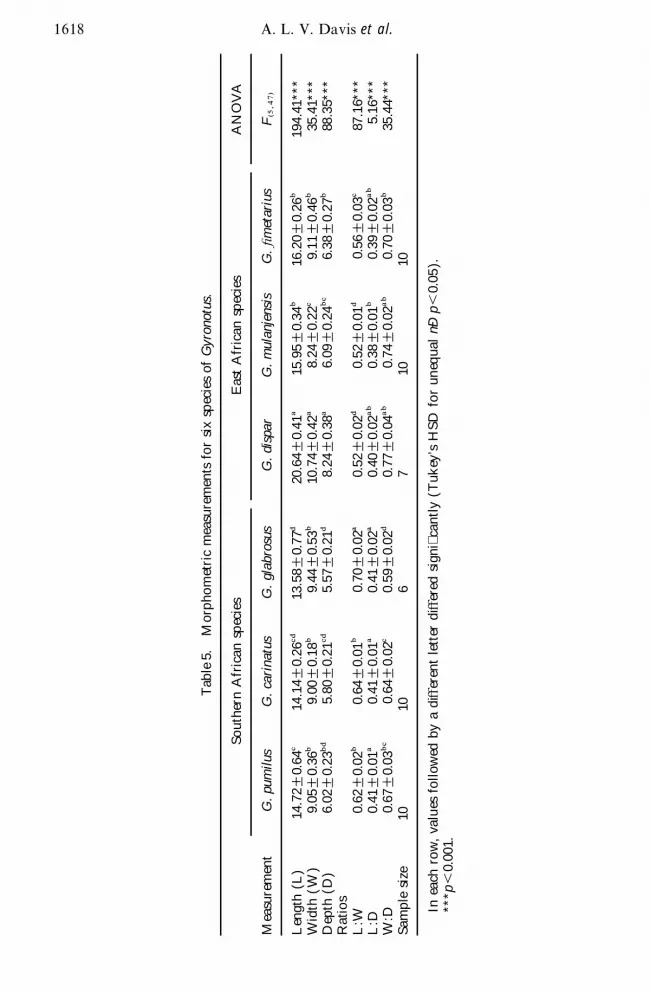
A L V Davis et al1618
Tab
le5
Mor
pho
met
ric
mea
sure
men
tsfo
rsi
xsp
ecie
sof
Gyro
notu
s
Sout
hern
Afr
ican
spec
ies
Eas
tA
fric
ansp
ecie
sA
NO
VA
Mea
sure
men
tG
pum
ilus
G
cari
natu
sG
gla
bro
sus
G
dis
par
G
mula
nje
nsi
sG
Wm
etari
us
F(5
47)
Len
gth
(L)
147
2Ocirc0
64c
141
4Ocirc0
26cd
135
8Ocirc0
77d
206
4Ocirc0
41a
159
5Ocirc0
34b
162
0Ocirc0
26b
194
41
W
idth
(W)
905
Ocirc0
36b
900
Ocirc0
18b
944
Ocirc0
53b
107
4Ocirc0
42a
824
Ocirc0
22c
911
Ocirc0
46b
354
1
Dep
th(D
)6
02Ocirc
023
bd5
80Ocirc
021
cd5
57Ocirc
021
d8
24Ocirc
038
a6
09Ocirc
024
bc6
38Ocirc
027
b88
35
R
atio
sL
W0
62Ocirc
002
b0
64Ocirc
001
b0
70Ocirc
002
a0
52Ocirc
002
d0
52Ocirc
001
d0
56Ocirc
003
c87
16
L
D0
41Ocirc
001
a0
41Ocirc
001
a0
41Ocirc
002
a0
40Ocirc
002
ab0
38Ocirc
001
b0
39Ocirc
002
ab5
16
W
D0
67Ocirc
003
bc0
64Ocirc
002
c0
59Ocirc
002
d0
77Ocirc
004
ab0
74Ocirc
002
ab0
70Ocirc
003
b35
44
Sa
mp
lesi
ze10
106
710
10
Inea
chro
w
valu
esfo
llow
edby
adi
Œer
ent
lett
erdi
Œer
edsi
gni
cant
ly(T
ukey
rsquosH
SD
for
uneq
ual
nETH
plt
005
)
plt
000
1
Cladistic and biogeographical analysis of Gyronotus 1619
Fig 4 Distal and ventral views of canthonine aedeagi showing diŒerences in parameremorphology between (A) Anachalcos procerus Gerstaecker (distal ) (B)Canthodimorpha lawrencei Davis Scholtz and Harrison (ventral) (C ) east AfricanGyronotus van Lansberge (ventral ) (C1 G Wmetarius Kolbe C2 G mulanjensis DavisScholtz and Harrison C3 G dispar (Felsche)) and (D) South African Gyronotus(ventral ) (D1 G carinatus Felsche D2 G glabrosus Scholtz and Howden D3G pumilus (Boheman)) Scale bars 5 1 mm
away to the left of the left paramere However in the southernmost species theasymmetry is more extreme in that each paramere is radically diŒerent in shape(Davis et al 1999) The left paramere is more or less straight and bears the leftwardsaligned terminal process the right paramere is curved towards the mid-point andboth are quite unlike those of the other ve species in which each paramere issimilarly curved close to the distal end Thus there is clear biogeographical polariza-tion of the character states of the aedeagi with greater plesiomorphy in eastAfrican species
A L V Davis et al1620
Parallels between morphology biogeography and phenetic analysis of GyronotusThe interspeci c relationships indicated by the cladograms derived from phenetic
analysis largely corresponded to relationships suggested by morphological charactersof the aedeagi ( gure 4 Davis et al 1999) and these relationships were consistentwith the biogeographical distribution of the species along the south-eastern seaboardof Africa The clear separation of the elongate east African and more ovoid southernAfrican species is re ected by the diŒering position of the terminal process on theleft paramere of the aedeagi ( gure 4) The shape of this process in G Wmetarius
and in G mulanjensis diŒers only slightly in that the one is rounded and the otherangular Although it appears that these two species are very close the angularprocess of G dispar is su ciently similar that it is di cult to determine which ofthe cladograms if any re ects the true phylogenetic relationships between the eastAfrican species those strongly re ecting size and shape ( gures 3A 3B) or thosemore biased by shape ( gures 3C 3D) It is also possible that evolution of aedeagiand morphometric character states are divergent rather than parallel The relation-ships between the southern African species are resolved with greater consistencyThe shape of the parameres is similar in the two northerly species which diŒerprincipally only in the mono d shape of the outward projecting terminal process inG carinatus and the bi d shape of this process in G glabrosus ( gure 4) The moredistant phenetic separation of the southernmost species G pumilus is very stronglysupported by the characters of its aedeagus which diŒer radically from those of theother species However the outward projection of the terminal process indicates itscloser relationships to the other South African species than to the east Africanspecies
Biogeographical climatic and habitat associations of GyronotusSingle Gyronotus species are characteristic of several southern sub-centres of
endemism identi ed in mainland eastern coastal forests of Africa by Burgess et al(1998) These sub-centres occur in the East UsambaraTanga (G Wmetarius) Lindi(G dispar) and UdzungwaMulanje mountain regions (G mulanjensis) The coastalforests of southern Tanzania (Lindi subcentre) contain the most unique assemblageof endemic taxa along the east coast (Burgess et al 1998) and also harbour thelargest of the six known Gyronotus species (G dispar)
In terms of lowland versus afromontane forest association the biogeographicala nities of Gyronotus are dependent on the system used to classify oral zonationIn diŒerent regions altitudinal temperature zonation is modi ed by a combinationof factors including latitude rainfall regime aspect and the relative in uence ofmaritime versus continental climate (Hamilton 1989) Therefore instead of usinga single in exible classi cation system we have used three regional systems whichmake allowance for geographical diŒerences in the altitudinal occurrence of thesame temperature zones and their related ora These classi cation systems are thoseproposed by Lovett (1993) for the Eastern Arc Chapman and White (1970) forMalawi and Low and Rebelo (1996) for southern Africa Thus the East Africanspecies of Gyronotus may be described as elements of tropical lowland forests whichoccur below 1370 m in Malawi (Chapman and White 1970) and below 800m in theeastern arc (Lovett 1993) In the more southerly latitudes of South Africa mostlowland and highland forest is classi ed as afromontane (Low and Rebelo 1996)except for that on coastal sandveld of recent origin (Lawes 1990) AlthoughGyronotus have been recorded in coastal regions around East London (33 szlig 00frac34 S
Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)
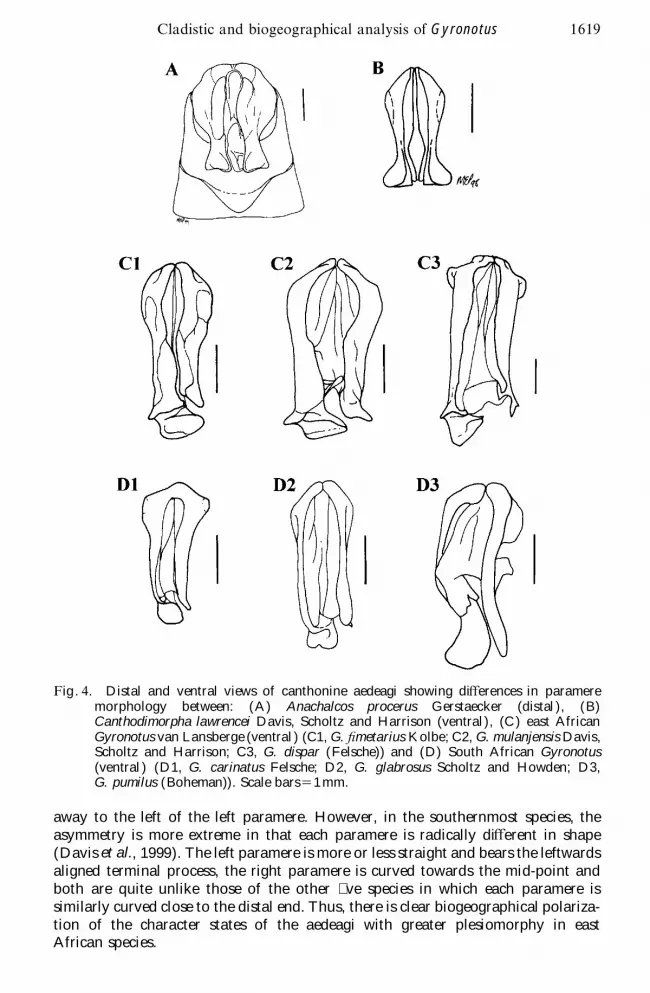
Cladistic and biogeographical analysis of Gyronotus 1619
Fig 4 Distal and ventral views of canthonine aedeagi showing diŒerences in parameremorphology between (A) Anachalcos procerus Gerstaecker (distal ) (B)Canthodimorpha lawrencei Davis Scholtz and Harrison (ventral) (C ) east AfricanGyronotus van Lansberge (ventral ) (C1 G Wmetarius Kolbe C2 G mulanjensis DavisScholtz and Harrison C3 G dispar (Felsche)) and (D) South African Gyronotus(ventral ) (D1 G carinatus Felsche D2 G glabrosus Scholtz and Howden D3G pumilus (Boheman)) Scale bars 5 1 mm
away to the left of the left paramere However in the southernmost species theasymmetry is more extreme in that each paramere is radically diŒerent in shape(Davis et al 1999) The left paramere is more or less straight and bears the leftwardsaligned terminal process the right paramere is curved towards the mid-point andboth are quite unlike those of the other ve species in which each paramere issimilarly curved close to the distal end Thus there is clear biogeographical polariza-tion of the character states of the aedeagi with greater plesiomorphy in eastAfrican species
A L V Davis et al1620
Parallels between morphology biogeography and phenetic analysis of GyronotusThe interspeci c relationships indicated by the cladograms derived from phenetic
analysis largely corresponded to relationships suggested by morphological charactersof the aedeagi ( gure 4 Davis et al 1999) and these relationships were consistentwith the biogeographical distribution of the species along the south-eastern seaboardof Africa The clear separation of the elongate east African and more ovoid southernAfrican species is re ected by the diŒering position of the terminal process on theleft paramere of the aedeagi ( gure 4) The shape of this process in G Wmetarius
and in G mulanjensis diŒers only slightly in that the one is rounded and the otherangular Although it appears that these two species are very close the angularprocess of G dispar is su ciently similar that it is di cult to determine which ofthe cladograms if any re ects the true phylogenetic relationships between the eastAfrican species those strongly re ecting size and shape ( gures 3A 3B) or thosemore biased by shape ( gures 3C 3D) It is also possible that evolution of aedeagiand morphometric character states are divergent rather than parallel The relation-ships between the southern African species are resolved with greater consistencyThe shape of the parameres is similar in the two northerly species which diŒerprincipally only in the mono d shape of the outward projecting terminal process inG carinatus and the bi d shape of this process in G glabrosus ( gure 4) The moredistant phenetic separation of the southernmost species G pumilus is very stronglysupported by the characters of its aedeagus which diŒer radically from those of theother species However the outward projection of the terminal process indicates itscloser relationships to the other South African species than to the east Africanspecies
Biogeographical climatic and habitat associations of GyronotusSingle Gyronotus species are characteristic of several southern sub-centres of
endemism identi ed in mainland eastern coastal forests of Africa by Burgess et al(1998) These sub-centres occur in the East UsambaraTanga (G Wmetarius) Lindi(G dispar) and UdzungwaMulanje mountain regions (G mulanjensis) The coastalforests of southern Tanzania (Lindi subcentre) contain the most unique assemblageof endemic taxa along the east coast (Burgess et al 1998) and also harbour thelargest of the six known Gyronotus species (G dispar)
In terms of lowland versus afromontane forest association the biogeographicala nities of Gyronotus are dependent on the system used to classify oral zonationIn diŒerent regions altitudinal temperature zonation is modi ed by a combinationof factors including latitude rainfall regime aspect and the relative in uence ofmaritime versus continental climate (Hamilton 1989) Therefore instead of usinga single in exible classi cation system we have used three regional systems whichmake allowance for geographical diŒerences in the altitudinal occurrence of thesame temperature zones and their related ora These classi cation systems are thoseproposed by Lovett (1993) for the Eastern Arc Chapman and White (1970) forMalawi and Low and Rebelo (1996) for southern Africa Thus the East Africanspecies of Gyronotus may be described as elements of tropical lowland forests whichoccur below 1370 m in Malawi (Chapman and White 1970) and below 800m in theeastern arc (Lovett 1993) In the more southerly latitudes of South Africa mostlowland and highland forest is classi ed as afromontane (Low and Rebelo 1996)except for that on coastal sandveld of recent origin (Lawes 1990) AlthoughGyronotus have been recorded in coastal regions around East London (33 szlig 00frac34 S
Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

A L V Davis et al1620
Parallels between morphology biogeography and phenetic analysis of GyronotusThe interspeci c relationships indicated by the cladograms derived from phenetic
analysis largely corresponded to relationships suggested by morphological charactersof the aedeagi ( gure 4 Davis et al 1999) and these relationships were consistentwith the biogeographical distribution of the species along the south-eastern seaboardof Africa The clear separation of the elongate east African and more ovoid southernAfrican species is re ected by the diŒering position of the terminal process on theleft paramere of the aedeagi ( gure 4) The shape of this process in G Wmetarius
and in G mulanjensis diŒers only slightly in that the one is rounded and the otherangular Although it appears that these two species are very close the angularprocess of G dispar is su ciently similar that it is di cult to determine which ofthe cladograms if any re ects the true phylogenetic relationships between the eastAfrican species those strongly re ecting size and shape ( gures 3A 3B) or thosemore biased by shape ( gures 3C 3D) It is also possible that evolution of aedeagiand morphometric character states are divergent rather than parallel The relation-ships between the southern African species are resolved with greater consistencyThe shape of the parameres is similar in the two northerly species which diŒerprincipally only in the mono d shape of the outward projecting terminal process inG carinatus and the bi d shape of this process in G glabrosus ( gure 4) The moredistant phenetic separation of the southernmost species G pumilus is very stronglysupported by the characters of its aedeagus which diŒer radically from those of theother species However the outward projection of the terminal process indicates itscloser relationships to the other South African species than to the east Africanspecies
Biogeographical climatic and habitat associations of GyronotusSingle Gyronotus species are characteristic of several southern sub-centres of
endemism identi ed in mainland eastern coastal forests of Africa by Burgess et al(1998) These sub-centres occur in the East UsambaraTanga (G Wmetarius) Lindi(G dispar) and UdzungwaMulanje mountain regions (G mulanjensis) The coastalforests of southern Tanzania (Lindi subcentre) contain the most unique assemblageof endemic taxa along the east coast (Burgess et al 1998) and also harbour thelargest of the six known Gyronotus species (G dispar)
In terms of lowland versus afromontane forest association the biogeographicala nities of Gyronotus are dependent on the system used to classify oral zonationIn diŒerent regions altitudinal temperature zonation is modi ed by a combinationof factors including latitude rainfall regime aspect and the relative in uence ofmaritime versus continental climate (Hamilton 1989) Therefore instead of usinga single in exible classi cation system we have used three regional systems whichmake allowance for geographical diŒerences in the altitudinal occurrence of thesame temperature zones and their related ora These classi cation systems are thoseproposed by Lovett (1993) for the Eastern Arc Chapman and White (1970) forMalawi and Low and Rebelo (1996) for southern Africa Thus the East Africanspecies of Gyronotus may be described as elements of tropical lowland forests whichoccur below 1370 m in Malawi (Chapman and White 1970) and below 800m in theeastern arc (Lovett 1993) In the more southerly latitudes of South Africa mostlowland and highland forest is classi ed as afromontane (Low and Rebelo 1996)except for that on coastal sandveld of recent origin (Lawes 1990) AlthoughGyronotus have been recorded in coastal regions around East London (33 szlig 00frac34 S
Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

Cladistic and biogeographical analysis of Gyronotus 1621
27 szlig 55frac34 E ) none has been recorded during extensive trapping in coastal sand forestin the north of KwaZulu-Natal (Van Rensburg et al 1999) However during veryrecent trapping (Davis Scholtz van Aarde and Chown unpublished ) Gyronotus
have been recorded in old coastal dune forest near Richards Bay (28 szlig 30ndash31 frac34 S32 szlig 23frac34 E ) in southern Maputaland Thus the South African species of Gyronotus
may be described as occurring in low-lying forests with either southern coastalnorthern sand or afromontane a nities (Low and Rebelo 1996)
However according to a GIS overlay of vegetation-type distribution (Low andRebelo 1996) there was a poor correlation between Gyronotus grid references andeither coastal or afromontane forest Most localities (22 out of 26) coincided withbushveld (17) or grass (5) This included some known forest localities VernonCrookes Nature Reserve (30 szlig 16frac34 S 30 szlig 29frac34 E ) Dwesa Forest (32 szlig 15frac34 S 28 szlig 49frac34 E ) SilakaForest (31 szlig 33frac34 S 29 szlig 30frac34 E ) Woodbush Forest (23 szlig 52frac34 S 29 szlig 56frac34 E ) Others (3) coincidedwith coastal forest Ntsubane Forest (31 szlig 27frac34 S 29 szlig 44frac34 E ) Dwesa Forest (32 szlig 17frac34 E28 szlig 50frac34 E ) Only one coincided with afromontane forest Ngoye Forest (28 szlig 50frac34 S31 szlig 43frac34 E ) This may be due to various factors including inaccurate locality labelsandor inaccurate grid references in relation to the patchy occurrence of forestfragments Clearance of forest subsequent to collection may also have in uencedresults since a number of the localities are for material collected in the late 1800s orearly 1900s
Available habitat data and circumstantial evidence suggest that Gyronotus com-prise entirely forest specialists (table 3) Furthermore trap data for Gyronotus Wmet-
arius G pumilus and G mulanjensis suggest that these species are also microhabitatspecialists occurring in cool or cool wet spots within the forest Traps placed atvarious sites in the East Usambara Mts failed to record G Wmetarius outside offorest or at drier sites within forest It was recorded at only one site adjoining astream (215m) (Newmark Davis and Rickart unpublished ) Similar data fromgrassland and forest in Vernon Crookes Nature Reserve shows that G pumilus isrestricted to forest patches where it was trapped only at wet spots next to a streamor in a patch characterized by a high density of worm casts (Davis Scholtz andChown unpublished ) Furthermore one of only two localities for G mulanjensis
occurs along Muloza Stream on Mt Mulanje in Malawi However very recentrecords (Davis Scholtz van Aarde and Chown unpublished ) show that G carinatus
occurs in relatively cool coastal dune forest in southern Maputaland as well as on ner-grained soils in forests of the coastal hills
Gyronotus species also show geographical diŒerences in body shape These diŒer-ences may be related to climate since the elongate Gyronotus species are found intropical east and east-central Africa whereas the ovoid species are found in lowlandand low-lying afromontane forest in South Africa A statistical test of correlationbetween body shape measurements and climate cannot be attempted at present inthe absence of suitable forest temperature data However the proportionally deepestand most elongate species (G dispar) occurs in low altitude tropical forest of south-east Tanzania The proportionally least deep and least elongate tropical species (G
Wmetarius) occurs primarily in the cool wet forests of the East Usambara Mts Theproportionally attest most ovoid of the southern African species (G glabrosus)shows a northerly inland distribution in forests with afromontane a nities Theproportionally deepest least ovoid of the southern African species (G pumilus)occupies a southerly but coastal distribution
A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

A L V Davis et al1622
Distributional hypotheses
Burgess et al (1998) have suggested ve hypotheses to account for the distribu-tional patterns of biota in east African coastal forests which they de ne as occurringbetween Somalia and Mozambique encompassing the Swahili centre of endemismand the transitional region between the Swahili and Maputaland centres of endem-ism These hypotheses are equally applicable to the relict distribution of Gyronotus
which links the Swahili centre of endemism to those of Maputaland and Pondolandfurther to the south Hypothesis one suggests that distributional patterns result fromcollecting artifacts which may or may not account for the between ZambeziLimpopodisjunction in Gyronotus records Hypothesis two suggests that climatic history isresponsible for distributional patterns In Gyronotus combined tectonic and orogenicin uences on climatic history are probably responsible for the isolation of theirancestors in eastern forests during the Miocene and perhaps for later north-southspecies group separation Hypothesis three suggests that vicariant species evolutionis responsible for distribution patterns In Gyronotus some within-group speciationmay result from more recent forest fragmentation during the Pleistocene InGyronotus it is probable that human disturbance (hypothesis four) is responsiblefor increasing fragmentation of range localization of survivors and threat to theirsurvival F inally hypothesis ve suggests past sea-level changes have in uenceddistributional patterns In Gyronotus the distinctive relict distribution pattern at themargin of coastal lowlands might possibly be in uenced by recent change in coastlineposition due to PlioPleistocene intensi cation of polar glaciation and subsequentmarine regression These hypotheses are discussed in more detail below
Gyronotus and south-east African historical and vicariance events
The geographical distribution of Gyronotus may describe the relict of an oldtropical lowland distribution pattern which links wet tropical lowland forest of EastAfrica (Chapman and White 1970 Lovett 1993) to low altitude forest with coastalor afromontane a nities (Low and Rebelo 1996) in South Africa The northernlimits of their polarized distribution lie in the East Usambara Mts whose age isuncertain but probably originated from 290ndash180 MY ago with further uplift at7 MY (Gri ths 1993) The East Usambaras comprise part of the ancient easternarc mountain system which is the eastern-most of ve principal centres of forestbiodiversity stretching across central Africa (Hamilton 1989) Their exclusion fromthe East African highlands Ethiopia and Somalia is probably related to the recentPlio-Pleistocene uplift of the East African mountains (Baker and Wollenberg 1971Gri ths 1993) and increasing aridity of the coastal regions In these more northerlyregions canthonines are represented only by the species of the two most widespreadgenera Anachalcos Hope and Odontoloma Boheman (Ferreira 1972 Howden andScholtz 1987) The southern limits of Gyronotus distribution lies at the southernmargins of subtropical climate in South Africa Their exclusion from the more south-westerly winter and bimodal rainfall regions would be related to the northwardexpansion of the westerly system of air currents in the Pliocene (3 MY ) whichresulted in cooler climate (Deacon 1983 Deacon et al 1992) This event would beresponsible for the geographically distinctive nature (Griswold 1991 Davis 1997)and high level of endemism (Davis 1993 1994 Picker and Samways 1996) in thearthropod fauna of the Western and Eastern Cape where canthonines have radiatedinto non-forest habitats There is a wide separation between east African andsouthern African Gyronotus species in geographical phenetic and systematic terms
Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

Cladistic and biogeographical analysis of Gyronotus 1623
( gures 1 4 table 3) Between the old barriers of the broad hot dry valleys of theLimpopo and the Zambezi rivers their apparent non-occurrence may be due toeither actual absence or absence of collection It is predicted that a survey of forestson the lower eastern slopes of the East Zimbabwecentral west Mozambique high-lands might discover further Gyronotus species assuming that such forests remainextant
The tropical Gyronotus species of east Africa mostly occur in lowland forests(below 1370m in MalawimdashChapman and White 1970 below 800m in the easternarcmdashLovett 1993) These east African forests are associated with the orographiccapture of moisture by Eastern Arc and Malawi Rift mountains or with condensationof maritime or lacustrine evaporation (Gillman 1949) Therefore they would besensitive to cyclic climatic oscillation from warmer wetter climate during interglacialsand cooler drier climate during glacials of the Pleistocene There is a continuingdebate on the eŒects of Pleistocene expansion and contraction cycles in East Africanforests (Lawes 1990 Bruhl 1997 Roy 1997) Pre-Pleistocene events are probablyresponsible for most of the distribution patterns recorded for arthropods in afromon-tane forests (Griswold 1991 Bruhl 1997) although montane regions have beenidenti ed as centres of Pleistocene speciation in afromontane birds with older ances-tral species occurring in the adjoining lowlands (Roy 1997) Some within regiondivisions between Gyronotus species may also be related to recent vicariance eventsAlthough recent northsouth separation between the east African G Wmetarius andG mulanjensis is supported by their closely-similar aedeagi it is not supported byany of the inconsistent results shown by the cladograms based on phenetic data Aclose relationship between G carinatus and G glabrosus in South Africa is supportedby both character states of the aedeagi and by phenetic data However separationbetween the two species would predate the late Pleistocene (before 100000BP) asreconstruction of past forest distribution (Lawes 1990) suggests that there havebeen no recent links between the forests of Northern Province and Zululand
Acknowledgements
The authors thank Mr Riaan Stals (National Collection of Insects Pretoria)Dr Hamish Robertson (South African Museum Cape Town) Mr Malcolm Kerley(The Natural History Museum London) Dr Cornell Dudley (Museums of MalawiChichiri Blantyre) Dr Tanza Crouch (Durban Natural Science Museum Durban)Dr Frederick Gess (Albany Museum Grahamstown) Dr John Irish (NasionaleMuseum Bloemfontein) the late Dr Sebastian Endrody-Younga (TransvaalMuseum Pretoria) Dr Manfred Uhlig (Museum fur Naturkunde HumboldtUniversitat Berlin) and Dr Lothar Zerche (Deutsches Entomologisches InstitutEberswalde) for the loan of Gyronotus material Dr Paulette Bloomer and MsBettine Jansen van Vuuren kindly analysed the cladistic matrices whereas Mr HeathHull kindly produced the GIS distribution map for south-eastern South Africa andMs Marguerite Pienaar did the drawings We also thank Mr John Crawford-Brunt(SAF COL) for permission to place traps for Gyronotus in Woodbush indigenousforest
References
Baker B H and Wollenberg J 1971 Structure and evolution of the Kenya rift valleyNature 229 538ndash542
A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

A L V Davis et al1624
Barnes R F W 1990 Deforestation trends in tropical Africa African Journal of Ecology28 161ndash173
Bruhl C A 1997 F lightless insectsmdasha test-case for historical relationships of Africanmountains Journal of Biogeography 24 233ndash250
Burgess N D Clarke G P and Rodgers W A 1998 Coastal forests of EasternAfricamdashstatus endemism patterns and their potential causes Biological Journal of theLinnean Society 64 337ndash367
Cambefort Y 1991 Biogeography and evolution in I Hanski and Y Cambefort (eds)Dung Beetle Ecology (Princeton Princeton University Press) pp 51ndash67
Chapill J A 1989 Quantitative characters in phylogenetic analysis Cladistics 5 217ndash234Chapman J D and White F 1970 The Evergreen Forests of Malawi (Oxford
Commonwealth Forestry Institute)Davis A L V 1993 Biogeographical groups in a southern African winter rainfall dung
beetle assemblage (Coleoptera Scarabaeidae)mdashconsequences of climatic history andhabitat fragmentation African Journal of Ecology 31 306ndash327
Davis A L V 1994 Habitat fragmentation in southern Africa and distributional responsepatterns in ve specialist or generalist dung beetle families (Coleoptera) African Journalof Ecology 32 192ndash207
Davis A L V 1997 Climatic and biogeographical associations of southern African dungbeetles (Coleoptera Scarabaeidae s str) African Journal of Ecology 35 10ndash38
Davis A L V Scholtz C H and Harrison J Du G 1999 New and threatenedAfrotropical dung beetle taxa in the Gondwanaland tribe Canthonini (ColeopteraScarabaeidae Scarabaeinae) African Entomology 7 77ndash84
Deacon H J 1983 An introduction to the fynbos region time scales and palaeoenviromentsin H J Deacon Q B Hendey and J J Lambrechts (eds) Fynbos Palaeoecology APreliminary Synthesis South African National Scienti c Report no 75 1ndash20(Pretoria CSIR )
Deacon H J Jury M R and Ellis F 1992 Selective regime and time in R M Cowling(ed) The Ecology of Fynbos Nutrients Fire and Diversity (Cape Town OxfordUniversity Press) pp 6ndash22
Fairbairn D J 1992 The origins of allometry size and shape polymorphism in the commonwaterstrider Gerris remigis Say (Heteroptera Gerridae) Biological Journal of theLinnaean Society 45 167ndash186
Felsenstein J 1993 PHYLIP Phylogeny Inference Package Version 35 (Seattle Universityof Washington)
Ferreira M C 1972 Os escarabotildedeos de Africa (sul do Saara) Revista Entomologica deMocEuml ambique 11 5ndash1088
G illman C 1949 A vegetation-types map of Tanganyika Territory The GeographicalReview 39 7ndash37
Griffiths C J 1993 The geological evolution of east Africa in J C Lovett and S KWasser (eds) Biogeography and Ecology of the Rainforests of Eastern Africa (CambridgeCambridge University Press) pp 9ndash21
Griswold C E 1991 Cladistic biogeography of afromontane spiders Australian SystematicBotany 4 73ndash89
Halffter G 1974 Elements anciens de lrsquoentomofaune neotropical ses implications biogeog-raphiques Quaestiones Entomologicae 10 223ndash262
Halffter G and Matthews E G 1966 The natural history of dung beetles of thesubfamily Scarabaeinae (Coleoptera Scarabaeidae) Folia EntomoloAcirc gica Mexicana12plusmn 14 1ndash312
Hamilton A C 1989 Afromontane forests in H Lieth and M J A Werger (eds) Ecosystemsof the World 14B Tropical Rain Forest Ecosystems (Amsterdam Elsevier) pp 155ndash182
Howden H F and Scholtz C H 1987 A revision of the African genus OdontolomaBoheman (Coleoptera Scarabaeidae Scarabaeinae) Journal of the EntomologicalSociety of Southern Africa 50 155ndash192
InternationalUnion for the Conservation of Nature Species Survival Commission 1994IUCN Red List Categories as Approved by the 40th Meeting of the IUCN CouncilGland Switzerland (Gland The World Conservation Union)
Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)

Cladistic and biogeographical analysis of Gyronotus 1625
Lawes M J 1990 The distribution of the Samango monkey (Cercopithecus mitis erythrarchusPeters 1852 and Cercopithecus mitis labiatus I GeoŒroy 1843) and forest history insouthern Africa Journal of Biogeography 17 669ndash680
Lovett J C 1993 Eastern Arc moist forest ora in J C Lovett and S K Wasser (eds)Biogeography and Ecology of the Rainforests of Eastern Africa (Cambridge CambridgeUniversity Press) pp 33ndash55
Low A B and Rebelo A G 1996 Vegetation of South Africa Lesotho and Swaziland(Pretoria Department of Environmental AŒairs and Tourism)
Picker M D and Samways M J 1996 Faunal diversity and endemicity of the CapePeninsula South Africamdasha rst assessment Biodiversity and Conservation 5 591ndash606
Prell W L Hutson W H Williams D F BeAacute A W H Geitzenhauer K andMalfino B 1980 Surface circulation of the Indian Ocean during the last glacialmaximum approximately 18000yr BP Quaternary Research 4 309ndash336
Roy M S 1997 Recent diversi cation in African greenbuls (Pycnotidae Andropadus)supports a montane speciation model Proceedings of the Royal Society of LondonSeries B 264 1337ndash1344
Scholtz C H and Howden H F 1987 A revision of the southern African Canthonina(Coleoptera Scarabaeidae Scarabaeinae) Journal of the Entomological Society ofSouthern Africa 50 75ndash119
Somers K M 1989 Allometry isometry and shape in principal components analysisSystematic Zoology 38 169ndash173
Swofford D L 1993 PAUP Phylogenetic Analysis Using Parsimony Version 31(Champaign Illinois Natural History Survey)
Thorpe R S 1984 Coding morphometric characters for constructing distance Wagnernetworks Evolution 38 244ndash255
Van Rensburg B McGeoch M A Chown S L and Van Jaarsveld A S 1999Conservation of heterogeneity among dung beetles in the Maputaland centre ofendemism South Africa Biological Conservation 88 145ndash153
Walter H and Lieth H 1964 Klimadiagramm-Weltatlas (Jena Gustav F ischer)Wiley E O 1981 Phylogenetics The Theory and Practice of Phylogenetic Systematics (New
York J Wiley amp Sons)