bioavailability in chicks of zinc from zinc propionate
TRANSCRIPT

DESCRIPTION OF PROBLEM
Zinc is an important mineral with many different roles in poultry metabolism. Noted among these are appetite control, regulation of the immune system, and the numerous effects it has on gene expression [1, 2]. In poultry, Zn de-ficiency leads to decreased feed intake and de-creased collagen formation, which in turn leads to lesions on the skin, delayed wound healing, long bone malformation, and poor feathering [3–5].
Zinc bioavailability is hindered by the lev-els of phytate (myoinositol hexaphosphate) in feeds. The zinc cations form strong, irreversible bonds with the phosphate groups in the hexa-phosphate, and poultry and other monogastric species lack natural phytase enzymes to break-down the phytate molecule [6]. This hindrance can be exacerbated even further with increased levels of calcium in the diet [7]. In recent re-search, it was shown that the levels of phytate and calcium in corn-soybean meal diets can in-crease the requirement of dietary Zn up to levels
© 2013 Poultry Science Association, Inc.
Bioavailability in chicks of zinc from zinc propionate
M. A. Brooks,* J. L. Grimes,† K. E. Lloyd,* S. Verissimo,† and J. W. Spears*1
*Department of Animal Science, and †Department of Poultry Science, North Carolina State University, Raleigh 27695-7608
Primary Audience: Nutritionists, Researchers
SUMMARY
The purpose of this experiment was to evaluate the relative bioavailability of Zn from Zn propionate relative to feed-grade Zn sulfate using BW gain and bone Zn as response criteria. For the first 7 d posthatching, chicks were fed a semipurified starter diet deficient in Zn (22 mg of Zn/kg). From d 8 to 21 birds were fed a control (20 mg of Zn/kg) semipurified diet containing ground corn, soy protein concentrate, and dextrose. The control diet was supplemented with 0, 6, or 12 mg of Zn/kg of diet from feed-grade Zn sulfate or organic Zn propionate. Using corn in the diet increased the level of phytate, an important Zn antagonist in nonruminant animals. As supplemental dietary Zn increased, a dose-dependent increase was observed in feed intake, weight gain, total Zn intake, tibia Zn concentration, and total tibia Zn. Zinc supplementation improved FE (feed:gain), but not in a dose-dependent manner. Relative bioavailability of Zn was determined using weight gain, tibia Zn concentration, and total tibia Zn, assuming a value of 100% for feed-grade Zn sulfate. Using relative slope assay, relative bioavailability of Zn pro-pionate was 119, 116, and 116% compared with Zn sulfate, respectively. We concluded, based on these results, that bioavailability of Zn from organic Zn propionate is greater than feed-grade Zn sulfate in the presence of dietary phytate.
Key words: chick, trace mineral, bioavailability, zinc propionate, zinc sulfate
2013 J. Appl. Poult. Res. 22 :153–159 http://dx.doi.org/ 10.3382/japr.2012-00525
1 Corresponding author: [email protected]

154 JAPR: Research Report
that are twice the current NRC recommendation of 40 mg of Zn/kg of diet [1, 8, 9]. In an effort to increase bioavailability and decrease the levels of Zn being excreted in the feces, interest has been growing in the use of organic forms of Zn due to work that has demonstrated an increased bioavailability in the presence of phytate [10–12].
For the current study, diets were formulated using both soy protein concentrate and corn, which contain approximately 1.52 and 1.35% phytate, respectively [13, 14], to assess the the-ory that organic Zn propionate (ZnProp) works in the same manner as other organic Zn sources when in the presence of the Zn antagonist phy-tate. The goal was to evaluate the relative bio-
availability (RB) of Zn, based on growth param-eters and bone Zn concentrations in chicks, from ZnProp compared with feed-grade Zn sulfate (ZnSO4).
MATERIALS AND METHODS
Care, handling, and sampling of birds were approved by the North Carolina State University Animal Care and Use Committee. One-hundred and ten 1-d-old Ross chicks were housed at the North Carolina State University Chicken Educa-tional Unit in Raleigh, NC, and fed a semipuri-fied starter diet deficient in Zn (Table 1) for 7 d posthatching. The diet contained isolated soy as the protein source and was formulated to meet
Table 1. Ingredient composition of the starter (0–7 d) and basal (8–21 d) zinc-deficient diets
Item Starter diet inclusion Basal diet inclusion
Ingredient (%) Ground corn 0.0 38.7 Soy protein concentrate (Arcon-F) 34.5 27.5 Dextrose 52.4 20.0 Corn oil 4.0 4.0 Ground corn-cob 1.8 3.0 Zinc-free mineral mix1 6.5 6.1 Vitamin mix2 0.2 0.2 dl-methionine 0.4 0.3 Choline chloride 0.2 0.2Calculated nutrient analysis ME (kcal/kg) 3,343 3,335 CP (%) 24.3 22.3 Ca (%) 1.27 1.19 Available P (%) 0.62 0.60 Zn (mg/kg) 22.33 20.511Provides per kilogram of diet: 12.6 g of Ca (from CaCO3), 5.2 g of P (from NaH2PO4∙H2O), 2.4 g of K (from KCl), 615 mg of Mg (from MgSO4∙H2O), 80 mg of Mn (from MnSO4∙H2O), 96 mg of Fe (from FeSO4∙H2O), 8.0 mg of Cu (from CuSO4∙5H2O), 0.20 mg of Se (from Na2SeO3), 0.50 mg of I [from Ca(IO3)2], 3.5 mg of Mo (from NaMoO4∙2H2O), 0.70 mg of B (from Na2B4O7∙5H2O).2Provides per kilogram of diet: 26,400 IU of vitamin A; 8,000 IU of vitamin D3; 132 IU of vitamin E; 79.2 μg of vitamin B12; 26.4 mg of riboflavin; 220 mg of niacin; 0.44 mg of d-pantothenate; 0.8 mg of menadione; 4.4 mg of folic acid; 8.0 mg of thiamine; 15.8 mg of pyridoxine; 506 μg of d-biotin; 200 mg of ethoxyquin; 398 mg of wheat middlings.
Table 2. Analyzed Zn concentrations in experimental diets
SourceZn added (mg/kg)
Analyzed Zn (mg/kg)
Realized supplemental Zn
(mg/kg)
None 0 20.51 0.00Zn sulfate1 6 26.93 6.42
12 32.36 11.85Zn propionate2 6 24.33 3.82
12 29.87 9.361Zinc sulfate, anhydrous [16].2KemTRACE brand Zn Organic Trace Mineral Supplement [17].

155BROOKS ET AL.: BIOAVAILABILITY OF ZINC
or exceed NRC [15] requirements for all nutri-ents of the young chick with the exception of Zn (Table 1). This diet was analyzed to contain 22 mg of Zn/kg, whereas the NRC requirement of the chick is 40 to 50 mg of Zn/kg [15].
At the end of the 7-d depletion period, chicks were weighed and 97 of the most uniform chicks (by BW) were randomly assigned to 1 of 5 treat-ments; chicks were then randomly assigned to 1 of 5 replicate pens within treatment (4 chicks per pen, with the exception of 3 pens in the con-trol treatment that contained only 3 chicks). The control diet shown in Table 1 was supplemented with 0, 6, or 12 mg of Zn/kg from feed-grade ZnSO4 [16] or ZnProp [17]. Analyzed con-centrations of Zn in the experimental diets are shown in Table 2. Chicks were housed in heated, thermostatically controlled Petersime batter-ies [18] with raised wire floors. Feed and water were offered ad libitum. Stainless steel feeders and waterers were used to minimize zinc con-tamination.
The control diet differs from the starter diet in that ground corn was added at 38.7% to re-place some of the dextrose and soy protein pres-ent in the semipurified starter diet. Using corn in the diet increased the level of phytate, an im-portant zinc antagonist in nonruminant animals. The control diet analyzed 20.51 mg of Zn/kg.
Experimental diets were fed for 14 d. Indi-vidual BW and feed intake (by pen) were mea-sured at 7-d intervals for determination of gain, feed intake, and FE (feed:gain). At the end of the study, chicks were killed by cervical dislocation and tibia bones were excised and used for Zn determination. Tibias were boiled for 10 min in
deionized H2O to remove all soft tissue, dried at 100°C, then ashed in a Thermolyne muffle fur-nace [19] at 550°C for 24 h. Bone ash was then dissolved in HNO3 and brought up to volume us-ing deionized H2O. Subsamples of experimental diets were taken weekly and composited at the end of the experiment. Diets were prepared for Zn analysis by wet ashing utilizing HNO3. Zinc was measured using atomic absorption spectro-photometry [20].
Data were analyzed as a completely random-ized design using the GLM procedure of SAS [21]. The data were analyzed by treatment with pen as the experimental unit. Zinc bioavailabil-ity was determined using ZnSO4 as a standard source by multiple linear regression and slope-ratio methodology [22]. Analyzed supplemental
Table 3. Effects of Zn level and dietary source on growth performance of 21-d-old chicks1
SourceLevel
(mg/kg)
Supplemental Zn
Intake (g)
Gain (g)
Feed:Gain (g/g)
None 0.00 321c 195c 1.64a
Zn sulfate2 6.42 462b 330ab 1.41b
11.85 518a 364a 1.42b
Zn propionate3 3.82 419b 305b 1.38b
9.36 511a 359a 1.43b
SEM 15 16 0.05a–cMeans with no superscripts in common within the same column are significantly different (P < 0.05).1Mean values are based on 5 replicate pens of 4 chicks per treatment during the 8- to 21-d posthatching period.2Zinc sulfate, anhydrous [16].3KemTRACE brand Zn Organic Trace Mineral Supplement [17].
Table 4. Effects of Zn level and dietary source on bone Zn of 21-d-old chicks1
SourceLevel
(mg/kg)
Supplemental Zn
Tibia Zn (μg/g)
Tibia Zn (μg/bone)
None 0.00 53.8c 18.7c
Zn sulfate2 6.42 75.5b 40.8b
11.85 103.3a 62.2a
Zn propionate3 3.82 70.5b 34.3b
9.36 97.4a 57.6a
SEM 3.6 2.4a–cMeans with no superscripts in common within the same column are significantly different (P < 0.01).1Mean values are based on 5 replicate pens of 4 chicks per treatment during the 8- to 21-d posthatching period.2Zinc sulfate, anhydrous [16].3KemTRACE brand Zn Organic Trace Mineral Supplement [17].

156 JAPR: Research Report
Table 5. Multiple linear regression of weight gain and tibia Zn composition on supplemental dietary Zn concentration (mg/kg)
Item Zn composition Slope ± SE P-value1 RBV2 (%)
Weight gain3 Zn sulfate4 15.29 ± 4.04 — 100Zn propionate5 18.22 ± 5.23 0.42 119
Tibia Zn concentration6 Zn sulfate 7.82 ± 0.65 — 100Zn propionate 9.11 ± 0.86 0.11 116
Total tibia Zn7 Zn sulfate 6.93 ± 0.43 — 100Zn propionate 8.03 ± 0.57 0.04 116
1P-value represents significance of comparison of slopes of the standard (Zn sulfate) versus the corresponding supplemental Zn source.2Relative bioavailability was calculated from the standard-curve regression, setting RBV of Zn in the Zn sulfate standard at 100%.3Multiple linear regression of weight gain (Y) on supplemental Zn from Zn sulfate (X1) and Zn propionate (X2) was Y = 275.56 + 15.29(X1) + 18.22(X2); R2 = 0.82.4Zinc sulfate, anhydrous [16].5KemTRACE brand Zn Organic Trace Mineral Supplement [17].6Multiple linear regression of tibia Zn concentration (Y) on supplemental Zn from Zn sulfate (X1) and Zn propionate (X2) was Y = 54.21 + 7.82(X1) + 9.11(X2); R2 = 0.87.7Multiple linear regression of total tibia Zn (Y) on supplemental Zn from Zn sulfate (X1) and Zn propionate (X2) was Y = 19.84 + 6.93(X1) + 8.03(X2); R2 = 0.93.
Figure 1. Total weight gain of 21-d-old chicks as affected by supplemental dietary Zn from feed-grade Zn sulfate (solid line) and Zn propionate (dashed line). Multiple linear regression of weight gain (Y) on supplemental Zn from Zn sulfate (X1) and Zn propionate (X2) was Y = 275.56 + 15.29(X1) + 18.22(X2); R2 = 0.82 based the 5 replicate pens per treatment level during the 8- to 21-d posthatching period.

157BROOKS ET AL.: BIOAVAILABILITY OF ZINC
Zn values of the experimental diets were used in the regression analysis. All statements of signifi-cance were based on P ≤ 0.05.
RESULTS AND DISCUSSION
Intake and weight gain (Table 3) were not af-fected by Zn source; however, intake and gain were negatively affected (P < 0.01) with de-creasing supplemental Zn level. Feed efficiency (feed:gain; Table 3) was poorer (P < 0.01) in the control diet (0 mg of supplemental Zn/kg) com-pared with all other treatments, but did not differ between the Zn-supplemented diets regardless of source or level. Previous studies have shown that inadequate dietary Zn decreases appetite [9, 23]; however, the cause of reduced appetite dur-ing Zn deficiency is still elusive [1]. Batal et al. [24] saw no further improvement in intake, gain, or FE when supplemental ZnSO4•H2O exceeded 13.5 mg of Zn/kg in birds fed a similar Zn-de-
ficient soy concentrate-dextrose-based diet. Our results are similar to those of Bao et al. [25], who reported that chicks will sacrifice growth when Zn concentrations in the diet are deficient.
To properly assess Zn bioavailability, low levels of supplemental Zn are used to prevent the response tissue, in this case bone, from reaching maximal concentrations; this assesses Zn status during a state of rapid accumulation. Further, the addition of corn to the soy protein concentrate-dextrose diet increases dietary phy-tate, which more closely simulates conventional diets than a purified test diet [11, 26]. Edwards and Baker [26] determined that the phytate con-tent of soy concentrate proteins can reduce the RB of dietary Zn from soy concentrate to 65% when compared with ZnSO4. Moreover, when inorganic Zn sources complex with dietary phy-tate, their absorption efficiency can be reduced by 40%; whereas, organic chelates have been shown to be less affected by phytate [27].
Figure 2. Tibia Zn concentration of 21-d-old chicks as affected by supplemental dietary Zn from feed-grade Zn sulfate (solid line) and Zn propionate (dashed line). Multiple linear regression of tibia Zn concentration (Y) on supplemental Zn from Zn sulfate (X1) and Zn propionate (X2) was Y = 54.21 + 7.82(X1) + 9.11(X2); R2 = 0.87 based the 5 replicate pens per treatment level during the 8- to 21-d posthatching period.
158 JAPR: Research Report
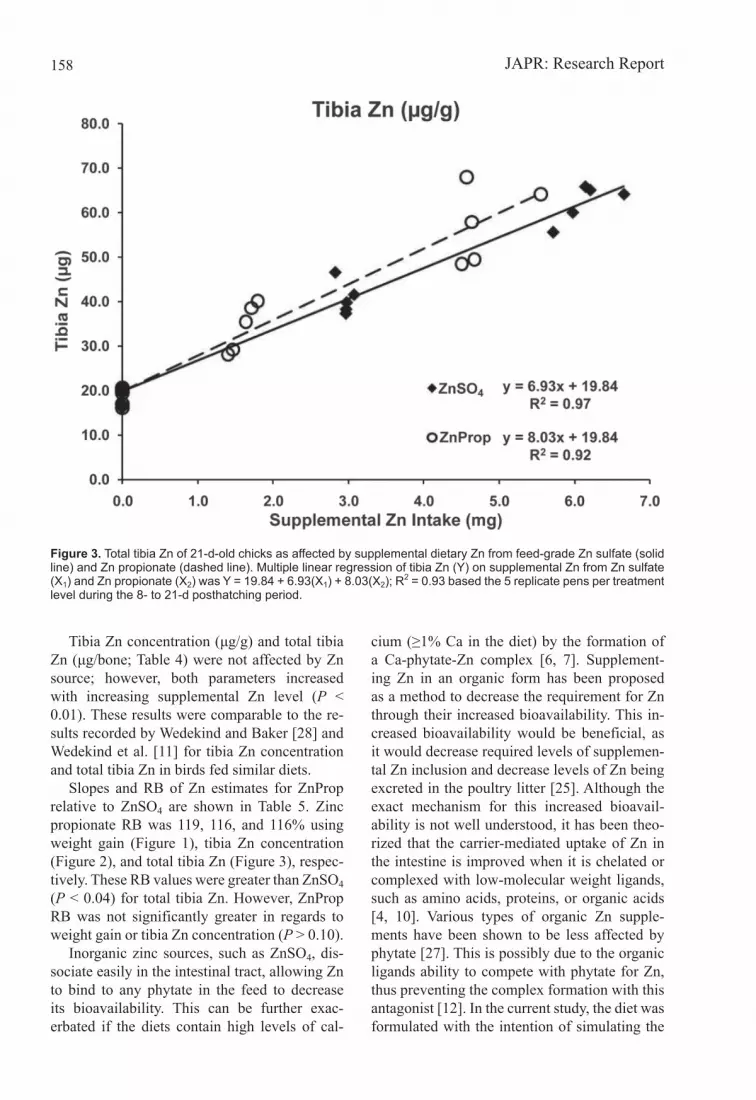
Tibia Zn concentration (μg/g) and total tibia Zn (μg/bone; Table 4) were not affected by Zn source; however, both parameters increased with increasing supplemental Zn level (P < 0.01). These results were comparable to the re-sults recorded by Wedekind and Baker [28] and Wedekind et al. [11] for tibia Zn concentration and total tibia Zn in birds fed similar diets.
Slopes and RB of Zn estimates for ZnProp relative to ZnSO4 are shown in Table 5. Zinc propionate RB was 119, 116, and 116% using weight gain (Figure 1), tibia Zn concentration (Figure 2), and total tibia Zn (Figure 3), respec-tively. These RB values were greater than ZnSO4 (P < 0.04) for total tibia Zn. However, ZnProp RB was not significantly greater in regards to weight gain or tibia Zn concentration (P > 0.10).
Inorganic zinc sources, such as ZnSO4, dis-sociate easily in the intestinal tract, allowing Zn to bind to any phytate in the feed to decrease its bioavailability. This can be further exac-erbated if the diets contain high levels of cal-
cium (≥1% Ca in the diet) by the formation of a Ca-phytate-Zn complex [6, 7]. Supplement-ing Zn in an organic form has been proposed as a method to decrease the requirement for Zn through their increased bioavailability. This in-creased bioavailability would be beneficial, as it would decrease required levels of supplemen-tal Zn inclusion and decrease levels of Zn being excreted in the poultry litter [25]. Although the exact mechanism for this increased bioavail-ability is not well understood, it has been theo-rized that the carrier-mediated uptake of Zn in the intestine is improved when it is chelated or complexed with low-molecular weight ligands, such as amino acids, proteins, or organic acids [4, 10]. Various types of organic Zn supple-ments have been shown to be less affected by phytate [27]. This is possibly due to the organic ligands ability to compete with phytate for Zn, thus preventing the complex formation with this antagonist [12]. In the current study, the diet was formulated with the intention of simulating the
Figure 3. Total tibia Zn of 21-d-old chicks as affected by supplemental dietary Zn from feed-grade Zn sulfate (solid line) and Zn propionate (dashed line). Multiple linear regression of tibia Zn (Y) on supplemental Zn from Zn sulfate (X1) and Zn propionate (X2) was Y = 19.84 + 6.93(X1) + 8.03(X2); R2 = 0.93 based the 5 replicate pens per treatment level during the 8- to 21-d posthatching period.

159BROOKS ET AL.: BIOAVAILABILITY OF ZINC
levels of phytate in a typical soy-corn-based diet to see if, when challenged with this antagonist, a difference exists between inorganic ZnSO4 and organic ZnProp. We concluded that the Zn from ZnProp is more bioavailable than from ZnSO4 when added at physiological concentrations to a soy-corn-based diet containing phytate.
CONCLUSIONS AND APPLICATIONS
1. Supplementation of Zn into a diet con-taining deficient levels of Zn (≤22 mg of Zn/kg of diet) will increase feed intake, weight gain, tibia Zn concentration, and improve FE.
2. When fed a corn-based diet containing the dietary antagonist phytate, RB of Zn from ZnProp was 116% of the standard feed-grade ZnSO4 source tested.
3. Increased bioavailability in a practical setting would allow for lesser inclusion of the mineral in the diet while providing similar concentrations of bioavailable Zn to the animal.
REFERENCES AND NOTES
1. Suttle, N. 2010. The Mineral Nutrition of Livestock. 4th ed. CABI Publishing, Cambridge, MA.
2. Chesters, J. 1997. Zinc. Pages 185–230 in Handbook of Nutritionally Essential Mineral Elements. B. O’Dell, and R. Sunde, ed. Marcel Dekker Inc., New York, NY.
3. Starcher, B. C., C. H. Hill, and J. G. Madaras. 1980. Effect of zinc deficiency on bone collagenase and collagen turnover. J. Nutr. 110:2095–2102.
4. Park, S. Y., S. Birkhold, L. Kubena, D. Nisbet, and S. Ricke. 2004. Review on the role of dietary zinc in poultry nutrition, immunity, and reproduction. Biol. Trace Elem. Res. 101:147–163.
5. Kienholz, E. W., D. E. Turk, M. L. Sunde, and W. G. Hoekstra. 1961. Effects of zinc deficiency in the diets of hens. J. Nutr. 75:211–221.
6. Lönnerdal, B. 2000. Dietary factors influencing zinc absorption. J. Nutr. 130:1378S–1383S.
7. Underwood, E., and N. Suttle. 1999. The Mineral Nu-trition of Livestock. 3rd ed. CABI Publishing, New York, NY.
8. Huang, Y. L., L. Lu, X. G. Luo, and B. Liu. 2007. An optimal dietary zinc level of broiler chicks fed a corn-soybean meal diet. Poult. Sci. 86:2582–2589.
9. Linares, L. B., J. N. Broomhead, E. A. Guaiume, D. R. Ledoux, T. L. Veum, and V. Raboy. 2007. Effects of low phytate barley (Hordeum vulgare L.) on zinc utilization in young broiler chicks. Poult. Sci. 86:299–308.
10. Cao, J., P. R. Henry, R. Guo, R. A. Holwerda, J. P. Toth, R. C. Littell, R. D. Miles, and C. B. Ammerman. 2000. Chemical characteristics and relative bioavailability of sup-plemental organic zinc sources for poultry and ruminants. J. Anim. Sci. 78:2039–2054.
11. Wedekind, K. J., A. E. Hortin, and D. H. Baker. 1992. Methodology for assessing zinc bioavailability: Efficacy es-timates for zinc-methionine, zinc sulfate, and zinc oxide. J. Anim. Sci. 70:178–187.
12. Schlegel, P., and W. Windisch. 2006. Bioavailability of zinc glycinate in comparison with zinc sulphate in the presence of dietary phytate in an animal model with 65zn labelled rats. J. Anim. Physiol. Anim. Nutr. (Berl.) 90:216–222.
13. de Boland, A. R., G. B. Garner, and B. L. O’Dell. 1975. Identification and properties of phytate in cereal grains and oilseed products. J. Agric. Food Chem. 23:1186–1189.
14. O’Dell, B. L., and A. De Boland. 1976. Complexation of phytate with proteins and cations in corn germ and oil seed meals. J. Agric. Food Chem. 24:804–808.
15. NRC. 1994. Nutrient Requirements of Poultry. 9th rev. ed. Natl. Acad. Press, Washington, DC.
16. Zinc sulfate, anhydrous, Eastern Minerals Inc., Hen-derson, NC.
17. KemTRACE brand Zinc Organic Trace Mineral Sup-plement, Kemin Agrifoods North America, Des Moines, IA.
18. Petersime batteries, Petersime Incubator Company, Gettysburg, OH.
19. Thermolyne muffle furnace, Sybron Corp., Dubuque, IA.
20. Model AA-6701F Atomic Absorption spectrometer, Shimadzu, Kyoto, Japan.
21. SAS Institute Inc. 2008. SAS/STAT 9.2 user’s guide. SAS Inst. Inc., Cary, NC.
22. Littell, R. C., A. J. Lewis, and P. R. Henry. 1995. Statistical evaluation of bioavailability assays. Pages 5–33 in Bioavailability of Nutrients for Animals: Amino Acids, Minerals, and Vitamins. C. B. Ammerman, D. H. Baker, and A. J. Lewis, ed. Academic Press Inc., San Diego, CA.
23. Edwards, H. M., 3rd, and D. H. Baker. 1999. Bio-availability of zinc in several sources of zinc oxide, zinc sul-fate, and zinc metal. J. Anim. Sci. 77:2730–2735.
24. Batal, A. B., T. Parr, and D. Baker. 2001. Zinc bio-availability in tetrabasic zinc chloride and the dietary zinc requirement of young chicks fed a soy concentrate diet. Poult. Sci. 80:87–90.
25. Bao, Y. M., M. Choct, P. A. Iji, and K. Bruerton. 2007. Effect of organically complexed copper, iron, manganese, and zinc on broiler performance, mineral excretion, and ac-cumulation in tissues. J. Appl. Poult. Res. 16:448–455.
26. Edwards, H. M., 3rd, and D. H. Baker. 2000. Zinc bioavailability in soybean meal. J. Anim. Sci. 78:1017–1021.
27. Yu, Y., L. Lu, R. L. Wang, L. Xi, X. G. Luo, and B. Liu. 2010. Effects of zinc source and phytate on zinc absorp-tion by in situ ligated intestinal loops of broilers. Poult. Sci. 89:2157–2165.
28. Wedekind, K. J., and D. H. Baker. 1990. Zinc bio-availability in feed-grade sources of zinc. J. Anim. Sci. 68:684–689.