changes in extracellular potassium during the spontaneous activity of medullary respiratory neurones
TRANSCRIPT

Pflfigers Arch. 376, 139-149 (1978) Pflfigers Archiv European Journal of Phys~ogy
�9 by Springer-Verlag 1978
Changes in Extracellular Potassium During the Spontaneous Activity of Medullary Respiratory Neurones
D. W. Richter, H. Camerer, and U. Sonnhof with technical assistance of Mrs. A. Bischoff
I. Physiologisches Institut, Universit/it Heidelberg, Im Neuenheimer Feld 326, D-6900 Heidelberg, Federal Republic of Germany
Abstract. In 34 cats, the changes in extracellular potassium ion activity (aK) and extracellular spike activity within the pool of respiratory neurones in the dorsormedial and ventrolateral medulla were recorded using microelectrodes filled with a liquid potassium ion exchange resin. Cyclic changes in a~ which parallel central respiratory activity were restricted to those regions where respiratory neurones are known to be localized. The largest changes in a K (0.1-0.3mmol - 1-1) were found within the ventral pool of inspiratory neurones. The aK increased during inspiration in paral- lel with the pattern of phrenic nerve activity. The smallest changes in aK (0.02-0.06 mmol �9 1-1) were observed within the ventral pool of expiratory neurones. Here, aK showed a transient increase during both inspiration and expiration. Within the dorsal pool of inspiratory neu- rones, small fluctuations of aK were observed parallel- ing phrenic nerve activity and the afferent discharge of the intact vagal nerves. After the vagal nerves were cut, the changes in aK then paralleled phrenic nerve activity. The variations in aK within the ventral pool of re- spiratory neurones did not change after bilateral sec- tion of vagal nerves.
Repetitive stimulation of the wgal nerves (0.1- 0.5V, 0.05 ms) produced an increase in aK only within the dorsal pool of inspiratory neurones, whereas re- petitive spinal cord stimulation (5-10V, 0.05 ms) resulted in an increase of aK within the ventral pool of respiratory neurones.
The amplitude of the cyclic changes in aK increased significantly whenever the electrode approached in- dividual respiratory neurones as verified by the ampli- tude and shape of the spikes recorded by the reference barrel. The maximal changes in aK then reached a peak amplitude of 1.3-1.5 mmol �9 1-1, the pattern of aK changes resembling that measured within the pools of neurones.
The a K started to rise prior to the discharge of action potentials, indicating that the efflux of K+-ions was
produced as a consequence of synaptic transmission. The functional importance of these changes in extracel- lular potassium is discussed.
Key words: Extracellular potassium activity - Medullary respiratory neurones - Spontaneous ac- tivity - Vagal nerve stimulation - Synchronization - Synaptic transmission.
Introduction
Medullary respiratory neurones periodically discharge a long-lasting burst of action potentials which appears to be synaptieally induced (Salmoiraghi and v. Baumgarten, 1961; Hildebrandt, 1974; Mitchell and Herbert, 1974; Richter and Heyde, 1975; Kreuter et al., 1977). This discharge has been assumed to be generated by a neuronal network which is capable of producing a rhythmic output when activated tonically (Salmoiraghi and Burns, 1960; Hugelin and Cohen, 1963; Cohen, 1970). This assumption does not, however, explain the mechanism of timing respiratory activity. Normally, synaptic interaction between neurones acts within a few milliseconds, whereas the timing cues for respiratory activity range in the order of seconds. There are, however, some bulbar interneurones which receive a weak respiratory-related synaptic input and their membrane potentials reach firing threshold only after a considerable delay from the onset of the respiratory phase (Richter et al., 1975). A similar inhibitory interneuronal activity possibly generates the 'off- switching' of inspiratory activity (v. Euler et al., 1973; Cohen, 1974, 1977; Richter et al., 1975). However, microstimulation techniques have failed to reveal the existence of axon collaterals of expiratory neurones (ENs) (Merrill, 1974). If the interpretation of these negative results is that ENs do not possess axon
0031-6768/78/0376/0139/$ 02.20

140 Pflfigers Arch. 376 (1978)
co l la te ra l s a t the b r a i n s t e m level, then the a u g m e n t i n g
p a t t e r n o f e x p i r a t o r y ac t iv i ty a n d the se l f - l imi t ing
d i scharge o f E N s c a n n o t be exp la ined by chemica l
a x o s o m a t i c o r a x o d e n d r i t i c synap t i c in t e rac t ion . In
c o n s e q u e n c e , d e n d r o d e n d r i t i c chemica l synap t i c in-
t e rac t ions a n d n o n - s y n a p t i c m e c h a n i s m s h a v e to be
i n v o k e d (v. B a u m g a r t e n , 1956; K r e u t e r et al., 1977).
' F a t i g u e ' has been c l a imed to p lay a ro le by
S a l m o i r a g h i a n d v. B a u m g a r t e n (1961), bu t ne i the r
a c c o m m o d a t i o n n o r a d a p t a t i o n o f b u l b o s p i n a l (BS)
r e sp i r a to ry n e u r o n e s seem to be s t rong e n o u g h to p l ay a
f unc t i ona l ro le ( R i c h t e r and Heyde , 1975). A di rec t
m o d i f i c a t i o n o f the exc i tab i l i ty o f a d j a c e n t n e u r o n a l
s t ruc tu res m a y exist , s ince p r e v i o u s i nves t iga t ions
( K r e u t e r et al., 1977) h a v e s h o w n tha t m a n y o f the
m e d u l l a r y r e s p i r a t o r y n e u r o n e s lie in c lose p r o x i m i t y
and h a v e r ichly o v e r l a p p i n g dendr i t i c trees. In such
a cond i t i on , bes ides d e n d r o d e n d r i t i c synap t i c in ter -
ac t ion , ex t r ace l l u l a r f ield po t en t i a l s (v. B a u m -
gar ten , 1956) a n d changes in t he ion ic e n v i r o n -
m e n t i.e. especia l ly changes in ex t race l lu la r concen -
t r a t i on o f p o t a s s i u m ions (K § m a y d i rec t ly effect
changes in the n e u r o n e s ' exc i tab i l i ty ( R i c h t e r et al.,
1977b). In o r d e r to s tudy the l a t t e r poss ibi l i ty , we h a v e
m e a s u r e d the changes in p o t a s s i u m ac t iv i ty (aK) wi th in
the ex t r ace l lu l a r space ( E C S ) o f r e s p i r a t o r y n e u r o n e s
us ing ion-sens i t ive mic roe l ec t rodes . In a f o l l o w i n g
p a p e r we will t ry to c lar i fy the f u n c t i o n a l s igni f icance o f
the changes in ex t race l lu la r a K, we obse rved .
A p r e l i m i n a r y r e p o r t has a l r eady been pub l i shed
e l sewhere ( R i c h t e r et al., 1977a).
Methods
The experiments were performed on 34 adult cats under chloralose anaesthesia (50rag. kg-1 i.v.). Artificial positive pressure ventilation was given and paralysis was induced by i.v. injection of Pancuronium bromide (0.5 mg. kg- 1 intial dose, and thereafter maintained with 0.2rag. kg 1 �9 h- 1). Respiratory movements were abolished bY bilat- eral pneumothorax, and complete collapse of the lungs was prevent- ed by an expiratory resistance of 0.5 - 1 cm H20. End-tidal CO2 was adjusted to 3 - 5 volume % by altering the rate of artificial venti- lation. The basic experimental preparation and the intracellular recording techniques have been fully described in a preceding paper (Richter et al., 1975). As a modification of the methods de- scribed there, the lower brain stem was superfused with an artificial cerebrospinal fluid (2.7mmol �9 1 1 KCI, 2.5mmol �9 1-1 CaCI2, 16mmol �9 1-1 NaHCOa ' 140mmol �9 1-~ NaC1, and 2.0mmol - 1-1 MgC12, pH 7.3).
In some experiments the vagal nerves were cut and their central ends prepared for electrical stimulation via bipolar platinum elec- trodes, Alternatively, bipolar stimulating and recording (Pt) elec- trodes were placed onto the intact vagal nerves. In these preparations it was possible to investigate the reflex effect of lung inflation and to record the afferent discharge of lung stretch receptors or to simulate lung inflation by electrical stimulation of the vagal nerves (rec- tangular pulses of 0.05 ms duration at 100-500 mV) and to record
the compound potential of the fibres excited. In addition to the original recording, phrenic nerve activity and the afferent discharge of the intact vagaI nerves were integrated by a 'leaky integrator' system.
The K+-ion-sensitive microelectrodes (ISMs) were positioned stereotactically in the brainstem from a dorsal approach either to a region of the ventrolateral nucleus of the solitary tract (dorsal population of respiratory neurones, 0.5 - 1.5 mm rostral to the Obex and 1.5 -2 .5 mm lateral to the midline) or to a region dorsomedial to the nucleus ambiguus or the lateral reticular nucleus (ventral pool of respiratory neurones, 1 mm caudal to 2 mm rostral to the Obex and 2.5-3.5 mm lateral to the midline). Bulbospinal respiratory neu- rones within the ventral and dorsal populations of respiratory neurones (Bianchi, 1971) were identified by antidromic excitation after stimulating the spinal cord at the C 2 - C3 level with two bipolar concentric stainless steel electrodes (outer diameter 200 pm). Laryngeal neurones were identified by vagal nerve stimulation and a high velocity of antidromic spike conduction. Neurones, which could not be antidromically activated either by bilateral spinal cord or vagal nerve stimulation (NAA-neurons, Richter et al., 1975; Kreuter et al., 1977) were sometimes recorded from within both the dorsal and ventral population of respiratory neurones. Some of the NAA- neurones within the dorsal population revealed a monosynaptic excitation following electrical excitation of afferent fibres within the vagal nerves conducting with 17- 57m. s -1 (Richter et al., 1978) as assumed by v. Euler et al. (1973).
K+-ISMs were manufactured from double barralled glass pi- pettes. Their tips were broken to an outer diameter of 2 - 3 pm. One barrel was filled with 150 mmol �9 1-1 NaC1 and was used both as a recording electrode and a reference electrode for the ion-sensitive barrel. The second barrel, after siliconization of the inner wall of the tip for a length of about 0.5mm, was filled from behind with 100 mmol. 1-1KCI solution. The tip of this barrel was then filled with Coming potassium exchange resin No.477317 for a length of 0.2-0.4ram by suction. Before and after measurements within neural tissue, the electrodes were calibrated in standard solutions containing 150 mmol-1-1 sodium chloride and known concen- trations of potassium chloride [KCL] varying between 0.5 and 80 mtnol - 1 1 The calibration curves were similar for all electrodes used and showed a sensitivity of 45 - 55 mV per a decade change in [KCL] measured at a temperature of 20~ At a [KCL] level below 5 - 10 mmol �9 1 - 1, the sensitivity of the ISMs continuously decreased due to Na + interference (Lux and Neher, 1973). At a [KCL] of 3 mmol �9 1 - 1, the slope of the calibration curve ranged between 30 - 40 mV per decade change in [KCL]. The K + sensitive barrel was connected to a DC preamplifier with an input resistance of 1013f2 and with capacitance neutralization. The signal, recorded by the reference barrel, was amplified by the same type of preamplifier and was differentially recorded against that of the sensitive barrel. This type of measuring potassium activity (aK) has a very short response time and is able to reproduce variations in aK with time courses longer than 20 ms (Neher and Lux, 1973). In addition to this 'K+-signal ', the potential of the reference barrel was taken to record focal potentials and neuronal spike activity at the same location where the extracel- lular a K was measured. The extracellular spike potentials were visualized on an oscilloscope and, after window-discrimination, written as normalized pulses on a direct writing system. The frequency of spontaneous spike discharge was integrated with a 'leaky integrator' system.
Results
Changes in Membrane Potential
M e d u l l a r y r e sp i r a to ry n e u r o n e s rece ive i n h i b i t o r y a n d
exc i t a t o ry synap t i c inpu t s (R ich t e r et al., 1975; K r e u t e r
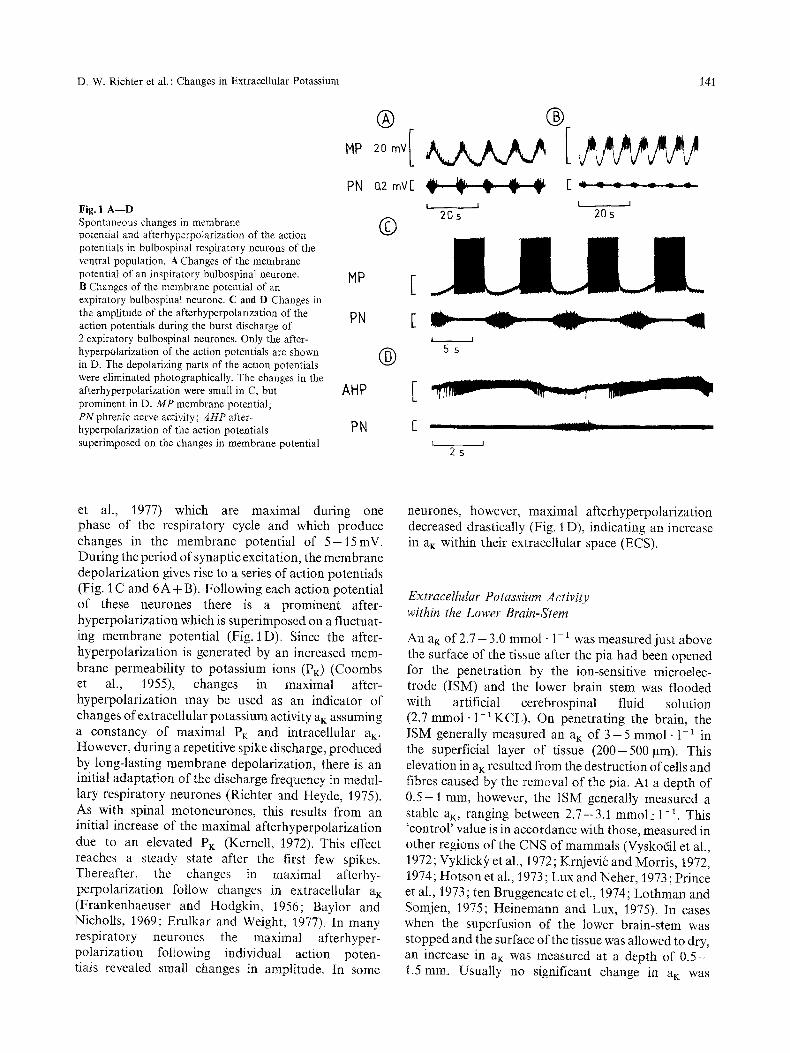
D. W. Richter et al. : Changes in Extracellular Potassium 141
HP
PN
| 20 rnV[ 0,2 mVE
Fig. 1 A - - D Spontaneous changes in membrane ( ~ potential and afterhyperpolarization of the action potentials in bulbospinal respiratory neurons of the ventral population. A Changes of the membrane potential of an inspiratory bulbospinal neurone. MP [- B Changes of the membrane potential of an k expiratory bulbospinal neurone. C and D Changes in the amplitude of the afterhyperpolarization of the PN [" action potentials during the burst discharge of 2 expiratory bulbospinal neurones. Only the after- hyperpolarization of the action potentials are shown ( ' ~ in D. The depolarizing parts of the action potentials were eliminated photographically. The changes in the afterhyperpolarization were small in C, but AN P r prominent in D. M P membrane potential;
[_
P N phrenic nerve activity; A H P after- hyperpolarization of the action potentials P N I- superimposed on the changes in membrane potential
|
y wlr " ~lr " ~ IV
I J t l
20s 20s
I ' q P " ~ , , , 1 ~ e r , - , ~ m w
I I
5s
L _ _ I
2 s
l |
et al., 1977) which are maximal during one phase of the respiratory cycle and which produce changes in the membrane potential of 5-15mV. During the period of synaptic excitation, the membrane depolarization gives rise to a series of action potentials (Fig. 1 C and 6 A + B). Following each action potential of these neurones there is a prominent after- hyperpolarization which is superimposed on a fluctuat- ing membrane potential (Fig. 1 D). Since the after- hyperpolarization is generated by an increased mem- brane permeability to potassium ions (PK) (Coombs et al., 1955), changes in maximal after- hyperpolarization may be used as an indicator of changes of extracellular potassium activity aK assuming a constancy of maximal PK and intracellular aK. However, during a repetitive spike discharge, produced by long-lasting membrane depolarization, there is an initial adaptation of the discharge frequency in medul- lary respiratory neurones (Richter and Heyde, 1975). As with spinal motoneurones, this results from an initial increase of the maximal afterhyperpolarization due to an elevated PK (Kernell, 1972). This effect reaches a steady state after the first few spikes. Thereafter, the changes in maximal afterhy- perpolarization follow changes in extracellular aK (Frankenhaeuser and Hodgkin, 1956; Baylor and Nicholls, 1969; Erulkar and Weigl~.t, 1977). In many respiratory neurones the maximal afterhyper- polarization following individual action poten- tials revealed small changes in amplitude. In some
neurones, however, maximal afterhyperpolarization decreased drastically (Fig. 1 D), indicating an increase in aK within their extracellular space (ECS).
Extracetlutar Potassium Activity within the Lower Brain-Stem
An aK of 2.7-3.0 mmol - 1-t was measured just above the surface of the tissue after the pia had been opened for the penetration by the ion-sensitive microelec- trode (ISM) and the lower brain stem was flooded with artificial cerebrospinal fluid solution (2.7 mmol. 1-1KCL). On penetrating the brain, the ISM generally measured an a K of 3 - 5 mmol �9 1-1 in the superficial layer of tissue (200-500 gin). This elevation in a~ resulted from the destruction of cells and fibres caused by the removal of the pia. At a depth of 0.5-1 ram, however, the ISM generally measured a stable aK, ranging between 2.7-3.1 mmol: 1-1. This 'control' value is in accordance with those, measured in other regions of the CNS of mammals (Vysko6il et al., 1972; Vyklick~ et al., 1972; Krnjevi6 and Morris, 1972, 1974; Hotson et al., 1973; Lux and Neher, 1973; Prince et al., 1973 ; ten Bruggencate et el., 1974; Lothman and Somjen, 1975; Heinemann and Lux, 1975). In cases when the superfusion of the lower brain-stein was stopped and the surface of the tissue was allowed to dry, an increase in a K was measured at a depth of 0 .5- 1.5 mm. Usually no significant change in a K was
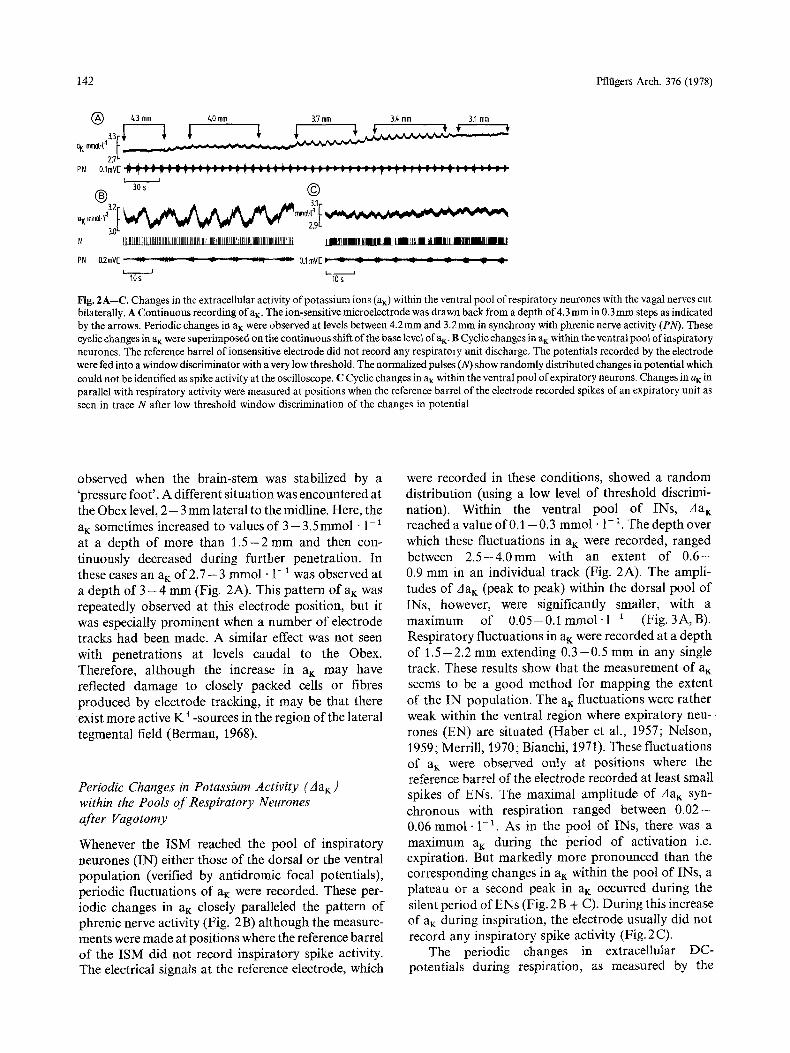
142 Pfliigers Arch. 376 (1978)
4.3 mm t,.O mm 3.7 mm 3.4 mm 3.1 mm
2] PN 0.1mVC ~ , , , , , ~ r . . ~ . . , ~ v , , ~ , , r ; : , ~ , . - , , . . , 1 ~ , r r r r . . - . . . , r , . . . . . . . . . . . . . . . . . . . . . . ~ ; ~
i i 30 s 0 @
3.0 L - " ~ ~ I t " ~ ~ " ' 2.9 c
~t IIHIIII[IIII[IIIII,IIIIIlilIIJ[ IIIIIII HIIIUh~llllllll~ltlllllllJFU~ltlllllll~lillllllll~l~]lil~tJ[BI I mlll[InlH i l l i l l l l n l i l i l l l l l �9 ill inl l l l mllllml[llmmmlll
PN 02mVE . . . . . . . . . . . . . . . . 0.1mV[; = ~- C- ~- :-7., -7. C- - .~
r l o s ~ ~ l o s '
Fig. 2A--C. Changes in the extracellular activity of potassium ions (aK) within the ventral pool of respiratory neurones with the vagal nerves cut bilaterally. A Continuous recording ofaK. The ion-sensitive microelectrode was drawn back from a depth of 4.3 mm in 0.3mm steps as indicated by the arrows. Periodic changes in aK were observed at levels between 4.2ram and 3.2mm in synchrony with phrenic nerve activity (PN). These cyclic changes in aK were superimposed on the continuous shift of the base level of aK- B Cyclic changes in aK within the ventral pool of inspiratory neurones. The reference barrel of ionsensitive electrode did not record any respiratory unit discharge. The potentials recorded by the electrode were fed into a window discriminator with a very low threshold. The normalized pulses (N) show randomly distributed changes in potential which could not be identified as spike activity at the oscilloscope. C Cyclic changes in aK within the ventral pool of expiratory neurons. Changes in aK in parallel with respiratory activity were measured at positions when the reference barrel of the electrode recorded spikes of an expiratory unit as seen in trace N after low threshold window discrimination of the changes in potential
observed when the brain-stem was stabilized by a 'pressure foot ' . A different situation was encountered at the Obex level, 2 - 3 mm lateral to the midline. Here, the aK sometimes increased to values of 3 - 3.5mmol �9 1- ~ at a depth of more than 1 . 5 - 2 mm and then con- tinuously decreased during further penetration. In these cases an aK o f 2 . 7 - 3 mmol �9 1-1 was observed at a depth of 3 - 4 mm (Fig. 2A). This pattern of a~ was repeatedly observed at this electrode position, but it was especially prominent when a number of electrode tracks had been made. A similar effect was not seen with penetrations at levels caudal to the Obex. Therefore, although the increase in aK may have reflected damage to closely packed cells or fibres produced by electrode tracking, it may be that there exist more active K +-sources in the region of the lateral tegmental field (Berman, 1968).
Periodic Changes in Potassium Activity (AaK ) within the Pools of Respiratory Neurones after Vagotomy
Whenever the ISM reached the pool of inspiratory neurones (IN) either those of the dorsal or the ventral population (verified by antidromic focal potentials), periodic fluctuations of aK were recorded. These per- iodic changes in ai~ closely paralleled the pattern of phrenic nerve activity (Fig. 2 B) although the measure- ments were made at positions where the reference barrel of the ISM did not record inspiratory spike activity. The electrical signals at the reference electrode, which
were recorded in these conditions, showed a random distribution (using a low level o f threshold discrimi- nation). Within the ventral pool of INs, Aa~ reached a value of0A - 0.3 mmol �9 1-1. The depth over which these fluctuations in aK were recorded, ranged between 2 . 5 - 4 . 0 m m with an extent of 0 . 6 - 0.9 m m in an individual track (Fig. 2A). The ampli- tudes of Aa K (peak to peak) within the dorsal pool of INs, however, were significantly smaller, with a maximum of 0 .05-0 .1 mmol- I - 1 (Fig. 3A, B). Respiratory fluctuations in aK were recorded at a depth of 1 .5 -2 .2 m m extending 0 . 3 - 0 . 5 m m in any single track. These results show that the measurement of a K seems to be a good method for mapping the extent of the IN population. The a K fluctuations were rather weak within the ventral region where expiratory neu-- tones (EN) are situated (Haber et al., 1957; Nelson, 1959; Merrill, 1970; Bianchi, 1971). These fluctuations of aK were observed only at positions where the reference barrel of the electrode recorded at least small spikes of ENs. The maximal amplitude of Aa K syn- chronous with respiration ranged between 0 . 0 2 - 0.06 mmol . 1-1. As in the pool of INs, there was a maximum a~: during the period of activation i.e. expiration. But markedly more pronounced than the corresponding changes in aK within the pool of INs, a plateau or a second peak in aK occurred during the silent period of ENs (Fig. 2 B + C). During this increase of aK during inspiration, the electrode usually did not record any inspiratory spike activity (Fig. 2 C).
The periodic changes in extracellular DC- potentials during respiration, as measured by the
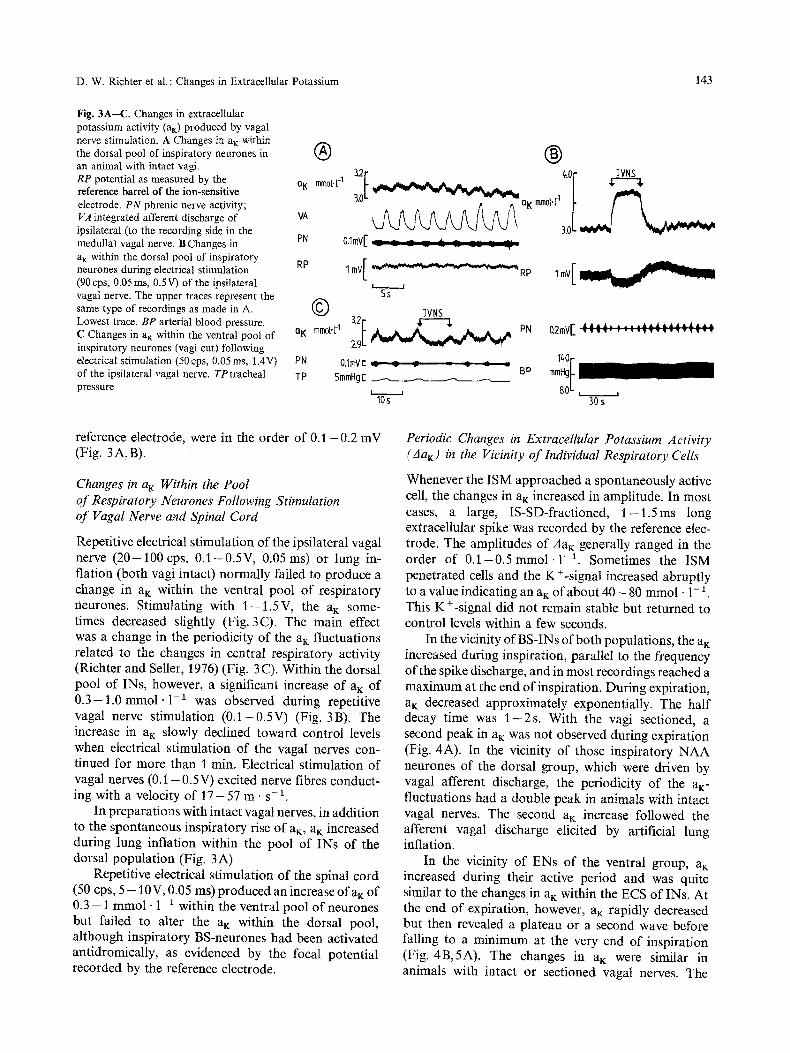
D. W. Richter et al. : Changes in Extracellular Potassium 143
Fig. 3A--C. Changes in extracellular potassium activity (a~) produced by vagal nerve stimulation. A Changes in aK within the dorsal pool of inspiratory neurones in an animal with intact vagi. RP potential as measured by the reference barrel of the ion-sensitive electrode. PN phrenic nerve activity; VA integrated afferent discharge of ipsilateral (to the recording side in the medulla) vagal nerve. B Changes in PN a K within the dorsal pool of inspiratory RP neurones during electrical stimulation (90 cps, 0.05 ms, 0.5V) of the ipsilateral vagal nerve. The upper traces represent the same type of recordings as made in A. Lowest trace. BP arterial blood pressure. C Changes in a K within the ventral pool of nK inspiratory neurones (vagi cut) following electrical stimulation (50cps, 0.05ms, 1.4V) PN of the ipsilateral vagal nerve. TP tracheal TP pressure
| | ,or
o K mm0t.t -1 3 . 0 L ~ a K m m 0 l . [ _ 1 [ ~
0.1mV[
�9 mmol.[ A
I mV[ ~ R P
Ss 3VNS 3.2[ ,F'-'---'$
29 ~ PN
0.1mVc ~ _ ~ = -z : BP
5mmHgE . . . .
10s J
1ovE..,, VE A A ~ ' ~ " " ~ J * A A A ' ~ A ~ A A A ~ A
30s
reference electrode, were in the order of 0 . 1 - 0 . 2 mV (Fig. 3 A, B).
Changes in a K Within the Pool of Respiratory Neurones Following Stimulation of Vagal Nerve and Spinal Cord
Repetitive electrical stimulation of the ipsilateral vagal nerve ( 2 0 - 1 0 0 cps, 0 .1 -0 .5V, 0.05 ms) or lung in- flation (both vagi intact) normally failed to produce a change in a~ within the ventral pool of respiratory neurones. Stimulating with 1 - 1 . 5 V , the aK some- times decreased slightly (Fig. 3C). The main effect was a change in the periodicity of the aK fluctuations related to the changes in central respiratory activity (Richter and Seller, 1976) (Fig. 3 C). Within the dorsal pool of INs, however, a significant increase of a K of 0 . 3 - 1 . 0 mmol -1 -1 was observed during repetitive vagal nerve stimulation (0 .1-0 .5V) (Fig. 3B). The increase in aK slowly declined toward control levels when electrical stimulation of the vagal nerves con- tinued for more than 1 min. Electrical stimulation of vagal nerves ( 0 . 1 - 0.5V) excited nerve fibres conduct- ing with a velocity of 1 7 - 5 7 m . s -1.
In preparations with intact vagal nerves, in addition to the spontaneous inspiratory rise of aK, aK increased during lung inflation within the pool of INs of the dorsal population (Fig. 3A)
Repetitive electrical stimulation of the spinal cord (50 cps, 5 - 1 0 V , 0.05 ms) produced an increase ofaK of 0.3 - 1 mmol . 1-1 within the ventral pool of neurones but failed to alter the aK within the dorsal pool, although inspiratory BS-neurones had been activated antidromicaUy, as evidenced by the focal potential recorded by the reference electrode.
Periodic Changes in Extracellular Potassium Activity (Aa~) in the Vicinity of Individual Respiratory Cells
Whenever the ISM approached a spontaneously active cell, the changes in aK increased in amplitude. In most cases, a large, IS-SD-fractioned, 1 - 1 . 5 m s long extracellular spike was recorded by the reference elec- trode. The amplitudes of AaK generally ranged in the order of 0 . 1 - 0 . 5 m m o l .1-1. Sometimes the ISM penetrated cells and the K +-signal increased abruptly to a value indicating an aK of about 4 0 - 80 mmol �9 1-1 This K+-signal did not remain stable but returned to control levels within a few seconds.
In the vicinity of BS-INs of both populations, the aK increased during inspiration, parallel to the frequency of the spike discharge, and in most recordings reached a maximum at the end of inspiration. During expiration, al~ decreased approximately exponentially. The half decay time was 1 - 2 s . With the vagi sectioned, a second peak in aK was not observed during expiration (Fig. 4A). In the vicinity of those inspiratory NAA neurones of the dorsal group, which were driven by vagal afferent discharge, the periodicity of the aK- fluctuations had a double peak in animals with intact vagal nerves. The second air increase followed the afferent vagal discharge elicited by artificial lung inflation.
In the vicinity of ENs of the ventral group, aK increased during their active period and was quite similar to the changes in aK within the ECS of INs. At the end of expiration, however, aK rapidly decreased but then revealed a plateau or a second wave before falling to a minimum at the very end of inspiration (Fig. 4B,5A). The changes in a K were similar in animals with intact or sectioned vagal nerves. The
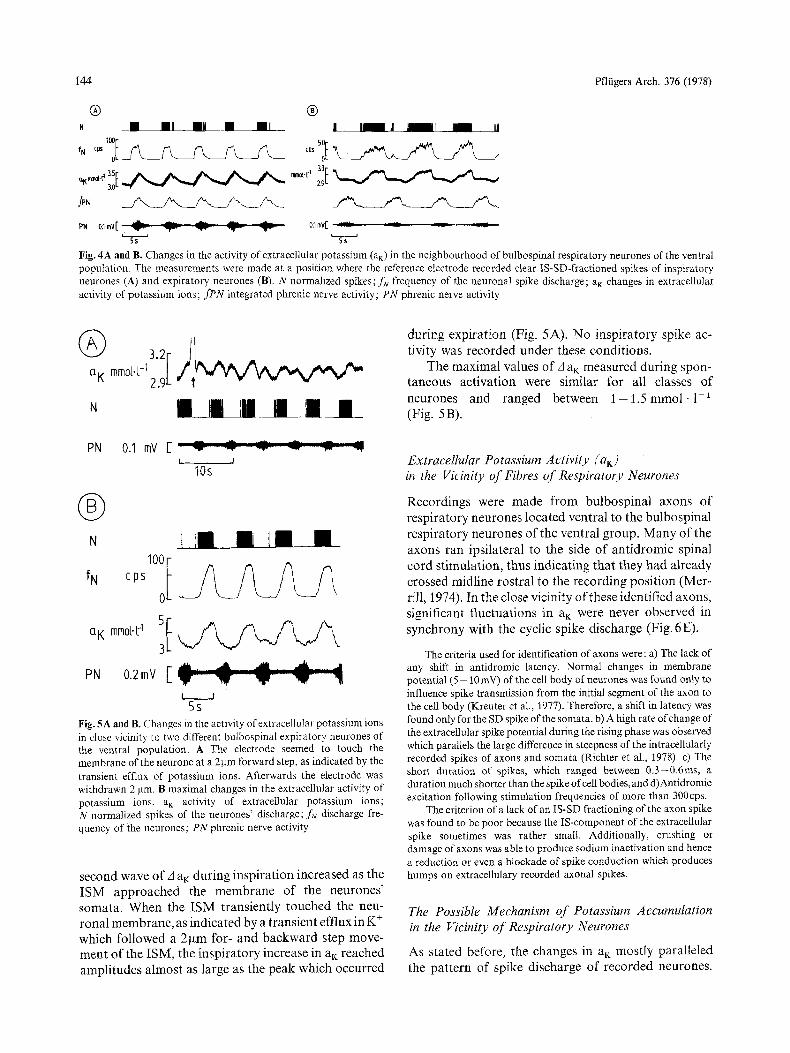
144 Pfltigers Arch. 376 (1978)
| | N �9 I l l UUU �9 H I I | I i i i II
, o o [ ~ so fN cps c~ o[ L ~ , "
0
3,0 L ~ ~ . . . .
PI',I 0:I m V [ ~ ~ ~ ~ ~ 0.1 rnv[ ~ ~ ~ . -
J 5 $ ~ L 5 s J
Fig. 4A and B. Changes in the activity of extracellular potassium (aK) in the neighbourhood of bulbospinal respiratory neurones of the ventral population. The measurements were made at a position where the reference electrode recorded clear IS-SD-fractioned spikes of inspiratory neurones (A) and expiratory neurones (B). N normalized spikes; fN frequency of the neuronal spike discharge; aK changes in extracellular activity of potassium ions; fPN integrated phrenic nerve activity; PN phrenic nerve activity
aK mm~ I 2.9
N | I I III I i I i I I
PN 0.1 mV [ :: ~,- ~- "," "~ ",- I I
tOs
| N i l l ! 1[] []
0a
ON 0.2mY [
t ~ J
5s Fig. 5A and B. Changes in the activity of extracellular potassium ions in dose vicinity to two different bulbospinaI expiratory neurones of the ventral population. A The electrode seemed to touch the membrane of the neurone at a 2 gm forward step, as indicated by the transient efflux of potassium ions. Afterwards the electrode was withdrawn 2 ~tm. B maximal changes in the extracellular activity of potassium ions. a K activity of extracellular potassium ions; N normalized spikes of the neurones' discharge; f~. discharge fre- quency of the neurones; PN phrenic nerve activity
second wave o f A a~: dur ing insp i ra t ion increased as the I S M app roached the m e m b r a n e of the neurones ' somata . When the I S M t rans ient ly touched the neu- rona l membrane , as indica ted by a t ransient efflux in K + which fol lowed a 2 g m for- and b a c k w a r d step move- men t o f the ISM, the insp i ra to ry increase in aK reached ampl i tudes a lmos t as large as the peak which occurred
dur ing expi ra t ion (Fig. 5A). N o insp i ra to ry spike ac- t ivi ty was recorded under these condi t ions .
The max imal values o f A a K measured dur ing spon- taneous ac t iva t ion were s imilar for all classes o f
neurones and ranged between 1 - 1 . 5 r e t o o l . 1 - 1 (Fig. 5B).
Extracellular Potassium Activity (aK) in the Vicinity of Fibres of Respiratory Neurones
Recordings were made f rom bu lbosp ina l axons o f resp i ra tory neurones loca ted ventral to the bu lbosp ina l resp i ra tory neurones o f the ventral group. M a n y o f the axons ran ipsi la teral to the side o f an t id romic spinal co rd s t imula t ion , thus indica t ing tha t they had a l ready crossed midl ine ros t ra l to the recording pos i t ion (Mer- rill, 1974). In the close vicinity o f these identif ied axons, significant f luctuat ions in a K were never observed in synchrony with the cyclic spike discharge (Fig. 6E).
The criteria used for identification of axons were: a) The lack of any shift in antidromic latency. Normal changes in membrane potential (5-- 10 mV) of the cell body of neurones was found only to influence spike transmission from the initial segment of the axon to the cell body (Kreuter et al., 1977). Therefore, a shift in latency was found only for the SD spike of the somata, b) A high rate of change of the extracellular spike potential during the rising phase was observed which parallels the large difference in steepness of the intracellularly recorded spikes of axons and somata (Richter et al., 1978). c) The short duration of spikes, which ranged between 0.3-0.6ms, a duration much shorter than the spike of cell bodies,and d)Antidromic excitation following stimulation frequencies of more than 300eps.
The criterion of a lack of an IS-SD fractioning of the axon spike was found to be poor because the IS-component of the extracellular spike sometimes was rather small. Additionally, crushing or damage of axons was able to produce sodium inactivation and hence a reduction or even a blockade of spike conduction which produces humps on extracellulary recorded axonal spikes.
The Possible Mechanism of Potassium Accumulation in the Vicinity of Respiratory Neurones
As stated before, the changes in aK most ly para l le led the pa t t e rn of spike discharge o f recorded neurones.
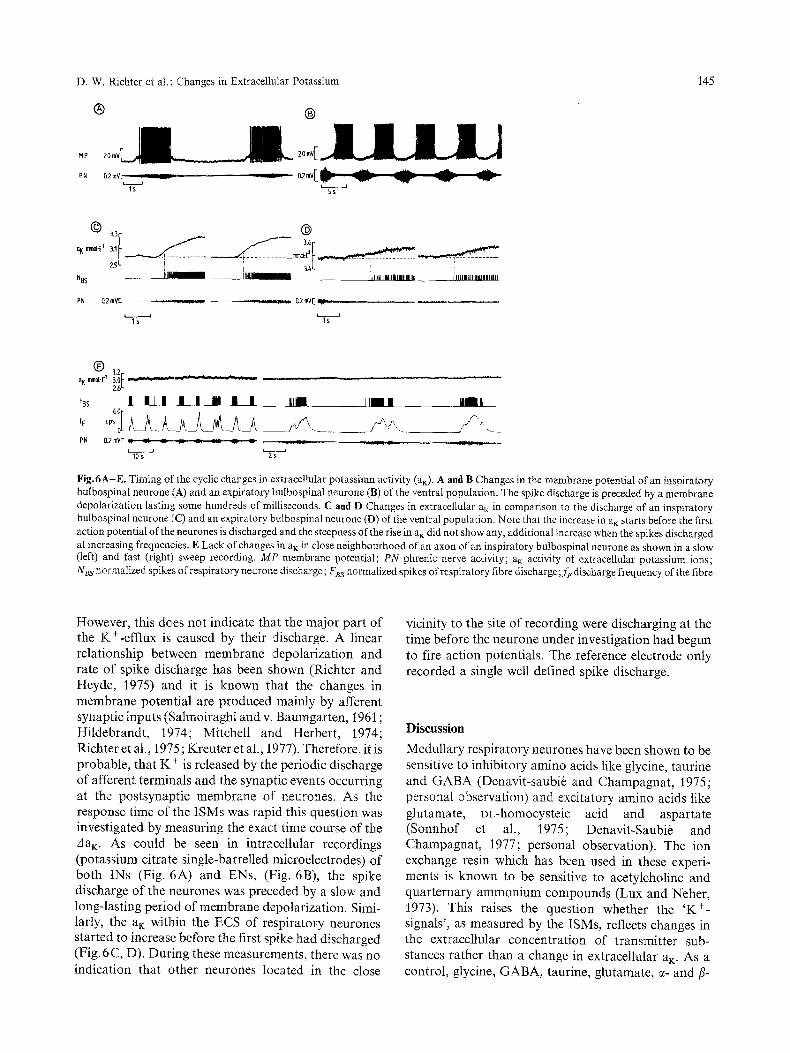
D. W. Richter et al. : Changes in Extracellular Potassium
| |
145
tip
PN ff2mV[ I U I J . - - 0.2 mVi- ~.-- ~ ~ ~
L ~ S J ' 5 S J ~ ~
ma-0 3.1 .... ~ r ~ 2s 3., '!
NB S N i l ] IIIii ....... Il l l(l l I l l III111 I I I I _ _ I l l l l UllllHllll I HIIIIII
PN 0.2 mVE : : : z . ; ; ; : ".._.._ . : . , , ~ 0.2 mV[ ~ ~-'~
i I I Is I Is
1 ~ 3.2 mraol.'F 1 3.01" . u ~ : ~ - . - - - _ _ = - - - - - ~ - ~ - _ ~ .v_ - - _ _ , . . . . I1K
2 3 k
FBS I II | I I Ill | I __llll l lIIII
+F cp, 0 j ~ . _ _ _ _ / . + ,
i i I IOs J 2s
/~-," t,
J ~ t~ r . . ~
Fig.6A-E. Timing of the cyclic changes in extracellular potassium activity (am). A and B Changes in the membrane potential of an inspiratory bulbospinal neurone (A) and an expiratory bulbospinal neurone (B) of the ventral population. The spike discharge is preceded by a membrane depolarization lasting some hundreds of milliseconds. C and D Changes in extracellular a m in comparison to the discharge of an inspiratory bulbospinal neurone (C) and an expiratory bulbospinal neurone (D) of the ventral population. Note that the increase in a m starts before the first action potential of the neurones is discharged and the steepness of the rise in a~ did not show any, additional increase when the spikes discharged at increasing frequencies. E Lack of changes in a m in close neighbourhood of an axon of an inspiratory bulbospinal neurone as shown in a slow (left) and fast (right) sweep recording. MP membrane potential; PN phrenic nerve activity; a K activity of extracellular potassium ions; NBs normalized spikes of respiratory neurone discharge; F~s normalized spikes of respiratory fibre discharge; f~ discharge frequency of the fibre
However, this does not indicate that the major part of the K+-efflux is caused by their discharge. A linear relationship between membrane depolarization and rate of spike discharge has been shown (Richter and Heyde, 1975) and it is known that the changes in membrane potential are produced mainly by afferent synaptic inputs (Salmoiraghi and v. Baumgarten, 1961 ; Hildebrandt, 1974; Mitchell and Herbert, 1974; Richter et al., 1975; Kreuter et al., 1977). Therefore, it is probable, that K + is released by the periodic discharge of afferent terminals and the synaptic events occurring at the postsynaptic membrane of neurones. As the response time of the ISMs was rapid this question was investigated by measuring the exact time course of the A a K. As could be seen in intracellular recordings (potassium citrate single-barrelled microelectrodes) of both INs (Fig. 6A) and ENs, (Fig. 6B), the spike discharge of the neurones was preceded by a slow and long-lasting period of membrane depolarization. Sinai- larly, the a K within the ECS of respiratory neurones started to increase before the first spike had discharged (Fig. 6C, D). During these measurements, there was no indication that other neurones located in the close
vicinity to the site of recording were discharging at the time before the neurone under investigation had begun to fire action potentials. The reference electrode only recorded a single well defined spike discharge.
Discussion
Medullary respiratory neurones have been shown to be sensitive to inhibitory amino acids like glycine, taurine and GABA (Denavit-saubi~ and Champagnat , 1975; personal observation) and excitatory amino acids like glutamate, DL-homocysteic acid and aspartate (Sonnhof et al., 1975; Denavit-Saubie and Champagnat , 1977; personal observation). The ion exchange resin which has been used in these experi- ments is known to be sensltive to acetylcholine and quarternary ammonium compounds (Lux and Neher, 1973). This raises the question whether the 'K +- signals', as measured by the ISMs, reflects changes in the extracellular concentration of transmitter sub- stances rather than a change in extracellular aK. As a control, glycine, GABA, taurine, glutamate, e- and fi-

146 Pflfigers Arch. 376 (1978)
alanine and aspartate have been tested and the ISMs were found to be insensitive to these substances (Sonnhof et al., 1977b). Therefore, only a release of acetylcholine by afferent nerve fibres, ending on medul- lary respiratory neurones might be able to simulate an increase in extracellular aK. However, an effect of acetylcholine upon BS-respiratory neurones seems to be absent (Salmoiraghi and Steiner, 1963). The parallel change in the ISM-signal and the reduction of the afterhyperpolarization of action potentials of bulbospinal respiratory neurones indicate an accumu- lation of the K* in addition to an eventual release and accumulation of acetylcholine. Likewise, the double peaked activity of the signal, measured by the ISM in the vicinity to expiratory neurones, cannot be explained by acetylcholine release alone because different trans- mitter substances would be expected to be released during neuronal excitation and inhibition. In general, the transmitter substances released at distinct synapses are effectively inactivated by specific enzyme systems and the transmitter would not be expected to diffuse in large quantities beyond the synaptic cleft. In the case of INs, however, changes in aK are measured at consider- able distances away from individual cells. If the poten- tials of the ISMs were generated by a small increase in acetylcholine concentration, the reference barrel of the ISMs would never reveal diffusion potentials as large as those seen in these experiments (Fig. 3A, B). l'his makes the assumption that the ISM-recorded signals reflect changes in extracellular ai( highly probable.
Medullary respiratory neurones generally discharge bursts of action potentials during one phase of respiration. The period of activity may last for several seconds and during that time, the membrane potential is depolarized. In this state, the pot+ntial differs significantly from the equilibrium potential for K +, as indicated by the afterhyperpolarization of the action potential, which is known to be generated by a K+-current (Coombs et al., 1955). Assuming a re- latively high PK also during 'resting' conditions (Richter et al., 1975; Kreuter et al., 1977), any long- lasting membrane depolarization should result in a continuous flow of K + as a consequence of an increased driving potential (Frankenhaeuser and Hodgkin, 1956; Alving, 1969; Neher and Lux, 1973; Lux and Eckert, 1974; Dubois and Bergman, 1975; Barrett and Barrett, 1976). This outward current of K + may diminish with time because of PK inactivation (Frankenhaeuser, 1963; Bergman, 1970; Lux and Heyer, 1975) or because of a shift of the equilibrium potential due to accumu- lation of extracellular K + (Alving, 1969; Eaton, 1972; Adelman, Jr. et al., 1973; Heyer and Lux, 1976).
In addition to such a 'passive' outward current at a 'resting' PK during each membrane depolarization,
K + can be expected to flow because of an increased PK resulting from the postsynaptic activity of re- spiratory neurones or by the excitation of afferent terminals impinging upon them. These synaptic events may be the most important mechanisms for K § from the respiratory neuronal network.
The behaviour of medullary respiratory neurones has been compared with pacemaker neurones (Brodie and Borison, 1956; Salmoiraghi and v. Baumgarten, 1961; B6hmer et al., 1976; Wyman, 1977). Although there is no evidence to confirm this idea, the spon- taneous activity of pacemaker and respiratory neu- rones is similar. Both types of cells discharge a long- lasting burst of action potentials which may continue for several seconds. Within the ECS of spontaneously bursting pacemaker neurones a slow increase in extra- cellular aK has been shown to occur during this dis- charge which rapidly declines to baseline during the post-burst membrane hyperpolarization (Lux and Heyer, 1975; Heyer and Lux, 1976). The amplitude of the AaK ranges between 0 .3- 0.5 mmol �9 1-1, which is only about one third of the amplitude of the change which can be measured in the close neighbourhood of respiratory neurones. In contrast to Aplysia burster neurones, the main portion of the K+-fiuxes seems to arise from synaptic activation of the respiratory neu- rones rather than from somato-dendritic spiking. This is consistent with the observation that repetitive anti- dromic excitation did not produce a significant increase in aK as seen at the level of the bulbospinal INs of the dor- sal group. In addition, the early increase in extracellular aK, preceded any change in the discharge of neighbour- ing respiratory neurones. However, this sort of obser- vations do not differentiate whether the discharge of presynaptic axon terminals or the postsynaptic pro- cesses generate the increase of a K.
Beside an increase of aK during neuronal excitation, expiratory neurones revealed an outward flow of K + during inspiration, an effect which could not be attri- buted to the discharge of adjacent INs. This result may suggest a synaptic inhibition of ENs during inspiration (Mitchell and Herbert, 1974; Richter et al., 1975; Kreuter et al., 1977), as a C1--inward-current (Lux, 1971), may be accompanied by a K+-outward current. As similar K +-fluxes were weak in the vicinity of INs, this indicates that reciprocal inhibition of INs arises at the dendrites whereas in ENs inhibition occurs near to the soma (Richter et al., 1975; Kreuter et al., 1977). If the synaptic inhibition of neurones is exclusively me- diated by a C1--current, the increase in a~: recorded during the period of inhibition would suggest that K + is released from axon terminals.
The finding, that cyclic changes in aK can b e measured anywhere within the area where INs are located, indicates that INs form a dense and uniformly

D. W. Richter et al. : Changes in Extracellular Potassium 147
distributed source of K +. This is particularly the case for the INs of the ventral group. In contrast, ENs re- semble dispersed and point-like sources as K +-signals were only recorded in the close vicinity of these cells. In the case of a point source, ejecting K + for about 2s, aK would decrease monotonically afterwards with a half decay time of 2 - 3 s and the asymptotic level of aK would be reached after 5 -10 s. Any prolongation of the time of K+-ejection or a dispersed distribution of many K+-sources would result in a slowing of K +- reequilibration. This has been shown by Lux and Neher (1973) who electrophoretically injected K + into the cerebral cortex of cats. In the neighbourhood of ENs, however, the initial fall of a K following activation of neurones, was found to be steeper than that in the neighbourhood of INs. As ENs revealed a K+-effiux also during inspiration, such a prolonged release of K + would be expected to slow K+-reequilibration within the ECS. Similarly, K + reequilibration should be slow within the ECS of the uniformly distributed K + sources of INs and as a consequence, K +-accumulation should occur. A half decay time of only 1 - 2 s for INs and the steepness of the initial fall of aK in the ECS of ENs, therefore indicate that an active K +-uptake by glial or neural elements dominates. An active uptake of K + by permanently bursting respiratory neurones is likely to be important in the maintenance of their intracellular concentration of K +.
The largest changes in ai~ were observed in the close vicinity of both INs and ENs. This change is likely to be underestimated, as the 2-31am tips of the ISMs undoubtedly damaged the area from which re- cordings were made, especially in a pulsating tissue like the lower brain-stem. A dead space of some 10-15 ~tm in 'diameter' might result (Herz et al., 1969). This would mean that the actual changes in external aK at the surface of the neuronal membranes probably exceed the values measured in both steepness and amplitude.
Accumulation of K + results in a depolariza- tion of excitable membranes (Huxley and Stgmpfli, 1951 ; Frankenhaeuser and Hodgkin, 1956; Sonnhofet al., 1977a; Erulkar and Weight, 1977; Richter et al., 1977 b), but it is difficult to calculate the strength of the K+-induced membrane depolarization in neurones receiving a permanent synaptic input. Electrical stimu- lation of the mesencephalic reticular formation has been shown to produce an increase of 2 - 3 mmol �9 1-1 in extracellular aK within the lateral geniculate nucleus (Singer and Lux, 1973). This increase was accompanied by a 1 - 3 mV depolarization of neurones with a similar time course. The same increase in aK within the spinal cord after tetanic stimulation of the sciatic nerve was predicted to result in a 15 mV depolarization of pri- mary afferents according to the Nernst-equation (Vyklick) et al., 1972). In myelinated nerve fibres of the
flog, however, Huxley and Stfimpfli (1951) measured a membrane depolarization of about 5 mV when elevat- ing extracellular a~ from 2.5 - 5 mmol �9 1- !. This is in the order of what can be measured in frog spinal moto- neurones after manganese application had blocked chemical synaptic transmission (Richter et al., in pre- paration). The changes in aK recorded in these experi- ments are of similar magnitude to those tested in the frog experiments and should depolarize the postsynap- tic elements of respiratory neurones by up to 5mV. In the intact nervous system, however, the effect of an a K- excess should vary with every change in membrane permeability of the neurones produced by synaptic inputs. Thus a synchronizing effect of activity may result when groups of neurones with a low level of activation are located together in close proximity.
In addition, K+-excess should depolarize the pre- synaptic elements producing presynaptic inhibition (Liley, 1956; Hagiwara and Tasaki, 1958; Hubbard and Willis, 1962; Takeuchi and Takeuchi, 1962; Llin/ts, 1968; Erulkar and Weight, 1977). In the spinal cord, primary afferent depolarization may also be explained in part by K +-accumulation (Vycklick) et al., 1972; ten Bruggencate et al., 1974; Kfi2 et al., 1975; Sykovit et al., 1976) and likewise in the lateral geniculate nucleus (Singer and Lux, 1973), the thalamocortical system (Gutnick and Prince, 1972; Lux, 1976) and the cuneate nucleus (Krnjevib and Morris, 1974). A similar type of presynaptic inhibition within the respiratory system would develop in parallel with neuronal excitation and may work with a time delay longer than the membrane time constants. This steady increase in presynaptic inhibition may represent one of the timing cues which have to be postulated for the mechanism generating the respiratory rhythm.
Another 'inhibitory' effect of an increased a,: is predicted for axons which undergo an increase in diameter (Grossman et al., 1973; Parnas et al., 1976; Hart and Smith, 1976; Smith and Hatt, 1976). This is the case at points of branching. The longitudinal current generated by an action potential of the parent axon is split between the daughter branches and has to charge an enlarged capacitance. A K+-accumulation would depolarize the membrane of the daughter branches and, in addition to the already lowered safety factor of conduction in this part of the axon, may disturb conduction and even may cause a conduction block due to sodium inactivation. This could result in a block of the high frequency spike transmission within the af- ferent fibre system of respiratory neurones which pro- gressively develops during the respiratory cycle.
Extracellular accumulation of K + in the vicinity of respiratory neurones may therefore produce antagonis- tic effects. The postsynaptic elements may reveal exci- tatory effects, which in a homogenous pool of adjacent

148 Pfliigers Arch. 376 (1978)
p o s t s y n a p t i c e l e m e n t s w o u l d c a u s e s y n c h r o n i z e d ac-
t ivi ty. A t t he p r e s y n a p t i c site, K + - a c c u m u l a t i o n m a y
r e s u l t in p r e s y n a p t i c i n h i b i t i o n .
Acknowledgements'. The authors express thanks to Dr. G. ten Bruggencate for kindly supplying potassium ion exchange resin, Mrs. G. Froelich for photographing and Mrs. K. Issbriicker for typing the manuscript.
This investigation was supported by the Deutsche Forschungs- gemeinschaft, Grant Ri 279/5.
References
Adelman, W. J. Jr., Palti, Y., Senft, J. P. : Potassium ion accumu- lation in a periaxonal space and its effects on the measurement of membrane potassium ion conductance. J. Membr. Biol. 13, 387-410 (1973)
Alving, B. O. : Differences between pacemaker and non-pacemaker neurones of Aplysia on voltage clamping. J. Gen. Physiol. 54, 512-531 (1969)
Barrett, E. F., Barrett, J. N.: Separation of two voltage-sensitive potassium currents, and demonstration of a tetrodotoxin-resis- taut calcium current in frog motoneurones. J. Physiol. (Loud.) 255, 737- 774 (1976)
von Baumgarten, R. : Koordinationsformen einzelner Ganglienzellen der rhombencephalen Atemzentren. Pfltigers Arch. ges. Physiol. 262, 573-594 (1965)
Baylor, D. A., Nicholls, J. G. : Changes in extracellular potassium concentration produced by neuronal activity in the central nervous system of the leech. J. Physiol. (Lond.) 203, 555-569 (1969)
Bergman, C. : Increase of sodium concentration near the inner surface of the nodal membrane. Pfl/igers Arch. 317, 287-302 (1970)
Berman, A. L. : The brain stem of the cat. A cytoarchitectonic atlas with stereotaxic coordinates. London: The University of Wisconsin Press 1968
Bianchi, A. L. : Localisation et ~tude des neurones respiratoires bulbaires. Mise en jeu antidromique par stimulation spinale ou vagale. J. Physiol. (Paris) 63, 5 - 4 0 (1971)
B6hmer, G., Chaplain, R. A., Fallert, M.: Metabolic control of respiratory neuronal activity and the accompanying changes in breathing movements of the rabbit. II. Manipulation of expi- ratory and inspiratory-expiratory neurons. Pfl/igers Arch. 365, 61 -68 (1976)
Brodie, D. A., Borison, H. L.: Evidence for a medullary inspiratory pacemaker. Functional concept of central regulation of re- spiration. Am. J. Physiol. 188, 347-354 (1957)
ten Bruggencate, G., Lux, H. D., Liebl, L.: Possible relationship between extracellular potassium activity and presynaptic in- hibition in the spinal cord of the cat. Pfliigers Arch. 349, 301 - 317 (1974)
Cohen, M. I.: How respiratory rhythm originates: Evidence from discharge patterns of brainstem respiratory neurones. In: Breathing: Hering-Breuer Centenary Symposium (R.Porter ed.), pp. 125-157. London: Churchill 1970
Cohen, M. I.: The genesis of respiratory rhythmicity. In: Central rhythmic and regulation (W. Umbach, H. P. Koepchen, eds.), pp. 15-35. Stuttgart: Hippokrates 1974
Cohen, M. I., Feldman, J. L. : Models of respiratory phase-switching. Fed. Proc. 36, 2367-2374 (1977)
Coombs, J. S., Eccles, J. C., Fatt, P. : The electrical properties of the motoneurone membrane. J. Physiol. (Lond.) 130, 291-325 (1955)
Denavit-SaubiO, M., Champagnat, J. : The effect of some depressing amino acids on bulbar respiratory and non-respiratory neurons. Brain Res. 97, 356-361 (1975)
Denavit-Saubi6, M., Champagnat, J. : Amino acids and rhythmic repiratory discharge. In: Iontophoresis and transmitter me- chanisms in the mammalian central nervous system, pp. 7 .7 - 7.9. Cambridge: Churchill College 1977
Dubois, J. M., Bergmann, C.: Potassium accumulation in the peri- nodal space of frog myelinated axons. Pflfigers Arch. 358, 1 l 1 - 124 (1975)
Eaton, D. C. : Potassium ion accumulation near a pace-making cell of Aplysia. J. Physiol. (Lond.) 224, 421-440 (1972)
Erulkar, S. D., Weight, F. F. : Extracellular potassium and transmit- ter release at the giant synapse of squid. J. Physiol. (Lond.) 266, 209--218 (1977)
von Euler, C., Hayward, J. N., Marttila, 1., Wyman, R. J.: Respiratory neurones of the ventrolateral nucleus of the solitary tract of cat: VagaI input, spinal connections and morphological identification. Brain Res. 61, 1 - 2 2 (1973)
Frankenhaeuser, B.: A quantitative description of potassium cur- rents in myelinated nerve fibres of Xenopns Laevis. J. Physiol. (Loud.) 169, 424-430 (1963)
Frankenhaeuser, B., Hodgkin, A. L. : The after-effects of impulses in the giant nerve fibres ofLoligo. J. Physiol. (Loud.) 131,341 - 376 (1956)
Grossman, Y., Spira, M. E., Parnas, I. : Differential flow of infor- mation into branches of a single axon. Brain Res. 64, 379-386 (1973)
Gutnick, M. J., Prince, D. A.: Thalamocortical relay neurons: Antidromic invasion of spikes from a cortical epileptogenic focus. Science 176, 424-425 (1972)
Haber, E., Kohn, K. W., Ngai, S. H., Holiday, D. A., Wang, S. C. : Localization of spontaneous respiratory neuronal activities in the medulla oblongata of the cat: a new location of the expiratory center. Am. J. Physiol. 190, 350-355 (1957)
Hagiwara, S., Tasaki, I. : A study on the mechanism of impulse transmission across the giant synapse of the squid. J. Physiol. (Lond.) 143, 114-137 (1958)
Hatt, H., Smith, D. O.: Synaptic depression related to presynaptic axon conduction block. J. Physiol. (Loud.) 259, 367-393 (1976)
Heinemann, U., Lux, H. D. : Undershoots following stimulus-in- duced rises of extracellular potassium concentration in cerebral cortex of cat. Brain Res. 93, 63 -76 (1975)
Herz, A., Ziegelg/insberger, W., Ffirber, G. : Microelectrophoretic studies concerning the spread of glutamic acid and GABA in the tissue. Exp. Brain Res. 9, 221-235 (1969)
Heyer, C. B., Lux, H. D. : Control of the delayed outward potassium currents in bursting pace-maker neurones of the snail, Helix Pomatia. J. Physiol. (Lond.) 262, 349--382 (I976)
Hildebrandt, J. R. : Intracellnlar activity of medullary respiratory neurons. Exp. Neurol. 45, 298-313 (1974)
Hotson, J. R., Sypert, G. W., Ward, A. A., Jr.: Extracellular potassium concentration changes during propagated seizures in neocortex. Exp. Neurol. 38, 20 -26 (1973)
Hubbard, J. I., Willis, W. D. : Reduction of transmitter output by depolarization. Nature (Lond.) 193, 1294-1295 (1962)
Hugelin, A., Cohen, M. I.: The reticular activating system and respiratory regulation in the cat. Ann. N. Y. Acad. Sci. 109, 586-603 (1963)
Huxley, A. F., St/impfli, R. : Direct determination of membrane resting potential and action potential in single myelinated fibres. J. Physiol. (Lond.) 112, 476-495 (1951)
Kernell, D. : The early phase of adaptation in repetitive impulse dis- charge of cat spinal motoneurones. Brain Res. 41, 184-186 (1972)
Kreuter, F., Richter, D. W., Camerer, H., Senekowitsch, R.: Morphological and electrical description of medullary respi- ratory neurons of the cat. Pfliigers Arch. 372, 7 - 1 6 (1977)
K'fi~, N., Sykov~, E., Vyklick~,, L. : Extracellular potassium changes in the spinal cord of the cat and their relation to slow potentials,

D. W. R{chter et ai.: Changes in Extracellular Potassium 149
active transport and impulse transmission, J. Physiol. (Lond.) 249, 167- t82 (1975)
Krnjevi6, K., Morris, M. E.: Extracellular K + activity and stow potential changes in spinal cord and medulla. Can. J. Physiol. Pharmacol. 50, 1214-1217 (1972)
Krnjevid, K., Morris, M. E.: Extracellular accumulation of K + evoked by activity of primary afferent fibers in the cuneate nuclens and dorsal horn of cats. Can. J. Physiol. Pharmacol. 52, 852- 871 (1974)
Liley, A. W. : The effects of presynaptic polarization on the spon- taneous activity at the mammalian neuromuscular junction. J, Physiol. (Lond.) t34, 427-443 (1956)
Llin/ts, R.: A possible mechanism for presynaptic inhibition. In: Structure and Function of Inhibitory Neuronal Mechanisms (C. yon Euler, B. Skoglund, U. S6derberg, eds.), pp. 249-250. London: Pergamon Press 1968
Lothman, E. W., Somjen, G. G. : Extracellular potassium activity, intracellular and extracellular potential responses in the spinal cord. J. Physiol. (Lond.) 252, 115-I36 (1975)
Lux, H. D. : Ammonium and chloride extrusion: Hyperpolarizing synaptic inhibition in ,spinal motoneurons. Science 173, 555- 557 (1971)
Lux, H. D. : Kaliumaktivit~it im Cortex der Katze : Untersuchungen zur experimentellen Epitepsie, Arch. Psychiat. Nervenkr. 221, 227-244 (1976)
Lux, H. D., Eckert, R.: Inferred slow inward current in snail neurones. Nature (Lond.) 250, 574-576 (1974)
Lux, H. D., Heyer, C. B.: Fast K + activity determinations during outward currents of the neuronal membrane of Helix Pomatia. Biochem. Bioenerg. 3, 169-182 (1975)
Lux, H. D., Neber, E_: The equilibration time course of [K +]o in cat cortex. Exp. Brain Res. 17, 190-205 (1973)
Merrill, E. G. : The lateral respiratory neurones of the medulla: their associations with nucleus ambiguus, nucleus retroambigualis, the spinal accessory nucleus and the spinal cord. Brain Res. 24, 11-28 (1970)
Merrill, E. G. : Finding a respiratory function for the medullary respiratory neurons. In: Essays on the nervous System (R. Bellairs, E. G. Gray, eds.), pp. 451--486. Oxford: Clarendon Press 1974
Michell, R. A., Herbert, D. A. : Synchronized high frequency synaptic potentials in medullary respiratory neurons. Brain Res. 75, 350- 355 (1974)
Neher, E., Lux, H. D. : Rapid changes of potassium concentration at the outer surface of exposed neurons during membrane current flow. J. Gen. Physiol. 61,385-399 (1973)
Nelson, J. R. : Single unit activity in medullary respiratory centers of cat. J. Neurophysiol. 22, 590-598 (1959)
Parnas, I., Hochstein, S., Parnas, H.: Theoretical analysis of parameters leading to frequency modulation along an in- homogenous axon. J. Neurophysiol. 39, 909-923 (1976)
Prince, D. A., Lux, H. D., Neher, E. : Measurements of extracellular potassium activity in cat cortex. Brain Res. 50, 489-495 (1973)
Richter, D. W., Heyde, F.: Accommodative reactions of medullary respiratory neurons of the cat. J. Neurophysioh 38, 1172-1180 (1975)
Richter, D. W., Seller, H.: Response of medullary respiratory neurons to lung inflation and vagal nerve stimulation. Pfifigers Arch. 362, R 40 (1976)
Richter, D. W., Camerer, H , Sonnhof, U. : Effect of extracellular potassium accumulation produced by medullary respiratory activity, (In preparation)
Richter, D. W., Heyde, F., Gabriel, M. : Intracellular recordings from different types of medullary respiratory neurons of the cat. ). Neurophysiol. 38, 1162-1171 (1975)
Richter, D. W., Sonnhof, U., Camerer, H. : Changes in extracellular potassium concentration evoked by medullary respiratory ac- tivity of cats. Pflfigers Arch. 368, R 38 (1977a)
Richter, D. W., Sonnhof, U., Camerer, H. : Effects of extracellular potassium accumulation produced by medullary respiratory activity. Proc. Int. Union Physiol. Sci. 13, 629 (1977b)
Richter, D. W., Camerer, H., R6hrig, N.: Medullary inspiratory interneurons receiving a monosynaptic input from lung stretch receptors. Pflfigers Arch. 373, R 75 (1978)
Salmoiraghi, G. C., yon Baumgarten, R.: Intracellular potentials from respiratory neurones in brain-stem of cat and mechanism of rhythmic respiration. J. Neurophysiol. 24, 203-218 (1961)
Salmoiraghi, G. C., Burns, B. D. : Notes on mechanisms of rhythmic respiration. J. Neurophysiol. 23, 14-26 (1960)
Salmoiraghi, G. C., Steiner, F. A. : Acetylcholine sensitivity of cat's medullary neurons. J. Neurophysiol. 26, 581- 597 (1963)
Singer, W., Lux, H. D. : Presynaptic depolarization and extracellular potassium in the cat lateral geniculate nucleus. Brain Res. 64, 17-33 (1973)
Smith, D. O., Hatt, H. : Axon conduction block in a region of dense connective tissue in crayfish. J. Neurophysiol. 39, 794-801 (1976)
Sonnhof, U., Richter, D. W., Steinberg, R.: A tungsten electrode insulated by a concentric arrangement of glasspipettes for iontophoresis. Pflfigers Arch. 360, 45-48 (I975)
Sonnhof, U., Richter, D. W., Taugner, R. : Electrotonic coupling between frog spinal motoneurons. An electrophysiological and morphological study. Brain Res. 138, 197-215 (1977a)
Sonnhof, U., Grafe, P., Richter, D. W., Parekh, N , Krumnikl, G., Linder, M. : Investigations of the effects of glutamate on motoneurons of the isolated frog spinal cord. In: I ontophoresis and transmitter mechanisms in the mammitian central nervous system, pp. 7.29-7.31. Cambridge: Churchill College 1977b
Sykovfi, E., Shirayev0 B., K?i2, N., Vyklick), L.: Accumulation of extracellular potassium in the spinal cord of frog. Brain Res. 106, 413-417 (1976)
Takeuchi0 A., Takeuchi, N. : Electrical changes in pre- and postsyn- aptic axons of the giant synapse of Loligo, J. Gen. Physiol. 45, 1181 - 1193 (1962)
Vysko~il, F., K~i2,, N., Bureg, J. : Potassium-selective microelectrodes used for measuring the extracellular brain potassium during spreading depression and anoxic depolarization in rats. Brain Res. 39, 255-259 (1972)
Vyklick~, L., Sykov~t, E., K?is N., Ujec, E.: Post-stimulation changes of extracellular potassium concentration in the spinal cord of the rat. Brain Res. 45, 608--611 (1972)
Wyman, R. J. : Neural generation of the breathing rhythm. Ann. Rev. Physiol. 39, 417-448 (1977)
Received March 14, 1978