catalytic properties of the lactate dehydrogenase isozyme “x” from mouse testis
TRANSCRIPT

Catalytic Properties of the Lactate Dehydrogenase Isozyme "X" from Mouse Testis
LUIS J. BATTELLINO AND ANTONIO BLANCO Cdtedra de Quimica Bial6gica, Facultades de Medicina y Odontologia, Universidad Nacional de Cdrdoba, Cdrdoba, Argentina
ABSTRACT Lactate dehydrogenase (LDH) isozyme 1 (B4), 5 (A4), and " X (Testis or Sperm type) have been partially purified from mouse tissues.
The following studies were carried out on the three isozymes: K, and optimum substrate concentration for pyruvate, a-0x0-butyrate, a-0x0-valerate, lactate, a-OH- butyrate, and a-OH-valerate, inhibition by substrate and product, effect of malate, N-( 4-carboxy-2-hydroxyphenyl) maleimide, some citric acid cycle metabolites, urea, trypsin and pH.
The mouse testicular LDH isozyme clearly differs from the common lactate dehy- drogenases and from the testicular isozymes from other species. It shows distinct sensitivity to inhibition by substrate or product whether the direct (pyruvate+ lac- tate) or reverse reactions are studied. There is no effect of increasing concentrations of pyruvate or lactate on the direct reaction, while a clear inhibition by lactate or pyruvate is demonstrated on the reverse reaction. Citric acid cycle metabolites, spe- cially malate and succinate, inhibit the direct reaction catalyzed by the " X isozyme.
These peculiar characteristics suggest a high degree of specialization for the prin- cipal testicular isozyme of lactate dehydrogenase.
The appearance of an additional lactate dehydrogenase (L-lactate : NAD oxidore- ductase, EC 1.1.1.27) isozyme in testis is a phenomenon observed in many species (Zinkham, Blanco and Clowry, '64). This testicular isozyme, originally designated LDH-X, differs in catalytic properties, mo- lecular structure, and genetic control from the other five lactate dehydrogenase frac- tions present in most tissues (Blanco and Zinkham, '63; Blanco, Zinkham and Kupchyk, '64).
Available evidence indicates that LDH-X is a tetramer composed of polypeptide units different from the A and B monomers comprising the five common is0zymes.l The synthesis of the additional polypeptide is activated in testis during sexual matu- ration. In the species in which sperm isozymic complement was studied, LDH-X appeared as the predominant fraction (Zinkham et al., '64; Goldberg, '65).
In an attempt to elucidate the biological significance of LDH-X we undertook an investigation of catalytic properties of iso- zymes from different species. A previous report on rabbit testicular lactate dehy- drogenase indicated that it is a unique isozyme and that its kinetic properties
J. EXP. ZOOL., 174: 173-186.
correlate with metabolic requirements of spermatozoa (Battellino, Ramos and Blan- co, '68).
Investigations of reactivity with several substrate and coenzyme analogues showed differences among testicular isozymes from diverse species (Zinkham et al., '64). These findings reflect structural differences and probably distinct functional abilities among LDH-Xs from different animals.
Mouse " X isozyme was one of the lac- tate dehydrogenases showing peculiar be- havior. For that reason, a more detailed study of its kinetic properties seemed justi- fied. This paper presents such a study.
MATERIAL AND METHODS
DL-lactic acid, a-0x0-butyrate, a-0x0- valerate, DL-a-OH-butyrate, DL-a-OH-val- erate, NADH, and NAD were purchased from Sigma (U. S. A.); sodium pyruvate, potassium oxalate, DL-malate, succinate,
1 LDH: Lactate dehydrogenase. The nomenclature originally proposed by Markert ('62) is used. The five common isozymes are tetramers formed by as- sociations of A and B chains: LDH-1: Bq, LDH-2: ArB3, LDH-3: AeB2, LDH-4: A,BI and LDH-5: A+ Cahn et al. ('62) designated the A chams by M (Muscle type) and the B chains by H (Heart type).
LDH-X is the additional isozyme in testis and its polypeptide units have been designated C (Zinkham, Blanco and Kupchyk, '63).
173

174 LUIS J. BATTELLINO AND ANTONIO BLANCO
citrate, L-glutamate, L-aspartate, urea, and trypsin, from Merck (Germany); hydro- lyzed starch for electrophoresis, from Connaught (Canada) ; N-( 4-carboxy-2-hy- droxyphenyl) maleimide was synthesized and kindly supplied by Dr. G. Carvajal (I. P. N., Mexico).
Swiss albino mice were killed by de- capitation, their tissues removed immedi- ately and processed at once or kept frozen at - 20°C for a few days until utilized. Results with fresh or frozen tissues were the same. Mouse LDH-1 (B4) was obtained from heart; LDH-5 (A4), from liver; and LDH-X, from pooled testes. Mouse sperm was collected by squeezing and washing excised epididymes with 0.9% NaC1. Mi- croscopic examination of this material revealed that it was composed almost ex- clusively of spermatozoa.
Homogenates and sperm lysates were prepared as previously described (Battel- lino et al., '68).
Electrophoresis. Electrophoresis was performed using the vertical technique proposed by Smithies ('59). Gels were usually made with 12 g of starch per 100 ml of 0.03 M borate buffer pH 8.6. Another buffer system frequently used was the Tris-borate-EDTA (Boyer, Fainer and Naughton, '63). A voltage gradient of 6 Volt/cm was applied during 14 hours at 4" C. After electrophoresis, the starch block was sliced and stained for lactate dehydrogenase with the same method used by Zinkham et al. ('64).
Eluates were obtained by the procedure previously outlined (Battellino et al., '68). A 100-fold purification was attained by that method.
Lactate dehydrogenase assay. The methods of Wrdblewski and LaDue ('55), and of Markert and Ursprung ('62) were used for the direct and reverse reactions respectively. Optical density change at 340 mp was read every minute during s i x minutes in a Beckman model DU spectro- photometer. Samples were diluted to ob- tain a AEa40 of 0.030 to 0.040 per minute under conditions assuring maximal ac- tivity. Assays were carried out at 20" C. One Unit of enzyme is the amount pro- ducing the conversion of 1 pM of NAD per minute.
Elution.
RESULTS
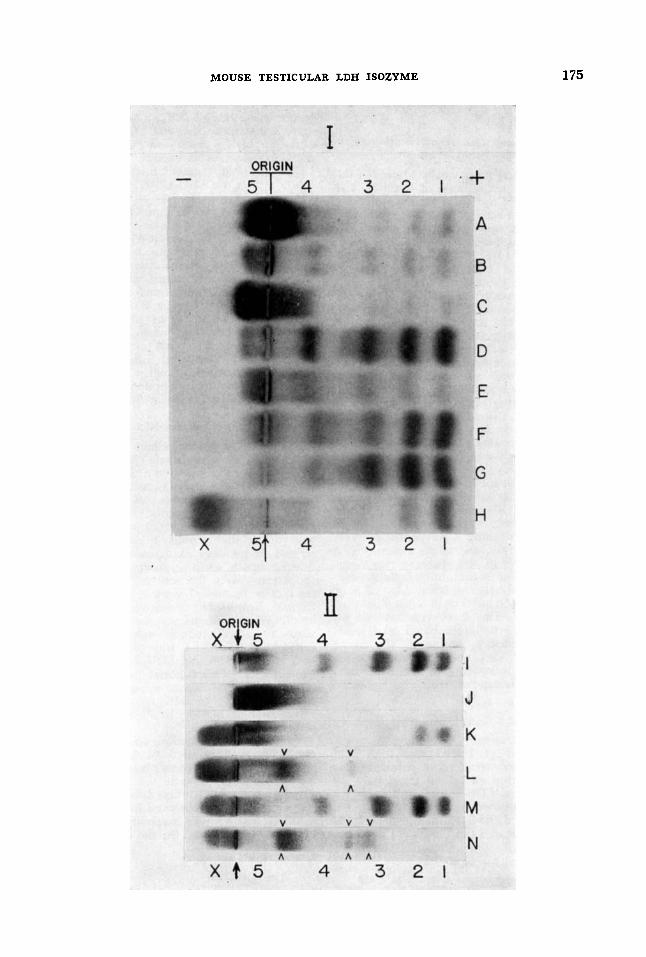
Electrophoretic patterns Figure 1 ,I shows lactate dehydrogenase
isozymic patterns of several mouse tissue extracts as revealed by starch gel electro- phoresis and histochemical staining. Ho- mogenates from heart, diaphragm, skeletal muscle, tongue, pharynx, esophagus, stom- ach, duodenum, large intestine, liver, kidney, lung, spleen and immature testis and epididymis, exhibited patterns similar to those previously described by other authors (Markert and Ursprung, '62). Relative distribution of activity among the five isozymes LDH-1 to LDH-5 was char- acteristic for each tissue.
Mature testis and epididymis presented an additional lactate dehydrogenase frac- tion not demonstrated in any other tissue. This fraction migrated nearer the cathode than did LDH-5. Most of the activity in patterns of adult testis was located in LDH-1 and LDH-X areas, while LDH-3, -4, and -5 were barely visible.
Appearance of LDH-X was detected in testis at the seventeenth day of life, in close agreement with previous ontogenetic studies by Goldberg and Hawtrey ('67). The relative amount of the sixth isozyme increased steadily to reach maximum values in 120 day old mice. No significant variations in the proportion of LDH-X were observed after that age. It accounted for about 40% of total lactate dehydro- genase activity in fully developed testis.
Zymograms of sperm lysates exhibited most of the activity on the site of LDH-X,
Fig. 1 ,I Electrophoretic patterns of lactate dehydrogenase from mouse tissue homogenates. Numbers and X indicate the position of corres- ponding isozymes. A, skeletal muscle (1:6, tissue to water, wt/vol); B, ovary (1:4); C, liver (1:6); D, kidney (1:6) ; E, uterus ( 1 : 4 ) ; F, diaphragm (1:s); G, heart ( 1 : 7 ) ; H, mature testis (1:4). All homogenates were subjected to electrophore- sis simultaneously in the same starch block. Gel prepared with 0.03 M borate buffer pH 8.6.
Fig. 1,II I, heart (1:7); J, liver (1:6); K, mature testis (1:4) . 0.5 M NaCl and 0.1 M phos- phate buffer pH 7.0 was added to all control homogenates. L, mature testis, same as K, after freezing and thawing twice; M, mixture of testis and heart homogenates ( 1 : 1 ) ; N, testis-heat mixture same as M after freezing and thawing twice. All homogenates were subjected to electro- phoresis simultaneously in the same starch block. Gel was prepared with Tris-borate-EDTA buffer.
MOUSE TESTICULAR LDH ISOZYME 175

176 LUIS J. BATTELLINO AND ANTONIO BLANCO
with very faint staining of LDH-1 and -2 bands.
Dissociation and recombination of polypeptide units
The method proposed by Markert ('63) was used. After addition of 4 mg of NAD per ml, whole testicular homogenates and mixtures of heart and testis extracts were subjected to two freeze-thaw cycles in a medium containing 0.5 M NaCl and 0.1 M phosphate buffer pH 7.0. After electro- phoresis and staining, the recombined preparations showed a decrease in LDH-1, -2, and sometimes In LDH-X. Simultane- ously, new bands appeared: one migrating between LDH-3 and LDH-4, and another, more intensely stained, with a mobility intermediate between LDH-4 and LDH-5. In mixtures of heart and testis homoge- nates, a third additional fraction was formed. It was visible as a faint and broad band migrating closely behind the site of LDH-3 (fig. 1,II). Experiments of recom- bination with heart-liver mixtures and with homogenates of tissues not contain- ing LDH-X showed only redistribution of activity among the five common isozymes, but no additional hybrids.
In testicular extracts and in heart-testis mixtures, LDH-1 and -X are the predomi- nant fractions, and the proportion of isozymes containing A polypeptides is very low. Apparently, the small amount of A units present in the mixture was dena- tured, since no A-containing hybrids were demonstrated after recombination of poly- peptide units in testis or in testis plus heart preparations. The three fractions shown in figure 1,II for recombined testis-heart extracts probably represent the three hybrids B3C1, BX,, and B I G , although the electrophoretic spacing of the first band does not correspond with the expected migration.
Effect of substrate concentration Direct reaction. (pyruvate + lactate).
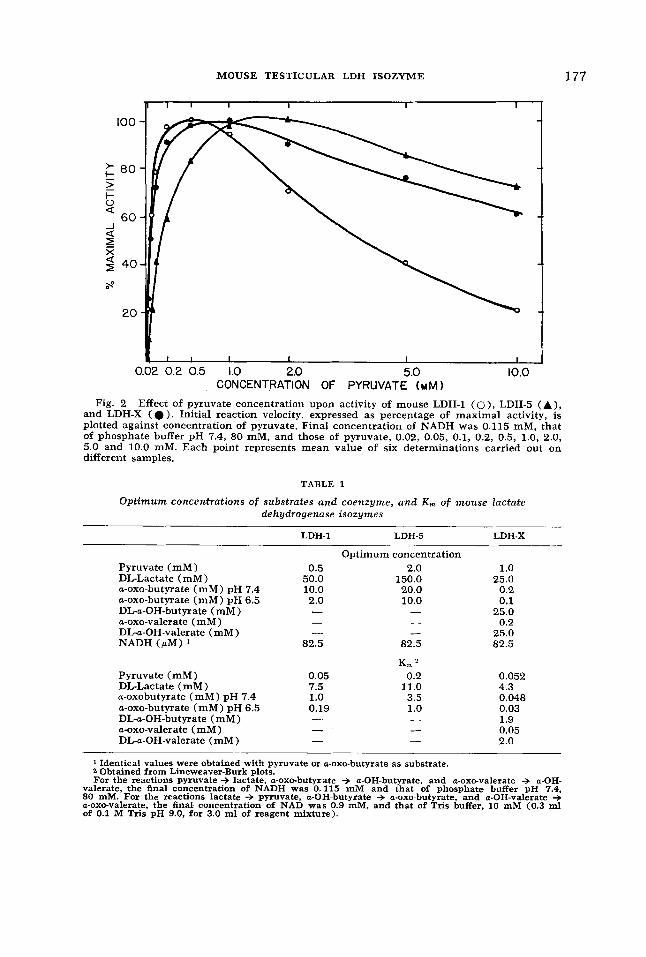
The behavior of LDH-1 and LDH-5 at increased concentrations of substrate was similar to that described previously by many authors. LDH-X had an affinity for pyruvate intermediate between those for LDH-1 and LDH-5, although its K, is closer to that of LDH-1 (table 1) . Figure
2 shows the effect of pyruvate concen- tration upon rate of activity of LDH-1, -5, and -X isozymes. The first portion of the curve for LDH-X resembled that for LDH- 1, but as substrate concentration increased, LDH-X showed greater resistance to in- hibition than did LDH-1. At 10 mM pyru- vate, LDH-5 presented 74% of its maximal activity, LDH-X, 60%, and LDH-1, 22%.
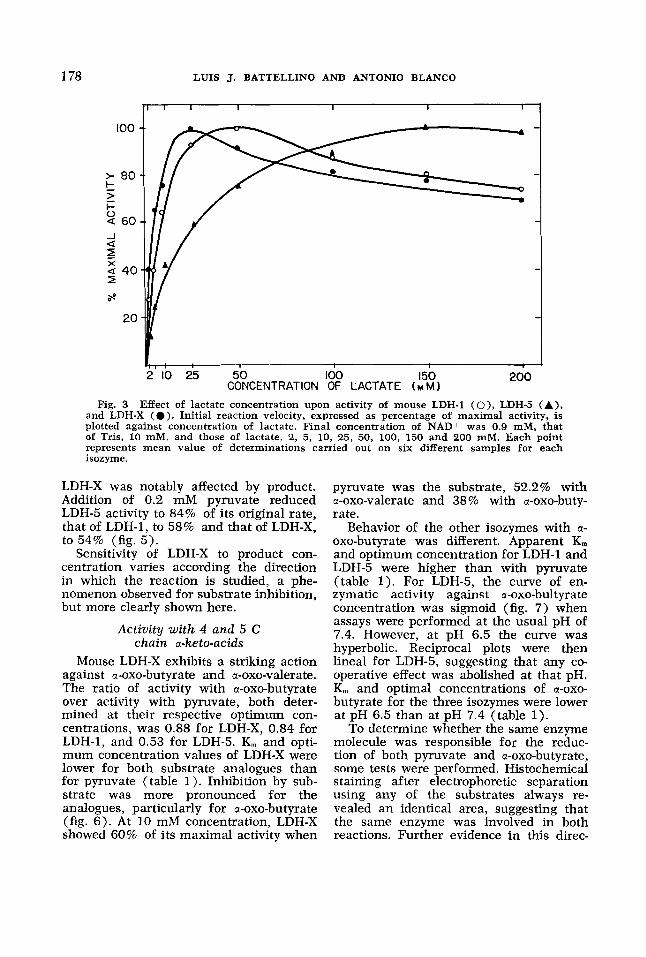
Reverse reaction (lactate + pyruvate). LDH-X showed the greatest affinity for lactate; K, and optimum concentration values for LDH-X are the lowest (table 1). At higher concentrations of substrate, LDH-X revealed inhibition, reproducing the behavior of LDH-1 (fig. 3).
Thus, the sensitivity of LDH-X to in- creased concentrations of substrate was different when the direct and reverse reactions were compared. Little effect of high concentrations of pyruvate was ob- served on the direct reaction, but there was inhibition by lactate in the reverse.
Effect of product The study of product effect was carried
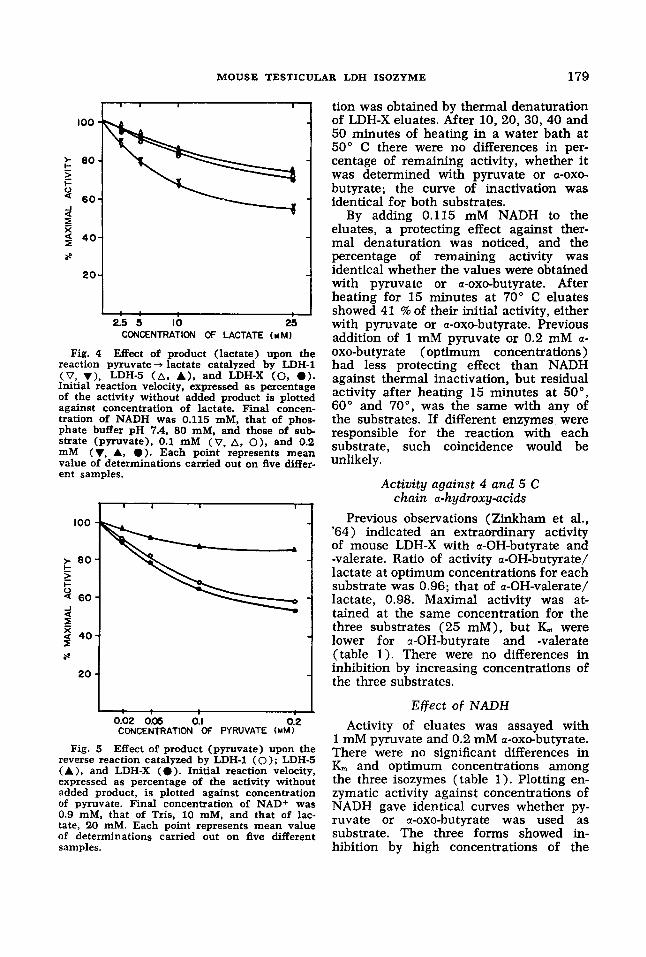
out under the conditions proposed by Stambaugh and Post ('66). The product was added to the assay mixture ten min- utes before the substrate was and incu- bated at 25" C . Substrate and product concentrations were selected within values well below the inhibitory levels demon- strated in previous experiments. For the direct reaction, concentrations of pyruvate (substrate) were 0.1 and 0.2 mM, those for lactate (product) were 2.5, 5, 10 and 25 mM. Results are presented in figure 4 by plotting activity as per cent of the rate without added product, against lactate concentration. LDH-1 was clearly inhib- ited, while LDH-5 and LDH-X were much less affected. With 0.1 mM pyruvate, ad- dition of 25 mM lactate reduced the ac- tivity of LDH-1 to 54% of the control value, that of LDH-X, to 73%, and that
For the reverse reaction, substrate (lac- tate) concentration used was 20 mM, and product (pyruvate) concentrations added were 0.02, 0.05, 0.1 and 0.2 mM. Effect of product on LDH-1 and LDH-5 activity was similar to that observed for the direct reaction: LDH-1 was markedly inhibited, while LDH-5 was not. On the other hand,
of LDH-5, to 76%.
MOUSE TESTICULAR LDH ISOZYME 177
100
* 80 k 1
a
a 2 9 40
.\"
+ 0
1 60
X
20
0.02 0.2 0.5 1.0 2.0 5.0 10.0 CONCENTRATION OF PYRUVATE (MM 1
Fig. 2 Effect of pyruvate concentration upon activity of mouse LDH-1 ( O ) , LDH-5 (A), and LDH-X ( 0 ) . Initial reaction velocity, expressed as percentage of maximal activity, is plotted against concentration of pyruvate. Final concentration of NADH was 0.115 mM, that of phosphate buffer pH 7.4, 80 mM, and those of pyruvate, 0.02, 0.05, 0.1, 0.2, 0.5, 1.0, 2.0, 5.0 and 10.0 mM. Each point represents mean value of six determinations carried out on different samples.
TABLE 1
Optimum concentrations of substrates and coenzyme, and K, of mouse lactate dehydrogenase isozymes
LDH-1 LDH-5 LDH-X
Optimum concentration Pyruvate (mM) 0.5 2.0
a-0x0-butyrate (mM) pH 7.4 10.0 20.0 a-0x0-butyrate (mM) pH 6.5 2.0 10.0 DL-a-OH-butyrate (mM) - - a-0x0-valerate (mM) - - DL-a-OH-valerate (mM) - -
DL-Lactate (mM) 50.0 150.0
NADH (pM) 82.5 82.5 K 2
Pyruvate (mM) 0.05 0.2 DL-Lactate (mM) 7.5 11.0 a-oxobutyrate (mM) pH 7.4 1.0 3.5 a-0x0-butyrate (mM) pH 6.5 0.19 1 .o DL-a-OH-butyrate (mM) - a-0x0-valerate (mM) - - DL-a-OH-valerate (mM) - -
-
1 .o 25.0
0.2 0.1
25.0 0.2
25.0 82.5
0.052 4.3 0.048 0.03 1.9 0.05 2.0
1 Identical values were obtained with pyruvate or a-0x0-butyrate as substrate. 2 Obtained from Lineweaver-Burk plots. For the reactions pyruvate + lactate, a-0x0-butyrate + a-OH-butyrate, and a-0x0-valerate + a-OH-
valerate, the final concentration of NADH was 0.115 mM and that of phosphate buffer pH 7.4, 80 mM. For the reactions lactate + pyruvate, a-OH-butyrate + a-0x0-butyrate, and a-OH-valerate + a-0x0-valerate, the final concentration of NAD was 0.9 mM, and that of Tris buffer, 10 mM (0.3 ml of 0.1 M Tris pH 9.0, for 3.0 ml of reagent mixture).
1 78
too
>- 80 k 1
a 60
a z 5 40 I $
I- 0
-I
20
LUIS J. BATTELLINO AND ANTONIO BLANCO
I , : 2 10 25 50 100 150 200
CONCENTRATION OF LACTATE ( M M 1 Fig. 3 Effect of lactate concentration upon activity of mouse LDH-1 ( O ) , LDH-5 ( A ) ,
and LDH-X ( 0 ) . Initial reaction velocity, expressed as percentage of maximal activity, is plotted against concentration of lactate. Final concentration of NAD+ was 0.9 mM, that of Tris, 10 mM, and those of lactate, 2, 5, 10, 25, 50, 100, 150 and 200 mM. Each point represents mean value of determinations carried out on six different samples for each isozyme.
LDH-X was notably affected by product. Addition of 0.2 mM pyruvate reduced LDH-5 activity to 84% of its original rate, that of LDH-1, to 58% and that of LDH-X,
Sensitivity of LDH-X to product con- centration varies according the direction in which the reaction is studied, a phe- nomenon observed for substrate inhibition, but more clearly shown here.
Activity wi th 4 and 5 C chain a-keto-acids
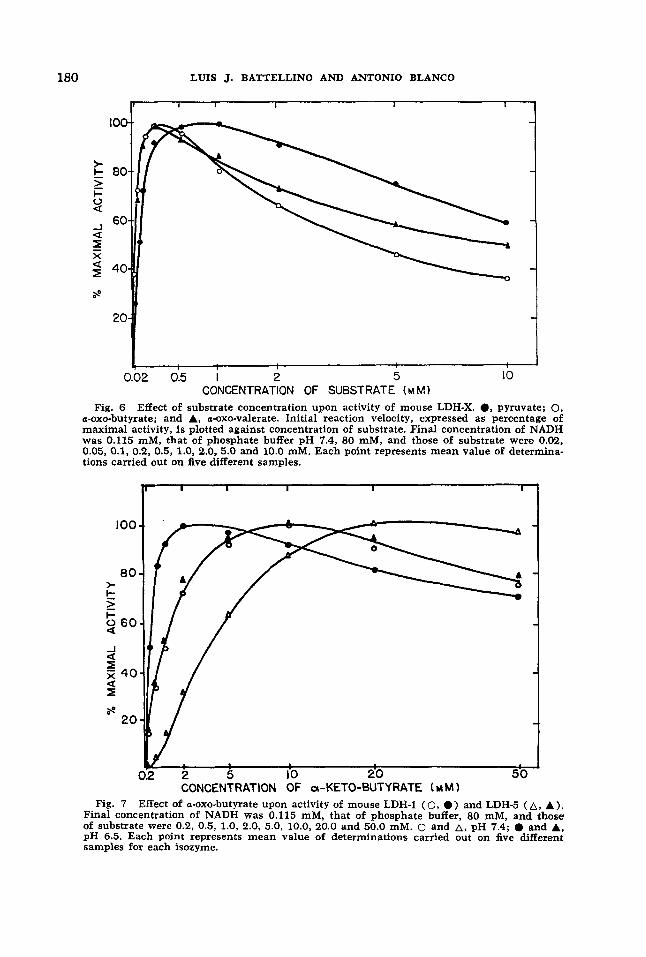
Mouse LDH-X exhibits a striking action against a-0x0-butyrate and a-0x0-valerate. The ratio of activity with a-0x0-butyrate over activity with pyruvate, both deter- mined at their respective optimum con- centrations, was 0.88 for LDH-X, 0.84 for LDH-1, and 0.53 for LDH-5. K, and opti- mum concentration values of LDH-X were lower for both substrate analogues than for pyruvate (table 1). Inhibition by sub- strate was more pronounced for the analogues, particularly for a-0x0-butyrate (fig. 6). At 10 mM concentration, LDH-X showed 60% of its maximal activity when
to 54% (fig. 5) .
pyruvate was the substrate, 52.2% with a-0x0-valerate and 38% with a-0x0-buty- rate.
Behavior of the other isozymes with a- 0x0-butyrate was different. Apparent K, and optimum concentration for LDH-1 and LDH-5 were higher than with pyruvate (table 1). For LDH-5, the curve of en- zymatic activity against a-0x0-bultyrate concentration was sigmoid (fig. 7) when assays were performed at the usual pH of 7.4. However, at pH 6.5 the curve was hyperbolic. Reciprocal plots were then lineal for LDH-5, suggesting that any co- operative effect was abolished at that pH. K, and optimal concentrations of U-OXO- butyrate for the three isozymes were lower at pH 6.5 than at pH 7.4 (table 1) .
To determine whether the same enzyme molecule was responsible for the reduc- tion of both pyruvate and a-0x0-butyrate, some tests were performed. Histochemical staining after electrophoretic separation using any of the substrates always re- vealed an identical area, suggesting that the same enzyme was involved in both reactions. Further evidence in this direc-
MOUSE TESTICULAR LDH ISOZYME 179
I , I
100 -
8 0 - - E ' 6 0 -
z a 9 40- - X
z 20 - -
I I I
z
I : 1 0.02 0.05 0.1 0.2 CONCENTRATION OF PYRUVATE (MM)
Effect of product (pyruvate) upon the reverse reaction catalyzed by LDH-1 (0 ) ; LDH-5 (A), and LDH-X (0 ) . Initial reaction velocity, expressed as percentage of the activity without added product, is plotted against concentration of pyruvate. Final concentration of NAD+ was 0.9 mM, that of Tris, 10 mM, and that of lac- tate, 20 mM. Each point represents mean value of determinations carried out on five different samples.
Fig. 5
tion was obtained by thermal denaturation of LDH-X eluates. After 10, 20, 30, 40 and 50 minutes of heating in a water bath at 50" C there were no differences in per- centage of remaining activity, whether it was determined with pyruvate or a-oxo- butyrate; the curve of inactivation was identical for both substrates.
By adding 0.115 mM NADH to the eluates, a protecting effect against ther- mal denaturation was noticed, and the percentage of remaining activity was identical whether the values were obtained with pyruvate or a-oxo-butyrate. After heating for 15 minutes at 70" C eluates showed 41 % of their initial activity, either with pyruvate or a-oxo-butyrate. Previous addition of 1 mM pyruvate or 0.2 mM a- 0x0-butyrate (optimum concentrations) had less protecting effect than NADH against thermal inactivation, but residual activity after heating 15 minutes at 50°, 60" and 70°, was the same with any of the substrates. If different enzymes were responsible for the reaction with each substrate, such coincidence would be unlikely.
Activity against 4 and 5 C chain a-hydroxy-acids
Previous observations (Zinkham et al., '64) indicated an extraordinary activity of mouse LDH-X with a-OH-butyrate and -valerate. Ratio of activity a-OH-butyrate/ lactate at optimum concentrations for each substrate was 0.96; that of a-OH-valerate/ lactate, 0.98. Maximal activity was at- tained at the same concentration for the three substrates (25 mM), but K, were lower for a-OH-butyrate and -valerate (table 1). There were no differences in inhibition by increasing concentrations of the three substrates.
ERect of NADH Activity of eluates was assayed with
1 mM pyruvate and 0.2 mM a-oxo-butyrate. There were no significant differences in K, and optimum concentrations among the three isozymes (table 1). Plotting en- zymatic activity against concentrations of NADH gave identical curves whether py- ruvate or a-0x0-butyrate was used as substrate. The three forms showed in- hibition by high concentrations of the
180 LUIS J. BATTELLINO AND ANTONIO BLANCO
r' 4u
O\" 20] l
4 0.02 0.5 I 2 5 10
CONCENTRATION OF SUBSTRATE ( M M 1 Fig. 6 Effect of substrate concentration upon activity of mouse LDH-X. 0, pyruvate; 0.
a-0x0-butyrate; and A, a-oxo-valerate. Initial reaction velocity, expressed as percentage of maximal activity, is plotted against concentration of substrate. Final concentration of NADH was 0.115 mM, that of phosphate buffer pH 7.4, 80 mM, and those of substrate were 0.02, 0.05, 0.1, 0.2, 0.5, 1.0, 2.0, 5.0 and 10.0 mM. Each point represents mean value of determina- tions carried out on five different samples.
-
-
-
CONCENTRATION OF a-KETO-BUTYRATE ( Y M 1 Fig. 7 Effect of a-0x0-butyrate upon activity of mouse LDH-1 (0 , 0 ) and LDH-5 ( A , A).
Final concentration of NADH was 0.115 mM, that of phosphate buffer, 80 mM, and those of substrate were 0.2, 0.5, 1.0, 2.0, 5.0, 10.0, 20.0 and 50.0 mM. 0 and A, pH 7.4; 0 and A, pH 6.5. Each point represents mean value of determinations carried out on five different samples for each isozyme.

MOUSE TESTICULAR LDH ISOZYME 181
coenzyme. Activity was 80% of the maxi- mal with 0.2 mM NADH.
Effect of metabolites DL-malate, succinate, citrate, L-gluta-
mate, or Caspartate, diluted in 0.2 M phos- phate buffer pH 7.4 were incorporated in the reagent mixture up to a final con- centration of 1.5 mM and incubated for ten minutes before the substrate was added. The pH deviation in the mixture was always less than 0.1 after addition of metabolites.
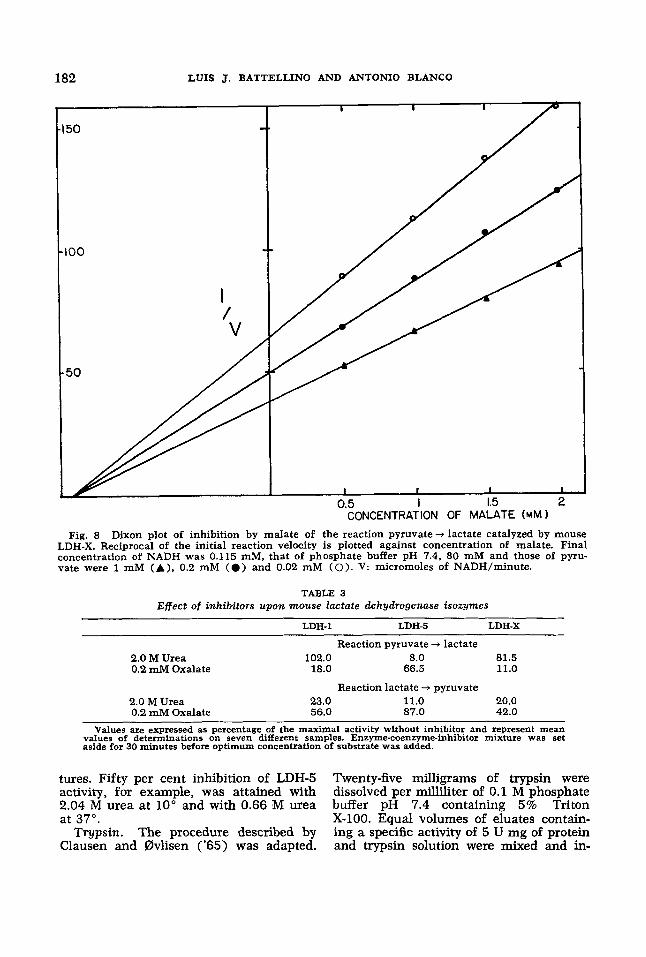
Activity of LDH-1 and LDH-5 against optimum concentrations of pyruvate was not affected by any of the metabolites. However, all compounds studied produced a significant inhibition upon activity of the testicular isozyme (table 2). Malate and succinate were the most effective in- hibitors, reducing LDH-X activity to 69.5% and 73% of the control value respectively. Inhibition is of the non-competitive type as shown by Dixon plots. Figure 8 repre- sents the effect of malate upon LDH-X.
The reverse reaction is absolutely not affected by the same metabolites. Neither is the reaction a-0x0-butyrate + a-OH-bu- tyrate catalyzed by LDH-X affected.
Effect of inhibitors Oxalate. The inhibitor was added as
indicated by Emerson and Wilkinson ('65). When the forward reaction was studied, LDH-1 and LDH-X were the most affected isozymes. Addition of 0.2 mM oxalate re- duced LDH-1 activity to 18% of that of control; that of LDH-X, to 11% (table 3). KI was 9.8 X mM for LDH-X, 7.8 X lo-' mM for LDH-1, and 0.63 mM for LDH-5. The inhibition was non-competi- tive for all the isozymes. The effect of oxalate was less intense for the three iso-
zymes when the reverse reaction was analyzed (table 3 ) .
N-( 4-carboxy-2-hydroxyphenyl) malei- mide. This compound attaches irreversi- bly to sulfhydryl groups at the active site and yields specific information about structural differences between LDH iso- zymes (Carvajal et al., '67).
The inhibitor was diluted in 0.1 M sod- ium phosphate buffer pH 7.4, incorporated in the reagent mixture and incubated 30 minutes before the substrate was added. LDH-5 was the most affected. Ki for LDH-5 was 9.2 X mM; for LDH-X, 4.4 mM; and for LDH-1, 12 mM. Dixon plots showed that inhibition is competitive for the three isozymes.
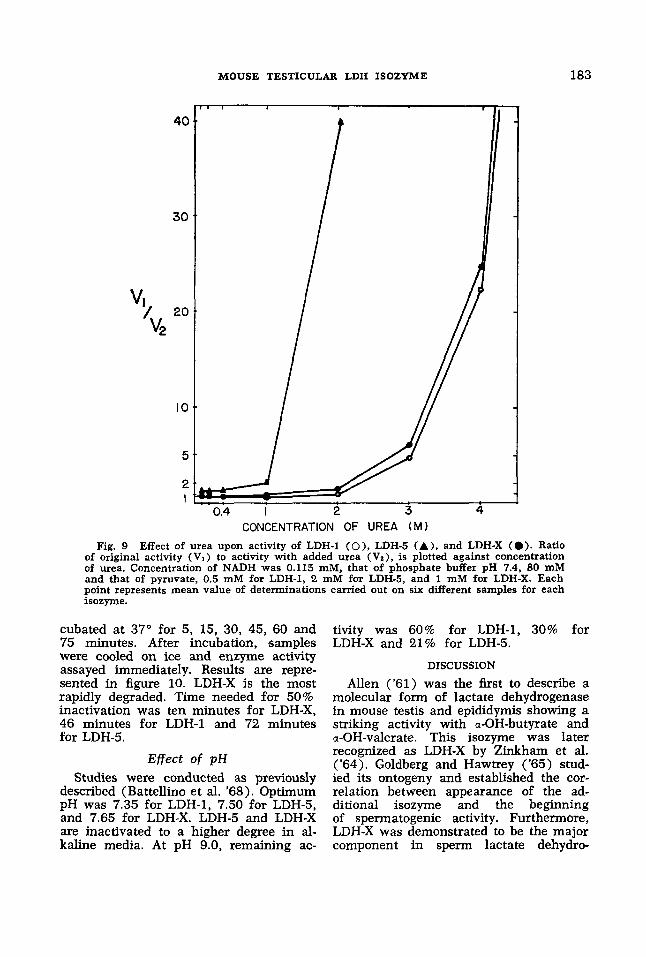
Urea. The inhibitory action of urea can be used for the differentiation of lac- tate dehydrogenase isozymes (Withy- combe, Plummer and Wilkinson, '65). Eluates were incubated with urea under the same conditions described for the other inhibitors. Two molar urea pro- duced a striking inactivation of the for- ward reaction catalyzed by LDH-5. LDH-1 was not affected and LDH-X showed an intermediate effect (table 3). Activity of LDH-5 was reduced to 85% of that of control by 0.1 M urea and completely in- activated by 3 M solutions. A concentra- tion higher than 4 M is needed to get a similar effect on LDH-1 and LDH-X. Ratio of original activity (V,) over activity with added inhibitor ( V2) has been plotted against concentration of urea in figure 9. Fifty per cent inhibition (the point where V1/V2 equals 2) occurs at a 2.33 M con- centration for LDH-1, 0.92 for LDH-5 and 2.10 M for LDH-X. These values varied markedly with the temperature of incuba- tion; the given figures were obtained at 20", but were different at other tempera-
TABLE 2
Effec t of metabolites upon mouse lactate dehydrogenase isozymes ~~ ~~
LDH-1 LDH-5 LDH-X
DL-Malate 97.5 (f3.81) 102.0 (f3.08) 69.5 ( f 9.50) Citrate 97.5 ( f 3.74) 98.5 ( e4.03) 78.0 ( f 3.89) Succinate 99.0 ( & 2.45) 100.0 ( f 5.47) 73.0 ( f 6.49)
I.-Aspartate 100.0 (24.86) 102.0 ( If: 3.52) 81.0 (e4.83) L-Glutamate 99.0 ( f 2.78) 101.0 (24.34) 81.0 (e5.12)
Numbers in parentheses represent standard deviations. Metabolites were added to the reagent mixture up to a final concentration of 1.5 mM and incubated
for 30 minutes before the substrate was added. Optimum concentration of pyruvate was used for the assays. Values are expressed as percentage of the activity without added metabolite, and represent averages of determinations on seven different samples.
182 LUIS J. BATTELLINO AND ANTONIO BLANCO
50
00
50
Fig. 8 Dixon plot of inhibition by malate of the reaction pyruvate + lactate catalyzed by mouse LDH-X. Reciprocal of the initial reaction velocity is plotted against concentration of malate. Final concentration of NADH was 0.115 mM, that of phosphate buffer pH 7.4, 80 mM and those of pyru- vate were 1 mM (A), 0.2 mM (e) and 0.02 mM (0). V: micromoles of NADH/minute.
TABLE 3 Effect of inhibitors upon mouse lactate dehydrogenase isozymes
LDH-1 LDH-5 LDH-X
Reaction pyruvate + lactate 2.0 M Urea 102.0 8.0 81.5 0.2 mM Oxalate 18.0 66.5 11.0
2.0 M Urea 23.0 11.0 20.0 0.2 mM Oxalate 56.0 87.0 42.0
Values are expressed as percentage of the maximal activity without inhibitor and represent mean values of determinations on seven different samples. Enzyme-coenzyme-inhibitor mlxture was set aside for 30 minutes before optimum concentration of substrate was added.
Reaction lactate -;r pyruvate
tures. Fifty per cent inhibition of LDH-5 Twenty-five milligrams of trypsin were activity, for example, was attained with dissolved per milliliter of 0.1 M phosphate 2.04 M urea at 10" and with 0.66 Tvl urea buffer pH 7.4 containing 5% Triton at 37". X-100. Equal volumes of eluates contain-
Trypsin. The procedure described by ing a specific activity of 5 U mg of protein Clausen and @vlisen ('65) was adapted. and trypsin solution were mixed and in-
MOUSE TESTICULAR LDH ISOZYME 183
CONCENTRATION OF UREA ( M I Fig. 9 Effect of urea upon activity of LDH-1 (O), LDH-5 (A), and LDH-X (0 ) . Ratio
of original activity (V,) to activity with added urea (VZ), is plotted against concentration of urea. Concentration of NADH was 0.115 mM, that of phosphate buffer pH 7.4, 80 mM and that of pyruvate, 0.5 mM for LDH-1, 2 mM for LDH-5, and 1 mM for LDH-X. Each point represents mean value of determinations carried out on six different samples for each isozyme.
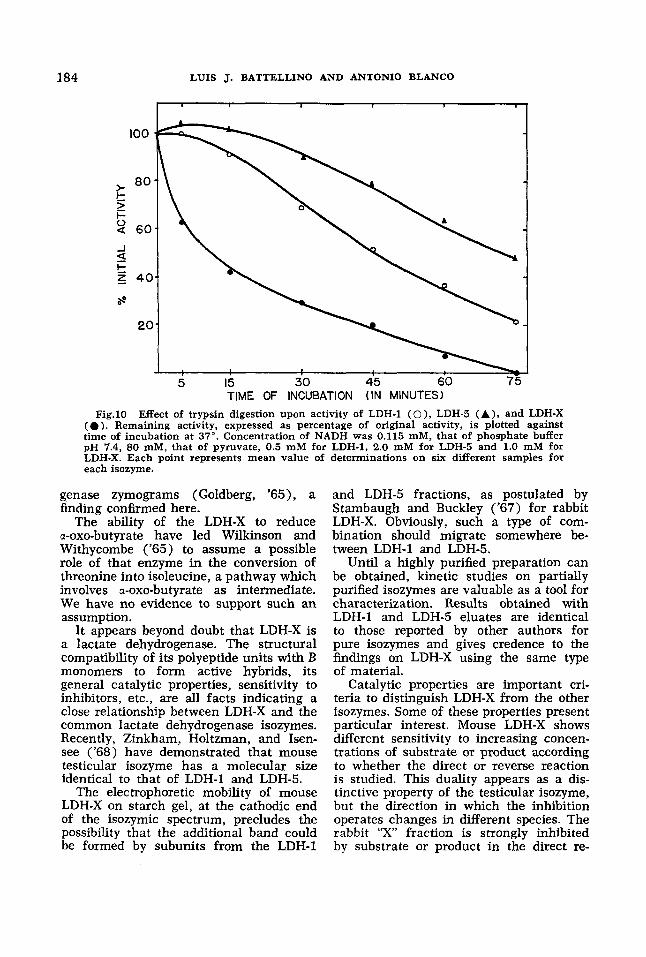
cubated at 37" for 5, 15, 30, 45, 60 and 75 minutes. After incubation, samples were cooled on ice and enzyme activity assayed immediately. Results are repre- sented in figure 10. LDH-X is the most rapidly degraded. Time needed for 50% inactivation was ten minutes for LDH-X, 46 minutes for LDH-1 and 72 minutes for LDH-5.
Effect of pH Studies were conducted as previously
described (Battellino et al. '68). Optimum pH was 7.35 for LDH-1, 7.50 for LDH-5, and 7.65 for LDH-X. LDH-5 and LDH-X are inactivated to a higher degree in al- kaline media. At pH 9.0, remaining ac-
tivity was 60% for LDH-1, 30% for LDH-X and 21% for LDH-5.
DISCUSSION
Allen ('61) was the f i s t to describe a molecular form of lactate dehydrogenase in mouse testis and epididymis showing a striking activity with a-OH-butyrate and a-OH-valerate. This isozyme was later recognized as LDH-X by Zinkham et al. ('64). Goldberg and Hawtrey ('65) stud- ied its ontogeny and established the cor- relation between appearance of the ad- ditional isozyme and the beginning of spermatogenic activity. Furthermore, LDH-X was demonstrated to be the major component in sperm lactate dehydro-
184 LUIS J. BATTELLINO AND ANTONIO BLANCO
80 - z 2
$ 60-
9 k g 40.
8
I-
-I
20-
- 5 15 30 45 60 75
TIME OF INCUBATION (IN MINUTES) Fig.10 Effect of trypsin digestion upon activity of LDH-1 ( O ) , LDH-5 (A) , and LDH-X
(0 ) . Remaining activity, expressed as percentage of original activity, is plotted against time of incubation at 37". Concentration of NADH was 0.115 mM, that of phosphate buffer pH 7.4, 80 mM, that of pyruvate, 0.5 mM for LDH-1, 2.0 mM for LDH-5 and 1.0 mM for LDH-X. Each point represents mean value of determinations on six different samples for each isozyme.
genase zymograms (Goldberg, '65), a finding confirmed here.
The ability of the LDH-X to reduce a-0x0-butyrate have led Wilkinson and Withycombe ('65) to assume a possible role of that enzyme in the conversion of threonine into isoleucine, a pathway which involves a-0x0-butyrate as intermediate. We have no evidence to support such an assumption.
It appears beyond doubt that LDH-X is a lactate dehydrogenase. The structural compatibility of its polyeptide units with B monomers to form active hybrids, its general catalytic properties, sensitivity to inhibitors, etc., are all facts indicating a close relationship between LDH-X and the common lactate dehydrogenase isozymes. Recently, Zinkham, Holtzman, and Isen- see ('68) have demonstrated that mouse testicular isozyme has a molecular size identical to that of LDH-1 and LDH-5.
The electrophoretic mobility of mouse LDH-X on starch gel, at the cathodic end of the isozymic spectrum, precludes the possibility that the additional band could be formed by subunits from the LDH-1
and LDH-5 fractions, as postulated by Stambaugh and Buckley ('67) for rabbit LDH-X. Obviously, such a type of com- bination should migrate somewhere be- tween LDH-1 and LDH-5.
Until a highly purified preparation can be obtained, kinetic studies on partially purified isozymes are valuable as a tool for characterization. Results obtained with LDH-1 and LDH-5 eluates are identical to those reported by other authors for pure isozymes and gives credence to the findings on LDH-X using the same type of material.
Catalytic properties are important cri- teria to distinguish LDH-X from the other isozymes. Some of these properties present particular interest. Mouse LDH-X shows different sensitivity to increasing concen- trations of substrate or product according to whether the direct or reverse reaction is studied. This duality appears as a dis- tinctive property of the testicular isozyme, but the direction in which the inhibition operates changes in different species. The rabbit "X" fraction is strongly inhibited by substrate or product in the direct re-

MOUSE TESTICULAR LDH ISOZYME 185
action (Battellino et al., '68), while mouse LDH-X is not. The reverse is true for the backward reaction.
Perhaps this fact has some physiologi- cal meaning. Properties of rabbit isozyme correlate with the metabolic requirements of spermatozoa (Battellino et al., '68). Un- fortunately, mouse sperm has not been studied from the metabolic point of view. The lack of sensitivity to high concentra- tions of substrate or product in the direct reaction suggests that mouse LDH-X is adapted to an anaerobic type of metabo- lism. It is able to accumulate lactate, which, in turn, inhibits the backward re- action and can not be oxidized, especially if pyruvate is in high concentration. Ex- periments of inhibition by product seem particularly demonstrative because they utilize concentrations of substrate or prod- uct similar to the physiological ones.
Another curious finding is the inhibi- tion of mouse LDH-X by some metabolites of the Krebs cycle, especially malate and succinate. Although the real importance of this phenomenon can not be ascertained at present, it could indicate a regulatory mechanism assuring that LDH-X accom- plishes its function only in anaerobic con- ditions. When oxygen supply is restored and citric acid cycle is normally working, complete utilization of pyruvate would be favored by inactivation of lactate dehydro- genase. On the other hand, oxidation of lactate is not affected under the same conditions.
In conclusion, the fhdings reported here indicate that catalytic properties of mouse LDH-X clearly differ from those of common lactate dehydrogenase isozymes and LDH-Xs of other species. Those prop- erties suggest a high degree of specializa- tion and metabolic adaptation for the ad- ditional isozyme.
ACKNOWLEDGMENTS
We thank Dr. Guillermo Carvajal of the Departmento de Bioquimica, Escuela Na- cional de Ciencias Biolcjgicas, I. P. N., MCxico, who kindly supplied the N-( 4- carboxy-2-hydroxyphenyl) maleimide used in this study, and Dr. Ernest0 E. Aeber- hard, who read the manuscript and offered helpful criticisms,
LITERATURE CITED
Allen, J. M. 1961 Multiple forms of lactic de- hydrogenase in tissues of mouse: Their speci- ficity. cellular localization, and resDonse to al- tered.physiological.conditions. Ann.N. Y. Acad. Sci., 94: 937-951.
Battellino, L. J., F. Ramos Jaime and A. Blanco 1968 Kinetic urouerties of rabbit testicular lactate dehydrogenise isozyme. J. Biol. Chem., 243: 5185-5192.
Blanco, A., and W. H. Zinkham 1963 Lactate dehydrogenases in human testes. Science, 139:
Blanco, A., W. H. Zinkham and L. Kupchyk 1964 Genetic control and ontogeny of lactate dehy- drogenase in pigeon testes. J. Exp. Zool., 156:
Boyer, S. H., D. F. Fainer and M. A. Naughton 1963 Myoglobin: Inherited structural varia- tion in man. Science, 140: 1228-1231.
Cahn, R. O., N. 0. Kaplan, L. Levine and E. Zwilling 1962 Nature and development of lactic dehydrogenases. Science, 136: 962-969.
Carvajal, G., E. J. Carvajal, R. YAfiez and V. Z. Medina 1967 Lactic dehydrogenase activity and aerobic glycolysis in tumors. In: Interna- tional Symposium: Enzymatic Aspects of Me- tabolic Regulation. M. P. Stulberg, ed. National Cancer Institute Monograph 27, Bethesda, Md.,
Clausen, J., and B. Ovlisen 1965 Lactate dehy. drogenase isoenzymes of human semen. Bio- chem. J., 97: 513-517.
Emerson, P. M., and J. H. Wilkinson 1965 Urea and oxalate inhibition of the serum lac- tate dehydrogenase. J. Clin. Path., 18: 803-807.
Goldberg, E. 1965 Lactate dehydrogenase in spermatozoa: subunits interactions in vitro. Arch. Biochem. Biophys., 109: 134-141.
Goldberg, E., and C. Hawtrey 1967 The onto- geny of sperm specific lactate dehydrogenase i n mice. J. Exp. Zool., 164: 309-316.
Markert, C. L. 1962 Isozymes in kidney de- velopment. In: Proceedings of the 13th. Annual Conference on Kidney. J. Metcoff, ed. North- western University Press, Evanston, 111. pp. 54-63.
1963 Lactate dehydrogenase isozymes: Dissociation and recombination of subunits. Science, 140: 1329-1330.
Markert, C. L., and H. Ursprung 1962 The on- togeny of isozyme patterns of lactate dehydro- genase in the mouse. Develop. Biol., 5: 363- 381.
Smithies, 0. 1959 An improved procedure for starch-gel electrophoresis: Further variations in the serum proteins of normal individuals. Biochem. J., 71: 585-587.
Stambaugh, R., and J. Buckley 1967 The en- zymic and molecular nature of the lactic de- hydrogenase subbands and X, isozyme. J. Biol. Chem., 242: 40534059.
Stambaugh, R., and D. Post 1966 Substrate and product inhibition of rabbit muscle lactic dehydrogenase heart (H,) and muscle (MI) isozymes. J. Biol. Chem., 241: 1462-1467.
601-602.
137-152.
pp. 111-124.

186 LUIS J. BATTELLINO AND ANTONIO BLANCO
Wilkinson, J. H., and W. A. Withycombe 1965 Organ specificity and lactate dehydrogenase ac- tivity. Some properties of human spermatozoa1 lactate dehydrogenase. Biochem. J., 97: 663- 668.
Withycombe, W. A., D, T. Plummer and J. H. Wilkinson 1965 Organ specificity and lac- tate dehydrogenase activity. Differential inhi- bition by urea and related compounds. Bio- chem. J., 94: 384-389.
Wr6blewski, F., and J. S. LaDue 1955 Lactic dehydrogenase activity in blood. Proc. SOC. Exp. Biol. Med., 90: 210-213.
Zinkham, W. H., A. Blanco and L. J. C l o ~ r y 1964 An unusual isozyme of lactate dehydro- genase in mature testes: Localization, onto- geny and kinetic properties. Ann. N. Y. Acad. Sci., 121: 571-588.
Zinkham, W. H., A. Blanco and L. Kupchyk 1963 Lactate dehydmgenase in testes: Disso- ciation and recombination of subunits. Science,
Zinkham, W. H., N. A. Holtzman and H. Isensee 1968 The molecular size of lactate dehydro- genase isozymes in mature testes. Biochim. Biophys. Acta, 160: 172-177.
142: 1303-1304.