bawitius, gen. nov., a giant polypterid (osteichthyes, actinopterygii) from the upper cretaceous...
TRANSCRIPT

Journal of Vertebrate Paleontology 32(1):17–26, January 2012© 2012 by the Society of Vertebrate Paleontology
ARTICLE
BAWITIUS, GEN. NOV., A GIANT POLYPTERID (OSTEICHTHYES, ACTINOPTERYGII) FROMTHE UPPER CRETACEOUS BAHARIYA FORMATION OF EGYPT
BARBARA S. GRANDSTAFF,*,1 JOSHUA B. SMITH,2 MATTHEW C. LAMANNA,3 KENNETH J. LACOVARA,4
and MEDHAT SAID ABDEL-GHANI5
1Laboratories of Anatomy, School of Veterinary Medicine, University of Pennsylvania, 3800 Spruce Street, Philadelphia,Pennsylvania 19104-6045, U.S.A., [email protected];
2American Institutes for Research, 1000 Thomas Jefferson Street NW, Washington, District of Columbia 20007-3835, U.S.A.,[email protected];
3Section of Vertebrate Paleontology, Carnegie Museum of Natural History, 4400 Forbes Avenue, Pittsburgh, Pennsylvania15213-4080, U.S.A., [email protected];
4Department of Biology, Drexel University, 32nd and Chestnut Streets, Philadelphia, Pennsylvania 19104, U.S.A.,[email protected];
5Egyptian Geological Museum, Athar El Nabi, Maadi, Cairo, Egypt, [email protected]
ABSTRACT—A newly discovered osteichthyan ectopterygoid from the Upper Cretaceous (lower Cenomanian) BahariyaFormation of the Bahariya Oasis, Egypt, is nearly identical to the holotypic specimen of Polypterus? bartheli from the samearea and geologic unit. The Bahariya ectopterygoids are referable to Polypteridae based on the presence of a robust, laterallydirected process that articulates with the maxilla. Additionally, ganoid scales from the Bahariya Formation have an isopedinelayer, a histological character of Polypteriformes; but differ from those of previously described members of this clade inhaving a discontinuous ganoin layer, rectilinear shape, and proportionally small articular processes. Both the ectopterygoidsand the scales are unusually large, and are the only polypteriform remains so far identified from the Bahariya Formation.The ectopterygoids and (tentatively) the scales are herein assigned to a single species of gigantic polypterid, the morphologyof which is sufficiently distinctive to warrant its placement in a new genus, Bawitius, gen. nov. Differences in scale anatomyand the enormous disparity in body size between Bawitius and Serenoichthys support the hypothesis (originally proposed onthe basis of varied fin spine morphologies) that a diversity of polypterid fishes inhabited North Africa during the early LateCretaceous.
INTRODUCTION
Polypteriform fishes (bichirs, reedfish, and their closest ex-tinct relatives) are classified as Actinopterygii (Jamieson, 1991;Grande and Bemis, 1996; Noack et al., 1996; Gayet et al., 2002;Inoue et al., 2003; Gardiner et al., 2005) and are positionedas a sister group to all other actinopterygians (Suzuki et al.,2010). There are two extant genera, which are both membersof Polypteridae. Polypterus Lacepede, 1803 (described but notLatinized by Geoffroy Saint-Hilaire, 1802), includes 11 validspecies (Agbayani, 2009a). Erpetoichthys Smith, 1865 (= Calam-oichthys Smith, 1866; see Swinney and Heppell, 1982), containsa single species: E. calabaricus Smith, 1865 (Agbayani, 2009b).Extant polypterids are found only in Africa, but fossil polypteri-forms are also known from South America (Gayet and Meunier,1991, 1992; Meunier and Gayet, 1996).
Polypterid autapomorphies include multiple dorsal finlets,fleshy pectoral fin bases with unique skeletal structure, andinfraorbital-maxillary fusion (Jamieson, 1991; Paxton and Es-chmeyer, 2003; Helfman et al., 2009). They are additionally char-acterized by a laterally directed process on the ectopterygoid,variously referred to as the lateral process (Allis, 1922; Schaal,1984; Grande, 2010), processus ectopterygoideus (Arratia andSchultze, 1991; Clemen et al., 1998), or labial process (Claesonet al., 2007). This process articulates with the maxilla (Traquair,1870; Gregory, 1933; Pehrson, 1947; Schaal, 1984; Clemen et al.,
*Corresponding author.
1998; Wacker et al., 2001). It will herein be referred to as thelateral process, as per Allis (1922), Schaal (1984), and Grande(2010).
Stromer (1925) was the first to report polypterid material fromthe Upper Cretaceous (lower Cenomanian) Bahariya Forma-tion of the Bahariya Oasis, Western Desert of Egypt. Weiler(1935) attributed distinctively ornamented ganoid scales from theBahariya Formation to Lepidotus (sic) aff. souzai Woodward,1908. Stromer (1936) redescribed these scales (PalaontologischeStaatssammlung Munchen [BSP] 1922 X 71, 1922 X 72, and 1922X 73a) and referred them to Polypteridae on the basis of theirhistology and gross similarity to the scales of Polypterus. He con-sidered the Bahariya scales anatomically distinct from those ofPolypterus, but did not erect a new taxon to receive them. Un-fortunately, most of the BSP fossil collection from the BahariyaFormation was destroyed during World War II (Smith et al.,2006b). A single scale, BSPG 1922 X 72a, survives in the col-lections of the Bayerische Staatssammlung fur Palaontologie undGeologie.
Nearly a half-century later, Schaal (1984) referred large scalesfrom the Bahariya Formation to Polypteridae (Bah 6/12-013 andBah 1/12-005; now catalogued as Technische Universitat Berlin[TU-B] SFB 69 Vb 328 and SFB 69 Vb 333, respectively). Likethe Bahariya scales reported on by Stromer (1925, 1936) andWeiler (1935), those described by Schaal (1984) are ornamentedwith pustules or ridges of ganoin. As had Stromer (1936), Schaal(1984) felt that the Bahariya scales might belong to a polypteridgenus that is distinct from Polypterus; however, like Stromer, hedid not place them within a new taxon.
17
Dow
nloa
ded
by [
Uni
vers
ity o
f Pe
nnsy
lvan
ia]
at 0
7:32
13
Janu
ary
2012

18 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 32, NO. 1, 2012
In addition, Schaal (1984) designated specimen Bah 5/12-016(now TU-B SFB 69 Vb 003), a large actinopterygian ectoptery-goid from Bahariya, as the holotype of Polypterus? bartheliSchaal, 1984. Schaal was able to confirm that this element be-longed to a polypterid because of the presence of a lateral pro-cess. However, because of its large size, and because of morpho-logical differences between it and the ectopterygoid of Polypterusbichir Lacepede, 1803, Schaal suspected that the Bahariya ec-topterygoid might pertain to a genus other than Polypterus.Therefore, his assignment of the Bahariya species to Polypteruswas tentative.
Recently, Smith et al. (2006a) attributed more than 300well-preserved ganoid scales from the Bahariya Formationto Polypteridae. The scales were collected by joint Egyp-tian Geological Museum–University of Pennsylvania expedi-tions to Bahariya under the auspices of the Bahariya DinosaurProject (BDP) in the winters of 2000 and 2001. Cataloguedunder CGM (Egyptian Geological Museum, Cairo) 81145 and81153, these scales were recovered from locality BDP 2000-19(28◦20.107′N latitude, 28◦59.047′E longitude; informally knownas the ‘Jon’s Birthday Site’), situated near the southwest-ern end of Gebel el Fagga in the northeastern part of theoasis basin (Fig. 1). Fossils collected from the BDP 2000-19 bone bed are dominated by fish remains, which consti-tute 1356 (85.5%) of the 1586 vertebrate fossils recoveredto date (Table 1). Polypterid remains make up the major-
FIGURE 1. Locations (indicated by stars) of polypterid discoveries inthe Bahariya Formation of the Bahariya Oasis basin. Basin floor in white;higher elevations shaded. The ectopterygoid of Bawitius bartheli, comb.nov., described herein (CGM 81151) was recovered from BDP 2000-19.Map modified from Dominik (1985). Inset (modified from Smith et al.,2006a) shows location of the Bahariya Oasis, approximately 320 kmsouthwest of Cairo, Egypt. Abbreviation: G, Gebel.
TABLE 1. Numbers of fossil vertebrate specimens recovered fromsite BDP 2000-19, with relative proportions of remains assigned to thepolypterid Bawitius bartheli, comb. nov.
Taxon Identifiable Unidentifiable Total
Selachii 3 51 54Batoidea 67 20 87Coelacanthiformes 46 0 46Dipnoi 24 0 24Polypteridae (Bawitius
bartheli)316 0 316
Osteichthyes, other 92 737 829Testudines 0 107 107Squamata 21 0 21Crocodyliformes 0 14 14Dinosauria 1 38 39Vertebrata indet. 0 49 49
Totals Identifiable Unidentifiable Total
Vertebrata 570 1016 1586Chondrichthyes +
Osteichthyes (i.e., ‘fishes’)548 808 1356
Percentages Identifiable Unidentifiable Total
Vertebrata,%Chondrichthyes +Osteichthyes
96.1% 79.5% 85.5%
Chondrichthyes +Osteichthyes,% Bawitius
57.7% N/A 23.3%
Vertebrata,% Bawitius 55.4% N/A 19.9%
Remains designated as ‘identifiable’ are those that are diagnostic to thegenus level.
ity of identifiable vertebrate material from BDP 2000-19, andthe polypterid has supplied 316 (57.7%) of 548 fish ele-ments from the site that have been identified to the genericlevel.
In addition to the scales reported by Smith et al. (2006a),polypterid fossils from BDP 2000-19 include a large leftectopterygoid, catalogued as CGM 81151. Like the holo-typic ectopterygoid of Polypterus? bartheli, this bone has awell-developed lateral process. Here we describe the new ec-topterygoid, demonstrate that it pertains to Polypterus? bartheli,and erect a new genus, Bawitius, gen. nov., to receive the species,yielding Bawitius bartheli, comb. nov. We also tentatively assignlarge polypteriform scales from Bahariya to B. bartheli, based ontheir occurrence with both known ectopterygoids, the unusuallylarge size of both ectopterygoids and scales, and the absenceto date of other polypteriform remains from the BahariyaFormation.
GEOLOGIC SETTING
The Bahariya Formation is well exposed in the Bahariya Oasisbasin, which is located approximately 320 km southwest of Cairo(Fig. 1). The formation constitutes the lower parts of the gebels(hills) within it and of the surrounding escarpment (Abdalla andEl-Bassyouni, 1985; Allam, 1986; Khalifa and Catuneanu, 2008).In the northern part of the basin, where the new polypterid mate-rial was recovered, the Bahariya Formation includes coastal sed-iments deposited during Late Cretaceous (Cenomanian) eustatichighstands of the Tethys Sea.
The Tethyan Bahariya coast developed on a gently slopingcoastal plain, producing a wide swath of paralic environmentsthat included lagoons, tidal flats and channels, oyster reefs (Exo-gyra sp.), mangals, and beaches (Abdalla and El-Bassyouni, 1985;Soliman and Khalifa, 1993; Smith et al., 2001; El Sisi et al., 2002;
Dow
nloa
ded
by [
Uni
vers
ity o
f Pe
nnsy
lvan
ia]
at 0
7:32
13
Janu
ary
2012
GRANDSTAFF ET AL.—GIANT CRETACEOUS POLYPTERID FROM EGYPT 19
Lacovara et al., 2003; Tanner and Khalifa, 2010). Fluvial depositshave also been described from the Bahariya Formation (Franks,1982; El Sisi et al., 2002; Catuneanu et al., 2006; Khalifa andCatuneanu, 2008), particularly in the southern part of the oasiswhere the polypterid material described by Schaal (1984) was col-lected.
The littoral zone was lined by a chain of low-lying sediment-starved transgressive barrier islands, separated by tidal inlets.Intertidal environments away from the surf zone were domi-nated by the halophytic tree fern Weichselia reticulata (Stokesand Webb, 1824) Fontaine, 1899 (Dominik, 1985; Smith et al.,2001; Lacovara et al., 2003). Tidal bundles composed of muddrapes alternating with angular foreset beds show neap-springtidal cyclicity at a number of sites, including BDP 2000-19. Theserhythmites record cycles that correspond to semi-diurnal tides(Lacovara et al., 2002a).
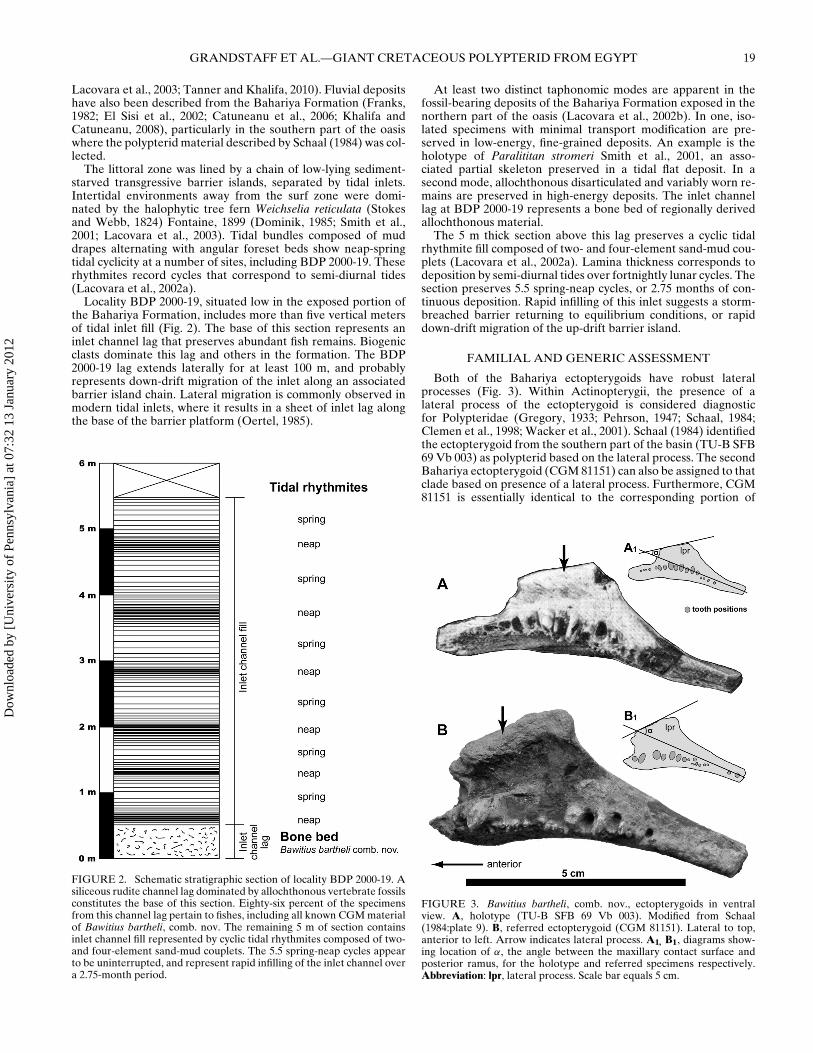
Locality BDP 2000-19, situated low in the exposed portion ofthe Bahariya Formation, includes more than five vertical metersof tidal inlet fill (Fig. 2). The base of this section represents aninlet channel lag that preserves abundant fish remains. Biogenicclasts dominate this lag and others in the formation. The BDP2000-19 lag extends laterally for at least 100 m, and probablyrepresents down-drift migration of the inlet along an associatedbarrier island chain. Lateral migration is commonly observed inmodern tidal inlets, where it results in a sheet of inlet lag alongthe base of the barrier platform (Oertel, 1985).
FIGURE 2. Schematic stratigraphic section of locality BDP 2000-19. Asiliceous rudite channel lag dominated by allochthonous vertebrate fossilsconstitutes the base of this section. Eighty-six percent of the specimensfrom this channel lag pertain to fishes, including all known CGM materialof Bawitius bartheli, comb. nov. The remaining 5 m of section containsinlet channel fill represented by cyclic tidal rhythmites composed of two-and four-element sand-mud couplets. The 5.5 spring-neap cycles appearto be uninterrupted, and represent rapid infilling of the inlet channel overa 2.75-month period.
At least two distinct taphonomic modes are apparent in thefossil-bearing deposits of the Bahariya Formation exposed in thenorthern part of the oasis (Lacovara et al., 2002b). In one, iso-lated specimens with minimal transport modification are pre-served in low-energy, fine-grained deposits. An example is theholotype of Paralititan stromeri Smith et al., 2001, an asso-ciated partial skeleton preserved in a tidal flat deposit. In asecond mode, allochthonous disarticulated and variably worn re-mains are preserved in high-energy deposits. The inlet channellag at BDP 2000-19 represents a bone bed of regionally derivedallochthonous material.
The 5 m thick section above this lag preserves a cyclic tidalrhythmite fill composed of two- and four-element sand-mud cou-plets (Lacovara et al., 2002a). Lamina thickness corresponds todeposition by semi-diurnal tides over fortnightly lunar cycles. Thesection preserves 5.5 spring-neap cycles, or 2.75 months of con-tinuous deposition. Rapid infilling of this inlet suggests a storm-breached barrier returning to equilibrium conditions, or rapiddown-drift migration of the up-drift barrier island.
FAMILIAL AND GENERIC ASSESSMENT
Both of the Bahariya ectopterygoids have robust lateralprocesses (Fig. 3). Within Actinopterygii, the presence of alateral process of the ectopterygoid is considered diagnosticfor Polypteridae (Gregory, 1933; Pehrson, 1947; Schaal, 1984;Clemen et al., 1998; Wacker et al., 2001). Schaal (1984) identifiedthe ectopterygoid from the southern part of the basin (TU-B SFB69 Vb 003) as polypterid based on the lateral process. The secondBahariya ectopterygoid (CGM 81151) can also be assigned to thatclade based on presence of a lateral process. Furthermore, CGM81151 is essentially identical to the corresponding portion of
FIGURE 3. Bawitius bartheli, comb. nov., ectopterygoids in ventralview. A, holotype (TU-B SFB 69 Vb 003). Modified from Schaal(1984:plate 9). B, referred ectopterygoid (CGM 81151). Lateral to top,anterior to left. Arrow indicates lateral process. A1, B1, diagrams show-ing location of α, the angle between the maxillary contact surface andposterior ramus, for the holotype and referred specimens respectively.Abbreviation: lpr, lateral process. Scale bar equals 5 cm.
Dow
nloa
ded
by [
Uni
vers
ity o
f Pe
nnsy
lvan
ia]
at 0
7:32
13
Janu
ary
2012
20 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 32, NO. 1, 2012
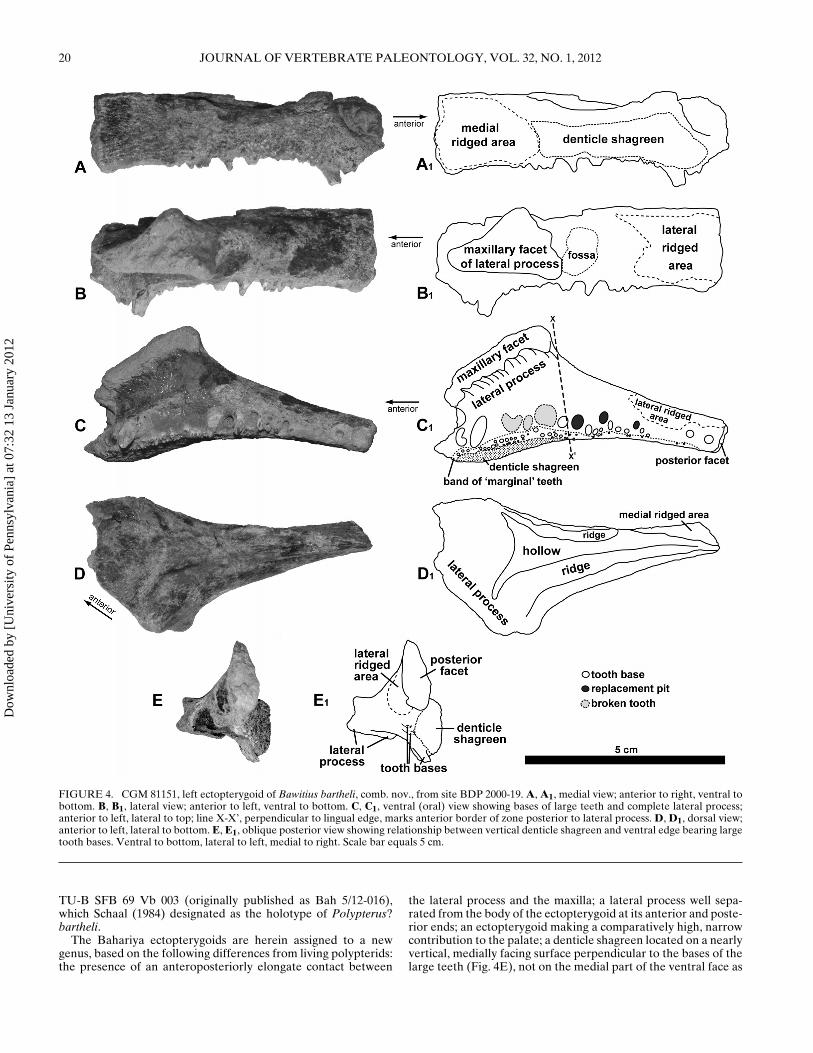
FIGURE 4. CGM 81151, left ectopterygoid of Bawitius bartheli, comb. nov., from site BDP 2000-19. A, A1, medial view; anterior to right, ventral tobottom. B, B1, lateral view; anterior to left, ventral to bottom. C, C1, ventral (oral) view showing bases of large teeth and complete lateral process;anterior to left, lateral to top; line X-X’, perpendicular to lingual edge, marks anterior border of zone posterior to lateral process. D, D1, dorsal view;anterior to left, lateral to bottom. E, E1, oblique posterior view showing relationship between vertical denticle shagreen and ventral edge bearing largetooth bases. Ventral to bottom, lateral to left, medial to right. Scale bar equals 5 cm.
TU-B SFB 69 Vb 003 (originally published as Bah 5/12-016),which Schaal (1984) designated as the holotype of Polypterus?bartheli.
The Bahariya ectopterygoids are herein assigned to a newgenus, based on the following differences from living polypterids:the presence of an anteroposteriorly elongate contact between
the lateral process and the maxilla; a lateral process well sepa-rated from the body of the ectopterygoid at its anterior and poste-rior ends; an ectopterygoid making a comparatively high, narrowcontribution to the palate; a denticle shagreen located on a nearlyvertical, medially facing surface perpendicular to the bases of thelarge teeth (Fig. 4E), not on the medial part of the ventral face as
Dow
nloa
ded
by [
Uni
vers
ity o
f Pe
nnsy
lvan
ia]
at 0
7:32
13
Janu
ary
2012

GRANDSTAFF ET AL.—GIANT CRETACEOUS POLYPTERID FROM EGYPT 21
in Polypterus senegalus Cuvier, 1829, P. bichir, and Erpetoichthys(Traquair, 1870; Allis, 1922; Clemen et al., 1998; Claeson et al.,2007); and an ectopterygoid narrowing posterior to the lateralprocess, rather than widening posterior to this process as in P.bichir (Traquair, 1870; Allis, 1922), P. senegalus (Clemen et al.,1998), and Erpetoichthys (Claeson et al., 2007). The morphologyof the Bahariya ectopterygoids is sufficiently distinct from thatof Polypterus and Erpetoichthys to justify their separation at thegeneric level.
SYSTEMATIC PALEONTOLOGY
OSTEICHTHYES Huxley, 1880ACTINOPTERYGII Cope, 1887
CLADISTIA Cope, 1871POLYPTERIFORMES Bleeker, 1859
POLYPTERIDAE Gunther, 1870BAWITIUS, gen. nov.
Etymology—For Bawiti, the largest population center in theBahariya Oasis basin.
Diagnosis—As for the type and only known species.
BAWITIUS BARTHELI (Schaal, 1984), comb. nov.
Polypterus? bartheli Schaal, 1984:51, fig. 25, pl. IX 1–3 (originaldescription).
Holotype—TU-B SFB 69 Vb 003 (identified by Schaal as Bah5/12-016), an isolated ectopterygoid (Fig. 3A). Although origi-nally described as being from the right side (Schaal, 1984), thisbone is actually a left ectopterygoid.
Referred Specimens—CGM 81151, an isolated left ectoptery-goid missing its anterior and dorsal portions (Figs. 3B, 4). In ad-dition, 315 isolated polypterid scales (Smith et al., 2006a), CGM81145 and CGM 81153 (Fig. 5A–D), are tentatively referred toBawitius bartheli, as are Bahariya polypterid scales described byWeiler (1935) and Stromer (1936) (BSP 1922 X 71, 1922 X 72,BSP 1922 X 73a), Schaal (1984) (TU-B SFB 69 Vb 328, TU-BSFB 69 Vb 333) and BSPG 1922 X 72a, the surviving scale fromStromer’s collection (Fig. 5E).
Type Locality—Bahariya Oasis basin, Western Desert ofEgypt (escarpment west of Ghorbiya [28◦05′N latitude, 28◦15′Elongitude; Schaal, 1984]) (Fig. 1).
Type Horizon—Bahariya Formation (Upper Cretaceous:Cenomanian).
Revised Diagnosis—Ectopterygoid with at least 14 teeth inmain tooth row; single row of large teeth extending to posteriorend of bone; largest teeth oval in cross-section and located me-dial to base of lateral process on narrow ventral (oral) face; an-terior end of medial face with shagreen of denticles; largest den-ticles appearing as narrow band of ventrally pointing marginalteeth, with band extending to posterior end of ectopterygoid; ro-bust lateral process that contacts maxilla; lateral process taper-ing only slightly toward its lateral end, lending it a rectilinearoutline in dorsal/ventral view; lateral end of process (maxillaryfacet) rough, elongate, triangular in lateral view; shallow fossaon lateral face just posterior to lateral process; ectopterygoid an-gled medially (i.e., away from maxilla) posterior to lateral pro-cess; gigantic body size (ectopterygoid ∼500% larger than that ofPolypterus). The presence of these diagnostic characters in bothBahariya ectopterygoids justifies their inclusion within a singlepolypterid taxon, Bawitius bartheli, comb. nov.
Proposed autapomorphies of B. bartheli include: ectopterygoidwith single row of large teeth extending to posterior end, in con-trast to the shorter rows of large teeth seen in extant polypterids(Traquair, 1870; Allis, 1922; Clemen et al., 1998); ectoptery-goid lateral process robust, elongate, and only slightly tapered,in contrast to the short, slender lateral process seen in extantpolypterids (Traquair, 1870; Allis, 1922; Arratia and Schultze,
FIGURE 5. Examples of scales tentatively referred to Bawitius bartheli,comb. nov. A–D, scales (catalogued under CGM 81153) from site BDP2000-19. A, D, scales from the front part of the body. B, C, scales fromthe back part of the body. Scales from the rear part of the body (B, C)have wide, squared anterior processes; short articular pegs are visible onscales shown in A and C. E, surviving scale from the Stromer collection(BSPG 1922 X 72a), showing both dorsal and ventral anterior processesand a short articular peg. Photography courtesy of A. Lopez-Arbarello.Dorsal to top for A, B, E; dorsal to bottom for C, D. Anterior to left forall scales. Abbreviations: ant. proc., anterior process; artic. peg, articularpeg. Scale bar equals 1 cm.
1991; Clemen et al., 1998; Claeson et al., 2007; Grande, 2010);shallow fossa on lateral face of ectopterygoid posterior to lat-eral process, unlike the “weak furrows” seen in extant Polypterus(Schaal, 1984:51); and posterior ramus of ectopterygoid angledmedially posterior to lateral process, rather than being essentiallyparallel to the cheek as in Polypterus (Traquair, 1870; Clemenet al., 1998) or Erpetoichthys (Claeson et al., 2007).
The two Bawitius ectopterygoids additionally share the follow-ing characteristics (of unclear taxonomic significance) that do notoccur in other polypterids for which this element is known: atleast 14 teeth in the main tooth row, the largest of which lie lin-gual to the lateral process and are oval in cross-section; anteriorpart of medial face bearing a denticle shagreen extending in aband along the ventral edge of the medial face, with the largestdenticles situated near the ventral edge and appearing as a sin-gle row of marginal teeth; and a roughened lateral facet for con-tact with the maxilla. This last character resembles the “flat andsomewhat spreading” lateral end of the lateral process reportedin Polypterus bichir (Allis, 1922:244). In the B. bartheli holo-type TU-B SFB 69 Vb 003, the medial face continues as a thinsheet of bone extending dorsally above the denticle shagreen.The Bahariya ectopterygoids are also characterized by giganticsize (Schaal [1984] observed that the Bawitius holotype is 500%larger than the ectopterygoid of Polypterus).
Dow
nloa
ded
by [
Uni
vers
ity o
f Pe
nnsy
lvan
ia]
at 0
7:32
13
Janu
ary
2012

22 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 32, NO. 1, 2012
TABLE 2. Measurements (mm) of CGM 81151, a left ectopterygoid ofBawitius bartheli, comb. nov., from locality BDP 2000-19.
Parameter measured mm
Anteroposterior length, maximum preserved 66∗Dorsoventral height, maximum preserved 19∗Mediolateral width, maximum (at lateral process) 30Anteroposterior length of lateral process 29Anterior dorsoventral height, maxillary facet of lateral process 5Posterior dorsoventral height, maxillary facet of lateral process 14Tooth basoapical height, maximum preserved 8∗
Measurements marked with an asterisk (∗) are ‘as preserved’ (measure-ment incomplete).
Description of New Material—The new polypterid ectoptery-goid from the Bahariya Formation, CGM 81151 (Figs. 3B, 4), iswell preserved but incomplete anteriorly and dorsally. Only thethicker ventral part of the ectopterygoid is preserved; the thinsheet of bone dorsal to the medial tooth shagreen in the holo-type is not preserved in CGM 81151. Measurements of the speci-men are provided in Table 2. The most prominent feature of theectopterygoid is the robust lateral process. This process is rec-tilinear in dorsal or ventral view, and well separated from thebody of the ectopterygoid at its anterior and posterior edges. Ittapers only slightly toward its lateral end, which bears an elon-gate facet that would have contacted the maxilla. The maxillaryfacet is triangular in lateral view, highest posteriorly, and gentlyconvex laterally in dorsal or ventral view. The lateral surface ofthe maxillary facet is rough, indicating that the joint between ec-topterygoid and maxilla was fibrous or cartilaginous rather thansynovial. The ectopterygoid tapers posterior to the lateral pro-cess in dorsal or ventral view. The narrow posterior end of theectopterygoid terminates in a flat, slightly roughened, posteriorlyfacing vertical surface.
There is a poorly preserved ridged region located posteriorlyon the lateral face of CGM 81151 (Fig. 4B, B1). This region sug-gests contact with a more laterally positioned bone or bones (pre-sumably the metapterygoid or quadrate). The angle between themaxillary contact surface at the end of the lateral process and theaxis of the posterior ramus (α in Fig. 3A1, B1; 35◦ in the holo-type, 49◦ in CGM 81151) suggests that there was an open spacebetween the ectopterygoid and maxilla posterior to their articula-tion in Bawitius. In living polypterids, this space, bounded later-ally by the preopercular (Claeson et al., 2007; the ‘dermal cheek’of Allis [1922] and Traquair [1870]), accommodates the jaw ad-ductors and coronoid process of the mandible.
The medial edge of the ventral face of CGM 81151 preservesthe bases of 13 conical teeth of varying sizes, plus three replace-ment pits and three positions where large teeth have completelybroken away. As preserved, there are thus 19 tooth positions onthe new ectopterygoid. The tooth row arches lingually in ventralview, following the curvature of the medial face. One tooth baselies mesial to the lateral process. Two preserved tooth bases andthree places where large teeth have broken away lie lingual tothe lateral process. Ten teeth and three replacement pits (13tooth positions) lie posterior to a line drawn perpendicular to themedial face of the ectopterygoid and passing just posterior to themaxillary facet (see Fig. 4C1, X-X′). The distal-most tooth liesjust anterior to the posterior end of the bone. The largest toothpreserved in CGM 81151 is 8 mm in basoapical height, is missingat least its apex, and is positioned lingual to the anterior edge ofthe lateral process. The surfaces of all preserved tooth bases areworn, and show no evidence of striations or facets. The row oflarge teeth is paralleled lingually by a row of smaller, marginalteeth.
The anterior half of the vertical medial face is covered by a sha-green of small, rounded, unornamented denticles, each of which
has a distinctly pointed, lingually facing apex (giving each den-ticle an ‘onion dome’ shape). The shagreen is deepest near itsmesial end (Fig. 4A, A1), and tapers toward the midpoint of theectopterygoid as preserved, ending just posterior to the plane ofthe posterior edge of the lateral process. The marginal teeth liealong the edge of this shagreen and are not well separated fromit, but are oriented so that their apices point ventrally. This nar-row band of marginal teeth extends along the entire medial edgeof the ventral face, nearly reaching the posterior end of the ec-topterygoid.
The medial face of CGM 81151 posterior to the denticle sha-green is ridged (Fig. 4A, A1), suggesting that the posterior por-tion of the medial face articulated via a fibrous or scarf jointwith a more medial element in the palate, possibly the entoptery-goid. The flat, roughened posterior end of CGM 81151 will havecontacted a bone of the suspensorium, such as the quadrate ormetapterygoid.
Total preserved length of CGM 81151 is 66 mm (Table 2),as compared to 75 mm in TU-B SFB 69 Vb 003 (Schaal, 1984).The latter is the more anteroposteriorly complete of the two ec-topterygoids (Fig. 3). The two Bawitius individuals to which theseectopterygoids pertained were comparable in body size.
Articular Relationships of the Ectopterygoid in Bawitius
In Polypterus and Erpetoichthys, the ectopterygoid con-tacts the entopterygoid and metapterygoid medially, and themetapterygoid and quadrate posteriorly (Fig. 6). The entoptery-goid contacts most of the medial edge of the ectopterygoid in ex-tant polypterids. The posterior part of the medial edge makes anoblique contact with the metapterygoid. The posteroventral tipof the ectopterygoid contacts the quadrate (Traquair, 1870; Al-lis, 1922; Arratia and Schultze, 1991; Clemen et al., 1998; Clae-son et al., 2007; Grande, 2010). The quadrate may contact the
FIGURE 6. Palatal elements of Polypterus bichir in ventral view,modified from Traquair (1870:plate 6, fig. 5). Note lateral process of ec-topterygoid that contacts maxilla. Large teeth shown by black circles; der-mopalatine, ectopterygoid, entopterygoid, parasphenoid, and vomer allbear shagreens of smaller teeth (not shown). Left ectopterygoid shadedto highlight location of known Bawitius ectopterygoids relative to otherbones in palate. Areas of palate occupied by soft tissues or cartilageshaded light grey. Anterior to the left. Abbreviations: bo, basioccipital;ck, cheek element (‘posterior maxillary segment’ of Grande, 2010); dp,dermopalatine; ect, ectopterygoid; enpt, entopterygoid; hm, hyomandibu-lar; lat.proc, lateral process of ectopterygoid; mpt, metapterygoid; ps,parasphenoid; qu, quadrate; vo, vomer. Scale bar equals 5 cm.
Dow
nloa
ded
by [
Uni
vers
ity o
f Pe
nnsy
lvan
ia]
at 0
7:32
13
Janu
ary
2012

GRANDSTAFF ET AL.—GIANT CRETACEOUS POLYPTERID FROM EGYPT 23
posterior part of the lateral face of the ectopterygoid (Arratiaand Schultze, 1991; Grande, 2010). As described above, the me-dial, lateral, and posterior surfaces of the posterior ramus of theectopterygoid in Bawitius bartheli bear facets for contact withother skull bones. We suspect the entopterygoid or metaptery-goid contacted the Bawitius ectopterygoid at the medially fac-ing scarf facet; articular relationships in extant polypterids sug-gest contact with the metapterygoid is more likely. The narrowposterior facet of the Bawitius ectopterygoid probably contactedonly the metapterygoid. We consider it likely that the quadrateof Bawitius articulated mainly with the ridged area on the lateralside of the ectopterygoid posterior to the lateral process ratherthan with the posterior end of the ectopterygoid. The Bawitiusectopterygoid differs from extant polypterids in having a rela-tively larger lateral contact with the quadrate.
As also noted above, the angle α (Fig. 3A1, B1) between themaxillary facet and the posterior ramus of the ectopterygoid (35◦
to 49◦ based on the two available specimens) suggests a sub-stantial gap between the posterior portions of the ectopterygoidand maxilla in Bawitius. In Polypterus, the space between theectopterygoid and maxilla posterior to the lateral process ac-commodates the ascending process of the prearticular, which fitsbehind the lateral process as the jaws close (Allis, 1922). Thisspace is also occupied by mandibular adductor muscles (M. ad-ductor mandibulae, M. temporalis, and M. pterygoideus), whichinsert on the medial aspect of the prearticular (Allis, 1922). In B.bartheli, the large space between the posterior parts of the max-illa and ectopterygoid would have allowed for robust mandibularadductors as well as the ascending process of the prearticular. Itwill also have accommodated the quadrate.
Additional Polypterid Material from Bahariya
Ganoid scales of distinctive morphology (Fig. 5) were recov-ered with CGM 81151, and have also been found elsewhere in theBahariya Formation (Stromer, 1925, 1936; Weiler, 1935; Schaal,1984). These scales exhibit the following autapomorphic char-acters: an incomplete ganoin layer expressed as highly variableridges and bosses on the free portion of the scale, in contrast tothe complete ganoin layer typical of most basal ray-finned fishes(Schultze, 1996); a generally rectilinear shape, with the anteriorand posterior edges nearly perpendicular to the dorsal and ven-tral edges, in contrast with the more rhomboidal scale shape inextant polypterids (Gemballa and Bartsch, 2002); scales from thefront part of the body may have a second, ventrally located an-terior process; the anterior process is located at the center of theanterior edge in some scales; the anterior process of scales fromthe rear part of the body is short and broad, in contrast to thenarrower and more pointed anterior processes in scales of extantpolypterids; and the articular pegs are short and articular socketscorrespondingly shallow on all scales.
The scales have an isopedine layer (sensu Daget et al., 2001;non Schultze, 1996), visible in thin-section under cross-polarizedlight as thin, alternating light and dark bands in the central area ofthe scale, between the basal bony plate and the overlying dentine(Smith et al., 2006a). The presence of isopedine in this position isconsidered a synapomorphy of Polypteriformes (Gayet and Me-unier, 1992; Daget et al., 2001; Sire et al., 2009). The Bahariyascales can confidently be referred to this clade, as first contendedby Stromer (1925, 1936).
Morphology of all known polypterid scales from the BahariyaFormation falls within the variations of scale morphology in dif-ferent regions of the body seen in Polypterus (Gemballa andBartsch, 2002), and is consistent with their belonging to a singlespecies. The only polypterid species yet identified in this forma-tion is Bawitius bartheli, a taxon definitively known only from ec-topterygoids. However, both ectopterygoids and scales suggesta fish of unusually large size (∼2.5–3 m in total body length;
Schaal, 1984), and it seems reasonable that only one species of gi-ant polypterid might have inhabited the paralic environment pre-served by the Bahariya Formation. Consequently, we tentativelyrefer all polypterid scales from this formation to B. bartheli.
The scales of the Bahariya taxon differ from those of mostother polypterids in bearing a discontinuous ganoin layer on theirfree surface. In B. bartheli, ganoin forms a highly variable orna-ment of ridges and bosses; the ganoin elevations are separatedby ganoin-free areas (Smith et al., 2006a). Although the ganoin-free areas appear to have formed through ganoin resorption,Bawitius lacks the evidence of ganoin remodeling reported inthe semionotid Lepidotyle enigmatica Meunier and Gayet, 1992.There is no evidence of secondary ganoin deposition in Bawitius,nor is there any evidence that ganoin resorption was cyclical.
The single articular peg in polypterid scales from Bahariya Oa-sis is located just posterior to the area of the scale covered by theadjacent scale. All Bahariya scales studied to date, even thosewith proportionally large anterior processes, have relatively smallarticular pegs. By contrast, in living Polypterus species scalesfrom the front part of the body, which have large anterior pro-cesses, also have relatively large articular pegs (Gemballa andBartsch, 2002). Bawitius scales from the back part of the bodyhave short, broad, rectilinear anterior processes; in Polypterus,the anterior processes of scales from all parts of the body aretapering, not rectilinear. Some Bahariya polypterid scales haveboth dorsal and ventral anterior processes (Fig. 5E) instead ofa single dorsal anterior process as in Polypterus. Among thebasal actinopterygian taxa studied by Cavin et al. (2009), onlysemionotids have a second, ventral anterior process. However,polypteriforms did not occur in the faunas studied by Cavin et al.(2009). The Bahariya scales, even those with two anterior pro-cesses, have a dentine layer (Stromer, 1936; Smith et al., 2006a)and are therefore histologically distinct from semionotid scales(Meunier, 2011).
DISCUSSION
Stratigraphic Distribution of Bahariya Polypterid Fossils
In geographic terms, polypterid material from the BahariyaFormation is widely distributed within the Bahariya Oasis basin(Fig. 1). Regrettably, stratigraphic relationships of these remainsare difficult to determine. The Bahariya Formation exhibitsconsiderable lithologic variability (Stromer, 1914; Dominik,1985; Lacovara et al., 2003; Catuneanu et al., 2006; Khalifa andCatuneanu, 2008), and stratigraphic sections from different out-crops (some preserved as isolated erosional remnants) may bearlittle resemblance to one another (see, for example, the sectionspublished by Catuneanu et al., 2006). Weiler (1935) reportedthat polypterid scales (erroneously identified as Lepidotus [sic]aff. souzai) were found in Stromer’s (1914) ‘layer n’ and ‘layero,’ at the base of Gebel el Dist near the northern margin of theoasis basin, and from the basin floor approximately 18 km eastof Ain Harra. According to Stromer (1936:45), the latter scaleswere actually recovered 10 km north of Ain Harra, probablyfrom his (1914) ‘layer e.’ Stromer (1936) reported multiplepolypterid scales from his (1914) ‘layer n’ near Gebel el Dist, andmentioned, but did not describe, indeterminate polypterid bonesfrom the same area (Stromer, 1936:23). He additionally reportedthat at least six polypterid scales were collected from the base ofGebel Maghrafa.
The holotypic ectopterygoid of Bawitius bartheli was collectedfrom the escarpment west of Ghorbiya, from Schaal’s (1984)‘stratum 3,’ which lies some 35–57 m above the base of the Ba-hariya Formation as it is exposed at Gebel el Dist (Schaal, 1984).Polypterid scales were also found in his ‘stratum 3,’ and rarelyin ‘stratum 6.’ Schaal (1984:54) stated that the scales describedby Weiler (1935) as Lepidotus (sic) aff. souzai, later reidenti-fied as polypterid by Stromer (1936), are also from ‘stratum 3’
Dow
nloa
ded
by [
Uni
vers
ity o
f Pe
nnsy
lvan
ia]
at 0
7:32
13
Janu
ary
2012

24 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 32, NO. 1, 2012
(as used by Schaal), thus establishing that at least some scalesfrom the northern part of the oasis come from the same levelof the Bahariya Formation as does the holotypic ectopterygoid.Unfortunately, we have not been able to determine either theprecise stratigraphic relationships of the polypterid-bearing lay-ers described by Stromer (1914, 1936), Weiler (1935), and Schaal(1984) to the bone bed at BDP 2000-19, or their relationships toeach other.
Late Cretaceous Polypterid Diversity in North Africa
Due to a dearth of overlapping material, the relationship ofBawitius bartheli to other Late Cretaceous polypterids fromNorth Africa remains unclear. The numerous species fromthe Cenomanian Wadi Milk Formation of Sudan and/orthe ?Coniacian-Santonian Ibeceten Formation of Niger(Bartschichthys arnoulti Gayet and Meunier, 1996; B. nap-atensis Werner and Gayet, 1997; B. tubularis Gayet and Meunier,1996; Inbecetemia torta Gayet and Meunier, 1996; I. tortissimaGayet and Meunier, 1996; Nagaia extrema Werner and Gayet,1997; Polypterus dageti Gayet and Meunier, 1996; P. sudanensisWerner and Gayet, 1997; Saharichthys africanus Gayet and Me-unier, 1996; S. nigeriensis Gayet and Meunier, 1996; Sainthilairiabeccusiformis Gayet and Meunier, 1996; S. elongata Werner andGayet, 1997; S. falciformis Gayet and Meunier, 1996; S. grandisGayet and Meunier, 1996; S. intermedia Werner and Gayet, 1997;Sudania gracilis Werner and Gayet, 1997; S. oblonga Werner andGayet, 1997) are diagnosed on the basis of fin spines (Werner,1991; Gayet and Meunier, 1996; Gayet et al., 1997, 2002; Wernerand Gayet, 1997), which have not yet been identified for theBahariya polypterid. The very small, stout-bodied polypteridSerenoichthys kemkemensis Dutheil, 1999a, is known from twolargely complete, articulated specimens from the CenomanianKem Kem beds of Morocco, but not yet from substantial cranialmaterial (Dutheil, 1999a, 1999b). Two other, as yet undescribedcladistian taxa have also been recovered from the upper KemKem beds (Dutheil, 2009; Cavin et al., 2010).
The scales of Serenoichthys have continuous ganoin, therebydiffering from those attributed to Bawitius. Known material ofthe two forms differs dramatically in size—for example, the ec-topterygoid of Bawitius is very probably longer than the entirebody of Serenoichthys (Table 2; Schaal, 1984; Dutheil, 1999a).It is unlikely that the diminutive size of known specimens ofthe latter is due to ontogenetic immaturity: as stated by Dutheil(1999a:245), “The presence of branched fin rays in the caudal finsuggests that, despite its small size, the specimens of S. kemke-mensis were post juveniles. . .” Bawitius and Serenoichthys there-fore appear distinct, even though it is not presently possible tocompare their cranial elements. Coniacian-Santonian polypteridscales from In Becetem in Niger have continuous ganoin (Gayetet al., 1988), thereby also differing from the Bahariya scales ten-tatively attributed to Bawitius.
With the addition of Bawitius, there are now nine gen-era (Bartschichthys, Bawitius, Inbecetemia, Nagaia, Polypterus,Saharichthys, Sainthilairia, Serenoichthys, and Sudania) ofPolypteridae known from an approximately 16 million-year span(Cenomanian-Santonian, 99.6–83.5 Ma; Ogg et al., 2008) of theLate Cretaceous in continental Africa, as compared to onlytwo genera living today (Erpetoichthys and Polypterus). Further-more, the extreme size disparity between some taxa (e.g., Baw-itius [∼250–300 cm in estimated total length; Schaal, 1984; thispaper] vs. Serenoichthys [∼6 cm total length; estimated fromDutheil, 1999a]) and the aberrant fin spine morphology of oth-ers (e.g., Inbecetemia, Nagaia; Gayet et al., 1997) indicates thatthe Cretaceous forms were anatomically diverse. It appears that,in both taxonomic and morphological terms, Late Cretaceouspolypterids were considerably more diverse than are their extantrelatives. The presence of two additional polypteriform genera
in South America (Gayet and Meunier, 1991, 1992; Meunier andGayet, 1996) further attests to this Late Cretaceous diversity.
Additional analyses of articulated or associated specimens areneeded before a more complete understanding of Late Creta-ceous polypterid diversity may be attained. As noted by Dutheil(1999a:245), studies of intraspecific variability of fin spines will becrucial to evaluating validity of the Cretaceous taxa from Nigerand Sudan based on these elements. Only studies of as yet undis-covered material preserving fin spines in association with the ec-topterygoid may potentially decipher the relationships of Baw-itius to these taxa.
Along with the eight genera currently known from the Ceno-manian of Morocco and Sudan (Bartschichthys, Inbecetemia, Na-gaia, Polypterus, Saharichthys, Sainthilairia, Serenoichthys, andSudania; Werner and Gayet, 1997; Dutheil, 1999a), Bawitius isamong the most ancient representatives of Polypteridae yet dis-covered. Interestingly, very large polypterid scales are knownfrom Eocene-aged beds elsewhere in Egypt (Stromer, 1905, 1925,1936; Schaal, 1984; Murray et al., 2010), suggesting that giganticpolypterids survived into the early Cenozoic in northern Africa.
ACKNOWLEDGMENTS
We thank P. Dodson, J. Poole, R. Giegengack, A. Swedan,K. Soleiman, Y. Attia, Y. Abdelrazik, J. R. Smith, J. Caton, A.Tumarkin-Deratzian, J. Harris, S. Kurth, P. Kane-Vanni, J. La-covara, K. Johnson, D. Nichols, M. Lyon, J. Losos, W. Dominik,P. Luger, X. Sun, A. Lopez-Arbarello, the Egyptian Mineral Re-sources Authority (formerly the Egyptian Geological Survey andMining Authority), and the El Beshmo Lodge for invaluable as-sistance both in and out of the field. We thank M. Friedman,A. Berta, and P. Barrett for editorial guidance, and A. Mur-ray and L. Cavin for their helpful reviews of an earlier versionof the manuscript. The BDP has been supported by grants toJ.B.S. from Cosmos Studios, MPH Entertainment, the Univer-sity of Pennsylvania, the Andrew W. Mellon Foundation, andthe Delaware Valley Paleontological Society, and to P. Dodson(Penn) from the late E. de Hellebranth and the University ofPennsylvania. This paper is BDP contribution 7.
LITERATURE CITED
Abdalla, A. Y., and A. A. El-Bassyouni. 1985. Primary sedimentarystructures and sedimentary environment of the Bahariya Formation‘Lower Cenomanian’, Bahariya Oasis, Egypt. Annals of the Geolog-ical Survey of Egypt 15:267–274.
Agbayani, E. 2009a. Valid species of the genus Polypterus. FishBase.Available at www.fishbase.org. Accessed November 24, 2009.
Agbayani, E. 2009b. Valid species of the genus Erpetoichthys. FishBase.Available at www.fishbase.org. Accessed November 24, 2009.
Allam, A. M. 1986. A regional and paleoenvironmental study on theUpper Cretaceous deposits of the Bahariya Oasis, Libyan Desert,Egypt. Journal of African Earth Sciences 5:407–412.
Allis, E. P. J. 1922. The cranial anatomy of Polypterus, with special refer-ence to Polypterus bichir. Journal of Anatomy 56:189–294.
Arratia, G., and H.-P. Schultze. 1991. Palatoquadrate and its ossifications:development and homology within osteichthyans. Journal of Mor-phology 208:1–81.
Bleeker, P. 1859. Enumeratio specierum piscium hucusque inArchipelago Indico observatarum, adjectis habitationibus cita-tionibusque, ubi descriptiones earum recentiores reperiuntur,nec non speciebus Musei Bleekeriani Bengalensibus, Japonicis,Capensibus, Tasmanicisque. Acta Societatis Regiae ScientiarumIndo-Neerlandicae 6:1–276.
Catuneanu, O., M. A. Khalifa, and H. A. Wanas. 2006. Sequence stratig-raphy of the Lower Cenomanian Bahariya Formation, Bahariya Oa-sis, Western Desert, Egypt. Sedimentary Geology 190:121–137.
Cavin, L., U. Deesri, and V. Suteethorn. 2009. The Jurassic and Creta-ceous bony fish record (Actinopterygii, Dipnoi) from Thailand; pp.125–139 in E. Buffetaut, G. Cuny, J. Le Leouff, and B. Suteethorn(eds.), Late Palaeozoic and Mesozoic Ecosystems in SE Asia.
Dow
nloa
ded
by [
Uni
vers
ity o
f Pe
nnsy
lvan
ia]
at 0
7:32
13
Janu
ary
2012

GRANDSTAFF ET AL.—GIANT CRETACEOUS POLYPTERID FROM EGYPT 25
Geological Society Special Publication 315. Geological Society,London, England.
Cavin, L., H. Tong, L. Boudad, C. Meister, A. Piuz, J. Tabouelle, M.Aarab, R. Amiot, E. Buffetaut, G. Dyke, S. Hua, and J. Le Loeuff.2010. Vertebrate assemblages from the early Late Cretaceous ofsoutheastern Morocco: an overview. Journal of African Earth Sci-ences 57:391–412.
Claeson, K. M., W. E. Bemis, and J. W. Hagadorn. 2007. New interpre-tations of the skull of a primitive bony fish Erpetoichthys calabar-icus (Actinopterygii: Cladistia). Journal of Morphology 268:1021–1039.
Clemen, G., P. Bartsch, and K. Wacker. 1998. Dentition and dentiger-ous bones in juveniles and adults of Polypterus senegalus (Cladistia,Actinopterygii). Annals of Anatomy 180:211–221.
Cope, E. D. 1871. Contribution to the ichthyology of the Lesser Antilles.Transactions of the American Philosophical Society 14:445–483.
Cope, E. D. 1887. Zittel’s Manual of Palaeontology. American Naturalist21:1014–1019.
Cuvier, G. 1829. Le Regne Animal, distribue d’Apres son Organisa-tion. 2nd edition, Volume 2 (Fishes, etc.). Deterville, Paris, France.406 pp.
Daget, J., M. Gayet, F. J. Meunier, and J.-Y. Sire. 2001. Major discov-eries on the dermal skeleton of fossil and Recent polypteriforms: areview. Fish and Fisheries 2:113–127.
Dominik, W. 1985. Stratigraphie und Sedimentologie (Geochemie, Schw-ermineralanalyse) der Oberkreide von Bahariya und ihre Korrela-tion zum Dakhla-Becken (Western Desert, Agypten). Berliner Ge-owissenschaftliche Abhandlungen Reihe A: Geologie und Palaon-tologie 62:1–173.
Dutheil, D. B. 1999a. The first articulated fossil cladistian: Serenoichthyskemkemensis, gen. et sp. nov., from the Cretaceous of Morocco.Journal of Vertebrate Paleontology 19:243–246.
Dutheil, D. B. 1999b. An overview of the freshwater fish fauna fromthe Kem Kem beds (Late Cretaceous: Cenomanian) of southeasternMorocco; pp. 553–563 in G. Arratia and H.-P. Schultze (eds.), Meso-zoic Fishes 2—Systematics and Fossil Record. Proceedings of the In-ternational Meeting, Buckow, 6–10 July 1997. Verlag Dr. FriedrichPfeil, Munich, Germany.
Dutheil, D. B. 2009. Two new short-bodies Cladistia (Actinopterii)from the Kem Kem beds (Cenomanian of Morocco), p. 23 inN.-E. Jalil (ed.), 1st International Congress on North AfricanVertebrate Palaeontology, Program & Abstracts, Marrakech,25–27 May 2009. Available at http://www.ucam.ac.ma/smh/ABSTRACTS NAVEP1.pdf
El Sisi, Z., M. Hassouba, M. J. Oldani, and J. C. Dolson. 2002. Field TripNo. 8, Cairo 2002 International Conference and Exhibition. ApacheEgypt Companies, Cairo, Egypt.
Fontaine, W. M. 1899. Notes on Lower Cretaceous plants from the HayCreek Coal Field, Crook County, Wyoming. United States Geolog-ical Survey Annual Report 19, Part 2:645–702.
Franks, G. D. 1982. Stratigraphic modeling of the Upper Cretaceous sed-iments of the Bahariya Oasis; pp. 93–105 in Sixth Egyptian Gen-eral Petroleum Corporation Exploration Seminar, Cairo, Egypt,7–10 March 1982. Egyptian General Petroleum Corporation, Cairo,Egypt.
Gardiner, B. G., B. Schaeffer, and J. A. Masserie. 2005. A review of thelower actinopterygian phylogeny. Zoological Journal of the LinneanSociety 144:511–525.
Gayet, M., and F. J. Meunier. 1991. First discovery of Polypteridae(Pisces, Cladistia) outside of Africa. Geobios 24:463–466.
Gayet, M., and F. J. Meunier. 1992. Polypteriformes (Pisces, Cladistia)du Maastrichtien et du Paleocene de Bolivie. Geobios (Memoirespecial) 14:159–168.
Gayet, M., and F. J. Meunier. 1996. Nouveaux Polypteriformes du gise-ment Coniacien-Senonien d’In Becetem (Niger). Comptes Rendusde l’Academie des Sciences Paris, Serie II, Sciences de la Terre etdes Planetes 322:701–707.
Gayet, M., F. J. Meunier, and V. Levrat-Calviac. 1988. Mise en evidencedes plus anciens Polypteridae dans le gisement senonien, d’In Be-cetem (Niger). Comptes Rendus de l’Academie Sciences Paris, SerieII, Sciences de la Terre et des Planetes 307:205–210.
Gayet, M., F. J. Meunier, and C. Werner. 1997. Strange Polypteri-formes from the Upper Cretaceous of In Becetem (Niger) andWadi Milk Formation (Sudan). Geobios (Memoire special) 20:249–255.
Gayet, M., F. J. Meunier, and C. Werner. 2002. Diversification inPolypteriformes and special comparison with the Lepisosteiformes.Palaeontology 45:361–376.
Gemballa, S., and P. Bartsch. 2002. Architecture of the integument inlower teleostomes: functional morphology and evolutionary impli-cations. Journal of Morphology 253:290–309.
Geoffroy Saint-Hilaire, E. 1802. Histoire naturelle et descriptionanatomique d’un nouveau genre de poisson du Nil, nommepolyptere. Annales du Museum d’Histoire Naturelle (Paris),Memoires divers 1802:57–68.
Grande, L. 2010. An empirical synthetic pattern study of gars (Lep-isosteiformes) and closely related species, based mostly on skeletalanatomy. The resurrection of holostei. American Society of Ichthy-ologists and Herpetologists Special Publication 6:i–x, 1–871. Copeia10(2A).
Grande, L., and W. E. Bemis. 1996. Interrelationships of Acipenseri-formes, with comments on “Chondrostei”; pp. 85–115 in M. L. J.Stiassny, L. R. Parenti, and G. D. Johnson (eds.), Interrelationshipsof Fishes. Academic Press, New York, New York.
Gregory, W. K. 1933. Fish skulls: a study of the evolution of naturalmechanisms. Transactions of the American Philosophical Society23:75–481.
Gunther, A. 1870. Catalogue of the Fishes in the British Museum, Vol-ume 8. London, England, 574 pp.
Helfman, G. S., B. B. Collette, D. E. Facey, and B.W Bowen. 2009. TheDiversity of Fishes: Biology, Evolution and Ecology, 2nd edition.Wiley-Blackwell, Chicester, West Sussex, U.K., 720 pp.
Huxley, T. H. 1880. On the applications of the laws of evolutionto the arrangement of the Vertebrata and more particularly ofthe Mammalia. Proceedings of the Zoological Society of London1880:649–662.
Inoue, J. G., M. Miya, K. Tsukamoto, and M. Nishida. 2003. Basalactinopterygian relationships: a mitogenomic perspective on thephylogeny of the “ancient fish.” Molecular Phylogenetics and Evo-lution 26:110–120.
Jamieson, B. G. M. 1991. Fish Evolution and Systematics: Evidence fromSpermatozoa. Cambridge University Press, New York, New York,319 pp.
Khalifa, M. A., and O. Catuneanu. 2008. Sedimentology of the fluvial andfluvio-marine facies of the Bahariya Formation (early Cenomanian),Bahariya Oasis, Western Desert, Egypt. Journal of African EarthSciences 51:89–103.
Lacepede, B. G. E. 1803. Histoire Naturelle des Poissons, Volume 5. Plas-san Paris, France, 803 pp.
Lacovara, K. J., J. R. Smith, J. B. Smith, and M. C. Lamanna. 2002a. Ev-idence of semi-diurnal tides along the African coast of the Creta-ceous Tethys Seaway: Bahariya Oasis, Egypt. Geological Society ofAmerica, Abstracts with Programs 34:32.
Lacovara, K. J., J. R. Smith, J. B. Smith, and M. C. Lamanna. 2003. TheTen Thousand Islands coast of Florida: a modern analog to low-energy mangrove coasts of Cretaceous epeiric seas; pp. 1773–1784in J. R. A. Davis (ed.), Proceedings of the 5th International Confer-ence on Coastal Sediments, Clearwater Beach, Florida, 18–23 May2003. World Scientific Publishing Company, Singapore.
Lacovara, K. J., M. C. Lamanna, J. B. Smith, B. S. Grandstaff, and J.R. Smith. 2002b. Concentration and preservation potential of ver-tebrate fossils within coastal lithosomes: examples from the UpperCretaceous Bahariya Formation of Egypt. Journal of Vertebrate Pa-leontology 22(3, Supplement):77A.
Meunier, F. J. 2011. The Osteichtyes, from the Paleozoic to the ex-tant time, through histology and palaeohistology of bony tissues.Comptes Rendus Palevol 10:347–355
Meunier, F. J., and M. Gayet. 1992. Nouveau remainiement de la ganoınechez un Semionotidae du Cretace Superieur de Bolivie: interetpaleobiologique. Geobios 25:767–774.
Meunier, F. J., and M. Gayet. 1996. A new polypteriform from the LateCretaceous and the middle Paleocene of South America; pp. 95–103in G. Arratia and G. Viohl (eds.), Mesozoic Fishes—Systematicsand Paleoecology. Proceedings of the International Meeting,Eichstatt, 9–12 August, 1993. Verlag Dr. Friedrich Pfeil, Munich,Germany.
Murray, A. M., T. D. Cook, Y. S. Attia, P. Chatrath, and E. Simons.2010. A freshwater ichthyofauna from the late Eocene BirketQarun Formation, Fayum, Egypt. Journal of Vertebrate Paleontol-ogy 30:665–680.
Dow
nloa
ded
by [
Uni
vers
ity o
f Pe
nnsy
lvan
ia]
at 0
7:32
13
Janu
ary
2012

26 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 32, NO. 1, 2012
Noack, K., R. Zardoya, and A. Meyer. 1996. The complete mitochondrialDNA sequence of the bichir (Polypterus ornatipinnis), a basal ray-finned fish: ancient establishment of the consensus vertebrate geneorder. Genetics 144:1165–1180.
Oertel, G. F. 1985. The barrier island system. Marine Geology 63:1–12.Ogg, J. G., G. Ogg, and F. M. Gradstein. 2008. The Concise Geo-
logic Time Scale. Cambridge University Press, Cambridge, U.K.,184 pp.
Paxton, J. R., and W. N. Eschmeyer. 2003. Encyclopedia of Fishes. 2ndEdition. Fog City Press, San Francisco, California, 240 pp.
Pehrson, T. 1947. Some new interpretations of the skull in Polypterus.Acta Zoologica 28:241–258.
Schaal, S. 1984. Oberkretazische Osteichthyes (Knochenfische) aus demBereich von Bahariya und Kharge, Agypten, und ihre Aussagen zurPalokologie und Stratigraphie. Berliner Geowissenschaftliche Ab-handlungen Reihe A 53:1–76.
Schultze, H.-P. 1996. The scales of Mesozoic actinopterygians; pp. 83–93in G. Arratia and G. Viohl (eds.), Mesozoic Fishes—Systematics andPaleoecology. Proceedings of the International Meeting, Eichstatt,9–12 August 1993. Verlag Dr. Friedrich Pfeil, Munich, Germany.
Sire, J.-Y., P. C. J. Donoghue, and M. K. Vickaryous. 2009. Originand evolution of the integumentary skeleton in non-tetrapod ver-tebrates. Journal of Anatomy 214:409–440.
Smith, J. A. 1865. Untitled. (reported in) Daily Review 1242:2.Smith, J. A. 1866. Description of Calamoichthys, a new genus of ganoid
fish from Old Calabar, western Africa. Annals and Magazine of Nat-ural History 3:112–114.
Smith, J. B., B. S. Grandstaff, and M. S. Abdel-Ghani. 2006a. Microstruc-ture of polypterid scales (Osteichthyes: Actinopterygii: Polypteri-dae) from the Upper Cretaceous Bahariya Formation, Bahariya Oa-sis, Egypt. Journal of Paleontology 80:1179–1185.
Smith, J. B., M. C. Lamanna, H. Mayr, and K. J. Lacovara. 2006b.New information regarding the holotype of Spinosaurus aegyptiacusStromer, 1915. Journal of Paleontology 80:400–406.
Smith, J. B., M. C. Lamanna, K. J. Lacovara, P. Dodson, J. R. Smith, J.C. Poole, R. Giegengack, and Y. Attia. 2001. A giant sauropod di-nosaur from an Upper Cretaceous mangrove deposit in Egypt. Sci-ence 292:1704–1706.
Soliman, H. E., and M. A. Khalifa. 1993. Stratigraphy, facies and depo-sitional environments of the Lower Cenomanian Bahariya Forma-tion, Bahariya Oasis, Western Desert, Egypt. Egyptian Journal ofGeology 37:193–209.
Stokes, C., and P. B. Webb. 1824. Description of some fossil vegetables ofthe Tilgate Forest in Sussex. Transactions of the Geological Societyof London Series 21:421–424, 3 plates.
Stromer, E. 1905. Die Fischreste des mittleren und oberen Eocans vonAgypten. Beitrage zur Palaontologie und Geologie Osterreichs-Ungarns und des Orients 18:163–192.
Stromer, E. 1914. Ergebnisse der Forschungsreisen Prof. E. Stromersin den Wusten Agyptens. I. Die Topographie und Geologie der
Strecke Gharaq-Baharıje nebst Ausfuhrangen uber die geologischeGeschichte-Agyptens. Abhandlungen der Koniglich BayerischenAkademie der Wissenschaften, Mathmatisch-physikalische Klasse16:1–78.
Stromer, E. 1925. Der Ruckgang der Ganoidfische von der Kreidezeit an.Zeitschrift der Deutsche Geologische Gesellschafte 77:348–371.
Stromer, E. 1936. Ergebnisse der Forschungsreisen Prof. E. Stromers inden Wusten Agyptens VII. Baharıja-Kessel und -Stufe mit derenFauna und Flora, Eine erganzende Zusammenfassung. Abhandlun-gen der Bayerischen Akademie der Wissenschaften Mathmatisch-naturwissenschaftliche Abteilung, Neue Folge 33:1–102.
Suzuki, D., M. C. Brandley, and M. Tokita. 2010. The mitochondrial phy-logeny of an ancient lineage of ray-finned fishes (Polypteridae) withimplications for the evolution of body elongation, pelvic fin loss, andcraniofacial morphology in Osteichthyes. BMC Evolutionary Biol-ogy 10:21.
Swinney, G. N., and D. Heppell. 1982. Erpetoichthys or Calamoichthys:the correct name for the African Reed-fish. Journal of Natural His-tory 16:95–100.
Tanner, L. H., and M. A. Khalifa. 2010. Origin of ferricretes in fluvial-marine deposits of the Lower Cenomanian Bahariya Formation, Ba-hariya Oasis, Western Desert, Egypt. Journal of African Earth Sci-ences 56:179–189.
Traquair, R. H. 1870. On the cranial osteology of Polypterus. Journal ofAnatomy and Physiology 5:166–184.
Wacker, K., P. Bartsch, and G. Clemen. 2001. The development ofthe tooth pattern and dentigerous bones in Polypterus senegalus(Cladistia, Actinopterygii). Annals of Anatomy 183:37–52.
Weiler, W. 1935. Ergebnisse der Forschungsneisen Prof. E. Stromers inden Wusten Agyptens. II. Wirbeltierreste der Baharıje-Stufe (un-terestes Cenoman). 16. Neue Untersuchungen an den Fischresten.Abhandlungen der Bayerischen Akademie der Wissenschaften,Mathmatisch-naturwissenschaftliche Abteilung 32:1–57.
Werner, C. 1991. Aspects on terrestrial Upper Cretaceous ecosystemsof Egypt and northern Sudan; pp. 71–72 in Z. Kielan-Jaworowska,N. Heintz, and H. A. Nakrem (eds.), Fifth Symposium on Meso-zoic Terrestrial Ecosystems and Biota, Oslo, Norway, 12–15 August1991. Contributions from the Paleontological Museum, Universityof Oslo.
Werner, C., and M. Gayet. 1997. New fossil Polypteridae from the Ceno-manian of Sudan, an evidence of their high diversity in the early LateCretaceous. Cybium 21:67–81.
Woodward, A. S. 1908. On some fossil fishes discovered by Prof. Ennes deSouza in the Cretaceous formation of Ilheos (State of Bahia), Brazil.Quarterly Journal of the Geological Society of London 64:358–362.
Submitted December 1, 2010; revisions received August 26, 2011;accepted September 20, 2011.Handling Editor: Matt Friedman.
Dow
nloa
ded
by [
Uni
vers
ity o
f Pe
nnsy
lvan
ia]
at 0
7:32
13
Janu
ary
2012