chemosynthetic bacterial mats at cold hydrocarbon seeps, gulf of mexico continental slope
TRANSCRIPT

Org. Geochem. Vol. 20, No. 1, pp. 77-89, 1993 0146-6380/93 $6.00 + 0.00 Printed in Great Britain. All rights reserved Copyright © 1993 Pergamon Press Ltd
Chemosynthetic bacterial mats at cold hydrocarbon seeps, Gulf of Mexico continental slope
ROGER SASSEN, 1 HARRY H. ROBERTS, 2 PAUL AHARON, 3 JOHN LARKIN, 4 ELIZABETH W. CHINN 3
and ROBERT CARNEY 5
L Geochemical and Environmental Research Group, Texas A&M University, College Station, TX 77845, ~Coastal Studies Institute and Department of Oceanography and Coastal Studies, Louisiana State University, Baton Rouge, LA 70803, 3Department of Geology and Geophysics, Louisiana State University, Baton Rouge, LA 70803, 4Department of Microbiology, Louisiana State University, Baton Rouge, LA 70803 and 5Coastal Ecology Laboratory, Center for Wetland Resources, Louisiana State
University, Baton Rouge, LA 70803, U.S.A.
(Received 27 December 1991; accepted in revised form 10 March 1992)
A~traet--White and pigmented filamentous bacterial mats dominated by several undescribed species of Beggiatoa were sampled during research submersible dives to cold hydrocarbon seep sites on the upper continental slope off Louisiana (130-550 m). Mats occur at the interface between reducing sediments and the oxygenated water column. They are localized at sea floor features related to seepage of biogenic methane and crude oil, but there is little evidence that the organisms utilize the hydrocarbons directly. Granules of elemental sulfur (S °) are visible within cells of Beggiatoa, and mat material is characterized by high contents of S O (up to 193,940 ppm). The Beggiatoa biomass is isotopically light (6 t3C = -27.9%0 PDB). Our geochemical data suggest that the Beggiatoa species are part of a complex bacterial assemblage in cold seep sediments. They oxidize H2S derived from the bacterial sulfate reduction that accompanies bacterial hydrocarbon oxidation when 02 is depleted in sediments, and fix isotopically light carbon from CO 2 that is the result of bacterial hydrocarbon oxidation. Beggiatoa mats appear to retard loss of hydrocarbons to the water column by physically retaining fluids in sediments, a function that could enhance production by other bacteria of the H2S and CO 2 needed by Beggiatoa.
Key words--Gulf of Mexico, Beggiatoa, bacterial mats, hydrocarbons, hydrogen sulfide, elemental sulfur
INTRODUCTION
Active crude oil and gas seepage is widely distributed across the Gulf of Mexico continental slope offshore Louisiana over shallow salt diapirs and near faults (Anderson et al., 1983; Brooks et al., 1984, 1986; Kennicutt et al., 1988). Geochemical correlation of crude oils indicates that significant vertical migration occurs from subsurface reservoirs to near-surface sediments in the Green Canyon area (Kennicutt et al., 1988). The large volumes of hydrocarbons in near- surface sediments in Green Canyon result in an unusual geochemical environment.
Thermogenic gas hydrates accumulate in near- surface sediments because of cold sea bot tom tem- peratures and the high pressures concomitant with deep water (Brooks et al., 1984, 1986; Davidson et al., 1986). Microbial oxidation of hydrocarbons in near surface sediments leads to formation of large volumes of authigenic carbonate minerals, often with ex- tremely light &~3C values (Anderson et al., 1983; Roberts et al., 1989, 1990a, b). This hydrocarbon- derived authigenic carbonate rock is present in suffi- ciently large volume to significantly influence sea bot tom topography over salt dome crests and near deep-seated faults (Behrens, 1988; Roberts et al.,
1989, 1990a, b; Kennicutt and Brooks, 1990). The authigenic carbonate presently forming over shallow salt sheets and diapirs on the Gulf slope is believed to represent the embryonic stage of development of older carbonate cap rocks with extremely light 6 ~3C values found over salt diapirs onshore in Texas and Louisiana (Sassen, 1987; Sassen et al., 1991).
Chemosynthetic tube worms that oxidize HzS were first discovered at deep-water hydrothermal vent environments on the Galapagos Rift and East Pacific Rise (Corliss et al., 1979). Similar organisms have since been discovered at "cold" hydrocarbon seeps of the deep-water Gulf of Mexico continental slope where HzS is abundant (Kennicutt et al., 1985; Brooks et al., 1987; Rosman et al., 1987; McDonald et al., 1989, 1990a, b, c, d). In particular, colonies of large tube worms (up to 2 m in length) with bacterial symbionts that oxidize H 2 S have been documented by MacDonald et al. (1989) on the crest of a gas hydrate mound on the Gulf slope at water depths in the 530-550m range (Bush Hill). MacDonald et al. (1990c) also describe an occurrence of seep mussels with methanotrophic bacterial symbionts at 650m water depth that surround a brine-filled pockmark that they interpreted as the result of gas erosion on the Gulf slope. Moreover, two distinct assemblages of
77

78 ROGER SASSEN et al.
Vesicomyid and Lucinid clams with bacterial sym- bionts that oxidize H2S have been described on the Gulf slope (MacDonald et al., 1990d).
Less attention, however, has been given to the geochemical significance of bacterial mats of Beggia- toa that are ubiquitous at cold hydrocarbon seeps of the Gulf slope (MacDonald et al., 1989, 1990c; Roberts et al., 1989, t990a, b; Roberts and Neu- rauter, 1990). Similar bacterial mats have been ob- served in association with hydrocarbon seeps in other areas, including offshore California (Davis and Spies, 1980) and in the North Sea (Hovland and Thomsen, 1989). Larkin and Polman (1988) note that this large, gliding filamentous bacterium generally oxidizes H 2 S and produces internal granules of S O which can be further oxidized to sulfate. Zobell (1963) was among the first to emphasize the important role of Beggiatoa and other H2 S oxidizing bacteria in the geochemical sulfur cycle of marine sediments. Most Beggiatoa require CO 2 for growth (Strohl et al., 1981; Nelson and Jannasch, 1983). Nevertheless, from the micro- biologic standpoint, there remains uncertainty as to the energy and carbon sources utilized by unde- scribed species of Beggiatoa in cold hydrocarbon seeps of the Gulf slope because these particular organisms have yet to be cultured in the laboratory. The main objectives of this preliminary study, there- fore, are (1) to better document the occurrence of Beggiatoa mats on the Gulf slope, and (2) to provide new geochemical insight to potential utilization of H2 S and CO2 by Beggiatoa.
EXPERIMENTAL
Samples and geologic setting
The Pisces H research submersible was used to obtain samples from cold hydrocarbon seep environ- ments of the Gulf slope at water depths in the 130-545 m range. Dive locations in Blocks 53, 140, and 185 of the Green Canyon (GC) protraction area offshore Louisiana are shown in Fig. 1.
Dives 88-21 and 88-23 focused on a highly-faulted area in the northeast corner of GC 53 with outcrop- ping authigenic carbonate rock, gas seeps, and Beg- giatoa mats. We sampled Beggiatoa mat material. The dives covered part of a discontinuous outcrop (about 3 km across) of authigenic carbonate with light ~13C derived from long-term microbial oxi- dation of hydrocarbons (Roberts et al., 1990a, b). Because of relatively shallow water depths on the upper Gulf slope (as shallow as 130 m on carbonate highs), carbonate hardgrounds are abundantly en- crusted by heterotrophic organisms of the deeper photic zone including octocorals, coralline algae, solitary corals, sponges, brachiopods, and crinoids (Roberts et al., 1990a). The area overlies the flanks of shallow salt near a subsurface structural trap contain- ing crude oil on GC 52-53 (Cook and D'Onfro, 1991).
We collected samples of Beggiatoa mat material, a gas bubble train issuing to the water column, and outcropping authigenic carbonate rock in southern GC 140 during 2 dives (88-24 and 88-25). The GC 140 cold seep sites (230-530m) are over the highly- faulted crest of a salt diapir (Cook and D'Onfro, 1991). The crestal area is overlain by a discontinuous outcrop of authigenic carbonate > 4 km across (with relief up to > 10m) exposed by sea floor erosion (Roberts et al., 1989). Various carbonate minerals are present (aragonite, magnesium-calcite, and dolomite) that show a wide range of di~3C values ( - 1 6 . 6 to -53.9%o PDB) consistent with origins from micro- bial oxidation of both biogenic and thermogenic hydrocarbons (Roberts et al., 1989). The authigenic carbonate appears to be related to long-term vertical migration of hydrocarbons along the flanks of the salt diapir. Kennicutt et al. (1988) showed that crude oil from reservoirs of Jolliet Field underlying GC 184-185 (2000-3000m), and crude oil in sea-floor cores and sea-surface slicks are geochemically related.
Mat material with oil-stained sediments and authigenic carbonate were sampled on Bush Hill in GC 185 at water depths in the 530-570m range (Kennicutt et al., 1985). Bush Hill is an isolated sea-floor mound with a relief of about 40m (MacDonald et al., 1989). Cores of sediments show oil staining, gas hydrates, and authigenic carbonate (Neurauter and Bryant, 1990). The origin of ther- mogenic hydrocarbons (MacDonald et al., 1989) at Bush Hill is related to vertical migration along faults near the eastern edge of the Jolliet Field which are described by Cook and D'Onfro (1991). Bush Hill is one of the best documented examples (MacDonald et al., 1989) of the roughly 45 chemosynthetic com- munities of tube worms, mussels, and clams thus far identified on the Gulf of Mexico continental slope (MacDonald et al., 1990d).
Methods
The robot arm of the research submersible was used to collect push-cores of surficial sediments (gen- erally <0.3 m) in bacterial mat areas, and to collect samples of outcropping authigenic carbonate rock. A suction sampling device on the research submersibles was also used to collect bacterial mats and associated sediments.
Sediment samples intended for analysis were transferred to metal cans intended for geochemical samples, treated with bactericide (lead azide), sealed, and then refrigerated until analyzed. Total organic carbon (TOC) contents of sediment samples were measured using a Rock-Eval II with TOC module. TOC measurements were only performed on samples that had already been solvent extracted to remove crude oil and elemental sulfur (S°). The interference of S O during pyrolysis has been described (Sassen and Chinn, 1989).
The methods used for extraction and separation of C~5+ extractable organic matter (EOM) into

Chemosynthetic bacterial mats at cold hydrocarbon seeps 79
saturated hydrocarbon (SAT), aromatic hydro- carbon (AROM), resin (NSO), and asphaltene (ASPH) fractions, for Ct0+ chromatography of satu- rated hydrocarbons, and for determination of 613C compositions of EOM are described elsewhere (Sassen et al., 1988). The method used for determi-
nation of carbonate 613C is described by Aharon et al. (1991). Concentrations of S O in Beggiatoa mat and in sediment samples were estimated during sol- vent extraction of samples by reaction with copper foil, and are reported as ppm relative to sample weight.
OF MEXICO
( ms. TEXAS ..i LOUISIANA_
CANYON
o SO
GREEN CANYON AREA o~
Fig. I. Regional map (top) showing simplified bathymetry (m) across the Gulf of Mexico. Note the research submersible area of operations (shaded) in the northwestern quadrant of the Green Canyon protraction area. Another map (below) shows detailed bathymetry (m) and identifies dive areas in which samples were collected (boxes). Individual blocks in the protraction area are 4.83 km (3 miles) on an edge.

80 ROGER SASSEN et al.
C1~C6+ hydrocarbons in headspace of canned samples were measured as described previously (Sassen et al., 1988). Results are reported as ppm relative to sample volume. The C~-C6÷ hydrocarbon concentrations are minimum values since some de- gassing occurred before the samples could be brought to the surface and canned. A large sample (about 1200 ml) of a gas bubble train was collected just above a sea-floor vent. The gas was allowed to displace sea water from a bell jar held by the robot arm, which was then sealed under water by a diver when the research submersible surfaced.
Conventional transmitted-light microscopy and scanning electron microscopy (SEM) was used to investigate selected samples of bacteria, authigenic carbonate, and solid crude oil residues from cold seep environments. SEM samples were dried at reduced pressure, coated with gold, and studied with a JEOL U.S.A. T300 scanning electron microscope in con- junction with a Tracor Northern TN 2000 energy dispersive X-ray system.
RESULTS AND DISCUSSION
Observations of the Beggiatoa habitat
Beggiatoa mats up to several meters across are ubiquitous in cold hydrocarbon seep environments at water depths in the 130-545 m range across the upper Gulf slope study areas (Fig. 2). Observations during research submersible dives suggest that Beggiatoa mats are more widely distributed than chemosyn- thetic macrofauna such as tube worms, mussels, and clams. For example, we observed Beggiatoa mats in water depths as shallow as 130 m, within the lower photic zone where carbonate-encrusting hetero- trophic organisms are abundant and chemosynthetic macrofauna are absent. Chemosynthetic macrofauna occur most commonly at Gulf cold seep sites in deeper waters (>350 m) below the photic zone (MacDonald et al., 1990d).
Beggiatoa appear to be so widely distributed because they are well adapted to colonization of the many small or transient cold seep sites that provide energy and carbon sources. During one submersible dive near Bush Hill (dive 88-27), a "rain" of white bacterial filaments was observed in the water column, suggesting that bacterial filaments are lifted into the water column by hydrocarbon venting or sediment erosion, and thus are dispersed to new sites by deep-water currents. Moreover, we observed onboard the ship that freshly collected Beggiatoa filaments held at sea-floor temperatures can glide short dis- tances along the sediment surface to optimize nutri- ent sources.
There is an obvious spatial association of Beggia- toa mats with deep-water Gulf sea floor physical features related to hydrocarbon seepage (Roberts et al., 1990a, b; Roberts and Neurauter, 1990). Such sea floor features include narrow linear fractures in
sediment (mm to cm wide), steeply-dipping fault scarps in sediments, burrows of unidentified animals, pockmarks or craters (meters to tens of m across) formed by episodic gas erosion (Prior et al., 1989), mud vents that episodically discharge hyrocarbons and brines to the water column, gas hydrate mounds, and fault-vents in outcropping masses (< 1 m to km across) of authigenic carbonate rock.
The most direct visual evidence of hydrocarbons at Beggiatoa mats consists of small bubble trains of gas, sometimes in linear arrays, that continuously or episodically enter the water column. A sample of bubble train gas recovered from the water column during dive 88-25 on GC 140 consists of pure biogenic C~ with a 613C value of -66.3%o PDB. The coring process itself frequently increases the rate of gas bubble discharge into the water column. In addition, we did not observe crude oil to enter the water column unless oil-stained sediments were mechanically disrupted by sampling activities. This observation has also been emphasized by MacDonald et al. (1989). If no barriers to migration existed at the interface between sediment and water, buoyancy differences should often result in continuous seepage of hydrocarbons from the sediment surface to the water column. It could be argued that Beggiatoa mats are physical barriers that retain hydrocarbons and other fluids in sedi- ments. This function could serve to enhance pro- duction by other bacteria of the H 2 S and CO2 needed by Beggiatoa.
Although our data could be interpreted to suggest a direct association between Beggiatoa and hydro- carbon seepage, this association nevertheless appears to be indirect. There is as yet no unambiguous evidence as that Beggiatoa utilize C~ or other hydro- carbons. Strohl et al. (1981) and Nelson and Jannasch (1983) suggest utilization of CO2 as a carbon source. The 6uC values of some authigenic carbonate rock samples from our dive localities provide insight to a source of CO2. For example, the 6uC of the authi- genic carbonate outcrop near one Beggiatoa mat we sampled on GC 140 (88-25-1) is -31.9%o PDB. Two samples of authigenic carbonate rock from GC 185 show 6J3C values of -23.3%o PDB (88-27-3) PDB and -21.8%o PDB (88-27-4). The large volumes of authigenic carbonate with light 6~3C at cold hydro- carbon seeps record long-term CO2 production from microbial hydrocarbon oxidation (Behrens, 1988; Roberts et al., 1989, 1990a, b; Kennicutt and Brooks, 1990). More directly, measurements of dissolved in- organic carbon in the water column during dives over cold hydrocarbon seeps at GC 53 and 143 (Aharon et al., 1991) imply that CO2 with a calculated mean 613C of -27.1%o PDB is available for utilization by Beggiatoa. It should be emphasized that our research submersible observations do not suggest a preferen- tial distribution of Beggiatoa occurrences with respect to either biogenic gas or oil-stained sediments. This should not be surprising if Beggiatoa utilize CO2

Fig. 2. Photographs of Beggiatoa mats at cold hydrocarbon seeps in Green Canyon. At top, scattered white and pigmented Beggiatoa mats can be seen on the sediment surface behind a colony of tube worms. At bottom, a large pigmented Beggiatoa mat is seen that drapes one edge of a buried gas hydrate mass. Two
small laser dots near the center of the images are 10 cm apart, and provide scale.
81

0 Z
~E ~E
.=_ .E
0 ~
o ' ~
.=
L~
82

Chemosynthetic bacterial mats at cold hydrocarbon seeps 83
derived from microbial oxidation of hydrocarbons instead of the hydrocarbons themselves.
Beggiatoa mats are ubiquitously associated with H2S, the odor of which is obvious during sample handling. Moreover, cold hydrocarbon seep sedi- ments and authigenic carbonate rock associated with lush Beggiatoa occurrences are sometimes colored black by hydrous iron sulfides. Although most Beg- giatoa mats are white, some are pigmented gray or black, also apparently because of hydrous iron sulfides. The H z S in cold seep sediments is suggested to result from the microbial sulfate reduction that often accompanies microbial hydrocarbon oxidation in cold hydrocarbon seeps when available 02 is depleted (Sassen, 1980). Although orange and yellow Beggiatoa mats are observed over oil-stained sedi- ment. there is as yet no clear explanation for the pigmentation.
Conventional photomicrographs of white Beggia- toa filaments from sediments near gas vents and an outcrop of authigenic carbonate rock on GC 140 (sample 88-25-1) provide insight to their dimensions (Fig. 2). The bacterial filaments are large. The largest bacterial filament shown is about 0.2 mm in diameter and could be several cm in length. The work of Larkin and Henk (1989) shows that Beggiatoa filaments are hollow. Differences in size and mor- phology between Beggiatoa filaments suggest that several undescribed species are present (Fig. 3). Gran- ules are sometimes visible in transmitted light within the cells of Beggiatoa samples, and they are inter- preted to consist of S °.
Geochemistry of Beggiatoa and associated sediments
We analyzed Beggiatoa and associated sediments to increase our insight to the nutrients utilized by these organisms, and the impact of bacterial life processes on sediment geochemistry. It should be noted that sampling pure Beggiatoa for geochemical analysis is often difficult because core samples of thin surficial mats are dominated by sediment, and even suction sampling devices usually recover sediment along with bacterial filaments. Some of our samples also contain significant amounts of EOM from crude oil. Nevertheless, patterns emerge from measure-
ments of low molecular weight (LMW) hydrocar- bons, EOM, (~13C, and S O (Table 1).
An unusually thick Beggiatoa mat from the sedi- ment surface near outcropping authigenic carbonate rock on GC 140 (dive 88-25), collected using a suction device, provides our best sample. TOC of the bulk- sample extraction residue is 9.37%, the concentration of EOM is 4288 ppm, and the EOM/TOC ratio is 0.05 (Table 1). Since the absence of a naphthenic hump in a Cl0+ chromatogram of the EOM saturated hydrocarbon fraction suggests that crude oil hydro- carbons are absent or in low abundance, the bulk of EOM appears to be bacterial lipids. The concen- tration of S o in the bulk sample is 193,940 ppm, the S°/TOC ratio is 2.07, and the S°/EOM ratio is 45.23 (Table 1). The 6J3C of Beggiatoa filaments further separated by suspension in water from an aliquot of the original sample is -27.9%0 PDB (Table 1).
Beggiatoa filaments are believed to represent the bulk of EOM-free TOC (0.16-0.57%) in 3 other samples from GC 53 and 140 (Table 1). Ct is the only LMW hydrocarbon detected in the canned samples from this group (Table 1). The low EOM/TOC ratios, as well as absence of naphthenic humps in Cl0+ chromatograms of saturated hydrocarbons, suggests that crude oil is absent or in low abundance. EOM is in the 87-395 ppm range, and EOM/TOC ratios are in the 0.03-0.07 range (Table 1). The EOM is inter- preted to consist largely of bacterial lipids. It was not possible to physically isolate sufficient Beggiatoa filaments from these samples for 6~3C analysis, but the 613C values of EOM are in the -27.5 to -30.9%0 PDB range (Table 1). Lipids from Beggiatoa, and possibly other unidentified bacteria in associated sediments, thus appear to be an in situ source of isotopically-light EOM in samples where crude oil is absent or in low abundance. The concentration of S O ranges from 90 to 668 ppm, S°/TOC ratios are in the 0.05-0.37 range, and S°/EOM ratios are in the 0.66-8.58 range (Table 1).
Visibly oil-stained samples of sediment (samples 88-21-2, 88-27-1, and -2) and authigenic carbonate (samples 88-27-3, and -4) from the vicinity of white and pigmented Beggiatoa mats were also analyzed (Table 1). Three samples of mat material were canned
Table 1. Results of geochemical analyses of Beggiatoa mat material from cold hydrocarbon seeps in Green Canyon
Sample Location C,, ppm C 2, ppm % TOC* EOM, ppm 6t3C EOM EOM/TOC S °, ppm S°/TOC S°/EOM
Crude oil absent or in low abundance 88-25-1 GC 140 t t 9.37 4288 -27.95 0.05 193,940 2.07 45.23 88-21-1 GC 53 2637 0.0 0.18 95 -27.5 0.05 668 0.37 7.03 88-23-1 GC 53 t t 0.29 87 -30.9 0.03 746 0.26 8.58 88-24-1 GC 140 491 0.0 0.57 395 27.7 0.07 259 0.05 0.66
Crude oil present 88-21-2 GC 53 86 0.0 0.32 7545 -28.2 2.36 27,767 8.68 3.68 88-27-1 GC 185 3528 16.7 3.97 24,807 -27.1 0.62 667 0.02 0.03 88-27-2 GC 185 37 0.0 1.08 23,011 -27 .4 2.13 2079 0.19 0.09 88-27-3 GC 185 + t 0.74 12,188 -27 .0 1.65 1349 0.18 0.11 88-27-4 GC 185 ~" t 0.77 10,540 26.8 1.37 756 0.01 0.07
*TOC OF EOM-free samples. tSamples not canned at collection. ~:Determined on bacterial biomass.

84 ROGER SASSEN el al.
and analyzed for LMW hydrocarbons. Although C~ is the dominant LMW hydrocarbon in the 3 canned samples from this group, some C2 occurs in 1 sample (Table 1). The absence or low abundance (below detection limits) of the oil-related C~E6+ hydrocar- bons is assumed to reflect microbial oxidation effects. EOM-free TOC values of oil-stained samples are in the 0.32-3.97% range. EOM is in the 7545-24,807 range, and EOM/TOC ratios are in the 0.62-2.36 range (Table 1). C~o+ chromatograms of saturated hydrocarbon fractions all show a naphthenic signa- ture lacking n-alkanes and isoprenoids as illustrated by the example in Fig. 4. The 613C values of EOM from the samples are in the -26 .8 to -28.2%o PDB range (Table 1).
Adequate sample weight allowed us to determine the bulk compositions of EOM from 3 of the oil seep samples from GC 185 (88-27-1, -3, and -4). These results can be compared to compositions of 6 crude oil samples from subsurface reservoirs (2000-3000 m) of nearby Jolliet Field (Table 2). There are large differences in C~. composition between crude oil in subsurface reservoirs and in seeps. Ratios of C~5÷ saturated to aromatic hydrocarbons in seep samples (0.37-0.52) are significantly lower than in relatively unaltered crude oil from subsurface reservoirs (0.91-1.54) of Jolliet Field (Table 2). Preferential depletion of C~+ saturated hydrocarbons and preser- vation of Cl~. aromatic hydrocarbons is character- istic of microbially oxidized crude oil (Sassen, 1980; Sassen et al., 1988). Moreover, the C~5+ NSO- compounds and asphaltenes occur in higher percent- ages in cold seep samples than in crude oils (Table 2), which is also consistent with microbial oxidation of crude oil (Sassen, 1980; Sassen et al., 1988). Cl0+ chromatograms of saturated hydrocarbons from cold seep samples and subsurface crude oils show that the n-alkanes and isoprenoid hydrocarbons (including pristane and phytane) originally present in seep oils have been preferentially metabolized by bacteria, leaving a naphthenic residue (Fig. 4).
The c5~3C values of EOM from oil-stained mat material and authigenic carbonate samples from GC 53 and GC 185 (Table 1 : -26 .8 to -28.2%o PDB) are isotopically lighter than the 613C of whole oil samples from Jolliet Field (Table 2: -26 .4 to -26.6%0 PDB). Beggiatoa mats ocur on authigenic carbonate as well as on sediments near vents. SEM shows that shrunken Beggiatoa filaments are associated with solid crude oil coatings on the carbonate rock samples from GC 185 that we extracted and analyzed (Fig. 5). It should not be surprising that extracts of oil-stained sediment and carbonate samples have lighter 6~3C values than Jolliet Field crude oils, because extracts also include some isotopically-lighter bacterial lipids. The 6~3C values of individual frac- tions of oil-stained sediment samples and subsurface crude oils are also shown in Table 2. The 6 ~3C values of saturated and aromatic hydrocarbons are similar• The greatest difference in 6 ~3C values is between the
l J
i 1 "2-
I
.-!
o
-1
81,d "1~
c7
.o
g k
O
8
O
O
o_o
0
2
~.~ ~ 0
E ~
e~
i .
~5 ,4 ._~

Fig. 5. Scanning electron micrograph of authigenic carbonate rock from GC 184. Bacterial filaments occur oll solid microbially oxidized crude oil. The SEM preparation process has shrunk the hollow filaments.
Note scale bar in itm.
85

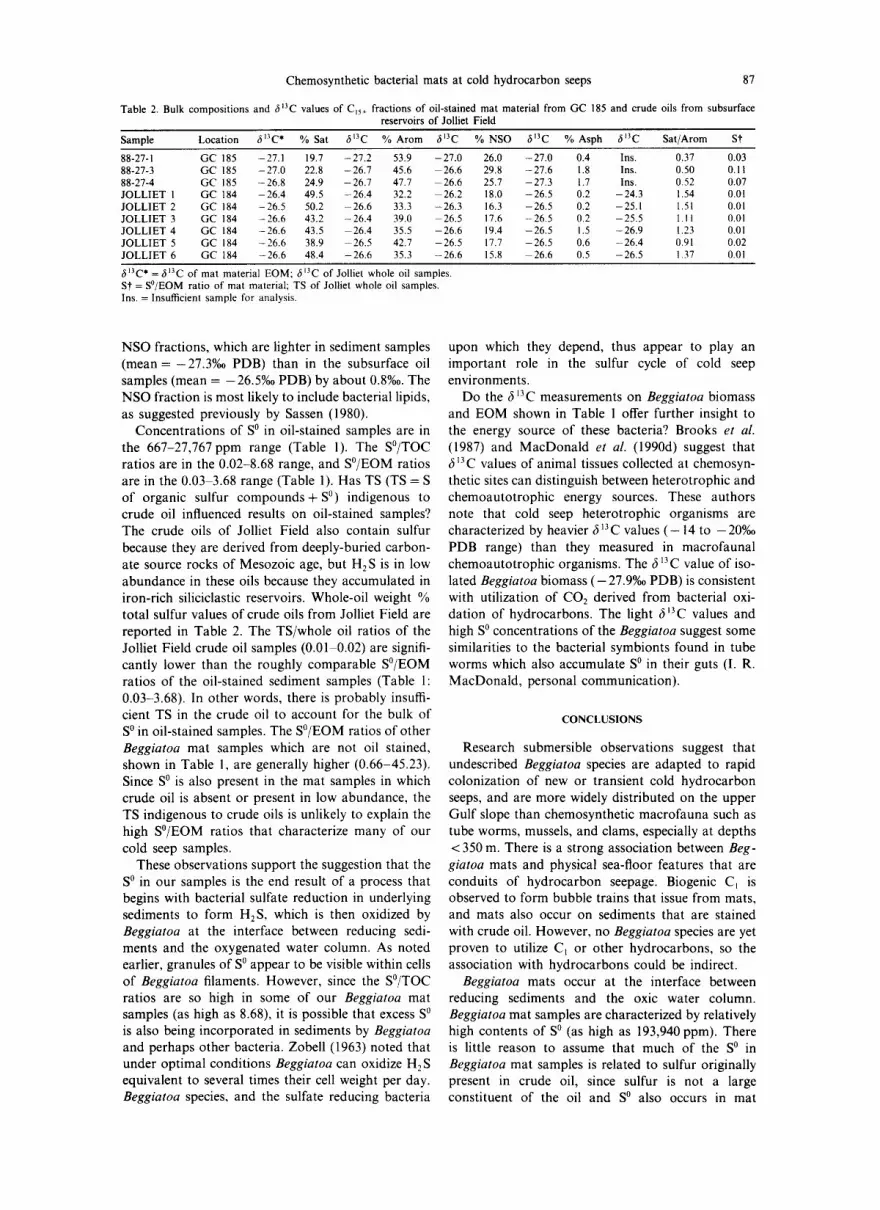
Table 2. Bulk compositions and
Chemosynthe t i c bacter ial mats at cold h y d ro ca rb o n seeps 87
~13C values of Cjs+ fractions of oil-stained mat material from GC 185 and crude oils from subsurface reservoirs of Jolliet Field
Sample Location 613C * % Sat 6t3C % Arom fit3C % NSO fil3C % Asph 6 1 3 C Sat/Arom St
88-27-1 GC 185 -27,1 19.7 -27.2 53.9 -27.0 26.0 -27.0 0.4 Ins. 0.37 0.03 88-27-3 GC 185 -27.0 22,8 -26.7 45.6 26.6 29.8 -27.6 1,8 Ins. 0.50 0.11 88-27-4 GC 185 -26.8 24.9 -26.7 47.7 26.6 25.7 -27.3 1.7 Ins. 0.52 0.07 JOLLIET 1 GC 184 -26.4 49.5 -26.4 32.2 26.2 18 .0 -26,5 0.2 -24.3 1.54 0.01 JOLLIET 2 GC 184 -26.5 50.2 26.6 33.3 -26.3 16.3 -26.5 0.2 -25.1 1.51 0.01 JOLLIET 3 GC 184 -26.6 43.2 -26.4 39.0 26.5 17 .6 --26.5 0.2 -25.5 1.11 0.01 JOLLIET 4 GC 184 -26.6 43.5 26.4 35.5 -26.6 19 .4 -26.5 1.5 -26.9 1.23 0.01 JOLLIET 5 GC 184 -26.6 38.9 26.5 42.7 -26.5 17.7 -26.5 0.6 -26.4 0.91 0.02 JOLLIET 6 GC 184 -26.6 48.4 -26.6 35.3 -26.6 15.8 26.6 0.5 -26.5 1.37 0.01
6~3C * = ,5~3C of mat material EOM; 6 ~3C of Jolliet whole oil samples. St = S°/EOM ratio of mat material; TS of Jolliet whole oil samples. Ins. = Insufficient sample for analysis.
NSO fractions, which are lighter in sediment samples (mean =-27.3%0 PDB) than in the subsurface oil samples (mean = -26.5%0 PDB) by about 0.8%0. The NSO fraction is most likely to include bacterial lipids, as suggested previously by Sassen (1980).
Concentrations of S o in oil-stained samples are in the 667-27,767ppm range (Table 1). The S°/TOC ratios are in the 0.02-8.68 range, and S°/EOM ratios are in the 0.03-3.68 range (Table 1). Has TS (TS = S of organic sulfur compounds+ S o ) indigenous to crude oil influenced results on oil-stained samples? The crude oils of Jolliet Field also contain sulfur because they are derived from deeply-buried carbon- ate source rocks of Mesozoic age, but H 2 S is in low abundance in these oils because they accumulated in iron-rich siliciclastic reservoirs. Whole-oil weight % total sulfur values of crude oils from Jolliet Field are reported in Table 2. The TS/whole oil ratios of the Jolliet Field crude oil samples (0.01 0.02) are signifi- cantly lower than the roughly comparable S°/EOM ratios of the oil-stained sediment samples (Table 1: 0.03-3.68). In other words, there is probably insuffi- cient TS in the crude oil to account for the bulk of S o in oil-stained samples. The S°/EOM ratios of other Beggiatoa mat samples which are not oil stained, shown in Table 1, are generally higher (0.66-45.23). Since S o is also present in the mat samples in which crude oil is absent or present in low abundance, the TS indigenous to crude oils is unlikely to explain the high S°/EOM ratios that characterize many of our cold seep samples.
These observations support the suggestion that the S o in our samples is the end result of a process that begins with bacterial sulfate reduction in underlying sediments to form H2S, which is then oxidized by Beggiatoa at the interface between reducing sedi- ments and the oxygenated water column. As noted earlier, granules of S o appear to be visible within cells of Beggiatoa filaments. However, since the S°/TOC ratios are so high in some of our Beggiatoa mat samples (as high as 8.68), it is possible that excess S o is also being incorporated in sediments by Beggiatoa and perhaps other bacteria. Zobell (1963) noted that under optimal conditions Beggiatoa can oxidize H 2 S equivalent to several times their cell weight per day. Beggiatoa species, and the sulfate reducing bacteria
upon which they depend, thus appear to play an important role in the sulfur cycle of cold seep environments.
Do the ~13C measurements on Beggiatoa biomass and EOM shown in Table 1 offer further insight to the energy source of these bacteria? Brooks et al. (1987) and MacDonald et al. (1990d) suggest that 6 I3C values of animal tissues collected at chemosyn- thetic sites can distinguish between heterotrophic and chemoautotrophic energy sources. These authors note that cold seep heterotrophic organisms are characterized by heavier ~i 13 C values ( - 14 to -20%o PDB range) than they measured in macrofaunal chemoautotrophic organisms. The 6 ~3C value of iso- lated Beggiatoa biomass ( - 27.9%0 PDB) is consistent with utilization of CO z derived from bacterial oxi- dation of hydrocarbons. The light 6~3C values and high S o concentrations of the Beggiatoa suggest some similarities to the bacterial symbionts found in tube worms which also accumulate S o in their guts (I. R. MacDonald, personal communication).
CONCLUSIONS
Research submersible observations suggest that undescribed Beggiatoa species are adapted to rapid colonization of new or transient cold hydrocarbon seeps, and are more widely distributed on the upper Gulf slope than chemosynthetic macrofauna such as tube worms, mussels, and clams, especially at depths < 350 m. There is a strong association between Beg- giatoa mats and physical sea-floor features that are conduits of hydrocarbon seepage. Biogenic C~ is observed to form bubble trains that issue from mats, and mats also occur on sediments that are stained with crude oil. However, no Beggiatoa species are yet proven to utilize C~ or other hydrocarbons, so the association with hydrocarbons could be indirect.
Beggiatoa mats occur at the interface between reducing sediments and the oxic water column. Beggiatoa mat samples are characterized by relatively high contents of S o (as high as 193,940 ppm). There is little reason to assume that much of the S o in Beggiatoa mat samples is related to sulfur originally present in crude oil, since sulfur is not a large constituent of the oil and S o also occurs in mat

88 ROGER SASSEN et al.
BEGGIATOA
OXIDATION
BACTERIAL SULFATE
REDUCTION ox,o_
I CARBON FIXATION
02 BACTERIAL [ \ * . . . . . . HYDROCARBON [ CaCO,, l DEPLETION OXIDATION
Fig. 6. Summary diagram illustrating the proposed geochemical role of Beggiatoa in cold hydrocarbon seeps.
material in which crude oil is absent or in low abundance. On this basis, the origin of S o is best explained geochemically by in situ oxidation of H2 S by Beggiatoa.
Beggiatoa species are known to utilize CO2. The light 613C value of Beggiatoa biomass (-27.9%o PDB) is consistent with chemoautotrophy. There is evidence that CO 2 is being formed presently at mat localities by microbial oxidation of hydrocarbons. The authigenic carbonate rock common at mat localities is characterized by light 613C values that reflect a slow origin from precipitation of CO2 derived by bacterial oxidation of seeping hydrocarbons. Based on the geochemistry of their environment, it is probable that the Beggiatoa species at cold hydro- carbon seeps utilize CO2 produced by hydrocarbon oxidizing bacteria.
This work emphasizes the complex relationships among bacteria at cold hydrocarbon seeps, much of which is not yet well understood. It appears that the Beggiatoa species are dependent on the activities of various bacteria that oxidize hydro- carbons, and which reduce sulfate when 02 is de- pleted in cold hydrocarbon seeps (Fig. 6). Our geochemical data are consistent with the hypothesis that they oxidize H2S to form S O and sulfate, and utilize CO2 as a carbon source. Beggiatoa thus play a significant role in the sulfur cycle at cold hydrocarbon seeps, but it should be noted that the carbon cycle in cold hydrocarbon seeps is per- turbed by net accumulation of carbonate minerals (Fig. 6). Beggiatoa mats appear to retard loss of hydrocarbons to the water column by physically retaining fluids in sediments. This function could serve to enhance production of the H2S and CO 2 needed by Beggiatoa, and also to indirectly enhance
the formation of authigenic carbonate minerals in sediments.
Acknowledgements--Analytical work was supported by the Basin Research Institute and The Center for Energy Studies, Louisiana State University, and research submersible time was provided through grants from the NOAA National Undersea Research Center, University of North Carolina at Wilmington. Discussions with Thomas Naurauter (ARCO), Douglas Cook (Conoco), and I. R. MacDonald (Texas A&M University) are greatly appreciated.
REFERENCES
Aharon P., Graber E. R. and Roberts H. H. (1991) Detec- tion of hydrocarbon venting on the Gulf sea floor from determination of dissolved inorganic carbon and 613C of the water column overlying seeps. Trans. Gulf Coast Assoc. Geol. Socs. 41, 24 .
Anderson R. K., Scalan R. S., Parker P. L. and Behrens E. W. (1983) Seep oil and gas in Gulf of Mexico slope sediment. Science 222, 619-621.
Behrens E. W. (1988) Geology of a continental slope oil seep, northern Gulf of Mexico. AAPG Bull. 72, 105-114.
Brooks J. M., Kennicutt II, M. C., Fay R. R., McDonald T. J. and Sassen R. 0984) Thermogenic gas hydrates in the Gulf of Mexico. Science 225, 409-411.
Brooks J. M., Cox B. H., Bryant W. R., Kennicutt II, M. C, Mann R. G. and McDonald T. J. (1986) Association of gas hydrates and oil seepage in the Gulf of Mexico. In Advances in Organic Geochemistry 1985 (Edited by Leythaeuser D. and Rullkotter J.). Org. Geochem. 10, 221-234.
Brooks J. M., Kennicutt II, M. C., Fisher C. R., Macko S. A., Cole K., Childress J. J., Bidigare R. R. and Vetter R. D. (1987) Deep-sea hydrocarbon communities: evi- dence for energy and nutritional sources. Science 238, 1138-1142.
Cook D. and D'Onfro P. (1991) Jolliet Field thrust fault structure and stratigraphy, Green Canyon Block 184, Offshore Louisiana. Trans Gulf Coast Assoc'. Geol. Socs. 41, 100-101.

Chemosynthetic bacterial mats at cold hydrocarbon seeps 89
Corliss J. B., Dymond J., Gordon L. I., Edmond J. M., von Herzen R. P., Banard R. D., Green K., Williams D., Bainbridge A., Crane K. and van Andel T. H. (1979) Submarine thermal springs on the Galopagos Rift. Science 203, 1073 1083.
Davidson D. W., Gard S. K., Gough S. R., Handa Y. P., Ratcliffe C., Ripmeester J. A., Tse J. S. and Lawson W. R. (1986) Laboratory analysis of a naturally occurring gas hydrate from sediment of the Gulf of Mexico. Geochim. Cosmochim. Acta 50, 619~23.
Davis P. H. and Spies R. B. (1980) Infaunal benthos of a natural petroleum seep. Study of community structure. Mar. Biol. 59, 31-41.
Hovland M. and Thomsen E. (1989) Hydrocarbon-based communities in the North Sea? Sarsia 74, 29-42.
Kennicutt II, M. C. and Brooks J. M. (1990) Recog- nition of areas effected by petroleum seepage: Northern Gulf of Mexico continental slope. Geo-Mar. Lett. 10, 221-224.
Kennicutt II, M. C., Brooks J. M. and Denoux G. J. (1988) Leakage ofdeep, reservoired petroleum to the near surface of the Gulf of Mexico continental slope. Mar. Chem. 24, 39 59.
Kennicutt 1I, M. C., Brooks J. M., Bidigare R. R., Wade T. L. and McDonald T. J. (1985) Vent-type taxa in a hydrocarbon seep region on the Louisiana slope. Nature (Lond.) 317, 351-353.
Larkin J. M. and Polman J. K. (1988) Beggiatoa in Louisiana rice fields. In Current Perspecth~e in Environmental Geo- chemistry (Edited by Giovanni-Sermanni G. and Nannip- ieri P.). C.N.R.-I.P.R.A., Rome, 329 339.
Larkin J. M. and Henk M. C. (1989) Is 'hollowness' an adaptation of large prokaryotes to their largeness? Micro- bios Left. 42, 69 72.
MacDonald 1. R., Boland G. S., Baker J. S., Brooks J. M., Kennicutt II, M. C. and Bidigare R. R. (1989) Gulf of Mexico hydrocarbon seep communities. Mar. Biol. 101, 235-247.
MacDonald I. R., Reilly II, J. F., Guinasso Jr N. L., Brooks J. M., Carney R. S., Bryant W. A. and Bright T. J. (1990a) Chemosynthetic mussels at a brine-filled pockmark in the northern Gulf of Mexico. Science 248, 1096-1099.
MacDonald I. R., Callender W. R., Burke Jr R. A., McDonald S. J. and Carney R. S. (1990b) Fine-scale distribution of methanotropic mussels at a Louisiana cold seep. Prog. Oceanogr. 24, 15 24.
MacDonald I. R., Reilly II, J. F., Guinasso Jr N. L., Brooks J. M., Carney R. S., Bryant W. A. and Bright T. J. (1990c) Chemosynthetic mussels at a brine-filled pockmark in the northern Gulf of Mexico. Science 248, 1096-1099.
MacDonald I. R., Guinasso Jr N. L., Reilly J. F., Brooks J. M., Callender W. R. and GabrieUe S. G. (1990d) Gulf of Mexico hydrocarbon seep communities: VI. Patterns of community structure and habitat. Geo-Mar. Lett 10, 244- 252.
Nelson D. C. and Jannasch H. W. (1983) Chemoautotrophic growth of a marine Beggiatoa in sulfide-gradient cultures. Arch. Microbiol. 136, 262-269.
Neurauter T. W. and Bryant W. R. (1990) Seismic expression of sedimentary volcanism on the continental slope, north- ern Gulf of Mexico. Geo-Mar. Lett. 10, 225-231.
Prior D. B., Doyle E. H. and Kaluza M. J. (1989) Evidence of sediment eruption on deep sea floor, Gulf of Mexico. Science 243, 517-519.
Roberts H. H. and Neurauter T. W. (1990) Direct obser- vations of a large active mud vent on the Louisiana continental slope. AAPG Bull. 74, 1508.
Roberts H. H., Sassen R., Carney R. and Aharon P. (1989) L3-C depleted authigenic carbonate buildups from hydro- carbon seeps, Louisiana continental slope. Trans. Gulf Coast Assoc. Geol. Soes. 39, 523-530.
Roberts H. H., Sassen, R., Carney R. and Aharon P. (1990a) The role of hydrocarbons in creating sediment and small-scale topography of the Louisiana continental slope. In Gulf Coast Oils and Gases, Proc. Ninth Ann. Res. Conf. Gulf Coast See. Soc. Econ. Paleont. Mon. Foun- dation (Edited by Schumacher D. and Perkins B. F.), 311-324.
Roberts H. H., Aharon P., Carney R., Larkin J. and Sassen R. (1990b) Seafloor responses to hydrocarbon seeps, Louisiana continental slope. Geo-Mar. Lett. 10, 232 243.
Rosman I., Boland G. S. and Baker J. S. (1987) Epifaunal aggregations of Vesicomyidae on the continental slope off Louisiana. Deep-Sea Res. 34, 1811-1820.
Sassen R. (1987) Organic geochemistry of salt dome cap rocks, Gulf Coast salt basin. In Dynamical Geology of Salt and Related Structures (Edited by Lerche I. and O'Brien J. J.), pp. 631~49. Academic Press, London.
Sassen R. (1980) Biodegradation of crude oil and mineral deposition in a shallow Gulf Coast salt dome. Org. Geochem. 2, 153 166.
Sassen R. and Chinn E. W. (1989) Effects of elemental sulfur during programmed pyrolysis. Org. Geochem. 14, 475-477.
Sassen R., Chinn E. W. and McCabe C. (1988) Recent hydrocarbon alteration, sulfate reduction and formation of elemental sulfur and metal sulfides in salt dome cap rock. Chem. Geol. 74, 57~6.
Sassen R., Grayson P., Cole G., Roberts H. H. and Aharon P. (1991) Hydrocarbon seepage and salt-dome related carbonate reservoir rocks of the Gulf Coast. Trans. Gulf Coast Assoc. Geol. Socs. 41, 570-578.
Strohl W. R., Cannon G. C., Shively J. M., Gude H., Hook L. A., Lane C. M. and Larkin J. M. (1981) Heterotrophic carbon metabolism by Beggiatoa alba. J. Bacteriol. 148, 572-583.
Zobell C. E. (1963) Organic Geochemistry of sulfur. In Organic Geochemistry (Edited by I. A. Breger), Mono- graph 16, Earth Science Series, 543-578. Macmillan, New York.