changes in bryozoan assemblages and bathymetric variations. examples from the messinian of northwest...
TRANSCRIPT

ELSEVIER Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326
Changes in bryozoan assemblages and bathymetric variations.Examples from the Messinian of northwest Algeria
Pierre Moissette *
Universite de Lyon I, UFR des Sciences de la Terre and FRE 2042 du CNRS, 27 Bd du 11 novembre, 69622 Villeurbanne Cedex, France
Received 1 February 1999; revised version received 23 June 1999; accepted 16 July 1999
Abstract
Studies of Recent bryozoan faunas have shown that there is a correlation between colonial growth forms and variousphysical factors of the environment, especially bathymetry. Each given assemblage of zoarial forms can thus be relatedto a certain depth interval. In marine Neogene deposits numerous extant bryozoan species occur and some of themhave more or less restricted depth ranges: they are used here to complement the indications given by the analysis ofgrowth forms. In the Messinian (uppermost Miocene) of northwestern Algeria a general feature is the occurrence ofwell-developed diatomaceous deposits interbedded with marl beds where benthic invertebrate remains are often abundant.In the five sections under study bryozoans are represented by 87 species belonging to nine different colonial growthforms. Four different assemblages of these zoarial forms, characterised by presence=absence and number of fragments, aredistinguished, corresponding to four depth intervals. The first and deepest assemblage (upper bathyal to lower circalittoral)occurs in the bottom part of the sections where colonies of a conescharelliniform species strongly predominate. The secondassemblage (lower circalittoral) is represented by several species belonging to various zoarial forms: conescharelliniform,cellariiform, vinculariiform, adeoniform, and reteporiform. The third assemblage (upper circalittoral) is characterised bythe absence of conescharelliniforms, but membraniporiforms and celleporiforms are abundantly represented, togetherwith numerous erect colony fragments. In the fourth assemblage (infralittoral) membraniporiforms and celleporiformspredominate, although a number of erect zoarial forms are sometimes associated in various proportions. The presenceof several extant bryozoan species associated with each of these four assemblages is used as a confirmation for thesuggested depth intervals. A gradual shallowing is thus indicated by the study of the bryozoan assemblages through thefive Messinian sections. The discussion shows that modern data on zoarial forms and species bathymetric distributionsare clearly applicable to Cenozoic faunas but that caution must be exercised when using results obtained from differentoceanographic regions. 2000 Elsevier Science B.V. All rights reserved.
Keywords: palaeobathymetry; palaeoecology; bryozoans; Cenozoic; Miocene; Algeria
1. Introduction
Marine sessile suspension feeders, bryozoans areconsidered as good palaeoecological and palaeo-
Ł E-mail: [email protected]
bathymetric tools (Stach, 1936; Lagaaij and Gautier,1965; Rucker, 1967; Schopf, 1969; Harmelin, 1988;McKinney and Jackson, 1989; Bone and James,1993; Smith, 1995; Hageman et al., 1997). Theyhave proved especially useful for the interpretation ofbathymetric variations in Tertiary geological sections
0031-0182/00/$ – see front matter 2000 Elsevier Science B.V. All rights reserved.PII: S 0 0 3 1 - 0 1 8 2 ( 9 9 ) 0 0 1 2 4 - 8

306 P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326
Fig. 1. Location map of the sections in the Messinian Chelif Basin (modified from Perrodon, 1957). Asterisks and numbers identify thesections: 1 D Arbal, 2 D Chabat el Haricha, 3 D Chabat Bou Seter, 4 D Hammar Semmoumet, 5 D Sig.
where changes in assemblages are more or less eas-ily evaluated (Labracherie and Prud’homme, 1966;Askren, 1968; Labracherie, 1970, 1973; Cheetham,1971; Braga, 1979; Nelson et al., 1988; Moissette,1988, 1993; James and Bone, 1991; Boreen andJames, 1995).
In the Messinian (uppermost Miocene) of theChelif Basin (northwest Algeria) as in other Mediter-ranean basins, the normal marine sedimentary suc-cession comprises marl deposits followed by analternation of diatomite beds with marl or clayeylimestones. This sequence, normally about 100 mthick, is succeeded by sands, sandstones and=orcoral and algal limestones (Anderson, 1933; Per-rodon, 1957; Gersonde, 1980; Rouchy, 1982; SaintMartin, 1990; Saint Martin et al., 1992). Gypsummay also be abundant near the top of some sections.Good outcrops are generally available in this region,with thick sedimentary sequences observable undervarious facies. In most marl and limestone bedsbryozoans are present and sometimes even abundantamong a fossil biota including coralline algae, fora-minifers, sponges, corals, serpulid worms, bivalves,
gastropods, cephalopods, brachiopods, decapods, os-tracods, echinoderms, and fishes (Moissette, 1988,1993).
The main purpose of this study is to assess thebathymetric variations occurring through a marinesedimentary succession using an analysis of the dis-tribution of bryozoan colonial morphotypes, com-plemented by information on known depth rangesof a number of extant species. In order to illus-trate this approach, an example of five sections fromthe Messinian of northwest Algeria has been chosen(Fig. 1). Bryozoans were found in 106 of the 120samples collected in these five sections. They be-long to 87 species of cyclostomes and cheilostomes,which have been divided into nine different colonial(or zoarial) growth forms.
2. Methods
Among the various ecological parameters govern-ing the distribution of bryozoans, water depth hasmainly been taken into account here but other factors
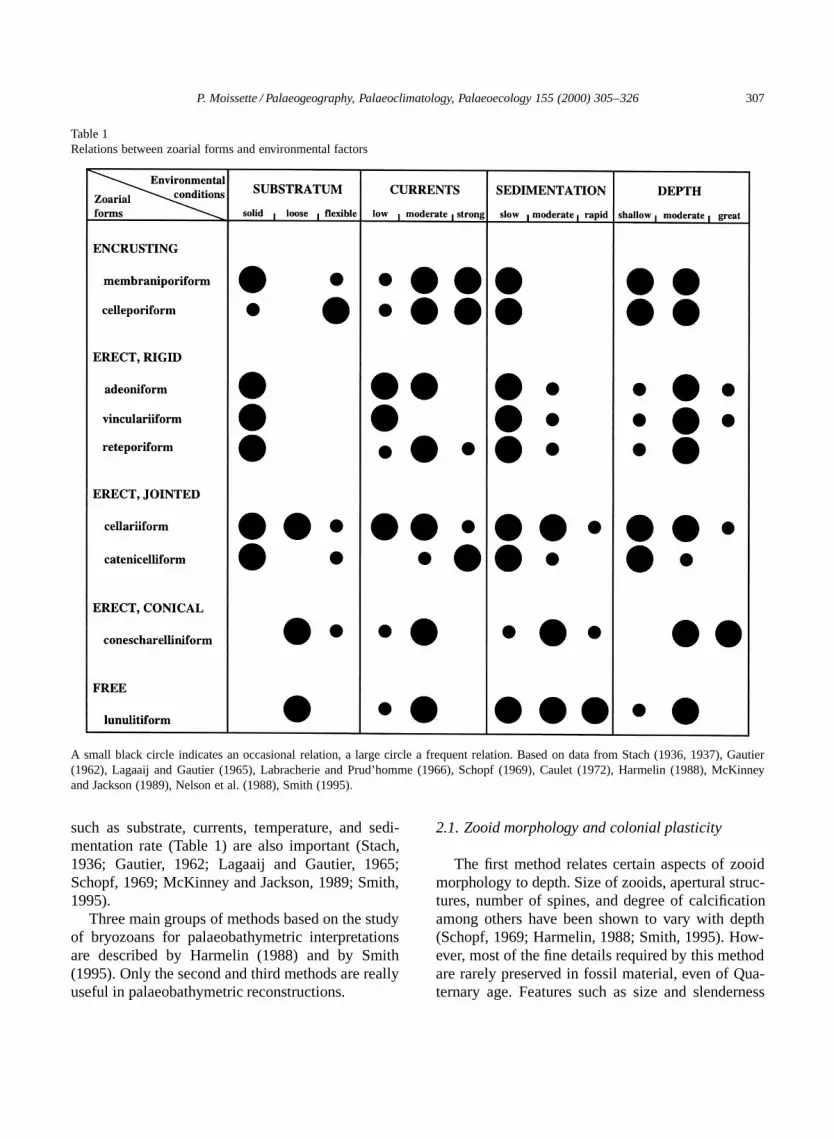
P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326 307
Table 1Relations between zoarial forms and environmental factors
A small black circle indicates an occasional relation, a large circle a frequent relation. Based on data from Stach (1936, 1937), Gautier(1962), Lagaaij and Gautier (1965), Labracherie and Prud’homme (1966), Schopf (1969), Caulet (1972), Harmelin (1988), McKinneyand Jackson (1989), Nelson et al. (1988), Smith (1995).
such as substrate, currents, temperature, and sedi-mentation rate (Table 1) are also important (Stach,1936; Gautier, 1962; Lagaaij and Gautier, 1965;Schopf, 1969; McKinney and Jackson, 1989; Smith,1995).
Three main groups of methods based on the studyof bryozoans for palaeobathymetric interpretationsare described by Harmelin (1988) and by Smith(1995). Only the second and third methods are reallyuseful in palaeobathymetric reconstructions.
2.1. Zooid morphology and colonial plasticity
The first method relates certain aspects of zooidmorphology to depth. Size of zooids, apertural struc-tures, number of spines, and degree of calcificationamong others have been shown to vary with depth(Schopf, 1969; Harmelin, 1988; Smith, 1995). How-ever, most of the fine details required by this methodare rarely preserved in fossil material, even of Qua-ternary age. Features such as size and slenderness

308 P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326
or robustness of colonies may also be used forpalaeobathymetric interpretation (Harmelin, 1988)and even in fossil, mostly fragmented, materialbranch diameters can still be determined in all butthe most heavily abraded specimens. Within-speciesvariations in branch diameter thus provide a reliablemeans of determining relative bathymetry.
2.2. Extant species
The second method is based on known depthranges of extant bryozoan species. These speciesare numerous in most upper Tertiary marine de-posits. This is also true for the five sections studiedin the Messinian of northwestern Algeria where 54(62%) of the 87 species identified still live in mod-ern seas. Their depth distribution, notably in theMediterranean, is generally well known from previ-ous studies (Gautier, 1962; Prenant and Bobin, 1966;Harmelin, 1969a,b, 1976; Hayward, 1974; Rylandand Hayward, 1977; Hayward and Ryland, 1979;Zabala, 1986; Rosso, 1996).
Although most of these species are nearly eu-rybathic (around 0 to 100–300 m), a number ofextant stenobathic species have more or less re-stricted bathymetric ranges (Table 2). Several speciesfound in the Messinian of northwest Algeria indicateshallow depths. On the contrary, other species arecharacteristic of deeper water. A number of speciesalso show an optimum bathymetric range that cangive some more indications whenever their frag-ments are abundant in a sample. Although perhapsyet to be demonstrated in bryozoans, the possibilityof changes through time in the depth distributions ofspecies should be considered, especially in view ofmore general observations implying offshore migra-tion of clades through time.
The most severe limitation for the application ofthis method is access to data on the depth distribu-tion of species. Indications given in the literature onvertical ranges are often old and thus very restricted,notably because of the mode of sampling (e.g. dredg-ing, not efficient on shallow rocky bottoms). Unfor-tunately, a large amount of valuable information onthe distribution of modern species, obtained by div-ing, remains unpublished (J.G. Harmelin, pers. com-mun., 1998; Moissette, 1999). Furthermore, depthsrecorded in the literature (and used in this paper)
Table 2Depth range and bathymetric optimum of some extant steno-bathic species
Species Depth range Bathymetric(m) optimum (m)
Calpensia nobilis 0–60Cryptosula pallasiana 0–50Haplopoma bimucronatum 20–40Hincksina flustroides 0–50Hippopodinella lata 0–30Scruparia ambigua 0–50Tubulipora plumosa 0–50Umbonula ovicellata 0–50
Gemellipora eburnea 75–1310Tervia irregularis 60–300Tessaradoma boreale 50–4300
Adeonella polystomella 20–130 30–60Cellaria salicornioides 0–250 ? 50–80Escharella variolosa 10–240 10–80Escharina dutertrei 10–280 100–150Exidmonea atlantica 10–850 40–100Hornera frondiculata 40–200 70–100Margaretta cereoides 10–90 10–45Nellia oculata 5–400 20–40Onychocella angulosa 0–80 30–50Porella cervicornis 30–120 40–60Schizoporella longirostris 0–150 20–60Schizotheca fissa 20–120 40–120Turbicellepora coronopus 20–200 40–60
should not be considered as very accurate. Theknown figures are likely to be more or less radicallyaltered by new oceanographic campaigns. This indi-cates that, more than isolated species, assemblagesshould be taken into account.
Another source of erroneous interpretation is thefact that many bryozoan species occupy cryptichabitats in fissures, caves and cavities where theyencounter environmental conditions normally pre-vailing in deeper, quieter, waters (Harmelin, 1969c,1985, 1997). In the Messinian of northwest Algeria,this is only the case for the limestones of the upperparts of the studied sections where corals and=orcoralline algae predominate, thus offering cavitieswhere erect colonies may find environments corre-sponding to deeper habitats.
Also, depth limits in a certain region may be dif-ferent in other regions of the same basin due to vari-ations in thermal regime and hydrological conditions(e.g. northern and southern parts of the Mediter-

P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326 309
ranean, or western and eastern Mediterranean). Thisis also true when one compares the depth distribu-tion of the same species in the Mediterranean andin the Atlantic (Harmelin and d’Hondt, 1993) whererecorded bathymetric ranges are notably deeper, es-pecially at their lower limit (e.g. Porella cervicornisfound between 30 and 120 m in the Mediterraneanhas a depth range of 30 to 400 m in the Atlantic).
2.3. Zoarial forms
The third, and probably the most promising,method is based on the known bathymetric distri-bution of colonial morphotypes, called zoarial forms.Developed by Stach (1936), the actualistic model us-ing these growth forms for palaeobathymetric recon-structions has been modified and applied by variousworkers (Stach, 1937; Lagaaij and Gautier, 1965;Labracherie and Prud’homme, 1966; Schopf, 1969;Cheetham, 1971; Braga, 1979; Nelson et al., 1988;Bianchi et al., 1988; Moissette, 1988, 1993; Boneand James, 1993; Moissette and Saint Martin, 1995;Smith, 1995; Hageman et al., 1998).
Fig. 2. Schematic diagrams of the zoarial forms used in this study.
The relations between zoarial forms and environ-mental conditions, especially depth, are now relativelywell established (Stach, 1936; Gautier, 1962; Lagaaijand Gautier, 1965; Labracherie and Prud’homme,1966; Schopf, 1969; Reguant et al., 1985, 1987; Har-melin, 1988; Nelson et al., 1988; Poluzzi and Rosso,1988; McKinney and Jackson, 1989; Bone and James,1993; Smith, 1995; Hageman et al., 1997).
Nine distinct bryozoan growth forms have beenidentified in this study (Fig. 2 and Table 1). Nodifference has been made here between cyclostomeand cheilostome colonial morphologies. The relativeabundance of each zoarial form is used as a measureof the depth interval at which a given assemblagelived, rather than simply using the parameters ofpresence or absence.
2.3.1. MembraniporiformThis encrusting unilaminar morphotype is at-
tached by cementation on many hard substrates (heremostly bivalve shells and other calcareous fragmentsof organisms). Although found also at greater depthsit predominates in shallow water. In the five sections

310 P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326
studied most species (49) belong to this zoarialform. There is no trace here of membraniporiform Bcolonies that encrust soft flexible substrates and aregenerally poorly calcified.
2.3.2. CelleporiformForming nodular massive or branching colonies
this morphotype is considered as living slightlydeeper than the previous one (Spjeldnaes and Mois-sette, 1997). Only five celleporiform species havebeen found in the sections under study. Fragmentsnot identifiable to genus or species (or even fam-ily) level are grouped under the general name of‘indeterminate celleporiform’.
2.3.3. AdeoniformForming erect, rigid bilaminate colonies, this mor-
photype lives at moderate depths on various solidsubstrates. Lagaaij and Gautier (1965) indicate thatit attains sizeable proportions (around 10% of thezoarial form assemblage) only at depths of ca 40–50m. Eight species belong to this zoarial form. Amongthem, Adeonella polystomella and Porella cervicor-nis have depth optima at about 30 to 60 m.
2.3.4. VinculariiformThis erect, rigid morphotype consists of sub-cyl-
indrical branches. Living mostly at moderate depthson a solid substrate it is also found in much deeper en-vironments. Eleven vinculariiform species have beenidentified in the five sections under study. At leasttwo of them (Tervia irregularis and Tessaradoma bo-reale) are deep stenobathic species (Rosso and DiGeronimo, 1998).
2.3.5. ReteporiformErect, rigid and fenestrate, this type of colony
lives on hard substrates at variable but mostly shal-low depths. Only three species belong here to thiszoarial form.
2.3.6. CellariiformErect articulated (by chitinous joints) and thus
flexible, each cylindrical segment being constitutedby numerous zooids, this zoarial form lives at variousdepths attached by its chitinous rootlets on differentsubstrates (loose and solid). As noted by Lagaaij andGautier (1965), Schopf (1969) and Reguant et al.
(1985) this is probably the colonial morphotype bestadapted to regions of high sedimentation rates. Inthe studied Messinian sections seven species belongto this morphotype represented in some samplesby thousands of fragments. Among them, the extantspecies Gemellipora eburnea is mostly found at greatdepths (Rosso and Di Geronimo, 1998), whereasMargaretta cereoides is a relatively shallow species.
2.3.7. CatenicelliformThis is also an erect articulated morphotype at-
tached by chitinous rootlets but each segment com-prises only one or two zooids. This morphotypeis considered as very shallow (Stach, 1937; Lagaaijand Gautier, 1965; Moissette and Saint Martin, 1995;Moissette, 1997). As noted by Lagaaij and Gautier(1965), these jointed forms “are generally so fragilethat their best chance of being preserved lies in areaswhere a moderate rate of deposition of fine sedimentprevails”. The only species of this group here is Scu-paria ambigua, represented in one limestone sampleby a very small number of specimens.
2.3.8. LunulitiformIn the Recent this free-living discoidal morpho-
type is abundant on particulate substrates at moder-ate depths: Cook and Chimonides (1983) indicate adepth range of 2–500 m with a preference for 20–70m, and Bone and James (1993) give a depth range of30–200 m. Only a very small number of specimensbelonging to two species have been found in two ofthe sections under study. This is probably due to theabsence of the sandy bottoms preferred by this zoar-ial form. Another factor contributing to the paucityof lunulitiform colonies may be dissolution of thesecharacteristically aragonitic bryozoans.
2.3.9. ConescharelliniformThis conical rooted morphotype is indicative of
muddy bottoms in moderately deep to deep environ-ments (Cook and Lagaaij, 1976; Cook, 1981; Gor-don, 1987). In the Messinian of Algeria, the onlyspecies belonging to this zoarial form is Batoporarosula which has also been found in numerous Neo-gene deposits of the Mediterranean (Moissette, 1996).

P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326 311
3. Analytical procedures
Since most samples were taken from marl levels,the procedure is fairly straightforward. For each sam-ple, 500 g of dry sediment was weighed, washed andsieved on a column of six sieves with diminishingmesh size (2, 1, 0.5, 0.25, 0.125, 0.063 mm). Bry-ozoan remains were found only in the four coarsersieves. For indurated calcareous levels, a number ofrock slabs have been sampled, washed and brushedin the laboratory. An estimate of the proportions ofthe various zoarial forms has then been made af-ter observing the prepared surfaces under the stereomicroscope.
For every sample of the five sections studied, eachbryozoan fragment has been counted as one unit.This is, of course, often a gross approximation sincesome of the colony types may break into numer-ous pieces (erect growth forms), whereas others aremostly preserved as whole colonies (membranipori-form and celleporiform). When using semi-quantita-tive representations (Figs. 3–7) it is, however, tol-erably precise, especially when considering mostlyvariations in relative abundance between verticallyspaced levels (Labracherie, 1973; Moissette, 1988).
4. Results
Based on the analysis of zoarial form distributionand complemented by data on extant species, themain results of this study are presented in Table 3and Figs. 3–7. The sedimentary succession observedin the five sections has been subdivided into fourparts. Each part is characterised by an assemblageof zoarial forms in various proportions (Table 3) andalso by the corresponding species and abundance offragments (Table 4).
The distribution of each species varies stronglybetween samples and only a small number of speciesis restricted to a single assemblage. Two species arethus restricted to assemblage 1 (Tessaradoma bore-ale and Gemellipora eburnea), none to assemblage2, seven to assemblage 3, and four to assemblage 4.Most species occur in two or three (adjacent) assem-blages. Only three species are found in all assem-blages (Cellaria salicornioides, Crisia eburnea andScrupocellaria elliptica). These three belong to the
cellariiform morphotype and their ubiquitous distri-bution on the continental platform may be controlledmainly by taphonomic processes.
At the base of the sections, one conescharellini-form species largely predominates and is associ-ated with very few fragments of erect colonies(mostly cellariiform). This first part is succeeded byan assemblage containing rarer conescharelliniformcolonies, numerous cellariiform, less numerous erectrigid morphotypes, and very few membraniporiformsand celleporiforms. The third assemblage containsno conescharelliniform, but numerous erect zoarialforms, celleporiforms and membraniporiforms. Inthe fourth assemblage membraniporiform, cellepori-form and cellariiform species predominate, some-times associated with erect rigid colonies.
This general pattern for the Messinian of north-west Algeria shows some variations between the fivesections studied in the region (Figs. 3–7).
4.1. Arbal section
A 30 m thick alternation of diatomites and marlsis overlain by about 20 m of clayey limestones(Fig. 3). All ten samples yielded bryozoans. A totalof twenty species has been identified, none beingvery abundant. The succession of zoarial form as-semblages follows the general pattern with the lastsample yielding only membraniporiform and celle-poriform colony fragments (but the complete ab-sence of erect colonies may possibly be due to aninsufficient amount of material being sampled andobserved in these indurated clayey limestones).
4.2. Chabat el Aricha section
A 100 m thick section has been logged and sam-pled. It shows an alternation of about 60 m of marlor clayey limestone and diatomite beds overlain by40 m of limestones (Fig. 4). No sample was takenfrom the top of the section, formed by hard recrys-tallised limestones. Among the 22 samples, 3 con-tained no bryozoans. The general pattern is patchierthan in the other sections and the bryozoan faunasshow relatively low abundance and diversity, withonly 30 identified species.

312P.M
oissette/Palaeogeography,
Palaeoclimatology,
Palaeoecology155
(2000)305–326
Table 3Distribution of the bryozoan species and their zoarial forms in the five sections under study
Species Sections
Arbal El Haricha Bou Seter Semmoumet Sig
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
MembraniporiformAlderina sp. žAnnectocyma major ž ž ž ž ž ž ž žAplousina bobiesi ž ž ž ž ž ž ž ž ž ž ž ž ž žBiflustra savarti ž ž ž ž ž ž žBuffonellaria divergens ž ž ž ž ž ž ž žCallopora fenestrata žCalloporina decorata ž žCalpensia gracilis žCalpensia nobilis ž ž žCheiloporina campanulata ? žChorizopora brongniarti ž ž ž ž ž žCribrilaria innominata ž ž ž ž ž ž ž žCryptosula pallasiana ž žDesmeplagioecia sp. ž ž žDiplosolen obelium žEscharella variolosa žEscharina dutertrei žEscharina vulgaris ž ž žEscharoides coccinea ž ž ž žEscharoides fulgurans žFigularia figularis ž ž ž žHaplopoma bimucronatum žHincksina flustroides ž ž ž žHincksina loxopora ž ž žHippopleurifera sp. žHippopodinella lata žLichenopora hispida ž ž žLichenopora prolifera žMicropora coriacea ž ž ž ž žMicroporella ciliata ž ž ž ž žOnychocella angulosa ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž žPentapora pertusa ž ž ž ž ž ž ž ž ž žPlagioecia patina žPlagioecia sarniensis ž žSchismoporella aculifera ? žSchizobrachiella sanguinea ž ž žSchizomavella auriculata ž ž ž žSchizomavella linearis ž ž ž

P.Moissette
/Palaeogeography,Palaeoclim
atology,Palaeoecology
155(2000)
305–326313
Table 3 (continued)
Species Sections
Arbal El Haricha Bou Seter Semmoumet Sig
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
Schizomavella rudis žSchizoporella longirostris ž ž ž ž ž ž ž žSchizotheca fissa ž ž ž ž ž žSmittina aff. marionensis žSmittina sp. žStomatopora granulata ž žStomatopora parvula ž žTeuchopora castrocarensis ž žTubulipora plumosa žUmbonula ovicellata žWatersipora (?) sp. žCelleporiformCelleporaria palmata ž ž ž ž ž žCellepora pumicosa ž žCelleporina costazi ž ž ž ž ž ž ž ž ž ž ž ž žTurbicellepora coronopus ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž žTurbicellepora cf. coarcta ž ž ž žIndeterminate celleporiform ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž žAdeoniformAdeonella polystomella ž ž ž ž ž žAdeonellopsis maghrebensis žMesenteripora meandrina ž ž ž ž ž ž žMetrarabdotos canui ž ž ž ž žOnychocella angulosa demarcqi žPorella cervicornis ž ž ž ž ž ž ž ž ž ž ž žSchizostomella (?) dubia ž ž ž ž žSchizotheca serratimargo ? ž ž žVinculariiformEntalophoroecia proboscidea ž ž ž ž ž ž ž ž ž ž ž žEntalophoroecia subverticellata ž ž ž ž ž ž ž ž ž ž žExidmonea atlantica ž ž ž ž ž ž ž ž ž ž ž ž žFrondipora verrucosa ž žHornera frondiculata ž žIdmidronea coronopus žPhoceana pliocenica ž ž ž ž ž žSmittina canavarii žTervia irregularis ž ž ž ž ž ž ž ž ž

314P.M
oissette/Palaeogeography,
Palaeoclimatology,
Palaeoecology155
(2000)305–326
Table 3 (continued)
Species Sections
Arbal El Haricha Bou Seter Semmoumet Sig
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
Tessaradoma boreale ž žYbselosoecia typica ž ž ž ž ž ž ž ž ž ž žReteporiformSertella beaniana žSertella cellulosa ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž žSchizoretepora aff. fungosa ž ž žCellariiformCellaria salicornioides ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž žCrisia eburnea ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž žCrisia hoernesi ž ž ž ž žGemellipora eburnea ž ž ž žMargaretta cereoides ž žNellia oculata ž žScrupocellaria elliptica ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž žCatenicelliformScruparia ambigua žLunulitiformAnoteropora persimplex žDiscoporella reussiana žConescharelliniformBatopora rosula ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž ž žFor each of the four assemblages, the number of black dots is proportional to the number of bryozoan fragments (resulting from an average of all samples in thecorresponding section and assemblage).

P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326 315
Fig. 3. Sedimentary log and zoarial form distribution in the Arbal section. Numbers indicate sampling levels.
4.3. Chabat Bou Seter section
The 40 m thick diatomite=marl alternation is over-lain by about 50 m of mostly algal and coral lime-stones (Fig. 5). From a total of 19 samples, only onewas taken from these hardened limestones (it con-tains the catenicelliform species not represented inthe other sections). Sample 5 is devoid of bryozoans,but the abundance and diversity of the bryozoansin the marl and clayey limestone levels is generallyhigh, with a total of 54 species.
4.4. Hammar Semmoumet section
Almost 100 m of clayey limestones, marls anddiatomites are overlain by sandstones at the top ofwhich appears a limestone bed formed by hermatypiccorals and coralline algae (Fig. 6). Although 10 sam-ples out of 44 — especially in the sandy levels —yielded no bryozoans, a total of 56 species have beenidentified in this section and the diversity is generallyhigh in samples 36 to 40. One lunulitiform species(Discoporella reussiana) is poorly represented in thesecond assemblage (level 21).

316 P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326
Fig. 4. Sedimentary log and zoarial form distribution in the Chabat el Aricha section. Numbers indicate sampling levels (samples markedwith an asterisk yielded no bryozoan).

P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326 317
Fig. 5. Sedimentary log and zoarial form distribution in the Chabat Bou Seter section. Numbers indicate sampling levels (sample 5yielded no bryozoan).

318 P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326

P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326 319
4.5. Sig section
A 100 m thick sedimentary succession that showsa well developed alternation of marl and diatomitebeds followed by sand levels containing only oys-ters (Fig. 7). All 26 samples yielded bryozoans. Thediversity is high (60 species), with some speciesabundant in a number of samples (notably betweenlevel 13 and 23). Another lunulitiform species (An-oteropora persimplex) is represented by some frag-ments at the base of the section. The main differencewith the general pattern is the somewhat highernumber of erect species (11) found in the first as-semblage. Compared to other sections, there is alsoan exceptionally large amount of conescharellini-form colonies in levels 10–11 which explains theprobably exaggerated average number of coloniesbelonging to this zoarial form in assemblage 2(Table 4). An additional difference is the absenceof membraniporiform colony remains in the lasttwo samples where erect (mostly cellariiform) mor-photypes predominate, together with celleporiforms.This is interpreted as resulting from an accumu-lation — probably in very shallow waters — ofthe disarticulated segments of cellariiform colonies,easily transported over long distances (Poluzzi etal., 1991, 1997). However, as noted by Lagaaij andGautier (1965), the cellariiform zoarial type is alsothe least sensitive to high sedimentation rates andits occurrence in great proportions in these levelscould possibly be linked to an increase in terrigenoussupply, accentuated in the overlying sandy sedi-ments (of fluviomarine origin?), completely barrenof bryozoans (this was also probably the case forsamples 41–43 of the Hammar Semmoumet sec-tion).
5. Palaeobathymetric reconstruction
In all five sections, four distinct depth-related as-semblages have been recognised. They are well char-acterised by different proportions of zoarial forms,numbers of species and of fragments (Table 4 and
Fig. 6. Sedimentary log and zoarial form distribution in the Hammar Semmoumet section. Numbers indicate sampling levels (samplesmarked with an asterisk yielded no bryozoan).
Fig. 8) and presence of several extant species corre-sponding to various depth intervals (Table 2).
The first assemblage is the deepest (Fig. 8). Itsmain feature is the almost exclusive presence ofthe fossil conescharelliniform Batopora rosula, ac-companied by rare cellariiforms and very rare vin-culariiforms (fragments of adeoniform and lunuliti-form colonies are here extremely scarce). The aver-age number of species and of fragments (or coloniesfor B. rosula) is here very small (2 species=sampleand 14 fragments=sample). The occurrence of a num-ber of erect deep stenobathic species (Gemelliporaeburnea, Tervia irregularis, Tessaradoma boreale)also suggests a minimum depth of 75 m, more prob-ably around 200 m or even more (for Tessaradomaboreale today the shallowest occurrences are in factfrom the Arctic). In terms of the bathymetric zonationused for the Mediterranean (Peres and Picard, 1964;Peres, 1967) this assemblage corresponds to the up-per bathyal and the lower circalittoral (D slope riseto outer shelf). A peak abundance of conescharellini-form colonies (level 2 at Arbal, level 3 at Chabat BouSeter, level 14 at Hammar Semmoumet) inside thisoligospecific assemblage is interpreted as indicatingthe greatest depth and thereby the maximum of theMessinian transgression (Moissette, 1996).
The second assemblage is more diverse (Fig. 8).The conescharelliniform species Batopora rosula isstill represented, but with fewer colonies (althoughthere is an exceptionally high number of coloniesof B. rosula in two levels of the Sig section), ac-companied by fairly abundant cellariiform segments(mostly belonging to Cellaria salicornioides) and asmall number of erect rigid colony fragments (withvery rare membraniporiforms, celleporiforms andlunulitiforms). This assemblage is moderately rich,both in systematic diversity (averaging 6 species persample) and number of fragments (36 per sample).Deep-water species such as Gemellipora eburneaand Tessaradoma boreale are absent, but Tervia ir-regularis is still present, indicating a minimum depthof about 60 m. A maximum depth of around 100to 120 m is suggested by the occurrence of specieslike Adeonella polystomella, Porella cervicornis or

320 P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326
Fig. 7. Sedimentary log and zoarial form distribution in the Sig section. Numbers indicate sampling levels.

P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326 321
Table 4Number of species and number of fragments in the four bryozoan assemblages
Assemblage 1 Assemblage 2 Assemblage 3 Assemblage 4
SpeciesMembraniporiform 0 7 44 23Celleporiform 0 3 4 4Adeoniform 3 3 6 2Vinculariiform 6 6 9 4Reteporifom 0 1 3 1Cellariiform 5 4 5 4Catenicelliform 0 0 0 1Conescharelliniform 1 1 0 0Lunulitiform 1 1 0 0Total number 16 26 71 39Average number=section 7 10 37 17Average number=sample 2 6 17 10
Number of fragments=sampleAverage 14 36 355 77Membraniporiform 0 1* 43 15Celleporiform 0 1* 40 16Adeoniform 1* 11 26 4Vinculariiform 1 2 63 2Reteporifom 0 2 23 0Cellariiform 3 30 166 26Catenicelliform 0 0 0 1*Conescharelliniform 9 7 0 0Lunulitiform 1* 1* 0 0
Asterisks indicate that the corresponding average numbers were less than one unit.
Schizotheca fissa. This assemblage was probablydeposited in the lower circalittoral zone (D outershelf).
Batopora rosula is absent in the third assemblagebut there is a strong increase in the number of mem-braniporiform and celleporiform colony fragmentsin all levels of the five sections (Fig. 8). They areaccompanied by numerous to very numerous debrisof adeoniforms, vinculariiforms, reteporiforms, andcellariiforms. The specific richness of this assem-blage is high (on average 17 species=sample) andso is the abundance (355 fragments=sample). Severalextant shallow-water species occur in this assem-blage: Calpensia nobilis, Haplopoma bimucronatum,Hincksina flustroides, Margaretta cereoides, Ony-chocella angulosa, Tubulipora plumosa, Umbonulaovicellata. Moreover, some species are particularlyabundant in a number of samples belonging to thisassemblage (Cellaria salicornioides, Exidmonea at-lantica, Onychocella angulosa, Porella cervicornis,Turbicellepora coronopus) and, from their known
optimum bathymetric ranges, depths of around 50to 80 m are suggested. This is corroborated by theobservations made by several authors in very dif-ferent settings that bryozoan abundance peaks at ca.40–90 m (Cuffey and Turner, 1987) or 50–100 m(Caulet, 1972; Theroux and Wigley, 1998). Thisrich assemblage indicates another decrease in waterdepth and characterises the upper circalittoral zone(D mid-shelf).
The fourth assemblage contains relatively numer-ous membraniporiform and celleporiform colonies,associated with rare adeoniform and vinculariiform,but abundant cellariiform fragments (Fig. 8). Itshows a decrease in the number of species (on av-erage 10 per sample) and in the average abundanceof bryozoan fragments (77 per sample) compared tothe previous assemblage. A maximum depth of lessthan about 30 to 50 m is suggested by the presenceof shallow to very shallow species such as Calpensianobilis, Cryptosula pallasiana, Hincksina flustroides,Hippopodinella lata, and Scruparia ambigua. This

322 P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326
Fig. 8. Distribution of the various zoarial form assemblages (in average number of fragments per sample) on the continental platform ofthe region under study (based on data from Table 4). Circled digits indicate the average number of species per section. Catenicelliformand lunulitiform colonies were always negligible and have been omitted; the same was done whenever the average number ofcelleporiform or membraniporiform colony fragments was less than one unit.
assemblage is the shallowest, characterising the in-fralittoral zone (D inner shelf to shoreface).
The overall pattern of the succession of zoar-ial form assemblages is thus strongly indicative ofan upward progressive reduction in palaeodepth,which is supported by the presence of a numberof extant stenobathic bryozoan species. Confirma-tory evidence is given by the observed sedimen-tary successions (marls–marls and diatomites–clayeylimestones–limestones and sandstones) and for theSig section by the analysis of benthic foraminifers(Poignant and Moissette, 1992) and diatom assem-blages (Mansour et al., 1995). Similar trends have alsobeen observed in the Messinian of other regions of theMediterranean (Gersonde, 1980; Rouchy, 1982).
6. Discussion
The bryozoan faunas of the five sections understudy and other coeval sections of the same region(Moissette, 1988, 1993) are all relatively diverse andindicate a marine environment with normal salinity.Although most extant species are eurythermal andlive now in the Mediterranean and the temperateAtlantic, the presence of a number of tropical tosub-tropical bryozoan taxa (here Biflustra savarti,Nellia oculata, Metrarabdotos; in other sections Ste-ginoporella, Tremogasterina, Tremopora radicifera)is indicative of warmer waters than in the modernMediterranean. This is corroborated by the great de-velopment of coral reefs in the Chelif Basin as inother Messinian basins of the Mediterranean and theRifian corridor (Rouchy, 1982; Saint Martin, 1990).

P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326 323
However, the influence of upwelling currents on thesedimentation and on the invertebrate faunas of thisregion has also been observed (Rouchy, 1982; SaintMartin, 1990; Moissette and Saint Martin, 1992;Mansour et al., 1995).
This study clearly shows that bryozoan mor-photypes occupy different bathymetric zones, eventhough their depth ranges are largely overlapping.The known bathymetric distribution of some of thezoarial forms has also been validated here. The en-crusting morphotypes (membraniporiform and celle-poriform) are absent to very rare in deep habitats(Gordon, 1987; Moissette and Spjeldnaes, 1995;Rosso and Di Geronimo, 1998), but cellepori-forms live somewhat deeper than membranipori-forms (Caulet, 1972; Spjeldnaes and Moissette,1997). Among erect rigid colonies the vinculari-iform morphotype occurs at greater depths thanadeoniforms, but is also more abundant at inter-mediate depths, whereas reteporiforms are absent atgreater depths and apparently excluded from veryshallow environments (Schopf, 1969; Smith, 1995).Although found deeper, the cellariiform morphotypeis abundant at moderate depths but can also predom-inate in shallow waters (Lagaaij and Gautier, 1965;Schopf, 1969).
A detailed comparison of bryozoan assemblages(species and=or zoarial forms) between sections withotherwise similar lithofacies may also help in corre-lating different ecostratigraphical units. This is no-tably the case in this study for the maximum ofabundance of Batopora rosula interpreted as themaximum depth of deposition and as the maximumof the Messinian transgression (Moissette, 1996).
As previously mentioned, there are, however,some differences in depth ranges of species betweenthe Mediterranean and the Atlantic. As for the depthdistributions of the zoarial forms, there are appar-ently also more or less important differences betweenthe Mediterranean=Atlantic and other regions such asthe swell-dominated cool-water continental shelvesof Australia (James et al., 1992; Bone and James,1993; Boreen et al., 1993) and New Zealand (Nelsonet al., 1988). This is especially true for the cateni-celliform morphotype which is the most widespreadand abundant bryozoan form on the southern Aus-tralian shelf and is found there from shallow waterto over 400 m and for the celleporiform morphotype
which is considered as never occurring at depths lessthan 90 m (Bone and James, 1993), or as abundantonly at depths of 200–300 m or more (Nelson et al.,1988). A complicating factor is, however, the occur-rence of relict skeletal deposits generally dominatingin deeper-water environments (Nelson et al., 1988;James et al., 1992). The presence of relicts is a resultof the continual reworking of uncemented fragmentson such a high-energy seafloor, where storm wavebase is �120 m or deeper. The uncemented nature isdue to the primary mineralogy being predominantlylow-Mg calcite to intermediate-Mg calcite. This iseasily seen in the modern, but difficult to distinguishin the Tertiary. Nevertheless, Nelson and Hancock(1984) also found that, for example, the peak abun-dance of modern celleporiform colonies was 20–50m in New Zealand. Caution must be exercised sincea number of assumptions made in this paper may notbe applicable for areas outside the Mediterranean.
Another potential problem is preservation bias.As noted by Smith and Nelson (1994) and by Smith(1995), taphonomic filters always operate betweena living community and the resulting fossil assem-blage. Nearly all bryozoans in this study are rep-resented by fragments of colonies and even thoughmost of them are fairly well preserved, some in-formation has probably been lost through variousseafloor and diagenetic processes. This is especiallythe case for the fragile colonies of membranipori-form B and catenicelliform morphotypes. Arago-nite skeletons are often dissolved rapidly before andduring burial (Smith and Nelson, 1994); this phe-nomenon may explain the almost complete lack oflunulitiform colonies (although edaphic factors prob-ably dominate here) and the very poor preservationof some erect bryozoans like the adeoniform speciescalled for convenience Schizostomella (?) dubia.
Transport and mixing of faunas from adjacentenvironments may also occur. This will distort theoriginal environmental signals (Smith, 1995). In allsections studied this phenomenon is most probablynot important (except perhaps for some cellariiformspecies) in the mostly marly sediments correspond-ing to low or moderate energy levels but may bemore important in the limestones of the upper parts.For most shelf environments the transport of bio-genic sediment is considered of relatively minorimportance and may even be considered as negligi-

324 P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326
ble in some cases (Lagaaij and Gautier, 1965; Caulet,1972; James et al., 1992). The accumulation of fossilremains near their habitats over some period of timemay even sometimes result in the integration of moreor less patchy and fluctuating living communitiesinto enriched and averaged death assemblages.
7. Conclusion
The results of the analysis of bryozoan assem-blages — relative abundances of the various zoarialforms and presence of a number of extant steno-bathic species — in the Messinian of northwestAlgeria demonstrate that much information can begained for palaeobathymetric reconstructions in Ter-tiary sedimentary successions by using a combina-tion of these two methods. They are of special valuein determining changes in water depth through avertical sequence where variations in assemblagesare more easily assessed than in isolated samples.Moreover, the zoarial form method is still usable byspecialists even when specimens are so badly pre-served that species cannot be identified. The same istrue for the identification of bryozoan colonial mor-photypes in thin sections of indurated limestones bynon-specialists, although supporting evidence fromcleaned macroscopic material may sometimes beneeded (Nelson et al., 1988).
The use of these methods for older levels ofthe Cenozoic and of course the Mesozoic and thePalaeozoic is more difficult since no extant speciesare present (although the use of extant stenobathicgenera and even families may still be possible).Moreover, changes with time in bathymetric distri-bution of zoarial forms could possibly have occurred(Kelly and Horowitz, 1987; Smith, 1995), just as areversal in the diversity of encrusting versus erectspecies between the Palaeozoic and the post-Palaeo-zoic was noted by McKinney and Jackson (1989).
In summary, an increase in knowledge of the mod-ern bathymetric distribution of both zoarial formsand species is still very much needed before moredetailed palaeobathymetric reconstructions can bepossible.
Acknowledgements
I am grateful to Arlette Armand (University ofLyon) for her help in the preparation of Figs. 1–7 andTable 1. I also wish to thank Y. Bone (University ofAdelaide) and P.D. Taylor (Natural History Museum,London) who reviewed the manuscript of this paperand contributed to its improvement.
References
Anderson, R.V.V., 1933. The diatomaceous and fish-bearingBeida stage of Algeria. J. Geol. 41 (7), 673–698.
Askren, L.T., 1968. Bryozoan paleoecology from the Tertiary ofAlabama. Southeast. Geol. 9 (3), 157–163.
Bianchi, M., Braga, G., Finotti, F., Manfredi, M., 1988. Ruolo deibriozoi nelle interpretazioni paleoambientali. Atti del QuartoSimposio di Ecologia e Paleoecologia delle Comunita Ben-toniche, Sorrente, pp. 21–33.
Bone, Y., James, N.P., 1993. Bryozoans as carbonate sedimentproducers on the cool-water Lacepede Shelf, southern Aus-tralia. Sediment. Geol. 86, 247–271.
Boreen, T., James, N., 1995. Stratigraphic sedimentology ofTertiary cool-water limestones, SE Australia. J. Sediment. Res.B65, 142–159.
Boreen, T., James, N., Wilson, C., Heggie, D., 1993. Surficialcool-water carbonate sediments on the Otway continental mar-gin, southeastern Australia. Mar. Geol. 112, 35–56.
Braga, G., 1979. Tentativo di interpretazione del paleoambientein una serie oligo–miocenica di Follina (Trevigniano orientale)per mezzo delle forme zoariali dei briozoi. Stud. Trentini Sci.Nat. 56, 125–138.
Caulet, M.J., 1972. Les sediments organogenes du precontinentalgerien. Mem. Mus. Natl. Hist. Nat., Paris, 25, 289 pp.
Cheetham, A.H., 1971. Functional morphology and biofaciesdistribution of cheilostome Bryozoa in the Danian stage (Pa-leocene) of southern Scandinavia. Smithsonian Contrib. Paleo-biol. 6, 1–87.
Cook, P.L., 1981. The potential of minute bryozoan colonies inthe analysis of deep sea sediments. Cah. Biol. Mar. 22, 89–106.
Cook, P.L., Chimonides, P.J., 1983. A short history of the lunuliteBryozoa. Bull. Mar. Sci. 33, 566–581.
Cook, P.L., Lagaaij, R., 1976. Some Tertiary and Recentconescharelliniform Bryozoa. Bull. Br. Mus. (Nat. Hist.) Zool.29, 317–376.
Cuffey, R.J., Turner, R.F., 1987. Modern bryozoans on the Ko-diak Shelf, southern Alaska. In: Ross, J.R.P. (Ed.), Bryozoa:Present and Past. Western Washington University, Bellingham,Washington, pp. 65–72.
Gautier, Y.V., 1962. Recherches ecologiques sur les BryozoairesCheilostomes en Mediterranee occidentale. Rec. Trav. Sta.Mar. Endoume, 24 (38), 434 pp.
Gersonde, R., 1980. Palaookologische und biostratigraphische

P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326 325
Auswertung von Diatomeenassoziationen aus dem Messiniumdes Caltanisetta-Beckens (Sizilien) und einiger Vergleichspro-file in SO-Spanien, NW-Algerien und auf Kreta. UnpublishedThesis, University of Kiel, 393 pp.
Gordon, D.P., 1987. The deep-sea Bryozoa of the New Zealandregion. In: Ross, J.R.P. (Ed.), Bryozoa: Present and Past.Western Washington University, Bellingham, Washington, pp.97–104.
Hageman, S.J., Bone, Y., McGowran, B., James, N.P., 1997. Bry-ozoan colonial growth-forms as paleoenvironmental indicators:evaluation of methodology. Palaios 12, 405–419.
Hageman, S.J., Bock, P.E., Bone, Y., McGowran, B., 1998. Bry-ozoan growth habits: classification and analysis. J. Paleontol.72 (3), 418–436.
Harmelin, J.-G., 1969a. Bryozoaires recoltes au cours de lacampagne du Jean Charcot en Mediterranee orientale (aout–septembre 1967), I. Dragages. Bull. Mus. Natl. Hist. Nat.,Paris 40 (6), 1179–1208.
Harmelin, J.-G., 1969b. Bryozoaires recoltes au cours de lacampagne du Jean Charcot en Mediterranee orientale (aout–septembre 1967), I. Dragages (suite). Bull. Mus. Natl. Hist.Nat., Paris 41 (1), 295–311.
Harmelin, J.-G., 1969c. Bryozoaires des grottes sous-marinesobscures de la region marseillaise. Faunistique et ecologie.Tethys 1 (3), 793–806.
Harmelin, J.-G., 1976. Le sous-ordre des Tubuliporina (Bry-ozoaires Cyclostomes) en Mediterranee. Mem. InstitutOceanogr. Monaco 10, 326 pp.
Harmelin, J.-G., 1985. Bryozoan dominated assemblages inMediterranean cryptic environments. In: Nielsen, C., Larwood,G.P. (Eds.), Bryozoan: Ordovician to Recent. Olsen and Olsen,Fredensborg, pp. 135–143.
Harmelin, J.-G., 1988. Les Bryozoaires, de bons indicateursbathymetriques en paleoecologie? Geol. Mediterr. 15 (1), 49–63.
Harmelin, J.-G., 1997. Diversity of bryozoans in a Mediterraneansublittoral cave with bathyal-like conditions: role of dispersalprocesses and local factors. Mar. Ecol. Progr. Ser. 153, 139–152.
Harmelin, J.-G., d’Hondt, J.-L., 1993. Transfers of bryozoanspecies between the Atlantic Ocean and the MediterraneanSea via the Strait of Gibraltar. Oceanol. Acta 16 (1), 63–72.
Hayward, P.J., 1974. Studies on the cheilostome bryozoan faunaof the Aegean island of Chios. J. Nat. Hist. 8, 369–402.
Hayward, P.J., Ryland, J.S., 1979. British Ascophoran Bryo-zoans. Academic Press, London, 312 pp.
James, N.P., Bone, Y., 1991. Origin of a cool-water, Oligo–Miocene deep shelf limestone, Eucla Platform, southern Aus-tralia. Sedimentology 38, 323–341.
James, N.P., Bone, Y., Von der Borch, C.C., Gostin, V.A., 1992.Modern carbonate and terrigenous clastic sediments on a coolwater, high energy, mid-latitude shelf: Laceped, southern Aus-tralia. Sedimentology 39, 877–903.
Kelly, S.M., Horowitz, A.S., 1987. Growth forms and paleoecol-ogy of Mississippian bryozoans: Critical application of Stach’s1936 model, eastern United States. In: Ross, J.R.P. (Ed.),
Bryozoa: Present and Past. Western Washington University,Bellingham, Washington, pp. 137–144.
Labracherie, M., 1970. Valeur de l’abondance et de la diversitedes Bryozoaires dans l’interpretation des sediments fossiles.Bull. Soc. Geol. Fr. 12, 134–136.
Labracherie, M., 1973. Functional morphology and habitat ofBryozoa in the Eocene of the northern Aquitaine Basin,France. In: Larwood, G.P. (Ed.), Living and Fossil Bryozoa,Recent Advances in Research. Academic Press, London, pp.129–138.
Labracherie, M., Prud’homme, J., 1966. Essai d’interpretation depaleomilieux grace a la methode de distribution des formeszoariales chez les Bryozoaires. Bull. Soc. Geol. Fr. 8, 102–106.
Lagaaij, R., Gautier, Y.V., 1965. Bryozoan assemblages from ma-rine sediments of the Rhone delta, France. Micropaleontology11 (1), 39–58.
Mansour, B., Moissette, P., Noel, D., Rouchy, J.M., 1995.L’enregistrement par les assemblages de diatomees des envi-ronnements messiniens: l’exemple de la coupe de Sig (Bassindu Chelif–Algerie). Geobios 28 (3), 261–279.
McKinney, F.K., Jackson, J.B.C., 1989. Bryozoan Evolution.Special Topics in Paleontology, Hyman, Boston, 238 pp.
Moissette, P., 1988. Faunes de bryozoaires du Messiniend’Algerie occidentale. Doc. Lab. Geol. Fac. Sci. Lyon 102,351 pp.
Moissette, P., 1993. Bryozoan assemblages in Messinian depositsof western Algeria. Lethaia 26, 247–259.
Moissette, P., 1996. The Cheilostomate bryozoan Batoporarosula (Reuss, 1848): a paleobathymetric indicator in theMediterranean Neogene. In: Gordon, D.P., Smith, A.M.,Grant-Mackie, J.A. (Eds.), Bryozoans in Space and Time.New Zealand National Institute of Water and AtmosphericResearch Ltd, pp. 193–198.
Moissette, P., 1997. Bryozoaires recoltes dans les unites messini-ennes de sondages offshore dans le golfe de Gabes (Tunisie).Rev. Micropaleontol. 40 (2), 181–203.
Moissette, P., 1999. The use of Neogene bryozoans for a betterunderstanding of the ecology of some Recent species. Proc.11th Int. Conf. Bryozoology (in press).
Moissette, P., Saint Martin, J.P., 1992. Upwelling and benthiccommunities in the Messinian of the western Mediterranean.Paleontol. Evoluc. 24–25, 245–254.
Moissette, P., Saint Martin, J.P., 1995. Bryozoaires des milieuxrecifaux miocenes du sillon sud-rifain au Maroc. Lethaia 28,271–283.
Moissette, P., Spjeldnaes, N., 1995. Plio–Pleistocene deep-waterbryozoans from Rhodes, Greece. Palaeontology 38, 771–799.
Nelson, C.S., Hancock, G.E., 1984. Composition and origin oftemperate skeletal carbonate sediments on South Maria Ridge,northern New Zealand. N.Z. J. Mar. Freshwater Res. 18, 221–239.
Nelson, C.S., Hyden, F.M., Keane, S.L., Leask, W.L., Gordon,D.P., 1988. Application of bryozoan zoarial growth-form stud-ies in facies analysis of non-tropical carbonate deposits inNew Zealand. Sediment. Geol. 60, 301–322.

326 P. Moissette / Palaeogeography, Palaeoclimatology, Palaeoecology 155 (2000) 305–326
Peres, J.M., 1967. Les biocoenoses benthiques dans le systemephytal. Rec. Trav. Sta. Mar. Endoume 58, 3–113.
Peres, J.M., Picard, J., 1964. Nouveau manuel de bionomie ben-thique de la Mer Mediterranee. Rec. Trav. Sta. Mar. Endoume31, 1–137.
Perrodon, A., 1957. Etude geologique des bassins neogeneslittoraux de l’Algerie occidentale. Bull. Serv. Carte Geol.,Algerie, 12, 323 pp.
Poignant, A., Moissette, P., 1992. Foraminiferes benthiquesdes intercalations marneuses d’une serie diatomitique duMessinien d’Algerie. Neues Jahrb. Geol. Palaontol. Monatsh.11, 657–674.
Poluzzi, A., Rosso, A., 1988. I tipi zoariali di un’associazionea Briozoi del Mar Ionio. Acta Naturalia Ateneo Parmense 24,83–108.
Poluzzi, A., Masini, F., Capozzi, R., 1991. I Briozoi eretti-artico-lati nello Spungone della Romagna (Pliocene inferiore, Nord-Italia). Boll. Soc. Paleontol. Ital. 30 (3), 329–347.
Poluzzi, A., Ligi, M., Badalini, M., 1997. Bryozoan transport inhigh-energy environments (Strait of Messina, Sicily). G. Geol.59 (1–2), 55–79.
Prenant, M., Bobin, G., 1966. Faune de France 68 — Bry-ozoaires, 2eme partie: Chilostomes Bryozoa. Lechevallier,Paris, 647 pp.
Reguant, S., Vazquez, A., Zamarreno, I., Maluquer, P., 1985.Significacion de los briozoos en los sedimentos superficialesde la plataforma continental del Cabo de Gata (Almerıa,Espana). Acta Geol. Hisp. 20 (1), 69–80.
Reguant, S., Rodriguez-Fernandez, J., Fernandez, J., 1987. Mor-fologıa de las colonias de Briozoos en relacion con la hidrod-inamica del medio sedimentario. Acta Geol. Hisp. 21–22,541–547.
Rosso, A., 1996. Popolamenti e tanatocenosi a Briozoi di fondimobili circalitorali del Golfo di Noto (Sicilia, Italia). Natural-ista Siciliana 4 (20), 189–225.
Rosso, A., Di Geronimo, I., 1998. Deep-sea Pleistocene Bryozoaof Southern Italy. Geobios 31 (3), 303–317.
Rouchy, J.-M., 1982. La genese des evaporites messiniennes deMediterranee. Mem. Mus. Natl. Hist. Nat., Paris 50, 267 pp.
Rucker, J.B., 1967. Paleoecological analysis of cheilostome Bry-
ozoa from Venezuela–British Guiana shelf sediments. Bull.Mar. Sci. 17 (4), 787–839.
Ryland, J.S., Hayward, P.J., 1977. British Anascan Bryozoans.Synopses of the British Fauna 10, The Linnean Society ofLondon, 188 pp.
Saint Martin, J.P., 1990. Les formations recifales coralliennesdu Miocene superieur d’Algerie et du Maroc. Aspects paleoe-cologiques et paleogeographiques. Mem. Mus. Natl. Hist. Nat.,Paris 56, 373 pp.
Saint Martin, J.P., Cornee, J.J., Conesa, G., Bessedik, M., Belke-bir, L., Mansour, B., Moissette, P., Anglada, R., 1992. Undispositif particulier de plate-forme carbonatee messinienne:la bordure meridionale du bassin du Bas-Chelif, Algerie. C. R.Acad. Sci., Paris 315 (II), 1365–1372.
Schopf, T.J.M., 1969. Paleoecology of Ectoprocts (bryozoans). J.Paleontol. 43 (2), 234–244.
Smith, A.M., 1995. Palaeoenvironmental interpretation usingbryozoans: a review. In: Bosence, D.W.J., Allison, P.A. (Eds.),Marine Palaeoenvironmental Analysis from Fossils. Geol. Soc.Spec. Publ. 83, 231–243.
Smith, A.M., Nelson, C.S., 1994. Selectivity in sea-floor pro-cesses: taphonomy of bryozoans. In: Hayward, P.J., Ryland,J.S., Taylor, P.D. (Eds.), Biology and Palaeobiology of Bryo-zoans. Olsen and Olsen, Fredensborg, pp. 177–180.
Spjeldnaes, N., Moissette, P., 1997. Celleporid (bryozoan) thick-ets from the upper Pliocene of the island of Rhodes, Greece.Soc. Econ. Paleontol. Mineral. Spec. Publ. 56, 263–270.
Stach, L.W., 1936. Correlation of zoarial form with habitat. J.Geol. 44, 60–66.
Stach, L.W., 1937. The application of the Bryozoa in Caino-zoic stratigraphy. Report of the 23rd Meeting, Australia–NewZealand Association of Advanced Science, 23, 80–83.
Theroux, R.B., Wigley, R.L., 1998. Quantitative composition ofthe macrobenthic invertebrate fauna of the continental shelfecosystems of the northeastern United States. U.S. Departmentof Commerce, Seattle, NOAA Tech. Rep. NMFS 140, 240 pp.
Zabala, M., 1986. Fauna dels Briozous del paısos Catalans.Institut d’Estudis Catalans, Arxius de la Seccio de Ciencies84, Barcelona, 294 pp.