building supertrees: an empirical assessment using the grass family (poaceae
TRANSCRIPT

Syst Biol 51(1)136ndash150 2002
Building Supertrees An Empirical Assessment Using the Grass Family(Poaceae)
NICOLAS SALAMIN12 TREVOR R HODKINSON1 AND VINCENT SAVOLAINEN2
1Department of Botany University of Dublin Trinity College Dublin 2 IrelandE-mail salaminntcdie trevorhodkinsontcdie
2Molecular Systematics Section Jodrell Laboratory Royal Botanic Gardens Kew Richmond Surrey TW9 3DSUnited Kingdom E-mail vsavolainenrbgkeworguk
AbstractmdashLarge and comprehensive phylogenetic trees are desirable for studying macroevolutionaryprocesses and for classication purposes Such trees can be obtained in two different ways Either thewidest possible range of taxa can be sampled and used in a phylogenetic analysis to produce a ldquobigtreerdquo or preexisting topologies can be used to create a supertree Although large multigene analysesare often favored combinable data are not always available and supertrees offer a suitable solutionThe most commonly used method of supertree reconstruction matrix representation with parsimony(MRP) is presented here We used a combined data set for the Poaceae to (1) assess the differencesbetween an approach that uses combined data and one that uses different MRP modications based onthe character partitions and (2) investigate the advantages and disadvantages of these modicationsBaum and Ragan and Purvis modications gave similar results Incorporating bootstrap supportassociated with pre-existing topologies improved Baum and Ragan modication and its similaritywith a combined analysis Finally we used the supertree reconstruction approach on 55 publishedphylogenies to build one of most comprehensive phylogenetic trees published for the grass familyincluding 403 taxa and discuss its strengths and weaknesses in relation to other published hypotheses[Matrix representation with parsimony phylogeny Poaceae supertree]
Comprehensive and well-resolved phylo-genetic trees that contain when possible es-timates of divergence dates underpin com-parative biology and allow powerful testsof a wide range of hypotheses (Felsenstein1985 Harvey and Pagel 1991 Pagel 1999)Phylogenetic trees representing a large sam-ple of taxa are also preferred to those basedon limited numbers of taxa for classica-tion purposes and for studying characterevolution
Two different approaches can be usedto obtain comprehensive phylogenetic treesThe rst uses characters gathered from thewidest possible range of taxa directly in ananalysis to produce a ldquobig treerdquo In this ap-proach phylogenetic analyses of true biolog-ical characters either molecular or pheno-typic (eg morphological) are performedand the meaning of the evolutionary hy-potheses underlining these characters can beinterpreted The second is the meta-analysisapproach (Arnqvist and Wooster 1995) usedin supertree-building methods The underly-ing idea of these methods is to combine thetopologies (or source trees) resulting frommultiple phylogenetic studies (Sandersonet al 1998) rather than their respective bi-ological data sets to produce a supertreeMatrix elements derived directly from thesepublished topologies and for which no real
phylogenetic interpretation can be obtainedrepresent the characters that areused to buildthe supertree or composite phylogenetictree (Bininda-Emonds and Bryant 1998)Although consensus techniques also workontopologies to produce a summarized phylo-genetic tree supertree reconstruction has theadvantageof not requiring identical terminaltaxa sets Only overlapping sets are neededwhich allows the method to produce morecomprehensive phylogenies than the origi-nal ones
A recent theoretical study (Hillis 1996)suggested that large phylogenetic trees canbe easier to analyze than previously thoughtand empirical analyses have demonstratedthat large combined multigene analyses cancorrectly infer large trees (Soltis et al 19992000 Savolainen et al 2000) Because ldquobigtreesrdquo may not be as easy to construct as sug-gested by these studies (Kim 1996 Purvisand Quick 1997a b Yang and Goldman1997) and because combinable data are notalways available the use of composite phy-logenies to study evolutionary patterns isone of the few choices left to the investiga-tors However choosing between the varioussupertree methods is not a straightforwardtask
Two basic algorithms have been devel-oped for supertree reconstruction The strict
136
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 137
supertree reconstructionmethod (Steel 1992Constantinescu and Sankoff 1995) directlyassembles the topologies of the source treesinto a supertree compatible trees (ie treeswithout conicting nodes) are required as in-put Incompatible trees cannot be incorpo-rated in the analysis a problem to which nosolutionhas yet been found (Steel et al 2000)This limitation precludes its use with realdata sets where incompatible nodes reect-ing either real evolutionary divergence be-tween data sets (hard incongruence) or sim-ply random error because of limited samplesize (soft incongruence) areoften the rule be-tween different trees
In contrast matrix representation withparsimony (MRP Baum 1992 Ragan 1992)can be used whether or not the source treesare compatible MRP uses additive binarycoding (Farris et al 1970) to represent the hi-erarchical structure of trees as a series of ma-trix elements (Baum and Ragan 1993) Everynode on each source tree is represented by abinary matrix element and parsimony anal-ysis of the matrix is used to retrieve the treeor trees that represent the hierarchical infor-mation in the source trees If multiple most-parsimonious composite trees are obtaineda strict or semistrict consensus can be used togenerate a consensus composite tree (Baum1992 Ragan 1992)
Several coding procedures have been pro-posed for the MRP method Baum (1992) andRagan (1992) independently rst proposedthat terminal taxa delimited by each nodeshould be coded as 1 in the binary matrixandall other taxaas 0Missing taxa from indi-vidual source trees are then coded as missingvalues (typically ) for the matrix elementsrepresenting these trees Purvis (1995a) ar-gued that the elements derived from sourcetrees lack independence and hence add re-dundant information to the matrix He pro-posed removing this apparent redundancyby allocating the value 0 only to taxa withinthe immediate sister group to the particu-lar clade under consideration and by assign-ing missing values to the other taxa of thesource tree (ie coding them as insteadof0)Subsequently Ronquist (1996) suggestedthat the bias would not be associated withredundant information as stated by Purvis(1995a) but with the relative sizes of thesource trees He argued that the differencein the amount of information contributed
by each source tree could be removed byinversely weighting each tree according tothe number of its nodes However he fa-vored other weighting schemes that werebased on the support for nodes which heargued would also compensate for any sizebias
Bininda-Emonds and Bryant (1998) dis-cussed some of the properties of MRP andinvestigated modications to the methodLittle is known about the merits of suchmodications or how they perform withreal data Bininda-Emonds and Bryant (1998)also discussed the issue that supertrees ob-tained from MRP are not always congruentwith those based on an approach that usescombined data and suggested that differentweighting schemes might help MRP betterapproximate the combined result Further-more matrix elements represent member-ship (character state coded as 1) or lack ofmembership (character state coded as 0) ofa particular taxon relative to a clade Allow-ing reversals in the parsimony analyses cantherefore produce clades in the compositetree that are supported by a lack of mem-bership in some components of the sourcetrees Bininda-Emonds and Bryant (1998) ad-vocated using irreversible character statesin a parsimony analysis to overcome thisshortcoming
In this paper we report the results of anempirical study aimed at assessing the rel-ative merits of the supertree approach us-ing the grass family as a case study We usedthe combined molecular and morphologicaldata set from the Grass Phylogeny WorkingGroup (GPWG 2000) to compare the variousproposed modications of the MRP meth-ods to investigate the effect of irreversiblecharacters on supertree reconstructions andto evaluate the differences between the su-pertrees we obtained and an approach usingcombined data Accurate and meaningfulcomparisons between the combined analy-sis and the supertree approach were madeby breaking down the GPWG combined dataset into its eight character partitions andthen rebuilding a phylogenetic tree by us-ing the various MRP modications We alsoused the same MRP modications to pro-duce different large supertrees that include403 genera from 55 published phylogeniesWe discuss the strengths and weaknessesof each MRP modication obtained in
138 SYSTEMATIC BIOLOGY VOL 51
relation to other evolutionary hypothesesconcerning the grass family
MATERIALS AND METHODS
Combined Phylogenetic Tree
Using maximum parsimony we reana-lyzed the GPWG data set (GPWG 2000)which combined for 61 species of grasseseight different data sets representing molec-ular as well as morphological data anddened well-supported clades according tobootstrap percentages (Felsenstein 1985)One thousand bootstrap replicates were per-formed (with the tree bisectionndashreconnectionswapping algorithm random addition oftaxa and keeping 20 trees at each step)by using PAUPcurren408b (Swofford 1998) TheGPWG matrix was also divided into its eightdata partitions representing three plastidgene sequences (ndhF rbcL and rpoC2) threenuclear DNA regions (gbss1 and phyB genesand the 58Sand internal transcribed spacer 2of the nrDNA hereafter termed ITS) plas-tid restriction site variations and morpho-logical data (Table 1) Parsimony analysesperformed on each character set were fol-lowed by 1000 bootstrap replicates with thesame heuristic search options as previouslydescribed
MRP ReconstructionsTwo different types of MRP analyses
were performed for the grass family Forthe two sets of supertree reconstructionswe built ve binary matrices using the pro-gram SuperTree 085b (available at httpwwwtcdieBotanyNSSuperTreehtml)First the eight different bootstrap trees ob-tained from each data partition of theGPWG matrix were used as source trees
TABLE 1 Details of the eight character partitions ofthe GPWG data set
Number of Number ofData type Genome characters taxa
ndhF plastid 2186 51rbcL plastid 1344 30rpoC2 plastid 855 29phyB nuclear 1182 39gbss1 nuclear 773 14ITS nuclear 424 42Restriction sites plastid 364 45Morphology NA 52 61
for supertree reconstructions [referred tohereafter as GPWG supertree(s)] with thesame 61 terminal taxa as in the GPWG com-bined tree Then we used the informationreported in 55 publications in supertreereconstructions (referred hereafter as largesupertrees) to produce matrices of 403 gen-era (from a total of 635 for the whole familyMabberley 1993) The publications chosendid not represent an exhaustive sample ofthe published literature concerning the grassfamily but that was not the goal of our studyGiven the homogeneity of characters andmethods used in the reviewed publications(indicated with an asterisk in the Referenceslist) we made no distinction between eachof the source trees Our decision followedBininda-Emonds et al (1999) and Purvis(1995b) who found that differential weight-ing of the source trees according to dataor tree search method (or both) had littleimpact on the composite phylogenetic tree
Supertree reconstruction requires an over-lap of taxa sampling between source treesBecause of the large size of the family(about 10000 species) however few taxawere in common between published stud-ies To overcome this problem we con-sidered only generic names in the largeanalyses Species were used when evi-dence against the monophyly of the genuswas demonstrated in the published studyThree taxonomicgroupsmdashpooidspanicoidsand chloridoidsmdashare consistently found asstrong monophyletic groups and no evi-dence has ever contradicted this view (Clarket al 1995 Duvall and Morton 1996Mason-Gamer et al 1998 Gaut et al 1999 Hiluet al 1999 Hsiao et al 1999 Mathews et al2000) To ease the heuristic searches we con-strained our large analyses of the 55 pub-lished phylogenetic trees by forcing eachof these three groups to form three mono-phyletic clades but allowed other taxa to beinserted within them (the ldquobackbonerdquo optionin PAUPcurren408b)
Because bootstrap percentages are miss-ing in most studies before 1993 or becausevalues lt50 are not specied for most ofthe published trees we used a transfor-med function of the bootstrap percentagesto weight the characters into the codedmatrices in both analyses All percent-ages lt50 or nodes with missing val-ues were given the weight of 1 Bootstrap
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 139
percentages cedil50 were weighted by us-ing an exponential transformation (James SFarris pers comm)
e[log(a )pound(b=100)]
where a represents the weight assigned to100 of node support and b representsthe bootstrap percentage This transforma-tion attempts to overcome the conservativebias found in bootstrap percentages IndeedHillis and Bull (1993) using simulationsshowed that bootstrap values gt70 in theirstudy usually indicated with gt95 proba-bility that the corresponding clade was realCorrections to bootstrap estimates have beenproposed (Rodrigo 1993 Zharkikh and Li1995 Effron et al 1996) but these correc-tions cannot compensate for lack of infor-mation in large analyses (Sanderson andWojciechowski 2000) As the number of taxaincreases the relative number of charactersavailable declines and phylogenetic accu-racy suffers Our transformation thus atte-mpts to linearize the bootstrap percent-ages allowing for a gradual increase in thecharacter weights in subsequent parsimonyanalyses
For every supertree reconstructionheuris-tic searches under maximum parsimonywere performed with BaumRagan andPurvis coding schemes alone (hereafterBR-alone and PU-alone respectively) Wealso used bootstrap percentages weight-ing schemes as described above on bothcoding schemes (hereafter BR C bootstrapand PU C bootstrap respectively) and theBaumRagan coding scheme weighted bythe inverse of the number of nodes presentin each source tree (hereafter BR C nodes)We performed 1000 replicates of random ad-dition sequence using the nearest-neighborinterchange swapping algorithm and keep-ing only 20 trees at each replicate Thestored trees were then swapped with thetree bisectionndashreconnection swapping algo-rithm and a maximum of 1000 trees werekept (the ldquomaxtreesrdquo option in PAUPcurren408b)Two different parsimony analyses were per-formed in each of the cases describedabove one considering characters as un-ordered (Fitch parsimony Fitch 1971) andone considering the same characters as ir-reversible (CaminndashSokal parsimony Caminand Sokal 1965) the result was a total of
ten GPWG supertrees and ten large su-pertrees Equally most-parsimonious solu-tions were summarized by using semistrictconsensus
Topological ComparisonsTwo incongruence indices expressed as
distances were used to compare the differ-ent topologies obtained The partition metric(PM symmetric difference in PAUPcurren408bRobinson and Foulds 1981) and the agree-ment subtree metric (D1 Gordon 1980)werecalculated by using PAUPcurren408b (Swofford1998) For the comparisons between thecombined analysis and the GPWG supertreereconstructions we performed KishinondashHasegawa(KH) test (Kishino and Hasegawa1989) and the ShimodairandashHasegawa (SH)test (Shimodaira and Hasegawa 1999) TheGPWG data set consisted of DNA sequencesrestriction site data and morphological char-acters (Table 1) and both tests were per-formed under the maximum parsimony cri-terion Two sets of P-values were thuscalculatedmdashone according to the default op-tions present in PAUPcurren408b (for the KH-test Swofford 1998) the other as describedin Shimodaira and Hasegawa (1999) to en-sure the validity of the test with topolo-gies specied a posteriori The latter testinvolved the creation of 500 bootstrappedreplicates of the GPWG matrix followedby the optimization of 1000 random treesas well as the trees under consideration onthese bootstrapped matrices (for details seeShimodaira and Hasegawa 1999 Goldmanet al 2000)
RESULTS
GPWG Combined TreeThe GPWG combined tree (Fig 1) is
one of the most comprehensive phyloge-netic hypotheses concerning the grass fam-ily Several features can be emphasizedand will serve as a reference for the su-pertree comparisons First and as proposedby GPWG (2000) a large clade (PACCADclade Fig 1) composed of six subfamiliesmdashpanicoids arundinoids chloridoids cen-tothecoids aristidoids and danthonioidsmdashforms a highly supported monophyleticgroup (100 bootstrap Fig 1) No boot-strap values gt52 support any group withinthe PACCAD clade Outside the PACCAD
140 SYSTEMATIC BIOLOGY VOL 51
BEPclade
PACCADclade
early-diverginglineages
outgroup
100
100100
100
100
100
100
100100
100
100
100
100
100
100
100
100
100
100
96
98
76
7787
63
60
86
5268
67
65
9887
98
87
81
91
9292
7890
pooids
bambusoids
ehrhartoids
chloridoids
panicoids +centothecoids
arundinoids
aristidoids +danthonioids
BromusTriticumAvenaBrachypodiumDiarrhenaMelicaGlyceriaNassellaStipaPiptatherumAmpelodesmosPhaenospermaAnisopogonLygeumNardusBrachyelytrumLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaOryzaLeersiaEhrhartaStreptogynaSpartinaSporobolusZoysiaDistichlisEragrostisUniolaPappophorumEriachneMicrairaAmphipogonArundoMoliniaPhragmitesAristidaStipagrostisDanthoniaPennisetumPanicumMiscanthusZeaDanthoniopsis
ChasmanthiumGynerium
ThysanolaenaZeugites
GuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
FIGURE 1 The ldquobigrdquo GPWG tree based on the analysis of eight different data sets (both molecular and morpho-logical) numbers above branches represent bootstrap percentages
clade the bambusoids ehrhartoids andthe pooids form another group called theBEP clade supported by a lower bootstrapvalue (87 bootstrap Fig 1) Finally three
cladesmdashthe anomochloids (Anomochloa Stre-ptochaeta) pharoids (Pharus) and puelioids(Guaduella Puelia)mdashform the basal taxaof thePoaceae (early diverging lineages Fig 1)
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 141
TABLE 2 Partition metrics for the ve MRP modications expressed as distances A) Comparisons withinmodications with unordered characters B) within modications with irreversible characters and C) betweenmodications with unordered characters and modications with irreversible characters
Unordered characters
A) BR-alone BR C bootstrap BR C nodes PU-alone PU C bootstrap
Unordered BR-alone ndash characters BR C bootstrap 0457 ndash
BR C nodes 0379 0388 ndash PU-alone 0440 0362 0422 ndash PU C bootstrap 0474 0379 0457 0362 ndash
B) Irreversible characters
Irreversible BR-alone ndash characters BR C bootstrap 0431 ndash
BR C nodes 0216 0491 ndash PU-alone 0293 0431 0216 ndash PU C bootstrap 0414 0379 0440 0379 ndash
C) Irreversible characters
Unordered BR-alone 0000 0431 0216 0293 0414characters BR C bootstrap 0457 0112 0552 0491 0457
BR C nodes 0379 0483 0319 0328 0483PU-alone 0440 0457 0500 0422 0474PU C bootstrap 0474 0422 0517 0457 0181
GPWG Supertrees
The proportion of nonidentical nodes be-tween the ten MRP reconstructions takenby pairs was calculated by using the PM(Table 2) Analyses using unordered char-acters resulted in topologies with pairwisedistances of between 0362 and 0474 (meandistance 0412 Table 2) Pairwise com-parisons for each MRP modication us-ing irreversible characters gave distances ofbetween 0216 and 0491 (mean distance0369 Table 2) Values for each unorderedBaumRagan modication are closer totheir irreversible counterparts than to anyother methods (mean distance unorderedBRirreversible BR 0328mean distance un-ordered BRirreversible PU 0411 Table 2)The same pattern is found for the un-ordered Purvis modications (mean dis-tance unordered PUirreversible PU 0386mean unordered PUirreversible BR 0468Table 2) Three modications gave very sim-ilar topologies with both types of charac-ters BR-alone gave the exact same topologywhereas the two BR C bootstrap and thetwo PU C bootstrap values were very closeto each other (0112 and 0181 respectivelyTable 2)
For comparisons between the GPWG com-bined tree and the GPWG supertrees D1 dis-tances ranged from 032 for BR C bootstrapwith irreversible characters to 059 for PU-
alone with unordered characters the othereight modications had values from 042 to052 (Fig 2) When considering distancesobtained with PM BR C bootstrap withirreversible characters again gave the topol-ogy the most similar to the GPWG combinedtree yielding a distance of 025 The worstmodication was the BR C nodes with ir-reversible characters for which the distancewas 053 whereas those in the eight othermodications ranged from 041 to 050
We further examined the placement ofthe major subclades dened in the GPWGcombined tree (Fig 3) Two modicationsBR C bootstrap and PU C bootstrap bothwith irreversible characters are the onlymodications to yield the same basalbranching pattern as the combined GPWGtree (Fig 3A) In all the other modicationsinvolving irreversible characters two basalgrasses Guaduella and Puelia are insertedinside the BEP clade BR-alone placed thepooids as sister group to the PACCAD cladewhile BR C nodes and PU-alone placed aclade of the pooids and the ehrhartoids assister group of the PACCAD clade (Fig 3A)With unordered characters and exceptfor the BR-alone modication which wasidentical to the BR-alone with irreversiblecharacters modication all modicationshad an odd basal branching pattern theoutgroup being inserted between two cladesof the early diverging lineages (Fig 3B)
142 SYSTEMATIC BIOLOGY VOL 51
FIGURE 2 D1 greatest subtree agreementmetrics (black) and PM partition metrics (white) expressed asdistancesto the GPWG tree for each of the GPWG supertrees build from separate phylogenies based on the eight characterpartitions of the GPWG combined data set as source trees with characters considered as irreversible (left) andunordered (right)
The two different implementations of theKH-test were used to estimate the validity ofthe ten GPWG supertrees as possible alter-native hypothesis to the GPWG combinedtree (Table 3) Based on the characters ofthe GPWG combined data set the classicalKH-test rejected all the GPWG supertrees(P lt 0001) as suitable alternative phyloge-netic trees However the modied KH-testfound only the PU-alone with unorderedcharacters signicantly different (P lt 005)from the GPWG combined tree (Table 3)None of the other GPWG supertrees couldbe rejected BR C bootstrap with both char-acters types being once again the modica-tion giving the topology closest to that ofthe GPWG combined topology (P D 0588and P D 0624 for unordered and irreversiblecharacters respectively)
Finally Figure 4 shows the supertree ob-tained with BR C bootstrap and irreversiblecharacters The discrepancies between thissupertree and the GPWG combined tree weresmall and concerned mainly single taxa thatwere always positioned close to the cladeto which they belonged in the GPWG tree(Fig 4) The main differences concerned theplacement of the ehrhartoids and bambu-soids which exchanged their position as sis-ter group of the pooids in the supertree incomparison with the GPWG tree and theplacement of the three subfamilies arundi-noids aristidoids and danthonioids
Large SupertreeWe analyzed the 55 publications the same
way as the eight character partitions fromthe GPWG combined data set For clar-ity we do not show the results of BR Cnodes a MRP modication that resultedin topologies incompatible with the place-ment of subfamilies suggested in the GPWGcombined tree When irreversible characterswere used (Fig 5A) BR-alone BR C boot-strap and PU C bootstrap obtained the samebasal branching pattern but the placementof subfamilies inside the PACCAD cladewas slightly different in each case (data notshown) PU-alone was the only modica-tion to place the three subfamilies of theBEP clade together as a sister group to thePACCAD clade agreeing with the GPWG su-pertrees and the GPWG combined tree Withunordered characters (Fig 5B) BR-alone BRC bootstrapand PU-alone obtained the samebasal branching pattern with the BEP cladebeing paraphyletic PU C bootstrap gave anodd combination in which the early diverg-ing lineages were embedded inside the bam-busoids at the base of the tree (Fig 5B)
DISCUSSION
The information given by the two incon-gruence indices helped pinpoint the differ-ences present in the topologies obtained fromthe ten GPWG supertrees built using the
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 143
FIGURE 3 Summary of the subfamilial relationships in the GPWG supertrees obtained with the ve MRP mod-ications with column A) characters considered as irreversible and column B) characters considered as unordered
eight character partitions from the combineddata set Analyses using unordered charac-ters produced supertrees that were less sim-ilar as measured by D1 to the combinedtree than those obtained in analyses usingirreversible characters The only exceptionis for BR-alone which obtained the sametopology with both character types How-ever unordered charactersproduced slightlybetter topologies than irreversible charactersaccording to the PM incongruence index D1is dened as the number of taxa needed tobe removed from both trees to obtain an
identical subtree whereas PM is the numberof taxa bipartitions found between the twotrees being compared (Johnson and Soltis1998) Having higher D1 values than PMvalues suggests that the topological differ-ences do not involve single taxa (in whichcase D1 would be smaller than PM) butrather a large set of taxa placed differentlyin the supertree and the GPWG combinedtree Similar values of D1 and PM wouldsuggest that a few discrepant taxa wereinvolved none being far from their properlocations The D1 distances were much
144 SYSTEMATIC BIOLOGY VOL 51
TABLE 3 Kishino-Hasegawa test based on the difference in length between the GPWG combined tree and the tenGPWG supertrees obtained after applying the different MRP modications The p-values were calculated using thedefault parameter of PAUPcurren408b (Swofford 2000 classic Kishino-Hasegawa [KH] test) and Goldman et al (2000)modication to ensure the validity of the test with a posteriori specied topologies (modied KH test) Asterisk (curren)indicates a signicant p-value (p lt 005)
p-value
Number of steps Classic KH test Modied KH test
Combined GPWG 9054 ndash ndash
Unordered characters BR-alone 9291 lt00001curren 0172BR C bootstrap 9116 lt00001curren 0588BR C nodes 9203 lt00001curren 0396PU-alone 9411 lt00001curren 0042curren
PU C bootstrap 9211 lt00001curren 0374Irreversible characters BR-alone 9291 lt00001curren 0172
BR C bootstrap 9105 00003curren 0624BR C nodes 9281 lt00001curren 0198PU-alone 9311 lt00001curren 0144PU C bootstrap 9191 lt00001curren 0416
greater with unordered than irreversiblecharacters and were much closer to thePM ones obtained with irreversible char-acters Following this logic we can em-phasize that using unordered characters re-sulted in topologies with more differencesfrom the GPWG combined tree as to theplacement of larger subclades than did theuse of irreversible characters The type ofcharacters (ie unordered or irreversible)used to reconstruct the supertrees did notgreatly inuence the placement of the ter-minal taxa in the GPWG supertrees There-fore topologies obtained by consideringmatrix elements as irreversible characterswere closer to the combined phylogenetictree represented by the GPWG combinedanalysis This effect is clearly visible in theplacement of the major subclades dened inthe GPWG combined tree (Fig 4) Unorderedcharacters produced topologies in which theearly diverging lineages were split into twogroupsmdashone including Puelia and Guadu-ella and one including Anomochloa Strep-tochaeta and Pharus This extremely odd andunrealistic pattern does not correspond toany published phylogenetic trees concerningthe grass phylogeny Bininda-Emonds andBryant (1998) found only a minor impactof irreversible characters compared to un-ordered characters in their analysis but wefound that topologies created by using irre-versible characters are closer to an approachusing combined data than are the topolo-gies created by using unordered charactersThe resolution in the large supertrees was ex-tremely dependent on the type of characters
used to perform the analysis such was notthe case with the GPWG supertrees whereboth types of characters gave similar reso-lution (data not shown) Using irreversiblecharacters produced only one or two largesupertrees depending on the modicationused in contrast the ldquomaxtreesrdquo option inPAUPcurren408b was always reached when un-ordered characters were used Of course us-ing irreversible characters should put moreconstraintson the parsimonyanalysis by pre-venting the reversion from state 1 to 0 (thusreducing the number of most-parsimonioustrees) but it is not clear why such a drasticdifference appeared only between the largesupertrees
The Purvis coding scheme has been pro-posed as an improvement to the BaumRagan procedure as an attempt to reducethe dependency and redundancy betweenelements coming from the same topology(Purvis 1995a Ronquist 1996 Bininda-Emonds and Bryant 1998) However our re-sults suggest that the Purvis coding schemedoes not have a large impact on the MRPreconstructions and that the BaumRaganmethod works just as well or even betterComparisons between the approach usingcombined data and BR-alone or combineddata and PU-alone gave similar values forboth PM and D1 (Fig 2) Comparisons be-tween MRP modications also indicated aclose relationship between the two modi-cations especially with irreversible charac-ters (Table 2) This conclusion is less wellsupported with unordered characters be-cause BR-alone is the only modication to
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 145
PACCADclade
BEPclade
early-diverginglineages
outgroup
NassellaStipaPiptatherumAmpelodesmosMelicaGlyceriaBromusTriticumAvenaBrachypodiumDiarrhenaAnisopogonLygeumNardusBrachyelytrumPhaenospermaOryzaLeersiaEhrhartaStreptogynaLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaPennisetumPanicumMiscanthusZeaGyneriumDanthoniopsisZeugitesChasmanthiumThysanolaenaAmphipogonArundoMoliniaPhragmitesSpartinaSporobolusZoysiaDistichlisEragrostisPappophorumUniolaDanthoniaMicrairaAristidaStipagrostisEriachneGuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
pooids
bambusoids
ehrhartoids
chloridoids +danthonioids
panicoids
arundinoids
aristidoids
centothecoids
FIGURE 4 Grass supertree obtained with Baum and Ragan coding scheme with bootstrap support weightingusing separate phylogenies built from the eight character partitions of the GPWG combined data set as source treesDashed lines indicate incongruent branches between the supertree and the GPWG combined tree
146 SYSTEMATIC BIOLOGY VOL 51
FIGURE 5 Summary of the subfamilial relationships in the large supertrees based on 55 published source trees(including 403 genera) obtained using the ve MRP modications with column A) characters considered as irre-versible and column B) characters considered as unordered
give a basal topology very different fromthat obtained with any other modication(Table 2 and Fig 2) Ronquistrsquos (1996) propo-sition to weight each character in the bi-nary matrix by the inverse of the number ofnodes in the corresponding source tree doesnot perform as well as the other modica-tions It yields the greatest partition metricdistance and the second largest agreementsubtree distance (Fig 2) Although the twoalternatives to the BaumRagan method arebased on logical and plain arguments (ienonindependence and redundancy of matrixelements and impact of larger trees) theireffects on MRP reconstruction are not obvi-ous and do not result in topologies closerto our combined reference Indeed PU-alonewith unordered characters was rejected as asuitable alternative hypothesis to the GPWGcombined tree with the modied KH-testand gave the second smallest P-value when
used with irreversible characters (Table 2)However conclusions from the large su-pertree analysis differed because PU-alonewas the only modication to produce thesame basal branching pattern as the GPWGcombined tree (Fig 5B) whereas BR-alonewith the pooids as sister group to the PAC-CAD clade corresponded to an alternativehypothesis for the grass family that is sup-ported by some data from nuclear (Adh Gautet al 1999) and plastid (rbcL Duvall andMorton 1996) genomes Because no refer-ence phylogenetic trees are available thatcontain a number of taxa similar to those ofour large supertrees it is difcult to assesstheir topologies and to determine whetherPU-alone with irreversible characters reallygave a more accurate phylogeny of largegrasses than did the other methods Theplacement of the pooids is possibly not thebest criterion by which to judge the methods
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 147
the evolutionary position of this subfamilywithin the grass family differs as inferredwith different DNA sequence data sets
Weighting by node support as suggestedby Ronquist (1996) improved the t betweenthe additive binary matrices and the GPWGcombined tree The benecial impact of thebootstrap weighting scheme is evident fromthe increased P-values obtained with themodied KH-test (Table 2) This trend is vis-ible for the BaumRagan coding schemebut is less obvious with the Purvis codingscheme (Fig 2) Bootstrap weighting wasproposed (Ronquist 1996) as an alternativeto the Purvis coding scheme both to reducethe greater impact of large source trees oversmalleronesand to improve the effect of well-supported nodes in the MRP analysis It isnot clear how bootstrap weighting could re-duce the impact of larger trees if the smallertrees do not have much greater support val-ues than those of the larger trees Howeverweighting of charactersby bootstrapsupportwithin the Purvis coding scheme appears tobe a redundant procedure unable to greatlyimprove the MRP reconstruction from theimportant information present in bootstrapsupport values (Fig 2) In the large supertreeanalysis PU C bootstrap with unorderedcharacters produced unrealistic topologieswith the early diverging lineages embed-ded in the bambusoids (Fig 5B) The Purviscoding schemes removes important restric-tive information from the matrix (Bininda-Emonds and Bryant 1998) possibly there-fore weighting by bootstrap support wouldrandomly assign high values to characters inthe matrix where this restrictive informationhas been removed and would prevent theweighting schemes from being as effectiveas with the BaumRagan coding schemesWeighting the matrix elements by node sup-port also poses problems Not all bootstrapanalyses can be considered identical and thenumber of replicates the type of searchesdone or both will inuence the supportfound Moreover when node support is notprovided for a source tree weights cannot beassigned and information from this tree willbe down-weighted in the MRP analysis Theeffect of this is great in supertree reconstruc-tions done with older publications which ingeneral do not report node support and arehence down-weighted in the MRP analyses
An important aspect affecting the large su-pertrees that is not resolved in our compar-
isons is the placement of some rarely sam-pled taxa Taxa that are present in only a fewpublications have been allocated a high pro-portion of missing character values whichmakes parsimony analyses much more dif-cult How many of these taxa are mis-placed is difcult to determine because noreference phylogeny for the 403 genera isavailable MRP reconstruction done withoutconstraining three clades (panicoids pooidsand chloridoids) to be monophyletic endedwith rarely sampled taxa being scattered allaround the supertrees (data not shown) Thatcan be an important problem when MRPanalyses are performed on a wide taxonomicgroup comprising large numbers of rarelysampled taxa
MRP coding especially when weighted bynode support can be considered an indica-tion of the signal in the primary data witheach node represented by one (weighted)synapomorphy (see Bininda-Emonds et al1999) However we stress that supertreescannot be viewed and interpreted in ex-actly the same way as phylogenetic analy-ses based on biological characters One ma-jor problem is the difculty of assigningnode support for supertrees Bootstrappingand other resampling procedures cannot beapplied given the clear nonindependenceof the characters present in the binary ma-trix and Bremer support (Bremer 1988) iscertainly equally affected by such noninde-pendence (as is the parsimony itself hencethe different MRP modications proposed)Moreoverbranch lengths associatedwith su-pertrees are treated in the same way as mor-phological branch lengths and unlike molec-ular branch length they cannot be used toestablish divergence times This weakness iseven more signicant in taxonomic groupssuch as the grasses for which dates are dif-cult to gather from other sources (eg fossilrecords)
CONCLUSIONS
We have demonstrated that supertreesoffer an easy way of producing phyloge-nies with a large number of taxa and thatthey can give good estimates of relation-ships within these groups (in this case thegrass family) Supertrees based on the eightcharacter partitions of the GPWG data setswere found to roughly match the combinedanalysis the discrepancies of the best MRP
148 SYSTEMATIC BIOLOGY VOL 51
modications being found mainly in weaklysupported branches of the combined treeThe BaumRagan and the Purvis modica-tions gave similar results but incorporatingbootstrap support associated with preexist-ing topologies improved the BaumRaganmodication Moreover supertrees can beuseful for comparative studies (Purvis1995b Bininda-Emonds et al 1999) whetherof adaptation coevolution rates of evo-lution cospeciation or rates of effectivecladogenesis where accuracy in the branchlength of phylogenetic reconstructions is notthe primary problem (Purvis et al 1994)Supertree reconstructions are also a usefulway to help highlight poor taxonomic sam-pling and identify where previous studiesareinconsistentThey can therefore be used as anexploratory tool capable of developing newhypotheses and indicating where future re-search should be focused
ACKNOWLEDGMENTS
We are grateful to Elizabeth Kellogg and LynnClark for providing the GPWG matrix and for their com-ments on the manuscript James Farris for helpful dis-cussion regarding bootstrap weighting schemes duringthe XIX Hennig Society meeting Stephen Renvoize andMark Chase for helpful discussion on the project OlafBininda-Emonds for his comments on the manuscriptand JonathanDavis for the testing of the SuperTree 085bprogram This study was supported by the Irish HigherEducation Authority the Roche Research Foundation(99-186) Enterprise Iceland (IC200129) the AgassizFoundation and the Swiss NSF (31-52378)
REFERENCES
(References marked curren were used as sources of phyloge-nies for the supertree reconstructions)
ARNQVIST G AND D WOOSTER 1995 Meta-analysisSynthesizing research ndings in ecology and evolu-tion Trends Ecol Evol 10236ndash240
currenBARKER N P 1997 The relationships of AmphipogonElytrophorus and Cyperochloa (Poaceae) as suggestedby rbcL sequence data Telopea 7205ndash213
currenBARKER N P H P LINDER AND E H HARLEY 1995Polyphyly of Arundinoids (Poaceae) Evidence fromrbcL sequence data Syst Bot 20423ndash435
currenBARKER N P H P LINDER and E H HARLEY 1999Sequences of the grass-specic insert in the chloro-plast rpoC2 gene elucidate generic relationships of theArundinoids (Poaceae) Syst Bot 23327ndash350
currenBAUM B R 1982 A phylogenetic analysis of the tribeTriticeae (Poaceae) based on morphological charactersof the genera Can J Bot 61518ndash535
BAUM B R 1992 Combining trees as a way of com-bining data sets for phylogenetic inference and thedesirability of combining gene trees Taxon 413ndash10
currenBAUM B R J R ESTES AND P K GUPTA 1986 Assess-ment of the genomic systematic classication in theTriticeae Am J Bot 741388ndash1395
BAUM B R AND M A RAGAN 1993 Reply to A GRodrigorsquos ldquoA comment on Baumrsquos method for com-bining phylogenetic treesrdquo Taxon 42637ndash640
BININD A-EMONDS O R P AND H N BRYANT 1998Properties of matrix representation with parsimonyanalyses Syst Biol 47497ndash508
BININD A-EMONDS O R P J L GITTLEMAN ANDA PURVIS 1999 Building large trees by combiningphylogenetic information A complete phylogenyof the extant Carnivora (Mammalia) Biol Rev 74143ndash175
BREMER K 1988 The limits of amino acid sequencedata in angiosperm phylogenetic reconstruction Evo-lution 42795ndash803
currenBUCKLER E S IV AND T P HOLTSFORD 1996 Zeasystematics Ribosomal ITS evidence Mol Biol Evol13612ndash622
CAMIN J H AND R R SOKAL 1965 A method for de-ducing branching sequences in phylogeny Evolution19311ndash326
currenCATALAN P E A KELLOGG AND R G OLMS TEAD1997 Phylogeny of Poaceae subfamily Pooideaebased on chloroplast ndhFgene sequences Mol Phy-logenet Evol 8150ndash166
currenCLARK L G W ZHANG AND J F WENDEL 1995 Aphylogenyof the grass family (Poaceae) basedon ndhFsequence data Syst Bot 20436ndash460
CONSTANTINES CU M AND D SANKOFF 1995 An ef-cient algorithm for supertrees J Classif 12101ndash112
currenCUMMINGS M P L M KING AND E A KELLOGG1994 Slipped-strand mispairing in a plastid generpoC2 in grasses (Poaceae) Mol Biol Evol 111ndash8
currenDARBYSHIRE S J AND S I WARWICK 1992 Phylogenyof North American Festuca (Poaceae) and related gen-era using chloroplast DNA restriction site variationCan J Bot 702415ndash2419
currenDAVIS J I AND R J SORENG 1993Phylogenetic struc-ture in the grass family (Poaceae) as inferred fromchloroplast DNA restriction site variation Am J Bot801444ndash1454
currenDOEBLEY J M DURBIN E M GOLENBERG M TCLEGG AND D P MA 1990 Evolutionary analysisof the large subunit of carboxylase (rbcL) nucleotidesequence among the grasses (Gramineae) Evolution441097ndash1108
currenDUVALL M R AND B R MORTON 1996 Molecularphylogenetics of Poaceae An expanded analysis ofrbcL sequence data Mol Phylogenet Evol 5352ndash358
currenDUVALL M R P M PETERSON E E TERRELL ANDA H CHRISTENS EN 1993 Phylogeny of NorthAmerican oryzoid grasses as constructed from mapsof plastid DNA restriction sites Am J Bot 8083ndash88
EFFRON B E HALLORAN AND S HOLMES 1996 Boot-strap condence levels for phylogenetic trees ProcNatl Acad Sci USA 937085ndash7090
currenESEN A AND K W HILU 1991 Electrophoretic andimmunological studies of prolamins in the PoaceaeII Phylogenetic afnities of the Atistideae Taxon 405ndash17
currenESEN A AND K W HILU 1993Prolamin and immuno-logical studies in the Poaceae IV SubfamilyPanicoidsCan J Bot 71315ndash322
FARRIS J S A G KLUGE AND M J ECKHARDT 1970 Anumerical approach to phylogenetic systematics SystZool 19172ndash191
FELSENSTEIN J 1985 Condence limits on phylogeniesAn approach using the bootstrap Evolution 39783ndash791
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 149
FITCH W M 1971 Toward dening the course of evolu-tion Minimal change for a specic tree topology SystZool 20406ndash416
currenFREDERIKSEN S AND O SEBERG 1991 Phylogeneticanalysis of the Triticeae (Poaceae) Hereditas 11515ndash19
currenGAUT B S A SPEEK BR MORTON AND M T CLEGG 1999Patterns of genetic diversication within the Adhgene family in the grasses (Poaceae) Mol Biol Evol161086ndash1097
GOLDMANN N J P ANDERSON AND A G RODRIGO2000 Likelihood-based tests of topologies in phyloge-netics Syst Biol 49652ndash670
GORDON A D 1980 On the assessment and compar-ison of classications Pages 149ndash160 in Analyse dedonnees et informatique (R Tomassone ed) INRALe Chesnay France
GPWG2000A phylogeny of the grass family (Poaceae)as inferred from eight character sets Pages 3ndash7 inGrasses Systematics and evolution (S W L Jacobsand J Everett eds) CSIRO Melbourne Australia
currenGREBENS TEIN B O GREBENS TEIN W SAUER ANDV HEMLEBEN 1996 Distribution and complex organi-zation of satellite DNA sequences in Aveneae speciesGenome 391045ndash1050
currenGREBENS TEIN B M ROSER W SAUER ANDV HEMLEBEN 1998 Molecular phylogenetic re-lationships in Aveneae (Poaceae) species and othergrasses as inferred from ITS1 and ITS2 rDNAsequences Plant Syst Evol 213233ndash250
currenHAMBY R K AND EA ZIMMER 1987Ribosomal RNAsequences for inferring phylogeny within the grassfamily (Poaceae) Plant Syst Evol 16029ndash37
HARVEY P H AND M D PAGEL 1991The comparativemethod in evolutionary biology Oxford Univ PressLondon
HILLIS D M 1996 Inferring complex phylogenies Na-ture 383130ndash131
HILLIS D M AND J J BULL 1993 An empirical test ofbootstrapping as a method for assessing condencein phylogenetic analysis Syst Biol 42182ndash192
currenHILU K W L A ALICE AND H LIANG 1999 Phy-logeny of Poaceae inferred from matK sequencesAnn Mo Bot Gard 86835ndash851
currenHILU K W AND A ESEN 1992Prolamin andimmuno-logical strudies in the Poaceae III Subfamily Chlori-doids Am J Bot 80104ndash113
currenHILU K W AND H LIANG 1997 The matK gene Se-quence variation and application in plant systematicsAm J Bot 84830ndash839
currenHILU K W AND K WRIGHT 1982 Systematics ofGramineae A cluster analysis study Taxon 319ndash37
currenHSIAO C N J CHATTERTON K H ASAY AND K BJENS EN 1993 Phylogenetic relationships of 10 grassspecies An assessment of phylogenetic utility of theinternal transcribed spacer region in nuclear riboso-mal DNA in monocots Genome 37112ndash120
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1994 Phylogenetic relationships of themonogenomic species of the wheat tribe Triticeae(Poaceae) inferred from nuclear rDNA (internal tran-scribed spacer) sequences Genome 38211ndash223
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1995Molecular phylogeny of the pooids(Poaceae) based on nuclear rDNA (ITS) sequencesTheor Appl Genet 90389ndash398
currenHSIAO C S W JACOBS N P BARKER AND N JCHATTERTON 1998 A molecular phylogeny of the
subfamily arundinoids (Poaceae) based on sequencesof rDNA Aust Syst Bot 1141ndash52
currenHSIAO C S W L JACOBS N J CHATTERTON AND KH ASAY 1999 A molecular phylogeny of the grassfamily (Poaceae) based on the sequences of nuclearribosomal DNA (ITS) Aust Syst Bot 11667ndash688
JOHNS ON L A AND D E SOLTIS 1998 Assessing con-gruence Empirical examples from molecular dataPages 297ndash348 in Molecular systematics of plants IIDNA sequencing (D E Soltis P S Soltis and J JDoyle eds) Kluwer Academic Publishers NorwellMassachusetts
currenKATAYAMA H AND Y OGIHARA 1996 Phylogeneticafnities of the grasses to other monocots as re-vealed by molecular analysis of chloroplast DNACurr Genet 29572ndash581
currenKELCHNER S A AND L G CLARK 1997 Molecu-lar evolution and phylogenetic utility of the chloro-plast rpl16 intron in Chusquea and the bambusoids(Poaceae) Mol Phylogenet Evol 8385ndash397
currenKELLOGG E A 1989 Comments on genomic genera inthe Triticeae (Poaceae) Am J Bot 76796ndash805
currenKELLOGG E A 1992Tools for studying the chloroplastgenome in the Triticeae (Gramineae) An ecoRI mapa diagnostic deletion and support for Bromus as anoutgroup Am J Bot 79186ndash197
currenKELLOGG E A 1998aRelationships of cereal crops andother grassesProc NatlAcad Sci USA952005ndash2010
currenKELLOGG E A 1998bWhorsquos related to whom Recentresults from molecular systematic studies Curr OpinPlant Biol 1149ndash158
currenKELLOGG E A AND R APPELS 1995 Intraspecic andinterspecic variation in 5S RNA genes are decoupledin diploid wheat relatives Genetics 140325ndash343
currenKELLOGG E A AND L WATSON 1993 Phyloge-netic studies of a large data set I bambusoids An-dropogonodae and pooids (Gramineae) Bot Rev59273ndash311
KIM J 1996 General inconsistency conditions for max-imum parsimony Effects of branch lengths and in-creasing numbers of taxa Syst Biol 45363ndash374
KISHINO H AND M HASEGAWA 1989 Evaluation ofthe maximum likelihood estimate of the evolution-ary tree topologies form DNA sequence data and thebranching order in Hominoidea J Mol Evol 29170ndash179
currenLIANG H AND K W HILU 1995 Application of thematK gene sequence to grass systematics Can J Bot74125ndash134
MABBERLEY D J 1993 The plant-book A portable dic-tionary of the vascular plants 2nd edition CambridgeUniv Press Cambridge
currenMACFARLANE T D AND L WATSON 1982The classi-cation of Poaceaesubfamily pooids Taxon31178ndash203
currenMASON-GAMER R J AND EA KELLOGG 1996Testingfor phylogenetic conict among molecular data sets inthe tribe Triticeae (Gramineae) Syst Biol 45524ndash545
currenMASON-GAMER R J C F WEIL AND E A KELLOGG1998Granule-bound starch synthase Structure func-tion andphylogenetic utility Mol Biol Evol 151658ndash1673
currenMATHEWS S AND R A SHARROCK 1996 The phy-tochrome gene family in grasses (Poaceae) A phy-logeny and evidence that grasses have a subset ofthe loci found in dicot angiosperms Mol Biol Evol131141ndash1150
currenMATHEWS S R C TSAL AND E A KELLOGG 2000Phylogenetic structure in the grass family (Poaceae)
150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel

2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 137
supertree reconstructionmethod (Steel 1992Constantinescu and Sankoff 1995) directlyassembles the topologies of the source treesinto a supertree compatible trees (ie treeswithout conicting nodes) are required as in-put Incompatible trees cannot be incorpo-rated in the analysis a problem to which nosolutionhas yet been found (Steel et al 2000)This limitation precludes its use with realdata sets where incompatible nodes reect-ing either real evolutionary divergence be-tween data sets (hard incongruence) or sim-ply random error because of limited samplesize (soft incongruence) areoften the rule be-tween different trees
In contrast matrix representation withparsimony (MRP Baum 1992 Ragan 1992)can be used whether or not the source treesare compatible MRP uses additive binarycoding (Farris et al 1970) to represent the hi-erarchical structure of trees as a series of ma-trix elements (Baum and Ragan 1993) Everynode on each source tree is represented by abinary matrix element and parsimony anal-ysis of the matrix is used to retrieve the treeor trees that represent the hierarchical infor-mation in the source trees If multiple most-parsimonious composite trees are obtaineda strict or semistrict consensus can be used togenerate a consensus composite tree (Baum1992 Ragan 1992)
Several coding procedures have been pro-posed for the MRP method Baum (1992) andRagan (1992) independently rst proposedthat terminal taxa delimited by each nodeshould be coded as 1 in the binary matrixandall other taxaas 0Missing taxa from indi-vidual source trees are then coded as missingvalues (typically ) for the matrix elementsrepresenting these trees Purvis (1995a) ar-gued that the elements derived from sourcetrees lack independence and hence add re-dundant information to the matrix He pro-posed removing this apparent redundancyby allocating the value 0 only to taxa withinthe immediate sister group to the particu-lar clade under consideration and by assign-ing missing values to the other taxa of thesource tree (ie coding them as insteadof0)Subsequently Ronquist (1996) suggestedthat the bias would not be associated withredundant information as stated by Purvis(1995a) but with the relative sizes of thesource trees He argued that the differencein the amount of information contributed
by each source tree could be removed byinversely weighting each tree according tothe number of its nodes However he fa-vored other weighting schemes that werebased on the support for nodes which heargued would also compensate for any sizebias
Bininda-Emonds and Bryant (1998) dis-cussed some of the properties of MRP andinvestigated modications to the methodLittle is known about the merits of suchmodications or how they perform withreal data Bininda-Emonds and Bryant (1998)also discussed the issue that supertrees ob-tained from MRP are not always congruentwith those based on an approach that usescombined data and suggested that differentweighting schemes might help MRP betterapproximate the combined result Further-more matrix elements represent member-ship (character state coded as 1) or lack ofmembership (character state coded as 0) ofa particular taxon relative to a clade Allow-ing reversals in the parsimony analyses cantherefore produce clades in the compositetree that are supported by a lack of mem-bership in some components of the sourcetrees Bininda-Emonds and Bryant (1998) ad-vocated using irreversible character statesin a parsimony analysis to overcome thisshortcoming
In this paper we report the results of anempirical study aimed at assessing the rel-ative merits of the supertree approach us-ing the grass family as a case study We usedthe combined molecular and morphologicaldata set from the Grass Phylogeny WorkingGroup (GPWG 2000) to compare the variousproposed modications of the MRP meth-ods to investigate the effect of irreversiblecharacters on supertree reconstructions andto evaluate the differences between the su-pertrees we obtained and an approach usingcombined data Accurate and meaningfulcomparisons between the combined analy-sis and the supertree approach were madeby breaking down the GPWG combined dataset into its eight character partitions andthen rebuilding a phylogenetic tree by us-ing the various MRP modications We alsoused the same MRP modications to pro-duce different large supertrees that include403 genera from 55 published phylogeniesWe discuss the strengths and weaknessesof each MRP modication obtained in
138 SYSTEMATIC BIOLOGY VOL 51
relation to other evolutionary hypothesesconcerning the grass family
MATERIALS AND METHODS
Combined Phylogenetic Tree
Using maximum parsimony we reana-lyzed the GPWG data set (GPWG 2000)which combined for 61 species of grasseseight different data sets representing molec-ular as well as morphological data anddened well-supported clades according tobootstrap percentages (Felsenstein 1985)One thousand bootstrap replicates were per-formed (with the tree bisectionndashreconnectionswapping algorithm random addition oftaxa and keeping 20 trees at each step)by using PAUPcurren408b (Swofford 1998) TheGPWG matrix was also divided into its eightdata partitions representing three plastidgene sequences (ndhF rbcL and rpoC2) threenuclear DNA regions (gbss1 and phyB genesand the 58Sand internal transcribed spacer 2of the nrDNA hereafter termed ITS) plas-tid restriction site variations and morpho-logical data (Table 1) Parsimony analysesperformed on each character set were fol-lowed by 1000 bootstrap replicates with thesame heuristic search options as previouslydescribed
MRP ReconstructionsTwo different types of MRP analyses
were performed for the grass family Forthe two sets of supertree reconstructionswe built ve binary matrices using the pro-gram SuperTree 085b (available at httpwwwtcdieBotanyNSSuperTreehtml)First the eight different bootstrap trees ob-tained from each data partition of theGPWG matrix were used as source trees
TABLE 1 Details of the eight character partitions ofthe GPWG data set
Number of Number ofData type Genome characters taxa
ndhF plastid 2186 51rbcL plastid 1344 30rpoC2 plastid 855 29phyB nuclear 1182 39gbss1 nuclear 773 14ITS nuclear 424 42Restriction sites plastid 364 45Morphology NA 52 61
for supertree reconstructions [referred tohereafter as GPWG supertree(s)] with thesame 61 terminal taxa as in the GPWG com-bined tree Then we used the informationreported in 55 publications in supertreereconstructions (referred hereafter as largesupertrees) to produce matrices of 403 gen-era (from a total of 635 for the whole familyMabberley 1993) The publications chosendid not represent an exhaustive sample ofthe published literature concerning the grassfamily but that was not the goal of our studyGiven the homogeneity of characters andmethods used in the reviewed publications(indicated with an asterisk in the Referenceslist) we made no distinction between eachof the source trees Our decision followedBininda-Emonds et al (1999) and Purvis(1995b) who found that differential weight-ing of the source trees according to dataor tree search method (or both) had littleimpact on the composite phylogenetic tree
Supertree reconstruction requires an over-lap of taxa sampling between source treesBecause of the large size of the family(about 10000 species) however few taxawere in common between published stud-ies To overcome this problem we con-sidered only generic names in the largeanalyses Species were used when evi-dence against the monophyly of the genuswas demonstrated in the published studyThree taxonomicgroupsmdashpooidspanicoidsand chloridoidsmdashare consistently found asstrong monophyletic groups and no evi-dence has ever contradicted this view (Clarket al 1995 Duvall and Morton 1996Mason-Gamer et al 1998 Gaut et al 1999 Hiluet al 1999 Hsiao et al 1999 Mathews et al2000) To ease the heuristic searches we con-strained our large analyses of the 55 pub-lished phylogenetic trees by forcing eachof these three groups to form three mono-phyletic clades but allowed other taxa to beinserted within them (the ldquobackbonerdquo optionin PAUPcurren408b)
Because bootstrap percentages are miss-ing in most studies before 1993 or becausevalues lt50 are not specied for most ofthe published trees we used a transfor-med function of the bootstrap percentagesto weight the characters into the codedmatrices in both analyses All percent-ages lt50 or nodes with missing val-ues were given the weight of 1 Bootstrap
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 139
percentages cedil50 were weighted by us-ing an exponential transformation (James SFarris pers comm)
e[log(a )pound(b=100)]
where a represents the weight assigned to100 of node support and b representsthe bootstrap percentage This transforma-tion attempts to overcome the conservativebias found in bootstrap percentages IndeedHillis and Bull (1993) using simulationsshowed that bootstrap values gt70 in theirstudy usually indicated with gt95 proba-bility that the corresponding clade was realCorrections to bootstrap estimates have beenproposed (Rodrigo 1993 Zharkikh and Li1995 Effron et al 1996) but these correc-tions cannot compensate for lack of infor-mation in large analyses (Sanderson andWojciechowski 2000) As the number of taxaincreases the relative number of charactersavailable declines and phylogenetic accu-racy suffers Our transformation thus atte-mpts to linearize the bootstrap percent-ages allowing for a gradual increase in thecharacter weights in subsequent parsimonyanalyses
For every supertree reconstructionheuris-tic searches under maximum parsimonywere performed with BaumRagan andPurvis coding schemes alone (hereafterBR-alone and PU-alone respectively) Wealso used bootstrap percentages weight-ing schemes as described above on bothcoding schemes (hereafter BR C bootstrapand PU C bootstrap respectively) and theBaumRagan coding scheme weighted bythe inverse of the number of nodes presentin each source tree (hereafter BR C nodes)We performed 1000 replicates of random ad-dition sequence using the nearest-neighborinterchange swapping algorithm and keep-ing only 20 trees at each replicate Thestored trees were then swapped with thetree bisectionndashreconnection swapping algo-rithm and a maximum of 1000 trees werekept (the ldquomaxtreesrdquo option in PAUPcurren408b)Two different parsimony analyses were per-formed in each of the cases describedabove one considering characters as un-ordered (Fitch parsimony Fitch 1971) andone considering the same characters as ir-reversible (CaminndashSokal parsimony Caminand Sokal 1965) the result was a total of
ten GPWG supertrees and ten large su-pertrees Equally most-parsimonious solu-tions were summarized by using semistrictconsensus
Topological ComparisonsTwo incongruence indices expressed as
distances were used to compare the differ-ent topologies obtained The partition metric(PM symmetric difference in PAUPcurren408bRobinson and Foulds 1981) and the agree-ment subtree metric (D1 Gordon 1980)werecalculated by using PAUPcurren408b (Swofford1998) For the comparisons between thecombined analysis and the GPWG supertreereconstructions we performed KishinondashHasegawa(KH) test (Kishino and Hasegawa1989) and the ShimodairandashHasegawa (SH)test (Shimodaira and Hasegawa 1999) TheGPWG data set consisted of DNA sequencesrestriction site data and morphological char-acters (Table 1) and both tests were per-formed under the maximum parsimony cri-terion Two sets of P-values were thuscalculatedmdashone according to the default op-tions present in PAUPcurren408b (for the KH-test Swofford 1998) the other as describedin Shimodaira and Hasegawa (1999) to en-sure the validity of the test with topolo-gies specied a posteriori The latter testinvolved the creation of 500 bootstrappedreplicates of the GPWG matrix followedby the optimization of 1000 random treesas well as the trees under consideration onthese bootstrapped matrices (for details seeShimodaira and Hasegawa 1999 Goldmanet al 2000)
RESULTS
GPWG Combined TreeThe GPWG combined tree (Fig 1) is
one of the most comprehensive phyloge-netic hypotheses concerning the grass fam-ily Several features can be emphasizedand will serve as a reference for the su-pertree comparisons First and as proposedby GPWG (2000) a large clade (PACCADclade Fig 1) composed of six subfamiliesmdashpanicoids arundinoids chloridoids cen-tothecoids aristidoids and danthonioidsmdashforms a highly supported monophyleticgroup (100 bootstrap Fig 1) No boot-strap values gt52 support any group withinthe PACCAD clade Outside the PACCAD
140 SYSTEMATIC BIOLOGY VOL 51
BEPclade
PACCADclade
early-diverginglineages
outgroup
100
100100
100
100
100
100
100100
100
100
100
100
100
100
100
100
100
100
96
98
76
7787
63
60
86
5268
67
65
9887
98
87
81
91
9292
7890
pooids
bambusoids
ehrhartoids
chloridoids
panicoids +centothecoids
arundinoids
aristidoids +danthonioids
BromusTriticumAvenaBrachypodiumDiarrhenaMelicaGlyceriaNassellaStipaPiptatherumAmpelodesmosPhaenospermaAnisopogonLygeumNardusBrachyelytrumLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaOryzaLeersiaEhrhartaStreptogynaSpartinaSporobolusZoysiaDistichlisEragrostisUniolaPappophorumEriachneMicrairaAmphipogonArundoMoliniaPhragmitesAristidaStipagrostisDanthoniaPennisetumPanicumMiscanthusZeaDanthoniopsis
ChasmanthiumGynerium
ThysanolaenaZeugites
GuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
FIGURE 1 The ldquobigrdquo GPWG tree based on the analysis of eight different data sets (both molecular and morpho-logical) numbers above branches represent bootstrap percentages
clade the bambusoids ehrhartoids andthe pooids form another group called theBEP clade supported by a lower bootstrapvalue (87 bootstrap Fig 1) Finally three
cladesmdashthe anomochloids (Anomochloa Stre-ptochaeta) pharoids (Pharus) and puelioids(Guaduella Puelia)mdashform the basal taxaof thePoaceae (early diverging lineages Fig 1)
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 141
TABLE 2 Partition metrics for the ve MRP modications expressed as distances A) Comparisons withinmodications with unordered characters B) within modications with irreversible characters and C) betweenmodications with unordered characters and modications with irreversible characters
Unordered characters
A) BR-alone BR C bootstrap BR C nodes PU-alone PU C bootstrap
Unordered BR-alone ndash characters BR C bootstrap 0457 ndash
BR C nodes 0379 0388 ndash PU-alone 0440 0362 0422 ndash PU C bootstrap 0474 0379 0457 0362 ndash
B) Irreversible characters
Irreversible BR-alone ndash characters BR C bootstrap 0431 ndash
BR C nodes 0216 0491 ndash PU-alone 0293 0431 0216 ndash PU C bootstrap 0414 0379 0440 0379 ndash
C) Irreversible characters
Unordered BR-alone 0000 0431 0216 0293 0414characters BR C bootstrap 0457 0112 0552 0491 0457
BR C nodes 0379 0483 0319 0328 0483PU-alone 0440 0457 0500 0422 0474PU C bootstrap 0474 0422 0517 0457 0181
GPWG Supertrees
The proportion of nonidentical nodes be-tween the ten MRP reconstructions takenby pairs was calculated by using the PM(Table 2) Analyses using unordered char-acters resulted in topologies with pairwisedistances of between 0362 and 0474 (meandistance 0412 Table 2) Pairwise com-parisons for each MRP modication us-ing irreversible characters gave distances ofbetween 0216 and 0491 (mean distance0369 Table 2) Values for each unorderedBaumRagan modication are closer totheir irreversible counterparts than to anyother methods (mean distance unorderedBRirreversible BR 0328mean distance un-ordered BRirreversible PU 0411 Table 2)The same pattern is found for the un-ordered Purvis modications (mean dis-tance unordered PUirreversible PU 0386mean unordered PUirreversible BR 0468Table 2) Three modications gave very sim-ilar topologies with both types of charac-ters BR-alone gave the exact same topologywhereas the two BR C bootstrap and thetwo PU C bootstrap values were very closeto each other (0112 and 0181 respectivelyTable 2)
For comparisons between the GPWG com-bined tree and the GPWG supertrees D1 dis-tances ranged from 032 for BR C bootstrapwith irreversible characters to 059 for PU-
alone with unordered characters the othereight modications had values from 042 to052 (Fig 2) When considering distancesobtained with PM BR C bootstrap withirreversible characters again gave the topol-ogy the most similar to the GPWG combinedtree yielding a distance of 025 The worstmodication was the BR C nodes with ir-reversible characters for which the distancewas 053 whereas those in the eight othermodications ranged from 041 to 050
We further examined the placement ofthe major subclades dened in the GPWGcombined tree (Fig 3) Two modicationsBR C bootstrap and PU C bootstrap bothwith irreversible characters are the onlymodications to yield the same basalbranching pattern as the combined GPWGtree (Fig 3A) In all the other modicationsinvolving irreversible characters two basalgrasses Guaduella and Puelia are insertedinside the BEP clade BR-alone placed thepooids as sister group to the PACCAD cladewhile BR C nodes and PU-alone placed aclade of the pooids and the ehrhartoids assister group of the PACCAD clade (Fig 3A)With unordered characters and exceptfor the BR-alone modication which wasidentical to the BR-alone with irreversiblecharacters modication all modicationshad an odd basal branching pattern theoutgroup being inserted between two cladesof the early diverging lineages (Fig 3B)
142 SYSTEMATIC BIOLOGY VOL 51
FIGURE 2 D1 greatest subtree agreementmetrics (black) and PM partition metrics (white) expressed asdistancesto the GPWG tree for each of the GPWG supertrees build from separate phylogenies based on the eight characterpartitions of the GPWG combined data set as source trees with characters considered as irreversible (left) andunordered (right)
The two different implementations of theKH-test were used to estimate the validity ofthe ten GPWG supertrees as possible alter-native hypothesis to the GPWG combinedtree (Table 3) Based on the characters ofthe GPWG combined data set the classicalKH-test rejected all the GPWG supertrees(P lt 0001) as suitable alternative phyloge-netic trees However the modied KH-testfound only the PU-alone with unorderedcharacters signicantly different (P lt 005)from the GPWG combined tree (Table 3)None of the other GPWG supertrees couldbe rejected BR C bootstrap with both char-acters types being once again the modica-tion giving the topology closest to that ofthe GPWG combined topology (P D 0588and P D 0624 for unordered and irreversiblecharacters respectively)
Finally Figure 4 shows the supertree ob-tained with BR C bootstrap and irreversiblecharacters The discrepancies between thissupertree and the GPWG combined tree weresmall and concerned mainly single taxa thatwere always positioned close to the cladeto which they belonged in the GPWG tree(Fig 4) The main differences concerned theplacement of the ehrhartoids and bambu-soids which exchanged their position as sis-ter group of the pooids in the supertree incomparison with the GPWG tree and theplacement of the three subfamilies arundi-noids aristidoids and danthonioids
Large SupertreeWe analyzed the 55 publications the same
way as the eight character partitions fromthe GPWG combined data set For clar-ity we do not show the results of BR Cnodes a MRP modication that resultedin topologies incompatible with the place-ment of subfamilies suggested in the GPWGcombined tree When irreversible characterswere used (Fig 5A) BR-alone BR C boot-strap and PU C bootstrap obtained the samebasal branching pattern but the placementof subfamilies inside the PACCAD cladewas slightly different in each case (data notshown) PU-alone was the only modica-tion to place the three subfamilies of theBEP clade together as a sister group to thePACCAD clade agreeing with the GPWG su-pertrees and the GPWG combined tree Withunordered characters (Fig 5B) BR-alone BRC bootstrapand PU-alone obtained the samebasal branching pattern with the BEP cladebeing paraphyletic PU C bootstrap gave anodd combination in which the early diverg-ing lineages were embedded inside the bam-busoids at the base of the tree (Fig 5B)
DISCUSSION
The information given by the two incon-gruence indices helped pinpoint the differ-ences present in the topologies obtained fromthe ten GPWG supertrees built using the
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 143
FIGURE 3 Summary of the subfamilial relationships in the GPWG supertrees obtained with the ve MRP mod-ications with column A) characters considered as irreversible and column B) characters considered as unordered
eight character partitions from the combineddata set Analyses using unordered charac-ters produced supertrees that were less sim-ilar as measured by D1 to the combinedtree than those obtained in analyses usingirreversible characters The only exceptionis for BR-alone which obtained the sametopology with both character types How-ever unordered charactersproduced slightlybetter topologies than irreversible charactersaccording to the PM incongruence index D1is dened as the number of taxa needed tobe removed from both trees to obtain an
identical subtree whereas PM is the numberof taxa bipartitions found between the twotrees being compared (Johnson and Soltis1998) Having higher D1 values than PMvalues suggests that the topological differ-ences do not involve single taxa (in whichcase D1 would be smaller than PM) butrather a large set of taxa placed differentlyin the supertree and the GPWG combinedtree Similar values of D1 and PM wouldsuggest that a few discrepant taxa wereinvolved none being far from their properlocations The D1 distances were much
144 SYSTEMATIC BIOLOGY VOL 51
TABLE 3 Kishino-Hasegawa test based on the difference in length between the GPWG combined tree and the tenGPWG supertrees obtained after applying the different MRP modications The p-values were calculated using thedefault parameter of PAUPcurren408b (Swofford 2000 classic Kishino-Hasegawa [KH] test) and Goldman et al (2000)modication to ensure the validity of the test with a posteriori specied topologies (modied KH test) Asterisk (curren)indicates a signicant p-value (p lt 005)
p-value
Number of steps Classic KH test Modied KH test
Combined GPWG 9054 ndash ndash
Unordered characters BR-alone 9291 lt00001curren 0172BR C bootstrap 9116 lt00001curren 0588BR C nodes 9203 lt00001curren 0396PU-alone 9411 lt00001curren 0042curren
PU C bootstrap 9211 lt00001curren 0374Irreversible characters BR-alone 9291 lt00001curren 0172
BR C bootstrap 9105 00003curren 0624BR C nodes 9281 lt00001curren 0198PU-alone 9311 lt00001curren 0144PU C bootstrap 9191 lt00001curren 0416
greater with unordered than irreversiblecharacters and were much closer to thePM ones obtained with irreversible char-acters Following this logic we can em-phasize that using unordered characters re-sulted in topologies with more differencesfrom the GPWG combined tree as to theplacement of larger subclades than did theuse of irreversible characters The type ofcharacters (ie unordered or irreversible)used to reconstruct the supertrees did notgreatly inuence the placement of the ter-minal taxa in the GPWG supertrees There-fore topologies obtained by consideringmatrix elements as irreversible characterswere closer to the combined phylogenetictree represented by the GPWG combinedanalysis This effect is clearly visible in theplacement of the major subclades dened inthe GPWG combined tree (Fig 4) Unorderedcharacters produced topologies in which theearly diverging lineages were split into twogroupsmdashone including Puelia and Guadu-ella and one including Anomochloa Strep-tochaeta and Pharus This extremely odd andunrealistic pattern does not correspond toany published phylogenetic trees concerningthe grass phylogeny Bininda-Emonds andBryant (1998) found only a minor impactof irreversible characters compared to un-ordered characters in their analysis but wefound that topologies created by using irre-versible characters are closer to an approachusing combined data than are the topolo-gies created by using unordered charactersThe resolution in the large supertrees was ex-tremely dependent on the type of characters
used to perform the analysis such was notthe case with the GPWG supertrees whereboth types of characters gave similar reso-lution (data not shown) Using irreversiblecharacters produced only one or two largesupertrees depending on the modicationused in contrast the ldquomaxtreesrdquo option inPAUPcurren408b was always reached when un-ordered characters were used Of course us-ing irreversible characters should put moreconstraintson the parsimonyanalysis by pre-venting the reversion from state 1 to 0 (thusreducing the number of most-parsimonioustrees) but it is not clear why such a drasticdifference appeared only between the largesupertrees
The Purvis coding scheme has been pro-posed as an improvement to the BaumRagan procedure as an attempt to reducethe dependency and redundancy betweenelements coming from the same topology(Purvis 1995a Ronquist 1996 Bininda-Emonds and Bryant 1998) However our re-sults suggest that the Purvis coding schemedoes not have a large impact on the MRPreconstructions and that the BaumRaganmethod works just as well or even betterComparisons between the approach usingcombined data and BR-alone or combineddata and PU-alone gave similar values forboth PM and D1 (Fig 2) Comparisons be-tween MRP modications also indicated aclose relationship between the two modi-cations especially with irreversible charac-ters (Table 2) This conclusion is less wellsupported with unordered characters be-cause BR-alone is the only modication to
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 145
PACCADclade
BEPclade
early-diverginglineages
outgroup
NassellaStipaPiptatherumAmpelodesmosMelicaGlyceriaBromusTriticumAvenaBrachypodiumDiarrhenaAnisopogonLygeumNardusBrachyelytrumPhaenospermaOryzaLeersiaEhrhartaStreptogynaLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaPennisetumPanicumMiscanthusZeaGyneriumDanthoniopsisZeugitesChasmanthiumThysanolaenaAmphipogonArundoMoliniaPhragmitesSpartinaSporobolusZoysiaDistichlisEragrostisPappophorumUniolaDanthoniaMicrairaAristidaStipagrostisEriachneGuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
pooids
bambusoids
ehrhartoids
chloridoids +danthonioids
panicoids
arundinoids
aristidoids
centothecoids
FIGURE 4 Grass supertree obtained with Baum and Ragan coding scheme with bootstrap support weightingusing separate phylogenies built from the eight character partitions of the GPWG combined data set as source treesDashed lines indicate incongruent branches between the supertree and the GPWG combined tree
146 SYSTEMATIC BIOLOGY VOL 51
FIGURE 5 Summary of the subfamilial relationships in the large supertrees based on 55 published source trees(including 403 genera) obtained using the ve MRP modications with column A) characters considered as irre-versible and column B) characters considered as unordered
give a basal topology very different fromthat obtained with any other modication(Table 2 and Fig 2) Ronquistrsquos (1996) propo-sition to weight each character in the bi-nary matrix by the inverse of the number ofnodes in the corresponding source tree doesnot perform as well as the other modica-tions It yields the greatest partition metricdistance and the second largest agreementsubtree distance (Fig 2) Although the twoalternatives to the BaumRagan method arebased on logical and plain arguments (ienonindependence and redundancy of matrixelements and impact of larger trees) theireffects on MRP reconstruction are not obvi-ous and do not result in topologies closerto our combined reference Indeed PU-alonewith unordered characters was rejected as asuitable alternative hypothesis to the GPWGcombined tree with the modied KH-testand gave the second smallest P-value when
used with irreversible characters (Table 2)However conclusions from the large su-pertree analysis differed because PU-alonewas the only modication to produce thesame basal branching pattern as the GPWGcombined tree (Fig 5B) whereas BR-alonewith the pooids as sister group to the PAC-CAD clade corresponded to an alternativehypothesis for the grass family that is sup-ported by some data from nuclear (Adh Gautet al 1999) and plastid (rbcL Duvall andMorton 1996) genomes Because no refer-ence phylogenetic trees are available thatcontain a number of taxa similar to those ofour large supertrees it is difcult to assesstheir topologies and to determine whetherPU-alone with irreversible characters reallygave a more accurate phylogeny of largegrasses than did the other methods Theplacement of the pooids is possibly not thebest criterion by which to judge the methods
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 147
the evolutionary position of this subfamilywithin the grass family differs as inferredwith different DNA sequence data sets
Weighting by node support as suggestedby Ronquist (1996) improved the t betweenthe additive binary matrices and the GPWGcombined tree The benecial impact of thebootstrap weighting scheme is evident fromthe increased P-values obtained with themodied KH-test (Table 2) This trend is vis-ible for the BaumRagan coding schemebut is less obvious with the Purvis codingscheme (Fig 2) Bootstrap weighting wasproposed (Ronquist 1996) as an alternativeto the Purvis coding scheme both to reducethe greater impact of large source trees oversmalleronesand to improve the effect of well-supported nodes in the MRP analysis It isnot clear how bootstrap weighting could re-duce the impact of larger trees if the smallertrees do not have much greater support val-ues than those of the larger trees Howeverweighting of charactersby bootstrapsupportwithin the Purvis coding scheme appears tobe a redundant procedure unable to greatlyimprove the MRP reconstruction from theimportant information present in bootstrapsupport values (Fig 2) In the large supertreeanalysis PU C bootstrap with unorderedcharacters produced unrealistic topologieswith the early diverging lineages embed-ded in the bambusoids (Fig 5B) The Purviscoding schemes removes important restric-tive information from the matrix (Bininda-Emonds and Bryant 1998) possibly there-fore weighting by bootstrap support wouldrandomly assign high values to characters inthe matrix where this restrictive informationhas been removed and would prevent theweighting schemes from being as effectiveas with the BaumRagan coding schemesWeighting the matrix elements by node sup-port also poses problems Not all bootstrapanalyses can be considered identical and thenumber of replicates the type of searchesdone or both will inuence the supportfound Moreover when node support is notprovided for a source tree weights cannot beassigned and information from this tree willbe down-weighted in the MRP analysis Theeffect of this is great in supertree reconstruc-tions done with older publications which ingeneral do not report node support and arehence down-weighted in the MRP analyses
An important aspect affecting the large su-pertrees that is not resolved in our compar-
isons is the placement of some rarely sam-pled taxa Taxa that are present in only a fewpublications have been allocated a high pro-portion of missing character values whichmakes parsimony analyses much more dif-cult How many of these taxa are mis-placed is difcult to determine because noreference phylogeny for the 403 genera isavailable MRP reconstruction done withoutconstraining three clades (panicoids pooidsand chloridoids) to be monophyletic endedwith rarely sampled taxa being scattered allaround the supertrees (data not shown) Thatcan be an important problem when MRPanalyses are performed on a wide taxonomicgroup comprising large numbers of rarelysampled taxa
MRP coding especially when weighted bynode support can be considered an indica-tion of the signal in the primary data witheach node represented by one (weighted)synapomorphy (see Bininda-Emonds et al1999) However we stress that supertreescannot be viewed and interpreted in ex-actly the same way as phylogenetic analy-ses based on biological characters One ma-jor problem is the difculty of assigningnode support for supertrees Bootstrappingand other resampling procedures cannot beapplied given the clear nonindependenceof the characters present in the binary ma-trix and Bremer support (Bremer 1988) iscertainly equally affected by such noninde-pendence (as is the parsimony itself hencethe different MRP modications proposed)Moreoverbranch lengths associatedwith su-pertrees are treated in the same way as mor-phological branch lengths and unlike molec-ular branch length they cannot be used toestablish divergence times This weakness iseven more signicant in taxonomic groupssuch as the grasses for which dates are dif-cult to gather from other sources (eg fossilrecords)
CONCLUSIONS
We have demonstrated that supertreesoffer an easy way of producing phyloge-nies with a large number of taxa and thatthey can give good estimates of relation-ships within these groups (in this case thegrass family) Supertrees based on the eightcharacter partitions of the GPWG data setswere found to roughly match the combinedanalysis the discrepancies of the best MRP
148 SYSTEMATIC BIOLOGY VOL 51
modications being found mainly in weaklysupported branches of the combined treeThe BaumRagan and the Purvis modica-tions gave similar results but incorporatingbootstrap support associated with preexist-ing topologies improved the BaumRaganmodication Moreover supertrees can beuseful for comparative studies (Purvis1995b Bininda-Emonds et al 1999) whetherof adaptation coevolution rates of evo-lution cospeciation or rates of effectivecladogenesis where accuracy in the branchlength of phylogenetic reconstructions is notthe primary problem (Purvis et al 1994)Supertree reconstructions are also a usefulway to help highlight poor taxonomic sam-pling and identify where previous studiesareinconsistentThey can therefore be used as anexploratory tool capable of developing newhypotheses and indicating where future re-search should be focused
ACKNOWLEDGMENTS
We are grateful to Elizabeth Kellogg and LynnClark for providing the GPWG matrix and for their com-ments on the manuscript James Farris for helpful dis-cussion regarding bootstrap weighting schemes duringthe XIX Hennig Society meeting Stephen Renvoize andMark Chase for helpful discussion on the project OlafBininda-Emonds for his comments on the manuscriptand JonathanDavis for the testing of the SuperTree 085bprogram This study was supported by the Irish HigherEducation Authority the Roche Research Foundation(99-186) Enterprise Iceland (IC200129) the AgassizFoundation and the Swiss NSF (31-52378)
REFERENCES
(References marked curren were used as sources of phyloge-nies for the supertree reconstructions)
ARNQVIST G AND D WOOSTER 1995 Meta-analysisSynthesizing research ndings in ecology and evolu-tion Trends Ecol Evol 10236ndash240
currenBARKER N P 1997 The relationships of AmphipogonElytrophorus and Cyperochloa (Poaceae) as suggestedby rbcL sequence data Telopea 7205ndash213
currenBARKER N P H P LINDER AND E H HARLEY 1995Polyphyly of Arundinoids (Poaceae) Evidence fromrbcL sequence data Syst Bot 20423ndash435
currenBARKER N P H P LINDER and E H HARLEY 1999Sequences of the grass-specic insert in the chloro-plast rpoC2 gene elucidate generic relationships of theArundinoids (Poaceae) Syst Bot 23327ndash350
currenBAUM B R 1982 A phylogenetic analysis of the tribeTriticeae (Poaceae) based on morphological charactersof the genera Can J Bot 61518ndash535
BAUM B R 1992 Combining trees as a way of com-bining data sets for phylogenetic inference and thedesirability of combining gene trees Taxon 413ndash10
currenBAUM B R J R ESTES AND P K GUPTA 1986 Assess-ment of the genomic systematic classication in theTriticeae Am J Bot 741388ndash1395
BAUM B R AND M A RAGAN 1993 Reply to A GRodrigorsquos ldquoA comment on Baumrsquos method for com-bining phylogenetic treesrdquo Taxon 42637ndash640
BININD A-EMONDS O R P AND H N BRYANT 1998Properties of matrix representation with parsimonyanalyses Syst Biol 47497ndash508
BININD A-EMONDS O R P J L GITTLEMAN ANDA PURVIS 1999 Building large trees by combiningphylogenetic information A complete phylogenyof the extant Carnivora (Mammalia) Biol Rev 74143ndash175
BREMER K 1988 The limits of amino acid sequencedata in angiosperm phylogenetic reconstruction Evo-lution 42795ndash803
currenBUCKLER E S IV AND T P HOLTSFORD 1996 Zeasystematics Ribosomal ITS evidence Mol Biol Evol13612ndash622
CAMIN J H AND R R SOKAL 1965 A method for de-ducing branching sequences in phylogeny Evolution19311ndash326
currenCATALAN P E A KELLOGG AND R G OLMS TEAD1997 Phylogeny of Poaceae subfamily Pooideaebased on chloroplast ndhFgene sequences Mol Phy-logenet Evol 8150ndash166
currenCLARK L G W ZHANG AND J F WENDEL 1995 Aphylogenyof the grass family (Poaceae) basedon ndhFsequence data Syst Bot 20436ndash460
CONSTANTINES CU M AND D SANKOFF 1995 An ef-cient algorithm for supertrees J Classif 12101ndash112
currenCUMMINGS M P L M KING AND E A KELLOGG1994 Slipped-strand mispairing in a plastid generpoC2 in grasses (Poaceae) Mol Biol Evol 111ndash8
currenDARBYSHIRE S J AND S I WARWICK 1992 Phylogenyof North American Festuca (Poaceae) and related gen-era using chloroplast DNA restriction site variationCan J Bot 702415ndash2419
currenDAVIS J I AND R J SORENG 1993Phylogenetic struc-ture in the grass family (Poaceae) as inferred fromchloroplast DNA restriction site variation Am J Bot801444ndash1454
currenDOEBLEY J M DURBIN E M GOLENBERG M TCLEGG AND D P MA 1990 Evolutionary analysisof the large subunit of carboxylase (rbcL) nucleotidesequence among the grasses (Gramineae) Evolution441097ndash1108
currenDUVALL M R AND B R MORTON 1996 Molecularphylogenetics of Poaceae An expanded analysis ofrbcL sequence data Mol Phylogenet Evol 5352ndash358
currenDUVALL M R P M PETERSON E E TERRELL ANDA H CHRISTENS EN 1993 Phylogeny of NorthAmerican oryzoid grasses as constructed from mapsof plastid DNA restriction sites Am J Bot 8083ndash88
EFFRON B E HALLORAN AND S HOLMES 1996 Boot-strap condence levels for phylogenetic trees ProcNatl Acad Sci USA 937085ndash7090
currenESEN A AND K W HILU 1991 Electrophoretic andimmunological studies of prolamins in the PoaceaeII Phylogenetic afnities of the Atistideae Taxon 405ndash17
currenESEN A AND K W HILU 1993Prolamin and immuno-logical studies in the Poaceae IV SubfamilyPanicoidsCan J Bot 71315ndash322
FARRIS J S A G KLUGE AND M J ECKHARDT 1970 Anumerical approach to phylogenetic systematics SystZool 19172ndash191
FELSENSTEIN J 1985 Condence limits on phylogeniesAn approach using the bootstrap Evolution 39783ndash791
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 149
FITCH W M 1971 Toward dening the course of evolu-tion Minimal change for a specic tree topology SystZool 20406ndash416
currenFREDERIKSEN S AND O SEBERG 1991 Phylogeneticanalysis of the Triticeae (Poaceae) Hereditas 11515ndash19
currenGAUT B S A SPEEK BR MORTON AND M T CLEGG 1999Patterns of genetic diversication within the Adhgene family in the grasses (Poaceae) Mol Biol Evol161086ndash1097
GOLDMANN N J P ANDERSON AND A G RODRIGO2000 Likelihood-based tests of topologies in phyloge-netics Syst Biol 49652ndash670
GORDON A D 1980 On the assessment and compar-ison of classications Pages 149ndash160 in Analyse dedonnees et informatique (R Tomassone ed) INRALe Chesnay France
GPWG2000A phylogeny of the grass family (Poaceae)as inferred from eight character sets Pages 3ndash7 inGrasses Systematics and evolution (S W L Jacobsand J Everett eds) CSIRO Melbourne Australia
currenGREBENS TEIN B O GREBENS TEIN W SAUER ANDV HEMLEBEN 1996 Distribution and complex organi-zation of satellite DNA sequences in Aveneae speciesGenome 391045ndash1050
currenGREBENS TEIN B M ROSER W SAUER ANDV HEMLEBEN 1998 Molecular phylogenetic re-lationships in Aveneae (Poaceae) species and othergrasses as inferred from ITS1 and ITS2 rDNAsequences Plant Syst Evol 213233ndash250
currenHAMBY R K AND EA ZIMMER 1987Ribosomal RNAsequences for inferring phylogeny within the grassfamily (Poaceae) Plant Syst Evol 16029ndash37
HARVEY P H AND M D PAGEL 1991The comparativemethod in evolutionary biology Oxford Univ PressLondon
HILLIS D M 1996 Inferring complex phylogenies Na-ture 383130ndash131
HILLIS D M AND J J BULL 1993 An empirical test ofbootstrapping as a method for assessing condencein phylogenetic analysis Syst Biol 42182ndash192
currenHILU K W L A ALICE AND H LIANG 1999 Phy-logeny of Poaceae inferred from matK sequencesAnn Mo Bot Gard 86835ndash851
currenHILU K W AND A ESEN 1992Prolamin andimmuno-logical strudies in the Poaceae III Subfamily Chlori-doids Am J Bot 80104ndash113
currenHILU K W AND H LIANG 1997 The matK gene Se-quence variation and application in plant systematicsAm J Bot 84830ndash839
currenHILU K W AND K WRIGHT 1982 Systematics ofGramineae A cluster analysis study Taxon 319ndash37
currenHSIAO C N J CHATTERTON K H ASAY AND K BJENS EN 1993 Phylogenetic relationships of 10 grassspecies An assessment of phylogenetic utility of theinternal transcribed spacer region in nuclear riboso-mal DNA in monocots Genome 37112ndash120
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1994 Phylogenetic relationships of themonogenomic species of the wheat tribe Triticeae(Poaceae) inferred from nuclear rDNA (internal tran-scribed spacer) sequences Genome 38211ndash223
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1995Molecular phylogeny of the pooids(Poaceae) based on nuclear rDNA (ITS) sequencesTheor Appl Genet 90389ndash398
currenHSIAO C S W JACOBS N P BARKER AND N JCHATTERTON 1998 A molecular phylogeny of the
subfamily arundinoids (Poaceae) based on sequencesof rDNA Aust Syst Bot 1141ndash52
currenHSIAO C S W L JACOBS N J CHATTERTON AND KH ASAY 1999 A molecular phylogeny of the grassfamily (Poaceae) based on the sequences of nuclearribosomal DNA (ITS) Aust Syst Bot 11667ndash688
JOHNS ON L A AND D E SOLTIS 1998 Assessing con-gruence Empirical examples from molecular dataPages 297ndash348 in Molecular systematics of plants IIDNA sequencing (D E Soltis P S Soltis and J JDoyle eds) Kluwer Academic Publishers NorwellMassachusetts
currenKATAYAMA H AND Y OGIHARA 1996 Phylogeneticafnities of the grasses to other monocots as re-vealed by molecular analysis of chloroplast DNACurr Genet 29572ndash581
currenKELCHNER S A AND L G CLARK 1997 Molecu-lar evolution and phylogenetic utility of the chloro-plast rpl16 intron in Chusquea and the bambusoids(Poaceae) Mol Phylogenet Evol 8385ndash397
currenKELLOGG E A 1989 Comments on genomic genera inthe Triticeae (Poaceae) Am J Bot 76796ndash805
currenKELLOGG E A 1992Tools for studying the chloroplastgenome in the Triticeae (Gramineae) An ecoRI mapa diagnostic deletion and support for Bromus as anoutgroup Am J Bot 79186ndash197
currenKELLOGG E A 1998aRelationships of cereal crops andother grassesProc NatlAcad Sci USA952005ndash2010
currenKELLOGG E A 1998bWhorsquos related to whom Recentresults from molecular systematic studies Curr OpinPlant Biol 1149ndash158
currenKELLOGG E A AND R APPELS 1995 Intraspecic andinterspecic variation in 5S RNA genes are decoupledin diploid wheat relatives Genetics 140325ndash343
currenKELLOGG E A AND L WATSON 1993 Phyloge-netic studies of a large data set I bambusoids An-dropogonodae and pooids (Gramineae) Bot Rev59273ndash311
KIM J 1996 General inconsistency conditions for max-imum parsimony Effects of branch lengths and in-creasing numbers of taxa Syst Biol 45363ndash374
KISHINO H AND M HASEGAWA 1989 Evaluation ofthe maximum likelihood estimate of the evolution-ary tree topologies form DNA sequence data and thebranching order in Hominoidea J Mol Evol 29170ndash179
currenLIANG H AND K W HILU 1995 Application of thematK gene sequence to grass systematics Can J Bot74125ndash134
MABBERLEY D J 1993 The plant-book A portable dic-tionary of the vascular plants 2nd edition CambridgeUniv Press Cambridge
currenMACFARLANE T D AND L WATSON 1982The classi-cation of Poaceaesubfamily pooids Taxon31178ndash203
currenMASON-GAMER R J AND EA KELLOGG 1996Testingfor phylogenetic conict among molecular data sets inthe tribe Triticeae (Gramineae) Syst Biol 45524ndash545
currenMASON-GAMER R J C F WEIL AND E A KELLOGG1998Granule-bound starch synthase Structure func-tion andphylogenetic utility Mol Biol Evol 151658ndash1673
currenMATHEWS S AND R A SHARROCK 1996 The phy-tochrome gene family in grasses (Poaceae) A phy-logeny and evidence that grasses have a subset ofthe loci found in dicot angiosperms Mol Biol Evol131141ndash1150
currenMATHEWS S R C TSAL AND E A KELLOGG 2000Phylogenetic structure in the grass family (Poaceae)
150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel

138 SYSTEMATIC BIOLOGY VOL 51
relation to other evolutionary hypothesesconcerning the grass family
MATERIALS AND METHODS
Combined Phylogenetic Tree
Using maximum parsimony we reana-lyzed the GPWG data set (GPWG 2000)which combined for 61 species of grasseseight different data sets representing molec-ular as well as morphological data anddened well-supported clades according tobootstrap percentages (Felsenstein 1985)One thousand bootstrap replicates were per-formed (with the tree bisectionndashreconnectionswapping algorithm random addition oftaxa and keeping 20 trees at each step)by using PAUPcurren408b (Swofford 1998) TheGPWG matrix was also divided into its eightdata partitions representing three plastidgene sequences (ndhF rbcL and rpoC2) threenuclear DNA regions (gbss1 and phyB genesand the 58Sand internal transcribed spacer 2of the nrDNA hereafter termed ITS) plas-tid restriction site variations and morpho-logical data (Table 1) Parsimony analysesperformed on each character set were fol-lowed by 1000 bootstrap replicates with thesame heuristic search options as previouslydescribed
MRP ReconstructionsTwo different types of MRP analyses
were performed for the grass family Forthe two sets of supertree reconstructionswe built ve binary matrices using the pro-gram SuperTree 085b (available at httpwwwtcdieBotanyNSSuperTreehtml)First the eight different bootstrap trees ob-tained from each data partition of theGPWG matrix were used as source trees
TABLE 1 Details of the eight character partitions ofthe GPWG data set
Number of Number ofData type Genome characters taxa
ndhF plastid 2186 51rbcL plastid 1344 30rpoC2 plastid 855 29phyB nuclear 1182 39gbss1 nuclear 773 14ITS nuclear 424 42Restriction sites plastid 364 45Morphology NA 52 61
for supertree reconstructions [referred tohereafter as GPWG supertree(s)] with thesame 61 terminal taxa as in the GPWG com-bined tree Then we used the informationreported in 55 publications in supertreereconstructions (referred hereafter as largesupertrees) to produce matrices of 403 gen-era (from a total of 635 for the whole familyMabberley 1993) The publications chosendid not represent an exhaustive sample ofthe published literature concerning the grassfamily but that was not the goal of our studyGiven the homogeneity of characters andmethods used in the reviewed publications(indicated with an asterisk in the Referenceslist) we made no distinction between eachof the source trees Our decision followedBininda-Emonds et al (1999) and Purvis(1995b) who found that differential weight-ing of the source trees according to dataor tree search method (or both) had littleimpact on the composite phylogenetic tree
Supertree reconstruction requires an over-lap of taxa sampling between source treesBecause of the large size of the family(about 10000 species) however few taxawere in common between published stud-ies To overcome this problem we con-sidered only generic names in the largeanalyses Species were used when evi-dence against the monophyly of the genuswas demonstrated in the published studyThree taxonomicgroupsmdashpooidspanicoidsand chloridoidsmdashare consistently found asstrong monophyletic groups and no evi-dence has ever contradicted this view (Clarket al 1995 Duvall and Morton 1996Mason-Gamer et al 1998 Gaut et al 1999 Hiluet al 1999 Hsiao et al 1999 Mathews et al2000) To ease the heuristic searches we con-strained our large analyses of the 55 pub-lished phylogenetic trees by forcing eachof these three groups to form three mono-phyletic clades but allowed other taxa to beinserted within them (the ldquobackbonerdquo optionin PAUPcurren408b)
Because bootstrap percentages are miss-ing in most studies before 1993 or becausevalues lt50 are not specied for most ofthe published trees we used a transfor-med function of the bootstrap percentagesto weight the characters into the codedmatrices in both analyses All percent-ages lt50 or nodes with missing val-ues were given the weight of 1 Bootstrap
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 139
percentages cedil50 were weighted by us-ing an exponential transformation (James SFarris pers comm)
e[log(a )pound(b=100)]
where a represents the weight assigned to100 of node support and b representsthe bootstrap percentage This transforma-tion attempts to overcome the conservativebias found in bootstrap percentages IndeedHillis and Bull (1993) using simulationsshowed that bootstrap values gt70 in theirstudy usually indicated with gt95 proba-bility that the corresponding clade was realCorrections to bootstrap estimates have beenproposed (Rodrigo 1993 Zharkikh and Li1995 Effron et al 1996) but these correc-tions cannot compensate for lack of infor-mation in large analyses (Sanderson andWojciechowski 2000) As the number of taxaincreases the relative number of charactersavailable declines and phylogenetic accu-racy suffers Our transformation thus atte-mpts to linearize the bootstrap percent-ages allowing for a gradual increase in thecharacter weights in subsequent parsimonyanalyses
For every supertree reconstructionheuris-tic searches under maximum parsimonywere performed with BaumRagan andPurvis coding schemes alone (hereafterBR-alone and PU-alone respectively) Wealso used bootstrap percentages weight-ing schemes as described above on bothcoding schemes (hereafter BR C bootstrapand PU C bootstrap respectively) and theBaumRagan coding scheme weighted bythe inverse of the number of nodes presentin each source tree (hereafter BR C nodes)We performed 1000 replicates of random ad-dition sequence using the nearest-neighborinterchange swapping algorithm and keep-ing only 20 trees at each replicate Thestored trees were then swapped with thetree bisectionndashreconnection swapping algo-rithm and a maximum of 1000 trees werekept (the ldquomaxtreesrdquo option in PAUPcurren408b)Two different parsimony analyses were per-formed in each of the cases describedabove one considering characters as un-ordered (Fitch parsimony Fitch 1971) andone considering the same characters as ir-reversible (CaminndashSokal parsimony Caminand Sokal 1965) the result was a total of
ten GPWG supertrees and ten large su-pertrees Equally most-parsimonious solu-tions were summarized by using semistrictconsensus
Topological ComparisonsTwo incongruence indices expressed as
distances were used to compare the differ-ent topologies obtained The partition metric(PM symmetric difference in PAUPcurren408bRobinson and Foulds 1981) and the agree-ment subtree metric (D1 Gordon 1980)werecalculated by using PAUPcurren408b (Swofford1998) For the comparisons between thecombined analysis and the GPWG supertreereconstructions we performed KishinondashHasegawa(KH) test (Kishino and Hasegawa1989) and the ShimodairandashHasegawa (SH)test (Shimodaira and Hasegawa 1999) TheGPWG data set consisted of DNA sequencesrestriction site data and morphological char-acters (Table 1) and both tests were per-formed under the maximum parsimony cri-terion Two sets of P-values were thuscalculatedmdashone according to the default op-tions present in PAUPcurren408b (for the KH-test Swofford 1998) the other as describedin Shimodaira and Hasegawa (1999) to en-sure the validity of the test with topolo-gies specied a posteriori The latter testinvolved the creation of 500 bootstrappedreplicates of the GPWG matrix followedby the optimization of 1000 random treesas well as the trees under consideration onthese bootstrapped matrices (for details seeShimodaira and Hasegawa 1999 Goldmanet al 2000)
RESULTS
GPWG Combined TreeThe GPWG combined tree (Fig 1) is
one of the most comprehensive phyloge-netic hypotheses concerning the grass fam-ily Several features can be emphasizedand will serve as a reference for the su-pertree comparisons First and as proposedby GPWG (2000) a large clade (PACCADclade Fig 1) composed of six subfamiliesmdashpanicoids arundinoids chloridoids cen-tothecoids aristidoids and danthonioidsmdashforms a highly supported monophyleticgroup (100 bootstrap Fig 1) No boot-strap values gt52 support any group withinthe PACCAD clade Outside the PACCAD
140 SYSTEMATIC BIOLOGY VOL 51
BEPclade
PACCADclade
early-diverginglineages
outgroup
100
100100
100
100
100
100
100100
100
100
100
100
100
100
100
100
100
100
96
98
76
7787
63
60
86
5268
67
65
9887
98
87
81
91
9292
7890
pooids
bambusoids
ehrhartoids
chloridoids
panicoids +centothecoids
arundinoids
aristidoids +danthonioids
BromusTriticumAvenaBrachypodiumDiarrhenaMelicaGlyceriaNassellaStipaPiptatherumAmpelodesmosPhaenospermaAnisopogonLygeumNardusBrachyelytrumLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaOryzaLeersiaEhrhartaStreptogynaSpartinaSporobolusZoysiaDistichlisEragrostisUniolaPappophorumEriachneMicrairaAmphipogonArundoMoliniaPhragmitesAristidaStipagrostisDanthoniaPennisetumPanicumMiscanthusZeaDanthoniopsis
ChasmanthiumGynerium
ThysanolaenaZeugites
GuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
FIGURE 1 The ldquobigrdquo GPWG tree based on the analysis of eight different data sets (both molecular and morpho-logical) numbers above branches represent bootstrap percentages
clade the bambusoids ehrhartoids andthe pooids form another group called theBEP clade supported by a lower bootstrapvalue (87 bootstrap Fig 1) Finally three
cladesmdashthe anomochloids (Anomochloa Stre-ptochaeta) pharoids (Pharus) and puelioids(Guaduella Puelia)mdashform the basal taxaof thePoaceae (early diverging lineages Fig 1)
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 141
TABLE 2 Partition metrics for the ve MRP modications expressed as distances A) Comparisons withinmodications with unordered characters B) within modications with irreversible characters and C) betweenmodications with unordered characters and modications with irreversible characters
Unordered characters
A) BR-alone BR C bootstrap BR C nodes PU-alone PU C bootstrap
Unordered BR-alone ndash characters BR C bootstrap 0457 ndash
BR C nodes 0379 0388 ndash PU-alone 0440 0362 0422 ndash PU C bootstrap 0474 0379 0457 0362 ndash
B) Irreversible characters
Irreversible BR-alone ndash characters BR C bootstrap 0431 ndash
BR C nodes 0216 0491 ndash PU-alone 0293 0431 0216 ndash PU C bootstrap 0414 0379 0440 0379 ndash
C) Irreversible characters
Unordered BR-alone 0000 0431 0216 0293 0414characters BR C bootstrap 0457 0112 0552 0491 0457
BR C nodes 0379 0483 0319 0328 0483PU-alone 0440 0457 0500 0422 0474PU C bootstrap 0474 0422 0517 0457 0181
GPWG Supertrees
The proportion of nonidentical nodes be-tween the ten MRP reconstructions takenby pairs was calculated by using the PM(Table 2) Analyses using unordered char-acters resulted in topologies with pairwisedistances of between 0362 and 0474 (meandistance 0412 Table 2) Pairwise com-parisons for each MRP modication us-ing irreversible characters gave distances ofbetween 0216 and 0491 (mean distance0369 Table 2) Values for each unorderedBaumRagan modication are closer totheir irreversible counterparts than to anyother methods (mean distance unorderedBRirreversible BR 0328mean distance un-ordered BRirreversible PU 0411 Table 2)The same pattern is found for the un-ordered Purvis modications (mean dis-tance unordered PUirreversible PU 0386mean unordered PUirreversible BR 0468Table 2) Three modications gave very sim-ilar topologies with both types of charac-ters BR-alone gave the exact same topologywhereas the two BR C bootstrap and thetwo PU C bootstrap values were very closeto each other (0112 and 0181 respectivelyTable 2)
For comparisons between the GPWG com-bined tree and the GPWG supertrees D1 dis-tances ranged from 032 for BR C bootstrapwith irreversible characters to 059 for PU-
alone with unordered characters the othereight modications had values from 042 to052 (Fig 2) When considering distancesobtained with PM BR C bootstrap withirreversible characters again gave the topol-ogy the most similar to the GPWG combinedtree yielding a distance of 025 The worstmodication was the BR C nodes with ir-reversible characters for which the distancewas 053 whereas those in the eight othermodications ranged from 041 to 050
We further examined the placement ofthe major subclades dened in the GPWGcombined tree (Fig 3) Two modicationsBR C bootstrap and PU C bootstrap bothwith irreversible characters are the onlymodications to yield the same basalbranching pattern as the combined GPWGtree (Fig 3A) In all the other modicationsinvolving irreversible characters two basalgrasses Guaduella and Puelia are insertedinside the BEP clade BR-alone placed thepooids as sister group to the PACCAD cladewhile BR C nodes and PU-alone placed aclade of the pooids and the ehrhartoids assister group of the PACCAD clade (Fig 3A)With unordered characters and exceptfor the BR-alone modication which wasidentical to the BR-alone with irreversiblecharacters modication all modicationshad an odd basal branching pattern theoutgroup being inserted between two cladesof the early diverging lineages (Fig 3B)
142 SYSTEMATIC BIOLOGY VOL 51
FIGURE 2 D1 greatest subtree agreementmetrics (black) and PM partition metrics (white) expressed asdistancesto the GPWG tree for each of the GPWG supertrees build from separate phylogenies based on the eight characterpartitions of the GPWG combined data set as source trees with characters considered as irreversible (left) andunordered (right)
The two different implementations of theKH-test were used to estimate the validity ofthe ten GPWG supertrees as possible alter-native hypothesis to the GPWG combinedtree (Table 3) Based on the characters ofthe GPWG combined data set the classicalKH-test rejected all the GPWG supertrees(P lt 0001) as suitable alternative phyloge-netic trees However the modied KH-testfound only the PU-alone with unorderedcharacters signicantly different (P lt 005)from the GPWG combined tree (Table 3)None of the other GPWG supertrees couldbe rejected BR C bootstrap with both char-acters types being once again the modica-tion giving the topology closest to that ofthe GPWG combined topology (P D 0588and P D 0624 for unordered and irreversiblecharacters respectively)
Finally Figure 4 shows the supertree ob-tained with BR C bootstrap and irreversiblecharacters The discrepancies between thissupertree and the GPWG combined tree weresmall and concerned mainly single taxa thatwere always positioned close to the cladeto which they belonged in the GPWG tree(Fig 4) The main differences concerned theplacement of the ehrhartoids and bambu-soids which exchanged their position as sis-ter group of the pooids in the supertree incomparison with the GPWG tree and theplacement of the three subfamilies arundi-noids aristidoids and danthonioids
Large SupertreeWe analyzed the 55 publications the same
way as the eight character partitions fromthe GPWG combined data set For clar-ity we do not show the results of BR Cnodes a MRP modication that resultedin topologies incompatible with the place-ment of subfamilies suggested in the GPWGcombined tree When irreversible characterswere used (Fig 5A) BR-alone BR C boot-strap and PU C bootstrap obtained the samebasal branching pattern but the placementof subfamilies inside the PACCAD cladewas slightly different in each case (data notshown) PU-alone was the only modica-tion to place the three subfamilies of theBEP clade together as a sister group to thePACCAD clade agreeing with the GPWG su-pertrees and the GPWG combined tree Withunordered characters (Fig 5B) BR-alone BRC bootstrapand PU-alone obtained the samebasal branching pattern with the BEP cladebeing paraphyletic PU C bootstrap gave anodd combination in which the early diverg-ing lineages were embedded inside the bam-busoids at the base of the tree (Fig 5B)
DISCUSSION
The information given by the two incon-gruence indices helped pinpoint the differ-ences present in the topologies obtained fromthe ten GPWG supertrees built using the
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 143
FIGURE 3 Summary of the subfamilial relationships in the GPWG supertrees obtained with the ve MRP mod-ications with column A) characters considered as irreversible and column B) characters considered as unordered
eight character partitions from the combineddata set Analyses using unordered charac-ters produced supertrees that were less sim-ilar as measured by D1 to the combinedtree than those obtained in analyses usingirreversible characters The only exceptionis for BR-alone which obtained the sametopology with both character types How-ever unordered charactersproduced slightlybetter topologies than irreversible charactersaccording to the PM incongruence index D1is dened as the number of taxa needed tobe removed from both trees to obtain an
identical subtree whereas PM is the numberof taxa bipartitions found between the twotrees being compared (Johnson and Soltis1998) Having higher D1 values than PMvalues suggests that the topological differ-ences do not involve single taxa (in whichcase D1 would be smaller than PM) butrather a large set of taxa placed differentlyin the supertree and the GPWG combinedtree Similar values of D1 and PM wouldsuggest that a few discrepant taxa wereinvolved none being far from their properlocations The D1 distances were much
144 SYSTEMATIC BIOLOGY VOL 51
TABLE 3 Kishino-Hasegawa test based on the difference in length between the GPWG combined tree and the tenGPWG supertrees obtained after applying the different MRP modications The p-values were calculated using thedefault parameter of PAUPcurren408b (Swofford 2000 classic Kishino-Hasegawa [KH] test) and Goldman et al (2000)modication to ensure the validity of the test with a posteriori specied topologies (modied KH test) Asterisk (curren)indicates a signicant p-value (p lt 005)
p-value
Number of steps Classic KH test Modied KH test
Combined GPWG 9054 ndash ndash
Unordered characters BR-alone 9291 lt00001curren 0172BR C bootstrap 9116 lt00001curren 0588BR C nodes 9203 lt00001curren 0396PU-alone 9411 lt00001curren 0042curren
PU C bootstrap 9211 lt00001curren 0374Irreversible characters BR-alone 9291 lt00001curren 0172
BR C bootstrap 9105 00003curren 0624BR C nodes 9281 lt00001curren 0198PU-alone 9311 lt00001curren 0144PU C bootstrap 9191 lt00001curren 0416
greater with unordered than irreversiblecharacters and were much closer to thePM ones obtained with irreversible char-acters Following this logic we can em-phasize that using unordered characters re-sulted in topologies with more differencesfrom the GPWG combined tree as to theplacement of larger subclades than did theuse of irreversible characters The type ofcharacters (ie unordered or irreversible)used to reconstruct the supertrees did notgreatly inuence the placement of the ter-minal taxa in the GPWG supertrees There-fore topologies obtained by consideringmatrix elements as irreversible characterswere closer to the combined phylogenetictree represented by the GPWG combinedanalysis This effect is clearly visible in theplacement of the major subclades dened inthe GPWG combined tree (Fig 4) Unorderedcharacters produced topologies in which theearly diverging lineages were split into twogroupsmdashone including Puelia and Guadu-ella and one including Anomochloa Strep-tochaeta and Pharus This extremely odd andunrealistic pattern does not correspond toany published phylogenetic trees concerningthe grass phylogeny Bininda-Emonds andBryant (1998) found only a minor impactof irreversible characters compared to un-ordered characters in their analysis but wefound that topologies created by using irre-versible characters are closer to an approachusing combined data than are the topolo-gies created by using unordered charactersThe resolution in the large supertrees was ex-tremely dependent on the type of characters
used to perform the analysis such was notthe case with the GPWG supertrees whereboth types of characters gave similar reso-lution (data not shown) Using irreversiblecharacters produced only one or two largesupertrees depending on the modicationused in contrast the ldquomaxtreesrdquo option inPAUPcurren408b was always reached when un-ordered characters were used Of course us-ing irreversible characters should put moreconstraintson the parsimonyanalysis by pre-venting the reversion from state 1 to 0 (thusreducing the number of most-parsimonioustrees) but it is not clear why such a drasticdifference appeared only between the largesupertrees
The Purvis coding scheme has been pro-posed as an improvement to the BaumRagan procedure as an attempt to reducethe dependency and redundancy betweenelements coming from the same topology(Purvis 1995a Ronquist 1996 Bininda-Emonds and Bryant 1998) However our re-sults suggest that the Purvis coding schemedoes not have a large impact on the MRPreconstructions and that the BaumRaganmethod works just as well or even betterComparisons between the approach usingcombined data and BR-alone or combineddata and PU-alone gave similar values forboth PM and D1 (Fig 2) Comparisons be-tween MRP modications also indicated aclose relationship between the two modi-cations especially with irreversible charac-ters (Table 2) This conclusion is less wellsupported with unordered characters be-cause BR-alone is the only modication to
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 145
PACCADclade
BEPclade
early-diverginglineages
outgroup
NassellaStipaPiptatherumAmpelodesmosMelicaGlyceriaBromusTriticumAvenaBrachypodiumDiarrhenaAnisopogonLygeumNardusBrachyelytrumPhaenospermaOryzaLeersiaEhrhartaStreptogynaLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaPennisetumPanicumMiscanthusZeaGyneriumDanthoniopsisZeugitesChasmanthiumThysanolaenaAmphipogonArundoMoliniaPhragmitesSpartinaSporobolusZoysiaDistichlisEragrostisPappophorumUniolaDanthoniaMicrairaAristidaStipagrostisEriachneGuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
pooids
bambusoids
ehrhartoids
chloridoids +danthonioids
panicoids
arundinoids
aristidoids
centothecoids
FIGURE 4 Grass supertree obtained with Baum and Ragan coding scheme with bootstrap support weightingusing separate phylogenies built from the eight character partitions of the GPWG combined data set as source treesDashed lines indicate incongruent branches between the supertree and the GPWG combined tree
146 SYSTEMATIC BIOLOGY VOL 51
FIGURE 5 Summary of the subfamilial relationships in the large supertrees based on 55 published source trees(including 403 genera) obtained using the ve MRP modications with column A) characters considered as irre-versible and column B) characters considered as unordered
give a basal topology very different fromthat obtained with any other modication(Table 2 and Fig 2) Ronquistrsquos (1996) propo-sition to weight each character in the bi-nary matrix by the inverse of the number ofnodes in the corresponding source tree doesnot perform as well as the other modica-tions It yields the greatest partition metricdistance and the second largest agreementsubtree distance (Fig 2) Although the twoalternatives to the BaumRagan method arebased on logical and plain arguments (ienonindependence and redundancy of matrixelements and impact of larger trees) theireffects on MRP reconstruction are not obvi-ous and do not result in topologies closerto our combined reference Indeed PU-alonewith unordered characters was rejected as asuitable alternative hypothesis to the GPWGcombined tree with the modied KH-testand gave the second smallest P-value when
used with irreversible characters (Table 2)However conclusions from the large su-pertree analysis differed because PU-alonewas the only modication to produce thesame basal branching pattern as the GPWGcombined tree (Fig 5B) whereas BR-alonewith the pooids as sister group to the PAC-CAD clade corresponded to an alternativehypothesis for the grass family that is sup-ported by some data from nuclear (Adh Gautet al 1999) and plastid (rbcL Duvall andMorton 1996) genomes Because no refer-ence phylogenetic trees are available thatcontain a number of taxa similar to those ofour large supertrees it is difcult to assesstheir topologies and to determine whetherPU-alone with irreversible characters reallygave a more accurate phylogeny of largegrasses than did the other methods Theplacement of the pooids is possibly not thebest criterion by which to judge the methods
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 147
the evolutionary position of this subfamilywithin the grass family differs as inferredwith different DNA sequence data sets
Weighting by node support as suggestedby Ronquist (1996) improved the t betweenthe additive binary matrices and the GPWGcombined tree The benecial impact of thebootstrap weighting scheme is evident fromthe increased P-values obtained with themodied KH-test (Table 2) This trend is vis-ible for the BaumRagan coding schemebut is less obvious with the Purvis codingscheme (Fig 2) Bootstrap weighting wasproposed (Ronquist 1996) as an alternativeto the Purvis coding scheme both to reducethe greater impact of large source trees oversmalleronesand to improve the effect of well-supported nodes in the MRP analysis It isnot clear how bootstrap weighting could re-duce the impact of larger trees if the smallertrees do not have much greater support val-ues than those of the larger trees Howeverweighting of charactersby bootstrapsupportwithin the Purvis coding scheme appears tobe a redundant procedure unable to greatlyimprove the MRP reconstruction from theimportant information present in bootstrapsupport values (Fig 2) In the large supertreeanalysis PU C bootstrap with unorderedcharacters produced unrealistic topologieswith the early diverging lineages embed-ded in the bambusoids (Fig 5B) The Purviscoding schemes removes important restric-tive information from the matrix (Bininda-Emonds and Bryant 1998) possibly there-fore weighting by bootstrap support wouldrandomly assign high values to characters inthe matrix where this restrictive informationhas been removed and would prevent theweighting schemes from being as effectiveas with the BaumRagan coding schemesWeighting the matrix elements by node sup-port also poses problems Not all bootstrapanalyses can be considered identical and thenumber of replicates the type of searchesdone or both will inuence the supportfound Moreover when node support is notprovided for a source tree weights cannot beassigned and information from this tree willbe down-weighted in the MRP analysis Theeffect of this is great in supertree reconstruc-tions done with older publications which ingeneral do not report node support and arehence down-weighted in the MRP analyses
An important aspect affecting the large su-pertrees that is not resolved in our compar-
isons is the placement of some rarely sam-pled taxa Taxa that are present in only a fewpublications have been allocated a high pro-portion of missing character values whichmakes parsimony analyses much more dif-cult How many of these taxa are mis-placed is difcult to determine because noreference phylogeny for the 403 genera isavailable MRP reconstruction done withoutconstraining three clades (panicoids pooidsand chloridoids) to be monophyletic endedwith rarely sampled taxa being scattered allaround the supertrees (data not shown) Thatcan be an important problem when MRPanalyses are performed on a wide taxonomicgroup comprising large numbers of rarelysampled taxa
MRP coding especially when weighted bynode support can be considered an indica-tion of the signal in the primary data witheach node represented by one (weighted)synapomorphy (see Bininda-Emonds et al1999) However we stress that supertreescannot be viewed and interpreted in ex-actly the same way as phylogenetic analy-ses based on biological characters One ma-jor problem is the difculty of assigningnode support for supertrees Bootstrappingand other resampling procedures cannot beapplied given the clear nonindependenceof the characters present in the binary ma-trix and Bremer support (Bremer 1988) iscertainly equally affected by such noninde-pendence (as is the parsimony itself hencethe different MRP modications proposed)Moreoverbranch lengths associatedwith su-pertrees are treated in the same way as mor-phological branch lengths and unlike molec-ular branch length they cannot be used toestablish divergence times This weakness iseven more signicant in taxonomic groupssuch as the grasses for which dates are dif-cult to gather from other sources (eg fossilrecords)
CONCLUSIONS
We have demonstrated that supertreesoffer an easy way of producing phyloge-nies with a large number of taxa and thatthey can give good estimates of relation-ships within these groups (in this case thegrass family) Supertrees based on the eightcharacter partitions of the GPWG data setswere found to roughly match the combinedanalysis the discrepancies of the best MRP
148 SYSTEMATIC BIOLOGY VOL 51
modications being found mainly in weaklysupported branches of the combined treeThe BaumRagan and the Purvis modica-tions gave similar results but incorporatingbootstrap support associated with preexist-ing topologies improved the BaumRaganmodication Moreover supertrees can beuseful for comparative studies (Purvis1995b Bininda-Emonds et al 1999) whetherof adaptation coevolution rates of evo-lution cospeciation or rates of effectivecladogenesis where accuracy in the branchlength of phylogenetic reconstructions is notthe primary problem (Purvis et al 1994)Supertree reconstructions are also a usefulway to help highlight poor taxonomic sam-pling and identify where previous studiesareinconsistentThey can therefore be used as anexploratory tool capable of developing newhypotheses and indicating where future re-search should be focused
ACKNOWLEDGMENTS
We are grateful to Elizabeth Kellogg and LynnClark for providing the GPWG matrix and for their com-ments on the manuscript James Farris for helpful dis-cussion regarding bootstrap weighting schemes duringthe XIX Hennig Society meeting Stephen Renvoize andMark Chase for helpful discussion on the project OlafBininda-Emonds for his comments on the manuscriptand JonathanDavis for the testing of the SuperTree 085bprogram This study was supported by the Irish HigherEducation Authority the Roche Research Foundation(99-186) Enterprise Iceland (IC200129) the AgassizFoundation and the Swiss NSF (31-52378)
REFERENCES
(References marked curren were used as sources of phyloge-nies for the supertree reconstructions)
ARNQVIST G AND D WOOSTER 1995 Meta-analysisSynthesizing research ndings in ecology and evolu-tion Trends Ecol Evol 10236ndash240
currenBARKER N P 1997 The relationships of AmphipogonElytrophorus and Cyperochloa (Poaceae) as suggestedby rbcL sequence data Telopea 7205ndash213
currenBARKER N P H P LINDER AND E H HARLEY 1995Polyphyly of Arundinoids (Poaceae) Evidence fromrbcL sequence data Syst Bot 20423ndash435
currenBARKER N P H P LINDER and E H HARLEY 1999Sequences of the grass-specic insert in the chloro-plast rpoC2 gene elucidate generic relationships of theArundinoids (Poaceae) Syst Bot 23327ndash350
currenBAUM B R 1982 A phylogenetic analysis of the tribeTriticeae (Poaceae) based on morphological charactersof the genera Can J Bot 61518ndash535
BAUM B R 1992 Combining trees as a way of com-bining data sets for phylogenetic inference and thedesirability of combining gene trees Taxon 413ndash10
currenBAUM B R J R ESTES AND P K GUPTA 1986 Assess-ment of the genomic systematic classication in theTriticeae Am J Bot 741388ndash1395
BAUM B R AND M A RAGAN 1993 Reply to A GRodrigorsquos ldquoA comment on Baumrsquos method for com-bining phylogenetic treesrdquo Taxon 42637ndash640
BININD A-EMONDS O R P AND H N BRYANT 1998Properties of matrix representation with parsimonyanalyses Syst Biol 47497ndash508
BININD A-EMONDS O R P J L GITTLEMAN ANDA PURVIS 1999 Building large trees by combiningphylogenetic information A complete phylogenyof the extant Carnivora (Mammalia) Biol Rev 74143ndash175
BREMER K 1988 The limits of amino acid sequencedata in angiosperm phylogenetic reconstruction Evo-lution 42795ndash803
currenBUCKLER E S IV AND T P HOLTSFORD 1996 Zeasystematics Ribosomal ITS evidence Mol Biol Evol13612ndash622
CAMIN J H AND R R SOKAL 1965 A method for de-ducing branching sequences in phylogeny Evolution19311ndash326
currenCATALAN P E A KELLOGG AND R G OLMS TEAD1997 Phylogeny of Poaceae subfamily Pooideaebased on chloroplast ndhFgene sequences Mol Phy-logenet Evol 8150ndash166
currenCLARK L G W ZHANG AND J F WENDEL 1995 Aphylogenyof the grass family (Poaceae) basedon ndhFsequence data Syst Bot 20436ndash460
CONSTANTINES CU M AND D SANKOFF 1995 An ef-cient algorithm for supertrees J Classif 12101ndash112
currenCUMMINGS M P L M KING AND E A KELLOGG1994 Slipped-strand mispairing in a plastid generpoC2 in grasses (Poaceae) Mol Biol Evol 111ndash8
currenDARBYSHIRE S J AND S I WARWICK 1992 Phylogenyof North American Festuca (Poaceae) and related gen-era using chloroplast DNA restriction site variationCan J Bot 702415ndash2419
currenDAVIS J I AND R J SORENG 1993Phylogenetic struc-ture in the grass family (Poaceae) as inferred fromchloroplast DNA restriction site variation Am J Bot801444ndash1454
currenDOEBLEY J M DURBIN E M GOLENBERG M TCLEGG AND D P MA 1990 Evolutionary analysisof the large subunit of carboxylase (rbcL) nucleotidesequence among the grasses (Gramineae) Evolution441097ndash1108
currenDUVALL M R AND B R MORTON 1996 Molecularphylogenetics of Poaceae An expanded analysis ofrbcL sequence data Mol Phylogenet Evol 5352ndash358
currenDUVALL M R P M PETERSON E E TERRELL ANDA H CHRISTENS EN 1993 Phylogeny of NorthAmerican oryzoid grasses as constructed from mapsof plastid DNA restriction sites Am J Bot 8083ndash88
EFFRON B E HALLORAN AND S HOLMES 1996 Boot-strap condence levels for phylogenetic trees ProcNatl Acad Sci USA 937085ndash7090
currenESEN A AND K W HILU 1991 Electrophoretic andimmunological studies of prolamins in the PoaceaeII Phylogenetic afnities of the Atistideae Taxon 405ndash17
currenESEN A AND K W HILU 1993Prolamin and immuno-logical studies in the Poaceae IV SubfamilyPanicoidsCan J Bot 71315ndash322
FARRIS J S A G KLUGE AND M J ECKHARDT 1970 Anumerical approach to phylogenetic systematics SystZool 19172ndash191
FELSENSTEIN J 1985 Condence limits on phylogeniesAn approach using the bootstrap Evolution 39783ndash791
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 149
FITCH W M 1971 Toward dening the course of evolu-tion Minimal change for a specic tree topology SystZool 20406ndash416
currenFREDERIKSEN S AND O SEBERG 1991 Phylogeneticanalysis of the Triticeae (Poaceae) Hereditas 11515ndash19
currenGAUT B S A SPEEK BR MORTON AND M T CLEGG 1999Patterns of genetic diversication within the Adhgene family in the grasses (Poaceae) Mol Biol Evol161086ndash1097
GOLDMANN N J P ANDERSON AND A G RODRIGO2000 Likelihood-based tests of topologies in phyloge-netics Syst Biol 49652ndash670
GORDON A D 1980 On the assessment and compar-ison of classications Pages 149ndash160 in Analyse dedonnees et informatique (R Tomassone ed) INRALe Chesnay France
GPWG2000A phylogeny of the grass family (Poaceae)as inferred from eight character sets Pages 3ndash7 inGrasses Systematics and evolution (S W L Jacobsand J Everett eds) CSIRO Melbourne Australia
currenGREBENS TEIN B O GREBENS TEIN W SAUER ANDV HEMLEBEN 1996 Distribution and complex organi-zation of satellite DNA sequences in Aveneae speciesGenome 391045ndash1050
currenGREBENS TEIN B M ROSER W SAUER ANDV HEMLEBEN 1998 Molecular phylogenetic re-lationships in Aveneae (Poaceae) species and othergrasses as inferred from ITS1 and ITS2 rDNAsequences Plant Syst Evol 213233ndash250
currenHAMBY R K AND EA ZIMMER 1987Ribosomal RNAsequences for inferring phylogeny within the grassfamily (Poaceae) Plant Syst Evol 16029ndash37
HARVEY P H AND M D PAGEL 1991The comparativemethod in evolutionary biology Oxford Univ PressLondon
HILLIS D M 1996 Inferring complex phylogenies Na-ture 383130ndash131
HILLIS D M AND J J BULL 1993 An empirical test ofbootstrapping as a method for assessing condencein phylogenetic analysis Syst Biol 42182ndash192
currenHILU K W L A ALICE AND H LIANG 1999 Phy-logeny of Poaceae inferred from matK sequencesAnn Mo Bot Gard 86835ndash851
currenHILU K W AND A ESEN 1992Prolamin andimmuno-logical strudies in the Poaceae III Subfamily Chlori-doids Am J Bot 80104ndash113
currenHILU K W AND H LIANG 1997 The matK gene Se-quence variation and application in plant systematicsAm J Bot 84830ndash839
currenHILU K W AND K WRIGHT 1982 Systematics ofGramineae A cluster analysis study Taxon 319ndash37
currenHSIAO C N J CHATTERTON K H ASAY AND K BJENS EN 1993 Phylogenetic relationships of 10 grassspecies An assessment of phylogenetic utility of theinternal transcribed spacer region in nuclear riboso-mal DNA in monocots Genome 37112ndash120
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1994 Phylogenetic relationships of themonogenomic species of the wheat tribe Triticeae(Poaceae) inferred from nuclear rDNA (internal tran-scribed spacer) sequences Genome 38211ndash223
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1995Molecular phylogeny of the pooids(Poaceae) based on nuclear rDNA (ITS) sequencesTheor Appl Genet 90389ndash398
currenHSIAO C S W JACOBS N P BARKER AND N JCHATTERTON 1998 A molecular phylogeny of the
subfamily arundinoids (Poaceae) based on sequencesof rDNA Aust Syst Bot 1141ndash52
currenHSIAO C S W L JACOBS N J CHATTERTON AND KH ASAY 1999 A molecular phylogeny of the grassfamily (Poaceae) based on the sequences of nuclearribosomal DNA (ITS) Aust Syst Bot 11667ndash688
JOHNS ON L A AND D E SOLTIS 1998 Assessing con-gruence Empirical examples from molecular dataPages 297ndash348 in Molecular systematics of plants IIDNA sequencing (D E Soltis P S Soltis and J JDoyle eds) Kluwer Academic Publishers NorwellMassachusetts
currenKATAYAMA H AND Y OGIHARA 1996 Phylogeneticafnities of the grasses to other monocots as re-vealed by molecular analysis of chloroplast DNACurr Genet 29572ndash581
currenKELCHNER S A AND L G CLARK 1997 Molecu-lar evolution and phylogenetic utility of the chloro-plast rpl16 intron in Chusquea and the bambusoids(Poaceae) Mol Phylogenet Evol 8385ndash397
currenKELLOGG E A 1989 Comments on genomic genera inthe Triticeae (Poaceae) Am J Bot 76796ndash805
currenKELLOGG E A 1992Tools for studying the chloroplastgenome in the Triticeae (Gramineae) An ecoRI mapa diagnostic deletion and support for Bromus as anoutgroup Am J Bot 79186ndash197
currenKELLOGG E A 1998aRelationships of cereal crops andother grassesProc NatlAcad Sci USA952005ndash2010
currenKELLOGG E A 1998bWhorsquos related to whom Recentresults from molecular systematic studies Curr OpinPlant Biol 1149ndash158
currenKELLOGG E A AND R APPELS 1995 Intraspecic andinterspecic variation in 5S RNA genes are decoupledin diploid wheat relatives Genetics 140325ndash343
currenKELLOGG E A AND L WATSON 1993 Phyloge-netic studies of a large data set I bambusoids An-dropogonodae and pooids (Gramineae) Bot Rev59273ndash311
KIM J 1996 General inconsistency conditions for max-imum parsimony Effects of branch lengths and in-creasing numbers of taxa Syst Biol 45363ndash374
KISHINO H AND M HASEGAWA 1989 Evaluation ofthe maximum likelihood estimate of the evolution-ary tree topologies form DNA sequence data and thebranching order in Hominoidea J Mol Evol 29170ndash179
currenLIANG H AND K W HILU 1995 Application of thematK gene sequence to grass systematics Can J Bot74125ndash134
MABBERLEY D J 1993 The plant-book A portable dic-tionary of the vascular plants 2nd edition CambridgeUniv Press Cambridge
currenMACFARLANE T D AND L WATSON 1982The classi-cation of Poaceaesubfamily pooids Taxon31178ndash203
currenMASON-GAMER R J AND EA KELLOGG 1996Testingfor phylogenetic conict among molecular data sets inthe tribe Triticeae (Gramineae) Syst Biol 45524ndash545
currenMASON-GAMER R J C F WEIL AND E A KELLOGG1998Granule-bound starch synthase Structure func-tion andphylogenetic utility Mol Biol Evol 151658ndash1673
currenMATHEWS S AND R A SHARROCK 1996 The phy-tochrome gene family in grasses (Poaceae) A phy-logeny and evidence that grasses have a subset ofthe loci found in dicot angiosperms Mol Biol Evol131141ndash1150
currenMATHEWS S R C TSAL AND E A KELLOGG 2000Phylogenetic structure in the grass family (Poaceae)
150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel

2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 139
percentages cedil50 were weighted by us-ing an exponential transformation (James SFarris pers comm)
e[log(a )pound(b=100)]
where a represents the weight assigned to100 of node support and b representsthe bootstrap percentage This transforma-tion attempts to overcome the conservativebias found in bootstrap percentages IndeedHillis and Bull (1993) using simulationsshowed that bootstrap values gt70 in theirstudy usually indicated with gt95 proba-bility that the corresponding clade was realCorrections to bootstrap estimates have beenproposed (Rodrigo 1993 Zharkikh and Li1995 Effron et al 1996) but these correc-tions cannot compensate for lack of infor-mation in large analyses (Sanderson andWojciechowski 2000) As the number of taxaincreases the relative number of charactersavailable declines and phylogenetic accu-racy suffers Our transformation thus atte-mpts to linearize the bootstrap percent-ages allowing for a gradual increase in thecharacter weights in subsequent parsimonyanalyses
For every supertree reconstructionheuris-tic searches under maximum parsimonywere performed with BaumRagan andPurvis coding schemes alone (hereafterBR-alone and PU-alone respectively) Wealso used bootstrap percentages weight-ing schemes as described above on bothcoding schemes (hereafter BR C bootstrapand PU C bootstrap respectively) and theBaumRagan coding scheme weighted bythe inverse of the number of nodes presentin each source tree (hereafter BR C nodes)We performed 1000 replicates of random ad-dition sequence using the nearest-neighborinterchange swapping algorithm and keep-ing only 20 trees at each replicate Thestored trees were then swapped with thetree bisectionndashreconnection swapping algo-rithm and a maximum of 1000 trees werekept (the ldquomaxtreesrdquo option in PAUPcurren408b)Two different parsimony analyses were per-formed in each of the cases describedabove one considering characters as un-ordered (Fitch parsimony Fitch 1971) andone considering the same characters as ir-reversible (CaminndashSokal parsimony Caminand Sokal 1965) the result was a total of
ten GPWG supertrees and ten large su-pertrees Equally most-parsimonious solu-tions were summarized by using semistrictconsensus
Topological ComparisonsTwo incongruence indices expressed as
distances were used to compare the differ-ent topologies obtained The partition metric(PM symmetric difference in PAUPcurren408bRobinson and Foulds 1981) and the agree-ment subtree metric (D1 Gordon 1980)werecalculated by using PAUPcurren408b (Swofford1998) For the comparisons between thecombined analysis and the GPWG supertreereconstructions we performed KishinondashHasegawa(KH) test (Kishino and Hasegawa1989) and the ShimodairandashHasegawa (SH)test (Shimodaira and Hasegawa 1999) TheGPWG data set consisted of DNA sequencesrestriction site data and morphological char-acters (Table 1) and both tests were per-formed under the maximum parsimony cri-terion Two sets of P-values were thuscalculatedmdashone according to the default op-tions present in PAUPcurren408b (for the KH-test Swofford 1998) the other as describedin Shimodaira and Hasegawa (1999) to en-sure the validity of the test with topolo-gies specied a posteriori The latter testinvolved the creation of 500 bootstrappedreplicates of the GPWG matrix followedby the optimization of 1000 random treesas well as the trees under consideration onthese bootstrapped matrices (for details seeShimodaira and Hasegawa 1999 Goldmanet al 2000)
RESULTS
GPWG Combined TreeThe GPWG combined tree (Fig 1) is
one of the most comprehensive phyloge-netic hypotheses concerning the grass fam-ily Several features can be emphasizedand will serve as a reference for the su-pertree comparisons First and as proposedby GPWG (2000) a large clade (PACCADclade Fig 1) composed of six subfamiliesmdashpanicoids arundinoids chloridoids cen-tothecoids aristidoids and danthonioidsmdashforms a highly supported monophyleticgroup (100 bootstrap Fig 1) No boot-strap values gt52 support any group withinthe PACCAD clade Outside the PACCAD
140 SYSTEMATIC BIOLOGY VOL 51
BEPclade
PACCADclade
early-diverginglineages
outgroup
100
100100
100
100
100
100
100100
100
100
100
100
100
100
100
100
100
100
96
98
76
7787
63
60
86
5268
67
65
9887
98
87
81
91
9292
7890
pooids
bambusoids
ehrhartoids
chloridoids
panicoids +centothecoids
arundinoids
aristidoids +danthonioids
BromusTriticumAvenaBrachypodiumDiarrhenaMelicaGlyceriaNassellaStipaPiptatherumAmpelodesmosPhaenospermaAnisopogonLygeumNardusBrachyelytrumLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaOryzaLeersiaEhrhartaStreptogynaSpartinaSporobolusZoysiaDistichlisEragrostisUniolaPappophorumEriachneMicrairaAmphipogonArundoMoliniaPhragmitesAristidaStipagrostisDanthoniaPennisetumPanicumMiscanthusZeaDanthoniopsis
ChasmanthiumGynerium
ThysanolaenaZeugites
GuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
FIGURE 1 The ldquobigrdquo GPWG tree based on the analysis of eight different data sets (both molecular and morpho-logical) numbers above branches represent bootstrap percentages
clade the bambusoids ehrhartoids andthe pooids form another group called theBEP clade supported by a lower bootstrapvalue (87 bootstrap Fig 1) Finally three
cladesmdashthe anomochloids (Anomochloa Stre-ptochaeta) pharoids (Pharus) and puelioids(Guaduella Puelia)mdashform the basal taxaof thePoaceae (early diverging lineages Fig 1)
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 141
TABLE 2 Partition metrics for the ve MRP modications expressed as distances A) Comparisons withinmodications with unordered characters B) within modications with irreversible characters and C) betweenmodications with unordered characters and modications with irreversible characters
Unordered characters
A) BR-alone BR C bootstrap BR C nodes PU-alone PU C bootstrap
Unordered BR-alone ndash characters BR C bootstrap 0457 ndash
BR C nodes 0379 0388 ndash PU-alone 0440 0362 0422 ndash PU C bootstrap 0474 0379 0457 0362 ndash
B) Irreversible characters
Irreversible BR-alone ndash characters BR C bootstrap 0431 ndash
BR C nodes 0216 0491 ndash PU-alone 0293 0431 0216 ndash PU C bootstrap 0414 0379 0440 0379 ndash
C) Irreversible characters
Unordered BR-alone 0000 0431 0216 0293 0414characters BR C bootstrap 0457 0112 0552 0491 0457
BR C nodes 0379 0483 0319 0328 0483PU-alone 0440 0457 0500 0422 0474PU C bootstrap 0474 0422 0517 0457 0181
GPWG Supertrees
The proportion of nonidentical nodes be-tween the ten MRP reconstructions takenby pairs was calculated by using the PM(Table 2) Analyses using unordered char-acters resulted in topologies with pairwisedistances of between 0362 and 0474 (meandistance 0412 Table 2) Pairwise com-parisons for each MRP modication us-ing irreversible characters gave distances ofbetween 0216 and 0491 (mean distance0369 Table 2) Values for each unorderedBaumRagan modication are closer totheir irreversible counterparts than to anyother methods (mean distance unorderedBRirreversible BR 0328mean distance un-ordered BRirreversible PU 0411 Table 2)The same pattern is found for the un-ordered Purvis modications (mean dis-tance unordered PUirreversible PU 0386mean unordered PUirreversible BR 0468Table 2) Three modications gave very sim-ilar topologies with both types of charac-ters BR-alone gave the exact same topologywhereas the two BR C bootstrap and thetwo PU C bootstrap values were very closeto each other (0112 and 0181 respectivelyTable 2)
For comparisons between the GPWG com-bined tree and the GPWG supertrees D1 dis-tances ranged from 032 for BR C bootstrapwith irreversible characters to 059 for PU-
alone with unordered characters the othereight modications had values from 042 to052 (Fig 2) When considering distancesobtained with PM BR C bootstrap withirreversible characters again gave the topol-ogy the most similar to the GPWG combinedtree yielding a distance of 025 The worstmodication was the BR C nodes with ir-reversible characters for which the distancewas 053 whereas those in the eight othermodications ranged from 041 to 050
We further examined the placement ofthe major subclades dened in the GPWGcombined tree (Fig 3) Two modicationsBR C bootstrap and PU C bootstrap bothwith irreversible characters are the onlymodications to yield the same basalbranching pattern as the combined GPWGtree (Fig 3A) In all the other modicationsinvolving irreversible characters two basalgrasses Guaduella and Puelia are insertedinside the BEP clade BR-alone placed thepooids as sister group to the PACCAD cladewhile BR C nodes and PU-alone placed aclade of the pooids and the ehrhartoids assister group of the PACCAD clade (Fig 3A)With unordered characters and exceptfor the BR-alone modication which wasidentical to the BR-alone with irreversiblecharacters modication all modicationshad an odd basal branching pattern theoutgroup being inserted between two cladesof the early diverging lineages (Fig 3B)
142 SYSTEMATIC BIOLOGY VOL 51
FIGURE 2 D1 greatest subtree agreementmetrics (black) and PM partition metrics (white) expressed asdistancesto the GPWG tree for each of the GPWG supertrees build from separate phylogenies based on the eight characterpartitions of the GPWG combined data set as source trees with characters considered as irreversible (left) andunordered (right)
The two different implementations of theKH-test were used to estimate the validity ofthe ten GPWG supertrees as possible alter-native hypothesis to the GPWG combinedtree (Table 3) Based on the characters ofthe GPWG combined data set the classicalKH-test rejected all the GPWG supertrees(P lt 0001) as suitable alternative phyloge-netic trees However the modied KH-testfound only the PU-alone with unorderedcharacters signicantly different (P lt 005)from the GPWG combined tree (Table 3)None of the other GPWG supertrees couldbe rejected BR C bootstrap with both char-acters types being once again the modica-tion giving the topology closest to that ofthe GPWG combined topology (P D 0588and P D 0624 for unordered and irreversiblecharacters respectively)
Finally Figure 4 shows the supertree ob-tained with BR C bootstrap and irreversiblecharacters The discrepancies between thissupertree and the GPWG combined tree weresmall and concerned mainly single taxa thatwere always positioned close to the cladeto which they belonged in the GPWG tree(Fig 4) The main differences concerned theplacement of the ehrhartoids and bambu-soids which exchanged their position as sis-ter group of the pooids in the supertree incomparison with the GPWG tree and theplacement of the three subfamilies arundi-noids aristidoids and danthonioids
Large SupertreeWe analyzed the 55 publications the same
way as the eight character partitions fromthe GPWG combined data set For clar-ity we do not show the results of BR Cnodes a MRP modication that resultedin topologies incompatible with the place-ment of subfamilies suggested in the GPWGcombined tree When irreversible characterswere used (Fig 5A) BR-alone BR C boot-strap and PU C bootstrap obtained the samebasal branching pattern but the placementof subfamilies inside the PACCAD cladewas slightly different in each case (data notshown) PU-alone was the only modica-tion to place the three subfamilies of theBEP clade together as a sister group to thePACCAD clade agreeing with the GPWG su-pertrees and the GPWG combined tree Withunordered characters (Fig 5B) BR-alone BRC bootstrapand PU-alone obtained the samebasal branching pattern with the BEP cladebeing paraphyletic PU C bootstrap gave anodd combination in which the early diverg-ing lineages were embedded inside the bam-busoids at the base of the tree (Fig 5B)
DISCUSSION
The information given by the two incon-gruence indices helped pinpoint the differ-ences present in the topologies obtained fromthe ten GPWG supertrees built using the
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 143
FIGURE 3 Summary of the subfamilial relationships in the GPWG supertrees obtained with the ve MRP mod-ications with column A) characters considered as irreversible and column B) characters considered as unordered
eight character partitions from the combineddata set Analyses using unordered charac-ters produced supertrees that were less sim-ilar as measured by D1 to the combinedtree than those obtained in analyses usingirreversible characters The only exceptionis for BR-alone which obtained the sametopology with both character types How-ever unordered charactersproduced slightlybetter topologies than irreversible charactersaccording to the PM incongruence index D1is dened as the number of taxa needed tobe removed from both trees to obtain an
identical subtree whereas PM is the numberof taxa bipartitions found between the twotrees being compared (Johnson and Soltis1998) Having higher D1 values than PMvalues suggests that the topological differ-ences do not involve single taxa (in whichcase D1 would be smaller than PM) butrather a large set of taxa placed differentlyin the supertree and the GPWG combinedtree Similar values of D1 and PM wouldsuggest that a few discrepant taxa wereinvolved none being far from their properlocations The D1 distances were much
144 SYSTEMATIC BIOLOGY VOL 51
TABLE 3 Kishino-Hasegawa test based on the difference in length between the GPWG combined tree and the tenGPWG supertrees obtained after applying the different MRP modications The p-values were calculated using thedefault parameter of PAUPcurren408b (Swofford 2000 classic Kishino-Hasegawa [KH] test) and Goldman et al (2000)modication to ensure the validity of the test with a posteriori specied topologies (modied KH test) Asterisk (curren)indicates a signicant p-value (p lt 005)
p-value
Number of steps Classic KH test Modied KH test
Combined GPWG 9054 ndash ndash
Unordered characters BR-alone 9291 lt00001curren 0172BR C bootstrap 9116 lt00001curren 0588BR C nodes 9203 lt00001curren 0396PU-alone 9411 lt00001curren 0042curren
PU C bootstrap 9211 lt00001curren 0374Irreversible characters BR-alone 9291 lt00001curren 0172
BR C bootstrap 9105 00003curren 0624BR C nodes 9281 lt00001curren 0198PU-alone 9311 lt00001curren 0144PU C bootstrap 9191 lt00001curren 0416
greater with unordered than irreversiblecharacters and were much closer to thePM ones obtained with irreversible char-acters Following this logic we can em-phasize that using unordered characters re-sulted in topologies with more differencesfrom the GPWG combined tree as to theplacement of larger subclades than did theuse of irreversible characters The type ofcharacters (ie unordered or irreversible)used to reconstruct the supertrees did notgreatly inuence the placement of the ter-minal taxa in the GPWG supertrees There-fore topologies obtained by consideringmatrix elements as irreversible characterswere closer to the combined phylogenetictree represented by the GPWG combinedanalysis This effect is clearly visible in theplacement of the major subclades dened inthe GPWG combined tree (Fig 4) Unorderedcharacters produced topologies in which theearly diverging lineages were split into twogroupsmdashone including Puelia and Guadu-ella and one including Anomochloa Strep-tochaeta and Pharus This extremely odd andunrealistic pattern does not correspond toany published phylogenetic trees concerningthe grass phylogeny Bininda-Emonds andBryant (1998) found only a minor impactof irreversible characters compared to un-ordered characters in their analysis but wefound that topologies created by using irre-versible characters are closer to an approachusing combined data than are the topolo-gies created by using unordered charactersThe resolution in the large supertrees was ex-tremely dependent on the type of characters
used to perform the analysis such was notthe case with the GPWG supertrees whereboth types of characters gave similar reso-lution (data not shown) Using irreversiblecharacters produced only one or two largesupertrees depending on the modicationused in contrast the ldquomaxtreesrdquo option inPAUPcurren408b was always reached when un-ordered characters were used Of course us-ing irreversible characters should put moreconstraintson the parsimonyanalysis by pre-venting the reversion from state 1 to 0 (thusreducing the number of most-parsimonioustrees) but it is not clear why such a drasticdifference appeared only between the largesupertrees
The Purvis coding scheme has been pro-posed as an improvement to the BaumRagan procedure as an attempt to reducethe dependency and redundancy betweenelements coming from the same topology(Purvis 1995a Ronquist 1996 Bininda-Emonds and Bryant 1998) However our re-sults suggest that the Purvis coding schemedoes not have a large impact on the MRPreconstructions and that the BaumRaganmethod works just as well or even betterComparisons between the approach usingcombined data and BR-alone or combineddata and PU-alone gave similar values forboth PM and D1 (Fig 2) Comparisons be-tween MRP modications also indicated aclose relationship between the two modi-cations especially with irreversible charac-ters (Table 2) This conclusion is less wellsupported with unordered characters be-cause BR-alone is the only modication to
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 145
PACCADclade
BEPclade
early-diverginglineages
outgroup
NassellaStipaPiptatherumAmpelodesmosMelicaGlyceriaBromusTriticumAvenaBrachypodiumDiarrhenaAnisopogonLygeumNardusBrachyelytrumPhaenospermaOryzaLeersiaEhrhartaStreptogynaLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaPennisetumPanicumMiscanthusZeaGyneriumDanthoniopsisZeugitesChasmanthiumThysanolaenaAmphipogonArundoMoliniaPhragmitesSpartinaSporobolusZoysiaDistichlisEragrostisPappophorumUniolaDanthoniaMicrairaAristidaStipagrostisEriachneGuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
pooids
bambusoids
ehrhartoids
chloridoids +danthonioids
panicoids
arundinoids
aristidoids
centothecoids
FIGURE 4 Grass supertree obtained with Baum and Ragan coding scheme with bootstrap support weightingusing separate phylogenies built from the eight character partitions of the GPWG combined data set as source treesDashed lines indicate incongruent branches between the supertree and the GPWG combined tree
146 SYSTEMATIC BIOLOGY VOL 51
FIGURE 5 Summary of the subfamilial relationships in the large supertrees based on 55 published source trees(including 403 genera) obtained using the ve MRP modications with column A) characters considered as irre-versible and column B) characters considered as unordered
give a basal topology very different fromthat obtained with any other modication(Table 2 and Fig 2) Ronquistrsquos (1996) propo-sition to weight each character in the bi-nary matrix by the inverse of the number ofnodes in the corresponding source tree doesnot perform as well as the other modica-tions It yields the greatest partition metricdistance and the second largest agreementsubtree distance (Fig 2) Although the twoalternatives to the BaumRagan method arebased on logical and plain arguments (ienonindependence and redundancy of matrixelements and impact of larger trees) theireffects on MRP reconstruction are not obvi-ous and do not result in topologies closerto our combined reference Indeed PU-alonewith unordered characters was rejected as asuitable alternative hypothesis to the GPWGcombined tree with the modied KH-testand gave the second smallest P-value when
used with irreversible characters (Table 2)However conclusions from the large su-pertree analysis differed because PU-alonewas the only modication to produce thesame basal branching pattern as the GPWGcombined tree (Fig 5B) whereas BR-alonewith the pooids as sister group to the PAC-CAD clade corresponded to an alternativehypothesis for the grass family that is sup-ported by some data from nuclear (Adh Gautet al 1999) and plastid (rbcL Duvall andMorton 1996) genomes Because no refer-ence phylogenetic trees are available thatcontain a number of taxa similar to those ofour large supertrees it is difcult to assesstheir topologies and to determine whetherPU-alone with irreversible characters reallygave a more accurate phylogeny of largegrasses than did the other methods Theplacement of the pooids is possibly not thebest criterion by which to judge the methods
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 147
the evolutionary position of this subfamilywithin the grass family differs as inferredwith different DNA sequence data sets
Weighting by node support as suggestedby Ronquist (1996) improved the t betweenthe additive binary matrices and the GPWGcombined tree The benecial impact of thebootstrap weighting scheme is evident fromthe increased P-values obtained with themodied KH-test (Table 2) This trend is vis-ible for the BaumRagan coding schemebut is less obvious with the Purvis codingscheme (Fig 2) Bootstrap weighting wasproposed (Ronquist 1996) as an alternativeto the Purvis coding scheme both to reducethe greater impact of large source trees oversmalleronesand to improve the effect of well-supported nodes in the MRP analysis It isnot clear how bootstrap weighting could re-duce the impact of larger trees if the smallertrees do not have much greater support val-ues than those of the larger trees Howeverweighting of charactersby bootstrapsupportwithin the Purvis coding scheme appears tobe a redundant procedure unable to greatlyimprove the MRP reconstruction from theimportant information present in bootstrapsupport values (Fig 2) In the large supertreeanalysis PU C bootstrap with unorderedcharacters produced unrealistic topologieswith the early diverging lineages embed-ded in the bambusoids (Fig 5B) The Purviscoding schemes removes important restric-tive information from the matrix (Bininda-Emonds and Bryant 1998) possibly there-fore weighting by bootstrap support wouldrandomly assign high values to characters inthe matrix where this restrictive informationhas been removed and would prevent theweighting schemes from being as effectiveas with the BaumRagan coding schemesWeighting the matrix elements by node sup-port also poses problems Not all bootstrapanalyses can be considered identical and thenumber of replicates the type of searchesdone or both will inuence the supportfound Moreover when node support is notprovided for a source tree weights cannot beassigned and information from this tree willbe down-weighted in the MRP analysis Theeffect of this is great in supertree reconstruc-tions done with older publications which ingeneral do not report node support and arehence down-weighted in the MRP analyses
An important aspect affecting the large su-pertrees that is not resolved in our compar-
isons is the placement of some rarely sam-pled taxa Taxa that are present in only a fewpublications have been allocated a high pro-portion of missing character values whichmakes parsimony analyses much more dif-cult How many of these taxa are mis-placed is difcult to determine because noreference phylogeny for the 403 genera isavailable MRP reconstruction done withoutconstraining three clades (panicoids pooidsand chloridoids) to be monophyletic endedwith rarely sampled taxa being scattered allaround the supertrees (data not shown) Thatcan be an important problem when MRPanalyses are performed on a wide taxonomicgroup comprising large numbers of rarelysampled taxa
MRP coding especially when weighted bynode support can be considered an indica-tion of the signal in the primary data witheach node represented by one (weighted)synapomorphy (see Bininda-Emonds et al1999) However we stress that supertreescannot be viewed and interpreted in ex-actly the same way as phylogenetic analy-ses based on biological characters One ma-jor problem is the difculty of assigningnode support for supertrees Bootstrappingand other resampling procedures cannot beapplied given the clear nonindependenceof the characters present in the binary ma-trix and Bremer support (Bremer 1988) iscertainly equally affected by such noninde-pendence (as is the parsimony itself hencethe different MRP modications proposed)Moreoverbranch lengths associatedwith su-pertrees are treated in the same way as mor-phological branch lengths and unlike molec-ular branch length they cannot be used toestablish divergence times This weakness iseven more signicant in taxonomic groupssuch as the grasses for which dates are dif-cult to gather from other sources (eg fossilrecords)
CONCLUSIONS
We have demonstrated that supertreesoffer an easy way of producing phyloge-nies with a large number of taxa and thatthey can give good estimates of relation-ships within these groups (in this case thegrass family) Supertrees based on the eightcharacter partitions of the GPWG data setswere found to roughly match the combinedanalysis the discrepancies of the best MRP
148 SYSTEMATIC BIOLOGY VOL 51
modications being found mainly in weaklysupported branches of the combined treeThe BaumRagan and the Purvis modica-tions gave similar results but incorporatingbootstrap support associated with preexist-ing topologies improved the BaumRaganmodication Moreover supertrees can beuseful for comparative studies (Purvis1995b Bininda-Emonds et al 1999) whetherof adaptation coevolution rates of evo-lution cospeciation or rates of effectivecladogenesis where accuracy in the branchlength of phylogenetic reconstructions is notthe primary problem (Purvis et al 1994)Supertree reconstructions are also a usefulway to help highlight poor taxonomic sam-pling and identify where previous studiesareinconsistentThey can therefore be used as anexploratory tool capable of developing newhypotheses and indicating where future re-search should be focused
ACKNOWLEDGMENTS
We are grateful to Elizabeth Kellogg and LynnClark for providing the GPWG matrix and for their com-ments on the manuscript James Farris for helpful dis-cussion regarding bootstrap weighting schemes duringthe XIX Hennig Society meeting Stephen Renvoize andMark Chase for helpful discussion on the project OlafBininda-Emonds for his comments on the manuscriptand JonathanDavis for the testing of the SuperTree 085bprogram This study was supported by the Irish HigherEducation Authority the Roche Research Foundation(99-186) Enterprise Iceland (IC200129) the AgassizFoundation and the Swiss NSF (31-52378)
REFERENCES
(References marked curren were used as sources of phyloge-nies for the supertree reconstructions)
ARNQVIST G AND D WOOSTER 1995 Meta-analysisSynthesizing research ndings in ecology and evolu-tion Trends Ecol Evol 10236ndash240
currenBARKER N P 1997 The relationships of AmphipogonElytrophorus and Cyperochloa (Poaceae) as suggestedby rbcL sequence data Telopea 7205ndash213
currenBARKER N P H P LINDER AND E H HARLEY 1995Polyphyly of Arundinoids (Poaceae) Evidence fromrbcL sequence data Syst Bot 20423ndash435
currenBARKER N P H P LINDER and E H HARLEY 1999Sequences of the grass-specic insert in the chloro-plast rpoC2 gene elucidate generic relationships of theArundinoids (Poaceae) Syst Bot 23327ndash350
currenBAUM B R 1982 A phylogenetic analysis of the tribeTriticeae (Poaceae) based on morphological charactersof the genera Can J Bot 61518ndash535
BAUM B R 1992 Combining trees as a way of com-bining data sets for phylogenetic inference and thedesirability of combining gene trees Taxon 413ndash10
currenBAUM B R J R ESTES AND P K GUPTA 1986 Assess-ment of the genomic systematic classication in theTriticeae Am J Bot 741388ndash1395
BAUM B R AND M A RAGAN 1993 Reply to A GRodrigorsquos ldquoA comment on Baumrsquos method for com-bining phylogenetic treesrdquo Taxon 42637ndash640
BININD A-EMONDS O R P AND H N BRYANT 1998Properties of matrix representation with parsimonyanalyses Syst Biol 47497ndash508
BININD A-EMONDS O R P J L GITTLEMAN ANDA PURVIS 1999 Building large trees by combiningphylogenetic information A complete phylogenyof the extant Carnivora (Mammalia) Biol Rev 74143ndash175
BREMER K 1988 The limits of amino acid sequencedata in angiosperm phylogenetic reconstruction Evo-lution 42795ndash803
currenBUCKLER E S IV AND T P HOLTSFORD 1996 Zeasystematics Ribosomal ITS evidence Mol Biol Evol13612ndash622
CAMIN J H AND R R SOKAL 1965 A method for de-ducing branching sequences in phylogeny Evolution19311ndash326
currenCATALAN P E A KELLOGG AND R G OLMS TEAD1997 Phylogeny of Poaceae subfamily Pooideaebased on chloroplast ndhFgene sequences Mol Phy-logenet Evol 8150ndash166
currenCLARK L G W ZHANG AND J F WENDEL 1995 Aphylogenyof the grass family (Poaceae) basedon ndhFsequence data Syst Bot 20436ndash460
CONSTANTINES CU M AND D SANKOFF 1995 An ef-cient algorithm for supertrees J Classif 12101ndash112
currenCUMMINGS M P L M KING AND E A KELLOGG1994 Slipped-strand mispairing in a plastid generpoC2 in grasses (Poaceae) Mol Biol Evol 111ndash8
currenDARBYSHIRE S J AND S I WARWICK 1992 Phylogenyof North American Festuca (Poaceae) and related gen-era using chloroplast DNA restriction site variationCan J Bot 702415ndash2419
currenDAVIS J I AND R J SORENG 1993Phylogenetic struc-ture in the grass family (Poaceae) as inferred fromchloroplast DNA restriction site variation Am J Bot801444ndash1454
currenDOEBLEY J M DURBIN E M GOLENBERG M TCLEGG AND D P MA 1990 Evolutionary analysisof the large subunit of carboxylase (rbcL) nucleotidesequence among the grasses (Gramineae) Evolution441097ndash1108
currenDUVALL M R AND B R MORTON 1996 Molecularphylogenetics of Poaceae An expanded analysis ofrbcL sequence data Mol Phylogenet Evol 5352ndash358
currenDUVALL M R P M PETERSON E E TERRELL ANDA H CHRISTENS EN 1993 Phylogeny of NorthAmerican oryzoid grasses as constructed from mapsof plastid DNA restriction sites Am J Bot 8083ndash88
EFFRON B E HALLORAN AND S HOLMES 1996 Boot-strap condence levels for phylogenetic trees ProcNatl Acad Sci USA 937085ndash7090
currenESEN A AND K W HILU 1991 Electrophoretic andimmunological studies of prolamins in the PoaceaeII Phylogenetic afnities of the Atistideae Taxon 405ndash17
currenESEN A AND K W HILU 1993Prolamin and immuno-logical studies in the Poaceae IV SubfamilyPanicoidsCan J Bot 71315ndash322
FARRIS J S A G KLUGE AND M J ECKHARDT 1970 Anumerical approach to phylogenetic systematics SystZool 19172ndash191
FELSENSTEIN J 1985 Condence limits on phylogeniesAn approach using the bootstrap Evolution 39783ndash791
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 149
FITCH W M 1971 Toward dening the course of evolu-tion Minimal change for a specic tree topology SystZool 20406ndash416
currenFREDERIKSEN S AND O SEBERG 1991 Phylogeneticanalysis of the Triticeae (Poaceae) Hereditas 11515ndash19
currenGAUT B S A SPEEK BR MORTON AND M T CLEGG 1999Patterns of genetic diversication within the Adhgene family in the grasses (Poaceae) Mol Biol Evol161086ndash1097
GOLDMANN N J P ANDERSON AND A G RODRIGO2000 Likelihood-based tests of topologies in phyloge-netics Syst Biol 49652ndash670
GORDON A D 1980 On the assessment and compar-ison of classications Pages 149ndash160 in Analyse dedonnees et informatique (R Tomassone ed) INRALe Chesnay France
GPWG2000A phylogeny of the grass family (Poaceae)as inferred from eight character sets Pages 3ndash7 inGrasses Systematics and evolution (S W L Jacobsand J Everett eds) CSIRO Melbourne Australia
currenGREBENS TEIN B O GREBENS TEIN W SAUER ANDV HEMLEBEN 1996 Distribution and complex organi-zation of satellite DNA sequences in Aveneae speciesGenome 391045ndash1050
currenGREBENS TEIN B M ROSER W SAUER ANDV HEMLEBEN 1998 Molecular phylogenetic re-lationships in Aveneae (Poaceae) species and othergrasses as inferred from ITS1 and ITS2 rDNAsequences Plant Syst Evol 213233ndash250
currenHAMBY R K AND EA ZIMMER 1987Ribosomal RNAsequences for inferring phylogeny within the grassfamily (Poaceae) Plant Syst Evol 16029ndash37
HARVEY P H AND M D PAGEL 1991The comparativemethod in evolutionary biology Oxford Univ PressLondon
HILLIS D M 1996 Inferring complex phylogenies Na-ture 383130ndash131
HILLIS D M AND J J BULL 1993 An empirical test ofbootstrapping as a method for assessing condencein phylogenetic analysis Syst Biol 42182ndash192
currenHILU K W L A ALICE AND H LIANG 1999 Phy-logeny of Poaceae inferred from matK sequencesAnn Mo Bot Gard 86835ndash851
currenHILU K W AND A ESEN 1992Prolamin andimmuno-logical strudies in the Poaceae III Subfamily Chlori-doids Am J Bot 80104ndash113
currenHILU K W AND H LIANG 1997 The matK gene Se-quence variation and application in plant systematicsAm J Bot 84830ndash839
currenHILU K W AND K WRIGHT 1982 Systematics ofGramineae A cluster analysis study Taxon 319ndash37
currenHSIAO C N J CHATTERTON K H ASAY AND K BJENS EN 1993 Phylogenetic relationships of 10 grassspecies An assessment of phylogenetic utility of theinternal transcribed spacer region in nuclear riboso-mal DNA in monocots Genome 37112ndash120
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1994 Phylogenetic relationships of themonogenomic species of the wheat tribe Triticeae(Poaceae) inferred from nuclear rDNA (internal tran-scribed spacer) sequences Genome 38211ndash223
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1995Molecular phylogeny of the pooids(Poaceae) based on nuclear rDNA (ITS) sequencesTheor Appl Genet 90389ndash398
currenHSIAO C S W JACOBS N P BARKER AND N JCHATTERTON 1998 A molecular phylogeny of the
subfamily arundinoids (Poaceae) based on sequencesof rDNA Aust Syst Bot 1141ndash52
currenHSIAO C S W L JACOBS N J CHATTERTON AND KH ASAY 1999 A molecular phylogeny of the grassfamily (Poaceae) based on the sequences of nuclearribosomal DNA (ITS) Aust Syst Bot 11667ndash688
JOHNS ON L A AND D E SOLTIS 1998 Assessing con-gruence Empirical examples from molecular dataPages 297ndash348 in Molecular systematics of plants IIDNA sequencing (D E Soltis P S Soltis and J JDoyle eds) Kluwer Academic Publishers NorwellMassachusetts
currenKATAYAMA H AND Y OGIHARA 1996 Phylogeneticafnities of the grasses to other monocots as re-vealed by molecular analysis of chloroplast DNACurr Genet 29572ndash581
currenKELCHNER S A AND L G CLARK 1997 Molecu-lar evolution and phylogenetic utility of the chloro-plast rpl16 intron in Chusquea and the bambusoids(Poaceae) Mol Phylogenet Evol 8385ndash397
currenKELLOGG E A 1989 Comments on genomic genera inthe Triticeae (Poaceae) Am J Bot 76796ndash805
currenKELLOGG E A 1992Tools for studying the chloroplastgenome in the Triticeae (Gramineae) An ecoRI mapa diagnostic deletion and support for Bromus as anoutgroup Am J Bot 79186ndash197
currenKELLOGG E A 1998aRelationships of cereal crops andother grassesProc NatlAcad Sci USA952005ndash2010
currenKELLOGG E A 1998bWhorsquos related to whom Recentresults from molecular systematic studies Curr OpinPlant Biol 1149ndash158
currenKELLOGG E A AND R APPELS 1995 Intraspecic andinterspecic variation in 5S RNA genes are decoupledin diploid wheat relatives Genetics 140325ndash343
currenKELLOGG E A AND L WATSON 1993 Phyloge-netic studies of a large data set I bambusoids An-dropogonodae and pooids (Gramineae) Bot Rev59273ndash311
KIM J 1996 General inconsistency conditions for max-imum parsimony Effects of branch lengths and in-creasing numbers of taxa Syst Biol 45363ndash374
KISHINO H AND M HASEGAWA 1989 Evaluation ofthe maximum likelihood estimate of the evolution-ary tree topologies form DNA sequence data and thebranching order in Hominoidea J Mol Evol 29170ndash179
currenLIANG H AND K W HILU 1995 Application of thematK gene sequence to grass systematics Can J Bot74125ndash134
MABBERLEY D J 1993 The plant-book A portable dic-tionary of the vascular plants 2nd edition CambridgeUniv Press Cambridge
currenMACFARLANE T D AND L WATSON 1982The classi-cation of Poaceaesubfamily pooids Taxon31178ndash203
currenMASON-GAMER R J AND EA KELLOGG 1996Testingfor phylogenetic conict among molecular data sets inthe tribe Triticeae (Gramineae) Syst Biol 45524ndash545
currenMASON-GAMER R J C F WEIL AND E A KELLOGG1998Granule-bound starch synthase Structure func-tion andphylogenetic utility Mol Biol Evol 151658ndash1673
currenMATHEWS S AND R A SHARROCK 1996 The phy-tochrome gene family in grasses (Poaceae) A phy-logeny and evidence that grasses have a subset ofthe loci found in dicot angiosperms Mol Biol Evol131141ndash1150
currenMATHEWS S R C TSAL AND E A KELLOGG 2000Phylogenetic structure in the grass family (Poaceae)
150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel
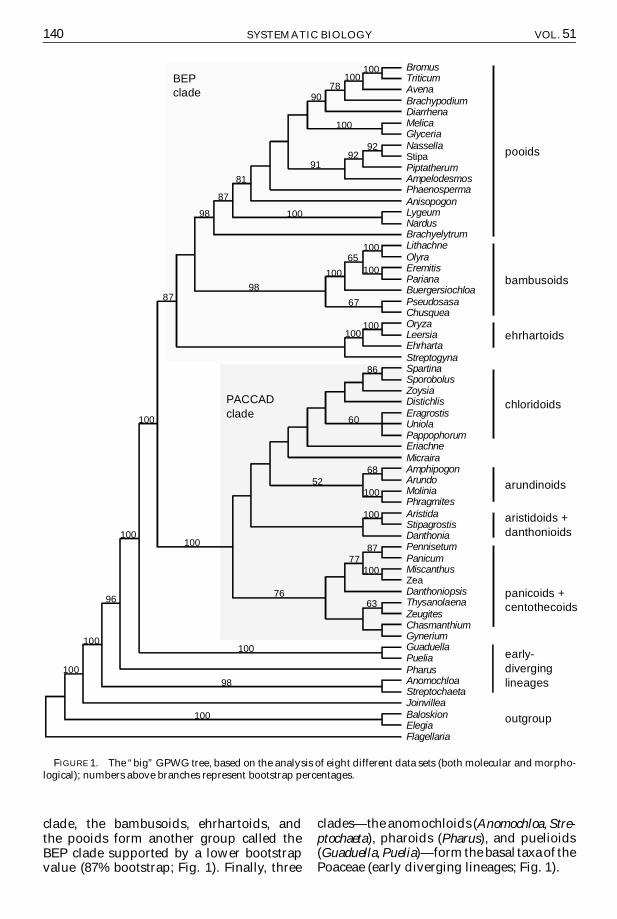
140 SYSTEMATIC BIOLOGY VOL 51
BEPclade
PACCADclade
early-diverginglineages
outgroup
100
100100
100
100
100
100
100100
100
100
100
100
100
100
100
100
100
100
96
98
76
7787
63
60
86
5268
67
65
9887
98
87
81
91
9292
7890
pooids
bambusoids
ehrhartoids
chloridoids
panicoids +centothecoids
arundinoids
aristidoids +danthonioids
BromusTriticumAvenaBrachypodiumDiarrhenaMelicaGlyceriaNassellaStipaPiptatherumAmpelodesmosPhaenospermaAnisopogonLygeumNardusBrachyelytrumLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaOryzaLeersiaEhrhartaStreptogynaSpartinaSporobolusZoysiaDistichlisEragrostisUniolaPappophorumEriachneMicrairaAmphipogonArundoMoliniaPhragmitesAristidaStipagrostisDanthoniaPennisetumPanicumMiscanthusZeaDanthoniopsis
ChasmanthiumGynerium
ThysanolaenaZeugites
GuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
FIGURE 1 The ldquobigrdquo GPWG tree based on the analysis of eight different data sets (both molecular and morpho-logical) numbers above branches represent bootstrap percentages
clade the bambusoids ehrhartoids andthe pooids form another group called theBEP clade supported by a lower bootstrapvalue (87 bootstrap Fig 1) Finally three
cladesmdashthe anomochloids (Anomochloa Stre-ptochaeta) pharoids (Pharus) and puelioids(Guaduella Puelia)mdashform the basal taxaof thePoaceae (early diverging lineages Fig 1)
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 141
TABLE 2 Partition metrics for the ve MRP modications expressed as distances A) Comparisons withinmodications with unordered characters B) within modications with irreversible characters and C) betweenmodications with unordered characters and modications with irreversible characters
Unordered characters
A) BR-alone BR C bootstrap BR C nodes PU-alone PU C bootstrap
Unordered BR-alone ndash characters BR C bootstrap 0457 ndash
BR C nodes 0379 0388 ndash PU-alone 0440 0362 0422 ndash PU C bootstrap 0474 0379 0457 0362 ndash
B) Irreversible characters
Irreversible BR-alone ndash characters BR C bootstrap 0431 ndash
BR C nodes 0216 0491 ndash PU-alone 0293 0431 0216 ndash PU C bootstrap 0414 0379 0440 0379 ndash
C) Irreversible characters
Unordered BR-alone 0000 0431 0216 0293 0414characters BR C bootstrap 0457 0112 0552 0491 0457
BR C nodes 0379 0483 0319 0328 0483PU-alone 0440 0457 0500 0422 0474PU C bootstrap 0474 0422 0517 0457 0181
GPWG Supertrees
The proportion of nonidentical nodes be-tween the ten MRP reconstructions takenby pairs was calculated by using the PM(Table 2) Analyses using unordered char-acters resulted in topologies with pairwisedistances of between 0362 and 0474 (meandistance 0412 Table 2) Pairwise com-parisons for each MRP modication us-ing irreversible characters gave distances ofbetween 0216 and 0491 (mean distance0369 Table 2) Values for each unorderedBaumRagan modication are closer totheir irreversible counterparts than to anyother methods (mean distance unorderedBRirreversible BR 0328mean distance un-ordered BRirreversible PU 0411 Table 2)The same pattern is found for the un-ordered Purvis modications (mean dis-tance unordered PUirreversible PU 0386mean unordered PUirreversible BR 0468Table 2) Three modications gave very sim-ilar topologies with both types of charac-ters BR-alone gave the exact same topologywhereas the two BR C bootstrap and thetwo PU C bootstrap values were very closeto each other (0112 and 0181 respectivelyTable 2)
For comparisons between the GPWG com-bined tree and the GPWG supertrees D1 dis-tances ranged from 032 for BR C bootstrapwith irreversible characters to 059 for PU-
alone with unordered characters the othereight modications had values from 042 to052 (Fig 2) When considering distancesobtained with PM BR C bootstrap withirreversible characters again gave the topol-ogy the most similar to the GPWG combinedtree yielding a distance of 025 The worstmodication was the BR C nodes with ir-reversible characters for which the distancewas 053 whereas those in the eight othermodications ranged from 041 to 050
We further examined the placement ofthe major subclades dened in the GPWGcombined tree (Fig 3) Two modicationsBR C bootstrap and PU C bootstrap bothwith irreversible characters are the onlymodications to yield the same basalbranching pattern as the combined GPWGtree (Fig 3A) In all the other modicationsinvolving irreversible characters two basalgrasses Guaduella and Puelia are insertedinside the BEP clade BR-alone placed thepooids as sister group to the PACCAD cladewhile BR C nodes and PU-alone placed aclade of the pooids and the ehrhartoids assister group of the PACCAD clade (Fig 3A)With unordered characters and exceptfor the BR-alone modication which wasidentical to the BR-alone with irreversiblecharacters modication all modicationshad an odd basal branching pattern theoutgroup being inserted between two cladesof the early diverging lineages (Fig 3B)
142 SYSTEMATIC BIOLOGY VOL 51
FIGURE 2 D1 greatest subtree agreementmetrics (black) and PM partition metrics (white) expressed asdistancesto the GPWG tree for each of the GPWG supertrees build from separate phylogenies based on the eight characterpartitions of the GPWG combined data set as source trees with characters considered as irreversible (left) andunordered (right)
The two different implementations of theKH-test were used to estimate the validity ofthe ten GPWG supertrees as possible alter-native hypothesis to the GPWG combinedtree (Table 3) Based on the characters ofthe GPWG combined data set the classicalKH-test rejected all the GPWG supertrees(P lt 0001) as suitable alternative phyloge-netic trees However the modied KH-testfound only the PU-alone with unorderedcharacters signicantly different (P lt 005)from the GPWG combined tree (Table 3)None of the other GPWG supertrees couldbe rejected BR C bootstrap with both char-acters types being once again the modica-tion giving the topology closest to that ofthe GPWG combined topology (P D 0588and P D 0624 for unordered and irreversiblecharacters respectively)
Finally Figure 4 shows the supertree ob-tained with BR C bootstrap and irreversiblecharacters The discrepancies between thissupertree and the GPWG combined tree weresmall and concerned mainly single taxa thatwere always positioned close to the cladeto which they belonged in the GPWG tree(Fig 4) The main differences concerned theplacement of the ehrhartoids and bambu-soids which exchanged their position as sis-ter group of the pooids in the supertree incomparison with the GPWG tree and theplacement of the three subfamilies arundi-noids aristidoids and danthonioids
Large SupertreeWe analyzed the 55 publications the same
way as the eight character partitions fromthe GPWG combined data set For clar-ity we do not show the results of BR Cnodes a MRP modication that resultedin topologies incompatible with the place-ment of subfamilies suggested in the GPWGcombined tree When irreversible characterswere used (Fig 5A) BR-alone BR C boot-strap and PU C bootstrap obtained the samebasal branching pattern but the placementof subfamilies inside the PACCAD cladewas slightly different in each case (data notshown) PU-alone was the only modica-tion to place the three subfamilies of theBEP clade together as a sister group to thePACCAD clade agreeing with the GPWG su-pertrees and the GPWG combined tree Withunordered characters (Fig 5B) BR-alone BRC bootstrapand PU-alone obtained the samebasal branching pattern with the BEP cladebeing paraphyletic PU C bootstrap gave anodd combination in which the early diverg-ing lineages were embedded inside the bam-busoids at the base of the tree (Fig 5B)
DISCUSSION
The information given by the two incon-gruence indices helped pinpoint the differ-ences present in the topologies obtained fromthe ten GPWG supertrees built using the
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 143
FIGURE 3 Summary of the subfamilial relationships in the GPWG supertrees obtained with the ve MRP mod-ications with column A) characters considered as irreversible and column B) characters considered as unordered
eight character partitions from the combineddata set Analyses using unordered charac-ters produced supertrees that were less sim-ilar as measured by D1 to the combinedtree than those obtained in analyses usingirreversible characters The only exceptionis for BR-alone which obtained the sametopology with both character types How-ever unordered charactersproduced slightlybetter topologies than irreversible charactersaccording to the PM incongruence index D1is dened as the number of taxa needed tobe removed from both trees to obtain an
identical subtree whereas PM is the numberof taxa bipartitions found between the twotrees being compared (Johnson and Soltis1998) Having higher D1 values than PMvalues suggests that the topological differ-ences do not involve single taxa (in whichcase D1 would be smaller than PM) butrather a large set of taxa placed differentlyin the supertree and the GPWG combinedtree Similar values of D1 and PM wouldsuggest that a few discrepant taxa wereinvolved none being far from their properlocations The D1 distances were much
144 SYSTEMATIC BIOLOGY VOL 51
TABLE 3 Kishino-Hasegawa test based on the difference in length between the GPWG combined tree and the tenGPWG supertrees obtained after applying the different MRP modications The p-values were calculated using thedefault parameter of PAUPcurren408b (Swofford 2000 classic Kishino-Hasegawa [KH] test) and Goldman et al (2000)modication to ensure the validity of the test with a posteriori specied topologies (modied KH test) Asterisk (curren)indicates a signicant p-value (p lt 005)
p-value
Number of steps Classic KH test Modied KH test
Combined GPWG 9054 ndash ndash
Unordered characters BR-alone 9291 lt00001curren 0172BR C bootstrap 9116 lt00001curren 0588BR C nodes 9203 lt00001curren 0396PU-alone 9411 lt00001curren 0042curren
PU C bootstrap 9211 lt00001curren 0374Irreversible characters BR-alone 9291 lt00001curren 0172
BR C bootstrap 9105 00003curren 0624BR C nodes 9281 lt00001curren 0198PU-alone 9311 lt00001curren 0144PU C bootstrap 9191 lt00001curren 0416
greater with unordered than irreversiblecharacters and were much closer to thePM ones obtained with irreversible char-acters Following this logic we can em-phasize that using unordered characters re-sulted in topologies with more differencesfrom the GPWG combined tree as to theplacement of larger subclades than did theuse of irreversible characters The type ofcharacters (ie unordered or irreversible)used to reconstruct the supertrees did notgreatly inuence the placement of the ter-minal taxa in the GPWG supertrees There-fore topologies obtained by consideringmatrix elements as irreversible characterswere closer to the combined phylogenetictree represented by the GPWG combinedanalysis This effect is clearly visible in theplacement of the major subclades dened inthe GPWG combined tree (Fig 4) Unorderedcharacters produced topologies in which theearly diverging lineages were split into twogroupsmdashone including Puelia and Guadu-ella and one including Anomochloa Strep-tochaeta and Pharus This extremely odd andunrealistic pattern does not correspond toany published phylogenetic trees concerningthe grass phylogeny Bininda-Emonds andBryant (1998) found only a minor impactof irreversible characters compared to un-ordered characters in their analysis but wefound that topologies created by using irre-versible characters are closer to an approachusing combined data than are the topolo-gies created by using unordered charactersThe resolution in the large supertrees was ex-tremely dependent on the type of characters
used to perform the analysis such was notthe case with the GPWG supertrees whereboth types of characters gave similar reso-lution (data not shown) Using irreversiblecharacters produced only one or two largesupertrees depending on the modicationused in contrast the ldquomaxtreesrdquo option inPAUPcurren408b was always reached when un-ordered characters were used Of course us-ing irreversible characters should put moreconstraintson the parsimonyanalysis by pre-venting the reversion from state 1 to 0 (thusreducing the number of most-parsimonioustrees) but it is not clear why such a drasticdifference appeared only between the largesupertrees
The Purvis coding scheme has been pro-posed as an improvement to the BaumRagan procedure as an attempt to reducethe dependency and redundancy betweenelements coming from the same topology(Purvis 1995a Ronquist 1996 Bininda-Emonds and Bryant 1998) However our re-sults suggest that the Purvis coding schemedoes not have a large impact on the MRPreconstructions and that the BaumRaganmethod works just as well or even betterComparisons between the approach usingcombined data and BR-alone or combineddata and PU-alone gave similar values forboth PM and D1 (Fig 2) Comparisons be-tween MRP modications also indicated aclose relationship between the two modi-cations especially with irreversible charac-ters (Table 2) This conclusion is less wellsupported with unordered characters be-cause BR-alone is the only modication to
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 145
PACCADclade
BEPclade
early-diverginglineages
outgroup
NassellaStipaPiptatherumAmpelodesmosMelicaGlyceriaBromusTriticumAvenaBrachypodiumDiarrhenaAnisopogonLygeumNardusBrachyelytrumPhaenospermaOryzaLeersiaEhrhartaStreptogynaLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaPennisetumPanicumMiscanthusZeaGyneriumDanthoniopsisZeugitesChasmanthiumThysanolaenaAmphipogonArundoMoliniaPhragmitesSpartinaSporobolusZoysiaDistichlisEragrostisPappophorumUniolaDanthoniaMicrairaAristidaStipagrostisEriachneGuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
pooids
bambusoids
ehrhartoids
chloridoids +danthonioids
panicoids
arundinoids
aristidoids
centothecoids
FIGURE 4 Grass supertree obtained with Baum and Ragan coding scheme with bootstrap support weightingusing separate phylogenies built from the eight character partitions of the GPWG combined data set as source treesDashed lines indicate incongruent branches between the supertree and the GPWG combined tree
146 SYSTEMATIC BIOLOGY VOL 51
FIGURE 5 Summary of the subfamilial relationships in the large supertrees based on 55 published source trees(including 403 genera) obtained using the ve MRP modications with column A) characters considered as irre-versible and column B) characters considered as unordered
give a basal topology very different fromthat obtained with any other modication(Table 2 and Fig 2) Ronquistrsquos (1996) propo-sition to weight each character in the bi-nary matrix by the inverse of the number ofnodes in the corresponding source tree doesnot perform as well as the other modica-tions It yields the greatest partition metricdistance and the second largest agreementsubtree distance (Fig 2) Although the twoalternatives to the BaumRagan method arebased on logical and plain arguments (ienonindependence and redundancy of matrixelements and impact of larger trees) theireffects on MRP reconstruction are not obvi-ous and do not result in topologies closerto our combined reference Indeed PU-alonewith unordered characters was rejected as asuitable alternative hypothesis to the GPWGcombined tree with the modied KH-testand gave the second smallest P-value when
used with irreversible characters (Table 2)However conclusions from the large su-pertree analysis differed because PU-alonewas the only modication to produce thesame basal branching pattern as the GPWGcombined tree (Fig 5B) whereas BR-alonewith the pooids as sister group to the PAC-CAD clade corresponded to an alternativehypothesis for the grass family that is sup-ported by some data from nuclear (Adh Gautet al 1999) and plastid (rbcL Duvall andMorton 1996) genomes Because no refer-ence phylogenetic trees are available thatcontain a number of taxa similar to those ofour large supertrees it is difcult to assesstheir topologies and to determine whetherPU-alone with irreversible characters reallygave a more accurate phylogeny of largegrasses than did the other methods Theplacement of the pooids is possibly not thebest criterion by which to judge the methods
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 147
the evolutionary position of this subfamilywithin the grass family differs as inferredwith different DNA sequence data sets
Weighting by node support as suggestedby Ronquist (1996) improved the t betweenthe additive binary matrices and the GPWGcombined tree The benecial impact of thebootstrap weighting scheme is evident fromthe increased P-values obtained with themodied KH-test (Table 2) This trend is vis-ible for the BaumRagan coding schemebut is less obvious with the Purvis codingscheme (Fig 2) Bootstrap weighting wasproposed (Ronquist 1996) as an alternativeto the Purvis coding scheme both to reducethe greater impact of large source trees oversmalleronesand to improve the effect of well-supported nodes in the MRP analysis It isnot clear how bootstrap weighting could re-duce the impact of larger trees if the smallertrees do not have much greater support val-ues than those of the larger trees Howeverweighting of charactersby bootstrapsupportwithin the Purvis coding scheme appears tobe a redundant procedure unable to greatlyimprove the MRP reconstruction from theimportant information present in bootstrapsupport values (Fig 2) In the large supertreeanalysis PU C bootstrap with unorderedcharacters produced unrealistic topologieswith the early diverging lineages embed-ded in the bambusoids (Fig 5B) The Purviscoding schemes removes important restric-tive information from the matrix (Bininda-Emonds and Bryant 1998) possibly there-fore weighting by bootstrap support wouldrandomly assign high values to characters inthe matrix where this restrictive informationhas been removed and would prevent theweighting schemes from being as effectiveas with the BaumRagan coding schemesWeighting the matrix elements by node sup-port also poses problems Not all bootstrapanalyses can be considered identical and thenumber of replicates the type of searchesdone or both will inuence the supportfound Moreover when node support is notprovided for a source tree weights cannot beassigned and information from this tree willbe down-weighted in the MRP analysis Theeffect of this is great in supertree reconstruc-tions done with older publications which ingeneral do not report node support and arehence down-weighted in the MRP analyses
An important aspect affecting the large su-pertrees that is not resolved in our compar-
isons is the placement of some rarely sam-pled taxa Taxa that are present in only a fewpublications have been allocated a high pro-portion of missing character values whichmakes parsimony analyses much more dif-cult How many of these taxa are mis-placed is difcult to determine because noreference phylogeny for the 403 genera isavailable MRP reconstruction done withoutconstraining three clades (panicoids pooidsand chloridoids) to be monophyletic endedwith rarely sampled taxa being scattered allaround the supertrees (data not shown) Thatcan be an important problem when MRPanalyses are performed on a wide taxonomicgroup comprising large numbers of rarelysampled taxa
MRP coding especially when weighted bynode support can be considered an indica-tion of the signal in the primary data witheach node represented by one (weighted)synapomorphy (see Bininda-Emonds et al1999) However we stress that supertreescannot be viewed and interpreted in ex-actly the same way as phylogenetic analy-ses based on biological characters One ma-jor problem is the difculty of assigningnode support for supertrees Bootstrappingand other resampling procedures cannot beapplied given the clear nonindependenceof the characters present in the binary ma-trix and Bremer support (Bremer 1988) iscertainly equally affected by such noninde-pendence (as is the parsimony itself hencethe different MRP modications proposed)Moreoverbranch lengths associatedwith su-pertrees are treated in the same way as mor-phological branch lengths and unlike molec-ular branch length they cannot be used toestablish divergence times This weakness iseven more signicant in taxonomic groupssuch as the grasses for which dates are dif-cult to gather from other sources (eg fossilrecords)
CONCLUSIONS
We have demonstrated that supertreesoffer an easy way of producing phyloge-nies with a large number of taxa and thatthey can give good estimates of relation-ships within these groups (in this case thegrass family) Supertrees based on the eightcharacter partitions of the GPWG data setswere found to roughly match the combinedanalysis the discrepancies of the best MRP
148 SYSTEMATIC BIOLOGY VOL 51
modications being found mainly in weaklysupported branches of the combined treeThe BaumRagan and the Purvis modica-tions gave similar results but incorporatingbootstrap support associated with preexist-ing topologies improved the BaumRaganmodication Moreover supertrees can beuseful for comparative studies (Purvis1995b Bininda-Emonds et al 1999) whetherof adaptation coevolution rates of evo-lution cospeciation or rates of effectivecladogenesis where accuracy in the branchlength of phylogenetic reconstructions is notthe primary problem (Purvis et al 1994)Supertree reconstructions are also a usefulway to help highlight poor taxonomic sam-pling and identify where previous studiesareinconsistentThey can therefore be used as anexploratory tool capable of developing newhypotheses and indicating where future re-search should be focused
ACKNOWLEDGMENTS
We are grateful to Elizabeth Kellogg and LynnClark for providing the GPWG matrix and for their com-ments on the manuscript James Farris for helpful dis-cussion regarding bootstrap weighting schemes duringthe XIX Hennig Society meeting Stephen Renvoize andMark Chase for helpful discussion on the project OlafBininda-Emonds for his comments on the manuscriptand JonathanDavis for the testing of the SuperTree 085bprogram This study was supported by the Irish HigherEducation Authority the Roche Research Foundation(99-186) Enterprise Iceland (IC200129) the AgassizFoundation and the Swiss NSF (31-52378)
REFERENCES
(References marked curren were used as sources of phyloge-nies for the supertree reconstructions)
ARNQVIST G AND D WOOSTER 1995 Meta-analysisSynthesizing research ndings in ecology and evolu-tion Trends Ecol Evol 10236ndash240
currenBARKER N P 1997 The relationships of AmphipogonElytrophorus and Cyperochloa (Poaceae) as suggestedby rbcL sequence data Telopea 7205ndash213
currenBARKER N P H P LINDER AND E H HARLEY 1995Polyphyly of Arundinoids (Poaceae) Evidence fromrbcL sequence data Syst Bot 20423ndash435
currenBARKER N P H P LINDER and E H HARLEY 1999Sequences of the grass-specic insert in the chloro-plast rpoC2 gene elucidate generic relationships of theArundinoids (Poaceae) Syst Bot 23327ndash350
currenBAUM B R 1982 A phylogenetic analysis of the tribeTriticeae (Poaceae) based on morphological charactersof the genera Can J Bot 61518ndash535
BAUM B R 1992 Combining trees as a way of com-bining data sets for phylogenetic inference and thedesirability of combining gene trees Taxon 413ndash10
currenBAUM B R J R ESTES AND P K GUPTA 1986 Assess-ment of the genomic systematic classication in theTriticeae Am J Bot 741388ndash1395
BAUM B R AND M A RAGAN 1993 Reply to A GRodrigorsquos ldquoA comment on Baumrsquos method for com-bining phylogenetic treesrdquo Taxon 42637ndash640
BININD A-EMONDS O R P AND H N BRYANT 1998Properties of matrix representation with parsimonyanalyses Syst Biol 47497ndash508
BININD A-EMONDS O R P J L GITTLEMAN ANDA PURVIS 1999 Building large trees by combiningphylogenetic information A complete phylogenyof the extant Carnivora (Mammalia) Biol Rev 74143ndash175
BREMER K 1988 The limits of amino acid sequencedata in angiosperm phylogenetic reconstruction Evo-lution 42795ndash803
currenBUCKLER E S IV AND T P HOLTSFORD 1996 Zeasystematics Ribosomal ITS evidence Mol Biol Evol13612ndash622
CAMIN J H AND R R SOKAL 1965 A method for de-ducing branching sequences in phylogeny Evolution19311ndash326
currenCATALAN P E A KELLOGG AND R G OLMS TEAD1997 Phylogeny of Poaceae subfamily Pooideaebased on chloroplast ndhFgene sequences Mol Phy-logenet Evol 8150ndash166
currenCLARK L G W ZHANG AND J F WENDEL 1995 Aphylogenyof the grass family (Poaceae) basedon ndhFsequence data Syst Bot 20436ndash460
CONSTANTINES CU M AND D SANKOFF 1995 An ef-cient algorithm for supertrees J Classif 12101ndash112
currenCUMMINGS M P L M KING AND E A KELLOGG1994 Slipped-strand mispairing in a plastid generpoC2 in grasses (Poaceae) Mol Biol Evol 111ndash8
currenDARBYSHIRE S J AND S I WARWICK 1992 Phylogenyof North American Festuca (Poaceae) and related gen-era using chloroplast DNA restriction site variationCan J Bot 702415ndash2419
currenDAVIS J I AND R J SORENG 1993Phylogenetic struc-ture in the grass family (Poaceae) as inferred fromchloroplast DNA restriction site variation Am J Bot801444ndash1454
currenDOEBLEY J M DURBIN E M GOLENBERG M TCLEGG AND D P MA 1990 Evolutionary analysisof the large subunit of carboxylase (rbcL) nucleotidesequence among the grasses (Gramineae) Evolution441097ndash1108
currenDUVALL M R AND B R MORTON 1996 Molecularphylogenetics of Poaceae An expanded analysis ofrbcL sequence data Mol Phylogenet Evol 5352ndash358
currenDUVALL M R P M PETERSON E E TERRELL ANDA H CHRISTENS EN 1993 Phylogeny of NorthAmerican oryzoid grasses as constructed from mapsof plastid DNA restriction sites Am J Bot 8083ndash88
EFFRON B E HALLORAN AND S HOLMES 1996 Boot-strap condence levels for phylogenetic trees ProcNatl Acad Sci USA 937085ndash7090
currenESEN A AND K W HILU 1991 Electrophoretic andimmunological studies of prolamins in the PoaceaeII Phylogenetic afnities of the Atistideae Taxon 405ndash17
currenESEN A AND K W HILU 1993Prolamin and immuno-logical studies in the Poaceae IV SubfamilyPanicoidsCan J Bot 71315ndash322
FARRIS J S A G KLUGE AND M J ECKHARDT 1970 Anumerical approach to phylogenetic systematics SystZool 19172ndash191
FELSENSTEIN J 1985 Condence limits on phylogeniesAn approach using the bootstrap Evolution 39783ndash791
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 149
FITCH W M 1971 Toward dening the course of evolu-tion Minimal change for a specic tree topology SystZool 20406ndash416
currenFREDERIKSEN S AND O SEBERG 1991 Phylogeneticanalysis of the Triticeae (Poaceae) Hereditas 11515ndash19
currenGAUT B S A SPEEK BR MORTON AND M T CLEGG 1999Patterns of genetic diversication within the Adhgene family in the grasses (Poaceae) Mol Biol Evol161086ndash1097
GOLDMANN N J P ANDERSON AND A G RODRIGO2000 Likelihood-based tests of topologies in phyloge-netics Syst Biol 49652ndash670
GORDON A D 1980 On the assessment and compar-ison of classications Pages 149ndash160 in Analyse dedonnees et informatique (R Tomassone ed) INRALe Chesnay France
GPWG2000A phylogeny of the grass family (Poaceae)as inferred from eight character sets Pages 3ndash7 inGrasses Systematics and evolution (S W L Jacobsand J Everett eds) CSIRO Melbourne Australia
currenGREBENS TEIN B O GREBENS TEIN W SAUER ANDV HEMLEBEN 1996 Distribution and complex organi-zation of satellite DNA sequences in Aveneae speciesGenome 391045ndash1050
currenGREBENS TEIN B M ROSER W SAUER ANDV HEMLEBEN 1998 Molecular phylogenetic re-lationships in Aveneae (Poaceae) species and othergrasses as inferred from ITS1 and ITS2 rDNAsequences Plant Syst Evol 213233ndash250
currenHAMBY R K AND EA ZIMMER 1987Ribosomal RNAsequences for inferring phylogeny within the grassfamily (Poaceae) Plant Syst Evol 16029ndash37
HARVEY P H AND M D PAGEL 1991The comparativemethod in evolutionary biology Oxford Univ PressLondon
HILLIS D M 1996 Inferring complex phylogenies Na-ture 383130ndash131
HILLIS D M AND J J BULL 1993 An empirical test ofbootstrapping as a method for assessing condencein phylogenetic analysis Syst Biol 42182ndash192
currenHILU K W L A ALICE AND H LIANG 1999 Phy-logeny of Poaceae inferred from matK sequencesAnn Mo Bot Gard 86835ndash851
currenHILU K W AND A ESEN 1992Prolamin andimmuno-logical strudies in the Poaceae III Subfamily Chlori-doids Am J Bot 80104ndash113
currenHILU K W AND H LIANG 1997 The matK gene Se-quence variation and application in plant systematicsAm J Bot 84830ndash839
currenHILU K W AND K WRIGHT 1982 Systematics ofGramineae A cluster analysis study Taxon 319ndash37
currenHSIAO C N J CHATTERTON K H ASAY AND K BJENS EN 1993 Phylogenetic relationships of 10 grassspecies An assessment of phylogenetic utility of theinternal transcribed spacer region in nuclear riboso-mal DNA in monocots Genome 37112ndash120
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1994 Phylogenetic relationships of themonogenomic species of the wheat tribe Triticeae(Poaceae) inferred from nuclear rDNA (internal tran-scribed spacer) sequences Genome 38211ndash223
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1995Molecular phylogeny of the pooids(Poaceae) based on nuclear rDNA (ITS) sequencesTheor Appl Genet 90389ndash398
currenHSIAO C S W JACOBS N P BARKER AND N JCHATTERTON 1998 A molecular phylogeny of the
subfamily arundinoids (Poaceae) based on sequencesof rDNA Aust Syst Bot 1141ndash52
currenHSIAO C S W L JACOBS N J CHATTERTON AND KH ASAY 1999 A molecular phylogeny of the grassfamily (Poaceae) based on the sequences of nuclearribosomal DNA (ITS) Aust Syst Bot 11667ndash688
JOHNS ON L A AND D E SOLTIS 1998 Assessing con-gruence Empirical examples from molecular dataPages 297ndash348 in Molecular systematics of plants IIDNA sequencing (D E Soltis P S Soltis and J JDoyle eds) Kluwer Academic Publishers NorwellMassachusetts
currenKATAYAMA H AND Y OGIHARA 1996 Phylogeneticafnities of the grasses to other monocots as re-vealed by molecular analysis of chloroplast DNACurr Genet 29572ndash581
currenKELCHNER S A AND L G CLARK 1997 Molecu-lar evolution and phylogenetic utility of the chloro-plast rpl16 intron in Chusquea and the bambusoids(Poaceae) Mol Phylogenet Evol 8385ndash397
currenKELLOGG E A 1989 Comments on genomic genera inthe Triticeae (Poaceae) Am J Bot 76796ndash805
currenKELLOGG E A 1992Tools for studying the chloroplastgenome in the Triticeae (Gramineae) An ecoRI mapa diagnostic deletion and support for Bromus as anoutgroup Am J Bot 79186ndash197
currenKELLOGG E A 1998aRelationships of cereal crops andother grassesProc NatlAcad Sci USA952005ndash2010
currenKELLOGG E A 1998bWhorsquos related to whom Recentresults from molecular systematic studies Curr OpinPlant Biol 1149ndash158
currenKELLOGG E A AND R APPELS 1995 Intraspecic andinterspecic variation in 5S RNA genes are decoupledin diploid wheat relatives Genetics 140325ndash343
currenKELLOGG E A AND L WATSON 1993 Phyloge-netic studies of a large data set I bambusoids An-dropogonodae and pooids (Gramineae) Bot Rev59273ndash311
KIM J 1996 General inconsistency conditions for max-imum parsimony Effects of branch lengths and in-creasing numbers of taxa Syst Biol 45363ndash374
KISHINO H AND M HASEGAWA 1989 Evaluation ofthe maximum likelihood estimate of the evolution-ary tree topologies form DNA sequence data and thebranching order in Hominoidea J Mol Evol 29170ndash179
currenLIANG H AND K W HILU 1995 Application of thematK gene sequence to grass systematics Can J Bot74125ndash134
MABBERLEY D J 1993 The plant-book A portable dic-tionary of the vascular plants 2nd edition CambridgeUniv Press Cambridge
currenMACFARLANE T D AND L WATSON 1982The classi-cation of Poaceaesubfamily pooids Taxon31178ndash203
currenMASON-GAMER R J AND EA KELLOGG 1996Testingfor phylogenetic conict among molecular data sets inthe tribe Triticeae (Gramineae) Syst Biol 45524ndash545
currenMASON-GAMER R J C F WEIL AND E A KELLOGG1998Granule-bound starch synthase Structure func-tion andphylogenetic utility Mol Biol Evol 151658ndash1673
currenMATHEWS S AND R A SHARROCK 1996 The phy-tochrome gene family in grasses (Poaceae) A phy-logeny and evidence that grasses have a subset ofthe loci found in dicot angiosperms Mol Biol Evol131141ndash1150
currenMATHEWS S R C TSAL AND E A KELLOGG 2000Phylogenetic structure in the grass family (Poaceae)
150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel
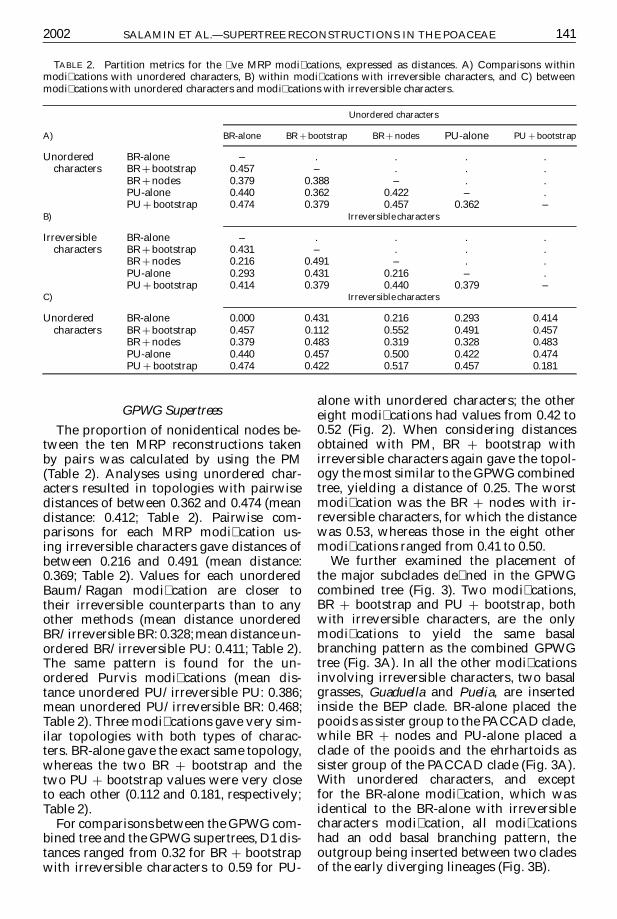
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 141
TABLE 2 Partition metrics for the ve MRP modications expressed as distances A) Comparisons withinmodications with unordered characters B) within modications with irreversible characters and C) betweenmodications with unordered characters and modications with irreversible characters
Unordered characters
A) BR-alone BR C bootstrap BR C nodes PU-alone PU C bootstrap
Unordered BR-alone ndash characters BR C bootstrap 0457 ndash
BR C nodes 0379 0388 ndash PU-alone 0440 0362 0422 ndash PU C bootstrap 0474 0379 0457 0362 ndash
B) Irreversible characters
Irreversible BR-alone ndash characters BR C bootstrap 0431 ndash
BR C nodes 0216 0491 ndash PU-alone 0293 0431 0216 ndash PU C bootstrap 0414 0379 0440 0379 ndash
C) Irreversible characters
Unordered BR-alone 0000 0431 0216 0293 0414characters BR C bootstrap 0457 0112 0552 0491 0457
BR C nodes 0379 0483 0319 0328 0483PU-alone 0440 0457 0500 0422 0474PU C bootstrap 0474 0422 0517 0457 0181
GPWG Supertrees
The proportion of nonidentical nodes be-tween the ten MRP reconstructions takenby pairs was calculated by using the PM(Table 2) Analyses using unordered char-acters resulted in topologies with pairwisedistances of between 0362 and 0474 (meandistance 0412 Table 2) Pairwise com-parisons for each MRP modication us-ing irreversible characters gave distances ofbetween 0216 and 0491 (mean distance0369 Table 2) Values for each unorderedBaumRagan modication are closer totheir irreversible counterparts than to anyother methods (mean distance unorderedBRirreversible BR 0328mean distance un-ordered BRirreversible PU 0411 Table 2)The same pattern is found for the un-ordered Purvis modications (mean dis-tance unordered PUirreversible PU 0386mean unordered PUirreversible BR 0468Table 2) Three modications gave very sim-ilar topologies with both types of charac-ters BR-alone gave the exact same topologywhereas the two BR C bootstrap and thetwo PU C bootstrap values were very closeto each other (0112 and 0181 respectivelyTable 2)
For comparisons between the GPWG com-bined tree and the GPWG supertrees D1 dis-tances ranged from 032 for BR C bootstrapwith irreversible characters to 059 for PU-
alone with unordered characters the othereight modications had values from 042 to052 (Fig 2) When considering distancesobtained with PM BR C bootstrap withirreversible characters again gave the topol-ogy the most similar to the GPWG combinedtree yielding a distance of 025 The worstmodication was the BR C nodes with ir-reversible characters for which the distancewas 053 whereas those in the eight othermodications ranged from 041 to 050
We further examined the placement ofthe major subclades dened in the GPWGcombined tree (Fig 3) Two modicationsBR C bootstrap and PU C bootstrap bothwith irreversible characters are the onlymodications to yield the same basalbranching pattern as the combined GPWGtree (Fig 3A) In all the other modicationsinvolving irreversible characters two basalgrasses Guaduella and Puelia are insertedinside the BEP clade BR-alone placed thepooids as sister group to the PACCAD cladewhile BR C nodes and PU-alone placed aclade of the pooids and the ehrhartoids assister group of the PACCAD clade (Fig 3A)With unordered characters and exceptfor the BR-alone modication which wasidentical to the BR-alone with irreversiblecharacters modication all modicationshad an odd basal branching pattern theoutgroup being inserted between two cladesof the early diverging lineages (Fig 3B)
142 SYSTEMATIC BIOLOGY VOL 51
FIGURE 2 D1 greatest subtree agreementmetrics (black) and PM partition metrics (white) expressed asdistancesto the GPWG tree for each of the GPWG supertrees build from separate phylogenies based on the eight characterpartitions of the GPWG combined data set as source trees with characters considered as irreversible (left) andunordered (right)
The two different implementations of theKH-test were used to estimate the validity ofthe ten GPWG supertrees as possible alter-native hypothesis to the GPWG combinedtree (Table 3) Based on the characters ofthe GPWG combined data set the classicalKH-test rejected all the GPWG supertrees(P lt 0001) as suitable alternative phyloge-netic trees However the modied KH-testfound only the PU-alone with unorderedcharacters signicantly different (P lt 005)from the GPWG combined tree (Table 3)None of the other GPWG supertrees couldbe rejected BR C bootstrap with both char-acters types being once again the modica-tion giving the topology closest to that ofthe GPWG combined topology (P D 0588and P D 0624 for unordered and irreversiblecharacters respectively)
Finally Figure 4 shows the supertree ob-tained with BR C bootstrap and irreversiblecharacters The discrepancies between thissupertree and the GPWG combined tree weresmall and concerned mainly single taxa thatwere always positioned close to the cladeto which they belonged in the GPWG tree(Fig 4) The main differences concerned theplacement of the ehrhartoids and bambu-soids which exchanged their position as sis-ter group of the pooids in the supertree incomparison with the GPWG tree and theplacement of the three subfamilies arundi-noids aristidoids and danthonioids
Large SupertreeWe analyzed the 55 publications the same
way as the eight character partitions fromthe GPWG combined data set For clar-ity we do not show the results of BR Cnodes a MRP modication that resultedin topologies incompatible with the place-ment of subfamilies suggested in the GPWGcombined tree When irreversible characterswere used (Fig 5A) BR-alone BR C boot-strap and PU C bootstrap obtained the samebasal branching pattern but the placementof subfamilies inside the PACCAD cladewas slightly different in each case (data notshown) PU-alone was the only modica-tion to place the three subfamilies of theBEP clade together as a sister group to thePACCAD clade agreeing with the GPWG su-pertrees and the GPWG combined tree Withunordered characters (Fig 5B) BR-alone BRC bootstrapand PU-alone obtained the samebasal branching pattern with the BEP cladebeing paraphyletic PU C bootstrap gave anodd combination in which the early diverg-ing lineages were embedded inside the bam-busoids at the base of the tree (Fig 5B)
DISCUSSION
The information given by the two incon-gruence indices helped pinpoint the differ-ences present in the topologies obtained fromthe ten GPWG supertrees built using the
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 143
FIGURE 3 Summary of the subfamilial relationships in the GPWG supertrees obtained with the ve MRP mod-ications with column A) characters considered as irreversible and column B) characters considered as unordered
eight character partitions from the combineddata set Analyses using unordered charac-ters produced supertrees that were less sim-ilar as measured by D1 to the combinedtree than those obtained in analyses usingirreversible characters The only exceptionis for BR-alone which obtained the sametopology with both character types How-ever unordered charactersproduced slightlybetter topologies than irreversible charactersaccording to the PM incongruence index D1is dened as the number of taxa needed tobe removed from both trees to obtain an
identical subtree whereas PM is the numberof taxa bipartitions found between the twotrees being compared (Johnson and Soltis1998) Having higher D1 values than PMvalues suggests that the topological differ-ences do not involve single taxa (in whichcase D1 would be smaller than PM) butrather a large set of taxa placed differentlyin the supertree and the GPWG combinedtree Similar values of D1 and PM wouldsuggest that a few discrepant taxa wereinvolved none being far from their properlocations The D1 distances were much
144 SYSTEMATIC BIOLOGY VOL 51
TABLE 3 Kishino-Hasegawa test based on the difference in length between the GPWG combined tree and the tenGPWG supertrees obtained after applying the different MRP modications The p-values were calculated using thedefault parameter of PAUPcurren408b (Swofford 2000 classic Kishino-Hasegawa [KH] test) and Goldman et al (2000)modication to ensure the validity of the test with a posteriori specied topologies (modied KH test) Asterisk (curren)indicates a signicant p-value (p lt 005)
p-value
Number of steps Classic KH test Modied KH test
Combined GPWG 9054 ndash ndash
Unordered characters BR-alone 9291 lt00001curren 0172BR C bootstrap 9116 lt00001curren 0588BR C nodes 9203 lt00001curren 0396PU-alone 9411 lt00001curren 0042curren
PU C bootstrap 9211 lt00001curren 0374Irreversible characters BR-alone 9291 lt00001curren 0172
BR C bootstrap 9105 00003curren 0624BR C nodes 9281 lt00001curren 0198PU-alone 9311 lt00001curren 0144PU C bootstrap 9191 lt00001curren 0416
greater with unordered than irreversiblecharacters and were much closer to thePM ones obtained with irreversible char-acters Following this logic we can em-phasize that using unordered characters re-sulted in topologies with more differencesfrom the GPWG combined tree as to theplacement of larger subclades than did theuse of irreversible characters The type ofcharacters (ie unordered or irreversible)used to reconstruct the supertrees did notgreatly inuence the placement of the ter-minal taxa in the GPWG supertrees There-fore topologies obtained by consideringmatrix elements as irreversible characterswere closer to the combined phylogenetictree represented by the GPWG combinedanalysis This effect is clearly visible in theplacement of the major subclades dened inthe GPWG combined tree (Fig 4) Unorderedcharacters produced topologies in which theearly diverging lineages were split into twogroupsmdashone including Puelia and Guadu-ella and one including Anomochloa Strep-tochaeta and Pharus This extremely odd andunrealistic pattern does not correspond toany published phylogenetic trees concerningthe grass phylogeny Bininda-Emonds andBryant (1998) found only a minor impactof irreversible characters compared to un-ordered characters in their analysis but wefound that topologies created by using irre-versible characters are closer to an approachusing combined data than are the topolo-gies created by using unordered charactersThe resolution in the large supertrees was ex-tremely dependent on the type of characters
used to perform the analysis such was notthe case with the GPWG supertrees whereboth types of characters gave similar reso-lution (data not shown) Using irreversiblecharacters produced only one or two largesupertrees depending on the modicationused in contrast the ldquomaxtreesrdquo option inPAUPcurren408b was always reached when un-ordered characters were used Of course us-ing irreversible characters should put moreconstraintson the parsimonyanalysis by pre-venting the reversion from state 1 to 0 (thusreducing the number of most-parsimonioustrees) but it is not clear why such a drasticdifference appeared only between the largesupertrees
The Purvis coding scheme has been pro-posed as an improvement to the BaumRagan procedure as an attempt to reducethe dependency and redundancy betweenelements coming from the same topology(Purvis 1995a Ronquist 1996 Bininda-Emonds and Bryant 1998) However our re-sults suggest that the Purvis coding schemedoes not have a large impact on the MRPreconstructions and that the BaumRaganmethod works just as well or even betterComparisons between the approach usingcombined data and BR-alone or combineddata and PU-alone gave similar values forboth PM and D1 (Fig 2) Comparisons be-tween MRP modications also indicated aclose relationship between the two modi-cations especially with irreversible charac-ters (Table 2) This conclusion is less wellsupported with unordered characters be-cause BR-alone is the only modication to
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 145
PACCADclade
BEPclade
early-diverginglineages
outgroup
NassellaStipaPiptatherumAmpelodesmosMelicaGlyceriaBromusTriticumAvenaBrachypodiumDiarrhenaAnisopogonLygeumNardusBrachyelytrumPhaenospermaOryzaLeersiaEhrhartaStreptogynaLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaPennisetumPanicumMiscanthusZeaGyneriumDanthoniopsisZeugitesChasmanthiumThysanolaenaAmphipogonArundoMoliniaPhragmitesSpartinaSporobolusZoysiaDistichlisEragrostisPappophorumUniolaDanthoniaMicrairaAristidaStipagrostisEriachneGuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
pooids
bambusoids
ehrhartoids
chloridoids +danthonioids
panicoids
arundinoids
aristidoids
centothecoids
FIGURE 4 Grass supertree obtained with Baum and Ragan coding scheme with bootstrap support weightingusing separate phylogenies built from the eight character partitions of the GPWG combined data set as source treesDashed lines indicate incongruent branches between the supertree and the GPWG combined tree
146 SYSTEMATIC BIOLOGY VOL 51
FIGURE 5 Summary of the subfamilial relationships in the large supertrees based on 55 published source trees(including 403 genera) obtained using the ve MRP modications with column A) characters considered as irre-versible and column B) characters considered as unordered
give a basal topology very different fromthat obtained with any other modication(Table 2 and Fig 2) Ronquistrsquos (1996) propo-sition to weight each character in the bi-nary matrix by the inverse of the number ofnodes in the corresponding source tree doesnot perform as well as the other modica-tions It yields the greatest partition metricdistance and the second largest agreementsubtree distance (Fig 2) Although the twoalternatives to the BaumRagan method arebased on logical and plain arguments (ienonindependence and redundancy of matrixelements and impact of larger trees) theireffects on MRP reconstruction are not obvi-ous and do not result in topologies closerto our combined reference Indeed PU-alonewith unordered characters was rejected as asuitable alternative hypothesis to the GPWGcombined tree with the modied KH-testand gave the second smallest P-value when
used with irreversible characters (Table 2)However conclusions from the large su-pertree analysis differed because PU-alonewas the only modication to produce thesame basal branching pattern as the GPWGcombined tree (Fig 5B) whereas BR-alonewith the pooids as sister group to the PAC-CAD clade corresponded to an alternativehypothesis for the grass family that is sup-ported by some data from nuclear (Adh Gautet al 1999) and plastid (rbcL Duvall andMorton 1996) genomes Because no refer-ence phylogenetic trees are available thatcontain a number of taxa similar to those ofour large supertrees it is difcult to assesstheir topologies and to determine whetherPU-alone with irreversible characters reallygave a more accurate phylogeny of largegrasses than did the other methods Theplacement of the pooids is possibly not thebest criterion by which to judge the methods
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 147
the evolutionary position of this subfamilywithin the grass family differs as inferredwith different DNA sequence data sets
Weighting by node support as suggestedby Ronquist (1996) improved the t betweenthe additive binary matrices and the GPWGcombined tree The benecial impact of thebootstrap weighting scheme is evident fromthe increased P-values obtained with themodied KH-test (Table 2) This trend is vis-ible for the BaumRagan coding schemebut is less obvious with the Purvis codingscheme (Fig 2) Bootstrap weighting wasproposed (Ronquist 1996) as an alternativeto the Purvis coding scheme both to reducethe greater impact of large source trees oversmalleronesand to improve the effect of well-supported nodes in the MRP analysis It isnot clear how bootstrap weighting could re-duce the impact of larger trees if the smallertrees do not have much greater support val-ues than those of the larger trees Howeverweighting of charactersby bootstrapsupportwithin the Purvis coding scheme appears tobe a redundant procedure unable to greatlyimprove the MRP reconstruction from theimportant information present in bootstrapsupport values (Fig 2) In the large supertreeanalysis PU C bootstrap with unorderedcharacters produced unrealistic topologieswith the early diverging lineages embed-ded in the bambusoids (Fig 5B) The Purviscoding schemes removes important restric-tive information from the matrix (Bininda-Emonds and Bryant 1998) possibly there-fore weighting by bootstrap support wouldrandomly assign high values to characters inthe matrix where this restrictive informationhas been removed and would prevent theweighting schemes from being as effectiveas with the BaumRagan coding schemesWeighting the matrix elements by node sup-port also poses problems Not all bootstrapanalyses can be considered identical and thenumber of replicates the type of searchesdone or both will inuence the supportfound Moreover when node support is notprovided for a source tree weights cannot beassigned and information from this tree willbe down-weighted in the MRP analysis Theeffect of this is great in supertree reconstruc-tions done with older publications which ingeneral do not report node support and arehence down-weighted in the MRP analyses
An important aspect affecting the large su-pertrees that is not resolved in our compar-
isons is the placement of some rarely sam-pled taxa Taxa that are present in only a fewpublications have been allocated a high pro-portion of missing character values whichmakes parsimony analyses much more dif-cult How many of these taxa are mis-placed is difcult to determine because noreference phylogeny for the 403 genera isavailable MRP reconstruction done withoutconstraining three clades (panicoids pooidsand chloridoids) to be monophyletic endedwith rarely sampled taxa being scattered allaround the supertrees (data not shown) Thatcan be an important problem when MRPanalyses are performed on a wide taxonomicgroup comprising large numbers of rarelysampled taxa
MRP coding especially when weighted bynode support can be considered an indica-tion of the signal in the primary data witheach node represented by one (weighted)synapomorphy (see Bininda-Emonds et al1999) However we stress that supertreescannot be viewed and interpreted in ex-actly the same way as phylogenetic analy-ses based on biological characters One ma-jor problem is the difculty of assigningnode support for supertrees Bootstrappingand other resampling procedures cannot beapplied given the clear nonindependenceof the characters present in the binary ma-trix and Bremer support (Bremer 1988) iscertainly equally affected by such noninde-pendence (as is the parsimony itself hencethe different MRP modications proposed)Moreoverbranch lengths associatedwith su-pertrees are treated in the same way as mor-phological branch lengths and unlike molec-ular branch length they cannot be used toestablish divergence times This weakness iseven more signicant in taxonomic groupssuch as the grasses for which dates are dif-cult to gather from other sources (eg fossilrecords)
CONCLUSIONS
We have demonstrated that supertreesoffer an easy way of producing phyloge-nies with a large number of taxa and thatthey can give good estimates of relation-ships within these groups (in this case thegrass family) Supertrees based on the eightcharacter partitions of the GPWG data setswere found to roughly match the combinedanalysis the discrepancies of the best MRP
148 SYSTEMATIC BIOLOGY VOL 51
modications being found mainly in weaklysupported branches of the combined treeThe BaumRagan and the Purvis modica-tions gave similar results but incorporatingbootstrap support associated with preexist-ing topologies improved the BaumRaganmodication Moreover supertrees can beuseful for comparative studies (Purvis1995b Bininda-Emonds et al 1999) whetherof adaptation coevolution rates of evo-lution cospeciation or rates of effectivecladogenesis where accuracy in the branchlength of phylogenetic reconstructions is notthe primary problem (Purvis et al 1994)Supertree reconstructions are also a usefulway to help highlight poor taxonomic sam-pling and identify where previous studiesareinconsistentThey can therefore be used as anexploratory tool capable of developing newhypotheses and indicating where future re-search should be focused
ACKNOWLEDGMENTS
We are grateful to Elizabeth Kellogg and LynnClark for providing the GPWG matrix and for their com-ments on the manuscript James Farris for helpful dis-cussion regarding bootstrap weighting schemes duringthe XIX Hennig Society meeting Stephen Renvoize andMark Chase for helpful discussion on the project OlafBininda-Emonds for his comments on the manuscriptand JonathanDavis for the testing of the SuperTree 085bprogram This study was supported by the Irish HigherEducation Authority the Roche Research Foundation(99-186) Enterprise Iceland (IC200129) the AgassizFoundation and the Swiss NSF (31-52378)
REFERENCES
(References marked curren were used as sources of phyloge-nies for the supertree reconstructions)
ARNQVIST G AND D WOOSTER 1995 Meta-analysisSynthesizing research ndings in ecology and evolu-tion Trends Ecol Evol 10236ndash240
currenBARKER N P 1997 The relationships of AmphipogonElytrophorus and Cyperochloa (Poaceae) as suggestedby rbcL sequence data Telopea 7205ndash213
currenBARKER N P H P LINDER AND E H HARLEY 1995Polyphyly of Arundinoids (Poaceae) Evidence fromrbcL sequence data Syst Bot 20423ndash435
currenBARKER N P H P LINDER and E H HARLEY 1999Sequences of the grass-specic insert in the chloro-plast rpoC2 gene elucidate generic relationships of theArundinoids (Poaceae) Syst Bot 23327ndash350
currenBAUM B R 1982 A phylogenetic analysis of the tribeTriticeae (Poaceae) based on morphological charactersof the genera Can J Bot 61518ndash535
BAUM B R 1992 Combining trees as a way of com-bining data sets for phylogenetic inference and thedesirability of combining gene trees Taxon 413ndash10
currenBAUM B R J R ESTES AND P K GUPTA 1986 Assess-ment of the genomic systematic classication in theTriticeae Am J Bot 741388ndash1395
BAUM B R AND M A RAGAN 1993 Reply to A GRodrigorsquos ldquoA comment on Baumrsquos method for com-bining phylogenetic treesrdquo Taxon 42637ndash640
BININD A-EMONDS O R P AND H N BRYANT 1998Properties of matrix representation with parsimonyanalyses Syst Biol 47497ndash508
BININD A-EMONDS O R P J L GITTLEMAN ANDA PURVIS 1999 Building large trees by combiningphylogenetic information A complete phylogenyof the extant Carnivora (Mammalia) Biol Rev 74143ndash175
BREMER K 1988 The limits of amino acid sequencedata in angiosperm phylogenetic reconstruction Evo-lution 42795ndash803
currenBUCKLER E S IV AND T P HOLTSFORD 1996 Zeasystematics Ribosomal ITS evidence Mol Biol Evol13612ndash622
CAMIN J H AND R R SOKAL 1965 A method for de-ducing branching sequences in phylogeny Evolution19311ndash326
currenCATALAN P E A KELLOGG AND R G OLMS TEAD1997 Phylogeny of Poaceae subfamily Pooideaebased on chloroplast ndhFgene sequences Mol Phy-logenet Evol 8150ndash166
currenCLARK L G W ZHANG AND J F WENDEL 1995 Aphylogenyof the grass family (Poaceae) basedon ndhFsequence data Syst Bot 20436ndash460
CONSTANTINES CU M AND D SANKOFF 1995 An ef-cient algorithm for supertrees J Classif 12101ndash112
currenCUMMINGS M P L M KING AND E A KELLOGG1994 Slipped-strand mispairing in a plastid generpoC2 in grasses (Poaceae) Mol Biol Evol 111ndash8
currenDARBYSHIRE S J AND S I WARWICK 1992 Phylogenyof North American Festuca (Poaceae) and related gen-era using chloroplast DNA restriction site variationCan J Bot 702415ndash2419
currenDAVIS J I AND R J SORENG 1993Phylogenetic struc-ture in the grass family (Poaceae) as inferred fromchloroplast DNA restriction site variation Am J Bot801444ndash1454
currenDOEBLEY J M DURBIN E M GOLENBERG M TCLEGG AND D P MA 1990 Evolutionary analysisof the large subunit of carboxylase (rbcL) nucleotidesequence among the grasses (Gramineae) Evolution441097ndash1108
currenDUVALL M R AND B R MORTON 1996 Molecularphylogenetics of Poaceae An expanded analysis ofrbcL sequence data Mol Phylogenet Evol 5352ndash358
currenDUVALL M R P M PETERSON E E TERRELL ANDA H CHRISTENS EN 1993 Phylogeny of NorthAmerican oryzoid grasses as constructed from mapsof plastid DNA restriction sites Am J Bot 8083ndash88
EFFRON B E HALLORAN AND S HOLMES 1996 Boot-strap condence levels for phylogenetic trees ProcNatl Acad Sci USA 937085ndash7090
currenESEN A AND K W HILU 1991 Electrophoretic andimmunological studies of prolamins in the PoaceaeII Phylogenetic afnities of the Atistideae Taxon 405ndash17
currenESEN A AND K W HILU 1993Prolamin and immuno-logical studies in the Poaceae IV SubfamilyPanicoidsCan J Bot 71315ndash322
FARRIS J S A G KLUGE AND M J ECKHARDT 1970 Anumerical approach to phylogenetic systematics SystZool 19172ndash191
FELSENSTEIN J 1985 Condence limits on phylogeniesAn approach using the bootstrap Evolution 39783ndash791
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 149
FITCH W M 1971 Toward dening the course of evolu-tion Minimal change for a specic tree topology SystZool 20406ndash416
currenFREDERIKSEN S AND O SEBERG 1991 Phylogeneticanalysis of the Triticeae (Poaceae) Hereditas 11515ndash19
currenGAUT B S A SPEEK BR MORTON AND M T CLEGG 1999Patterns of genetic diversication within the Adhgene family in the grasses (Poaceae) Mol Biol Evol161086ndash1097
GOLDMANN N J P ANDERSON AND A G RODRIGO2000 Likelihood-based tests of topologies in phyloge-netics Syst Biol 49652ndash670
GORDON A D 1980 On the assessment and compar-ison of classications Pages 149ndash160 in Analyse dedonnees et informatique (R Tomassone ed) INRALe Chesnay France
GPWG2000A phylogeny of the grass family (Poaceae)as inferred from eight character sets Pages 3ndash7 inGrasses Systematics and evolution (S W L Jacobsand J Everett eds) CSIRO Melbourne Australia
currenGREBENS TEIN B O GREBENS TEIN W SAUER ANDV HEMLEBEN 1996 Distribution and complex organi-zation of satellite DNA sequences in Aveneae speciesGenome 391045ndash1050
currenGREBENS TEIN B M ROSER W SAUER ANDV HEMLEBEN 1998 Molecular phylogenetic re-lationships in Aveneae (Poaceae) species and othergrasses as inferred from ITS1 and ITS2 rDNAsequences Plant Syst Evol 213233ndash250
currenHAMBY R K AND EA ZIMMER 1987Ribosomal RNAsequences for inferring phylogeny within the grassfamily (Poaceae) Plant Syst Evol 16029ndash37
HARVEY P H AND M D PAGEL 1991The comparativemethod in evolutionary biology Oxford Univ PressLondon
HILLIS D M 1996 Inferring complex phylogenies Na-ture 383130ndash131
HILLIS D M AND J J BULL 1993 An empirical test ofbootstrapping as a method for assessing condencein phylogenetic analysis Syst Biol 42182ndash192
currenHILU K W L A ALICE AND H LIANG 1999 Phy-logeny of Poaceae inferred from matK sequencesAnn Mo Bot Gard 86835ndash851
currenHILU K W AND A ESEN 1992Prolamin andimmuno-logical strudies in the Poaceae III Subfamily Chlori-doids Am J Bot 80104ndash113
currenHILU K W AND H LIANG 1997 The matK gene Se-quence variation and application in plant systematicsAm J Bot 84830ndash839
currenHILU K W AND K WRIGHT 1982 Systematics ofGramineae A cluster analysis study Taxon 319ndash37
currenHSIAO C N J CHATTERTON K H ASAY AND K BJENS EN 1993 Phylogenetic relationships of 10 grassspecies An assessment of phylogenetic utility of theinternal transcribed spacer region in nuclear riboso-mal DNA in monocots Genome 37112ndash120
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1994 Phylogenetic relationships of themonogenomic species of the wheat tribe Triticeae(Poaceae) inferred from nuclear rDNA (internal tran-scribed spacer) sequences Genome 38211ndash223
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1995Molecular phylogeny of the pooids(Poaceae) based on nuclear rDNA (ITS) sequencesTheor Appl Genet 90389ndash398
currenHSIAO C S W JACOBS N P BARKER AND N JCHATTERTON 1998 A molecular phylogeny of the
subfamily arundinoids (Poaceae) based on sequencesof rDNA Aust Syst Bot 1141ndash52
currenHSIAO C S W L JACOBS N J CHATTERTON AND KH ASAY 1999 A molecular phylogeny of the grassfamily (Poaceae) based on the sequences of nuclearribosomal DNA (ITS) Aust Syst Bot 11667ndash688
JOHNS ON L A AND D E SOLTIS 1998 Assessing con-gruence Empirical examples from molecular dataPages 297ndash348 in Molecular systematics of plants IIDNA sequencing (D E Soltis P S Soltis and J JDoyle eds) Kluwer Academic Publishers NorwellMassachusetts
currenKATAYAMA H AND Y OGIHARA 1996 Phylogeneticafnities of the grasses to other monocots as re-vealed by molecular analysis of chloroplast DNACurr Genet 29572ndash581
currenKELCHNER S A AND L G CLARK 1997 Molecu-lar evolution and phylogenetic utility of the chloro-plast rpl16 intron in Chusquea and the bambusoids(Poaceae) Mol Phylogenet Evol 8385ndash397
currenKELLOGG E A 1989 Comments on genomic genera inthe Triticeae (Poaceae) Am J Bot 76796ndash805
currenKELLOGG E A 1992Tools for studying the chloroplastgenome in the Triticeae (Gramineae) An ecoRI mapa diagnostic deletion and support for Bromus as anoutgroup Am J Bot 79186ndash197
currenKELLOGG E A 1998aRelationships of cereal crops andother grassesProc NatlAcad Sci USA952005ndash2010
currenKELLOGG E A 1998bWhorsquos related to whom Recentresults from molecular systematic studies Curr OpinPlant Biol 1149ndash158
currenKELLOGG E A AND R APPELS 1995 Intraspecic andinterspecic variation in 5S RNA genes are decoupledin diploid wheat relatives Genetics 140325ndash343
currenKELLOGG E A AND L WATSON 1993 Phyloge-netic studies of a large data set I bambusoids An-dropogonodae and pooids (Gramineae) Bot Rev59273ndash311
KIM J 1996 General inconsistency conditions for max-imum parsimony Effects of branch lengths and in-creasing numbers of taxa Syst Biol 45363ndash374
KISHINO H AND M HASEGAWA 1989 Evaluation ofthe maximum likelihood estimate of the evolution-ary tree topologies form DNA sequence data and thebranching order in Hominoidea J Mol Evol 29170ndash179
currenLIANG H AND K W HILU 1995 Application of thematK gene sequence to grass systematics Can J Bot74125ndash134
MABBERLEY D J 1993 The plant-book A portable dic-tionary of the vascular plants 2nd edition CambridgeUniv Press Cambridge
currenMACFARLANE T D AND L WATSON 1982The classi-cation of Poaceaesubfamily pooids Taxon31178ndash203
currenMASON-GAMER R J AND EA KELLOGG 1996Testingfor phylogenetic conict among molecular data sets inthe tribe Triticeae (Gramineae) Syst Biol 45524ndash545
currenMASON-GAMER R J C F WEIL AND E A KELLOGG1998Granule-bound starch synthase Structure func-tion andphylogenetic utility Mol Biol Evol 151658ndash1673
currenMATHEWS S AND R A SHARROCK 1996 The phy-tochrome gene family in grasses (Poaceae) A phy-logeny and evidence that grasses have a subset ofthe loci found in dicot angiosperms Mol Biol Evol131141ndash1150
currenMATHEWS S R C TSAL AND E A KELLOGG 2000Phylogenetic structure in the grass family (Poaceae)
150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel
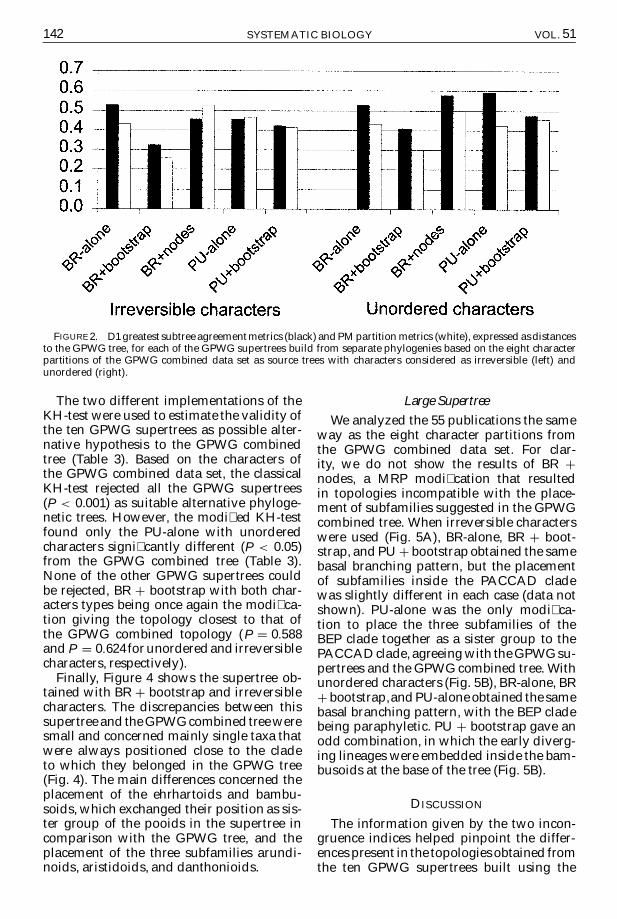
142 SYSTEMATIC BIOLOGY VOL 51
FIGURE 2 D1 greatest subtree agreementmetrics (black) and PM partition metrics (white) expressed asdistancesto the GPWG tree for each of the GPWG supertrees build from separate phylogenies based on the eight characterpartitions of the GPWG combined data set as source trees with characters considered as irreversible (left) andunordered (right)
The two different implementations of theKH-test were used to estimate the validity ofthe ten GPWG supertrees as possible alter-native hypothesis to the GPWG combinedtree (Table 3) Based on the characters ofthe GPWG combined data set the classicalKH-test rejected all the GPWG supertrees(P lt 0001) as suitable alternative phyloge-netic trees However the modied KH-testfound only the PU-alone with unorderedcharacters signicantly different (P lt 005)from the GPWG combined tree (Table 3)None of the other GPWG supertrees couldbe rejected BR C bootstrap with both char-acters types being once again the modica-tion giving the topology closest to that ofthe GPWG combined topology (P D 0588and P D 0624 for unordered and irreversiblecharacters respectively)
Finally Figure 4 shows the supertree ob-tained with BR C bootstrap and irreversiblecharacters The discrepancies between thissupertree and the GPWG combined tree weresmall and concerned mainly single taxa thatwere always positioned close to the cladeto which they belonged in the GPWG tree(Fig 4) The main differences concerned theplacement of the ehrhartoids and bambu-soids which exchanged their position as sis-ter group of the pooids in the supertree incomparison with the GPWG tree and theplacement of the three subfamilies arundi-noids aristidoids and danthonioids
Large SupertreeWe analyzed the 55 publications the same
way as the eight character partitions fromthe GPWG combined data set For clar-ity we do not show the results of BR Cnodes a MRP modication that resultedin topologies incompatible with the place-ment of subfamilies suggested in the GPWGcombined tree When irreversible characterswere used (Fig 5A) BR-alone BR C boot-strap and PU C bootstrap obtained the samebasal branching pattern but the placementof subfamilies inside the PACCAD cladewas slightly different in each case (data notshown) PU-alone was the only modica-tion to place the three subfamilies of theBEP clade together as a sister group to thePACCAD clade agreeing with the GPWG su-pertrees and the GPWG combined tree Withunordered characters (Fig 5B) BR-alone BRC bootstrapand PU-alone obtained the samebasal branching pattern with the BEP cladebeing paraphyletic PU C bootstrap gave anodd combination in which the early diverg-ing lineages were embedded inside the bam-busoids at the base of the tree (Fig 5B)
DISCUSSION
The information given by the two incon-gruence indices helped pinpoint the differ-ences present in the topologies obtained fromthe ten GPWG supertrees built using the
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 143
FIGURE 3 Summary of the subfamilial relationships in the GPWG supertrees obtained with the ve MRP mod-ications with column A) characters considered as irreversible and column B) characters considered as unordered
eight character partitions from the combineddata set Analyses using unordered charac-ters produced supertrees that were less sim-ilar as measured by D1 to the combinedtree than those obtained in analyses usingirreversible characters The only exceptionis for BR-alone which obtained the sametopology with both character types How-ever unordered charactersproduced slightlybetter topologies than irreversible charactersaccording to the PM incongruence index D1is dened as the number of taxa needed tobe removed from both trees to obtain an
identical subtree whereas PM is the numberof taxa bipartitions found between the twotrees being compared (Johnson and Soltis1998) Having higher D1 values than PMvalues suggests that the topological differ-ences do not involve single taxa (in whichcase D1 would be smaller than PM) butrather a large set of taxa placed differentlyin the supertree and the GPWG combinedtree Similar values of D1 and PM wouldsuggest that a few discrepant taxa wereinvolved none being far from their properlocations The D1 distances were much
144 SYSTEMATIC BIOLOGY VOL 51
TABLE 3 Kishino-Hasegawa test based on the difference in length between the GPWG combined tree and the tenGPWG supertrees obtained after applying the different MRP modications The p-values were calculated using thedefault parameter of PAUPcurren408b (Swofford 2000 classic Kishino-Hasegawa [KH] test) and Goldman et al (2000)modication to ensure the validity of the test with a posteriori specied topologies (modied KH test) Asterisk (curren)indicates a signicant p-value (p lt 005)
p-value
Number of steps Classic KH test Modied KH test
Combined GPWG 9054 ndash ndash
Unordered characters BR-alone 9291 lt00001curren 0172BR C bootstrap 9116 lt00001curren 0588BR C nodes 9203 lt00001curren 0396PU-alone 9411 lt00001curren 0042curren
PU C bootstrap 9211 lt00001curren 0374Irreversible characters BR-alone 9291 lt00001curren 0172
BR C bootstrap 9105 00003curren 0624BR C nodes 9281 lt00001curren 0198PU-alone 9311 lt00001curren 0144PU C bootstrap 9191 lt00001curren 0416
greater with unordered than irreversiblecharacters and were much closer to thePM ones obtained with irreversible char-acters Following this logic we can em-phasize that using unordered characters re-sulted in topologies with more differencesfrom the GPWG combined tree as to theplacement of larger subclades than did theuse of irreversible characters The type ofcharacters (ie unordered or irreversible)used to reconstruct the supertrees did notgreatly inuence the placement of the ter-minal taxa in the GPWG supertrees There-fore topologies obtained by consideringmatrix elements as irreversible characterswere closer to the combined phylogenetictree represented by the GPWG combinedanalysis This effect is clearly visible in theplacement of the major subclades dened inthe GPWG combined tree (Fig 4) Unorderedcharacters produced topologies in which theearly diverging lineages were split into twogroupsmdashone including Puelia and Guadu-ella and one including Anomochloa Strep-tochaeta and Pharus This extremely odd andunrealistic pattern does not correspond toany published phylogenetic trees concerningthe grass phylogeny Bininda-Emonds andBryant (1998) found only a minor impactof irreversible characters compared to un-ordered characters in their analysis but wefound that topologies created by using irre-versible characters are closer to an approachusing combined data than are the topolo-gies created by using unordered charactersThe resolution in the large supertrees was ex-tremely dependent on the type of characters
used to perform the analysis such was notthe case with the GPWG supertrees whereboth types of characters gave similar reso-lution (data not shown) Using irreversiblecharacters produced only one or two largesupertrees depending on the modicationused in contrast the ldquomaxtreesrdquo option inPAUPcurren408b was always reached when un-ordered characters were used Of course us-ing irreversible characters should put moreconstraintson the parsimonyanalysis by pre-venting the reversion from state 1 to 0 (thusreducing the number of most-parsimonioustrees) but it is not clear why such a drasticdifference appeared only between the largesupertrees
The Purvis coding scheme has been pro-posed as an improvement to the BaumRagan procedure as an attempt to reducethe dependency and redundancy betweenelements coming from the same topology(Purvis 1995a Ronquist 1996 Bininda-Emonds and Bryant 1998) However our re-sults suggest that the Purvis coding schemedoes not have a large impact on the MRPreconstructions and that the BaumRaganmethod works just as well or even betterComparisons between the approach usingcombined data and BR-alone or combineddata and PU-alone gave similar values forboth PM and D1 (Fig 2) Comparisons be-tween MRP modications also indicated aclose relationship between the two modi-cations especially with irreversible charac-ters (Table 2) This conclusion is less wellsupported with unordered characters be-cause BR-alone is the only modication to
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 145
PACCADclade
BEPclade
early-diverginglineages
outgroup
NassellaStipaPiptatherumAmpelodesmosMelicaGlyceriaBromusTriticumAvenaBrachypodiumDiarrhenaAnisopogonLygeumNardusBrachyelytrumPhaenospermaOryzaLeersiaEhrhartaStreptogynaLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaPennisetumPanicumMiscanthusZeaGyneriumDanthoniopsisZeugitesChasmanthiumThysanolaenaAmphipogonArundoMoliniaPhragmitesSpartinaSporobolusZoysiaDistichlisEragrostisPappophorumUniolaDanthoniaMicrairaAristidaStipagrostisEriachneGuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
pooids
bambusoids
ehrhartoids
chloridoids +danthonioids
panicoids
arundinoids
aristidoids
centothecoids
FIGURE 4 Grass supertree obtained with Baum and Ragan coding scheme with bootstrap support weightingusing separate phylogenies built from the eight character partitions of the GPWG combined data set as source treesDashed lines indicate incongruent branches between the supertree and the GPWG combined tree
146 SYSTEMATIC BIOLOGY VOL 51
FIGURE 5 Summary of the subfamilial relationships in the large supertrees based on 55 published source trees(including 403 genera) obtained using the ve MRP modications with column A) characters considered as irre-versible and column B) characters considered as unordered
give a basal topology very different fromthat obtained with any other modication(Table 2 and Fig 2) Ronquistrsquos (1996) propo-sition to weight each character in the bi-nary matrix by the inverse of the number ofnodes in the corresponding source tree doesnot perform as well as the other modica-tions It yields the greatest partition metricdistance and the second largest agreementsubtree distance (Fig 2) Although the twoalternatives to the BaumRagan method arebased on logical and plain arguments (ienonindependence and redundancy of matrixelements and impact of larger trees) theireffects on MRP reconstruction are not obvi-ous and do not result in topologies closerto our combined reference Indeed PU-alonewith unordered characters was rejected as asuitable alternative hypothesis to the GPWGcombined tree with the modied KH-testand gave the second smallest P-value when
used with irreversible characters (Table 2)However conclusions from the large su-pertree analysis differed because PU-alonewas the only modication to produce thesame basal branching pattern as the GPWGcombined tree (Fig 5B) whereas BR-alonewith the pooids as sister group to the PAC-CAD clade corresponded to an alternativehypothesis for the grass family that is sup-ported by some data from nuclear (Adh Gautet al 1999) and plastid (rbcL Duvall andMorton 1996) genomes Because no refer-ence phylogenetic trees are available thatcontain a number of taxa similar to those ofour large supertrees it is difcult to assesstheir topologies and to determine whetherPU-alone with irreversible characters reallygave a more accurate phylogeny of largegrasses than did the other methods Theplacement of the pooids is possibly not thebest criterion by which to judge the methods
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 147
the evolutionary position of this subfamilywithin the grass family differs as inferredwith different DNA sequence data sets
Weighting by node support as suggestedby Ronquist (1996) improved the t betweenthe additive binary matrices and the GPWGcombined tree The benecial impact of thebootstrap weighting scheme is evident fromthe increased P-values obtained with themodied KH-test (Table 2) This trend is vis-ible for the BaumRagan coding schemebut is less obvious with the Purvis codingscheme (Fig 2) Bootstrap weighting wasproposed (Ronquist 1996) as an alternativeto the Purvis coding scheme both to reducethe greater impact of large source trees oversmalleronesand to improve the effect of well-supported nodes in the MRP analysis It isnot clear how bootstrap weighting could re-duce the impact of larger trees if the smallertrees do not have much greater support val-ues than those of the larger trees Howeverweighting of charactersby bootstrapsupportwithin the Purvis coding scheme appears tobe a redundant procedure unable to greatlyimprove the MRP reconstruction from theimportant information present in bootstrapsupport values (Fig 2) In the large supertreeanalysis PU C bootstrap with unorderedcharacters produced unrealistic topologieswith the early diverging lineages embed-ded in the bambusoids (Fig 5B) The Purviscoding schemes removes important restric-tive information from the matrix (Bininda-Emonds and Bryant 1998) possibly there-fore weighting by bootstrap support wouldrandomly assign high values to characters inthe matrix where this restrictive informationhas been removed and would prevent theweighting schemes from being as effectiveas with the BaumRagan coding schemesWeighting the matrix elements by node sup-port also poses problems Not all bootstrapanalyses can be considered identical and thenumber of replicates the type of searchesdone or both will inuence the supportfound Moreover when node support is notprovided for a source tree weights cannot beassigned and information from this tree willbe down-weighted in the MRP analysis Theeffect of this is great in supertree reconstruc-tions done with older publications which ingeneral do not report node support and arehence down-weighted in the MRP analyses
An important aspect affecting the large su-pertrees that is not resolved in our compar-
isons is the placement of some rarely sam-pled taxa Taxa that are present in only a fewpublications have been allocated a high pro-portion of missing character values whichmakes parsimony analyses much more dif-cult How many of these taxa are mis-placed is difcult to determine because noreference phylogeny for the 403 genera isavailable MRP reconstruction done withoutconstraining three clades (panicoids pooidsand chloridoids) to be monophyletic endedwith rarely sampled taxa being scattered allaround the supertrees (data not shown) Thatcan be an important problem when MRPanalyses are performed on a wide taxonomicgroup comprising large numbers of rarelysampled taxa
MRP coding especially when weighted bynode support can be considered an indica-tion of the signal in the primary data witheach node represented by one (weighted)synapomorphy (see Bininda-Emonds et al1999) However we stress that supertreescannot be viewed and interpreted in ex-actly the same way as phylogenetic analy-ses based on biological characters One ma-jor problem is the difculty of assigningnode support for supertrees Bootstrappingand other resampling procedures cannot beapplied given the clear nonindependenceof the characters present in the binary ma-trix and Bremer support (Bremer 1988) iscertainly equally affected by such noninde-pendence (as is the parsimony itself hencethe different MRP modications proposed)Moreoverbranch lengths associatedwith su-pertrees are treated in the same way as mor-phological branch lengths and unlike molec-ular branch length they cannot be used toestablish divergence times This weakness iseven more signicant in taxonomic groupssuch as the grasses for which dates are dif-cult to gather from other sources (eg fossilrecords)
CONCLUSIONS
We have demonstrated that supertreesoffer an easy way of producing phyloge-nies with a large number of taxa and thatthey can give good estimates of relation-ships within these groups (in this case thegrass family) Supertrees based on the eightcharacter partitions of the GPWG data setswere found to roughly match the combinedanalysis the discrepancies of the best MRP
148 SYSTEMATIC BIOLOGY VOL 51
modications being found mainly in weaklysupported branches of the combined treeThe BaumRagan and the Purvis modica-tions gave similar results but incorporatingbootstrap support associated with preexist-ing topologies improved the BaumRaganmodication Moreover supertrees can beuseful for comparative studies (Purvis1995b Bininda-Emonds et al 1999) whetherof adaptation coevolution rates of evo-lution cospeciation or rates of effectivecladogenesis where accuracy in the branchlength of phylogenetic reconstructions is notthe primary problem (Purvis et al 1994)Supertree reconstructions are also a usefulway to help highlight poor taxonomic sam-pling and identify where previous studiesareinconsistentThey can therefore be used as anexploratory tool capable of developing newhypotheses and indicating where future re-search should be focused
ACKNOWLEDGMENTS
We are grateful to Elizabeth Kellogg and LynnClark for providing the GPWG matrix and for their com-ments on the manuscript James Farris for helpful dis-cussion regarding bootstrap weighting schemes duringthe XIX Hennig Society meeting Stephen Renvoize andMark Chase for helpful discussion on the project OlafBininda-Emonds for his comments on the manuscriptand JonathanDavis for the testing of the SuperTree 085bprogram This study was supported by the Irish HigherEducation Authority the Roche Research Foundation(99-186) Enterprise Iceland (IC200129) the AgassizFoundation and the Swiss NSF (31-52378)
REFERENCES
(References marked curren were used as sources of phyloge-nies for the supertree reconstructions)
ARNQVIST G AND D WOOSTER 1995 Meta-analysisSynthesizing research ndings in ecology and evolu-tion Trends Ecol Evol 10236ndash240
currenBARKER N P 1997 The relationships of AmphipogonElytrophorus and Cyperochloa (Poaceae) as suggestedby rbcL sequence data Telopea 7205ndash213
currenBARKER N P H P LINDER AND E H HARLEY 1995Polyphyly of Arundinoids (Poaceae) Evidence fromrbcL sequence data Syst Bot 20423ndash435
currenBARKER N P H P LINDER and E H HARLEY 1999Sequences of the grass-specic insert in the chloro-plast rpoC2 gene elucidate generic relationships of theArundinoids (Poaceae) Syst Bot 23327ndash350
currenBAUM B R 1982 A phylogenetic analysis of the tribeTriticeae (Poaceae) based on morphological charactersof the genera Can J Bot 61518ndash535
BAUM B R 1992 Combining trees as a way of com-bining data sets for phylogenetic inference and thedesirability of combining gene trees Taxon 413ndash10
currenBAUM B R J R ESTES AND P K GUPTA 1986 Assess-ment of the genomic systematic classication in theTriticeae Am J Bot 741388ndash1395
BAUM B R AND M A RAGAN 1993 Reply to A GRodrigorsquos ldquoA comment on Baumrsquos method for com-bining phylogenetic treesrdquo Taxon 42637ndash640
BININD A-EMONDS O R P AND H N BRYANT 1998Properties of matrix representation with parsimonyanalyses Syst Biol 47497ndash508
BININD A-EMONDS O R P J L GITTLEMAN ANDA PURVIS 1999 Building large trees by combiningphylogenetic information A complete phylogenyof the extant Carnivora (Mammalia) Biol Rev 74143ndash175
BREMER K 1988 The limits of amino acid sequencedata in angiosperm phylogenetic reconstruction Evo-lution 42795ndash803
currenBUCKLER E S IV AND T P HOLTSFORD 1996 Zeasystematics Ribosomal ITS evidence Mol Biol Evol13612ndash622
CAMIN J H AND R R SOKAL 1965 A method for de-ducing branching sequences in phylogeny Evolution19311ndash326
currenCATALAN P E A KELLOGG AND R G OLMS TEAD1997 Phylogeny of Poaceae subfamily Pooideaebased on chloroplast ndhFgene sequences Mol Phy-logenet Evol 8150ndash166
currenCLARK L G W ZHANG AND J F WENDEL 1995 Aphylogenyof the grass family (Poaceae) basedon ndhFsequence data Syst Bot 20436ndash460
CONSTANTINES CU M AND D SANKOFF 1995 An ef-cient algorithm for supertrees J Classif 12101ndash112
currenCUMMINGS M P L M KING AND E A KELLOGG1994 Slipped-strand mispairing in a plastid generpoC2 in grasses (Poaceae) Mol Biol Evol 111ndash8
currenDARBYSHIRE S J AND S I WARWICK 1992 Phylogenyof North American Festuca (Poaceae) and related gen-era using chloroplast DNA restriction site variationCan J Bot 702415ndash2419
currenDAVIS J I AND R J SORENG 1993Phylogenetic struc-ture in the grass family (Poaceae) as inferred fromchloroplast DNA restriction site variation Am J Bot801444ndash1454
currenDOEBLEY J M DURBIN E M GOLENBERG M TCLEGG AND D P MA 1990 Evolutionary analysisof the large subunit of carboxylase (rbcL) nucleotidesequence among the grasses (Gramineae) Evolution441097ndash1108
currenDUVALL M R AND B R MORTON 1996 Molecularphylogenetics of Poaceae An expanded analysis ofrbcL sequence data Mol Phylogenet Evol 5352ndash358
currenDUVALL M R P M PETERSON E E TERRELL ANDA H CHRISTENS EN 1993 Phylogeny of NorthAmerican oryzoid grasses as constructed from mapsof plastid DNA restriction sites Am J Bot 8083ndash88
EFFRON B E HALLORAN AND S HOLMES 1996 Boot-strap condence levels for phylogenetic trees ProcNatl Acad Sci USA 937085ndash7090
currenESEN A AND K W HILU 1991 Electrophoretic andimmunological studies of prolamins in the PoaceaeII Phylogenetic afnities of the Atistideae Taxon 405ndash17
currenESEN A AND K W HILU 1993Prolamin and immuno-logical studies in the Poaceae IV SubfamilyPanicoidsCan J Bot 71315ndash322
FARRIS J S A G KLUGE AND M J ECKHARDT 1970 Anumerical approach to phylogenetic systematics SystZool 19172ndash191
FELSENSTEIN J 1985 Condence limits on phylogeniesAn approach using the bootstrap Evolution 39783ndash791
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 149
FITCH W M 1971 Toward dening the course of evolu-tion Minimal change for a specic tree topology SystZool 20406ndash416
currenFREDERIKSEN S AND O SEBERG 1991 Phylogeneticanalysis of the Triticeae (Poaceae) Hereditas 11515ndash19
currenGAUT B S A SPEEK BR MORTON AND M T CLEGG 1999Patterns of genetic diversication within the Adhgene family in the grasses (Poaceae) Mol Biol Evol161086ndash1097
GOLDMANN N J P ANDERSON AND A G RODRIGO2000 Likelihood-based tests of topologies in phyloge-netics Syst Biol 49652ndash670
GORDON A D 1980 On the assessment and compar-ison of classications Pages 149ndash160 in Analyse dedonnees et informatique (R Tomassone ed) INRALe Chesnay France
GPWG2000A phylogeny of the grass family (Poaceae)as inferred from eight character sets Pages 3ndash7 inGrasses Systematics and evolution (S W L Jacobsand J Everett eds) CSIRO Melbourne Australia
currenGREBENS TEIN B O GREBENS TEIN W SAUER ANDV HEMLEBEN 1996 Distribution and complex organi-zation of satellite DNA sequences in Aveneae speciesGenome 391045ndash1050
currenGREBENS TEIN B M ROSER W SAUER ANDV HEMLEBEN 1998 Molecular phylogenetic re-lationships in Aveneae (Poaceae) species and othergrasses as inferred from ITS1 and ITS2 rDNAsequences Plant Syst Evol 213233ndash250
currenHAMBY R K AND EA ZIMMER 1987Ribosomal RNAsequences for inferring phylogeny within the grassfamily (Poaceae) Plant Syst Evol 16029ndash37
HARVEY P H AND M D PAGEL 1991The comparativemethod in evolutionary biology Oxford Univ PressLondon
HILLIS D M 1996 Inferring complex phylogenies Na-ture 383130ndash131
HILLIS D M AND J J BULL 1993 An empirical test ofbootstrapping as a method for assessing condencein phylogenetic analysis Syst Biol 42182ndash192
currenHILU K W L A ALICE AND H LIANG 1999 Phy-logeny of Poaceae inferred from matK sequencesAnn Mo Bot Gard 86835ndash851
currenHILU K W AND A ESEN 1992Prolamin andimmuno-logical strudies in the Poaceae III Subfamily Chlori-doids Am J Bot 80104ndash113
currenHILU K W AND H LIANG 1997 The matK gene Se-quence variation and application in plant systematicsAm J Bot 84830ndash839
currenHILU K W AND K WRIGHT 1982 Systematics ofGramineae A cluster analysis study Taxon 319ndash37
currenHSIAO C N J CHATTERTON K H ASAY AND K BJENS EN 1993 Phylogenetic relationships of 10 grassspecies An assessment of phylogenetic utility of theinternal transcribed spacer region in nuclear riboso-mal DNA in monocots Genome 37112ndash120
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1994 Phylogenetic relationships of themonogenomic species of the wheat tribe Triticeae(Poaceae) inferred from nuclear rDNA (internal tran-scribed spacer) sequences Genome 38211ndash223
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1995Molecular phylogeny of the pooids(Poaceae) based on nuclear rDNA (ITS) sequencesTheor Appl Genet 90389ndash398
currenHSIAO C S W JACOBS N P BARKER AND N JCHATTERTON 1998 A molecular phylogeny of the
subfamily arundinoids (Poaceae) based on sequencesof rDNA Aust Syst Bot 1141ndash52
currenHSIAO C S W L JACOBS N J CHATTERTON AND KH ASAY 1999 A molecular phylogeny of the grassfamily (Poaceae) based on the sequences of nuclearribosomal DNA (ITS) Aust Syst Bot 11667ndash688
JOHNS ON L A AND D E SOLTIS 1998 Assessing con-gruence Empirical examples from molecular dataPages 297ndash348 in Molecular systematics of plants IIDNA sequencing (D E Soltis P S Soltis and J JDoyle eds) Kluwer Academic Publishers NorwellMassachusetts
currenKATAYAMA H AND Y OGIHARA 1996 Phylogeneticafnities of the grasses to other monocots as re-vealed by molecular analysis of chloroplast DNACurr Genet 29572ndash581
currenKELCHNER S A AND L G CLARK 1997 Molecu-lar evolution and phylogenetic utility of the chloro-plast rpl16 intron in Chusquea and the bambusoids(Poaceae) Mol Phylogenet Evol 8385ndash397
currenKELLOGG E A 1989 Comments on genomic genera inthe Triticeae (Poaceae) Am J Bot 76796ndash805
currenKELLOGG E A 1992Tools for studying the chloroplastgenome in the Triticeae (Gramineae) An ecoRI mapa diagnostic deletion and support for Bromus as anoutgroup Am J Bot 79186ndash197
currenKELLOGG E A 1998aRelationships of cereal crops andother grassesProc NatlAcad Sci USA952005ndash2010
currenKELLOGG E A 1998bWhorsquos related to whom Recentresults from molecular systematic studies Curr OpinPlant Biol 1149ndash158
currenKELLOGG E A AND R APPELS 1995 Intraspecic andinterspecic variation in 5S RNA genes are decoupledin diploid wheat relatives Genetics 140325ndash343
currenKELLOGG E A AND L WATSON 1993 Phyloge-netic studies of a large data set I bambusoids An-dropogonodae and pooids (Gramineae) Bot Rev59273ndash311
KIM J 1996 General inconsistency conditions for max-imum parsimony Effects of branch lengths and in-creasing numbers of taxa Syst Biol 45363ndash374
KISHINO H AND M HASEGAWA 1989 Evaluation ofthe maximum likelihood estimate of the evolution-ary tree topologies form DNA sequence data and thebranching order in Hominoidea J Mol Evol 29170ndash179
currenLIANG H AND K W HILU 1995 Application of thematK gene sequence to grass systematics Can J Bot74125ndash134
MABBERLEY D J 1993 The plant-book A portable dic-tionary of the vascular plants 2nd edition CambridgeUniv Press Cambridge
currenMACFARLANE T D AND L WATSON 1982The classi-cation of Poaceaesubfamily pooids Taxon31178ndash203
currenMASON-GAMER R J AND EA KELLOGG 1996Testingfor phylogenetic conict among molecular data sets inthe tribe Triticeae (Gramineae) Syst Biol 45524ndash545
currenMASON-GAMER R J C F WEIL AND E A KELLOGG1998Granule-bound starch synthase Structure func-tion andphylogenetic utility Mol Biol Evol 151658ndash1673
currenMATHEWS S AND R A SHARROCK 1996 The phy-tochrome gene family in grasses (Poaceae) A phy-logeny and evidence that grasses have a subset ofthe loci found in dicot angiosperms Mol Biol Evol131141ndash1150
currenMATHEWS S R C TSAL AND E A KELLOGG 2000Phylogenetic structure in the grass family (Poaceae)
150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel

2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 143
FIGURE 3 Summary of the subfamilial relationships in the GPWG supertrees obtained with the ve MRP mod-ications with column A) characters considered as irreversible and column B) characters considered as unordered
eight character partitions from the combineddata set Analyses using unordered charac-ters produced supertrees that were less sim-ilar as measured by D1 to the combinedtree than those obtained in analyses usingirreversible characters The only exceptionis for BR-alone which obtained the sametopology with both character types How-ever unordered charactersproduced slightlybetter topologies than irreversible charactersaccording to the PM incongruence index D1is dened as the number of taxa needed tobe removed from both trees to obtain an
identical subtree whereas PM is the numberof taxa bipartitions found between the twotrees being compared (Johnson and Soltis1998) Having higher D1 values than PMvalues suggests that the topological differ-ences do not involve single taxa (in whichcase D1 would be smaller than PM) butrather a large set of taxa placed differentlyin the supertree and the GPWG combinedtree Similar values of D1 and PM wouldsuggest that a few discrepant taxa wereinvolved none being far from their properlocations The D1 distances were much
144 SYSTEMATIC BIOLOGY VOL 51
TABLE 3 Kishino-Hasegawa test based on the difference in length between the GPWG combined tree and the tenGPWG supertrees obtained after applying the different MRP modications The p-values were calculated using thedefault parameter of PAUPcurren408b (Swofford 2000 classic Kishino-Hasegawa [KH] test) and Goldman et al (2000)modication to ensure the validity of the test with a posteriori specied topologies (modied KH test) Asterisk (curren)indicates a signicant p-value (p lt 005)
p-value
Number of steps Classic KH test Modied KH test
Combined GPWG 9054 ndash ndash
Unordered characters BR-alone 9291 lt00001curren 0172BR C bootstrap 9116 lt00001curren 0588BR C nodes 9203 lt00001curren 0396PU-alone 9411 lt00001curren 0042curren
PU C bootstrap 9211 lt00001curren 0374Irreversible characters BR-alone 9291 lt00001curren 0172
BR C bootstrap 9105 00003curren 0624BR C nodes 9281 lt00001curren 0198PU-alone 9311 lt00001curren 0144PU C bootstrap 9191 lt00001curren 0416
greater with unordered than irreversiblecharacters and were much closer to thePM ones obtained with irreversible char-acters Following this logic we can em-phasize that using unordered characters re-sulted in topologies with more differencesfrom the GPWG combined tree as to theplacement of larger subclades than did theuse of irreversible characters The type ofcharacters (ie unordered or irreversible)used to reconstruct the supertrees did notgreatly inuence the placement of the ter-minal taxa in the GPWG supertrees There-fore topologies obtained by consideringmatrix elements as irreversible characterswere closer to the combined phylogenetictree represented by the GPWG combinedanalysis This effect is clearly visible in theplacement of the major subclades dened inthe GPWG combined tree (Fig 4) Unorderedcharacters produced topologies in which theearly diverging lineages were split into twogroupsmdashone including Puelia and Guadu-ella and one including Anomochloa Strep-tochaeta and Pharus This extremely odd andunrealistic pattern does not correspond toany published phylogenetic trees concerningthe grass phylogeny Bininda-Emonds andBryant (1998) found only a minor impactof irreversible characters compared to un-ordered characters in their analysis but wefound that topologies created by using irre-versible characters are closer to an approachusing combined data than are the topolo-gies created by using unordered charactersThe resolution in the large supertrees was ex-tremely dependent on the type of characters
used to perform the analysis such was notthe case with the GPWG supertrees whereboth types of characters gave similar reso-lution (data not shown) Using irreversiblecharacters produced only one or two largesupertrees depending on the modicationused in contrast the ldquomaxtreesrdquo option inPAUPcurren408b was always reached when un-ordered characters were used Of course us-ing irreversible characters should put moreconstraintson the parsimonyanalysis by pre-venting the reversion from state 1 to 0 (thusreducing the number of most-parsimonioustrees) but it is not clear why such a drasticdifference appeared only between the largesupertrees
The Purvis coding scheme has been pro-posed as an improvement to the BaumRagan procedure as an attempt to reducethe dependency and redundancy betweenelements coming from the same topology(Purvis 1995a Ronquist 1996 Bininda-Emonds and Bryant 1998) However our re-sults suggest that the Purvis coding schemedoes not have a large impact on the MRPreconstructions and that the BaumRaganmethod works just as well or even betterComparisons between the approach usingcombined data and BR-alone or combineddata and PU-alone gave similar values forboth PM and D1 (Fig 2) Comparisons be-tween MRP modications also indicated aclose relationship between the two modi-cations especially with irreversible charac-ters (Table 2) This conclusion is less wellsupported with unordered characters be-cause BR-alone is the only modication to
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 145
PACCADclade
BEPclade
early-diverginglineages
outgroup
NassellaStipaPiptatherumAmpelodesmosMelicaGlyceriaBromusTriticumAvenaBrachypodiumDiarrhenaAnisopogonLygeumNardusBrachyelytrumPhaenospermaOryzaLeersiaEhrhartaStreptogynaLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaPennisetumPanicumMiscanthusZeaGyneriumDanthoniopsisZeugitesChasmanthiumThysanolaenaAmphipogonArundoMoliniaPhragmitesSpartinaSporobolusZoysiaDistichlisEragrostisPappophorumUniolaDanthoniaMicrairaAristidaStipagrostisEriachneGuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
pooids
bambusoids
ehrhartoids
chloridoids +danthonioids
panicoids
arundinoids
aristidoids
centothecoids
FIGURE 4 Grass supertree obtained with Baum and Ragan coding scheme with bootstrap support weightingusing separate phylogenies built from the eight character partitions of the GPWG combined data set as source treesDashed lines indicate incongruent branches between the supertree and the GPWG combined tree
146 SYSTEMATIC BIOLOGY VOL 51
FIGURE 5 Summary of the subfamilial relationships in the large supertrees based on 55 published source trees(including 403 genera) obtained using the ve MRP modications with column A) characters considered as irre-versible and column B) characters considered as unordered
give a basal topology very different fromthat obtained with any other modication(Table 2 and Fig 2) Ronquistrsquos (1996) propo-sition to weight each character in the bi-nary matrix by the inverse of the number ofnodes in the corresponding source tree doesnot perform as well as the other modica-tions It yields the greatest partition metricdistance and the second largest agreementsubtree distance (Fig 2) Although the twoalternatives to the BaumRagan method arebased on logical and plain arguments (ienonindependence and redundancy of matrixelements and impact of larger trees) theireffects on MRP reconstruction are not obvi-ous and do not result in topologies closerto our combined reference Indeed PU-alonewith unordered characters was rejected as asuitable alternative hypothesis to the GPWGcombined tree with the modied KH-testand gave the second smallest P-value when
used with irreversible characters (Table 2)However conclusions from the large su-pertree analysis differed because PU-alonewas the only modication to produce thesame basal branching pattern as the GPWGcombined tree (Fig 5B) whereas BR-alonewith the pooids as sister group to the PAC-CAD clade corresponded to an alternativehypothesis for the grass family that is sup-ported by some data from nuclear (Adh Gautet al 1999) and plastid (rbcL Duvall andMorton 1996) genomes Because no refer-ence phylogenetic trees are available thatcontain a number of taxa similar to those ofour large supertrees it is difcult to assesstheir topologies and to determine whetherPU-alone with irreversible characters reallygave a more accurate phylogeny of largegrasses than did the other methods Theplacement of the pooids is possibly not thebest criterion by which to judge the methods
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 147
the evolutionary position of this subfamilywithin the grass family differs as inferredwith different DNA sequence data sets
Weighting by node support as suggestedby Ronquist (1996) improved the t betweenthe additive binary matrices and the GPWGcombined tree The benecial impact of thebootstrap weighting scheme is evident fromthe increased P-values obtained with themodied KH-test (Table 2) This trend is vis-ible for the BaumRagan coding schemebut is less obvious with the Purvis codingscheme (Fig 2) Bootstrap weighting wasproposed (Ronquist 1996) as an alternativeto the Purvis coding scheme both to reducethe greater impact of large source trees oversmalleronesand to improve the effect of well-supported nodes in the MRP analysis It isnot clear how bootstrap weighting could re-duce the impact of larger trees if the smallertrees do not have much greater support val-ues than those of the larger trees Howeverweighting of charactersby bootstrapsupportwithin the Purvis coding scheme appears tobe a redundant procedure unable to greatlyimprove the MRP reconstruction from theimportant information present in bootstrapsupport values (Fig 2) In the large supertreeanalysis PU C bootstrap with unorderedcharacters produced unrealistic topologieswith the early diverging lineages embed-ded in the bambusoids (Fig 5B) The Purviscoding schemes removes important restric-tive information from the matrix (Bininda-Emonds and Bryant 1998) possibly there-fore weighting by bootstrap support wouldrandomly assign high values to characters inthe matrix where this restrictive informationhas been removed and would prevent theweighting schemes from being as effectiveas with the BaumRagan coding schemesWeighting the matrix elements by node sup-port also poses problems Not all bootstrapanalyses can be considered identical and thenumber of replicates the type of searchesdone or both will inuence the supportfound Moreover when node support is notprovided for a source tree weights cannot beassigned and information from this tree willbe down-weighted in the MRP analysis Theeffect of this is great in supertree reconstruc-tions done with older publications which ingeneral do not report node support and arehence down-weighted in the MRP analyses
An important aspect affecting the large su-pertrees that is not resolved in our compar-
isons is the placement of some rarely sam-pled taxa Taxa that are present in only a fewpublications have been allocated a high pro-portion of missing character values whichmakes parsimony analyses much more dif-cult How many of these taxa are mis-placed is difcult to determine because noreference phylogeny for the 403 genera isavailable MRP reconstruction done withoutconstraining three clades (panicoids pooidsand chloridoids) to be monophyletic endedwith rarely sampled taxa being scattered allaround the supertrees (data not shown) Thatcan be an important problem when MRPanalyses are performed on a wide taxonomicgroup comprising large numbers of rarelysampled taxa
MRP coding especially when weighted bynode support can be considered an indica-tion of the signal in the primary data witheach node represented by one (weighted)synapomorphy (see Bininda-Emonds et al1999) However we stress that supertreescannot be viewed and interpreted in ex-actly the same way as phylogenetic analy-ses based on biological characters One ma-jor problem is the difculty of assigningnode support for supertrees Bootstrappingand other resampling procedures cannot beapplied given the clear nonindependenceof the characters present in the binary ma-trix and Bremer support (Bremer 1988) iscertainly equally affected by such noninde-pendence (as is the parsimony itself hencethe different MRP modications proposed)Moreoverbranch lengths associatedwith su-pertrees are treated in the same way as mor-phological branch lengths and unlike molec-ular branch length they cannot be used toestablish divergence times This weakness iseven more signicant in taxonomic groupssuch as the grasses for which dates are dif-cult to gather from other sources (eg fossilrecords)
CONCLUSIONS
We have demonstrated that supertreesoffer an easy way of producing phyloge-nies with a large number of taxa and thatthey can give good estimates of relation-ships within these groups (in this case thegrass family) Supertrees based on the eightcharacter partitions of the GPWG data setswere found to roughly match the combinedanalysis the discrepancies of the best MRP
148 SYSTEMATIC BIOLOGY VOL 51
modications being found mainly in weaklysupported branches of the combined treeThe BaumRagan and the Purvis modica-tions gave similar results but incorporatingbootstrap support associated with preexist-ing topologies improved the BaumRaganmodication Moreover supertrees can beuseful for comparative studies (Purvis1995b Bininda-Emonds et al 1999) whetherof adaptation coevolution rates of evo-lution cospeciation or rates of effectivecladogenesis where accuracy in the branchlength of phylogenetic reconstructions is notthe primary problem (Purvis et al 1994)Supertree reconstructions are also a usefulway to help highlight poor taxonomic sam-pling and identify where previous studiesareinconsistentThey can therefore be used as anexploratory tool capable of developing newhypotheses and indicating where future re-search should be focused
ACKNOWLEDGMENTS
We are grateful to Elizabeth Kellogg and LynnClark for providing the GPWG matrix and for their com-ments on the manuscript James Farris for helpful dis-cussion regarding bootstrap weighting schemes duringthe XIX Hennig Society meeting Stephen Renvoize andMark Chase for helpful discussion on the project OlafBininda-Emonds for his comments on the manuscriptand JonathanDavis for the testing of the SuperTree 085bprogram This study was supported by the Irish HigherEducation Authority the Roche Research Foundation(99-186) Enterprise Iceland (IC200129) the AgassizFoundation and the Swiss NSF (31-52378)
REFERENCES
(References marked curren were used as sources of phyloge-nies for the supertree reconstructions)
ARNQVIST G AND D WOOSTER 1995 Meta-analysisSynthesizing research ndings in ecology and evolu-tion Trends Ecol Evol 10236ndash240
currenBARKER N P 1997 The relationships of AmphipogonElytrophorus and Cyperochloa (Poaceae) as suggestedby rbcL sequence data Telopea 7205ndash213
currenBARKER N P H P LINDER AND E H HARLEY 1995Polyphyly of Arundinoids (Poaceae) Evidence fromrbcL sequence data Syst Bot 20423ndash435
currenBARKER N P H P LINDER and E H HARLEY 1999Sequences of the grass-specic insert in the chloro-plast rpoC2 gene elucidate generic relationships of theArundinoids (Poaceae) Syst Bot 23327ndash350
currenBAUM B R 1982 A phylogenetic analysis of the tribeTriticeae (Poaceae) based on morphological charactersof the genera Can J Bot 61518ndash535
BAUM B R 1992 Combining trees as a way of com-bining data sets for phylogenetic inference and thedesirability of combining gene trees Taxon 413ndash10
currenBAUM B R J R ESTES AND P K GUPTA 1986 Assess-ment of the genomic systematic classication in theTriticeae Am J Bot 741388ndash1395
BAUM B R AND M A RAGAN 1993 Reply to A GRodrigorsquos ldquoA comment on Baumrsquos method for com-bining phylogenetic treesrdquo Taxon 42637ndash640
BININD A-EMONDS O R P AND H N BRYANT 1998Properties of matrix representation with parsimonyanalyses Syst Biol 47497ndash508
BININD A-EMONDS O R P J L GITTLEMAN ANDA PURVIS 1999 Building large trees by combiningphylogenetic information A complete phylogenyof the extant Carnivora (Mammalia) Biol Rev 74143ndash175
BREMER K 1988 The limits of amino acid sequencedata in angiosperm phylogenetic reconstruction Evo-lution 42795ndash803
currenBUCKLER E S IV AND T P HOLTSFORD 1996 Zeasystematics Ribosomal ITS evidence Mol Biol Evol13612ndash622
CAMIN J H AND R R SOKAL 1965 A method for de-ducing branching sequences in phylogeny Evolution19311ndash326
currenCATALAN P E A KELLOGG AND R G OLMS TEAD1997 Phylogeny of Poaceae subfamily Pooideaebased on chloroplast ndhFgene sequences Mol Phy-logenet Evol 8150ndash166
currenCLARK L G W ZHANG AND J F WENDEL 1995 Aphylogenyof the grass family (Poaceae) basedon ndhFsequence data Syst Bot 20436ndash460
CONSTANTINES CU M AND D SANKOFF 1995 An ef-cient algorithm for supertrees J Classif 12101ndash112
currenCUMMINGS M P L M KING AND E A KELLOGG1994 Slipped-strand mispairing in a plastid generpoC2 in grasses (Poaceae) Mol Biol Evol 111ndash8
currenDARBYSHIRE S J AND S I WARWICK 1992 Phylogenyof North American Festuca (Poaceae) and related gen-era using chloroplast DNA restriction site variationCan J Bot 702415ndash2419
currenDAVIS J I AND R J SORENG 1993Phylogenetic struc-ture in the grass family (Poaceae) as inferred fromchloroplast DNA restriction site variation Am J Bot801444ndash1454
currenDOEBLEY J M DURBIN E M GOLENBERG M TCLEGG AND D P MA 1990 Evolutionary analysisof the large subunit of carboxylase (rbcL) nucleotidesequence among the grasses (Gramineae) Evolution441097ndash1108
currenDUVALL M R AND B R MORTON 1996 Molecularphylogenetics of Poaceae An expanded analysis ofrbcL sequence data Mol Phylogenet Evol 5352ndash358
currenDUVALL M R P M PETERSON E E TERRELL ANDA H CHRISTENS EN 1993 Phylogeny of NorthAmerican oryzoid grasses as constructed from mapsof plastid DNA restriction sites Am J Bot 8083ndash88
EFFRON B E HALLORAN AND S HOLMES 1996 Boot-strap condence levels for phylogenetic trees ProcNatl Acad Sci USA 937085ndash7090
currenESEN A AND K W HILU 1991 Electrophoretic andimmunological studies of prolamins in the PoaceaeII Phylogenetic afnities of the Atistideae Taxon 405ndash17
currenESEN A AND K W HILU 1993Prolamin and immuno-logical studies in the Poaceae IV SubfamilyPanicoidsCan J Bot 71315ndash322
FARRIS J S A G KLUGE AND M J ECKHARDT 1970 Anumerical approach to phylogenetic systematics SystZool 19172ndash191
FELSENSTEIN J 1985 Condence limits on phylogeniesAn approach using the bootstrap Evolution 39783ndash791
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 149
FITCH W M 1971 Toward dening the course of evolu-tion Minimal change for a specic tree topology SystZool 20406ndash416
currenFREDERIKSEN S AND O SEBERG 1991 Phylogeneticanalysis of the Triticeae (Poaceae) Hereditas 11515ndash19
currenGAUT B S A SPEEK BR MORTON AND M T CLEGG 1999Patterns of genetic diversication within the Adhgene family in the grasses (Poaceae) Mol Biol Evol161086ndash1097
GOLDMANN N J P ANDERSON AND A G RODRIGO2000 Likelihood-based tests of topologies in phyloge-netics Syst Biol 49652ndash670
GORDON A D 1980 On the assessment and compar-ison of classications Pages 149ndash160 in Analyse dedonnees et informatique (R Tomassone ed) INRALe Chesnay France
GPWG2000A phylogeny of the grass family (Poaceae)as inferred from eight character sets Pages 3ndash7 inGrasses Systematics and evolution (S W L Jacobsand J Everett eds) CSIRO Melbourne Australia
currenGREBENS TEIN B O GREBENS TEIN W SAUER ANDV HEMLEBEN 1996 Distribution and complex organi-zation of satellite DNA sequences in Aveneae speciesGenome 391045ndash1050
currenGREBENS TEIN B M ROSER W SAUER ANDV HEMLEBEN 1998 Molecular phylogenetic re-lationships in Aveneae (Poaceae) species and othergrasses as inferred from ITS1 and ITS2 rDNAsequences Plant Syst Evol 213233ndash250
currenHAMBY R K AND EA ZIMMER 1987Ribosomal RNAsequences for inferring phylogeny within the grassfamily (Poaceae) Plant Syst Evol 16029ndash37
HARVEY P H AND M D PAGEL 1991The comparativemethod in evolutionary biology Oxford Univ PressLondon
HILLIS D M 1996 Inferring complex phylogenies Na-ture 383130ndash131
HILLIS D M AND J J BULL 1993 An empirical test ofbootstrapping as a method for assessing condencein phylogenetic analysis Syst Biol 42182ndash192
currenHILU K W L A ALICE AND H LIANG 1999 Phy-logeny of Poaceae inferred from matK sequencesAnn Mo Bot Gard 86835ndash851
currenHILU K W AND A ESEN 1992Prolamin andimmuno-logical strudies in the Poaceae III Subfamily Chlori-doids Am J Bot 80104ndash113
currenHILU K W AND H LIANG 1997 The matK gene Se-quence variation and application in plant systematicsAm J Bot 84830ndash839
currenHILU K W AND K WRIGHT 1982 Systematics ofGramineae A cluster analysis study Taxon 319ndash37
currenHSIAO C N J CHATTERTON K H ASAY AND K BJENS EN 1993 Phylogenetic relationships of 10 grassspecies An assessment of phylogenetic utility of theinternal transcribed spacer region in nuclear riboso-mal DNA in monocots Genome 37112ndash120
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1994 Phylogenetic relationships of themonogenomic species of the wheat tribe Triticeae(Poaceae) inferred from nuclear rDNA (internal tran-scribed spacer) sequences Genome 38211ndash223
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1995Molecular phylogeny of the pooids(Poaceae) based on nuclear rDNA (ITS) sequencesTheor Appl Genet 90389ndash398
currenHSIAO C S W JACOBS N P BARKER AND N JCHATTERTON 1998 A molecular phylogeny of the
subfamily arundinoids (Poaceae) based on sequencesof rDNA Aust Syst Bot 1141ndash52
currenHSIAO C S W L JACOBS N J CHATTERTON AND KH ASAY 1999 A molecular phylogeny of the grassfamily (Poaceae) based on the sequences of nuclearribosomal DNA (ITS) Aust Syst Bot 11667ndash688
JOHNS ON L A AND D E SOLTIS 1998 Assessing con-gruence Empirical examples from molecular dataPages 297ndash348 in Molecular systematics of plants IIDNA sequencing (D E Soltis P S Soltis and J JDoyle eds) Kluwer Academic Publishers NorwellMassachusetts
currenKATAYAMA H AND Y OGIHARA 1996 Phylogeneticafnities of the grasses to other monocots as re-vealed by molecular analysis of chloroplast DNACurr Genet 29572ndash581
currenKELCHNER S A AND L G CLARK 1997 Molecu-lar evolution and phylogenetic utility of the chloro-plast rpl16 intron in Chusquea and the bambusoids(Poaceae) Mol Phylogenet Evol 8385ndash397
currenKELLOGG E A 1989 Comments on genomic genera inthe Triticeae (Poaceae) Am J Bot 76796ndash805
currenKELLOGG E A 1992Tools for studying the chloroplastgenome in the Triticeae (Gramineae) An ecoRI mapa diagnostic deletion and support for Bromus as anoutgroup Am J Bot 79186ndash197
currenKELLOGG E A 1998aRelationships of cereal crops andother grassesProc NatlAcad Sci USA952005ndash2010
currenKELLOGG E A 1998bWhorsquos related to whom Recentresults from molecular systematic studies Curr OpinPlant Biol 1149ndash158
currenKELLOGG E A AND R APPELS 1995 Intraspecic andinterspecic variation in 5S RNA genes are decoupledin diploid wheat relatives Genetics 140325ndash343
currenKELLOGG E A AND L WATSON 1993 Phyloge-netic studies of a large data set I bambusoids An-dropogonodae and pooids (Gramineae) Bot Rev59273ndash311
KIM J 1996 General inconsistency conditions for max-imum parsimony Effects of branch lengths and in-creasing numbers of taxa Syst Biol 45363ndash374
KISHINO H AND M HASEGAWA 1989 Evaluation ofthe maximum likelihood estimate of the evolution-ary tree topologies form DNA sequence data and thebranching order in Hominoidea J Mol Evol 29170ndash179
currenLIANG H AND K W HILU 1995 Application of thematK gene sequence to grass systematics Can J Bot74125ndash134
MABBERLEY D J 1993 The plant-book A portable dic-tionary of the vascular plants 2nd edition CambridgeUniv Press Cambridge
currenMACFARLANE T D AND L WATSON 1982The classi-cation of Poaceaesubfamily pooids Taxon31178ndash203
currenMASON-GAMER R J AND EA KELLOGG 1996Testingfor phylogenetic conict among molecular data sets inthe tribe Triticeae (Gramineae) Syst Biol 45524ndash545
currenMASON-GAMER R J C F WEIL AND E A KELLOGG1998Granule-bound starch synthase Structure func-tion andphylogenetic utility Mol Biol Evol 151658ndash1673
currenMATHEWS S AND R A SHARROCK 1996 The phy-tochrome gene family in grasses (Poaceae) A phy-logeny and evidence that grasses have a subset ofthe loci found in dicot angiosperms Mol Biol Evol131141ndash1150
currenMATHEWS S R C TSAL AND E A KELLOGG 2000Phylogenetic structure in the grass family (Poaceae)
150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel

144 SYSTEMATIC BIOLOGY VOL 51
TABLE 3 Kishino-Hasegawa test based on the difference in length between the GPWG combined tree and the tenGPWG supertrees obtained after applying the different MRP modications The p-values were calculated using thedefault parameter of PAUPcurren408b (Swofford 2000 classic Kishino-Hasegawa [KH] test) and Goldman et al (2000)modication to ensure the validity of the test with a posteriori specied topologies (modied KH test) Asterisk (curren)indicates a signicant p-value (p lt 005)
p-value
Number of steps Classic KH test Modied KH test
Combined GPWG 9054 ndash ndash
Unordered characters BR-alone 9291 lt00001curren 0172BR C bootstrap 9116 lt00001curren 0588BR C nodes 9203 lt00001curren 0396PU-alone 9411 lt00001curren 0042curren
PU C bootstrap 9211 lt00001curren 0374Irreversible characters BR-alone 9291 lt00001curren 0172
BR C bootstrap 9105 00003curren 0624BR C nodes 9281 lt00001curren 0198PU-alone 9311 lt00001curren 0144PU C bootstrap 9191 lt00001curren 0416
greater with unordered than irreversiblecharacters and were much closer to thePM ones obtained with irreversible char-acters Following this logic we can em-phasize that using unordered characters re-sulted in topologies with more differencesfrom the GPWG combined tree as to theplacement of larger subclades than did theuse of irreversible characters The type ofcharacters (ie unordered or irreversible)used to reconstruct the supertrees did notgreatly inuence the placement of the ter-minal taxa in the GPWG supertrees There-fore topologies obtained by consideringmatrix elements as irreversible characterswere closer to the combined phylogenetictree represented by the GPWG combinedanalysis This effect is clearly visible in theplacement of the major subclades dened inthe GPWG combined tree (Fig 4) Unorderedcharacters produced topologies in which theearly diverging lineages were split into twogroupsmdashone including Puelia and Guadu-ella and one including Anomochloa Strep-tochaeta and Pharus This extremely odd andunrealistic pattern does not correspond toany published phylogenetic trees concerningthe grass phylogeny Bininda-Emonds andBryant (1998) found only a minor impactof irreversible characters compared to un-ordered characters in their analysis but wefound that topologies created by using irre-versible characters are closer to an approachusing combined data than are the topolo-gies created by using unordered charactersThe resolution in the large supertrees was ex-tremely dependent on the type of characters
used to perform the analysis such was notthe case with the GPWG supertrees whereboth types of characters gave similar reso-lution (data not shown) Using irreversiblecharacters produced only one or two largesupertrees depending on the modicationused in contrast the ldquomaxtreesrdquo option inPAUPcurren408b was always reached when un-ordered characters were used Of course us-ing irreversible characters should put moreconstraintson the parsimonyanalysis by pre-venting the reversion from state 1 to 0 (thusreducing the number of most-parsimonioustrees) but it is not clear why such a drasticdifference appeared only between the largesupertrees
The Purvis coding scheme has been pro-posed as an improvement to the BaumRagan procedure as an attempt to reducethe dependency and redundancy betweenelements coming from the same topology(Purvis 1995a Ronquist 1996 Bininda-Emonds and Bryant 1998) However our re-sults suggest that the Purvis coding schemedoes not have a large impact on the MRPreconstructions and that the BaumRaganmethod works just as well or even betterComparisons between the approach usingcombined data and BR-alone or combineddata and PU-alone gave similar values forboth PM and D1 (Fig 2) Comparisons be-tween MRP modications also indicated aclose relationship between the two modi-cations especially with irreversible charac-ters (Table 2) This conclusion is less wellsupported with unordered characters be-cause BR-alone is the only modication to
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 145
PACCADclade
BEPclade
early-diverginglineages
outgroup
NassellaStipaPiptatherumAmpelodesmosMelicaGlyceriaBromusTriticumAvenaBrachypodiumDiarrhenaAnisopogonLygeumNardusBrachyelytrumPhaenospermaOryzaLeersiaEhrhartaStreptogynaLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaPennisetumPanicumMiscanthusZeaGyneriumDanthoniopsisZeugitesChasmanthiumThysanolaenaAmphipogonArundoMoliniaPhragmitesSpartinaSporobolusZoysiaDistichlisEragrostisPappophorumUniolaDanthoniaMicrairaAristidaStipagrostisEriachneGuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
pooids
bambusoids
ehrhartoids
chloridoids +danthonioids
panicoids
arundinoids
aristidoids
centothecoids
FIGURE 4 Grass supertree obtained with Baum and Ragan coding scheme with bootstrap support weightingusing separate phylogenies built from the eight character partitions of the GPWG combined data set as source treesDashed lines indicate incongruent branches between the supertree and the GPWG combined tree
146 SYSTEMATIC BIOLOGY VOL 51
FIGURE 5 Summary of the subfamilial relationships in the large supertrees based on 55 published source trees(including 403 genera) obtained using the ve MRP modications with column A) characters considered as irre-versible and column B) characters considered as unordered
give a basal topology very different fromthat obtained with any other modication(Table 2 and Fig 2) Ronquistrsquos (1996) propo-sition to weight each character in the bi-nary matrix by the inverse of the number ofnodes in the corresponding source tree doesnot perform as well as the other modica-tions It yields the greatest partition metricdistance and the second largest agreementsubtree distance (Fig 2) Although the twoalternatives to the BaumRagan method arebased on logical and plain arguments (ienonindependence and redundancy of matrixelements and impact of larger trees) theireffects on MRP reconstruction are not obvi-ous and do not result in topologies closerto our combined reference Indeed PU-alonewith unordered characters was rejected as asuitable alternative hypothesis to the GPWGcombined tree with the modied KH-testand gave the second smallest P-value when
used with irreversible characters (Table 2)However conclusions from the large su-pertree analysis differed because PU-alonewas the only modication to produce thesame basal branching pattern as the GPWGcombined tree (Fig 5B) whereas BR-alonewith the pooids as sister group to the PAC-CAD clade corresponded to an alternativehypothesis for the grass family that is sup-ported by some data from nuclear (Adh Gautet al 1999) and plastid (rbcL Duvall andMorton 1996) genomes Because no refer-ence phylogenetic trees are available thatcontain a number of taxa similar to those ofour large supertrees it is difcult to assesstheir topologies and to determine whetherPU-alone with irreversible characters reallygave a more accurate phylogeny of largegrasses than did the other methods Theplacement of the pooids is possibly not thebest criterion by which to judge the methods
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 147
the evolutionary position of this subfamilywithin the grass family differs as inferredwith different DNA sequence data sets
Weighting by node support as suggestedby Ronquist (1996) improved the t betweenthe additive binary matrices and the GPWGcombined tree The benecial impact of thebootstrap weighting scheme is evident fromthe increased P-values obtained with themodied KH-test (Table 2) This trend is vis-ible for the BaumRagan coding schemebut is less obvious with the Purvis codingscheme (Fig 2) Bootstrap weighting wasproposed (Ronquist 1996) as an alternativeto the Purvis coding scheme both to reducethe greater impact of large source trees oversmalleronesand to improve the effect of well-supported nodes in the MRP analysis It isnot clear how bootstrap weighting could re-duce the impact of larger trees if the smallertrees do not have much greater support val-ues than those of the larger trees Howeverweighting of charactersby bootstrapsupportwithin the Purvis coding scheme appears tobe a redundant procedure unable to greatlyimprove the MRP reconstruction from theimportant information present in bootstrapsupport values (Fig 2) In the large supertreeanalysis PU C bootstrap with unorderedcharacters produced unrealistic topologieswith the early diverging lineages embed-ded in the bambusoids (Fig 5B) The Purviscoding schemes removes important restric-tive information from the matrix (Bininda-Emonds and Bryant 1998) possibly there-fore weighting by bootstrap support wouldrandomly assign high values to characters inthe matrix where this restrictive informationhas been removed and would prevent theweighting schemes from being as effectiveas with the BaumRagan coding schemesWeighting the matrix elements by node sup-port also poses problems Not all bootstrapanalyses can be considered identical and thenumber of replicates the type of searchesdone or both will inuence the supportfound Moreover when node support is notprovided for a source tree weights cannot beassigned and information from this tree willbe down-weighted in the MRP analysis Theeffect of this is great in supertree reconstruc-tions done with older publications which ingeneral do not report node support and arehence down-weighted in the MRP analyses
An important aspect affecting the large su-pertrees that is not resolved in our compar-
isons is the placement of some rarely sam-pled taxa Taxa that are present in only a fewpublications have been allocated a high pro-portion of missing character values whichmakes parsimony analyses much more dif-cult How many of these taxa are mis-placed is difcult to determine because noreference phylogeny for the 403 genera isavailable MRP reconstruction done withoutconstraining three clades (panicoids pooidsand chloridoids) to be monophyletic endedwith rarely sampled taxa being scattered allaround the supertrees (data not shown) Thatcan be an important problem when MRPanalyses are performed on a wide taxonomicgroup comprising large numbers of rarelysampled taxa
MRP coding especially when weighted bynode support can be considered an indica-tion of the signal in the primary data witheach node represented by one (weighted)synapomorphy (see Bininda-Emonds et al1999) However we stress that supertreescannot be viewed and interpreted in ex-actly the same way as phylogenetic analy-ses based on biological characters One ma-jor problem is the difculty of assigningnode support for supertrees Bootstrappingand other resampling procedures cannot beapplied given the clear nonindependenceof the characters present in the binary ma-trix and Bremer support (Bremer 1988) iscertainly equally affected by such noninde-pendence (as is the parsimony itself hencethe different MRP modications proposed)Moreoverbranch lengths associatedwith su-pertrees are treated in the same way as mor-phological branch lengths and unlike molec-ular branch length they cannot be used toestablish divergence times This weakness iseven more signicant in taxonomic groupssuch as the grasses for which dates are dif-cult to gather from other sources (eg fossilrecords)
CONCLUSIONS
We have demonstrated that supertreesoffer an easy way of producing phyloge-nies with a large number of taxa and thatthey can give good estimates of relation-ships within these groups (in this case thegrass family) Supertrees based on the eightcharacter partitions of the GPWG data setswere found to roughly match the combinedanalysis the discrepancies of the best MRP
148 SYSTEMATIC BIOLOGY VOL 51
modications being found mainly in weaklysupported branches of the combined treeThe BaumRagan and the Purvis modica-tions gave similar results but incorporatingbootstrap support associated with preexist-ing topologies improved the BaumRaganmodication Moreover supertrees can beuseful for comparative studies (Purvis1995b Bininda-Emonds et al 1999) whetherof adaptation coevolution rates of evo-lution cospeciation or rates of effectivecladogenesis where accuracy in the branchlength of phylogenetic reconstructions is notthe primary problem (Purvis et al 1994)Supertree reconstructions are also a usefulway to help highlight poor taxonomic sam-pling and identify where previous studiesareinconsistentThey can therefore be used as anexploratory tool capable of developing newhypotheses and indicating where future re-search should be focused
ACKNOWLEDGMENTS
We are grateful to Elizabeth Kellogg and LynnClark for providing the GPWG matrix and for their com-ments on the manuscript James Farris for helpful dis-cussion regarding bootstrap weighting schemes duringthe XIX Hennig Society meeting Stephen Renvoize andMark Chase for helpful discussion on the project OlafBininda-Emonds for his comments on the manuscriptand JonathanDavis for the testing of the SuperTree 085bprogram This study was supported by the Irish HigherEducation Authority the Roche Research Foundation(99-186) Enterprise Iceland (IC200129) the AgassizFoundation and the Swiss NSF (31-52378)
REFERENCES
(References marked curren were used as sources of phyloge-nies for the supertree reconstructions)
ARNQVIST G AND D WOOSTER 1995 Meta-analysisSynthesizing research ndings in ecology and evolu-tion Trends Ecol Evol 10236ndash240
currenBARKER N P 1997 The relationships of AmphipogonElytrophorus and Cyperochloa (Poaceae) as suggestedby rbcL sequence data Telopea 7205ndash213
currenBARKER N P H P LINDER AND E H HARLEY 1995Polyphyly of Arundinoids (Poaceae) Evidence fromrbcL sequence data Syst Bot 20423ndash435
currenBARKER N P H P LINDER and E H HARLEY 1999Sequences of the grass-specic insert in the chloro-plast rpoC2 gene elucidate generic relationships of theArundinoids (Poaceae) Syst Bot 23327ndash350
currenBAUM B R 1982 A phylogenetic analysis of the tribeTriticeae (Poaceae) based on morphological charactersof the genera Can J Bot 61518ndash535
BAUM B R 1992 Combining trees as a way of com-bining data sets for phylogenetic inference and thedesirability of combining gene trees Taxon 413ndash10
currenBAUM B R J R ESTES AND P K GUPTA 1986 Assess-ment of the genomic systematic classication in theTriticeae Am J Bot 741388ndash1395
BAUM B R AND M A RAGAN 1993 Reply to A GRodrigorsquos ldquoA comment on Baumrsquos method for com-bining phylogenetic treesrdquo Taxon 42637ndash640
BININD A-EMONDS O R P AND H N BRYANT 1998Properties of matrix representation with parsimonyanalyses Syst Biol 47497ndash508
BININD A-EMONDS O R P J L GITTLEMAN ANDA PURVIS 1999 Building large trees by combiningphylogenetic information A complete phylogenyof the extant Carnivora (Mammalia) Biol Rev 74143ndash175
BREMER K 1988 The limits of amino acid sequencedata in angiosperm phylogenetic reconstruction Evo-lution 42795ndash803
currenBUCKLER E S IV AND T P HOLTSFORD 1996 Zeasystematics Ribosomal ITS evidence Mol Biol Evol13612ndash622
CAMIN J H AND R R SOKAL 1965 A method for de-ducing branching sequences in phylogeny Evolution19311ndash326
currenCATALAN P E A KELLOGG AND R G OLMS TEAD1997 Phylogeny of Poaceae subfamily Pooideaebased on chloroplast ndhFgene sequences Mol Phy-logenet Evol 8150ndash166
currenCLARK L G W ZHANG AND J F WENDEL 1995 Aphylogenyof the grass family (Poaceae) basedon ndhFsequence data Syst Bot 20436ndash460
CONSTANTINES CU M AND D SANKOFF 1995 An ef-cient algorithm for supertrees J Classif 12101ndash112
currenCUMMINGS M P L M KING AND E A KELLOGG1994 Slipped-strand mispairing in a plastid generpoC2 in grasses (Poaceae) Mol Biol Evol 111ndash8
currenDARBYSHIRE S J AND S I WARWICK 1992 Phylogenyof North American Festuca (Poaceae) and related gen-era using chloroplast DNA restriction site variationCan J Bot 702415ndash2419
currenDAVIS J I AND R J SORENG 1993Phylogenetic struc-ture in the grass family (Poaceae) as inferred fromchloroplast DNA restriction site variation Am J Bot801444ndash1454
currenDOEBLEY J M DURBIN E M GOLENBERG M TCLEGG AND D P MA 1990 Evolutionary analysisof the large subunit of carboxylase (rbcL) nucleotidesequence among the grasses (Gramineae) Evolution441097ndash1108
currenDUVALL M R AND B R MORTON 1996 Molecularphylogenetics of Poaceae An expanded analysis ofrbcL sequence data Mol Phylogenet Evol 5352ndash358
currenDUVALL M R P M PETERSON E E TERRELL ANDA H CHRISTENS EN 1993 Phylogeny of NorthAmerican oryzoid grasses as constructed from mapsof plastid DNA restriction sites Am J Bot 8083ndash88
EFFRON B E HALLORAN AND S HOLMES 1996 Boot-strap condence levels for phylogenetic trees ProcNatl Acad Sci USA 937085ndash7090
currenESEN A AND K W HILU 1991 Electrophoretic andimmunological studies of prolamins in the PoaceaeII Phylogenetic afnities of the Atistideae Taxon 405ndash17
currenESEN A AND K W HILU 1993Prolamin and immuno-logical studies in the Poaceae IV SubfamilyPanicoidsCan J Bot 71315ndash322
FARRIS J S A G KLUGE AND M J ECKHARDT 1970 Anumerical approach to phylogenetic systematics SystZool 19172ndash191
FELSENSTEIN J 1985 Condence limits on phylogeniesAn approach using the bootstrap Evolution 39783ndash791
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 149
FITCH W M 1971 Toward dening the course of evolu-tion Minimal change for a specic tree topology SystZool 20406ndash416
currenFREDERIKSEN S AND O SEBERG 1991 Phylogeneticanalysis of the Triticeae (Poaceae) Hereditas 11515ndash19
currenGAUT B S A SPEEK BR MORTON AND M T CLEGG 1999Patterns of genetic diversication within the Adhgene family in the grasses (Poaceae) Mol Biol Evol161086ndash1097
GOLDMANN N J P ANDERSON AND A G RODRIGO2000 Likelihood-based tests of topologies in phyloge-netics Syst Biol 49652ndash670
GORDON A D 1980 On the assessment and compar-ison of classications Pages 149ndash160 in Analyse dedonnees et informatique (R Tomassone ed) INRALe Chesnay France
GPWG2000A phylogeny of the grass family (Poaceae)as inferred from eight character sets Pages 3ndash7 inGrasses Systematics and evolution (S W L Jacobsand J Everett eds) CSIRO Melbourne Australia
currenGREBENS TEIN B O GREBENS TEIN W SAUER ANDV HEMLEBEN 1996 Distribution and complex organi-zation of satellite DNA sequences in Aveneae speciesGenome 391045ndash1050
currenGREBENS TEIN B M ROSER W SAUER ANDV HEMLEBEN 1998 Molecular phylogenetic re-lationships in Aveneae (Poaceae) species and othergrasses as inferred from ITS1 and ITS2 rDNAsequences Plant Syst Evol 213233ndash250
currenHAMBY R K AND EA ZIMMER 1987Ribosomal RNAsequences for inferring phylogeny within the grassfamily (Poaceae) Plant Syst Evol 16029ndash37
HARVEY P H AND M D PAGEL 1991The comparativemethod in evolutionary biology Oxford Univ PressLondon
HILLIS D M 1996 Inferring complex phylogenies Na-ture 383130ndash131
HILLIS D M AND J J BULL 1993 An empirical test ofbootstrapping as a method for assessing condencein phylogenetic analysis Syst Biol 42182ndash192
currenHILU K W L A ALICE AND H LIANG 1999 Phy-logeny of Poaceae inferred from matK sequencesAnn Mo Bot Gard 86835ndash851
currenHILU K W AND A ESEN 1992Prolamin andimmuno-logical strudies in the Poaceae III Subfamily Chlori-doids Am J Bot 80104ndash113
currenHILU K W AND H LIANG 1997 The matK gene Se-quence variation and application in plant systematicsAm J Bot 84830ndash839
currenHILU K W AND K WRIGHT 1982 Systematics ofGramineae A cluster analysis study Taxon 319ndash37
currenHSIAO C N J CHATTERTON K H ASAY AND K BJENS EN 1993 Phylogenetic relationships of 10 grassspecies An assessment of phylogenetic utility of theinternal transcribed spacer region in nuclear riboso-mal DNA in monocots Genome 37112ndash120
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1994 Phylogenetic relationships of themonogenomic species of the wheat tribe Triticeae(Poaceae) inferred from nuclear rDNA (internal tran-scribed spacer) sequences Genome 38211ndash223
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1995Molecular phylogeny of the pooids(Poaceae) based on nuclear rDNA (ITS) sequencesTheor Appl Genet 90389ndash398
currenHSIAO C S W JACOBS N P BARKER AND N JCHATTERTON 1998 A molecular phylogeny of the
subfamily arundinoids (Poaceae) based on sequencesof rDNA Aust Syst Bot 1141ndash52
currenHSIAO C S W L JACOBS N J CHATTERTON AND KH ASAY 1999 A molecular phylogeny of the grassfamily (Poaceae) based on the sequences of nuclearribosomal DNA (ITS) Aust Syst Bot 11667ndash688
JOHNS ON L A AND D E SOLTIS 1998 Assessing con-gruence Empirical examples from molecular dataPages 297ndash348 in Molecular systematics of plants IIDNA sequencing (D E Soltis P S Soltis and J JDoyle eds) Kluwer Academic Publishers NorwellMassachusetts
currenKATAYAMA H AND Y OGIHARA 1996 Phylogeneticafnities of the grasses to other monocots as re-vealed by molecular analysis of chloroplast DNACurr Genet 29572ndash581
currenKELCHNER S A AND L G CLARK 1997 Molecu-lar evolution and phylogenetic utility of the chloro-plast rpl16 intron in Chusquea and the bambusoids(Poaceae) Mol Phylogenet Evol 8385ndash397
currenKELLOGG E A 1989 Comments on genomic genera inthe Triticeae (Poaceae) Am J Bot 76796ndash805
currenKELLOGG E A 1992Tools for studying the chloroplastgenome in the Triticeae (Gramineae) An ecoRI mapa diagnostic deletion and support for Bromus as anoutgroup Am J Bot 79186ndash197
currenKELLOGG E A 1998aRelationships of cereal crops andother grassesProc NatlAcad Sci USA952005ndash2010
currenKELLOGG E A 1998bWhorsquos related to whom Recentresults from molecular systematic studies Curr OpinPlant Biol 1149ndash158
currenKELLOGG E A AND R APPELS 1995 Intraspecic andinterspecic variation in 5S RNA genes are decoupledin diploid wheat relatives Genetics 140325ndash343
currenKELLOGG E A AND L WATSON 1993 Phyloge-netic studies of a large data set I bambusoids An-dropogonodae and pooids (Gramineae) Bot Rev59273ndash311
KIM J 1996 General inconsistency conditions for max-imum parsimony Effects of branch lengths and in-creasing numbers of taxa Syst Biol 45363ndash374
KISHINO H AND M HASEGAWA 1989 Evaluation ofthe maximum likelihood estimate of the evolution-ary tree topologies form DNA sequence data and thebranching order in Hominoidea J Mol Evol 29170ndash179
currenLIANG H AND K W HILU 1995 Application of thematK gene sequence to grass systematics Can J Bot74125ndash134
MABBERLEY D J 1993 The plant-book A portable dic-tionary of the vascular plants 2nd edition CambridgeUniv Press Cambridge
currenMACFARLANE T D AND L WATSON 1982The classi-cation of Poaceaesubfamily pooids Taxon31178ndash203
currenMASON-GAMER R J AND EA KELLOGG 1996Testingfor phylogenetic conict among molecular data sets inthe tribe Triticeae (Gramineae) Syst Biol 45524ndash545
currenMASON-GAMER R J C F WEIL AND E A KELLOGG1998Granule-bound starch synthase Structure func-tion andphylogenetic utility Mol Biol Evol 151658ndash1673
currenMATHEWS S AND R A SHARROCK 1996 The phy-tochrome gene family in grasses (Poaceae) A phy-logeny and evidence that grasses have a subset ofthe loci found in dicot angiosperms Mol Biol Evol131141ndash1150
currenMATHEWS S R C TSAL AND E A KELLOGG 2000Phylogenetic structure in the grass family (Poaceae)
150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel
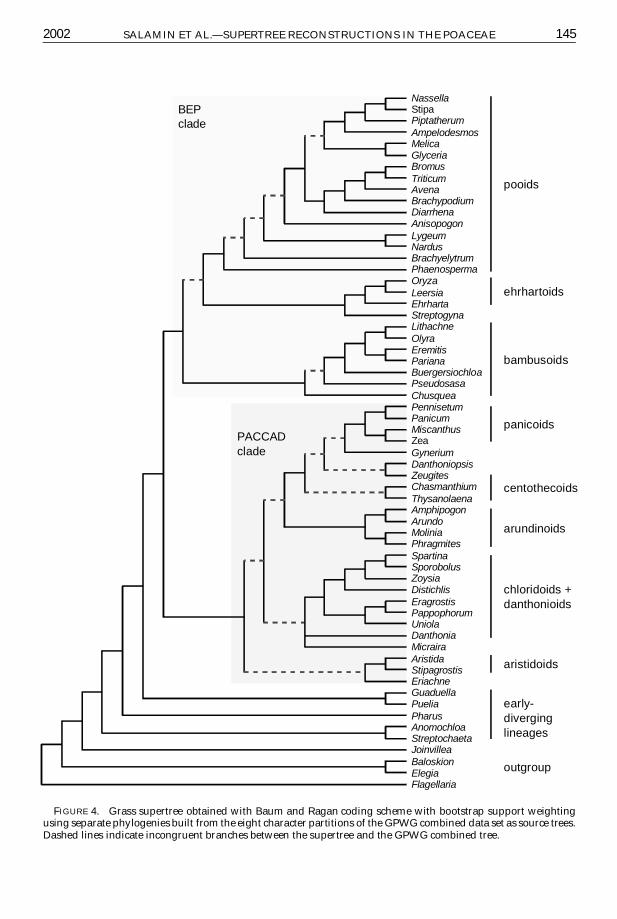
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 145
PACCADclade
BEPclade
early-diverginglineages
outgroup
NassellaStipaPiptatherumAmpelodesmosMelicaGlyceriaBromusTriticumAvenaBrachypodiumDiarrhenaAnisopogonLygeumNardusBrachyelytrumPhaenospermaOryzaLeersiaEhrhartaStreptogynaLithachneOlyraEremitisParianaBuergersiochloaPseudosasaChusqueaPennisetumPanicumMiscanthusZeaGyneriumDanthoniopsisZeugitesChasmanthiumThysanolaenaAmphipogonArundoMoliniaPhragmitesSpartinaSporobolusZoysiaDistichlisEragrostisPappophorumUniolaDanthoniaMicrairaAristidaStipagrostisEriachneGuaduellaPueliaPharusAnomochloaStreptochaetaJoinvilleaBaloskionElegiaFlagellaria
pooids
bambusoids
ehrhartoids
chloridoids +danthonioids
panicoids
arundinoids
aristidoids
centothecoids
FIGURE 4 Grass supertree obtained with Baum and Ragan coding scheme with bootstrap support weightingusing separate phylogenies built from the eight character partitions of the GPWG combined data set as source treesDashed lines indicate incongruent branches between the supertree and the GPWG combined tree
146 SYSTEMATIC BIOLOGY VOL 51
FIGURE 5 Summary of the subfamilial relationships in the large supertrees based on 55 published source trees(including 403 genera) obtained using the ve MRP modications with column A) characters considered as irre-versible and column B) characters considered as unordered
give a basal topology very different fromthat obtained with any other modication(Table 2 and Fig 2) Ronquistrsquos (1996) propo-sition to weight each character in the bi-nary matrix by the inverse of the number ofnodes in the corresponding source tree doesnot perform as well as the other modica-tions It yields the greatest partition metricdistance and the second largest agreementsubtree distance (Fig 2) Although the twoalternatives to the BaumRagan method arebased on logical and plain arguments (ienonindependence and redundancy of matrixelements and impact of larger trees) theireffects on MRP reconstruction are not obvi-ous and do not result in topologies closerto our combined reference Indeed PU-alonewith unordered characters was rejected as asuitable alternative hypothesis to the GPWGcombined tree with the modied KH-testand gave the second smallest P-value when
used with irreversible characters (Table 2)However conclusions from the large su-pertree analysis differed because PU-alonewas the only modication to produce thesame basal branching pattern as the GPWGcombined tree (Fig 5B) whereas BR-alonewith the pooids as sister group to the PAC-CAD clade corresponded to an alternativehypothesis for the grass family that is sup-ported by some data from nuclear (Adh Gautet al 1999) and plastid (rbcL Duvall andMorton 1996) genomes Because no refer-ence phylogenetic trees are available thatcontain a number of taxa similar to those ofour large supertrees it is difcult to assesstheir topologies and to determine whetherPU-alone with irreversible characters reallygave a more accurate phylogeny of largegrasses than did the other methods Theplacement of the pooids is possibly not thebest criterion by which to judge the methods
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 147
the evolutionary position of this subfamilywithin the grass family differs as inferredwith different DNA sequence data sets
Weighting by node support as suggestedby Ronquist (1996) improved the t betweenthe additive binary matrices and the GPWGcombined tree The benecial impact of thebootstrap weighting scheme is evident fromthe increased P-values obtained with themodied KH-test (Table 2) This trend is vis-ible for the BaumRagan coding schemebut is less obvious with the Purvis codingscheme (Fig 2) Bootstrap weighting wasproposed (Ronquist 1996) as an alternativeto the Purvis coding scheme both to reducethe greater impact of large source trees oversmalleronesand to improve the effect of well-supported nodes in the MRP analysis It isnot clear how bootstrap weighting could re-duce the impact of larger trees if the smallertrees do not have much greater support val-ues than those of the larger trees Howeverweighting of charactersby bootstrapsupportwithin the Purvis coding scheme appears tobe a redundant procedure unable to greatlyimprove the MRP reconstruction from theimportant information present in bootstrapsupport values (Fig 2) In the large supertreeanalysis PU C bootstrap with unorderedcharacters produced unrealistic topologieswith the early diverging lineages embed-ded in the bambusoids (Fig 5B) The Purviscoding schemes removes important restric-tive information from the matrix (Bininda-Emonds and Bryant 1998) possibly there-fore weighting by bootstrap support wouldrandomly assign high values to characters inthe matrix where this restrictive informationhas been removed and would prevent theweighting schemes from being as effectiveas with the BaumRagan coding schemesWeighting the matrix elements by node sup-port also poses problems Not all bootstrapanalyses can be considered identical and thenumber of replicates the type of searchesdone or both will inuence the supportfound Moreover when node support is notprovided for a source tree weights cannot beassigned and information from this tree willbe down-weighted in the MRP analysis Theeffect of this is great in supertree reconstruc-tions done with older publications which ingeneral do not report node support and arehence down-weighted in the MRP analyses
An important aspect affecting the large su-pertrees that is not resolved in our compar-
isons is the placement of some rarely sam-pled taxa Taxa that are present in only a fewpublications have been allocated a high pro-portion of missing character values whichmakes parsimony analyses much more dif-cult How many of these taxa are mis-placed is difcult to determine because noreference phylogeny for the 403 genera isavailable MRP reconstruction done withoutconstraining three clades (panicoids pooidsand chloridoids) to be monophyletic endedwith rarely sampled taxa being scattered allaround the supertrees (data not shown) Thatcan be an important problem when MRPanalyses are performed on a wide taxonomicgroup comprising large numbers of rarelysampled taxa
MRP coding especially when weighted bynode support can be considered an indica-tion of the signal in the primary data witheach node represented by one (weighted)synapomorphy (see Bininda-Emonds et al1999) However we stress that supertreescannot be viewed and interpreted in ex-actly the same way as phylogenetic analy-ses based on biological characters One ma-jor problem is the difculty of assigningnode support for supertrees Bootstrappingand other resampling procedures cannot beapplied given the clear nonindependenceof the characters present in the binary ma-trix and Bremer support (Bremer 1988) iscertainly equally affected by such noninde-pendence (as is the parsimony itself hencethe different MRP modications proposed)Moreoverbranch lengths associatedwith su-pertrees are treated in the same way as mor-phological branch lengths and unlike molec-ular branch length they cannot be used toestablish divergence times This weakness iseven more signicant in taxonomic groupssuch as the grasses for which dates are dif-cult to gather from other sources (eg fossilrecords)
CONCLUSIONS
We have demonstrated that supertreesoffer an easy way of producing phyloge-nies with a large number of taxa and thatthey can give good estimates of relation-ships within these groups (in this case thegrass family) Supertrees based on the eightcharacter partitions of the GPWG data setswere found to roughly match the combinedanalysis the discrepancies of the best MRP
148 SYSTEMATIC BIOLOGY VOL 51
modications being found mainly in weaklysupported branches of the combined treeThe BaumRagan and the Purvis modica-tions gave similar results but incorporatingbootstrap support associated with preexist-ing topologies improved the BaumRaganmodication Moreover supertrees can beuseful for comparative studies (Purvis1995b Bininda-Emonds et al 1999) whetherof adaptation coevolution rates of evo-lution cospeciation or rates of effectivecladogenesis where accuracy in the branchlength of phylogenetic reconstructions is notthe primary problem (Purvis et al 1994)Supertree reconstructions are also a usefulway to help highlight poor taxonomic sam-pling and identify where previous studiesareinconsistentThey can therefore be used as anexploratory tool capable of developing newhypotheses and indicating where future re-search should be focused
ACKNOWLEDGMENTS
We are grateful to Elizabeth Kellogg and LynnClark for providing the GPWG matrix and for their com-ments on the manuscript James Farris for helpful dis-cussion regarding bootstrap weighting schemes duringthe XIX Hennig Society meeting Stephen Renvoize andMark Chase for helpful discussion on the project OlafBininda-Emonds for his comments on the manuscriptand JonathanDavis for the testing of the SuperTree 085bprogram This study was supported by the Irish HigherEducation Authority the Roche Research Foundation(99-186) Enterprise Iceland (IC200129) the AgassizFoundation and the Swiss NSF (31-52378)
REFERENCES
(References marked curren were used as sources of phyloge-nies for the supertree reconstructions)
ARNQVIST G AND D WOOSTER 1995 Meta-analysisSynthesizing research ndings in ecology and evolu-tion Trends Ecol Evol 10236ndash240
currenBARKER N P 1997 The relationships of AmphipogonElytrophorus and Cyperochloa (Poaceae) as suggestedby rbcL sequence data Telopea 7205ndash213
currenBARKER N P H P LINDER AND E H HARLEY 1995Polyphyly of Arundinoids (Poaceae) Evidence fromrbcL sequence data Syst Bot 20423ndash435
currenBARKER N P H P LINDER and E H HARLEY 1999Sequences of the grass-specic insert in the chloro-plast rpoC2 gene elucidate generic relationships of theArundinoids (Poaceae) Syst Bot 23327ndash350
currenBAUM B R 1982 A phylogenetic analysis of the tribeTriticeae (Poaceae) based on morphological charactersof the genera Can J Bot 61518ndash535
BAUM B R 1992 Combining trees as a way of com-bining data sets for phylogenetic inference and thedesirability of combining gene trees Taxon 413ndash10
currenBAUM B R J R ESTES AND P K GUPTA 1986 Assess-ment of the genomic systematic classication in theTriticeae Am J Bot 741388ndash1395
BAUM B R AND M A RAGAN 1993 Reply to A GRodrigorsquos ldquoA comment on Baumrsquos method for com-bining phylogenetic treesrdquo Taxon 42637ndash640
BININD A-EMONDS O R P AND H N BRYANT 1998Properties of matrix representation with parsimonyanalyses Syst Biol 47497ndash508
BININD A-EMONDS O R P J L GITTLEMAN ANDA PURVIS 1999 Building large trees by combiningphylogenetic information A complete phylogenyof the extant Carnivora (Mammalia) Biol Rev 74143ndash175
BREMER K 1988 The limits of amino acid sequencedata in angiosperm phylogenetic reconstruction Evo-lution 42795ndash803
currenBUCKLER E S IV AND T P HOLTSFORD 1996 Zeasystematics Ribosomal ITS evidence Mol Biol Evol13612ndash622
CAMIN J H AND R R SOKAL 1965 A method for de-ducing branching sequences in phylogeny Evolution19311ndash326
currenCATALAN P E A KELLOGG AND R G OLMS TEAD1997 Phylogeny of Poaceae subfamily Pooideaebased on chloroplast ndhFgene sequences Mol Phy-logenet Evol 8150ndash166
currenCLARK L G W ZHANG AND J F WENDEL 1995 Aphylogenyof the grass family (Poaceae) basedon ndhFsequence data Syst Bot 20436ndash460
CONSTANTINES CU M AND D SANKOFF 1995 An ef-cient algorithm for supertrees J Classif 12101ndash112
currenCUMMINGS M P L M KING AND E A KELLOGG1994 Slipped-strand mispairing in a plastid generpoC2 in grasses (Poaceae) Mol Biol Evol 111ndash8
currenDARBYSHIRE S J AND S I WARWICK 1992 Phylogenyof North American Festuca (Poaceae) and related gen-era using chloroplast DNA restriction site variationCan J Bot 702415ndash2419
currenDAVIS J I AND R J SORENG 1993Phylogenetic struc-ture in the grass family (Poaceae) as inferred fromchloroplast DNA restriction site variation Am J Bot801444ndash1454
currenDOEBLEY J M DURBIN E M GOLENBERG M TCLEGG AND D P MA 1990 Evolutionary analysisof the large subunit of carboxylase (rbcL) nucleotidesequence among the grasses (Gramineae) Evolution441097ndash1108
currenDUVALL M R AND B R MORTON 1996 Molecularphylogenetics of Poaceae An expanded analysis ofrbcL sequence data Mol Phylogenet Evol 5352ndash358
currenDUVALL M R P M PETERSON E E TERRELL ANDA H CHRISTENS EN 1993 Phylogeny of NorthAmerican oryzoid grasses as constructed from mapsof plastid DNA restriction sites Am J Bot 8083ndash88
EFFRON B E HALLORAN AND S HOLMES 1996 Boot-strap condence levels for phylogenetic trees ProcNatl Acad Sci USA 937085ndash7090
currenESEN A AND K W HILU 1991 Electrophoretic andimmunological studies of prolamins in the PoaceaeII Phylogenetic afnities of the Atistideae Taxon 405ndash17
currenESEN A AND K W HILU 1993Prolamin and immuno-logical studies in the Poaceae IV SubfamilyPanicoidsCan J Bot 71315ndash322
FARRIS J S A G KLUGE AND M J ECKHARDT 1970 Anumerical approach to phylogenetic systematics SystZool 19172ndash191
FELSENSTEIN J 1985 Condence limits on phylogeniesAn approach using the bootstrap Evolution 39783ndash791
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 149
FITCH W M 1971 Toward dening the course of evolu-tion Minimal change for a specic tree topology SystZool 20406ndash416
currenFREDERIKSEN S AND O SEBERG 1991 Phylogeneticanalysis of the Triticeae (Poaceae) Hereditas 11515ndash19
currenGAUT B S A SPEEK BR MORTON AND M T CLEGG 1999Patterns of genetic diversication within the Adhgene family in the grasses (Poaceae) Mol Biol Evol161086ndash1097
GOLDMANN N J P ANDERSON AND A G RODRIGO2000 Likelihood-based tests of topologies in phyloge-netics Syst Biol 49652ndash670
GORDON A D 1980 On the assessment and compar-ison of classications Pages 149ndash160 in Analyse dedonnees et informatique (R Tomassone ed) INRALe Chesnay France
GPWG2000A phylogeny of the grass family (Poaceae)as inferred from eight character sets Pages 3ndash7 inGrasses Systematics and evolution (S W L Jacobsand J Everett eds) CSIRO Melbourne Australia
currenGREBENS TEIN B O GREBENS TEIN W SAUER ANDV HEMLEBEN 1996 Distribution and complex organi-zation of satellite DNA sequences in Aveneae speciesGenome 391045ndash1050
currenGREBENS TEIN B M ROSER W SAUER ANDV HEMLEBEN 1998 Molecular phylogenetic re-lationships in Aveneae (Poaceae) species and othergrasses as inferred from ITS1 and ITS2 rDNAsequences Plant Syst Evol 213233ndash250
currenHAMBY R K AND EA ZIMMER 1987Ribosomal RNAsequences for inferring phylogeny within the grassfamily (Poaceae) Plant Syst Evol 16029ndash37
HARVEY P H AND M D PAGEL 1991The comparativemethod in evolutionary biology Oxford Univ PressLondon
HILLIS D M 1996 Inferring complex phylogenies Na-ture 383130ndash131
HILLIS D M AND J J BULL 1993 An empirical test ofbootstrapping as a method for assessing condencein phylogenetic analysis Syst Biol 42182ndash192
currenHILU K W L A ALICE AND H LIANG 1999 Phy-logeny of Poaceae inferred from matK sequencesAnn Mo Bot Gard 86835ndash851
currenHILU K W AND A ESEN 1992Prolamin andimmuno-logical strudies in the Poaceae III Subfamily Chlori-doids Am J Bot 80104ndash113
currenHILU K W AND H LIANG 1997 The matK gene Se-quence variation and application in plant systematicsAm J Bot 84830ndash839
currenHILU K W AND K WRIGHT 1982 Systematics ofGramineae A cluster analysis study Taxon 319ndash37
currenHSIAO C N J CHATTERTON K H ASAY AND K BJENS EN 1993 Phylogenetic relationships of 10 grassspecies An assessment of phylogenetic utility of theinternal transcribed spacer region in nuclear riboso-mal DNA in monocots Genome 37112ndash120
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1994 Phylogenetic relationships of themonogenomic species of the wheat tribe Triticeae(Poaceae) inferred from nuclear rDNA (internal tran-scribed spacer) sequences Genome 38211ndash223
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1995Molecular phylogeny of the pooids(Poaceae) based on nuclear rDNA (ITS) sequencesTheor Appl Genet 90389ndash398
currenHSIAO C S W JACOBS N P BARKER AND N JCHATTERTON 1998 A molecular phylogeny of the
subfamily arundinoids (Poaceae) based on sequencesof rDNA Aust Syst Bot 1141ndash52
currenHSIAO C S W L JACOBS N J CHATTERTON AND KH ASAY 1999 A molecular phylogeny of the grassfamily (Poaceae) based on the sequences of nuclearribosomal DNA (ITS) Aust Syst Bot 11667ndash688
JOHNS ON L A AND D E SOLTIS 1998 Assessing con-gruence Empirical examples from molecular dataPages 297ndash348 in Molecular systematics of plants IIDNA sequencing (D E Soltis P S Soltis and J JDoyle eds) Kluwer Academic Publishers NorwellMassachusetts
currenKATAYAMA H AND Y OGIHARA 1996 Phylogeneticafnities of the grasses to other monocots as re-vealed by molecular analysis of chloroplast DNACurr Genet 29572ndash581
currenKELCHNER S A AND L G CLARK 1997 Molecu-lar evolution and phylogenetic utility of the chloro-plast rpl16 intron in Chusquea and the bambusoids(Poaceae) Mol Phylogenet Evol 8385ndash397
currenKELLOGG E A 1989 Comments on genomic genera inthe Triticeae (Poaceae) Am J Bot 76796ndash805
currenKELLOGG E A 1992Tools for studying the chloroplastgenome in the Triticeae (Gramineae) An ecoRI mapa diagnostic deletion and support for Bromus as anoutgroup Am J Bot 79186ndash197
currenKELLOGG E A 1998aRelationships of cereal crops andother grassesProc NatlAcad Sci USA952005ndash2010
currenKELLOGG E A 1998bWhorsquos related to whom Recentresults from molecular systematic studies Curr OpinPlant Biol 1149ndash158
currenKELLOGG E A AND R APPELS 1995 Intraspecic andinterspecic variation in 5S RNA genes are decoupledin diploid wheat relatives Genetics 140325ndash343
currenKELLOGG E A AND L WATSON 1993 Phyloge-netic studies of a large data set I bambusoids An-dropogonodae and pooids (Gramineae) Bot Rev59273ndash311
KIM J 1996 General inconsistency conditions for max-imum parsimony Effects of branch lengths and in-creasing numbers of taxa Syst Biol 45363ndash374
KISHINO H AND M HASEGAWA 1989 Evaluation ofthe maximum likelihood estimate of the evolution-ary tree topologies form DNA sequence data and thebranching order in Hominoidea J Mol Evol 29170ndash179
currenLIANG H AND K W HILU 1995 Application of thematK gene sequence to grass systematics Can J Bot74125ndash134
MABBERLEY D J 1993 The plant-book A portable dic-tionary of the vascular plants 2nd edition CambridgeUniv Press Cambridge
currenMACFARLANE T D AND L WATSON 1982The classi-cation of Poaceaesubfamily pooids Taxon31178ndash203
currenMASON-GAMER R J AND EA KELLOGG 1996Testingfor phylogenetic conict among molecular data sets inthe tribe Triticeae (Gramineae) Syst Biol 45524ndash545
currenMASON-GAMER R J C F WEIL AND E A KELLOGG1998Granule-bound starch synthase Structure func-tion andphylogenetic utility Mol Biol Evol 151658ndash1673
currenMATHEWS S AND R A SHARROCK 1996 The phy-tochrome gene family in grasses (Poaceae) A phy-logeny and evidence that grasses have a subset ofthe loci found in dicot angiosperms Mol Biol Evol131141ndash1150
currenMATHEWS S R C TSAL AND E A KELLOGG 2000Phylogenetic structure in the grass family (Poaceae)
150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel

146 SYSTEMATIC BIOLOGY VOL 51
FIGURE 5 Summary of the subfamilial relationships in the large supertrees based on 55 published source trees(including 403 genera) obtained using the ve MRP modications with column A) characters considered as irre-versible and column B) characters considered as unordered
give a basal topology very different fromthat obtained with any other modication(Table 2 and Fig 2) Ronquistrsquos (1996) propo-sition to weight each character in the bi-nary matrix by the inverse of the number ofnodes in the corresponding source tree doesnot perform as well as the other modica-tions It yields the greatest partition metricdistance and the second largest agreementsubtree distance (Fig 2) Although the twoalternatives to the BaumRagan method arebased on logical and plain arguments (ienonindependence and redundancy of matrixelements and impact of larger trees) theireffects on MRP reconstruction are not obvi-ous and do not result in topologies closerto our combined reference Indeed PU-alonewith unordered characters was rejected as asuitable alternative hypothesis to the GPWGcombined tree with the modied KH-testand gave the second smallest P-value when
used with irreversible characters (Table 2)However conclusions from the large su-pertree analysis differed because PU-alonewas the only modication to produce thesame basal branching pattern as the GPWGcombined tree (Fig 5B) whereas BR-alonewith the pooids as sister group to the PAC-CAD clade corresponded to an alternativehypothesis for the grass family that is sup-ported by some data from nuclear (Adh Gautet al 1999) and plastid (rbcL Duvall andMorton 1996) genomes Because no refer-ence phylogenetic trees are available thatcontain a number of taxa similar to those ofour large supertrees it is difcult to assesstheir topologies and to determine whetherPU-alone with irreversible characters reallygave a more accurate phylogeny of largegrasses than did the other methods Theplacement of the pooids is possibly not thebest criterion by which to judge the methods
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 147
the evolutionary position of this subfamilywithin the grass family differs as inferredwith different DNA sequence data sets
Weighting by node support as suggestedby Ronquist (1996) improved the t betweenthe additive binary matrices and the GPWGcombined tree The benecial impact of thebootstrap weighting scheme is evident fromthe increased P-values obtained with themodied KH-test (Table 2) This trend is vis-ible for the BaumRagan coding schemebut is less obvious with the Purvis codingscheme (Fig 2) Bootstrap weighting wasproposed (Ronquist 1996) as an alternativeto the Purvis coding scheme both to reducethe greater impact of large source trees oversmalleronesand to improve the effect of well-supported nodes in the MRP analysis It isnot clear how bootstrap weighting could re-duce the impact of larger trees if the smallertrees do not have much greater support val-ues than those of the larger trees Howeverweighting of charactersby bootstrapsupportwithin the Purvis coding scheme appears tobe a redundant procedure unable to greatlyimprove the MRP reconstruction from theimportant information present in bootstrapsupport values (Fig 2) In the large supertreeanalysis PU C bootstrap with unorderedcharacters produced unrealistic topologieswith the early diverging lineages embed-ded in the bambusoids (Fig 5B) The Purviscoding schemes removes important restric-tive information from the matrix (Bininda-Emonds and Bryant 1998) possibly there-fore weighting by bootstrap support wouldrandomly assign high values to characters inthe matrix where this restrictive informationhas been removed and would prevent theweighting schemes from being as effectiveas with the BaumRagan coding schemesWeighting the matrix elements by node sup-port also poses problems Not all bootstrapanalyses can be considered identical and thenumber of replicates the type of searchesdone or both will inuence the supportfound Moreover when node support is notprovided for a source tree weights cannot beassigned and information from this tree willbe down-weighted in the MRP analysis Theeffect of this is great in supertree reconstruc-tions done with older publications which ingeneral do not report node support and arehence down-weighted in the MRP analyses
An important aspect affecting the large su-pertrees that is not resolved in our compar-
isons is the placement of some rarely sam-pled taxa Taxa that are present in only a fewpublications have been allocated a high pro-portion of missing character values whichmakes parsimony analyses much more dif-cult How many of these taxa are mis-placed is difcult to determine because noreference phylogeny for the 403 genera isavailable MRP reconstruction done withoutconstraining three clades (panicoids pooidsand chloridoids) to be monophyletic endedwith rarely sampled taxa being scattered allaround the supertrees (data not shown) Thatcan be an important problem when MRPanalyses are performed on a wide taxonomicgroup comprising large numbers of rarelysampled taxa
MRP coding especially when weighted bynode support can be considered an indica-tion of the signal in the primary data witheach node represented by one (weighted)synapomorphy (see Bininda-Emonds et al1999) However we stress that supertreescannot be viewed and interpreted in ex-actly the same way as phylogenetic analy-ses based on biological characters One ma-jor problem is the difculty of assigningnode support for supertrees Bootstrappingand other resampling procedures cannot beapplied given the clear nonindependenceof the characters present in the binary ma-trix and Bremer support (Bremer 1988) iscertainly equally affected by such noninde-pendence (as is the parsimony itself hencethe different MRP modications proposed)Moreoverbranch lengths associatedwith su-pertrees are treated in the same way as mor-phological branch lengths and unlike molec-ular branch length they cannot be used toestablish divergence times This weakness iseven more signicant in taxonomic groupssuch as the grasses for which dates are dif-cult to gather from other sources (eg fossilrecords)
CONCLUSIONS
We have demonstrated that supertreesoffer an easy way of producing phyloge-nies with a large number of taxa and thatthey can give good estimates of relation-ships within these groups (in this case thegrass family) Supertrees based on the eightcharacter partitions of the GPWG data setswere found to roughly match the combinedanalysis the discrepancies of the best MRP
148 SYSTEMATIC BIOLOGY VOL 51
modications being found mainly in weaklysupported branches of the combined treeThe BaumRagan and the Purvis modica-tions gave similar results but incorporatingbootstrap support associated with preexist-ing topologies improved the BaumRaganmodication Moreover supertrees can beuseful for comparative studies (Purvis1995b Bininda-Emonds et al 1999) whetherof adaptation coevolution rates of evo-lution cospeciation or rates of effectivecladogenesis where accuracy in the branchlength of phylogenetic reconstructions is notthe primary problem (Purvis et al 1994)Supertree reconstructions are also a usefulway to help highlight poor taxonomic sam-pling and identify where previous studiesareinconsistentThey can therefore be used as anexploratory tool capable of developing newhypotheses and indicating where future re-search should be focused
ACKNOWLEDGMENTS
We are grateful to Elizabeth Kellogg and LynnClark for providing the GPWG matrix and for their com-ments on the manuscript James Farris for helpful dis-cussion regarding bootstrap weighting schemes duringthe XIX Hennig Society meeting Stephen Renvoize andMark Chase for helpful discussion on the project OlafBininda-Emonds for his comments on the manuscriptand JonathanDavis for the testing of the SuperTree 085bprogram This study was supported by the Irish HigherEducation Authority the Roche Research Foundation(99-186) Enterprise Iceland (IC200129) the AgassizFoundation and the Swiss NSF (31-52378)
REFERENCES
(References marked curren were used as sources of phyloge-nies for the supertree reconstructions)
ARNQVIST G AND D WOOSTER 1995 Meta-analysisSynthesizing research ndings in ecology and evolu-tion Trends Ecol Evol 10236ndash240
currenBARKER N P 1997 The relationships of AmphipogonElytrophorus and Cyperochloa (Poaceae) as suggestedby rbcL sequence data Telopea 7205ndash213
currenBARKER N P H P LINDER AND E H HARLEY 1995Polyphyly of Arundinoids (Poaceae) Evidence fromrbcL sequence data Syst Bot 20423ndash435
currenBARKER N P H P LINDER and E H HARLEY 1999Sequences of the grass-specic insert in the chloro-plast rpoC2 gene elucidate generic relationships of theArundinoids (Poaceae) Syst Bot 23327ndash350
currenBAUM B R 1982 A phylogenetic analysis of the tribeTriticeae (Poaceae) based on morphological charactersof the genera Can J Bot 61518ndash535
BAUM B R 1992 Combining trees as a way of com-bining data sets for phylogenetic inference and thedesirability of combining gene trees Taxon 413ndash10
currenBAUM B R J R ESTES AND P K GUPTA 1986 Assess-ment of the genomic systematic classication in theTriticeae Am J Bot 741388ndash1395
BAUM B R AND M A RAGAN 1993 Reply to A GRodrigorsquos ldquoA comment on Baumrsquos method for com-bining phylogenetic treesrdquo Taxon 42637ndash640
BININD A-EMONDS O R P AND H N BRYANT 1998Properties of matrix representation with parsimonyanalyses Syst Biol 47497ndash508
BININD A-EMONDS O R P J L GITTLEMAN ANDA PURVIS 1999 Building large trees by combiningphylogenetic information A complete phylogenyof the extant Carnivora (Mammalia) Biol Rev 74143ndash175
BREMER K 1988 The limits of amino acid sequencedata in angiosperm phylogenetic reconstruction Evo-lution 42795ndash803
currenBUCKLER E S IV AND T P HOLTSFORD 1996 Zeasystematics Ribosomal ITS evidence Mol Biol Evol13612ndash622
CAMIN J H AND R R SOKAL 1965 A method for de-ducing branching sequences in phylogeny Evolution19311ndash326
currenCATALAN P E A KELLOGG AND R G OLMS TEAD1997 Phylogeny of Poaceae subfamily Pooideaebased on chloroplast ndhFgene sequences Mol Phy-logenet Evol 8150ndash166
currenCLARK L G W ZHANG AND J F WENDEL 1995 Aphylogenyof the grass family (Poaceae) basedon ndhFsequence data Syst Bot 20436ndash460
CONSTANTINES CU M AND D SANKOFF 1995 An ef-cient algorithm for supertrees J Classif 12101ndash112
currenCUMMINGS M P L M KING AND E A KELLOGG1994 Slipped-strand mispairing in a plastid generpoC2 in grasses (Poaceae) Mol Biol Evol 111ndash8
currenDARBYSHIRE S J AND S I WARWICK 1992 Phylogenyof North American Festuca (Poaceae) and related gen-era using chloroplast DNA restriction site variationCan J Bot 702415ndash2419
currenDAVIS J I AND R J SORENG 1993Phylogenetic struc-ture in the grass family (Poaceae) as inferred fromchloroplast DNA restriction site variation Am J Bot801444ndash1454
currenDOEBLEY J M DURBIN E M GOLENBERG M TCLEGG AND D P MA 1990 Evolutionary analysisof the large subunit of carboxylase (rbcL) nucleotidesequence among the grasses (Gramineae) Evolution441097ndash1108
currenDUVALL M R AND B R MORTON 1996 Molecularphylogenetics of Poaceae An expanded analysis ofrbcL sequence data Mol Phylogenet Evol 5352ndash358
currenDUVALL M R P M PETERSON E E TERRELL ANDA H CHRISTENS EN 1993 Phylogeny of NorthAmerican oryzoid grasses as constructed from mapsof plastid DNA restriction sites Am J Bot 8083ndash88
EFFRON B E HALLORAN AND S HOLMES 1996 Boot-strap condence levels for phylogenetic trees ProcNatl Acad Sci USA 937085ndash7090
currenESEN A AND K W HILU 1991 Electrophoretic andimmunological studies of prolamins in the PoaceaeII Phylogenetic afnities of the Atistideae Taxon 405ndash17
currenESEN A AND K W HILU 1993Prolamin and immuno-logical studies in the Poaceae IV SubfamilyPanicoidsCan J Bot 71315ndash322
FARRIS J S A G KLUGE AND M J ECKHARDT 1970 Anumerical approach to phylogenetic systematics SystZool 19172ndash191
FELSENSTEIN J 1985 Condence limits on phylogeniesAn approach using the bootstrap Evolution 39783ndash791
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 149
FITCH W M 1971 Toward dening the course of evolu-tion Minimal change for a specic tree topology SystZool 20406ndash416
currenFREDERIKSEN S AND O SEBERG 1991 Phylogeneticanalysis of the Triticeae (Poaceae) Hereditas 11515ndash19
currenGAUT B S A SPEEK BR MORTON AND M T CLEGG 1999Patterns of genetic diversication within the Adhgene family in the grasses (Poaceae) Mol Biol Evol161086ndash1097
GOLDMANN N J P ANDERSON AND A G RODRIGO2000 Likelihood-based tests of topologies in phyloge-netics Syst Biol 49652ndash670
GORDON A D 1980 On the assessment and compar-ison of classications Pages 149ndash160 in Analyse dedonnees et informatique (R Tomassone ed) INRALe Chesnay France
GPWG2000A phylogeny of the grass family (Poaceae)as inferred from eight character sets Pages 3ndash7 inGrasses Systematics and evolution (S W L Jacobsand J Everett eds) CSIRO Melbourne Australia
currenGREBENS TEIN B O GREBENS TEIN W SAUER ANDV HEMLEBEN 1996 Distribution and complex organi-zation of satellite DNA sequences in Aveneae speciesGenome 391045ndash1050
currenGREBENS TEIN B M ROSER W SAUER ANDV HEMLEBEN 1998 Molecular phylogenetic re-lationships in Aveneae (Poaceae) species and othergrasses as inferred from ITS1 and ITS2 rDNAsequences Plant Syst Evol 213233ndash250
currenHAMBY R K AND EA ZIMMER 1987Ribosomal RNAsequences for inferring phylogeny within the grassfamily (Poaceae) Plant Syst Evol 16029ndash37
HARVEY P H AND M D PAGEL 1991The comparativemethod in evolutionary biology Oxford Univ PressLondon
HILLIS D M 1996 Inferring complex phylogenies Na-ture 383130ndash131
HILLIS D M AND J J BULL 1993 An empirical test ofbootstrapping as a method for assessing condencein phylogenetic analysis Syst Biol 42182ndash192
currenHILU K W L A ALICE AND H LIANG 1999 Phy-logeny of Poaceae inferred from matK sequencesAnn Mo Bot Gard 86835ndash851
currenHILU K W AND A ESEN 1992Prolamin andimmuno-logical strudies in the Poaceae III Subfamily Chlori-doids Am J Bot 80104ndash113
currenHILU K W AND H LIANG 1997 The matK gene Se-quence variation and application in plant systematicsAm J Bot 84830ndash839
currenHILU K W AND K WRIGHT 1982 Systematics ofGramineae A cluster analysis study Taxon 319ndash37
currenHSIAO C N J CHATTERTON K H ASAY AND K BJENS EN 1993 Phylogenetic relationships of 10 grassspecies An assessment of phylogenetic utility of theinternal transcribed spacer region in nuclear riboso-mal DNA in monocots Genome 37112ndash120
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1994 Phylogenetic relationships of themonogenomic species of the wheat tribe Triticeae(Poaceae) inferred from nuclear rDNA (internal tran-scribed spacer) sequences Genome 38211ndash223
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1995Molecular phylogeny of the pooids(Poaceae) based on nuclear rDNA (ITS) sequencesTheor Appl Genet 90389ndash398
currenHSIAO C S W JACOBS N P BARKER AND N JCHATTERTON 1998 A molecular phylogeny of the
subfamily arundinoids (Poaceae) based on sequencesof rDNA Aust Syst Bot 1141ndash52
currenHSIAO C S W L JACOBS N J CHATTERTON AND KH ASAY 1999 A molecular phylogeny of the grassfamily (Poaceae) based on the sequences of nuclearribosomal DNA (ITS) Aust Syst Bot 11667ndash688
JOHNS ON L A AND D E SOLTIS 1998 Assessing con-gruence Empirical examples from molecular dataPages 297ndash348 in Molecular systematics of plants IIDNA sequencing (D E Soltis P S Soltis and J JDoyle eds) Kluwer Academic Publishers NorwellMassachusetts
currenKATAYAMA H AND Y OGIHARA 1996 Phylogeneticafnities of the grasses to other monocots as re-vealed by molecular analysis of chloroplast DNACurr Genet 29572ndash581
currenKELCHNER S A AND L G CLARK 1997 Molecu-lar evolution and phylogenetic utility of the chloro-plast rpl16 intron in Chusquea and the bambusoids(Poaceae) Mol Phylogenet Evol 8385ndash397
currenKELLOGG E A 1989 Comments on genomic genera inthe Triticeae (Poaceae) Am J Bot 76796ndash805
currenKELLOGG E A 1992Tools for studying the chloroplastgenome in the Triticeae (Gramineae) An ecoRI mapa diagnostic deletion and support for Bromus as anoutgroup Am J Bot 79186ndash197
currenKELLOGG E A 1998aRelationships of cereal crops andother grassesProc NatlAcad Sci USA952005ndash2010
currenKELLOGG E A 1998bWhorsquos related to whom Recentresults from molecular systematic studies Curr OpinPlant Biol 1149ndash158
currenKELLOGG E A AND R APPELS 1995 Intraspecic andinterspecic variation in 5S RNA genes are decoupledin diploid wheat relatives Genetics 140325ndash343
currenKELLOGG E A AND L WATSON 1993 Phyloge-netic studies of a large data set I bambusoids An-dropogonodae and pooids (Gramineae) Bot Rev59273ndash311
KIM J 1996 General inconsistency conditions for max-imum parsimony Effects of branch lengths and in-creasing numbers of taxa Syst Biol 45363ndash374
KISHINO H AND M HASEGAWA 1989 Evaluation ofthe maximum likelihood estimate of the evolution-ary tree topologies form DNA sequence data and thebranching order in Hominoidea J Mol Evol 29170ndash179
currenLIANG H AND K W HILU 1995 Application of thematK gene sequence to grass systematics Can J Bot74125ndash134
MABBERLEY D J 1993 The plant-book A portable dic-tionary of the vascular plants 2nd edition CambridgeUniv Press Cambridge
currenMACFARLANE T D AND L WATSON 1982The classi-cation of Poaceaesubfamily pooids Taxon31178ndash203
currenMASON-GAMER R J AND EA KELLOGG 1996Testingfor phylogenetic conict among molecular data sets inthe tribe Triticeae (Gramineae) Syst Biol 45524ndash545
currenMASON-GAMER R J C F WEIL AND E A KELLOGG1998Granule-bound starch synthase Structure func-tion andphylogenetic utility Mol Biol Evol 151658ndash1673
currenMATHEWS S AND R A SHARROCK 1996 The phy-tochrome gene family in grasses (Poaceae) A phy-logeny and evidence that grasses have a subset ofthe loci found in dicot angiosperms Mol Biol Evol131141ndash1150
currenMATHEWS S R C TSAL AND E A KELLOGG 2000Phylogenetic structure in the grass family (Poaceae)
150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel

2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 147
the evolutionary position of this subfamilywithin the grass family differs as inferredwith different DNA sequence data sets
Weighting by node support as suggestedby Ronquist (1996) improved the t betweenthe additive binary matrices and the GPWGcombined tree The benecial impact of thebootstrap weighting scheme is evident fromthe increased P-values obtained with themodied KH-test (Table 2) This trend is vis-ible for the BaumRagan coding schemebut is less obvious with the Purvis codingscheme (Fig 2) Bootstrap weighting wasproposed (Ronquist 1996) as an alternativeto the Purvis coding scheme both to reducethe greater impact of large source trees oversmalleronesand to improve the effect of well-supported nodes in the MRP analysis It isnot clear how bootstrap weighting could re-duce the impact of larger trees if the smallertrees do not have much greater support val-ues than those of the larger trees Howeverweighting of charactersby bootstrapsupportwithin the Purvis coding scheme appears tobe a redundant procedure unable to greatlyimprove the MRP reconstruction from theimportant information present in bootstrapsupport values (Fig 2) In the large supertreeanalysis PU C bootstrap with unorderedcharacters produced unrealistic topologieswith the early diverging lineages embed-ded in the bambusoids (Fig 5B) The Purviscoding schemes removes important restric-tive information from the matrix (Bininda-Emonds and Bryant 1998) possibly there-fore weighting by bootstrap support wouldrandomly assign high values to characters inthe matrix where this restrictive informationhas been removed and would prevent theweighting schemes from being as effectiveas with the BaumRagan coding schemesWeighting the matrix elements by node sup-port also poses problems Not all bootstrapanalyses can be considered identical and thenumber of replicates the type of searchesdone or both will inuence the supportfound Moreover when node support is notprovided for a source tree weights cannot beassigned and information from this tree willbe down-weighted in the MRP analysis Theeffect of this is great in supertree reconstruc-tions done with older publications which ingeneral do not report node support and arehence down-weighted in the MRP analyses
An important aspect affecting the large su-pertrees that is not resolved in our compar-
isons is the placement of some rarely sam-pled taxa Taxa that are present in only a fewpublications have been allocated a high pro-portion of missing character values whichmakes parsimony analyses much more dif-cult How many of these taxa are mis-placed is difcult to determine because noreference phylogeny for the 403 genera isavailable MRP reconstruction done withoutconstraining three clades (panicoids pooidsand chloridoids) to be monophyletic endedwith rarely sampled taxa being scattered allaround the supertrees (data not shown) Thatcan be an important problem when MRPanalyses are performed on a wide taxonomicgroup comprising large numbers of rarelysampled taxa
MRP coding especially when weighted bynode support can be considered an indica-tion of the signal in the primary data witheach node represented by one (weighted)synapomorphy (see Bininda-Emonds et al1999) However we stress that supertreescannot be viewed and interpreted in ex-actly the same way as phylogenetic analy-ses based on biological characters One ma-jor problem is the difculty of assigningnode support for supertrees Bootstrappingand other resampling procedures cannot beapplied given the clear nonindependenceof the characters present in the binary ma-trix and Bremer support (Bremer 1988) iscertainly equally affected by such noninde-pendence (as is the parsimony itself hencethe different MRP modications proposed)Moreoverbranch lengths associatedwith su-pertrees are treated in the same way as mor-phological branch lengths and unlike molec-ular branch length they cannot be used toestablish divergence times This weakness iseven more signicant in taxonomic groupssuch as the grasses for which dates are dif-cult to gather from other sources (eg fossilrecords)
CONCLUSIONS
We have demonstrated that supertreesoffer an easy way of producing phyloge-nies with a large number of taxa and thatthey can give good estimates of relation-ships within these groups (in this case thegrass family) Supertrees based on the eightcharacter partitions of the GPWG data setswere found to roughly match the combinedanalysis the discrepancies of the best MRP
148 SYSTEMATIC BIOLOGY VOL 51
modications being found mainly in weaklysupported branches of the combined treeThe BaumRagan and the Purvis modica-tions gave similar results but incorporatingbootstrap support associated with preexist-ing topologies improved the BaumRaganmodication Moreover supertrees can beuseful for comparative studies (Purvis1995b Bininda-Emonds et al 1999) whetherof adaptation coevolution rates of evo-lution cospeciation or rates of effectivecladogenesis where accuracy in the branchlength of phylogenetic reconstructions is notthe primary problem (Purvis et al 1994)Supertree reconstructions are also a usefulway to help highlight poor taxonomic sam-pling and identify where previous studiesareinconsistentThey can therefore be used as anexploratory tool capable of developing newhypotheses and indicating where future re-search should be focused
ACKNOWLEDGMENTS
We are grateful to Elizabeth Kellogg and LynnClark for providing the GPWG matrix and for their com-ments on the manuscript James Farris for helpful dis-cussion regarding bootstrap weighting schemes duringthe XIX Hennig Society meeting Stephen Renvoize andMark Chase for helpful discussion on the project OlafBininda-Emonds for his comments on the manuscriptand JonathanDavis for the testing of the SuperTree 085bprogram This study was supported by the Irish HigherEducation Authority the Roche Research Foundation(99-186) Enterprise Iceland (IC200129) the AgassizFoundation and the Swiss NSF (31-52378)
REFERENCES
(References marked curren were used as sources of phyloge-nies for the supertree reconstructions)
ARNQVIST G AND D WOOSTER 1995 Meta-analysisSynthesizing research ndings in ecology and evolu-tion Trends Ecol Evol 10236ndash240
currenBARKER N P 1997 The relationships of AmphipogonElytrophorus and Cyperochloa (Poaceae) as suggestedby rbcL sequence data Telopea 7205ndash213
currenBARKER N P H P LINDER AND E H HARLEY 1995Polyphyly of Arundinoids (Poaceae) Evidence fromrbcL sequence data Syst Bot 20423ndash435
currenBARKER N P H P LINDER and E H HARLEY 1999Sequences of the grass-specic insert in the chloro-plast rpoC2 gene elucidate generic relationships of theArundinoids (Poaceae) Syst Bot 23327ndash350
currenBAUM B R 1982 A phylogenetic analysis of the tribeTriticeae (Poaceae) based on morphological charactersof the genera Can J Bot 61518ndash535
BAUM B R 1992 Combining trees as a way of com-bining data sets for phylogenetic inference and thedesirability of combining gene trees Taxon 413ndash10
currenBAUM B R J R ESTES AND P K GUPTA 1986 Assess-ment of the genomic systematic classication in theTriticeae Am J Bot 741388ndash1395
BAUM B R AND M A RAGAN 1993 Reply to A GRodrigorsquos ldquoA comment on Baumrsquos method for com-bining phylogenetic treesrdquo Taxon 42637ndash640
BININD A-EMONDS O R P AND H N BRYANT 1998Properties of matrix representation with parsimonyanalyses Syst Biol 47497ndash508
BININD A-EMONDS O R P J L GITTLEMAN ANDA PURVIS 1999 Building large trees by combiningphylogenetic information A complete phylogenyof the extant Carnivora (Mammalia) Biol Rev 74143ndash175
BREMER K 1988 The limits of amino acid sequencedata in angiosperm phylogenetic reconstruction Evo-lution 42795ndash803
currenBUCKLER E S IV AND T P HOLTSFORD 1996 Zeasystematics Ribosomal ITS evidence Mol Biol Evol13612ndash622
CAMIN J H AND R R SOKAL 1965 A method for de-ducing branching sequences in phylogeny Evolution19311ndash326
currenCATALAN P E A KELLOGG AND R G OLMS TEAD1997 Phylogeny of Poaceae subfamily Pooideaebased on chloroplast ndhFgene sequences Mol Phy-logenet Evol 8150ndash166
currenCLARK L G W ZHANG AND J F WENDEL 1995 Aphylogenyof the grass family (Poaceae) basedon ndhFsequence data Syst Bot 20436ndash460
CONSTANTINES CU M AND D SANKOFF 1995 An ef-cient algorithm for supertrees J Classif 12101ndash112
currenCUMMINGS M P L M KING AND E A KELLOGG1994 Slipped-strand mispairing in a plastid generpoC2 in grasses (Poaceae) Mol Biol Evol 111ndash8
currenDARBYSHIRE S J AND S I WARWICK 1992 Phylogenyof North American Festuca (Poaceae) and related gen-era using chloroplast DNA restriction site variationCan J Bot 702415ndash2419
currenDAVIS J I AND R J SORENG 1993Phylogenetic struc-ture in the grass family (Poaceae) as inferred fromchloroplast DNA restriction site variation Am J Bot801444ndash1454
currenDOEBLEY J M DURBIN E M GOLENBERG M TCLEGG AND D P MA 1990 Evolutionary analysisof the large subunit of carboxylase (rbcL) nucleotidesequence among the grasses (Gramineae) Evolution441097ndash1108
currenDUVALL M R AND B R MORTON 1996 Molecularphylogenetics of Poaceae An expanded analysis ofrbcL sequence data Mol Phylogenet Evol 5352ndash358
currenDUVALL M R P M PETERSON E E TERRELL ANDA H CHRISTENS EN 1993 Phylogeny of NorthAmerican oryzoid grasses as constructed from mapsof plastid DNA restriction sites Am J Bot 8083ndash88
EFFRON B E HALLORAN AND S HOLMES 1996 Boot-strap condence levels for phylogenetic trees ProcNatl Acad Sci USA 937085ndash7090
currenESEN A AND K W HILU 1991 Electrophoretic andimmunological studies of prolamins in the PoaceaeII Phylogenetic afnities of the Atistideae Taxon 405ndash17
currenESEN A AND K W HILU 1993Prolamin and immuno-logical studies in the Poaceae IV SubfamilyPanicoidsCan J Bot 71315ndash322
FARRIS J S A G KLUGE AND M J ECKHARDT 1970 Anumerical approach to phylogenetic systematics SystZool 19172ndash191
FELSENSTEIN J 1985 Condence limits on phylogeniesAn approach using the bootstrap Evolution 39783ndash791
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 149
FITCH W M 1971 Toward dening the course of evolu-tion Minimal change for a specic tree topology SystZool 20406ndash416
currenFREDERIKSEN S AND O SEBERG 1991 Phylogeneticanalysis of the Triticeae (Poaceae) Hereditas 11515ndash19
currenGAUT B S A SPEEK BR MORTON AND M T CLEGG 1999Patterns of genetic diversication within the Adhgene family in the grasses (Poaceae) Mol Biol Evol161086ndash1097
GOLDMANN N J P ANDERSON AND A G RODRIGO2000 Likelihood-based tests of topologies in phyloge-netics Syst Biol 49652ndash670
GORDON A D 1980 On the assessment and compar-ison of classications Pages 149ndash160 in Analyse dedonnees et informatique (R Tomassone ed) INRALe Chesnay France
GPWG2000A phylogeny of the grass family (Poaceae)as inferred from eight character sets Pages 3ndash7 inGrasses Systematics and evolution (S W L Jacobsand J Everett eds) CSIRO Melbourne Australia
currenGREBENS TEIN B O GREBENS TEIN W SAUER ANDV HEMLEBEN 1996 Distribution and complex organi-zation of satellite DNA sequences in Aveneae speciesGenome 391045ndash1050
currenGREBENS TEIN B M ROSER W SAUER ANDV HEMLEBEN 1998 Molecular phylogenetic re-lationships in Aveneae (Poaceae) species and othergrasses as inferred from ITS1 and ITS2 rDNAsequences Plant Syst Evol 213233ndash250
currenHAMBY R K AND EA ZIMMER 1987Ribosomal RNAsequences for inferring phylogeny within the grassfamily (Poaceae) Plant Syst Evol 16029ndash37
HARVEY P H AND M D PAGEL 1991The comparativemethod in evolutionary biology Oxford Univ PressLondon
HILLIS D M 1996 Inferring complex phylogenies Na-ture 383130ndash131
HILLIS D M AND J J BULL 1993 An empirical test ofbootstrapping as a method for assessing condencein phylogenetic analysis Syst Biol 42182ndash192
currenHILU K W L A ALICE AND H LIANG 1999 Phy-logeny of Poaceae inferred from matK sequencesAnn Mo Bot Gard 86835ndash851
currenHILU K W AND A ESEN 1992Prolamin andimmuno-logical strudies in the Poaceae III Subfamily Chlori-doids Am J Bot 80104ndash113
currenHILU K W AND H LIANG 1997 The matK gene Se-quence variation and application in plant systematicsAm J Bot 84830ndash839
currenHILU K W AND K WRIGHT 1982 Systematics ofGramineae A cluster analysis study Taxon 319ndash37
currenHSIAO C N J CHATTERTON K H ASAY AND K BJENS EN 1993 Phylogenetic relationships of 10 grassspecies An assessment of phylogenetic utility of theinternal transcribed spacer region in nuclear riboso-mal DNA in monocots Genome 37112ndash120
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1994 Phylogenetic relationships of themonogenomic species of the wheat tribe Triticeae(Poaceae) inferred from nuclear rDNA (internal tran-scribed spacer) sequences Genome 38211ndash223
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1995Molecular phylogeny of the pooids(Poaceae) based on nuclear rDNA (ITS) sequencesTheor Appl Genet 90389ndash398
currenHSIAO C S W JACOBS N P BARKER AND N JCHATTERTON 1998 A molecular phylogeny of the
subfamily arundinoids (Poaceae) based on sequencesof rDNA Aust Syst Bot 1141ndash52
currenHSIAO C S W L JACOBS N J CHATTERTON AND KH ASAY 1999 A molecular phylogeny of the grassfamily (Poaceae) based on the sequences of nuclearribosomal DNA (ITS) Aust Syst Bot 11667ndash688
JOHNS ON L A AND D E SOLTIS 1998 Assessing con-gruence Empirical examples from molecular dataPages 297ndash348 in Molecular systematics of plants IIDNA sequencing (D E Soltis P S Soltis and J JDoyle eds) Kluwer Academic Publishers NorwellMassachusetts
currenKATAYAMA H AND Y OGIHARA 1996 Phylogeneticafnities of the grasses to other monocots as re-vealed by molecular analysis of chloroplast DNACurr Genet 29572ndash581
currenKELCHNER S A AND L G CLARK 1997 Molecu-lar evolution and phylogenetic utility of the chloro-plast rpl16 intron in Chusquea and the bambusoids(Poaceae) Mol Phylogenet Evol 8385ndash397
currenKELLOGG E A 1989 Comments on genomic genera inthe Triticeae (Poaceae) Am J Bot 76796ndash805
currenKELLOGG E A 1992Tools for studying the chloroplastgenome in the Triticeae (Gramineae) An ecoRI mapa diagnostic deletion and support for Bromus as anoutgroup Am J Bot 79186ndash197
currenKELLOGG E A 1998aRelationships of cereal crops andother grassesProc NatlAcad Sci USA952005ndash2010
currenKELLOGG E A 1998bWhorsquos related to whom Recentresults from molecular systematic studies Curr OpinPlant Biol 1149ndash158
currenKELLOGG E A AND R APPELS 1995 Intraspecic andinterspecic variation in 5S RNA genes are decoupledin diploid wheat relatives Genetics 140325ndash343
currenKELLOGG E A AND L WATSON 1993 Phyloge-netic studies of a large data set I bambusoids An-dropogonodae and pooids (Gramineae) Bot Rev59273ndash311
KIM J 1996 General inconsistency conditions for max-imum parsimony Effects of branch lengths and in-creasing numbers of taxa Syst Biol 45363ndash374
KISHINO H AND M HASEGAWA 1989 Evaluation ofthe maximum likelihood estimate of the evolution-ary tree topologies form DNA sequence data and thebranching order in Hominoidea J Mol Evol 29170ndash179
currenLIANG H AND K W HILU 1995 Application of thematK gene sequence to grass systematics Can J Bot74125ndash134
MABBERLEY D J 1993 The plant-book A portable dic-tionary of the vascular plants 2nd edition CambridgeUniv Press Cambridge
currenMACFARLANE T D AND L WATSON 1982The classi-cation of Poaceaesubfamily pooids Taxon31178ndash203
currenMASON-GAMER R J AND EA KELLOGG 1996Testingfor phylogenetic conict among molecular data sets inthe tribe Triticeae (Gramineae) Syst Biol 45524ndash545
currenMASON-GAMER R J C F WEIL AND E A KELLOGG1998Granule-bound starch synthase Structure func-tion andphylogenetic utility Mol Biol Evol 151658ndash1673
currenMATHEWS S AND R A SHARROCK 1996 The phy-tochrome gene family in grasses (Poaceae) A phy-logeny and evidence that grasses have a subset ofthe loci found in dicot angiosperms Mol Biol Evol131141ndash1150
currenMATHEWS S R C TSAL AND E A KELLOGG 2000Phylogenetic structure in the grass family (Poaceae)
150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel

148 SYSTEMATIC BIOLOGY VOL 51
modications being found mainly in weaklysupported branches of the combined treeThe BaumRagan and the Purvis modica-tions gave similar results but incorporatingbootstrap support associated with preexist-ing topologies improved the BaumRaganmodication Moreover supertrees can beuseful for comparative studies (Purvis1995b Bininda-Emonds et al 1999) whetherof adaptation coevolution rates of evo-lution cospeciation or rates of effectivecladogenesis where accuracy in the branchlength of phylogenetic reconstructions is notthe primary problem (Purvis et al 1994)Supertree reconstructions are also a usefulway to help highlight poor taxonomic sam-pling and identify where previous studiesareinconsistentThey can therefore be used as anexploratory tool capable of developing newhypotheses and indicating where future re-search should be focused
ACKNOWLEDGMENTS
We are grateful to Elizabeth Kellogg and LynnClark for providing the GPWG matrix and for their com-ments on the manuscript James Farris for helpful dis-cussion regarding bootstrap weighting schemes duringthe XIX Hennig Society meeting Stephen Renvoize andMark Chase for helpful discussion on the project OlafBininda-Emonds for his comments on the manuscriptand JonathanDavis for the testing of the SuperTree 085bprogram This study was supported by the Irish HigherEducation Authority the Roche Research Foundation(99-186) Enterprise Iceland (IC200129) the AgassizFoundation and the Swiss NSF (31-52378)
REFERENCES
(References marked curren were used as sources of phyloge-nies for the supertree reconstructions)
ARNQVIST G AND D WOOSTER 1995 Meta-analysisSynthesizing research ndings in ecology and evolu-tion Trends Ecol Evol 10236ndash240
currenBARKER N P 1997 The relationships of AmphipogonElytrophorus and Cyperochloa (Poaceae) as suggestedby rbcL sequence data Telopea 7205ndash213
currenBARKER N P H P LINDER AND E H HARLEY 1995Polyphyly of Arundinoids (Poaceae) Evidence fromrbcL sequence data Syst Bot 20423ndash435
currenBARKER N P H P LINDER and E H HARLEY 1999Sequences of the grass-specic insert in the chloro-plast rpoC2 gene elucidate generic relationships of theArundinoids (Poaceae) Syst Bot 23327ndash350
currenBAUM B R 1982 A phylogenetic analysis of the tribeTriticeae (Poaceae) based on morphological charactersof the genera Can J Bot 61518ndash535
BAUM B R 1992 Combining trees as a way of com-bining data sets for phylogenetic inference and thedesirability of combining gene trees Taxon 413ndash10
currenBAUM B R J R ESTES AND P K GUPTA 1986 Assess-ment of the genomic systematic classication in theTriticeae Am J Bot 741388ndash1395
BAUM B R AND M A RAGAN 1993 Reply to A GRodrigorsquos ldquoA comment on Baumrsquos method for com-bining phylogenetic treesrdquo Taxon 42637ndash640
BININD A-EMONDS O R P AND H N BRYANT 1998Properties of matrix representation with parsimonyanalyses Syst Biol 47497ndash508
BININD A-EMONDS O R P J L GITTLEMAN ANDA PURVIS 1999 Building large trees by combiningphylogenetic information A complete phylogenyof the extant Carnivora (Mammalia) Biol Rev 74143ndash175
BREMER K 1988 The limits of amino acid sequencedata in angiosperm phylogenetic reconstruction Evo-lution 42795ndash803
currenBUCKLER E S IV AND T P HOLTSFORD 1996 Zeasystematics Ribosomal ITS evidence Mol Biol Evol13612ndash622
CAMIN J H AND R R SOKAL 1965 A method for de-ducing branching sequences in phylogeny Evolution19311ndash326
currenCATALAN P E A KELLOGG AND R G OLMS TEAD1997 Phylogeny of Poaceae subfamily Pooideaebased on chloroplast ndhFgene sequences Mol Phy-logenet Evol 8150ndash166
currenCLARK L G W ZHANG AND J F WENDEL 1995 Aphylogenyof the grass family (Poaceae) basedon ndhFsequence data Syst Bot 20436ndash460
CONSTANTINES CU M AND D SANKOFF 1995 An ef-cient algorithm for supertrees J Classif 12101ndash112
currenCUMMINGS M P L M KING AND E A KELLOGG1994 Slipped-strand mispairing in a plastid generpoC2 in grasses (Poaceae) Mol Biol Evol 111ndash8
currenDARBYSHIRE S J AND S I WARWICK 1992 Phylogenyof North American Festuca (Poaceae) and related gen-era using chloroplast DNA restriction site variationCan J Bot 702415ndash2419
currenDAVIS J I AND R J SORENG 1993Phylogenetic struc-ture in the grass family (Poaceae) as inferred fromchloroplast DNA restriction site variation Am J Bot801444ndash1454
currenDOEBLEY J M DURBIN E M GOLENBERG M TCLEGG AND D P MA 1990 Evolutionary analysisof the large subunit of carboxylase (rbcL) nucleotidesequence among the grasses (Gramineae) Evolution441097ndash1108
currenDUVALL M R AND B R MORTON 1996 Molecularphylogenetics of Poaceae An expanded analysis ofrbcL sequence data Mol Phylogenet Evol 5352ndash358
currenDUVALL M R P M PETERSON E E TERRELL ANDA H CHRISTENS EN 1993 Phylogeny of NorthAmerican oryzoid grasses as constructed from mapsof plastid DNA restriction sites Am J Bot 8083ndash88
EFFRON B E HALLORAN AND S HOLMES 1996 Boot-strap condence levels for phylogenetic trees ProcNatl Acad Sci USA 937085ndash7090
currenESEN A AND K W HILU 1991 Electrophoretic andimmunological studies of prolamins in the PoaceaeII Phylogenetic afnities of the Atistideae Taxon 405ndash17
currenESEN A AND K W HILU 1993Prolamin and immuno-logical studies in the Poaceae IV SubfamilyPanicoidsCan J Bot 71315ndash322
FARRIS J S A G KLUGE AND M J ECKHARDT 1970 Anumerical approach to phylogenetic systematics SystZool 19172ndash191
FELSENSTEIN J 1985 Condence limits on phylogeniesAn approach using the bootstrap Evolution 39783ndash791
2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 149
FITCH W M 1971 Toward dening the course of evolu-tion Minimal change for a specic tree topology SystZool 20406ndash416
currenFREDERIKSEN S AND O SEBERG 1991 Phylogeneticanalysis of the Triticeae (Poaceae) Hereditas 11515ndash19
currenGAUT B S A SPEEK BR MORTON AND M T CLEGG 1999Patterns of genetic diversication within the Adhgene family in the grasses (Poaceae) Mol Biol Evol161086ndash1097
GOLDMANN N J P ANDERSON AND A G RODRIGO2000 Likelihood-based tests of topologies in phyloge-netics Syst Biol 49652ndash670
GORDON A D 1980 On the assessment and compar-ison of classications Pages 149ndash160 in Analyse dedonnees et informatique (R Tomassone ed) INRALe Chesnay France
GPWG2000A phylogeny of the grass family (Poaceae)as inferred from eight character sets Pages 3ndash7 inGrasses Systematics and evolution (S W L Jacobsand J Everett eds) CSIRO Melbourne Australia
currenGREBENS TEIN B O GREBENS TEIN W SAUER ANDV HEMLEBEN 1996 Distribution and complex organi-zation of satellite DNA sequences in Aveneae speciesGenome 391045ndash1050
currenGREBENS TEIN B M ROSER W SAUER ANDV HEMLEBEN 1998 Molecular phylogenetic re-lationships in Aveneae (Poaceae) species and othergrasses as inferred from ITS1 and ITS2 rDNAsequences Plant Syst Evol 213233ndash250
currenHAMBY R K AND EA ZIMMER 1987Ribosomal RNAsequences for inferring phylogeny within the grassfamily (Poaceae) Plant Syst Evol 16029ndash37
HARVEY P H AND M D PAGEL 1991The comparativemethod in evolutionary biology Oxford Univ PressLondon
HILLIS D M 1996 Inferring complex phylogenies Na-ture 383130ndash131
HILLIS D M AND J J BULL 1993 An empirical test ofbootstrapping as a method for assessing condencein phylogenetic analysis Syst Biol 42182ndash192
currenHILU K W L A ALICE AND H LIANG 1999 Phy-logeny of Poaceae inferred from matK sequencesAnn Mo Bot Gard 86835ndash851
currenHILU K W AND A ESEN 1992Prolamin andimmuno-logical strudies in the Poaceae III Subfamily Chlori-doids Am J Bot 80104ndash113
currenHILU K W AND H LIANG 1997 The matK gene Se-quence variation and application in plant systematicsAm J Bot 84830ndash839
currenHILU K W AND K WRIGHT 1982 Systematics ofGramineae A cluster analysis study Taxon 319ndash37
currenHSIAO C N J CHATTERTON K H ASAY AND K BJENS EN 1993 Phylogenetic relationships of 10 grassspecies An assessment of phylogenetic utility of theinternal transcribed spacer region in nuclear riboso-mal DNA in monocots Genome 37112ndash120
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1994 Phylogenetic relationships of themonogenomic species of the wheat tribe Triticeae(Poaceae) inferred from nuclear rDNA (internal tran-scribed spacer) sequences Genome 38211ndash223
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1995Molecular phylogeny of the pooids(Poaceae) based on nuclear rDNA (ITS) sequencesTheor Appl Genet 90389ndash398
currenHSIAO C S W JACOBS N P BARKER AND N JCHATTERTON 1998 A molecular phylogeny of the
subfamily arundinoids (Poaceae) based on sequencesof rDNA Aust Syst Bot 1141ndash52
currenHSIAO C S W L JACOBS N J CHATTERTON AND KH ASAY 1999 A molecular phylogeny of the grassfamily (Poaceae) based on the sequences of nuclearribosomal DNA (ITS) Aust Syst Bot 11667ndash688
JOHNS ON L A AND D E SOLTIS 1998 Assessing con-gruence Empirical examples from molecular dataPages 297ndash348 in Molecular systematics of plants IIDNA sequencing (D E Soltis P S Soltis and J JDoyle eds) Kluwer Academic Publishers NorwellMassachusetts
currenKATAYAMA H AND Y OGIHARA 1996 Phylogeneticafnities of the grasses to other monocots as re-vealed by molecular analysis of chloroplast DNACurr Genet 29572ndash581
currenKELCHNER S A AND L G CLARK 1997 Molecu-lar evolution and phylogenetic utility of the chloro-plast rpl16 intron in Chusquea and the bambusoids(Poaceae) Mol Phylogenet Evol 8385ndash397
currenKELLOGG E A 1989 Comments on genomic genera inthe Triticeae (Poaceae) Am J Bot 76796ndash805
currenKELLOGG E A 1992Tools for studying the chloroplastgenome in the Triticeae (Gramineae) An ecoRI mapa diagnostic deletion and support for Bromus as anoutgroup Am J Bot 79186ndash197
currenKELLOGG E A 1998aRelationships of cereal crops andother grassesProc NatlAcad Sci USA952005ndash2010
currenKELLOGG E A 1998bWhorsquos related to whom Recentresults from molecular systematic studies Curr OpinPlant Biol 1149ndash158
currenKELLOGG E A AND R APPELS 1995 Intraspecic andinterspecic variation in 5S RNA genes are decoupledin diploid wheat relatives Genetics 140325ndash343
currenKELLOGG E A AND L WATSON 1993 Phyloge-netic studies of a large data set I bambusoids An-dropogonodae and pooids (Gramineae) Bot Rev59273ndash311
KIM J 1996 General inconsistency conditions for max-imum parsimony Effects of branch lengths and in-creasing numbers of taxa Syst Biol 45363ndash374
KISHINO H AND M HASEGAWA 1989 Evaluation ofthe maximum likelihood estimate of the evolution-ary tree topologies form DNA sequence data and thebranching order in Hominoidea J Mol Evol 29170ndash179
currenLIANG H AND K W HILU 1995 Application of thematK gene sequence to grass systematics Can J Bot74125ndash134
MABBERLEY D J 1993 The plant-book A portable dic-tionary of the vascular plants 2nd edition CambridgeUniv Press Cambridge
currenMACFARLANE T D AND L WATSON 1982The classi-cation of Poaceaesubfamily pooids Taxon31178ndash203
currenMASON-GAMER R J AND EA KELLOGG 1996Testingfor phylogenetic conict among molecular data sets inthe tribe Triticeae (Gramineae) Syst Biol 45524ndash545
currenMASON-GAMER R J C F WEIL AND E A KELLOGG1998Granule-bound starch synthase Structure func-tion andphylogenetic utility Mol Biol Evol 151658ndash1673
currenMATHEWS S AND R A SHARROCK 1996 The phy-tochrome gene family in grasses (Poaceae) A phy-logeny and evidence that grasses have a subset ofthe loci found in dicot angiosperms Mol Biol Evol131141ndash1150
currenMATHEWS S R C TSAL AND E A KELLOGG 2000Phylogenetic structure in the grass family (Poaceae)
150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel

2002 SALAMIN ET ALmdashSUPERTREE RECONSTRUCTIONS IN THE POACEAE 149
FITCH W M 1971 Toward dening the course of evolu-tion Minimal change for a specic tree topology SystZool 20406ndash416
currenFREDERIKSEN S AND O SEBERG 1991 Phylogeneticanalysis of the Triticeae (Poaceae) Hereditas 11515ndash19
currenGAUT B S A SPEEK BR MORTON AND M T CLEGG 1999Patterns of genetic diversication within the Adhgene family in the grasses (Poaceae) Mol Biol Evol161086ndash1097
GOLDMANN N J P ANDERSON AND A G RODRIGO2000 Likelihood-based tests of topologies in phyloge-netics Syst Biol 49652ndash670
GORDON A D 1980 On the assessment and compar-ison of classications Pages 149ndash160 in Analyse dedonnees et informatique (R Tomassone ed) INRALe Chesnay France
GPWG2000A phylogeny of the grass family (Poaceae)as inferred from eight character sets Pages 3ndash7 inGrasses Systematics and evolution (S W L Jacobsand J Everett eds) CSIRO Melbourne Australia
currenGREBENS TEIN B O GREBENS TEIN W SAUER ANDV HEMLEBEN 1996 Distribution and complex organi-zation of satellite DNA sequences in Aveneae speciesGenome 391045ndash1050
currenGREBENS TEIN B M ROSER W SAUER ANDV HEMLEBEN 1998 Molecular phylogenetic re-lationships in Aveneae (Poaceae) species and othergrasses as inferred from ITS1 and ITS2 rDNAsequences Plant Syst Evol 213233ndash250
currenHAMBY R K AND EA ZIMMER 1987Ribosomal RNAsequences for inferring phylogeny within the grassfamily (Poaceae) Plant Syst Evol 16029ndash37
HARVEY P H AND M D PAGEL 1991The comparativemethod in evolutionary biology Oxford Univ PressLondon
HILLIS D M 1996 Inferring complex phylogenies Na-ture 383130ndash131
HILLIS D M AND J J BULL 1993 An empirical test ofbootstrapping as a method for assessing condencein phylogenetic analysis Syst Biol 42182ndash192
currenHILU K W L A ALICE AND H LIANG 1999 Phy-logeny of Poaceae inferred from matK sequencesAnn Mo Bot Gard 86835ndash851
currenHILU K W AND A ESEN 1992Prolamin andimmuno-logical strudies in the Poaceae III Subfamily Chlori-doids Am J Bot 80104ndash113
currenHILU K W AND H LIANG 1997 The matK gene Se-quence variation and application in plant systematicsAm J Bot 84830ndash839
currenHILU K W AND K WRIGHT 1982 Systematics ofGramineae A cluster analysis study Taxon 319ndash37
currenHSIAO C N J CHATTERTON K H ASAY AND K BJENS EN 1993 Phylogenetic relationships of 10 grassspecies An assessment of phylogenetic utility of theinternal transcribed spacer region in nuclear riboso-mal DNA in monocots Genome 37112ndash120
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1994 Phylogenetic relationships of themonogenomic species of the wheat tribe Triticeae(Poaceae) inferred from nuclear rDNA (internal tran-scribed spacer) sequences Genome 38211ndash223
currenHSIAO C N J CHATTERTON K H ASAY ANDK B JENSEN 1995Molecular phylogeny of the pooids(Poaceae) based on nuclear rDNA (ITS) sequencesTheor Appl Genet 90389ndash398
currenHSIAO C S W JACOBS N P BARKER AND N JCHATTERTON 1998 A molecular phylogeny of the
subfamily arundinoids (Poaceae) based on sequencesof rDNA Aust Syst Bot 1141ndash52
currenHSIAO C S W L JACOBS N J CHATTERTON AND KH ASAY 1999 A molecular phylogeny of the grassfamily (Poaceae) based on the sequences of nuclearribosomal DNA (ITS) Aust Syst Bot 11667ndash688
JOHNS ON L A AND D E SOLTIS 1998 Assessing con-gruence Empirical examples from molecular dataPages 297ndash348 in Molecular systematics of plants IIDNA sequencing (D E Soltis P S Soltis and J JDoyle eds) Kluwer Academic Publishers NorwellMassachusetts
currenKATAYAMA H AND Y OGIHARA 1996 Phylogeneticafnities of the grasses to other monocots as re-vealed by molecular analysis of chloroplast DNACurr Genet 29572ndash581
currenKELCHNER S A AND L G CLARK 1997 Molecu-lar evolution and phylogenetic utility of the chloro-plast rpl16 intron in Chusquea and the bambusoids(Poaceae) Mol Phylogenet Evol 8385ndash397
currenKELLOGG E A 1989 Comments on genomic genera inthe Triticeae (Poaceae) Am J Bot 76796ndash805
currenKELLOGG E A 1992Tools for studying the chloroplastgenome in the Triticeae (Gramineae) An ecoRI mapa diagnostic deletion and support for Bromus as anoutgroup Am J Bot 79186ndash197
currenKELLOGG E A 1998aRelationships of cereal crops andother grassesProc NatlAcad Sci USA952005ndash2010
currenKELLOGG E A 1998bWhorsquos related to whom Recentresults from molecular systematic studies Curr OpinPlant Biol 1149ndash158
currenKELLOGG E A AND R APPELS 1995 Intraspecic andinterspecic variation in 5S RNA genes are decoupledin diploid wheat relatives Genetics 140325ndash343
currenKELLOGG E A AND L WATSON 1993 Phyloge-netic studies of a large data set I bambusoids An-dropogonodae and pooids (Gramineae) Bot Rev59273ndash311
KIM J 1996 General inconsistency conditions for max-imum parsimony Effects of branch lengths and in-creasing numbers of taxa Syst Biol 45363ndash374
KISHINO H AND M HASEGAWA 1989 Evaluation ofthe maximum likelihood estimate of the evolution-ary tree topologies form DNA sequence data and thebranching order in Hominoidea J Mol Evol 29170ndash179
currenLIANG H AND K W HILU 1995 Application of thematK gene sequence to grass systematics Can J Bot74125ndash134
MABBERLEY D J 1993 The plant-book A portable dic-tionary of the vascular plants 2nd edition CambridgeUniv Press Cambridge
currenMACFARLANE T D AND L WATSON 1982The classi-cation of Poaceaesubfamily pooids Taxon31178ndash203
currenMASON-GAMER R J AND EA KELLOGG 1996Testingfor phylogenetic conict among molecular data sets inthe tribe Triticeae (Gramineae) Syst Biol 45524ndash545
currenMASON-GAMER R J C F WEIL AND E A KELLOGG1998Granule-bound starch synthase Structure func-tion andphylogenetic utility Mol Biol Evol 151658ndash1673
currenMATHEWS S AND R A SHARROCK 1996 The phy-tochrome gene family in grasses (Poaceae) A phy-logeny and evidence that grasses have a subset ofthe loci found in dicot angiosperms Mol Biol Evol131141ndash1150
currenMATHEWS S R C TSAL AND E A KELLOGG 2000Phylogenetic structure in the grass family (Poaceae)
150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel

150 SYSTEMATIC BIOLOGY VOL 51
Evidence from the nuclear gene phytochrome B AmJ Bot 8796ndash107
currenMONTE J V C L MCINTYRE AND J P GUSTAFSON1993 Analysis of phylogenetic relationships in theTriticeae tribe using RFLPs Theor Appl Genet86649ndash655
currenMORTON B R AND M T CLEGG 1993 A chloro-plast DNA mutation hotspot and gene conversion in anoncoding region near rbcL in the grass family(Poaceae) Curr Genet 24357ndash365
currenNADOT S R BAJON AND B LEJEUNE 1994 The chloro-plast gene rps4 as a tool for the study of Poaceaephylogeny Plant Syst Evol 19127ndash38
currenNADOT S G BITTAR L CARTER R LACROIX ANDB LEJEUNE 1995 A phylogenetic analysis of Mono-cotyledons based on the chloroplast gene rps4 usingparsimony and a new numerical phenetics methodMol Phylogenet Evol 4257ndash282
PAGEL M 1999 Inferring the historical patterns of bio-logical evolution Nature 401877ndash884
currenPETERSEN G AND O SEBERG 1997 Phylogenetic anal-ysis of the Triticeae (Poaceae) based on rpoA sequencedata Mol Phylogenet Evol 7217ndash230
PURVIS A 1995a A modication to Baum and Raganrsquosmethod for combining phylogenetic trees Syst Biol44251ndash255
PURVIS A 1995bA composite estimate of primate phy-logeny Philos Trans R Soc London B 348405ndash421
PURVIS A J J GITTLEMAN AND H K LUH 1994 Truthor consequencesmdashndashEffects of phylogenetic accuracyon two comparative methods J Theor Biol 167293ndash300
PURVIS A AND D L J QUICKE 1997a Building phylo-genies Are the big easy Trends Ecol Evol 1249ndash50
PURVIS A AND D L J QUICKE 1997b Are the big in-deed easy Reply from A Purvis and D L J QuickeCorrespondence Trends Ecol Evol 12 357ndash358
RAGAN M A 1992 Phylogenetic inference based onmatrix representation of trees Mol Phylogenet Evol153ndash58
ROBINS ON D F AND L R FOULDS 1981 Comparisonof phylogenetic trees Math Biol 59131ndash144
RODRIGO A G 1993 Calibrating the bootstrap test ofmonophyly Int J Parasitol 23507ndash514
RONQUIST F 1996 Matrix representation of trees re-dundancy and weighting Syst Biol 45247ndash253
SANDERSON M J A PURVIS AND C HENZE 1998Phylogenetic supertrees Assembling the trees of lifeTrends Ecol Evol 13105ndash110
SANDERSON M J AND M F WOJCIECHOWSKI 2000Improved bootstrap condence limits in large-scalephylogenies with an example from Neo-Astragalus(leguminosae) Syst Biol 49671ndash685
SAVOLAINEN V M W CHASE S B HOOT C MMORTON D E SOLTIS C BAYER M F FAY A Y DEBRUIJN S SULLIVAN AND Y-LQIU 2000Phylogenet-ics of owering plants based on combined analysis
of plastid atpB and rbcL gene sequences Syst Biol49306ndash362
SHIMODAIRA H AND M HASEGAWA 1999 Multi-ple comparisons of log-likelihoods with applicationsto phylogenetic inference Mol Biol Evol 161114ndash1116
SOLTIS D E P S SOLTIS M W CHASE M E MORTD C ALBACH M ZANIS V SAVOLAINEN W HHAHN S B HOOT M F FAY M AXTELL S MSWENSEN L M PRINCE W J KRESS K C NIXONAND J S FARRIS 2000 Angiosperm phylogeny in-ferred from 18S rDNA rbcL and atpB sequences BotJ Linn Soc 133381ndash461
SOLTIS D E P S SOLTIS M E MORT M W CHASEV SAVOLAINEN SB HOOT AND CMMORTON 1999Inferring complex phylogenies using parsimony Anempirical approach using three large DNA data setsfor angiosperms Syst Biol 4732ndash42
currenSORENG R J J I DAVIS AND J J DOYLE 1990 A phy-logenetic analysis of chloroplast DNA restriction sitevariation in Poaceae subfam Pooids Plant Syst Evol17283ndash97
STEEL M 1992 The complexity of reconstructing treesfrom qualitative characters and subtrees J Classif991ndash116
STEEL M A W M DRESS AND S BOCKER 2000 Simplebut fundamental limitations on supertree and consen-sus tree methods Syst Biol 49363ndash368
SWOFFORD D L 1998PAUPcurren Phylogenetic analysis us-ing parsimony (currenand other methods) Version 4 Sin-auer Associates Sunderland Massachusetts
currenVAN DEN BORRE A AND L WATSON 1997 On theclassication of the chloridoids (Poaceae) Aust SystBot 10491ndash531
currenVERBOOM G A H P LINDER AND N P BARKER1994Haustorial synergids An important character inthe systematics of danthonioid grasses (arundinoidsPoaceae) Am J Bot 811601ndash1610
currenWATANABE M M ITO AND S KURITA 1994 Chloro-plast DNA phylogeny of Asian bamboos (bambu-soids Poaceae) and its systematic implication J PlantRes 107253ndash261
currenWATSON L HT CLIFFORD AND MJ DALLWITZ 1985The classication of Poaceae Subfamilies and super-tribes Aust J Bot 33433ndash483
YANG Z AND N GOLDMAN 1997 Are the big indeedeasy Correspondance Trends Ecol Evol 12357ndash358
currenZHANG W 2000 Phylogeny of the grass family(Poaceae) from rpl16 intron sequence data Mol Phy-logenet Evol 15135ndash146
ZHARKIKH A AND W-H LI 1995 Estimation ofcondence in phylogeny The complete-and-partialbootstrap technique Mol Phylogenet Evol 444ndash63
Received 2 March 2001 accepted 26 September 2001Associate Editor Mike Steel