citric acid assisted phytoremediation of cadmium by brassica napus l
TRANSCRIPT

Citric acid assisted phytoremediation of cadmium by Brassica napus L
Sana Ehsan a, Shafaqat Ali a,n, Shamaila Noureen a, Khalid Mahmood b, Mujahid Farid a,Wajid Ishaque b, Muhammad Bilal Shakoor a, Muhammad Rizwan a
a Department of Environmental Sciences, Government College University, Allama Iqbal Road, Faisalabad 38000, Pakistanb Nuclear Institute for Agriculture and Biology (NIAB), P.O. Box 128, Jhang Road, Faisalabad, Pakistan
a r t i c l e i n f o
Article history:Received 13 August 2013Received in revised form6 March 2014Accepted 11 March 2014
Keywords:Antioxidant enzymesBiomassCadmiumCitric acidGrowthPhytoremediation
a b s t r a c t
Phytoextraction is an eco-friendly and cost-effective technique for removal of toxins, especially heavymetals and metalloids from contaminated soils by the roots of high biomass producing plant specieswith subsequent transport to aerial parts. Lower metal bioavailability often limits the phytoextraction.Organic chelators can help to improve this biological technique by increasing metal solubility. The aim ofthe present study was to investigate the possibility of improving the phytoextraction of Cd by theapplication of citric acid. For this purpose, plants were grown in hydroponics under controlledconditions. Results indicated that Cd supply significantly decreased the plant growth, biomass, pigments,photosynthetic characteristics and protein contents which were accompanied by a significant increase inCd concentration, hydrogen peroxide (H2O2), electrolyte leakage, malondialdehyde (MDA) accumulationand decrease in antioxidant capacity. The effects were dose dependent with obvious effects at higher Cdconcentration. Application of CA significantly enhanced Cd uptake and its accumulation in plant roots,stems and leaves. Citric acid alleviated Cd toxicity by increasing plant biomass and photosynthetic andgrowth parameters alone and in combination with Cd and by reducing oxidative stress as observed byreduction in MDA and H2O2 production and decreased electrolyte leakage induced by Cd stress.Application of CA also enhanced the antioxidant enzymes activity alone and under Cd stress. Thus,the data indicate that exogenous CA application can increase Cd uptake and minimize Cd stress in plantsand may be beneficial in accelerating the phytoextraction of Cd through hyper-accumulating plants suchas Brassica napus L.
& 2014 Elsevier Inc. All rights reserved.
1. Introduction
Soil has been habitually the most common sink for wastes contain-ing heavy metals (Jadia and Fulekar, 2008). Anthropogenic activitiesare the main source of heavy metal pollution worldwide (Ali et al.,2013a; Qu et al., 2011). Among heavy metals, Cd is a highly toxic, non-essential and carcinogenic element (Alloway, 1995). Cadmium iscommonly released from electroplating, burning of fossil fuels,nickel–cadmium batteries, raw city effluents and sewage sludge etc.(Shakoor et al., 2013; Zhang et al., 2006). In plants, Cd enters mainlythrough root uptake and is transported to above ground plant parts(Farid et al., 2013; Liu et al., 2010). Higher Cd concentration in plantscaused several physiological and biochemical disorders includingreduced growth and yield, nutrient uptake, changes in chloroplastultrastructure and initiation of oxidative stress (Wang et al., 2009; Gillet al., 2011). Food chain Cd contamination is the main source of Cdentry to human especially non-smoking general population and is the
main constraint for food safety and agricultural land quality (Atafar etal., 2010). Therefore, different actions can be undertaken to remediateCd contaminated soils including in situ remediation techniques.
Among remediation techniques, plant based remediation tech-niques are becoming more widespread as these are environmentalfriendly and cost effective (Raziuddin et al., 2011). Phytoremedia-tion is a biological technique, considered for cleanup of pollutedsites because of its economical, visual advantages and extensiveapplicability (Su and Wong, 2004; Boonyapookana et al., 2005).Phytoextraction is a technique which uses plants to hyper-accumulate metals into harvestable plants. The degree of metaltranslocation from root to aerial plant parts depends upon plantspecies, metals and environmental conditions (Jadia and Fulekar,2008; Farooq et al., 2013). Phytoextraction is commonly recog-nized as a superior type of phytoremediation technologies(Weerakoon and Somaratne, 2009). However, the success ofphytoextration depends upon plant species and metal forms andavailability in the soil (Ali et al., 2013b).
Due to inherent capability of Brassica species to hyper-accumulate metals, many of the species such as Brassica oleraceaL., Brassica juncea L., Brassica rapa L. and Brassica napus L. have
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/ecoenv
Ecotoxicology and Environmental Safety
http://dx.doi.org/10.1016/j.ecoenv.2014.03.0070147-6513/& 2014 Elsevier Inc. All rights reserved.
n Corresponding author. Fax: þ92 41 9200671.E-mail address: [email protected] (S. Ali).
Ecotoxicology and Environmental Safety 106 (2014) 164–172

been used for phytoextraction strategies (Szczygłowska et al., 2011;Bareen, 2012). Recently, it has been reported that B. napus L.deserves special attention among Brassica species becauseB. napus L. is one of the most common oil sources and is cultivatedworldwide among oil seed crops (Szczygłowska et al., 2011; Park etal., 2012). B. napus L. can also be grown on contaminated soils forremediation purposes and the oil obtained from this plant can beused as a source of biodiesel (Park et al., 2008) because this plantcontains 40–44 percent of oil in its seeds (Laaniste et al., 2004). Ithas been reported that B. napus L. can be a very useful candidate forphytoextraction of many heavy metals because of its faster growth,high above ground biomass and greater metal uptake (Vamerali etal., 2010). Therefore, phytoextraction using B. napus L. is feasibleand important for decontamination of heavy metals from the soil.
Heavy metals mobility and solubility in the soil are mostimportant for their bioavailability. For this purpose, differentchelating agents have been used to enhance metal solubility inthe soil. These chelating agents include synthetic chelants such asdiethylene triamine pentaacetic acid (DTPA) and ethylene diaminetetraacetic acid (EDTA) and organic chelants such as citric acid(Sinhal et al., 2010; Szczygłowska et al., 2011; Bareen, 2012; Yeh etal., 2012; Chigbo and Batty, 2013). Synthetic chelators are capableof phytoextraction but cause ground water contamination and arenon-biodegradable (Anwer et al., 2012; Bareen, 2012). Organicacids including citric acid have higher degree of biodegradabilityand less leaching hazard as compared to synthetic chelating agents(Bareen, 2012). So synthetic chelating agents should be replacedby organic acids because organic acids reduce metal leachinghazards and are environmentally favorable due to the high rateof biodegradability (Melo et al., 2008; Wuana, et al., 2010). Amongorganic acids, citric acid significantly enhances metal solubilityand uptake by plants and can increase uptake of other nutrients byplants (Yeh et al., 2012; Freitas et al., 2013). Plant uptake of heavymetals can considerably be improved by adding smaller amount ofCA, whereas higher concentration of CA pretends intense phyto-toxic consequence in particular plant species (Turgut et al., 2004).Recently, it has been reported that CA is useful for mobilizationand phytoextraction of Cd (Sinhal et al., 2010). The application ofCA may reduce the phytotoxicity and environmental problems.However, there is a need to study the plant physiological responseunder metal stress with CA application for better understandingand use of B. napus L. for remediation purposes because plantphysiological response with CA application under metal stress hasbeen rarely studied (Najeeb et al., 2011).
Based upon the above discussion, the objectives of this study wereaimed to evaluate the ability of B. napus L. to uptake and accumulateCd alone and in combination with CA in culture media. Theparameters regarding plant growth, biomass, gas exchange attributesphotosynthesis efficiency and activities of anti-oxidant enzymes werealso studied under Cd stress with and without CA application.
2. Material and methods
2.1. Growth conditions and treatments
Mature seeds of B. napus L. genotype (Faisal canola) were thoroughly washed withdistilled water. Seeds were sown in trays containing sterilized sand of about twoinches layer and incubated at 20–22 1C in a growth chamber. The morphologicallyuniform seedlings were wrapped, after four weeks of germination, with foam at root–shoot junction and then transferred in thermopore sheets with holes in them floatingon iron tubs of 40 L capacity with complete randomized design (CRD). Hoaglandnutrient solution was used in the experiment which consists of KNO3 3000 mM; Ca(NO3)2 2000 mM; KH2PO4 100 mM; MgSO4 1000 mM; H3BO3 50 mM; MnCl2 � 4H2O0.05 mM; ZnSO4 �7H2O 0.8 mM; CuSO4 � 5H2O 0.3 mM; H2MO4 �H2O 0.10 mM andFeNa–CA 12.5 mM. Iron tubs were lined with polythene sheets. Continuous aerationwas done with the help of an air pump. The solution was changed every seven days.After two weeks of transplanting, uniform plants were treated with CdCl2 and citric
acid as T1: Control, T2: Cd (10 mM), T3: Cd (50 mM), T4: CA (2.5 mM), T5: Cd (10 mM)þCA (2.5 mM) and T6: Cd (50 mM)þCA (2.5mM) with three replications. In controlplants, no CdCl2 and CA were applied. The pH was maintained at 6.070.1 during thewhole experiment when required with 1 M H2SO4 or NaOH.
2.2. Determination of plant growth parameters
Plants were harvested after eight weeks of treatment application. Plant growthparameters including plant height, root length, number of leaves, fresh and dryweight of root, stem and leaf were determined.
2.3. Estimation of chlorophyll contents and SPAD value
Chlorophyll (a & b) and carotenoids contents were extracted in the dark with85 percent (v/v) aqueous acetone solution by continuous shaking until color hadcompletely disappeared from the leaves. The assay mixture was centrifuged at4000 rpm for 10 min. Then supernatant was removed and mixed with 85 percentaqueous acetone solution to an appropriate concentration. Light absorbance at 663,644 and 452.5 nm was determined by spectrophotometer (Metzner et al., 1965)beside the blank of a pure 85 percent aqueous acetone solution. Chlorophyll a, b,total chlorophyll and carotenoids were estimated by using subsequent calculations:
Chlorophyll a (mg/ml)¼10.3�E663–0.98�E644Chlorophyll b (mg/ml)¼19.7�E644–3:87�E663Total chlorophyll¼chlorophyll aþchlorophyll bTotal carotenoids (mg/ml)¼4.2�E452.5�{(0.0264� chl a)þ(0.426� chl b)}Finally, these pigments divisions were considered as mg g�1 fresh weight.
SPAD value was determined with meter (SPAD-502 m).
2.4. Gas exchange characteristics
After eight weeks of treatment application, Infrared Gas Analyzer (IRGA, CI-340,Analytical Development Company, Hoddesdon, England) was used for the mea-surements of photosynthetic rate (A), transpiration rate (E), stomatal conductance(Gs) and water use efficiency (A/E).
2.5. Antioxidative enzyme assays and protein content measurement
Anti-oxidant enzymes including superoxide dismutases (SOD), peroxidase(POD), catalase (CAT), ascorbate peroxidase (APX) and soluble protein in rootsand leaves were evaluated spectrophotometrically. The soluble protein content wasanalyzed according to Bradford (1976) assay using coomassie brilliant blue G-250as dye and albumin as a standard.
After eight weeks of treatment fully expanded leaves of plants and rootssamples were taken for enzymatic analysis. Leaves and roots were firstly choppedwith mortar and pestle under chilled condition with liquid nitrogen. This patternwas standardized in 0.05 M phosphate buffer (maintaining pH at 7.8) and filteredthrough four layers of muslin cloth and centrifuged for 10 min at 4 1C at 12,000g.Finally, this enzyme extract was used for quantification of SOD and POD activitiesfollowing Zhang (1992).
Catalase (CAT, EC 1.11.1.6) activity was determined according to Aebi (1984). Theassay mixture (3.0 ml) consisted of 100 μl enzyme extract, 100 μl H2O2 (300 mM)and 2.8 ml 50 mM phosphate buffer with 2 mM CA (pH 7.0). The CAT activity wasassayed by measuring the decrease in absorbance at 240 nm as a consequence ofH2O2 disappearance (ε¼39.4 mM�1 cm�1).
Ascorbate peroxidase (APX, EC 1.11.1.11) activity was assayed according to themethod of Nakano and Asada (1981). The reaction mixture consisted of 100 μlenzyme extract, 100 μl ascorbate (7.5 mM), 100 μl H2O2 (300 mM) and 2.7 ml25 mM potassium phosphate buffer with 2 mM CA (pH 7.0). The oxidation activityof ascorbate was determined by measuring the change in wavelength at 290 nm(ε¼2.8 mM�1 cm�1).
2.6. Determination of electrolyte leakage, hydrogen peroxide and malondialdehyde
Electrolyte outflow was checked by the method described by Dionisio-Sese andTobita (1998). After eight weeks of treatments, the uppermost fully expandedleaves were cut into small parts of 5 mm length and positioned in test tubescontaining 8 ml deionized and distilled water. The tubes were processed in anincubator in a water bath at 32 1C for two hours and then electrical conductivity ofinitial medium (EC1) was assessed. After this, all samples were placed in anautoclave at 121 1C for 20 min so that all electrolytes could release, then thesesamples were cooled to 25 1C then again electrical conductivity (EC2) wasmeasured and computed with the following formula:
EL¼ ðEC1=EC2Þ � 100:
The level of lipid peroxidation in leaf tissue was measured in terms of malondial-dehyde (MDA, a product of lipid peroxidation) content determined by the thiobar-bituric acid (TBA) reaction using the method of Heath and Packer (1968), with minor
S. Ehsan et al. / Ecotoxicology and Environmental Safety 106 (2014) 164–172 165

modifications as described by Dhindsa et al. (1981) and Zhang and Kirkham (1994).A 0.25 g leaf sample was homogenized in 5 ml 0.1 percent TCA. The homogenatewas centrifuged at 10,000g for 5 min. To 1 ml aliquot of the supernatant, 4 ml oftwenty percent TCA containing 0.5 percent TBA was added. The mixture was heatedat 95 1C for 30 min and then quickly cooled in an ice bath. After centrifugation at10,000g for 10 min, the absorbance of the supernatant at 532 nmwas measured andthe value for the nonspecific absorption at 600 nm was subtracted. The MDAcontent was calculated by using an extinction coefficient of 155 mM�1 cm�1.
Hydrogen peroxide was extracted by homogenizing 50 mg leaf or root tissueswith 3 ml of phosphate buffer (50 mM, pH 6.5). Then, the homogenate wascentrifuged at 6000g for 25 min. To measure H2O2 content, 3 ml of extractedsolution was mixed with 1 ml of 0.1 percent titanium sulfate in twenty percent (v/v) H2SO4 and the mixture was then centrifuged at 6000g for 15 min. The intensityof the yellow color of the supernatant was measured at 410 nm. H2O2 content wascomputed by using the extinction coefficient of 0.28 mmol�1 cm�1.
2.7. Measurement heavy metal content and statistical analysis
Plant sample (0.5 g) was taken in a 100 ml flask and then 15 ml of concentratedHNO3 was added. After mixing, the flasks were put on a hot plate whosetemperature was gradually increased up to 275 1C and dense yellow fumes appearedfrom the flask. When quantity of dense yellow fumes became low then hydrogenperoxide was added until dense yellow fumes disappeared. When samples becamecolorless, the flasks were removed from hot plate and shifted to lab where itsvolume was made up to 25 ml using distilled water and Cd contents in root, stemand leaf were determined by using the flame atomic absorption spectrometry.
The concentration of Cd in plant root, stem and leaf was measured by thefollowing formula:
Cd concentration (mg kg�1)¼reading�dilution factor/dry wt. of plant partThe accumulation of Cd in plant shoot and root was estimated by the following
formula:Cadmium accumulation (mg plant�1)¼conc. of Cd�dry wt. of plant organAll values described in this study are the means of three replicates. Analysis of
variance (ANOVA) was done by using a statistical package, SPSS version 16.0 (SPSS,Chicago, IL) followed by Tukey's post hoc test between the means of treatments todetermine the significant difference.
3. Results
3.1. Effect of cadmium and citric acid on plant growth and biomass
B. napus L. plant height, root length, leaf area and number ofleaves significantly decreased with Cd application as compared tocontrol plants (Fig. 1). Furthermore, the reduction was more obviousat higher Cd treatment (50 μM). Addition of citric acid (2.5 mM) inthe culture medium significantly increased these plant parameters ascompared to respective Cd treatments without CA addition (Fig. 1).Increase in plant growth characteristic was the highest in only CAtreatment and was the lowest in Cd50 treatment as compared tocontrol. The addition of CA improved growth characteristics byreducing the inhibitory effects of Cd at both levels of stress showingthat CA obviously ameliorated Cd toxicity symptoms in B. napus L.
The response of plant biomass parameters such as root, stemand fresh leaf and dry weights per plant is given in Table 1.Application of Cd reduced root, stem and leaves both fresh and dryweights as compared to control plants. Plant biomass parametersgradually decreased with increasing Cd treatments. The additionof CA significantly increased fresh and dry weights of leaves, stemsand roots under both 10 and 50 mM Cd treatments as compared torespective treatments without CA application.
3.2. Cadmium concentration and uptake by plant
Cadmium concentrations in shoots (leaves and stem) and rootsof B. napus L. seedlings were significantly increased when plantswere exposed to 10 and 50 mM Cd relative to control plants(Table 2). Cadmium concentration in shoots and roots gradually
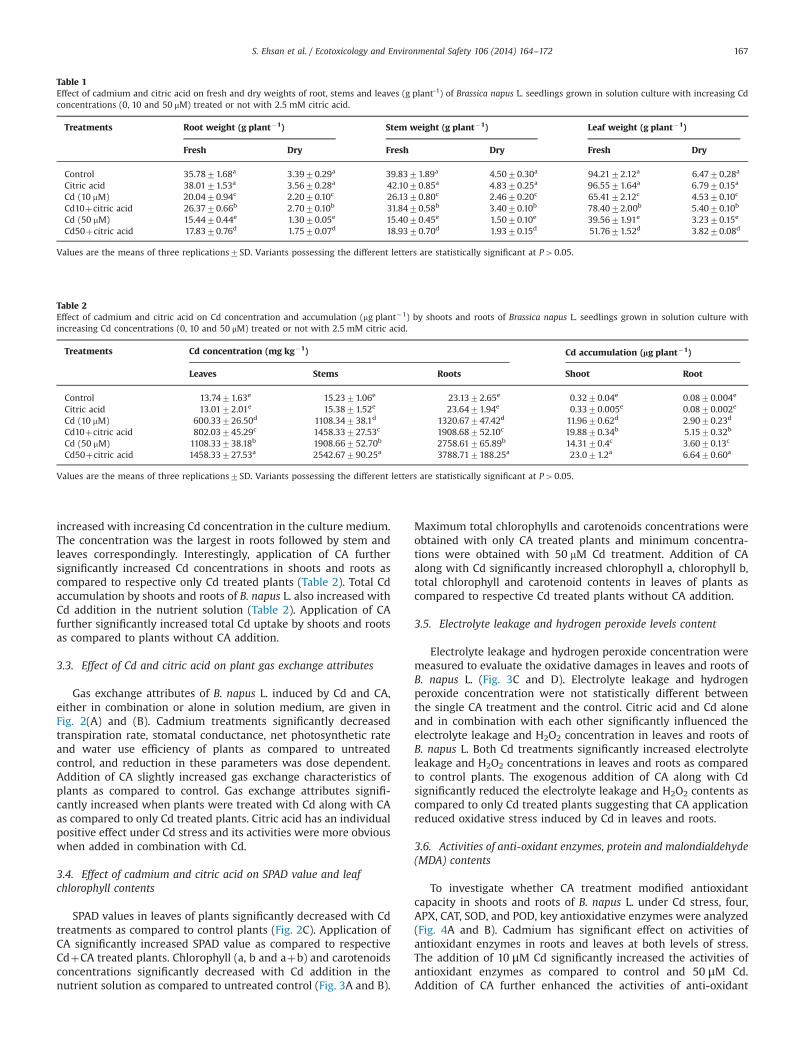
Fig. 1. Changes in root length and shoot height (A), leaf area (B) and number of leaves plant�1 (C) of Brassica napus L. seedlings grown in solution culture with different Cdtreatments (0, 10 and 50 μM) treated or not with 2.5 mM citric acid. Values are expressed as means of three replicates with standard deviations. Different letters indicate thatvalues are significantly different at Po0.05.
S. Ehsan et al. / Ecotoxicology and Environmental Safety 106 (2014) 164–172166
increased with increasing Cd concentration in the culture medium.The concentration was the largest in roots followed by stem andleaves correspondingly. Interestingly, application of CA furthersignificantly increased Cd concentrations in shoots and roots ascompared to respective only Cd treated plants (Table 2). Total Cdaccumulation by shoots and roots of B. napus L. also increased withCd addition in the nutrient solution (Table 2). Application of CAfurther significantly increased total Cd uptake by shoots and rootsas compared to plants without CA addition.
3.3. Effect of Cd and citric acid on plant gas exchange attributes
Gas exchange attributes of B. napus L. induced by Cd and CA,either in combination or alone in solution medium, are given inFig. 2(A) and (B). Cadmium treatments significantly decreasedtranspiration rate, stomatal conductance, net photosynthetic rateand water use efficiency of plants as compared to untreatedcontrol, and reduction in these parameters was dose dependent.Addition of CA slightly increased gas exchange characteristics ofplants as compared to control. Gas exchange attributes signifi-cantly increased when plants were treated with Cd along with CAas compared to only Cd treated plants. Citric acid has an individualpositive effect under Cd stress and its activities were more obviouswhen added in combination with Cd.
3.4. Effect of cadmium and citric acid on SPAD value and leafchlorophyll contents
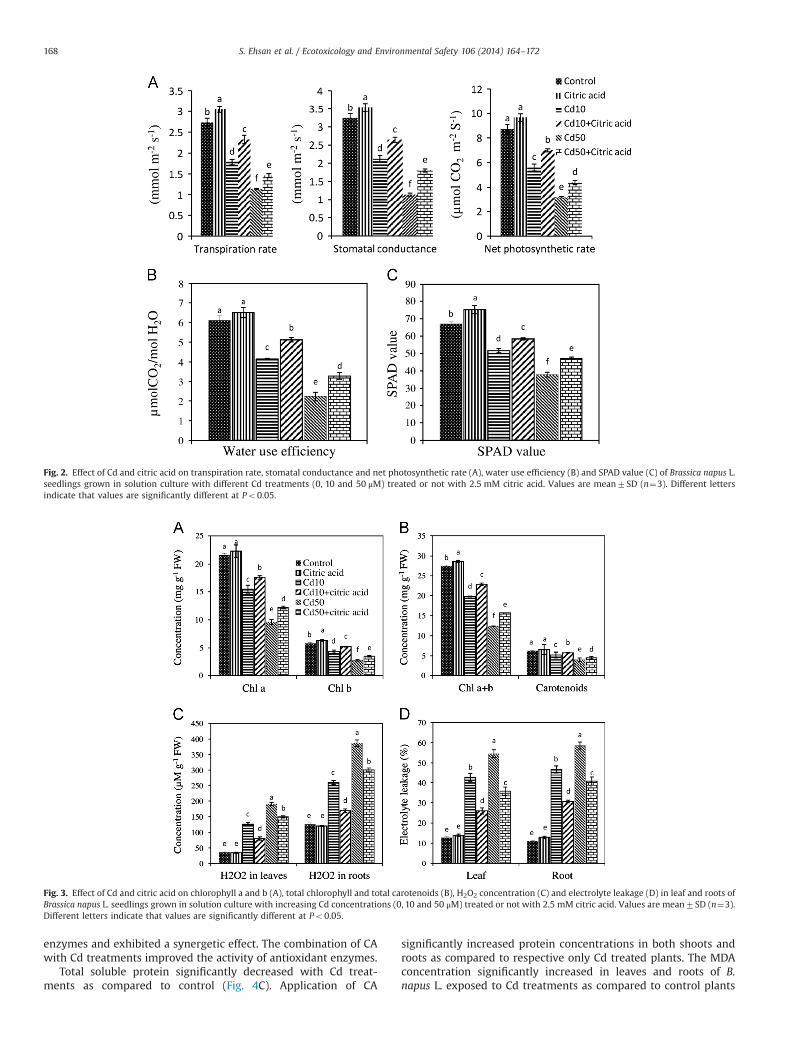
SPAD values in leaves of plants significantly decreased with Cdtreatments as compared to control plants (Fig. 2C). Application ofCA significantly increased SPAD value as compared to respectiveCdþCA treated plants. Chlorophyll (a, b and aþb) and carotenoidsconcentrations significantly decreased with Cd addition in thenutrient solution as compared to untreated control (Fig. 3A and B).
Maximum total chlorophylls and carotenoids concentrations wereobtained with only CA treated plants and minimum concentra-tions were obtained with 50 mM Cd treatment. Addition of CAalong with Cd significantly increased chlorophyll a, chlorophyll b,total chlorophyll and carotenoid contents in leaves of plants ascompared to respective Cd treated plants without CA addition.
3.5. Electrolyte leakage and hydrogen peroxide levels content
Electrolyte leakage and hydrogen peroxide concentration weremeasured to evaluate the oxidative damages in leaves and roots ofB. napus L. (Fig. 3C and D). Electrolyte leakage and hydrogenperoxide concentration were not statistically different betweenthe single CA treatment and the control. Citric acid and Cd aloneand in combination with each other significantly influenced theelectrolyte leakage and H2O2 concentration in leaves and roots ofB. napus L. Both Cd treatments significantly increased electrolyteleakage and H2O2 concentrations in leaves and roots as comparedto control plants. The exogenous addition of CA along with Cdsignificantly reduced the electrolyte leakage and H2O2 contents ascompared to only Cd treated plants suggesting that CA applicationreduced oxidative stress induced by Cd in leaves and roots.
3.6. Activities of anti-oxidant enzymes, protein and malondialdehyde(MDA) contents
To investigate whether CA treatment modified antioxidantcapacity in shoots and roots of B. napus L. under Cd stress, four,APX, CAT, SOD, and POD, key antioxidative enzymes were analyzed(Fig. 4A and B). Cadmium has significant effect on activities ofantioxidant enzymes in roots and leaves at both levels of stress.The addition of 10 μM Cd significantly increased the activities ofantioxidant enzymes as compared to control and 50 μM Cd.Addition of CA further enhanced the activities of anti-oxidant
Table 1Effect of cadmium and citric acid on fresh and dry weights of root, stems and leaves (g plant-1) of Brassica napus L. seedlings grown in solution culture with increasing Cdconcentrations (0, 10 and 50 μM) treated or not with 2.5 mM citric acid.
Treatments Root weight (g plant�1) Stem weight (g plant�1) Leaf weight (g plant�1)
Fresh Dry Fresh Dry Fresh Dry
Control 35.7871.68a 3.3970.29a 39.8371.89a 4.5070.30a 94.2172.12a 6.4770.28a
Citric acid 38.0171.53a 3.5670.28a 42.1070.85a 4.8370.25a 96.5571.64a 6.7970.15a
Cd (10 mM) 20.0470.94c 2.2070.10c 26.1370.80c 2.4670.20c 65.4172.12c 4.5370.10c
Cd10þcitric acid 26.3770.66b 2.7070.10b 31.8470.58b 3.4070.10b 78.4072.00b 5.4070.10b
Cd (50 mM) 15.4470.44e 1.3070.05e 15.4070.45e 1.5070.10e 39.5671.91e 3.2370.15e
Cd50þcitric acid 17.8370.76d 1.7570.07d 18.9370.70d 1.9370.15d 51.7671.52d 3.8270.08d
Values are the means of three replications7SD. Variants possessing the different letters are statistically significant at P40.05.
Table 2Effect of cadmium and citric acid on Cd concentration and accumulation (mg plant�1) by shoots and roots of Brassica napus L. seedlings grown in solution culture withincreasing Cd concentrations (0, 10 and 50 μM) treated or not with 2.5 mM citric acid.
Treatments Cd concentration (mg kg�1) Cd accumulation (lg plant�1)
Leaves Stems Roots Shoot Root
Control 13.7471.63e 15.2371.06e 23.1372.65e 0.3270.04e 0.0870.004e
Citric acid 13.0172.01e 15.3871.52e 23.6471.94e 0.3370.005e 0.0870.002e
Cd (10 mM) 600.33726.50d 1108.34738.1d 1320.67747.42d 11.9670.62d 2.9070.23d
Cd10þcitric acid 802.03745.29c 1458.33727.53c 1908.68752.10c 19.8870.34b 5.1570.32b
Cd (50 mM) 1108.33738.18b 1908.66752.70b 2758.61765.89b 14.3170.4c 3.6070.13c
Cd50þcitric acid 1458.33727.53a 2542.67790.25a 3788.717188.25a 23.071.2a 6.6470.60a
Values are the means of three replications7SD. Variants possessing the different letters are statistically significant at P40.05.
S. Ehsan et al. / Ecotoxicology and Environmental Safety 106 (2014) 164–172 167
enzymes and exhibited a synergetic effect. The combination of CAwith Cd treatments improved the activity of antioxidant enzymes.
Total soluble protein significantly decreased with Cd treat-ments as compared to control (Fig. 4C). Application of CA
significantly increased protein concentrations in both shoots androots as compared to respective only Cd treated plants. The MDAconcentration significantly increased in leaves and roots of B.napus L. exposed to Cd treatments as compared to control plants
Fig. 2. Effect of Cd and citric acid on transpiration rate, stomatal conductance and net photosynthetic rate (A), water use efficiency (B) and SPAD value (C) of Brassica napus L.seedlings grown in solution culture with different Cd treatments (0, 10 and 50 μM) treated or not with 2.5 mM citric acid. Values are mean7SD (n¼3). Different lettersindicate that values are significantly different at Po0.05.
Fig. 3. Effect of Cd and citric acid on chlorophyll a and b (A), total chlorophyll and total carotenoids (B), H2O2 concentration (C) and electrolyte leakage (D) in leaf and roots ofBrassica napus L. seedlings grown in solution culture with increasing Cd concentrations (0, 10 and 50 μM) treated or not with 2.5 mM citric acid. Values are mean7SD (n¼3).Different letters indicate that values are significantly different at Po0.05.
S. Ehsan et al. / Ecotoxicology and Environmental Safety 106 (2014) 164–172168
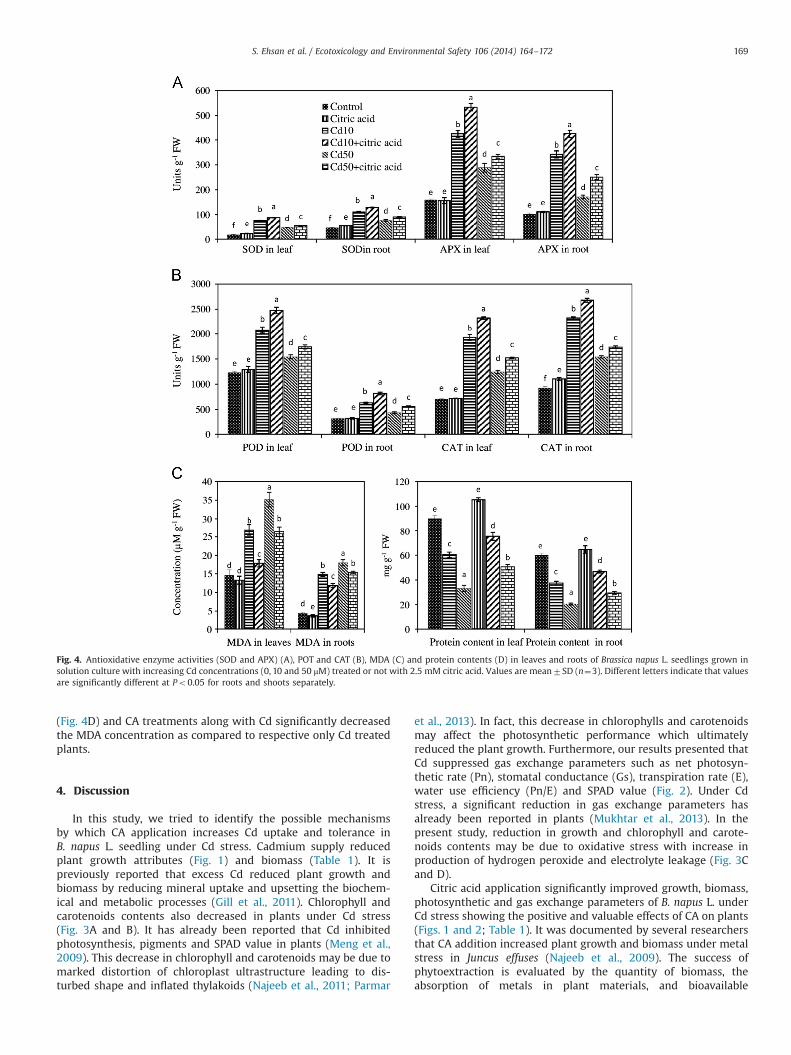
(Fig. 4D) and CA treatments along with Cd significantly decreasedthe MDA concentration as compared to respective only Cd treatedplants.
4. Discussion
In this study, we tried to identify the possible mechanismsby which CA application increases Cd uptake and tolerance inB. napus L. seedling under Cd stress. Cadmium supply reducedplant growth attributes (Fig. 1) and biomass (Table 1). It ispreviously reported that excess Cd reduced plant growth andbiomass by reducing mineral uptake and upsetting the biochem-ical and metabolic processes (Gill et al., 2011). Chlorophyll andcarotenoids contents also decreased in plants under Cd stress(Fig. 3A and B). It has already been reported that Cd inhibitedphotosynthesis, pigments and SPAD value in plants (Meng et al.,2009). This decrease in chlorophyll and carotenoids may be due tomarked distortion of chloroplast ultrastructure leading to dis-turbed shape and inflated thylakoids (Najeeb et al., 2011; Parmar
et al., 2013). In fact, this decrease in chlorophylls and carotenoidsmay affect the photosynthetic performance which ultimatelyreduced the plant growth. Furthermore, our results presented thatCd suppressed gas exchange parameters such as net photosyn-thetic rate (Pn), stomatal conductance (Gs), transpiration rate (E),water use efficiency (Pn/E) and SPAD value (Fig. 2). Under Cdstress, a significant reduction in gas exchange parameters hasalready been reported in plants (Mukhtar et al., 2013). In thepresent study, reduction in growth and chlorophyll and carote-noids contents may be due to oxidative stress with increase inproduction of hydrogen peroxide and electrolyte leakage (Fig. 3Cand D).
Citric acid application significantly improved growth, biomass,photosynthetic and gas exchange parameters of B. napus L. underCd stress showing the positive and valuable effects of CA on plants(Figs. 1 and 2; Table 1). It was documented by several researchersthat CA addition increased plant growth and biomass under metalstress in Juncus effuses (Najeeb et al., 2009). The success ofphytoextraction is evaluated by the quantity of biomass, theabsorption of metals in plant materials, and bioavailable
Fig. 4. Antioxidative enzyme activities (SOD and APX) (A), POT and CAT (B), MDA (C) and protein contents (D) in leaves and roots of Brassica napus L. seedlings grown insolution culture with increasing Cd concentrations (0, 10 and 50 μM) treated or not with 2.5 mM citric acid. Values are mean7SD (n¼3). Different letters indicate that valuesare significantly different at Po0.05 for roots and shoots separately.
S. Ehsan et al. / Ecotoxicology and Environmental Safety 106 (2014) 164–172 169

percentage of heavy metals in the rooting portion (McGrath, 1998).Increase in plant biomass may be due to enhanced nutrient uptakeby plants (Najeeb et al., 2011) or an efficient sulfate uptake andassimilation (Sun et al., 2007) or due to synthesis of phytochela-tion (PCs) in plants (Muhammad et al., 2009) or due to the abilityof plant species to detoxify Cd (Ghani, 2011). In the present study,however, increase in plant growth and biomass, photosyntheticand gas exchange parameters of B. napus L. under Cd stress may bedue to enhanced antioxidant enzymes activity (Fig. 4A and B),which decreased the production of H2O2 and electrolyte leakage(Fig. 3C and D).
In the present study, Cd induced oxidative stress as shown byincrease in hydrogen peroxide production and electrolyte leakage(Fig. 3C and D) and MDA accumulation data (Fig. 4C). Largerproduction of MDA under Cd stress has also been reported inB. napus L. seedlings (Meng et al., 2009) and in tomato (Haouari etal., 2012). Our results suggested that reactive oxygen species (ROS)might be produced in B. napus L. seedlings under Cd stress andcause lipid peroxidation, as evidenced by the increased levels ofH2O2 and MDA. By contrast, CA plus Cd addition significantlyprevented H2O2 and MDA accumulation compared with the Cdtreatment alone, suggesting a protective role of CA in preventingoxidative stress in B. napus L. Cd cannot directly produce ROS butcan bind and interfere with targets or compete for binding sitesultimately altering target protein functions which involves theproduction of ROS (Zhang et al., 2009). Plants usually faceoxidative damage when in contact with Cd and other metals(Erdei et al., 2002; Macfarlane, 2003). ROS production in plantsunder metal stress needs to be tightly regulated by efficient ROS-scavenging mechanisms. Plants have protective mechanisms ofmitigating and repairing ROS damage including the activation ofantioxidant enzymes (Mittler et al., 2004). The SOD, POD, CAT andAPX are important antioxidant enzymes in plants (Zhang et al.,2009). In the present study, the activity of SOD, POD, CAT and APXin shoots and roots significantly increased under 10 mM Cd stressthen decreased with higher Cd 50 mM concentration in thenutrient solution. This indicated overexpression of antioxidantenzymes in plants might be a powerful tool for the survival ofplants with the highest metal accumulation capacity (Haouari etal., 2012; Ali et al., 2011; Muhammad et al., 2009). Conversely,increase in antioxidant enzymes activity has been reported inB. napus L. under Cd 50 mM stress as compared to without Cdtreatments (Meng et al., 2009). In present study, the reduction inantioxidant enzymes at higher Cd applications may be due tosevere stress of ROS to antioxidant enzymes (Mishra et al., 2006).Citric acid application significantly increased the activities of theseenzymes in shoots and roots as compared to Cd only treated plants(Fig. 4A and B). It has been reported that CA helped plants toovercome oxidative stress by enhancing their antioxidant enzymeactivities under metal stress (Meng et al., 2009; Najeeb et al., 2009,2011). These results suggest that CA application scavenges oxida-tive stress, increase in H2O2 production and electrolyte leakageand MDA accumulation, in B. napus L. seedlings by enhancing theactivity of antioxidant enzymes.
Our present results showed that Cd reduced soluble protein inboth roots and leaves of B. napus L. (Fig. 4D). It might be due tomore oxidative destruction that suppressed the protein contents(Gupta et al., 2009). Application of CA increased protein contentsin both shoots and roots of B. napus L. Increase in soluble proteincontents with CA application may be due to increase in antiox-idant activity and decrease in H2O2 production and electrolyteleakage which protected plants from damage under Cd stress.
The Cd concentration and uptake by B. napus L. increased inroots and above ground parts with increasing Cd levels in thenutrient solution (Table 2). These results are in line with theprevious findings in B. napus L. (Nouairi et al., 2006; Ghani, 2011;
Park et al., 2012). Interestingly, application of CA furtherincreased Cd concentration and uptake by the plants as com-pared to control and Cd treated plants (Table 2). Citric acidapplication also increased Cd concentrations in other plants suchas J. effuses (Najeeb et al., 2009, 2011) and Sedum alfredii (Lu et al.,2013). Contrarily, the application of citric acid decreased Cduptake by maize plants (Anwar et al., 2012). In the present study,addition of CA enhanced heavy metal availability because of itschelating ability (Turgut et al., 2004; Duarte et al., 2007).Increased Cd uptake in the presence of chelator might be dueto the organometallic complex formation in the solution and atthe root surface that increased their disconnection into free Cdwhich may be certainly immersed or taken up by roots (Wuana etal., 2010). In the present study, higher Cd concentration in plantsin the presence of CA may be due to ameliorative effect of CA onroot structure and shape as proposed by Najeeb et al. (2011) in J.effuses with CA application under Cd stress. Enhanced Cd uptakemay be due to activation of ATPases in the root plasma mem-brane by the addition of chelator, citric acid, which changed thetransport of ions through membrane and increased Cd uptakethrough symplasmic or apoplasmic pathways (Han et al., 2005).The distribution of metals within plant tissues is an importantproperty that can act as an indirect indicator of detoxificationmechanism.
In present study, however, Cd concentrations were larger, morethan 1000 mg kg�1 with the highest Cd, 50 mM, level in thenutrient solution, in all parts of B. napus L. plants and furtherincreased with CA application along with Cd stress for eight weeks(Table 2). Cadmium concentration in shoots of B. napus L. has beenreported up to 555 mg g�1 dry weight only exposure for fifteendays (Nouairi et al., 2006) and about 240 mg kg�1 dry weight onlyexposure for ten days in solution (Meng et al., 2009). Cadmiumconcentration in plants varies with genotype and duration of Cdexposure to plants of B. napus L. (Ghani, 2011) and B. juncea (Gill etal., 2011). It has been reported that B. napus L. may have nolimitation of Cd uptake until Cd caused significant damage toplants (Park et al., 2012). Upper threshold of Cd accumulation in B.napus L. has not been shown and further study is required in orderto investigate the limitation of Cd accumulation in plants. Inpresent study, duration of plant exposure to Cd was long, eightweeks, and CA application enhanced antioxidant enzyme activityand gas exchange parameters which may increase Cd in shootsand roots. Moreover, B. napus L. is a hyperaccumulator and mayaccumulate a relatively high Cd in shoot and roots after a long-term exposure. Recently, Park et al. (2012) reported that more thanhalf the Cd was left in the residues during oil extraction from theseeds grown in contaminated soil so this crop is acceptable as afuture energy source. However, further research is required toevaluate the B. napus L. ability to uptake Cd from the soil especiallywith CA application and its utilization as a biofuel source.
5. Conclusion
In conclusion, our results indicated that Cd supply significantlydecreased plant growth, biomass, pigments, photosynthetic char-acteristics and protein contents which were accompanied by asignificant increase in Cd concentration, H2O2, electrolyte leakage,MDA accumulation and decrease in antioxidant capacity. Bycontrast, addition of CA alleviated Cd toxicity by reducing oxida-tive stress (H2O2 and MDA accumulations) through CA chelatingproperty and increased antioxidant capacity when compared withthe Cd treatment alone which increased plant biomass, photo-synthetic pigments and maintained gas exchange characteristics ofplants. Thus, the data indicate that exogenous CA application canminimize Cd stress in plants and increase Cd uptake and may be
S. Ehsan et al. / Ecotoxicology and Environmental Safety 106 (2014) 164–172170

beneficial in accelerating the phytoextraction of Cd through hyper-accumulating plants such as B. napus L. However, soil basedstudies under different environmental conditions are required tovalidate our results on a large scale and in real field conditions.
References
Aebi, H., 1984. Catalase in vitro. Methods Enzymol. 105, 121–126.Ali, S., Farooq, M.A., Yasmeen, T., Hussain, S., Arif, M.S., Abbas, F., Bharwana, S.A.,
Zhang, G., 2013a. The influence of silicon on barley growth, photosynthesis andultra-structure under chromium stress. Ecotox. Environ. Safe 89, 66–72.
Ali, S., Farooq, M.A., Jahangir, M.M., Abbas, F., Bharwana, S.A., Zhang, G.P., 2013b.Effect of chromium and nitrogen form on photosynthesis and anti-oxidativesystem in barley. Biol. Plant. 57, 785 (763).
Ali, S., Zeng, F., Cai, S., Qiu, B., Zhang, G.P., 2011. The interaction of salinity andchromium in the influence of barley growth and oxidative stress. Plant SoilEnviron. 57, 153–159.
Alloway, B.J., 1995. Heavy metals in soil. J. Exp. Bot., 53. Blackie Academic &Professional, London, UK, pp. 1–11.
Anwer, S., Ashraf, M.Y., Hussain, M., Ashraf, M., Jamil, A., 2012. Citric acid mediatedphytoextraction of cadmium by Maize (Zea Mays L.) Pak. J. Bot. 44 (6),1831–1836.
Atafar, Z., Mesdaghinia, A.R., Nouri, J., Homaee, M., Yunesian, M., Ahmadimoghad-dam, M., Mahvi, A.H., 2010. Effect of fertilizer application on soil heavy metalconcentration. Environ. Monitor. Assess 160, 83–89.
Bareen, F.E., 2012. Chelate assisted phytoextraction using oilseed brassicas. Environ.Pollut. 21, 289–311.
Boonyapookana, B., Parkpian, P., Techapinyawat, S., Delaune, R.D., Jugsujinda, A.,2005. Phytoaccumulation of lead by Sunflower (Helianthusannus), Tobacco(Nicotianatabacum), and Vetiver (Vetiveriazizanioides). J. Environ. Sci. Health40, 117–137.
Bradford, M.M., 1976. A rapid and sensitive method for the quantification ofmicrogram quantities of protein utilizing the principle of protein–dye binding.Anal. Biochem. 72, 248–254.
Chigbo, C., Batty, L., 2013. Effect of EDTA and citric acid on phytoremediation of Cr-B[a] P-co-contaminated soil. Environ. Sci. Pollut. Res., 1–9
Dhindsa, R.S., Plumb-Dhindsa, P., Thorne, T.A., 1981. Leaf senescence correlatedwith increased levels of membrane permeability and lipid peroxidation anddecreased levels of superoxide dismutase and catalase. J. Exp. Bot. 32, 93–101.
Dionisio-Sese, M.L., Tobita, S., 1998. Antioxidant responses of rice seedlings tosalinity stress. Plant Sci. 135, 1–9.
Duarte, B., Delgado, M., Cacador, I., 2007. The role of citric acid in cadmium andnickel uptake and translocation, in Halimioneportulacoides. Chemosphere 69,836–840.
Erdei, S., Hegedus, A., Hauptmann, G., Szalai, J., Horvath, G., 2002. Heavy metalinduced physiological changes in the antioxidative response system, In:Proceedings of the Seventh Hungarian Congress on Plant Physiology, pp. 89–90.
Farid, M., Shakoor, M.B., Ehsan, A., Ali, S., Zubair, M., Hanif, M.S., 2013. Morpholo-gical, physiological and biochemical responses of different plant species to Cdstress. Int. J. Chem. Biochem. Sci. 3, 53–60.
Farooq, M.A., Ali, S., Hameed, A., Ishaque, W., Mahmood, K., Iqbal, Z., 2013.Alleviation of cadmium toxicity by silicon is related to elevated photosynthesis,antioxidant enzymes; suppressed cadmium uptake and oxidative stress incotton. Ecotoxicol. Environ. Safe 96, 242–249.
Freitas, E.V., Nascimento, C.W., Souza, A., Silva, F.B., 2013. Citric acid-assistedphytoextraction of lead: a field experiment. Chemosphere 92, 213–217.
Ghani, A., 2011. Varietal differences in canola (Brassica napus L.) for the growth,yield and yield components exposed to cadmium stress. J. Anim. Plant Sci. 21,57–59.
Gill, S.S., Khan, N.A., Tuteja, N., 2011. Differential cadmium stress tolerance in fiveIndian mustard (Brassica juncea L.) cultivars an evaluation of the role ofantioxidant machinery. Plant Signal. Behav. 6, 293–300.
Gupta, D.K., Nicolosoa, F.T., Schetingerb, M.R.C., Rossatoa, L.V., Pereirab, L.B., Castroa,G.Y., Srivastavac, S., Tripathi, R.D., 2009. Antioxidant defense mechanism inhydroponically grown Zea mays seedlings under moderate lead stress. J.Hazard. Mater. 172, 479–484.
Han, F., Shan, X.Q., Zhang, J., Xie, Y.N., Pei, Z.G., Zhang, S.Z., Zhu, Y.G., Wen, B., 2005.Organic acids promote the uptake of lanthanum by barley roots. New Phytol.165, 481–492.
Haouari, C.C., Nasraoui, A.H., Bouthour, D., Houda, M.D., Daieb, C.B., Mnai, J., Gouia,H., 2012. Response of tomato (Solanum lycopersicon) to cadmium toxicity:growth, element uptake, chlorophyll content and photosynthesis rate. Afr. J.Plant Sci. 6, 1–7.
Heath, R.L., Packer, L., 1968. Photoperoxidation in isolated chloroplasts. I. Kineticsand stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 125,189–198.
Jadia, C.D., Fulekar, M.H., 2008. Phytoremediation: the application of vermicompostto remove zinc, cadmium, copper, nickel and lead by sunflower plant. Environ.Eng. Manag. J. 7, 547–558.
Laaniste, P., Joudu, J., Eremeev, V., 2004. Oil content of spring oilseed rape seedsaccording to fertilization. Agron. Res. 2, 83–86.
Liu, F., Tang, Y., Du, R., Yang, H., Wu, Q., Qiu, R., 2010. Rootforaging for zinc andcadmium requirement in the Zn/Cd hyperaccumulator plant Sedum alfredii.Plant 34, 112–114.
Lu, L., Tian, S., Zhang, J., Yang, X., Labavitch, J.M., Webb, S.M., Latimer, M., Brown, P.H.,2013. Improved cadmium uptake and accumulation in the hyperaccumulator Sedumalfredii: the impact of citric acid and tartaric acid. J. Zhejiang Univ. 14, 106–114.
Macfarlane, G.R., 2003. Chlorophyll a fluorescence as a potential biomarker of zincstress in the grey mangrove, Avicenniamarina. Bull. Environ. Contam. Toxicol.70, 90–96.
McGrath, S.P., 1998. Phytoextraction for soil remediation. In: Brooks, R.R. (Ed.),Plants that Hyperaccumulate Heavy Metals: Their Role in Phytoremediation,Microbiology, Archeology, Mineral Exploration and Phytomining. CAN Interna-tional, New York, pp. 261–288.
Melo, E.E.C., Nascimento, C.W.A., Accioly, A.M.A., Santos, A.C.Q., 2008. Phytoextrac-tion and fractionation of heavy metals in soil after multiple applications ofnatural chelants. Agric. Sci. 65, 61–68.
Meng, H., Hua, S., Shamsi, I.H., Jilani, G., Li, Y., Jiang, L., 2009. Cd-induced stress onthe seed germination and seedling growth of Brassica napus L. and itsalleviation through exogenous plant growth regulators. Plant Growth Regul.58, 47–59.
Metzner, H., Rau, R., Senger, H., 1965. Untersuchungen zur synchronisierbarkeiteinzelner pigmentmangel-mutanten von Chlorella. Planta 65, 186–194.
Mishra, S., Srivastava, S., Tripathi, R.D., Govindarajan, R., Kuriakose, S.C., Prasad, M.N.V., 2006. Phytochelatin synthesis and response of antioxidants duringcadmium stress in Bacopamonnieri L. Plant Physiol. Biochem. 44, 25–37.
Mittler, R., Vanderauwera, S., Gollery, M., Breusegem, F.V., 2004. Reactive oxygengene network of plants. Trends Plant Sci. 9, 490–498.
Muhammad, D., Chen, F., Zhao, J., Zhang, G., Wu, F., 2009. Comparison of EDTA- andcitric acid-enhanced phytoextraction of heavy metals in artificially metalcontaminated soil by Typha angustifolia. Int. J. Phytoremediation 11, 558–574.
Mukhtar, N., Hameed, M., Ashraf, M., Ahmed, R., 2013. Modifications in stomatalstructure and function in Cenchrus ciliaris L. and Cynodon dactylon (L.) Pers. inresponse to cadmium stress. Pak. J. Bot. 45, 351–357.
Najeeb, U., Jilani, G., Ali, S., Sarwar, M., Xu, L., Zhou, W., 2011. Insights into cadmiuminduced physiological and ultra-structural disorders in Juncus effusus L. and itsremediation through exogenous citric acid. J. Hazard. Mater. 186, 565–574.
Najeeb, U., Xua, L., Ali, S., Jilani, G., Gong, H.J., Shen, W.Q., Zhou, W.J., 2009. Citricacid enhances the phytoextraction of manganese and plant growth by alleviat-ing the ultrastructural damages in Juncus effuses L. J. Hazard. Mater. 170,1156–1163.
Nakano, Y., Asada, K., 1981. Hydrogen peroxide scavenged by ascorbate specificperoxidase in spinach chloroplasts. Plant Cell Physiol. 22, 867–880.
Nouairi, I., Ammar, W.B., Youssef, N.B., Daoud, D.B.M., Ghorbal, M.H., Zarrouk, M.,2006. Comparative study of cadmium effects on membrane lipid compositionof Brassica juncea and Brassica napus leaves. Plant Sci. 170, 511–519.
Park, H., McGinn, P.J., Morel, F.M.M., 2008. Expression of cadmium carbonicanhydrase of diatoms in seawater. Aquat. Microb. Ecol. 51, 183–193.
Park, J., Kim, J.Y., Kim, K.W., 2012. Phytoremediation of soil contaminated withheavy metals using Brassica napus. Geosyst. Eng. 15, 10–18.
Parmar, P., Kumari, N., Sharma, V., 2013. Structural and functional alterations inphotosynthetic apparatus of plants under cadmium stress. Bot. Stud. 54, 45.
Qu, J., Lou, C.Q., Yuan, X., Wang, X.H., Cong, Q., Wang, L., 2011. The effect of sodiumhydrogen phosphate/citric acid mixtures on phytoremediation by alfalfa &metals availability in soil. J. Soil Sci. Plant Nutr. 11 (2), 85–95.
Raziuddin, Farhatullah, Hassan, G., Akmal, M., Shah, S.S., Mohammad, F., Shafi, M.,Bakht, J., Zhou, W., 2011. Effects of cadmium and salinity on growth andphotosynthesis parameters of brassica species. Pak. J. Bot. 43, 333–340.
Shakoor, M.B., Ali, S., Farid, M., Farooq, M.A., Tauqeer, H.M., Iftikhar, U., Hannan, F.,Bharwana, S.A., 2013. Heavy metal pollution, a global problem and itsremediation by chemically enhanced phytoremediation: a review. J. Biol.Environ. Sci. 3, 12–20.
Sinhal, V.K., Srivastava, A., Singh, V.P., 2010. EDTA and citric acid mediatedphytoextraction of Zn, Cu, Pb and Cd through marigold (Tageteserecta). J.Environ. Biol. 31, 255–259.
Su, D.C., Wong, J.W.C., 2004. Selection of Mustard Oilseed Rape (Brassica juncea L.)for phytoremediation of cadmium contaminated soil. Bull. Environ. Contam.Toxicol. 72, 991–998.
Sun, Q., Ye, Z.H., Wang, X.R., Wong, M.H., 2007. Cadmium hyperaccumulation leadsto an increase of glutathione rather than phytochelatins in the cadmiumhyperaccumulator Sedum alfredii. J. Plant Physiol. 164, 1489–1498.
Szczygłowska, M., Piekarska, A., Konieczka, P., Namiesnik, J., 2011. Use of brassicaplants in the phytoremediation and biofumigation processes. Int. J. Mol. Sci. 12,7760–7771.
Turgut, C., Pepe, M.K., Cutright, T.J., 2004. The effect of EDTA and citric acid onphytoremediation of Cd, Cr, and Ni from soil using Helianthus annuus. Environ.Pollut. 131, 147–154.
Vamerali, T., Bandiera, M., Mosca, G., 2010. Field crops for phytoremediation ofmetal- contaminated land. Environ. Chem. Lett. 8, 1–17.
Wang, C., Sun, Q., Wang, L., 2009. Cadmium toxicity and phytochelatins productionin a rooted-submerged macrophyte Vallisneria spiral is exposed to lowconcentrations of cadmium. Environ. Toxicol. 24, 271–278.
Weerakoon, S.R., Somaratne, S., 2009. phytoextractive potential among mustard(brassica juncea) genotypes in srilanka. Ceylon. J. Sci. Biol. Sci. 38 (2), 85–93.
Wuana, R.A., Okieimen, F.A., Imborvungu, J.A., 2010. Removal of heavy metals froma contaminated soil using organic chelating acids. Int. J. Environ. Sci. Tech. 7 (3),485–496.
S. Ehsan et al. / Ecotoxicology and Environmental Safety 106 (2014) 164–172 171

Yeh, T.Y., Lin, C.F., Chuang, C.C., Pan, C.T., 2012. The Effect of Varying Soil OrganicLevels on Phytoextraction of Cu and Zn uptake, enhanced by chelator EDTA,DTPA, EDDS and Citric Acid, in Sunflower (Helianthus annuus), Chinese Cabbage(Brassica campestris), Cattail (Typha latifolia), and Reed (Phragmites commu-nis). Environ. Anal. Toxicol. 2, 5.
Zhang, J., Kirkham, M.B., 1994. Drought-stress induced changes in activities ofsuperoxide dismutase, catalase and peroxidases in wheat leaves. Plant CellPhysiol. 35, 785–791.
Zhang, G., Wu, K., Dong, Q., Dai, F., Chen, F., Yang, J., 2006. Cd stress in higher plants.In: KhanandSamiullah, N.A. (Ed.), Cd toxicity and Tolerance in Plants. Narosapublishers, pp. 87–101.
Zhang, X.Z., 1992. The measurement and mechanism of lipid peroxidation andSOD, POD and CAT activities in biological system. In: Zhang, X.Z. (Ed.),Research Methodology of Crop Physiology. Agriculture Press, Beijing,pp. 208–211.
Zhang, F.Q., Zhang, H.X., Wang, G.P., Xu, L.L., Shen, Z.G., 2009. Cadmium-inducedaccumulation of hydrogen peroxide in the leaf apoplast of Phaseolus aureus andVicia sativa and the roles of different antioxidant enzymes. J. Hazard. Mater.168, 76–84.
S. Ehsan et al. / Ecotoxicology and Environmental Safety 106 (2014) 164–172172