association between endogenous feline leukemia virus loads and exogenous feline leukemia virus...
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Virus Research 135 (2008) 136–143
Contents lists available at ScienceDirect
Virus Research
journa l homepage: www.e lsev ier .com/ locate /v i rusres
Association between endogenous feline leukemia virus loads and exogenousfeline leukemia virus infection in domestic cats
Ravi Tandona, Valentino Cattori a, Andrea C. Pepina, Barbara Rionda, Marina L. Meli a,Mike McDonaldb, Marcus G. Doherrc, Hans Lutza, Regina Hofmann-Lehmanna,∗
a Clinical Laboratory, Vetsuisse Faculty, University of Zurich, Winterthurerstrasse 260, 8057 Zurich, Switzerlandb Department of Veterinary Pathology, Companion Animal Diagnostics, Institute of Comparative Medicine, Bearsden Road, Glasgow, Scotland, UKc Department of Clinical Veterinary Medicine, Vetsuisse Faculty, University of Bern, Switzerland
a r t i c l e i n f o
Article history:Received 12 December 2007Received in revised form 28 February 2008Accepted 29 February 2008Available online 14 April 2008
Keywords:Endogenous feline leukemia virusLong terminal repeatsSpecific pathogen freeReceptor binding domain
a b s t r a c t
Recently, we demonstrated that endogenous feline leukemia virus (enFeLV) loads may vary among cats ofdifferent populations and that FeLV-infected cats have higher enFeLV loads than uninfected cats. Thus, wehypothesized that enFeLV might influence the pathogenesis and outcome of FeLV infection. No significantdifference in the infection outcome (regressive versus progressive infection) was observed between groupsof cats with high or low enFeLV loads following FeLV-A challenge. However, cats with high enFeLV loadsshowed higher viral replication (plasma viral RNA and p27 antigen levels) than cats with low enFeLV loadsin the early phase of the infection. The enFeLV transcription level varied at different time points, but noclear-cut pattern was observed. In conclusion, our results demonstrated an association between enFeLVloads and FeLV replication but not outcome of infection. enFeLV should be considered as an importantconfounder in experimental FeLV infection or vaccination studies.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Endogenous feline leukemia viruses (enFeLV) are replicationdefective viruses of felids (Benveniste et al., 1975; Benveniste andTodaro, 1975; Roca et al., 2004, 2005). They were most probablyacquired by cats via trans-species infection with a virus of rodentorigin (Benveniste et al., 1975) and hence represent a footprint ofancient retrovirus infection. The enFeLV retain the primer bindingsites, packaging signal, leader sequences (Berry et al., 1988) andopen reading frames in the pol and env gene sequences (Kumaret al., 1989; Pandey et al., 1991). The main sequence differencesbetween enFeLV and their exogenous counterpart, the felineleukemia virus (FeLV), are located in the U3 region of the longterminal repeats (LTR) and in parts of env (Berry et al., 1988;Casey et al., 1981; Kumar et al., 1989). Recombination of FeLV-Awith enFeLV gives rise to FeLV-B that has an altered host range(Overbaugh et al., 1988; Roy-Burman, 1995; Stewart et al., 1986).
No infectious enFeLV virions have been isolated so far althougha subset of defective enFeLV proviruses containing gag, env andLTR or env and LTR is highly expressed in lymphoma cell lines andin a variety of tissues during certain disease states, such as autoim-
∗ Corresponding author. Tel.: +41 44 635 83 22; fax: +41 44 635 89 23.E-mail addresses: [email protected], [email protected]
(R. Hofmann-Lehmann).
mune and malignant conditions (Perl, 2003; Stoye, 2001; Taruscioand Mantovani, 1998). In addition, defective proviruses are alsoexpressed in lymphoid tissues from healthy specific pathogen free(SPF) cats (McDougall et al., 1994). In general, a wide diversity offunctions has been assigned to endogenous retroviruses. While insome studies endogenous retroviruses were supposed to induceimmunological tolerance (Arshad et al., 1997; Cooper et al., 2004;Crittenden et al., 1982, 1984; Payne, 1998; Venugopal, 1999;Woodland et al., 1991) in other cases they were believed to havea protecting role by receptor interference (Boeke and Stoye, 1997;Mura et al., 2004; Spencer et al., 2003). The latter scenario has alsobeen described for FeLV-B where receptor blockage by a truncatedenFeLV envelope protein was suggested to mediate the naturalresistance of cats to infection with FeLV-B in the absence of FeLV-A(McDougall et al., 1994). In contrast, for FeLV-T, a T-cell tropic FeLVvariant with a disrupted receptor binding domain (RBD), Andersonet al. (2000) demonstrated that an enFeLV envelope protein, FeLVinfectivity X-essory protein (FeLIX) together with its appropriatereceptor, Pit1, allows for productive FeLV-T infection. The enFeLVenvelope protein or the RBD thereof seem to behave as a cofactorand in combination with the matching receptor help overcomerestrictions given by the disrupted RBD of FeLV-T. Recently, otherenvelope proteins (of FeLV-A and -C) or receptor binding domainsthereof in combination with their specific receptors have beenshown to permit FeLV-T infection (Anderson et al., 2000; Cheng etal., 2007).
0168-1702/$ – see front matter © 2008 Elsevier B.V. All rights reserved.doi:10.1016/j.virusres.2008.02.016

Author's personal copy
R. Tandon et al. / Virus Research 135 (2008) 136–143 137
We have recently demonstrated differences in enFeLV loadsamong cats of different breeds and between domestic cats andEuropean wildcats (Tandon et al., 2007). In addition, we observedthat FeLV-positive cats had higher enFeLV loads than FeLV-negativecats (Tandon et al., 2007). Based on these results, we hypothesizedthat enFeLV may be involved in the pathogenesis and disease out-come of FeLV infections. To address this hypothesis, we conductedan in vivo FeLV challenge experiment using SPF cats with high andlow enFeLV loads, respectively. The cats were followed throughoutFeLV infection and viral, proviral and FeLV antigen loads and thehumoral immune response were determined. In addition, consid-ering endogenous retroviruses as a class of transposable elements,enFeLV loads were measured throughout FeLV infection to evalu-ate their retrotransposition activity and the influence of FeLV onenFeLV.
2. Materials and methods
2.1. Animals and experimental design
An in vivo experiment was conducted using 20 domestic shorthair SPF cats originating from two different SPF catteries: 10 kittens(3 females, 7 males) from Charles River Laboratories, Lyon, France(cattery A) and 10 kittens (3 females, 7 males) from Liberty ResearchLaboratory, Waverly, NY, USA (cattery B). The latter kittens had beenselected from a larger group of cats for their high enFeLV proviralloads. All kittens were 11 weeks of age at the time of arrival. Thecats were challenged intraperitoneally with 5 × 105 focus-formingunits (FFU) of FeLV-A/Glasgow-1 virus (Jarrett et al., 1973) at theage of 18 weeks. Male cats were castrated and female cats werespayed at the age of 22 weeks. Cats were housed in the faculty’sbarrier facility in two groups in large rooms under optimal etholog-ical and hygienic conditions. For blood collection all cats received amidazolam–ketamine sedoanalgesic mixture; each cat was admin-istered intramuscular doses of 0.1 mg/kg of midazolam (Roche,Reinach, Switzerland) and 10 mg/kg of ketamine (Vetoquinol AG,Bern, Switzerland). Blood samples were collected prior to, at thetime point of challenge (week 0) and weekly thereafter until week15 by jugular venipuncture using evacuated tubes (Becton Dickin-son, Plymouth, UK) containing K3EDTA or Lithium Heparin. Viraland proviral FeLV and enFeLV loads were determined, as well asFeLV antigen loads and the humoral immune response againstFeLV.
2.2. Serology, virus isolation and virus neutralizing antibodies
The presence of plasma FeLV p27 antigen as a measure forviremia was determined using a double antibody sandwich ELISA(Lutz et al., 1983). Samples reaching ≥4% of a defined positive con-trol were designated positive. In agreement with the EuropeanPharmacopoeia (2005), a cat was considered persistently infected(characterized by persistent antigenemia) when it was FeLV p27-positive for three consecutive weeks or on five occasions betweenweeks 3 and 15 after virus exposure. Samples for virus isolationwere collected from all cats at weeks 0, 3, 6, 11 or 12 and 15 andin addition from cats with high enFeLV loads at week 9. The pres-ence of infectious virus was determined in vitro by inoculation ofheparinized plasma samples onto QN10S cells (Jarrett and Ganiere,1996). To detect latency, bone marrow was collected on week 17after challenge and cultured in the presence of 10−6 mol/l hydrocor-tisone for 4 weeks (Lehmann et al., 1991). Plasma virus neutralizingantibodies were measured by focus reduction of FeLV-A/Glasgow-1in QN10S cells (Jarrett and Ganiere, 1996). ELISA-reactive anti-FeLVantibody responses were measured using 0.2 �g of gradient puri-fied FeLV antigen per well as described (Lutz et al., 1980, 1988).
2.3. Quantification of enFeLV proviral and cell-associated RNAloads
Loads of enFeLV provirus and feline glyceraldehyde-3-phosphate dehydrogenase (fGAPDH), of which one copy of apseudogene is present in the genomic DNA of feline cells (Molia etal., 2004), were determined prior to and during the course of theinfection applying quantitative real-time TaqMan PCR assays on anABI PRISM 7700 Sequence Detection System (Applied Biosystems,Rotkreuz, Switzerland) as reported (Tandon et al., 2007). Thefollowing enFeLV targets were measured: two of the assays specif-ically detect the unique region, U3 designated as enFeLV-U3-1and enFeLV-U3-2, respectively. The enFeLV-U3-1 assay recog-nizes five of the nine enFeLV LTR U3 sequences present in theGenBank database (AF226623, M25582, M25425, M21479, andM21480), while the enFeLV-U3-2 assay was designed to amplifythe four remaining sequences (AY364319, AY364318, AY364322,and M21481), the third assay amplifies part of the enFeLV-envgene (Tandon et al., 2007). The latter assay also detects FeLV-B. Forabsolute quantification, enFeLV and fGAPDH DNA standards wereused (Tandon et al., 2007). Copy numbers of enFeLV were dividedby fGAPDH copy numbers to calculate the amount of enFeLV percell. Total enFeLV-U3 copy numbers were calculated by addition ofenFeLV-U3-1 and -U3-2 copy numbers.
In addition, cell-associated enFeLV RNA from the group of catswith low and high enFeLV load was quantified at weeks −4 or−2, 1, 2, 3, 6 and 14 or 15. For absolute quantification of enFeLV-U3-1, a RNA standard was produced as follows: a NotI linearizedenFeLV-U3-1 plasmid (Tandon et al., 2007) was subjected to in vitrotranscription using the large-scale SP6 transcription kit (Novagen,Juro Supply, Lucerne, Switzerland) followed by purification byRNeasy® Mini Kit (Qiagen, Hombrechtikon, Switzerland). In vitrotranscribed RNA was quantified, the RNA copy number was calcu-lated as described (Tandon et al., 2005) and 10-fold serial dilutionswere prepared in PCR grade water with 30 �g/ml of carrier yeasttRNA (Sigma, Fluka GmbH, Buchs, Switzerland). The RNA was storedin 50 �l aliquots at −70 ◦C until use. For absolute quantification,enFeLV-U3-1 copy numbers were divided by fGAPDH mRNA copiesas described (Pepin et al., 2007). For enFeLV-U3-2 and enFeLV-envRNA, no RNA standards were available; the comparative CT methodwas used (Leutenegger et al., 1999; Livak and Schmittgen, 2001)and fGAPDH was used as endogenous reference.
2.4. Quantification of FeLV proviral and plasma viral RNA loads
For determination of FeLV proviral loads, the blood volume con-taining 106 white blood cells was subjected to DNA extraction usingthe MagNa Pure LC Total Nucleic Acid Isolation Kit (Roche diagnos-tics, Basel, Switzerland) as described (Tandon et al., 2005). Viral RNAwas extracted from 140 �l of sterile plasma using the QIAamp ViralRNA Mini Kit (Qiagen). RNA samples were stored at −70 ◦C untiluse. Proviral and plasma viral RNA loads were quantified by real-time TaqMan PCR and RT-PCR as described (Tandon et al., 2005).The FeLV assay was designed to amplify the unique region of theLTR and amplify FeLV-A, -B, and -C, but not enFeLV.
2.5. Detection of FeLV-B
The FeLV-B-specific PCR system was designed to amplifya 2.0 kb portion belonging to the envelope and the 3′LTRof FeLV-B. The specificity of the system was validated usinggenomic DNA prepared from FEA cells carrying molecularly clonedFeLV-A, -B, or -C or from uninfected FEA cells as described(Tandon et al., 2005). The PCR reactions contained 0.4 U ofPhusion High Fidelity DNA Polymerase (Finnzymes, Keilaranta,
Author's personal copy
138 R. Tandon et al. / Virus Research 135 (2008) 136–143
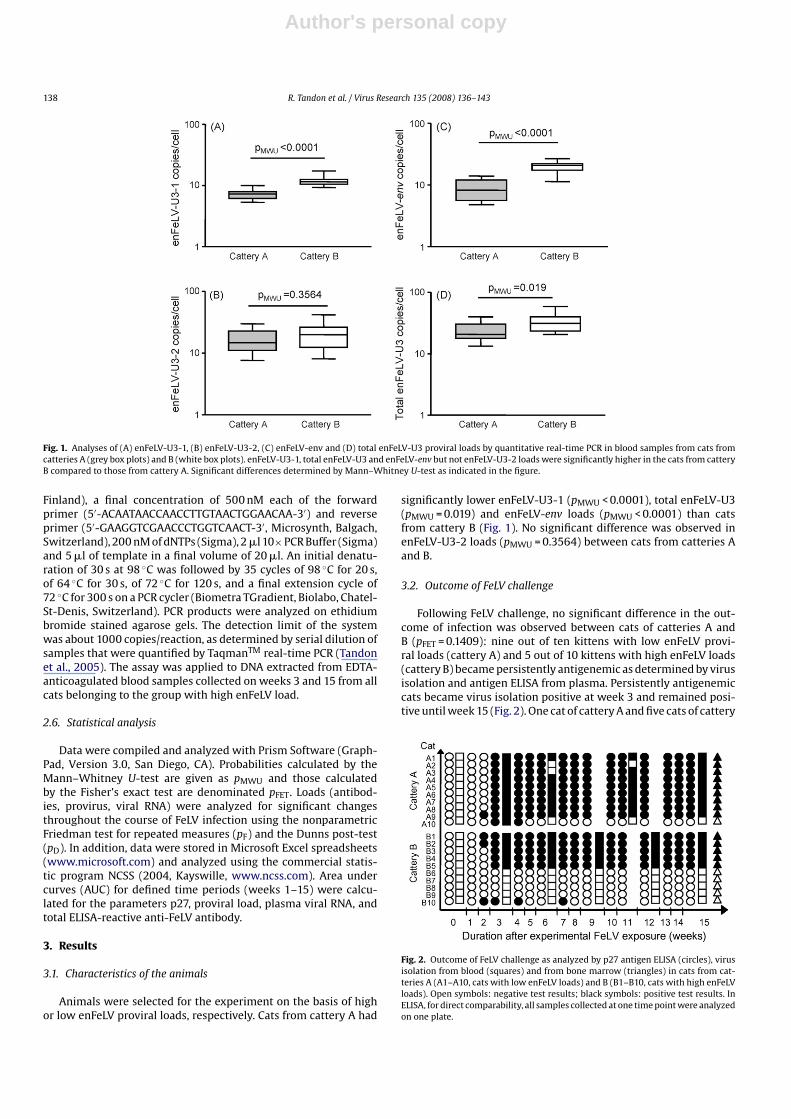
Fig. 1. Analyses of (A) enFeLV-U3-1, (B) enFeLV-U3-2, (C) enFeLV-env and (D) total enFeLV-U3 proviral loads by quantitative real-time PCR in blood samples from cats fromcatteries A (grey box plots) and B (white box plots). enFeLV-U3-1, total enFeLV-U3 and enFeLV-env but not enFeLV-U3-2 loads were significantly higher in the cats from catteryB compared to those from cattery A. Significant differences determined by Mann–Whitney U-test as indicated in the figure.
Finland), a final concentration of 500 nM each of the forwardprimer (5′-ACAATAACCAACCTTGTAACTGGAACAA-3′) and reverseprimer (5′-GAAGGTCGAACCCTGGTCAACT-3′, Microsynth, Balgach,Switzerland), 200 nM of dNTPs (Sigma), 2 �l 10× PCR Buffer (Sigma)and 5 �l of template in a final volume of 20 �l. An initial denatu-ration of 30 s at 98 ◦C was followed by 35 cycles of 98 ◦C for 20 s,of 64 ◦C for 30 s, of 72 ◦C for 120 s, and a final extension cycle of72 ◦C for 300 s on a PCR cycler (Biometra TGradient, Biolabo, Chatel-St-Denis, Switzerland). PCR products were analyzed on ethidiumbromide stained agarose gels. The detection limit of the systemwas about 1000 copies/reaction, as determined by serial dilution ofsamples that were quantified by TaqmanTM real-time PCR (Tandonet al., 2005). The assay was applied to DNA extracted from EDTA-anticoagulated blood samples collected on weeks 3 and 15 from allcats belonging to the group with high enFeLV load.
2.6. Statistical analysis
Data were compiled and analyzed with Prism Software (Graph-Pad, Version 3.0, San Diego, CA). Probabilities calculated by theMann–Whitney U-test are given as pMWU and those calculatedby the Fisher’s exact test are denominated pFET. Loads (antibod-ies, provirus, viral RNA) were analyzed for significant changesthroughout the course of FeLV infection using the nonparametricFriedman test for repeated measures (pF) and the Dunns post-test(pD). In addition, data were stored in Microsoft Excel spreadsheets(www.microsoft.com) and analyzed using the commercial statis-tic program NCSS (2004, Kayswille, www.ncss.com). Area undercurves (AUC) for defined time periods (weeks 1–15) were calcu-lated for the parameters p27, proviral load, plasma viral RNA, andtotal ELISA-reactive anti-FeLV antibody.
3. Results
3.1. Characteristics of the animals
Animals were selected for the experiment on the basis of highor low enFeLV proviral loads, respectively. Cats from cattery A had
significantly lower enFeLV-U3-1 (pMWU < 0.0001), total enFeLV-U3(pMWU = 0.019) and enFeLV-env loads (pMWU < 0.0001) than catsfrom cattery B (Fig. 1). No significant difference was observed inenFeLV-U3-2 loads (pMWU = 0.3564) between cats from catteries Aand B.
3.2. Outcome of FeLV challenge
Following FeLV challenge, no significant difference in the out-come of infection was observed between cats of catteries A andB (pFET = 0.1409): nine out of ten kittens with low enFeLV provi-ral loads (cattery A) and 5 out of 10 kittens with high enFeLV loads(cattery B) became persistently antigenemic as determined by virusisolation and antigen ELISA from plasma. Persistently antigenemiccats became virus isolation positive at week 3 and remained posi-tive until week 15 (Fig. 2). One cat of cattery A and five cats of cattery
Fig. 2. Outcome of FeLV challenge as analyzed by p27 antigen ELISA (circles), virusisolation from blood (squares) and from bone marrow (triangles) in cats from cat-teries A (A1–A10, cats with low enFeLV loads) and B (B1–B10, cats with high enFeLVloads). Open symbols: negative test results; black symbols: positive test results. InELISA, for direct comparability, all samples collected at one time point were analyzedon one plate.

Author's personal copy
R. Tandon et al. / Virus Research 135 (2008) 136–143 139
B were found to have a regressive FeLV infection: one cat of eachcattery showed transient viremia/antigenemia, while four cats ofcattery B never had detectable FeLV p27 antigen and virus isolationwas found negative at all time points tested (Fig. 2). Nonetheless,all cats became FeLV provirus positive indicating that FeLV infectionhad occurred in all cats.
FeLV loads and immune responses were compared between(I) all cats of catteries A and B, and (II) the persistently antigen-emic cats of the two catteries only (progressive infection). Catswith regressive infection could not be statistically analyzed becauseonly one cat with low enFeLV load developed regressive infec-tion.
3.3. High enFeLV loads associated with high viral replication inthe early phase of FeLV infection
No difference was observed in FeLV p27 antigen and plasmaviral RNA loads between all cats from cattery A (low enFeLV loads)and those from cattery B (high enFeLV loads) during the course ofFeLV infection. However, when only persistently antigenemic catswere compared, cats with high enFeLV load had significantly higherFeLV p27 antigen blood levels and FeLV plasma viral RNA loads thancats with low enFeLV loads at several time points throughout theobservation period (Fig. 3A and C). AUC analysis also showed sig-nificant higher level of plasma FeLV p27 antigen and FeLV plasmaviral RNA loads in persistently infected cats with high enFeLV loadsthan in cats with low enFeLV loads. Comparing FeLV proviral loads,all cats from cattery B had significantly higher loads in weeks 1and 2 than those from cattery A; the same was seen when onlypersistently FeLV antigenemic cats were included in the analysis(Fig. 3B). However, no significant difference was observed in theAUC analysis.
3.4. Humoral immune response
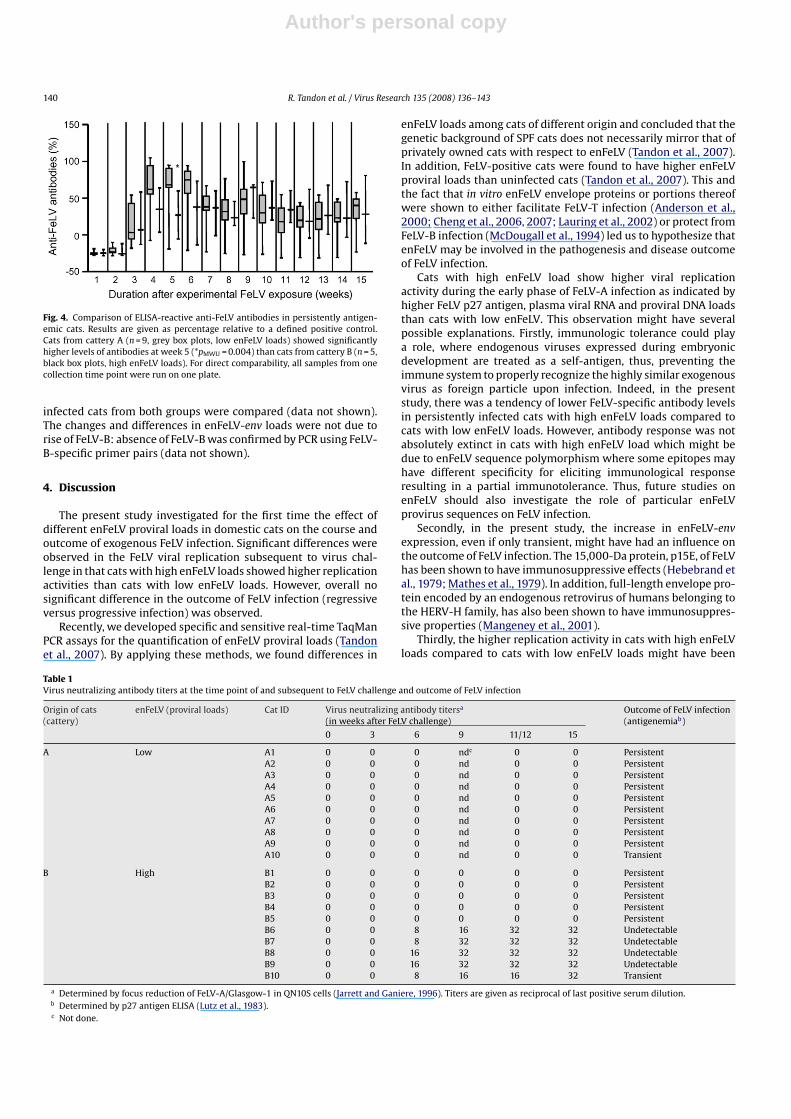
When ELISA-reactive anti-FeLV antibody responses were mea-sured, persistently antigenemic cats from cattery A (low enFeLVloads) showed significantly higher levels at week 5 (pMWU = 0.004)than persistently antigenemic cats from cattery B (high enFeLVloads; Fig. 4). Whereas when all cats from catteries A and B weretaken for analysis, no significant difference in anti-FeLV antibodylevels was observed. In addition, no significant difference wasobserved when AUC analysis was performed either on all cats orpersistently infected cats only.
Virus neutralizing antibody titers were tested at several timepoints (Table 1). The group of cats from cattery A (low enFeLV loads)did not show any virus neutralizing antibody responses althoughthere was one cat with a transient infection. In cats from catteryB (high enFeLV loads), all five cats with regressive infection pro-duced virus neutralizing antibody titers; persistently antigenemiccats remained negative (Table 1).
3.5. Quantification of enFeLV proviral and cell-associated viralRNA loads throughout the course of FeLV infection
enFeLV-U3-1, -U3-2 and -env proviral loads were quantifiedprior to and after FeLV challenge at different time points. Thecats from cattery A (low enFeLV loads) did not show any signifi-cant changes in enFeLV-U3-1, enFeLV-U3-2 and enFeLV-env proviralloads. When cats from cattery B (high enFeLV loads) were observed,there was no significant change in enFeLV-U3-1 and -env proviralloads during the course of FeLV infection. enFeLV-U3-2 loads weresignificantly lower at week 1 than at week 12 (data not shown);however, this difference was due to one cat, which had a very lowenFeLV-U3-2 load at week 1.
Fig. 3. Comparison of (A) FeLV p27 antigen as determined by ELISA, (B) FeLV proviraland (C) plasma viral RNA loads determined by quantitative real-time PCR and RT-PCR, respectively, in persistently antigenemic cats from cattery A (n = 9, grey boxplots, cats with low enFeLV loads) and cattery B (n = 5, black box plots, cats with highenFeLV loads) throughout the first 15 weeks of FeLV infection. Significant differencesin p27 antigen levels between cats from catteries A and B were observed at weeks 2, 3,7, 8, 10, 11, 12 and 13 after infection (*pMWU < 0.01). In FeLV proviral loads, significantdifferences between cats of the two catteries were observed at weeks 1 and 2 afterinfection (*pMWU < 0.01) and in plasma viral RNA loads significant differences werefound at weeks 3, 4, 7, 8, 9 and 10 after infection (*pMWU < 0.0001).
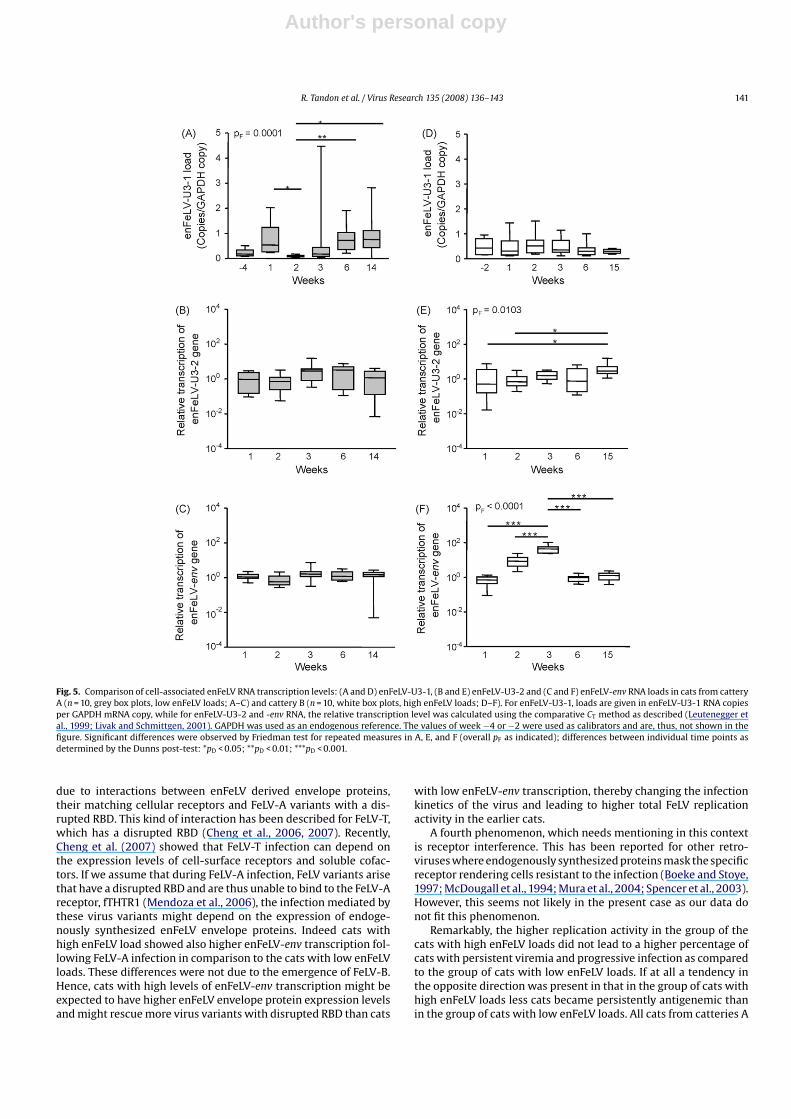
Cell-associated enFeLV RNA was quantified prior to and follow-ing FeLV challenge. enFeLV RNA was detectable in all cats alreadyprior to FeLV infection. Cats from cattery A (low enFeLV loads)showed significantly lower enFeLV-U3-1 RNA loads at week 2 thanat weeks 1, 6 and 14 (Fig. 5A). No significant changes were observedin enFeLV-U3-2 and enFeLV-env RNA loads (Fig. 5B and C) in thesecats and in enFeLV-U3-1 RNA loads in cats from cattery B (highenFeLV loads; Fig. 5D). However, there was significant variation overtime in enFeLV-U3-2 and enFeLV-env RNA loads in cats from cat-tery B (Fig. 5E and F): enFeLV-U3-2 RNA loads were significantlyhigher at week 15 compared to weeks 1 and 2; and enFeLV-envRNA loads increased from weeks 1 to 2 and 3, respectively, beforethey decreased again (Fig. 5F). In addition, cats from cattery B(high enFeLV loads) had significantly higher enFeLV-env RNA loadsat weeks 2 and 3 in comparison to the cats from cattery A (lowenFeLV loads). Similar results were obtained when only persistently
Author's personal copy
140 R. Tandon et al. / Virus Research 135 (2008) 136–143
Fig. 4. Comparison of ELISA-reactive anti-FeLV antibodies in persistently antigen-emic cats. Results are given as percentage relative to a defined positive control.Cats from cattery A (n = 9, grey box plots, low enFeLV loads) showed significantlyhigher levels of antibodies at week 5 (*pMWU = 0.004) than cats from cattery B (n = 5,black box plots, high enFeLV loads). For direct comparability, all samples from onecollection time point were run on one plate.
infected cats from both groups were compared (data not shown).The changes and differences in enFeLV-env loads were not due torise of FeLV-B: absence of FeLV-B was confirmed by PCR using FeLV-B-specific primer pairs (data not shown).
4. Discussion
The present study investigated for the first time the effect ofdifferent enFeLV proviral loads in domestic cats on the course andoutcome of exogenous FeLV infection. Significant differences wereobserved in the FeLV viral replication subsequent to virus chal-lenge in that cats with high enFeLV loads showed higher replicationactivities than cats with low enFeLV loads. However, overall nosignificant difference in the outcome of FeLV infection (regressiveversus progressive infection) was observed.
Recently, we developed specific and sensitive real-time TaqManPCR assays for the quantification of enFeLV proviral loads (Tandonet al., 2007). By applying these methods, we found differences in
enFeLV loads among cats of different origin and concluded that thegenetic background of SPF cats does not necessarily mirror that ofprivately owned cats with respect to enFeLV (Tandon et al., 2007).In addition, FeLV-positive cats were found to have higher enFeLVproviral loads than uninfected cats (Tandon et al., 2007). This andthe fact that in vitro enFeLV envelope proteins or portions thereofwere shown to either facilitate FeLV-T infection (Anderson et al.,2000; Cheng et al., 2006, 2007; Lauring et al., 2002) or protect fromFeLV-B infection (McDougall et al., 1994) led us to hypothesize thatenFeLV may be involved in the pathogenesis and disease outcomeof FeLV infection.
Cats with high enFeLV load show higher viral replicationactivity during the early phase of FeLV-A infection as indicated byhigher FeLV p27 antigen, plasma viral RNA and proviral DNA loadsthan cats with low enFeLV. This observation might have severalpossible explanations. Firstly, immunologic tolerance could playa role, where endogenous viruses expressed during embryonicdevelopment are treated as a self-antigen, thus, preventing theimmune system to properly recognize the highly similar exogenousvirus as foreign particle upon infection. Indeed, in the presentstudy, there was a tendency of lower FeLV-specific antibody levelsin persistently infected cats with high enFeLV loads compared tocats with low enFeLV loads. However, antibody response was notabsolutely extinct in cats with high enFeLV load which might bedue to enFeLV sequence polymorphism where some epitopes mayhave different specificity for eliciting immunological responseresulting in a partial immunotolerance. Thus, future studies onenFeLV should also investigate the role of particular enFeLVprovirus sequences on FeLV infection.
Secondly, in the present study, the increase in enFeLV-envexpression, even if only transient, might have had an influence onthe outcome of FeLV infection. The 15,000-Da protein, p15E, of FeLVhas been shown to have immunosuppressive effects (Hebebrand etal., 1979; Mathes et al., 1979). In addition, full-length envelope pro-tein encoded by an endogenous retrovirus of humans belonging tothe HERV-H family, has also been shown to have immunosuppres-sive properties (Mangeney et al., 2001).
Thirdly, the higher replication activity in cats with high enFeLVloads compared to cats with low enFeLV loads might have been
Table 1Virus neutralizing antibody titers at the time point of and subsequent to FeLV challenge and outcome of FeLV infection
Origin of cats(cattery)
enFeLV (proviral loads) Cat ID Virus neutralizing antibody titersa
(in weeks after FeLV challenge)Outcome of FeLV infection(antigenemiab)
0 3 6 9 11/12 15
A Low A1 0 0 0 ndc 0 0 PersistentA2 0 0 0 nd 0 0 PersistentA3 0 0 0 nd 0 0 PersistentA4 0 0 0 nd 0 0 PersistentA5 0 0 0 nd 0 0 PersistentA6 0 0 0 nd 0 0 PersistentA7 0 0 0 nd 0 0 PersistentA8 0 0 0 nd 0 0 PersistentA9 0 0 0 nd 0 0 PersistentA10 0 0 0 nd 0 0 Transient
B High B1 0 0 0 0 0 0 PersistentB2 0 0 0 0 0 0 PersistentB3 0 0 0 0 0 0 PersistentB4 0 0 0 0 0 0 PersistentB5 0 0 0 0 0 0 PersistentB6 0 0 8 16 32 32 UndetectableB7 0 0 8 32 32 32 UndetectableB8 0 0 16 32 32 32 UndetectableB9 0 0 16 32 32 32 UndetectableB10 0 0 8 16 16 32 Transient
a Determined by focus reduction of FeLV-A/Glasgow-1 in QN10S cells (Jarrett and Ganiere, 1996). Titers are given as reciprocal of last positive serum dilution.b Determined by p27 antigen ELISA (Lutz et al., 1983).c Not done.
Author's personal copy
R. Tandon et al. / Virus Research 135 (2008) 136–143 141
Fig. 5. Comparison of cell-associated enFeLV RNA transcription levels: (A and D) enFeLV-U3-1, (B and E) enFeLV-U3-2 and (C and F) enFeLV-env RNA loads in cats from catteryA (n = 10, grey box plots, low enFeLV loads; A–C) and cattery B (n = 10, white box plots, high enFeLV loads; D–F). For enFeLV-U3-1, loads are given in enFeLV-U3-1 RNA copiesper GAPDH mRNA copy, while for enFeLV-U3-2 and -env RNA, the relative transcription level was calculated using the comparative CT method as described (Leutenegger etal., 1999; Livak and Schmittgen, 2001). GAPDH was used as an endogenous reference. The values of week −4 or −2 were used as calibrators and are, thus, not shown in thefigure. Significant differences were observed by Friedman test for repeated measures in A, E, and F (overall pF as indicated); differences between individual time points asdetermined by the Dunns post-test: *pD < 0.05; **pD < 0.01; ***pD < 0.001.
due to interactions between enFeLV derived envelope proteins,their matching cellular receptors and FeLV-A variants with a dis-rupted RBD. This kind of interaction has been described for FeLV-T,which has a disrupted RBD (Cheng et al., 2006, 2007). Recently,Cheng et al. (2007) showed that FeLV-T infection can depend onthe expression levels of cell-surface receptors and soluble cofac-tors. If we assume that during FeLV-A infection, FeLV variants arisethat have a disrupted RBD and are thus unable to bind to the FeLV-Areceptor, fTHTR1 (Mendoza et al., 2006), the infection mediated bythese virus variants might depend on the expression of endoge-nously synthesized enFeLV envelope proteins. Indeed cats withhigh enFeLV load showed also higher enFeLV-env transcription fol-lowing FeLV-A infection in comparison to the cats with low enFeLVloads. These differences were not due to the emergence of FeLV-B.Hence, cats with high levels of enFeLV-env transcription might beexpected to have higher enFeLV envelope protein expression levelsand might rescue more virus variants with disrupted RBD than cats
with low enFeLV-env transcription, thereby changing the infectionkinetics of the virus and leading to higher total FeLV replicationactivity in the earlier cats.
A fourth phenomenon, which needs mentioning in this contextis receptor interference. This has been reported for other retro-viruses where endogenously synthesized proteins mask the specificreceptor rendering cells resistant to the infection (Boeke and Stoye,1997; McDougall et al., 1994; Mura et al., 2004; Spencer et al., 2003).However, this seems not likely in the present case as our data donot fit this phenomenon.
Remarkably, the higher replication activity in the group of thecats with high enFeLV loads did not lead to a higher percentage ofcats with persistent viremia and progressive infection as comparedto the group of cats with low enFeLV loads. If at all a tendency inthe opposite direction was present in that in the group of cats withhigh enFeLV loads less cats became persistently antigenemic thanin the group of cats with low enFeLV loads. All cats from catteries A

Author's personal copy
142 R. Tandon et al. / Virus Research 135 (2008) 136–143
and B became provirus positive indicating the successful infection.FeLV provirus positivity has been shown to be associated with virusintegration in the genome which is one of the most critical stepsof retroviral replication (Cattori et al., 2006). The failure to detect aclear significant effect might be due to the limited numbers of catsper group or to the rather small difference in enFeLV loads betweenthe two groups of cats. To have groups of cats with significantly dif-ferent enFeLV loads, we had also to obtain them from two differentSPF catteries. This fact added a variation to the already heterogenicgenetic background of the individual cats (which are not available,e.g. like mice as inbred strains) and might have masked part of theeffect due to enFeLV. Nonetheless, to the best of our knowledge, thisis the first study to compare groups of cats with different geneticbackground after experimental FeLV infection within one experi-ment. The male to female ratio was identical in the two groups ofcats, excluding gender as a factor influencing the immune response(Ahmed et al., 1985; Butterworth et al., 1967; London and Drew,1977).
FeLV antigen and plasma viral RNA levels were intermittentlyhigher in cats with high enFeLV loads between 2 and 13 weeks aftervirus challenge, while the proviral load was only increased in thevery early infection (weeks 1 and 2). In week 3, proviral loads hadreached a plateau in all cats independent of the enFeLV load. The dif-ference might be ascribed to the fact that proviruses are integratedinto the genome of the cell up to a certain copy number, when asteady state is reached. In contrast, virus production including tran-scription and shedding of viral particles in the plasma are dynamicprocesses and may be expected to vary in a more pronouncedly waythan the proviral load.
Some differences in the humoral immune response (ELISA-reactive anti-FeLV antibodies) were observed between persistentlyantigenemic cats with low and high enFeLV loads, respectively.However, none of the cats with persistent infection developed virusneutralizing antibodies. Thus, we assume that other factors mayalso play a role in differential modulation of FeLV replication, suchas virus-specific cytotoxic T lymphocyte responses, which mightbe expected as early as 1 week after virus exposure (Flynn et al.,2002)—exactly the time point we started seeing differences in themagnitude of FeLV replication activity. We attempted to measurethe cellular immune responses to FeLV in these cats but we were notsuccessful due to technical problems. As generally known for FeLVinfection, in accordance with earlier reports (Hofmann-Lehmann etal., 2001), most of the cats with regressive infection developed highlevels of virus neutralizing antibodies, but as only one cat with lowenFeLV loads developed regressive infection no comparison waspossible between the two groups of cats with low and high enFeLV.
The variation observed in enFeLV cell-associated RNA could havebeen induced by chemokines, which had been shown to influ-ence the up- or downregulation of other genes (Lee et al., 2007;Ranganathan et al., 2007). The changes observed were not dueto the castration or spaying of the cats: six age matched unchal-lenged control cats, which were treated identical to FeLV exposedanimals, did not show any changes in enFeLV mRNA loads (datanot shown). Thus, no clear explanation can be offered for thedifferences observed in enFeLV RNA expression at this time. Never-theless, it should be worth looking at different genes of enFeLV fortheir ability to translate their RNA into proteins.
In conclusion, the present study was conducted to monitor theeffect of enFeLV loads on FeLV infection. Our results indicate thatthere is no difference in the outcome of the FeLV infection betweenthe cats with high and low enFeLV loads. However, significant differ-ences in viral replication following experimental FeLV-A challengewere observed in association with differences in enFeLV loads inthe two groups of cats. Whether the main influencing factor wasthe enFeLV load or other components of the genetic background
of these two groups of cats cannot be determined at the moment.Nonetheless, our observation that differences in FeLV replicationcan be found between cats of different origin (SPF cattery) shouldbe accounted for when designing experimental FeLV infection andvaccination studies. Finally, in the long run, analyses of more geneticmarkers and their influence on FeLV might lead to breeding of catswith some resistance to FeLV infection.
Acknowledgements
We are grateful to E. Gonczi, V. Fornera, B. Weibel, T. Meili Pro-dan, E. Rogg, E. Schuler, U. Egger, E. Grassli, M. Huder, B. Lange, M.Rios and J. Walchli for excellent technical assistance. The study wasperformed using the logistics of the Centre for Clinical Studies atthe Vetsuisse Faculty of the University of Zurich. This work was sup-ported by a research grant of the Swiss National Science Foundation(31-65231). A.C.P. was supported by a research grant of the Univer-sity of Zurich. R.H.-L. is the recipient of a professorship by the SwissNational Science Foundation (PP00B-102866). These studies wereconducted by R. Tandon as partial fulfilment of the requirementsfor a Ph.D. degree at the University of Zurich.
References
Ahmed, S.A., Penhale, W.J., Talal, N., 1985. Sex hormones and autoimmune disease:mechanisms of sex hormones action. Am. J. Pathol. 121, 531–559.
Anderson, M.M., Lauring, A.S., Burns, C.C., Overbaugh, J., 2000. Identification of acellular cofactor required for infection by feline leukemia virus. Science 287,1828–1830.
Arshad, S.S., Howes, K., Barron, G.S., Smith, L.M., Russell, P.H., Payne, L.N., 1997. Tis-sue tropism of the HPRS-103 strain of J subgroup avian leukosis virus and of aderivative acutely transforming virus. Vet. Pathol. 34, 127–137.
Benveniste, R.E., Sherr, C.J., Todaro, G.J., 1975. Evolution of type C viral genes: originof feline leukemia virus. Science 190, 886–888.
Benveniste, R.E., Todaro, G.J., 1975. Segregation of RD-114 and FeL-V-relatedsequences in crosses between domestic cat and leopard cat. Nature 257,506–508.
Berry, B.T., Ghosh, A.K., Kumar, D.V., Spodick, D.A., Roy-Burman, P., 1988. Structureand function of endogenous feline leukemia virus long terminal repeats andadjoining regions. J. Virol. 62, 3631–3641.
Boeke, J.D., Stoye, J.P., 1997. Retrotransposons, endogenous retroviruses, and theevolution of retroelements. In: Coffin, J.M., Hughes, S.H., Varmus, H.E. (Eds.),Retroviruses. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Butterworth, M.B., McClelland, B., Alan-Smith, M., 1967. Influence of sex onimmunoglobulin levels. Nature 214, 1224–1228.
Casey, J.W., Roach, A., Mullins, J.I., Burck, K.B., Nicolson, M.O., Gardner, M.B., Davidson,N., 1981. The U3 portion of feline leukemia virus DNA identifies horizontallyacquired proviruses in leukemic cats. Proc. Natl. Acad. Sci. U.S.A. 78, 7778–7782.
Cattori, V., Tandon, R., Pepin, A., Lutz, H., Hofmann-Lehmann, R., 2006. Rapid detec-tion of feline leukemia virus provirus integration into feline genomic DNA. Mol.Cell. Probes 20, 172–181.
Cheng, H.H., Anderson, M.M., Hankenson, F.C., Johnston, L., Kotwaliwale, C.V., Over-baugh, J., 2006. Envelope determinants for dual-receptor specificity in felineleukemia virus subgroup A and T variants. J. Virol. 80, 1619–1628.
Cheng, H.H., Anderson, M.M., Overbaugh, J., 2007. Feline leukemia virus T entry isdependent on both expression levels and specific interactions between cofactorand receptor. Virology 359, 170–178.
Cooper, C.J., Turk, G.L., Sun, M., Farr, A.G., Fink, P.J., 2004. Cutting edge: TCR revisionoccurs in germinal centers. J. Immunol. 173, 6532–6536.
Crittenden, L.B., Fadly, A.M., Smith, E.J., 1982. Effect of endogenous leukosis virusgenes on response to infection with avian leukosis and reticuloendotheliosisviruses. Avian Dis. 26, 279–294.
Crittenden, L.B., Smith, E.J., Fadly, A.M., 1984. Influence of endogenous viral (ev)gene expression and strain of exogenous avian leukosis virus (ALV) on mor-tality and ALV infection and shedding in chickens. Avian Dis. 28, 1037–1056.
Feline leukemia vaccine (inactivated) 2005. Monograph of the European Pharma-copoeia Commission, 5th ed., p. 761.
Flynn, J.N., Dunham, S.P., Watson, V., Jarrett, O., 2002. Longitudinal analysis of felineleukemia virus-specific cytotoxic T lymphocytes: correlation with recovery frominfection. J. Virol. 76, 2306–2315.
Hebebrand, L.C., Olsen, R.G., Mathes, L.E., Nichols, W.S., 1979. Inhibition of humanlymphocyte mitogen and antigen response by a 15,000-Da protein from felineleukemia virus. Cancer Res. 39, 443–447.
Hofmann-Lehmann, R., Huder, J.B., Gruber, S., Boretti, F., Sigrist, B., Lutz, H., 2001.Feline leukaemia provirus load during the course of experimental infection andin naturally infected cats. J. Gen. Virol. 82, 1589–1596.

Author's personal copy
R. Tandon et al. / Virus Research 135 (2008) 136–143 143
Jarrett, O., Ganiere, J.P., 1996. Comparative studies of the efficacy of a recombinantfeline leukaemia virus vaccine. Vet. Rec. 138, 7–11.
Jarrett, O., Laird, H.M., Hay, D., 1973. Determinants of the host range of felineleukaemia viruses. J. Gen. Virol. 20, 169–175.
Kumar, D.V., Berry, B.T., Roy-Burman, P., 1989. Nucleotide sequence and distinctivecharacteristics of the env gene of endogenous feline leukemia provirus. J. Virol.63, 2379–2384.
Lauring, A.S., Cheng, H.H., Eiden, M.V., Overbaugh, J., 2002. Genetic and biochemicalanalyses of receptor and cofactor determinants for T-cell-tropic feline leukemiavirus infection. J. Virol. 76, 8069–8078.
Lee, Y.R., Yu, H.N., Noh, E.M., Youn, H.J., Song, E.K., Han, M.K., Park, C.S., Kim, B.S.,Park, Y.S., Park, B.K., Lee, S.H., Kim, J.S., 2007. TNF-alpha upregulates PTEN via NF-kappaB signaling pathways in human leukemic cells. Exp. Mol. Med. 39, 121–127.
Lehmann, R., Franchini, M., Aubert, A., Wolfensberger, C., Cronier, J., Lutz, H., 1991.Vaccination of cats experimentally infected with feline immunodeficiency virus,using a recombinant feline leukemia virus vaccine. J. Am. Vet. Med. Assoc. 199,1446–1452.
Leutenegger, C.M., Mislin, C.N., Sigrist, B., Ehrengruber, M.U., Hofmann-Lehmann, R.,Lutz, H., 1999. Quantitative real-time PCR for the measurement of feline cytokinemRNA. Vet. Immunol. Immunopathol. 71, 291–305.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods 25,402–408.
London, W.T., Drew, J.R., 1977. Antibody response to hepatitis B infection amongpatients receiving chronic dialysis treatment. Proc. Natl. Acad. Sci. U.S.A. 74,2561–2563.
Lutz, H., Arnold, P., Hubscher, U., Egberink, H., Pedersen, N., Horzinek, M.C., 1988.Specificity assessment of feline T-lymphotropic lentivirus serology. Z. Vet. B 35,773–778.
Lutz, H., Pedersen, N., Higgins, J., Hubscher, U., Troy, F.A., Theilen, G.H., 1980. Humoralimmune reactivity to feline leukemia virus and associated antigens in cats nat-urally infected with feline leukemia virus. Cancer Res. 40, 3642–3651.
Lutz, H., Pedersen, N.C., Durbin, R., Theilen, G.H., 1983. Monoclonal antibodies tothree epitopic regions of feline leukemia virus p27 and their use in enzyme-linked immunosorbent assay of p27. J. Immunol. Methods 56, 209–220.
Mangeney, M., de Parseval, N., Thomas, G., Heidmann, T., 2001. The full-lengthenvelope of an HERV-H human endogenous retrovirus has immunosuppressiveproperties. J. Gen. Virol. 82, 2515–2518.
Mathes, L.E., Olsen, R.G., Hebebrand, L.C., Hoover, E.A., Schaller, J.P., Adams, P.W.,Nichols, W.S., 1979. Immunosuppressive properties of a virion polypeptide, a15,000-Da protein, from feline leukemia virus. Cancer Res. 39, 950–955.
McDougall, A.S., Terry, A., Tzavaras, T., Cheney, C., Rojko, J., Neil, J.C., 1994. Defectiveendogenous proviruses are expressed in feline lymphoid cells: evidence for arole in natural resistance to subgroup B feline leukemia viruses. J. Virol. 68,2151–2160.
Mendoza, R., Anderson, M.M., Overbaugh, J., 2006. A putative thiamine transportprotein is a receptor for feline leukemia virus subgroup A. J. Virol. 80, 3378–3385.
Molia, S., Chomel, B.B., Kasten, R.W., Leutenegger, C.M., Steele, B.R., Marker, L.,Martenson, J.S., Keet, D.F., Bengis, R.G., Peterson, R.P., Munson, L., O’Brien, S.J.,2004. Prevalence of Bartonella infection in wild African lions (Panthera leo) andcheetahs (Acinonyx jubatus). Vet. Microbiol. 100, 31–41.
Mura, M., Murcia, P., Caporale, M., Spencer, T.E., Nagashima, K., Rein, A., Palmarini, M.,2004. Late viral interference induced by transdominant Gag of an endogenousretrovirus. Proc. Natl. Acad. Sci. U.S.A. 101, 11117–11122.
Overbaugh, J., Riedel, N., Hoover, E.A., Mullins, J.I., 1988. Transduction of endogenousenvelope genes by feline leukaemia virus in vitro. Nature 332, 731–734.
Pandey, R., Ghosh, A.K., Kumar, D.V., Bachman, B.A., Shibata, D., Roy-Burman, P.,1991. Recombination between feline leukemia virus subgroup B or C andendogenous env elements alters the in vitro biological activities of the viruses[published erratum appears in J. Virol. 1992 June;66(6):3976]. J. Virol. 65,6495–6508.
Payne, L.N., 1998. HPRS-103: a retrovirus strikes back. The emergence of subgroop Javian leukosis virus. Avian Pathol. 27, S36–S45.
Pepin, A.C., Tandon, R., Cattori, V., Niederer, E., Riond, B., Willi, B., Lutz, H., Hofmann-Lehmann, R., 2007. Cellular segregation of feline leukemia provirus and viralRNA in leukocyte subsets of long-term experimentally infected cats. Virus Res.127, 9–16.
Perl, A., 2003. Role of endogenous retroviruses in autoimmune diseases. Rheum. Dis.Clin. North Am. 29, 123–143, vii.
Ranganathan, P., Agrawal, A., Bhushan, R., Chavalmane, A.K., Kalathur, R.K., Taka-hashi, T., Kondaiah, P., 2007. Expression profiling of genes regulated by TGF-beta:differential regulation in normal and tumour cells. BMC Genom. 8, 98.
Roca, A.L., Nash, W.G., Menninger, J.C., Murphy, W.J., O’Brien, S.J., 2005. Insertionalpolymorphisms of endogenous feline leukemia viruses. J. Virol. 79, 3979–3986.
Roca, A.L., Pecon-Slattery, J., O’Brien, S.J., 2004. Genomically intact endogenous felineleukemia viruses of recent origin. J. Virol. 78, 4370–4375.
Roy-Burman, P., 1995. Endogenous env elements: partners in generation ofpathogenic feline leukemia viruses. Virus Genes 11, 147–161.
Spencer, T.E., Mura, M., Gray, C.A., Griebel, P.J., Palmarini, M., 2003. Receptorusage and fetal expression of ovine endogenous betaretroviruses: implica-tions for coevolution of endogenous and exogenous retroviruses. J. Virol. 77,749–753.
Stewart, M.A., Warnock, M., Wheeler, A., Wilkie, N., Mullins, J.I., Onions, D.E., Neil,J.C., 1986. Nucleotide sequences of a feline leukemia virus subgroup A envelopegene and long terminal repeat and evidence for the recombinational origin ofsubgroup B viruses. J. Virol. 58, 825–834.
Stoye, J.P., 2001. Endogenous retroviruses: still active after all these years? Curr. Biol.11, R914–R916.
Tandon, R., Cattori, V., Gomes-Keller, M.A., Meli, M.L., Golder, M.C., Lutz, H., Hofmann-Lehmann, R., 2005. Quantitation of feline leukaemia virus viral and proviral loadsby TaqMan real-time polymerase chain reaction. J. Virol. Methods 130, 124–132.
Tandon, R., Cattori, V., Willi, B., Meli, M.L., Gomes-Keller, M.A., Lutz, H., Hofmann-Lehmann, R., 2007. Copy number polymorphism of endogenous feline leukemiavirus-like sequences. Mol. Cell. Probes 21, 257–266.
Taruscio, D., Mantovani, A., 1998. Human endogenous retroviral sequences: possibleroles in reproductive physiopathology. Biol. Reprod. 59, 713–724.
Venugopal, K., 1999. Avian leukosis virus subgroup J: a rapidly evolving group ofoncogenic retroviruses. Res. Vet. Sci. 67, 113–119.
Woodland, D.L., Happ, M.P., Gollob, K.J., Palmer, E., 1991. An endogenous retrovirusmediating deletion of alpha beta T cells? Nature 349, 529–530.