cell proliferation and gill morphology in anoxic crucian carp
TRANSCRIPT

Cell proliferation and gill morphology in anoxic crucian carp
Jørund Sollid1,*, Aina Kjernsli1, Paula M. De Angelis2, Åsmund K. Røhr1 and
Göran E. Nilsson1
1Dept. of Molecular Biosciences, University of Oslo, PO Box 1041, 0316 Oslo, Norway
2Institute of Pathology, Rikshospitalet, Oslo, Norway
*Author of correspondence
Jørund Sollid
Department of Molecular Biosciences
University of Oslo
Blindernveien 31
P.O. Box 1041
0316 Oslo
Norway
Phone: +4722854732
Fax: +4722854664
E-mail: [email protected]
Running head: cell proliferation gill morphology anoxic crucian. carp
Keywords: PCNA, BrdU, ribonucleotide reductase, RNR, intestine, liver
Abbreviations: BrdU - 5’-bromo-2’-deoxyuridine, PCNA - proliferating cell nuclear
antigen, RNR - , ribonucleotide reductase, ILCM - interlamellar cell mass
Articles in PresS. Am J Physiol Regul Integr Comp Physiol (May 26, 2005). doi:10.1152/ajpregu.00267.2005
Copyright © 2005 by the American Physiological Society.

Abstract
Is DNA replication/ cell proliferation in vertebrates possible during anoxia? The
oxygen dependence of ribonucleotide reductase (RNR) could lead to a stop in DNA
synthesis, thereby making anoxic DNA replication impossible. We have studied this
question in an anoxia tolerant vertebrate, the crucian carp (Carassius carassius), by
examining 5’-bromo-2’-deoxyuridine (BrdU) incorporation and proliferating cell
nuclear antigen (PCNA) levels in the gills, intestinal crypts and liver. We exposed
crucian carp to 1 and 7 days of anoxia followed by 7 days of reoxygenation. There was a
reduced incidence of S-phase cells (from 12.2% to 5.0%) in gills during anoxia, which
coincided with a concomitant increase of G0-cells. Anoxia also decreased the number of
S-phase cells in intestine (from 8.1% to 1.8%). No change in the fraction of S-phase cells
(~1%) in liver was found. Thus, new S-phase cells after 7 days of anoxia were present in
all tissues revealing a considerable rate of DNA synthesis. Subsequently, the oxygen
dependent subunit of crucian carp RNR (RNRR2) was cloned. We found no differences
in amino acids involved in radical generation and availability of the iron centre
compared to mouse, which could have explained reduced oxygen dependence. Further,
the amount of RNRR2 mRNA in gills did not decrease throughout anoxia exposure.
These results indicate that crucian carp is able to sustain some cell proliferation in
anoxia possibly because RNRR2 retains its tyrosyl radical in anoxia, and that the
replication machinery is still maintained. We have previously observed that hypoxia
triggers a 7.5 fold increase of respiratory surface area in crucian carp. This response
was not triggered in anoxia.

Introduction
Mammals, and most other vertebrates, are intolerant to prolonged oxygen depravation
(8). However, there are a few vertebrates and invertebrates that are anoxia tolerant.
Invertebrates like Caenorhabditis elegans, Drosophila melanogaster and Artemia
franciscana are able to survive anoxia through suspended animation and arrest of cell
division (5, 7, 10, 27). Further, this response to anoxia is also found in some vertebrates
like zebrafish (Danio rerio) embryo and turtles of the genera Trachemys and Chrysemys (6,
28, 36). Crucian carp can survive anoxia for several months at 4°C (13). However, crucian
carp is, unlike the other species, still active in anoxia (21, 22, 35). An active species will
have a need for basic maintenance of organs, which is likely to demand maintained cell
proliferation, and therefore DNA synthesis.
Functional RNR enzymes are needed for DNA synthesis. They convert ribonucleotides to
deoxyribonucleotides through a radical based reaction. The RNR enzymes are divided into
3 main classes based on their utilization of metal cofactors (17). In vertebrates, RNR
consists of 4 subunits (α2β2) where the oxygen dependent radical formation occurs in the
two β subunits (R2). Previous findings on vertebrate RNR has shown radical half-life,
which is essential for catalytic activity, of 30-60 min (4, 24). This suggests that DNA
synthesis will stop in anoxia. Indeed, this has found to be the case in cultured mammalian
cells (11, 29, 30)
In the crucian carp’s habitat in Northern Europe, seasonal severe hypoxia and anoxia are
common during winter. The anoxia is the result of stop in photosynthesis combined with
thick ice coverage that blocks oxygen diffusion. The crucian carp has evolved behavioral,
physiological and molecular adaptations to survive this environment. These include

reduced brain activity and locomotion (20, 23), maintained cardiac activity (35), increased
glycolysis (37) and avoidance of lactate self-pollution through generation and excretion of
ethanol as a major glycolytic end-product (15). Especially the latter is critical since it
enables the fish to survive on anaerobic metabolism without suffering from acidosis, for as
long as there is glycogen present. Nevertheless, survival is ultimately dependent on the
glycogen stores. Hence saving energy, whenever possible, will be important.
We have previously found that crucian carp living in normoxic water have gills lacking
protruding lamellae, the primary site of O2 uptake in fish (34). Such an unusual trait leads
to a very small respiratory surface area. This reduces water and ion fluxes, thereby
minimizing the cost of osmoregulation. However, if crucian carp experiences hypoxia, a
large increase of the respiratory surface area occurs, enhancing oxygen uptake capabilities
(34). The morphological alteration is due to hypoxia induced apoptosis and cell cycle arrest
in a tissue, named interlamellar cell mass (ILCM), which fills the space between the
lamellae in normoxia. The crucian carp is then able to cover its energy needs through
aerobic metabolism at a lower oxygen tension, allowing it to postpone depletion of
precious glycogen.
We have previously found that ILCM in normoxic gills have relatively high rate of cell
proliferation (34). Preliminary investigations have shown that anoxic crucian carp has gills
resembling those seen in normoxia. Cell proliferation could thus still be present in anoxia,
which, on the other hand, appears unlikely due to the oxygen dependence of RNR.
We have here exposed crucian carp up to 7 days of anoxia, and investigated gill
morphology and cell proliferation in gills, intestine and liver by using both BrdU and
staining for PCNA. BrdU is a thymidine analogue incorporated into DNA during DNA

synthesis revealing cells in the S-phase of cell cycle. PCNA is a DNA sliding clamp for
replicative DNA polymerases. Subsequently, we partially cloned and characterized crucian
carp RNRR2 to investigate possible differences of crucian carp RNRR2 compared to
mammalian RNRR2, and measured its expression with real-time PCR.
Materials and methods
Animals
Crucian carp (Carassius carassius L.; weighing 12.5-31.5 g; all adults) were caught June
2003 in Tjernsrud pond, Oslo community. They were kept in tanks (~100 fish/ 500 l)
continuously supplied with aerated and dechlorinated Oslo tap water (10ºC), on a 12:12 h
diurnal regime and fed daily with commercial carp food (Tetra Pond, Tetra, Melle,
Germany).
Exposures
During anoxia exposures, fish were kept in dark sealed tanks continuously supplied with
N2-bubbeled dechlorinated Oslo tap water. The oxygen level was monitored with an
oxygen electrode (Oxi340i, WTW, Weilheim, Germany). The oxygen level was kept < 0.01
mg O2 l-1 (=<0.1% of air saturation), which is the electrode’s detection limit. Control fish
were kept in an identical tank, but supplied with aerated water. The exposure groups, all
consisting of 7 fish, were 7 days normoxia, 1 and 7 days of anoxia and 7 days of anoxia
followed by 7 days of reoxygenation.

Scanning electron microscopy (SEM)
The SEM was done as previously described (34). Briefly, for SEM, gills were fixated in
3% glutaraldehyde in 0.1 M Na-cacodylate buffer before dried and AuPd coated. Gills were
investigated using a JSM 6400 electron microscope (JEOL, Peabody, USA).
Measurement of mitotically active cells with BrdU
BrdU staining was done as previously described (34) with one modification, cross-
sections were treated with boiling citric acid buffer in a microwave for 25 min at 750W
before staining. Briefly, BrdU was injected intraperitonally (100 µg g-1 fish) 24 h before
tissue sampling. Gills were fixated in 4% formaldehyde, paraffin embedded and serially
cross-sectioned before rehydration and citric acid treatment. Primary antibody, mouse IgG
anti-BrdU (Becton Dickinson, Franklin Lakes, USA), was diluted 1:5 in 0.5 % blocking
solution (BS) (0.5% bovine serum albumine, 0.3% Triton X-100 in PBS pH 7.2).
Secondary antibody was a biotinylated goat anti-mouse IgG, diluted 1:20 in 0.5% BS. The
peroxidase conjugate was diluted 1:20 in 0.5% BS and applied before slides where
incubated with diaminobenzidine solution. All chemicals were from Sigma (St. Louis, MI,
USA).
Measurement of mitotically active cells by staining for PCNA
PCNA has been used previously with success in other fish studies (19, 26). The PCNA
staining was done as described for BrdU staining except primary antibody, anti PCNA
(Sigma), was diluted 1:10 in 0.5% BS.

Counting of cells
Stained (BrdU or PCNA) and unstained cells in gills where counted as previously
described (34). In intestine, the number of stained and unstained cells in the lower ¾ of the
crypts where counted. PCNA is present in most phases of cell cycle (G1, S, G2 or the M-
phase), but is not present in the G0-phase. We therefore used lack of PCNA staining to
quantify number of G0-cells in gills and intestine. However, in liver, the low number of
PCNA positive cells made us chose to present data for this tissue as PCNA positive cells.
Partially cloning and characterization of the oxygen dependent subunit of RNR
(RNRR2)
RNA was extracted from normoxic crucian carp gills using TRIzol® reagent (Invitrogen,
Carlsbad, CA, USA), before reverse transcription was performed with poly-dT primers and
SuperscriptTM III RT (Invitrogen).
We did an alignment between known RNRR2 gene sequences in the vertebrate lineage to
find conserved regions using ClustalX v.1.81 (40) and Genedoc v. 2.6.002
(http://www.psc.edu/biomed/genedoc). Gene specific primers (5’ ccatcgctttgtcattttcc and 5’
tctcgaagaagttggtcttgc) were designed from these conserved regions using the free net based
Primer3 program (32).
The gene fragment of RNRR2 was obtained with PCR using the Platinum Taq
Polymerase Kit (Invitrogen). The success of the PCR reaction was examined on a 1% EtBr-
agarose gel, before ligated into pGEM®-T Easy Vector System I (Promega, Madison, WI,
USA). Plasmids were then incorporated into electro-competent E.coli DH5α-strain
(Invitrogen). Positive colonies were screened for correct insert through PCR with plasmid

specific primers (T7 and SP6). Plasmids from 10 positive clones were purified (QIAprep®
Spin Miniprep Kit, VWR International, West Chester, PA, USA) and sequenced.
The 3D-model of the iron "cluster" of RNRR2 was made using the PyMOL Molecular
Graphics System (2002) (DeLano Scientific, San Carlos, CA, USA), and the model was
based on "the structure obtained under oxidizing conditions" (38).
RNA extraction and cDNA synthesis for Real-Time PCR
25 mg of crucian carp gill tissue from each individual was homogenized with TRIzolTM
(Invitrogen). Extracted RNA was quantified with a NanoDrop (NanoDrop Technologies,
Rockland, DE, USA) and the RNA quality was confirmed by a BioAnalyzer (Agilent
Technologies, Palo Alto, CA, USA). The RNA was DNAse I treated (Sigma) before cDNA
synthesis (SuperscriptTM III, Invitrogen) of 2 µg RNA.
Set-up and quantification of real time PCR
A Lightcycler and a LC FastStart DNA MasterPLUS SYBR Green I kit was used (Roche
Diagnostics Corp., Indianapolis, IN, USA). The protocol was that recommended by
supplier. RNRR2 primers (5’ tgaaggagaaccccac and 5’gtaaatcgctccaccaa) were picked from
the consensus sequence using Lightcycler Probe Design Software v. 1.0 (Roche
Diagnostics) and had a final concentration of 0.5 µM. The following PCR amplification
program was applied: 10 min of denaturation at 95°C before a 40 cycles of a three-segment
program were accomplished by: I) 10 s at 95°C for denaturation, II) 10 s at 60°C for
annealing and III) elongation time set to 25bp s-1 at 72°C. Efficiency of each single reaction
was analyzed using the LinRegPCR program (31). The crossing point (Cp) of each reaction

was determined by using the second derivative method (SDM) in the Roche Lighcycler
Software Version 3.5. The latter program was also used to analyzing melting point of
products to assure specificity and purity of the PCR reactions.
Statistics
Statistical significance was tested with one-way ANOVA with Tukey´s post-test where
P<0.05 was considered significant. Data were analyzed by using GraphPad InStat
(GraphPad, San Diego, CA, USA).
Results
Gill morphology
The scanning micrographs revealed that after 7 days in anoxia, crucian carp gills were
indistinguishable from normoxic gills (Fig. 1a-b). Thus, ILCM remained intact during
anoxia.
Cell proliferation in the gills
During anoxia exposure the fraction of S-phase cells stained with BrdU decreased
significantly (P<0.0001) in ILCM, from 12.2±1.3% in normoxia to 5.0±0.9% after 7 days
in anoxia (Fig. 2a). After 7 days of reoxygenation there was a non-significant increase to
6.9±0.5%. Both in normoxia and anoxia, BrdU positive cells were only found in ILCM,
confirming previous findings showing a low mitotic activity in other gill structures (34).
The PCNA staining also indicated a reduced, but continued, mitotic activity. The number
of G0-cells (PCNA negative) increased significantly (P<0.01) from 6.8±1.2% in normoxia

to 11.1±0.5% after 7 days of anoxia (Fig. 2a). There was no significant change in the
number of G0-cells after 7 days of reoxygenation (11.5±0.3%). Like observed for BrdU
positive cells, PCNA positive cells were only found in ILCM (Fig. 3c-d).
Cell proliferation in the intestine
In intestine, the number of S-phase cells decreased significantly (P<0.01) from 8.1±0.7%
in normoxia to 1.8±0.3 after 7 days in anoxia. After reoxygentation the amount of S-phase
cells had increased significantly to 10.2±2.3% (Fig. 2b). BrdU positive cells were only
found in intestinal crypts (Fig. 3e-f). PCNA staining indicated no significant change in
number of G0-cells during anoxia (Fig. 2b).
Cell proliferation in the liver
The amount of BrdU and PCNA positive cells remained constant at approximately 1% in
liver throughout anoxia (Fig. 2c).
RNRR2 characterization and regulation
A sequence alignment between RNRR2 from several species demonstrated a very high
sequence identity (Fig. 4). From the 3D structure of mouse RNRR2 and crucian carp
sequence we could determine that all amino acids involved in coordinating the diiron-
oxygen cluster, transporting electrons and surrounding the tyrosyl radical were conserved
(Fig. 5). Furthermore, amino acids suggested to influence accessibility of the diiron-oxygen
cluster and the tyrosyl radical (16) was also found to be conserved between mouse and
crucian carp (Fig. 4).

The real-time PCR revealed no significant change in the amount of RNRR2 mRNA
between normoxia and 7 days of anoxia (Fig. 6).
Discussion
The results show that anoxia induces a reduction of cell proliferation in ILCM and in
intestinal crypts, two tissues with a relatively high proliferative activity. In liver, with a
more moderate mitotic activity, no changes in the number of S-phase or G0-phase cells
were detected. Most importantly, the results indicate that DNA synthesis continue after 7
days in anoxia. Surprisingly, amino acid residues in RNRR2, especially those surrounding
the iron center, were identical in crucian carp and mouse. There was no change in the
amount of RNRR2 mRNA after anoxia exposure. Finally, the gill morphology did not
change in response to anoxia, in contrast to previous findings in hypoxia.
The reduction in number of S-phase cells mirrored the increase of G0-cells in ILCM (Fig.
2a). In normoxia, the mitotic activity in ILCM was very high, whereas the number of cells
in G0-phase was only 7%. During anoxia exposure the number of S-phase cells fell from
12% to approximately 5%. The drop in S-phase cells is approximately equal as previously
found in hypoxia exposed crucian carp (34), indicating similar proliferative response
situations. Still, ILCM remained intact after 7 days of anoxia, while a previous study
showed 50% reduction after the same time period in hypoxia (34). This indicate that
anoxia, unlike hypoxia, does not induce apoptosis. The maintenance of a large ILCM in
anoxia should help keeping osmoregulatory costs low. Because no oxygen is present, there
would not be any advantage of increasing the respiratory surface area.

Over the recent years it has become clear that PCNA not only functions as a DNA sliding
clamp for replicative DNA polymerases, but also has a marked ability to interact with
multiple partners involved in metabolic pathways, DNA repair and cell cycle regulation
(18). The results in the present paper show that about 90% of the cells in ILCM still had
PCNA present after 7 days in anoxia (Fig. 2a, 3d). This indicates that cell cycle slows
down, but the cells remain prepared to proceed through mitosis when conditions improve.
This conclusion is supported by the concomitant sustained expression of RNRR2 (Fig. 6).
However, the number of S-phase and G0-phase cells had not returned to pre-anoxic levels
after 7 days of reoxygenation. This could indicate that there was no immediate need to
replace ILCM cells after 7 days in anoxia. Indeed, no reduction in the ILCM was apparent
at the end of the anoxic episode (Fig. 1b).
There was a significant fall in the number of S-phase cells in intestine, which returned to
normoxic levels during reoxygentation. This is in contrast to what was observed in gills.
This may indicate a need for new intestinal cells for getting the organ back into a fully
functional state. In the crucian carp’s natural habitat, the increase of water oxygen tension
during spring, after a period of anoxia, is likely to trigger feeding behaviour, since they
then have very low energy reserves (14).
As previously mentioned, the oxygen dependent oxido-reductase activity of RNR is
located on the R2 subunit. Previously, mammalian RNRR2 was only found to be expressed
in the S-phase of cell cycle (1, 9). However, as shown in Fig. 2a and Fig. 6, the significant
reduction of S-phase ILCM cells in anoxia was not correlated with a reduction of RNRR2
mRNA. Thus, not only does RNRR2 appear not to follow the number of S-phase cells, but
its expression is maintained for 7 days in anoxia. This suggests either that a reduction of

cell proliferation in anoxia does not necessarily affect expression of RNRR2 gene, or that
there is, in conjunction with a reduction of cell cycle related RNRR2, an increased
expression of p53 induced R2 (12, 39), believed to be important in DNA damage repair.
However, RNRR2 protein degradation is dependent on transition into mitosis and the level
of RNRR2 protein is not affected by DNA damage or replication block (3).
The partial sequence of crucian carp RNRR2 indicated no important differences in key
amino acids between crucian carp and mouse RNRR2. This seems puzzling as mammalian
RNRR2 is inactivated in anoxia (2, 4). We still found new S-phase cells after 7 days of
anoxia exposure revealing RNR activity in anoxic crucian carp. The iron center of RNRR2
is essential for production and stabilization of the tyrosyl radical. If iron is lost, the radical
is lost and must be regenerated through an oxygen demanding reaction. The amino acid
configuration has been used to explain the low stability of the diiron-oxygen cluster and
tyrosyl radical in mouse RNRR2 (16). The sequence homology between crucian carp and
mouse RNRR2 could suggest that the enzymes have similar properties regarding radical
stability. The mouse RNRR2 radical is readily inactivated by radical scavengers and has a
low stability (25). However, since no structural or spectroscopic data are presently
available for crucian carp RNR, no direct measurements of the radical stability in this
species exists. In mammals, 60% of the iron is lost after 30 min at 37°C, and 30% is lost
after 30 min at 0°C (24). When extrapolated, the data suggests that virtually no iron is left
in the R2 subunit after approximately 2 hours at 0°C. Thus, even at the relatively low
temperature (10°C) in the present study, the iron centre is lost and no radical should be
present- if crucian carp RNR function as mammalian RNR. Since we found no differences
in key amino acids between mouse and crucian carp R2, it is possible that crucian carp has

other means of ensuring radical stability and RNR activity. This could involve other parts
of the enzyme or supporting cellular systems. However, we can not exclude that some of
the differences seen in amino acids outside areas with known importance affect RNR
function in anoxia.
There are two methodological considerations that must be discussed in the present paper.
First, could there be traces of oxygen in the water allowing crucian carp to regenerate the
RNR radical? The oxygen electrode sensitivity is 0.1 % O2, and it is possible that water
oxygen level was just below this detection limit. However, it has been shown that below
0.1% O2 the radical is abolished in mammalian cell cultures exposed for 5 hours (2).
Second, when the fish is injected with BrdU, it is exposed to air for a few seconds.
Although crucian carp is not an air breather, the short contact with air could supply tissues
with some oxygen that could reconstitute the RNR radical and promote some DNA
synthesis. We find this unlikely since such a short and minute supply of oxygen would
probably not produce radicals in more than a fraction of the RNRs. We did not find any
change of cell proliferation in liver throughout the exposures or any incidences of weak
staining indicating partial DNA synthesis. Summed up, it is unlikely that new S-phase cells
are present in anoxia due to a small, undetectable, transient supply of oxygen.
Acknowledgements
We would like to thank Guro Sandvik, Aasa R. Schjølberg and Liza Kravik for technical
assistance, and Lars Thelander and Kristoffer K. Andersson for valuable input. This work
was funded by The Research Council of Norway.

References
1. Bjorklund S, Skog S, Tribukait B, and Thelander L. S-phase-specific expression of mammalian ribonucleotide reductase-R1 and reductase-R2 subunit messenger-RNAs. Biochem 29: 5452-5458, 1990.
2. Brischwein K, Engelcke M, Riedinger HJ, and Probst H. Role of ribonucleotide reductase and deoxynucleotide pools in the oxygen-dependent control of DNA replication in Ehrlich ascites cells. Eur J Biochem 244: 286-293, 1997.
3. Chabes A and Thelander L. Controlled protein degradation regulates ribonucleotide reductase activity in proliferating mammalian cells during the normal cell cycle and in response to DNA damage and replication blocks. J Biol Chem 275: 17747-17753, 2000.
4. Chimploy K, Tassotto ML, and Mathews CK. Ribonucleotide reductase, a possible agent in deoxyribonucleotide pool asymmetries induced by hypoxia. J Biol Chem 275: 39267-39271, 2000.
5. Clegg JS. Embryos of Artemia franciscana survive four years of continuous anoxia: The case for complete metabolic rate depression. J Exp Biol 200: 467-475, 1997.
6. Cowan KJ and Storey KB. Protein kinase and phosphatase responses to anoxia in crayfish, Orconectes virilis: purification and characterization of cAMP-dependent protein kinase. Comp Biochem Physiol B-Biochem & Mol Biol 130: 565-577, 2001.
7. Douglas RM, Farahani R, Morcillo P, Kanaan A, Xu T, and Haddad GG.Hypoxia induces major effects on cell cycle kinetics and protein expression in Drosophila melanogaster embryos. Am Jof Physiol-Reg Int Comp Physiol 288: R511-R521, 2005.
8. Douglas RM and Haddad GG. Genetic models in applied physiology - Invited review: Effect of oxygen deprivation on cell cycle activity: a profile of delay and arrest. J App Physiol 94: 2068-2083, 2003.
9. Engstrom Y, Eriksson S, Jildevik I, Skog S, Thelander L, and Tribukait B. Cell cycle-dependent expression of mammalian ribonucleotide reductase - differentialregulation of the 2 Subunits. J Biol Chem 260: 9114-9116, 1985.
10. Foe VE and Alberts BM. Reversible chromosome condensation induced in Drosophila embryos by anoxia - Visualization of interphase nuclear-organization. J Cell Biol 100: 1623-1636, 1985.
11. Graff P, Seim J, Amellem O, Arakawa H, Nakamura Y, Andersson KK, Stokke T, and Pettersen EO. Counteraction of pRb-dependent protection after extreme hypoxia by elevated ribonucleotide reductase. Cell Prolif 37: 367-383, 2004.
12. Guittet O, Hakansson P, Voevodskaya N, Fridd S, Graslund A, Arakawa H, Nakamura Y, and Thelander L. Mammalian p53R2 protein forms an active ribonucleotide reductase in vitro with the R1 protein, which is expressed both in resting cells in response to DNA damage and in proliferating cells. J Biol Chem276: 40647-40651, 2001.

13. Holopainen IJ, Hyvärinen,H.,Piironen,J. Anaerobic wintering of crucian carp (Carassius carassius L.)--II. Metabolic products. Comp Biochem Physiol 83: 239-242, 1986.
14. Hyvärinen H, Holopainen,J., Piironen,J. Anaerobic wintering of crucian carp -annual dynamics of glycogen reserves in nature. Comp Biochem Physiol 82A: 797-803, 1985.
15. Johnston IA and Bernard LM. Utilization of the ethanol pathway in carp following exposure to anoxia. J Exp Biol 104: 73-78, 1983.
16. Kauppi B, Nielsen BB, Ramaswamy S, Kjøller IL, Thelander M, Thelander L, and Eklund H. The three-dimensional structure of mammalian ribonucleotide reductase protein R2 reveals a more-accessible iron-radical site than Escherichia coli R2. J Mol Biol 262: 706-720, 1996.
17. Kolberg M, Strand KR, Graff P, and Andersson KK. Structure, function, and mechanism of ribonucleotide reductases. Biochim Biophys Acta 1699: 1-34, 2004.
18. Maga G and Hübscher U. Proliferating cell nuclear antigen (PCNA): a dancer with many partners. J Cell Sci 116: 3051-3060, 2003.19. Negishi K, Stell WK, Teranishi T, Karkhanis A, Owusu-Yaw V, and Takasal
Y. Induction of proliferating cell nuclear antigen (PCNA)-immunoreactive cells in goldfish retina following intravitreal injection with 6-hydroxydopamine. Cell Mol Neurobiol 11: 639-659, 1991.
20. Nilsson GE. Evidence for a role of GABA in metabolic depression during anoxia in crucian carp (Carassius carassius). J Exp Biol 164: 243-259, 1992.
21. Nilsson GE. Surviving anoxia with the brain turned on. News Physiol Sci 16: 217-221, 2001.
22. Nilsson GE and Lutz PL. Anoxia tolerant brains. J Cereb Blood Flow Metab 24: 475-486, 2004.
23. Nilsson GE, Rosen,P.,Johansson,D. Anoxic depression of spontaneous locomotor activity in Crucian carp quantified by a computerized imaging technique. J Exp Biol 180: 153-162, 1993.
24. Nyholm S, Mann GJ, Johansson AG, Bergeron RJ, Gräslund A, and Thelander L. Role of ribonucleotide reductase in inhibition of mammalian cell growth by potent iron chelators. J Biol Chem 268: 26200-26205, 1993.
25. Nyholm S, Thelander L, and Graslund A. Reduction and loss of the iron center in the reaction of the small subunit of mouse ribonucleotide reductase with hydroxyurea. Biochemistry-Us 32: 11569-11574, 1993.
26. Ortego LS, Hawkins WE, Walker WW, Krol RM, and Benson WH. Detection of proliferating cell nuclear antigen in tissues of three small fish species. Biotech Histochem 69: 317-323, 1994.
27. Padilla PA, Nystul TG, Zager RA, Johnson ACM, and Roth MB.Dephosphorylation of cell cycle-regulated proteins correlates with anoxia-induced suspended animation in Caenorhabditis elegans. Mol Biol Cell 13: 1473-1483, 2002.
28. Padilla PA and Roth MB. Oxygen deprivation causes suspended animation in the zebrafish embryo. P Natl Acad Sci USA 98: 7331-7335, 2001.
29. Probst H, Gekeler V, and Helftenbein E. Oxygen Dependence of Nuclear-DNA Replication in Ehrlich Ascites-Cells. Exp Cell Res 154: 327-341, 1984.

30. Probst H, Schiffer H, Gekeler V, Kienzlepfeilsticker H, Stropp U, Stotzer KE, and Frenzelstotzer I. Oxygen Dependent Regulation of DNA-Synthesis and Growth of Ehrlich Ascites Tumor-Cells Invitro and Invivo. Cancer Res 48: 2053-2060, 1988.
31. Ramakers C, Ruijter JM, Deprez RHL, and Moorman AFM. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci Lett 339: 62-66, 2003.
32. Rozen S and Skaletsky HJ. Primer3 on the WWW for general users and for biologist programmers. Totowa, NJ: Humana Press, 2000.
33. Sjoberg BM. Ribonucleotide reductases - A group of enzymes with different metallosites and a similar reaction mechanism. Struct Bond 88: 139-173, 1997.
34. Sollid J, De Angelis P, Gundersen K, and Nilsson GE. Hypoxia induces adaptive and reversible gross morphological changes in crucian carp gills. J Exp Biol 206: 3667-3673, 2003.
35. Stecyk JA, Stenslokken KO, Farrell AP, and Nilsson GE. Maintained cardiac pumping in anoxic crucian carp. Science 306: 77, 2004.
36. Storey KB. Metabolic adaptations supporting anoxia tolerance in reptiles: Recent advances. Comp Biochem Phys B 113: 23-35, 1996.
37. Storey KB. Tissue-specific controls on carbohydrate catabolism during anoxia in goldfish. Physiol Zool 60: 601-607, 1987.
38. Strand KR, Karlsen S, Kolberg M, Rohr AK, Gorbitz CH, and Andersson KK. Crystal structural studies of changes in the native dinuclear iron center of ribonucleotide reductase protein R2 from mouse. J Biol Chem 279: 46794-46801, 2004.
39. Tanaka H, Arakawa H, Yamaguchi T, Shiraishi K, Fukuda S, Matsui K, Takei Y, and Nakamura Y. A ribonucleotide reductase gene involved in a p53-dependent cell-cycle checkpoint for DNA damage. Nature 404: 42-49, 2000.
40. Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, and Higgins DG. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25: 4876-4882, 1997.

Fig. 1a-b. Scanning micrographs of crucian carp gill filament in normoxia (a) and after 7
days of anoxia exposure (b). There were no apparent differences in gill morphology
between the two exposure groups, as both groups displayed gill filaments with no
protruding lamellae. Four fish from each group were examined. Scale bar = 50 µm
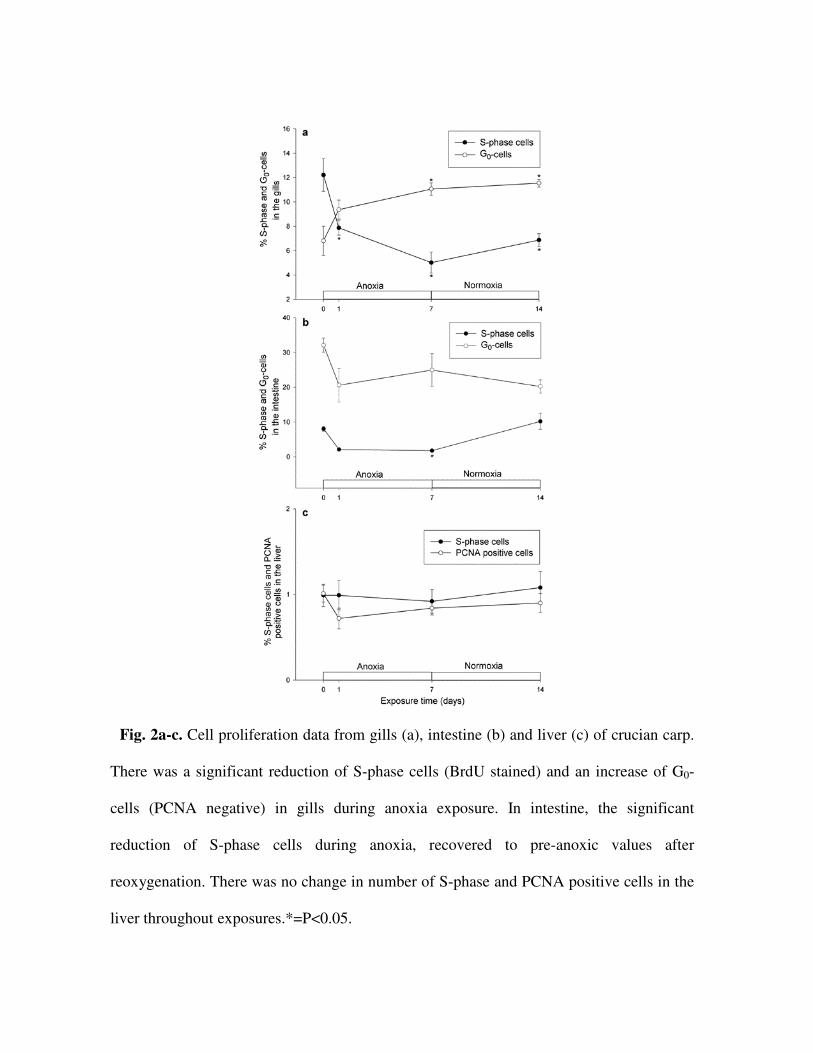
Fig. 2a-c. Cell proliferation data from gills (a), intestine (b) and liver (c) of crucian carp.
There was a significant reduction of S-phase cells (BrdU stained) and an increase of G0-
cells (PCNA negative) in gills during anoxia exposure. In intestine, the significant
reduction of S-phase cells during anoxia, recovered to pre-anoxic values after
reoxygenation. There was no change in number of S-phase and PCNA positive cells in the
liver throughout exposures.*=P<0.05.

Fig. 3a-f. Staining of cell proliferation in gills (a-d) and intestinal crypts (e-f) of normoxic
and 7 day anoxic crucian carp. Pictures (a) and (b) show BrdU positive S-phase cells
(stained dark brown, exemplified with arrowheads) in gill filament of normoxic and anoxic
crucian carp. PCNA positive cells (stained dark brown) are shown in pictures (c) and (d),
where lack of PCNA positive cells in the lamellae makes these stand out by the blue
(haematoxylin) counter staining. Scale bars = 50µm.
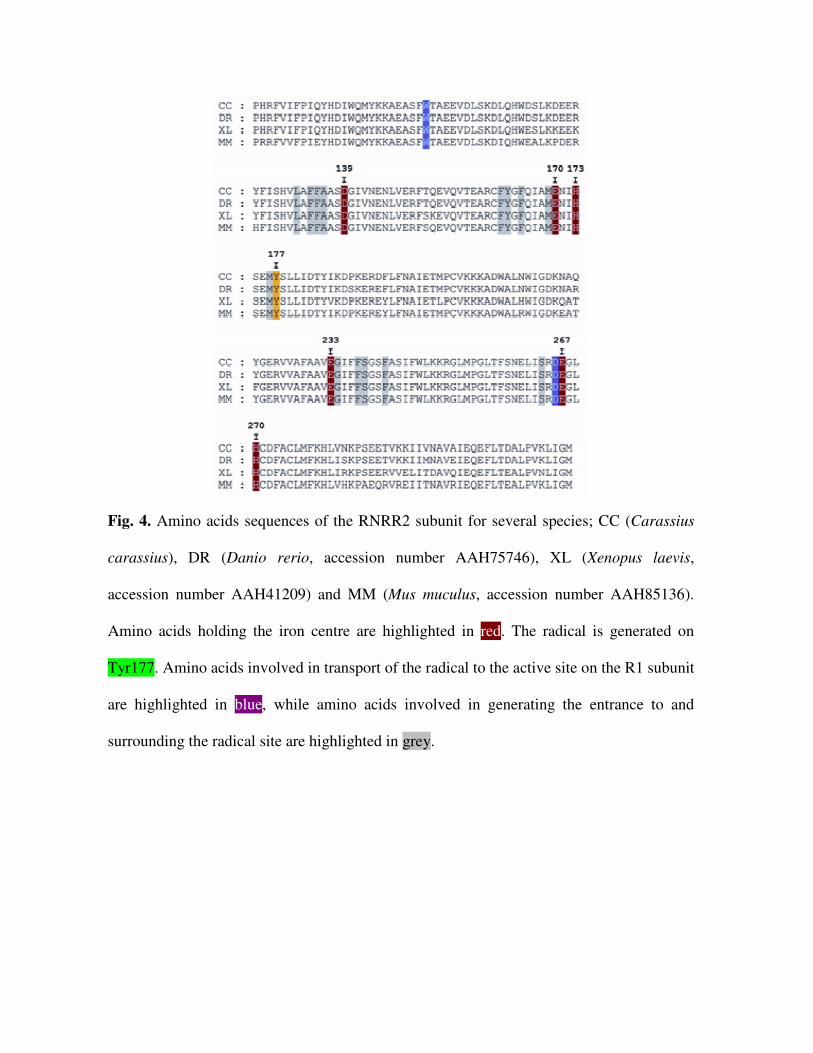
Fig. 4. Amino acids sequences of the RNRR2 subunit for several species; CC (Carassius
carassius), DR (Danio rerio, accession number AAH75746), XL (Xenopus laevis,
accession number AAH41209) and MM (Mus muculus, accession number AAH85136).
Amino acids holding the iron centre are highlighted in red. The radical is generated on
Tyr177. Amino acids involved in transport of the radical to the active site on the R1 subunit
are highlighted in blue, while amino acids involved in generating the entrance to and
surrounding the radical site are highlighted in grey.
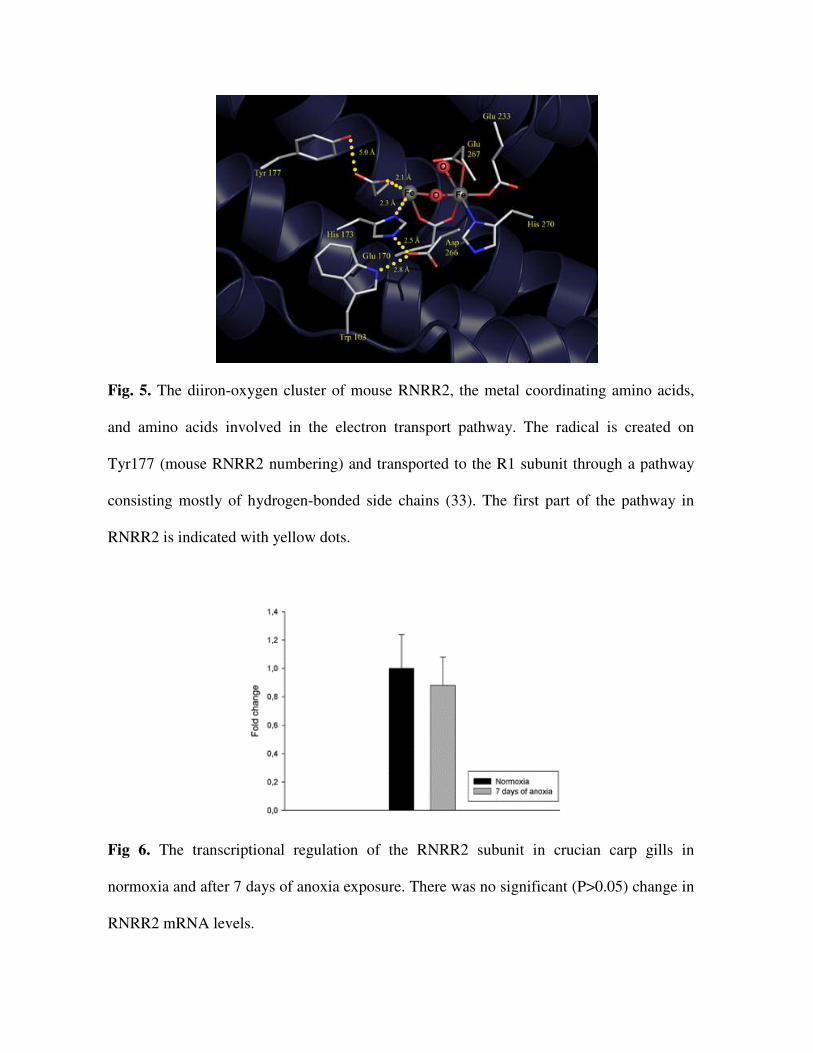
Fig. 5. The diiron-oxygen cluster of mouse RNRR2, the metal coordinating amino acids,
and amino acids involved in the electron transport pathway. The radical is created on
Tyr177 (mouse RNRR2 numbering) and transported to the R1 subunit through a pathway
consisting mostly of hydrogen-bonded side chains (33). The first part of the pathway in
RNRR2 is indicated with yellow dots.
Fig 6. The transcriptional regulation of the RNRR2 subunit in crucian carp gills in
normoxia and after 7 days of anoxia exposure. There was no significant (P>0.05) change in
RNRR2 mRNA levels.