bacterial opportunism in fallow and meadow soils
TRANSCRIPT

8
Phytopedon (Bratislava), Vol. 9, 2010/2, p. 8-17. DOMINIKA CHMOLOWSKA1, VÁCLAV KRIŠTŮFEK2, DANA ELHOTTOVÁ2, RYSZARD LASKOWSKI1, MACIEJ KOZAK3
1Jagiellonian University, Institute of Environmental Sciences, Gronostajowa 7, 30-387 Kraków, Poland; 2Biology Centre AS CR, v.v.i. – Institute of Soil Biology, Na Sádkách 7, 370 05 České Budějovice, Czech Republic 3Jagiellonian University, Institute of Botany, Lubicz 46, 31-512 Kraków, Poland
BACTERIAL OPPORTUNISM IN FALLOW AND MEADOW SOILS Abstract Chmolowska, D., Krištůfek, V., Elhottová, D., Laskowski R., Kozak M., 2010: Bacterial opportunism in fallow and meadows soils. Phytopedon (Bratislava), Vol. 9, 2010/2: p. 8-17.
Opportunism, or the r strategy, in the case of soil microbial communities refers to the ability of being cultivated and the rate of growth on culture media. It is generally assumed that younger soil is inhabited by more opportunistic (r-selected) communities. In our research we examined the opportunism of soil bacterial communities from six fallow fields, which represent a young ecosystem under succession, six meadows, which represent a stable ecosystem, and four sites transitional between fallow and meadow fields. Culturability was calculated from the ratio of Colony Forming Units from agar R2A plates to the total number of cells from epifluorescence microscopy. To compare the rate of growth, colonies were counted on plates every day for seven days. Additionally, the results obtained from agar inoculations were compared with soil microbial communities PLFA profiles. There were no differences in the culturability ratio of soil bacterial communities between ecosystems. However, fallow soil communities exhibited small but significantly higher growth activity than meadow soils communities, which was observed both in the case of the number of colonies (repeated measures ANOVA: p = 0.03, F(1, 10) = 5.97) and also in the time of proliferation. Fallow soil bacterial communities showed significantly faster growth in the first two days of incubation than the meadow and transitional ones. Differences in the number of CFU between the two types of fields did not follow the pattern of PLFA profiles.
Key words: soil microbial community, r/K strategy, opportunism, culturability, fallow, meadow
INTRODUCTION
In the last 20 years considerable land use changes have taken place in the countries undergoing socio-economical transformations. In Poland, due to lower cost-effectiveness of agricultural production, a substantial increase in the area of abandoned farmlands has been
observed. The area of fallow fields was six times higher in 2006 than in 1990 (GUS 2006, 1990). The GUS database does not report the real status of these fields: whether they are under basic care by owners, e.g. are mowed, or abandoned. The increase of farmland area taken by ruderal plants has been seen across the country.

9
Abandoned fields undergo secondary plant succession. Not only do soil properties follow plant community changes in terms of physicochemical properties, such as increased organic matter (BILLINGS 1941) or ammonium content (MALICKI et PODSTAWKA-CHMIELEWSKA 1998), but also in soil microbial communities, e.g. an increase of the fungi:bacteria ratio (VAN DER WAL et al. 2006, PLASSART et al. 2008). All these processes are desirable from the ecological point of view.
Together with the enlarged area of fallows, a decrease in the total area of meadows was reported (GUS 2000, 2006). Scientists have also noted a decrease in meadow plant diversity (e.g. KAŹMIERCZAKOWA et al. 1990, KOTAŃSKA 1995, BARABASZ 1997, KOZAK 2007), which is caused by cessation of meadow management. Also, the expansion of cities to surrounding villages and grasslands poses a threat to these human-dependent but biologically diverse ecosystems. For example, Kraków, the city with a population of 800 000, has expanded to the meadows giving shelter to populations of rare butterflies (SKÓRKA et al. 2006). The fresh meadows chosen for our research, representing communities within the Arrhenatherion alliance, are protected by Natura 2000 as a valuable, endangered and rapidly disappearing habitat in the UE.
The processes listed above show how important it is to study these ecosystems. Soil microorganisms and soil processes are particularly interesting and important because, being crucial for organic matter turnover, they constitute the base for functioning of whole ecosystems.
Opportunism means a "behaviour in which you use every situation to try to get power or an advantage" (dictionary.cambridge.org). Microbiologists use this term to describe the ability of microbial communities to grow on culture media. According to SIGLER et al. (2002), an index of opportunism is a ratio of culturable to total cells. Other authors expressed the ability of an microorganism to use an artificial nutrient source from agar media as the r-strategy, that is a strategy of organisms living and multiplying in the environment which is uncrowded and rich in nutrients (DELEJI et al. 1993). However, the assumption of r- and K-selection is inextricably linked with a "speed strategy", r-selected organisms being those proliferating
quicker, and the term is also used by microbiologists in such a meaning. This may lead to some confusion. To avoid this, one has to accept that the first criterion of opportunism, or the r strategy, is the ability to grow on an artificial medium, and the second is the speed with which microbial cells proliferate.
The criteria for r- and K-strategists in microbial communities proposed by DELEJI et al. (1993), according to the evolutionary hypothesis by MACARTHUR et WILSON (1967), assume that r-selected microorganisms grow easier and quicker in a new environment (or medium), rich in easily degradable C source, while K-selected ones grow slower, but their cell metabolism is more efficient: they should be more effective in utilisation of recalcitrant substrates and more resistant to toxins in a saturated environment. DELEJI et al. (1993) was right that the younger the soil the more r-selected are microorganisms. Later research by SIGLER et ZEYER (2004) showed, however, that fast growing bacterial communities are more stress tolerant (nutrient and temperature stress) and more frequently resistant to antibiotics.
The already well proved assumption about the relationship between the age of soil and microbial opportunism (DELEJI et al.1993, SIGLER et al. 2002, SIGLER et ZEYER 2004) led us to check whether soils of fallows and meadows exhibit different cultivable to total cell ratios and growth rates of microbial colonies. The hypothesis was that soils obtained from fallows, which are only a few years old, should contain more r-selected bacterial communities than soils from meadows, which were consistently managed for many years. Ecosystems transitional between fallow and meadow were also included and their soil microbial communities were expected to exhibit in-between opportunism. Additionally, a general comparison of soil microbial communities was performed.
MATERIALS AND METHODS
A set of six fallows, six meadows, and four transitional fallow-to-meadow fields was chosen in the Beskidy Mountains, Poland. All sites were less than a few kilometers apart to avoid large environmental differences which might influence the results. They were chosen

10
according to their floristic composition. For fallows, the indicators were: presence of ruderal and segetal weeds (especially Elymus repens) and less than 20 species in 25 m2 plots; for meadows - high richness (30-50 plant species) and high proportion of species characteristic for the Molinio-Arrhenatheretea class. The transitional fields consisted of 20-30 plant species. Each site within a given field type group was treated as an independent replicate. Ten 10 cm deep soil cores were taken from each site. Each ten was mixed to obtain one combined sample per site. Soil
samples were sieved through a 4 mm mesh and all animals were removed. pH in H2O was measured as 1:5, soil mass:extractant volume. Content of organic matter was estimated through overnight combustion in 550 Cْ. C:N ratio was calculated from organic carbon and nitrogen measured on an elemental analyser (Elemental System GmbH). Soil properties and plant community indexes are summarized in Table 1. Soil for PLFA analysis was frozen at -20°C and soil for inoculation and DAPI staining was stored at 4°C 4 months prior to measurements.
Table 1. Organic matter content (OM), pHH2O, C:N ratio, plant taxa number and Shanon-Wiener diversity index of the examined fields
Field OM [%] pHH2O C:N number of plant taxa
Shannon-Wiener diversity index
fallow 1 8.4 5.7 9.9 22 2.26 fallow 2 5.8 5.6 10.3 13 1.08 fallow 3 5.0 5.8 10.3 23 1.96 fallow 4 6.5 5.5 10.1 26 2.22 fallow 5 5.2 6.6 10.5 19 1.56 fallow 6 7.4 6.5 10.2 16 1.62
transitional 1 6.5 5.6 9.9 35 2.89 transitional 2 5.0 5.2 10.4 36 2.83 transitional 3 4.5 5.4 10.2 33 2.90 transitional 4 6.7 5.4 10.0 31 2.75
meadow 1 8.3 6.7 10.6 47 3.38 meadow 2 9.4 5.9 11.0 42 3.41 meadow 3 11.8 6.9 11.0 50 3.45 meadow 4 7.4 5.8 10.8 41 3.08 meadow 5 6.4 7.7 12.3 37 3.13 meadow 6 6.5 7.6 10.8 46 3.36
For agar plates inoculations, 5 g of soil
was suspended in 45 ml of sodium hexametaphoshpate buffer (10-1 dilution). The suspensions were left in a sonic bath for 4 minutes. 0.2 ml of 10-5 dilution in sterile tap water was inoculated on R2A agar plates . For each soil sample at least 5 replicate plates were used. Growing colonies were counted for
seven days, beginning on the first day after inoculation. Colonies growing in the first three days were considered as representing r-strategists and those appearing and growing in days 4-7 were considered as K-strategists.
To estimate the total number of bacteria, DAPI stained cells were counted with epi-fluorescence microscopy according to BLOEM

11
(1995). The ratio of culturable to total cells was calculated.
To compare the structure of soil bacterial communities, Phoshoplipid Fatty Acid (PLFA) analysis was performed with the MIDI system (MIDI Inc, Newark, USA). Soils corres-ponding to 5 g of dw were extracted according to manufacturer guidelines, in line with Frouz et al. (2006).
Statistical analyses were performed in Statgraphics (normality validation, nonlinear regression for asymptote estimation), STATISTICA (Repeated Measures ANOVA) and Canoco (Principal Component Analysis).
RESULTS
There were no statistically significant differences in the number of bacteria (Tab. 2) and Actinomycetes (Tab. 2) cultivated on agar medium. The final number of colonies obtained in fallow soils ranged from 40*106 g-1 dry weight (dw), to 72*106, while in meadows the minimum was 27*106, and maximum 60*106 g-1 dw, which indicates a trend for higher colony number in fallows. Transitional fields were more similar to meadows, with a range of 28*106 - 60*106 g-1 dw. The average number of Actinomycetes from fallow soils was 1.9*106, transitional 2.1*106, and from meadow soils 2.3*106 g-1 dw (Tab. 2).
Table 2. Total bacteria number, bacteria CFU, Actinomycetes CFU and culturability fallow meadow transitory mean ± SD mean ± SD mean ± SD Total bacteria (cell*g dw-1) 11.8 *108 ± 2.0*108 9.6 *108 ± 2.0*108 11.7 *108 ± 6.1*108 Culturable bacteria (CFU*g dw-1) 5.3 *107 ± 1.2*107 4.2 *107 ± 1.4*107 4.6 *108 ± 1.6*109 Culturable Actinomycetes (CFU*g dw-1) 1.9*106 ± 1.0*106 2.3*106 ± 1.8*106 2.1*106 ± 1.6*106 Culturability (%) 4.7 ± 0.9 4.7 ± 1.4 4.5 ± 1.4
There was no statistically significant
difference in the total number (DAPI) of bacteria among ecosystems (Tab. 2), however, a trend of higher numbers in fallows than in meadows was seen (p=0.07, one-way ANOVA). The average number of total bacterial cells in fallows and transitional fields was 1.2*109 g-1 dw, whereas in meadow it was 1.0*109 g-1 dw (Tab. 2). There were no differences in the ratio of cultivable to total
bacteria between fallow, meadow and transitional soils. The average culturability (sum of bacterial and Actinomycetes) was 4.7% for fallow soils, 4.5% for transitional and 4.7% for meadow soils (Tab. 2). The number of CFU were correlated with the total number of bacterial cells (simple regression, p=0.005) and the model explained 45% of variation in the CFU and showed moderate relationship strength (corr. coeff.=0.67).
Table 3. Results of the asymptotic function fitted to the cumulated CFU (A-asymptote, k-sloping, SE – standard error, 95% confidence intervals lower and upper)
Estimate
SE
95% Interval Lower
95% Interval Upper
Estimate
SE
95% Interval Lower
95% Interval Upper
R2 adj
fallow k 0.42 0.09 0.25 0.59 A [*106 CFU] 57 4.76 47.8 67 81% transitional k 0.14 0.09 -0.04 0.33 A [*106 CFU] 81.8 36.5 6.74 157 79% meadow k 0.18 0.10 -0.02 0.37 A [*106 CFU] 64 23.6 16.3 112 70%

12
Regarding the rate of colonies growth on medium, a trend for higher growth activity was seen on plates inoculated with solutions from fallow soils. An asymptotic function was fitted to the cumulated counts of the colony forming units (CFUs) for each biocenosis (Fig. 1, Fig. 2, Tab. 3), but high variance between replicates led to wide confidence intervals, resulting in non-significant differences between the soils in regression parameters. Also, the repeated measures ANOVA for cumulated number of CFUs did not reveal statistically significant differences (p=0.08) between the three biocenoses (Fig. 3). However, when transitional fields were excluded, the repeated measures ANOVA showed a significant difference between fallows and meadows: F(1,10)=5.97, p=0.03 (Fig 3).
Figure 1. Asymptotic function of cumulated CFU: fallow
To avoid the possible influence of the total
number of cells on the number of colonies, the cumulated CFU was calculated as percentage of the final colonies number in each ecosystem (Fig. 4). Again fallow soil communities tended to be faster. Transitional fields were similar to meadows in this regard. Nontheless, a repeated measures ANOVA didn't prove different r- or K-selection in fallow, transitional and meadow soil microbial communities (day 1-3 vs. 4-7).
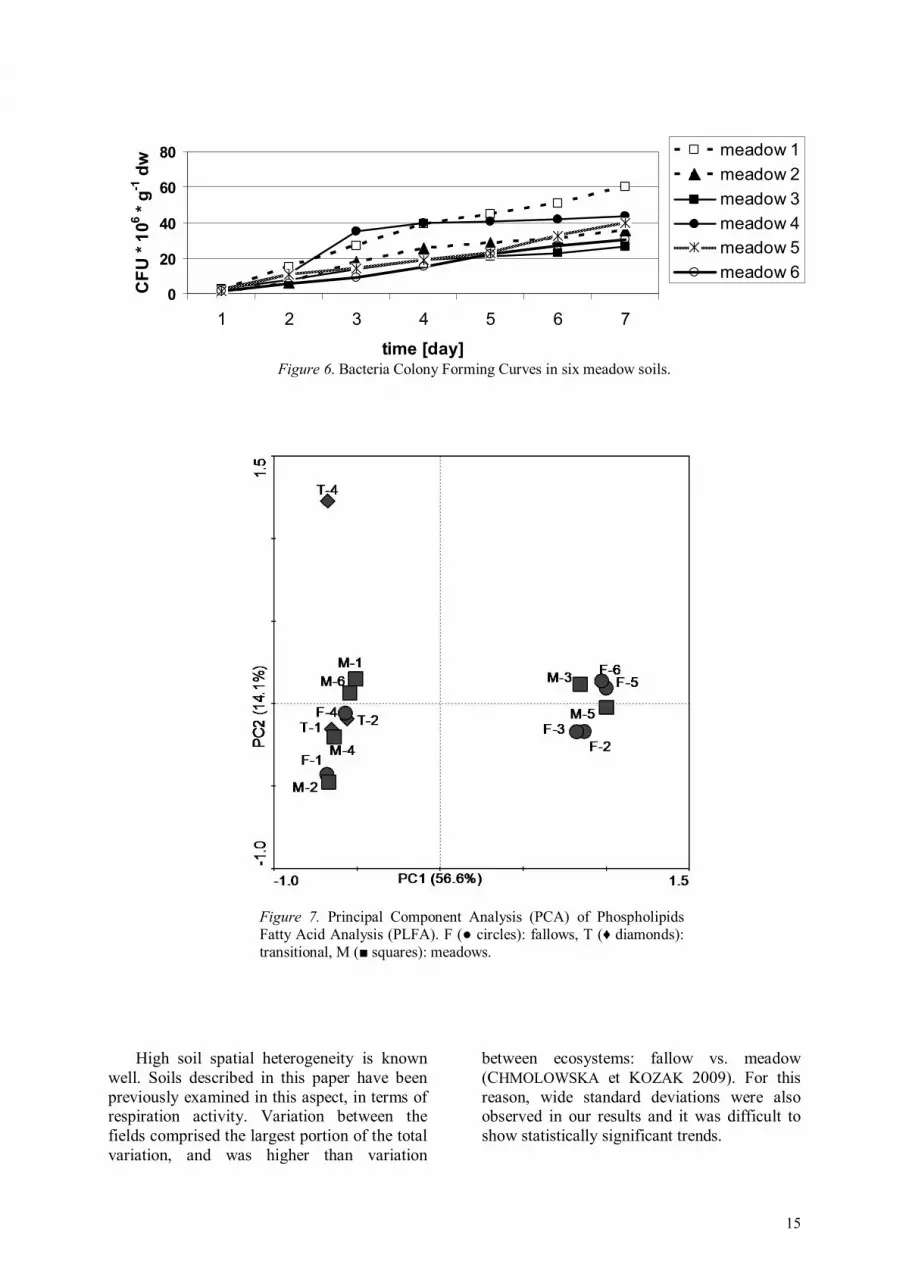
Examination of Colony Forming Curves for each set of fields showed that while cumulative curves for fallow (Fig. 5) and transitional had consistent shapes, meadow
bacterial communities showed higher inconstancy, with some soils dominated even by fast-growers, e.g. in meadow 4 (Fig. 6). Interestingly, this meadow had the highest dominance index of plant community, which was also reflected in the lowest Shannon-Wiener diversity index (Table 2). A PCA analysis of bacterial community structure (PLFA) (Fig. 7) did not relate to differences in CFUs activity of particular fields.
Figure 2. Asymptotic function of cumulated CFU: meadow
DISSCUSSION
It is often stated that younger soils exhibit higher opportunism, that is higher culturability and/or faster growth rate of bacterial colonies. However, this conclusion was drawn mainly from research on soils of very different properties. For instance, pot rhizosphere soil had more fast growing bacteria than field rhizosphere soil after two months from sieving, but the effect did not last long and a month later the differences disappeared (DELEJI et al. 1993). A comparison of 60 year old and 150 year old tree rhizosphere showed a higher number of bacterial CFUs for the older trees (KUSKE et al. 2003). Ten year old soil (time since deglaciation) had higher culturable:total ratio (C:T) than virgin soil (zero years old). After this initial increase, C:T remained constant for up to 70 years and then decreased again or, in the case of another chronosequence

13
from that publication, declined right after 10 years. These results indicate that there is some inconsistency in the accepted view: some soils may be inhabited by fast growing opportunistic bacteria, but some not, and this may depend on plant growth stage, successional stage and, possibly, other still unrecognized factors. With such a diversity of results obtained by other authors, it does not seem surprising that no significant differences between meadow and fallow soil were found in this study for the counts of colonies and C:T ratio, as actual differences between these two ecosystems are not high. Nevertheless, the method based on Colony Growing Curves was sensitive enough
to differentiate meadows from fallows, although this could have resulted from somewhat higher number of total cells in fallow soils. This is supported by the significant correlation of CFUs with the total number of bacterial cells. Results for transitional fields were, as expected, intermediate between fallows and meadows. These findings are in agreement with BRODIE et al. (2002), who obtained higher number of colonies from agriculturally improved but less plant rich field than from a semi-natural meadow. Interestingly, the difference in colonies number was up to one order of magnitude.
transitory fallows meadows
1 2 3 4 5 6 7
time [day]
0
20
40
60
CFU
* 10
6 * g
-1 dw
Figure 3. Repeated measure ANOVA of bacteria CFU. Bars denote 0.95 confidence intervals. Effects: fallow*transitional*meadow: p=0.08, day*'fallow*transitional*meadow': p>>0.05; Effects: fallow*meadow: p=0.03, F(1,10)=5,97, day* 'fallow*meadow': p=0.06
Nonetheless, fallow soil colonies proved their higher opportunism by visibly faster growth in the two first days of incubation. Because growth rate did not follow the common partitioning for days 1-3 days and 4-7, we cannot unequivocally state that fallow communities are r-strategists, but we may conclude that they tend to be more r-selected than meadow and transitional fields soil communities are.
The storage of soils for 4 months before inoculation to agar plates does not seem to affect the viability of soil communities. The obtained numbers of CFUs matched similar published data. SIGLER et al. (2002) reported approximately 1.4*107 and 2.8*107 CFUs g-1 dw in a ten years old soil, and 3.8 *107 and 8.3*107 CFU g-1 dw in a 46-years old soil (time since deglaciation). In our study, on average 4.7*107 CFU g-1 dw was obtained.

14
BRODIE et al. (2002) counted 1.2*107 CFU g-1 dw at a semi-natural meadow, 4.6*107 and 13.8*107 CFU g-1 dw at two transect sites and 141*107 CFU g-1 dw at an agriculturally managed meadow. The fields from our study were most similar to a transect site close the to semi-natural grassland. The semi-natural meadow reported by BRODIE et al. (2002) had very low pH (3.9), compared to 6.3 in the managed field, and organic matter content was 26% and 18%, respectively. The differences in NH4+, NO3-, P, K and Ca content were large. The differences between fallow and meadow fields from this study were not as high: soil
organic matter comprised 8% in meadows and 6% in fallows, and the content of most nutrients was not significantly different between the two, which can be a reason for small differences in colony counts. It was the initial assumption of the study to compare ecosystems similar in terms of abiotic properties, but differing in history, age and diversity, in order to observe if this might have an impact on soil microbial communities. These relationships appear to be subtle and differences are less obvious and more difficult to prove.
0
20
40
60
80
100
1 2 3 4 5 6 7
time [day]
%
FALLOW
TRANSITORY
MEADOW
Figure 4. Bacteria Colony Forming Curves in fallow, transitional and meadow soils (average +/- SD).
0
20
40
60
80
1 2 3 4 5 6 7
time [day]
CFU
* 10
6 * g-1
dw fallow 1
fallow 2fallow 3fallow 4fallow 5fallow 6
Figure 5. Bacteria Colony Forming Curves in six fallows.
15
0
20
40
60
80
1 2 3 4 5 6 7
time [day]
CFU
* 10
6 * g-1
dw meadow 1
meadow 2meadow 3meadow 4meadow 5meadow 6
Figure 6. Bacteria Colony Forming Curves in six meadow soils.
Figure 7. Principal Component Analysis (PCA) of Phospholipids Fatty Acid Analysis (PLFA). F (● circles): fallows, T (♦ diamonds): transitional, M (■ squares): meadows.
High soil spatial heterogeneity is known well. Soils described in this paper have been previously examined in this aspect, in terms of respiration activity. Variation between the fields comprised the largest portion of the total variation, and was higher than variation
between ecosystems: fallow vs. meadow (CHMOLOWSKA et KOZAK 2009). For this reason, wide standard deviations were also observed in our results and it was difficult to show statistically significant trends.

16
CONCLUSIONS
Because of the high within-group variation, trends in the data were difficult to prove statistically. Nevertheless, meadow soils differed from fallow soils significantly in terms of Colony Forming Curves and exhibited lower bacterial colonies growth activity. There were no differences in the culturable:total ratio or clear dominance of either the r or K growing strategy. However, the significantly higher
growth rate of fallow soil colonies at the beginning indicated higher opportunism of its soil bacterial communities. Meadows chosen for this research differ from fallows in terms of age, history and biodiversity but plant biomass and most abiotic properties are similar. Comparison of our data with literature data leads to a conclusion that shifts in bacterial opportunism are driven by considerable changes in soil chemical properties and in conditions on the ground surface.
Translated by the authors and corrected by otostrona.pl/tlumaczenia
ACKNOWLEDGEMENTS This research was supported by Jagiellonian University funds: DS/BiNoZ/INoS/759/2009, LC06066 Centre for Environmental Microbiology, MŠMT ČR (2006-2010) and by Research Plan of Biology Centre v.v.i., Academy of Sciences of the Czech Republic – ISB (AVOZ 60660521).
REFERENCES
BARABASZ, B., 1997: Changes in the meadows of the northern part of the Niepołomice Forest during
twenty years (in Polish). Studia Nat. 22: p. 1-99. BILLINGS, W.D., 1941: Quantitative Correlations Between Vegetational Changes and Soil
Development. Ecology 22, 4: p. 448-456. BLOEM, J., 1995: Fluorescent staining of microbes for total direct counts. In: Akkerman, A.D.L., van
Elsas, J.D., de Bruijn, F.J. (Eds.), Molecular Microbial Ecology Manual, vol. 4.1.8. Kluwer Academic Publ., Dordrecht. Boston, London, pp. 1-12.
BRODIE, E., EDWARDS S., CLIPSON, N., 2002: Bacterial Community Dynamics across a Floristic Gradient in a Temperate Grassland Ecosystem. Microbial Ecology 44: p. 260-270.
CHMOLOWSKA, D., KOZAK, M., 2009: Nomolt impact on soil respiration of fallows and meadows – do their microbial communities differ in resistance? SETAC Europe 19th Annual Meeting, Göteborg, Sweden 25-29.05.2009.
DELEJI, F.A.A.M., WHIPPS, J.M., LYNCH, J.M. 1993: The Use of Colony Development for the Characterization of Bacterial Communities in Soil and on Roots. Microbial Ecology 27: p. 81-97.
FROUZ, J.D., ELHOTTOVÁ, D., KURÁŽ, V., ŠOURKOVÁ, M., 2006: Effects of soil macrofauna on other soil biota and soil formation in reclaimed and unreclaimed post mining sites: Results of a field microcosm experiment. Applied Soil Ecology 33: p. 308 – 320.
GUS (Central Statistical Office in Poland): 1990, 2001, 2006: The Statistical Yearbook of Agriculture (in Polish). Zakł. Wyd. Statyst., Warszawa.
KAŹMIERCZAKOWA, R., KUSIŃSKA, M., KWIATKOWSKA, A., POZNAŃSKA, Z., RAMS, B., 1990: Productivity of meadow communities in the Tatra glades (in Polish). Studia Nat., Ser. A 34: p. 77-111.
KOTAŃSKA, M., 1995: Fresh meadow dynamics in agriculture landscape (in Polish). Conferences and symposiums materials, 50th Meeting of Polish Botany Society PTB – Kraków, Poland.

17
KOZAK, M., 2007: Differentiation of the meadow communities in the Gorce Mountain Range (Polish Western Carpathians) (in Polish). Prace Bot. 41: p. 1-174.
KUSKE, C.R., TICKNOR, L.O., BUSCH, J.D., GEHRING, C.A., WHITHAM, T.G., 2003: The Pinyon Rhizosphere, Plant Stress, and Herbivory Affect the Abundance of Microbial Decomposers in Soils. Microbial Ecology 45: p. 340-352.
MALICKI, L., PODSTAWKA-CHMIELEWSKA, E., 1998: Zmiany fitocenozy i niektórych właściwości gleby zachodzące podczas odłogowania oraz będące efektem zagospodarowania wieloletniego odłogu. Biblioth. Fragm. Agron., 5: p. 97-114.
MACARTHUR, R.H., WILSON, E.0., 1967: The theory of island biogeography. Princetoll Univ. Press. Princeton, N.J.
PLASSART, P., VINCESLAS, M.A., GANGNEUX, CH., MERCIER, A., BARRAY, S., LAVAL, K., 2008: Molecular and functional responses of soil microbial communities under grassland restoration. Agr. Ecosys. Env. 127: p. 286-293.
SIGLER, W.V., ZEYER, J., 2004: Colony-Forming Analysis of Bacterial Community Succession in Deglaciated Soils Indicates Pioneer Stress-Tolerant Opportunists. Microbial Ecology 48: p. 316-323.
SIGLER, W.V., CRIVII, S., ZEYER, J., 2004: Bacterial Succession in Glacial Forefield Soils Characterized by Community Structure, Activity and Opportunistic Growth Dynamics. Microbial Ecology 48: p. 316-323.
SKÓRKA, P., SETTELE, J., WOYCIECHOWSKI M., 2007: Effects of management cessation on grassland butterflies in southern Poland. Agric. Eco Ecosyst. Environ., 121, 4: p. 319-324.
VAN DER WAL, A.., VAN VEEN, J.A., VAN VEEN W., BOSCHKER, H.T.S., BLOEM, J., KARDOL, P., VAN DER PUTTEN W.H., DE BOER, W., 2006: Fungal biomass development in a chronosequence of land abandonment. Soil Biology & Biochemistry, 38: p. 51-60.
Souhrn Chmolowska, D., Krištůfek, V., Elhottová, D., Laskowski R., Kozak M., 2010: Přizpůsobivost bakterií v půdách úhorů a luk.
Porovnávali jsme r-strategii (oportunismus) půdních heterotrofních bakterií v půdách opuštěných polí - úhorů, lučních půd a půdách přechodových mezi luční půdou a úhorem. Předpokládali jsme, že půdy úhorů, které jsou mladší, budou obsahovat více r-stratégů. Luční půdy se liší od půd úhoru zejména věkem založení, historií hospodaření a biodiverzitou rostlin avšak biomasa rostlin a většina abiotických charakteristik je podobná. Na základě ekologického konceptu r- a K-strategie byla kvantifikována bakteriální populace půd plotnovou metodou s denním odečtem bakterií po dobu 7 dnů. Celkové počty bakterií ve vzorku byly sledovány epifluorescenční mikroskopií barvením DAPI. Ze získaných dat byl stanoven tzv. index sukcesního stavu půdy C/T (poměr kultivovatelných a celkových počtů bakterií) a tzv. Colony-Forming Curves - CFCs (počet bakterií pro každý den kultivace vyjádřený jako procento z celkového počtu bakterií narostlých na Petriho misce). Rozdíly mezi sledovanými půdami úhor - přechodová půda – luční půda nebyly statisticky významné i když určité trendy byly zřejmé. Výsledky naznačují vyšší zastoupení oportunistů v půdách úhoru než v lučních půdách.