benthic invertebrate fauna, small streams - northern
TRANSCRIPT

Benthic Invertebrate Fauna, Small StreamsJ Bruce Wallace, University of Georgia, Athens, GA, USAS L Eggert, USDA Forest Service, Northern Research Station, Grand Rapids, MN, USA
ã 2009 Elsevier Inc. All rights reserved.
Introduction
Small streams (first- through third-order streams)make up >98% of the total number of stream seg-ments and >86% of stream length in many drainagenetworks. Small streams occur over a wide array ofclimates, geology, and biomes, which influence tem-perature, hydrologic regimes, water chemistry, light,substrate, stream permanence, a basin’s terrestrialplant cover, and food base of a given stream. Smallstreams are generally most abundant in the upperreaches of a basin, but they can also be found through-out the basin and may enter directly into larger rivers.They have maximum interface with the terrestrialenvironment, and in most temperate and tropical cli-mates they may receive large inputs of terrestrial, orallochthonous, organic matter (e.g., leaves, wood)from the surrounding plant communities. In locationswith open canopies such as grasslands and deserts,autochthonous or primary production in the form ofalgae, or higher aquatic plants, may serve as the mainfood base. Hence, headwater streams display a diversefauna, which is often adapted to physical, chemical,and biotic conditions of the region.
Diversity of Benthic Invertebrates inSmall Streams
The benthic invertebrate fauna of small streams is com-posed primarily of aquatic insects, crustaceans, mol-lusks, and various other invertebrate taxa. The insectfauna consists primarily of Odonata (dragonflies anddamselflies), Ephemeroptera (mayflies), Plecoptera(stoneflies), Megaloptera (alderflies and dobsonflies),Coleoptera (beetles), Trichoptera (caddisflies), occa-sional Lepidoptera (moths), and Diptera (true flies).Crustaceans (including amphipods, isopods, and cray-fish) can also be found in small streams as well asmicrocrustaceans such as cladocerans, ostracods, andcopepods. Other common invertebrates found in smallstreams include nematodes, oligochaetes, turbellarians,and mollusks such as snails, limpets, and finger-nailclams. Total invertebrate diversity in small streamscan be quite high. The Breitenbach, a first-order streamin Germany, contains at least 1004 described inverte-brate taxa. At least 293 invertebrate taxa have beenfound in headwater streams in the southern Appala-chianmountains of the United States. Over 182 known
Encyclopedia of Inland Wat
invertebrate taxa have been recorded in a mountainstream on Bougainville Island, Papua New Guinea.Incredibly, there are many headwater invertebrate spe-cies that remain undescribed in both isolated and popu-lated regions of the world.
With the great diversity of foods available for con-sumption by invertebrates (i.e., deposited and retainedon substrates, or suspended in the water column), it isnot surprising that invertebrates have evolved diversemorphobehavioral mechanisms for exploiting foodresources. Their diverse feeding behaviors have beenlumped into a broad functional classification scheme,which is based onmechanisms used by invertebrates toacquire foods. These functional groups are as follows:scrapers, animals adapted to graze or scrape materials(periphyton, or attached algae, fine particulate organicmatter, and its associated microbiota) from mineraland organic substrates; shredders, organisms that com-minute large pieces of decomposing vascular plant tis-sue such as leaf detritus (>1mm diameter) along withits associated microflora and fauna, or feed directly onliving vascular hydrophytes, or gouge decomposingwood; gatherers, animals that feed primarily on depos-ited fine particulate organic matter (FPOM� 1mmdiameter); filterers, animals that have specialized ana-tomical structures (e.g., setae, mouth brushes, or fans)or silk and silk-like secretions that act as sieves toremove particulate matter from suspension; and pre-dators, those organisms that feed primarily on animaltissue by either engulfing their prey, or piercing preyand sucking body contents.
Functional feeding groups refer primarily to modesof feeding and not type of food per se. For example,many filter-feeding insects of high gradient streamsare primarily carnivores, whereas scrapers consumequantities of what must be characterized as epilithon,a matrix of polysaccharide exudates, detritus, micro-flora, and microfauna associated with stone surfaces,and not solely attached algae. Shredders may selectthose leaves that have been ‘microbially conditioned’by colonizing fungi and bacteria. Shredders also ingestattached algal cells, protozoans, and various othercomponents of the fauna during feeding. Some ‘shred-ders’ have been shown to grow by harvesting primar-ily the epixylic biofilm, the matrix of exudates,detritus, microflora, and microfauna found on woodsurfaces. Although it appears valid to separate benthicinvertebrates according to these mechanisms usedto obtain foods, many questions remain concerning
173ers (2009), vol. 2, pp. 173-190
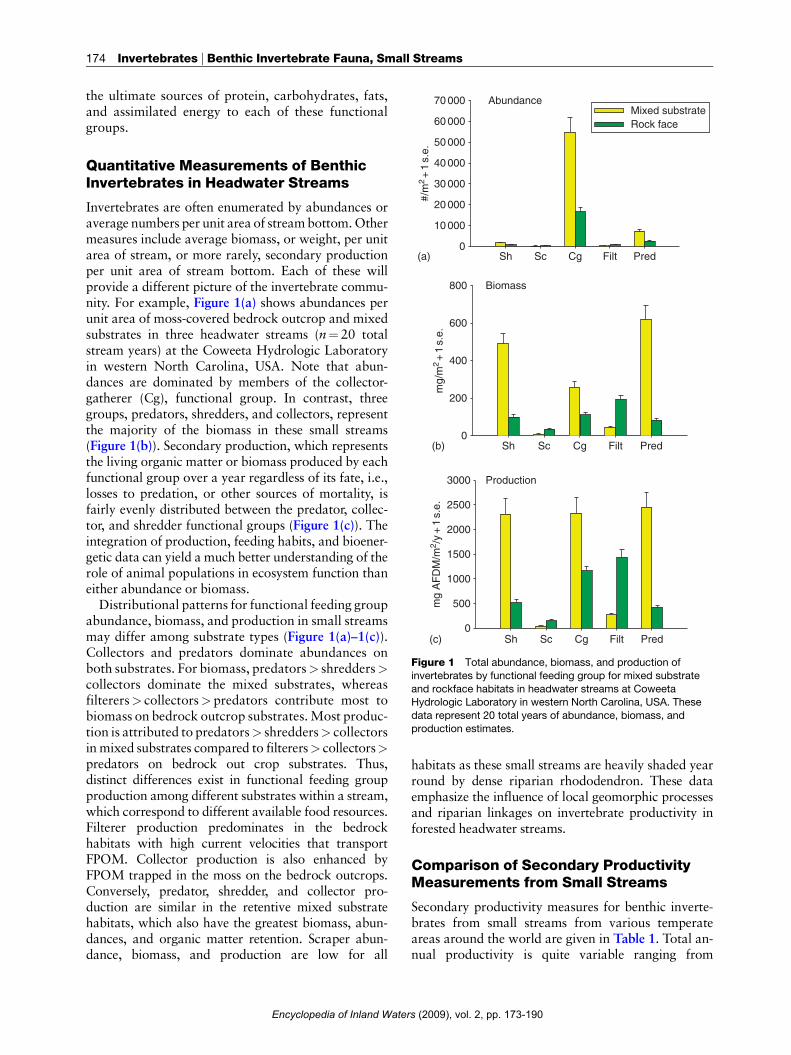
60 000
70 000 AbundanceMixed substrateRock face
174 Invertebrates _ Benthic Invertebrate Fauna, Small Streams
the ultimate sources of protein, carbohydrates, fats,and assimilated energy to each of these functionalgroups.
0
500
1000
1500
2000
2500
3000
0
200
400
600
800
mg
AF
DM
/m2 /
y+
1s.
e.
Sh Sc Cg Filt Pred
Sh Sc Cg Filt Pred
Sh Sc Cg Filt Pred
#/m
2+
1s.
e.m
g/m
2+
1s.
e.
0
10 000
20 000
30 000
40 000
50 000
Biomass
Production
(a)
(b)
(c)
Figure 1 Total abundance, biomass, and production of
invertebrates by functional feeding group for mixed substrate
and rockface habitats in headwater streams at CoweetaHydrologic Laboratory in western North Carolina, USA. These
data represent 20 total years of abundance, biomass, and
production estimates.
Quantitative Measurements of BenthicInvertebrates in Headwater Streams
Invertebrates are often enumerated by abundances oraverage numbers per unit area of streambottom.Othermeasures include average biomass, or weight, per unitarea of stream, or more rarely, secondary productionper unit area of stream bottom. Each of these willprovide a different picture of the invertebrate commu-nity. For example, Figure 1(a) shows abundances perunit area of moss-covered bedrock outcrop and mixedsubstrates in three headwater streams (n¼ 20 totalstream years) at the Coweeta Hydrologic Laboratoryin western North Carolina, USA. Note that abun-dances are dominated by members of the collector-gatherer (Cg), functional group. In contrast, threegroups, predators, shredders, and collectors, representthe majority of the biomass in these small streams(Figure 1(b)). Secondary production, which representsthe living organic matter or biomass produced by eachfunctional group over a year regardless of its fate, i.e.,losses to predation, or other sources of mortality, isfairly evenly distributed between the predator, collec-tor, and shredder functional groups (Figure 1(c)). Theintegration of production, feeding habits, and bioener-getic data can yield a much better understanding of therole of animal populations in ecosystem function thaneither abundance or biomass.Distributional patterns for functional feeding group
abundance, biomass, and production in small streamsmay differ among substrate types (Figure 1(a)–1(c)).Collectors and predators dominate abundances onboth substrates. For biomass, predators> shredders>collectors dominate the mixed substrates, whereasfilterers> collectors> predators contribute most tobiomass on bedrock outcrop substrates.Most produc-tion is attributed to predators> shredders> collectorsin mixed substrates compared to filterers> collectors>predators on bedrock out crop substrates. Thus,distinct differences exist in functional feeding groupproduction among different substrates within a stream,which correspond to different available food resources.Filterer production predominates in the bedrockhabitats with high current velocities that transportFPOM. Collector production is also enhanced byFPOM trapped in the moss on the bedrock outcrops.Conversely, predator, shredder, and collector pro-duction are similar in the retentive mixed substratehabitats, which also have the greatest biomass, abun-dances, and organic matter retention. Scraper abun-dance, biomass, and production are low for all
Encyclopedia of Inland Water
habitats as these small streams are heavily shaded yearround by dense riparian rhododendron. These dataemphasize the influence of local geomorphic processesand riparian linkages on invertebrate productivity inforested headwater streams.
Comparison of Secondary ProductivityMeasurements from Small Streams
Secondary productivity measures for benthic inverte-brates from small streams from various temperateareas around the world are given in Table 1. Total an-nual productivity is quite variable ranging from
s (2009), vol. 2, pp. 173-190
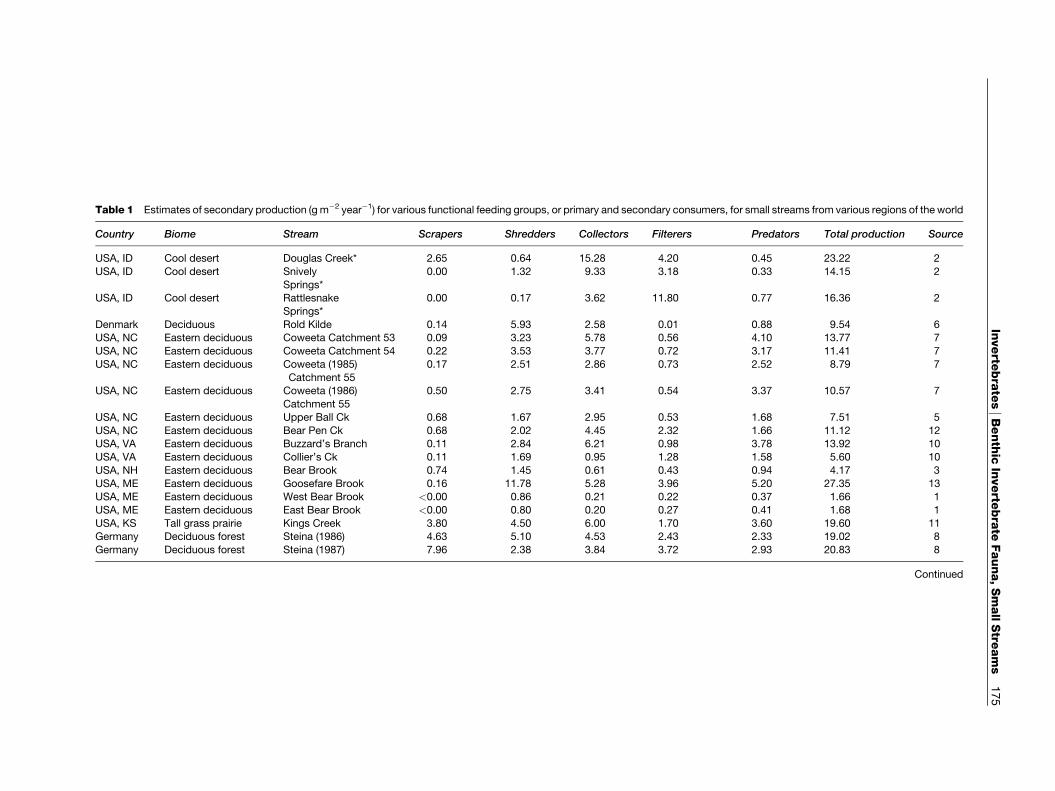
Table 1 Estimates of secondary production (g m�2 year�1) for various functional feeding groups, or primary and secondary consu ers, for small streams from various regions of the world
Country Biome Stream Scrapers Shredders Collectors Filterers Predators Total production Source
USA, ID Cool desert Douglas Creek* 2.65 0.64 15.28 4.20 0.45 23.22 2
USA, ID Cool desert Snively 0.00 1.32 9.33 3.18 0.33 14.15 2
Springs*USA, ID Cool desert Rattlesnake 0.00 0.17 3.62 11.80 0.77 16.36 2
Springs*
Denmark Deciduous Rold Kilde 0.14 5.93 2.58 0.01 0.88 9.54 6USA, NC Eastern deciduous Coweeta Catchment 53 0.09 3.23 5.78 0.56 4.10 13.77 7
USA, NC Eastern deciduous Coweeta Catchment 54 0.22 3.53 3.77 0.72 3.17 11.41 7
USA, NC Eastern deciduous Coweeta (1985) 0.17 2.51 2.86 0.73 2.52 8.79 7
Catchment 55USA, NC Eastern deciduous Coweeta (1986) 0.50 2.75 3.41 0.54 3.37 10.57 7
Catchment 55
USA, NC Eastern deciduous Upper Ball Ck 0.68 1.67 2.95 0.53 1.68 7.51 5
USA, NC Eastern deciduous Bear Pen Ck 0.68 2.02 4.45 2.32 1.66 11.12 12USA, VA Eastern deciduous Buzzard’s Branch 0.11 2.84 6.21 0.98 3.78 13.92 10
USA, VA Eastern deciduous Collier’s Ck 0.11 1.69 0.95 1.28 1.58 5.60 10
USA, NH Eastern deciduous Bear Brook 0.74 1.45 0.61 0.43 0.94 4.17 3
USA, ME Eastern deciduous Goosefare Brook 0.16 11.78 5.28 3.96 5.20 27.35 13USA, ME Eastern deciduous West Bear Brook <0.00 0.86 0.21 0.22 0.37 1.66 1
USA, ME Eastern deciduous East Bear Brook <0.00 0.80 0.20 0.27 0.41 1.68 1
USA, KS Tall grass prairie Kings Creek 3.80 4.50 6.00 1.70 3.60 19.60 11Germany Deciduous forest Steina (1986) 4.63 5.10 4.53 2.43 2.33 19.02 8
Germany Deciduous forest Steina (1987) 7.96 2.38 3.84 3.72 2.93 20.83 8
Continued
Inverte
bra
tes_B
enth
icInverte
bra
teFauna,SmallStre
ams
175
Encyclopedia of Inland Waters (2009), vol. 2, pp. 173-190
m
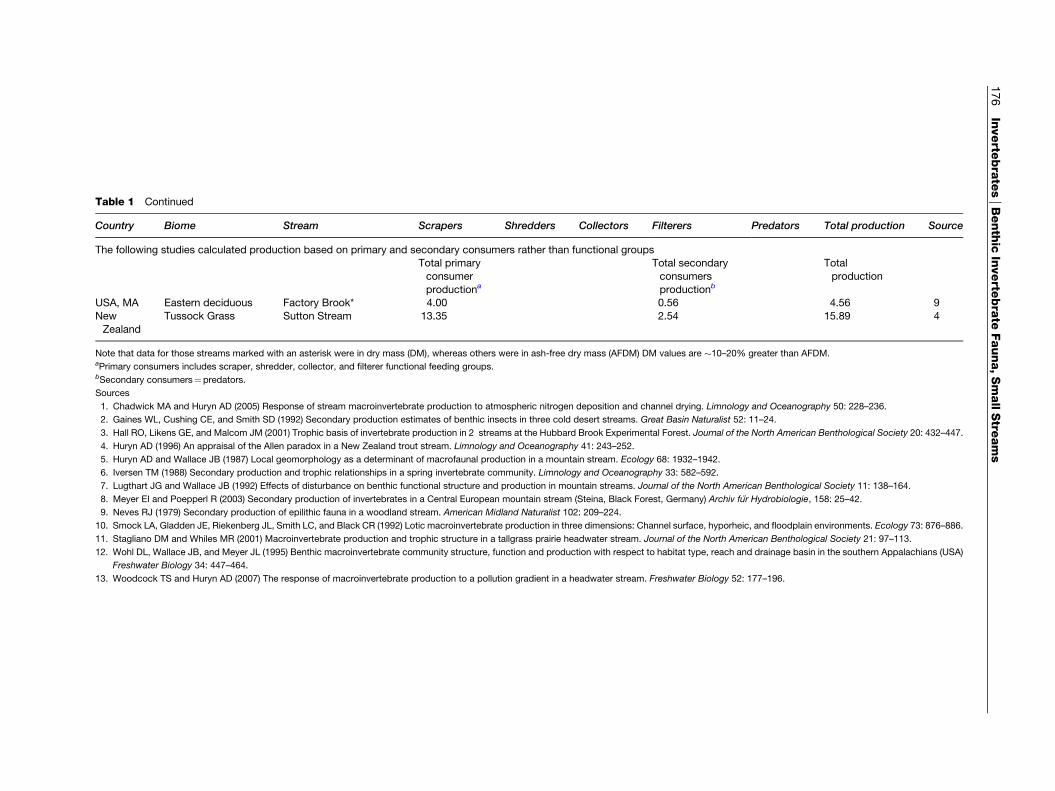
Table 1 Continued
Country Biome Stream Scrapers Shredders Collectors Filterers Predators Total production Source
The following studies calculated production based on primary and secondary consumers rather than functional groupsTotal primary
consumer
productiona
Total secondary
consumers
productionb
Total
production
USA, MA Eastern deciduous Factory Brook* 4.00 0.56 4.56 9
New
Zealand
Tussock Grass Sutton Stream 13.35 2.54 15.89 4
Note that data for those streams marked with an asterisk were in dry mass (DM), whereas others were in ash-free dry mass (AFDM) DM values are �10–20% greater than AFDM.aPrimary consumers includes scraper, shredder, collector, and filterer functional feeding groups.bSecondary consumers¼predators.
Sources
1. Chadwick MA and Huryn AD (2005) Response of stream macroinvertebrate production to atmospheric nitrogen deposition and channel drying. Limnology and Oceanography 50: 228–236.
2. Gaines WL, Cushing CE, and Smith SD (1992) Secondary production estimates of benthic insects in three cold desert streams. Great Basin Naturalist 52: 11–24.
3. Hall RO, Likens GE, and Malcom JM (2001) Trophic basis of invertebrate production in 2 streams at the Hubbard Brook Experimental Forest. Journal of the North American Benthological Society 20: 432–447.
4. Huryn AD (1996) An appraisal of the Allen paradox in a New Zealand trout stream. Limnology and Oceanography 41: 243–252.
5. Huryn AD and Wallace JB (1987) Local geomorphology as a determinant of macrofaunal production in a mountain stream. Ecology 68: 1932–1942.
6. Iversen TM (1988) Secondary production and trophic relationships in a spring invertebrate community. Limnology and Oceanography 33: 582–592.
7. Lugthart JG and Wallace JB (1992) Effects of disturbance on benthic functional structure and production in mountain streams. Journal of the North American Benthological Society 11: 138–164.
8. Meyer EI and Poepperl R (2003) Secondary production of invertebrates in a Central European mountain stream (Steina, Black Forest, Germany) Archiv fur Hydrobiologie, 158: 25–42.
9. Neves RJ (1979) Secondary production of epilithic fauna in a woodland stream. American Midland Naturalist 102: 209–224.
10. Smock LA, Gladden JE, Riekenberg JL, Smith LC, and Black CR (1992) Lotic macroinvertebrate production in three dimensions: Channel surface, hyporheic, and floodplain environments. Ecology 73: 876–886.
11. Stagliano DM and Whiles MR (2001) Macroinvertebrate production and trophic structure in a tallgrass prairie headwater stream. Journal of the North American Benthological Society 21: 97–113.
12. Wohl DL, Wallace JB, and Meyer JL (1995) Benthic macroinvertebrate community structure, function and production with respect to habitat type, reach and drainage basin in the southern Appalachians (USA)
Freshwater Biology 34: 447–464.
13. Woodcock TS and Huryn AD (2007) The response of macroinvertebrate production to a pollution gradient in a headwater stream. Freshwater Biology 52: 177–196.
176
Inverte
bra
tes_B
enth
icInverte
bra
teFauna,SmallStre
ams
Encyclopedia of Inland Waters (2009), vol. 2, pp. 173-190

Invertebrates _ Benthic Invertebrate Fauna, Small Streams 177
<2 to >27g m�2 year�1 for streams in a variety oflandscapes (Figures 2–10). The following is anindication of the average secondary production (per-centage of total) in invertebrates: collectors (31.2%,range 11.9–65.9%)> shredders (avg.¼ 27.0%, range
Figure 2 A steep-gradient headwater stream in the
Appalachians of western NC. Note the dense rhododendron
understory, which shades the stream. (Photo by S. L. Eggert.)
Figure 3 Headwater stream draining catchment 53 at the CoweetaNote large amounts of leaves and woody debris in the stream chann
Encyclopedia of Inland Wat
1–62.2%)> predators (avg.¼ 19.0%, range 1.9–31.9%)> filterers (avg.¼ 15.1%, range 0.1–72.1%)> scrapers (avg.¼ 7.4%, range¼ 0.0–38.2%). Scra-per production as a percent of total production wasgreatest in the Steina, Germany (deciduous forest),followed by Kings Creek, a tall grass prairie streamin Kansas, and Bear Brook, NH (deciduous forest).The differences in scraper production in small streamsin the eastern deciduous forest are striking. In BearBrook NH, scraper production comprised >17%of total production, compared with 0.7–9% for theCoweeta streams in NC, which are heavily shaded bydense riparian rhododendron. With the exception ofthree cold desert streams in southeasternWashington,USA, shredder production was always greater than10%of the total production. Percent collector–filtererproduction was lowest in a Danish Spring and highestin a cool desert stream. These data demonstrate thatinvertebrate production in small streams can be quitevariable among various temperate regions. Given theusefulness of this integrative measure for comparingsmall stream functioning in natural and disturbedenvironments, additional efforts at quantifying totalsecondary production in small streams are badlyneeded.
Factors that Influence Invertebrates inSmall Streams
Small stream invertebrates are influenced by physical,chemical, and biological factors (Table 2). Physicalfactors include climate, (e.g., temperature and
Hydrologic Laboratory in western North Carolina during autumn.el. (Photo by S. L. Eggert.)
ers (2009), vol. 2, pp. 173-190

Figure 4 Headwater stream draining watershed 6 in the Hubbard Brook Experimental Forest in central New Hampshire, USA.
(Photo by R. O. Hall.)
Figure 5 Intermittent stream in the Huron Mountains of Michigan’s upper peninsula, USA. (Photo by S. L. Eggert.)
178 Invertebrates _ Benthic Invertebrate Fauna, Small Streams
precipitation), hydrology, and geology. Hydrologyand the frequency of flooding or drying can alsoinfluence community structure and productivity. Forexample, the lowest annual productivity shown inTable 1 occurs in two intermittent streams, EastBear Book and West Bear Brook in Maine, USA.Geology influences both stream substratum andchemistry. Substratum and the proportions of erodingand depositional substratum within a given streamcan have an important effect on invertebrate func-tional distribution and production. The faster flow-ing erosional reaches in small streams are often
Encyclopedia of Inland Water
dominated by filter-feeders and scrapers, whereasdepositional reaches with greater amounts of retainedorganic matter are often dominated by shredders andcollectors. Stream chemistries can be strikingly differ-ent from those in nearby streams if they have differentunderlying geologies. For example, in the southernAppalachians, streams draining limestone regionssuch as the ridge and valley province have highernutrients, conductivity, pH, and primary productionthan those draining the crystalline Appalachians.Geology and climate also influence the vegetationof catchments, including the abundance and type of
s (2009), vol. 2, pp. 173-190

Figure 6 Bison grazing in the riparian area of the headwaters
of King’s Creek, at the Konza Prairie LTER in Kansas, USA.
(Photo by Walter Dodd.)
Figure 7 Douglas Creek, a cold desert stream in eastern
Washington, USA. (Photo by C. E. Cushing.)
Figure 8 Rattlesnake Creek, a cold desert stream in eastern
Washington, USA. (Photo by C. E. Cushing.)
Figure 9 A spring-fed tundra stream in Alaska, USA. (Photo
by A. D. Huryn.)
Invertebrates _ Benthic Invertebrate Fauna, Small Streams 179
riparian vegetation. Streams that are open, and whichreceive large solar inputs compared with those drain-ing dense forested catchments, may have a very dif-ferent food base (autochthonous) compared withthose receiving primarily allochthonous inputs fromforested catchments. Depending on the food base,small streams may display large differences in func-tional group abundance, biomass, and production.
Encyclopedia of Inland Waters (2009), vol. 2, pp. 173-190

Figure 10 A stream draining tall tussock grass on SouthIsland, New Zealand. (Photo by A. D. Huryn.)
180 Invertebrates _ Benthic Invertebrate Fauna, Small Streams
Ecological Roles of Invertebrates inSmall Streams
Feeding activities of invertebrates in small streams linkheadwater streams to larger rivers downstream byaltering resource quantity, size, and shape (Table 2).For example, the shredding of leaf detritus and woodydebris by shredders in headwater streams increases therate of coarse organic matter breakdown to fineorganic matter, which is transported by the current todownstream reaches. Scrapers, through their feedingactivities and dislodging of epilithon, can enhance themovement of downstream organic particles. Heavygrazing by scrapers results in periphyton mats withadnate, or closely attached forms of diatoms that areless susceptible to scouring during disturbances such aslarge storms and also promotes nutrient turnover inperiphyton communities. Thus, both shredding andgrazing activities may result in a consistent, prolongedrelease of materials to downstream reaches, in contrastto large storms that induce pulsed massive export overshort time intervals. The role of gatherers in FPOMtransport has been implicated in an Idaho stream,which exhibited continuous deposition and resuspen-sion as particles moved downstream. In montanePuerto Rican streams, feeding activities of atyid shrimpreduce organic matter accrual on benthic substrates.Other invertebrate gatherers such as Ptychoptera
Encyclopedia of Inland Water
(Diptera: Ptychopteridae) and sericostomatid (Tri-choptera) larvae may transfer fine organic matterburied in depositional areas to substratum surfaces asfeces.
Filter-feeding stream invertebrates enhance reten-tion of organic matter and nutrients. Certain inverte-brates can transport superficial organic matter todeeper sediments, which reduces downstream trans-port. However, direct removal of transported mate-rial by filter-feeders has received the bulk of attentionand has been shown to have variable effects on reten-tion depending on size of stream, abundance of fil-terers, and taxon-specific differences in feeding, (e.g.,feeding on extremely fine organic particles or driftinginvertebrates). Studies using radioactive tracers inAlaskan streams have also suggested that particlesgenerated by invertebrate scrapers such as baetidmayflies and chironomid larvae were instrumentalin supplying fine particles to downstream black flies.Microfilterers such as the caddisfly family Philopota-midae, and black flies (Diptera: Simuliidae), andbivalve mollusks increase particle sizes by ingestingminute particles and egesting compacted fecal parti-cles larger than those originally consumed. Suchmicrofilterers perform two very important functionsin streams. First, they remove FPOM from suspension(which would otherwise pass unused through thestream segment) and second, they defecate largerparticles, which are available to deposit-feedingdetritivores.
Predators can play numerous roles at scales rangingfrom individuals to ecosystems and invertebrate pre-dators in small streams are no exception. Predatorscan influence export and retention of energy andnutrients through their effects on the standing stocksof other functional groups. Other mechanismsinclude decreasing rates of nutrient cycling by immo-bilizing nutrients in long-lived predator taxa versusshort-lived prey. Besides direct consumption, foragingby invertebrate predators can enhance invertebratedrift and suspended FPOM, which also increases ex-port of nutrients. Invertebrate predators can enhanceretention of organic matter by retarding breakdownrates of leaf litter as well as subsequent generation ofFPOM. Predaceous stoneflies and caddisflies cansignificantly decrease the rate of leaf litter processingby reducing shredder populations in leaf packs. Inver-tebrate predators can also increase the rate of down-stream movement of organisms and sediment. Manystream invertebrates exhibit different responses tofish and invertebrate predators, and the local impactof invertebrate predation on benthic prey may exceedthe impact of fish predators. In the presence of fish,invertebrate prey often reduces movement and seeksrefuge in the substrate. In contrast, invertebrate
s (2009), vol. 2, pp. 173-190
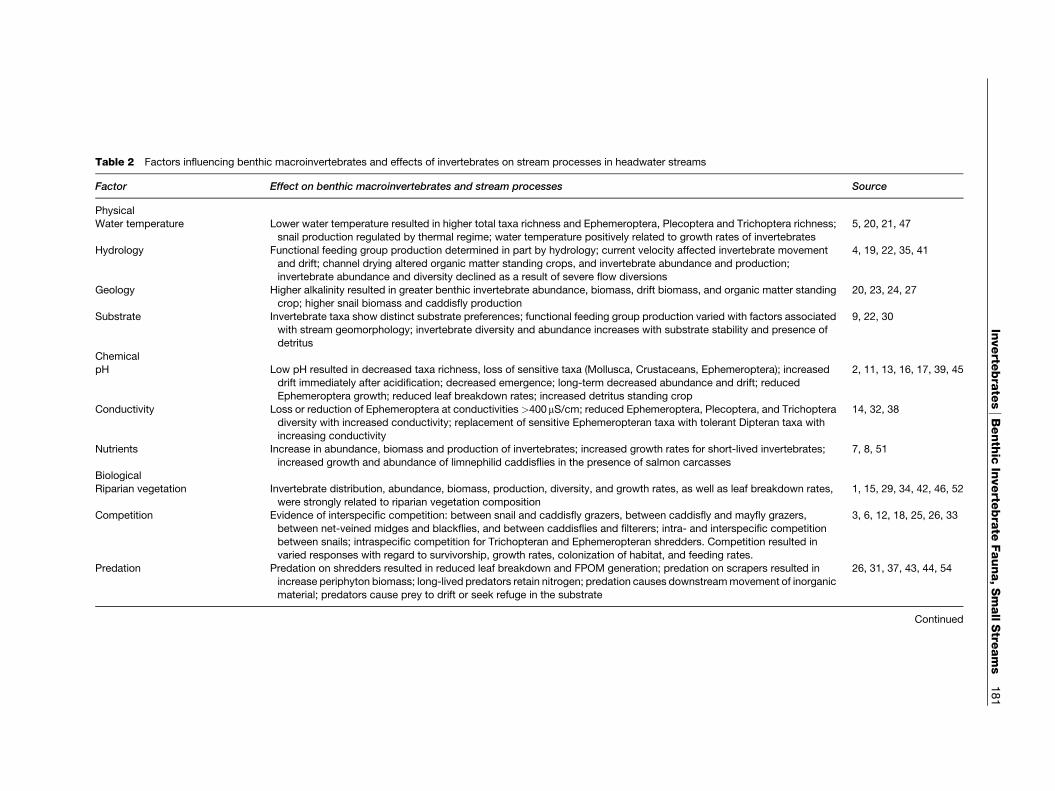
Table 2 Factors influencing benthic macroinvertebrates and effects of invertebrates on stream processes in headwater stream
Factor Effect on benthic macroinvertebrates and stream processes Source
PhysicalWater temperature Lower water temperature resulted in higher total taxa richness and Ephemeroptera, Plecopte and Trichoptera richness;
snail production regulated by thermal regime; water temperature positively related to grow rates of invertebrates
5, 20, 21, 47
Hydrology Functional feeding group production determined in part by hydrology; current velocity affec d invertebrate movement
and drift; channel drying altered organic matter standing crops, and invertebrate abundan and production;invertebrate abundance and diversity declined as a result of severe flow diversions
4, 19, 22, 35, 41
Geology Higher alkalinity resulted in greater benthic invertebrate abundance, biomass, drift biomass, nd organic matter standing
crop; higher snail biomass and caddisfly production
20, 23, 24, 27
Substrate Invertebrate taxa show distinct substrate preferences; functional feeding group production v ied with factors associatedwith stream geomorphology; invertebrate diversity and abundance increases with substra stability and presence of
detritus
9, 22, 30
ChemicalpH Low pH resulted in decreased taxa richness, loss of sensitive taxa (Mollusca, Crustaceans, hemeroptera); increased
drift immediately after acidification; decreased emergence; long-term decreased abundan and drift; reduced
Ephemeroptera growth; reduced leaf breakdown rates; increased detritus standing crop
2, 11, 13, 16, 17, 39, 45
Conductivity Loss or reduction of Ephemeroptera at conductivities >400mS/cm; reduced Ephemeroptera lecoptera, and Trichopteradiversity with increased conductivity; replacement of sensitive Ephemeropteran taxa with lerant Dipteran taxa with
increasing conductivity
14, 32, 38
Nutrients Increase in abundance, biomass and production of invertebrates; increased growth rates fo hort-lived invertebrates;
increased growth and abundance of limnephilid caddisflies in the presence of salmon carc sses
7, 8, 51
Biological
Riparian vegetation Invertebrate distribution, abundance, biomass, production, diversity, and growth rates, as w ll as leaf breakdown rates,
were strongly related to riparian vegetation composition
1, 15, 29, 34, 42, 46, 52
Competition Evidence of interspecific competition: between snail and caddisfly grazers, between caddis and mayfly grazers,
between net-veined midges and blackflies, and between caddisflies and filterers; intra- an interspecific competition
between snails; intraspecific competition for Trichopteran and Ephemeropteran shredders ompetition resulted in
varied responses with regard to survivorship, growth rates, colonization of habitat, and fee ing rates.
3, 6, 12, 18, 25, 26, 33
Predation Predation on shredders resulted in reduced leaf breakdown and FPOM generation; predatio on scrapers resulted in
increase periphyton biomass; long-lived predators retain nitrogen; predation causes downs ammovement of inorganic
material; predators cause prey to drift or seek refuge in the substrate
26, 31, 37, 43, 44, 54
Continued
Inverte
bra
tes_B
enth
icInverte
bra
teFauna,SmallStre
ams
181
Encyclopedia of Inland Waters (2009), vol. 2, pp. 173-190
s
ra
th
te
ce
a
arte
Ep
ce
, Pto
r s
a
e
fly
d
. C
dn
tre
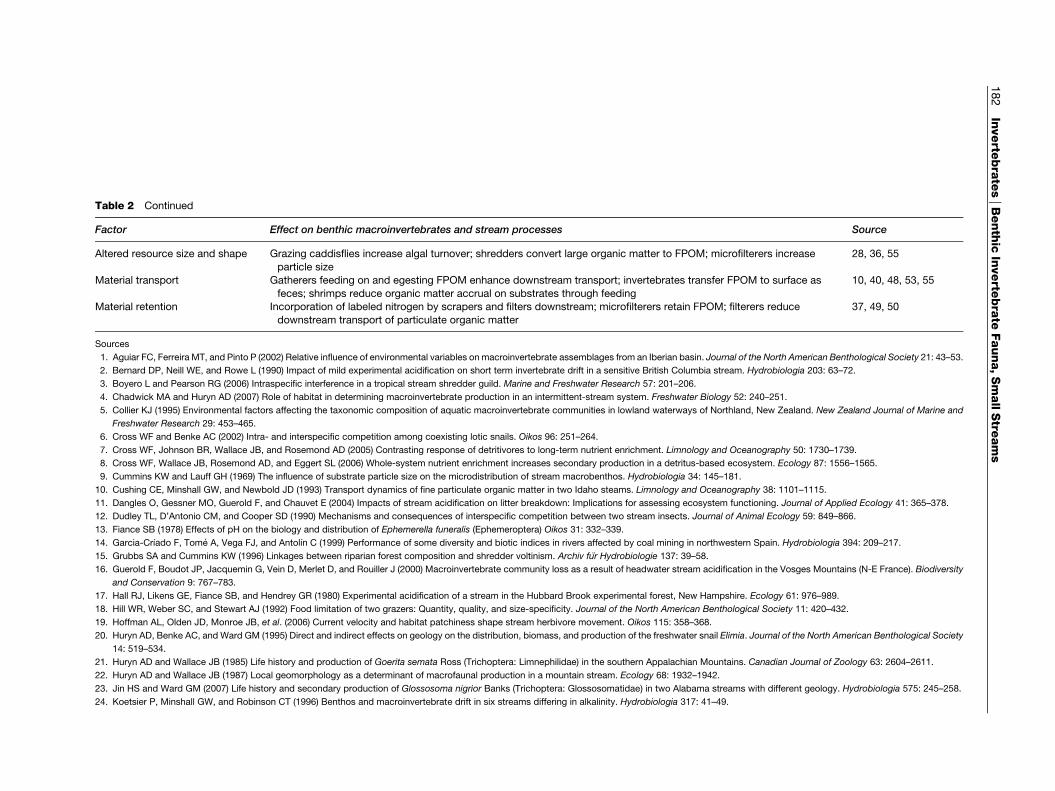
2 Continued
r Effect on benthic macroinvertebrates and stream processes Source
d resource size and shape Grazing caddisflies increase algal turnover; shredders convert large organic matter to FPOM; microfilterers increase
particle size
28, 36, 55
ial transport Gatherers feeding on and egesting FPOM enhance downstream transport; invertebrates transfer FPOM to surface as
feces; shrimps reduce organic matter accrual on substrates through feeding
10, 40, 48, 53, 55
ial retention Incorporation of labeled nitrogen by scrapers and filters downstream; microfilterers retain FPOM; filterers reduce
downstream transport of particulate organic matter
37, 49, 50
s
uiar FC, Ferreira MT, and Pinto P (2002) Relative influence of environmental variables on macroinvertebrate assemblages from an Iberian basin. Journal of the North American Benthological Society 21: 43–53.
rnard DP, Neill WE, and Rowe L (1990) Impact of mild experimental acidification on short term invertebrate drift in a sensitive British Columbia stream. Hydrobiologia 203: 63–72.
yero L and Pearson RG (2006) Intraspecific interference in a tropical stream shredder guild. Marine and Freshwater Research 57: 201–206.
adwick MA and Huryn AD (2007) Role of habitat in determining macroinvertebrate production in an intermittent-stream system. Freshwater Biology 52: 240–251.
llier KJ (1995) Environmental factors affecting the taxonomic composition of aquatic macroinvertebrate communities in lowland waterways of Northland, New Zealand. New Zealand Journal of Marine and
eshwater Research 29: 453–465.
oss WF and Benke AC (2002) Intra- and interspecific competition among coexisting lotic snails. Oikos 96: 251–264.
oss WF, Johnson BR, Wallace JB, and Rosemond AD (2005) Contrasting response of detritivores to long-term nutrient enrichment. Limnology and Oceanography 50: 1730–1739.
oss WF, Wallace JB, Rosemond AD, and Eggert SL (2006) Whole-system nutrient enrichment increases secondary production in a detritus-based ecosystem. Ecology 87: 1556–1565.
mmins KW and Lauff GH (1969) The influence of substrate particle size on the microdistribution of stream macrobenthos. Hydrobiologia 34: 145–181.
shing CE, Minshall GW, and Newbold JD (1993) Transport dynamics of fine particulate organic matter in two Idaho steams. Limnology and Oceanography 38: 1101–1115.
ngles O, Gessner MO, Guerold F, and Chauvet E (2004) Impacts of stream acidification on litter breakdown: Implications for assessing ecosystem functioning. Journal of Applied Ecology 41: 365–378.
dley TL, D’Antonio CM, and Cooper SD (1990) Mechanisms and consequences of interspecific competition between two stream insects. Journal of Animal Ecology 59: 849–866.
ance SB (1978) Effects of pH on the biology and distribution of Ephemerella funeralis (Ephemeroptera) Oikos 31: 332–339.
rcia-Crıado F, Tome A, Vega FJ, and Antolın C (1999) Performance of some diversity and biotic indices in rivers affected by coal mining in northwestern Spain. Hydrobiologia 394: 209–217.
ubbs SA and Cummins KW (1996) Linkages between riparian forest composition and shredder voltinism. Archiv fur Hydrobiologie 137: 39–58.
erold F, Boudot JP, Jacquemin G, Vein D, Merlet D, and Rouiller J (2000) Macroinvertebrate community loss as a result of headwater stream acidification in the Vosges Mountains (N-E France). Biodiversity
d Conservation 9: 767–783.
ll RJ, Likens GE, Fiance SB, and Hendrey GR (1980) Experimental acidification of a stream in the Hubbard Brook experimental forest, New Hampshire. Ecology 61: 976–989.
ll WR, Weber SC, and Stewart AJ (1992) Food limitation of two grazers: Quantity, quality, and size-specificity. Journal of the North American Benthological Society 11: 420–432.
ffman AL, Olden JD, Monroe JB, et al. (2006) Current velocity and habitat patchiness shape stream herbivore movement. Oikos 115: 358–368.
ryn AD, Benke AC, and Ward GM (1995) Direct and indirect effects on geology on the distribution, biomass, and production of the freshwater snail Elimia. Journal of the North American Benthological Society
: 519–534.
ryn AD and Wallace JB (1985) Life history and production of Goerita semata Ross (Trichoptera: Limnephilidae) in the southern Appalachian Mountains. Canadian Journal of Zoology 63: 2604–2611.
ryn AD and Wallace JB (1987) Local geomorphology as a determinant of macrofaunal production in a mountain stream. Ecology 68: 1932–1942.
HS and Ward GM (2007) Life history and secondary production of Glossosoma nigrior Banks (Trichoptera: Glossosomatidae) in two Alabama streams with different geology. Hydrobiologia 575: 245–258.
etsier P, Minshall GW, and Robinson CT (1996) Benthos and macroinvertebrate drift in six streams differing in alkalinity. Hydrobiologia 317: 41–49.
182
Inverte
bra
tes_B
enth
icInverte
bra
teFauna,SmallStre
ams
Encyclopedia of Inland Waters (2009), vol. 2, pp. 173-190
Table
Facto
Altere
Mater
Mater
Source
1. Ag
2. Be
3. Bo
4. Ch
5. Co
Fr
6. Cr
7. Cr
8. Cr
9. Cu
10. Cu
11. Da
12. Du
13. Fi
14. Ga
15. Gr
16. Gu
an
17. Ha
18. Hi
19. Ho
20. Hu
14
21. Hu
22. Hu
23. Jin
24. Ko

25. Kohler SL (1992) Competition and the structure of a benthic stream community. Ecological Monographs 62: 165–188.
26. Kohler SL and Wiley MJ (1997) Pathogen outbreaks reveal large-scale effects of competition in stream communities. Ecology 78: 2164–2176.
27. Krueger CC and Waters TF (1983) Annual production of macroinvertebrates in three streams of different water quality. Ecology 64: 840–850.
28. Lamberti GA and Resh VH (1983) Stream periphyton and insect herbivores: An experimental study of grazing by a caddisfly population. Ecology 64: 1124–1135.
29. Lecerf A, Dobson M, Dang CK, and Chauvet E (2005) Riparian plant species loss alters trophic dynamics in detritus-based stream ecosystems. Oecologia 146: 432–442.
30. Mackay RJ and Kalff J (1969) Seasonal variation in standing crop and species diversity of insect communities in a small Quebec stream. Ecology 50: 101–109.
31. Malmqvist B (1993) Interactions in stream leaf packs: Effects of a stonefly predator on detritivores and organic matter processing. Oikos 66: 454–462.
32. Marques MJ, Martınez-Conde E, and Rovira JV (2003) Effects of zinc and lead mining on the benthic macroinvertebrates of a fluvial ecosystem. Water, Air, and Soil Pollution 148: 363–388.
33. McAuliffe JR (1984) Resource depression by a stream herbivore: Effects on distributions and abundances of other grazers. Oikos 42: 327–333.
34. Milner AM and Gloyne-Phillips IT (2005) The role of riparian vegetation and woody debris in the development of macroinvertebrate assemblages in streams. River Research and Applications 21: 403–420.
35. Minshall GW and Winger PV (1968) The effect of reduction in stream flow on invertebrate drift. Ecology 49: 580–582.
36. Petersen RC and Cummins KW (1974) Leaf processing in a woodland stream. Freshwater Biology 4: 343–368.
37. Peterson BJ, Bahr M, and Kling GW (1997) A tracer investigation of nitrogen cycling in a pristine tundra river. Canadian Journal of Fisheries & Aquatic Sciences 54: 2361–2367.
38. Pond GJ and McMurray SE (2002) A macroinvertebrate bioassessment index for headwater streams in the eastern coalfield region, Kentucky. Kentucky Department for Environmental Protection, Division of
Water, Frankfort, KY.
39. Pretty JL, Gibberson DJ, and Dobson M (2005) Resource dynamics and detritivore production in an acid stream. Freshwater Biology 50: 578–591.
40. Pringle CM, Blake GA, Covich AP, Buzby KM, and Finley A (1993) Effects of omnivorous shrimp in a montane tropical stream: Sediment removal, disturbance of sessile invertebrates and enhancement of
understory algal biomass. Oecolgia 93: 1–11.
41. Rader RB and Belish TA (1999) Influence of mild to severe flow alterations on invertebrates in three mountain streams. Regulated Rivers: Research & Management 15: 353–363.
42. Ross HH (1963) Stream communities and terrestrial biomes. Archiv fur Hydrobiologie 59: 235–242.
43. Sih A and Wooster D (1994) Prey behavior, prey dispersal, and predator impacts on stream prey. Ecology 75: 1199–1207.
44. Statzner B, Fuchs U, and Higler LWG. (1996) Sand erosion by mobile predaceous stream insects: Implications for ecology and hydrology. Water Resources Research 32: 2279–2287.
45. Sutcliffe DW and Carrick TR (1973) Studies on mountain streams in the English Lake District. I. pH, calcium and the distribution of invertebrates in the River Duddon. Freshwater Biology 3: 437–462.
46. Synder CD, Young JA, Lemarie DP, and Smith DR (2002) Influence of eastern hemlock (Tsuga canadensis) forests on aquatic invertebrate assemblages in headwater streams. Canadian Journal of Fisheries &
Aquatic Sciences 59: 262–275.
47. Vannote RL and Sweeney BW (1980) Geographic analysis of thermal equilibria: A conceptual model for evaluating the effect of natural and modified thermal regimes on aquatic insect communities. The
American Naturalist 115: 667–695.
48. Wagner R (1991) The influence of the diel activity pattern of the larvae of Sericostoma personatum (Kirby and Spence) (Trichoptera) on organic matter distribution in stream-bed sediments – A laboratory study.
Hydrobiologia 224: 65–70.
49. Wallace JB and Webster JR (1996) The role of macroinvertebrates in stream ecosystem function. Annual Review of Entomology 41: 115–139.
50. Wallace JB, Webster JR, and Woodall WR (1977) The role of filter-feeders in flowing waters. Archiv fur Hydrobiologie 79: 506–532.
51. Walter JK, Bilby RE, and Fransen BR (2006) Effects of Pacific salmon spawning and carcass availability on the caddisfly Ecclisomyia conspersa (Trichoptera: Limnephilidae) Freshwater Biology 51: 1211–1218.
52. Whiles MR and Wallace JB (1997) Leaf litter decomposition and macroinvertebrate communities in headwater streams draining pine and hardwood catchments. Hydrobiologia 353: 107–119.
53. Wolf B, Zwick P, and Marxsen J (1997) Feeding ecology of the freshwater detritivore Ptychoptera paludosa (Diptera, Nematocera). Freshwater Biology 38: 375–386.
54. Wooster D (1994) Predator impacts on stream benthic prey. Oecologia 99: 7–15.
55. Wotton RS, Malmqvist B, Muotka T, and Larsson K (1998) Fecal pellets from a dense aggregation of suspension-feeders in a stream: An example of ecosystem engineering. Limnology and Oceanography 43:
719–725.
Inverte
bra
tes_B
enth
icInverte
bra
teFauna,SmallStre
ams
183
Encyclopedia of Inland Waters (2009), vol. 2, pp. 173-190

184 Invertebrates _ Benthic Invertebrate Fauna, Small Streams
predators have the ability to search in sites similar tothose being used by their prey, and the latter mayrespond by actively entering the water column anddrifting downstream. Foraging by invertebrate preda-tors can also influence the downstream movement ofinorganic material through their physical activities.Several studies have suggested that the foraging activ-ities increase erosion and downstream transport ofsand and fine sediments. Furthermore, some specia-lized parasites (a subcategory of predators) of scrap-ing Glossosoma caddisflies have been shown toinfluence periphyton biomass in Michigan streams.
Anthropogenic and Natural Disturbancesto Invertebrate Productivity in SmallStreams
Many unique fauna are found in small streams. Unfor-tunately, invertebrate fauna in these streams are underassault by anthropogenic and natural disturbances suchas invasive species, agriculture, development, logging,mining, recreational activities, global climate change,and wildfires (Table 3). Macroinvertebrate commu-nities and productivity can be altered, which can affecthigher trophic levels (e.g., fish production) and otherstream processes (e.g., organic matter processing).
Invasive Species
Invasive species within riparian habitats can havelasting effects on headwater stream functioningbecause of the tight linkage between riparian forestsand stream processes. Macroinvertebrates abundanceand diversity in small streams can be altered bychanges in microclimate, energy availability, and hab-itat that results from loss of tree species within theriparian forest. Outbreaks of terrestrial invaders suchas the balsam and hemlock woolly adelgids and thegypsy moth result in losses of some riparian tree spe-cies, pulses of slow decaying wood inputs, increases inother tree species, and increases of pesticides used tocontrol invading pests. Indirectly, these changescan affect headwater stream functioning throughreductions in the survival, growth, and emergence ofmacroinvertebrate shredders and detrital processing.The effects of terrestrial invasive species on streamecosystems are expected to increase in the future.Nonnative scale and fungal diseases such as dogwoodanthracnose and beech bark disease have invaded for-ests of the eastern United States, and the fungus caus-ing butternut canker is beginning to spread rapidly.The fungus causing chestnut blight eliminated theAmerican chestnut from eastern forests and resultedin decreased leaf litter processing, decreased quality of
Encyclopedia of Inland Water
litter inputs, and decreased invertebrate growth ratesin headwater streams. The input of dead chestnut logsinto streams also facilitated the retention of sedimentand served to stabilize stream channels. Few examplesof exotic aquatic species invading small streams havebeen documented in the literature. One species thatsuccessfully invaded first- and second-order streams,Gammarus pulex, resulted in spatial and temporalreductions in macroinvertebrate diversity.
Agriculture
The filling of former wetlands and headwater streamsfor agriculture has greatly reduced surface water areaworldwide. As an example, 96.6% of the originalsurface water area of the Kavlinge River catchmentin Sweden has been lost due to channelization anddrainage of streams for agriculture over a 141-yearperiod. Along with the loss of small streams, intensiveagriculture results in excessive nitrate levels in streamwater. Overfertilization of agricultural land in low-order sections of river networks affects downstreamriver reaches. It has been estimated that agriculturalsources in Illinois contribute 10–15% of nitrogen andphosphorus loads to the Mississippi River. Nutrientenrichment in small streams can stimulate primaryproduction and higher trophic levels such as scrapersthat feed on the abundant periphyton. In detritus-based streams, increased nutrients can lead toincreases in microbial production on organic matter,which improves the quality of the food resource forshredder invertebrates. With higher food quality,macroinvertebrate production, particularly thosetaxa with short life cycles, can increase dramaticallyin nutrient-enriched streams.
Shifts in the invertebrate community associatedwith increased sedimentation have been observed inheadwater reaches of agriculturally impacted streams.As the percent fine sediment increases, there is usuallya shift from clinging and crawling taxa to burrowers.Insecticide runoff from agricultural fields into head-water streams can have more deleterious impacts onmacroinvertebrate communities. Pesticides introduc-ed into headwater streams can result in the loss ofinvertebrate species, cause shifts in functional produc-tion of invertebrates, and negatively impact ecosystemprocesses such as leaf litter breakdown and FPOMexport.
Urbanization and Roads
Urban growth scenarios predict substantial increasesin population and growth for many regions of theworld. The replacement of forested land and riparianhabitats with impervious surfaces such as roads,
s (2009), vol. 2, pp. 173-190

Table 3 Examples of disturbances and their effects on benthic invertebrates in headwater streams
Disturbance Effect on invertebrates and stream function Source
Invasive species
Gypsy moth defoliation Accelerated detritus processing 27
Decline in eastern hemlock forests due to hemlock woolly adelgid Reduction in alpha and gamma diversity of inverte ates and changes in trophiccomposition; pesticide inputs caused decline in i ertebrate emergence; increased
inputs of slow decaying wood; higher hydrologic ariability
19, 20, 51
Loss of American chestnut trees as a result of chestnut blight Decrease in leaf litter processing, quality of litter in uts, and invertebrate growth rates;
increase in wood inputs and sediment stabilizatio
50, 57
Invasion of G. pulex Increased predation on native invertebrates 28, 29
Agriculture
Filling and tiling of streams Reduced drainage density of stream network 59
Overapplication of fertilizer Increased nitrogen alter food resources for inverteb tes; increase in abundance, biomassand production of invertebrates; increased growt rates for short-lived invertebrates
13, 14, 16
Sediment runoff Decline in Ephemeropteran, Plecopteran, and Trich pteran, and Coleopteran taxa;
increases in chironomids, oligochaetes and mollu cs
2, 6, 7, 24,
39, 44, 53Insecticide runoff Decline in invertebrate abundance, biomass, and p oduction; loss of species; shifts in
functional structure; decline in organic matter exp rt and leaf breakdown rates
15, 31, 32, 33
Increased water temperature Decline in Ephemeroptera, Plecoptera, and Trichop era 24, 44
Urbanization and roadsAltered hydrology and geomorphology, increased bacterial populations and
turbidity, increases of pesticide, herbicide, and fertilizer runoff; decline in
habitat
Decline in invertebrate diversity; decline in Epheme optera, Plecoptera, and Trichoptera;
increase in number of pollution-tolerant taxa; dec e in invertebrate production; decline
in leaf breakdown rates
4, 9, 30, 39,
40, 48,
58, 60
Increased number of culverts Reduced adult caddisfly diversity and abundance ove culverts 5Forestry practices
Increased stream temperature, discharge, nutrients, and primary
production; reduced organic matter inputs
Shift from allochthonous to autochthonous energy; ncrease in abundance, biomass, and
production of taxa with short life cycles; leaf litter reakdown altered; significantreduction in invertebrate production with decline detrital inputs
1, 3, 21, 23,
41, 42, 52,55, 56
Sediment runoff from logging roads Decline in total richness and abundance of all inve ebrate taxa 22, 54
Mining
Acid mine drainage and metal uptake Reductions in abundances of sensitive invertebrate axa; increase in tolerant taxa; declinein species diversity; increased drift; reduced com unity respiration; reduced secondary
production
8, 11, 12, 18,34, 45, 49
Mountaintop mining/Valley fill
Burial of headwater streams; increased sedimentation, conductivity,and metals
Elimination of all biota in buried streams; downstre m declines in Ephemeropterarichness, decline in abundances of Ephemeropte , Odonata, Coleoptera; decline in
scraper and shredder abundance
25, 43
Global climate change
Channel drying Altered organic matter standing crops, and inverteb ate abundance and production; shiftsfrom large-bodied, long-lived taxa to small-bodie , short-lived taxa
10, 17
Continued
Inverte
bra
tes_B
enth
icInverte
bra
teFauna,SmallStre
ams
185
Encyclopedia of Inland Waters (2009), vol. 2, pp. 173-190
brnv
v
p
n
rah
o
sr
o
t
r
lin
ab
i
bin
rt
tm
ara
rd
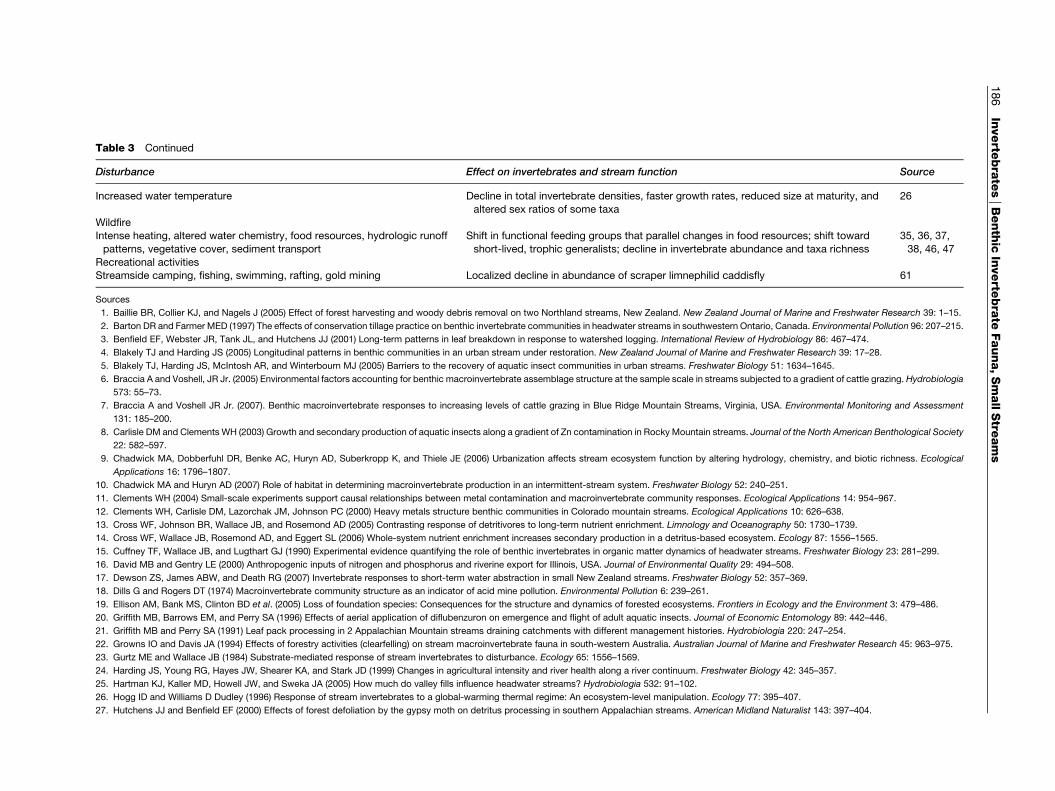
Table 3 Continued
Disturbance Effect on invertebrates and stream function Source
Increased water temperature Decline in total invertebrate densities, faster growth rates, reduced size at maturity, and
altered sex ratios of some taxa
26
WildfireIntense heating, altered water chemistry, food resources, hydrologic runoff
patterns, vegetative cover, sediment transport
Shift in functional feeding groups that parallel changes in food resources; shift toward
short-lived, trophic generalists; decline in invertebrate abundance and taxa richness
35, 36, 37,
38, 46, 47
Recreational activitiesStreamside camping, fishing, swimming, rafting, gold mining Localized decline in abundance of scraper limnephilid caddisfly 61
Sources
1. Baillie BR, Collier KJ, and Nagels J (2005) Effect of forest harvesting and woody debris removal on two Northland streams, New Zealand. New Zealand Journal of Marine and Freshwater Research 39: 1–15.
2. Barton DR and Farmer MED (1997) The effects of conservation tillage practice on benthic invertebrate communities in headwater streams in southwestern Ontario, Canada. Environmental Pollution 96: 207–215.
3. Benfield EF, Webster JR, Tank JL, and Hutchens JJ (2001) Long-term patterns in leaf breakdown in response to watershed logging. International Review of Hydrobiology 86: 467–474.
4. Blakely TJ and Harding JS (2005) Longitudinal patterns in benthic communities in an urban stream under restoration. New Zealand Journal of Marine and Freshwater Research 39: 17–28.
5. Blakely TJ, Harding JS, McIntosh AR, and Winterbourn MJ (2005) Barriers to the recovery of aquatic insect communities in urban streams. Freshwater Biology 51: 1634–1645.
6. Braccia A and Voshell, JR Jr. (2005) Environmental factors accounting for benthic macroinvertebrate assemblage structure at the sample scale in streams subjected to a gradient of cattle grazing. Hydrobiologia
573: 55–73.
7. Braccia A and Voshell JR Jr. (2007). Benthic macroinvertebrate responses to increasing levels of cattle grazing in Blue Ridge Mountain Streams, Virginia, USA. Environmental Monitoring and Assessment
131: 185–200.
8. Carlisle DM and Clements WH (2003) Growth and secondary production of aquatic insects along a gradient of Zn contamination in Rocky Mountain streams. Journal of the North American Benthological Society
22: 582–597.
9. Chadwick MA, Dobberfuhl DR, Benke AC, Huryn AD, Suberkropp K, and Thiele JE (2006) Urbanization affects stream ecosystem function by altering hydrology, chemistry, and biotic richness. Ecological
Applications 16: 1796–1807.
10. Chadwick MA and Huryn AD (2007) Role of habitat in determining macroinvertebrate production in an intermittent-stream system. Freshwater Biology 52: 240–251.
11. Clements WH (2004) Small-scale experiments support causal relationships between metal contamination and macroinvertebrate community responses. Ecological Applications 14: 954–967.
12. Clements WH, Carlisle DM, Lazorchak JM, Johnson PC (2000) Heavy metals structure benthic communities in Colorado mountain streams. Ecological Applications 10: 626–638.
13. Cross WF, Johnson BR, Wallace JB, and Rosemond AD (2005) Contrasting response of detritivores to long-term nutrient enrichment. Limnology and Oceanography 50: 1730–1739.
14. Cross WF, Wallace JB, Rosemond AD, and Eggert SL (2006) Whole-system nutrient enrichment increases secondary production in a detritus-based ecosystem. Ecology 87: 1556–1565.
15. Cuffney TF, Wallace JB, and Lugthart GJ (1990) Experimental evidence quantifying the role of benthic invertebrates in organic matter dynamics of headwater streams. Freshwater Biology 23: 281–299.
16. David MB and Gentry LE (2000) Anthropogenic inputs of nitrogen and phosphorus and riverine export for Illinois, USA. Journal of Environmental Quality 29: 494–508.
17. Dewson ZS, James ABW, and Death RG (2007) Invertebrate responses to short-term water abstraction in small New Zealand streams. Freshwater Biology 52: 357–369.
18. Dills G and Rogers DT (1974) Macroinvertebrate community structure as an indicator of acid mine pollution. Environmental Pollution 6: 239–261.
19. Ellison AM, Bank MS, Clinton BD et al. (2005) Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Frontiers in Ecology and the Environment 3: 479–486.
20. Griffith MB, Barrows EM, and Perry SA (1996) Effects of aerial application of diflubenzuron on emergence and flight of adult aquatic insects. Journal of Economic Entomology 89: 442–446.
21. Griffith MB and Perry SA (1991) Leaf pack processing in 2 Appalachian Mountain streams draining catchments with different management histories. Hydrobiologia 220: 247–254.
22. Growns IO and Davis JA (1994) Effects of forestry activities (clearfelling) on stream macroinvertebrate fauna in south-western Australia. Australian Journal of Marine and Freshwater Research 45: 963–975.
23. Gurtz ME and Wallace JB (1984) Substrate-mediated response of stream invertebrates to disturbance. Ecology 65: 1556–1569.
24. Harding JS, Young RG, Hayes JW, Shearer KA, and Stark JD (1999) Changes in agricultural intensity and river health along a river continuum. Freshwater Biology 42: 345–357.
25. Hartman KJ, Kaller MD, Howell JW, and Sweka JA (2005) How much do valley fills influence headwater streams? Hydrobiologia 532: 91–102.
26. Hogg ID and Williams D Dudley (1996) Response of stream invertebrates to a global-warming thermal regime: An ecosystem-level manipulation. Ecology 77: 395–407.
27. Hutchens JJ and Benfield EF (2000) Effects of forest defoliation by the gypsy moth on detritus processing in southern Appalachian streams. American Midland Naturalist 143: 397–404.
186
Inverte
bra
tes_B
enth
icInverte
bra
teFauna,SmallStre
ams
Encyclopedia of Inland Waters (2009), vol. 2, pp. 173-190

28. Kelly DW and Dick JTA (2005) Effects of environment and an introduced invertebrate species on the structure of benthic macroinvertebrate spec s at the catchment level. Archiv fur Hydrobiologie 164: 69–88.
29. Kelly DW, Dick JTA, Montgomery WI, and MacNeil C (2003) Differences in composition of macroinvertebrate communities with invasive and nativ Gammarus spp. (Crustacea: Amphipoda) Freshwater Biology
48: 306–315.
30. Kemp SJ and Spotila JR (1997) Effects of urbanization on brown trout (Salmo trutta), other fishes and macroinvertebrates in Valley Creek, Valley orge, Pennsylvania. American Midland Naturalist 138: 55–68.
31. Liess M and Schulz R (1999) Linking insecticide contamination and population response in an agricultural stream. Environmental Toxicology an Chemistry 18: 1948–1955.
32. Lugthart GJ and Wallace JB (1992) Effects of disturbance on benthic functional structure and production in mountain streams. Journal of the N rth American Benthological Society 11: 138–164.
33. Lugthart GJ, Wallace JB, and Huryn AD (1990) Secondary production of chironomid communities in insecticide-treated and untreated headwat r streams. Freshwater Biology 24: 417–427.
34. Malmqvist B and Hoffsten P (1999) Influence of drainage from old mine deposits on benthic macroinvertebrate communities in central Swedish treams. Water Research 33: 2415–2423.
35. Mihuc TB and Minshall GW (2005) The trophic basis of reference and post-fire stream food webs 10 years after wildfire in Yellowstone Nationa ark. Aquatic Sciences 67: 541–548.
36. Minshall GW (2003) Responses of stream benthic macroinvertebrates to fire. Forest Ecology and Management 178: 155–161.
37. Minshall GW, Brock JT, Andrews DA, and Robinson CT (2001) Water quality, substratum and biotic responses of five central Idaho (USA) streams uring the first year following the Mortar Creek fire. International
Journal of Wildland Fire 10: 185–199.
38. Minshall GW, Royer TV, and Robinson CT (2001) Response of the Cache Creek macroinvertebrates during the first 10 years following disturbance the 1988 Yellowstone wildfires.Canadian Journal of Fisheries
& Aquatic Sciences 58: 1077–1088.
39. Moore AA and Palmer MA (2005) Invertebrate biodiversity in agricultural and urban headwater streams: Implications for conservation and mana ement. Ecological Applications 15: 1169–1177.
40. Morse CC, Huryn AD, and Cronan C (2003) Impervious surface area as a predictor of the effects of urbanization on stream insect communities Maine, USA. Environmental Monitoring and Assessment 89:
95–127.
41. Newbold JD, Erman DC, and Roby KB (1980) Effects of logging on macroinvertebrates in streams with and without buffer strips. Canadian Jou al of Fisheries & Aquatic Sciences 37: 1076–1085.
42. Noel DS, Martin CW, and Federer CA (1986) Effects of forest clearcutting in New England on stream macroinvertebrates and periphyton. Enviro mental Management 10: 661–670.
43. Pond GJ and McMurray SE (2002) A macroinvertebrate bioassessment index for headwater streams in the eastern coalfield region, Kentucky. entucky Department for Environmental Protection, Division of
Water, Frankfort, KY.
44. Quinn JM, Williamson RB, Smith RK, and Vickers ML (1992) Effects of riparian grazing and channelisation on streams in Southland, New Zealan 2. Benthic invertebrates. New Zealand Journal of Marine and
Freshwater Research 26: 259–273.
45. Roback SS and Richardson JW (1969) The effects of acid mine drainage on aquatic insects. Proceedings of the Academy of Natural Sciences P iladelphia 121: 81–107.
46. Robinson CT, Uehlinger U, and Minshall GW (2005) Functional characteristics of wilderness streams twenty years following wildfire. Western N th American Naturalist 65: 1–10.
47. Roby KB and Azuma DL (1995) Changes in a reach of a northern California stream following wildfire. Environmental Management 19: 591–600.
48. Roy AH, Rosemond AD, Paul MJ, Leigh DS, and Wallace JB (2003) Stream macroinvertebrate response to catchment urbanisation (Georgia, U A). Freshwater Biology 48: 329–346.
49. Short TM, Black JA, and Birge WJ (1990) Effects of acid-mine drainage on the chemical and biological character of an alkaline headwater stream Archives of Environmental Contamination and Toxicology 19:
241–248.
50. Smock LA and MacGregor CM (1988) Impact of the American chestnut blight on aquatic shredding macroinvertebrates. Journal of the North A erican Benthological Society 7: 212–221.
51. Snyder CD, Young JA, Lemarie DP, and Smith DR (2002) Influence of eastern hemlock (Tsuga canadensis) forests on aquatic invertebrate assemb ages in headwater streams. Canadian Journal of Fisheries and
Aquatic Sciences 59: 262–275.
52. StoneMK andWallace JB (1998) Long-term recovery of amountain stream from clear-cut logging: The effects of forest succession on benthic inve tebrate community structure. Freshwater Biology 39: 141–169.
53. Stone ML, Whiles MR, Webber JA, Williard KWJ, and Reeve JD (2005) Macroinvertebrate communities in agriculturally impacted southern Illinois reams: Patterns with riparian vegetation, water quality, and in-
stream habitat quality. Journal of Environmental Quality 34: 907–917.
54. Tebo LB Jr. (1955) Effects of siltation, resulting from improper logging, on the bottom fauna of a small trout stream in the southern Appalachian . Progressive Fish-Culturist 17: 64–70.
55. Wallace JB, Eggert SL, Meyer JL, and Webster JR (1999) Effects of resource limitation on a detrital-based ecosystem. Ecological Monographs 9: 409–442.
56. Wallace JB and Gurtz ME (1986) Response of Baetis mayflies (Ephemeroptera) to catchment logging. American Midland Naturalist 115: 25–41.
57. Wallace JB, Webster JR, Eggert SL, Meyer JL, and Siler ER (2001) Large woody debris in a headwater stream: Long-term legacies of forest dis rbance. International Review of Hydrobiology 86: 501–513.
58. Whiting ER and Clifford HF (1983) Invertebrates and urban runoff in a small northern stream, Edmonton, Alberta, Canada. Hydrobiologia 102: 7 80.
59. Wolf P (1956) Utdikad Civilisation. (Drained Civilization). Malmo, Sweden: Gleerups.
60. Woodcock TS and Huryn AD (2007) The reponse of macroinvertebrate production to a pollution gradient in a headwater stream. Freshwater Bio ogy 52: 177–196.
61. Wright KK and Li JL (1998) Effects of recreational activities on the distribution of Dicosmoecus gilvipes in a mountain stream. Journal of the No American Benthological Society 17: 535–543.
Inverte
bra
tes_B
enth
icInverte
bra
teFauna,SmallStre
ams
187
Encyclopedia of Inland Waters (2009), vol. 2, pp. 173-190
ie
e
F
d
o
e
s
l P
d
by
g
in
rn
n
K
d.
h
or
S
.
m
l
r
st
s
6
tu
3–
l
rth

188 Invertebrates _ Benthic Invertebrate Fauna, Small Streams
rooftops, and lawns alters the hydrology and geomor-phology of streams. Increases in surface runoff asso-ciated with storm flow lead to declines in waterquality, increases in bacterial populations and turbid-ity, and increases of pesticide, herbicide, and fertilizerrunoff into nearby streams. Sediment runoff fromconstruction sites and erosion due to downcuttingresult in habitat loss for aquatic life. Measurableaquatic degradation occurs with 6–10% impervi-ous area and has been long been associated withdecreased water quality of nearby streams. Urbaniza-tion in watersheds containing small streams usuallyresults in less diverse invertebrate communities con-sisting of pollution-tolerant species. Culverts havebeen shown to act as barriers to upstream migrationof adult caddisflies. Since many stressors associatedwith urbanization act synergistically, it is difficult toseparate cause and effect of individual stressors on theinvertebrate communities in small urban streams.
Forestry Practices
Logging results in changes in stream temperatureregimes, increased discharge and altered hydrology,increased nutrient export and increased solar radia-tion and primary production, increased sedimentexport, and changes in dissolved organic matterderived from the terrestrial ecosystem. These changesare accompanied by substantial changes in the energybase of headwater streams, with a shift from allo-chthonous detritus to autochthonous production.The physical and energy base changes can lead tolarge changes in macroinvertebrate community struc-ture. An experimental long-term reduction of organicmatter inputs to a small stream in the southern Appa-lachians resulted in a significant decline in total inver-tebrate production. Invertebrate taxa with short lifecycles and the ability to exploit increases in primaryproduction greatly increase in abundance, biomass,and productivity. Studies in the central and southernAppalachians show that long-term patterns of leaflitter breakdown can be altered for many years fol-lowing logging. However, depending upon the extentof terrestrial succession, invertebrate assemblages canrevert back toward their prelogged condition.
Mining
Mining has severe consequences for benthic inverte-brates in small streams worldwide. Effects of miningon macroinvertebrates in small streams are causedby acid mine drainage, sediments, or burial ofthe streams themselves. Acid mine drainage and theassociated problems of heavy metal contaminationusually results in reductions of numbers of sensitive
Encyclopedia of Inland Water
taxa in the orders of Ephemeroptera (particularlythose of the family Heptageniidae), Plecoptera,Trichoptera, Megaloptera, Odonata, and Dipteraand an overall decline in species diversity. Some stud-ies have shown that functional measures of ben-thic invertebrates such as drift and communityrespiration are also negatively affected by miningimpacts. In recent years, the practice of mountain-top removal and valley fill mining has resulted in thefilling and permanent burial of at least 1450 km ofsmall streams in the Appalachian Mountains. Theburial of multiple small streams destroys all aquaticlife in these streams and results in declines of sensitiveinvertebrate taxa immediately below valley fills. Thecumulative effects of burying multiple headwaterstreams on the water quality in downstream riversshould be evaluated.
Recreational Activities
Little information regarding the effects of recrea-tional activities (e.g., horseback riding, cycling, allterrain vehicle use (ATV)) on small streams has beenreported in the primary literature. One study sug-gested that populations of Dicosmeocus gilvipes, ascraping limnephilid caddisfly, in a fifth-order streamwere affected by localized disturbances associatedwith multiple recreational activities such as goldmining, streamside camping, swimming, and fishing.With growing public demand for access to undevel-oped land harboring networks of small streams forrecreational activities such as off-highway vehicleuse, there is an urgent need for more research exam-ining the impacts of such use and ways to mitigatepotential negative effects.
Global Climate Change
Consequences of global climate change on inverte-brates in small streams will vary greatly spatiallyand temporally, thus making it difficult to predictpotential effects. Generally, precipitation and evapo-ration are expected to become more variable overtime. Some regions of the world will become wetter,while others will become drier, affecting runoff pat-terns. Increased temperatures as a result of globalclimate change will reduce snow cover and also affecthydrologic patterns in small streams. Shifts in hydro-logic patterns (e.g., flooding, drying) will impacttransport of nutrients, organic matter, and habitatsavailable for colonization by benthic invertebrates.Changes in riparian vegetation may alter the qualityand quantity of detrital inputs to headwater stre-ams, thereby altering ecosystem processes (e.g.,production, respiration, organic matter breakdown)
s (2009), vol. 2, pp. 173-190

Invertebrates _ Benthic Invertebrate Fauna, Small Streams 189
within small stream reaches and longitudinally-linkeddownstream reaches, as well as invertebrate life his-tories and species composition. There is some evi-dence in the literature that the timing and durationof small stream channel drying results in alteredorganic matter standing crops, and invertebrate pro-duction. Furthermore, extended channel dryingresults in shifts from large-bodied, long-lived taxa tosmall-bodied, short-lived taxa. An experimentalmanipulation of thermal regime (2.5–3.0 �C increasein water temperature) in a small stream near Ontario,Canada, resulted in a reduction of total invertebratedensities, faster growth rates, reduced size at matu-rity, and altered sex ratios of some invertebrates.
Wildfire
Invertebrates in small streams are more susceptible tofire disturbance than those in larger streams. Intenseheating, severely altered water chemistry, and thesmothering of food resources by ash in smallerstreams can kill invertebrates directly. Over longertime periods, changes in hydrologic runoff patterns,vegetative cover, channel morphology, and sedimenttransport also affect invertebrates in fire impactedstreams. Changes in food resources over time resultin changes in the functional characteristics of themacroinvertebrate community. Initially, scraper den-sities increase following a fire because of increasedprimary productivity associated with canopy openingand increased available nutrients. As transportableorganic matter levels increase in the stream, abun-dances of collectors increase. Shredder populationsare usually the last to recover since they depend ondetrital inputs from the riparian habitat. Recovery ofmacroinvertebrate communities in intact, normallyfunctioning small streams prior to fire usually occursquickly (5–10 years) following fire disturbance andparallels the regeneration of the terrestrial vegetation.Short-lived invertebrate taxa that are trophic general-ists, and have wide habitat preferences generallyrecover quicker.
Summary and Knowledge Gaps
The functional contributions of benthic invertebratesto small streams are well known. Hundreds of inver-tebrate species may be found in a small stream. Sinceheadwater streams make up such a large proportionof total stream length in river networks, total inverte-brate production in small stream segments mayexceed that in large rivers. Invertebrates also repre-sent an important link between terrestrial and aquaticecosystems due to the close proximity of the two
Encyclopedia of Inland Wat
systems. A variety of environmental factors influencethe types and productivity of invertebrates in smallstreams. Benthic invertebrates are also good indica-tors of the health of small streams. Human and natu-ral disturbances alter typical macroinvertebrateassemblages in small streams which may have indirecteffects on higher trophic levels and small streamprocesses.
In the last two decades scientists have begun tostudy macroinvertebrate communities and the eco-logical processes affected by invertebrates along lon-gitudinal reaches spanning multiple stream orders.However, little information is known about the quan-titative and qualitative contribution of headwaterbenthic fauna to the functioning of downstream eco-systems. In some cases, entire benthic invertebratecommunities are being destroyed by burial or streampiping before the true diversity of organisms found insmall streams is known. Furthermore, with anincreasing number of disturbances that are largescale in magnitude, it is critical that scientists becomebetter able to predict threshold levels of disturbancewithin headwaters of river networks such that down-stream water quality and ecosystem functions are notirrevocably damaged.
See also: Acidification; Agriculture; Aquatic Insects –Ecology, Feeding, and Life History; Aquatic Insects,Classification; Aquatic Plants: A General Introduction;Aquatic Plants and Attached Algae; Benthic InvertebrateFauna, Small Streams; Benthic Invertebrate Fauna;Bioassessment of Aquatic Ecosystems; Biodiversity ofAquatic Ecosystems; Biological Interactions in RiverEcosystems; Climate and Rivers; Coarse Woody Debrisin Lakes and Streams; Comparative Primary Production;Conservation of Aquatic Ecosystems; Decapoda; Defor-estation and Nutrient Loading to Fresh Waters; Diptera(Biting Flies); Diptera (Non-Biting Flies); Ecology and Roleof Headwater Streams; Effects of Recreation andCommercial Shipping; Ephemeroptera (Mayflies); Fires;Flatworms (Turbellarians); Floods; Fluvial Export; GasExchange at the Air-water Interface; Gastrotricha; Geo-morphology of Streams and Rivers; Hemiptera (TrueBugs); Hydrachnida (Water Mites); Invasive Species;Isopoda (Aquatic Sowbugs); Littoral Zone; Megaloptera(Alderflies, Dobsonflies); Mercury Pollution in RemoteFreshwaters; Microbial Food Webs; Mollusca; Mosses(Bryophytes); Natural Organic Matter; Nematoda; Nema-tomorpha (Horsehair Worms); Neuston in Aquatic Eco-systems; Odonata (Dragonflies and Damselflies);Plecoptera (Stoneflies); Regulators of Biotic Processesin Stream and River Ecosystems; Restoration Ecology ofRivers; Restoration of Acidic Drainage; Riparian Zones;Streams; Subterranean Aquatic Ecosystems - Ground-water Ecology; Tardigrada (Water Bears); Trichoptera(Caddisflies); Trophic Dynamics in Aquatic Ecosystems;Urban Aquatic Ecosystems.
ers (2009), vol. 2, pp. 173-190

190 Invertebrates _ Benthic Invertebrate Fauna, Small Streams
Further Reading
Allan JD (1995) Stream Ecology: Structure and Function of Run-ning Waters. New York, NY: Chapman & Hall.
Cushing CE, Cummins KW, and Minshall GW (2006) River andStream Ecosystems of the World. Berkeley, CA: University of
California Press.Freeman MC, Pringle CM, and Jackson CR (2007) Hydrologic
connectivity and the contribution of steam headwaters to ecolog-
ical integrity at regional scales. Journal of the American WaterResources Association 43: 5–14.
Heino J, Muotka T, Mykra H, Paavola R, Hamalainen H, and
Koskenniemi E (2003) Defining macroinvertebrate assemblage
types of headwater streams: Implications for bioassessment andconservation. Ecological Applications 13: 842–852.
Huryn AD and Wallace JB (2000) Life history and production of
stream insects. Annual Review of Entomology 45: 83–110.
Merritt RW, Cummins KW, and Berg MB (2007) An Introductionto the Aquatic Insects of North America, 4th edn. Dubuque, IA:
Kendall/Hunt.
Merritt RW and Cummins KW (2006) Trophic relationships of
macroinvertebrates. In: Hauer FR and Lamberti GA (eds.)Meth-ods in Stream Ecology, 2nd edn., pp. 585–609. UK: Elsevier.
Meyer JL, Strayer DL, Wallace JB, Eggert SL, Helfman GS, and
Leonard NE (2007) The contribution of headwater streams to
biodiversity in river networks. Journal of the American WaterResources Association 43: 86–103.
Meyer JL and Wallace JB (2001) Lost linkages and lotic ecology:
Rediscovering small streams. In: PressMC,HuntlyNJ, andLevin S(eds.) Ecology: Achievement and Challenge, pp. 295–317.
Oxford, UK: Blackwell Science.
Nadeau TL and Rains MC (2007) Hydrological connectivity be-
tween headwater streams and downstream waters: How science
Encyclopedia of Inland Water
can inform policy. Journal of the American Water ResourcesAssociation 43: 118–133.
Resh VH and Rosenberg DM (1984) The Ecology of AquaticInsects. New York: Praeger.
Thorp JH and Covich AP (eds.) (2001) Ecology and Classificationof North American Freshwater Invertebrates, 2nd edn. SanDiego, CA: Academic Press.
Vannote RL, Minshall GW, Cummins KW, Sedell JR, and Cushing
CE (1980) The river continuum concept. Canadian Journal ofFisheries and Aquatic Sciences 37: 130–137.
Wallace JB andMerritt RW (1980) Filter-feeding ecology of aquatic
insects. Annual Review of Entomology 25: 103–132.
Wallace JB and Webster JR (1996) The role of macroinvertebrates
in stream ecosystem function.Annual Review of Entomology 41:115–139.
Wipfli MS, Richardson JS, and Naiman RJ (2007) Ecological lin-
kages between headwaters and downstream ecosystems: Trans-port of organic matter, invertebrates, and wood down headwater
channels. Journal of the American Water Resources Association43: 72–85.
Relevant Websites
http://www.americanrivers.org/site/DocServer/WhereRiversAre
Born1.pdf?docID ¼ 182 – American Rivers: Where Rivers are
Born.http://www.benthos.org – North American Benthological Society.
http://www.tolweb.org/tree/phylogeny.html – Tree of Life.
http://www.epa.state.oh.us/dsw/wqs/ – Ohio EPA, Division of Sur-face Water, Ohio Primary Headwater Habitat Streams.
http://www.stroudcenter.org/index.htm – Stroud Water Research
Center.
s (2009), vol. 2, pp. 173-190