butyrylcholinesterase and the cholinergic system
TRANSCRIPT

Neuroscience 234 (2013) 53–68
BUTYRYLCHOLINESTERASE AND THE CHOLINERGIC SYSTEM
G. A. REID, a N. CHILUKURI b AND S. DARVESH a,c*
aDepartment of Medical Neuroscience Dalhousie University, Halifax,
Nova Scotia, CanadabResearch Division, Physiology & Immunology Branch, US
Army Medical Research, Institute of Chemical Defense,
Aberdeen Proving Ground, MD 21010, USA
cDepartment of Medicine (Neurology and Geriatric Medicine),
Dalhousie University, Halifax, Nova Scotia, Canada
Abstract—The cholinergic system plays important roles in
neurotransmission inboth theperipheral andcentral nervous
systems. The cholinergic neurotransmitter acetylcholine is
synthesized by choline acetyltransferase (ChAT) and its
action terminated by acetylcholinesterase (AChE) and butyr-
ylcholinesterase (BuChE). The predominance of AChE has
focused much attention on understanding the relationship
of this enzyme to ChAT-positive cholinergic neurons. How-
ever, there is ample evidence thatBuChEalsoplays an impor-
tant role in cholinergic regulation. To elucidate the
relationship of BuChE to neural elements that are producing
acetylcholine, the distribution of this enzyme was compared
to that of ChAT in the mouse CNS. Brain tissues from
0306-4522 � 2013 IBRO. Published by Elsevier Ltd.http://dx.doi.org/10.1016/j.neuroscience.2012.12.054
*Correspondence to: S. Darvesh, Department of Medical Neuroscience, DaVeterans’ Memorial Lane, Halifax, Nova Scotia, Canada B3H 2E1. Tel: +1-
E-mail address: [email protected] (S. Darvesh).Abbreviations: AChE, acetylcholinesterase; Ad-moBuChE, adenovirus containAA, anterior amygdaloid area; ac, anterior commissure; ACo, anterior corticalAHP, anterior posterior hypothalamic nucleus; APT, anterior pretectal nucleusauditory cortex; BL, basolateral amygdala; BM, basomedial amygdala; BST,butyrylcholinesterase; CPu, caudate putamen nucleus; Ce, central amygdala;cortex; Cl, claustrum; cc, corpus collosum; DpMe, deep mesencephalic nuclegeniculate nucleus; 10N, dorsal motor nucleus of the vagus; DR, dorsal raimmunosorbent assay; En, endopiriform cortex; ec, external capsule; ECu, exfasciculus retroflexus; FBS, fetal bovine serum; fi, fimbria; f, fornix; g7, genupallidus; H, hippocampal formation; HC, histochemical; HDB, horizontabutyrylcholinesterase; 12N, hypoglossal nucleus; IgG, immunoglobulin G; icpnucleus; IS, inferior salivatory nucleus; I, insular cortex; InG, intermediate graintermediate white layer of the superior colliculus; ic, internal capsule; IP, intCalleja; La, lateral amygdala; LDTg, lateral dorsal tegmental nucleus; LD,hypothalamus; LPGi, lateral paragigantocellular nucleus; LP, lateral posterior tlateral vestibular nucleus; LC, locus coeruleus; MCPO, magnocellular preopticMD, medial dorsal thalamus; MEnt, medial entorhinal cortex; MG, medial geniclongitudinal fasciculus; MS, medial septal nucleus; MT, medial terminal nucleuraphe nucleus; MiTg, microcellular tegmental nucleus; M, motor cortex; Mo5,nigrostriatal bundle; A5, noradrenaline cell group A5; LL, nucleus of lateralhypothalamic nucleus; PPTg, pedunculopontine tegmental nucleus; PAG, periaPir, piriform cortex; PoDG, polymorphic cell layer of the dentate gyrus; Pn, pcommissure; PH, posterior hypothalamic nucleus; PMCo, posteromedial corticanucleus, ventral; PO, preoptic nucleus; Pr, prepositus nucleus; Pr5, principal stract; rmoBuChE, recombinant mouse BuChE; R, red nucleus; Rt, reticular ncortex; Re, reuniens; RI, rostral interstitial nucleus of mlf; RPO, rostral paraosodium dodecyl sulfate–polyacrylamide gel electrophoresis; sol, solitary tract;Sp5OVL, spinal trigeminal nucleus, ventrolateral oralis; sp5, spinal trigeminamedullaris; st, stria terminalis; SubC, subcoeruleus nucleus; SMT, submammillosuperior cerebellar peduncle; str, superior thalamic radiation; SuVe, superitectospinal tract; TeA, temporal association cortex; TMB, 3,30,5,50-tetramethylbeanterior thalamic nucleus; VLG, ventral lateral geniculate; VP, ventral pallidumband of Broca; V, visual cortex; ZI, zona incerta.
53
Open access under CC BY-NC-N
129S1/SvImJ mice were stained for BuChE and ChAT using
histochemical, immunohistochemical and immunofluorescent
techniques. Both BuChE and ChAT were found in neural
elements throughout theCNS.BuChEstainingwith histochem-
istry and immunohistochemistry produced the same distribu-
tion of labeling throughout the brain and spinal cord.
Immunofluorescent double labeling demonstrated that many
nuclei in themedulla oblongata, aswell as regions of the spinal
cord, had neurons that contained both BuChE and ChAT.
BuChE-positive neurons without ChAT were found in close
proximitywithChAT-positive neuropil in areas suchas the thal-
amus and amygdala. BuChE-positive neuropil was also found
closely associated with ChAT-positive neurons, particularly
in tegmental nuclei of the pons. These observations
provide further neuroanatomical evidenceof a role forBuChE
in the regulation of acetylcholine levels in the CNS.
� 2013 IBRO. Published by Elsevier Ltd.
Key words: cholinesterases, acetylcholinesterase, pseudo-
cholinesterase, choline acetyltransferase, 129S1/SvImJ mice,
acetylcholine.
Open access under CC BY-NC-ND license.
lhousie University, Room 1308, Camp Hill Veterans’ Memorial, 5955902-473-2490; fax: +1-902-473-7133.
ing the gene for mouse butyrylcholinesterase; Amb, ambiguus nucleus;amygdala; AD, anterior dorsal thalamus; AO, anterior olfactory cortex;; ATg, anterior tegmental nucleus; AV, anterior ventral thalamus; Au,bed nucleus of stria terminalis; BSA, bovine serum albumin; BuChE,cp, cerebral peduncle; ChAT, choline acetyltransferase; Cg, cingulateus; DAB, 3,30-diaminobenzidine tetrahydrochloride; DLG, dorsal lateralphe nucleus; DTg, dorsal tegmental nucleus; ELISA, enzyme-linkedternal cuneate; eml, external medullary lamina; 7N, facial nucleus; fr,of the facial nucleus; Gi, gigantocellular reticular nucleus; GP, globusl limb nuclei of the diagonal band of Broca; hBuChE, human, inferior cerebellar peduncle; IC, inferior colliculus; IO, inferior olivaryy layer of superior colliculus; IRt, intermediate reticular nucleus; InWh,erpeduncular nucleus; InC, interstitial nucleus of Cajal; ICj, islands oflateral dorsal thalamus; LEnt, lateral entorhinal cortex; LH, lateral
halamus; LRt, lateral reticular nucleus; LS, lateral septal nucleus; LVe,nucleus; mt, mammillothalamic tract; MeA, medial anterior amygdala;
ulate nucleus; MHb, medial habenula; ml, medial lemniscus; mlf, medials of the accessory tract; MVe, medial vestibular nucleus; MnR, medianmotor trigeminal nucleus; moBuChE, mouse butyrylcholinesterase; ns,lemniscus; Tu, olfactory tubercle; opt, optic tract; Pa, paraventricularqueductal gray; PB, phosphate buffer; PBS, phosphate-buffered saline;ontine nuclei; PnO, pontine reticular nucleus, oral part; pc, posteriorl amygdala; PMD, premammillary nucleus, dorsal; PMV, premammillaryensory trigeminal nucleus; Po, pulvinar thalamic nucleus; py, pyramidalucleus; RtTg, reticulotegmental nucleus of the pons; RS, retrospleniallivary nucleus; rs, rubral spinal tract; S, sensory cortex; SDS–PAGE,Sol, solitary tract nucleus; Sp5I, spinal trigeminal nucleus, interpolaris;l tract; SVe, spinal vestibular nucleus; Or, stratum oriens; sm, striathalamic nucleus; SI, substantia innominata; SN, substantia nigra; scp,or vestibular nucleus; SuMM, supramammillary, medial nucleus; ts,nzidine; m5, trigeminal nerve root; TBS, tris-buffered saline; VA, ventral; VTA, ventral tegmental area; VDB, vertical limb nuclei of the diagonal
D license.

54 G. A. Reid et al. / Neuroscience 234 (2013) 53–68
INTRODUCTION
Cholinergic neurotransmission in the mammalian CNS is
regulated predominantly by the enzyme
acetylcholinesterase (AChE, EC 3.1.1.7) by catalyzing
the hydrolysis of the cholinergic neurotransmitter
acetylcholine (Silver, 1974). Improved histochemical
techniques (Koelle and Friedenwald, 1949) for detecting
this enzyme led to the first map of the distribution of
AChE in the rodent brain (Shute and Lewis, 1963).
Subsequently, antibodies were developed to detect
choline acetyltransferase (ChAT, EC 2.3.1.6), the
enzyme that catalyzes the synthesis of acetylcholine
(Eng et al., 1974). This permitted elucidating the
organization of cholinergic neurons (Kimura et al., 1980;
Armstrong et al., 1983). Combined AChE histochemical
and ChAT immunohistochemical staining studies
demonstrated that most ChAT-positive neurons were
also AChE-positive. (Eckenstein and Sofroniew, 1983;
Levey et al., 1983, 1984).
In addition to AChE, the enzyme butyrylcholinesterase
(BuChE, EC 3.1.1.8) is important in the regulation of the
cholinergic system (Darvesh et al., 1998, 2003;
Mesulam et al., 2002b; Giacobini, 2003; Duysen et al.,
2007). Like AChE, BuChE is able to efficiently catalyze
the hydrolysis of acetylcholine (Silver, 1974). BuChE is
expressed in distinct populations of neurons, some of
which also contain AChE (Friede, 1967; Tago et al.,
1992; Darvesh et al., 1998; Darvesh and Hopkins, 2003;
Geula and Nagykery, 2007). The importance of BuChE
in cholinergic neurotransmission is further supported by
the observation that AChE-knockout mice survive to
adulthood. This indicates BuChE is able to compensate
for the lack of AChE, allowing the continued regulation
of cholinergic neurotransmission (Li et al., 2000; Xie
et al., 2000; Mesulam et al., 2002a).
To date, study of the colocalization of BuChE and
ChAT in the mammalian CNS has been limited to the
spinal cord in the rat (Mis, 2005). Because of the earlier
indications of BuChE co-regulation of cholinergic
neurotransmission (Xie et al., 2000; Mesulam et al.,
2002a,b), the present work was undertaken to examine
the organization of BuChE-expressing neural elements
as they relate to the ChAT-defined cholinergic system in
the mouse CNS.
This work has not been presented elsewhere except
in abstract form (Darvesh et al., 2012b).
EXPERIMENTAL PROCEDURES
Animals
Twenty male, wild-type (129S1/SvImJ) mice were purchased
from The Jackson Laboratory (USA). This mouse strain was
chosen because it has been utilized to examine components of
the cholinergic system in other studies (Li et al., 2000;
Mesulam et al., 2002a; Duysen et al., 2007). Animals were
cared for according to the guidelines set by the Canadian
Council on Animal Care. Formal approval to conduct the
experiments was obtained from the Dalhousie University
Committee on Laboratory Animals.
Materials
Unless otherwise stated, all reagents were purchased from
Sigma–Aldrich (St. Louis, MO).
Preparation of brain tissue
Mice (between 10 and 18 weeks old) were deeply anesthetized
with an intra-peritoneal injection of sodium pentobarbital
(200 mg/kg) and perfused with approximately 25 ml of 0.9%
saline solution containing 0.1% sodium nitrite followed by 50 ml
of 0.1 M phosphate buffer (PB, pH 7.4) containing 4%
paraformaldehyde. Brains were removed and post-fixed in PB
with 4% paraformaldehyde for 1–2 h, cryoprotected and stored
in PB with 30% sucrose and 0.05% sodium azide. Brains were
cut in 40-lm serial sections in a coronal plane on a Leica
SM2000R microtome with Physitemp freezing stage and BFS-
30TC controller. Sections were stained for BuChE or AChE by
histochemical (HC) technique, and for BuChE and ChAT using
immunohistochemical (IHC) methods. Double labeling for
BuChE and ChAT was performed using immunofluorescence
(IF).
Cholinesterase histochemistry
Cholinesterase histochemical staining was performed using a
modified (Darvesh et al., 2012a) Karnovsky–Roots method
(Karnovsky and Roots, 1964). Briefly, tissue sections were
rinsed in 0.1 M maleate buffer (pH 7.4) for 30 min and
incubated for 1 h 45 min in 0.1 M maleate buffer (pH 8.0)
containing 0.5 mM sodium citrate, 0.47 mM cupric sulfate,
0.05 mM potassium ferricyanide, 0.8 mM butyrylthiocholine
iodide and 0.01 mM BW 284 C 51 (to inhibit AChE). All
sections were then rinsed with gentle agitation for 30 min in
dH2O and placed in 0.1% cobalt chloride in water for 10 min.
After further rinsing in dH2O, sections were placed in PB
containing 1.39 mM 3,30-diaminobenzidine tetrahydrochloride
(DAB). After 5 min in the DAB solution, 50 ll of 0.15% H2O2 in
dH2O was added per ml of DAB solution, and the reaction was
carried out for approximately 3 min. Sections were then washed
in 0.01 M acetate buffer (pH 3.3), mounted on slides, cleared in
xylene and cover-slipped. Control experiments to demonstrate
specificity of BuChE staining were performed as described
previously (Darvesh et al., 1998) and indicated the staining
pattern observed was specific for BuChE activity.
The procedure for the visualization of AChE activity was
similar to that for BuChE except that the reaction solution was
24.99 mM sodium citrate, 14.72 mM cupric sulfate, 2.43 mM
potassium ferricyanide, 2.46 mM acetylthiocholine iodide and
0.13 mM ethopropazine (to inhibit BuChE), at room temperature
in 0.1 M maleate buffer (pH 6.0) for 15 min with gentle agitation.
Generation and characterization of polyclonalantibodies to BuChE
Rabbit polyclonal antibodies to recombinant mouse BuChE
(rmoBuChE) were generated for this study. Production of
rmoBuChE (immunogen) was accomplished in human
embryonic kidney epithelial cells (293A cells) using an
adenovirus containing the gene for mouse BuChE (Ad-
moBuChE) (Parikh et al., 2011). The mouse BuChE
(moBuChE) gene contained a 6x histidine tag at its carboxyl
terminus suggesting that the rmoBuChE used was a fusion
protein. Large-scale expression of rmoBuChE was
accomplished by overnight culturing of 293A cells (10 � 106) in
150-cm2 tissue culture dishes and infecting them for 1 h with
10 ll of 4th cycle crude viral lysate (high titer CVL) of Ad-
moBuChE in 10 ml of infection medium (DMEM containing 2%
fetal bovine serum (FBS), antibiotics penicillin and streptomycin

G. A. Reid et al. / Neuroscience 234 (2013) 53–68 55
and sodium pyruvate) at 37 �C. Fifteen ml of growth medium
(DMEM containing 10% FBS, 50 lg/ml penicillin and
streptomycin, and 50 lg/ml sodium pyruvate) was then added
and plates were returned to the incubator for 5–7 days.
Seventy to one hundred culture dishes were infected in a single
experiment. When BuChE activity in the culture medium
reached between 3–4 units/ml, it was collected and cleared of
cells and debris by centrifugation at 2500 rpm for 15 min, 4 �C.Culture media containing �60,000 units (or 70 mg) of the
recombinant enzyme was used for purification of rmoBuChE.
Ellman assay (Ellman et al., 1961), using butyrylthiocholine as
substrate was employed to monitor moBuChE activity during
various steps of the enzyme purification.
The purification scheme for rmoBuChE involved ammonium
sulfate fractionation followed by affinity chromatography using
procainamide and nickel-affinity resins. The culture media was
fractionated with ammonium sulfate (0–50% saturation). The
precipitate was collected by centrifugation at 10,000 rpm for
15 min and discarded as the supernatant contained all the
enzyme activity. Ammonium sulfate was added to the
supernatant to 80% saturation and the precipitate containing
the entire recombinant enzyme was collected as described
above. The enzyme was dissolved in 100 ml of 50 mM PB (pH
8.0), desalted by dialysis against 30 volumes of the same
buffer and the solution was then applied to a 50-ml column of
procainamide Sepharose equilibrated with the same buffer. The
bound enzyme was eluted with 50 mM PB (pH 8.0) containing
1 M NaCl and 0.2 M choline chloride. The active fractions were
combined and dialyzed against 25 mM PB (pH 8.0) containing
500 mM NaCl and 2.5 mM imidazole and loaded onto a 10-ml
column of nickel Sepharose resin equilibrated against the same
buffer. The bound enzyme was eluted with 25 mM PB (pH 8.0)
containing 500 mM NaCl and 500 mM imidazole. The enzyme
fractions were combined and dialyzed against 25 mM PB (pH
8.0). For long-term storage of the enzyme, glycerol was added
(50%) and stored in �20 �C freezer.
Polyclonal antibodies to rmoBuChE were produced in two
New Zealand white rabbits according to standard polyclonal
antibody production protocols (Washington Biotechnology Inc.,
6200 Seaforth Street, Baltimore, MD 21224). Three mg of the
purified rmoBuChE was supplied to Washington Biotechnology
as lyophilized powder for antibody development. The protocol
was approved by the Institutional Animal Care and Use
Committee of the Johns Hopkins University, Baltimore,
Maryland. Sera collected after the booster injections were used
for purification of anti-moBuChE immunoglobulin G (IgG) by
protein A/G chromatography. Sera were diluted with
phosphate-buffered saline (PBS, 1:10 dilution; pH 7.4) and
loaded onto a 5-ml column of protein A/G Sepharose beads.
The bound IgG were eluted with 50 mM glycine–HCl (pH 2.7)
and fractions containing IgG were neutralized using 1 M dibasic
PB (pH 7.5). Later, sodium azide and glycerol were added to
final concentrations of 0.05% and 10%, respectively for long-
term storage of the antibody.
The reactivity of anti-moBuChE IgG toward rmoBuChE or
human BuChE (hBuChE) was determined by Western blotting.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis
(SDS–PAGE) was carried out for 100 ng of each enzyme using
precast 10% Tris–HCl gels. One microgram of bovine serum
albumin (BSA) was included in the sample to prevent non-
specific loss of the enzyme. After electrophoresis, proteins
were transferred to nitrocellulose membrane using IBlot gel
transfer apparatus (InVitrogen, CA). The membrane was
blocked in blocking buffer (Licor Inc., NE) for 2 h at 24 �C,washed once with 50 mM phosphate buffer containing 0.05%
Tween 20 (washing buffer) and kept overnight in blocking buffer
containing anti-moBChE antibody (1:20,000 dilution). The
membrane was then washed five times with intermittent
shaking for 5 min and incubated with secondary antibody
conjugated with infra red dye 680 (Licor Inc., NE, 1:10,000
dilution) made in blocking buffer for 1 h, and protein bands
were detected using Infrared Imager (Licor Inc. NE). The
reactivity of anti-moBuChE IgG toward recombinant moBuChE
or hBuChE was also determined by direct enzyme-linked
immunosorbent assay (ELISA). One hundred ll of recombinant
moBuChE or hBuChE in PBS (5 lg/ml) was added to each well
of a 96-well plate and incubated overnight at 4 �C. After
washing the wells with PBS containing 0.05% Tween 20
(washing buffer), they were blocked with 3% BSA in PBS for
2 h at 24 �C. After washing the plate three times with washing
buffer, 100 ll each of five or fourfold serial dilutions (ranging
from 1:1000 to 1:4,000,000) of anti-moBChE IgG in PBS
containing 0.1% BSA was added and incubated for 2 h at
24 �C, followed by five washes with washing buffer. One
hundred ll of goat anti-rabbit IgG (H + L) conjugated with
horseradish peroxidase diluted 1:10,000 in PBS containing
0.1% BSA was then added to each well and incubated for 1 h
at 24 �C. After eight to 10 washes with washing buffer, 100 llof substrate solution containing 3,30,5,50-tetramethylbenzidine
(TMB) was added and incubated for 15 min in the dark at
24 �C. The reaction was stopped with the addition of 100 ll of2 N sulfuric acid and end point absorbance of each well was
measured in a plate reader at 450 nm. BSA blocked wells were
used as reagent controls.
Butyrylcholinesterase immunohistochemistry
For BuChE immunohistochemistry (IHC), sections were rinsed in
PB for 30 min and placed in 0.3% H2O2 in PB for 30 min to
quench endogenous peroxidase activity. Sections were rinsed
again in PB for 30 min and then incubated in PB containing
0.1% Triton X-100, normal goat serum (NGS, 1:100) and rabbit
anti-BuChE primary antibody (1:10,000) for approximately 16 h
at room temperature. After rinsing, sections were incubated in
PB with 0.1% Triton X-100, NGS (1:1000) and biotinylated goat
anti-rabbit secondary antibody (1:500; Vector) for 1 h. Sections
were rinsed in PB and then placed in PB with 0.1% Triton X-
100 and Vectastain� Elite ABC kit (Vector) according to the
manufacturer’s instructions. After rinsing the sections in PB,
they were placed in a solution of PB containing 1.39 mM DAB.
After 5 min, 50 ll of 0.3% aqueous H2O2 per mL of DAB
staining solution was added and incubated for 5 min. The
reaction was stopped by rinsing the sections in 0.01 M acetate
buffer (pH 3.3). Sections were mounted on slides, cleared in
xylene and cover-slipped.
Choline acetyltransferase immunohistochemistry
For ChAT-IHC, sections were rinsed in 0.05 M tris-buffered saline
(TBS; pH 7.6) for 30 min and placed in 0.3% H2O2 in TBS for
30 min to quench endogenous peroxidase activity. Sections
were rinsed again for 30 min and incubated in TBS containing
0.3% Triton X-100, normal rabbit serum (NRS, 1:100) and goat
anti-ChAT primary antibody (1:1000; Millipore, AB144P) for
about 16 h at room temperature. After rinsing, sections were
incubated in TBS containing 0.3% Triton X-100, normal rabbit
serum (1:1000) and biotinylated rabbit anti-goat secondary
antibody (1:500; Vector) for 1 h. After another rinse, sections
were placed in TBS containing 0.3% Triton X-100 and
Vectastain� Elite ABC kit according to the manufacturer’s
instructions. After rinsing, sections were placed in a solution of
TBS (pH 8.0) containing 1.39 mM DAB and 0.6% nickel
ammonium sulfate. After 5 min, 50 ll of 0.3% H2O2 in dH2O
was added per mL of DAB staining solution and the mixture
was incubated for 2–5 min. The reaction was stopped by
rinsing the sections in TBS. Sections were mounted on slides,
cleared in xylene and cover-slipped. Control experiments were
performed by omitting the primary antibody and no staining was
observed.

56 G. A. Reid et al. / Neuroscience 234 (2013) 53–68
Butyrylcholinesterase and choline acetyltransferaseimmunofluorescence double labeling
Double labeling for BuChE and ChAT was done on the same
sections using an immunofluorescent method. Sections were
rinsed in TBS for 30 min and incubated for approximately 16 h
at room temperature in TBS containing 0.3% Triton X-100,
normal donkey serum (1:100) and both primary antibodies,
rabbit anti-BuChE (1:7000) and goat anti-ChAT (1:1000). After
rinsing in TBS, sections were incubated for 1 h in TBS
containing 0.3% Triton X-100, donkey anti-goat Alexa Fluor�
488 (1:350; Molecular Probes, A11055) and donkey anti-rabbit
Alexa Fluor� 555 (1:350; Molecular probes, A31572). Sections
were rinsed, mounted on slides, cleared in xylene and cover-
slipped.
Data analysis
Sections stained by HC, IHC or IF were analyzed on a Zeiss
Axioplan 2 microscope and photographed with a Zeiss Axiocam
HRc digital camera and AxioVision 4.6 software.
Photomicrographs of IF sections were also acquired on a Zeiss
LSM 510 laser scanning-confocal microscope. Orthogonal
images were rendered and edited with LSM imaging software
(Zeiss). Photographs were assembled using Adobe�
Photoshop� CS5. Images were contrast enhanced and the
brightness adjusted to match the background from different
images. Images of IF labeling for BuChE and ChAT were
merged to illustrate the relationship between these two
markers. BuChE-IF appeared green while ChAT appeared red.
Neural elements with both BuChE and ChAT appeared yellow
in merged images.
Anatomical maps of the distribution of BuChE or ChAT were
produced by plotting the neuronal distribution on a Wacom
drawing tablet with Microbrightfield Neuroleucida software
linked to a Microfire digital camera on an Olympus BMAX
microscope with Heidenhain linear potentiometers and Prior
Optiscan controller. The same sections were then
photographed at low power and assembled into a single image
using Adobe� Photoshop� CS5. The anatomical parcellation,
based on the mouse brain atlas by Paxinos and Franklin
(2001), and neuronal distributions were then redrawn using
Adobe� Illustrator� CS5 by overlaying and aligning the
neuroleucida drawings on the low power photomicrographs.
RESULTS
The present investigation was undertaken to elucidate the
relationships between BuChE-positive neural elements
and ChAT in the mouse CNS. Using HC and IHC
methods, the distributions of BuChE and ChAT neural
elements were analyzed. Colocalization of these two
markers was confirmed with immunofluorescent (IF)
double labeling.
Characterization of mouse BuChE antibodies
Analysis of rmoBuChE by reducing SDS–PAGE revealed
that it was a pure protein and the molecular weight of
85 kDa for its subunit was similar to that for purified
native hBuChE (Fig. 1A). Polyclonal moBuChE antibody
was characterized for its binding and specificity to
recombinant moBuChE and hBuChE by Western
blotting (Fig. 1B) and direct ELISA (Fig. 1C). Both
assays show that moBuChE antibody preferred binding
to moBuChE than hBuChE. By Western blotting,
antibody binding to moBuChE was 10-fold greater than
its binding to hBuChE (Fig. 1B). ELISA results also
showed a similar trend. Certain dilutions of the antibody
(1:25,000, 1:125,000, and 1:500,000) produced a
fourfold higher absorbance values with moBuChE
compared to hBuChE (Fig. 1C). The specificity of
moBuChE antibody was also confirmed by conducting
BuChE-IHC with the antibody following incubation with
BuChE from mouse serum. No staining was observed
indicating specificity of this antibody (Fig. 1D). All
together, these results demonstrate the specificity of this
antibody toward moBuChE.
Validation of BuChE immunohistochemical staining
The mouse BuChE antibody was evaluated for its ability
to recapitulate the distribution of BuChE activity through
established histochemical analysis. To this end,
sections, at representative levels throughout the brain,
were stained by BuChE-HC, BuChE-IHC or AChE-HC
and compared. Fig. 2 provides two examples, the
piriform cortex (Pir; Fig. 2A–C) and the motor trigeminal
nucleus (Mo5; Fig. 2D–F), that illustrate the
comparability of BuChE-HC (Fig. 2A, D) and BuChE-
IHC (Fig. 2B, E) staining and emphasizes differences in
distribution of this enzyme relative to that of AChE-HC
staining (Fig. 2C, F). These areas were chosen as
examples because of the distinct distribution of BuChE
and AChE neural elements that occur therein.
Throughout the brain, the patterns of staining for BuChE
were comparable whether HC or IHC was used, except
that neuropil staining by BuChE-HC was generally more
diffuse than that revealed by BuChE-IHC. These
experiments confirmed the ability of the BuChE antibody
to accurately visualize the location of BuChE that is
conventionally obtained by HC analysis.
Distribution of butyrylcholinesterase and cholineacetyltransferase
Within the mouse CNS, BuChE staining labeled distinct
populations of neurons, neuropil, glia and white matter
while ChAT labeled neurons and neuropil (see Fig. 3 for
examples). The glial perikarya were distinguished from
the neuronal perikarya based on their size and
morphology (Kettenmann and Ransom, 2005). Staining
patterns for each marker were consistent in all brains
examined. Comparative photomicrographs and
corresponding schematic maps (Fig. 4) of BuChE-HC
(left) and ChAT-IHC (right), show the presence of
stained neural elements in each region, proceeding from
the rostral to the caudal levels of the brain. In these
maps, each neuron stained for BuChE or ChAT is
represented by a black circle. Stained neuropil and
white matter, not dealt with in the maps, can be seen
directly for each marker in the corresponding
photomicrographs (Fig. 4). A summary of structures in
the mouse brain containing neuronal somata or neuropil
stained for BuChE and ChAT is presented in Table 1
with an estimate of soma numbers and density of
neuropil. In this tables, boxes shaded in gray indicate
nuclei that contain neurons stained for both BuChE and
ChAT as demonstrated by IF double labeling. The
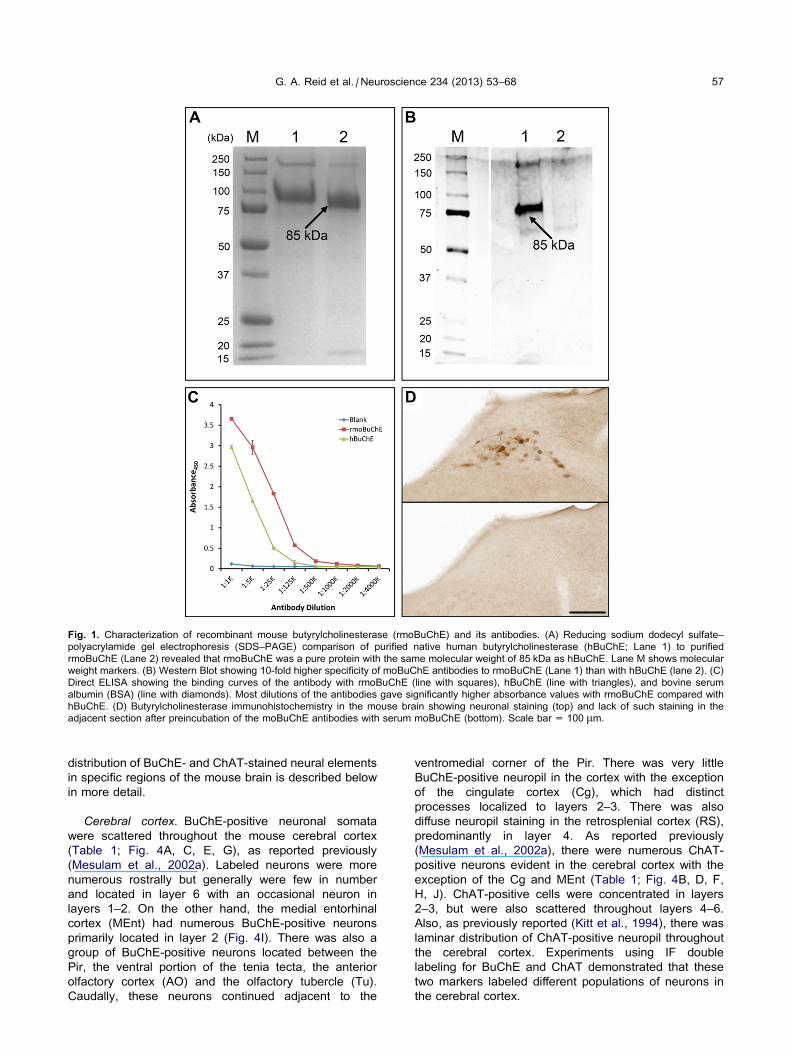
Fig. 1. Characterization of recombinant mouse butyrylcholinesterase (rmoBuChE) and its antibodies. (A) Reducing sodium dodecyl sulfate–
polyacrylamide gel electrophoresis (SDS–PAGE) comparison of purified native human butyrylcholinesterase (hBuChE; Lane 1) to purified
rmoBuChE (Lane 2) revealed that rmoBuChE was a pure protein with the same molecular weight of 85 kDa as hBuChE. Lane M shows molecular
weight markers. (B) Western Blot showing 10-fold higher specificity of moBuChE antibodies to rmoBuChE (Lane 1) than with hBuChE (lane 2). (C)
Direct ELISA showing the binding curves of the antibody with rmoBuChE (line with squares), hBuChE (line with triangles), and bovine serum
albumin (BSA) (line with diamonds). Most dilutions of the antibodies gave significantly higher absorbance values with rmoBuChE compared with
hBuChE. (D) Butyrylcholinesterase immunohistochemistry in the mouse brain showing neuronal staining (top) and lack of such staining in the
adjacent section after preincubation of the moBuChE antibodies with serum moBuChE (bottom). Scale bar = 100 lm.
G. A. Reid et al. / Neuroscience 234 (2013) 53–68 57
distribution of BuChE- and ChAT-stained neural elements
in specific regions of the mouse brain is described below
in more detail.
Cerebral cortex. BuChE-positive neuronal somata
were scattered throughout the mouse cerebral cortex
(Table 1; Fig. 4A, C, E, G), as reported previously
(Mesulam et al., 2002a). Labeled neurons were more
numerous rostrally but generally were few in number
and located in layer 6 with an occasional neuron in
layers 1–2. On the other hand, the medial entorhinal
cortex (MEnt) had numerous BuChE-positive neurons
primarily located in layer 2 (Fig. 4I). There was also a
group of BuChE-positive neurons located between the
Pir, the ventral portion of the tenia tecta, the anterior
olfactory cortex (AO) and the olfactory tubercle (Tu).
Caudally, these neurons continued adjacent to the
ventromedial corner of the Pir. There was very little
BuChE-positive neuropil in the cortex with the exception
of the cingulate cortex (Cg), which had distinct
processes localized to layers 2–3. There was also
diffuse neuropil staining in the retrosplenial cortex (RS),
predominantly in layer 4. As reported previously
(Mesulam et al., 2002a), there were numerous ChAT-
positive neurons evident in the cerebral cortex with the
exception of the Cg and MEnt (Table 1; Fig. 4B, D, F,
H, J). ChAT-positive cells were concentrated in layers
2–3, but were also scattered throughout layers 4–6.
Also, as previously reported (Kitt et al., 1994), there was
laminar distribution of ChAT-positive neuropil throughout
the cerebral cortex. Experiments using IF double
labeling for BuChE and ChAT demonstrated that these
two markers labeled different populations of neurons in
the cerebral cortex.

Fig. 2. Staining in the piriform cortex (Pir; A–C) and the trigeminal motor nucleus (Mo5; D–F) comparing butyrylcholinesterase (BuChE)
histochemistry (A, D), BuChE immunohistochemistry (B, E) and acetylcholinesterase (AChE) histochemistry (C, F) demonstrating that BuChE
histochemical and immunohistochemical staining methods are comparable, in contrast with AChE histochemistry staining. Scale bars = 200 lmand 25 lm (inset).
58 G. A. Reid et al. / Neuroscience 234 (2013) 53–68
Basal ganglia. BuChE-positive neurons were very
sparse within the basal ganglia with only a few scattered
neurons in the caudate putamen nucleus (CPu) and the
ventral pallidum (VP) (Table 1; Fig. 4A, C). In contrast,
there were numerous ChAT-positive neurons and
neuropil labeling in these regions of the basal ganglia
(Fig. 4B, D), as reported previously (Kitt et al., 1994;
Mesulam et al., 2002a). There were no BuChE/ChAT
double-labeled neurons in this region.
Basal forebrain. Within the basal forebrain there were
scattered BuChE-positive neuronal somata (Table 1,
Fig. 4A) in the medial septal nucleus (MS), substantia
innominata (SI) and vertical (VDB) and horizontal (HDB)
limb nuclei of the diagonal band of Broca. There was
generally sparse BuChE neuropil staining in these
regions except for islands of Calleja (ICj), which had
intense labeling. In contrast, ChAT staining within the
basal forebrain was much more prominent (Table 1;
Fig. 4B), as shown previously (Kitt et al., 1994).
Immunofluorescent double labeling showed that BuChE
and ChAT were located in different populations of
neuronal somata in the basal forebrain.
Amygdala. The amygdala had several nuclei with
BuChE-positive neurons (see Table 1). There were
numerous BuChE-positive neuronal somata in the
anterior amygdaloid area (AA), medial anterior
amygdala (MeA; Fig. 4C) and in the most caudal tip of
the posteromedial cortical amygdala (PMCo). The
remaining nuclei either had a few scattered BuChE-
positive neurons (Fig. 4C), as in the anterior cortical
(ACo), basolateral (BL), basomedial (BM) and lateral
(La) amygdaloid nuclei or were not positive for BuChE,
as in the central amygdala (Ce). ChAT staining within
the amygdala was limited to neuropil (Fig. 4D), as
reported previously (Kitt, 1994). There were no BuChE-
positive neurons stained for ChAT in this region.
Hypothalamus. A number of nuclei of the
hypothalamus contained BuChE-positive neuronal
somata (Table 1; Fig. 4A, C, E) including the medial
supramammillary (SuMM), ventral premammillary
(PMV), lateroanterior hypothalamic (LA), posterior part
of the anterior hypothalamic (AHP), dorsal
premammillary (PMD), lateral hypothalamic (LH),
magnocellular preoptic (MCPO), terete hypothalamic,
submammillothalamic (SMT), lateral preoptic,
paraventricular hypothalamic (Pa), perifornical and
posterior hypothalamic (PH) nuclei (Fig. 4A, C, E).
Distinct BuChE-positive neuropil was labeled in the
SuMM, SMT, LH, PH with sparse staining in the
mammillary nucleus. As observed previously in the
mouse (Bobkova et al., 2001), the MCPO had ChAT-
positive perikarya (Fig. 4B). In addition, other
hypothalamic nuclei had ChAT-positive neuronal somata
and neuropil, including the LH and SuMM (Fig. 4D, F).
There were no neurons within the hypothalamus that
were positive for both BuChE and ChAT labeling.
Thalamus. There was extensive BuChE staining
associated with neuronal somata and neuropil within the
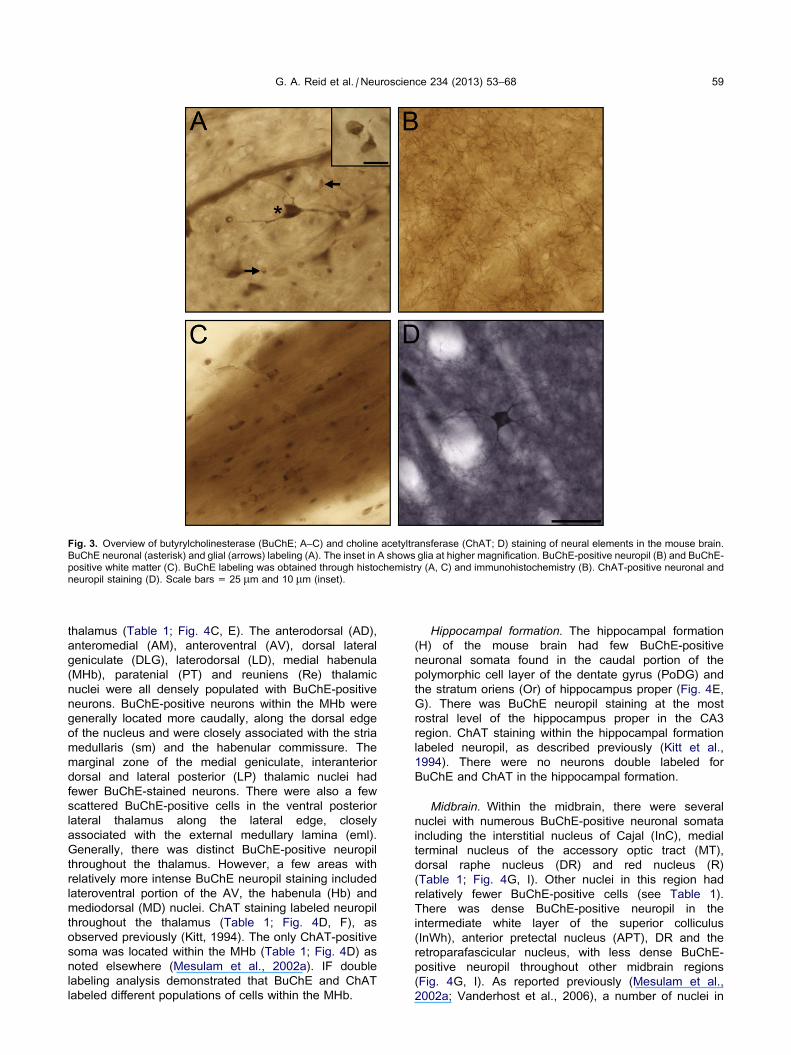
Fig. 3. Overview of butyrylcholinesterase (BuChE; A–C) and choline acetyltransferase (ChAT; D) staining of neural elements in the mouse brain.
BuChE neuronal (asterisk) and glial (arrows) labeling (A). The inset in A shows glia at higher magnification. BuChE-positive neuropil (B) and BuChE-
positive white matter (C). BuChE labeling was obtained through histochemistry (A, C) and immunohistochemistry (B). ChAT-positive neuronal and
neuropil staining (D). Scale bars = 25 lm and 10 lm (inset).
G. A. Reid et al. / Neuroscience 234 (2013) 53–68 59
thalamus (Table 1; Fig. 4C, E). The anterodorsal (AD),
anteromedial (AM), anteroventral (AV), dorsal lateral
geniculate (DLG), laterodorsal (LD), medial habenula
(MHb), paratenial (PT) and reuniens (Re) thalamic
nuclei were all densely populated with BuChE-positive
neurons. BuChE-positive neurons within the MHb were
generally located more caudally, along the dorsal edge
of the nucleus and were closely associated with the stria
medullaris (sm) and the habenular commissure. The
marginal zone of the medial geniculate, interanterior
dorsal and lateral posterior (LP) thalamic nuclei had
fewer BuChE-stained neurons. There were also a few
scattered BuChE-positive cells in the ventral posterior
lateral thalamus along the lateral edge, closely
associated with the external medullary lamina (eml).
Generally, there was distinct BuChE-positive neuropil
throughout the thalamus. However, a few areas with
relatively more intense BuChE neuropil staining included
lateroventral portion of the AV, the habenula (Hb) and
mediodorsal (MD) nuclei. ChAT staining labeled neuropil
throughout the thalamus (Table 1; Fig. 4D, F), as
observed previously (Kitt, 1994). The only ChAT-positive
soma was located within the MHb (Table 1; Fig. 4D) as
noted elsewhere (Mesulam et al., 2002a). IF double
labeling analysis demonstrated that BuChE and ChAT
labeled different populations of cells within the MHb.
Hippocampal formation. The hippocampal formation
(H) of the mouse brain had few BuChE-positive
neuronal somata found in the caudal portion of the
polymorphic cell layer of the dentate gyrus (PoDG) and
the stratum oriens (Or) of hippocampus proper (Fig. 4E,
G). There was BuChE neuropil staining at the most
rostral level of the hippocampus proper in the CA3
region. ChAT staining within the hippocampal formation
labeled neuropil, as described previously (Kitt et al.,
1994). There were no neurons double labeled for
BuChE and ChAT in the hippocampal formation.
Midbrain. Within the midbrain, there were several
nuclei with numerous BuChE-positive neuronal somata
including the interstitial nucleus of Cajal (InC), medial
terminal nucleus of the accessory optic tract (MT),
dorsal raphe nucleus (DR) and red nucleus (R)
(Table 1; Fig. 4G, I). Other nuclei in this region had
relatively fewer BuChE-positive cells (see Table 1).
There was dense BuChE-positive neuropil in the
intermediate white layer of the superior colliculus
(InWh), anterior pretectal nucleus (APT), DR and the
retroparafascicular nucleus, with less dense BuChE-
positive neuropil throughout other midbrain regions
(Fig. 4G, I). As reported previously (Mesulam et al.,
2002a; Vanderhost et al., 2006), a number of nuclei in
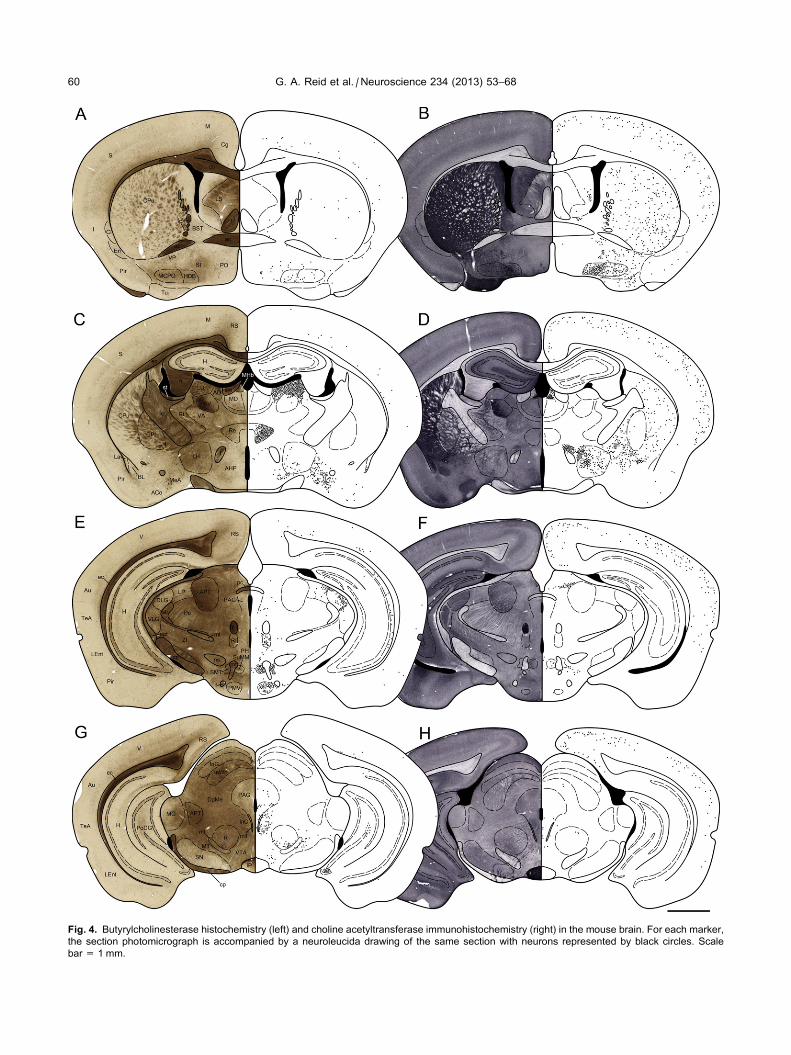
Fig. 4. Butyrylcholinesterase histochemistry (left) and choline acetyltransferase immunohistochemistry (right) in the mouse brain. For each marker,
the section photomicrograph is accompanied by a neuroleucida drawing of the same section with neurons represented by black circles. Scale
bar = 1 mm.
60 G. A. Reid et al. / Neuroscience 234 (2013) 53–68
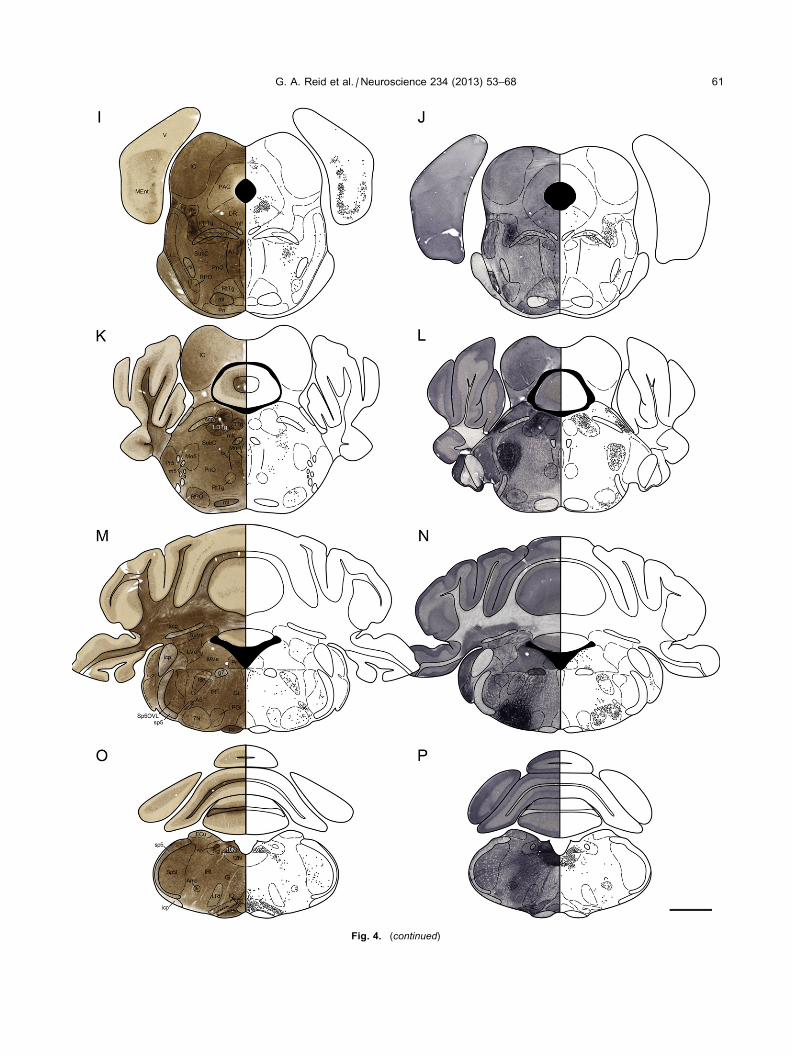
Fig. 4. (continued)
G. A. Reid et al. / Neuroscience 234 (2013) 53–68 61
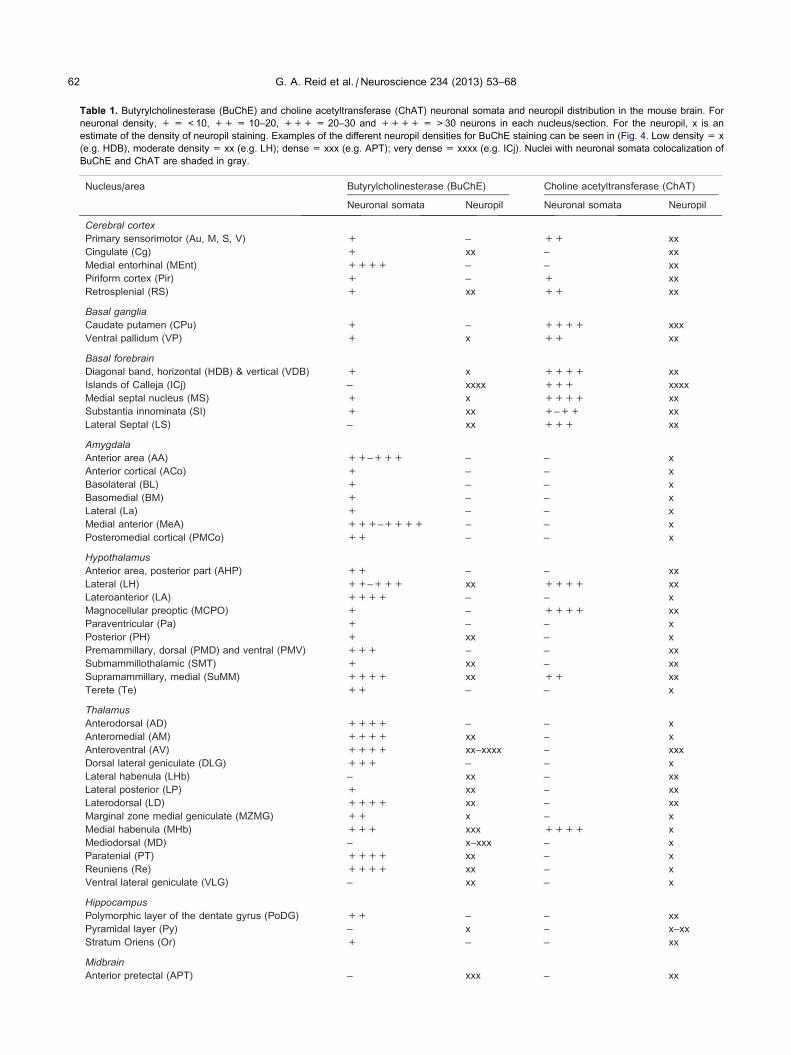
Table 1. Butyrylcholinesterase (BuChE) and choline acetyltransferase (ChAT) neuronal somata and neuropil distribution in the mouse brain. For
neuronal density, + = <10, ++ = 10–20, +++ = 20–30 and ++++ = >30 neurons in each nucleus/section. For the neuropil, x is an
estimate of the density of neuropil staining. Examples of the different neuropil densities for BuChE staining can be seen in (Fig. 4. Low density = x
(e.g. HDB), moderate density = xx (e.g. LH); dense = xxx (e.g. APT); very dense = xxxx (e.g. ICj). Nuclei with neuronal somata colocalization of
BuChE and ChAT are shaded in gray.
62 G. A. Reid et al. / Neuroscience 234 (2013) 53–68
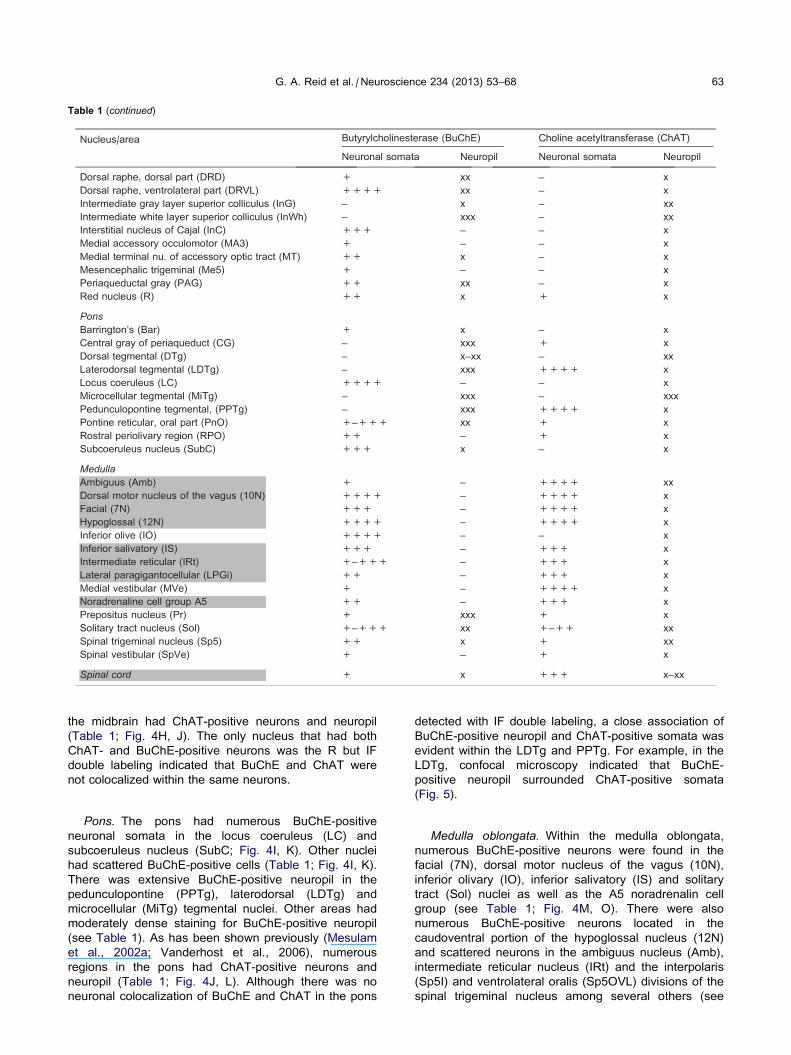
Table 1 (continued)
G. A. Reid et al. / Neuroscience 234 (2013) 53–68 63
the midbrain had ChAT-positive neurons and neuropil
(Table 1; Fig. 4H, J). The only nucleus that had both
ChAT- and BuChE-positive neurons was the R but IF
double labeling indicated that BuChE and ChAT were
not colocalized within the same neurons.
Pons. The pons had numerous BuChE-positive
neuronal somata in the locus coeruleus (LC) and
subcoeruleus nucleus (SubC; Fig. 4I, K). Other nuclei
had scattered BuChE-positive cells (Table 1; Fig. 4I, K).
There was extensive BuChE-positive neuropil in the
pedunculopontine (PPTg), laterodorsal (LDTg) and
microcellular (MiTg) tegmental nuclei. Other areas had
moderately dense staining for BuChE-positive neuropil
(see Table 1). As has been shown previously (Mesulam
et al., 2002a; Vanderhost et al., 2006), numerous
regions in the pons had ChAT-positive neurons and
neuropil (Table 1; Fig. 4J, L). Although there was no
neuronal colocalization of BuChE and ChAT in the pons
detected with IF double labeling, a close association of
BuChE-positive neuropil and ChAT-positive somata was
evident within the LDTg and PPTg. For example, in the
LDTg, confocal microscopy indicated that BuChE-
positive neuropil surrounded ChAT-positive somata
(Fig. 5).
Medulla oblongata. Within the medulla oblongata,
numerous BuChE-positive neurons were found in the
facial (7N), dorsal motor nucleus of the vagus (10N),
inferior olivary (IO), inferior salivatory (IS) and solitary
tract (Sol) nuclei as well as the A5 noradrenalin cell
group (see Table 1; Fig. 4M, O). There were also
numerous BuChE-positive neurons located in the
caudoventral portion of the hypoglossal nucleus (12N)
and scattered neurons in the ambiguus nucleus (Amb),
intermediate reticular nucleus (IRt) and the interpolaris
(Sp5I) and ventrolateral oralis (Sp5OVL) divisions of the
spinal trigeminal nucleus among several others (see
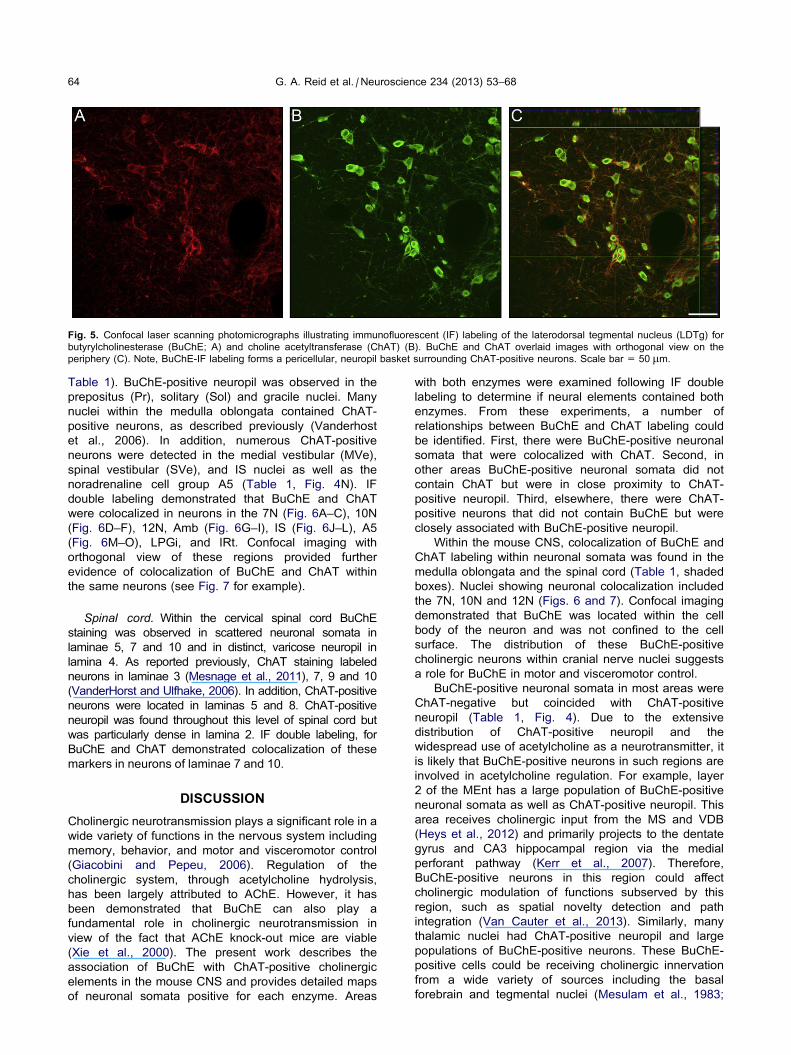
Fig. 5. Confocal laser scanning photomicrographs illustrating immunofluorescent (IF) labeling of the laterodorsal tegmental nucleus (LDTg) for
butyrylcholinesterase (BuChE; A) and choline acetyltransferase (ChAT) (B). BuChE and ChAT overlaid images with orthogonal view on the
periphery (C). Note, BuChE-IF labeling forms a pericellular, neuropil basket surrounding ChAT-positive neurons. Scale bar = 50 lm.
64 G. A. Reid et al. / Neuroscience 234 (2013) 53–68
Table 1). BuChE-positive neuropil was observed in the
prepositus (Pr), solitary (Sol) and gracile nuclei. Many
nuclei within the medulla oblongata contained ChAT-
positive neurons, as described previously (Vanderhost
et al., 2006). In addition, numerous ChAT-positive
neurons were detected in the medial vestibular (MVe),
spinal vestibular (SVe), and IS nuclei as well as the
noradrenaline cell group A5 (Table 1, Fig. 4N). IF
double labeling demonstrated that BuChE and ChAT
were colocalized in neurons in the 7N (Fig. 6A–C), 10N
(Fig. 6D–F), 12N, Amb (Fig. 6G–I), IS (Fig. 6J–L), A5
(Fig. 6M–O), LPGi, and IRt. Confocal imaging with
orthogonal view of these regions provided further
evidence of colocalization of BuChE and ChAT within
the same neurons (see Fig. 7 for example).
Spinal cord. Within the cervical spinal cord BuChE
staining was observed in scattered neuronal somata in
laminae 5, 7 and 10 and in distinct, varicose neuropil in
lamina 4. As reported previously, ChAT staining labeled
neurons in laminae 3 (Mesnage et al., 2011), 7, 9 and 10
(VanderHorst and Ulfhake, 2006). In addition, ChAT-positive
neurons were located in laminas 5 and 8. ChAT-positive
neuropil was found throughout this level of spinal cord but
was particularly dense in lamina 2. IF double labeling, for
BuChE and ChAT demonstrated colocalization of these
markers in neurons of laminae 7 and 10.
DISCUSSION
Cholinergic neurotransmission plays a significant role in a
wide variety of functions in the nervous system including
memory, behavior, and motor and visceromotor control
(Giacobini and Pepeu, 2006). Regulation of the
cholinergic system, through acetylcholine hydrolysis,
has been largely attributed to AChE. However, it has
been demonstrated that BuChE can also play a
fundamental role in cholinergic neurotransmission in
view of the fact that AChE knock-out mice are viable
(Xie et al., 2000). The present work describes the
association of BuChE with ChAT-positive cholinergic
elements in the mouse CNS and provides detailed maps
of neuronal somata positive for each enzyme. Areas
with both enzymes were examined following IF double
labeling to determine if neural elements contained both
enzymes. From these experiments, a number of
relationships between BuChE and ChAT labeling could
be identified. First, there were BuChE-positive neuronal
somata that were colocalized with ChAT. Second, in
other areas BuChE-positive neuronal somata did not
contain ChAT but were in close proximity to ChAT-
positive neuropil. Third, elsewhere, there were ChAT-
positive neurons that did not contain BuChE but were
closely associated with BuChE-positive neuropil.
Within the mouse CNS, colocalization of BuChE and
ChAT labeling within neuronal somata was found in the
medulla oblongata and the spinal cord (Table 1, shaded
boxes). Nuclei showing neuronal colocalization included
the 7N, 10N and 12N (Figs. 6 and 7). Confocal imaging
demonstrated that BuChE was located within the cell
body of the neuron and was not confined to the cell
surface. The distribution of these BuChE-positive
cholinergic neurons within cranial nerve nuclei suggests
a role for BuChE in motor and visceromotor control.
BuChE-positive neuronal somata in most areas were
ChAT-negative but coincided with ChAT-positive
neuropil (Table 1, Fig. 4). Due to the extensive
distribution of ChAT-positive neuropil and the
widespread use of acetylcholine as a neurotransmitter, it
is likely that BuChE-positive neurons in such regions are
involved in acetylcholine regulation. For example, layer
2 of the MEnt has a large population of BuChE-positive
neuronal somata as well as ChAT-positive neuropil. This
area receives cholinergic input from the MS and VDB
(Heys et al., 2012) and primarily projects to the dentate
gyrus and CA3 hippocampal region via the medial
perforant pathway (Kerr et al., 2007). Therefore,
BuChE-positive neurons in this region could affect
cholinergic modulation of functions subserved by this
region, such as spatial novelty detection and path
integration (Van Cauter et al., 2013). Similarly, many
thalamic nuclei had ChAT-positive neuropil and large
populations of BuChE-positive neurons. These BuChE-
positive cells could be receiving cholinergic innervation
from a wide variety of sources including the basal
forebrain and tegmental nuclei (Mesulam et al., 1983;
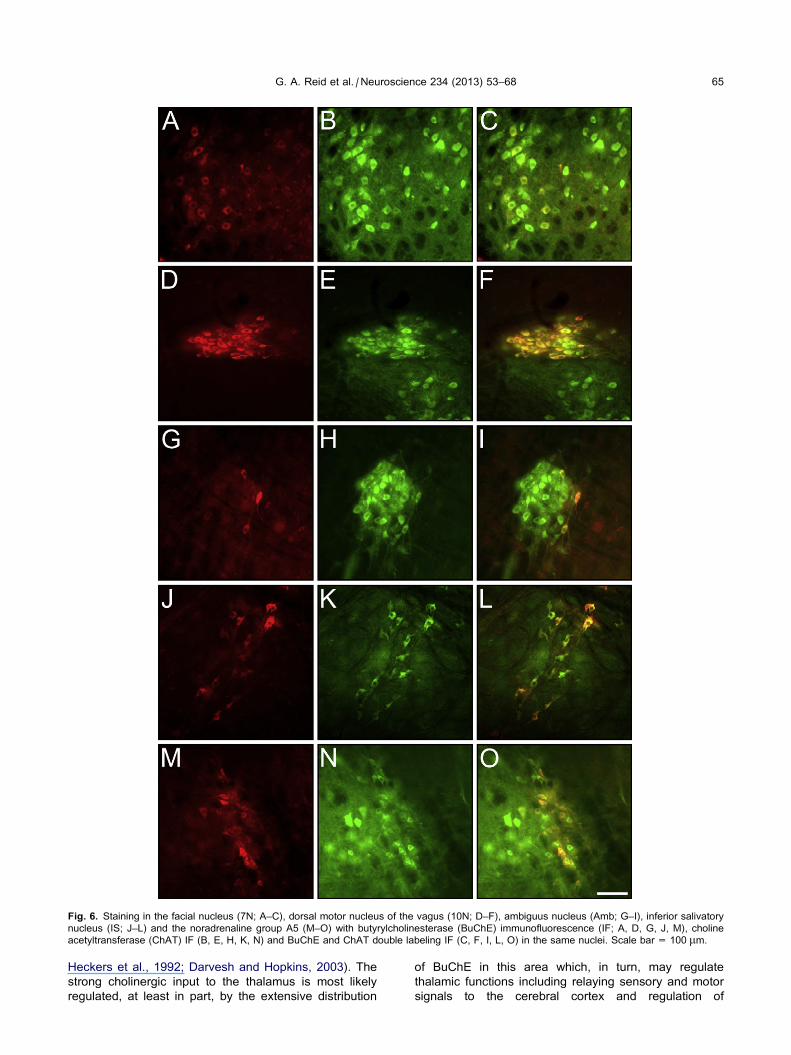
Fig. 6. Staining in the facial nucleus (7N; A–C), dorsal motor nucleus of the vagus (10N; D–F), ambiguus nucleus (Amb; G–I), inferior salivatory
nucleus (IS; J–L) and the noradrenaline group A5 (M–O) with butyrylcholinesterase (BuChE) immunofluorescence (IF; A, D, G, J, M), choline
acetyltransferase (ChAT) IF (B, E, H, K, N) and BuChE and ChAT double labeling IF (C, F, I, L, O) in the same nuclei. Scale bar = 100 lm.
G. A. Reid et al. / Neuroscience 234 (2013) 53–68 65
Heckers et al., 1992; Darvesh and Hopkins, 2003). The
strong cholinergic input to the thalamus is most likely
regulated, at least in part, by the extensive distribution
of BuChE in this area which, in turn, may regulate
thalamic functions including relaying sensory and motor
signals to the cerebral cortex and regulation of
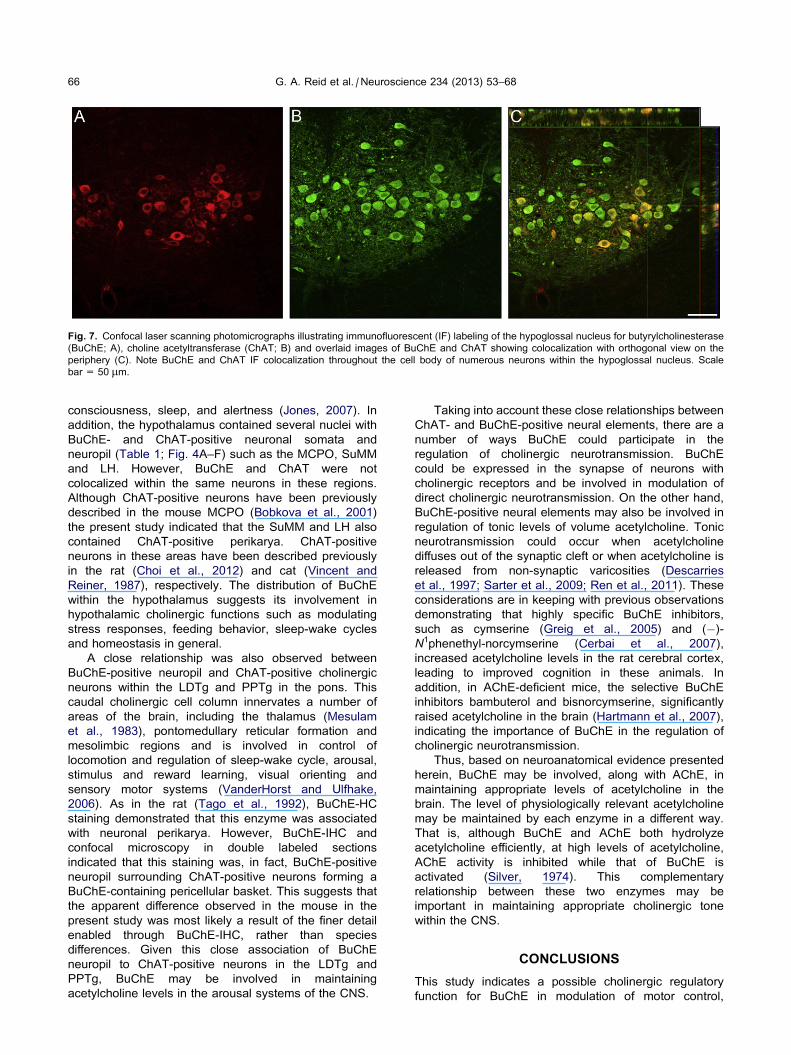
Fig. 7. Confocal laser scanning photomicrographs illustrating immunofluorescent (IF) labeling of the hypoglossal nucleus for butyrylcholinesterase
(BuChE; A), choline acetyltransferase (ChAT; B) and overlaid images of BuChE and ChAT showing colocalization with orthogonal view on the
periphery (C). Note BuChE and ChAT IF colocalization throughout the cell body of numerous neurons within the hypoglossal nucleus. Scale
bar = 50 lm.
66 G. A. Reid et al. / Neuroscience 234 (2013) 53–68
consciousness, sleep, and alertness (Jones, 2007). In
addition, the hypothalamus contained several nuclei with
BuChE- and ChAT-positive neuronal somata and
neuropil (Table 1; Fig. 4A–F) such as the MCPO, SuMM
and LH. However, BuChE and ChAT were not
colocalized within the same neurons in these regions.
Although ChAT-positive neurons have been previously
described in the mouse MCPO (Bobkova et al., 2001)
the present study indicated that the SuMM and LH also
contained ChAT-positive perikarya. ChAT-positive
neurons in these areas have been described previously
in the rat (Choi et al., 2012) and cat (Vincent and
Reiner, 1987), respectively. The distribution of BuChE
within the hypothalamus suggests its involvement in
hypothalamic cholinergic functions such as modulating
stress responses, feeding behavior, sleep-wake cycles
and homeostasis in general.
A close relationship was also observed between
BuChE-positive neuropil and ChAT-positive cholinergic
neurons within the LDTg and PPTg in the pons. This
caudal cholinergic cell column innervates a number of
areas of the brain, including the thalamus (Mesulam
et al., 1983), pontomedullary reticular formation and
mesolimbic regions and is involved in control of
locomotion and regulation of sleep-wake cycle, arousal,
stimulus and reward learning, visual orienting and
sensory motor systems (VanderHorst and Ulfhake,
2006). As in the rat (Tago et al., 1992), BuChE-HC
staining demonstrated that this enzyme was associated
with neuronal perikarya. However, BuChE-IHC and
confocal microscopy in double labeled sections
indicated that this staining was, in fact, BuChE-positive
neuropil surrounding ChAT-positive neurons forming a
BuChE-containing pericellular basket. This suggests that
the apparent difference observed in the mouse in the
present study was most likely a result of the finer detail
enabled through BuChE-IHC, rather than species
differences. Given this close association of BuChE
neuropil to ChAT-positive neurons in the LDTg and
PPTg, BuChE may be involved in maintaining
acetylcholine levels in the arousal systems of the CNS.
Taking into account these close relationships between
ChAT- and BuChE-positive neural elements, there are a
number of ways BuChE could participate in the
regulation of cholinergic neurotransmission. BuChE
could be expressed in the synapse of neurons with
cholinergic receptors and be involved in modulation of
direct cholinergic neurotransmission. On the other hand,
BuChE-positive neural elements may also be involved in
regulation of tonic levels of volume acetylcholine. Tonic
neurotransmission could occur when acetylcholine
diffuses out of the synaptic cleft or when acetylcholine is
released from non-synaptic varicosities (Descarries
et al., 1997; Sarter et al., 2009; Ren et al., 2011). These
considerations are in keeping with previous observations
demonstrating that highly specific BuChE inhibitors,
such as cymserine (Greig et al., 2005) and (�)-N1phenethyl-norcymserine (Cerbai et al., 2007),
increased acetylcholine levels in the rat cerebral cortex,
leading to improved cognition in these animals. In
addition, in AChE-deficient mice, the selective BuChE
inhibitors bambuterol and bisnorcymserine, significantly
raised acetylcholine in the brain (Hartmann et al., 2007),
indicating the importance of BuChE in the regulation of
cholinergic neurotransmission.
Thus, based on neuroanatomical evidence presented
herein, BuChE may be involved, along with AChE, in
maintaining appropriate levels of acetylcholine in the
brain. The level of physiologically relevant acetylcholine
may be maintained by each enzyme in a different way.
That is, although BuChE and AChE both hydrolyze
acetylcholine efficiently, at high levels of acetylcholine,
AChE activity is inhibited while that of BuChE is
activated (Silver, 1974). This complementary
relationship between these two enzymes may be
important in maintaining appropriate cholinergic tone
within the CNS.
CONCLUSIONS
This study indicates a possible cholinergic regulatory
function for BuChE in modulation of motor control,

G. A. Reid et al. / Neuroscience 234 (2013) 53–68 67
awareness, cognition and behavior. Similar to what has been
observed for the distribution of AChE in the brain, not all
BuChE-positive neuronal somata contain ChAT. BuChE
and ChAT colocalization within neurons was found to be
limited to particular nuclei within the medulla oblongata and
spinal cord. However, there were other relationships
evident between these two enzymes, including BuChE-
positive neuronal somata in areas of ChAT neuropil and
BuChE neuropil that formed pericellular baskets around
ChAT-positive, cholinergic neurons.
This work provides further neuroanatomical evidence
that BuChE plays a role in the modulation of cholinergic
neurotransmission.
Acknowledgments—This work was supported in part by Cana-
dian Institutes of Health Research, Capital District Health Author-
ity Research Fund, Nova Scotia Health Research Foundation,
Faculty of Medicine (Clinician-Scientist Program), Dalhousie Uni-
versity Department of Medicine (University Internal Medicine Re-
search Fund) and Dalhousie Medical Research Foundation
(S.D.) and Defense Threat Reduction Agency (DTRA), Joint Sci-
ence and Technology Office, Medical S&T Division (N.C.). We
would like to thank Professor Earl Martin for critically reviewing
this manuscript.
REFERENCES
Armstrong DM, Saper CB, Levey AI, Wainer BH, Terry RD (1983)
Distribution of cholinergic neurons in rat brain: demonstrated by
the immunocytochemical localization of choline acetyltransferase.
J Comp Neurol 216(1):53–68.
Bobkova NV, Nesterova IV, Nesterov VV (2001) The state of
cholinergic structures in forebrain of bulbectomized mice. Bull
Exp Biol Med 131(5):427–431.
Cerbai F, Giovannini MG, Melani C, Enz A, Pepeu G (2007)
N1phenethyl-norcymserine, a selective butyrylcholinesterase
inhibitor, increases acetylcholine release in rat cerebral cortex: a
comparison with donepezil and rivastigmine. Eur J Pharmacol
572(2–3):142–150.
Choi WK, Wirtshafter D, Park HJ, Lee MS, Her S, Shim I (2012) The
characteristics of supramammillary cells projecting to the
hippocampus in stress response in the rat. Korean J Physiol
Pharmacol 16(1):17–24.
Darvesh S, Hopkins DA (2003) Differential distribution of
butyrylcholinesterase and acetylcholinesterase in the human
thalamus. J Comp Neurol 463:25–43.
Darvesh S, Grantham DL, Hopkins DA (1998) Distribution of
butyrylcholinesterase in the human amygdala and hippocampal
formation. J Comp Neurol 393:374–390.
Darvesh S, Hopkins DA, Geula C (2003) Neurobiology of
butyrylcholinesterase. Nat Rev Neurosci 4(2):131–138.
Darvesh S, Cash MK, Reid GA, Martin E, Mitnitski A, Geula C
(2012a) Butyrylcholinesterase is associated with b-amyloid
plaques in the transgenic APPSWE/PSEN1dE9 mouse model of
Alzheimer disease. J Neuropathol Exp Neurol 71(1):2–14.
Darvesh S, Reid GA, Chilukuri N (2012b) Butyrylcholinesterase and
cholinergic neural elements in the mouse brain. Soc Neurosci
Abstr. Program#/Poster#:840.15/B5.
Descarries L, Gisiger V, Steriade M (1997) Diffuse transmission by
acetylcholine in the CNS. Prog Neurobiol 53(5):603–625.
Duysen EG, Li B, Darvesh S, Lockridge O (2007) Sensitivity of
butyrylcholinesterase knockout mice to (�)-huperzine A and
donepezil suggests humans with butyrylcholinesterase
deficiency may not tolerate these Alzheimer’s disease drugs
and indicates butyrylcholinesterase function in
neurotransmission. Toxicology 233(1–3):60–69.
Eckenstein F, Sofroniew MV (1983) Identification of central
cholinergic neurons containing both choline acetyltransferase
and acetylcholinesterase and of central neurons containing only
acetylcholinesterase. J Neurosci 3(11):2286–2291.
Ellman GL, Courtney KD, Andres Jr V, Feather-Stone RM (1961) A
new and rapid colorimetric determination of acetylcholinesterase
activity. Biochem Pharmacol 7:88–95.
Eng LF, Uyeda CT, Chao LP, Wolfgram F (1974) Antibody to bovine
choline acetyltransferase and immunofluorescent localisation of
the enzyme in neurons. Nature 250(463):243–245.
Friede RL (1967) A comparative histochemical mapping of the
distribution of butyryl cholinesterase in the brains of four species
of mammals, including man. Acta Anat (Basel) 66(2):161–177.
Geula C, Nagykery N (2007) Butyrylcholinesterase activity in the rat
forebrain and upper brainstem: postnatal development and adult
distribution. Exp Neurol 204(2):640–657.
Giacobini E (2003) Butyrylcholinesterase: its function and
inhibitors. United Kingdom: Martin Dunitz.
Giacobini E, Pepeu G (2006) The brain cholinergic system: in health
and disease. United Kingdom: Informa Healthcare.
Greig NH, Utsuki T, Ingram DK, Wang Y, Pepeu G, Scali C, Yu QS,
Mamczarz J, Holloway HW, Giordano T, Chen D, Furukawa K,
Sambamurti K, Brossi A, Lahiri DK (2005) Selective
butyrylcholinesterase inhibition elevates brain acetylcholine,
augments learning and lowers Alzheimer beta-amyloid peptide
in rodent. Proc Natl Acad Sci USA 102(47):17213–17218.
Hartmann J, Kiewert C, Duysen EG, Lockridge O, Greig NH, Klein J
(2007) Excessive hippocampal acetylcholine levels in
acetylcholinesterase-deficient mice are moderated by
butyrylcholinesterase activity. J Neurochem 100(5):1421–1429.
Heckers S, Geula C, Mesulam MM (1992) Cholinergic innervation of
the human thalamus: dual origin and differential nuclear
distribution. J Comp Neurol 325(1):68–82.
Heys JG, Schultheiss NW, Shay CF, Tsuno Y, Hasselmo ME (2012)
Effects of acetylcholine on neuronal properties in entorhinal
cortex. Front Behav Neurosci 6(32) (Epub Jul 24).
Jones E (2007) The thalamus. second ed. Cambridge, United
Kingdom: Cambridge University Press.
Karnovsky MJ, Roots L (1964) A direct-coloring thiocholine method
for cholinesterases. J Histochem Cytochem 12:219–221.
Kerr KM, Agster KL, Furtak SC, Burwell RD (2007) Functional
neuroanatomy of the parahippocampal region: the lateral and
medial entorhinal areas. Hippocampus 17(9):697–708.
Kettenmann H, Ransom BR (2005) Neuroglia. New York: Oxford
University Press.
Kimura H, McGeer PL, Peng F, McGeer EG (1980) Choline
acetyltransferase-containing neurons in rodent brain demonstrated by
immunohistochemistry. Science 208(4447):1057–1059.
Kitt CA, Hohmann C, Coyle JT, Price DL (1994) Cholinergic
innervation of mouse forebrain structures. J Comp Neurol
341(1):117–129.
Koelle GB, Friedenwald JA (1949) A histochemical method for
localizing cholinesterase activity. Proc Soc Exp Biol Med
70(4):617–622.
Levey AI, Wainer BH, Mufson EJ, Mesulam MM (1983) Co-
localization of acetylcholinesterase and choline
acetyltransferase in the rat cerebrum. Neuroscience 9(1):9–22.
Levey AI, Wainer BH, Rye DB, Mufson EJ, Mesulam MM (1984)
Choline acetyltransferase-immunoreactive neurons intrinsic to
rodent cortex and distinction from acetylcholinesterase-positive
neurons. Neuroscience 13(2):341–353.
Li B, Stribley JA, Ticu A, Xie W, Schopfer LM, Hammond P, Brimijoin
S, Hinrichs SH, Lockridge O (2000) Abundant tissue
butyrylcholinesterase and its possible function in the
acetylcholinesterase knockout mouse. J Neurochem
75(3):1320–1331.
Mesnage B, Gaillard S, Godin AG, Rodeau JL, Hammer M, Von
Engelhardt J, Wiseman PW, De Koninck Y, Schlichter R,
Cordero-Erausquin M (2011) Morphological and functional
characterization of cholinergic interneurons in the dorsal horn of
the mouse spinal cord. J Comp Neurol 519(16):3139–3158.

68 G. A. Reid et al. / Neuroscience 234 (2013) 53–68
Mesulam MM, Mufson EJ, Wainer BH, Levey AI (1983) Central
cholinergic pathways in the rat: an overview based on an
alternative nomenclature (Ch1–Ch6). Neuroscience
10(4):1185–1201.
Mesulam MM, Guillozet A, Shaw P, Levey A, Duysen EG, Lockridge
O (2002a) Acetylcholinesterase knockouts establish central
cholinergic pathways and can use butyrylcholinesterase to
hydrolyze acetylcholine. Neuroscience 110(4):627–639.
Mesulam M, Guillozet A, Shaw P, Quinn B (2002b) Widely spread
butyrylcholinesterase can hydrolyze acetylcholine in the normal
and Alzheimer brain. Neurobiol Dis 9(1):88–93.
Mis K (2005) Colocalization of acetylcholinesterase,
butyrylcholinesterase and choline acetyltransferase in rat spinal
cord. Hum Exp Toxicol 24(10):543–545.
Parikh K, Duysen EG, Snow B, Jensen NS, Manne V, Lockridge O,
Chilukuri N (2011) Gene-delivered BuChE Is Prophylactic against
the toxicity of chemical warfare nerve agents and
organophosphorus compounds. J Pharmacol Exp Ther
337:92–101.
Paxinos G, Franklin KBJ (2001) The mouse brain in stereotaxic
coordinates. second ed. San Diego: Academic Press.
Ren J, Qin C, Hu F, Tan J, Qiu L, Zhao S, Feng G, Luo M (2011)
Habenula ‘‘cholinergic’’ neurons co-release glutamate and
acetylcholine and activate postsynaptic neurons via distinct
transmission modes. Neuron 69(3):445–452.
Sarter M, Parikh V, Howe WM (2009) Phasic acetylcholine release
and the volume transmission hypothesis: time to move on. Nat
Rev Neurosci 10(5):383–390.
Shute CC, Lewis PR (1963) Cholinesterase-containing systems of
the brain of the rat. Nature 199:1160–1164.
Silver A (1974) The biology of cholinesterases. Amsterdam: Elsevier.
Tago H, Maeda T, McGeer PL, Kimura H (1992)
Butyrylcholinesterase-rich neurons in rat brain demonstrated by
a sensitive histochemical method. J Comp Neurol
325(2):301–312.
Van Cauter T, Camon J, Alvernhe A, Elduayen C, Sargolini F, Save E
(2013) Distinct roles of medial and lateral entorhinal cortex in
spatial cognition. Cereb Cortex 23(2):451–459.
VanderHorst VG, Ulfhake BJ (2006) The organization of the
brainstem and spinal cord of the mouse: relationships between
monoaminergic, cholinergic, and spinal projection systems. Chem
Neuroanat 31(1):2–36.
Vincent SR, Reiner PB (1987) The immunohistochemical localization
of choline acetyltransferase in the cat brain. Brain Res Bull
18(3):371–415.
Xie W, Stribley JA, Chatonnet A, Wilder PJ, Rizzino A, McComb RD,
Taylor P, Hinrichs SH, Lockridge O (2000) Postnatal
developmental delay and supersensitivity to organophosphate in
gene-targeted mice lacking acetylcholinesterase. J Pharmacol
Exp Ther 293(3):896–902.
(Accepted 21 December 2012)(Available online 7 January 2013)