biosynthesis of polyhydroxyalkanoates containing 2-hydroxybutyrate from unrelated carbon source by...
TRANSCRIPT

APPLIED GENETICS AND MOLECULAR BIOTECHNOLOGY
Biosynthesis of polyhydroxyalkanoates containing2-hydroxybutyrate from unrelated carbon sourceby metabolically engineered Escherichia coli
Si Jae Park & Tae Woo Lee & Sung-Chul Lim &
Tae Wan Kim & Hyuk Lee & Min Kyung Kim &
Seung Hwan Lee & Bong Keun Song & Sang Yup Lee
Received: 19 June 2011 /Revised: 24 July 2011 /Accepted: 3 August 2011 /Published online: 14 August 2011# Springer-Verlag 2011
Abstract We have previously reported in vivo biosynthesis ofpolylactic acid (PLA) and poly(3-hydroxybutyrate-co-lactate)[P(3HB-co-LA)] employing metabolically engineered Escher-ichia coli strains by the introduction of evolved Clostridiumpropionicum propionyl-CoA transferase (PctCp) and Pseudo-monas sp. MBEL 6-19 polyhydroxyalkanoate (PHA) synthase1 (PhaC1Ps6-19). Using this in vivo PLA biosynthesis system,we presently report the biosynthesis of PHAs containing 2-hydroxybutyrate (2HB) monomer by direct fermentation of ametabolically engineered E. coli strain. The recombinant E.coli ldhA mutant XLdh strain expressing PhaC1Ps6-19 andPctCp was developed and cultured in a chemically definedmedium containing 20 g/L of glucose and varying concen-trations of 2HB and 3HB. PHAs consisting of 2HB, 3HB, anda small fraction of lactate were synthesized. Their monomer
compositions were dependent on the concentrations of 2HBand 3HB added to the culture medium. Even though the ldhAgene was completely deleted in the chromosome of E. coli, upto 6 mol% of lactate was found to be incorporated into thepolymer depending on the culture condition. In order tosynthesize PHAs containing 2HB monomer without feeding2HB into the culture medium, a heterologous metabolicpathway for the generation of 2HB from glucose wasconstructed via the citramalate pathway, in which 2-ketobutyrate is synthesized directly from pyruvate andacetyl-CoA. Introduction of the Lactococcus lactis subsp.lactis Il1403 2HB dehydrogenase gene (panE) into E. coliallowed in vivo conversion of 2-ketobutyrate to 2HB. Themetabolically engineered E. coli XLdh strain expressing thephaC1437, pct540, cimA3.7, and leuBCD genes together with
S. J. Park (*) : S. H. Lee : B. K. SongChemical Biotechnology Research Center, Korea ResearchInstitute of Chemical Technology,P.O. Box 107, Sinseongno 19, Yuseong-gu,Daejeon 305-600, Republic of Koreae-mail: [email protected]
T. W. Lee :M. K. Kim : S. Y. Lee (*)Metabolic and Biomolecular Engineering National ResearchLaboratory, Department of Chemical and BiomolecularEngineering (BK21 Program), Center for Systems and SyntheticBiotechnology, and Institute for the BioCentury,KAIST, 291 Daehak-ro, Yuseong-gu,Daejeon 305-701, Republic of Koreae-mail: [email protected]
S. Y. LeeDepartment of Bio and Brain Engineering, Departmentof Biological Sciences, BioProcess Engineering Research Centerand Bioinformatics Research Center,KAIST, 291 Daehak-ro, Yuseong-gu,Daejeon 305-701, Republic of Korea
S.-C. LimCorporate R&D, LG Chem Research Park,104-1 Moonji-dong, Yuseong-gu,Daejeon 305-380, Republic of Korea
T. W. KimMarine Biotechnology Research Center, Korea Ocean Research& Development Institute,PO Box 29, Ansan 425-600, Republic of Korea
H. LeeMedicinal Chemistry Research Center, Korea Research Instituteof Chemical Technology,P.O.Box 107, Sinseongno 19, Yuseong-gu,Daejeon 305-600, Republic of Korea
Appl Microbiol Biotechnol (2012) 93:273–283DOI 10.1007/s00253-011-3530-x

the L. lactis Il1403 panE gene successfully produced PHAsconsisting of 2HB, 3HB, and a small fraction of lactate byvarying the 3HB concentration in the culture medium. As the3HB concentration in the medium increased the 3HBmonomerfraction in the polymer, the polymer content increased. WhenRalstonia eutropha phaAB genes were additionally expressedin this recombinant E. coli XLdh strain, P(2HB-co-3HB-co-LA) having small amounts of 2HB and LA monomers couldalso be produced from glucose as a sole carbon source. Themetabolic engineering strategy reported here should be usefulfor the production of PHAs containing 2HB monomer.
Keywords Polyhydroxyalkanoate (PHA) . 2-hydroxybutyrate(2HB) . PHA synthase . Recombinant E. coli
Introduction
Polyhydroxyalkanoates (PHAs) are biodegradable and bio-compatible polyesters consisting of various hydroxycarboxylicacids, which accumulate in many bacteria as carbon (energy)and/or reducing power storage material (Lee 1996; Madisonand Huisman 1999). PHAs are polymerized by PHAsynthase, which uses various hydroxyacyl-CoAs as itssubstrates. If hydroxyacyl-CoAs contain an asymmetric centerat the carbon position of hydroxyl group, they are all in the(R)-configuration. Among various hydroxyacyl-CoAs, naturalPHA synthases generally accept 3-hydroxyacyl-CoAs (3HA-CoAs) as the most favorable substrates, but 4-, 5-, and 6-hydroxyacyl-CoAs can also be accepted (Lee 1996; Madisonand Huisman 1999). However, there has been no report on thein vivo biosynthesis of PHAs containing 2-hydroxyacid asmonomer by natural PHA-producing bacteria. Natural PHAsynthases screened to date have shown very low or negligiblein vitro activities towards 2-hydroxyacyl-CoAs comparedwith other HA-CoAs (Yuan et al. 2001; Zhang et al. 2001).Recently, we developed metabolically engineered E. colistrains capable of producing polylactic acid (PLA) and itscopolymers consisting of lactate and other monomers includ-ing 3-hydroxypropionate (3HP), 3-hydroxybutyrate (3HB), 3-hydroxyvalerate (3HV), 3-hydroxyhexanoate (3HHx), 3-hydroxyoctanoate (3HO), 3-hydroxydecanoate (3HD), 3-hydroxydodecanoate (3HDD), and 4-hydroxybutyrate (4HB)by a direct fermentation (Jung et al. 2010; Jung and Lee 2011;Park et al. 2008a, b; Yang et al. 2010, 2011). In thismetabolically engineered E. coli strain, PLA is synthesizedby in vivo generation of (D)-lactyl-CoA followed by itspolymerization. We have demonstrated that (D)-lactyl-CoAcan be synthesized in recombinant E. coli by employing theengineered Clostridium propionicum propionyl-CoA transfer-ase (PctCp) that converts (D)-lactate to (D)-lactyl-CoA usingacetyl-CoA as a CoA donor. For the polymerization of (D)-lactyl-CoA to (D)-PLA, Pseudomonas sp. 6-19 PHA synthase
(PhaC1Ps6-19) was engineered to efficiently utilize (D)-lactyl-CoA as a substrate because natural PHA synthases havenegligible activity towards (D)-lactyl-CoA (Park et al. 2008a,b; Yang et al. 2010; Jung et al. 2010). Also, the efficiency ofPLA synthesis could be enhanced by genome engineering ofE. coli XL1-Blue to strengthen (D)-lactyl-CoA biosynthesisflux based on the systems-level metabolic pathway analysisand by enhancing the PctCp activity by random mutagenesis(Jung et al. 2010; Yang et al. 2010). In a different study,Taguchi and colleagues also reported the biosynthesis of poly(3-hydroxybutyrate-co-lactate) [P(3HB-co-LA)] using a simi-lar system (Shozui et al. 2010; Taguchi et al. 2008; Yamada etal. 2009, 2010, 2011). As the engineered PhaC1Ps6-19 has amuch broad substrate specificity and lactate is also themember of 2-hydroxyacid, it was reasoned that other 2-hydroxyacyl-CoAs might also be used as substrates forPhaC1Ps6-19 resulting in the biosynthesis of novel PHAscontaining 2-hydroxyacid monomer. Because 2-hydroxybutyrate (2HB) can be derived from the amino acidbiosynthetic pathway, it was thought that 2HB might not havetoxic effects to E. coli. Thus, 2HB was examined as a proof-of-concept monomer among various 2-hydroxyacids for thebiosynthesis of PHAs containing 2-hydroxyacids.
Recently, the citramalate pathway for the production of1-propanol and 1-butanol from glucose, which directlyconverts pyruvate to 2-ketobutyrate, was developed byemploying the evolved Methanococcus jannaschii citrama-late synthase (CimA) (Atsumi and Liao 2008a). Theprecursor for 2HB, 2-ketobutyrate, could be produced fromthis metabolic pathway. Thus, E. coli was metabolicallyengineered to produce 2-hydroxybutyryl-CoA (2HB-CoA),the substrate for PhaC1Ps6-19, by additional expression ofthe 2-hydroxyacid dehydrogenase gene and propionyl-CoAtransferase gene together with the genes corresponding tocitramalate pathway (Fig. 1). Using this system, wepresently report for the first time the microbial biosynthesisand characterization of novel PHA copolymers containing2HB as monomer by metabolically engineered E. coli.
Materials and methods
Bacterial strains and plasmids
All bacterial strains and plasmids used in this study arelisted in Table 1. E. coli XL1-Blue (Stratagene CloningSystems, La Jolla, CA, USA) was used for general genecloning studies. For the production of PHA copolymerscontaining 2HB, a recombinant E. coli ldhA mutant, XLdhstrain, was used as a host strain. Plasmid p619C1437-pct540,which expresses the Pseudomonas sp. 6-19 PHA synthasegene containing quadruple mutations of E130D, S325T,S477G, and Q481K (PhaC1437) and the C. propionicum
274 Appl Microbiol Biotechnol (2012) 93:273–283
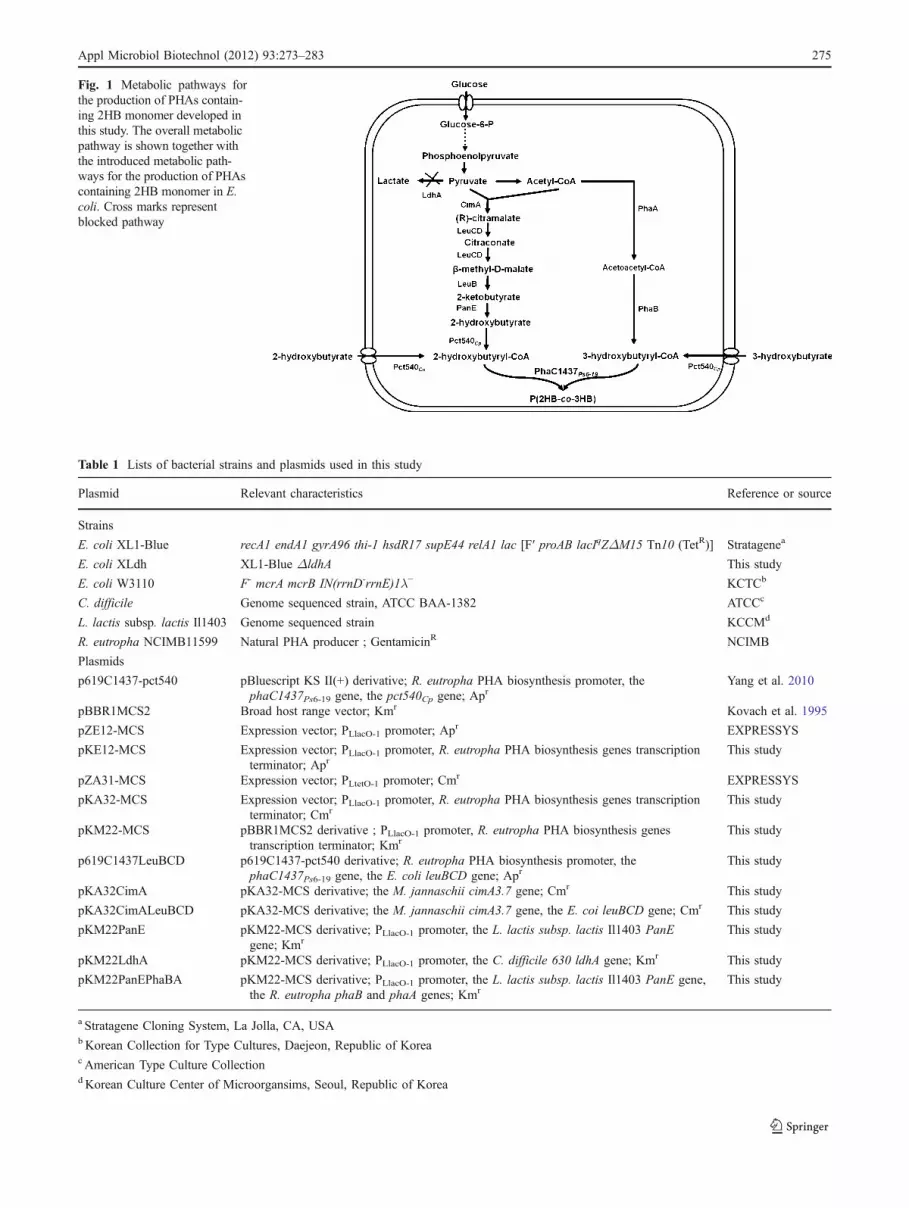
Fig. 1 Metabolic pathways forthe production of PHAs contain-ing 2HB monomer developed inthis study. The overall metabolicpathway is shown together withthe introduced metabolic path-ways for the production of PHAscontaining 2HB monomer in E.coli. Cross marks representblocked pathway
Table 1 Lists of bacterial strains and plasmids used in this study
Plasmid Relevant characteristics Reference or source
Strains
E. coli XL1-Blue recA1 endA1 gyrA96 thi-1 hsdR17 supE44 relA1 lac [F′ proAB lacIqZΔM15 Tn10 (TetR)] Stratagenea
E. coli XLdh XL1-Blue ΔldhA This study
E. coli W3110 F- mcrA mcrB IN(rrnD-rrnE)1λ− KCTCb
C. difficile Genome sequenced strain, ATCC BAA-1382 ATCCc
L. lactis subsp. lactis Il1403 Genome sequenced strain KCCMd
R. eutropha NCIMB11599 Natural PHA producer ; GentamicinR NCIMB
Plasmids
p619C1437-pct540 pBluescript KS II(+) derivative; R. eutropha PHA biosynthesis promoter, thephaC1437Ps6-19 gene, the pct540Cp gene; Ap
rYang et al. 2010
pBBR1MCS2 Broad host range vector; Kmr Kovach et al. 1995
pZE12-MCS Expression vector; PLlacO-1 promoter; Apr EXPRESSYS
pKE12-MCS Expression vector; PLlacO-1 promoter, R. eutropha PHA biosynthesis genes transcriptionterminator; Apr
This study
pZA31-MCS Expression vector; PLtetO-1 promoter; Cmr EXPRESSYS
pKA32-MCS Expression vector; PLlacO-1 promoter, R. eutropha PHA biosynthesis genes transcriptionterminator; Cmr
This study
pKM22-MCS pBBR1MCS2 derivative ; PLlacO-1 promoter, R. eutropha PHA biosynthesis genestranscription terminator; Kmr
This study
p619C1437LeuBCD p619C1437-pct540 derivative; R. eutropha PHA biosynthesis promoter, thephaC1437Ps6-19 gene, the E. coli leuBCD gene; Apr
This study
pKA32CimA pKA32-MCS derivative; the M. jannaschii cimA3.7 gene; Cmr This study
pKA32CimALeuBCD pKA32-MCS derivative; the M. jannaschii cimA3.7 gene, the E. coi leuBCD gene; Cmr This study
pKM22PanE pKM22-MCS derivative; PLlacO-1 promoter, the L. lactis subsp. lactis Il1403 PanEgene; Kmr
This study
pKM22LdhA pKM22-MCS derivative; PLlacO-1 promoter, the C. difficile 630 ldhA gene; Kmr This study
pKM22PanEPhaBA pKM22-MCS derivative; PLlacO-1 promoter, the L. lactis subsp. lactis Il1403 PanE gene,the R. eutropha phaB and phaA genes; Kmr
This study
a Stratagene Cloning System, La Jolla, CA, USAbKorean Collection for Type Cultures, Daejeon, Republic of Koreac American Type Culture Collectiond Korean Culture Center of Microorgansims, Seoul, Republic of Korea
Appl Microbiol Biotechnol (2012) 93:273–283 275

propionyl-CoA transferase mutant gene containing V193Aand four silent nucleotide mutations of T78C, T669C,A1125G, and T1158C (Pct540) under the Ralstonia eutrophaPHA biosynthesis operon promoter, has been previouslydescribed (Yang et al. 2010). Also, the M. jannaschiicimA3.7 gene (Atsumi and Liao 2008a) was synthesizedfrom GenScript (www.genscript.com) based on the reportedsequence. Plasmids pZE12-MCS and pZA31-MCS werepurchased from the EXPRESSYS (www.expressys.com).
Construction of plasmids
All DNA manipulations were performed following standardprocedures (Sambrook and Russell 2001). Polymerasechain reaction (PCR) was performed with the C1000Thermal Cycler (Bio-Rad, Hercules, CA, USA). Primersused in this study (Table 2) were synthesized at Bioneer(Daejeon, Korea). The expression vector, pZE12-MCS,was modified to have multiple-cloning sites (MCS) ofpTacLac (Lee et al. 2008) and the transcription terminatorof the R. eutropha PHA biosynthesis operon. First, theterminator of the R. eutropha PHA biosynthesis operonwas obtained by PCR with the primers 8 and 9 usingp619C1437-pct540 as a template. PCR was performedwith primers 7 and 9 using this gene fragment as atemplate to obtain the gene fragment containing RBS andpTacLac MCS, and the R. eutropha transcription termina-tor. This PCR product was digested with MfeI and AvrIIand was cloned into the EcoRI/AvrII digested pZE12-MCSto obtain pKE12-MCS. Plasmid pKA32-MCS was con-structed by replacing the PLtetO-1 promoter in pKA31-MCSwith the PLlacO-1 promoter obtained from pKE12-MCS byXhoI/EcoRI digestion.
Genes involved in the generation of 2HB-CoA wereamplified from the chromosomal DNAs of E. coli W3110,Clostridium difficile 630 and Lactococcus lactis subsp. lactisIl1403, and from the plasmid p619C1437-pct540. R. eutrophaphaAB genes were amplified from the chromosomal DNAsof R. eutropha NCIMB11599. Plasmid p619C1437LeuBCDwas constructed by replacing the pct540 gene inp619C1437-pct540 with the PCR-amplified E. colileuBCD genes (using the primers 1 and 2) at SbfI andNdeI sites. Plasmid pKA32-CimA was constructed bycloning the synthesized cimA3.7 gene (Atsumi and Liao2008a) into pKA32-MCS at EcoRI and SbfI sites. Then,pKA32-CimALeuBCD was constructed by cloning the E.coli leuBCD genes obtained from p619C1437LeuBCD bydigestion with SbfI and HindIII into pKA32-CimA. Thegene fragment containing PLlacO-1 promoter, MCS, and R.eutropha transcription terminator was amplified frompKA32-MCS using the primers 10 and 11, and then wascloned into the SspI of pBBR1MCS2 to make pKM22-MCS. Plasmids pKM22PanE and pKM22LdhA wereconstructed by cloning the L. lactis subsp. lactis Il1403panE gene and the C. difficile 630 ldhA gene into pKM22-MCS at EcoRI and KpnI sites, respectively. PlasmidpKM22PanEPhaB was constructed by cloning the R.eutropha phaB gene at KpnI and BamHI sites. Then, theR. eutropha phaA gene was cloned into pKM22PanEPhaBat BamHI and PstI sites to make pKM22PanEPhaBA.
Culture condition
E. coli XL1-Blue was cultured at 37°C in lysogeny broth(LB) medium (containing per liter 10 g tryptone, 5 g yeastextract, and 5 g NaCl). For the synthesis of PHAs
Table 2 List of primers used in this study
Primer Primer sequence Target gene
Primer 1 CCTGCAGGTTTCACACAGGAAACA ATGTCGAAGAATTACCATATTG E. coli leuBCD genesPrimer 2 CATATGTTAATTCATAAACGCAGGTTG
Primer 3 GAATTCATGAGAATTACAATTGCCGG L. lactis subsp. lactis Il1403 panE genePrimer 4 GGTACCTTATTTTGCTTTTAATAACTC
Primer 5 GAATTCATGAAAATACTAGTATTTGG C. difficile 630 ldhA genePrimer 6 GGTACCCTAATTTACTCTATTAGTAG
Primer 7 CAATTGATTAAAGAGGAGAAA GAATTCGAGCTCGGTACCCGGGGATCCTCTAGAGTCGACCTGCAGGCATGCAAGCTT
RBS and MCS for pKE12-MCS
Primer 8 GAGTCGACCTGCAGGCATGCAAGCTT CCTGCCGGCCTGGTTCAACC R. eutropha PHA biosynthesis genestranscription terminatorPrimer 9 CCTAGG GCCTCGCCCCCGCGAGGGCC
Primer 10 AATATT AATTGTGAGCGGATAACAAT PLlacO-1 promoter, MCS, and R. eutrophaPHA biosynthesis genes transcriptionterminator for pKM22-MCS
Primer 11 AATATT GCCTCGCCCCCGCGAGGGCC
Primer 12 GGTACCTTTCACACAGGAAACA ATGACTCAGCGCATTGCGTA R. eutropha phaB genePrimer 13 GGATCCTTATCAGCCCATGTGCAGGC CGCCGTTGAGCGAG
Primer 14 GCAGGCCTGCAGGGGGATCC TTCCCTCCCGTTTCCATTGAAAG R. eutropha phaA genePrimer 15 AACTGCAGGTCCACTCCTTGATTGGCTTC
276 Appl Microbiol Biotechnol (2012) 93:273–283

containing 2HB monomer, recombinant E. coli XLdhstrains were cultured in MR medium supplemented with20 g/L of glucose and desired concentrations of 2HB and3HB at 30°C in a rotary shaker at 250 rpm for 72 h. MRmedium (pH 7.0) contains (per liter) 6.67 g KH2PO4, 4 g(NH4)2HPO4, 0.8 g MgSO4·7H2O, 0.8 g citric acid, and5 ml trace metal solution. The trace metal solution contains(per liter of 0.5 M HCl) 10 g FeSO4·7H2O, 2 g CaCl2, 2.2 gZnSO4·7H2O, 0.5 g MnSO4·4H2O, 1 g CuSO4·5H2O, 0.1 g(NH4)6Mo7O24·4H2O, and 0.02 g Na2B4O7·10H2O. Glu-cose, MgSO4·7H2O, 2HB, and 3HB (Acros Organics, Geel,Belgium) were sterilized separately. 2HB and 3HB wereadded into the culture medium to different concentrations.Recombinant E. coli XLdh strain was grown to an opticaldensity (600 nm) of 0.5 before induction with 1 mM ofisopropyl-beta-D-thiogalactopyranoside (IPTG) for theexpression of the cimA, leuBCD, phaAB, and panE genes.Ampicillin (Ap, 50 μg/mL), kanamycin (Km, 30 μg/mL),and chloramphenicol (Cm, 34 μg/mL) were added to themedium depending on the resistance marker of theemployed plasmid.
Genome engineering
Deletion of the ldhA gene in the chromosome of E. coliXL1-Blue to reduce lactate synthesis was carried out usingthe one-step inactivation method as previously reported(Jung et al. 2010; Datsenko and Wanner 2000).
Polymer analysis
The content and monomer composition of the synthesizedpolymer were determined by gas chromatography (GC)(Braunegg et al. 1978). Polymers were purified from thecells by the solvent extraction method (Jacquel et al. 2008).The structure, molecular weight, and thermal properties ofthe polymers were determined by nuclear magnetic reso-nance (NMR) spectroscopy, gel permeation chromatogra-phy (GPC), and differential scanning calorimetry (DSC) aspreviously described (Yang et al. 2010; Jung et al. 2010;Lim et al. 2008).
Results
Biosynthesis of PHAs containing 2HB monomerin recombinant E. coli XLdh by adding2HB and 3HB into the culture medium
We previously reported the production of P(3HB-co-LA) inrecombinant E. coli employing the engineered PhaC1Ps6-19and PctCp (Yang et al. 2010; Jung et al. 2010). As theengineered PhaC1Ps6-19 was able to accept lactate (2-
hydroxypropionate) and exhibited rather broad substratespecificity, it was reasoned that 2HB-CoA can also be usedas its substrate. To facilitate the incorporation of 2HBmonomer into PHA in recombinant E. coli, 3HB wasselected as the second monomer since 3HB-CoA acts as aninitiator for lactate containing polymer synthesis (Jung et al.2010; Taguchi et al. 2008; Yamada et al. 2009; Yang et al.2010). Among the variants of PhaC1Ps6-19 and PctCp,PhaC1437 and Pct540 were chosen because these enzymesefficiently produce P(3HB-co-LA) copolymers with thehighest lactate fraction in recombinant E. coli (Yang et al.2010). It was thought that this could be also the case in theincorporation of 2HB monomer into poly(2-hydroxybuty-rate-co-3-hydroxybutyrate) [(2HB-co-3HB)]. To make P(2HB-co-3HB) without incorporation of lactate monomer,E. coli XLdh strain, in which the E. coli ldhA gene has beencompletely deleted from the chromosome, was constructedand used as host strain for polymer synthesis. RecombinantE. coli XLdh (p619C1437-pct540) was cultured in MRmedium supplemented with 20 g/L of glucose and differentconcentrations of 2HB and 3HB to examine the synthesis ofP(2HB-co-3HB). Recombinant E. coli XLdh expressingPhaC1437 and Pct540 was able to produce PHA randomcopolymers consisting of 2HB, 3HB, and a small fractionof lactate. As shown in Fig. 2, PHAs with different 2HBmonomer fractions, ranging from 10 to 60 mol%, weresynthesized depending on the culture condition. Thehighest 2HB fraction in the copolymer was obtainedby adding 2 g/L of 2HB and 0.5 g/L of 3HB to theculture medium, which resulted in the production of P(61 mol%2HB-co-36 mol%3HB-co-3 mol%LA) withthe PHA content of 26.7 wt.%. Even though the ldhAgene was completely deleted in the chromosome of E.coli, GC analysis revealed that lactate was incorporatedinto the polymer up to 6 mol% depending on the culturecondition. Monomer compositions of the PHA copoly-mer could be modulated by changing the concentrationsof 2HB and 3HB added to the culture medium. The molefraction of 2HB in the copolymer was proportional to theconcentration of 2HB added to the culture medium. Thepolymer content was proportional to the total concen-tration of added 2HB and 3HB, and the highest polymercontent of 68 wt.% was obtained when 2 g/L each of2HB and 3HB were added. Biosynthesis of PHAsconsisting of 2HB and a small amount of LA was alsoexamined by the cultivation of recombinant E. coliXLdh (p619C1437-pct540) in MR medium supple-mented with 20 g/L of glucose and 2 g/L of 2HB.However, PHA was not produced suggesting that aninitiator for polymer synthesis such as 3HB-CoA is stillnecessary for 2HB containing polymer. Thus, 2HB-CoAis also not a favorable substrate for PHA synthase, aswith lactyl-CoA.
Appl Microbiol Biotechnol (2012) 93:273–283 277

Construction of metabolic pathways for the productionof PHAs containing 2HB monomer from glucose
Recent studies reported the production of higher alcoholsvia the keto-acid intermediates of native amino-acid path-ways in microorganisms (Atsumi and Liao 2008a, b;Atsumi et al. 2008; Shen and Liao 2008). Among thevarious recombinant pathways for higher alcohols, the 1-propanol biosynthetic pathway using 2-ketobutyrate as aprecursor is of particular interest because 2-ketobutyrate isalso the possible precursor for 2HB. Between two possiblepathways—the threonine degradation pathway and thecitramalate pathway—the latter was chosen to synthesize2-ketobutyrate because it has been suggested to be the mostdirect route for 2-ketobutyrate (Atsumi and Liao 2008a).The 2-ketobutyrate synthesis pathway was constructed bythe expression of the evolved M. jannaschii cimA3.7 geneand E. coli leuBCD genes as previously described (Atsumiand Liao 2008a). To construct the metabolic pathway forthe synthesis of copolymer containing 2HB monomer fromglucose, 2-ketobutyrate synthesized through the citramalatepathway should be converted into 2HB. There have beenreports on the molecular characterization of (D)-2-hydrox-yacid dehydrogenases from several microorganisms, whichcatalyze the enantioselective reduction of branched chain 2-keto acids to (D)-2-hydroxyacids using NADH as a cofactor(Bernard et al. 1994; Chambellon et al. 2009; Kim et al. 2005).(D)-2-Hydroxyacid dehydrogenase has broad specificities tosubstrates including 2-ketoisocaproate, 2-ketoisovalerate, 2-ketocaproate, 2-ketovalerate, 2-ketobutyrate, and mandelate.Among various (D)-2-hydroxyacid dehydrogenases, two 2-hydroxyacid dehydrogenases encoded by the L. lactis subsp.lactis Il1403 panE gene (Chambellon et al. 2009) and the C.difficile 630 ldhA gene (Kim et al. 2005) were examined for
the synthesis of 2HB. However, the expression of the C.difficile ldhA gene that encodes a putative (D)-2-hydroxyaciddehydrogenase, especially 2-ketoisocaproate dehydrogenasein the L-leucine fermentation pathway of C. difficile (Kim etal. 2005), did not allow production of PHAs containing the2HB monomer in recombinant E. coli. Thus, the conversionof 2-ketobutyrate into 2HB was carried out by expressing theL. lactis Il1403 panE gene encoding the 2HB dehydrogenase.The metabolically engineered E. coli XLdh strain expressingthe phaC1437, pct540, cimA3.7, and leuBCD genes togetherwith the L. lactis Il1403 panE gene successfully producedPHAs consisting of 2HB, 3HB, and a small fraction oflactate from glucose and 3HB added to the culture medium(Fig. 3). This result could be justified by the previous studyshowing that PanE has a substrate specificity toward 2-ketobutyrate by in vitro enzyme activity measurement(Chambellon et al. 2009). To synthesize PHA copolymerscontaining 2HB and 3HB as monomers using this recombi-nant E. coli strain, in which 2HB was endogenously generatedfrom glucose, 3HB was added to the culture medium toprovide 3HB-CoA. As the 3HB concentration increased, boththe 3HB monomer fraction and the polymer content increased(Fig. 3). When this recombinant E. coli XLdh strain wascultured on glucose as a sole carbon source, no PHA wasproduced, suggesting that 2HB-CoA cannot be efficientlyincorporated into PHA (as in the lactyl-CoA case) without aninitiator for PHA synthesis such as 3HB-CoA. Finally, weconstructed a pathway to generate 3HB-CoA from glucose inE. coli by employing β-ketothiolase and acetoacetyl-CoAreductase encoded by R. eutropha phaAB genes. Themetabolically engineered E. coli XLdh strain expressing thephaC1437, pct540, cimA3.7, and leuBCD genes together withthe L. lactis Il1403 panE gene and R. eutropha phaAB genesproduced P(3 mol%2HB-co-96 mol%3HB-co-1 mol%LA)
Fig. 2 Biosynthesis of PHAsconsisting of 2HB, 3HB, and asmall fraction of lactate inrecombinant E. coli XLdh(p619C1437-pct540) by additionof 2HB and 3HB into theculture medium
278 Appl Microbiol Biotechnol (2012) 93:273–283

with the PHA content of up to 74 wt.% from glucose as a solecarbon source.
Characterization of PHAs containing 2HB and 3HBmonomers synthesized in recombinant E. coli
The composition and the sequence distribution of PHAconsisting of 22 mol% 2HB, 76 mol% 3HB, and a smallfraction of lactate (2 mol%) were investigated with 1D (1Hand 13C) NMR and 2D (1H–1H) COSY NMR spectroscopy(Fig. 4a–c). The exact lactate mole fraction in thecopolymer could not be resolved in 1H NMR analysis dueto the low integral value of lactate and peaks overlap oflactate and 2HB, even though the methyl proton of lactatewas observed at 1.6 ppm, and the oxymethine proton oflactate was observed at around 4.9–5.2 ppm. The molefractions of 2HB and 3HB units in the copolymer obtainedfrom 1H NMR were in good agreement with those from GCanalysis except the lactate fraction. The complete assign-ments of 1H NMR spectra in CDCl3 for P(22 mol%2HB-co-76 mol%3HB-co-2 mol%LA) are shown in Fig. 4a. Themethyl protons and methylene protons of 2HB are assignedat the regions of δ0.7–1.2 and 1.6–2.1 ppm, respectively.The methylene protons of 3HB are assigned at the region ofδ2.3–2.9 ppm, while the oxymethine protons of 2HB and3HB are at around δ4.5–5.7 ppm.
Figure 4b shows the COSY spectrum, which highlightsthe intra-three bond coupling of protons, and as a result, thedelicate configurational structure of P(22 mol%2HB-co-76 mol%3HB-co-2 mol%LA). The signal ① between theoxymethine proton of 2HB and the methylene protons of2HB appears at δ4.9 ppm/2.0 ppm. The coupling ②
between the methylene protons of 2HB and the methylprotons of 2HB can be assigned as a cross peak at
δ1.8 ppm/1.0 ppm. The coupling ③ between the oxy-methine proton of 3HB and the methylene protons of3HB can be seen as a cross peak at δ5.4 ppm/2.6 ppm,while the coupling ④ between oxymethine proton of3HB and the remaining methyl protons of 3HB is seenat δ5.2 ppm/1.3 ppm.
Figure 4c shows the 125-MHz 13C-NMR spectrum ofP(22 mol%2HB-co-76 mol%3HB-co-2 mol%LA) with thechemical shift assignments and an expanded spectrum ofcarbonyl carbons region. This region (168.0–170.1 ppm)is clearly resolved into three groups of peaks showingdifferent diad bonding sequences of 2HB and 3HB. Thepeak at 168.7 ppm is assigned to carbonyl carbons in the2HB*–2HB sequence. The peak at 169.1 ppm is assignedto the carbonyl resonance in the 3HB*–3HB and 3HB–3HB* sequences. The peak at 169.5 ppm is assigned to the3HB*–2HB+2HB–3HB* sequences. The diad-sequencedistribution of 2HB and 3HB units was determined fromthe peak areas of carbonyl resonances (Table 3). The2HB–3HB diad-sequence data were compared with theBernoullian statistics applicable to a statistically randomcopolymerization.
DSC and GPC studies on P(47 mol%2HB-co-50 mol%3HB-co-3 mol%LA) showed that amorphous polymer wassynthesized by recombinant E. coli. The molecular weightsof P(47 mol%2HB-co-50 mol%3HB-co-3 mol%LA) were20,000 (Mn) and 33,800 (Mw) with the polydispersity indexof 1.69. The glass transition temperature was 9.7°C, but themelting temperature and the enthalpy of fusion of P(47 mol%2HB-co-50 mol%3HB-co-3 mol%LA) were not detected.
Discussion
PLA is a carbon-neutral alternative to petroleum-based plasticbecause it can be produced from renewable biomass. PLA hasthe potential in a wide range of environmentally friendlyapplications due to its biodegradable, biocompatible, compo-stable properties. To improve the material properties of PLA,copolymerization with a range of other monomers includingglycolide, ε-caprolactone, δ-valerolactone, and trimethylenecarbonate has been employed (Rasal et al. 2010). Also,chemical syntheses of poly(phenyllactate) and polymande-late, which contain aromatic rings as side-chain residues,were performed to provide novel PLA analogs withimproved material properties (Liu et al. 2007; Simmonsand Baker 2001). Recently, poly(2-hydroxybutyrate)[P(2HB)], the analog of PLA in which the methyl group ofPLA is substituted with an ethyl group, was used for thegeneration of P[(S)-2HB]/P[(R)-2HB] stereocomplex toenhance thermal and material properties (Tsuji and Okumura2009). Therefore, development of a microbial system for theproduction of PHA containing 2HB as a monomer should be
Fig. 3 Biosynthesis of PHAs consisting of 2HB, 3HB, and a smallfraction of lactate in recombinant E. coli XLdh (p619C1437-pct540+pKA32CimALeuBCD+pKM22PanE) by addition of 3HB into theculture medium
Appl Microbiol Biotechnol (2012) 93:273–283 279

Fig. 4 NMR analysis (1D 1H,2D COSY, and 1D 13C) of PHAsconsisting of 2HB, 3HB, and asmall fraction of lactate synthe-sized in recombinant E. coliXLdh (p619C1437-pct540). a500 MHz 1H NMR spectrum, b1H–1H COSY spectrum, and c125 MHz 13C NMR spectrumwith the chemical shift assign-ments and an expanded spec-trum of carbonyl carbons region
280 Appl Microbiol Biotechnol (2012) 93:273–283

useful for the synthesis of a polymer with novel materialproperty.
As previously reported, efficient production of PHAs canbe achieved by enhancing the metabolic capacity of hoststrains to supply enough precursors for PHA synthesisalong with the expression of PHA synthase having highsubstrate specificity towards generated substrates (Fukui etal. 1999; Park et al. 2005). Activation of hydroxyacid intohydroxyacyl-CoA by enzymes such as CoA synthetase andCoA transferase has been employed for the production ofPHA from structure-related carbon sources added intoculture medium. PHAs containing lactate, 4HB and 3-hydroxypropionate, and 3-mercaptopropionate as a mono-mer constituent have successfully been synthesized by thisstrategy (Hein et al. 1997; Liu and Steinbüchel 2000;Lütke-Eversloh et al. 2002; Valentin et al 2000; Yang et al2011). At first, recombinant E. coli strains expressingPhaC1437 and Pct540 were cultured in the mediumcontaining 2HB following this strategy to provide enoughprecursors for 2HB-CoA. 3HB was also added to themedium to facilitate the incorporation of 2HB monomerinto PHA as an initiator for 2HB containing polymersynthesis. As shown in Fig. 2, PHAs having different 2HBmonomer compositions were synthesized depending on theconcentration of 2HB and 3HB added to the medium. 2HBmonomer fractions in PHA copolymer were increased up to60 mol% by enhancing the metabolic capacity of E. colistrain to provide more 2HB-CoA rather than 3HB-CoA,which was modulated by increasing the concentration of2HB and reducing the concentration of 3HB added to theculture medium. Because PhaC1437 can also accept lactyl-CoA as a substrate, an E. coli mutant strain in which theldhA gene is completely deleted in the chromosome wasused as a host strain to prevent lactate incorporation intoPHA copolymer. However, lactate was incorporated up to6 mol% in the PHA copolymer depending on the culturecondition. This seemed to be due to the synthesis of lactateby the remaining lactate dehydrogenase activities present inE. coli, despite the complete deletion of the ldhA gene asshown previously (Hong and Lee 2001; Zhang et al. 2009).
It is important to supply precursors for PHA monomersfrom renewable resources to achieve economical produc-tion of PHAs having different monomer composition. Thus,metabolic pathways of E. coli were engineered to provide2HB monomer from glucose for the efficient production of
PHAs containing 2HB monomer. In this study, weemployed the citramalate pathway, the most direct routefor the 2-ketobutyrate precursor of 2HB, by the expressionof the evolved M. jannaschii cimA3.7 gene and E. colileuBCD genes as previously described (Atsumi and Liao2008a). Conversion of 2-ketobutyrate into 2HB is mediatedby (D)-2-hydroxyacid dehydrogenase of the L. lactis subsp.lactis Il1403 (Chambellon et al. 2009). The monomercomposition of PHA copolymer was also dependent on theratio of the precursor concentrations for the 2HB-CoA and3HB-CoA. As the 3HB concentration in the culturemedium increased, the 2HB monomer fraction decreased(Fig. 3), similar to that observed during P(3HB-co-LA)production studies we previously reported ( Jung et al.2010; Yang et al. 2010). It was also found that expression ofthe R. eutropha phaAB genes to supply 3HB-CoA fromglucose significantly reduced the 2HB monomer fraction inthe copolymer, even though a high PHA content of 74 wt.%was achieved. The significant reduction of 2HB monomerfraction in PHA copolymer was likely due to the fact thatexpression of R. eutropha phaAB genes supplying 3HB-CoA from acetyl-CoA accelerated incorporation of 3HB-CoA, the most preferred substrate of PHA synthase, intoPHAs; this is due to that the metabolic pathways forsupplying 2HB, 3HB, and lactate monomer compete foracetyl-CoAs, as shown in Fig. 1. The relatively lower PHAcontent when 2HB was the major monomer suggests thatstrategies for supplying more 2HB monomers from glucoseand improving the substrate specificity towards 2HB-CoAshould be developed to further enhance the production ofPHA-containing 2HB monomer. For example, Atsumi et al.(2008a) reported that removal of IlvI and IlvB, whichcompete for 2-ketobutyrate, resulted in increased produc-tion of 1-propanol and 1-butanol in recombinant E. coli.These strategies will be examined in our future studies.
Very recently, chemo-enzymatic synthesis of 2HB-containing PHA by R. eutropha PHA synthase was reported(Han et al. 2011). Because PhaC1437 used in this study hasthe ability to accept lactyl-CoA as a substrate, PHAcopolymer consisting of only 2HB and 3HB could not besynthesized in recombinant E. coli. Based on this afore-mentioned study, application of R. eutropha PHA synthasein our system should be useful for the production of P(2HB-co-3HB) with a very minute fraction of lactatemonomer for the characterization of polymer properties
Table 3 Diad sequence distri-butions of P(23 mol%2HB-co-77 mol%3HB)
Relative intensity Diad sequence distribution
3HB–3HB (3HB–2HB+2HB–3HB) 2HB–2HB
Observed 57 37 6
Calculated 60 35 5
Appl Microbiol Biotechnol (2012) 93:273–283 281

because R. eutropha PHA synthase cannot efficiently acceptlactyl-CoA as substrate compared with the type II PHAsynthase mutants from Pseudomonas sp. (Taguchi et al.2008; Yang et al. 2010, 2011).
Presently, we report for the first time the biosynthesis ofPHAs containing 2HB monomer from glucose by metabol-ically engineered E. coli strains. PHA copolymers of 2HBand 3HB could be produced by feeding 3HB, which can beeasily generated by the β-ketothiolase and reductase path-ways. With the increase of the fraction of 3HB, the mostfavorable substrate for PHA synthase, the PHA content alsoincreased, reaching a comparable level to that obtained byrecombinant E. coli producing P(3HB) homopolymer. Thestrategies described here should be useful for the develop-ment of new strategies for the production of various PHAsconsisting of other 2-hydroxyacids by direct microbialfermentation from renewable resources.
Acknowledgements This work was supported by the KoreanSystems Biology Research Project (20090065571) of the Ministry ofEducation, Science and Technology (MEST) through the NationalResearch Foundation (NRF) of Korea. Further support by World ClassUniversity program (R32-2008-000-10142-0) and by the AdvancedBiomass R&D Center of Korea (ABC-2010-0029799) through theGlobal Frontier Research Program of MEST is appreciated. B.K.S, S.H.L, and S.J.P appreciate the financial supports from the R&DProgram of MKE/KEIT (10032001) and KRICT.
References
Atsumi S, Hanai T, Liao JC (2008) Non-fermentative pathways forsynthesis of branched-chain higher alcohols as biofuels. Nature451:86–89
Atsumi S, Liao JC (2008a) Directed evolution of Methanococcusjannaschii citramalate synthase for biosynthesis of 1-propanoland 1-butanol by Escherichia coli. Appl Environ Microbiol74:7802–7808
Atsumi S, Liao JC (2008b) Metabolic engineering for advancedbiofuels production from Escherichia coli. Curr Opin Biotechnol19:414–419
Bernard N, Johnsen K, Ferain T, GarmynD,Hols P, Holbrook JJ, DelcourJ (1994) NAD(+)-dependent D-2-hydroxyisocaproate dehydroge-nase of Lactobacillus delbrueckii subsp. bulgaricus. Gene cloningand enzyme characterization. Eur J Biochem 224:439–446
Braunegg G, Sonnleitner B, Lafferty RM (1978) A rapid gas chromato-graphic method for the determination of poly-β-hydroxybutyric acidin microbial biomass. Eur J Appl Microbiol Biotechnol 6:29–37
Chambellon E, Rijnen L, Lorquet F, Gitton C, van Hylckama Vlieg JE,Wouters JA, Yvon M (2009) The D-2-hydroxyacid dehydrogenaseincorrectly annotated PanE is the sole reduction system for branched-chain 2-keto acids in Lactococcus lactis. J Bacteriol 191:873–881
Datsenko KA, Wanner BL (2000) One-step inactivation of chromo-somal genes in Escherichia coli K-12 using PCR products. ProcNatl Acad Sci U S A 97:6640–6645
Fukui T, Yokomizo S, Kobayashi G, Doi Y (1999) Co-expression ofpolyhydroxyalkanoate synthase and (R)-enoyl-CoA hydratasegenes of Aeromonas caviae establishes copolyester biosynthesispathway in Escherichia coli. FEMS Microbiol Lett 170:69–75
Han X, Satoh Y, Satoh T, Matsumoto K, Kakuchi T, Taguchi S, Dairi T,Munekata M, Tajima K (2011) Chemo-enzymatic synthesis ofpolyhydroxyalkanoate (PHA) incorporating 2-hydroxybutyrateby wild-type class I PHAsynthase from Ralstonia eutropha. ApplMicrobiol Biotechnol. doi:10.1007/s00253-011-3362-8
Hein S, Sohling B, Gottschalk G, Steinbüchel A (1997) Biosynthesisof poly(4-hydroxybutyric acid) by recombinant strains ofEscherichia coli. FEMS Microbiol Lett 153:411–418
Hong SH, Lee SY (2001) Metabolic flux analysis for succinic acidproduction by recombinant Escherichia coli with amplified malicenzyme activity. Biotechnol Bioeng 74:89–95
Jacquel N, Lo CW, Wei YH, Wu HS, Wang SS (2008) Isolation andpurification of bacterial poly (3-hydroxyalkanoates). BiochemEng J 39:15–27
Jung YK, Kim TY, Park SJ, Lee SY (2010) Metabolic engineering ofEscherichia coli for the production of polylactic acid and itscopolymers. Biotechnol Bioeng 105:161–171
Jung YK, Lee SY (2011) Efficient production of polylactic acid and itscopolymers by metabolically engineered Escherichia coli. JBiotechnol 151:94–101
Kim J, Darley D, Buckel W (2005) 2-Hydroxyisocaproyl-CoAdehydratase and its activator from Clostridium difficile. FEBS J272:550–561
Kovach ME, Elzer PH, Hill DS, Robertson GT, Farris MA, Roop RMII, Peterson KM (1995) Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 166:175–176
Lee SH, Park SJ, Lee SY, Hong SH (2008) Biosynthesis of enantiopure(S)-3-hydroxybutyric acid in metabolically engineered Escherichiacoli. Appl Microbiol Biotechnol 79:633–641
Lee SY (1996) Bacterial polyhydroxyalkanoates. Biotechnol Bioeng49:1–14
Lim SC, Kim SW, Jung MH, Cho MK, Kim JH, Cho HS, Ok JH(2008) Complete NMR spectral assignments of siloxanol basedcopolycarbonate including the configurational copolymer struc-ture and the determination of each monomer conversion. J MolStruct 886:166–174
Liu SJ, Steinbüchel A (2000) A novel genetically engineered pathwayfor synthesis of poly(hydroxyalkanoic acids) in Escherichia coli.Appl Environ Microbiol 66:739–743
Liu T, Simmons TL, Bohnsack DA, Mackay ME, Smith MR, BakerGL (2007) Synthesis of polymandelide: a degradable polylactidederivative with polystyrene-like properties. Macromolecules40:6040–6047
Lütke-Eversloh T, Fischer A, Remminghorst U, Kawada J,Marchessault RH, Bögershausen A, Kalwei M, Eckert H,Reichelt R, Liu SJ, Steinbüchel A (2002) Biosynthesis ofnovel thermoplastic polythioesters by engineered Escherichiacoli. Nat Mater 1:236–240
Madison LL, Huisman GW (1999) Metabolic engineering of poly(3-hydroxyalkanoates): from DNA to plastic. Microbiol Mol BiolRev 63:21–53
Park SJ, Choi J, Lee SY (2005) Engineering of Escherichia coli fattyacid metabolism for the production of polyhydroxyalkanoates.Enzyme Microb Technol 36:579–588
Park SJ, Yang TH, Kang HO, Lee EJ, Lee SH, Kim TW, Lee SY(2008a) Copolymer containing 3-hydroxyalkanoate unit andlactate unit, and its manufacturing method. WO08/062996
Park SJ, Yang TH, Kang HO, Lee EJ, Lee SH, Kim TW, Lee SY(2008b) Copolymer containing 4-hydroxybutyrate unit andlactate unit, and its manufacturing method. WO08/062995
Rasal RM, Janorkar AV, Hirt DE (2010) Poly(lactic acid) modifica-tions. Prog Polym Sci 35:338–356
Sambrook J, Russell DW (2001) Molecular cloning—a laboratorymanual. 3rd edn. New York: Cold Spring Harbor LaboratoryPress
282 Appl Microbiol Biotechnol (2012) 93:273–283

Shen CR, Liao JC (2008) Metabolic engineering of Escherichia colifor 1-butanol and 1-propanol production via the keto-acidpathways. Metab Eng 10:312–320
Shozui F, Matsumoto K, Nakai T, Yamada M, Taguchi S (2010)Biosynthesis of novel terpolymers poly(lactate-co-3-hydroxybu-tyrate-co-3-hydroxyvalerate)s in lactate-overproducing mutantEscherichia coli JW0885 by feeding propionate as a precursorof 3-hydroxyvalerate. Appl Microbiol Biotechnol 85:949–954
Simmons TL, Baker GL (2001) Poly(phenyllactide): synthesis,characterization, and hydrolytic degradation. Biomacromolecules2:658–663
Taguchi S, Yamada M, Matsumoto K, Tajima K, Satoh Y, Munekata M,Ohno K, Kohda K, Shimamura T, Kambe H, Obata S (2008)A microbial factory for lactate-based polyesters using alactate-polymerizing enzyme. Proc Natl Acad Sci U S A105:17323–17327
Tsuji H, Okumura A (2009) Stereocomplex formation betweenenantiomeric substituted poly(lactide)s: blends of poly[(S)-2-hydroxybutyrate] and poly[(R)-2-hydroxybutyrate]. Macromole-cules 42:7263–7266
Valentin HE, Mitsky TA, Mahadeo DA, Tran M, Gruys KJ (2000)Application of a propionyl coenzyme A synthetase for poly(3-hydroxypropionate-co-3-hydroxybutyrate) accumulation in recom-binant Escherichia coli. Appl Environ Microbiol 66:5253–5258
YamadaM,MatsumotoK, Nakai T, Taguchi S (2009)Microbial productionof lactate-enriched poly[(R)-lactate-co-(R)-3-hydroxybutyrate] withnovel thermal properties. Biomacromolecules 10:677–681
Yamada M, Matsumoto K, Shimizu K, Uramoto S, Nakai T,Shozui F, Taguchi S (2010) Adjustable mutations in lactate
(LA)-polymerizing enzyme for the microbial production ofLA-based polyesters with tailor-made monomer composition.Biomacromolecules 11:815–819
Yamada M, Matsumoto K, Uramoto S, Motohashi R, Abe H, TaguchiS (2011) Lactate fraction dependent mechanical properties ofsemitransparent poly(lactate-co-3-hydroxybutyrate)s produced bycontrol of lactyl-CoA monomer fluxes in recombinant Escher-ichia coli. J Biotechnol. doi:10.1016/j.jbiotec.2011.05.011
Yang TH, Kim TW, Kang HO, Lee SH, Lee EJ, Lim SC, Oh SO, SongAJ, Park SJ, Lee SY (2010) Biosynthesis of polylactic acid andits copolymers using evolved propionate CoA transferase andPHA synthase. Biotechnol Bioeng 105:150–160
Yang TH, Jung YK, Kang HO, Kim TW, Park SJ, Lee SY (2011)Tailor-made type II Pseudomonas PHA synthases and their usefor the biosynthesis of polylactic acid and its copolymer inrecombinant Escherichia coli. Appl Microbiol Biotechnol 90:603–614
Yuan W, Jia Y, Tian J, Snell KD, Muh U, Sinskey AJ, Lambalot RH,Walsh CT, Stubbe J (2001) Class I and III polyhydroxyalkanoatesynthases from Ralstonia eutropha and Allochromatium vinosum:characterization and substrate specificity studies. Arch BiochemBiophys 394:87–98
Zhang S, Kamachi M, Takagi Y, Lenz RW, Goodwin S (2001)Comparative study of the relationship between monomer structureand reactivity for two polyhydroxyalkanoate synthases. ApplMicrobiol Biotechnol 56:131–136
Zhang X, Jantama K, Shanmugam KT, Ingram LO (2009) Reengin-eering Escherichia coli for succinate production in mineral saltsmedium. Appl Environ Microbiol 75:7807–7813
Appl Microbiol Biotechnol (2012) 93:273–283 283