ccr7 deficiency on dendritic cells enhances fungal clearance in a murine model of pulmonary invasive...
TRANSCRIPT

of March 3, 2016.This information is current as
Model of Pulmonary Invasive AspergillosisEnhances Fungal Clearance in a Murine CCR7 Deficiency on Dendritic Cells
HogaboamAdam J. Hartigan, John Westwick, Gabor Jarai and Cory M.
http://www.jimmunol.org/content/183/8/5171doi: 10.4049/jimmunol.0901027September 2009;
2009; 183:5171-5179; Prepublished online 25J Immunol
Referenceshttp://www.jimmunol.org/content/183/8/5171.full#ref-list-1
, 20 of which you can access for free at: cites 47 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2009 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on March 3, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on March 3, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

CCR7 Deficiency on Dendritic Cells Enhances FungalClearance in a Murine Model of Pulmonary InvasiveAspergillosis1
Adam J. Hartigan,* John Westwick,† Gabor Jarai,† and Cory M. Hogaboam2*
Aspergillus fumigatus is a sporulating fungus found ubiquitously in the environment and is easily cleared from immunocompetenthosts. Invasive aspergillosis develops in immunocompromised patients, and is a leading cause of mortality in hematopoietic stemcell transplant recipients. CCR7 and its ligands, CCL19 and CCL21, are responsible for the migration of dendritic cells from sitesof infection and inflammation to secondary lymphoid organs. To investigate the role of CCR7 during invasive aspergillosis, we useda well-characterized neutropenic murine model. During invasive aspergillosis, mice with a CCR7 deficiency in the hematopoieticcompartment exhibited increased survival and less pulmonary injury compared with the appropriate wild-type control. Flowcytometric analysis of the chimeric mice revealed an increase in the number of dendritic cells present in the lungs of CCR7-deficient chimeras following infection with Aspergillus conidia. An adoptive transfer of dendritic cells into neutropenic miceprovided a protective effect during invasive aspergillosis, which was further enhanced with the adoptive transfer of CCR7-deficientdendritic cells. Additionally, CCR7-deficient dendritic cells activated in vitro with Aspergillus conidia expressed higher TNF-�,CXCL10, and CXCL2 levels, indicating a more activated cellular response to the fungus. Our results suggest that the absence ofCCR7 is protective during invasive aspergillosis in neutropenic mice. Collectively, these data demonstrate a potential deleteriousrole for CCR7 during primary immune responses directed against A. fumigatus. The Journal of Immunology, 2009, 183: 5171–5179.
A spergillus fumigatus is a sporulating fungus present ubiq-uitously in the environment (1). Although most individ-uals are not affected by exposure to Aspergillus, the fun-
gus can cause a broad spectrum of diseases, ranging fromhypersensitivity reactions, such as allergic bronchopulmonary as-pergillosis, to serious opportunisitic infections, including chronicpulmonary necrotizing aspergillosis and invasive aspergillosis (2).Invasive aspergillosis is a rapidly progressive disease, often orig-inating in the pulmonary system, in which inhaled conidia from A.fumigatus germinate into hyphae and invade the lung parenchyma,leading to pneumonia and massive inflammation (3). This severe,usually fatal disease disproportionally affects immunocompro-mised individuals and is a leading cause of mortality in hemato-poietic stem cell transplant recipients (4–7). Neutropenia was ini-tially described as the most significant risk factor for thedevelopment of invasive aspergillosis (8), but mounting evidencehas shown the importance of several additional effector cell pop-ulations, most notably myeloid dendritic cells (DCs)3 (9–11).
DCs play a crucial role during the immune response to A. fu-migatus. Studies have shown that DCs become activated by A.fumigatus conidia and hyphae, leading to DC maturation and pro-duction of inflammatory cytokines, such as TNF-�, IL-12, IL-6,and IL-10 (9). Mature DCs have been shown to clear both A. fu-migatus conidia and hyphal elements, indicating their importancebefore, and following, the onset of invasive disease (12). DCs alsoparticipate in the adaptive immune response: when conidia-acti-vated DCs were adoptively transferred, they activated IFN-�-pro-ducing T cells, leading to fungal resistance.
In addition to effector cell production of cytokines in response toA. fumigatus, several chemokines and their receptors have beenreported to play a significant role during invasive aspergillosis (10,13–15). A key element of DC function is the ability to migrate tosites of inflammation and infection, which is mediated by CCR6expressed on immature DCs (16, 17). A recent study has shownthat CCR6�/� mice fail to recruit adequate DC numbers to thelung following conidia challenge, and they consequently had sig-nificantly enhanced morbidity and mortality compared with wild-type mice (15). Another study, using human DCs, showed thatupon internalization of Aspergillus conidia, DCs significantly up-regulated their expression of CCR7, in addition to antifungal cy-tokines (18, 19).
In our studies, we were interested in the role of CCR7 on DCsduring invasive aspergillosis. CCR7 is present on several cell sub-sets in addition to DCs, most notably T cells and B cells (20–22).CCR7 binds the ligands CCL19 and CCL21, which are expressedconstitutively in secondary lymphoid organs (20, 23–25). CCR7 isup-regulated on DCs after exposure to a maturing stimulus, allow-ing DCs to migrate away from sites of inflammation to secondarylymphoid organs. Here, the fully mature DC, expressing costimu-latory molecules, is able to prime an adaptive immune response(26).
*Department of Pathology, University of Michigan Medical School, Ann Arbor, MI48109; and †Novartis Institutes of Biomedical Research, Respiratory Disease Area,Horsham, West Sussex, United Kingdom
Received for publication March 31, 2009. Accepted for publication August 6, 2009.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by National Institutes of Health Grant HL069865 (toC.M.H.) and Novartis Institutes for Biomedical Research.2 Address correspondence and reprint requests to Dr. Cory M. Hogaboam, Depart-ment of Pathology, University of Michigan Medical School, Room 4057, BSRB,109 Zina Pitcher Place, Ann Arbor, MI 48109. E-mail address: [email protected] Abbreviations used in this paper: DC, myeloid dendritic cell; BMDC, bone marrow-derived dendritic cell; GMS, Gomori methenamine silver; i.t., intratracheal(ly).
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0901027
by guest on March 3, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

Herein we address the role of CCR7 in a well-characterizedmurine model of invasive aspergillosis. We found that CCR7 de-ficiency (CCR7�/�) in the hematopoietic compartment markedlyenhanced survival and fungal clearance following intratracheal(i.t.) administration of A. fumigatus conidia into neutropenic mice.This protective effect was associated with a significantly higherinflux of DCs into the lungs of CCR7�/� bone marrow chimeramice than into the lungs of CCR7-sufficient chimeras. Addition-ally, it was found that CCR7�/� bone marrow-derived DCs (BM-DCs) had a more activated cellular response to Aspergillus conidiathan did wild-type BMDCs, and adoptive transfer of CCR7�/�
BMDCs more effectively protected neutropenic wild-type micefrom invasive aspergillosis than did wild-type BMDCs. Thus,CCR7 expression negatively regulates the innate response to As-pergillus conidia, thereby permitting the development of invasivepulmonary aspergillosis.
Materials and MethodsMice
Wild-type female C57BL/6 mice (6–8 wk of age) were purchased fromThe Jackson Laboratory. CCR7�/� mice, generated on a C57BL/6 back-ground by Dr. M. Lipp’s group (Max Delbruck Center for Molecular Med-icine, Berlin, Germany), as previously described (27), were a kind gift fromS. Lira (Mount Sinai School of Medicine, New York, NY), and breedingcolonies of these mice were established and maintained in the University ofMichigan Medical School Laboratory of Animal Medicine facility. For allexperiments, mice were sex-matched and used between 8 and 10 wk of age.All animals were used in accordance with regulations mandated by theUniversity Committee on Use and Care of Animals at the University ofMichigan.
Generation of bone marrow chimeras
Wild-type mice were lethally irradiated with 1000 cGy using a cesiumsource. Within 12 h of irradiation, recipient mice were reconstituted with5.0 � 106 whole bone marrow cells in PBS from wild-type or CCR7�/�
mice via tail vein injection. Mice were considered fully chimeric 8–10 wkfollowing bone marrow transplantation.
Murine model of invasive aspergillosis
Mice were depleted of neutrophils with an i.p. injection of 100 �g ofRB6–8C5 (anti-Gr-1) as previously described (28). One day after the in-duction of neutropenia, mice were anesthetized with a mixture of ketamineand xylazine and given an i.t. injection of 6.0 � 106 A. fumigatus conidiasuspended in 30 �l of 0.1% Tween 80 in PBS. The highly virulent A.fumigatus strain 13073 (American Type Culture Collection) was used toelicit a reproducible form of invasive aspergillosis in anti-Gr-1-treatedmice (28).
Lung histology
Lungs were collected 2 days after conidial challenge from wild-type andCCR7�/� mice or wild-type and CCR7�/� chimeras. Whole lung samplesfor histologic analysis were excised, perfused with 10% formalin, andplaced in fresh formalin for an additional 24–48 h. Routine histologictechniques were used to paraffin-embed this tissue, and 5-�m sections ofwhole lung were stained with H&E or with Gomori methenamine silver(GMS) stain to visualize A. fumigatus conidia and hyphal elements (blackstaining). Image capture was conducted with an Olympus BX40F micro-scope equipped with �20/0.5 and �100/1.3 objective lenses and a �10eyepiece (Olympus). Digital photographs were obtained with a Sony 3CCD color video camera (model no. DXC-960MD), and IP Lab Spectrumsoftware was used for image acquisition (Scanalytics).
RNA isolation and real-time TaqMan PCR analysis
Total RNA was isolated from homogenized mouse lungs or BMDCs withthe TRIzol reagent (Invitrogen). Approximately 5 �g of purified RNA wasreverse transcribed to yield cDNA. CCR7, CCL19, IL-12p35, IL-17a,TNF-�, CXCL10, CXCL2, CCL5, IL-10, MHC II, CD86, CD40, TLR2,TLR4, and dectin-1 gene expression was analyzed by real-time quantitativeRT-PCR using an ABI Prism 7500 sequence detection system (AppliedBiosystems). For quantitative TaqMan analysis of CCL21, primers werecustom made as previously described (29). GAPDH was analyzed as an
internal control, and gene expression was normalized to GAPDH. Foldchanges in gene expression levels were calculated by comparison of thegene expression in unchallenged samples or mice, which were assigned anarbitrary value of 1. Additional analysis was performed by assigning wild-type challenged mice or BMDCs a value of 1 and comparing this withCCR7�/� challenged samples.
Protein analysis of lung cytokine levels
Whole lung cytokine and chemokine levels from wild-type or CCR7�/�
chimeras were measured using a Bio-Plex bead-based cytokine assay pur-chased from Invitrogen or Bio-Rad and analyzed on the Bio-Rad Bio-Plex200 system according to the manufacturer’s protocol.
Flow cytometry
Whole lung samples were minced using surgical scissors and incubated inRPMI 1640 supplemented with 5% FCS, type IV collagenase (Sigma-Al-drich), and DNase for 45 min. Cells were flushed through a nylon meshfilter and washed with FACS buffer, consisting of Ca2�- and Mg2�-freePBS with 0.1% azide, 1% BSA, and 5 mM EDTA as described (30). Beforesurface staining with labeled Abs, nonspecific binding was blocked byincubating cells with purified rat anti-mouse CD16/CD32 (Fc�III/II recep-tor) mAb from eBioscience. Flow cytometry analysis was performed witha Beckman Coulter Cytomics FC 500, and data were analyzed usingFlowJo 8.2 software.
Chitin assay
Chitin, a major constituent of the hyphal cell wall, is absent in mammaliantissue and thus can be used as a method to determine the fungal burden inthe lungs of infected mice. The chitin assay was performed as described(10).
In vitro BMDC culture
Bone marrow cells were flushed from the femurs and tibiae of wild-type orCCR7�/� mice with sterile media. A single-cell suspension was obtainedby filtering bone marrow cells through a 70-�m nylon mesh filter. Bonemarrow cells were plated at a concentration of 3.0 � 106 cells/10 ml inRPMI 1640 media containing 20 ng/ml murine GM-CSF (R&D Systems).Media was replenished at 3 days and BMDCs were purified at day 6 usinganti-CD11c magnetic beads (Miltenyi Biotec). Cells were replated in freshmedia, without GM-CSF, and incubated with live A. fumigatus conidia ata 1:5 ratio. Using the TRIzol reagent, RNA was extracted from the BMDCsafter 2 h of culture with conidia to prevent fungal overgrowth and to pre-vent the use of antifungals, which can affect DC function and maturation(18). To determine T cell stimulatory capacity and BMDC surface expres-sion of CD86, wild-type and CCR7�/� BMDCs were cultured with conidiaat a 1:5 ratio for 2 h, at which time amphotericin B was added to allcultures. At 24 h, BMDCs were analyzed by flow cytometry for CD86expression or were used to stimulate BALB/c T cells in an allogeneicMLR. At 72 h, T cell proliferation was measured by [3H]thymidineincorporation.
DC transfer and tracking
In vitro-derived wild-type or CCR7�/� BMDCs were cultured as de-scribed, and in some experiments were labeled with CFSE. BMDCs (1.0 �106) were injected 6 h before conidia challenge or were coinjected withconidia (6.0 � 106 to 107) into wild-type mice 1 day after administrationof 100 �g of anti-Gr-1. One set of mice was euthanized 24 h after theinjection, and lungs were analyzed for CFSE expression. The remainingmice were sacrificed at 48 h for lung histology, protein, and chitin analysis.
Statistical analysis
All results are expressed as the means � SE representative of two or moreexperiments. A Student’s t test or ANOVA was used to determine statis-tical significance between wild-type and CCR7�/� mice, chimeras, orBMDCs. Survival rates were expressed as percentages, and a log-rank test(�2 test) was used to detect differences in mouse survival. Values of p �0.05 were considered statistically significant.
ResultsCCR7 and its ligands, CCL19 and CCL21, were highlyexpressed in the lung during invasive aspergillosis
During the past decade it has become increasingly clear that che-mokines and their receptors play an important role in the antifun-gal response during invasive aspergillosis (10, 13, 15, 28, 31). To
5172 ROLE OF CCR7 DURING ASPERGILLOSIS
by guest on March 3, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
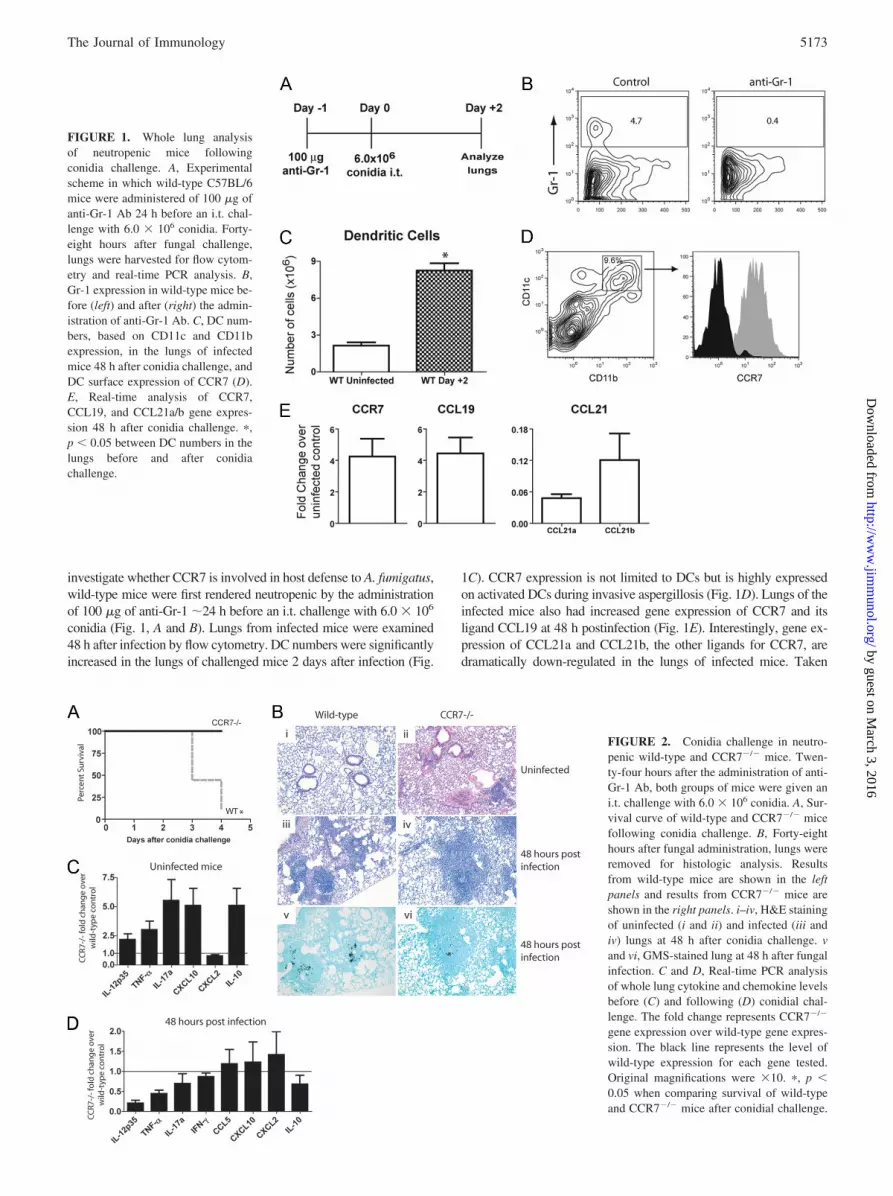
investigate whether CCR7 is involved in host defense to A. fumigatus,wild-type mice were first rendered neutropenic by the administrationof 100 �g of anti-Gr-1 �24 h before an i.t. challenge with 6.0 � 106
conidia (Fig. 1, A and B). Lungs from infected mice were examined48 h after infection by flow cytometry. DC numbers were significantlyincreased in the lungs of challenged mice 2 days after infection (Fig.
1C). CCR7 expression is not limited to DCs but is highly expressedon activated DCs during invasive aspergillosis (Fig. 1D). Lungs of theinfected mice also had increased gene expression of CCR7 and itsligand CCL19 at 48 h postinfection (Fig. 1E). Interestingly, gene ex-pression of CCL21a and CCL21b, the other ligands for CCR7, aredramatically down-regulated in the lungs of infected mice. Taken
FIGURE 2. Conidia challenge in neutro-penic wild-type and CCR7�/� mice. Twen-ty-four hours after the administration of anti-Gr-1 Ab, both groups of mice were given ani.t. challenge with 6.0 � 106 conidia. A, Sur-vival curve of wild-type and CCR7�/� micefollowing conidia challenge. B, Forty-eighthours after fungal administration, lungs wereremoved for histologic analysis. Resultsfrom wild-type mice are shown in the leftpanels and results from CCR7�/� mice areshown in the right panels. i–iv, H&E stainingof uninfected (i and ii) and infected (iii andiv) lungs at 48 h after conidia challenge. vand vi, GMS-stained lung at 48 h after fungalinfection. C and D, Real-time PCR analysisof whole lung cytokine and chemokine levelsbefore (C) and following (D) conidial chal-lenge. The fold change represents CCR7�/�
gene expression over wild-type gene expres-sion. The black line represents the level ofwild-type expression for each gene tested.Original magnifications were �10. �, p �0.05 when comparing survival of wild-typeand CCR7�/� mice after conidial challenge.
FIGURE 1. Whole lung analysisof neutropenic mice followingconidia challenge. A, Experimentalscheme in which wild-type C57BL/6mice were administered of 100 �g ofanti-Gr-1 Ab 24 h before an i.t. chal-lenge with 6.0 � 106 conidia. Forty-eight hours after fungal challenge,lungs were harvested for flow cytom-etry and real-time PCR analysis. B,Gr-1 expression in wild-type mice be-fore (left) and after (right) the admin-istration of anti-Gr-1 Ab. C, DC num-bers, based on CD11c and CD11bexpression, in the lungs of infectedmice 48 h after conidia challenge, andDC surface expression of CCR7 (D).E, Real-time analysis of CCR7,CCL19, and CCL21a/b gene expres-sion 48 h after conidia challenge. �,p � 0.05 between DC numbers in thelungs before and after conidiachallenge.
5173The Journal of Immunology
by guest on March 3, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

together, these results suggest that CCR7 and its ligands play an im-portant role in the immune response to A. fumigatus.
CCR7�/� mice are protected from invasive aspergillosis
To examine the effect of CCR7 during invasive pulmonary as-pergillosis, we infected neutropenic wild-type and CCR7�/� micewith 6.0 � 106 conidia. CCR7�/� mice were less susceptible todeath following fungal challenge when compared with their wild-type counterparts (Fig. 2A). It was observed that the lungs of un-infected CCR7�/� mice had significant cellular infiltrate, whichincluded the formation of lymphoid aggregates (Fig. 2B, i and ii).Although there was substantial leukocytic infiltration in both wild-type and CCR7�/� mice 48 h following conidial challenge, theCCR7�/� mice demonstrated a decrease in pulmonary fungal el-ements as determined by GMS staining (Fig. 2B, iii–vi). Usingreal-time PCR, the gene expression of whole lung cytokine levelswas determined in uninfected mice and in mice 48 h after A. fu-migatus infection (Fig. 2, C and D). Uninfected CCR7�/� micehad elevated levels of IL-12p35, TNF-�, IL-17a, CXCL10, andIL-10, when compared with unchallenged wild-type mice. Addi-tionally, CCR7�/� mice had significantly higher gene expressionof IFN-� before infection (data not shown). Interestingly, 48 hfollowing conidial challenge, CCR7�/� mice had lower expres-sion of IL-12p35, TNF-�, �L-17a, IFN-�, and IL-10 RNA than didwild-type mice (Fig. 2D). Taken together, these data indicate thatCCR7�/� mice are protected from invasive aspergillosis and havea regulated inflammatory response due to the accelerated clearanceof A. fumigatus.
Deficiency of CCR7 on hematopoietic cells significantlyenhanced survival and decreased fungal burden followingA. fumigatus conidia challenge
Previous studies, and work in our laboratory, have demonstratedthat CCR7�/� mice exhibit anatomical abnormalities, such as al-terations in secondary lymphoid organs, and generalized autoim-
munity (27, 32). Given this feature of CCR7�/� mice, we electedto make bone marrow chimeras to examine the effect of CCR7solely on the hematopoietic system during invasive aspergillosis.CCR7�/� chimeras had significantly lower mortality (10%) thandid their wild-type counterparts (66%) at day 5 postinfection withA. fumigatus (Fig. 3A). Analysis of whole lung chitin levels re-vealed that CCR7�/� chimeric mice had a significantly lower fun-gal burden compared with wild-type chimeras (Fig. 3B). Consis-tent with these data, GMS staining of whole lung sections showedsignificant fungal growth and hyphal formation in the wild-typechimera lung, which was widely absent in the CCR7�/� chimeras(Fig. 3, C–H). The lungs of uninfected CCR7�/� chimeras exhib-ited some lymphoid aggregates made up predominantly of B cellsand T cells as well as additional cell infiltrate, which was notobserved in the wild-type chimeras (Fig. 3, C and D). Interestingly,this infiltrate was markedly less than that observed in naiveCCR7�/� mice of the same age. Two days after conidia challenge,H&E staining showed significant cell infiltrate in the lungs of bothmice, but to a greater extent in the wild-type chimeras, when com-pared with CCR7�/� chimeras and to the unchallenged controls(Fig. 3, C–F). This cellular infiltrate corresponded to a massiveinflammatory response to A. fumigatus. Despite the influx of cellsin both groups of mice, wild-type chimeras were unable to controlthe growth of the fungus, while the knockout animals did not de-velop invasive aspergillosis (Fig. 3, G and H). These data clearlydemonstrate that neutropenic CCR7�/� chimeric mice are pro-tected from invasive aspergillosis.
Lung leukocyte cell recruitment following invasive aspergillosischallenge
Since treatment of mice with anti-Gr-1 eliminates neutrophils, amajor effector cell type in the aspergillus-infected lung, we exam-ined whole lung samples by flow cytometery to identify the cellsproviding the protective effect observed in CCR7�/� chimeras.
FIGURE 3. Survival, lung chitin content, and histology of wild-type and CCR7�/� bone marrow chimeras challenged with A. fumigatus conidia.Wild-type or CCR7�/� chimeric mice were given 100 �g of anti-Gr-1 Ab 24 h before an i.t. challenge with 6.0 � 106 conidia and followed for survivalor analyzed at 48 h. A, Survival of wild-type and CCR7�/� chimeras; n � 10 for wild-type mice and n � 12 for CCR7�/� mice. B, Lung chitin contentdetermined 48 h after the onset of infection. C–H, Histologic analysis of lung tissue 2 days after conidia challenge. H&E staining of uninfected lungs (Cand D) or 48 h after conidia challenge (E and F). G and H, Representative GMS-stained sections from wild-type (G) or CCR7�/� chimeras (H) followingconidial challenge; fungal elements are stained in black. Original magnification was �20 for H&E-stained sections and �40 for GMS sections. �, p � 0.05when comparing survival and chitin in wild-type and CCR7�/� chimeras.
5174 ROLE OF CCR7 DURING ASPERGILLOSIS
by guest on March 3, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

We saw a dramatic increase in myeloid DCs and immature my-eloid cells (monocytes and recruited macrophages) at 48 h afteraspergillus challenge in both the wild-type and CCR7�/� groups(Fig. 4). Conversely, the numbers of T cells, B cells, and NK cellswere decreased 2 days after conidia challenge. While immaturemyeloid cell numbers were elevated in both groups, flow cytomet-ric data revealed higher percentages of myeloid DCs in the lungsof CCR7�/� chimeric mice, which, in combination with higherabsolute number of cells in the lungs of the knockout animals,resulted in a significantly greater number of DCs present (Fig. 4).It has been previously reported that DCs provide protection againstinvasive aspergillosis and that mice lacking adequate numbers ofDCs are susceptible to disease (15). Therefore, antifungal re-sponses in CCR7�/� chimeras appear to be the result of an en-hanced accumulation of DCs.
Whole lung protein analysis 48 h after onset of invasiveaspergillosis
Inflammation is required for an appropriate antifungal immune re-sponse, but it has been reported that an uncontrolled inflammatoryresponse is detrimental to the host (33, 34). We found that proteinlevels were significantly higher in both groups of mice 2 days afterinfection when compared with naive controls (Fig. 5). Unlike wild-type and CCR7�/� mice, there is no significant difference in thebaseline cytokine expression between uninfected wild-type chime-ras and CCR7�/� chimeras (Figs. 2C and 5). The reduction ofproinflammatory cytokine expression in uninfected CCR7�/� chi-meras compared with CCR7�/� mice may be due to the reduction
in lymphoid aggregates in the lung (Figs. 2Bii and 3D). Similarly toinfected wild-type mice, conidial challenge in wild-type chimeras re-sults in significantly higher expression of TNF-�, CCL2, CCL5, andCCL3, but lower levels of CXCL9 and CXCL10, when comparedwith CCR7�/� chimeras. While CCR7�/� chimeras have lower lev-els of inflammatory mediators, both groups produce the same amountof the antiinflammatory cytokine IL-10. Taken together, these find-ings suggest that the inflammatory response in the lung of CCR7�/�
chimeric mice is appropriately regulated before and during infection,leading to the rapid clearance of A. fumigatus.
Characterization of wild-type and CCR7�/� DCs followingaspergillus challenge in vitro
Survival, histological, and fungal growth differences betweenwild-type and CCR7�/� chimeras appear to be the result of theaccumulation of DCs in the lungs of the knockout mice. However,these differences could also be the consequence of differential ac-tivation and maturation by CCR7�/� DCs, or due to a difference infungal recognition by these chemokine receptor-deficient DCs. Todetermine whether differences exist between wild-type andCCR7�/� BMDCs, bone marrow from both mice was grown invitro for 8 days with GM-CSF and the resulting BMDCs werechallenged with Aspergillus conidia. Two hours after fungal chal-lenge, CCR7�/� and wild-type BMDCs had very similar gene ex-pression of the costimulatory molecules MHC II, CD86, and CD40(Fig. 6A). When RNA was analyzed for expression of antifungal,proinflammatory cytokines at 2 h, CCR7�/� BMDCs showedhigher levels of TNF-�, CXCL10, CCL2, and CXCL2 and similar
FIGURE 4. Leukocyte differentials in the lung at 48 h after A. fumigatus challenge in wild-type and CCR7�/� bone marrow chimeras. The left lung lobefrom wild-type or CCR7�/� chimeras was analyzed by flow cytometery 48 h after fungal challenge. Myeloid DCs were CD11chigh, CD11bhigh cells (asshown in the dot plots). Monocytes/macrophages were CD11b�, F4/80�, and CD11c�. NK cells were NK1.1�, T cells were CD3� and CD4� or CD8�,and B cells were CD19�. �, p � 0.05 when comparing DCs in wild-type and CCR7�/� mice on day 2 following infection.
5175The Journal of Immunology
by guest on March 3, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
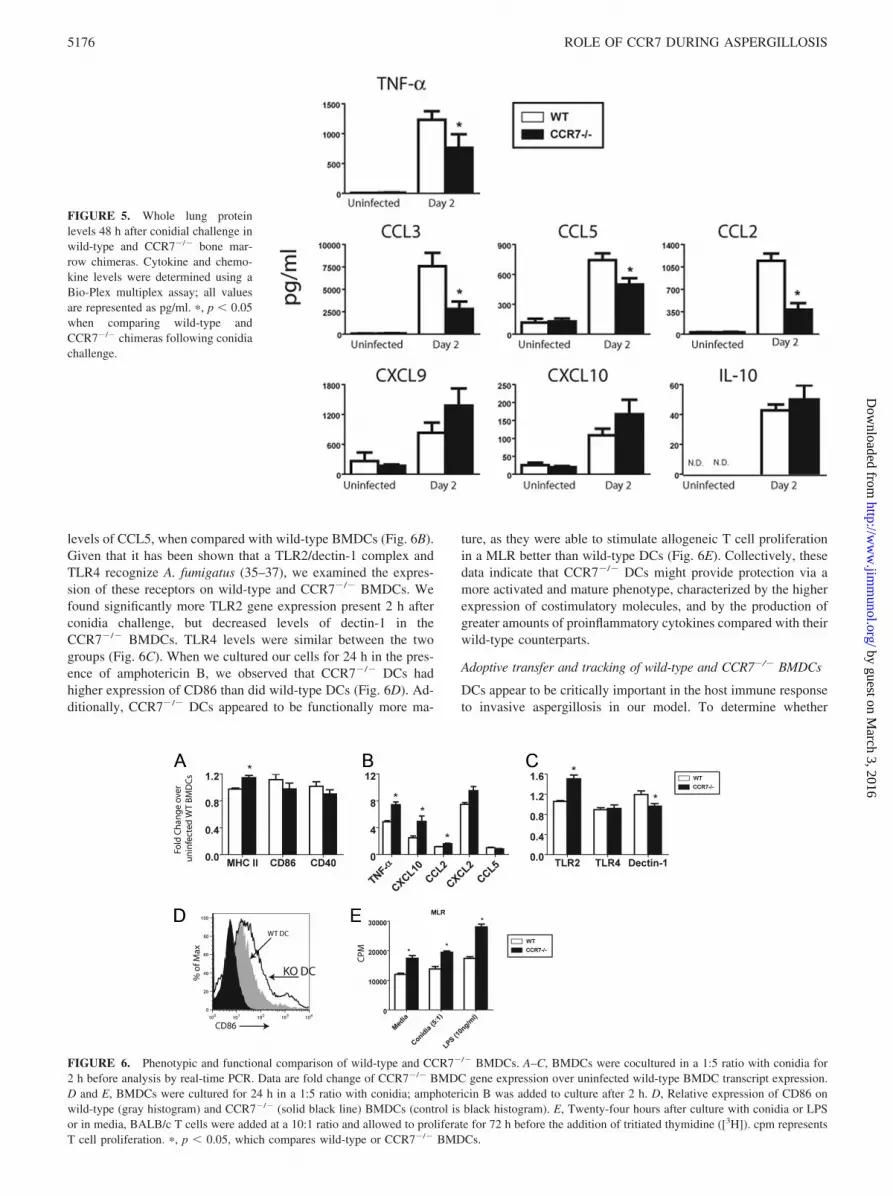
levels of CCL5, when compared with wild-type BMDCs (Fig. 6B).Given that it has been shown that a TLR2/dectin-1 complex andTLR4 recognize A. fumigatus (35–37), we examined the expres-sion of these receptors on wild-type and CCR7�/� BMDCs. Wefound significantly more TLR2 gene expression present 2 h afterconidia challenge, but decreased levels of dectin-1 in theCCR7�/� BMDCs. TLR4 levels were similar between the twogroups (Fig. 6C). When we cultured our cells for 24 h in the pres-ence of amphotericin B, we observed that CCR7�/� DCs hadhigher expression of CD86 than did wild-type DCs (Fig. 6D). Ad-ditionally, CCR7�/� DCs appeared to be functionally more ma-
ture, as they were able to stimulate allogeneic T cell proliferationin a MLR better than wild-type DCs (Fig. 6E). Collectively, thesedata indicate that CCR7�/� DCs might provide protection via amore activated and mature phenotype, characterized by the higherexpression of costimulatory molecules, and by the production ofgreater amounts of proinflammatory cytokines compared with theirwild-type counterparts.
Adoptive transfer and tracking of wild-type and CCR7�/� BMDCs
DCs appear to be critically important in the host immune responseto invasive aspergillosis in our model. To determine whether
FIGURE 5. Whole lung proteinlevels 48 h after conidial challenge inwild-type and CCR7�/� bone mar-row chimeras. Cytokine and chemo-kine levels were determined using aBio-Plex multiplex assay; all valuesare represented as pg/ml. �, p � 0.05when comparing wild-type andCCR7�/� chimeras following conidiachallenge.
FIGURE 6. Phenotypic and functional comparison of wild-type and CCR7�/� BMDCs. A–C, BMDCs were cocultured in a 1:5 ratio with conidia for2 h before analysis by real-time PCR. Data are fold change of CCR7�/� BMDC gene expression over uninfected wild-type BMDC transcript expression.D and E, BMDCs were cultured for 24 h in a 1:5 ratio with conidia; amphotericin B was added to culture after 2 h. D, Relative expression of CD86 onwild-type (gray histogram) and CCR7�/� (solid black line) BMDCs (control is black histogram). E, Twenty-four hours after culture with conidia or LPSor in media, BALB/c T cells were added at a 10:1 ratio and allowed to proliferate for 72 h before the addition of tritiated thymidine ([3H]). cpm representsT cell proliferation. �, p � 0.05, which compares wild-type or CCR7�/� BMDCs.
5176 ROLE OF CCR7 DURING ASPERGILLOSIS
by guest on March 3, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
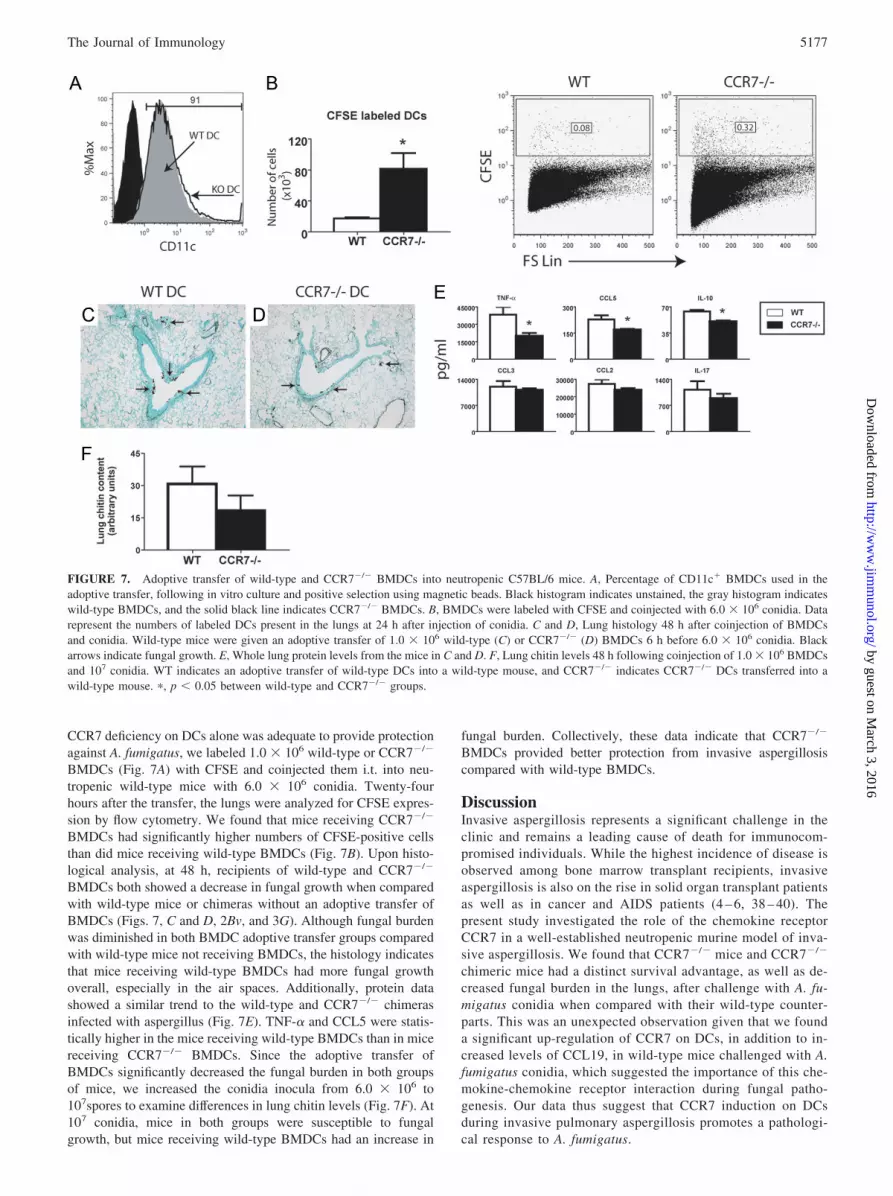
CCR7 deficiency on DCs alone was adequate to provide protectionagainst A. fumigatus, we labeled 1.0 � 106 wild-type or CCR7�/�
BMDCs (Fig. 7A) with CFSE and coinjected them i.t. into neu-tropenic wild-type mice with 6.0 � 106 conidia. Twenty-fourhours after the transfer, the lungs were analyzed for CFSE expres-sion by flow cytometry. We found that mice receiving CCR7�/�
BMDCs had significantly higher numbers of CFSE-positive cellsthan did mice receiving wild-type BMDCs (Fig. 7B). Upon histo-logical analysis, at 48 h, recipients of wild-type and CCR7�/�
BMDCs both showed a decrease in fungal growth when comparedwith wild-type mice or chimeras without an adoptive transfer ofBMDCs (Figs. 7, C and D, 2Bv, and 3G). Although fungal burdenwas diminished in both BMDC adoptive transfer groups comparedwith wild-type mice not receiving BMDCs, the histology indicatesthat mice receiving wild-type BMDCs had more fungal growthoverall, especially in the air spaces. Additionally, protein datashowed a similar trend to the wild-type and CCR7�/� chimerasinfected with aspergillus (Fig. 7E). TNF-� and CCL5 were statis-tically higher in the mice receiving wild-type BMDCs than in micereceiving CCR7�/� BMDCs. Since the adoptive transfer ofBMDCs significantly decreased the fungal burden in both groupsof mice, we increased the conidia inocula from 6.0 � 106 to107spores to examine differences in lung chitin levels (Fig. 7F). At107 conidia, mice in both groups were susceptible to fungalgrowth, but mice receiving wild-type BMDCs had an increase in
fungal burden. Collectively, these data indicate that CCR7�/�
BMDCs provided better protection from invasive aspergillosiscompared with wild-type BMDCs.
DiscussionInvasive aspergillosis represents a significant challenge in theclinic and remains a leading cause of death for immunocom-promised individuals. While the highest incidence of disease isobserved among bone marrow transplant recipients, invasiveaspergillosis is also on the rise in solid organ transplant patientsas well as in cancer and AIDS patients (4 – 6, 38 – 40). Thepresent study investigated the role of the chemokine receptorCCR7 in a well-established neutropenic murine model of inva-sive aspergillosis. We found that CCR7�/� mice and CCR7�/�
chimeric mice had a distinct survival advantage, as well as de-creased fungal burden in the lungs, after challenge with A. fu-migatus conidia when compared with their wild-type counter-parts. This was an unexpected observation given that we founda significant up-regulation of CCR7 on DCs, in addition to in-creased levels of CCL19, in wild-type mice challenged with A.fumigatus conidia, which suggested the importance of this che-mokine-chemokine receptor interaction during fungal patho-genesis. Our data thus suggest that CCR7 induction on DCsduring invasive pulmonary aspergillosis promotes a pathologi-cal response to A. fumigatus.
FIGURE 7. Adoptive transfer of wild-type and CCR7�/� BMDCs into neutropenic C57BL/6 mice. A, Percentage of CD11c� BMDCs used in theadoptive transfer, following in vitro culture and positive selection using magnetic beads. Black histogram indicates unstained, the gray histogram indicateswild-type BMDCs, and the solid black line indicates CCR7�/� BMDCs. B, BMDCs were labeled with CFSE and coinjected with 6.0 � 106 conidia. Datarepresent the numbers of labeled DCs present in the lungs at 24 h after injection of conidia. C and D, Lung histology 48 h after coinjection of BMDCsand conidia. Wild-type mice were given an adoptive transfer of 1.0 � 106 wild-type (C) or CCR7�/� (D) BMDCs 6 h before 6.0 � 106 conidia. Blackarrows indicate fungal growth. E, Whole lung protein levels from the mice in C and D. F, Lung chitin levels 48 h following coinjection of 1.0 � 106 BMDCsand 107 conidia. WT indicates an adoptive transfer of wild-type DCs into a wild-type mouse, and CCR7�/� indicates CCR7�/� DCs transferred into awild-type mouse. �, p � 0.05 between wild-type and CCR7�/� groups.
5177The Journal of Immunology
by guest on March 3, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

DCs are an important effector cell during invasive aspergillosis.The Mehrad group showed that CCR6-deficient mice were signif-icantly more susceptible to invasive aspergillosis than were CCR6-sufficient mice (15). CCR6 and its ligand, CCL20, are responsiblefor the migration of immature DCs to sights of inflammation, in-cluding the lung (16, 17, 41, 42). Consequently, DCs lacking theexpression of CCR6 were unable to efficiently traffic into the in-fected lung, thus leaving a neutropenic host susceptible to invasiveaspergillosis. As originally reported by Dieu et al. in human DCs,when DCs encounter a maturing stimulus, such as A. fumigatus,they down-regulate their expression of CCR6 and up-regulate theirexpression of CCR7 (43). DC up-regulation of CCR7 expressionpermits trafficking away from sites of infection and inflammationto secondary lymphoid organs, as the cells migrate in response tothe chemokines CCL19 and CCL21 (20, 23–25).
Based on these observations, we hypothesized that CCR7�/�
DCs, with functional CCR6 expression, would be able to effi-ciently traffic to the lungs of infected mice; once there, however,the DCs would accumulate as they would be unable to respond tothe ligands, CCL19 and CCL21, and effectively traffic out of thelung to the draining lymph node. Indeed, our results showed that48 h after infection with A. fumigatus conidia, there was a signif-icant increase in the number of DCs in the CCR7�/� chimeraswhen compared with wild-type chimeric mice. We had similarfindings in an adoptive transfer experiment where CCR7�/� orwild-type BMDCs are injected along with conidia into the lungs ofwild-type neutropenic mice. Here we saw many-fold higher num-bers of CCR7�/� BMDCs than wild-type BMDCs in the lungs ofthe neutropenic mice. These data support the hypothesis thatCCR7�/� DCs remain in the lungs of infected animals, where theyultimately provide protection against invasive aspergillosis.
New evidence suggests that an improperly regulated inflamma-tory response in the lung during invasive aspergillosis is detrimen-tal to the survival of the host (33, 34). When TNF-�, an importantantifungal cytokine, was neutralized in a murine model of invasiveaspergillosis, there was a significant increase in mortality associatedwith increased fungal burden (44). Consequently, significant levelsof TNF-� are required for the clearance of Aspergillus, but dan-gerously high levels of TNF-� are implicated in severe inflamma-tory conditions such as sepsis (44–47). Thus, a balance betweenpro- and antiinflammatory signals is required for appropriate fun-gal clearance, without excessive tissue injury. In our model, wefound significantly higher production of TNF-� as well as theproinflammatory mediators CCL2, CCL5, and CCL3 in wild-typechimeras. Conversely, CCR7�/� chimeric mice produced signifi-cantly higher CXCL10, a chemokine recently shown to render thehost susceptible to invasive aspergillosis if inappropriately ex-pressed (31). Additionally, both wild-type and CCR7�/� chimerasshowed similar expression of the potent antiinflammatory cytokineIL-10 after conidia challenge. Taken together, our results suggestthat the cytokine milieu demonstrated by CCR7�/� chimeras maycreate a more adequate balance between pro- and antiinflammatorycytokines, which is required for appropriate fungal clearance. Al-ternatively, the CCR7�/� chimeras may simply have less inflam-mation, as these animals showed significantly less fungal growth intheir lungs. The resulting decrease in fungal burden eliminated theneed for a prolonged acute inflammatory response, ultimately lead-ing to lower cytokine levels in the CCR7�/� mice.
While the accumulation of DCs in the lungs of CCR7�/� chi-meras is likely playing an important role in the clearance of thefungus, it is possible that differences in DC phenotype or functionalso play a pivotal role during invasive aspergillosis. Previous re-ports have shown that human DCs exposed to A. fumigatus conidiain vitro up-regulate their expression of costimulatory molecules
and their production of proinflammatory cytokines (18, 19). In thepresent study, we observed a trend indicating greater expression ofthe costimulatory molecules MHC II and CD86 on CCR7�/�
BMDCs following conidia challenge. Additionally, CCR7�/�
BMDCs had statistically higher gene expression of TNF-�,CXCL10, CCL2, and CXCL2 compared with wild-type BMDCs.Due to the importance of proinflammatory cytokines in the anti-fungal response, the protective phenotype provided by CCR7�/�
DCs might derive not only from their accumulation in the lung, butalso from their more robust cytokine response against A. fumiga-tus. A complication of these in vitro experiments is the 2 h timepoint, chosen to prevent the use of antifungal reagents, which havebeen shown to mature BMDCs in vitro (18). When amphotericin Bwas used in the MLR, we observed maturation of our media controlBMDCs, leading to significant T cell proliferation. Importantly, T cellexpansion was always greater when CCR7�/� BMDCs were used asT cell stimulators, regardless of the maturing stimulus used. Col-lectively, these data indicate that CCR7�/� BMDCs have a moreactivated and mature phenotype than do wild-type BMDCs duringchallenge with A. fumigatus conidia, which may augment the pro-tective phenotype seen in vivo.
In summary, invasive aspergillosis is a severe and often fataldisease primarily affecting immunocompromised individuals. Newtherapies are required for the effective prevention and treatment ofthis opportunistic fungal infection. The present study demonstratesa deleterious role for CCR7 in a murine model of experimentalinvasive aspergillosis, indicating that selective targeting of CCR7might provide a new course of therapies for invasive fungal dis-eases. More study is required to fully understand the beneficialmechanism of CCR7 deficiency during invasive aspergillosis, andto determine whether this receptor is clinically relevant to humandisease.
AcknowledgmentsWe thank Ms. Holly Evanoff and Ms. Lisa Riggs for their outstandingtechnical assistance.
DisclosuresThe authors have no financial conflict of interest.
References1. Mullins, J., R. Harvey, and A. Seaton. 1976. Sources and incidence of airborne
Aspergillus fumigatus (Fres). Clin. Allergy 6: 209–217.2. Thompson, G. R., 3rd, and T. F. Patterson. 2008. Pulmonary aspergillosis. Semin.
Respir. Crit. Care Med. 29: 103–110.3. Soubani, A. O., and P. H. Chandrasekar. 2002. The clinical spectrum of pulmo-
nary aspergillosis. Chest 121: 1988–1999.4. Garcia-Vidal, C., A. Upton, K. A. Kirby, and K. A. Marr. 2008. Epidemiology of
invasive mold infections in allogeneic stem cell transplant recipients: biologicalrisk factors for infection according to time after transplantation. Clin. Infect. Dis.47: 1041–1050.
5. Jantunen, E., V. J. Anttila, and T. Ruutu. 2002. Aspergillus infections in alloge-neic stem cell transplant recipients: have we made any progress? Bone MarrowTransplant. 30: 925–929.
6. Post, M. J., C. Lass-Floerl, G. Gastl, and D. Nachbaur. 2007. Invasive fungalinfections in allogeneic and autologous stem cell transplant recipients: a single-center study of 166 transplanted patients. Transpl. Infect. Dis. 9: 189–195.
7. Romani, L. 2004. Immunity to fungal infections. Nat. Rev. Immunol. 4: 1–23.8. Gerson, S. L., G. H. Talbot, S. Hurwitz, B. L. Strom, E. J. Lusk, and
P. A. Cassileth. 1984. Prolonged granulocytopenia: the major risk factor for in-vasive pulmonary aspergillosis in patients with acute leukemia. Ann. Intern. Med.100: 345–351.
9. Bozza, S., K. Perruccio, C. Montagnoli, R. Gaziano, S. Bellocchio, E. Burchielli,G. Nkwanyuo, L. Pitzurra, A. Velardi, and L. Romani. 2003. A dendritic cellvaccine against invasive aspergillosis in allogeneic hematopoietic transplantation.Blood 102: 3807–3814.
10. Morrison, B. E., S. J. Park, J. M. Mooney, and B. Mehrad. 2003. Chemokine-mediated recruitment of NK cells is a critical host defense mechanism in invasiveaspergillosis. J. Clin. Invest. 112: 1862–1870.
11. Rivera, A., G. Ro, H. L. Van Epps, T. Simpson, I. Leiner, D. B. Sant’Angelo, andE. G. Pamer. 2006. Innate immune activation and CD4� T cell priming duringrespiratory fungal infection. Immunity 25: 665–675.
5178 ROLE OF CCR7 DURING ASPERGILLOSIS
by guest on March 3, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

12. Bozza, S., R. Gaziano, A. Spreca, A. Bacci, C. Montagnoli, P. di Francesco, andL. Romani. 2002. Dendritic cells transport conidia and hyphae of Aspergillusfumigatus from the airways to the draining lymph nodes and initiate disparate Thresponses to the fungus. J. Immunol. 168: 1362–1371.
13. Carpenter, K. J., and C. M. Hogaboam. 2005. Immunosuppressive effects ofCCL17 on pulmonary antifungal responses during pulmonary invasive aspergil-losis. Infect. Immun. 73: 7198–7207.
14. Mehrad, B., M. Wiekowski, B. E. Morrison, S. C. Chen, E. C. Coronel,D. J. Manfra, and S. A. Lira. 2002. Transient lung-specific expression of thechemokine KC improves outcome in invasive aspergillosis. Am. J. Respir. Crit.Care Med. 166: 1263–1268.
15. Phadke, A. P., G. Akangire, S. J. Park, S. A. Lira, and B. Mehrad. 2007. The roleof CC chemokine receptor 6 in host defense in a model of invasive pulmonaryaspergillosis. Am. J. Respir. Crit. Care Med. 175: 1165–1172.
16. Greaves, D. R., W. Wang, D. J. Dairaghi, M. C. Dieu, B. Saint-Vis,K. Franz-Bacon, D. Rossi, C. Caux, T. McClanahan, S. Gordon, et al. 1997.CCR6, a CC chemokine receptor that interacts with macrophage inflammatoryprotein 3� and is highly expressed in human dendritic cells. J. Exp. Med. 186:837–844.
17. Le Borgne, M., N. Etchart, A. Goubier, S. A. Lira, J. C. Sirard, N. van Rooijen,C. Caux, S. Ait-Yahia, A. Vicari, D. Kaiserlian, and B. Dubois. 2006. Dendriticcells rapidly recruited into epithelial tissues via CCR6/CCL20 are responsible forCD8� T cell crosspriming in vivo. Immunity 24: 191–201.
18. Gafa, V., R. Lande, M. C. Gagliardi, M. Severa, E. Giacomini, M. E. Remoli,R. Nisini, C. Ramoni, P. Di Francesco, D. Aldebert, et al. 2006. Human dendriticcells following Aspergillus fumigatus infection express the CCR7 receptor and adifferential pattern of interleukin-12 (IL-12), IL-23, and IL-27 cytokines, whichlead to a Th1 response. Infect. Immun. 74: 1480–1489.
19. Gafa, V., M. E. Remoli, E. Giacomini, M. C. Gagliardi, R. Lande, M. Severa,R. Grillot, and E. M. Coccia. 2007. In vitro infection of human dendritic cells byAspergillus fumigatus conidia triggers the secretion of chemokines for neutrophiland Th1 lymphocyte recruitment. Microbes Infect. 9: 971–980.
20. Saeki, H., A. M. Moore, M. J. Brown, and S. T. Hwang. 1999. Cutting edge:secondary lymphoid-tissue chemokine (SLC) and CC chemokine receptor 7(CCR7) participate in the emigration pathway of mature dendritic cells from theskin to regional lymph nodes. J. Immunol. 162: 2472–2475.
21. Sallusto, F., D. Lenig, R. Forster, M. Lipp, and A. Lanzavecchia. 1999. Twosubsets of memory T lymphocytes with distinct homing potentials and effectorfunctions. Nature 401: 708–712.
22. Sanchez-Sanchez, N., L. Riol-Blanco, G. de la Rosa, A. Puig-Kroger,J. Garcia-Bordas, D. Martin, N. Longo, A. Cuadrado, C. Cabanas, A. L. Corbi,et al. 2004. Chemokine receptor CCR7 induces intracellular signaling that inhib-its apoptosis of mature dendritic cells. Blood 104: 619–625.
23. Gunn, M. D., K. Tangemann, C. Tam, J. G. Cyster, S. D. Rosen, andL. T. Williams. 1998. A chemokine expressed in lymphoid high endothelialvenules promotes the adhesion and chemotaxis of naive T lymphocytes. Proc.Natl. Acad. Sci. USA 95: 258–263.
24. Randolph, G. J., V. Angeli, and M. A. Swartz. 2005. Dendritic-cell trafficking tolymph nodes through lymphatic vessels. Nat. Rev. Immunol. 5: 617–628.
25. Forster, R., A. C. Davalos-Misslitz, and A. Rot. 2008. CCR7 and its ligands:balancing immunity and tolerance. Nat. Rev. Immunol. 8: 362–371.
26. Banchereau, J., and R. M. Steinman. 1998. Dendritic cells and the control ofimmunity. Nature 392: 245–252.
27. Forster, R., A. Schubel, D. Breitfeld, E. Kremmer, I. Renner-Muller, E. Wolf, andM. Lipp. 1999. CCR7 coordinates the primary immune response by establishingfunctional microenvironments in secondary lymphoid organs. Cell 99: 23–33.
28. Mehrad, B., R. M. Strieter, T. A. Moore, W. C. Tsai, S. A. Lira, andT. J. Standiford. 1999. CXC chemokine receptor-2 ligands are necessary com-ponents of neutrophil-mediated host defense in invasive pulmonary aspergillosis.J. Immunol. 163: 6086–6094.
29. Chen, S. C., G. Vassileva, D. Kinsley, S. Holzmann, D. Manfra,M. T. Wiekowski, N. Romani, and S. A. Lira. 2002. Ectopic expression of themurine chemokines CCL21a and CCL21b induces the formation of lymph node-like structures in pancreas, but not skin, of transgenic mice. J. Immunol. 168:1001–1008.
30. Vermaelen, K., and R. Pauwels. 2004. Accurate and simple discrimination ofmouse pulmonary dendritic cell and macrophage populations by flow cytometry:methodology and new insights. Cytometry A 61: 170–177.
31. Mezger, M., M. Steffens, M. Beyer, C. Manger, J. Eberle, M. R. Toliat,T. F. Wienker, P. Ljungman, H. Hebart, H. J. Dornbusch, H. Einsele, andJ. Loeffler. 2008. Polymorphisms in the chemokine (C-X-C motif) ligand 10 areassociated with invasive aspergillosis after allogeneic stem-cell transplantationand influence CXCL10 expression in monocyte-derived dendritic cells. Blood111: 534–536.
32. Davalos-Misslitz, A. C., J. Rieckenberg, S. Willenzon, T. Worbs, E. Kremmer,G. Bernhardt, and R. Forster. 2007. Generalized multi-organ autoimmunity inCCR7-deficient mice. Eur. J. Immunol. 37: 613–622.
33. Romani, L., F. Bistoni, K. Perruccio, C. Montagnoli, R. Gaziano, S. Bozza,P. Bonifazi, G. Bistoni, G. Rasi, A. Velardi, F. Fallarino, E. Garaci, andP. Puccetti. 2006. Thymosin �1 activates dendritic cell tryptophan catabolism andestablishes a regulatory environment for balance of inflammation and tolerance.Blood 108: 2265–2274.
34. Zelante, T., A. De Luca, P. Bonifazi, C. Montagnoli, S. Bozza, S. Moretti,M. L. Belladonna, C. Vacca, C. Conte, P. Mosci, et al. 2007. IL-23 and the Th17pathway promote inflammation and impair antifungal immune resistance. Eur.J. Immunol. 37: 2695–2706.
35. Mambula, S. S., K. Sau, P. Henneke, D. T. Golenbock, and S. M. Levitz. 2002.Toll-like receptor (TLR) signaling in response to Aspergillus fumigatus. J. Biol.Chem. 277: 39320–39326.
36. Meier, A., C. J. Kirschning, T. Nikolaus, H. Wagner, J. Heesemann, and F. Ebel.2003. Toll-like receptor (TLR) 2 and TLR4 are essential for Aspergillus-inducedactivation of murine macrophages. Cell. Microbiol. 5: 561–570.
37. Steele, C., R. R. Rapaka, A. Metz, S. M. Pop, D. L. Williams, S. Gordon,J. K. Kolls, and G. D. Brown. 2005. The �-glucan receptor dectin-1 recognizesspecific morphologies of Aspergillus fumigatus. PLoS Pathog. 1: e42.
38. Ali, R., F. Ozkalemkas, T. Ozcelik, V. Ozkocaman, A. Ozkan, S. Bayram,B. Ener, A. Ursavas, G. Ozal, and A. Tunali. 2006. Invasive pulmonary aspergil-losis: role of early diagnosis and surgical treatment in patients with acute leuke-mia. Ann. Clin. Microbiol. Antimicrob. 5: 17.
39. Morgan, J., K. A. Wannemuehler, K. A. Marr, S. Hadley, D. P. Kontoyiannis,T. J. Walsh, S. K. Fridkin, P. G. Pappas, and D. W. Warnock. 2005. Incidence ofinvasive aspergillosis following hematopoietic stem cell and solid organ trans-plantation: interim results of a prospective multicenter surveillance program.Med. Mycol. 43(Suppl. 1): S49–S58.
40. Mylonakis, E., T. F. Barlam, T. Flanigan, and J. D. Rich. 1998. Pulmonary as-pergillosis and invasive disease in AIDS: review of 342 cases. Chest 114:251–262.
41. Dieu-Nosjean, M. C., C. Massacrier, B. Homey, B. Vanbervliet, J. J. Pin,A. Vicari, S. Lebecque, C. Dezutter-Dambuyant, D. Schmitt, A. Zlotnik, andC. Caux. 2000. Macrophage inflammatory protein 3� is expressed at inflamedepithelial surfaces and is the most potent chemokine known in attracting Lang-erhans cell precursors. J. Exp. Med. 192: 705–718.
42. Osterholzer, J. J., T. Ames, T. Polak, J. Sonstein, B. B. Moore, S. W. Chensue,G. B. Toews, and J. L. Curtis. 2005. CCR2 and CCR6, but not endothelial se-lectins, mediate the accumulation of immature dendritic cells within the lungs ofmice in response to particulate antigen. J. Immunol. 175: 874–883.
43. Dieu, M. C., B. Vanbervliet, A. Vicari, J. M. Bridon, E. Oldham, S. Ait-Yahia,F. Briere, A. Zlotnik, S. Lebecque, and C. Caux. 1998. Selective recruitment ofimmature and mature dendritic cells by distinct chemokines expressed in differentanatomic sites. J. Exp. Med. 188: 373–386.
44. Mehrad, B., R. M. Strieter, and T. J. Standiford. 1999. Role of TNF-� in pul-monary host defense in murine invasive aspergillosis. J. Immunol. 162:1633–1640.
45. Bone, R. C. 1991. The pathogenesis of sepsis. Ann. Intern. Med. 115: 457–469.46. Michie, H. R., K. R. Manogue, D. R. Spriggs, A. Revhaug, S. O’Dwyer,
C. A. Dinarello, A. Cerami, S. M. Wolff, and D. W. Wilmore. 1988. Detection ofcirculating tumor necrosis factor after endotoxin administration. N. Engl. J. Med.318: 1481–1486.
47. Nagai, H., J. Guo, H. Choi, and V. Kurup. 1995. Interferon-� and tumor necrosisfactor-alpha protect mice from invasive aspergillosis. J. Infect. Dis. 172:1554–1560.
5179The Journal of Immunology
by guest on March 3, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from