apports fluviaux en zone côtière et réseaux trophiques marins benthiques
TRANSCRIPT

UNIVERSITE DE LA MEDITERRANEE (AIX-MARSEILLE II) - CENTRE D'OCEANOLOGIE DE MARSEILLE -
THESE de DOCTORAT spécialité "Biosciences de l'Environnement, Chimie et Santé"
présentée par : Audrey DARNAUDE
Soutenue à Marseille, le 14 mars
Pr. Charles-François BOUDPr. Ivan DEKEYSER, présiDr. Mireille HARMELIN-VDr. Françoise LAGARDERDr. Capucine MELLON, exPr. Nicholas POLUNIN, rapDr. Chantal SALEN-PICAR
- STATION MARINE D'ENDOU
APPORTS FLUVIAUX EN ZTROPHIQUES MA
TRANSFERT DE MATIERE TERRIGENE JUSQU'AUX POISSO
N° attribué par la bibliothèque : ……..………...
ONE COTIERE ET RESEAUXRINS BENTHIQUES :
ORGANIQUE PARTICULAIRENS PLATS AU LARGE DU RHONE
2003, devant le jury composé de :
OURESQUE, examinateurdent du jury IVIEN, directrice de thèseE, rapporteuraminateurporteurD, directrice de thèse
ME, UMR CNRS DIMAR 6540 -


Photo de couverture : Rolland Graille


UNIVERSITE DE LA MEDITERRANEE (AIX-MARSEILLE II) - CENTRE D'OCEANOLOGIE DE MARSEILLE -
THESE de DOCTORAT spécialité " Biosciences de l'Environnement, Chimie et Santé "
présentée par : Audrey DARNAUDE
Soutenue à Marseille, le 14 mars
Pr. Charles-François BOUPr. Ivan DEKEYSER, préDr. Mireille HARMELINDr. Françoise LAGARDEDr. Capucine MELLON, Pr. Nicholas POLUNIN, rDr. Chantal SALEN-PICA
- STATION MARINE D'ENDOU
APPORTS FLUVIAUX EN Z
TROPHIQUES MA
TRANSFERT DE MATIERE
TERRIGENE JUSQU'AUX POISSO
ONE COTIERE ET RESEAUX
RINS BENTHIQUES :
ORGANIQUE PARTICULAIRE
NS PLATS AU LARGE DU RHONE
2003, devant le jury composé de :
DOURESQUE, examinateursident du jury-VIVIEN, directrice de thèseRE, rapporteurexaminateurapporteurRD, directrice de thèse
ME, UMR CNRS DIMAR 6540 -


La vérité que je révère, c'est la
modeste vérité de la science, la vérité
relative, fragmentaire, provisoire,
toujours sujette à retouche, à
correction, à repentir : la vérité à
notre échelle…
- Jean Rostand -


REMERCIEMENTS
Avant de présenter les résultats de ce travail, je désire remercier un certain nombre de
personnes qui, de près ou de loin, ont contribué à sa réalisation.
Cette thèse a été essentiellement effectuée au Centre d'Océanologie de Marseille (COM), au
sein de l'unité mixte de recherche CNRS DIMAR 6540 basée à la Station Marine d'Endoume. Je tiens
donc à remercier ici Messieurs Charles-François Boudouresque, directeur de l'unité DIMAR, Lucien
Laubier, ancien directeur du COM, et Ivan Dekeyser, son récent successeur, de m'avoir accueilli au
sein de leurs structures de recherche. Mais c'est avant tout un grand plaisir pour moi que de remercier
tout particulièrement Mesdames Chantal Salen-Picard et Mireille Harmelin-Vivien, mes deux
directrices de thèse, qui sont à l'origine de ces recherches et qui ont su, pendant toute leur durée, non
seulement me guider et m'enrichir de leurs conseils, mais également me soutenir et m'entourer de
beaucoup de chaleur et de sympathie. J'ai énormément appris à leur contact et leur suis extrêmement
reconnaissante de la gentillesse, de la patience et de la disponibilité dont elles ont fait preuve à mon
égard depuis notre première rencontre, il y a maintenant quatre ans.
Ce travail a fait l'objet, pour les années 2000-2001, d'un financement partiel du Programme
National "Environnement Côtier" (PNEC, ART II) qui a permis la mise en place, dans le cadre de cette
thèse, d'une double collaboration scientifique avec le CREMA-L'HOUMEAU (UMR 10 CNRS-
IFREMER) et l’Université de Newcastle-upon-Tyne (U.K.). A ce propos, je souhaite transmettre ici
mes très sincères remerciements à Mesdames Françoise Lagardère et Gillian Taylor ainsi qu'à
Monsieur Nicholas Polunin pour leurs accueils chaleureux et le temps qu'ils ont bien voulu me
consacrer au cours des séjours successifs que j'ai effectués au sein de ces deux laboratoires.
Je tiens également à remercier Mesdames Françoise Lagardère et Capucine Mellon ainsi que
Messieurs Nicholas Polunin, Charles-François Boudouresque et Ivan Dekeyser d'avoir accepté de
juger ce travail, en dépit d'emplois du temps souvent fort chargés. Je suis particulièrement honorée de
la présence, dans ce jury, de Madame Françoise Lagardère et de Monsieur Nicholas Polunin. Leurs
expériences respectives de l'écologie des poissons plats et de l'utilisation des isotopes stables en milieu
marin les désignaient tout naturellement comme rapporteurs de cette thèse. Je leur sais gré d'avoir
accepté ce rôle malgré la charge de travail que cela représente.
Les conditions d’échantillonnage au large de l'embouchure du Rhône ne sont pas vraiment
paradisiaques, surtout lorsque l'on est, comme moi, particulièrement sensible au mal de mer. Le
soutien apporté, souvent dans des conditions extrêmes, par les membres d'équipage du Pr Georges
Petit, les plongeurs de la Station Marine d'Endoume, et les nombreux volontaires qui ont bien voulu
m’assister au cours de cette étape de ma thèse restera longtemps gravé en moi. Merci à vous tous pour
votre efficacité, votre professionnalisme et surtout pour votre extrême gentillesse.

La réalisation d'une étude pluridisciplinaire résulte avant tout d'un travail d'équipe. Je tiens donc
à remercier ici tous ceux qui, par leur aide logistique, leurs conseils ou leurs connaissances m'ont
permis de mener à bien ce travail. Merci notamment à Françoise Cubizolles, Christian Marschall,
Laurent Cortijos, David Mouillot, Jean-Antoine Tomasini et Yolande Bentosela pour leurs aides
précieuses dans la récolte et le traitement des échantillons, l'analyse statistique des données et la
recherche des articles scientifiques. J'en profite pour associer à ces remerciements, tous les membres
de la Station Marine d'Endoume qui, par un quelconque moyen, m'ont aidé à arriver jusqu'ici…
Enfin, un énorme merci à tous mes proches, parents et amis, qui m'ont supportée et soutenue sur
le plan affectif et humain, notamment pendant la longue et rude période de rédaction et de mise en
page de ce manuscrit. Quelque part, ce travail est aussi un peu le leur et je leur suis profondément
reconnaissante pour l'aide, tant morale que matérielle, qu'ils ont bien voulu m'apporter au cours de ces
trois années d'efforts continus.



SOMMAIREINTRODUCTION GENERALE ……………………………………………………………………..……1
PARTIE I : ZONE D’ETUDE ET ECHANTILLONNAGE ………………………………………5
I.1 - MILIEU ET PEUPLEMENTS MACROBENTHIQUES : CARACTERISTIQUES GENERALES ……………………………………………………………………………………………..6
I.1.1 - FACTEURS ABIOTIQUES ………………………………………………………………………………..6I.1.2 - PRODUCTION PLANCTONIQUE …………………………………………………….………… ……….9I.1.3 - COMMUNAUTES MACROBENTHIQUES …………………………….…………………………………...10I.1.4 - CONCLUSION SUR LA ZONE D'ETUDE …………………………………...…………………………….12
I.2 - POISSONS : CHOIX DES ESPECES ETUDIEES …………………………….…………………….13
I.2.1 - GENERALITES SUR LES CINQ ESPECES .…………………………….………………………..13
I.2.1.1- Solea solea ……………………….…………………………………………………………………14I.2.1.2 - Solea impar …………………………………………………………………..……………………..17I.2.1.3 - Buglossidium luteum ……………………………………………………………………………….19I.2.1.4 - Arnoglossus laterna …………………………………………………………….…………………..21I.2.1.5 - Citharus linguatula ……………………………………………………………..…………………..23
I.2.2 - POURQUOI CES CINQ POISSONS PLATS ? …………………………..………………………………….25
I.3 - ECHANTILLONNAGE ………………………………………………...……………………………..26
I.3.1 - STRATEGIE D'ECHANTILLONNAGE ……………………………………….………………………….27I.3.2 - CAPTURE DES POISSONS ………………………………………………….………………………….29I.3.3 - ECHANTILLONNAGE POUR L'ETUDE DES RESEAUX TROPHIQUES ……………..…………………….33
I.3.3.1 Récolte d'eau et de sédiment …………………………………………………………...……………33I.3.3.2 Récolte des proies benthiques ……………………………………………………….………………35
I.3.4 – CONCLUSION SUR L'ECHANTILLONNAGE REALISE …………………………..………….38
PARTIE II : REPARTITION BATHYMETRIQUE, ALIMENTATION ETINTERACTIONS SPATIO-TEMPORELLES DES POISSONS PLATS AU LARGEDU RHONE ……………………………………………..………...……………..……………………………..39
CHAPITRE I : REPARTITION SPATIO-TEMPORELLE DES 5 ESPECES ETUDIEES ………..39
II/I.1 – INTRODUCTION …………………………………………………………………………..….…..39II/I.2 – MATERIEL ET METHODES ……..…………………………………………….………………..39
II/I.2.1 – DENSITES ET ABONDANCES RELATIVES ………….…..………………………………………….39II/I.2.2 – POPULATIONS ……………………………………………………………..………………………41
II/I.3 – RESULTATS …..……………………………………………………………..…………………….44
II/I.3.1 – DENSITES ET ABONDANCES RELATIVES DES CINQ ESPECES ……………………………….….44II/I.3.2 – POPULATIONS DES CINQ ESPECES : STRUCTURE ET REPARTITION SPATIO-TEMPORELLE …..…………………………………………………………………...……47
II/I.4 – DISCUSSION ………………………………………………………………………………………..56II/I.5 – CONCLUSION …………………………………………………….……………………………...58
CHAPITRE II : STRATEGIES ALIMENTAIRES DES 5 ESPECES ..……………….………….……59

II/II.1 – INTRODUCTION …………………………………………………….……………………….…..59II/II.2 – MATERIEL ET METHODES …………………………………..…….……………………….…60II/II.3 – ALIMENTATION DES CINQ ESPECES ………………………….….………………………...68
II/II.3.1 – SOLEA SOLEA ………………………………………………..……….…………………….…..68.
II/II.3.1.1 – Alimentation globale ……...……………………………………………...……………….…68II/II.3.1.2 – Modifications du régime alimentaire ……..…………………………………………….…..70II/II.3.1.3 – Discussion sur l'alimentation de S. solea …………………………………………………..79
II/II.3.2 – SOLEA IMPAR ….……………….…………………………………...…………………….…..84
II/II.3.2.1 – Alimentation globale ………………….……………………………………….….…….……84II/II.3.2.2 – Modifications du régime alimentaire ……………………..……………….…………….…..86II/II.3.2.3 – Discussion sur l'alimentation de S. impar ………..…………….………………… …….…..93
II/II.3.3 – BUGLOSSIDIUM LUTEUM .…….……………………………….……………………….…....96
II/II.3.3.1 – Alimentation globale …....………………………………………………………….….…..…96II/II.3.3.2 – Modifications du régime alimentaire .……………………………….………...……….…....98II/II.3.3.3 – Discussion sur l'alimentation de B. luteum ….….…………..……………….……….….….107
II/II.3.4 – ARNOGLOSSUS LATERNA ……………………………………..…………………..……..…..110
II/II.3.4.1 – Alimentation globale …………………………………………….…………. …….……..…110II/II.3.4.2 – Modifications du régime alimentaire ….……………………….…………………....….…..112II/II.3.4.3 – Discussion sur l'alimentation d'A. laterna ……… ……………..……………..………..…..119
II/II.3.5 – CITHARUS LINGUATULA …………..…………………………….……………………………..122
II/II.3.5.1 – Alimentation globale …………………..……………………………………………………122II/II.3.5.2 – Modifications du régime alimentaire ……………..………………………...…….………..125II/II.3.5.3 – Discussion sur l'alimentation de C. linguatula …………..………………………….……...132
II/II.4 – CONCLUSION ……………………...………………………...………………………………….134
CHAPITRE III : PARTAGE DES RESSOURCES ALIMENTAIRES ………….……………………139
II/III.1 – INTRODUCTION ……………………………………..…………………..……………………139II/III.2 – MATERIEL ET METHODES ………………………....……………………………………….140
II/III.2.1 – CALCULS DES LARGEURS DE NICHES ALIMENTAIRES …..……………………………………140II/III.2.2 – RECOUVREMENT DES NICHES ALIMENTAIRES ……………..……………………………...…142II/III.2.3 – PRISE EN COMPTE DE LA TAILLE DES PROIES INGEREES
.…………………………….………145
II/III.3 – PARTAGE DES RESSOURCES ALIMENTAIRES ENTRE ESPECES ...…..……….……145
II/III.3.1 – COMPETITION POTENTIELLE POUR LES RESSOURCES ALIMENTAIRES …………….…………145II/III.3.2 – PARTAGE EFFECTIF DES RESSOURCES ALIMENTAIRES .....…………………………….…..…148
II/III.3.2.1 – Partage des ressources alimentaires sur les petits fonds sableux (0-20 m) ..………….…..149II/III.3.2.2 – Partage des ressources alimentaires sur les vases terrigènes de haut niveau (30-50 m) …154II/III.3.2.3 – Partage des ressources alimentaires sur les vases terrigènes profondes (70-100 m) ..…....158
II/III.4 – CONCLUSION .………………………………………………………………………………….161
PARTIE III : RESEAUX TROPHIQUES ET SUIVI DE LA MATIERE ORGANIQUEPARTICULAIRE TERRIGENE ..………………………..…………………….…......…….…….…….163

III.1 – INTRODUCTION ….…………………………………………..……………………….…...…….…163III.2 - MATERIEL ET METHODES ……….…………………………..……...….………….…...………165
.
III.2.1 - L'OUTIL "ISOTOPES STABLES" …………………………………..……… . ….……….…….……165III.2.2 - PREPARATION DES ECHANTILLONS ……….……………………..………………….…………….170III.2.3 - ANALYSES ISOTOPIQUES ….……………………………………..………………….……………...171III.2.4 - EXPLOITATION DES DONNEES …….……………………………………..………….………..……172
III.3 – SOURCE ET COMPOSITION DE LA MATIERE ORGANIQUE AU LARGE DU RHONE ..177III.4 - MOP TERRIGENE ET RESEAUX TROPHIQUES BENTHIQUES ……………...….…………184
III.4.1 - INCORPORATION DE MOP TERRIGENE DANS LES RESEAUX TROPHIQUES ? ………………….…....186III.4.2 - TRANSFERT DE MOP TERRIGENE JUSQU'AUX POISSONS ? ……………….…………………..……190
III.4.2.1 - Solea solea ………………………………… …………………………………………………192III.4.2.2 - Solea impar ……………………………………… ……………………………………………198III.4.2.3 - Buglossidium luteum ………………………………………..……………………..……………203III.4.2.4 - Arnoglossus laterna …….……………………………………………………………………….211III.4.2.5 - Citharus linguatula ……………………………………………………....……………………..218
III.5 – DISCUSSION ………………………………………………………………….……………….…….223
III.5.1 - SOURCES POTENTIELLES ET ORIGINE DE LA MATIERE ORGANIQUE ………………………………223III.5.2 - RESEAUX TROPHIQUES BENTHIQUES ………………………………………………………...……233III.5.3 - POISSONS PLATS : ALIMENTATION, SIGNATURE ISOTOPIQUE ET EXPLOITATION DE MOP
TERRIGENE …………………………………………………………………..………………….…242
III.6 - CONCLUSION …………………………………...………………………………………….……….248
PARTIE IV : DISCUSSION SUR L'INFLUENCE DES APPORTS RHODANIENS ETCONCLUSIONS GENERALES …………………………………………………………..………….….251
IV.1 - STRATEGIES DE VIE BENTHIQUE ET IMPACT POTENTIEL DES APPORTSRHODANIENS EN MOP TERRIGENE SUR LES CINQ ESPECES ………………..…………………253
IV.1.1 - STRATEGIES DE VIES ET PARTAGE DES RESSOURCES ALIMENTAIRES ……………….……………253IV.1.2 - STRATEGIES DE VIE ET INTENSITE DU TRANSFERT DE MOP TERRIGENE ………………………….260
IV.2 - CONSEQUENCES POTENTIELLES DES CRUES DU FLEUVE SUR LE CYCLE DE VIEET LA DYNAMIQUE DE POPULATION DES POISSONS PLATS …………………..………………265
IV.2.1 - APPORTS RHODANIENS EN MATERIEL DISSOUS …………………………………………….…..…266IV.2.2 - POLLUTION ASSOCIEE ……………………………………………………………………….……..269IV.2.3 - BIOLOGIE ET DELAIS DE REPONSE DES ESPECES ………………………………………..….…….270
IV - CONCLUSION GENERALE ET PERSPECTIVES ……………………………………….……….278
REFERENCES BIBLIOGRAPHIQUES ……………………………………….………………….…281
LISTE DES FIGURES ET TABLEAUX ……………………………………….………………..…...299
ANNEXES………………………..……………………………………………………………………………313
PUBLICATIONS, COMMUNICATIONS ORALES ET POSTERS DIRECTEMENTISSUS DE CE TRAVAIL ..………………………….……………………………………….……………373


INTRODUCTION GENERALE


Introduction générale
1
INTRODUCTION GENERALE
Les zones marines côtières situées à l’embouchure des grands fleuves présentent un intérêt
biologique majeur. Comme beaucoup de zones de fronts hydrologiques, elles sont caractérisées par
d'importantes concentrations en nutriments et une forte production primaire (Largier, 1993). Leur
niveau de production secondaire est très élevé, les communautés animales étant le plus souvent
composées d'espèces à croissance rapide et à forte productivité (Wolff, 1983). De plus, elles constituent
un lieu de passage obligatoire pour de nombreux organismes marins à un moment particulier de leur
cycle biologique (Day et al., 1989), ce qui contribue encore à accroître l'importance de la biomasse
présente. L'ensemble de ces caractéristiques place les zones marines côtières situées à l'embouchure des
fleuves parmi les écosystèmes les plus productifs de la planète (Mann, 1982) et explique l'intense
exploitation dont elles font l'objet dans le monde entier. Dans le contexte socio-économique actuel, avec
l'émergence d'une conscience collective de la nécessité d'une gestion rationnelle des ressources vivantes
marines, mieux comprendre leur fonctionnement est devenu une priorité. Sur le plan scientifique, cette
problématique s'inscrit dans le cadre plus vaste de l'étude du transfert de la matière organique aux
interfaces continent-océan qui constitue actuellement l'un des problèmes majeurs posés aux écologistes.
En effet, dans les zones marines sous influence fluviale, l'origine de la matière organique est complexe
et très variable (Mann, 1982). Les apports continentaux s'y superposent aux éléments nutritifs déjà
présents dans le milieu marin et influencent de façon considérable la productivité et le fonctionnement
des écosystèmes. Les crues saisonnières injectent également, au niveau du fond, de fortes quantités de
matière organique particulaire et divers débris végétaux d’origine terrestre. L’eutrophisation1 qui
découle de l’ensemble de ces processus a un impact certain sur la quantité de nourriture disponible dans
le milieu. Son influence sur la dynamique temporelle des communautés marines à l’embouchure peut
s’avérer primordiale, notamment dans le cas de mers semi-ouvertes ou oligotrophes comme la
Méditerranée (Caddy, 1993, 2000).
Apports fluviaux et pêcheries côtières
Le rôle joué par les apports fluviaux dans l’augmentation de la production des pêcheries côtières
a été reconnu dans de nombreuses régions (Kerr et Ryder, 1992) et des relations positives ont pu, à
moyen terme, être mises en évidence entre le débit de certains grands fleuves et les captures d’espèces
1 Eutrophisation : enrichissement du milieu en matière organique.

Introduction générale
2
d'intérêt commercial (Sutcliffe et al., 1976 ; Lloret et al., 2001). L’abondance des poissons est
généralement reliée à la production primaire, elle même liée en partie à des fluctuations globales du
climat océanique (Cushing, 1982 ; Nielsen et Richardson, 1996). Bien que complexes, des relations
entre production primaire et pêcheries pélagiques ont été démontrées dans différentes régions du globe
(Cushing, 1995 ; Yáñez et al., 1998). Le lien entre production primaire et fluctuations des pêcheries
démersales est moins évident. Si l'importance de la production planctonique influence les stades de vie
larvaire des poissons démersaux et conditionne le succès de leur recrutement (Leggett et DeBlois, 1994
; Bailey et al., 1995), d’autres facteurs sont susceptibles d'intervenir dans la dynamique de population de
ces espèces. L’augmentation des ressources alimentaires au niveau du benthos peut favoriser, dans
certains cas, les stades de vie benthiques des poissons (Salen-Picard et al., 2002). Par un impact positif
sur la condition des adultes et donc le succès reproducteur des espèces concernées, elle aurait une
influence sur leur taux de recrutement et, de là, sur leur dynamique de population. L’hypothèse selon
laquelle les fluctuations des pêcheries démersales seraient en partie liées aux fluctuations des
communautés benthiques a été suggérée par différents auteurs (Horwood, 1993 ; Millner et Whiting,
1996). En général, les variations d'abondance du macrobenthos marin sont reliées à des fluctuations de
la production phytoplanctonique in situ (Beukema, 1991 ; Josefson et al., 1993 ; Frid et al., 1996 ;
Josefson et Conley, 1997). Peu d'études (Salen-Picard et al., 1997 ; Salen-Picard et al., in press)
prennent en compte la contribution de la matière organique particulaire apportée par les fleuves dans les
fluctuations de ces communautés. Elle semble pourtant avoir une influence majeure sur la composition
et l'abondance du macrobenthos au large des embouchures (Salen-Picard et Arlhac, 2002).
Le contexte du golfe du Lion
Le golfe du Lion fournit, à lui seul, 90% des débarquements des pêcheries françaises
méditerranéennes (Campillo et al., 1989). Cette importance halieutique découle essentiellement de la
productivité relativement élevée des marges côtières de la zone (Campillo et Bigot, 1992), sous
l'influence directe des apports rhodaniens. Principal fleuve méditerranéen depuis la construction du
barrage d’Assouan sur le Nil, le Rhône est responsable de 50% de la production primaire du golfe du
Lion et joue un rôle prépondérant dans la productivité du bassin méditerranéen Nord-Occidental (Lochet
et Leveau, 1990). Ses apports en sels nutritifs et en matière organique dissoute sont élevés (Blanc et al.,
1969 ; Moutin et al., 1998) et favorisent, chaque année, à la fois la production planctonique (Blanc et
Leveau, 1971 ; Gaudy et al., 1996) et la production benthique (Salen-Picard et al., 1997 ; Massé, 1998)
au large de son delta. Cependant, les apports du Rhône en carbone organique particulaire, nettement
plus variables d’une année sur l’autre que les apports en matériel dissous, constituent l’essentiel du pool
de matière organique véhiculé par le fleuve en période de forte crue (Cauwet, 1996 ; Sempéré et al.,
2000). Les fluctuations inter-annuelles de débit du Rhône se répercutent donc principalement sur ses
apports en matière organique particulaire (MOP). L'essentiel de la MOP d'origine terrestre sédimente au

Introduction générale
3
niveau du prodelta2 du fleuve (Bouloubassi et al., 1997) où son assimilation par les organismes
dépositivores et son incorporation dans les réseaux trophiques benthiques pourraient intervenir pour une
grande part dans les fluctuations inter-annuelles du macrobenthos. Les variations d’abondance des
communautés benthiques à l’embouchure, ont été reliées aux variations hydrologiques du Rhône (Salen-
Picard et Arlhac, 2002) et seraient susceptibles d'avoir des répercussions sur l'abondance des poissons
démersaux situés en bout de chaîne alimentaire. Ce phénomène permettrait d’expliquer la relation
positive observée entre les fluctuations du débit du Rhône et les débarquements de certains poissons
plats dans le golfe du Lion (Salen-Picard et al., 2002).
L’étude réalisée
Le but de ce travail de thèse est d'étudier le devenir de la matière organique particulaire (MOP)
apportée par le Rhône au sein des communautés macrobenthiques à son embouchure afin d'estimer les
réactions potentielles des principales espèces de poissons plats (Solea solea, Solea impar, Buglossidium
luteum, Arnoglossus laterna, Citharus linguatula) vivant au large du fleuve à une augmentation de son
débit. Il a pour objectif de déterminer si la MOP terrestre est ou non incorporée au sein des réseaux
trophiques benthiques aboutissant à ces cinq espèces et, si tel est le cas, de décrire comment se fait son
transfert le long des réseaux trophiques :
- existe-t-il une voie de transfert trophique privilégiée, associée à un régime alimentaire
précis des poissons ?
- à quel(s) niveau(x) du cycle de vie des espèces (juvénile / adultes) l'incorporation de
matériel d'origine terrestre intervient-elle ?
- existe-t-il des différences de transfert selon la répartition bathymétrique des poissons ?
La réponse à ces questions devrait permettre d'estimer les sensibilités des différentes espèces de
poissons plats étudiées à une augmentation des apports du Rhône et d'émettre des hypothèses sur les
réponses possibles de leurs populations suite à une période de fortes crues du fleuve.
Cette étude nécessite non seulement de connaître les modalités du partage de l'habitat et des
ressources alimentaires entre individus des cinq espèces au large de l'embouchure, mais également de
pouvoir reconstituer le cheminement de la MOP terrigène au sein des réseaux trophiques de ces
poissons. Elle a été menée suivant deux approches complémentaires, reprises ici pour l'exposé des
résultats après une rapide présentation de la zone d'étude et de l'échantillonnage (Partie I). Dans un
premier temps (Partie II), l'étude de la répartition spatio-temporelle (Chapitre I) et de l’alimentation
(Chapitre II) des juvéniles et adultes des cinq poissons plats a été réalisée à l'embouchure afin de
déterminer les interactions spatio-temporelles des différents stades de vie benthique des cinq poissons et
2 Prodelta : partie d'un delta alluvial située sous le niveau de la mer, sur le plateau continental.

Introduction générale
4
leur compétition trophique (Chapitre III). Dans un second temps (Partie III), le suivi du devenir du
carbone autochtone (produit par le phytoplancton marin) et du carbone allochtone (d’origine terrestre
apporté par le fleuve) au sein des réseaux trophiques benthiques aboutissant aux juvéniles et aux adultes
des cinq espèces de poissons plats étudiées a été réalisé à l'aide des isotopes stables du carbone (δ13C)
et de l'azote (δ15N). Les résultats de ces deux approches ont ensuite été confrontés (Partie IV) afin de
déterminer l'influence des stratégies de vie des poissons sur leur sensibilité aux apports du Rhône et
l'impact de ces derniers sur leurs populations.

PARTIE I
ZONE D'ETUDE ET ECHANTILLONAGE


Zone d’étude et échantillonnage
5
FRANCE
Rhône
Golfede Fos
Marseille
They de laGracieuse
Iles duFrioul
MEDITERRANEE (golfe du Lion)
Zoned'étude
ZONE D’ETUDE ET ECHANTILLONNAGE
La zone géographique retenue pour cette étude est située dans la partie Nord-Est du golfe du
Lion (Méditerranée nord-occidentale), à environ 50 km à l’Ouest de Marseille (Fig. I.1). Elle s'étend à
l'Est de l'embouchure du Grand Rhône, entre 4°52’ et 5°01’ de longitude Est et entre 43°14’ et 43°22’
de latitude Nord. L'aire d'échantillonnage couvre les fonds marins de 0 à 100 m de profondeur situés au
large du they de la Gracieuse, à proximité du golfe de Fos. La détermination précise de la localisation de
l'aire d'échantillonnage par rapport à l'embouchure découle essentiellement des résultats d’une mission
préliminaire de chalutage, effectuée début 2000 à différentes profondeurs (0-100 m) de part et d’autre
du Grand Rhône. Les prises de poissons s’étant avérées beaucoup plus diversifiées et, pour un même
effort de pêche, environ deux fois plus importantes à l’Est qu’à l’Ouest de l’embouchure, la partie
proximale du prodelta du fleuve a été préférentiellement retenue pour cette étude.
Figure I.1 – Situation géographique de la zone retenue pour cette étude.

Zone d’étude et échantillonnage
6
I.1 MILIEU ET PEUPLEMENTS MACROBENTHIQUES1 : CARACTERISTIQUES
GENERALES
La zone d’étude a également été choisie d’après les conclusions de différents travaux
préexistants sur l’étendue du panache rhodanien, la sédimentation au large du delta du Rhône, la
production planctonique dans le golfe du Lion et la composition des communautés benthiques à
l'embouchure. L’essentiel de ces données, exposé au sein des trois sous-chapitres suivants, résume les
principales caractéristiques biotiques et abiotiques du golfe du Lion et décrit les particularités physiques
et biologiques qui font de la partie proximale du prodelta du fleuve, une des zones les plus appropriées
pour cette étude.
I.1.1 FACTEURS ABIOTIQUES
Le Rhône : apports en matière d’origine continentale
Depuis la construction du barrage d’Assouan sur le Nil, le Rhône constitue la composante
majeure, en terme de débits liquide et solide, des apports fluviatiles en Méditerranée. Son bassin
versant, d'une superficie de 97800 km2, englobe une vaste région montagneuse soumise en partie aux
influences méditerranéennes (Pont, 1997), ce qui lui confère une variété de régimes (pluvial, nival,
glaciaire, etc) exceptionnelle. Il se jette en Méditerranée nord-occidentale par l’intermédiaire d’un vaste
delta dont les deux bras principaux, le "Petit" et le "Grand" Rhône, débouchent dans le golfe du Lion de
part et d'autre de la Camargue. Le Grand Rhône, dont l'embouchure se situe à l'Ouest du golfe de Fos,
reçoit la grande majorité (90%) des apports fluviatiles (Sempéré et al., 2000).
Le débit liquide du fleuve fluctue entre 500 et 10 000 m3 s-1 autour d’une moyenne de 1715 m3
s-1 (Broche et al., 1998). Il varie à court terme selon la saison, à moyen terme selon la périodicité des
crues et à long terme selon les grandes périodes climatiques affectant l’Europe de l’Ouest (Probst, 1989
; Vivian, 1989). Les apports en matière organique dissoute ne suivent que modérément les fluctuations
du débit liquide et enrichissent de façon continue l’ensemble du golfe du Lion (Cauwet, 1996). De
l’ordre de 1,8 à 2,5 mg l-1 en moyenne, les teneurs en carbone organique dissous représentent un apport
de 9 à 13×104 t a–1. Elles varient relativement peu au cours de l’année, le flux de carbone associé ne
passant que de 2 kg C s–1, en période d’étiage, à 40 kg C s–1 au maximum pendant les crues. Les apports
du fleuve en sels nutritifs sont quantitativement moins élevés que ceux en carbone dissous. Ils
constituent néanmoins l'essentiel des apports en sels nutritifs dans le golfe du Lion (Pinazo et al., 2001),
avec des flux annuels d'environ 5,5×104 t a–1 pour l’azote et le silicium et de 0,5×104 t a-1 pour le
phosphore (Coste et Raimbault, 1993).
1 Macrobenthique : qui appartient au macrobenthos (organismes de taille supérieure à 1 mm vivant au niveau du fond).

Zone d’étude et échantillonnage
7
Le débit solide du Rhône, étroitement lié au débit liquide, varie de 3 à 14×106 t a-1 (Cauwet et
al., 1990 ; Pont, 1997), l’essentiel des apports solides étant déchargé pendant les courtes périodes de
crues (Cauwet, 1996). Les deux principaux pôles qui contribuent à la matière organique particulaire
(MOP) sont les détritus arrachés aux sols par l’érosion et les organismes et débris issus de la production
dulçaquicole. Les concentrations en carbone et en azote organiques particulaires montrent la même
tendance que la charge solide. Ainsi, la concentration en carbone organique particulaire, de 2 à 4 mg C l-1
en période d’étiage, peut atteindre pendant les crues jusqu’à plus de 65 mg C l-1, soit un flux de carbone
de plus de 600 kg s-1. Les apports du Rhône en carbone organique particulaire constituent donc
l’essentiel du pool de matière organique véhiculé par le fleuve en période de forte crue. Les fluctuations
interannuelles de débit du fleuve se répercutent principalement sur ses apports en matière organique
particulaire.
Vents, courants et étendue du panache rhodanien
Le Rhône génère à son embouchure une structure hydrologique très instable du point de vue
salinité mais aussi température (Blanc et al., 1969). En effet, le mélange entre les eaux douces
continentales et l’eau de mer ne se fait pas de façon continue. Ceci provoque au niveau des eaux de
surface un refroidissement et une dessalure non homogènes. Ainsi, au cours de l’année, on observe dans
cette zone d’importantes variations de salinité et de température liées à la fois à la situation
hydrologique (intensité des courants et débit du fleuve) et aux conditions météorologiques (vents et
précipitations). D’une façon globale, la circulation des masses d’eau dans le golfe du Lion subit
l’influence du courant cyclonique Liguro-Provençal-Catalan dont la branche Nord (courant Liguro-
Provençal), s’écoulant de Nord-Est en Sud-Ouest au large des côtes françaises, détermine le sens de
circulation général à l’échelle du golfe (Bekers et al., 1997). Cependant, les épisodes de vent, nombreux
et intenses, y constituent le moteur principal de la circulation des masses d'eau. La circulation en surface
peut ainsi être totalement distincte du flux global de circulation du courant Liguro-Provençal, et ce, tout
particulièrement à proximité des côtes (Bekers et al., 1997). Le régime des vents dominants sur
l’ensemble du golfe entraîne ainsi, dans certaines conditions, la formation, au large, de gyres de
recirculation responsables alors d’une déviation du panache rhodanien vers l’Est (Cruzado et Velasquez,
1990 ; Bekers et al., 1997). C'est souvent le cas au printemps et en été (Fig. I.2a), alors que la situation
hivernale engendre généralement une déviation du panache vers l'Ouest (Fig. I.2b). Au niveau de
l'embouchure du Rhône, la forme et l'étendue du panache sont essentiellement déterminées par la force
et la direction des vents locaux (Broche et al., 1998). Au dessus du delta, les vents de Nord/Nord-Ouest
soufflent en moyenne 54 % du temps (Ascensio et al., 1977) avec un maximum de fréquence et
d'intensité en hiver (Millot, 1990). Moins fréquents, les épisodes de vent de Sud-Est (17 % du temps) ou
d'Ouest (8 %) peuvent néanmoins être violents et durer plusieurs jours. Lorsque le vent est faible, le
panache s’élargit rapidement et forme une vaste zone dessalée de 6 km de large légèrement décalée vers

Zone d’étude et échantillonnage
8
l’Est de l’embouchure, puis est globalement dévié vers l’Ouest par le courant Liguro-Provençal. La
situation est similaire en cas de vent du Nord, bien que la déviation vers l'Ouest soit plus rapide, alors
que de forts vents d’Ouest provoquent, en l’espace de quelques heures, une importante déviation vers
l’Est du panache. D'une façon générale, le panache est initialement dévié vers l'Est la plupart du temps
(> 60 %) et s'étend au Sud-Est de l'embouchure avant d'être (ou non) dévié vers l'Ouest plus au large
(Demarcq, 1985). Il recouvre alors, en surface, l'ensemble de la zone d'étude entre 20 et 100 m de
profondeur (Broche et al., 1998).
Figure I.2 - Directions des courants de surface dans le golfe du Lion et forme et étendue du panache rhodanien :principaux cas de figure observés au printemps-été (a) et en automne-hiver (b). D'après Bekers et al.(1997),modifié.
Transport, sédimentation et zone d’influence du fleuve
Les distances de transport et la dispersion en mer des éléments apportés par le Rhône diffèrent
de beaucoup suivant la nature, dissoute ou particulaire, des composés. D'une façon générale, les
éléments sous forme dissoute sont transportés sur des distances nettement plus grandes que le matériel
particulaire qui sédimente à proximité de l'embouchure. Le carbone organique dissous suit les masses
d’eau du panache et peut être transporté au large sur des distances importantes suivant les conditions
météorologiques et le débit du fleuve (Cauwet et al., 1997). Les concentrations en phosphates et en
nitrates dissous sont maximales en face de l’embouchure et décroissent au fur et à mesure que l’on
s’éloigne vers le large (Blanc et al., 1969). Cependant, l’influence du Rhône dans les eaux de surface est
parfois sensible assez loin des côtes et favorise la production planctonique dans l’ensemble du golfe du
Lion (Cauwet et al., 1997).
Nitrate d'origine rhodanienne (échelle en g.l-1)
(a) situation fin Mars 1996 (b) situation fin Octobre 1996
Vitesse de référence16 m/s
Profondeur : 10 m
Vitesse de référence16 m/s
Profondeur : 10 m

Zone d’étude et échantillonnage
9
La majorité des apports en matière organique particulaire (MOP) du Rhône sédimente au niveau
du prodelta où le taux d’accumulation atteint environ 1,4×106 t par an (Bouloubassi et al., 1997). Le
taux moyen annuel d’accumulation est maximum (30-50 g cm-2 a-1) en face du Rhône (Durrieu de
Madron et al., 2000 ; Touzani et Giresse, 2002). Le carbone d’origine terrestre y participe pour plus de
50 % à la composition du carbone sédimentaire, la fraction d’origine marine ne représentant jamais plus
de 30 % du poids total (Bouloubassi et al., 1997). La quantité de carbone total (COT) dans les sédiments
au large du Rhône varie de 1 à 2 % du poids total (Durrieu de Madron et al., 2000). Elle est maximale
(2,0 %) à l’embouchure même (Bouloubassi et al., 1997), du fait de la présence de débris végétaux
riches en lignine. Elle diminue ensuite quand on s’éloigne vers le large, bien que les teneurs en carbone
organique soient plus élevées dans les dépôts boueux (0,6 %) que dans les sables côtiers (0,2-0,5 %).
L’accumulation de MOP rhodanienne est plus intense à l’Est de l’embouchure, dans la partie proximale
du prodelta (Durrieu de Madron et al., 2000) où le pourcentage de carbone organique particulaire (COP)
en surface (2 %), le taux d’accumulation moyen annuel (25-40 g cm-2 a-1) et les flux moyens annuels de
MOP (2,5-4,0 × 106 t a-1) et de COP (5,0-8,0 × 104 t a-1) sont maximums. Il semble y avoir, entre 30 et
50 m de profondeur sur la zone d’étude, une aire d’accumulation privilégiée de MOP terrestre. Le
pourcentage de COT est ainsi plus fort à 30 m de profondeur à l’Est du panache (1,72 %) qu’entre 20 et
100 m en face de l’embouchure (0,81 à 1,58 %) (Bouloubassi et al., 1997).
I.1.2 PRODUCTION PLANCTONIQUE
Bien que la Méditerranée soit reconnue comme une mer globalement oligotrophe, certaines de
ses zones côtières abritent des phénomènes d’upwelling non négligeables sur une échelle mondiale
(Bakun et Agostini, 2001). Le golfe du Lion, soumis au double impact de la Tramontane et du Mistral,
est caractérisé par une zone d’upwelling intense qui inclut le delta du Rhône et s’étend de Montpellier à
Toulon. Cette structure est maximale en hiver avec des vitesses de remontée des eaux dépassant 1 mètre
par jour. Mais, à cette saison, les faibles concentrations en sels nutritifs de l'eau remontant en surface et
le fort coefficient de mélange induit par les vents dans la zone sont peu favorables au développement de
blooms planctoniques (Blanc et al., 1969). L'upwelling hivernal n’aboutit donc pas à de fortes
concentrations en chlorophylle (Bakun et Agostini, 2001). La production primaire planctonique du golfe
du Lion, maximale au printemps (Blanc et Leveau, 1971), dépend essentiellement de la diffusion directe
des apports du Rhône en sels nutritifs dissous (Denis et al., 2001). Elle est beaucoup plus intense à la
côte (436 mg C m-2 j-1) qu'au large (233 mg C m-2 j-1) (Tusseau-Vuillemain, 1998), varie peu d'une
année sur l'autre, de 78 à 142 g C m-2 a-1 (Lefèvre et al., 1997), et semble relativement indépendante des
fluctuations interannuelles du débit du Fleuve.
Le panache du Rhône, du fait de ses concentrations élevées en nutriments (Moutin et al., 1998),
est une zone de forte production phytoplanctonique (Blanc et al., 1969 ; Lefèvre et al., 1997),
zooplanctonique (Gaudy et al., 1996) et bactérienne (Kirchman et al., 1989). L'eutrophisation du milieu

Zone d’étude et échantillonnage
10
permet le développement intense de quelques espèces phytoplanctoniques euryhalines qui, dans la zone
du panache, s’ajoutent aux cellules phytoplanctoniques d’eau douce apportées par le Rhône pour aboutir
à des concentrations cellulaires de plus de 460 cell. l-1 à l’embouchure (Blanc et al., 1969). La zone de
dilution du panache rhodanien présente, dans sa partie proximale, la plus forte production primaire (86 -
142 g C m-2 a-1) du golfe du Lion (Lefèvre et al., 1997). La production bactérienne y est également
importante, avec des densités de l'ordre de 1,1×109 cell. l-1 à l'intérieur du panache (Kirchman et al.,
1989). Ceci aboutit à des biomasses de zooplancton jusqu’à 10 fois supérieures à celles relevées dans le
golfe de Marseille, avec une dominance de copépodes et de cladocères et un maximum d’abondance
situé dans la zone Est de dilution, légèrement au large de l’embouchure (Blanc et al., 1969).
I.1.3 COMMUNAUTES MACROBENTHIQUES
A proximité de l’embouchure et au large du delta du Rhône, le dépôt des sédiments se fait en
fonction de leur granulométrie (Cauwet et al., 1990). Les sables sont déposés à la côte, les silts et les
argiles plus au large. Ce phénomène entraîne l'installation de deux communautés distinctes d’invertébrés
benthiques sur les fonds de 0 à 20 m (communauté des sables) et de 30 à 100 m de profondeur
(communauté des vases). Ce phénomène est plus marqué à l'Est de l'embouchure (Abou-Hamdan,
2001), où la largeur de la bande sableuse côtière est plus importante et la différence de composition des
deux communautés (sables / vases) maximale.
Les fonds sableux (0 - 20 m de profondeur)
Entre 0 et 10 m de profondeur, le sédiment est constitué par un sable fin, bien calibré mais
légèrement envasé (4 - 7 %) (Massé et al., 1977). Plus au large, le pourcentage de vase dans le sédiment
augmente pour atteindre 74 % à 20 m, profondeur qui délimite la zone d'extension des vases terrigènes
côtières. Cette tranche bathymétrique est soumise à d’importantes variations saisonnières des
caractéristiques physico-chimiques du milieu. Au dessus de 20 m, profondeur moyenne de la
thermocline méditerranéenne, les eaux de surface subissent d’importantes variations de température au
cours de l’année. Leur température ne dépasse pas 13°C en hiver, alors que le réchauffement des eaux
en été aboutit à des températures de 18 - 21°C (Blanc et Leveau, 1971). La zone, sous la double
influence de la dessalure rhodanienne (Blanc et al., 1969) et des remontées d’eaux profondes plus salées
induites par les upwellings hivernaux (Bakun et Agostini, 2001), subit également de fortes variations de
salinité. Enfin, la double action des houles et des vagues venant du large est à l'origine, dans cette
tranche bathymétrique, d'un fort hydrodynamisme responsable notamment de l’absence de macrophytes2
benthiques sur ces fonds. Etant donné la très faible contribution du microphytobenthos3 à la production
2 Macrophyte : végétal (phanérogame, algue) macroscopique de taille importante.3 Microphytobenthos : végétaux (ici, algues) de très petite taille ou microscopiques présents au niveau du fond.

Zone d’étude et échantillonnage
11
primaire dans cette zone (Bodoy et Plante-Cuny, 1980), la quasi totalité des entrées en matière
organique dans les réseaux trophiques benthiques est assurée par la sédimentation des populations
planctoniques de surface et du matériel terrigène d'origine rhodanienne (matière organique particulaire
et divers débris charriés par le fleuve). En période de crue, les débris végétaux, essentiellement des
fragments de feuilles et de branches de gymnospermes riches en lignine (Bouloubassi et al., 1997),
peuvent y former par endroits un épais tapis recouvrant le fond (Massé et al., 1977).
La biocénose des sables fins bien calibrés (S.F.B.C.), décrite par Picard (1965), occupe les
fonds sableux de la zone entre 0 et 10 mètres de profondeur (Febvre-Chevalier, 1969). Elle correspond à
un peuplement macrobenthique purement animal riche en invertébrés suspensivores et dépositivores de
surface. Les mollusques (surtout des bivalves) y dominent (Bodoy et Plante-Cuny, 1980). Viennent
ensuite, par ordre d’importance, les crustacés, suivis des polychètes. Enfin, s’y ajoutent quelques
groupes minoritaires comme les phoronidiens, les échinodermes, les poissons et les némertes. Plus au
large (10-20 m de profondeur), cette biocénose se modifie légèrement avec l'envasement du sédiment
(Gautier, 1957) : la proportion d'échinodermes augmente avec, par endroits, de très fortes densités
d'ophiures. Une étude de la macrofaune d'invertébrés benthiques des fonds sableux à l'Est de
l'embouchure (Abou-Hamdan, 2001) a montré une dominance globale des mollusques (surtout les
bivalves) qui représentent plus de 60 % de la biomasse entre 0 et 20 m. Viennent ensuite, par ordre
décroissant, les polychètes (12 %), les crustacés (9 %) et les échinodermes (3 %). Les fluctuations
temporelles de la biomasse, la densité et la composition de cette communauté macrobenthique sont
étroitement liées à celles des apports du fleuve (Massé, 2000).
Une trentaine d'espèces de poissons fréquentent les zones sableuses côtières à l’Est de
l’embouchure (Letourneur et al., 2001). Les gobiidés (surtout Pomatoschistus minutus et P.
marmoratus), présents sur ces fonds tout au long de l'année, constituent l'essentiel de la biomasse
ichtyique (3724 g ha-1) entre 0 et 20 m de profondeur. S'y ajoute, du début du printemps à la fin de l'été,
de nombreux juvéniles de poissons plats pour lesquels la bande de sable côtière sert de nurserie (Le
Direac'h-Boursier, 1990). Ceux-ci constituent alors l'essentiel de la biomasse ichtyique sur ces fonds
(Letourneur et al., 2001).
Les fonds vaseux ( > 20 m de profondeur)
A l'est de l'embouchure du Rhône, les vases terrigènes côtières s'étendent entre 20 et 100 m de
profondeur (Picard, 1971). Le sédiment est composé d'une vase fluide réduite riche en eau, avec une
fine couche oxydée à l'interface eau-sédiment (Blanc, 1974). Du fait de sa profondeur, cette tranche
bathymétrique est relativement stable du point de vue salinité, température de l’eau et hydrodynamisme
(Denis, 1999). Elle est cependant soumise à un fort taux de sédimentation et la turbidité des eaux y
empêche le développement de macrophytes benthiques (Salen-Picard, 1982).

Zone d’étude et échantillonnage
12
Le peuplement, biocénose des vases terrigènes côtières, est entièrement animal et se caractérise
par la présence de 88 % d'espèces vasicoles (Picard, 1965), essentiellement des annélides polychètes
dépositivores de surface ou de subsurface (Salen-Picard et al., in press). Les polychètes y sont
prédominants. Viennent ensuite les mollusques, les échinodermes et les crustacés (Picard, 1965). La
richesse faunistique et la densité diminuent avec la profondeur (Salen-Picard, 1982). Quatre groupes
zoologiques (polychètes, mollusques, crustacés et échinodermes) sont, en nombre d'espèces et en
nombre d'individus, particulièrement bien représentés sur l’ensemble de la tranche bathymétrique
concernée. Les polychètes prédominent partout et représentent plus de 50 % des espèces et 80 % des
individus quelle que soit la profondeur (Salen-Picard, 1982). Les mollusques, deuxième groupe après
les polychètes par l'abondance et le nombre d'espèces entre 30 et 50 m, sont remplacés après 70 m par
les crustacés et les échinodermes qui prennent la deuxième place, respectivement en terme de diversité
spécifique et d'abondance (Salen-Picard, 1982). Les fluctuations temporelles de la composition de ce
peuplement macrobenthique sont en étroite relation avec les variations hydrologiques du Rhône (Salen-
Picard et al., 1997 ; Massé, 1998), le débit du fleuve constituant le principal facteur structurant la
communauté (Salen-Picard et al., in press).
Le peuplement ichthyique des vases terrigènes au large du delta du Rhône est riche en poissons
démersaux et benthiques (Maurin, 1968). Il est dominé en abondance par les sélaciens (Scylliorhinus
caniculus, Mustellus mustellus, Raia circularis, Torpedo marmorata, etc), auxquels s'ajoutent
principalement des capelans (Gadus capelanus), des aloses (Alosa fallax) et des juvéniles de merlu et
d'anchois. On y trouve également de nombreuses autres espèces, essentiellement démersales, au sein
desquelles les poissons plats (Citharus linguatula, Buglossidium luteum, Solea solea, S. impar,
Arnoglossus laterna, A. thori, Platichthys flesus et Scophtalmus rhombus) occupent une place
prépondérante.
I.1.4 CONCLUSION SUR LA ZONE D'ETUDE
Située à l'Est de l'embouchure du Rhône, la partie proximale du prodelta est soumise de façon
importante aux apports du fleuve. Presque toujours recouverte en surface par les eaux dessalées du
panache rhodanien, cette zone possède le plus fort taux de sédimentation du golfe du Lion. Le dépôt de
matière organique particulaire d'origine continentale y est maximal, surtout sur les fonds de 30-50 m de
profondeur. La production planctonique, liée aux apports rhodaniens en matériel dissous, y est élevée.
Au niveau du fond, les macrophytes marines sont absentes et la production microphytobenthique faible.
L'essentiel des entrées en matière organique dans les réseaux trophiques benthiques est donc assuré par
la sédimentation des populations planctoniques de surface et du matériel d'origine rhodanienne. La zone
est riche du point de vue faunistique et abrite une communauté ichthyique conséquente, avec beaucoup
d'espèces démersales. Parmi elles, on compte de nombreux poissons plats pour lesquels les fonds
sableux côtiers constituent une importante zone de nurserie. L'ensemble de ces caractéristiques biotiques

Zone d’étude et échantillonnage
13
et abiotiques font de cette région du golfe du Lion une zone idéale pour étudier l'influence du Rhône sur
les communautés benthiques. Ceci explique son choix pour la réalisation du présent travail.
I.2 POISSONS : CHOIX DES ESPECES ETUDIEES
I.2.1 GENERALITES SUR LES CINQ ESPECES
Les cinq espèces retenues pour cette étude (Solea solea, Solea impar, Buglossidium luteum,
Arnoglossus laterna et Citharus linguatula) appartiennent à l'ordre Pleuronectiformes (ou "poissons
plats"). L'essentiel de leur cycle de vie, dit "démersal", se fait sur le fond, au sein des communautés
benthiques, après une courte période de vie larvaire pélagique passée dans le plancton.
Comme l'indique leur nom, les poissons plats sont caractérisés par un fort aplatissement latéral
du corps. Ils ont pour particularité la possession d’un flanc oculifère coloré, où se situent les deux yeux,
et d’un flanc aveugle généralement blanc (Quéro et Vayne, 1997). A cela près, l’organisation générale
du corps n’est pas très différente de celle des autres poissons osseux. A l’éclosion, la larve présente un
aspect normal, symétrique, avec un œil sur chacun de ses côtés. La métamorphose se produit au moment
du passage de la vie larvaire pélagique à la vie benthique qui caractérise les juvéniles et les adultes. Elle
se traduit principalement par la migration de l’un des deux yeux qui passe du flanc sur lequel le poisson
se couche à celui tourné vers la surface. Au sein des Pleuronectiformes, on observe, suivant les familles,
deux grands cas de figure traduisant des différences fondamentales au niveau des processus de la
métamorphose. Les poissons "dextres", dont font partie les Soléidés Solea solea, S. impar et
Buglossidium luteum, ont les deux yeux positionnés sur le côté droit du corps. A l'inverse, chez les
poissons "sénestres", les yeux sont situés sur le côté gauche : c'est le cas des Bothidés comme
Arnoglossus laterna et des Citharidés comme Citharus linguatula.
Au delà de ces considérations morphologiques, les différences entre les cinq poissons plats
étudiés sont nombreuses et touchent tant à leur physiologie qu’à leur répartition géographique et
bathymétrique, leur éthologie et leur mode d’alimentation. Ces différences seront exposées au sein des 5
sous-chapitres suivants afin de démontrer la particularité et l’originalité de chacune des espèces choisies
pour cette étude.

Zone d’étude et échantillonnage
14
I.2.1.1 SOLEA SOLEA (Linnaeus, 1758)
Pleuronectes solea LINNAEUS, 1758Solea vulgaris QUENSEL, 1806Solea solea JORDAN & GOSS, 1889Solea vulgaris typica CHABANAUD, 1927Solea vulgaris vulgaris TORCHIO, 1971
Noms : Fr. sole, sole commune Famille : Soleidae GB. sole, common sole, dover sole
Description :
Le corps est ovale. Les yeux sont sur le côté droit et la bouche se situe dans la partieinférieure du corps. Les deux nageoires pectorales sont bien développées. Celle de la faceoculifère porte une tache noire arrondie bien délimitée. La narine de la face aveugle estsimple, non dilatée.
Du fait de son fort intérêt économique, les travaux concernant la sole commune Solea solea
(Fig. I.3) sont nombreux et concernent l'ensemble de son aire de répartition géographique (e. g.
Horwood, 1993). L'espèce a cependant été moins étudiée en Méditerranée où les travaux concernant son
alimentation sont peu nombreux (Mendez de Elguezabal, 1978 ; Molinero et al., 1991 ; Molinero et
Flos, 1992a, 1992b).
10 cm
Figure I.3 – Adulte de Solea solea. D'après Fisher et al. (1987).

Zone d’étude et échantillonnage
15
maximales (6 individus ha-1) à proximité de l'embouchure du Rhône (Campillo et al., 1989). Cette
répartition est non seulement valable pour les adultes mais également pour les juvéniles dont les
nurseries se répartissent le long des côtes bordant le delta du fleuve. Le stock de sole du golfe du Lion
serait composé de près de 12 millions d'individus, dont 90 % de juvéniles (Campillo et al., 1989). Les
fourchettes de biomasse exploitable se situeraient entre 1500 et 2000 tonnes (Farrugio et Le Corre,
1987). L'essentiel des individus capturés en mer sont âgés de 2 ans ou plus (Benouada, 1985).
Croissance et reproduction
La croissance est forte au cours de la première année et aboutit, en Méditerranée, à une taille
moyenne de 18-20 cm à un an (Shéhata, 1984). Elle est plus rapide chez les femelles que chez les mâles.
Ils mesurent respectivement 33 et 29 cm à 5 ans, 38 et 34 cm à 10 ans et 40 et 36 cm à 15 ans. La
longévité serait de 27 ans pour les femelles et de 24 ans pour les mâles. La taille maximale atteinte est
de 70 cm. La première maturité sexuelle est acquise entre 3 et 5 ans suivant les zones (Quéro et Vayne,
1997). Dans le golfe du Lion, Shéhata (1984) signale son apparition chez les femelles à un âge de 3 ans
et une taille minimale de 30 cm. Elle serait plus précoce chez les mâles et se ferait à une taille d'environ
25-27 cm (Sorbe, 1972 ; Mendez de Elguezabal, 1978). La période de ponte varie selon les régions. Elle
se situe généralement en hiver et au début du printemps (décembre-avril), sauf en mer du Nord où elle
s'étale d’avril à juin (Fonds, 1979). Dans la région marseillaise, elle a été observée de décembre à mars
(Shéhata, 1984). Les œufs sont pélagiques et l’éclosion a lieu après 5 à 11 jours d’incubation, les larves
Distribution géographique et habitat
La sole commune se répartit
géographiquement en Atlantique Est, du Sud de
la Norvège au Sénégal (Quéro et Vayne, 1997).
On la trouve aussi en Méditerranée, dans la Mer
de Marmara et en Mer Noire (Fig. I.4). C’est une
espèce côtière qui occupe en général les fonds
meubles sableux et sablo-vaseux situés entre 0 et
130 m de profondeur. Elle est également présente
dans les milieux estuariens où ses juvéniles
pénètrent jusqu’à la limite des eaux douces.
Dans le golfe du Lion, la distribution
spatiale de l’espèce semble liée aux arrivées
d’eaux rhodaniennes. La densité de soles est
considérablement plus élevée à l'est du golfe du
Lion (Gaertner, 1999) et atteint des valeurs Figure I.4 – Répartition géographique de Solea solea.
D'après Deniel (1981), modifié.

Zone d’étude et échantillonnage
16
mesurant 3 à 3,7 mm de long (Quéro et Vayne, 1997). Celles-ci subissent la métamorphose après 3 à 4
semaines de vie pélagique à une taille de 12 à 15 mm. Les juvéniles s’adaptent alors à la vie benthique
sur des fonds de moins de 10 mètres de profondeur.
Alimentation
La sole commune est décrite comme présentant une alimentation surtout nocturne et un mode de
nutrition essentiellement basé sur l’olfaction (De Groot, 1971). Son régime alimentaire est généralement
dominé par les polychètes et les mollusques (De Groot, 1971; Darnaude, 2001) mais, en fonction de la
composition faunistique du milieu, l’espèce consomme également des quantités variables de crustacés et
d'échinodermes (Sorbe, 1972 ; Mendez de Elguezabal, 1978).

Zone d’étude et échantillonnage
17
I.2.1.2 SOLEA IMPAR Benett, 1831
Solea impar BENNETT, 1831Solea impar ALDEBERT, 1968Pegusa impar BINI, 1968Pegusa impar impar TORTONESE, 1971Pegusa nasuta ALILI, 1981
Noms : Fr. sole adriatique Famille : Soleidae GB. adriatic sole
Description :Le corps est ovale et brun, légèrement tacheté de brun plus intense. Les yeux sont sur lecôté droit et la bouche se situe dans la partie inférieure du corps. Les deux nageoirespectorales sont bien développées. La petite taille maximale de l'espèce et la formecaractéristique de la narine de sa face aveugle, fortement dilatée et plissée en forme derosette, permettent de la distinguer aisément des juvéniles de sole commune. Lacoloration assez uniforme de son corps et, surtout, le nombre de rayons de ses nageoiresdorsales et anales (D: moy. 74; A: moy. 59) la distinguent de la sole pôle, Solea(Pegusa) lascaris (D : moy. 82; A: moy. : 66), avec laquelle elle a été longtempsconfondue.
Les travaux sur Solea impar (Fig. I.5) sont rares et assez récents (Marinaro et Bouabib, 1983 ;
Marinaro et al., 1983 ; Deniel et al., 1989 ; Marinaro, 1991 ; Allam, 1995 ; Rodriguez, 1996), l'espèce
ayant été longtemps confondue avec Solea lascaris (c.f. Borsa et Quignard, 2001).
Figure I.5 – Adulte de Solea impar. D'après Fisher et al. (1987).
10 cm

Zone d’étude et échantillonnage
18
?
?
?
Croissance et reproduction
La taille maximale observée pour l'espèce est de l'ordre de 30 cm (Quéro et Vayne, 1997). Dans
le golfe du Lion, les juvéniles atteignent une taille d'environ 10 cm à 1 an (Marinaro, 1991). La
croissance de la femelle est plus rapide que celle du mâle (Fisher et al., 1987). Ils mesurent
respectivement 18 et 19 cm à 2 ans, 22 et 25 cm à 5 ans et 24 et 27 cm à 8 ans (Marinaro, 1991). Dans le
golfe du Lion, la première maturité sexuelle est acquise au cours de la deuxième année à une longueur
totale de 16-18 cm (Shéhata, 1984). La période de ponte varie selon les régions. Elle se situe
généralement au printemps et en été, avec un maximum en mai (Quéro et Vayne, 1997). Dans le golfe
du Lion, elle s'étend d'avril à octobre (Marinaro, 1991) avec un maximum en mai-juin (Shéhata, 1984).
Les œufs sont pélagiques. La métamorphose survient à environ 2 cm de longueur, taille où les juvéniles
s'installent sur le fond.
Alimentation
Solea impar se nourrit majoritairement de petits crustacés (Marinaro et Bouabib, 1983), surtout
des amphipodes et des mysidacés, et de mollusques bivalves auxquels s’ajoutent quelques annélides
polychètes et échinodermes (Shéhata, 1984 ; Rodriguez, 1996).
Distribution géographique et habitat
Solea impar se répartit géographiquement
essentiellement en Méditerranée et dans
l’Adriatique (Quéro et Vayne, 1997). Elle est
signalée également en Atlantique Est, de la manche
jusqu’en Afrique du Nord (Fig. I.6), mais sa
possible confusion avec S. lascaris rend la
localisation précise de son aire de répartition
malaisée. En général, l'espèce, très côtière, vit sur
des fonds sableux et vaseux, de 5 à 100 m de
profondeur.
Dans le golfe du lion, elle vit entre 0 et 25 m
(Marinaro, 1991). Très côtière, elle ne semble pas
dépasser vers le large la profondeur limite de 50 m.
Les jeunes passent leurs deux premières années sur
les plages sableuses bordant le littoral. Figure I.6 – Répartition géographique de Solea impar.D'après Shéhata (1984), modifié.

Zone d’étude et échantillonnage
19
I.2.1.3 BUGLOSSIDIUM LUTEUM (RISSO, 1810)
Pleuronectes luteus RISSO, 1810Rhombus luteus RISSO, 1826Solea lutea BONAPARTE, 1833Monochirus minutus PARNELL, 1837Microchirus luteus BONAPARTE, 1846Monochirus luteus COSTA, 1847Solea boscanion CHABANAUD, 1926Microchirus boscanion CHABANAUD, 1927Microchirus minutus CHABANAUD, 1927Buglossidium luteum CHABANAUD, 1930Microchiropsis boscanion CHABANAUD 1956
Noms : Fr. petite sole jaune, solenette Famille : Soleidae GB. solenette
Description :Le corps est ovale et charnu, de couleur brun-orangée. Les yeux sont sur le côté droitet la bouche se situe dans la partie inférieure du corps. La taille réduite de la nageoirepectorale gauche (face aveugle), la pigmentation noire et régulière des rayons del'anale et de la dorsale et le nanisme des adultes permettent une détermination rapidede l'espèce, fréquemment confondue avec les très jeunes soles.
Les travaux concernant B. luteum (Fig. I.7) sont peu abondants (Giovanardi et Piccinetti, 1981 ;
Nottage et Perkins, 1983 ; Tito de Morais, 1983, 1984 ; Tito de Morais et Bodiou, 1984), peut être du
fait de l'intérêt économique moindre de ce poisson plat de faible taille maximale.
Figure I.7 – Adulte de Buglossidium luteum. D'après Fisher et al. (1987)
10 cm

Zone d’étude et échantillonnage
20
Croissance et reproduction
Avec une taille adulte de 10 à 13 cm, B. luteum est le plus petit représentant des soléidés
(Nichols, 1976). La longévité de l'espèce est de 14 ans pour les femelles et de 12 ans chez les mâles
(Quéro et Vayne, 1997). Dans le golfe du Lion, les juvéniles mesurent environ 5 cm à la fin de leur
première année de vie (Tito de Morais, 1983). Les mâles atteignent leur taille maximale (11,5 cm de
longueur totale) à 5 ans et les femelles (13,5 cm) à 8 ans (Shéhata, 1984). La première maturité sexuelle
survient à une taille de 7-8 cm et un âge de 3 ans (Deniel, 1981). La période de ponte est généralement
étalée sur 4 à 8 mois. Dans notre région, elle s’étend d'avril à juin (Bourquard, 1985). Les œufs sont
pélagiques. Le développement larvaire de l’espèce s’accomplirait en 5 jours, la métamorphose ayant
lieu à une taille d'environ 8-9 mm (Shéhata, 1984). Ceci expliquerait l’installation benthique précoce
des juvéniles de B. luteum qui se produit, à Banyuls, essentiellement d’avril à mai sur les fonds sableux
de 5 à 20 m de profondeur (Tito de Morais, 1983).
Alimentation
B. luteum est décrit comme ayant une alimentation essentiellement nocturne et un mode de
capture des proies basé sur l’olfaction (De Groot, 1971). Son régime alimentaire se compose
majoritairement de polychètes auxquels s’ajoutent des bivalves et des crustacés de petite taille
(amphipodes, copépodes, cumacés, pagures, mysidacés…) (Tito de Morais, 1986 ; Darnaude et al.,
2001).
Distribution géographique et habitat
L’aire de répartition géographique de
Buglossidium luteum s’étend en Atlantique, de
la Norvège à l’Angola (Fig. I.8). L'espèce est
également commune en Méditerranée et se
retrouve jusque en mer Noire (Shéhata, 1984).
Elle vit entre 5 et 450 m de profondeur et
semble préférer les fonds de sable fin (Quéro
et Vayne, 1997).
Dans le golfe du Lion, l’espèce a une
répartition essentiellement côtière, de 5 à 100
m de profondeur, avec de fortes densités
autour de l’embouchure du Rhône (Gaertner,
1999).Figure I.8 – Répartition géographique deBuglossidium luteum. D'après Deniel (1981),modifié.

Zone d’étude et échantillonnage
21
I.2.1.4 ARNOGLOSSUS LATERNA (WALBAUM, 1792)
Pleuronectes laterna WALBAUM, 1792Pleuronectes diaphanus SHAW, 1803Solea arnoglossa RAFINESQUE, 1810Rhombus nudus CUVIER, 1817Arnoglossus laterna GUNTHER, 1862Arnoglossus laterna microstoma KILE, 1913Arnoglossus laterna conspersus BUEN, 1919Arnoglossus (Arnoglossus ) laterna CHABANAUD, 1933
Noms : Fr. arnoglosse Famille : Bothidae GB. scaldfish
Description :Petit et jaunâtre, le corps sort souvent pelé du chalut, du fait de la fragilité de la peauchez cette espèce. Les yeux sont sur le côté gauche, séparés l'un de l'autre par unedistance inférieure à leur diamètre. La bouche est terminale, dans le prolongement ducorps. Sur la face aveugle, la ligne latérale est absente et la base de la nageoirespelvienne plus courte que sur la face oculifère. La taille normale des 2 premiers rayonsde la nageoire dorsale permet de différencier rapidement l'espèce d'Arnoglossus thori,autre arnoglosse présent sur les fonds prospectés.
Les travaux sur A. laterna (Fig. I.9) sont peu abondants (Gibson et Ezzi, 1980 ; Deniel, 1983 ;
Avsar, 1994), peut-être parce que ce poisson a une importance commerciale réduite en raison de sa
faible taille maximale.
Figure I.9 – Adulte d'Arnoglossus laterna. D'après Deniel (1981).
10 cm

Zone d’étude et échantillonnage
22
Croissance et reproduction
En Méditerranée, l’espèce dépasse rarement les 15 cm mais peut atteindre une taille maximale
de 19 cm (Fisher et al., 1987). La longévité est de 8 ans pour le mâle et la femelle (Quéro et Vayne,
1997). En fin de première année de vie, les juvéniles mesurent environ 5 cm (Tito de Morais, 1983). La
première reproduction a lieu entre 1 et 2 ans (Giovanardi et Piccinetti, 1984) à une taille d'environ 6-7
cm (Gibson et Ezzi, 1980). L’espèce se reproduit au printemps et en été, de juin à août en Ecosse
(Gibson et Ezzi, 1980) et de mai à septembre avec un maximum en juin-juillet dans le golfe de
Gascogne (Deniel, 1983). En Méditerranée, la reproduction a lieu essentiellement de mars à juin
(Giovanardi et Piccinetti, 1984). Les œufs sont pélagiques et, à l'éclosion, la larve mesure 2,5 mm. La
métamorphose se fait entre 2 et 4 cm, taille où les juvéniles commencent leur vie benthique.
Alimentation
A. laterna est décrit comme étant essentiellement actif le jour (Gibson et Ezzi, 1980) et se
nourrissant par prédation visuelle (De Groot, 1971). Son régime alimentaire est principalement composé
de crustacés (mysidacés, macroures, amphipodes) et de bivalves (Darnaude et al., 2001 ; Tito de Morais
et Bodiou, 1984).
Distribution géographique et habitat
Arnoglossus laterna est présent en
Atlantique, de la Norvège jusqu’à l’Afrique (Fig.
I.10), en Méditerranée, et en mer Noire (Quéro et
Vayne, 1997). L'espèce vit entre 0 et 400 m de
profondeur sur les fonds sablo-vaseux et vaseux
du plateau continental et du bord du talus, avec
un maximum d’abondance entre 10 et 70 m (Poll,
1947 ; Wheeler, 1969).
Dans le golfe du Lion, elle est
principalement associée aux fonds de 80 à 100 m
de profondeur de la partie Ouest du golfe
(Gaertner, 1999). Les densités d’A. laterna sont
néanmoins élevées à l’embouchure du Rhône
(Darnaude et al., 2001 ; Letourneur et al., 2001). Figure I.10 – Répartition géographique d'Arnoglossuslaterna. D'après Deniel (1981), modifié.

Zone d’étude et échantillonnage
23
I.2.1.5 CITHARUS LINGUATULA (LINNAEUS, 1758)
Pleuronectes linguatula LINNAEUS, 1758Citharus macrolepidotus BLOCH, 1787Pleuronectes citharus SPINOLA, 1807Pleuronectes patarachia NARDO, 1847Citharus linguatula NIELSEN, 1986
Noms : Fr. cithare, feuille Famille : Citharidae GB. spotted flounder
10 cm
Description :Le corps est brun-jaune. Les yeux sont sur le côté gauche, l'œil supérieur situé un peu enavant de l'œil inférieur. La bouche est terminale, dans le prolongement du corps. Desdeux côtés du corps, la ligne latérale est présente et la base des nageoires pelviennes,courte, est sensiblement de la même taille, ce qui permet de distinguer les juvéniles del'espèce des individus d'A. laterna.
Peu de données sont disponibles sur C. linguatula (Fig. I.11). La plupart d'entre elles (Marinaro et
al., 1978 ; Sabatés, 1988 ; Vassilopoulou et Papaconstantinou, 1994 ; Garcia-Rodriguez et Eesteban,
2000) concernent la Méditerranée.
Distribution géographique et habitat
L’aire de répartition géographique de C. linguatula couvre l’ensemble de la Méditerranée (Fig
I.12) et s’étend en Atlantique Est, de Gibraltar jusqu’en Afrique du Sud (Quéro et Vayne, 1997).
Figure I.11 – Adulte de Citharus linguatula. D'après Fisher et al. (1987).

Zone d’étude et échantillonnage
24
L’espèce vit de préférence sur les fonds
meubles du plateau continental, depuis la
côte jusqu’à 450 m de profondeur, et est
présente dans l’ensemble du bassin
méditerranéen (Campillo et Bigot, 1992).
Dans le golfe du Lion, elle se
répartit de 70 à 130 m de profondeur mais
est exceptionnellement capturée à 20-30 m
(Campillo et Bigot, 1992). Les biomasses
estimées fluctuent de 140 à 180 tonnes.
Elles sont en moyenne 5 fois plus élevées
dans la partie Est du golfe où les densités
moyennes peuvent dépasser, au large du
Rhône, les 1,5 kg (soit 20 à 30 individus)
par hectare. Le quartier de Martigues
représente l'essentiel (> 63%) des prises
commerciales qui dépassent les 150
tonnes par an depuis 1989.
Croissance et reproduction
La taille maximale observée pour l’espèce est de 30 cm (Quéro et Vayne, 1997). La croissance,
rapide la première année (LT = 7,5 - 10 cm à un an), se ralentit ensuite (Garcia-Rodriguez et Eesteban,
2000) et diffère selon le sexe (Vassilopoulou et Papaconstantinou, 1994). En Méditerranée, mâles et
femelles mesurent respectivement 14 et 14,5 cm à 2 ans, 16 et 17 cm à 3 ans, 20 et 21 cm à 5 ans. Chez
les femelles, la première maturité sexuelle est acquise à 2 ans pour une longueur totale de 19 cm
(Garcia-Rodriguez et Eesteban, 2000). Chez les mâles, elle est plus précoce et survient dès l'âge de 1 an
à environ 15 cm de LT (Vassilopoulou et Papaconstantinou, 1994). En Méditerranée, la période de ponte
se situe d'août à novembre, avec un maximum en août-septembre (Sabatés, 1988). Les œufs sont
pélagiques. A l'éclosion, la larve mesure 3 mm (Marinaro et al., 1978). La métamorphose survient entre
8 et 9,5 mm de longueur, taille où les juvéniles s'installent sur le fond (Sabatés, 1988).
Régime alimentaire
L’espèce est décrite comme ayant une activité trophique diurne (Belghyti et al., 1995). Elle
chasse à vue, essentiellement des proies pélagiques et necto-benthiques. Son régime alimentaire est
composé principalement de crustacés (mysidacés, euphausiacés, macroures, etc) et de petits poissons.
Figure I.12 – Répartition géographique de Citharuslinguatula. D'après Deniel (1981), modifié.

Zone d’étude et échantillonnage
25
I.2.2 POURQUOI CES CINQ POISSONS PLATS ?
Le choix de cinq espèces de Pleuronectiformes pour ce travail est en partie lié à leur intérêt
commercial dans la région, les prises de poissons plats représentant plus de 10 % de la valeur
économique des débarquements à Martigues et à Sète (Campillo et Bigot, 1992). Cependant, les
Pleuronectiformes présentent également un intérêt biologique majeur. Leur cycle de vie, complexe,
comprend une courte période de vie larvaire planctonique suivie d'une longue période de vie benthique.
De ce fait, la dynamique de population de ces espèces est sous la double dépendance des variations de
production planctonique et d'abondance des proies benthiques, elles-mêmes respectivement liées aux
apports du fleuve en matériel dissous et en matière organique particulaire (Blanc et al., 1969 ; Salen-
Picard et Arlhac, 2002). Cette particularité fait des Pleuronectiformes l'un des groupes de poissons les
plus susceptibles d'être influencés par une augmentation notoire du débit du fleuve.
Les cinq espèces retenues pour cette étude sont les plus abondantes dans nos captures. Elles
représentent, groupées, plus de 95% des poissons plats vivant à l’embouchure du Rhône (Letourneur et
al., 2001). Elles sont donc globalement représentatives de l'ensemble de cette communauté ichtyique.
De plus, leur répartition géographique dans le golfe du Lion, majoritairement centrée autour de
l’embouchure du Rhône, suggère, chez ces poissons, l'existence d'une sensibilité accrue aux apports du
fleuve. Enfin, ces cinq espèces, bien que proches du point de vue systématique, présentent d'importantes
différences sur les plans biologique et éthologique (Tableau I.1). Celles-ci vont permettre d’explorer
plusieurs cas de figures et d’estimer les importances relatives de divers facteurs (répartition
bathymétrique, alimentation, période de reproduction, etc) sur la "sensibilité" potentielle des poissons
aux fluctuations du débit du fleuve.
Tableau I.1 – Principales caractéristiques biologiques et écologiques des cinq espèces de poissons plats choisiespour cette étude. P = polychètes, M = mollusques, C = crustacés, Po = poissons.
Espèces Répartition
bathymétrique
Alimentation
(proies principales)
Taille
maximale
Période de reproduction
J F M A M J J A S O N D
Solea solea 0-130 m P + M 70 cm ----------------- --
Solea impar 0-25 m C + M 30 cm ----------------------
Buglossidium luteum 5-100 m P + M + C 13 cm ----------
Arnoglossus laterna 10-100 m C + M 15 cm --------------
Citharus linguatula 30-130 m C + Po 30 cm --------------

Zone d’étude et échantillonnage
26
I.3 ECHANTILLONNAGE
Entre mars 2000 et novembre 2001, 19 missions à la mer ont été réalisées afin de récolter les
différents poissons, invertébrés benthiques et échantillons d'eau et de sédiment et nécessaires pour cette
étude (Tableau I.2).
Tableau I.2 – Récapitulatif des principaux types de missions réalisées entre mars 2000 et novembre 2001. "*" =la mission SOLMAR 4 (novembre 2000) a été interrompue à cause d'une importante quantité de filets calés par lespêcheurs sur la zone d'échantillonnage. Non complets, les quelques échantillons récoltés au cours de cettemission ont finalement été écartés des résultats.
Missions : nom et n° Navire Durée Saisons Type d'échantillons récoltés
SOLMAR 1 à 7N.O. Pr. Georges
Petit4-6 jours
Hiver
Printemps
Eté
Automne*
Poissons, invertébrés benthiques,
eau de surface, sédiment
(10-100 m de profondeur)
NURSERIES 1 à 5 N.O. Antédon 1 jourPrintemps
Eté
Juvéniles de poisson
(5-20 m de profondeur)
EAU-SED 1 N.O. Antédon 1 jour HiverEau de surface, sédiment
(10-100 m de profondeur)
PROIES 1 et 2 Zodiac 1 jourPrintemps
Ete
Invertébrés benthiques
(2-5 m de profondeur)
SENNES 1 à 4 1 jourPrintemps
Ete
Juvéniles de poisson
(0-2 m de profondeur)
Compte-tenu des variations de nature du substrat observées avec la profondeur à l'embouchure
du fleuve, la zone d'étude a été divisée en trois tranches bathymétriques distinctes. Les fonds de 0 à 20
m de profondeur, d'une part, et ceux situés en dessous de 30 m, de l'autre, ont été échantillonnés
séparément afin de reproduire les conditions existant respectivement sur les fonds sableux côtiers et au
niveau des vases terrigènes plus profondes. L'existence, au sein de ces dernières, d'une zone de
sédimentation particulière aux environs de 30 m de fond, a justifié leur découpage en deux tranches
bathymétriques. Au final, l'échantillonnage a donc été réalisé de façon systématique suivant 3 tranches
de profondeur : 0-20, 30-50 et 70-100 mètres.

Zone d’étude et échantillonnage
27
I.3.1 STRATEGIE D'ECHANTILLONNAGE
La mise au point d'une stratégie d'échantillonnage globale et la planification des différentes
missions de prélèvements se sont avérées délicates, compte tenu de la diversité des objectifs visés, de la
forte variabilité spatio-temporelle de la zone d'étude et de la variété des types de prélèvements à
effectuer. Du point de vue de l'expérimentation, l'étude menée peut être scindée en trois parties :
1. l'étude de l'alimentation des cinq poissons par l'analyse de leurs contenus stomacaux,
2. la détermination de l'influence potentielle du Rhône sur ces cinq espèces, par le biais de leurs
signatures isotopiques en carbone et en azote,
3. la reconstitution du cheminement éventuel de la MOP terrestre le long de leurs réseaux
trophiques, également à l'aide de l'outil "isotopes stables".
Les types d'échantillons à récolter, l'effectif minimum nécessaire et la fréquence des
prélèvements à réaliser diffèrent d'une partie à l'autre et seront exposées au sein des trois sous chapitres
suivants.
Alimentation des cinq poissons
L'échantillonnage lié à l'étude de l'alimentation des poissons doit prendre en compte un certain
nombre de paramètres. L'étude des régimes alimentaires par analyse des contenus stomacaux nécessite
l'ouverture d'un nombre conséquent d'estomacs afin de garantir une bonne représentativité de
l'alimentation réelle du groupe étudié au travers de l'échantillon. L'effectif minimum nécessaire varie
suivant que l'on compare les alimentations soit d'espèces différentes, soit de groupes d'individus au sein
d'une même espèce : de 30 estomacs pleins par espèce dans le premier cas, il descend à un minimum de
10 estomacs pleins par groupe dans le second (Randall et Myers, 2001). Dans la présente étude, ces
valeurs minimales ont été respectées et l'échantillonnage a été réalisé en conséquence. Chez beaucoup
de poissons plats, le régime alimentaire est variable et dépend essentiellement de l'abondance des proies
disponibles dans le milieu (Tito de Morais, 1984 ; Darnaude et al., 2001). Les différences
bathymétriques de composition du macrobenthos et les fluctuations saisonnières des communautés
d'invertébrés benthiques à l'embouchure (Massé, 1972a ; Salen-Picard, 1982) sont ainsi susceptibles de
se répercuter sur la composition du régime alimentaire des cinq espèces. Afin de prendre en compte ce
paramètre, la récolte des poissons a été réalisé séparément à 0-20 m, 30-50 m et 70-100 m de
profondeur, avec des missions d'échantillonnage initialement prévues à chaque saison (hiver, printemps,
été, automne). Divers problèmes techniques et de mauvaises conditions météorologiques ayant annulé
les missions programmées à l’automne en 2000 et 2001, les poissons n'ont pas été récoltés à cette
saison. L'échantillonnage couvre néanmoins les deux grandes périodes climatiques de l'année (saison
froide / saison chaude) et les premiers mois d'installation benthique des juvéniles avec des prélèvements
de début mars à fin septembre. Chez la plupart des poissons, on observe des modifications
ontogénétiques de l'alimentation pouvant entraîner d’importantes différence de régime alimentaire entre

Zone d’étude et échantillonnage
28
juvéniles et adultes d’une même espèce (Harmelin-Vivien et al., 1989). Afin d’obtenir, pour chaque
espèce, un échantillon d’individus représentatif de l'ensemble des classes d'âge de la population,
juvéniles et adultes ont été scindés en classes de taille de 2 cm pour S. solea, S. impar et C. linguatula
(LT max. ≥ 20 cm) et de 1cm pour A. laterna et B. luteum (LT max. ≤ 15 cm). Dans la mesure du
possible, un minimum de 10 individus a été récolté par classe de taille et pour chacune des saisons et
des tranches bathymétriques échantillonnées.
Signature isotopique des poissons (influence du Rhône sur les 5 espèces)
Du fait des différences de sédimentation et de composition du macrobenthos observées avec la
profondeur, le transfert de matière organique terrigène est susceptible de varier d'une tranche
bathymétrique à l'autre, influant à terme sur la signature isotopique de la chair des poissons. Les
variations ontogénétiques de l'alimentation résultent également, chez certains poissons, en une
différence de signature isotopique entre juvéniles et adultes (Pinnegar et Polunin, 2000) liée à une
modification des sources de matière organique utilisées pour la croissance. Pour ces raisons, la collecte
des poissons destinés aux analyses isotopiques nécessite, dans l'absolu, de couvrir l'ensemble des classes
de taille de la population et d'englober, à chacune des profondeurs étudiées, non seulement les adultes
mais aussi les juvéniles des cinq espèces. Cependant, le coût élevé des analyses a fortement limité les
effectifs destinés à cette partie de l'étude. Un compromis entre représentativité statistique de
l'échantillon et limitations budgétaires a conduit à la récolte d'un maximum de 20 individus par stade
(adulte / juvénile) avec, dans la mesure du possible, 6 individus minimum par tranche bathymétrique et
une gamme de tailles représentative de l'ensemble des effectifs récoltés à chaque profondeur. Du fait de
son fort intérêt économique, la sole commune, Solea solea, a fait l'objet d'une étude plus détaillée. Pour
cette espèce, les juvéniles ont été scindés en deux groupes, les G0 (juvéniles de l'année) et les G1+
(autres juvéniles), et, dans la mesure du possible, un minimum de 10 individus de chaque stade de vie
(juvéniles G0, juvéniles G1+ et adultes) a été récolté par profondeur. Chez les poissons, le délai assez
long observé entre un changement net d'alimentation et la modification de signature isotopique qui en
résulte (Hesslein et al., 1993) suggère une absence de variation des signatures isotopiques à l'échelle
saisonnière. Cependant, la longueur du délai dépend essentiellement de la vitesse de croissance du
poisson considéré. Afin de réduire les éventuelles différences de signature isotopique liées à ce facteur,
les individus analysés au sein de chaque groupe ont été sélectionnés, pour chacune des cinq espèces, de
façon à couvrir l'ensemble des saisons échantillonnées.
Réseaux trophiques et cheminement de la MOP terrestre
Pour l'étude des réseaux trophiques, l'échantillonnage a nécessité la prise en compte des
variations spatio-temporelles de la composition des communautés benthiques et des apports des deux
principales sources de matière organique (MOP rhodanienne et phytoplancton marin) à la base de la

Zone d’étude et échantillonnage
29
chaîne alimentaire au large de l'embouchure. Les différences de sédimentation du matériel rhodanien en
fonction de la profondeur ont des conséquences majeures sur la nature des sédiments et la composition
des peuplements d'invertébrés benthiques au large du delta du fleuve. Afin de tenir compte de la
variabilité spatiale du milieu et de récolter l'ensemble des proies ingérées par les cinq poissons, les
différents échelons des réseaux trophiques benthiques ont été échantillonnés séparément à 0-20 m, 30-
50 m et 70-100 m de profondeur. La matière organique particulaire en provenance du Rhône varie
quantitativement en fonction du débit du fleuve. Il est également probable qu'en périodes de crues elle
varie qualitativement en fonction de la typologie de ces dernières, c'est-à-dire selon la partie du bassin
versant dont elles proviennent (Pont, 1997). Il en est de même pour le phytoplancton marin dont
l'abondance et la composition diffèrent suivant la saison (Blanc et Leveau, 1971). Afin de tenir compte
de la variabilité des apports des deux sources, terrestre et marine, l'échantillonnage a été prévu à chaque
saison de l'année. Des prélèvements d'eau de surface dans le Rhône, en pleine mer et à l'embouchure ont
ainsi été programmés à intervalles réguliers, en période d'étiage et au cours des fortes crues du fleuve.
Les missions prévues en automne 2000 et 2001 ayant été annulées, aucun échantillonnage n'a été réalisé
d'octobre à décembre. L'échantillonnage couvre néanmoins l'ensemble de l'année, avec des
prélèvements trimestriels réguliers en mars, juin, septembre et décembre. Il englobe les deux saisons les
plus contrastées en terme de débit du fleuve : la période printemps-hiver (fortes crues liées aux
précipitations hivernales et à la fonte des neiges) et l'été (période d'étiage). Compte tenu du coût des
analyses isotopiques, la stratégie d'échantillonnage a été limitée, pour chaque saison, à l'obtention de 3
échantillons par catégorie (sources potentielles de matière organique et proies benthiques récoltées) et
par profondeur.
I.3.2 CAPTURE DES POISSONS
La récolte des juvéniles et des adultes des cinq espèces au large de l'embouchure (Planche I.1,
Fig. I.13) a été réalisée au cours de 15 missions en mer effectuées entre mars 2000 et septembre 2001
(Tableau I.3). La majorité des poissons a été capturée au cours des missions SOLMAR réalisées à bord
du NO Pr. Georges Petit en 2000 et 2001 (mars, juin, septembre). Ces missions ont consisté en des
cycles de chalutages réalisés à différentes profondeurs (10, 30 et 90 m) au large de l'embouchure. En
raison du rythmes d’alimentation des cinq poissons, diurne ou nocturne suivant les espèces,
l'échantillonnage a été effectué suivant des cycles de 24 heures. Toutes les 1 à 2 heures, des traits de
chalut, d'une durée effective de 30 minutes chacun, ont été réalisés à une vitesse constante de 2 nœuds.
La poche du chalut à panneaux expérimental utilisé (Fig. I.14) mesure 34 m de long et possède une
ouverture en pêche de 4 mètres par 3, lestée dans sa partie inférieure par une lourde chaîne qui racle le
fond et déloge les poissons enfouis dans le sédiment. Le vide de ses mailles, de 100 mm à l'ouverture
(ailes et dos du chalut), se réduit progressivement au niveau du corps du filet (60 puis 40 mm) pour
atteindre 20 mm en cul de chalut. Ce maillage relativement lâche a permis d'éviter, dans la majeure

Zone d’étude et échantillonnage
30
10 m
30 m
90 m
20 m
70 m
50 m
100 m
golfede Fos
Figure I.13 – Localisation (en rouge) des prélèvements depoissons effectués à l'embouchure en 2000-2001.
= traits de senne de plage ; chalutages.
Figure I.18 – Senne de plage utilisée.Schéma du constructeur
Figure I.19 – Juvéniles de soles récoltés à la senne deplage sur 0-2 m de profondeur (photo : M. Harmelin)
PLANCHE I.1 –RECOLTE DES POISSONS
CAPTURE DES JUVENILES :
MISSIONS "SENNE"
Figure I.17 – Juvéniles récoltés au petitgangui sur 5-10 m de profondeur.
CAPTUREDES
JUVENILES : MISSIONS
"NURSERIES"
Figure I.16 – Schéma dupetit gangui utilisé.
CAPTURE DES JUVENILES ETDES ADULTES : MISSIONS "SOLMAR"
Figure I.15 – pêche réalisée sur 30m de fond au chalut à panneaux.
Figure I.14 – Schéma du chalut àpanneaux utilisé.
Cul duchalut
Ailes
poche
bras

Zone d’étude et échantillonnage
31
partie des cas, le colmatage de la poche par le sédiment sur les fonds de 30-50 m, particulièrement
envasés (Fig. I.15). Les dimensions de ce chalut réduisent l’échappement des poissons. Cependant, les
adultes de soles, de plus de 26 cm de LT, se sont avérés peu vulnérables vis à vis de cet engin.
Tableau I.3 – Récapitulatif des principales missions d'échantillonnage des poissons réalisées. Engins utilisés :CP = chalut à panneaux, G = gangui, S = senne de plage.
Missions(N°) Nom Date Engin Saison Profondeur
Captures :stade(s) de vie récolté(s)
(1) SOLMAR 1 11-12/03/2000 CP + G hiver 10-100 m Juvéniles et adultes
(2) NURSERIES 1 11/05/2000 G printemps 5-10 m Juvéniles
(3) NURSERIES 2 15/05/2000 G printemps 5-10 m Juvéniles
(4) SENNES 1 17/05/2000 S printemps 0-2 m Juvéniles de l'année
(5) NURSERIES 3 29/05/2000 G printemps 5-10 m Juvéniles
(6) SOLMAR 2 6-7/06/2000 CP + G printemps 10-100 m Juvéniles et adultes
(7) SENNES 2 15/06/2000 S printemps 0-2 m Juvéniles de l'année
(8) NURSERIES 4 10/07/2000 G été 10-15 m Juvéniles
(9) SOLMAR 3 15-16/09/2000 CP + G été 10-100 m Juvéniles et adultes
(12) SOLMAR 5 25-26/03/2001 CP + G hiver 10-100 m Juvéniles et adultes
(13) SENNES 3 02/04/2001 S printemps 0-2 m Juvéniles de l'année
(14) SENNES 4 27/04/2001 S printemps 0-2 m Juvéniles de l'année
(15) NURSERIES 5 11/05/2001 G printemps 10-15 m Juvéniles
(16) SOLMAR 6 24-25/05/2001 CP + G printemps 10-100 m Juvéniles et adultes
(19) SOLMAR 7 18/09/2001 CP + G été 10-100 m Juvéniles et adultes
Capture des juvéniles de l'année et des adultes
Le maillage (20 mm) du cul du chalut à panneaux utilisé à bord du NO Pr. Georges Petit
(missions SOLMAR) ne permet pas la capture des plus petits individus, d'ailleurs souvent localisés sur
les fonds sableux de moins de 10 m inaccessibles avec ce navire de fort tirant d'eau. La récolte des
juvéniles de l’année (G0) a donc fait l’objet de missions journalières d'appoint, effectuées à bord de
l'Antédon au printemps et à l’été. Les traits de chalut, de 30 minutes chacun (v = 2 nœuds), ont été
réalisés de jour sur les fonds sableux de 5 à 15 m de profondeur. Le petit chalut à perche expérimental
utilisé (Fig. I.16), décrit par Harmelin-Vivien (1981), est caractérisé par une ouverture de poche de 0,5
m sur 1,5 m et une longueur de filet de 5 m. Le Nylon tressé formant la poche présente un vide de
maille de 10 mm sur une longueur de 3 mètres puis de 8 mm sur les 2 mètres formant le cul du chalut

Zone d’étude et échantillonnage
32
d’environ 0,6 mètres de largeur. La petite taille, la grande maniabilité (Harmelin-Vivien, 1981) et la
faible sélectivité de ce chalut ont permis de réaliser, sans trop perturber les zones côtières, un bon
échantillonnage des juvéniles de poissons par la capture d’individus mesurant jusqu'à moins de 40 mm
de LT (Fig. I.17). Afin de récolter les plus petits juvéniles de certaines espèces qui séjournent à très
faible profondeur (< 4 m), ces missions ont été complétées par 4 séries journalières de senne de plage
réalisées au printemps et en début d'été sur les plages bordant le They de la Gracieuse (0-2 m de fond).
L'échantillonnage a consisté en une série de 15 à 25 traits de 5-10 minutes chacun réalisés de jour
depuis 1,5 m de fond en remontant perpendiculairement à la côte jusqu'au rivage. La senne de plage
utilisée (Fig. I.18) mesure 10 m de large par 1,5 m de haut. Lestée en bas par une corde plombée et
maintenue à la surface en haut par une succession de flotteurs, elle présente l'avantage, du fait de ses
petites dimensions, d'être manœuvrable par seulement 4 personnes. La taille réduite de sa maille (8 mm)
permet la capture des plus petits juvéniles (Fig. I.19). Elle a également permis la récolte de petits
crustacés nectobenthiques (macroures, mysidacés, amphipodes) et d'une grande quantité de débris de
végétaux terrestres, conservés, une fois triés, pour analyse ultérieure de leur composition isotopique. Les
soles de grande taille (LT ≥ 250 mm) s'étant avérées peu vulnérables aux engins de pêches utilisés au
cours des missions scientifiques, leur récolte a fait l'objet d'une collaboration avec la Coordination de
Pêcheurs de l'Etang de Berre et de la Région. Cet accord, impliquant trois des pêcheurs professionnels
travaillant à l'embouchure du fleuve, a permis, de décembre 2000 à mars 2001, l'achat de 30 soles
adultes (285 mm ≤ LT ≤ 365 mm) capturées au filet trémail sur la zone d'étude. Immédiatement congelés
dès leur mise à bord, ces individus ont permis de compléter les effectifs de Solea solea et d'obtenir le
nombre d'adultes de l'espèce nécessaire à l'étude de leur alimentation.
Conditionnement des échantillons
Les poissons récoltés ont été déterminés à bord, triés par espèce et comptabilisés pour chaque
trait de chalut. Ils ont ensuite été pesés (poids : P en grammes) et leurs longueurs totale (LT) et standard
(LS) ont été mesurées au mm près. La majorité d'entre eux a été immédiatement plongée dans du formol
(Formaldéhyde 10 %) afin d'arrêter la digestion et conservée, groupée dans des bidons par trait de chalut
ou de senne de plage, pour ensuite servir aux études alimentaires. A chaque saison et pour chaque
tranche bathymétrique échantillonnée, un maximum de 10 individus par classe de taille de 20 mm a été
traité de façon individuelle et conditionné séparément après dissection. Sur chacun de ces poissons, un
échantillon de muscle blanc natatoire dorsal a été prélevé pour les analyses isotopiques. Les échantillons
ainsi obtenus ont été congelé à -30°C et conservés dans des cryotubes individualisés et hermétiquement
scellés. Le reste du corps a été stocké dans le formol pour compléter l'effectif des analyses alimentaires.

Zone d’étude et échantillonnage
33
ECHANTILLONNAGE POUR L'ETUDE DES RESEAUX TROPHIQUES
I.3.2.1 Récolte d'eau et de sédiment
Déterminer les importances respectives de la MOP rhodanienne et du phytoplancton marin dans
la composition des masses d'eau de surface et des sédiments à l'embouchure du Rhône constitue la
première étape de l'étude de l'influence du fleuve sur les communautés benthiques. Pour ce faire, des
échantillons d'eau de surface et de sédiment superficiel ont été récoltés entre mai 2000 et septembre
2001 à différentes stations (Planche I.2, Fig. I.20). Afin de tenir compte de la variabilité temporelle des
apports des deux principales sources de matière organique, MOP terrestre et phytoplancton marin,
l'échantillonnage a été réalisé à chaque saison de l'année, en période d'étiage et au cours des différentes
crues (débit > 3000 m3 s-1) du fleuve (Tableau I.4).
Tableau I.4 – Missions d'échantillonnage de l'eau de surface (dans le Rhône, en mer et à l'embouchure) et dusédiment supérieur (à l'embouchure). Les crues du fleuve sont surlignées en gris.
Missions(N°) Nom Date Saison Débit du fleuve Type de prélèvement
(2) NURSERIES 1 09 / 05 / 2000 printemps 1983 m3 s-1 Eau + Sédiment
(6) SOLMAR 2 08 / 06 / 2000 printemps 1281 m3 s-1 Sédiment
(11) EAU-SED 1 04 / 12 / 2000 hiver 3474 m3 s-1 Eau + Sédiment
(12) SOLMAR 5 24 / 03 / 2001 hiver 6646 m3 s-1 Eau + Sédiment
(16) SOLMAR 6 20 / 05 / 2001 printemps 3131 m3 s-1 Eau + Sédiment
(19) SOLMAR 7 18 / 09 / 2001 été 983 m3 s-1 Eau + Sédiment
Echantillonnage de l'eau de surface
A chaque mission, la température et la salinité ont été mesurées et 20 litres d'eau prélevés à une
profondeur de 1,5 m sous la surface, ceci :
- dans le cours du fleuve, à l'entrée de Port-Saint-Louis du Rhône (6 km en amont de l’embouchure),
- dans un environnement strictement marin, au large de l'île du Frioul (43° 15' 504 N / 05° 18' 080 E),
- et dans la zone d'étude, au dessus des fonds de 10, 30 et 90 m de profondeur.
L'eau récoltée à chaque point de prélèvement a été filtrée sur un tamis de 250 µm afin d'en
retirer les organismes zooplanctoniques et divers débris végétaux (surtout présents dans l'eau du Rhône).
Le filtrat obtenu, régulièrement agité pour maintenir son homogénéité, a ensuite été récolté sur des
filtres GF/F préalablement calcinés (4 heures au four à 500°C) et pré-pesés au dixième de mg près.
Cette deuxième filtration a été réalisée à l'aide d'une pompe à vide et a permis l'obtention, au minimum,
de 4 filtres colmatés par point de prélèvement et par mission. Dans certains cas, la forte teneur de l'eau
en seston ayant provoqué un colmatage rapide des filtres, seulement 10 litres d'eau ont été utilisés.

Zone d’étude et échantillonnage
34
Figure I.20 - Localisation des prélèvements d'eau de surface et de sédimentsuperficiel réalisés en 2000-2001. Les prélèvements de sédiment n'ont étéréalisés que sur R1.
Figure I.22 – Carottier-benne Fluchautilisé : mise à bord après récolte dusédiment (photo : C. Salen).
Figure I.23 – Remontée de la drague spatangueutilisée. Les premiers cm superficiels du sédimentont été écrémés grâce à la fente située sur la partiemétallique puis piégés dans la poche qui prolongel'engin.
PLANCHE I.2 –RECOLTE D'EAU ET DE SEDIMENT
Figure I.21 – Carottier multitube utilisé : mise à bord aprèsprélèvement sur le fond de 4 carottes de sédiment.
golfede Fos
Marseille
Iles duFrioul
Etang deBerre
10 m
30 m
90 m
R1
Rhône

Zone d’étude et échantillonnage
35
Les filtres GF/F colmatés obtenus ont été congelés, lyophilisés (24 heures), et pesés afin de
déterminer la charge en seston à chaque point de prélèvement en fonction de la saison. Ils ont ensuite été
conservés à l'abri de l'humidité dans un dessiccateur en attendant d'être préparés en vue de l'analyse
isotopique du matériel les recouvrant.
Echantillonnage du sédiment
Suivant la profondeur, le type de fond et le navire océanographique disponible, le sédiment
superficiel a été récolté à l'aide de différents engins de prélèvement (Planche I.2). Les fonds vaseux,
compacts, ont été, autant que possible, échantillonnés à l'aide d'un carottier multitube (Fig. I.21) ou d'un
carottier-benne Flucha (Fig. I.22) qui garantissent une interface eau/sédiment correcte et la récolte
précise des premiers centimètres de sédiment superficiel. Dans ce cas, seuls les 5 premiers centimètres à
la surface du sédiment ont été prélevés. Pour les fonds sableux, moins compacts, le sédiment a été
collecté à la drague spatangue (Fig. I.23), seule efficace sur ce type de substrat bien que moins précise
que la benne ou le carottier. Ce type d'engin permet un écrémage du fond sur les 2- 4 premiers
centimètres à l'interface eau/sédiment. Il garantit ainsi la récolte de la partie superficielle du sédiment
mais sans réelle précision sur la hauteur échantillonnée. La benne et le carottier multitube n'étant pas
utilisables à bord de l'Antédon, l'ensemble des prélèvements de sédiment effectués avec ce navire ont
également été réalisés à la spatangue. Dans tous les cas, la faune et la flore visibles à l'œil nu ont été
soigneusement retirées du sédiment collecté. Les débris de macrophytes marines (rares fragments de
Chlorophycées uniquement présents dans les prélèvements de 0-20 m de profondeur) et/ou de végétaux
terrestres (essentiellement des fragments de bois ou débris de feuilles de chêne, hêtre et peuplier) ainsi
obtenus ont été rincés à l'eau distillée et conservés congelés séparément pour analyse de leur
composition isotopique. Pour ce qui est du sédiment, chaque échantillon obtenu a ensuite été
homogénéisé, congelé, lyophilisé et conservé à l'abri de l'humidité en attendant sa préparation en vue
des analyses isotopiques.
I.3.2.2 Récolte des proies benthiques
Afin de reconstituer le cheminement du carbone d'origine autochtone (phytoplancton marin) et
allochtone (MOP rhodanienne) au sein des réseaux trophiques aboutissant aux cinq poissons étudiés, le
macrobenthos a été collecté à différentes profondeurs (Planche I.3, Fig. I.24), tout au long de l'année
(Tableau I.5). La majorité des prélèvements a été effectuée à la spatangue (Fig. I.25). Par son écrémage
des fonds échantillonnés, ce type d'engin permet la récolte sélective de la faune benthique sur une
épaisseur de 2-4 cm autour de la surface du sédiment. Il est par contre peu efficace pour la capture des
crustacés nectobenthiques. L'échantillonnage de ces proies a été complété par plusieurs prélèvements
réalisés à la suceuse pneumatique entre 0 et 20 m de profondeur (missions PROIES 1 et 2) et, plus au large,
par des traits de drague épibenthique (Fig. I.26) effectués à 30 et 90 m de fond (SOLMAR 7).

Zone d’étude et échantillonnage
36
PLANCHE I.3 –RECOLTE DES PROIES BENTHIQUES
10 m
30 m
90 m
20 m
70 m
50 m
100 m
golfede Fos
Figure I.24 – Localisation des prélèvements defaune benthique réalisés à l'embouchure de juin2000 à septembre 2001. Figure I.27 – Utilisation de la suceuse pneumatique
en plongée pour la récolte des proies benthiqueset des crustacés nectobenthiques sur 0-20 m defond (photo R. Graille).
Figure I.26 – Drague Waren utilisée pour la récolte descrustacés nectobenthiques sur 30-100 m de fond (photoC. Marschal).
Figure I.25 – Spatangue utilisée pour la récolte desproies benthiques sur 10-100 m de fond.

Zone d’étude et échantillonnage
37
La suceuse à benthos utilisée (Fig. I.27) est manipulée sous l'eau en plongée (Massé, 1970). Elle
fonctionne par injection d'air sous pression (délivré par une bouteille de plongée) dans un tuyau
galvanisé de 15 cm de diamètre et de 1,5 m de long qui constitue son tube d'aspiration. Un grand sac de
récolte est fixé dans le prolongement du tuyau pendant la durée du prélèvement, puis fermé et remonté à
bord une fois plein. La taille de ses mailles (800 µm) assure à la fois la récolte de la macrofaune et sa
séparation du sédiment qui s'échappe du filet. L'utilisation de cet engin assure une capture efficace des
petits crustacés des fonds sableux (amphipodes, cumacés, etc) (Massé et al., 1977). Sur les fonds de plus
de 30 m de profondeur, l'utilisation de la drague épibenthique a permis la capture des mysidacés et des
macroures jusque là quasiment absents des captures des vases.
Tableau I.5 – Récapitulatif des principales missions d'échantillonnage des communautés macrobenthiques.
Missions(N°) Nom Date Saison Profondeur Engin de prélèvement
(6) SOLMAR 2 02/ 06 / 2000 printemps 10-100 m Spatangue
(11) EAU-SED 1 04 / 12 / 2000 hiver 10-100 m Spatangue
(12) SOLMAR 5 24 / 03 / 2001 hiver 10-100 m Spatangue
(15) PROIES 1 09 / 05 /2001 printemps 2-4 m Suceuse pneumatique
(16) SOLMAR 6 27 / 05 / 2001 printemps 10-100 m Spatangue
(18) PROIES 2 06/ 07 / 2001 été 2-4 m Suceuse pneumatique
(19) SOLMAR 7 18 / 09 / 2001 été 10-100 m Spatangue
Conditionnement des échantillons
Pour chaque profondeur, les invertébrés benthiques ont été récoltés par tamisage du sédiment
sur des tamis successifs de 5 mm, 1 mm, 800 µm et 300 µm. Ils ont ensuite été conservés vivants
pendant 24 heures dans de l'eau de mer filtrée, afin de vider leur tube digestif du matériel ingéré avant
leur capture. Enfin, ils ont été triés sous la loupe binoculaire, rincés à l'eau distillée et congelés, groupés
par taxon pour la plupart (amphipodes, mysidacés, bivalves, gastéropodes, etc). Pour les polychètes, les
individus récoltés ont été triés en fonction de leur éthologie alimentaire et congelés séparés en 4
catégories différentes : carnivores, suspensivores, dépositivores de surface et dépositivores de sub-
surface. L'échantillonnage des réseaux trophiques a également été complété par une collecte de
zooplancton, réalisée à différents points au large de l'embouchure (mission NURSERIES 1, mai 2000) à
l'aide d'un filet à plancton de maille de 250 µm tiré verticalement depuis la thermocline (-20 m) jusqu'en
surface. Débarrassés de toute trace de débris végétaux et soigneusement rincés à l'eau distillée, les
organismes zooplanctoniques récoltés ont été congelés puis lyophilisés groupés, avant d'être conservés
au sec, en vue de leur préparation pour les analyses isotopiques.

Zone d’étude et échantillonnage
38
I.3.3 CONCLUSION SUR L'ECHANTILLONNAGE REALISE
Le tableau I.6 dresse la liste des différents types de prélèvements effectués au cours des 19
campagnes d’échantillonnage et indique leurs utilisations respectives. Les missions initialement prévues
en automne 2000 et 2001 et en été 2001 (septembre) n'ont pu être réalisées. L'échantillonnage est donc
moins important en été et inexistant en automne. Les 19 missions réalisées ont néanmoins permis la
récolte de plus de 3500 poissons plats dont 484 Solea solea, 138 Solea impar, 437 Buglossidium luteum,
2089 Arnoglossus laterna et 300 Citharus linguatula. Elles ont également permis, à intervalles réguliers
la collecte d'échantillons d'eau et de sédiment à chacune des stations choisies et l'obtention, à chaque
profondeur, d'invertébrés benthiques appartenant à 19 catégories de proies représentatives de l'ensemble
des régimes alimentaires des cinq espèces. Effectué au cours des deux principales périodes climatiques
de l'année (saison froide/saison chaude), l'échantillonnage est représentatif des variations intra annuelles
des conditions à l'embouchure. Le découpage de la zone d'étude en tranches bathymétriques
correspondant aux différents types de milieux présents à l'embouchure et leur échantillonnage de façon
distincte garantissent une bonne représentativité de l'ensemble des écosystèmes présents au large du
Rhône. Le traitement des échantillons récoltés à chaque saison et au sein de chaque tranche de
profondeur étudiée devrait ainsi fournir une vision réaliste de la situation à l'embouchure.
Tableau I.6 – Utilisation et nombre des prélèvements de chaque type (eau de surface, sédiment, macrobenthos,poissons) effectués à chaque saison et à chacune des différentes stations échantillonnées.
Hiver Printemps Eté Total Utilisation
EAU DE SURFACE Frioul (phytoplancton marin) Rhône (MOP terrestre) Embouchure 0-20 m 30-50 m
70-100 m Total
22222
22222
11111
33333
15
Analysesisotopiques
SEDIMENT SUPERFICIEL Embouchure 0-20 m 30-50 m
70-100 m Total
222
233
111
566
17
Analysesisotopiques
PROIES BENTHIQUES Embouchure 0-2 m 5-20 m 30-50 m
70-100 m Total
0222
1222
1111
2555
17
Analysesisotopiques
POISSONS Embouchure 0-2 m 5-20 m 30-50 m
70-100 m Total
01186
81281117
02066
81592529
194
Alimentationet
analysesisotopiques

PARTIE II
REPARTITION BATHYMETRIQUE,
ALIMENTATION ET INTERACTIONS
SPATIO-TEMPORELLES DES POISSONS
PLATS AU LARGE DU RHONE


PARTIE II – CHAPITRE I
REPARTITION BATHYMETRIQUEDES CINQ ESPECES ETUDIEES


Répartition spatio-temporelle
39
REPARTITION SPATIO-TEMPORELLE
DES CINQ ESPECES ETUDIEES
II/I.1 - INTRODUCTION
L'étude de l'influence des apports rhodaniens sur les populations de Solea solea, Solea impar,
Buglossidium luteum, Arnoglossus laterna et Citharus linguatula nécessite la connaissance préalable de
leur répartition spatio-temporelle au large du Rhône. En effet, dans la mesure où des différences
bathymétriques de sédimentation du matériel rhodanien et de composition de la macrofaune benthique
existent au large de l'embouchure du fleuve (Massé, 1972 ; Bouloubassi et al., 1997 ; Salen-Picard et
al., 1997), la répartition spatiale de ces cinq espèces de poissons plats va conditionner leur sensibilité
potentielle aux apports en matière organique d'origine continentale. De même, l'utilisation temporelle
des différentes tranches bathymétriques par les poissons (migrations saisonnières, installation benthique
des juvéniles) va déterminer leur capacité de réaction aux variations saisonnières des ressources
benthiques liées à celles des apports du Rhône. Les travaux portant sur les densités et les répartitions de
ces cinq espèces dans le golfe du Lion sont rares (Campillo et al., 1989 ; Marinaro, 1991 ; Campillo et
Bigot, 1992 ; Gaertner, 1999, 2000 ; Letourneur et al., 2001) et concernent essentiellement la partie
occidentale du golfe, la zone située en face de l'embouchure du Grand Rhône ayant été moins étudiée.
La majorité des données disponibles ne concernent qu'une voire deux espèce(s) et peu d'informations
existent sur leurs juvéniles (Campillo et al., 1989) ainsi que sur le peuplement des petits fonds sableux
côtiers (Letourneur et al., 2001). Ce chapitre a pour but de préciser la répartition spatio-temporelle des
individus de S. solea, S. impar, B. luteum, A. laterna et C. linguatula sur les fonds marins côtiers de 0 à
100 m de profondeur situés au large du Grand Rhône. Pour cela, les résultats concernant l’abondance et
la structure en taille des individus des cinq espèces seront exposés succinctement afin d'indiquer, pour
chacune d’elles, la ou les tranche(s) bathymétrique(s) où se déroule l'essentiel de la phase de vie
benthique et préciser les espèces et les stades de vie (juvénile, adulte) en interaction directe pour
l'utilisation des ressources alimentaires à chaque profondeur et chaque saison.
II/I.2 - MATERIEL ET METHODES
II/I.2.1 - DENSITES ET ABONDANCES RELATIVES
Le calcul des densités et des abondances relatives des cinq espèces de poissons plats a été
réalisé à partir des captures des missions SOLMAR, pour lesquelles des efforts de pêche (nombre de

Répartition spatio-temporelle
40
traits de chalut à panneaux) voisins ont été déployés par tranche bathymétrique (10-20 m = 30, 30-50 m
= 25, 70-100 m = 29), saison (hiver = 24 , printemps = 37, été = 23) et période du nycthémère (jour =
43, nuit = 32), ce qui simplifie les comparaisons croisées de ces différents paramètres.
Densités
Pour chaque trait de chalut, la densité de poissons par hectare a été calculée à l'aide de la
formule :
D =
où N représente l'effectif capturé pour chaque trait d'une demi-heure réalisé à 2nœuds (2 × 1852 m h-1) avec le chalut à panneaux (4 m d'ouverture au sol en pêche).
Les données ainsi obtenues ont servi à déterminer les densités moyennes par espèce ainsi que
celle des cinq espèces groupées. Les variations spatio-temporelles de ces densités ont ensuite été
étudiées en employant différents tests statistiques. Dans chaque cas, la Normalité des données brutes et
l'homogénéité des variances ont été testées au préalable par les tests de Shapiro-Wilks et de Bartlett au
seuil α=0,05. La majorité des espèces ne satisfaisant pas à ces deux critères, plusieurs transformations
mathématiques (X' = ln (X+1), X' = log (X+1), X' = √ (X+ 0,5), etc) ont été appliquées aux données afin
de normaliser la distribution des données et d'obtenir l'homoscédaticité nécessaire à l'utilisation des
méthodes paramétriques de comparaison d'échantillons, plus puissantes que leurs analogues non
paramétriques (Zar, 1999). Les fluctuations spatio-temporelles de la densité totale des cinq espèces ont
ainsi été testées par une ANOVA à deux facteurs croisés fixes (saison × profondeur) réalisée sur les
densités totales/trait de chalut après transformation X' = √ (X+ 0,5). Un test de Newman-Keuls a été
réalisé a posteriori afin de déterminer les profondeurs et les saisons responsables des différences
observées. De même, les fluctuations bathymétriques des densités de C. linguatula et d'A. laterna ont
été testées respectivement par un test T de Student et par une ANOVA à un facteur (profondeur) réalisés
sur les densités/trait de chalut après transformation X' = log (X+1). Dans le cas d'A. laterna (ANOVA),
un test de Newman-Keuls a été réalisé a posteriori afin de déterminer la(les) profondeur(s)
responsable(s) des différences observées. La normalité et l'homoscédaticité n'étant vérifiées ni pour S.
solea ni pour B. luteum (même après transformation des données), les différences de densité selon la
profondeur ont été testées, chez ces deux espèces, par une ANOVA non paramétrique Kruskal-Wallis
complétée par un test a posteriori de Dunn, proposé par Zar (1999) dans le cas d'échantillons d'effectifs
différents.
Abondances relatives
Les abondances relatives des cinq poissons plats dans les captures ont été déterminées au niveau
global ainsi qu'à chaque saison et chaque profondeur afin de déceler d'éventuelles modifications spatio-
N × 10 000
4 × 1852

Répartition spatio-temporelle
41
temporelles de la composition du peuplement formé par ces espèces. La normalité et l'homoscédaticité
des données n'étant pas vérifiées, les fluctuations spatio-temporelles de la composition du peuplement
de poissons plats ont été testées par une MANOVA non paramétrique à deux facteurs croisés fixes
(saison×profondeur) réalisée sur les effectifs capturés à chaque saison (S1= printemps, S2 = été, S3 =
hiver) pour chaque tranche bathymétrique (P1 = 0-20 m, P2 = 30-50 m, P3 = 70-100 m). Le logiciel
utilisé (NP MANOVA) fournit une approche alternative à l'analyse multivariée paramétrique en cas de
non normalité des données (Mc Ardle et Anderson, 2001). La méthode, basée sur le principe des tests de
permutations, calcule la probabilité d'obtenir une valeur égale ou supérieure à la valeur observée d'un
test statistique sous une hypothèse nulle donnée en recalculant la statistique du test après réarrangement
(permutation) des observations (Anderson, 2001). Le test statistique utilisé est essentiellement un F-
ratio, calculé comme le ratio de la somme du carré des distances inter-groupes divisée par la somme du
carré des distances intra-groupes (Anderson, 2000). Afin de disposer d'un nombre équivalent
d'observations par case (Pi × Sj), 6 traits de chalut ont été sous-échantillonnés de façon aléatoire pour
chaque doublet "saison × profondeur". L'analyse a été réalisée après transformation (X' = log (X+1)) des
données, afin d'assurer au mieux l'homogénéité des variances entre les différents groupes testés
(Anderson, 2000). Compte tenu du nombre élevé de zéros présents dans la matrice de données brutes, la
distance de Bray-Curtis a été utilisée pour l'obtention de la matrice de distances entre échantillons. Peu
sensible aux zéros, cette distance souligne les changements au sein des assemblages d'espèces en
donnant un poids identique aux espèces abondantes et aux espèces rares, contrairement aux distances
Euclidienne ou du χ2, qui accordent une importance maximale aux espèces rares et traduisent ainsi
davantage des changements de composition qualitative que de composition quantitative (Legendre et
Legendre, 1998 ; Legendre et Gallagher, 2001). Comme conseillé par Anderson (2000), le test a été
réalisé par permutation des résidus sous modèle réduit et avec un nombre élevé (4999) de permutations
afin d'assurer une puissance du test et une précision de l'erreur α maximales (Anderson et Legendre,
1999). Les tests de comparaisons a posteriori fournis par le programme ont été utilisés afin de préciser
le résultat obtenu pour la MANOVA. Basés sur un équivalant multivarié du test T de Student, ils ont
permis de déterminer les saisons et les profondeurs responsables des différences obtenues.
II/I.2.2 - POPULATIONS
Les modifications spatio-temporelles de la structure des populations ont été étudiées en fonction
de la longueur totale (LT en mm) des individus. Les équations reliant cette variable à la longueur
standard (LS en mm) et au poids (P en g) des individus ont néanmoins été établies pour chaque espèce
afin de permettre la comparaison des résultats obtenus ici avec ceux d'autres travaux utilisant ces deux
dernières mesures comme références (Annexe 1).

Répartition spatio-temporelle
42
Stades de vie benthiques
Afin de préciser l'utilisation spatio-temporelle de la zone côtière au large du Rhône au cours du
cycle de vie des espèces, les poissons récoltés ont été séparés en trois groupes ("stades") de vie
benthiques : les juvéniles de l'année (Juv. G0), les juvéniles de plus d'un an (Juv. G1+) et les sub-
adultes/adultes (Adultes). Cette répartition des individus a été réalisée suivant leur taille, à l'aide des
données bibliographiques disponibles sur la croissance des cinq espèces en Méditerranée (Tableau
II/I.1). Les tailles limites retenues pour chaque espèce ont été vérifiées par l'observation de l'état de
maturité des gonades des adultes en période de reproduction et par l'étude sous la loupe binoculaire de
la présence/absence du premier annulus à la marge des otolithes (sagittae) des juvéniles de taille proche
de la limite Juv. G0 / Juv. G1+.
Tableau II/I.1 – Données bibliographiques utilisées pour l'établissement des groupes de taille (LT, mm) correspondantaux trois "stades" de vie benthique (juvéniles G0, juvéniles G1+, adultes) étudiés chez Solea solea, S. impar,Buglossidium luteum, Arnoglossus laterna et Citharus linguatula au large du Rhône.
LT moyenne en fin de
première année- zone d'étude (référence)
Taille minimale d'acquisition
de la maturité sexuelle- zone d'étude (référence)
Limites de taille retenues
pour chaque groupe
S. solea
• 180 mm - golfe du Lion (Mendez de Elguezabal, 1978)
• 180-220 mm - golfe du Lion(Shéhata, 1984)
• 199 mm (Benouada, 1985)
• 300 mm (♀), 260 mm (♂) -
golfe du Lion (Mendez de
Elguezabal, 1978)
• 320 mm (♀) - golfe du Lion (Shéhata, 1984)
• Juv. G0 : 0-200 mm
• Juv. G1+ : 200-260 mm
• Adultes : LT > 260 mm
S. impar• 100 mm - golfe du Lion
(Marinaro, 1991)
• 160 mm (♀) - golfe du Lion (Shéhata, 1984 ; Marinaro, 1991)
• Juv. G0 : 0-100 mm
• Juv. G1+ : 100-160 mm
• Adultes : LT > 160 mm
B. luteum • 50 mm - golfe du Lion (Tito de Morais, 1983)
• 75 mm (♀/♂) - golfe du Lion(Shéhata, 1984)
• Juv. G0 : 0-50 mm
• Juv. G1+ : 50-70 mm
• Adultes : LT > 70 mm
A. laterna • 50 mm - golfe du Lion (Tito de Morais, 1983)
• 80 mm (♀) - Méditerranée (Avsar, 1994)
• 60-70 mm (♀/♂) - Atlantique(Gibson et Ezzi, 1980)
• Juv. G0 : 0-50 mm
• Juv. G1+ : 50-60 mm
• Adultes : LT > 60 mm
C. linguatula• 90 mm - Méditerranée
(Vassilopoulou et
Papaconstantinou, 1994)
• 160 mm (♀/♂) - Méditerranée(Vassilopoulou et Papaconstantinou,
1994)
• Juv. G0 : 0-100 mm
• Juv. G1+ : 100-160 mm
• Adultes : LT > 160 mm

Répartition spatio-temporelle
43
Exploitation des données
Afin de minimiser le biais d'échantillonnage lié à l'utilisation d'engins de capture différents
suivant la profondeur (senne de plage, gangui, chalut à panneaux) tout en conservant un maximum
d'information pour chaque espèce, deux méthodes d'exploitation des captures ont été appliquées.
Répartition spatio-temporelle des populations
Afin de décrire la stratégie d'occupation spatio-temporelle du milieu pour chaque espèce
(tranches bathymétriques fréquentées, migrations, période et profondeur de recrutement, etc.), des
histogrammes de fréquence de taille ont été établis à partir de la totalité des effectifs capturés à chaque
profondeur et saison. Pour cela, la première des trois tranches bathymétriques étudiées (0-20 m) a été
scindée en deux parties : les fonds entre 0 et 2 m de profondeur (échantillonnés à la senne de plage) et
ceux de 5-20 m (échantillonnés au chalut). Dans chacune des tranches de profondeur ainsi
individualisées (0-2 m, 5-20 m, 30-50 m, 70-100 m), les effectifs ont été regroupés en classes de LT de
20 mm (pour les poissons de grande taille : S. solea, S. impar et C. linguatula) ou de 10 mm (pour ceux
de petite taille : B. luteum et A. laterna) et les fréquences relatives des différentes classes de taille ont
été calculées à chaque saison.
Evolution des tailles en fonction de la profondeur
Pour suivre l'évolution de la structure en taille des populations selon la profondeur, les
longueurs totales moyennes des individus capturés à 0-20 m, à 30-50 m et à 70-100 m ont été calculées
pour chaque espèce. Pour ces calculs, seules les données des campagnes SOLMAR (réalisées avec le
chalut à panneaux) ont été utilisées afin d'éviter le biais lié à la sélectivité des engins de capture et
garantir un effort de pêche comparable par profondeur. L'influence de la profondeur sur la taille des
poissons a été testée pour chaque espèce excepté pour Solea impar, présente uniquement à 0-20 m. Pour
C. linguatula, un test T de Student a été réalisé sur les données de LT brutes qui, chez cette espèce,
satisfaisaient aux critères de normalité et d'homoscédaticité. Pour S. solea, B. luteum et A. laterna, la
normalité et l'homoscédaticité des données n'étant pas vérifiées même après transformation des données,
l'effet de la profondeur sur la taille des individus a été testé par une ANOVA non paramétrique de
Kruskal-Wallis, complétée par un test a posteriori de comparaisons multiples de Dunn (Zar, 1999). Les
pourcentages relatifs d'individus de chacun des stades de vie benthique définis précédemment (juvénile
G0, juvénile G1+, adulte) ont également été déterminés afin d'estimer l'importance de chaque tranche
bathymétrique dans le déroulement du cycle de vie de ces espèces.

Répartition spatio-temporelle
44
II/I.3 - RESULTATS
II/I.3.1 - DENSITES ET ABONDANCES RELATIVES DES CINQ ESPECES
Sur l'ensemble de la zone d'étude, la densité moyenne des cinq espèces groupées est de 138,6 ±
23,8 ind. ha-1. Le peuplement de poissons plats est dominé par A. laterna, suivi de B. luteum, les
densités de S. solea, C. linguatula, et S. impar étant moindres à cette échelle (Tableau II/I.2).
Tableau II/I.2 – Densités moyennes globales (moyenne ± erreur-type) obtenues sur l'ensemble de la zoned'étude (0-100 m de profondeur) pour les cinq espèces étudiées. Calculs réalisés à partir des 84 traits dechalut à panneaux des missions SOLMAR (voir Partie I). Les barres horizontales grises indiquent lesdensités moyennes non statistiquement différentes (P ≥ 0,05).
A. laterna B. luteum S. solea C. linguatula S. impar
Densité globale (ind. ha-1)
63 ± 9 41 ± 16 15 ± 4 11 ± 4 3 ± 1
% du total descaptures de poissons
plats47 % 31 % 11 % 8 % 2 %
Les faibles densités globales obtenues pour S. impar et C. linguatula reflètent en partie la
distribution bathymétrique réduite de leurs populations. En effet, les individus de ces deux espèces se
répartissent respectivement entre 0 et 20 m et entre 30 et 100 m de profondeur, alors que celles des trois
autres espèces (S. solea, B. luteum et A. laterna) couvrent l'ensemble de l'aire bathymétrique
échantillonnée (0-100 m). Citharus linguatula, dont la densité moyenne sur les fonds vaseux (30-100 m)
atteint 26 ± 3 ind. ha-1, est ainsi plus abondante sur son aire de répartition bathymétrique que la sole
commune S. solea. Par contre, la densité moyenne de S. impar reste faible, même ramenée aux seuls
fonds de 0-20 m (6 ± 1 ind. ha-1). Cette espèce est donc, de loin, la moins abondante sur la zone d'étude.
Pour l'ensemble des espèces étudiées, l'abondance des individus dépend de la tranche
bathymétrique considérée (Tableau II/I.3). Ce phénomène est particulièrement marqué chez S. impar et
C. linguatula, totalement absentes des captures à certaines profondeurs. Mais la densité moyenne des
individus diffère également selon la profondeur pour S. solea (Kruskal-Wallis : H(2, N = 72) = 28,4 ; P <
0,001), B. luteum (Kruskal-Wallis : H(2, N = 72) = 28,9 ; P < 0,001) et A. laterna (ANOVA : F(2, 69) = 9,3 ; P
< 0,001), avec des valeurs beaucoup plus élevées sur les petits fonds sableux côtiers qu'au niveau des
vases terrigènes plus profondes. Chez C. linguatula, la densité est plus élevée à 70-100 m qu'à 30-50 m
(Student : T(42) = -2,8 ; P < 0,01). Ces modifications bathymétriques de l'abondance des individus de
chaque espèce ont d'importantes conséquences sur la composition du peuplement de poissons plats à
chaque profondeur, avec une co-dominance d'A. laterna et B. luteum à 0-20 m, une majorité d'A. laterna
à 30-50 m et une légère prépondérance de C. linguatula à 70-100 m.

Répartition spatio-temporelle
45
Tableau II/I.3 – Fluctuations bathymétriques des densités (D moy. ± erreur-type, en ind. ha-1) des cinqespèces de poissons plats étudiées au large du Rhône. Les barres horizontales grises indiquent lesdensités moyennes non statistiquement différentes (P ≥ 0,05). "" = espèce absente à cette profondeur.N = nombre de traits de chaluts à panneaux réalisés à chaque profondeur (toutes saisons confondues) etayant servi pour les calculs.
Profondeur
Espèce
0-20 m (N = 30)
30-50 m (N = 25)
70-100 m (N = 29)
S. solea 38 ± 9 4 ± 2 1 ± 0,2
S. impar 6 ± 1
B. luteum 98 ± 38 3 ± 1 6 ± 3
A. laterna 110 ± 19 37 ± 6 29 ± 4
C. linguatula 14 ± 3 36 ± 7
Tableau II/I.4 – Fluctuations spatio-temporelles au large du Rhône de la densité totale (DT, en ind. ha-1)des cinq espèces groupées. N = nombre de traits de chalut réalisé à chaque profondeur et chaque saisonet utilisé pour les calculs (missions SOLMAR, c.f. Partie I).
DT : moyenne ± erreur-type N
Petits fonds sableux (0-20 m)
Hiver 50 ± 15 10Printemps 594 ± 173 9Eté 174 ± 28 11
Total 0-20 m 253 ± 54 30
Vases terrigènes supérieures (30-50 m)
Hiver 65 ± 15 8Printemps 52 ± 8 11Eté 88 ± 43 6
Total 30-50 m 61 ± 8 25
Vases terrigènes profondes (70-100 m)
Hiver 40 ± 16 6Printemps 76 ± 11 17Eté 77 ± 13 6
Total 70-100 m 69 ± 11 29

Répartition spatio-temporelle
46
HIV
ERPR
INTEM
PSÉTÉ
0-20 m
30-50 m
70-100 m
S. impar
2,6% S. solea10,5%
B. luteum20 ,2%
A. laterna 66,7%
A. laterna 32,8%
B. luteum62,9%
S. solea1,2%
S. impar
3,0%S. im
par1,7%
S. solea 29,3%
B. luteum 17,2%
A. laterna 51,8%
A. laterna 70,7%
B. luteum 12,6%
S. solea12,0%
C. linguatula
4,7%
A. laterna 62,7%
B. luteum2,6%
S. solea1,3%
C. linguatula
33,3%
C. linguatula
41,8%
S. solea 5,8%
B. luteum12,0%
A. laterna 40,4%
C. linguatula
13,1 %
S. solea3,4%
B. luteum11,1%
A. laterna 72,4%
C. linguatula
57,8% S. solea 0,6%
B. luteum4,6%
A. laterna37,0%
C. linguatula
56,8%
S. solea 1,0%
B. luteum 2,5%
A. laterna 39,7%
Figure II/I.1 – Fluctuations spatio-temporelles des abondances relatives des 5 espèces étudiées au large du R
hône. Les effectifs par espèce sont exprimés en pourcentages
relatifs du nombre total d'individus des cinq espèces capturés à chaque saison pour chaque profondeur.

Répartition spatio-temporelle
47
Au large du Rhône, la densité totale et la composition du peuplement de poissons plats
dépendent, non seulement de la tranche bathymétrique, mais également de la période de l'année
considérées. La densité totale des cinq espèces diffère de façon significative suivant la profondeur et la
saison (P < 0,001 ; Annexe 2), avec des valeurs plus élevées à 0-20 m qu'entre 30 et 100 m de
profondeur (P < 0,001) et une réduction générale des effectifs en hiver (P < 0,05) (Tableau II/I.4). Les
variations saisonnières concernent uniquement les petits fonds sableux côtiers où la densité des cinq
espèces varie significativement entre l'hiver, le printemps et l'été (P < 0,05). Sur les vases terrigènes
plus profondes, les modifications de densité totale ne sont pas significatives (P > 0,1) malgré des
valeurs plus élevées en été. La densité totale est maximale au printemps à 0-20 m (P < 0,001) mais reste
importante en été à cette profondeur (P < 0,05).
Les abondances relatives des cinq espèces diffèrent également en fonction de la profondeur et
de la saison (P < 0,001 ; Annexe 3) (Fig. II/I.1). A 0-20 m, le peuplement de poissons plats comprend, à
toutes les saisons, une quantité importante d'individus d'A. laterna et un nombre réduit de soles
adriatiques. Sa composition diffère cependant de façon significative (P < 0,05) entre les trois saisons
avec une dominance de B. luteum en été et une majorité d'A. laterna en hiver et au printemps, saisons où
S. solea est également proportionnellement plus abondante. A 30-50 m, les variations saisonnières du
peuplement sont peu marquées et non significatives (P > 0,07). Quelle que soit la saison, A. laterna
domine, suivi de C. linguatula et B. luteum. A 70-100 m, le peuplement est dominé par les individus de
C. linguatula et d'A. laterna quelle que soit la saison considérée mais sa composition diffère de façon
significative (P < 0,05) entre l'hiver, où A. laterna domine largement, et l'été et le printemps, où C.
linguatula est l'espèce la plus abondante.
II/I.3.2 - POPULATIONS DES CINQ ESPECES : STRUCTURE ET REPARTITION SPATIO-
TEMPORELLE
Au large du Rhône, les individus capturés sont essentiellement des juvéniles G0 pour S. solea et
S impar, des juvéniles G1+ pour C. linguatula et des adultes pour B. luteum et A. laterna (Tableau
II/I.5).
Tableau II/I.5 – Pourcentages des différents stades de vie benthique retenus (juvéniles G0, juvéniles G1+,adultes) dans les captures des cinq espèces de poissons plats étudiées au large du Rhône. Pour chaque espèce,le pourcentage le plus élevé est indiqué en gras.
% S. solea S. impar B. luteum A. laterna C. linguatula
Juvéniles G0 49 43 8 4 12
Juvéniles G1+ 23 37 12 13 50
Adultes 28 20 80 85 38
Total 100 100 100 100 100

Répartition spatio-temporelle
48
La structure en taille des populations, reflétée par celle des captures, varie cependant suivant la
saison et la profondeur considérées.
Modifications bathymétriques
Solea impar n'ayant été capturée qu'à 0-20 de profondeur, les modifications bathymétriques de
la structure des populations ne concernent que S. solea, B. luteum, A. laterna et C. linguatula. Pour C.
linguatula, la composition de la population est relativement stable d'une profondeur à l'autre. La taille
moyenne des individus ne diffère pas de façon significative (Student : T(298) = -1,4 ; P > 0,1) entre 30-50
m et 70-100 m de profondeur (Tableau II/I.6) et, mis à part une quantité légèrement plus importante de
G0 à 70-100 m, les individus de l'espèce se répartissent de façon homogène sur l'ensemble des fonds
vaseux étudiés (Tableau II/I.7). Par contre, la composition de la population dépend fortement de la
profondeur considérée pour S. solea, B. luteum et A. laterna. Pour ces trois espèces, la longueur totale
des individus capturés diffère de façon significative suivant la profondeur (Kruskal-Wallis : H(2, N=487) =
196,8 et P < 0,001 pour S. solea ; H(2, N=437) = 99,6 et P < 0,001 pour B. luteum ; H(2, N=2083) = 218,3 et P <
0,001 pour A. laterna), avec une augmentation de la taille moyenne des individus entre les fonds
sableux côtiers et les fonds vaseux plus profonds. Les différents stades de vie benthiques de ces trois
espèces se répartissent ainsi différemment en fonction du gradient de profondeur (Tableau II/I.7). Chez
S. solea, la densité des juvéniles G0 est maximale à 0-20 m et celle des juvéniles G1+ à 30-50 m, alors
que les individus adultes se répartissent de façon similaire entre 30-50 m et 70-100 m. Chez B. luteum et
A. laterna, les densités de juvéniles G0, et de juvéniles G1+ sont maximales à 0-20 m alors que les
individus adultes se répartissent principalement sur les fonds vaseux entre 30 et 100 m de profondeur,
avec un maximum d'abondance à 30-50 m.
Tableau II/I.6 – Tailles moyennes (LT en mm) et erreurs-types des individus des cinqespèces à chaque profondeur étudiée au large du Rhône. Les barres horizontalesgrises indiquent les tailles moyennes non statistiquement différentes (P ≥ 0,05). ""= espèce absente à cette profondeur.
LT (mm) 0-20 m 30-50 m 70-100 m
S. solea 138,6 ± 2,9 224,7 ± 3,8 271,3 ± 13,3
S. impar 101,9 ± 4,1
B. luteum 80,2 ± 1,6 99,1± 1,1 99,5 ± 0,9
A. laterna 76,5 ± 0,8 94,7 ± 0,4 95,2 ± 0,4
C. linguatula 146,1 ± 0,5 152,3 ± 0,4

Répartition spatio-temporelle
49
Tableau II/I.7 – Répartition bathymétrique des différents stades de vie benthique (juvéniles G0,juvéniles G1+ et adultes) des cinq espèces étudiées au large du Rhône. Dans chaque cas, lepourcentage maximal d'individus obtenu pour chaque stade est indiqué en gras. "" = stadeabsent à cette profondeur.
LT (mm) 0-20 m 30-50 m 70-100 m
S. solea Juv. G0Juv. G1+
Adultes
94,67,9
10,5
5,484,546,9
7,6
42,6
S. impar Juv. G0 Juv. G1+
Adultes
100,0100,0100,0
B. luteum Juv. G0 Juv. G1+
Adultes
100,083,415,9
16,662,5
21,6
A. laterna Juv. G0 Juv. G1+
Adultes
100,067,510,4
25,256,0
7,3
33,6
C. linguatula Juv. G0 Juv. G1+
Adultes
31,149,743,0
68,950,357,0
Modifications saisonnières
Pour chacune des cinq espèces de poissons plats étudiées, la structure en taille des effectifs
capturés dépend de la saison considérée (Fig. II/I.2 à II/I.6). Pour S. solea, les plus petits juvéniles (G0
de LT < 100 mm) sont capturés au printemps (avril, mai, juin), principalement à 0-2 m et jamais au delà
de 20 m de profondeur (Fig. II/I.2). Les juvéniles G0 de 100 à 200 mm de LT présentent un maximum
d'abondance entre 5 et 20 m en été mais sont capturés à toutes les saisons entre 5 et 50 m de profondeur.
Les individus de LT > 200 mm (juvéniles G1+ et adultes) sont présents à toutes les saisons entre 5 et 100
m de profondeur, avec cependant une abondance maximale en hiver sur les fonds vaseux (30-100 m).
Chez S. impar, les plus petits juvéniles (G0 : LT < 100 mm) ne sont capturés qu'au printemps
(mai-juin), principalement entre 0 et 2 m de profondeur (Fig. II/I.3). Les individus de plus de 100 mm
de LT (juvéniles G1+ et adultes) sont présents à toutes les saisons, essentiellement à 5-20 m de
profondeur. Ainsi, bien que l'ensemble des stades de vie benthiques de S. impar se répartissent
exclusivement entre 0 et 20 m, l'installation benthique des juvéniles se fait sur des zones moins
profondes (0-2 m) que celles occupées par les adultes, présents uniquement à partir de 5 m de
profondeur.

Répartition spatio-temporelle
50
Pour B. luteum, les plus petits individus (G0 : LT < 50 mm) sont capturés uniquement à 5-20 m
de profondeur au printemps (juin) et en été (juillet, août, septembre) (Fig. II/I.4). Les juvéniles de 50-70
mm sont surtout présents entre 5 et 20 m de profondeur où ils sont essentiellement capturés au
printemps et en été. Les individus adultes (LT > 70 mm) sont présents quelle que soit la saison au niveau
des trois tranches bathymétriques.
Chez A. laterna, les juvéniles G0 (LT < 50 mm) sont capturés entre 5 et 20 m, essentiellement au
printemps (mai - juin) mais également en été (Fig. II/I.5). Les juvéniles G1+ (50-60 mm) sont présents à
toutes les profondeurs, avec un maximum d'abondance entre 5 et 20 m, surtout au printemps et en été.
Les adultes (LT > 60 mm) sont toujours abondants, quelles que soient la saison et la profondeur.
Pour C. linguatula, les plus petits G0 (LT < 100 mm) sont principalement capturés au printemps
entre 70 et 100 m de profondeur (Fig. II/I.6). Les juvéniles G1+ (100-160 mm) sont capturés à 30-50 m
et à 70-100 m à toutes les saisons. Les adultes (LT > 160 mm) sont présents sur l'ensemble des fonds
entre 30 et 100 m de profondeur excepté en été à 30-50 m.
Dans chaque cas, les modifications saisonnières de la structure de taille des effectifs capturés
sont surtout liées au recrutement des juvéniles, sur les petits fonds sableux côtiers de 0-20 m pour S.
solea, S. impar, B. luteum, A. laterna et sur les vases terrigènes situées entre 30 et 100 m de profondeur
pour C. linguatula. Les migrations saisonnières semblent réduites sur la zone d'étude, la répartition
bathymétrique et la densité des juvéniles G1+ et des adultes changeant peu au cours de l'année.
II/I.4 - DISCUSSION
Densités et abondances relatives des cinq espèces
Quelle que soit la saison considérée, les juvéniles et les adultes des cinq poissons plats sont
présents en grand nombre au large du Rhône. La densité totale des cinq espèces est élevée, surtout à 0-
20 m (253 ind. ha-1), où elle représente environ les 2/3 de la densité moyenne de poissons signalée sur
cette zone (401 ind. ha-1) (Letourneur et al., 2001). A elles seules, A. laterna, C. linguatula et S. solea
représentent près de 11% en abondance et 10% en biomasse des poissons démersaux côtiers du golfe du
Lion (Blanchard, 2001). Les densités obtenues pour les cinq espèces sont en accord avec les données
disponibles sur leurs abondances au large du Rhône. Les valeurs trouvées à 0-20 m pour B. luteum (99
ind. ha-1) et A. laterna (110 ind. ha-1) sont proches de celles proposées pour ces deux espèces sur ces
fonds par Letourneur et al. (2001) (respectivement 89 et 114 ind. ha-1). De même, la densité de C.
linguatula entre 30 et 100 m de profondeur (26 ind. ha-1) est voisine de celles indiquées pour l'espèce
dans la partie Est du golfe (20-30 ind. ha-1) (Campillo et al., 1989). Les densités de S. solea observées
sur les petits fonds sableux côtiers (38 ind. ha-1) et sur les vases terrigènes plus profondes (1-4 ind. ha-1)
sont en accord avec les données fournies par Letourneur et al. (2001) et Campillo et al. (1989) pour ces
tranches bathymétriques.
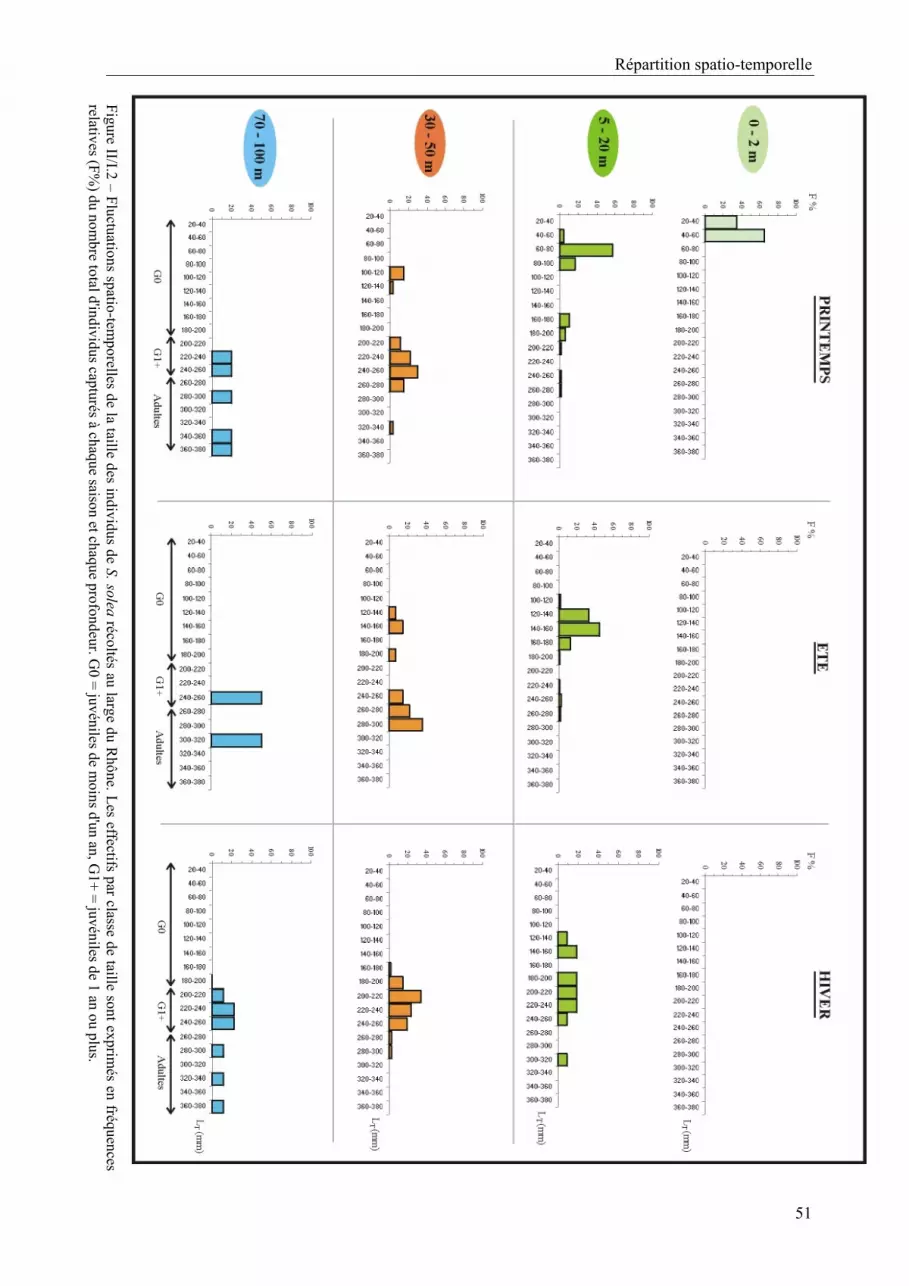
Répartition spatio-temporelle
51
Figure II/I.2 – Fluctuations spatio-temporelles de la taille des individus de S. solea récoltés au large du R
hône. Les effectifs par classe de taille sont exprimés en fréquences
relatives (F%) du nom
bre total d'individus capturés à chaque saison et chaque profondeur. G0 = juvéniles de m
oins d'un an, G1+ = juvéniles de 1 an ou plus.

Répartition spatio-temporelle
52
Figure II/I.3 – Fluctuations spatio-temporelles de la taille des individus de S. im
par récoltés au large du Rhône. Les effectifs par classe de taille sont exprim
és en fréquencesrelatives (F%
) du nombre total d'individus capturés à chaque saison et chaque profondeur. G
0 = juvéniles de moins d'un an, G
1+ = juvéniles de 1 an ou plus.

Répartition spatio-temporelle
53
Figure II/I.4 – Fluctuations spatio-temporelles de la taille des individus de B. luteum
récoltés au large du Rhône. Les effectifs par classe de taille sont exprim
és enfréquences relatives (F%
) du nombre total d'individus capturés à chaque saison et chaque profondeur. G
0 = juvéniles de moins d'un an, G
1+ = juvéniles de 1 an ou plus.

Répartition spatio-temporelle
54
Figure II/I.5 – Fluctuations spatio-temporelles de la taille des individus d'A. laterna récoltés au large du R
hône. Les effectifs par classe de taille sont exprimés en fréquences
relatives (F%) du nom
bre total d'individus capturés à chaque saison et chaque profondeur. G0 = juvéniles de m
oins d'un an, G1+ = juvéniles de 1 an ou plus.

Répartition spatio-temporelle
55
Figure II/I.6 – Fluctuations spatio-temporelles de la taille des individus de C
. linguatula récoltés au large du Rhône. Les effectifs par classe de taille sont exprim
és enfréquences relatives (F%
) du nombre total d'individus capturés à chaque saison et chaque profondeur. G
0 = juvéniles de moins d'un an, G
1+ = juvéniles de 1 an ou plus.

Répartition spatio-temporelle
56
Seule la densité de S. impar (6 ind. ha-1) est inférieure aux valeurs précédemment obtenues (14
ind. ha-1) (Letourneur et al., 2001). Parmi les cinq espèces, A. laterna et B. luteum dominent dans les
captures. Les abondances des trois autres espèces (S. solea, S. impar et C. linguatula) sont plus faibles,
avec un minimum pour S. impar. Cette situation a déjà été décrite sur les fond de 0-20 m de profondeur,
où les densités annuelles de B. luteum et A. laterna dépassent largement celles de S. solea et S. impar
(Darnaude et al., 2001 ; Letourneur et al., 2001), mais ces données doivent néanmoins être relativisées,
les densités relatives obtenues étant en partie biaisées par la sélectivité des engins employés. En effet,
les G0 de B. luteum et d'A. laterna (LTmax.= 50 mm) sont moins bien retenus par les chaluts utilisés
(vides de maille de 20 et 8 mm minimum) que ceux des trois autres espèces, de taille plus importante
(LTmax.= 160-200 mm). Il est également probable que les adultes des espèces de grande taille (LTmax.>
200 mm) échappent plus facilement aux engins de capture du fait de capacités de fuite plus importantes.
Ceci expliquerait en partie que les prises de S. solea, S. impar et C. linguatula comprennent une
majorité de juvéniles (LT.= 60-160 mm) alors que celles de B. luteum et A. laterna sont dominées par les
adultes (LT.= 60-130 mm). Cependant, la dominance des poissons de petite taille et l'importance des
juvéniles dans les populations des espèces de grande taille peuvent également être considérées comme
une conséquence de l’exploitation halieutique de la zone. L'impact de la pêche sur la structure des
peuplements a été démontré pour de nombreux assemblages démersaux (Bianchi et al., 2000). Il se
traduit le plus souvent par une réduction globale de la taille des poissons résultant non seulement d'une
réduction des effectifs des espèces cibles (en général de grande taille) au profit des autres espèces (de
taille plus réduite) mais également d'une altération de la structure démographique des espèces capturées
en faveur des juvéniles. Un tel phénomène a été mis en évidence pour les stocks de poissons démersaux
du golfe du Lion (Blanchard et Boucher, 2001). L'exploitation commerciale des adultes de S. solea et, à
moindre échelle, de C. linguatula et S. impar dans la région expliquerait donc également en partie le
schéma observé pour ces espèces au large du Rhône.
Répartitions bathymétriques
Les différences de répartition bathymétrique des cinq espèces confirment les données
préexistantes sur leur distribution spatiale dans le golfe du Lion, avec une répartition très côtière (0-15
m) chez S. impar (Marinaro, 1991), relativement profonde (> 30 m) chez C. linguatula (Campillo et al.,
1989 ; Sartor et al., 2002) et plus étendue chez S. solea, B. luteum et A. laterna, dont les populations se
répartissent généralement entre 0 et 100 m de profondeur (Campillo et al., 1989 ; Campillo et Bigot,
1992 ; Gaertner, 1997, 1999). L'enfoncement bathymétrique des individus avec la taille observé chez S.
solea, B. luteum et A. laterna est commun à de nombreuses espèces démersales (Macpherson et Duarte,
1991). Il correspondrait à un comportement évolutif inné visant à augmenter la longévité des adultes par
une migration vers les eaux profondes où les températures, plus faibles, garantissent des taux
métaboliques inférieurs et donc un vieillissement cellulaire moindre. Les modifications bathymétriques

Répartition spatio-temporelle
57
des densités de S. solea, B. luteum, A. laterna et C. linguatula sont conformes aux données de la
littérature. Dans le golfe du Lion, B. luteum est décrite comme essentiellement côtière et A. laterna
comme très abondante à la côte mais également dans la partie centrale du plateau continental (Gaertner,
1997, 1999). Les concentrations de C. linguatula et de S. solea dans le golfe du Lion sont
respectivement maximales entre à 70-130 m et 0-20 m de profondeur (Campillo et al., 1989 ; Sartor et
al., 2002), ce qui correspond à nos observations.
Modifications saisonnières
Le maillage réduit (8 mm) de la senne de plage et du gangui utilisés, adapté à la capture des
juvéniles dès 20 mm de LT, a permis une estimation fiable des périodes de présence des plus petits
juvéniles de S. solea, S. impar, B. luteum et A. laterna sur les fonds sableux où se fait leur installation
benthique (Darnaude, 1999 ; Letourneur et al., 2001). Ces périodes sont légèrement décalées dans le
temps. Les plus petits juvéniles sont essentiellement capturés d'avril à mai pour S. solea, de mai à juin
pour S. impar et A. laterna et de juin à juillet pour B. luteum, ce qui est en accord avec les données
disponibles sur la reproduction et l'installation benthique de ces espèces dans le golfe du Lion (Tito de
Morais, 1983 ; Shéhata, 1984 ; Marinaro, 1991). Le maillage plus lâche (20 mm) du chalut utilisé sur les
vases ne permettant pas la capture des plus petits poissons, les dates de récolte des juvéniles G0 en
profondeur sont par contre susceptibles d'être décalées de plusieurs mois par rapport à celles de
l'installation benthique. Ainsi, pour C. linguatula, dont la reproduction et l’installation benthique
s'effectuent à l'automne en Méditerranée (Sabatés, 1988), les juvéniles n'apparaissent dans nos captures
qu'au printemps, à une taille minimale de 60 mm. L'augmentation, au printemps et en été, des prises de
juvéniles G0 des cinq espèces correspond donc moins à une arrivée massive des plus juvéniles sur la
zone à ces deux saisons qu'au franchissement, par ces individus, du seuil de sélectivité des engins de
pêche utilisés. Il est ainsi probable que des densités de poissons plats beaucoup plus fortes soient
atteintes ponctuellement au large du Rhône lors de l'installation benthique des juvéniles des cinq
espèces, en fin d'hiver et au printemps à 0-20 m (S. solea, S. impar, B. luteum, A. laterna) ou en
automne à 30-100 m (C. linguatula). Ceci a d'importantes conséquences potentielles sur les peuplements
de poissons plats au large du Rhône. En effet, sur la zone d'étude, les migrations saisonnières des cinq
espèces sont peu marquées (ce résultat sera repris et discuté plus loin, c. f. partie IV). Leur influence sur
la densité et la composition du peuplement est donc négligeable et l'essentiel des modifications spatio-
temporelles observées au cours de cette étude sont imputables à l'augmentation d'abondance des
juvéniles G0 dans les captures. A 0-20 m, les pics successifs d'abondance des plus petits juvéniles de S.
solea, S. impar, B. luteum et A. laterna expliquent non seulement les densités de poissons plats élevées
mais très variables obtenues au printemps et en été, mais aussi les modifications saisonnières de
composition du peuplement. De même, à 70-100 m, de fortes abondances de juvéniles de C. linguatula
au printemps et en été expliquent la dominance numérique de cette espèce dans les captures de ces deux

Répartition spatio-temporelle
58
saisons. En dehors des périodes de recrutement, la composition du peuplement est plut stable, avec une
large dominance d'adultes d'A. laterna quelle que soit la profondeur. Les nombres différents d'espèces
recrutant à 0-20 m (4) et à 30-100 m de profondeur (1) entraînent une séparation nette entre fonds
sableux côtiers, où la densité moyenne des espèces étudiées est élevée mais varie fortement d'une saison
à l'autre, et vases terrigènes plus profondes, où les variations saisonnières sont réduites mais la densité
moyenne plus faible. Ces fluctuations spatio-temporelles d'abondance sont susceptibles d'avoir
d'importantes conséquences pour l'ensemble de la communauté ichtyique benthique au large du Rhône.
En effet, l'installation sur le fond des juvéniles de S. solea, S. impar, B. luteum et A. laterna est
responsable de l'essentiel des modifications saisonnières de la densité totale de poissons sur les petits
fonds sableux de la zone d'étude (Letourneur et al., 2001).
II/I.5 - CONCLUSION
Les populations de S. solea, S. impar, B. luteum, A. laterna et C. linguatula exploitent les fonds
marins côtiers situés au large du Rhône de façon permanente, avec un maximum d'abondance au
printemps et en été suite à l'installation benthique des jeunes recrues (à 0-2 m pour S. solea et S. impar,
5-20 m pour B. luteum et A. laterna et 30-100 m pour C. linguatula). L'importance de cette zone côtière
dans le cycle de vie de ces poissons plats est donc primordiale, ce secteur servant non seulement de
nurserie aux juvéniles mais supportant également de fortes densités d'adultes, tant pendant leurs
périodes de reproduction qu'au cours du repos sexuel. Cependant, les cinq espèces se répartissent
différemment dans l'espace. Pour S. solea, B. luteum et A. laterna, le recrutement et l'essentiel de la
croissance se font à 0-20 m, les adultes se répartissant ensuite surtout entre 30 et 50 m de profondeur.
On assiste donc, chez ces trois espèces, à un changement de répartition bathymétrique entre juvéniles et
adultes qui limite les interactions entre ces deux stades de vie alors que, chez S. impar et C. linguatula,
juvéniles et adultes occupent les mêmes fonds, entre 0 et 20 m de profondeur chez S. impar, et au delà
de 30 m de profondeur chez C. linguatula. Ces différences de répartition bathymétrique vont déterminer
en partie la sensibilité des poissons aux apports du fleuve, compte tenu des différences de sédimentation
de la MOP terrigène observées avec la profondeur. Elles vont également conditionner l'intensité des
interactions trophiques entre stades de vie benthique et espèces, soit par ségrégation bathymétrique
(pour S. impar et C. linguatula par exemple), soit par les différences de densité qu'elles entraînent. Les
interactions entre individus sont ainsi potentiellement maximales à 0-20 m au printemps et en été, où les
densités totales sont les plus fortes et où l'ensemble des stades de vie benthique de S. solea, S. impar, B.
luteum et A. laterna sont réunis. Elles sont en revanche minimales en hiver à 70-100 m, où la densité
totale est réduite et où seuls les adultes de S. solea, B. luteum, A. laterna et C. linguatula sont présents.
Les implications de ces différences de répartition et d'abondance seront discutées en termes de partage
des ressources et de stratégies adaptatives vis-à-vis des conditions du milieu dans la Partie IV.

PARTIE II – CHAPITRE II
STRATEGIES ALIMENTAIRESDES CINQ ESPECES


Stratégies alimentaires
59
STRATEGIES ALIMENTAIRES
DES CINQ ESPECES
II/II.1 - INTRODUCTION
L'étude de l'influence des crues du Rhône sur les populations de Solea solea, Solea impar,
Buglossidium luteum, Arnoglossus laterna et Citharus linguatula nécessite une connaissance précise de
la composition de leur régime alimentaire au large de l'embouchure. En effet, la capacité de chaque
prédateur à tirer ou non profit d'une modification de la nature ou de l'abondance des proies disponibles
dans son environnement dépend en grande partie de ses preferenda alimentaires et de la plasticité de son
alimentation. Les stratégies alimentaires des cinq espèces vont ainsi conditionner leurs sensibilités
respectives vis-à-vis des modifications du benthos, elles-même liées aux variations des apports
rhodaniens en matière organique au niveau du fond. Solea solea et B. luteum sont généralement décrits
comme présentant une alimentation surtout nocturne (Rogers et Jinadasa, 1989), des modes de détection
des proies similaires basés sur l'olfaction (De Groot, 1969) et des régimes alimentaires proches,
essentiellement composés de polychètes et de bivalves (De Groot, 1971). L'alimentation de S. impar, A.
laterna et C. linguatula est le plus souvent dominée par les crustacés (Marinaro et Bouabib, 1983 ; Tito
de Morais et Bodiou, 1984 ; Belghyti et al., 1993), A. laterna et C. linguatula présentant des rythmes et
des modes de nutrition similaires, avec une alimentation surtout diurne (Gibson et Ezzi, 1980 ; Belghyti
et al., 1995) associée à une détection visuelle des proies (De Groot, 1969). Les données disponibles sur
l'alimentation de ces cinq espèces en Méditerranée sont cependant peu abondantes. Le régime
alimentaire de S. solea a été étudiée par un grand nombre d'auteurs mais principalement en Atlantique
(Steven, 1930 ; Sorbe, 1972 ; Braber et De Groot, 1973 ; Castel et Lasserre, 1982 ; Lê, 1983 ;
Lagardère, 1987 ; Amara et al., 2001), les études réalisées en Méditerranée étant moins nombreuses
(Reys, 1960 ; Mendez de Elguezabal, 1978 ; Molinero et al., 1991 ; Molinero et Flos, 1992a, 1992b ;
Darnaude et al., 2001). Les travaux portant sur l'alimentation de S. impar (Marinaro et al., 1983 ; Allam,
1995 ; Rodriguez, 1996), B. luteum (Tito de Morais, 1983, 1984 ; Tito de Morais et Bodiou, 1984 ; Tito
de Morais, 1986 ; Darnaude et al., 2001), A. laterna (Gibson et Ezzi, 1980 ; Rogers, 1991 ; Avsar, 1994
; Darnaude et al., 2001) et C. linguatula (Belghyti et al., 1993 ; Redon et al., 1994 ; Belghyti et al.,
1995) sont encore moins abondants, probablement du fait de l'importance commerciale réduite de ces
quatre espèces. Parmi eux, rares sont ceux qui concernent la Méditerranée. Il est donc possible que les
régimes alimentaires de S. solea, S. impar, B. luteum, A. laterna et C. linguatula au large du Rhône
diffèrent de ceux communement admis pour ces espèces.

Stratégies alimentaires
60
Même au sein d'une zone géographique réduite, plusieurs facteurs déterminent et modifient la
composition du régime alimentaire des poissons. Celle-ci subit souvent d'importantes modifications au
cours de la croissance et diffère entre juvéniles et adultes d'une même espèce (Nikolskii, 1969). De
telles modifications de l'alimentation avec la taille des individus ont été décrites chez S. solea
(Lagardère, 1989 ; Molinero et Flos, 1992a), S. impar (Allam, 1995 ; Rodriguez, 1996), B. luteum (Tito
de Morais et Bodiou, 1984), A. laterna (Avsar, 1994) et C. linguatula (Belghyti et al., 1993 ; Redon et
al., 1994). Chez de nombreux Pleuronectiformes, ces variations ontogéniques de l'alimentation résultent
en partie du fait que les zones de nourricerie sont différentes des aires où vivent les adultes. C'est le cas
pour S. solea, B. luteum et A. laterna, dont les juvéniles se répartissent majoritairement au niveau des
zones sableuses de faible profondeur, alors que les adultes sont essentiellement capturés sur les fonds
vaseux plus profonds (Campillo et al., 1989 ; Gaertner, 1999 ; Letourneur et al., 2001; présente étude).
Les effets de la taille des individus et de la profondeur sur l'alimentation sont donc étroitement liés chez
ces trois espèces, pour lesquelles la composition du régime alimentaire est très variable et dépend
fortement du milieu dans lequel les individus se répartissent (De Groot, 1971 ; Darnaude et al., 2001).
Enfin, l'alimentation des poissons peut varier selon la saison considérée, ce qui a déjà été démontré pour
les cinq espèces étudiées (Molinero et Flos, 1992b ; Redon et al., 1994 ; Rodriguez, 1996 ; Darnaude et
al., 2001). Des différences saisonnières de l'intensité de la nutrition et de la composition du régime
alimentaire sont donc susceptibles de s'ajouter aux variations de l'alimentation liées à la taille et/ou à la
profondeur. L'ensemble de ces facteurs de variation doit être pris en compte pour l'étude de
l'alimentation des cinq espèces, afin de décrire au mieux les voies potentielles de cheminement de la
MOP terrigène vers les poissons. De plus, les apports en MOP rhodanienne au niveau du fond variant
suivant la profondeur et la saison (Cauwet, 1996 ; Bouloubassi et al., 1997), la quantité de matière
organique d'origine terrestre disponible à la base des réseaux trophiques benthiques (et donc l'impact du
Rhône sur la composition et l'abondance du macrobenthos) varie suivant la tranche bathymétrique et la
période de l'année, ce qui est susceptible d'influencer de façon différente les divers stades de vie
benthique des poissons via leur alimentation.
Ce chapitre a pour but de décrire de façon précise l'alimentation de S. solea, S. impar, B. luteum,
A. laterna et C. linguatula au large du Rhône. Pour cela, la composition du régime alimentaire global et
les modifications de l'alimentation selon la taille des individus, la saison et la profondeur ont été
étudiées. La méthode employée et les principaux résultats obtenus pour chaque espèce seront détaillés
avant de conclure sur les différences interspécifiques de stratégies alimentaires.
II/II.2 - MATERIEL ET METHODES
Pour chaque espèce, le régime alimentaire a été déterminé à partir des estomacs pleins (i.e.
contenant de la nourriture) d'un minimum de 10 individus par stade de vie benthique (juvéniles G0,
juvéniles G1+, adultes) présent à chaque saison et à chaque profondeur. Afin d'étudier les rythmes

Stratégies alimentaires
61
nycthéméraux et saisonniers de l'alimentation, le coefficient de vacuité CV (pourcentage d’estomacs
vides par rapport au nombre total d’estomacs examinés) a été déterminé pour chaque période. Le niveau
de réplétion des estomacs a également été estimé suivant un indice de réplétion (IR) variant de 0 à 4 (0
= vide; 1 = occupation de 1 à 25% du volume; 2 = de 26 à 75% du volume; 3 = de 76 à 99% du volume;
4 = totalement rempli voire distendu). Le côté subjectif de cette méthode ne peut être nié mais elle
présente l’avantage d’être rapide et facile à réaliser tout en fournissant une information suffisante pour
estimer par la suite les rythmes d’alimentation.
Pour chaque estomac contenant de la nourriture, les proies ingérées ont été identifiées et triées
par type de proie. A ce stade, une partie du matériel présent dans certains estomacs a été volontairement
négligée. C'est le cas des débris végétaux d'origine terrestre qui forment par endroits un épais tapis à la
surface du sédiment (Massé et al., 1977) et sont essentiellement avalés lors de la capture des proies
benthiques. Du fait de leur fort taux en lignine, ils sont peu assimilables par les poissons et n'ont donc
pas été comptabilisés en tant que catégorie de proie, tout comme les grains de sable et les particules
fines de sédiment rencontrés. Pour chaque catégorie de proie retenue, le nombre d’individus ingérés par
estomac a été évalué le plus précisément possible, bien que le degré élevé de digestion de certaines
proies (notamment les polychètes et les phoronidiens) ait parfois rendu cette estimation délicate. Le
poids représenté par les individus de chaque catégorie de proie n’a pu être déterminé pour chaque
estomac, la très petite taille de certaines proies rencontrées (notamment les copépodes) rendant la
mesure irréalisable pour des raisons de précision de la balance utilisée. Le poids total en matière sèche
(pièces dures comprises) de l'ensemble des individus récoltés pour un même type de proie a donc été
mesuré par classe de taille de poissons, pour chaque saison, profondeur et période de la journée. Un
poids moyen a ensuite été calculé par individu. Pour cela, les différents lots de proies ont été séchés en
étuve (60°C) pendant 24 h avant d’être pesés, au µg près. Pour les proies de poids inférieur au
microgramme, les individus des différentes classes de LT ont été pesés groupés et un poids moyen par
individu a ensuite été calculé.
Traitement des données : indices alimentaires
Les résultats de l’analyse des contenus stomacaux ont été exploités suivant la méthode mixte,
numérique et pondérale, mise au point par Hureau (1970). Cette méthode s’applique essentiellement à
l’étude des carnivores ingérant des invertébrés non coloniaux dont les individus peuvent être aisément
comptés. Elle apporte alors le maximum de renseignements sur l’alimentation d’une espèce (Harmelin-
Vivien, 1979). Pour chaque groupe de poissons étudié, les indices suivants ont été calculés :
- l’indice de fréquence F de chaque type de proie (rapport entre le nombre n d’estomacs
renfermant cette proie et le nombre N’ d’estomacs contenant de la nourriture) :
F = n / N’

Stratégies alimentaires
62
- le pourcentage numérique Cn (rapport entre le nombre N d’individus recensés pour un type de
proie donné et le nombre total NT des différentes proies rencontrées) :
Cn = N × 100 / NT
- le pourcentage pondéral Cp (rapport entre le poids P des individus recensés pour un type de
proie donné et le poids total PT de toutes les proies rencontrées) :
Cp = P × 100 / PT
Ces trois indicateurs (F, Cn et Cp) fournissent le maximum d’information sur la nourriture et le
comportement alimentaire des poissons (Hyslop, 1980).
L'efficacité des indices composés pour décrire les régimes alimentaires a souvent été contestée
(Tirasin et Jorgensen, 1999) du fait des phénomènes de redondance existant entre le nombre, le poids et
la fréquence des différentes proies (Macdonald et Green, 1983). Cependant, seule l'utilisation d'un
coefficient composé tenant compte, au moins, du nombre et du poids des proies ingérées peut permettre
une bonne appréciation de leur importance relative dans le régime alimentaire de poissons présentant
des tailles aussi différentes que celles rencontrées ici (variant de 3 à plus de 30 cm). Afin de pouvoir
quantifier de façon plus précise l'importance de chaque type de proie au sein du régime alimentaire des
poissons, le coefficient alimentaire Q (Hureau, 1970) de chaque proie a donc été calculé, suivant la
formule :
Q = Cn × Cp
Suivant Hureau (1970), les proies ont été classées en trois catégories selon la valeur de leur
coefficient alimentaire : proies "préférentielles" (Q ≥ 200), proies "secondaires" (20 ≤ Q < 200) et proies
"occasionelles" (Q < 20). Ces valeurs limites sont arbitraires comme le souligne Hureau (1970) mais en
accord avec l’importance relative des proies ingérées par les cinq espèces étudiées ici. Une dernière
catégorie de proies, dites "accidentelles", a été créée pour les proies de Q < 2 et/ou F < 0,01.
Exploitation des données
Pour chaque espèce, l'étude des fluctuations de l'indice de réplétion stomacal au cours du
nycthémère a permis de préciser la période d'alimentation principale des poissons. La composition
globale du régime alimentaire a ensuite été établie afin de préciser les proies les plus consommées.
Enfin, les variations ontogéniques et spatio-temporelles de l'alimentation ont été déterminées du point
de vue quantitatif (indice de réplétion stomacal, nombre et poids des proies) et qualitatif (composition
du régime alimentaire).

Stratégies alimentaires
63
Variations de l'intensité de l'activité alimentaire
Les variations de l'intensité de l'activité alimentaire suivant la période du nycthémère, la saison
et la profondeur ont été étudiées pour chacune des cinq espèces. Pour cela, les indices de réplétion
stomacaux (IR) obtenus pour les poissons capturés (1) de jour ou de nuit, (2) au printemps, en été ou en
hiver, (3) à 0-20 m, 30-50 m ou 70-100 m, ont été comparés après sous-échantillonnage des individus
afin d'éliminer, dans chaque cas, le biais lié à un effet potentiel des deux autres facteurs (par exemple,
pour l'étude des variations nycthémérales, un nombre similaire de poissons de chaque saison /
profondeur a été extrait au hasard parmi les effectifs récoltés de jour et ceux récoltés de nuit). La
normalité (test de Shapiro-Wilk, seuil α = 0,05) et l'homoscédaticité (test de Bartlett, seuil α = 0,05)
n'étant vérifiées pour aucune espèce et aucun des facteurs étudiés (période, saison, profondeur), même
après transformation des données (X' = ln (X+1), X' = log (X+1), X' = √ (X+ 0,5), etc), des méthodes
non paramétriques de comparaison d'échantillons ont été utilisées. L'existence ou non, d'une différence
significative de l'indice de réplétion stomacal des poissons entre les périodes diurne et nocturne a ainsi
été déterminée pour S. solea, B. luteum, A. laterna et C. linguatula par un test U de Mann-Whitney∗. De
même, les différences saisonnières et bathymétriques de l'indice de réplétion stomacal ont été testées par
une ANOVA non paramétrique de Kruskal-Wallis, complétée, le cas échéant, par un test à posteriori de
Dunn, proposé par Zar (1999) dans le cas d'échantillons d'effectifs différents. Les variations de l'indice
de réplétion stomacal selon la profondeur n'étant significatives pour aucune des cinq espèces étudiées,
seules les variations saisonnières de cet indice seront présentées, illustrées par des boites de Tukey (voir
ci-dessous).
Nombres et poids moyens globaux des proies ingérées
Pour chaque espèce, le nombre moyen (Np) et le poids moyen total (Pp) de proies par estomac
contenant de la nourriture ont été calculés et la distribution statistique des valeurs a été illustrée par des
boites de Tukey. Cette représentation graphique permet de visualiser les distributions des données
recueillies (homogénéité, valeur médiane, points exceptionnels) et ainsi de décrire les tendances pour
chaque échantillon. Elle présente en outre l'avantage d'être applicable à tous les types d'échantillons,
quel que soient leur effectif et la distribution (Normale ou non) de leurs données. Dans le cas présent,
seuls les points exceptionnels ("marginaux") ont été conservés pour la représentation graphique, les
points "extrêmes" étant éliminés car ne correspondant qu'à de très rares individus. Pour chaque facteur
étudié, la représentation fournie permet d'estimer la variabilité totale observée pour l'ensemble de la
population et indique la gamme de variation correspondant à la majorité des individus de l'espèce
(valeurs entourant la médiane et représentant 50% des individus analysés).
∗ Ce test n'a pas été réalisé pour S. impar chez qui l'indice de réplétion stomacal nocturne n'a pu être déterminé.

Stratégies alimentaires
64
Variations qualitatives de l'alimentation avec la taille des individus
Le regroupement des individus en classes de taille d'alimentation homogène a été réalisé grâce à
l'utilisation conjointe de méthodes d'ordination et de classification. Par leurs natures complémentaires,
ces deux types de méthodes constituent un outil très performant pour l'identification des groupes
structurels au sein d'un ensemble d'échantillons (Ludwig et Reynolds, 1988). Pour chaque espèce
étudiée, l'ordination et la classification des individus en fonction de leur alimentation ont permis de
déterminer les tailles correspondant à une modification significative du régime alimentaire et de répartir
les individus de chaque population en différents groupes de taille d'alimentation homogène. Dans un
souci de simplification et afin de limiter le nombre de zéros dans les matrices d'origine, les variables
rares (= proies "accidentelles" pour l'ensemble des classes de taille de chaque espèce) ont été exclues de
ces deux types d'analyses.
L'ordination des classes de tailles a été réalisée par N.M.S. ("Non metric Multidimentional
Scaling"). Cette technique vise à fournir la représentation visuelle la plus fiable possible d'un ensemble
complexe de relations entre observations afin qu'il puisse être apprécié d'un simple coup d'œil (Clarke,
1993). La méthode, largement employée en écologie depuis les années 60, surtout pour décrire les
relations au sein des assemblages d'espèces (e. g. Kenkel et Orloci, 1986 ; Ohman et Rajasuriya, 1998),
se situe parmi les méthodes d'ordination les plus performantes qui existent à l'heure actuelle (Warwick
et Clarke, 1991, 1993 ; Cao et al., 1996). Son utilisation a été récemment élargie à l'étude des
différences de composition de régimes alimentaires (e. g. Linke et al., 2001), domaine dans lequel elle
fournit de très bons résultats. Elle consiste en une recherche itérative de l'ordre et de la disposition de n
entités (observations) sur k dimensions (axes) qui minimise au maximum le stress de la configuration à
k dimensions. Les calculs sont basés sur une matrice de distances n × n, calculée à partir de la matrice
d'origine de n lignes (observations = groupes de poissons) et p colonnes (variables = catégories de
proies ingérées). La méthode préserve les distances réelles entre observations et présente l'avantage
d'être applicable à tous les types de données, même non-normales ou mesurées suivant des échelles
arbitraires ou encore discontinues (Clarke, 1993). Elle permet en outre le choix, en fonction de
l'hypothèse écologique ou biologique testée, de méthodes de standardisation/transformation des données
et de coefficients de similarité véritablement appropriés. Le "stress" associé à chaque représentation
graphique fournie par l’ordination (N.M.S.), permet d’évaluer la qualité de l'image proposée. Il mesure
l'écart à la monotonie existant entre les différences (distances) entre individus dans l'espace originel à p
dimensions et dans l'espace réduit à k dimensions. Il varie de 0, pour une représentation parfaite, à 1,
dans le cas inverse (Kruskal, 1964). La représentation graphique proposée par l'ordination peut être
considérée comme fiable dès lors que le stress est inférieur à 0,2 (Clarke, 1993). Elle est de bonne
qualité lorsqu'il est inférieur ou proche de 0,1 et excellente lorsqu'il est inférieur à 0,05. Dans le cas
présent, l'ordination des différentes classes de taille en fonction de leur alimentation a été réalisée à
partir des poids moyens des différentes catégories de proies par estomac (Pm, en mg de matière sèche).

Stratégies alimentaires
65
Afin de ne pas surcharger les graphes obtenus et de réduire la variabilité interindividuelle et le nombre
de zéros dans les matrices d'origine, les contenus stomacaux de plusieurs poissons ont été regroupés de
façon à obtenir, au maximum, 10 "individus moyens" (correspondant chacun à un groupe de 3 individus
minimum) par classe de taille. La distance de Bray-Curtis a été utilisée pour l'obtention de la matrice de
distances entre échantillons. Contrairement aux distances Euclidiennes ou du χ2, qui accordent une
importance maximale aux variables rares (et traduisent ainsi plus des changements qualitatifs, même
minimes, du régime alimentaire que de réelles modifications de l'abondance des proies consommées), ce
coefficient de similarité, peu sensible aux zéros, donne un même poids statistique aux variables (proies)
abondantes qu'aux variables rares (Legendre et Legendre, 1998 ; Legendre et Gallagher, 2001). Son
utilisation devrait assurer une description optimale des différences d'alimentation entre individus.
Suivant Clarke (1993), chaque N.M.S. a été répétée 7 à 8 fois afin de vérifier la stabilité de la
représentation graphique proposée. De même, seules les représentations pour lesquelles le stress associé
était inférieur à 0,15 ont été retenues. Réalisée dans ces conditions, l'utilisation de cette méthode
d'ordination, a permis de révéler les ressemblances trophiques entre classes de tailles et de décrire ainsi
les modalités de la modification ontogénique de l'alimentation chez chacune des cinq espèces.
La classification des différentes classes de tailles en fonction de leur alimentation, a été
effectuée par groupement agglomératif ("Cluster Analysis"). La méthode, dite "hiérarchique" et
"agglomérative", procède par regroupement progressif des individus (ici les classes de taille) en fonction
de leurs similarités (Greig-Smith, 1983) et fournit un arbre de classification des groupes indiquant les
différences (distances) entre eux. Elle peut être réalisée à l'aide de différents indices de similarité et
méthodes de regroupement, laissés au choix de l'utilisateur. Dans le cas présent, les classifications ont
été réalisées à partir du poids moyen (Pm, en mg) des différentes catégories de proies ingérées par
chaque classe de taille. Afin d'obtenir des résultats comparables à ceux de l'ordination par N.M.S., la
distance de Bray-Curtis a été utilisée pour le calcul des distances intergroupes. La construction de l'arbre
a été réalisée par la méthode du "Group Average" (UPGMA), couramment employée en complément de
la distance de Bray-Curtis. Pour chaque espèce, les groupes de tailles d'alimentation homogène ont été
établis par troncature de l'arbre de classification obtenu au seuil de 65% de l'information totale
représentée. Le choix arbitraire de ce seuil découle de l'étude des arbres obtenus pour les cinq espèces et
a été établi de façon à conjuguer au mieux précision de l'analyse et cohérence des groupes obtenus sur le
plan biologique.
Les résultats combinés de l'ordination et de la classification des classes de taille en fonction du
régime alimentaire ont permis de séparer, pour chaque espèce, plusieurs groupes d'alimentations
différentes. L'analyse des modifications du régime alimentaire d'un groupe à l'autre a ensuite permis de
déterminer les variations ontogéniques de l'alimentation.

Stratégies alimentaires
66
Variations quantitatives de l'alimentation avec la taille des individus
Les variations quantitatives de l'alimentation suivant la taille des individus ont été étudiées en
utilisant le poids total et le nombre de proies par estomac. Pour chacun de ces deux paramètres, la
normalité des données et l'homogénéité des variances ont été testées au préalable, respectivement par les
tests de Shapiro-Wilk et de Bartlett au seuil α = 0,05. Dans la mesure du possible, divers types de
transformation (X' = ln (X+1), X' = log (X+1), X' = √ (X+ 0,5), etc) ont été appliqués sur les données
brutes ne satisfaisant pas à ces deux critères, afin de normaliser leur distribution et d'obtenir
l'homoscédaticité nécessaire à l'utilisation des méthodes paramétriques de comparaison d'échantillons,
plus puissantes que leurs analogues non paramétriques (Zar, 1999). Chez S. impar, les différences
ontogéniques du nombre et du poids moyen des proies par estomac ont ainsi été testées par une
ANOVA à un facteur (stade de vie) réalisée après transformation X' = log (X+1). Un test de Newman-
Keuls a été réalisé a posteriori afin de déterminer les classes de taille concernées par les différences
observées. La normalité et l'homoscédaticité n'étant pas vérifiées pour les quatre autres espèces, les
différences ontogéniques du nombre et du poids moyen des proies par estomac ont été testées par une
ANOVA non paramétrique de Kruskal-Wallis. La réalisation a posteriori d'un test de Dunn a permis de
déterminer les groupes de tailles statistiquement différents. Le poids total des proies par estomac
augmentant régulièrement avec la taille pour l'ensemble des cinq poissons plats étudiés (observation
prévisible compte-tenu de l'élévation des besoins énergétiques au cours de la croissance), seuls les
résultats concernant le nombre total de proies par estomac seront présentés. Pour chaque espèce et
chaque groupe de taille, les distributions statistiques de ce facteur seront illustrées par des boites de
Tukey.
La relation entre la taille des poissons (LT, mm) et le poids moyen individuel de chaque proie
ingérée (Pm, mg) a également été étudiée. L'existence ou non d'une corrélation significative entre ces
deux facteurs a été déterminée. Pour cela, divers types de régressions (linéaire, exponentielle, puissance)
et différentes transformations des données (Y' = log (Y), Y' = log (Y+1), X' = √ (X+ 0,5), etc) ont été
testées. Dans chaque cas, la meilleure corrélation a été obtenue par régression linéaire entre les valeurs
brutes de LT et les valeurs log-transformées de Pm, le coefficient de corrélation (r) dépassant alors
toujours 0,94. Les corrélations linéaires entre données brutes, significatives pour les cinq espèces (P <
0,001), ont néanmoins été choisies pour figurer dans ce chapitre car elles permettent une estimation
rapide et visuelle du poids moyen des proies correspondant à chaque taille.
Variations spatio-temporelles de la composition du régime alimentaire
Les variations spatio-temporelles de la composition du régime alimentaire de chaque stade de
vie benthique ont été testées par des MANOVA non paramétriques à deux facteurs croisés fixes ("stade
de vie × saison", "stade de vie × profondeur" et/ou "profondeur × saison" pour chaque stade de vie)

Stratégies alimentaires
67
réalisées sur les poids moyens (en mg) des proies ingérées en fonction de la saison (S1= printemps, S2 =
été, S3 = hiver) et de la profondeur (P1 = 0-20 m, P2 = 30-50 m, P3 = 70-100 m). Des MANOVA non
paramétriques à un facteur (stade de vie) ont également été réalisées sur les poids moyens (en mg) des
différentes proies présentes dans les estomacs des stades de vie benthique capturés à une saison et/ou
une profondeur donnée(s). Le logiciel utilisé pour ces analyses (NP MANOVA) fournit une approche
alternative à l'analyse multivariée paramétrique en cas de non normalité des données (Mc Ardle et
Anderson, 2001). Basée sur le principe des tests de permutations, la méthode calcule la probabilité
d'obtenir une valeur égale ou supérieure à la valeur observée d'un test statistique sous une hypothèse
nulle donnée en recalculant la statistique du test après réarrangement (permutation) des observations
(Anderson, 2001)∗. Pour chaque analyse, les variables rares (= proies "accidentelles" pour l'ensemble
des stades de vie de chaque espèce) ont été exclues. Suivant le ou les facteur(s) testé(s) et l'espèce
étudiée, 10 à 19 individus (contenus stomacaux) ont été sous-échantillonnés par saison, profondeur ou
doublets "stade × saison" ou "stade × profondeur" afin de disposer d'un nombre équivalent
d'observations par case. Les analyses ont été réalisées après transformation (X' = log (X+1)) des
données, afin d'assurer au mieux l'homogénéité des variances entre les différents groupes testés
(Anderson, 2000). Compte tenu de la présence de nombreux zéros dans la matrice de données brutes et
afin de donner un même poids statistique aux espèces abondantes qu'aux espèces rares, la distance de
Bray-Curtis a été utilisée pour l'obtention de la matrice de distances entre échantillons. Comme conseillé
par Anderson (2000), les MANOVA à deux facteurs croisés ont été réalisées par permutation des
résidus sous modèle réduit et avec un nombre élevé (4999) de permutations afin d'assurer une puissance
du test et une précision de l'erreur α fournie maximales (Anderson et Legendre, 1999). Dans chaque cas,
les tests de comparaison a posteriori fournis par le programme ont été utilisés afin de préciser le résultat
obtenu par la MANOVA.
Résumé des variations de la composition du régime alimentaire au large du Rhône
Une ordination par N.M.S. (voir plus haut) a été utilisée afin d'estimer l'importance de la taille
des individus, de la saison et de la profondeur dans les modifications de l'alimentation de chaque espèce.
Comme pour l'ordination des classes de taille, les poids moyens par estomac des différentes catégories
de proies (à l'exception des proies "accidentelles") ont été utilisés, les contenus stomacaux de plusieurs
poissons ayant été regroupés de façon à obtenir, au maximum, 3 "individus moyens" (correspondant
chacun à un groupe de 3 individus minimum) par classe de taille présente à chaque saison et chaque
profondeur. La distance de Bray-Curtis a servi pour l'élaboration de la matrice de distances et chaque
N.M.S. a été répétée 7 à 8 fois. Seules les représentations pour lesquelles le stress associé était inférieur
à 0,15 ont été retenues dans chaque cas. Réalisée dans ces conditions, l'utilisation de cette méthode
∗ Cette méthode est décrite plus en détails au sein de la partie "matériel et méthodes" du Chapitre I de la Partie II.

Stratégies alimentaires
68
Indice de réplétionStomacal (IR)
d'ordination a permis d'identifier le(s) facteur(s) responsable(s) de la majorité des modifications de
l'alimentation des cinq espèces étudiées.
II/II.3 - ALIMENTATION DES CINQ ESPECES
II/II.3.1 - SOLEA SOLEA
II/II.3.1.1 - Alimentation globale
L'étude de l'alimentation de S. solea a été réalisée à partir des contenus stomacaux de 467
individus de longueur totale (LT) comprise entre 30 et 374 mm. La période de nutrition de la sole
commune est essentiellement nocturne, le remplissage stomacal étant significativement plus élevé
(Mann-Whitney : U(257,202) = 17336,5 ; P < 0,001) la nuit (IRmoy.= 1,9 ± 1,4∗) que le jour (IRmoy.= 1,0 ±
1,2) (Fig. II/II.1). La prise de nourriture s'intensifie à la tombée de la nuit et l'indice de réplétion
stomacal est maximal (IR = 2,8) entre 5:00 et 7:00 heures du matin. Au large du Rhône, S. solea
présente donc un seul pic d'alimentation, survenant juste avant l'aube, après une période d'alimentation
nocturne plus intense.
Figure II/II.1 - Variations nycthémérales de l'indice de réplétion stomacal moyen (IR) chez S.solea (juin 2000 ; heure locale = heure GMT +1). CS = coucher du soleil ; LS = lever du soleil.
Sur les 467 estomacs de S. solea analysés, 160 ne contenaient pas de nourriture (CV = 34,3%).
Le régime alimentaire global de l'espèce a donc été établi à partir des contenus stomacaux de 307 soles
∗ Les valeurs moyennes indiquées dans le texte sont suivies des erreurs-types correspondantes.

Stratégies alimentaires
69
de LT comprise entre 32 et 374 mm et comprenant 64% de juvéniles G0, 20% de juvéniles G1+ et 16%
de sub-adultes et adultes. Il se caractérise par l'ingestion d'un grand nombre de proies de tailles
différentes. Le nombre et le poids des proies par estomac fluctuent autour de valeurs médianes de 4
proies et 7,4 mg (Fig. II/II.2 a et b) mais sont très variables, ce qui résulte en des valeurs moyennes
élevées (Npmoy.= 20,6 ± 3,2 ind. est.-1; Ppmoy.= 29,4 ± 3,8 mg est.-1).
Figure II/II.2 - Représentation en Boites de Tukey des nombres (a) et des poids totaux (b) de proies par estomacchez S. solea au large du Rhône.
D'un point de vue qualitatif, S. solea présente un spectre alimentaire large (Tableau II/II.1). Son
régime alimentaire est cependant fortement dominé par les annélides polychètes qui forment la seule
catégorie de proies préférentielles. De nombreuses espèces de polychètes, appartenant à 20 familles
différentes, sont consommées avec une nette dominance numérique des dépositivores de surface
(Magelonidae, Spionidae) ou de sub-surface (Capitellidae, Sternaspidae). Les mollusques bivalves ont
une importance secondaire mais sont présents de façon régulière dans les estomacs. Cette catégorie de
proies englobe surtout des juvéniles consommés entiers (essentiellement des Tellinidae et des
Solenidae), mais également des fragments de siphons prélevés sur des bivalves adultes. L'importance
des autres catégories de proies est négligeable, celles-ci n'étant rencontrées dans les estomacs que de
façon occasionnelle voire accidentelle. On note également la présence, dans plus de 33% des estomacs
de l'espèce, de débris végétaux d'origine terrestre, souvent associés à des grains de sable ou des
particules fines de sédiment.
0
5
10
15
20
25
30
35
40
45
05
1015202530354045505560657075
(a) Np (b) Pp en mg
Non-marginaux max
Non-marginaux min 75% 25% Médiane Points marginaux

Stratégies alimentaires
70
Tableau II/II.1- Régime alimentaire global de S. solea au large du Rhône. F = fréquence ; N = nombre deproies ; Cn = pourcentage numérique ; P = poids sec des proies (mg) ; Cp = pourcentage pondéral ; Q =coefficient alimentaire (Q = Cn × Cp). "+" = valeur inférieure à 0,01.
F N Cn P Cp Q
Annélides :Oligochètes 0,01 6 0,10% 2,27 0,03% +Polychètes 0,77 5515 89,20% 5369,69 70,98% 6330,96
Siponcles + 1 0,02% 125,46 1,66% 0,03
Crustacés :Copépodes 0,01 3 0,05% 1,83 0,02% +Leptostracés 0,01 2 0,03% 7,73 0,10% +Mysidacés 0,01 4 0,06% 9,22 0,12% 0,01Cumacés 0,09 166 2,68% 20,52 0,27% 0,73Isopodes 0,01 2 0,03% 4,64 0,06% +Amphipodes 0,20 89 1,44% 44,85 0,59% 0,85Macroures 0,02 7 0,11% 204,26 2,70% 0,31Anomoures 0,03 8 0,13% 49,38 0,65% 0,08Brachyoures 0,04 24 0,39% 80,29 1,06% 0,41Crustacés indét. 0,02 5 0,08% 31,35 0,41% 0,03
Mollusques :Bivalves 0,57 287 4,64% 627,23 8,29% 38,48Gastéropodes 0,04 15 0,24% 274,32 3,63% 0,88Mollusque indét. 0,01 2 0,03% 9,44 0,12% +
Echinodermes :Ophiures 0,03 22 0,36% 202,42 2,68% 0,95Holothuries 0,02 8 0,13% 319,99 4,23% 0,55
Phoronidiens + 2 0,03% 2,21 0,03% +Poissons 0,03 9 0,15% 31,07 0,41% 0,06Proies indet. 0,02 6 0,10% 147,14 1,94% 0,19
Total proies 6183 100,0% 7565,31 100,0%
II/II.3.1.2 - Modifications du régime alimentaire
Variations en fonction de la taille
Chez S. solea, l'alimentation subit des modifications au cours de la croissance. Le changement
de régime alimentaire avec la taille des individus est progressif (Fig. II/II.3). La faible valeur de stress
obtenue pour la N.M.S. (0,06) garantit une représentation fidèle, dans le plan proposé, des distances
alimentaires réelles entre les points obtenus pour l'ensemble des classes de taille. Ceux-ci s'organisent
globalement suivant un gradient continu de taille croissante. Les poissons de 160-180 mm se détachent
cependant de façon significative du reste des individus, ce qui suggère une alimentation différente pour
cette classe de taille.

Stratégies alimentaires
71
Figure II/II.3 - Ordination (NMS, solution à 2 dimensions) des différentes classes de taille (LT, mm)de S. solea en fonction de leur régime alimentaire (Poids moyen des proies par estomac).
Figure II/II.4 - Classification par groupement agglomératif des classes de taille de S. solea en fonction de leurrégime alimentaire (poids moyens des proies par estomac). La flèche verticale rouge indique le pourcentaged'information (65 %) retenu comme valeur seuil pour l'établissement des groupes alimentaires (A, B, B' et C).
Stress : 0,055
-2
0
2
-2 0 2
Axe 1
Axe
2
< 60 mm
60-100 mm
100-140 mm
140-160 mm
160-180 mm
180-200 mm
200-220 mm
220-240 mm
240-260 mm
> 260 mm
Taille (LT, mm)
A
B
B'
C

Stratégies alimentaires
72
Non-Outlier MaxNon-Outlier Min75%25%MedianOutliers
Np
0
20
40
60
80
100
120
140
160
LT < 100 mm 100-200 mm LT > 200 mm
Non-marginaux maxNon-marginaux min
75%25%
MédianePoints marginaux
La classification par groupement agglomératif sépare quatre grands groupes d'alimentation : les
juvéniles G0 de moins de 100 mm de LT (groupe A), les juvéniles G0 de 100-200 mm (groupe B) -
avec, à part, les individus de 160-180 mm (groupe B') - et les juvéniles G1+ et adultes de plus de 200
mm (groupe C) (Fig. II/II.4). L'étude détaillée de l'alimentation de ces quatre groupes montre que le
positionnement alimentaire particulier des poissons de 160-180 mm (groupe B') résulte d'une ingestion
de gros gastéropodes par un nombre réduit d'individus de cette classe de taille (F = 0,09 ; Cn = 1,0% ;
Cp = 37,1%) (Annexe 4), les alimentations des groupes B et B' étant, par ailleurs, similaires. La
séparation entre ces deux groupes n'est qu'un artéfact lié à l'utilisation du poids moyen des proies par
estomac pour la classification. Les individus de S. solea se répartissent donc en trois groupes d'individus
d'alimentation homogène : les juvéniles G0 de moins de 100 mm de LT (groupe A), les juvéniles G0 de
100-200 mm (groupe B) et les juvéniles G1+ et adultes de plus de 200 mm (groupe C).
Les modifications ontogéniques de l'alimentation sont à la fois quantitatives et qualitatives.
L'augmentation significative de la ration alimentaire (poids total par estomac, Pp) avec la taille des
individus (Kruskall-Wallis : H(2,307) = 95,19 ; P < 0,001) s'accompagne d'une élévation du nombre de
proies par estomac, significative à partir de 200 mm de LT (P < 0,01 ; Annexe 5) (Fig. II/II.5). Les
nombres de proies par estomac oscillent principalement de 1 à 8 pour les juvéniles G0 des groupes A et
B (LT < 200 mm) et de 3 à 49 pour les juvéniles G1+ et adultes du groupe C (LT ≥ 200 mm), autour de
valeurs médianes respectives de 3, 3,5 et 11 ind. est.-1.
Figure II/II.5 - Représentation en Boites de Tukey des nombres de proies (Np, ind. est.-1) rencontrés dansles estomacs des individus de chaque groupe de taille (LT, mm) chez S. solea au large du Rhône.

Stratégies alimentaires
73
L'augmentation du nombre des proies ingérées est complétée par une modification de leur taille
et de leur nature. Le poids individuel moyen des proies consommées (Pm, mg) est positivement corrélé
(r = 0,629; r2 = 0,395; P < 0,001) à la longueur totale (LT, mm) des individus (Fig. II/II.6). Il passe de 1 à
4,9 mg entre les plus petits juvéniles et les adultes de l'espèce, soit une augmentation d'un facteur 5
environ. L'élargissement progressif de la gamme de poids des proies observée indique la persistance de
proies de petite taille dans l'alimentation des adultes. Néanmoins, chez ces derniers, la gamme de poids
moyen des proies couvre un intervalle de 4 à 11 mg alors qu'elle ne dépasse pas 3,8 mg chez les plus
petits juvéniles (LT < 100 mm).
Figure II/II.6 - Relation linéaire entre la taille (LT, mm) des individus de S. solea et le poids moyen (Pm,mg) des proies ingérées (r = 0,629; r2 = 0,395; P < 0,001).
La composition du régime alimentaire se modifie au cours de la croissance. Les proies ingérées
diffèrent d'un groupe de LT à l'autre (Tableau II/II.2). Chez les plus jeunes soles (LT < 100 mm), les
cumacés constituent, avec les polychètes, l'essentiel de l'alimentation. Le fort pourcentage numérique
(Cn = 77,7%) des premiers et le poids élevé des seconds (Cp = 34,5%) en font deux catégories de proies
préférentielles consommées par plus de 50% des individus (F = 0,58 et F = 0,52, respectivement). Les
bivalves (F = 0,39) et les amphipodes (F = 0,12) sont des proies secondaires, le régime alimentaire étant
principalement complété de petits mysidacés, de phoronidiens et d'ophiures. A partir de 100 mm de LT,
l'alimentation est largement dominée par les polychètes qui forment la seule catégorie de proies
préférentielles. Ces derniers sont présents dans les estomacs de plus de 75% des individus des groupes B
et C et constituent plus de 66% en poids et plus de 91% en nombre des proies ingérées. Les différences
de composition du régime alimentaire entre les juvéniles G0 de 100-200 mm (groupe B) et les juvéniles
0
2
4
6
8
10
12
14
0 20 40 60 80 100 120 140 160 180 200 220 240 260 280 300 320 340 360 380 LT(mm)
Pm (mg)
Pm = 0,0182 × LT – 0,8353

Stratégies alimentaires
74
G1+ et adultes de plus de 200 mm (groupe C) sont essentiellement liées à une diminution de
l'importance pondérale des bivalves dans l'alimentation. Ceux-ci constituent un groupe de proies
d'importance secondaire (F = 0,65 ; Cp = 18,0%) pour les individus de 100-200 mm, et une catégorie de
proies occasionnelle (F = 0,47 ; Cp = 5,4%) pour les soles de LT > 200 mm.
Tableau II/II.2 - Variations de l'alimentation de S. solea en fonction de la taille. Q = coefficient alimentaire."+" = valeur inférieure à 0,01 ; "" = valeur absente. LT = longueur totale des individus (en mm).
Juvéniles G0LT < 100 mm
Juvéniles G0100 < LT < 200 mm
Juvéniles G1+ et adultesLT > 200 mm
Nb estomacs pleins 33 177 97
LT moyenne (min.-max.) 58,9 (32-92) 148,5 (105-199) 259,8 (200-374)
PROIES Q Q Q
AnnélidesOligochètes +Polychètes 489,9 5995,9 7154,0
Siponcles +
CrustacésCopépodes +Leptostracés + Mysidacés 1,6 + +Cumacés 1374,4 0,1 +Isopodes + +Amphipodes 22,2 3,9 0,1Macroures 0,1 0,4Anomoures + +Brachyoures + 0,8Crustacés indét. + +
MollusquesBivalves 130,9 102,1 12,6Gastéropodes 1,7 0,6Mollusque indét. +
EchinodermesOphiures 0,4 0,3 1,2Holothuries 1,3
Phoronidiens +
Poissons + +
Proies indét. + + +

Stratégies alimentaires
75
0%
25%
50%
75%
100%
< 100 100-200 > 200Classesde taille
Cp
polychètes oligochètes bivalves gastéropodes amphipodes isopodes
macroures mysidacés brachyoures anomoures leptostracés cumacés
copépodes ophiures poissons siponcles holothuries phoronidiens
< 100 mm 100-200 mm > 200 mm
La largeur du spectre alimentaire augmente légèrement avec la taille des individus. Cet
élargissement correspond à l'ingestion de proies de taille de plus en plus importante. Les gastéropodes,
les macroures, les anomoures et les brachyoures ne sont consommés que par les individus de LT > 100
mm, et les siponcles et les holothuries que par ceux de plus de 200 mm. Cependant, ces proies restent
occasionelles et l'essentiel des modifications ontogéniques du régime alimentaire concerne l'importance
pondérale des polychètes dans l'alimentation, qui s'accroît avec la taille des poissons (Fig. II/II.7). Le
spectre des familles de polychètes consommées, très étroit chez les juvéniles G0 de LT < 100 mm (deux
familles), s'élargit chez les G0 de 100-200 mm et les juvéniles G1+ et adultes de plus de 200 mm
(respectivement 18 et 17 familles). Les polychètes ingérés sont en majorité des dépositivores :
dépositivores de surface chez les plus petits G0 (Paraonidae et Spionidae), dépositivores de surface
(60% en poids) et de sub-surface (27% en poids) chez les G0 de 100-200 mm et dépositivores de sub-
surface (Capitellidae et Sternaspidae) chez les juvéniles G1+ et les adultes.
Figure II/II.7 - Variations de la composition pondérale (Cp) du régime alimentaire de Solea solea au largedu Rhône en fonction de la taille des individus (LT, mm).
Variations spatio-temporelles
Les variations spatio-temporelles de l'alimentation ont été étudiées pour chacun des groupes
alimentaires mis en évidence pour S. solea au large du Rhône. Cependant, les fluctuations spatio-
temporelles de la distribution des individus de l'espèce empêchant une analyse statistique conjointe des
effets de la profondeur et de la saison sur la composition du régime alimentaire, les variations
saisonnières et bathymétriques de l'alimentation ont été traitées séparément.

Stratégies alimentaires
76
Printemps
Eté
Hiver
Juvéniles G0 L
T < 100 mm
Juvéniles G0 100-200 m
mJuvéniles G
1+ et adultes L
T > 200 mm
(N = 10)
polychète86,0%
poisson3,0%
mysidacé
1,9%
amphipode0,8%
ophiure2,2%
cumacé
0,3%anom
oure0,8%
bivalve4,9%
(N = 156)
polychète62,5%
bivalve19,5%
gastéropode9,6%
indét.1,6%
poisson0,8%
ophiure1,2%
cumacé
0,4%
leptostracé 0,5%
anomoure
0,3%brachyoure
0,4%
amphipode1,8%
macroure1,1%
(N = 12)
holothurie8,9%
siponcle10,74%
ophiure12,0%
macroure
3,5%bivalve5,3%
amphipode
0,3%
brachyoure0,2%
anomoure
0,3%polychète58,7%
(N = 19)
polychète55,4%
bivalve14,9%
gastéropode9,8%
indét.10,6%
holothurie2,1%
ophiure3,1%
cumacé
0,3%
brachyoure3,0%
macroure0,6%
(N = 62)
bivalve2,1%
polychète88,6%
holothurie5,8%
poisson0,4%
brachyoure0,8%
cumacé
0,1%
anomoure
1,3%
amphipode0,1%
gastéropode0,5%
(N = 11)
polychète 57,0%
bivalve9,9%
amphipode17,9%
isopode1,3%
ophiure13,9%
(N = 33)
13,1%
mysidacé
6,7%
amphipode9,2%
bivalve32,1%
polychète31,8%
crust. indét.1,3%
ophiure1,8%
phoronidien4,0%
cumacé
Figure II/II.9 - Variations de l'alim
entation (pourcentages pondéraux des proies ingérées) de S. solea en fonction de la saison et de la taille des individus (longueur totale LT ,
en mm
). N = nom
bre d'estomacs pleins analysé pour chaque groupe de poissons. D
ans chaque cas, les proies surlignées en gris sont rencontrées dans plus de 20% des
estomacs et/ou représentent plus de 10%
du poids total des proies ingérées.

Stratégies alimentaires
77
Non-Outlier MaxNon-Outlier Min75%25%Median
IR
-1
0
1
2
3
4
5
printemps été hiver
CV = 25,9% CV = 18,8% CV = 42,5%
Non-marginaux max Non-marginaux min 75% 25% Médiane
Variations saisonnières
L'intensité de la nutrition de S. solea varie au cours de l'année. L'indice de réplétion stomacal
diffère suivant la saison (Krukall-Wallis : H (2, N=432) = 7,72 ; P < 0,05) et est significativement plus élevé
en été (IRmoy.= 1,8 ± 0,1) qu'au printemps (IRmoy.= 1,7 ± 0,1) et en hiver (IRmoy.= 1,3 ± 0,2) (Annexe 6).
En hiver, l'activité nutritionnelle est fortement réduite avec un coefficient de vacuité élevé et un indice
de réplétion stomacal généralement faible (Fig. II/II.8). L'intensité de nutrition augmente au printemps
et est maximale en été, où le nombre d'estomacs vides est faible.
Figure II/II.8 - Représentation en Boites de Tukey des indices de réplétion stomacale (IR) observés auxtrois saisons étudiées chez S. solea au large du Rhône. CV = coefficient de vacuité (% d'estomacs vides).
La nature des proies ingérées varie également selon la saison considérée. Cependant, les
modifications observées dépendent de la taille des individus (Fig. II/II.9). Les plus petits juvéniles (G0 :
LT < 100 mm) n'ayant été capturés qu'au printemps, les variations saisonnières de l'alimentation mises
en évidence ne concernent que les soles de plus de 100 mm. Pour ces dernières, mis à part pour les
individus de 100-200 mm au printemps, le régime alimentaire reste globalement le même quels que
soient le groupe de taille et la saison (Annexe 7), les modifications observées ne concernant que des
catégories de proies occasionnelles ou accidentelles.
Au printemps, période où toutes les classes de taille sont représentées dans les captures, les
régimes alimentaires des trois groupes de taille sont statistiquement différents (Annexe 8). Les plus
petits juvéniles (G0 : LT < 100 mm) ont le régime le plus diversifié, avec une codominance pondérale de
polychètes, de bivalves et de crustacés (cumacés, amphipodes et mysidacés). L'alimentation de

Stratégies alimentaires
78
0 - 20 m
30 - 50 m
70 - 100 m
Juvéniles G0 L
T < 100 mm
Juvéniles G0 100-200 m
mJuvéniles G
1+ et adultes L
T > 200 mm
(N = 33)
phoronidien 4,0%
ophiure 1,8%
crust. indét.1,3%
polychète31,8%
bivalve32,1%
amphipode9,2%
mysidacé6,7% cum
acé13,1%
(N = 13)
anomoure
0,6%
polychète 82,9%
cumacé
0,3%m
ysidacé1,6%
ophiure0,6%poisson
1,2%
amphipode0,7%
bivalve12,2%
(N = 61)
polychète 88,3%
holothurie6,2%
poisson 0,2%
brachyoure1,6%
anomoure
0,9%m
ysidacé0,1%
amphipode0,1%
macroure1,2%oligochète
0,1%
bivalve1,2%
(N = 13)
polychète73,5%
holothurie9,2%
siponcle5,1%
poisson0,1%
bivalve5,0%
brachyoure0,2%
macroure6,0%
gastéropode0,8%
(N = 18)
brachyoure0,7%
anomoure
1,3%
amphipode0,5%
mol. indét.0,5%
poisson0,4%
polychète37,8%
bivalve23,6%
ophiure20,0%
gastéropode13,9%
macroure0,8% cum
acé0,5%
(N = 164)
macroure
1,1% anomoure
0,3%
amphipode2,0%
brachyoure0,4%
cumacé
0,5%
crust. indét.2,1%
ophiure1,5%
poisson 1,0%
indét.1,5%
gastéropode9,5% bivalve
18,6%polychète
60,9%
Figure II/II.10 - Variations de l'alim
entation (pourcentages pondéraux des proies ingérées) de S. solea en fonction de la profondeur et de la taille des individus (longueurtotale L
T , en mm
). N = nom
bre d'estomacs pleins analysé pour chaque groupe de poissons. D
ans chaque cas, les proies surlignées en gris sont rencontrées dans plus de20%
des estomacs et/ou représentent plus de 10%
du poids total des proies ingérées.

Stratégies alimentaires
79
l'ensemble des individus de plus de 100 mm de LT est dominée par les polychètes mais les juvéniles G0
de 100-200 mm ingèrent une quantité non négligeable d'amphipodes (F = 0,45), de bivalves (F = 0,32)
et d'ophiures (F = 0,24), alors que, chez les juvéniles G1+ et adultes de plus de 200 mm, ces proies ne
sont consommées que de façon occasionnelle (F < 0,10 ; Cn < 3,0%).
En été et en hiver, périodes où seuls les poissons de plus de 100 mm sont présents, la composition
saisonnière de l'alimentation ne diffère pas, quantitativement parlant, d'un groupe de LT à l'autre
(Annexe 7). Le régime alimentaire comprend, aux deux saisons, une nette majorité de polychètes,
auxquels s'ajoutent, en été, quelques siphons de bivalves ingérés par les individus des deux groupes (F <
0,12). Le reste des proies est consommé de façon accidentelle, soit par un seul des deux groupes
(mysidacés chez les individus de 100-200 mm; holothuries chez ceux de plus de 200 mm) soit par les
deux (anomoures, brachyoures, amphipodes, etc), à une seule saison (gastéropodes, macroures et
leptostracés en été; mysidacés en hiver) ou aux deux (holothurie, brachyoures, ophiures, anomoures,
poissons, etc).
Variations en fonction de la profondeur
L'intensité de la nutrition de S. solea ne varie pas de façon significative d'une tranche
bathymétrique à l'autre (Kruskall-Wallis : H(2, N = 326) = 5,20 ; P > 0,07). Par contre, la composition du
régime alimentaire subit d'importantes modifications selon la profondeur. Les plus petits juvéniles de S.
solea n'étant présents qu'à 0-20 m de profondeur, ces variations ne concernent que les individus de taille
supérieure à 100 mm (Fig. II/II.10). Pour ces derniers, le régime alimentaire diffère significativement (P
< 0,05 ; Annexe 9 et 10) entre les fonds sableux côtiers (0-20 m) et les vases terrigènes plus profondes
(30-100 m), avec une alimentation nettement plus diversifiée à 0-20 m. L'alimentation des soles de plus
de 200 mm, seules présentes jusqu'à 100 m de profondeur, ne subit par contre pas de modification
significative entre 30-50 m et 70-100 m de profondeur (Annexe 10) où le régime alimentaire reste
largement dominé par les polychètes (Cp >73%).
La nature des proies ingérées à chaque profondeur varie selon la taille des individus. A 0-20 m, seule
profondeur où toutes les classes de taille de S. solea sont présentes, le régime alimentaire diffère de
façon significative suivant la taille des poissons (Annexe 11). Les juvéniles G0 de moins de 100 mm
présentent un régime plus diversifié (P < 0,01) que les autres soles, avec une importance accrue des
crustacés (F = 0,72) et des bivalves (F = 0,52), aux dépends des polychètes (F = 0,58) qui ne
représentent que 32% en poids des proies ingérées (Fig. II/II.10). A cette profondeur, la composition des
régimes alimentaires des juvéniles G0 de 100-200 mm ne diffère pas de façon significative de celle des
juvéniles G1+ et adultes de plus de 200 mm (Annexe 11). L'alimentation de ces deux groupes est
dominée par les polychètes (F > 0,68), mais comprend également une quantité non négligeable de
bivalves (F > 0,22), de gastéropodes (F > 0,15), et d'ophiures (F > 0,09).

Stratégies alimentaires
80
Sur les fonds de 30-50 m de profondeur, les polychètes sont les seules proies préférentielles de
S. solea (Q > 7500) mais la composition du régime alimentaire varie de façon significative suivant la
taille des individus (P < 0,05; Annexe 12), du fait de différences importantes au niveau des catégories
de proies occasionelles ingérées (bivalves chez les G0 de 100-200 mm ; holothuries et brachyoures chez
les juvéniles G1+ et adultes de LT > 200 mm). Sur les fonds de 70-100 m de profondeur, où seul les
individus de plus de 200 mm ont été capturés, le régime alimentaire est clairement dominé par les
polychètes (F = 0,87) auxquels s'ajoutent des bivalves (F = 0,20), des macroures (F = 0,12) et des
holothuries (F = 0,08), ingérés de façon occasionnelle.
A ces modifications générales du régime alimentaire avec la profondeur s'ajoutent des
différences de nature des proies ingérées beaucoup plus fines. Ainsi, les espèces de polychètes les plus
consommées ne sont pas les mêmes aux trois profondeurs (dépositivores de surface à 0-20 m ;
dépositivores de sub-surface à 30-50 m ; carnivores, dépositivores de surface et dépositivores de sub-
surface à 70-100 m) (Annexe 13). La nature des bivalves ingérés par les soles de plus de 200 mm de LT
diffère également d'une profondeur à l'autre (siphons d'adultes à 0-20 m ; juvéniles à 30-100 m). Il en
est de même pour d'autres catégories de proies, notamment les macroures, avec essentiellement des
Crangonidae à 0-20 m et des Alpheidae à 30-100 m de profondeur.
II/II.3.1.3 - Discussion sur l'alimentation de S. solea
Comportement alimentaire
Au large du Rhône, la sole commune présente une alimentation essentiellement nocturne, avec
un pic unique de nutrition maximale en fin de nuit, juste avant le lever du soleil. Ces observations sont
en accord avec les rythmes d'alimentation habituellement décrits pour cette espèce, S. solea s'alimentant
essentiellement de nuit (Krüuk, 1963 ; De Groot, 1969 ; Lagardère, 1987 ; Darnaude et al., 2001).
L'alimentation de la population étudiée se caractérise par un poids total de proies par estomac
élevé associé à un nombre variable de proies de tailles différentes. Ce phénomène reflète l'ingestion
alternative, chez S. solea, soit d'un nombre élevé de proies de petite taille, soit d'un nombre réduit de
proies volumineuses. Ces données sont conformes à celles de la littérature. L'espèce est décrite comme
ayant des besoins nutritionnels élevés, tant au stade juvénile (Lasserre et Lasserre, 1979 ; Lagardère,
1987) qu'au stade adulte (Horwood, 1993). Elle s'alimente suivant le principe du "peu mais souvent", à
partir de proies peu nombreuses et de taille réduite (De Groot, 1971). Cependant, la taille de sa bouche
lui permet, à l'âge adulte, une ingestion occasionnelle de proies relativement volumineuses et elle
consomme de façon opportuniste un nombre élevé de proies de petite taille (Molinero et Flos, 1992a ;
Darnaude, 1999). Le régime alimentaire global obtenu pour S. solea au large du Rhône est surtout

Stratégies alimentaires
81
représentatif de l'alimentation des juvéniles G0 (LT < 200 mm), ceux-ci constituant la majorité des
effectifs récoltés. Il est essentiellement composé de polychètes dépositivores et, à une moindre échelle,
de mollusques bivalves. Il correspond à l'alimentation observée pour l'espèce sur l'ensemble de son aire
de répartition géographique (Todd, 1907 ; Steven, 1930 ; Sorbe, 1972 ; Mendez de Elguezabal, 1978 ;
Rogers et Jinadasa, 1989 ; Molinero et al., 1991 ; Horwood, 1993). Solea solea est décrite comme se
nourrissant essentiellement de polychètes, auxquels s'ajoutent des quantités variables de bivalves, de
crustacés (amphipodes et macroures) et d'échinodermes (ophiures et holothuries). Cependant,
l'importance des bivalves dans le régime alimentaire de l'espèce au large du Rhône (second rang après
les polychètes), déjà soulignée par plusieurs auteurs en Méditerranée (Reys, 1960 ; Mendez de
Elguezabal, 1978 ; Darnaude, 1999 ; Darnaude et al., 2001), ne correspond, en Mer du Nord, qu'aux
données de De Groot (1971), cette catégorie de proie ayant généralement une importance plus faible que
les crustacés. La présente étude confirme donc la forte adaptabilité trophique de S. solea, dont
l'alimentation varie de façon importante suivant la région et la nature des proies disponibles dans le
milieu (Molinero et Flos, 1992b ). Elle peut ainsi être dominée tantôt par les polychètes (Todd, 1907 ;
Steven, 1930 ; Braber et De Groot, 1973 ; Rogers et Jinadasa, 1989 ; Beyst et al., 1999), tantôt par les
crustacés (Sorbe, 1972 ; Molinero et al., 1991 ; Molinero et Flos, 1992a, 1992b).
Modifications de l'alimentation
La prise de nourriture varie suivant la saison : elle est réduite en hiver et maximale en été. Cette
observation s'explique par une forte réduction de l'alimentation des adultes pendant la période de
reproduction (Horwood, 1993), située en fin d'hiver en Méditerranée nord-occidentale (Shéhata, 1984).
Elle résulte également des forts écarts de température existant entre l'été et l'hiver à 0-20 m (Younes,
2000), profondeur où se répartissent les juvéniles de S. solea. En effet, la température a une influence
marquée sur le métabolisme de l'espèce (Fonds, 1975 ; Fonds et Saksena, 1977 ; Lagardère, 1989) qui
présente une pause de croissance et une réduction de son alimentation en hiver, compensée par une forte
croissance et une alimentation intense en été (Woodhead, 1964 ; Fonds, 1975 ; Fonds et Saksena, 1977 ;
Fonds, 1979 ; Darnaude, 1999 ; Darnaude et al., 2001).
La composition du régime alimentaire de S. solea au large du Rhône varie selon la saison mais
surtout suivant la taille des individus et la profondeur (Fig. II/II.11). Le régime alimentaire des juvéniles
G0 de moins de 100 mm s'individualise totalement de ceux des individus de 100-200 mm et de plus de
200 mm qui se chevauchent en partie. Pour ces deux derniers groupes, l'alimentation est cependant
distincte à chaque doublet saison × profondeur, mis à part à 0-20 m en été. Bien que les polychètes
constituent l'essentiel de l'alimentation quel que soit le stade de vie benthique, le poids et la nature des
proies ingérées diffèrent entre juvéniles G0 et adultes de l'espèce. Le régime alimentaire des premiers,
plus diversifié, est essentiellement composé de proies de taille réduite (cumacés, petits polychètes

Stratégies alimentaires
82
P
P
P
P
EE E
H
PP
EE
H
H HE
H
H
P
E
-2
0
2
-2 0 2
Axe 1
Axe
2
Stress = 0,040
< 100 mm
100- 200 mm
> 200 mm
70 - 100 m
30 - 50 m
0 - 20 m
Profondeur
juvéniles G0L T < 100 mm
juvéniles G1+ et adultes(LT > 200 mm)
juvénilesG0100-200 mm
Stade de vie
dépositivores de surface, siphons de bivalves, amphipodes et juvéniles d'ophiures). Celui des seconds
est largement dominé par les polychètes dépositivores de sub-surface auxquels s'ajoutent, suivant la
saison, diverses proies de taille importante (holothuries, macroures, gastéropodes). Ces observations
correspondent au schéma de modification ontogénique de l'alimentation observé pour S. solea sur
l'ensemble de son aire de répartition. Au cours de la croissance, la taille des proies augmente, avec un
remplacement progressif des petites proies par de plus grandes (Lagardère, 1987 ; Molinero et Flos,
1992a ; Darnaude, 1999). Le régime alimentaire des plus petits individus comprend une part importante
de petits crustacés (Amara et al., 2001) et de mollusques (Lagardère, 1987) dont l'importance diminue
avec la croissance au profit des polychètes et de proies volumineuses comme les gastéropodes, les
macroures, les anomoures et les brachyoures (Mendez de Elguezabal, 1978 ; Molinero et Flos, 1992a ;
Darnaude, 1999), ce qui correspond à nos observations.
Figure II/II.11 - Variations de l'alimentation de S. solea en fonction de la profondeur (0-20 m, 30-50m, 70-100 m), de la saison (P = printemps, E = été, H = hiver), et de la taille (LT, mm) des individus.Ordination (N.M.S., solution à 2 dimensions) réalisée sur les poids moyens des proies par estomac.

Stratégies alimentaires
83
La composition du régime alimentaire de S. solea varie peu suivant la saison et la profondeur.
La variabilité interindividuelle de l'alimentation augmente cependant avec la taille des individus,
essentiellement du fait d'une répartition bathymétrique plus large (Fig. II/II.11). Le régime alimentaire
est très homogène chez les juvéniles G0 de moins de 100 mm, présents uniquement à 0-20 m au
printemps. Sa variabilité augmente chez les G0 de 100-200 mm, présents de 0 à 50 m et est maximale
chez les juvéniles G1+ et adultes de plus de 200 mm qui fréquentent l'ensemble des tranches
bathymétriques prospectées (0-100 m de profondeur). Quel que soit le stade de vie benthique, le régime
alimentaire est plus diversifié sur les fonds sableux côtiers (polychètes, mollusques) que sur les vases
terrigènes plus profondes (polychètes) et reflète les modifications bathymétriques des peuplements
d'invertébrés benthiques (Massé, 1972a ; Salen-Picard et al., 1997), ce qui confirme la forte adaptabilité
trophique de S. solea (Molinero et Flos, 1992b). Les modifications saisonnières consistent en une
diversification de l'alimentation au printemps et en été avec une ingestion secondaire de bivalves,
d'amphipodes, de gastéropodes et/ou d'ophiures. Les polychètes sont cependant consommés de façon
préférentielle par les juvéniles et les adultes de l'espèce, quelles que soient la saison et/ou la profondeur
considérées et constituent, quantitativement parlant, l'essentiel des proies ingérées par S. solea au large
du Rhône.

Stratégies alimentaires
84
II/II.3.2 - SOLEA IMPAR
II/II.3.2.1 - Alimentation globale
L'alimentation de S. impar au large du Rhône a été étudiée à partir des contenus stomacaux de
143 individus de LT comprise entre 49 et 212 mm. Un seul de ces individus ayant été capturé de nuit, la
période de nutrition maximale de l'espèce n'a pu être déterminée. La sole adriatique semble néanmoins
avoir une alimentation essentiellement diurne (Fig. II/II.12). Le taux de réplétition des estomacs est
élevé pendant le jour (IRmoy.= 2,3 ± 0,1), avec un pic d'alimentation vers midi où l'indice de réplétition
moyen est maximal (IR = 3) et les estomacs pleins à plus de 75% de leur volume.
Figure II/II.12 - Variations nycthémérales de l'indice de réplétion stomacal moyen (IR) chez S.impar (juin 2000; heure locale = heure GMT + 1). CS = coucher du soleil; LS = lever du soleil.
Sur les 143 estomacs de S. impar analysés, 42 ne contenaient pas de nourriture (CV = 29,4%).
Le régime alimentaire global de l'espèce a donc été établi à partir des contenus stomacaux de 101
individus de LT comprise entre 33 et 212 mm, comprenant 47% de juvéniles G0, 32% de juvéniles G1+
et 21% d'adultes. Il est caractérisé par l'ingestion d'un nombre élevé de proies de tailles différentes. Le
nombre et le poids des proies par estomac sont très variables et fluctuent respectivement de 1 à 185 ind.
est.-1*et de 0,3 à 76,0 mg est –1 ∗, autour de valeurs médianes de 11,5 proies et 6,9 mg par estomac (Fig.
II/II.13 a et b). Cette forte variabilité explique les moyennes élevées obtenues pour l'ensemble des
individus étudiés (Npmoy.= 21,4 ± 2,9 ind. est.-1 et Ppmoy.= 10,0 ± 1,3 mg est -1) bien que la majorité
d'entre eux n'ingère que de 5 à 26 proies pour un poids par estomac de 2,9 à 11,6 mg.
∗ valeurs extrêmes ne concernant que quelques individus et donc non représentées sur la Figure II/II.13

Stratégies alimentaires
85
0
51015202530354045505560657075808590
0
5
10
15
20
25
30
35
Figure II/II.13 - Représentation en Boites de Tukey des nombres (a) et des poids totaux (b) des proies par estomacchez S. impar au large du Rhône.
Tableau II/II.3 - Régime alimentaire global de S. impar au large du Rhône. F = fréquence ; N = nombre deproies ; Cn = pourcentage numérique ; P = poids sec des proies (mg) ; Cp = pourcentage pondéral ; Q =coefficient alimentaire des proies (Q = Cn × Cp).
F N Cn P Cp Q
AnnélidesPolychètes 0,55 239 12,67% 144,03 16,27% 206,14
CrustacésCopépodes 0,11 16 0,85% 13,35 1,51% 1,28Mysidacés 0,17 37 1,96% 58,95 6,67% 13,08Cumacés 0,58 835 44,27% 77,71 8,79% 389,06Amphipodes 0,55 579 30,70% 92,53 10,46% 321,23Macroures (crevettes) 0,01 1 0,05% 3,26 0,37% 0,02Crustacés indét. 0,01 1 0,05% 3,87 0,44% 0,02
MollusquesBivalves 0,49 168 8,91% 468,30 52,96% 471,72Gastéropodes 0,01 1 0,05% 6,33 0,72% 0,04
EchinodermesOphiures 0,05 9 0,48% 15,98 1,81% 0,86
Total proies 1886 100,0% 884,31 100,0%
Non marginaux max
Non marginaux min
75%25%
Médiane
Points marginaux
(a) Np (b) Pp en mg

Stratégies alimentaires
86
Le spectre alimentaire de S. impar est relativement étroit (Tableau II/II.3). Trois phylums sont
consommés en abondance : les mollusques, les crustacés et les polychètes. Du fait de leur fort
pourcentage pondéral, les bivalves se rangent au premier rang des proies préférentielles. Les cumacés et
les amphipodes, ingérés en grandes quantités et présents dans plus de 50% des estomacs analysés,
complètent cette catégorie. Les polychètes, essentiellement de petits dépositivores de surface
(Spionidae) et quelques carnivores de grande taille (Glycéridae, Nephthydae), sont consommés de façon
régulière et se situent à la limite des proies préférentielles. Les autres catégories de proies consommées
sont occasionnelles (mysidacés) ou accidentelles (copépodes, ophiures, macroures et gastéropodes). Des
débris de végétaux terrestre ont également été trouvés dans 10% des estomacs analysés.
II/II.3.2.2 - Modifications du régime alimentaire
Variations en fonction de la taille
Chez S. impar, les variations interindividuelles de l'alimentation, relativement réduites chez les
juvéniles G0 (LT < 100 mm) et les adultes (LT > 160 mm), augmentent pour les classes de taille
intermédiaires (100-160 mm), avec un maximum entre 140 et 160 mm de LT (Fig. II/II.14). La faible
valeur de stress (0,088) obtenue pour la N.M.S. garantit une représentation fidèle dans le plan des
distances réelles entre individus, distances qui augmentent de façon très nette entre individus des classes
de 100 à 160 mm de LT. La composition du régime alimentaire se modifie cependant de façon
progressive avec la taille des individus, les points obtenus pour l'ensemble des classes de taille
s'organisant suivant un gradient continu de taille croissante. La classification par groupement
agglomératif sépare les individus de S. impar en quatre grands groupes alimentaires : les juvéniles G0
de moins de 100 mm de LT (groupe A), les juvéniles G1+ de 100-140 mm (groupe B), ceux de 140-160
mm (groupe C) et les adultes de plus de 160 mm (groupe D) (Fig. II/II.15).
L'alimentation varie de façon importante suivant la taille des individus, tant sur le plan
quantitatif que qualitatif. L'augmentation significative de la ration alimentaire (poids total par estomac,
Pp) au cours de la croissance (ANOVA : F(3, 97) = 25,78 ; P < 0,001) s'accompagne d'importantes
modifications du nombre de proies par estomac (ANOVA : F(3, 97) = 4,22 ; P < 0,01). Ce dernier oscille
principalement entre 4 et 20 ind est-1 pour les juvéniles G0 (LT < 100 mm) et les adultes (LT > 160 mm)
et entre 6 et 54 ind est-1 pour les juvéniles G1+ des groupes B (100-140 mm) et C (140-160 mm), autour
de valeurs médianes respectives de 11, 8, 31 et 24 ind. est-1. Le nombre de proies par estomac et sa
variabilité augmentent donc de façon considérable chez les juvéniles de 100-160 mm de LT pour
reprendre ensuite, chez les adultes, des valeurs comparables à celles des plus petits juvéniles (Fig.
II/II.16). Cependant, du fait de la très forte variabilité interindividuelle du groupe B (juvéniles G1+ de
100-140 mm), l'accroissement du nombre de proies par estomac n'est significatif (P < 0,05 ; Annexe 14)
que pour le groupe C (juvéniles G1+ de 140-160 mm).

Stratégies alimentaires
87
Figure II/II.14 - Ordination (NMS, solution à 2 dimensions) des différentes classes de taille (LT, mm)de S. impar en fonction de leur régime alimentaire (Poids moyen des proies par estomac).
Figure II/II.5 - Classification par groupement agglomératif des classes de taille de S. impar en fonction deleur régime alimentaire (poids moyens des proies par estomac). La flèche verticale rouge indique lepourcentage d'information (65%) retenu comme valeur seuil pour l'établissement des groupes alimentaires(A,B,C et D).
Stress : 0,088-2
0
2
-2 0 2
Axe 1
Axe
2
< 80 mm
80-100 mm
100-120 mm
120-140 mm
140-160 mm
160-180 mm
> 180 mm
Taille (LT, mm)
A
B
C
D

Stratégies alimentaires
88
Non-Outlier MaxNon-Outlier Min75%25%MedianOutliers
Np
0
20
40
60
80
100
120
140
160
180
200
LT < 100 mm 100-140 mm 140-160 mm LT > 160 mm
Non-marginaux maxNon-marginaux min
75%25%
MédianePoints marginaux
Figure II/II.16 - Représentation en Boites de Tukey des nombres de proies (Np, ind. est.-1) rencontrés dansles estomacs des individus de chaque groupe de taille (LT, mm) chez S. impar au large du Rhône.
Figure II/II.17 - Relation linéaire entre la taille (LT, mm) des individus de S. impar et le poids moyen (Pm,mg) des proies ingérées (r = 0,602; r2 = 0,362; P < 0,001).
0
1
2
3
4
5
6
7
8
9
0 20 40 60 80 100 120 140 160 180 200 220 240LT(mm)
Pm (mg)Pm = 0,0178 × LT – 1,0613
Npmoy.= 15,3±2,7 Npmoy = 40,8±14,3 Npmoy =35,6±8,1 Npmoy =13,5±3,0

Stratégies alimentaires
89
La nature des proies ingérées varie également au cours de la croissance, avec un passage
progressif de proies de petite taille à des proies beaucoup plus volumineuses. Le poids moyen individuel
des proies consommées est positivement corrélé (r = 0,602; r2 = 0,362; P < 0,001) avec la longueur totale
des individus (Fig. II/II.17). Il augmente fortement entre les plus petits juvéniles (G0 : LT < 100 mm) et
les adultes de l'espèce (LT > 160 mm), passant de 0,4 mg pour les premiers à 2,5 mg pour les seconds.
Ce phénomène s'accompagne d'un élargissement progressif de la gamme de poids des proies
consommées, traduisant la persistance de proies de petite taille dans l'alimentation des adultes malgré
l'ingestion croissante de proies de plus en plus volumineuses avec la taille des poissons.
La composition du régime alimentaire se modifie d'un groupe de LT à l'autre (Tableau II/II.4).
Chez les plus jeunes individus (LT < 100 mm), les polychètes (Spionidae), constituent, avec les
cumacés, l'essentiel de l'alimentation. Le poids élevé des premiers (Cp = 65,6%) et le fort pourcentage
numérique des seconds (Cn = 58,1%) en font deux catégories de proies préférentielles, consommées par
plus de 60% des individus. Les amphipodes (F = 0,35) sont des proies secondaires et les mysidacés et
les bivalves des proies occasionnelles. L'importance des amphipodes augmente fortement dans le régime
alimentaire des juvéniles G1+ de 100-140 mm. Avec les cumacés (F = 0,69), ils constituent le groupe de
proies préférentielles de ces individus qui ingèrent également un nombre élevé de polychètes
(Spionidae) et de bivalves (F > 0,23) et, occasionnellement, quelques mysidacés (F = 0,13). A partir de
140 mm de LT, les bivalves (juvéniles entiers et pieds ou siphons d'adultes) dominent en poids (Cp >
47%) et en fréquence (F > 0,83). Ils constituent, avec les amphipodes (F = 1,00) et les cumacés (F =
0,68), le groupe de proies préférentielles des juvéniles G1+ de 140-160 mm, les autres proies ingérées
n'étant plus qu'occasionnelles (polychètes, copépodes) voire accidentelles (juvéniles d'ophiures, de
macroures et de mysidacés). Chez les adultes (LT > 160 mm), les bivalves sont les seules proies
préférentielles (F = 0,85). Les amphipodes, les mysidacés et les cumacés sont des proies secondaires (F
= 0,35 - 0,64) et les autres catégories d'invertébrés ingérées ne sont qu'occasionelles (polychètes -
surtout des carnivores -, juvéniles d'ophiure) voire accidentelles (copépodes, gastéropodes).
L'augmentation de taille s'accompagne d'un léger élargissement du spectre alimentaire. Celui-ci
ne correspond cependant qu'à une ingestion occasionelle croissante, par les individus de plus de 100
mm, de proies de tailles différentes (copépodes, macroures, gastéropodes, ophiures). L'essentiel des
modifications du régime alimentaire au cours de la croissance consiste donc en un remplacement
progressif des polychètes par les bivalves, avec une ingestion transitoire, entre 100 et 160 mm de LT, de
fortes quantités de petits crustacés (cumacés, amphipodes) (Fig. II/II.18).

Stratégies alimentaires
90
Tableau II/II.4 - Variations de l'alimentation de S. impar en fonction de la taille. Q = coefficient alimentaire desproies (Q = Cn × Cp). "" = valeur absente. LT = longueur totale (en mm).
Juvéniles G0(LT < 100 mm)
Juvéniles G1+
100 - 140 mmJuvéniles G1+
140 - 160 mm Adultes (LT > 160 mm)
Estomacs pleins 47 20 12 22
LT moyenne (min. - max.) 70,7 (49-96) 118,1 (100-139) 148,6 (142-159) 179,6 (160 – 212)
PROIES Q Q Q Q
AnnélidesPolychètes 2110,0 56,0 3,0 8,5
CrustacésCopépodes 1,8 8,9 1,7Mysidacés 13,9 8,9 0,4 43,0Cumacés 792,8 1441,0 265,7 37,1Amphipodes 36,4 999,0 1526,3 135,3Macroures (crevettes) 0,6 Crustacés indét. 0,1
MollusquesBivalves 15,4 36,4 395,6 2990,9Gastéropodes 0,5
EchinodermesOphiures 0,8 4,6
Figure II/II.18 - Variations de la composition pondérale (Cp) du régime alimentaire de S. impar au large duRhône en fonction de la taille des individus (LT, mm).
0%
25%
50%
75%
100%
< 100 mm 100-120 mm 140-160 mm > 160 mmClassesde taille
Cp
polychètes bivalves gastéropodes amphipodes macroures
mysidacés cumacés copépodes ophiures

Stratégies alimentaires
91
(CV = 61, 6%) (CV = 16,7%) (CV = 5,9%)
IR
1
2
3
4
Hiver Printemps Eté
Non-marginaux maxNon-marginaux min
75%25%
Médiane
Variations spatio-temporelles
S. impar n'ayant été capturée qu'entre 0 et 20 m de profondeur, seules les variations saisonnières
de son alimentation ont été étudiées. Chez cette espèce, l'intensité de la nutrition varie de façon
importante au cours de l'année. L'indice de réplétion stomacal diffère suivant la saison (Krukall-Wallis :
H(2, N=101) = 8,61 ; P < 0,05) (Fig. II/II.19) et est significativement plus faible en hiver (IRmoy.= 1,2 ± 0,3)
qu'au printemps (IRmoy.= 2,5 ± 0,2) et en été (IRmoy.= 2,4 ± 0,2) (Annexe 15). L'activité nutritionnelle de
S. impar est donc nettement réduite pendant la saison froide, avec un faible indice de réplétion et un fort
coefficient de vacuité (CV > 60%) en hiver, tandis qu'elle est importante au printemps et en été,
périodes où l'indice de réplétion est élevé et le coefficient de vacuité réduit (CV < 17%).
Figure II/II.19 - Représentation en Boites de Tukey des indices de réplétion stomacale (IR) observés àchacune des trois saisons étudiées chez les individus de S. impar au large du Rhône. CV = coefficient devacuité (% d'estomacs vides).
Les estomacs des quelques individus s'étant alimentés en hiver (N = 5) étant pratiquement vides
(IRmed.= 1), le régime alimentaire de S. impar n'a pu être déterminé qu'au printemps et en été. Afin
d'obtenir un nombre suffisant d'estomacs pleins par classe de taille à chaque saison, les individus ont été
regroupés en trois classes : LT < 100 mm (juvéniles G0), 100-160 mm (juvéniles G1+) et LT > 160 mm
(adultes). Chez S. impar, les variations saisonnières de l'alimentation dépendent de la taille des individus
(Fig. II/II.20). Les juvéniles G0 n'étant présents dans les captures qu'au printemps, les variations
saisonnières de l'alimentation observées ne concernent que les juvéniles G1+ et adultes (LT ≥ 100 mm).
Chez les juvéniles G1+ (100-160 mm), la composition du régime alimentaire diffère statistiquement (P <
0,001) entre le printemps et l'été alors que, chez les adultes de l'espèce (LT > 160 mm), elle est stable
d'une saison à l'autre (Annexe 16).

Stratégies alimentaires
92
Figure II/II.20 - Variations de l'alim
entation (pourcentages pondéraux des proies ingérées) de S. impar en fonction de la saison et de la taille des individus (longueur totale
LT , en m
m). N
= nombre d'estom
acs pleins analysé pour chaque groupe de poissons. Dans chaque cas, les proies surlignées en gris sont rencontrées dans plus de 20%
desestom
acs et/ou représentent plus de 10% du poids total des proies ingérées.
Printemps
Eté
Hiver
Juvéniles G0 L
T < 100 mm
Juvéniles G1
+ 100-160 mm
Adultes L
T > 160 mm
(N = 50)
polychètes65,6%
bivalves7,8%
amphipodes6,4% m
ysidacés6,5%
cumacés
13,7%
(N = 15)
polychètes16,4%
bivalves28,4%
amphipodes
18,8%
ophiures1,8%
crust. indét.3,0%
copépodes2,1%
cumacés
23,1%
mysidacés6,4%
(N = 12)
bivalves36,9%
copépodes6,5%
cumacés
15,1%
amphipodes
38,1%
mcroures3,4%
(N = 14)
ophiures
4,8%copépodes
1,0%cum
acés1,9%
bivalves68,7%
amphipodes5,9%
mysidacés14,0%
polychètes3,6%
(N = 10)
polychètes1,8%
bivalves89,6%
gastéropodes3,3%
cumacés
2,6%
copépodes0,8%
amphipodes1,8%

Stratégies alimentaires
93
Au printemps, période où toutes les classes de taille sont présentes dans les captures, les régimes
alimentaires des trois groupes de LT sont statistiquement différents (P < 0,001 ; Annexe 17) (Fig.
II/II.20). Les juvéniles G0 (LT < 100 mm) ont un régime clairement dominé par les polychètes (Cp =
65,6% ; F = 0,81), les juvéniles G1+ (100-160 mm) consomment de fortes quantités de bivalves (F =
0,64), de cumacés (F = 0,67), d'amphipodes (F = 0,87) et de polychètes (F = 0,25), alors que les adultes
(LT > 160 mm) ingèrent surtout des bivalves (F = 0,85). De même, en été, période où seuls les individus
de plus de 100 mm de LT sont présents, la nature des proies ingérées diffère entre juvéniles G1+ et
adultes (P < 0,001 ; Annexe 16). L'importance alimentaire des bivalves est réduite chez les juvéniles
(100-160 mm), où ils co-dominent avec les amphipodes (Q = 1233; Cp = 38%) et les cumacés (Q = 633
; Cp = 15,1%), alors que, chez les adultes (LT > 160 mm), ils sont nettement prépondérants (Q > 2990 ;
Cp > 89%). Le reste des proies est consommé de façon occasionnelle, soit par un seul des deux groupes
(macroures, gastéropodes, polychètes) soit par les deux (copépodes, cumacés).
II/II.3.2.3 - Discussion sur l'alimentation de Solea impar
Comportement alimentaire
A notre connaissance, le rythme d'alimentation de S. impar n'a encore jamais été décrit de façon
précise. Il n'a pas pu être déterminé en totalité au cours de la présente étude, du fait des très faibles
effectifs capturés la nuit pour cette espèce. Cependant, l'intensité de l'alimentation observée le jour
plaide en faveur d'une période d'alimentation maximale diurne chez S. impar, l'enfouissement des
individus inactifs pouvant expliquer la réduction des captures en dehors de la période de nutrition (c.a.d.
la nuit), comme cela a été suggéré pour la sole commune (De Groot, 1971). La présence de villosités
très développées au niveau de la narine de la face aveugle de S. impar, généralement considérée comme
une adaptation à la prédation par chémoréception elle-même généralement associée à une alimentation
nocturne (De Groot, 1969 ; Batty et Hoyt, 1995), met en doute cette conclusion. Il est possible que les
individus de S. impar échappent au chalut la nuit parce que chassant au niveau de la colonne d'eau
pendant cette période. Cependant, la présence exclusive de proies benthiques (bivalves, polychètes,
amphipodes) dans les tubes digestifs (rectum compris) des individus de l'espèce en début de matinée ne
va pas dans le sens d'une alimentation pélagique nocturne. De plus, la présence d'une papille sensorielle
très développée au niveau de la narine gauche de S. impar n'implique pas forcément une activité
alimentaire nocturne. Elle peut être associée au développement intensif d'un mode de capture "aveugle"
des proies enfouies dans le sédiment, hypothèse déjà proposée pour cette espèce (Marinaro et Bouabid,
1983). Pour ces raisons, S. impar sera considérée comme essentiellement active le jour, avec un pic
d'alimentation maximale vers midi.
Au large du Rhône, S. impar se nourrit essentiellement de proies de petites tailles ingérées en
grandes quantités. Cette observation est en accord avec la biologie de l'espèce et concorde avec la

Stratégies alimentaires
94
morphologie de son appareil digestif. En effet, S. impar est décrite comme une espèce vorace (Marinaro
et al., 1983) mais présentant, comme la majorité des soléidés, un œsophage et un estomac réduits,
adaptés à l'ingestion de proies de petite taille (De Groot, 1971). Ne pouvant pas, de par la morphologie
de son appareil digestif, consommer de proies volumineuses, l'espèce doit, pour satisfaire ses besoins
énergétiques élevés, cumuler un nombre important de proies de taille réduite. Le régime alimentaire
global obtenu est dominé par les bivalves et complété de crustacés de petite taille (cumacés,
amphipodes, mysidacés) et de polychètes (dépositivores de surface et carnivores). Il est surtout
représentatif de l'alimentation des juvéniles de moins de 140 mm de LT qui représentent plus de 66% des
estomacs analysés et correspond en grande partie aux observations de Rodriguez (1996) en Atlantique et
de Marinaro et Bouabid (1983) et Allam (1995) en Méditerranée. L'ingestion de polychètes et de
cumacés, importante pour l'espèce au large du Rhône, n'a été signalée qu'en Atlantique (Rodriguez,
1996) où ces proies ont une importance alimentaire réduite. Les différences observées entre
l'alimentation de S. impar au large du Rhône et sur les côtes égyptiennes, où le régime alimentaire se
caractérise par une dominance des macroures et des gastéropodes, confirme la forte adaptabilité et le
caractère euryphage de cette espèce, déjà soulignés par Allam (1995).
Modifications de l'alimentation
La prise de nourriture de S. impar, importante au printemps et maximale en été, semble
fortement réduite en hiver. Ce phénomène correspond aux résultats des travaux précédents (Marinaro et
Bouabib, 1983 ; Allam, 1995 ; Rodriguez, 1996) et est directement lié aux forts écarts de température
observés entre l'été (18-21°C) et l'hiver (6-13°C) à 0-20 m (Younes, 2000), profondeur où se
répartissent les individus de S. impar. En effet, la température a une influence marquée sur le
métabolisme et la croissance de l'espèce (Marinaro, 1991) qui présente, de façon générale, une pause de
croissance et une réduction d'activité en hiver, compensée par une forte croissance et une alimentation
intense en été (Marinaro et Bouabib, 1983 ; Marinaro et al., 1983). La prise importante de nourriture au
printemps s'explique, de plus, par les besoins métaboliques accrus des femelles pendant la vitellogénèse
(Marinaro et Bouabib, 1983), située à cette période de l'année dans le golfe du Lion (Shéhata, 1984).
La composition du régime alimentaire varie suivant la saison et surtout selon la taille des
individus (Fig. II/II.21). Les régimes alimentaires des juvéniles G0, des juvéniles G1+ et des adultes de
S. impar sont totalement distincts les uns des autres. Ils diffèrent non seulement par le poids des proies
consommées (qui augmente avec la taille), mais également par leur nature : polychètes dépositivores de
surface chez les juvéniles G0, amphipodes et cumacés chez les juvéniles G1+, bivalves chez les adultes.
L'augmentation, avec la taille des individus, de l'importance des bivalves dans le régime alimentaire des
soles adriatiques a été établie par plusieurs études préexistantes (Marinaro et Bouabib, 1983 ; Allam,
1995 ; Rodriguez, 1996) qui confirment également la prédominance des amphipodes dans l'alimentation
des juvéniles de plus de 100 mm.

Stratégies alimentaires
95
P
P
P
P
P
EE
P
P
EP
P
-2
0
2
-2 0 2
Axe 1
Axe 2
Stress = 0,033
< 100 mm
100- 160 mm
> 160 mm
70 - 100 m
30 - 50 m
0 - 20 m
Profondeur
juvéniles G0(LT < 100 mm)
adultes(LT > 160 mm)
juvéniles G1+
(100-160 mm)
Stade de vie
Figure II/II.21 - Variations de l'alimentation de S. impar en fonction de la saison (P = printemps, E =été, H = hiver), et de la taille (LT, mm) des individus à 0-20 m de profondeur. Ordination (N.M.S.,solution à 2 dimensions) réalisée sur les poids moyens des proies par estomac.
L'alimentation des plus petits juvéniles de l'espèce (G0 : LT < 100 mm) n'a pas, à notre
connaissance, été décrite avant cette étude. La consommation par ces individus de fortes quantités de
polychètes n'est cependant pas étonnante dans la mesure où les polychètes font partie du régime
alimentaire des autres stades de vie benthique de l'espèce (Rodriguez, 1996 ; présente étude). De plus,
les familles de polychètes consommées (petits dépositivores de surface) sont très abondantes sur les
fonds sableux où vit S. impar (Abou-Hamdan, 2001) et sont facilement capturables par ce poisson plat
chez qui la présence d'une papille sensorielle très développée sur la face aveugle permet une chasse
endogée active (Marinaro et Bouabib, 1983).
En ce qui concerne la saison, les modifications sont peu marquées. Chez les adultes de l'espèce,
l'alimentation est relativement homogène (bivalves) et ne diffère pas, quantitativement parlant, d'une
saison à l'autre. La variabilité de l'alimentation est plus importante chez les juvéniles, même chez les G0
pourtant uniquement présents au printemps (Fig. II/II.21). Elle est maximale pour les G1+, pour qui les
alimentations printanière et estivale se distinguent l'une de l'autre, avec un régime alimentaire
relativement homogène en été (bivalves et amphipodes) et, au contraire, très variable au printemps
(bivalves, amphipodes, polychètes et/ou cumacés). Ces observations sont en accord avec les variations
saisonnières de l'alimentation précédemment décrites pour S. impar en Atlantique (Rodriguez, 1996) et
en Méditerranée (Allam, 1995), avec une ingestion maximale de bivalves et d'amphipodes en été.

Stratégies alimentaires
96
II/II.3.3 - BUGLOSSIDIUM LUTEUM
II/II.3.3.1 - Alimentation globale
L'alimentation de B. luteum au large du Rhône a été décrite à partir des contenus stomacaux de
206 individus de taille comprise entre 30 et 130 mm (LT moy.= 88,6 ± 0,88 mm). Chez cette espèce, la
prise de nourriture est essentiellement nocturne, le remplissage stomacal étant significativement plus
élevé (Mann-Whitney : U(122,53) = 2519,5 ; P < 0,05) la nuit (IRmoy. = 2,3 ± 0,2) que le jour (IRmoy. = 1,7 ±
0,1) (Fig. II/II.22). La prise de nourriture s'intensifie en fin d'après midi et est maximale (IR = 3,5) entre
1:00 et 3:00 heures du matin. L'indice de réplétion est minimal au lever du soleil et reste faible pendant
la journée, malgré un pic secondaire d'alimentation vers midi (IR = 2,5). Au large du Rhône, B. luteum
présente donc deux pics d'alimentation : le plus important est situé en fin de nuit après une période
nocturne d'alimentation intense et l'autre, plus faible, vers midi, au milieu d'une période diurne
d'alimentation réduite.
Figure II/II.22 - Variations nycthémérales de l'indice de réplétion stomacal moyen (IR) chez B.luteum (juin 2000; heure locale = heure GMT + 1). CS = coucher du soleil; LS = lever du soleil.
Sur les 206 estomacs de B. luteum analysés, 60 ne contenaient pas de nourriture (CV = 29,1%).
Le régime alimentaire global de l'espèce a donc été établi à partir des contenus stomacaux de 146
individus de LT comprise entre 32 et 126 mm, englobant 15% de juvéniles G0, 22% de juvéniles G1+ et
63 % d'adultes. Il est caractérisé par l'ingestion fréquente d'un nombre réduit de proies de petite taille.
Les nombres de proies et les poids par estomac fluctuent respectivement autour de valeurs médianes de
3 proies et 2,4 mg (Fig. II/II.23 a et b). Ils sont variables mais les valeurs moyennes obtenues pour
l'espèce (Npmoy. = 5,6 ± 0,6 proies ; Ppmoy. = 4,0 ± 0,5 mg) reflètent bien les nombres et les poids
observés pour la majorité des estomacs analysés (2 à 7 proies et 1,1 à 4,4 mg).

Stratégies alimentaires
97
Figure II/II.23 - Représentation en Boites de Tukey des nombres (a) et des poids totaux (b) des proies par estomacchez B. luteum au large du Rhône.
Le spectre alimentaire de B. luteum est large (Tableau II/II.5). Les polychètes sont les proies
préférentielles de l'espèce, avec 10 familles représentées dans les estomacs. Les petits dépositivores de
sub-surface (Capitellidae) dominent en nombre, suivis de carnivores de taille moyenne (Nephtydae).
Les crustacés tiennent également une place importante dans le régime alimentaire de B. luteum, surtout
les amphipodes (Pontoporeidés et Oedicerotidés) qui constituent la seconde catégorie de proies
préférentielles de l'espèce. Les copépodes et les cumacés, consommés de façon régulière, forment, avec
les bivalves (petits juvéniles avalés entiers et siphons ou pieds d'individus plus gros) un groupe de
proies secondaires. Les autres proies ne contribuent que faiblement à l'alimentation (Q < 15). Les
mysidacés, les macroures, les brachyoures et les gastéropodes sont consommés de façon occasionnelle
alors que les ophiures, les poissons, les isopodes et les leptostracés peuvent être considérés comme des
proies accidentelles. On note également la présence, dans plus de 25% des estomacs, de débris végétaux
d'origine terrestre, souvent accompagnés de grains de sable ou de particules fines de sédiment.
(a) Np (b) Pp en mg
0
5
10
15
20
25
01234567
89
101112131415
Non marginaux max
Non marginaux min
75%25%
Médiane
Points marginaux

Stratégies alimentaires
98
Tableau II/II.5 - Régime alimentaire global de B. luteum au large du Rhône. F = fréquence ; N = nombre deproies ; Cn = pourcentage numérique ; P = poids sec des proies (mg) ; Cp = pourcentage pondéral ; Q =coefficient alimentaire des proies (Q = Cn × Cp). "+" = valeur inférieure à 0,01.
F N Cn P Cp Q
AnnélidesPolychètes 0,61 235 33,10% 241,35 47,28% 1564,93
CrustacésCopépodes 0,21 121 17,04% 16,17 3,17% 53,99Leptostracés + 1 0,14% 2,66 0,52% 0,07Mysidacés 0,08 14 1,97% 28,40 5,56% 10,97Cumacés 0,20 69 9,72% 26,96 5,28% 51,33Isopodes + 1 0,14% 1,34 0,26% 0,04Amphipodes 0,60 205 28,87% 62,22 12,19% 351,94Macroures (crevettes) 0,04 5 0,70% 28,82 5,65% 3,98Brachyoures 0,03 4 0,56% 10,66 2,09% 1,18
MollusquesBivalves 0,02 48 6,76% 62,23 12,19% 82,42Gastéropodes 0,03 4 0,56% 10,12 1,98% 1,12
EchinodermesOphiures 0,01 2 0,28% 4,55 0,89% 0,25
Poissons + 1 0,14% 14,98 2,93% 0,41
Total proies 710 100,0% 510,46 100,0%
II/II.3.3.2 - Modifications du régime alimentaire
Variations en fonction de la taille
L'alimentation de B. luteum se modifie au cours de la croissance. La faible valeur de stress
obtenue pour la N.M.S. (0,13) garantit une bonne représentation, dans le plan proposé, des distances
alimentaires réelles entre individus (Fig. II/II.24). Ces distances sont parfois importantes entre points
d'une même classe de LT mais les points obtenus pour l'ensemble des classes de taille s'organisent
suivant un gradient de taille croissante. La composition du régime alimentaire, bien que très variable
d'un individu à l'autre (surtout chez les adultes de LT > 70 mm), se modifie donc de façon progressive
avec la taille des individus. La classification par groupement agglomératif des classes de taille sépare
quatre grands groupes d'alimentation : les juvéniles G0 de moins de 50 mm de LT (groupe A), les
juvéniles G1+ de 50-70 mm (groupe B), les sub-adultes et adultes de 70-110 mm (groupe C) et les
adultes de plus de 110 mm (groupe C') (Fig. II/II.25). Cependant, les groupes C et C' se chevauchant en
partie sur le graphe de la N.M.S. et présentant des régimes alimentaires proches (dominés par les
polychètes et les amphipodes ; Annexe 18), trois grands groupes alimentaires seront retenus pour le
reste de l'étude : les juvéniles G0 de moins de 50 mm de LT (groupe A), les juvéniles G1+ de 50-70 mm
(groupe B) et les sub-adultes et adultes de plus de 70 mm (groupe C).

Stratégies alimentaires
99
Figure II/II.24 - Ordination (NMS, solution à 2 dimensions) des différentes classes de taille (LT,mm) de B. luteum en fonction de leur régime alimentaire (Poids moyen des proies par estomac).
Figure II/II.25 - Classification par groupement agglomératif des classes de taille de B. luteum en fonction deleur régime alimentaire (poids moyens des proies par estomac). La flèche verticale rouge indique lepourcentage d'information (65 %) retenu comme valeur seuil pour l'établissement des groupes alimentaires(A, B, C et C').
Stress = 0,130-2
0
2
-2 0 2
Axe 1
Axe
2
30-40 mm
40-50 mm
50-60 mm
60-70 mm
70-80 mm
80-90 mm
90-100 mm
100-110 mm
110-120 mm
>120 mm
Taille (LT, mm)
A
B
C
C'

Stratégies alimentaires
100
Les modifications de l'alimentation au cours de la croissance sont quantitatives et qualitatives.
Le nombre de proies par estomac est très variable et ne subit pas de modification significative avec la
taille des poissons (Kruskal-Wallis : H(2, N=128) = 1,5 ; P > 0,45). L'augmentation de ration alimentaire
(poids total par estomac, Pp) observée avec la taille des poissons (Kruskal-Wallis : H(2, N=128) = 22,1 ; P <
0,001) correspond donc essentiellement à une élévation du poids individuel des proies ingérées. Le
poids moyen des proies consommées est positivement corrélé à la longueur totale des individus (r =
0,410; r2 = 0,168; P < 0,001) (Fig. II/II.26). Il augmente de façon significative au cours de la croissance
et passe de 0,4 à 1,2 mg entre 40 et 120 mm de LT, c'est à dire entre juvéniles G0 et adultes de l'espèce.
On observe, à partir de 70 mm de LT, un élargissement progressif de la gamme de poids des proies
consommées qui indique la persistance de proies de petite taille dans l'alimentation des adultes. La
gamme de poids des proies ingérées passe néanmoins de 0,01-1,1 mg par proie chez les juvéniles G0 à
0,5-4,1 mg chez les adultes.
Figure II/II.26 - Relation linéaire entre la taille (LT, mm) des individus de B. luteum et le poids moyen (Pm,mg) des proies ingérées (r = 0,410; r2 = 0,168; P < 0,001).
Les catégories de proies ingérées diffèrent également d'un groupe de LT à l'autre (Tableau
II/II.6). Chez les juvéniles G0 (LT < 50 mm), le régime alimentaire est clairement dominé par les
copépodes harpacticoïdes (F = 0,67) et les cumacés (F = 0,44). Il comprend également des amphipodes
(surtout des Pontoporeidae), des polychètes (essentiellement de petits Nephtydae carnivores), des
gastéropodes et des bivalves, ingérés de façon secondaire (F = 0,11-0,44).
Pm = 0,0107 × LT – 0,0578
0
0,5
1
1,5
2
2,5
3
3,5
4
20 30 40 50 60 70 80 90 100 110 120 130 LT(mm)
Pm (mg)

Stratégies alimentaires
101
Tableau II/II.6 - Variations de l'alimentation de B. luteum en fonction de la taille. Q = coefficientalimentaire. "+" = valeur inférieure à 0,01 ; "" = valeur absente. LT = longueur totale (en mm).
Juvéniles G0LT < 50 mm
Juvéniles G1+
50 < LT < 70 mmSub-adultes et adultes
LT > 70 mm
Estomacs pleins 22 32 92
LT moy (min. - max.) 39,5 (32 – 48) 63,0 (51-76) 99,1 (71-126)
PROIES Q Q Q
AnnélidesPolychètes 111,5 303,0 2132,5
CrustacésCopépodes 1157,6 508,1 2,9Leptostracés + 0,1Mysidacés 4,9 14,5Cumacés 231,2 150,6 32,0Isopodes 0,1Amphipodes 124,5 259,4 410,0Macroures 8,1Brachyoures 1,8
MollusquesBivalves 21,2 202,0 79,2Gastéropodes 33,6 10,4 0,1
EchinodermesOphiures 5,0 0,1
Poissons 0,6
Chez les juvéniles G1+ (50-70 mm), le régime alimentaire, plus diversifié, est caractérisé par
une codominance (F = 0,41-0,65) de copépodes harpacticoïdes, d'amphipodes, de polychètes
(Nephtydae et Spionidae) et de bivalves qui représentent, groupés, plus de 64% du poids total des proies
ingérées. Les cumacés sont des proies secondaires et les gastéropodes, les ophiures et les mysidacés des
proies occasionelles. A partir de 70 mm de LT, l'alimentation est dominée par les polychètes (F = 0,75),
qui constituent plus de 42% en nombre et 50% en poids des proies ingérées. Les adultes consomment
également, mais en quantités moindres, des amphipodes (F = 0,47), des bivalves (F = 0,14) et des
cumacés (F = 0,05). S'y ajoutent de nombreuses proies occasionnelles (mysidacés, copépodes,
macroures, brachyoures) ou accidentelles (poissons, gastéropodes, isopodes, leptostracés, ophiures).
Le spectre alimentaire de B. luteum s'élargit au cours de la croissance, le nombre moyen de
catégories de proies consommées étant multiplié par 2 (il passe de 6 à 13) entre juvéniles G0 (LT < 50
mm) et adultes de l'espèce (LT > 70 mm). Cet élargissement correspond essentiellement à une ingestion
croissante de proies de plus en plus volumineuses par les adultes (mysidacés, macroures, brachyoures,
poissons, ophiures). Cependant, l'essentiel des modifications ontogéniques du régime alimentaire de B.

Stratégies alimentaires
102
luteum concerne l'importance pondérale des polychètes dans l'alimentation, qui s'accroît avec la taille
des poissons au détriment des copépodes, des cumacés et des gastéropodes (Fig. II/II.27). Le spectre des
familles de polychètes consommées, très étroit chez les juvéniles G0 (une famille), s'élargit chez les
juvéniles G1+ et les adultes (respectivement 3 et 10 familles). Les polychètes ingérés sont
essentiellement des carnivores (Nephtydae chez les juvéniles G0; Nephtydae et Lumbrineridae chez les
juvéniles G1+; Nephtydae, Glyceridae, Polynoidae et Phyllodocidae chez les adultes), auxquels
s'ajoutent principalement des dépositivores de surface (Spionidae) chez les juvéniles G1+ et des
dépositivores de sub-surface (Capitellidae) chez les adultes.
Figure II/II.27 - Variations de la composition pondérale (Cp) du régime alimentaire de B. luteum au large duRhône en fonction de la taille des individus (LT, mm).
Variations spatio-temporelles
Les fluctuations spatio-temporelles des captures de B. luteum ont empêché l'analyse croisée des
effets de la taille, de la profondeur et de la saison sur l'alimentation, le nombre d'estomacs pleins par
classe de LT étant parfois inférieur à 10. Les effets de la saison et de la profondeur sur l'alimentation de
l'espèce ont donc été traitées séparément.
����������������������������������������������������������������������������������������������������������������������������������������������������
���������������������������������������������������������������������������������������������������������������
�������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������
���������������������������������������������������������������������������������������������������������������
��������������������������������������������������������������������������
����������������������������������������������������������������������������������������������������������������������������������������������������
���������������������������������������������������������������������������������������������������������������
���������������������������������������������������������������������������������������������������������������
��������������������������������������������������������������������������
��������������������������������������������������������������������������
����������������������������������������������������������������������������������������������������������������������������������������������������
���������������������������������������������������������������������������������������������������������������
���������������������������������������������������������������������������������������������������������������
����������������������������������������������������������������������������������������������������������������������������������������������������
��������������������������������������������������������������������������
��������������������������������������������������������������������������
��������������������������������������������������������������������������
�������������������������������������
0%
25%
50%
75%
100%
LT < 50 mm 50-70 mm LT > 70 mmClasses de taille
Cp
������polychètes bivalves
������gastéropodes
������amphipodes
������isopodes
������ macroures ������� mysidacés ������ brachyoures ������ leptostracés ������ cumacés������������ copépodes
�������������� ophiures poissons

Stratégies alimentaires
103
IR
0
1
2
3
4
hiver printemps été
Non-marginaux max Non-marginaux min
75% 25%
Médiane
(CV = 60, 1%) (CV = 14,6%) (CV = 19,6%)
Variations saisonnières
Chez B. luteum, l'intensité de la nutrition varie au cours de l'année (Fig. II/II.28). L'indice de
réplétion stomacal varie suivant la saison (Krukall-Wallis : H (2, N=122) = 12,31 ; P < 0,01), avec des
valeurs significativement moins élevées (Annexe 19) en hiver (IRmoy.= 0,8 ± 0,2) qu'au printemps
(IRmoy.= 2,1 ± 0,2) et en été (IRmoy.= 1,8 ± 0,3). Ceci confirme la réduction hivernale de l'activité
nutritionnelle de B. luteum suggérée par l'évolution saisonnière du coefficient de vacuité, fort en hiver
(CV = 60%) et réduit aux autres saisons (CV < 20%).
Figure II/II.28 - Représentation en Boites de Tukey des indices de réplétion stomacale (IR) observésaux trois saisons étudiées chez B. luteum au large du Rhône. CV = coefficient de vacuité (%d'estomacs vides).
La composition du régime alimentaire varie également au cours de l'année. Les modifications
saisonnières dépendent de la taille des individus (Fig. II/II.29). Les juvéniles G0 (LT < 50 mm) n'ayant
pratiquement été capturés qu'en été, les variations saisonnières de l'alimentation mises en évidence ne
concernent que les individus de plus de 50 mm (juvéniles G1+ et adultes). Pour ces derniers, le régime
alimentaire reste globalement le même quels que soient le groupe et la saison, mis à part en été où les
alimentations des juvéniles G1+ (50-70 mm) et des adultes (LT > 70 mm) diffèrent de façon significative
(Annexe 20). La composition du régime alimentaire des adultes (LT < 70 mm), seul groupe de taille
présent aux trois saisons, ne diffère pas de façon significative d'une saison à l'autre (Annexe 21), les
seules modifications observées concernant les catégories de proies accidentelles (gastéropodes, isopodes
et ophiures au printemps; leptostracés et juvéniles de poissons en été; brachyoures en hiver).

Stratégies alimentaires
104
Figure II/II.29 - Variations de l'alim
entation (pourcentages pondéraux des proies ingérées) de B. luteum en fonction de la saison et de la taille des individus (longueur totale L
T ,en m
m). N
= nombre d'estom
acs pleins analysé pour chaque groupe de poissons. Dans chaque cas, les proies surlignées en gris sont rencontrées dans plus de 20%
des estomacs
et/ou représentent plus de 10% du poids total des proies ingérées.
Printemps
Eté
Hiver
Adultes L
T > 70 mm
Juvéniles G1
+ 50-70 mm
Juvéniles G0 L
T < 50 mm
(N = 20)
cumacés
17,4%amphipodes17,4%
gastéropodes16,4%
bivalves10,4%
polychètes21,9%
copépodes16,4%
(N = 12)
mysidacés6,8%
cumacés
15,3%
copépodes13,4%
gastéropodes14,7%
ophiures7,1%
bivalves17,3%
polychètes15,9%
amphipodes9,5%
(N = 22)
polychètes36,7%
poissons15,8%
copépodes2,9%
leptostracés2,8%
mysidacés3,4%
amphipodes
11,0%
macroures14,6%
bivalves12,8%
(N = 41)
macroures 6,8%
amphipodes9,1%
bivalves8,4%
polychètes57,0%
mysidacés 9,3%
brachyoures4,8%
cumacés
4,6%
(N = 39)
polychètes52,0%
bivalves16,5%
gastéropodes1,3%
ophiures1,6%
copépodes3,2%
cumacés
5,1%m
ysidacés1,6%
amphipodes
17,7%
isopodes1,1%
(N = 12)
copépodes11,2%
polychètes38,0%
bivalves19,6%
amphipodes19,2%
cumacés
12,0%

Stratégies alimentaires
105
En été, période où toutes les classes de taille sont représentées dans les captures, les régimes
alimentaires des trois groupes de LT diffèrent de façon significative (P < 0,05 ; Annexe 22) malgré un
nombre important de proies en commun (polychètes, bivalves, amphipodes, copépodes). Les juvéniles
G0 (LT < 50 mm) ont une alimentation diversifiée, où les copépodes (F = 0,67), les cumacés (F = 0,44),
les amphipodes (F = 0,44), les polychètes (F = 0,28), les bivalves (F = 0,17) et les gastéropodes (F =
0,11) codominent (Fig. II/II.29). La composition en poids du régime alimentaire des juvéniles G1+ (50-
70 mm) est, à quelques proies accidentelles près (mysidacés, ophiures), similaire à celle de
l'alimentation des G0. On note cependant, par rapport à ces derniers, une augmentation des quantités de
bivalves (F = 0,44) et de polychètes (F = 0,48) consommées, au détriment des amphipodes (F = 0,14) et,
surtout, des gastéropodes (F < 0,01 malgré leur Cp élevé). Les polychètes (Q > 600 ; F = 0,58) dominent
de façon plus nette dans l'alimentation des adultes (LT > 70 mm), où amphipodes (F = 0,24) et bivalves
(F = 0,14) sont les deux seules autres catégories de proies ingérées en quantités non négligeables. Au
printemps, période où seuls les poissons de plus de 50 mm sont présents, la composition de
l'alimentation ne diffère pas, quantitativement parlant, d'un groupe de LT à l'autre (Annexe 20). Le
régime alimentaire des deux groupes est principalement constitué de polychètes (Q > 900 ; F > 0,65),
d'amphipodes (Q > 202 ; F > 0,36) et de bivalves (Q > 162 ; F > 0,34), les autres proies consommées à
cette saison n'étant qu'occasionnelles (cumacés, copépodes) ou accidentelles (gastéropodes, mysidacés,
ophiures, isopodes). En hiver, saison où seuls les adultes (LT > 70 mm) sont présents, le régime
alimentaire est largement dominé par les polychètes (Q > 3100), auxquels s'ajoutent principalement des
amphipodes (Q = 916; F = 0,36) et des bivalves (Q = 151 ; F = 0,34).
Variations en fonction de la profondeur
Au large du Rhône, l'intensité de nutrition de B. luteum ne varie pas de façon significative d'une
tranche bathymétrique à l'autre (IR, Kruskall-Wallis : H(2, N = 61) = 2,51 ; P > 0,28). Par contre, la
composition du régime alimentaire subit des modifications non négligeables suivant la profondeur. Les
juvéniles (LT ≤ 70 mm) n'étant présents qu'à 0-20 m, les variations observées ne concernent que les sub-
adultes et adultes de l'espèce (LT > 70 mm) (Fig. II/II.30). Pour ces derniers, le régime alimentaire des
individus capturés à 70-100 m diffère significativement (P < 0,05 ; Annexe 23) de celui des individus
pêchés moins profond (0-20 m et 30-50 m), par une importance réduite des polychètes au profit des
crustacés.
Entre 0 et 20 m, seule tranche bathymétrique où toutes les classes de taille de B. luteum sont
présentes, le régime alimentaire diffère statistiquement suivant la taille des poissons (P < 0,05 ; Annexe
24). Les juvéniles G0 (LT < 50 mm) présentent un régime alimentaire diversifié, avec une codominance
pondérale de copépodes (F = 0,67), de cumacés (F = 0,44), d'amphipodes (F = 0,44), de polychètes (F =
0,28), de bivalves (F = 0,17) et de gastéropodes (F = 0,11) (Fig. II/II.30).

Stratégies alimentaires
106
Figure II/II.30 - Variations de l'alim
entation (pourcentages pondéraux des proies ingérées) de B. luteum en fonction de la profondeur et de la taille des individus (longueur
totale LT , en m
m). N
= nombre d'estom
acs pleins analysé pour chaque groupe de poissons. Dans chaque cas, les proies surlignées en gris sont rencontrées dans plus de
20% des estom
acs et/ou représentent plus de 10% du poids total des proies ingérées.
0- 20 m
30 - 50 m
70 - 100 m
Juvéniles G0 L
T < 50 mm
Juvéniles G1
+ 50-70 mm
Adultes L
T > 70 mm
(N = 35)
amphipodes
23,5%
macroures25,8%
mysidacés20,1% cum
acés9,2%
copépodes3,3%
polychètes18,1%
(N = 32)
brachyoures2,9%
amphipodes11,8%
poissons6,8%
polychètes46,0%
bivalves17,2% ophiures
0,9%
macroures4,7%
copépodes2,2%
cumacés
3,2%
mysidacés3,1%
leptostracés1,2%
(N = 35)
polychètes69,9%
brachyoures2,6%
cumacés
2,6%
mysidacés4,5%
amphipodes8,1%
macroures2,2%
isopodes0,8%
gastéropodes1,0%
bivalves8,3%
(N = 22)
polychètes21,4%bivalves
17,8%
gastéropodes11,0%
amphipodes
12,0%
mysidacés5,1%
cumacés
14,5% copépodes12,8%
ophiures5,3%
(N = 22)
polychètes21,9%bivalves
10,4%
gastéropodes16,4%
copépodes16,4%
cumacés
17,4%amphipodes
17,4%

Stratégies alimentaires
107
Chez les juvéniles G1+ (50-70 mm), l'importance pondérale des amphipodes et des cumacés
diminue au profit des polychètes et des bivalves, consommés de façon plus régulière (F > 0,53). Les
polychètes (F = 0,58) dominent l'alimentation des adultes (LT > 70 mm), suivis des amphipodes (F =
0,34) et des bivalves (F = 0,22). Sur les fonds de 30-50 m de profondeur, où seuls les adultes sont
capturés, le régime alimentaire est clairement dominé par les polychètes (F = 0,78) qui représentent plus
de 69% en poids et 64% en nombre des proies ingérées et constituent la seule catégorie de proies
préférentielles de l'espèce. A cette profondeur, l'alimentation comprend également un nombre non
négligeable d'amphipodes (F = 0,38), de bivalves (F = 0,27) et de cumacés (F = 0,21). A 70-100 m,
l'alimentation des adultes (LT > 70 mm), est largement dominée par les crustacés avec une importance
majeure des amphipodes (F = 0,71) qui constituent, avec les polychètes (F = 0,56), le groupe de proies
préférentielles. Les mysidacés, les macroures et les cumacés sont des proies secondaires et les
copépodes des proies occasionnelles.
A ces modifications générales du régime alimentaire avec la profondeur s'ajoutent des
différences plus fines dans la nature des proies ingérées, notamment en ce qui concerne les Polychètes.
Les espèces les plus consommées ne sont pas les mêmes aux trois profondeurs (Annexe 25). Il s'agit
principalement de carnivores à 0-20 m (Nephtydae pour les juvéniles, Lumbrineridae et Glyceridae pour
les adultes), de dépositivores de sub-surface (Capitellidae) à 30-50 m et de suspensivores
(Ampharetidae) et de carnivores (Lumbrineridae) à 70-100 m.
II/II.3.3.3 - Discussion sur l'alimentation de Buglossidium luteum
Comportement alimentaire
Au large du Rhône, B. luteum s'alimente pendant toute la durée du nycthémère. L'activité de
nutrition est cependant plus intense la nuit, avec un pic d'alimentation maximale vers 2-3 heures du
matin, ce qui confirme les résultats préexistants sur la nutrition de cette espèce, décrite comme
essentiellement nocturne (De Groot, 1969 ; Tito de Morais, 1983 ; Darnaude, 1999 ; Darnaude et al.,
2001). La faiblesse des nombres et des poids de proies observés par estomac traduit l'ingestion fréquente
d'un petit nombre de proies de taille réduite. Ces observations sont en accord avec les résultats de Tito
de Morais (1983) et Darnaude et al. (2001) et expliquent la prise de nourriture diurne de l'espèce. En
effet, B. luteum se nourrit suivant le principe du "peu mais souvent", le faible nombre de proies par
estomac étant compensé par une prise de nourriture quasi ininterrompue durant toute la durée du
nycthémère (De Groot, 1969 ; Tito de Morais, 1984 ; Darnaude et al., 2001).
Le régime alimentaire global obtenu est surtout représentatif de l'alimentation des adultes de B.
luteum (63% des estomacs pleins analysés). Il est dominé par les polychètes (carnivores et dépositivores
de sub-surface) mais comprend également une quantité importante de crustacés de petite taille

Stratégies alimentaires
108
(amphipodes, cumacés, copépodes) et de bivalves. Ces résultats confirment l'importance des crustacés et
des polychètes dans l'alimentation de cette espèce, déjà décrite en Atlantique (Nottage et Perkins, 1983)
et en Méditerranée (Tito de Morais, 1983 ; Tito de Morais et Bodiou, 1984 ; Darnaude, 1999 ; Darnaude
et al., 2001). Cependant, la prépondérance des polychètes dans le régime alimentaire de B. luteum au
large du Rhône (Darnaude, 1999; Darnaude et al., 2001; présente étude) ne correspond pas aux
observations de Tito de Morais et Bodiou (1983, 1984), qui suggèrent une nette dominance des
crustacés dans l'alimentation à Banyuls-sur-Mer. Ce phénomène souligne la forte adaptabilité trophique
de l'espèce, capable de modifier la nature des proies qu'elle ingère en fonction de l'accessibilité des
ressources disponibles dans son environnement (Nottage et Perkins, 1983 ; Darnaude et al., 2001).
Modifications de l'alimentation
La prise de nourriture de B. luteum, fortement réduite en hiver, est importante au printemps et
maximale en été. Ceci résulte en partie des faibles températures hivernales et des fortes températures
estivales observés à l'embouchure du Rhône au dessus de 20 m de profondeur (Younes, 2000), zone
bathymétrique où se répartissent les juvéniles de l'espèce. En effet, la température a une influence
marquée sur le métabolisme et la croissance de B. luteum (Tito de Morais, 1986) qui présente, de façon
générale, une pause de croissance et une réduction de l'alimentation en hiver, compensée par une forte
croissance et une alimentation intense en été (Tito de Morais, 1983, 1984, 1986 ; Darnaude, 1999).
L'augmentation des besoins métaboliques des femelles pendant la phase de vitellogénèse (Deniel, 1984)
explique la prise importante de nourriture de l'espèce au printemps, la reproduction de B. luteum se
faisant en fin de printemps-début d'été dans le golfe du Lion (Shéhata, 1984).
Au large du Rhône, la composition du régime alimentaire de B. luteum varie selon la taille des
individus, la profondeur et la saison, les deux premiers facteurs étant les plus déterminants (Fig.
II/II.31). Le régime alimentaire des juvéniles de l'année (LT < 50 mm) s'individualise totalement de celui
des autres classes de taille. Les alimentations des juvéniles G1+ (50-70 mm) et des adultes (LT > 70 mm)
sont également distinctes, mais se rapprochent à 0-20 m au printemps, où l'ensemble des individus de
plus de 50 mm se nourrit essentiellement de polychètes, d'amphipodes et de bivalves. Bien que
l'alimentation de l'espèce comprenne une majorité de polychètes et d'amphipodes quel que soit le stade
de vie benthique, le poids et la nature des proies consommées diffèrent entre juvéniles et adultes. Le
régime alimentaire des premiers contient plus de petits crustacés (copépodes, cumacés) et de juvéniles
de mollusques (surtout des gastéropodes) que celui des seconds, plutôt basé sur l'ingestion de proies
volumineuses (polychètes, macroures). Ces résultats rejoignent ceux de Nottage et Perkins (1983), Tito
de Morais (1983, 1986) et Darnaude (1999) qui notent, avec la croissance des individus, un
remplacement progressif des crustacés de petite taille (copépodes, cumacés) par des proies de taille plus
importante (polychètes, bivalves).

Stratégies alimentaires
109
Figure II/II.31 - Variations de l'alimentation de B. luteum en fonction de la profondeur (0-20 m, 30-50 m, 70-100m), de la saison (P = printemps, E = été, H = hiver), et de la taille (LT, mm) des individus. Ordination (N.M.S.,solution à 2 dimensions) réalisée sur les poids moyens des proies par estomac.
La variabilité de l'alimentation, réduite chez les juvéniles (LT < 70 mm), augmente de façon
importante chez les adultes, qui présentent un régime alimentaire différent suivant la saison (printemps,
été ou hiver) et la tranche bathymétrique (0-20, 30-50 ou 70-100 m) (Fig. II/II.31). Les polychètes sont
des proies préférentielles à toutes les saisons et toutes les profondeurs mais la composition du régime
alimentaire subit un certain nombre de modifications en fonction de ces deux facteurs. L'alimentation,
largement dominée par les polychètes en été, est complétée par une ingestion élevée d'amphipodes au
printemps et en hiver. En ce qui concerne la profondeur, la consommation de polychètes, maximale à
30-50 m, est complétée par l'ingestion d'importantes quantités d'amphipodes et de bivalves à 0-20 m et
d'amphipodes à 70-100 m. De telles variations spatio-temporelles de la composition du régime
alimentaire ont déjà été décrites chez B. luteum au large du Rhône (Darnaude, 1999 ; Darnaude et al.,
2001). Elles sont fréquentes chez cette espèce qui présente une alimentation variée et très opportuniste
(Nottage et Perkins, 1983). Les polychètes et les crustacés constituent cependant l'essentiel de
l'alimentation de l'espèce au large du Rhône, tant au stade juvénile qu'au stade adulte.
E
E
E E
EP
P
PP
P P
EP
P P
EE
HH
H
H
H
H
E
-2
0
2
-2 0 2
Axe 1
Axe
2
70 - 100 m
30 - 50 m
0 - 20 m
Profondeur
juvéniles G0(LT < 50 mm)
adultes(LT > 70 mm)
juvéniles G1 +
50-70 mm
Stade de vie
LT < 50 mmLT > 70 mm50- 70 mm
Stress = 0, 095

Stratégies alimentaires
110
II/II.3.4 - ARNOGLOSSUS LATERNA
II/II.3.4.1 - Alimentation globale
L'alimentation d'Arnoglossus laterna au large du Rhône a été établie à partir des contenus
stomacaux de 906 individus, de LT comprise entre 20 et 132 mm (LT moy.= 87,9 ± 0,87). La période de
nutrition de l'espèce est essentiellement diurne, l'indice de réplétion stomacal étant significativement
plus élevé (Mann-Whitney : U(827, 81) = 28204,0 ; P < 0,05) le jour (IRmoy. = 1,9 ± 0,03) que la nuit (IRmoy.
= 1,6 ± 0,2) (Fig. II/II.32). La prise de nourriture est maximale l'après-midi (IR > 3,2) avec un léger pic
juste avant le coucher du soleil, entre 18:00 et 20:00 heures (IR = 3,8). Elle diminue fortement la nuit,
pour atteindre un minimum (IR = 0,2) juste avant le lever du soleil. L'indice de réplétion stomacal croît
progressivement à l'aube pour reprendre une valeur moyenne supérieure à 2 en milieu de matinée.
Arnoglossus laterna présente donc un rythme d'alimentation très contrasté, avec une prise de nourriture
qui se fait presque exclusivement de jour et un pic d'alimentation unique avant le coucher du soleil.
Figure II/II/32 - Variations nycthémérales de l'indice de réplétion stomacal moyen (IR) chez A.laterna (juin 2000; heure locale = heure GMT +1). CS = coucher du soleil; LS = lever du soleil.
Sur les 906 estomacs d'A. laterna analysés, 165 ne contenaient pas de nourriture (CV = 18,2%).
Le régime alimentaire global de l'espèce a donc été établi à partir des contenus stomacaux de 741
individus de LT comprise entre 20 et 132 mm, dont 8% de juvéniles G0, 12% de juvéniles G1+ et 80%
d'adultes. Il est caractérisé par l'ingestion d'un nombre variable de proies de tailles différentes.

Stratégies alimentaires
111
Le nombre et le poids des proies par estomac fluctuent respectivement autour de valeurs
médianes de 8 proies et de 4,2 mg (Fig II/II.33 a et b). Ils sont très variables (de 1 à 230 proies* et de
0,08 à 60,3 mg∗ par estomac) mais les valeurs moyennes obtenues (Npmoy.= 12,9 ± 0,7 ind. est.-1 et Ppmoy.
= 6,3 ± 0,3 mg est.-1) se situent dans les fourchettes de valeurs observées pour la plupart des estomacs
analysés (respectivement 3-14 ind. et 1,5-8,1 mg).
Figure II/II.33 - Représentation en Boites de Tukey des nombres (a) et des poids totaux (b) des proies par estomacrencontrés chez A. laterna au large du Rhône.
Le régime alimentaire global d'A. laterna est relativement diversifié mais dominé par les
crustacés (Tableau II/II.7). Les mysidacés, essentiellement des juvéniles de Leptomysis sp., sont les
proies préférentielles de l'espèce. Les amphipodes (surtout des Ampeliscidae et des Caprellidae), sont
consommés de façon régulière et forment l'essentiel des proies secondaires, que complètent des
macroures (Crangonidae) et des polychètes. Au total, 19 familles de polychètes ont été recensées dans
les estomacs d'A. laterna, avec une prédominance de petits suspensivores (Ampharetidae). Les
phoronidiens (fragments de parties antérieures) se rangent également parmi les proies secondaires mais
leur consommation concerne peu d'individus (F = 0,05). En dépit de leur coefficient alimentaire élevé,
leur importance dans le régime alimentaire est donc comparable à celles des proies occasionnelles
(brachyoures, bivalves et cumacés). Le reste des crustacés consommés (copépodes, leptostracés,
anomoures), les juvéniles de poissons (Gobiidae) et les ophiures sont des proies accidentelles. A. laterna
ingère également des débris végétaux d'origine terrestre, recensés dans 13% des estomacs analysés.
∗ valeurs extrêmes ne concernant que quelques individus et donc non représentées sur la Figure II/II.33
0
5
10
15
20
25
30
35
40
45
50
0
5
10
15
20
25
30
Non marginaux max
Non marginaux min
75% 25%
Médiane
Points marginaux
(a) Np (b) Pp en mg
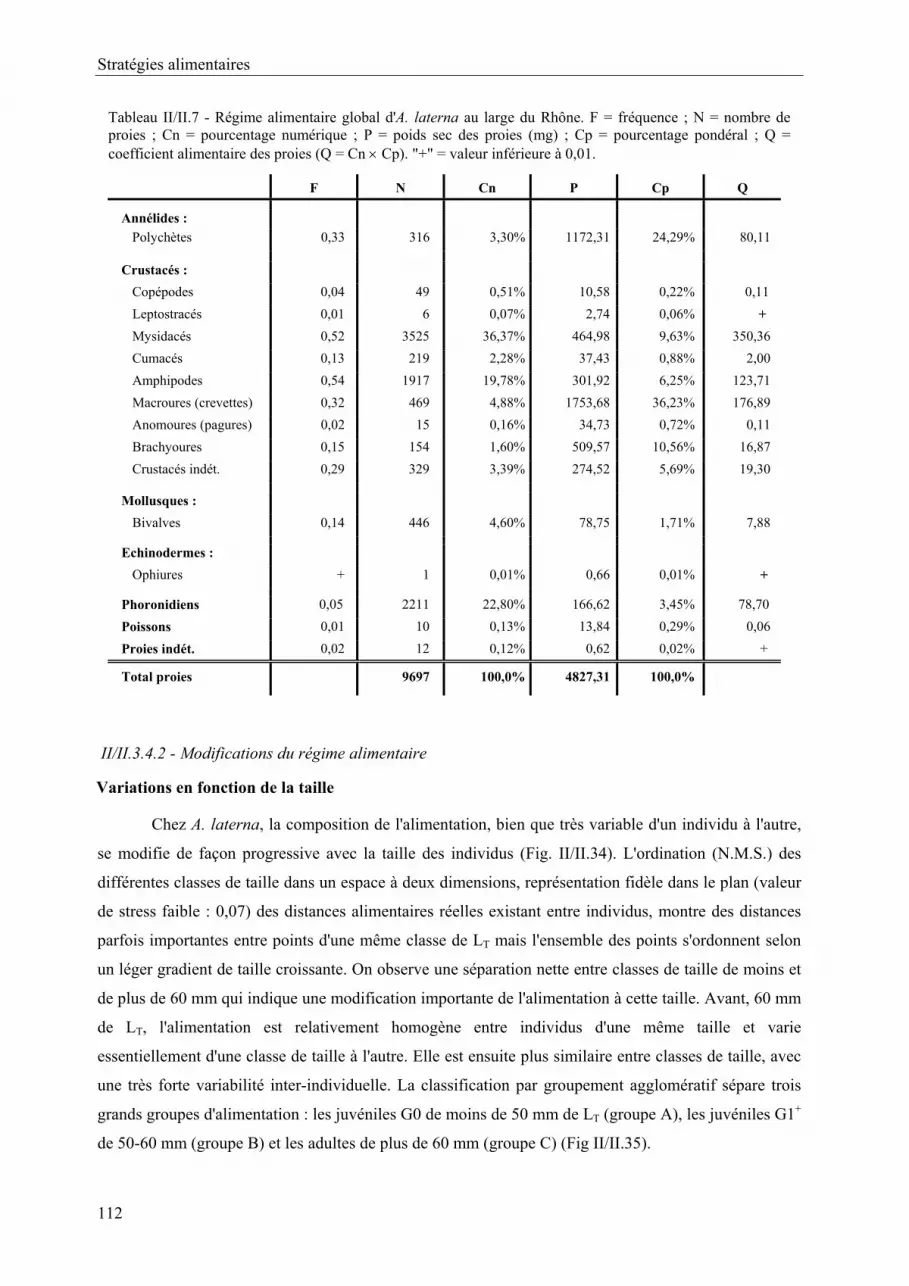
Stratégies alimentaires
112
Tableau II/II.7 - Régime alimentaire global d'A. laterna au large du Rhône. F = fréquence ; N = nombre deproies ; Cn = pourcentage numérique ; P = poids sec des proies (mg) ; Cp = pourcentage pondéral ; Q =coefficient alimentaire des proies (Q = Cn × Cp). "+" = valeur inférieure à 0,01.
F N Cn P Cp Q
Annélides :Polychètes 0,33 316 3,30% 1172,31 24,29% 80,11
Crustacés :Copépodes 0,04 49 0,51% 10,58 0,22% 0,11Leptostracés 0,01 6 0,07% 2,74 0,06% +Mysidacés 0,52 3525 36,37% 464,98 9,63% 350,36Cumacés 0,13 219 2,28% 37,43 0,88% 2,00Amphipodes 0,54 1917 19,78% 301,92 6,25% 123,71Macroures (crevettes) 0,32 469 4,88% 1753,68 36,23% 176,89Anomoures (pagures) 0,02 15 0,16% 34,73 0,72% 0,11Brachyoures 0,15 154 1,60% 509,57 10,56% 16,87Crustacés indét. 0,29 329 3,39% 274,52 5,69% 19,30
Mollusques :Bivalves 0,14 446 4,60% 78,75 1,71% 7,88
Echinodermes :Ophiures + 1 0,01% 0,66 0,01% +
Phoronidiens 0,05 2211 22,80% 166,62 3,45% 78,70Poissons 0,01 10 0,13% 13,84 0,29% 0,06Proies indét. 0,02 12 0,12% 0,62 0,02% +
Total proies 9697 100,0% 4827,31 100,0%
II/II.3.4.2 - Modifications du régime alimentaire
Variations en fonction de la taille
Chez A. laterna, la composition de l'alimentation, bien que très variable d'un individu à l'autre,
se modifie de façon progressive avec la taille des individus (Fig. II/II.34). L'ordination (N.M.S.) des
différentes classes de taille dans un espace à deux dimensions, représentation fidèle dans le plan (valeur
de stress faible : 0,07) des distances alimentaires réelles existant entre individus, montre des distances
parfois importantes entre points d'une même classe de LT mais l'ensemble des points s'ordonnent selon
un léger gradient de taille croissante. On observe une séparation nette entre classes de taille de moins et
de plus de 60 mm qui indique une modification importante de l'alimentation à cette taille. Avant, 60 mm
de LT, l'alimentation est relativement homogène entre individus d'une même taille et varie
essentiellement d'une classe de taille à l'autre. Elle est ensuite plus similaire entre classes de taille, avec
une très forte variabilité inter-individuelle. La classification par groupement agglomératif sépare trois
grands groupes d'alimentation : les juvéniles G0 de moins de 50 mm de LT (groupe A), les juvéniles G1+
de 50-60 mm (groupe B) et les adultes de plus de 60 mm (groupe C) (Fig II/II.35).

Stratégies alimentaires
113
Figure II/II.34 - Ordination (NMS, solution à 2 dimensions) des différentes classes de taille(LT, mm) d'A. laterna en fonction de leur régime alimentaire (Poids moyen des proies parestomac).
Figure II/II.35 - Classification par groupement agglomératif des différentes classes de taille d'A. laterna enfonction de leur régime alimentaire (poids moyens des proies par estomac). La flèche verticale rouge indique lepourcentage d'information (65 %) retenu comme valeur seuil pour l'établissement des groupes alimentaires (A,B et C).
Stress : 0,069-2
0
2
-2 0 2Axe 1
Axe
2
< 40 mm
40-50 mm
50-60 mm
60-70 mm
70-80 mm
80-90 mm
90-100 mm
> 100 mm
Taille (LT, mm)
A
B
C

Stratégies alimentaires
114
Non-Outlier MaxNon-Outlier Min75%25%MedianOutliers
Np
0
5
10
15
20
25
30
35
40
45
50
LT < 50 mm 50 - 60 mm LT > 60 mm
L'alimentation subit des modifications quantitatives et qualitatives d'un groupe à l'autre. Le
poids total par estomac augmente de façon significative avec la taille des poissons (Kruskal-Wallis : H(2,
N=722) = 90,82 ; P < 0,001). Cependant, cette augmentation de la ration alimentaire ne s'accompagne pas
d'une modification significative du nombre de proies par estomac (Kruskal-Wallis : H(2, N=722) = 4,64 ; P
= 0,098). Ce dernier est globalement plus élevé chez les individus de 50-60 mm (Fig. II/II.36) mais la
faiblesse des effectifs récoltés pour ce groupe (N = 18) et la très forte variabilité inter-individuelle
observée pour l'ensemble des individus quelle que soit leur taille, ne permettent pas de confirmer cette
tendance. L'accroissement du poids total par estomac avec la taille des individus résulte donc
principalement d'une augmentation du poids des proies consommées.
Figure II/II.36 - Représentation en Boites de Tukey des nombres de proies (Np, ind. est.-1) rencontrés dans lesestomacs des individus de chaque groupe de taille (LT, mm) chez A. laterna au large du Rhône.
Le poids individuel moyen de ces dernières (Pm) est positivement corrélé (r = 0,402; r2 = 0,161;
P < 0,001) avec la longueur totale (LT, mm) des individus (Fig II/II.37). Entre 50 mm et 100 mm de LT,
c'est à dire entre juvéniles G0 et adultes de l'espèce, le poids moyen des proies ingérées passe de 0,13 à
1,3 mg, soit une augmentation d'un facteur 10 environ. A 60-70 mm de LT, la gamme de poids des
proies consommées s'élargit brusquement, ce qui indique, à cette taille, un changement net
d'alimentation avec une ingestion de proies beaucoup plus volumineuses. Cependant, la limite inférieure
de la gamme de poids des proies est identique quelle que soit la taille (Pm = 0,1 mg), traduisant la
persistance de petite proies dans l'alimentation des adultes.
Non-marginaux maxNon-marginaux min75%25%
MédianePoints marginaux
Npmoy.= 8,4±0,9 Npmoy = 13,2±2,4 Npmoy =13,3±0,8

Stratégies alimentaires
115
Figure II/II.37 - Relation linéaire entre la taille (LT, mm) des individus d'A. laterna et le poids moyen (Pm,mg) des proies ingérées (r = 0,402; r2 = 0,161; P < 0,001).
La largeur du spectre alimentaire varie peu suivant la taille (Tableau II/II.8). La nature des
proies ingérées diffère cependant d'un groupe à l'autre. Chez les juvéniles G0 (LT < 50 mm), les
amphipodes (F = 0,76) et les cumacés (F = 0,51), proies préférentielles consommées par plus de 50%
des individus, représentent l'essentiel de l'alimentation, complétée de polychètes (F = 0,21),
principalement des carnivores (Nephtydae et Onuphidae). L'alimentation est globalement la même pour
les juvéniles G1+ de 50-60 mm mis à part une augmentation de l'importance des polychètes (F = 0,25 ;
Cp = 25,4%), surtout des suspensivores (Sabellidae) et des carnivores (Polynoidae), aux dépends des
cumacés. Chez les adultes (LT > 60 mm), l'alimentation est dominée par les mysidacés et les macroures.
Le fort pourcentage numérique des premiers (Cn = 40,7%) et le poids élevé des seconds (Cp = 39,1%)
en font deux catégories de proies préférentielles pour ce groupe de poissons qui ingère de façon
secondaire des polychètes (surtout des suspensivores : Ampharetidae, Terrebellidae), des amphipodes et
des phoronidiens.
L'essentiel des modifications ontogéniques de l'alimentation consiste en une diminution de
l'importance alimentaire des crustacés de petite taille (amphipodes, cumacés) au profit de ceux de
grande taille (macroures, mysidacés) et en une augmentation des quantités de polychètes consommés
(Fig II/II.38). Pour cette dernière catégorie de proies, le nombre de familles ingérées augmente avec la
taille des poissons par une consommation croissante d'espèces suspensivores qui supplantent les
carnivores dès 50 mm de LT. Il passe ainsi de 5 familles chez les G0 (LT < 50 mm), à 8 familles chez les
juvéniles de 50-60 mm, pour atteindre un maximum de 19 familles chez les adultes (LT > 60 mm).
0
1
2
3
4
5
6
0 10 20 30 40 50 60 70 80 90 100 110 120 LT(mm)
Pm (mg)
Pm = 0,0246 × LT – 1,0952

Stratégies alimentaires
116
Tableau II/II.8 - Variations de l'alimentation d'A. laterna en fonction de la taille. Q = coefficientalimentaire. "+" = valeur inférieure à 0,01 ; "" = valeur absente. LT = longueur totale des individus (enmm).
Juvéniles G0(LT < 50 mm)
Juvéniles G1+
(50 - 60 mm)Adultes
(LT > 60 mm)
Estomacs pleins 58 65 596
LT moy (min. - max.) 43,5 (20-49) 56,5 (50-59) 96,0 (60-132)
PROIES Q Q Q
AnnélidesPolychètes 38,8 47,3 89,3
CrustacésCopépodes 0,5 18,2 0,1Leptostracés 0,6 Mysidacés 0,8 4,6 425,0Cumacés 228,4 81,7 0,8Amphipodes 3615,4 3356,6 92,1Macroures (crevettes) 11,0 10,1 208,3Anomoures 0,2Brachyoures 0,3 5,4 19,9Crustacés indét. 0,9 +
MollusquesBivalves 1,6 0,3 8,9
EchinodermesOphiures +
Phoronidiens 97,3Poissons 7,8 0,4 +Proies indét. + +
Figure II/II.38 - Variations de la composition pondérale (Cp) du régime alimentaire d'A. laterna au large duRhône en fonction de la taille des individus (LT, mm).
0%
25%
50%
75%
100%
LT < 50 mm 50-60 mm LT > 60 mmClassesde taille
Cp
polychètes bivalves amphipodes macroures mysidacés brachyoures anomoures
leptostracés cumacés copépodes ophiures poissons phoronidiens

Stratégies alimentaires
117
Variations spatio-temporelles
L'importance des effectifs d'A. laterna capturés (N > 2000 individus) a rendu possible l'analyse
croisée des effets de la taille des individus, de la profondeur et de la saison sur l'alimentation de cette
espèce, à condition de regrouper entre eux les juvéniles G0 (LT < 50 mm) et G1+ (50-60 mm) qui
présentent des régimes alimentaires proches. Les variations spatio-temporelles de l'alimentation ont
ainsi été étudiées de façon précise tant pour les juvéniles (LT ≤ 60 mm) que pour les adultes (LT > 60
mm) de l'espèce.
Variations de l'intensité de la nutrition
Chez A. laterna, l'intensité de la nutrition ne subit pas de modification significative suivant la
profondeur (IR, Kruskall-Wallis : H(2, N=319) = 4,02 : P > 0,13). Elle varie par contre au cours de l'année.
L'indice de réplétition stomacal diffère suivant la saison (Kruskall-Wallis : H(2, N=711) = 44,25 ; P <
0,001). Il est significativement plus faible (P < 0,001 ; Annexe 26) au printemps (IRmoy= 1,2 ± 0,06)
mais ne diffère pas de façon significative entre l'été (IRmoy = 2,0 ± 0,1) et l'hiver (IRmoy.= 1,9 ± 0,9),
périodes où il est très variable d'un individu à l'autre (Fig. II/II.39). La prise de nourriture est cependant
plus importante en été, avec un nombre minimal d'estomacs vides à cette saison.
Figure II/II.39 - Représentation en Boites de Tukey des indices de réplétion stomacale (IR) observés àchaque saison chez A. laterna au large du Rhône. CV = coefficient de vacuité (% d'estomacs vides).
IR
0
1
2
3
4
hiver printemps été
Non-marginaux maxNon-marginaux min
75%25%
Médiane
(CV = 24,5%) (CV = 26,9%) (CV = 16,2%)

Stratégies alimentaires
118
30 - 50 m
70 - 100 m
PRIN
TE
MPS
ET
É
0- 20 m
Juvéniles LT < 60 m
m(N
= 19)
leptostracés3,4%
crust. indét.0,3%
copépode s4,5%
polychètes21,5%
bivalves7,2%
amphipodes
15,8 %
cumacés
47,3%
Juvéniles LT < 60 m
m (N
= 58)
mysidacés2,1%
brachyoures1,8%
macroure s9,6%
cumacé s
6,8%
crust. indét.0,9%
poissons10,2%
amphipodes
57,0 % bivalves1,4%
polychètes9,7%
Adultes L
T > 60 mm
(N = 24)
bivalves1,4%
brachyoures18,4 %
macroures6,6%
cumacé s
1,5%
crust. indét.3,4%
amphipodes14,0%
mysidacés2,3%
phoronidiens4,8%
polychètes47,5%
Adultes L
T > 60 mm
(N = 24)
phoronidiens11,9%
crust. indét.4,3%
macroure s0,5%
cumacés
5,4%
bivalves3,8%
amphipodes
12,4%
mysidacés8,1%
brachyoures1,1%
polychètes51,9 %
(N = 148)
polychètes45,9 %
macroures34,4%
cumacés
0,6%
mysidacé s1,6%
crust. indét.1,5%
amphipodes3,4%
brachyoures10,5%
bivalves1,9%
Adultes L
T > 60 mm
(N = 27)
amphipodes2,9%
polychètes4,0%
macroures2,9%
crust. indét .3,7%
mysidacés85,6%
leptostracés0,8%
Adultes L
T > 60 mm
(N = 133)
amphipodes3,5%
macroures
56,1 %
mysidacés21,0%
brachyoures9,1%
bivalves0,2%
cumacés
0,4% copépode s0,6%
crust. indét.1,2%
poissons0,9%
polychètes6,9%
Adultes L
T > 60 mm
(N = 11)
mysidacés37,5 % polychètes
19,6%
crust. indét .2,5%
brachyoures35,5%
macroures4,8%
HIV
ER
Adultes L
T > 60 mm
(N = 83)
polychètes30,4%
bivalves6,7%
phoronidiens12,4%
ophiures0,1%
cumacés
1,3%
brachyoures4,5%
mysidacés
4,1%
amphipodes
21,1%
macroures17,2%
Adultes L
T > 60 m
m (N
= 110)
polychètes4,7%
bivalves0,3%
poissons0,3%
crust. indét.3,0%
cumacés
0,1%
brachyoures15,1%
mysidacés5,3%
amphipodes2,8%
anomoures
3,6%
macroures64,8%
Adultes L
T > 60 mm
(N = 104)
macroures24,6%
amphipodes3,3%
mysidacés44,6%
copépodes1,2%
brachyoures16,2%
crust. indét.4,7%
polychètes5,4%
Adultes L
T > 60 mm
Figure II/II.40 - Variations de l'alim
entation (pourcentages pondéraux des proies ingérées) d'A. laterna en fonction de la saison, de la profondeurr et de la taille des individus(longueur totale L
T , en mm
). N = nom
bre d'estomacs pleins analysé pour chaque groupe de poissons. D
ans chaque cas, les proies surlignées en gris sont rencontrées dans plusde 20%
des estomacs et/ou représentent plus de 10%
du poids total des proies ingérées.

Stratégies alimentaires
119
Taille des individus et modifications spatio-temporelles de l'alimentation
Chez A. laterna, les variations spatio-temporelles de l'alimentation dépendent de la taille des
individus (Fig. II/II.40). Elles concernent essentiellement les adultes (LT > 60 mm), seuls présents de 0 à
100 m de profondeur et capturés en hiver. Chez les juvéniles (LT ≤ 60 mm), essentiellement présents à 0-
20 m au printemps et en été, la composition de l'alimentation diffère statistiquement (P < 0,001; Annexe
27) entre ces deux saisons, avec une ingestion accrue de cumacés au printemps et d'amphipodes en été.
Chez les adultes, le régime alimentaire se modifie de façon significative (P < 0,05 ; Annexe 28) entre
l'hiver, le printemps et l'été et est statistiquement diffèrent (P < 0,001; Annexe 28) sur les trois tranches
bathymétriques étudiées. Il diffère significativement, quelle que soit la saison, entre les fonds sableux
côtiers (0-20 m) et les vases terrigènes plus profondes (30-100 m) (P < 0,05 ; Annexe 28).
L'alimentation des adultes est, par contre, similaire à 30-50 m et à 70-100 m en été et ne varie de façon
significative entre ces deux profondeurs qu'en hiver (P < 0,001) et au printemps (P < 0,05). A 0-20 m,
seule tranche bathymétrique où l'ensemble des classes de taille coexistent, le régime alimentaire des
adultes est stable tout au long de l'année (P > 0,24; Annexe 28) et diffère significativement de celui des
juvéniles au printemps et en été (P < 0,001; Annexe 27). A cette profondeur, l'alimentation des juvéniles
est basée sur les petits crustacés (cumacés, amphipodes) et celle des adultes sur les polychètes et les
crustacés de plus grande taille (macroures, brachyoures, mysidacés). La composition du régime
alimentaire des adultes est plus variable sur les vases terrigènes (Annexe 28). A 30-50 m, l'alimentation
diffère de façon significative suivant la saison (P < 0,05) et est dominée par les polychètes et les
macroures au printemps, les mysidacés en été et les macroures en hiver. A 70-100 m, l'alimentation
comprend de fortes quantités de mysidacés quelle que soit la saison mais diffère de façon significative
(P < 0,05) entre l'été (mysidacés) et la période printemps-hiver (mysidacés, macroures, brachyoures).
II/II.3.4.3 - Discussion sur l'alimentation d'Arnoglossus laterna
Comportement alimentaire
Au large du Rhône, Arnoglossus laterna présente une activité nutritionnelle essentiellement
diurne, avec un pic unique d'alimentation en fin d'après-midi. Ces observations sont en accord avec la
littérature, cette espèce étant décrite comme s'alimentant surtout de jour et présentant un indice de
réplétion stomacal maximal l'après-midi (Avsar, 1994). La population d'A. laterna étudiée consomme
un nombre élevé (bien que variable) de proies de taille réduite. Ces observations sont en accord avec les
travaux préexistants (Tito de Morais, 1983 ; Tito de Morais et Bodiou, 1984 ; Darnaude et al., 2001) et
s'expliquent par la taille de la bouche et la morphologie de l'appareil digestif de l'espèce, adaptés à
l'ingestion de proies de petite taille (De Groot, 1969).
Le régime alimentaire global d'A. laterna au large du Rhône est dominé par les crustacés
(mysidacés, amphipodes, macroures), mais l'espèce ingère également d'importantes quantités de

Stratégies alimentaires
120
polychètes (suspensivores et carnivores) et, ponctuellement, de phoronidiens. Ce régime alimentaire est
surtout représentatif de l'alimentation des adultes de l'espèce (plus de 80% des estomacs pleins analysés)
et correspond, dans ses grandes lignes, au régime alimentaire observé pour l'espèce sur l'ensemble de
son aire de répartition géographique (Sorbe, 1972 ; Gibson et Ezzi, 1980 ; Tito de Morais et Bodiou,
1984 ; Rogers, 1991 ; Avsar, 1994). Cependant, la consommation de juvéniles de poissons par les
adultes, décrite par Gibson et Ezzi (1980), Rogers (1991) et Avsar (1994), est réduite au large du Rhône
(Darnaude et al., 2001; présente étude). En revanche, l'ingestion de phoronidiens, importante entre 0 et
20 m de profondeur, semble limitée à notre zone d'étude.
Modifications de l'alimentation
Chez A. laterna, la prise de nourriture est réduite au printemps mais élevée en hiver et surtout en
été, période où elle est maximale. Ces observations concordent avec les données disponibles sur la
biologie et la physiologie de l'espèce. Arnoglossus laterna est peu sensible aux baisses de température
hivernales (Tito de Morais, 1983) mais sa reproduction, précédée d'une période d'alimentation intense
(associée à la vitellogénèse), s'accompagne, au moment de la ponte, d'une diminution de l'activité
nutritionnelle chez les adultes (Deniel, 1981, 1983). L'espèce se reproduisant de mars à août en
Méditerranée (Giovanardi et Piccinetti, 1984), ceci explique le minimum d'alimentation observé au
printemps.
La composition du régime alimentaire d'A. laterna varie selon la taille des individus, la
profondeur et la saison, les deux premiers facteurs étant les plus importants (Fig. II/II.41).
L'alimentation des adultes (LT > 60 mm) s'individualise nettement de celles des juvéniles G0 (LT < 50
mm) et G1+ (50-60 mm), qui sont, elles, très proches. Ce phénomène est lié au poids et à la nature des
proies consommmées qui diffèrent de façon importante entre juvéniles et adultes. Le régime alimentaire
des premiers est essentiellement constitué de crustacés de petite taille (cumacés, amphipodes) alors que
celui des seconds, plus diversifié, comprend de fortes quantités de polychètes et de crustacés plus
volumineux (macroures, mysidacés, brachyoures). Ces observations sont en accord avec celles d'Avsar
(1994), Tito de Morais (1983) et Tito de Morais et Bodiou (1984) qui confirment l'existence, chez A.
laterna, d'une modification de la taille et de la nature des proies ingérées entre les juvéniles et les
adultes, accompagnée d'un élargissement du régime alimentaire.
Les variations spatio-temporelles de l'alimentation concernent surtout les adultes de l'espèce
chez qui la composition du régime alimentaire varie essentiellement avec la profondeur. La variabilité
de l'alimentation, réduite chez les juvéniles (LT ≤ 60 mm), présents uniquement à 0-20 m, augmente de
façon importante chez les adultes (LT > 60 mm), du fait d'une répartition bathymétrique plus large (Fig.
II/II.41). Le régime alimentaire de ces derniers est principalement basé sur l'ingestion d'amphipodes et
de phoronidiens à 0-20 m, de macroures et de polychètes à 30-50 m et de mysidacés à 70-100 m. La

Stratégies alimentaires
121
P
P
EE E
E
P
PE
E
H
H
H
H
PPP
P
P
P
E
H
H HH
H
PP
P
P
P
P
EE
H
H
H
H
H
H
-2
0
2
-2 0 2Axe 1
Axe
2
50-60 mm
70 - 100 m
30 - 50 m
0 - 20 m
Profondeur
juvéniles G0(LT < 50 mm)
adultes(LT > 60 mm)
juvéniles G1 +
50-60 mm
Stade de vie
Stress : 0,090
LT < 50 mm
LT > 60 mm
variabilité saisonnière de l'alimentation est réduite et concerne surtout les individus présents sur les
fonds vaseux de 30-100 m de profondeur où le régime alimentaire estival (mysidacés) se distingue des
alimentations printanière et hivernale quelle que soit la tranche bathymétrique concernée (30-50 m ou
70-100 m). Elle est maximale à 30-50 m de profondeur où la composition du régime alimentaire diffère
en outre entre l'hiver (macroures) et le printemps (polychètes et macroures). Quel que soit le stade de
vie benthique, le régime alimentaire reste cependant dominé par les crustacés (cumacés, amphipodes,
macroures et/ou mysidacés). Ceux-ci sont ingérés de façon préférentielle quelles que soient la période
de l'année et/ou la tranche bathymétrique et constituent, quantitativement parlant, l'essentiel des proies
consommées par A. laterna au large du Rhône.
Figure II/II.41 - Variations de l'alimentation de A. laterna en fonction de la profondeur (0-20 m, 30-50m, 70-100 m), de la saison (P = printemps, E = été, H = hiver), et de la taille (LT, mm) des individus.Ordination (N.M.S., solution à 2 dimensions) réalisée sur les poids moyen des proies par estomac.

Stratégies alimentaires
122
II/II.3.5 - CITHARUS LINGUATULA
II/II.3.5.1 - Alimentation globale
L'alimentation de C. linguatula a été étudiée à partir des contenus stomacaux de 174 individus
de LT comprise entre 63 et 240 mm. Cette espèce s'alimente essentiellement pendant le jour. L'indice de
réplétion stomacal, bien que relativement important quelle que soit la période (IR > 1,4 ; Fig. II/II.42),
est significativement plus élevé (Mann-Whitney : U = 3024,5 ; P < 0,05) le jour (IRmoy.= 2,1 ± 0,2) que
la nuit (IRmoy.= 1,6 ± 0,2). L'activité alimentaire, constante au cours de la matinée, augmente pendant
l'après-midi pour atteindre un maximum (IR = 3,7) entre 16:00 et 18:00 heures. L'indice de réplétion
décroît ensuite au coucher du soleil et reste faible pendant la nuit. Au large du Rhône, C. linguatula
s'alimente donc de façon ininterrompue sur toute la durée du nycthémère (bien que plus intensément de
jour), avec un pic d'alimentation maximale unique, situé en milieu d'après-midi.
Figure II/II.42 - Variations nycthémérales de l'indice de réplétion stomacal moyen (IR) chez C.linguatula (juin 2000; heure locale = heure GMT + 1). CS = coucher du soleil; LS = lever du soleil.
Sur les 174 estomacs de C. linguatula analysés, 52 ne contenaient pas de nourriture (CV =
29,9%). Le régime alimentaire global de l'espèce a donc été établi à partir des contenus stomacaux de
122 individus de LT comprise entre 63 et 237 mm, dont 12% de juvéniles G0, 46% de juvéniles G1+ et
42% d'adultes. Il est caractérisé par l'ingestion d'un nombre variable de proies de tailles très différentes.
Le nombre et le poids des proies par estomac fluctuent respectivement autour de 2 proies et 5,1 mg
(valeurs médianes, Fig II/II.43 a et b). Ils sont cependant très variables (1 à 100 individus* et 0,2 à 172,0
mg∗ par estomac), ce qui explique les moyennes élevées (Npmoy.= 9,6 ± 1,8 ; Ppmoy.= 30,7 ± 4,2)
obtenues pour l'espèce alors que la majorité des individus ingère de 1 à 8 proies pour un poids total par
estomac de 0,8 à 43,1 mg.
∗ valeurs extrêmes ne concernant que quelques individus exceptionnels et non représentées sur la Figure II/II.43

Stratégies alimentaires
123
Figure II/II.43 - Représentation en Boites de Tukey des nombres (a) et des poids totaux (b) des proies par estomacrencontrés chez C. linguatula au large du Rhône.
Le régime alimentaire de C. linguatula au large du Rhône est peu diversifié (Tableau II/II.9). Il
est largement dominé par les crustacés et les poissons. Avec une fréquence d'ingestion et un
pourcentage numérique élevés, les mysidacés, essentiellement des individus de Leptomysis gracilis, sont
les proies préférentielles de l'espèce. En raison de leur pourcentage pondéral élevé, les poissons
(essentiellement de petits juvéniles de Gobiidae avalés entiers) tiennent également une place
prépondérante dans l'alimentation malgré une fréquence d'ingestion moindre. Les macroures,
essentiellement des juvéniles ou adultes du genre Alpheus, constituent la seule catégorie de proies
d'importance secondaire de l'espèce, les autres proies ingérées (polychètes, juvéniles de bivalves) étant
accidentelles.
Tableau II/II.9 - Régime alimentaire global de C. linguatula au large du Rhône. F = fréquence ; N =nombre de proies ; Cn = pourcentage numérique ; P = poids sec des proies (mg) ; Cp = pourcentagepondéral ; Q = coefficient alimentaire des proies (Q = Cn × Cp). "+" = valeur inférieure à 0,01.
F N Cn P Cp Q
AnnélidesPolychètes 0,03 3 0,36% 64,70 0,97% 0,35
CrustacésMysidacés 0,61 752 89,74% 498,81 7,48% 671,67Macroures (crevettes) 0,25 33 3,94% 1004,36 15,07% 59,35
MollusquesBivalves 0,01 1 0,12% 2,58 0,04% +
Poissons 0,39 47 5,61% 5045,40 75,71% 424,61Proies indét. 0,02 2 0,23% 48,46 0,73% 0,17
Total proies 838 100,0% 6664,31 100,0%
Non marginaux maxNon marginaux min
75%25%
Médiane
Points marginaux
(a) Np (b) Pp en mg
0
5
10
15
20
25
30
01020304050607080
90100110120130140150160170

Stratégies alimentaires
124
Figure II/II.44 - Ordination (N.M.S., solution à 2 dimensions) des différentes classes de taille(LT, mm) de C. linguatula en fonction de leur régime alimentaire (poids moyen des proies parestomac).
Figure II/II.45 - Classification par groupement agglomératif (Cluster Analysis) des différentes classes detaille de C. linguatula en fonction de leur régime alimentaire (poids moyens des différentes proies ingéréespar estomac). La flèche verticale rouge indique le pourcentage d'information (65 %) retenu comme valeurseuil pour l'établissement des groupes alimentaires (A, B et C).
A
B
C
Taille (LT, mm)
Stress : 0,059-2
0
2
-2 0 2
Axe 1
Axe
2
< 80
80-100
100-120
120-140
140-160
160-180
180-200
> 200

Stratégies alimentaires
125
II/II.3.5.2 - Modifications du régime alimentaire
Variations en fonction de la taille
La composition de l'alimentation se modifie au cours de la croissance mais est très variable d'un
individu à l'autre. L'ordination (N.M.S.) des différentes classes de taille fournit une représentation fidèle
(valeur de stress faible : 0,059), dans le plan proposé, des distances alimentaires réelles entre individus,
distances parfois importantes entre points d'une même classe de LT (Fig. II/II.44). L'existence d'une
séparation nette entre individus de moins et de plus de 100 mm indique une modification du régime
alimentaire à cette taille. La composition de l'alimentation est ensuite plus homogène avec un léger
gradient de taille croissante traduisant une modification progressive de l'alimentation avec la taille des
individus. La classification par groupement agglomératif sépare trois grands groupes de taille
d'alimentation distincte : les juvéniles G0 de LT < 100 mm (groupe A), les juvéniles G1+ de 100-160
mm (groupe B) et les adultes de LT > 160 mm (groupe C) (Fig. II/II.45).
La nature des proies ingérées varie peu d'un groupe de LT à l'autre (Tableau II/II.10). Quelle que
soit la classe de taille, les mysidacés (Cn > 88% ; F > 0,43) et les poissons (Cp > 50%), constituent
l'essentiel de l'alimentation (Fig. II/II.46). Cependant, leurs importances relatives diminuent au cours de
la croissance, principalement entre juvéniles G0 (LT < 100 mm) et G1+ (100-160 mm) pour les
mysidacés et entre juvéniles G1+ et adultes (LT > 160 mm) pour les poissons. L'importance alimentaire
des macroures augmente avec la taille des individus : ce sont des proies occasionnelles chez les
juvéniles (LT ≤ 160 mm), et secondaires chez les adultes (LT > 160 mm).
Tableau II/II.10 - Variations de l'alimentation de C. linguatula en fonction de la taille. Q =coefficient alimentaire. "+" = valeur inférieure à 0,01 ; "" = valeur absente. LT = longueur totaledes individus (en mm).
LT < 100 mm 100 < LT < 160 mm LT > 160 mm
Estomacs pleins 15 56 51
LT moy. (min. – max.) 77,1 (63-99) 127,7 (100-159) 200,6 (161-237)
PROIES Q Q Q
Annélides :Polychètes + +
Crustacés :Mysidacés 4297,9 1180,0 863,4Macroures (crevettes) 4,2 14,8 44,3
Mollusques :Bivalves +
Poissons 292,0 604,2 228,1Proies indet. 0,9 +

Stratégies alimentaires
126
Figure II/II.46 - Variations de la composition pondérale (Cp) du régime alimentaire de C. linguatula aularge du Rhône en fonction de la taille des individus (LT, mm).
Les polychètes (Spionidae et Sternaspidae) sont uniquement ingérés par les individus de plus de
100 mm (juvéniles G1+ et adultes) et les bivalves par les juvéniles G1+ (100-160 mm). Ces deux
catégories de proies restent cependant accidentelles quel que soit le groupe de LT. L'essentiel des
modifications ontogéniques de la composition du régime alimentaire concerne donc les mysidacés, les
poissons et les macroures avec, à partir de 100 mm de LT, une forte réduction de l'importance
pondérales des mysidacés au profit des poissons et, au delà de 160 mm, une nette augmentation de la
proportion de macroures ingérés.
La taille des proies consommées augmente par contre au cours de la croissance. Le poids
individuel moyen des proies ingérées est positivement corrélé (r = 0,438; r2 = 0,192; P < 0,001) avec la
longueur totale des individus (Fig. II/II.47). Entre 80 et 200 cm de LT, c'est à dire entre juvéniles G0 et
adultes de l'espèce, il passe de 1,4 à 54,1 mg, soit une augmentation d'un facteur 40 environ.
L'élargissement avec la taille de la gamme de poids individuel des proies consommées (elle passe de
0,1-1,4 mg chez les juvéniles G0 à 0,1-171,2 mg chez les adultes) indique une ingestion de proies de
plus en plus volumineuses mais également la persistance des proies de petite taille dans l'alimentation
des adultes. Le nombre de proies par estomac, très variable d'un individu à l'autre (surtout chez les
adultes de LT > 160 mm), ne subit pas de modification significative d'un groupe de LT à l'autre (Kruskal-
Wallis : H(2,N=122) = 2,96 ; P > 0,23) (Fig. II/II.48). L'augmentation significative de la ration alimentaire
(poids total par estomac) avec la taille des individus (Kruskal-Wallis : H(2,N=122) = 292,96 ; P < 0,001)
résulte donc principalement d'une augmentation de la taille des proies ingérées.
0%
25%
50%
75%
100%
LT < 100 mm 100-160 mm LT > 160 mmClassesde taille
Cp
polychètes bivalves macroures mysidacés poisson

Stratégies alimentaires
127
Non-Outlier MaxNon-Outlier Min75%25%MedianOutliers0
5
10
15
20
25
30
35
40
LT < 100 mm 100-160 mm LT > 160 mm
Figure II/II.47 - Relation linéaire entre la taille (LT, mm) des individus de C. linguatula et le poids moyen(Pm, mg) des proies ingérées (r = 0,438; r2 = 0,192; P < 0,001).
Figure II/II.48 - Représentation en Boites de Tukey des nombres de proies (Np, ind. est.-1) rencontrés dansles estomacs des individus de chaque groupe de taille (LT, mm) chez C. linguatula au large du Rhône.
0
20
40
60
80
100
120
140
160
180
200
220
240
60 80 100 120 140 160 180 200 220 240 260 LT(mm)
Pm (mg)Pm = 0,439 × LT – 33,745
Non-marginaux maxNon-marginaux min75%25%
MédianePoints marginaux
Npmoy.= 5,7±0,9 Npmoy = 5,2±1,0 Npmoy =13,7±3,5

Stratégies alimentaires
128
Figure II/II.50 - Variations de l'alim
entation (pourcentages pondéraux des proies ingérées) de C. linguatula en fonction de la saison et de la taille des individus(longueur totale L
T , en mm
). N = nom
bre d'estomacs pleins analysé pour chaque groupe de poissons. D
ans chaque cas, les proies surlignées en gris sontrencontrées dans plus de 20%
des estomacs et/ou représentent plus de 10%
du poids total des proies ingérées.
Juvéniles G0 L
T < 100 mm
Juvéniles G1 + 100-160 m
mA
dultes LT > 160 m
m
Printemps
Eté
Hiver
(N = 11)
mysidacés24,9%
poissons72,5% m
acroures2,6%(N
= 20) polychètes
0,4%bivalves
0,6%
poissons75,6%
mysidacés7,8%
macroures15,6%
(N = 15)
indét.0,3%
poissons38,0%
mysidacés6,9%
macroures54,9%
(N = 26)
mysidacés11,9%
macroures0,7%
poissons87,4%
(N = 11)
macroures54,7%
mysidacés43,5%
poissons1,8%
(N = 13)
mysidacés13,2%
poissons84,1% indét.
2,6%(N = 19)m
acroures3,9%
poissons94,3%
polychètes1,8%

Stratégies alimentaires
129
Non-marginaux max Non-marginaux min
75% 25%
Médiane
CV = 37,5% CV = 18,2% CV = 10,3%
IR
0
1
2
3
4
hiver printemps été
Variations spatio-temporelles de l'alimentation
Les fluctuations spatio-temporelles des effectifs de C. linguatula ont empêché l'analyse croisée
des effets de la taille des individus, la profondeur et la saison sur l'alimentation de C. linguatula, le
nombre d'estomacs pleins par stade de vie benthique étant parfois inférieur à dix. Les variations
saisonnières et bathymétriques de l'alimentation ont donc été traitées séparément pour cette espèce.
Variations saisonnières
L'intensité de nutrition de C. linguatula, bien que relativement élevée (IRmoy.> 1,6) tout au long de
l'année, diffère suivant la saison (Kruskall-Wallis : H(2, N=102) = 12,44 ; P < 0,01) et augmente de façon
progressive de l'hiver à l'été (Annexe 29). En hiver, l'indice de réplétion stomacal est globalement faible
(IRméd.= 1,5) bien que très variable d'un individu à l'autre (Fig. II/II.49). Il augmente au printemps
(IRmoy.= 2,0 ± 0,3) et est maximal en été (IRmoy.= 3,0 ± 0,2). Le coefficient de vacuité, réduit au
printemps et en été (CV < 19%), est relativement élevé (CV > 37%) en hiver, ce qui confirme la
diminution de l'intensité de nutrition de cette espèce pendant la saison froide.
Figure II/II.49 - Représentation en Boites de Tukey des indices de réplétion stomacale (IR) observés àchacune des trois saisons étudiées chez C. linguatula au large du Rhône. CV = coefficient de vacuité (%d'estomacs vides).
La composition du régime alimentaire de C. linguatula varie au cours de l'année. Les
modifications saisonnières dépendent de la taille des individus (Fig. II/II.50). Les juvéniles G0 (LT <
100 mm) n'ayant été capturés qu'au printemps, les variations mises en évidence ne concernent que les
individus de plus de 100 mm (juvéniles G1+ et adultes). Pour ces derniers, les modifications
saisonnières dépendent de la taille des individus : le régime alimentaire des juvéniles G1+ (100-160 mm)
reste globalement stable tout au long de l'année (P > 0,08 ; Annexe 30) alors que celui des adultes (LT >
160 mm) varie fortement d'une saison à l'autre (P < 0,001; Annexe 30) (Fig. II/II.50).

Stratégies alimentaires
130
Figure II/II.51 - Variations de l'alim
entation (pourcentages pondéraux des proies ingérées) de C. linguatula en fonction de la profondeur et de la taille desindividus (longueur totale L
T , en mm
). N = nom
bre d'estomacs pleins analysé pour chaque groupe de poissons. D
ans chaque cas, les proies surlignées en gris sontrencontrées dans plus de 20%
des estomacs et/ou représentent plus de 10%
du poids total des proies ingérées.
Juvéniles G0 L
T < 100 mm
0- 20 m
30 - 50 m
70 - 100 m
(N = 28)
macroures15,6%
mysidacés7,8%
poissons75,6%
bivalves0,6%
polychètes0,4%
(N = 13)
indét. 0,3%
poissons37,9
mysidacés6,9%
macroures54,9%
(N = 11)
mysidacés 29,8%
macroures5,4%
poissons 64,8%
(N = 36)
polychètes0,3%
indét. 0,7%
poissons73,3%
mysidacés
19,7%
macroures5,7%
bivalves0,4%
(N = 30)
macroures24,2%m
ysidacés9,0%
poissons 64,5%
polychètes2,2%
Juvéniles G1
+ 100-160 mm
Adultes L
T > 160 mm
?

Stratégies alimentaires
131
Quelle que soit la saison, l'alimentation diffère de façon significative d'un groupe de LT à l'autre
(P < 0,05 ; Annexe 30 et 31). Au printemps, période où toutes les classes de taille sont présentes dans
les captures, l'alimentation des adultes (LT > 160 mm) est dominée par les macroures et les poissons.
Les régimes alimentaires des juvéniles G0 (LT < 100 mm) et G1+ (100-160 mm), très proches du point
de vue qualitatif, sont caractérisés par une nette prépondérance des poissons (Cp > 72%) (Fig. II/II.50).
Cependant, les alimentations de ces deux classes de taille se séparent nettement du point de vue
quantitatif, le poids total par estomac différant de façon significative (Mann-Whitney : U(15,10) = 1025,4 ;
P < 0,001) à cette saison entre juvéniles de moins de 100 mm (Ppmoy.= 0,7 ± 0,2 mg) et de 100-160 mm
(Ppmoy.= 19,9 ± 3,6 mg). En été et en hiver, périodes où seuls les individus de plus de 100 mm sont
présents, le régime alimentaire des juvéniles G1+ (100-160 mm) reste dominé par les poissons (Q >
820), et les mysidacés (Q > 650) alors que le régime alimentaire des adultes (LT >160 mm) diffère d'une
saison à l'autre, avec une dominance de poissons (Q = 3572) en hiver et de mysidacés (Q = 1028) et
macroures (Q = 572) en été.
Modifications en fonction de la profondeur
L'intensité de nutrition de C. linguatula ne se modifie pas avec la profondeur : l'indice de
réplétion stomacal (IR) ne diffère pas de façon significative suivant la tranche bathymétrique fréquentée
(Kruskall-Wallis : H(1, N=76) = 0,052 ; P > 0,81). Par contre, la composition du régime alimentaire se
modifie avec la profondeur. Les modifications bathymétriques de l'alimentation dépendent de la taille
des individus (Fig. II/II.51). Le nombre réduit de juvéniles G0 (LT < 100 mm) présents à 30-50 m n'a
pas permis d'étudier les modifications bathymétriques de l'alimentation de ce groupe d'individus, surtout
capturé à 70-100 m de profondeur. Pour les individus de plus de 100 mm de LT (juvéniles G1+ et
adultes), le régime alimentaire est similaire quel que soit le groupe ou la profondeur considérés, mis à
part pour les adultes (LT > 160 mm) à 30-50 m (P < 0,05 ; Annexe 32).
A 70-100 m, le régime alimentaire diffère statistiquement suivant la taille des poissons (P <
0,05 ; Annexe 33). Chez juvéniles G0 (LT < 100 mm), l'importance alimentaire des mysidacés est
nettement plus élevée (Q = 897) que chez les juvéniles G1+ et les adultes (Q < 202) dont le régime
alimentaire est largement dominé par les poissons (Q > 5700). Sur les fonds de 30-50 m de profondeur,
le régime alimentaire diffère significativement (P < 0,05; Annexe 32), entre les juvéniles G1+ de 100-
160 mm (poissons) et les adultes de LT > 160 mm (macroures et poissons).
II/II.3.5.3 - Discussion sur l'alimentation de C. linguatula
Comportement alimentaire
Au large du Rhône, C. linguatula présente une activité alimentaire essentiellement diurne, avec
un pic d'alimentation maximale en milieu d'après-midi. Ces observations sont en accord avec les travaux

Stratégies alimentaires
132
précédents, C. linguatula étant décrite comme un chasseur à vue s'alimentent pendant le jour (Belghyti
et al., 1993 ; Belghyti et al., 1995). L'alimentation de la population étudiée est caractérisée par un poids
total des proies par estomac élevé et un nombre variable de proies de tailles très différentes. Ce
phénomène reflète l'ingestion alternative, soit d'un nombre très élevé de proies de petite taille, soit d'un
nombre réduit de proies volumineuses. Ces données sont conformes à celles de la littérature : Citharus
linguatula est décrite comme un prédateur vorace consommant généralement un nombre très élevé de
proies de petite taille (Belghyti et al., 1993) mais ingérant également de façon régulière des proies
volumineuses (de 11 à 85% de sa taille) (Jardas, 1984).
Le régime alimentaire obtenu pour C. linguatula au large du Rhône, largement dominé par les
crustacés et les poissons, est en accord avec les résultats des travaux préexistants sur l'alimentation de
cette espèce, tant en Atlantique (Belghyti et al., 1993) qu'en Méditerranée (Jardas, 1984 ; Redon et al.,
1994). Par contre, les brachyoures et les céphalopodes, ingérés de façon occasionnelle dans les autres
sites (Jardas, 1984 ; Belghyti et al., 1993 ; Redon et al., 1994 ; Belghyti et al., 1995), ne sont pas
consommés au large du Rhône, où ils sont remplacés par des polychètes et des bivalves. D'une façon
générale, l'espèce est décrite comme un carnivore relativement sténophage, se nourrissant à partir d'un
nombre réduit de catégories de proies necto-benthiques (Belghyti et al., 1993 ; Belghyti et al., 1995), ce
qui correspond à nos observations.
Modifications de l'alimentation
La prise de nourriture de C. linguatula est importante au cours des trois saisons étudiées (hiver,
printemps, été), ce qui confirme les résultats précédents : l'activité nutritionnelle de l'espèce, intense tout
au long de l'année, ne se réduit véritablement que pendant sa période de frai (Jardas, 1984 ; Belghyti et
al., 1993 ; Redon et al., 1994). Citharus linguatula se reproduisant au début de l'automne en
Méditerranée, la réduction de l'alimentation correspondante n'a pu être décelée au cours du présent
travail. On observe néanmoins, pour cette espèce une légère augmentation de l'activité trophique de
l'espèce de l'hiver à l'été qui peut être attribuée à l'accumulation croissante de réserves par les femelles
en vue de la vitellogénèse.
La composition du régime alimentaire varie selon la taille des individus, la saison et la
profondeur, les deux premiers facteurs étant les plus déterminants (Fig. II/II.52). Quel que soit le stade
de vie benthique, l'alimentation de C. linguatula au large du Rhône est dominée par les poissons et les
mysidacés. Cependant les juvéniles G0, les juvéniles G1+ et les adultes de l'espèce présentent des
alimentation distinctes (Fig. II/II.52), du fait d'une forte augmentation du poids des proies consommées
et des quantités de macroures ingérées avec la taille des individus. Ces résultats sont en accord avec les
travaux préexistants sur l'alimentation de l'espèce qui notent, avec la croissance des individus, une
augmentation intraspécifique de la taille des proies ingérées (Belghyti et al., 1993) accompagnée d'une

Stratégies alimentaires
133
diminution de l'importance alimentaire des mysidacés au profit des poissons et des macroures, proies
plus volumineuses (Belghyti et al., 1993 ; Redon et al., 1994).
Les modifications spatio-temporelles de l'alimentation de C. linguatula sont peu marquées au
large du Rhône et dépendent essentiellement de la taille des individus. La composition du régime
alimentaire des juvéniles G0 (LT < 100 mm) est très homogène (poissons et mysidacés) quelle que soit
la profondeur. Chez les juvéniles G1+ (100-160 mm), les régimes alimentaires des deux tranches
bathymétriques se chevauchent (poissons, mysidacés) et seule la saison semble avoir une influence,
avec une individualisation de l'alimentation estivale à 30-50 m, due à une augmentation de l'importance
pondérale des macroures aux dépends des mysidacés. La variabilité de l'alimentation est maximale chez
les adultes (LT > 160 mm) du fait de modifications de la nature des proies ingérées entre l'hiver
(poissons), le printemps (macroures et poissons) et l'été (mysidacés et macroures). De telles
modifications saisonnières ont déjà été décrites pour cette espèce avec une ingestion accrue de poissons
en hiver (Belghyti et al., 1993) et de macroures au printemps (Redon et al., 1994) ainsi qu'une
augmentation de l'importance alimentaire des mysidacés en été et au début de l'automne (Belghyti et al.,
1993 ; Redon et al., 1994).
Figure II/II.52 - Variations de l'alimentation de C. linguatula en fonction de la profondeur (30-50 m, 70-100 m), de la saison (P = printemps, E = été, H = hiver), et de la taille (LT, mm) des individus. Ordination(N.M.S., solution à 2 dimensions) réalisée sur les poids moyen des proies par estomac.
PP
P
P
P
P
PP
P
EE
E
E
E EE
HH
H
H
PP
P
P
EEE
HH
H
H
-2
0
2
-2 0 2Axe 1
Axe
2
Stress : 0,087
LT < 100 mm
100-160 mm
LT > 160 mm
70 - 100 m
30 - 50 m
0 - 20 m
Profondeur
juvéniles G0(LT < 100 mm)
adultes(LT > 160 mm)
juvéniles G1+
100-160 mm
Stade de vie

Stratégies alimentaires
134
II/II.4 - CONCLUSION SUR LES STRATEGIES ALIMENTAIRES DES CINQ
ESPECES
Rythme nycthéméral d'alimentation
Parmi les cinq espèces étudiées, deux (Solea solea et Buglossidium luteum) s'alimentent
principalement de nuit et trois (Arnoglossus laterna, Citharus linguatula et vraisemblablement Solea
impar) de jour. Ceci permet une meilleure exploitation de l'ensemble des ressources disponibles dans le
milieu et réduit la compétition alimentaire directe entre les cinq espèces, d'autant plus que les pics
alimentaires sont décalés dans le temps entre espèces nocturnes d'une part et entre espèces diurnes de
l'autre (Fig. II/II.53).
Figure II/II.53 - Rythmes nycthéméraux d'alimentation observés chez les cinq espèces étudiées au largedu Rhône (juin 2000 ; heure locale = heure GMT + 1). AL = Arnoglossus laterna, BL = Buglossidium luteum, CL= Citharus linguatula, SI = Solea impar, SS = Solea solea.
Nombre et poids des proies
Le nombre total de proies par estomac et leur poids individuel moyen diffèrent entre les cinq
poissons plats étudiés, ce qui traduit l'existence de stratégies alimentaires distinctes, même entre espèces
de taille voisine (Tableau II/II.11). Arnoglossus laterna et B. luteum, bien que se nourrissant tous deux
de proies de tailles réduites et ayant des besoins énergétiques proches, présentent des stratégies
alimentaires différentes. Buglossidium luteum se nourrit suivant le principe du "peu mais souvent", le
faible nombre de proies par estomac étant, chez cette espèce, compensé par une prise de nourriture quasi
ininterrompue durant toute la durée du nycthémère. Au contraire, A. laterna, avec un nombre de proies
par estomac élevé et un indice de réplétion fort le jour, s'alimente beaucoup pendant sa période de
nutrition maximale mais peu le reste du temps. Parmi les espèces de taille importante, S. impar présente

Stratégies alimentaires
135
une stratégie alimentaire particulière. Elle se nourrit essentiellement de proies de petite taille, ingérées
en grandes quantités afin de satisfaire ses besoins énergétiques élevés. Les deux autres poissons de
grande taille, S. solea et C. linguatula, ingèrent, selon les circonstances et les proies disponibles, soit un
nombre très élevé d'organismes de petite taille soit un nombre réduit de proies volumineuses.
Tableau II/II.11 - Stratégies alimentaires des cinq espèces au large du Rhône. Np = nombre total desproies/estomac ; Pm proies = poids individuel moyen des proies ; med. = médiane.
Taille maximaleLT (mm)
Np / estomacmed. (min-max)
Pm proies (mg)med. (min-max)
S. soleaS. imparB. luteum
A. laternaC. linguatula
374212126132237
4 (1-323)
11,5 (1-185)
3 (1-40)
8 (1-238)
2 (1-104)
1,9 (0,01-11,49)
0,6 (0,06-8,21)
0,2 (0,01-4,62)
0,4 (0,08-6,12)
2,5 (0,05-171,21)
Régimes alimentaires
Au large du Rhône, l'alimentation des cinq espèces est globalement centrée sur les trois mêmes
groupes zoologiques (polychètes, mollusques, crustacés). Cependant les importances respectives de ces
proies diffèrent d'une espèce à l'autre. Les polychètes tiennent une place prépondérante dans
l'alimentation de S. solea et B. luteum qui ingèrent également de grandes quantités de mollusques
(bivalves), et pour B. luteum, de crustacés (amphipodes). Le régime alimentaire de S. impar est dominé
par les bivalves et les crustacés (amphipodes). Chez A. laterna et C. linguatula, les mysidacés et les
macroures dominent l'alimentation mais leur ingestion est complétée par une forte consommation
d'amphipodes chez A. laterna et de juvéniles de poissons chez C. linguatula. Ces preferenda
alimentaires correspondent, dans leurs grandes lignes, à ceux trouvés dans la littérature pour ces cinq
espèces mais traduisent également l'existence de particularités alimentaires propres aux populations
vivant au large du Rhône (consommation accrue de polychètes et de bivalves).
Modifications ontogéniques de l'alimentation
Chez les cinq espèces étudiées, l'alimentation subit des modifications quantitatives et
qualitatives au cours de la croissance (Tableau II/II.12). Deux principales stratégies peuvent être
dégagées suivant le système de sélection des proies. La première stratégie (relai intraspécifique),
observée chez C. linguatula, consiste en une simple augmentation de la taille des individus ingérés sans
réelle modification de la composition du régime alimentaire. La seconde (relai interspécifique),
observée chez les quatre autres espèces, combine une modification de la nature, de la taille et du nombre
des proies ingérées, avec une consommation de proies de taille réduite chez les juvéniles et de proies
plus volumineuses chez les adultes.

Stratégies alimentaires
136
Tableau II/II.12 - Modifications ontogéniques de l'alimentation des cinq espèces au large du Rhône. Pm proies =poids moyen des proies ingérées ; Pp = poids total des proies/estomac ; Np = nombre total des proies/estomac ;med. = médiane. A = amphipodes ; B = bivalves ; Co = copépodes ; Cu = cumacés ; Ma = macroures ; My =mysidacés ; P = polychètes ; Ph = phoronidiens ; Po = poissons. DSS = dépositivores de sub-surface ; DS =dépositivores de surface ; C = carnivores ; S = suspensivores.
Proies principalesPm proies (mg)med. (min-max)
Pp / estomac (mg)med. (min-max)
Np / estomacmed. (min-max)
Solea solea
Juvéniles G0 LT < 100 mm
LT > 100 mm
Juvéniles G1+, sub-adulteset adultes
Solea impar
Juvéniles G0
G1+
Sub-adultes et adultes
Buglossidium luteum
Juvéniles G0
G1+
Sub-adultes et adultes
Arnoglossus laterna
Juvéniles G0
G1+
Sub-adultes et adultes
Citharus linguatula
Juvéniles G0
G1+
Sub-adultes et adultes
Cu + P (DS) +B + A
P (DS) + B
P (DSS)
P (DS) + Cu + A
A + Cu + B
B + A + My + Cu
Co + Cu + A + P (C) +B + G
Co + P (C + DS) + A +B + Cu
P (C + DSS) + A + B
A + Cu + P (C +S)
A + Cu + P (s +C)
My + Ma + P (s) + Ph
My+ Po
My+ Po
My + Po + Ma
0,84 (0,01-1,88)
1,81 (0,32-5,25)
1,85 (0,36-11,49)
0,30 (0,06-0,88)
0,29 (0,06-1,49)
2,25 (0,18-8,21)
0,32 (0,01-1,11)
0,63 (0,38-1,12)
0,88 (0,51-4,62)
0,13 (0,08-1,22)
0,24 (0,11-0,81)
0,58 (0,18-6,12)
0,11 (0,05-1,41)
1,11 (0,05-62,4)
15,71 (0,12-171,21)
1,69 (0,44-5,65)
6,34 (4,21-60,21)
20,44 (7,24-457,69)
3,35 (0,26-13,50)
8,22 (0,95-21,81)
18,01 (6,19-75,95)
0,98 (0,02-2,73)
2,52 (0,82-6,52)
2,64 (0,51-33,49)
0,78 (0,08-4,08)
2,89 (0,48-6,65)
4,64 (0,13-60,32)
0,67 (0,16-1,08)
2,21 (0,16-115,6)
31,42 (0,68-172,01)
2 (1-18)
3,5 (1-32)
11 (1-323)
11 (1-27)
28 (1-185)
8 (1-57)
3 (1-24)
4 (1-17)
3 (1-40)
6 (1-30)
12 (1-45)
8 (1-238)
6 (1-13)
2 (1-43)
2 (1-104)
Modifications spatio-temporelles de l'alimentation
Chez les cinq espèces étudiées, l'alimentation subit des modifications spatio-temporelles à la
fois quantitatives et qualitatives. Sur le plan quantitatif, l'alimentation varie uniquement suivant la
saison, avec une prise de nourriture maximale en été quelle que soit l'espèce considérée et une
alimentation également intense au printemps chez B. luteum et S. impar. Elle est réduite en hiver chez
toutes les espèces, à l'exception d'A. laterna, pour qui l'alimentation est minimale au printemps.

Stratégies alimentaires
137
Sur le plan qualitatif, la composition du régime alimentaire se modifie suivant la tranche
bathymétrique et la saison considérées (Tableaux II/II.13 et II/II.14). L'essentiel des modifications avec
la profondeur concerne les espèces présentes aux trois tranches bathymétriques étudiées (S. solea, B.
luteum et A. laterna), avec un régime alimentaire plus diversifié à 0-20 m. Les polychètes sont les proies
préférentielles de S. solea et B. luteum à toutes les profondeurs et celles d'A. laterna à 30-50 m,
profondeur où ils sont le plus abondamment consommés par ces trois espèces. Les bivalves, les
amphipodes et les cumacés sont surtout consommés à 0-20 m, où ils sont les proies préférentielles de S.
impar, B. luteum et A. laterna. Les macroures et les mysidacés sont principalement ingérés à 70-100 m,
où ils forment deux catégories de proies préférentielles pour B. luteum, A. laterna et C. linguatula. En
ce qui concerne la saison, la catégorie de proie la plus abondamment consommée diffère généralement
d'une espèce à l'autre quelle que soit la période de l'année. Cependant, certaines catégories de proies
sont consommées de façon importante par plusieurs des cinq espèces à certaines saisons. Les polychètes
sont ingérés de façon préférentielle par S. solea, B. luteum et S. impar au printemps et par S. solea et B.
luteum en hiver et en été. Les mysidacés occupent une place importante dans le régime alimentaire d'A.
laterna et de C. linguatula quelle que soit la saison. Les amphipodes sont abondamment consommés par
S. impar, B. luteum et A. laterna tout au long de l'année, avec un maximum en été où ils constituent un
groupe de proies préférentielles pour S. impar et A. laterna. Enfin, les bivalves sont ingérés en quantités
non négligeables par S. solea, S. impar et B. luteum, surtout au printemps et en été. D'autres catégories
de proies ne sont abondamment consommées qu'à certaines saisons qui diffèrent d'une espèce à l'autre.
Les macroures sont ainsi les proies préférentielles d'A. laterna en hiver et de C. linguatula au printemps.
De même l'ingestion de cumacés est maximale au printemps pour S. impar et en été pour B. luteum.
Tableau II/II.13 - Principales proies (Cp ≥ 10%) des cinq espèces aux 3 profondeurs étudiées au large duRhône. A = amphipodes ; B = bivalves ; Br = brachyoures ; Co = copépodes ; Cu = cumacés ; Ma = macroures ; My =mysidacés ; P = polychètes ; Ph = phoronidiens ; Po = poissons. Les proies préférentielles sont indiquées en gras pourchaque espèce.
S. solea S. impar B. luteum A. laterna C. linguatula
0-20 m P + BB + A +
Cu + P
P + A + B +
Co + Cu
Ph + A +
P + B + Ma
30-50 m P P +
A + B + Cu
Ma + P +
My + A + BrPo + My + Ma
70-100 m P A + P +
My + Ma + CuMy + Ma + A My + Po +Ma

Stratégies alimentaires
138
Tableau II/II.14 - Principales proies (Cp ≥ 10%) observées à chaque saison pour les cinq espèces étudiées aularge du Rhône. A = amphipodes ; B = bivalves ; Br = brachyoures; Cu = cumacés ; Ma = macroures ; My = mysidacés ;P = polychètes ; Po = poissons. Les proies préférentielles sont indiquées en gras pour chaque espèce.
S. solea S. impar B. luteum A. laterna C. linguatula
Hiver P P + A +
Cu + B + My
My + Ma +
A +Br + P
Po + My + Ma
Printemps P + B + O Cu + P + B +
My + A
P + A +
B + Cu + Co
My + P +
Ma +A
My + Ma + Po
Eté P + B A + B + Cu P + A + Co +
B + Ma
My + A +
P + Br
My + Po
Conclusion
Bien qu'ayant des alimentations principalement basées sur l'ingestion des trois même groupes de
proies (crustacés, polychètes et mollusques), les cinq espèces de poissons plats étudiées au large du
Rhône présentent des rythmes d'activité, des stratégies et des régimes alimentaires différents. Leurs
réponses respectives à une augmentation potentielle des ressources alimentaires liée aux apports du
fleuve vont donc dépendre fortement de la nature des proies favorisées par l'augmentation en matière
organique disponible au niveau du fond. Parmi les proies ingérées par les cinq espèces, les polychètes,
les amphipodes, les bivalves et les mysidacés tiennent une place prépondérante. Les sensibilités de ces
quatre catégories de proies aux modifications du milieu liées aux crues du Rhône vont donc déterminer,
pour l'essentiel, l'influence des apports du fleuve sur les populations de S. solea, S. impar, B. luteum, A.
laterna et C. linguatula présentes au large de l'embouchure. Cependant, du fait de la variabilité
temporelle des apports rhodaniens (crues en hiver et au printemps, étiage en été), les modifications
saisonnières (quantitatives et qualitatives) de l'alimentation vont conditionner la capacité de chaque
espèce à tirer profit ou non de l'augmentation des ressources au niveau du fond. De plus, à l'exception de
C. linguatula, chez qui la nature des proies ingérées est relativement stable quel que soit le stade de vie
benthique et/ou la profondeur, l'alimentation varie de façon importante entre juvéniles et adultes et selon
la répartition bathymétrique des individus, ce qui peut avoir d'importantes conséquences quant à
l'intensité du transfert de MOP terrestre vers chaque espèce.

PARTIE II – CHAPITRE III
PARTAGE DES RESSOURCES ALIMENTAIRES


Partage des ressources alimentaires
139
PARTAGE DES RESSOURCES ALIMENTAIRES
II/III.1 - INTRODUCTION
La sensibilité d'une espèce aux fluctuations de son environnement dépend non seulement de la
nature des changements auxquels elle est confrontée et de sa propre capacité d'adaptation ou de réaction
à ces variations mais aussi des relations qu'elle entretient avec les autres espèces qui partagent son
milieu. Ainsi, l'aptitude de chaque poisson à tirer profit d'une augmentation des proies disponibles dans
le milieu va dépendre de ses interactions avec les autres prédateurs en présence. La façon dont s'effectue
le partage des ressources alimentaires entre les cinq espèces de poissons plats étudiées est donc
susceptible de modifier en partie l'intensité de leurs réponses aux modifications du débit du Rhône.
Le concept de "partage des ressources alimentaires" désigne l'utilisation différentielle des proies
par deux espèces ou plus, ou par différentes classes d'âge au sein d'une même espèce, de façon à
exploiter au mieux l'ensemble des ressources alimentaires disponibles (Gerking, 1994). La répartition de
la ressource trophique totale entre les différentes espèces reflète donc leurs exigences et leurs capacités
d'adaptation alimentaire respectives. Le partage des ressources alimentaires est fréquemment observé
dans les assemblages de poissons (Mc Arthur, 1972 ; Schoener, 1974 ; Thorman et Wiederholm, 1986 ;
Gibson et Ezzi, 1987 ; Labropoulou et Markakis, 1998 ; Darnaude et al., 2001). Il n’entraîne cependant
pas nécessairement de compétition alimentaire si la ressource considérée se trouve en abondance
suffisante pour satisfaire aux besoins spécifiques des différents prédateurs présents dans le milieu
(Thorman et Wiederholm, 1983, 1986). Son rôle structurant est donc contesté au sein des assemblages
de poissons, certains auteurs accordant une importance prépondérante au partage de l'habitat (Sale, 1977
; Robertson, 1980). Cependant, chez les poissons de substrats meubles, le partage de la nourriture
jouerait un rôle plus important que celui de l’habitat, du fait de la relative homogénéité de ce dernier
(Schoener, 1974 ; Ross, 1986 ; Piet et al., 1999). Dans ce chapitre, l'étude du partage des ressources au
large du Rhône entre Solea solea, Solea impar, Buglossidium luteum, Arnoglossus laterna et Citharus
linguatula sera donc centrée sur la description de leurs interactions trophiques. La répartition
bathymétrique des individus des cinq espèces sera néanmoins prise en compte dans le calcul des
largeurs et des recouvrements des niches alimentaires afin de ne pas négliger l'importance du partage de
l'habitat dans la réduction de la compétition trophique intra et interspécifique.

Partage des ressources alimentaires
140
II/III.2 - MATERIEL ET METHODES
Il ne peut y avoir de compétition entre espèces que si ces dernières exploitent des ressources
dont la quantité totale disponible est inférieure aux besoins de toutes les espèces en présence (Blondel,
1995). L'existence ou non d'une compétition effective pour les ressources alimentaires est donc en partie
liée à l'abondance et la disponibilité des proies dans le milieu. Cependant, l'intensité des interactions
trophiques entre espèces dépend de deux principaux paramètres qui déterminent, en amont, la possibilité
ou non d'une compétition : (1) la capacité d'adaptation de chaque espèce aux modifications de nature et
d'abondance des proies disponibles, estimée par la largeur de sa niche alimentaire, et (2) ses interactions
potentielles avec les autres prédateurs en présence, estimées par le taux de recouvrement de sa niche
alimentaire par celles des autres espèces. Les largeurs et les recouvrement des niches alimentaires,
seront donc estimés et comparés pour les cinq poissons plats étudiés afin de décrire et de quantifier le
partage des ressources alimentaires entre leurs populations.
L'utilisation des ressources alimentaires, exprimée par la largeur et le recouvrement des niches
alimentaires, ne peut être comparée pour plusieurs populations que si le biais lié à l'échantillonnage est
éliminé et si les modifications de l'alimentation existant pour chaque espèce sont prises en compte (Piet
et al., 1999). Au large du Rhône, le partage des ressources alimentaires entre les cinq espèces de
poissons plats étudiées est susceptible de varier fortement suivant la tranche bathymétrique et la saison
considérées, étant donné les modifications ontogéniques de l'alimentation et les variations spatio-
temporelles de répartition bathymétrique des différents stades de vie. Pour ces raisons, les relations
alimentaires intra et interspécifiques seront détaillées pour chaque saison et chaque profondeur, ce
dernier facteur étant retenu comme principal étant donné les différences importantes de densité de
poissons et de composition du peuplement entre les petits fonds sableux côtiers et les vases terrigènes
plus profondes. De plus, le facteur "profondeur" sépare totalement Solea impar et Citharus linguatula
entre qui toute compétition pour les ressources est impossible au large du Rhône du fait de la
ségrégation bathymétrique de leurs deux populations (respectivement réparties à 0-20 m et 30-100 m de
profondeur).
II/III.2.1 - CALCULS DES LARGEURS DE NICHES ALIMENTAIRES
La notion de "niche alimentaire" découle directement de celle de "niche écologique". Cette
dernière à donné lieu à de nombreuses discussions et publications. Nous retiendrons ici comme
définition celle de Blondel (1995) : "La niche est un hypervolume à n dimensions (Hutchinson, 1958 in
Blondel, 1995) qui rassemble la totalité des relations qu’une espèce entretient avec les paramètres de
son habitat et avec les autres espèces de la biocénose" (Fig. II/III.1). La niche écologique englobe non
seulement l'ensemble des besoins requis par un organisme pour survivre dans son milieu mais
également les actions réalisées pour les satisfaire. A ce niveau, il convient de séparer, d'une part, la

Partage des ressources alimentaires
141
niche fondamentale d'une espèce, définie par Solbrig et Solbrig (1979) comme "l'hyperespace limité par
les valeurs des variables dont les organismes ont besoin pour vivre dans un espace écologique non
limité", et, de l'autre, sa niche réalisée, nécessairement plus réduite en raison de contraintes comme la
compétition ou la limitation des ressources.
Figure II/III.1 - Modèle illustrant le concept de la niche écologique : exemple de laposition d'un hypervolume-niche dans un espace à trois dimensions et projection decet hypervolume sur les plans définis par ces dimensions (d'après Pianka, 1974 inBlondel, 1995).
L'alimentation est, tout comme l'habitat, l'une des dimensions de la niche écologique. Les
concepts définis pour cette dernière peuvent lui être transposés. Ainsi, la niche alimentaire d'une espèce
englobe à la fois ses exigences et ses potentialités d'adaptation d'ordre nutritionnel. On peut de plus
distinguer, pour chaque espèce, une niche alimentaire fondamentale, "théorique" (qui correspond à ce
que serait l'alimentation de l'espèce dans un espace écologique non limité, si elle n'était soumise à
aucune modification spatio-temporelle des ressources et en l'absence d'interaction avec les autres
espèces présentes dans le milieu), et une niche alimentaire réalisée, correspondant à celle observée de
façon effective pour sa population dans le milieu étudié.
aptitude optimale
aptitude moindre

Partage des ressources alimentaires
142
Dans la présente étude, seule les largeurs de niches alimentaires réalisées ont été estimées. Elles
fournissent une idée de la capacité d'adaptation des différents groupes de poissons en présence en cas de
modification dans le milieu des ressources alimentaires qu'ils exploitent. Pour chaque espèce, deux
types de largeurs de niche ont été calculées. Dans un premier temps, les largeurs des niches alimentaires
réalisées (B) des différents stades de vie benthique présents à chaque saison et chaque profondeur, ont
été calculées suivant la formule (Pielou, 1969) :
n
B = e H’ , H’= - ∑ pi × log2 pi i = 1
où pi est la fraction représentée par le type de proie i dans le poids total des n proieslistées dans le régime alimentaire du groupe de poissons considéré.
La proportion en poids a été utilisée plutôt que celle en nombre d’individus car elle reflète
mieux l’utilisation de la nourriture par le prédateur (Harmelin-Vivien et al., 1989). Afin de comparer les
cinq espèces entre elles, une largeur de niche alimentaire réalisée globale (BG) a ensuite été calculée
pour chaque population en pondérant la largeur de niche réalisée (B) obtenue pour chaque groupe de
poissons par l'effectif correspondant dans les captures (Piet et al., 1999) :
N
BG = ∑ B j × E j
i = 1
où Bj est la largeur de niche alimentaire réalisée (B) obtenue pour le groupe de poissons j etEj la fraction de l'effectif total de l'espèce représenté par ce même groupe de poissons.
II/III.2.2 - RECOUVREMENT DES NICHES ALIMENTAIRES
Le recouvrement des niches correspond à la fraction de l'ensemble des ressources exploitées par
deux espèces qui est partagée entre leurs individus. Il fournit une idée de la compétition potentielle pour
les ressources existant entre les différentes espèces, prises deux à deux. Dans le cas de l'alimentation, il
permet d'évaluer le degré de similitude des régimes alimentaires et de quantifier l'importance de la
compétition alimentaire potentielle entre les différents groupes de poissons, pris deux à deux.
Il existe de nombreux indices mesurant le recouvrement des niches écologiques, et par
extension celui des régimes alimentaires. Pour calculer, à chaque profondeur et chaque saison, le
chevauchement des spectres alimentaires des différents groupes de poissons individualisés pour chaque
espèce, l’indice (T) de Schoener (1970) a été utilisé. Couramment employé, ce dernier fournit, dans la
majorité des cas, une estimation convenable du recouvrement (Wallace, 1981). Il se calcule de la façon
suivante :

Partage des ressources alimentaires
143
n
T = 1 – 0,5 ∑ Pxi - Pyi i = 1
où Pxi et Pyi sont les fractions du poids total représentées par le type de proie i pour chacun desdeux poissons x et y.
l'indice T varie en théorie de zéro, quand les deux groupes de poissons s’alimentent à partir de
proies totalement différentes, à un, quand ils utilisent les mêmes proies dans les mêmes proportions. Un
recouvrement supérieur ou égal à 0,6 sera considéré comme significatif (Keast, 1978). De même que
pour le calcul des largeurs de niches alimentaires, les proportions en poids ont été utilisées.
Afin d'estimer le degré de similitude entre les alimentations des cinq espèces au large du Rhône,
un indice de recouvrement global théorique (TT) des niches alimentaires a été calculé pour chaque paire
de poissons, suivant la formule (Piet et al., 1999) :
Nx Ny
TT (x, y) = 0,5 ∑ ∑ (T(i, j) × E i × E j ) (1) i = 1 j = 1
où T(i, j) représente le recouvrement des niches alimentaires obtenu entre chaque groupe i de l'espèce x etchaque groupe j de l'espèce y, Ei et Ey les fractions respectives des effectifs totaux de l'espèce x et del'espèce y représentés par le groupe i et le groupe j, et Nx et Ny le nombre total de groupes de poissons(stade de vie benthique/profondeur/saison) respectivement individualisés pour l'espèce x et l'espèce y.
Cet indice de recouvrement n'a de valeur que théorique. Il permet de déterminer les espèces
ayant les régimes alimentaires les plus similaires, tous stades de vie benthique, toutes saisons et toutes
profondeurs confondus et donne une idée de la compétition interspécifique maximale possible dans le
milieu étudié, dans l'hypothèse où l'ensemble des individus des différentes espèces se trouvent
confrontés les uns aux autres dans le temps et dans l'espace. Comme l'indice T (dont il est dérivé), cet
indice varie de zéro (les populations des deux espèces s’alimentent à partir de proies totalement
différentes) à un (l'ensemble de leurs individus utilisent les mêmes proies dans les mêmes proportions).
Il sera considéré comme significatif lorsque supérieur ou égal à 0,6 (Keast, 1978).
Au large du Rhône, les différences de répartition spatio-temporelle des populations des cinq
espèces réduisent de façon importante les interactions potentielles entre individus, ce qui diminue la
compétition effective entre espèces (Fig. II/III.2) et peut même l'exclure totalement. C'est le cas pour
Solea impar et Citharus linguatula entre qui toute compétition alimentaire est écartée du fait de leurs
distributions bathymétriques totalement distinctes (respectivement entre 0 et 20 m et entre 30 et 100 m
de profondeur).

Partage des ressources alimentaires
144
Figure II/III.2 - Projection de la niche de cinq espèces types sur deux dimensions de l'hypervolume-niche.Les espèces dont les niches se chevauchent sur une dimension (la niche de l'espèce 3 est totalement incluse dans cellede l'espèce 4 sur la dimension I) peuvent être isolées par une deuxième dimension. (D'après Pianka, 1974 in Blondel,1995). Dans le cas présent, si les dimensions I et II correspondent respectivement à l'alimentation et l'habitat(profondeur), le cas de S. impar et C. linguatula est assimilable à celui des espèces 4 et 3 ou 4 et 2.
Pour cette raison, les indices de recouvrement effectif des niches alimentaires des cinq espèces
ont été calculés selon la formule suivante, version adaptée de la formule (1) qui ne croise que
l'alimentation des stades de vie benthique de deux espèces présents de façon simultanée à une
profondeur et une saison données :
3 3 NPSx NPSy
TE (x, y) = ∑ ∑ ∑ ∑ (TPS (i, j) × EPS i × EPS j ) P = 1 S = 1 i = 1 j = 1
où TPS(i, j) représente le recouvrement des niches alimentaires obtenu entre chaque groupe i de l'espècex et chaque groupe j de l'espèce y présents à chaque doublet " profondeur (P) × saison (S)", EPS i et EPS jles fractions respectives des effectifs totaux de l'espèce x et de l'espèce y représentés à chaque doublet" P × S " par le groupe i et le groupe j, et NPSx et NPSy le nombre total de groupes de stade de viebenthique respectivement individualisés pour l'espèce x et l'espèce y à chaque doublet " P × S ".
Cet indice varie de zéro, quand les individus des populations des deux espèces ne sont jamais
présents de façon simultanée ou s’alimentent toujours à partir de proies totalement différentes, à un,
quand ils sont présents aux mêmes saisons à chaque profondeur et s'y nourrissent des mêmes proies
dans les mêmes proportions. De même que pour les autres indices utilisés, un recouvrement supérieur
ou égal à 0,6 sera considéré comme significatif (Keast, 1978).
dimension II
dimension I
1
11
2 3 4 5
5
3
4
2
4
5
3
2

Partage des ressources alimentaires
145
II/III.2.3 - PRISE EN COMPTE DE LA TAILLE DES PROIES INGEREES
Si elle possède l'avantage de quantifier de façon mathématique le recouvrement des niches
alimentaires entre les groupes de poissons étudiés et de disposer d'une valeur seuil pour déterminer la
présence ou non d'une compétition alimentaire entre eux, l'utilisation des indices de recouvrement des
niches présente également d'importantes lacunes. En effet, ce type de calcul, réalisé sur les
pourcentages pondéraux des proies dans l'alimentation (donc sur des proportions ramenées à 100 afin
être comparées) et non sur les poids réels des différentes proies dans les estomacs de chaque espèce, ne
tient pas compte des variations intra et interspécifiques du poids total des proies ingérées par chacun des
groupes de poissons étudiés. Ce paramètre est pourtant susceptible de réduire fortement la compétition
entre les individus des espèces de poissons plats au large du Rhône, pour qui la taille (et donc le poids)
des proies consommées varie de façon importante suivant l'espèce et le stade de vie benthique considéré
(c.f. Partie II - Chapitre II). L'ordination par N.M.S. des différents groupes de poissons individualisés
(espèce / stade de vie benthique / saison / profondeur), réalisée à partir du poids moyen des proies
ingérées, permet de combler cette lacune et d'estimer, au mieux, les interactions d'ordre trophique entre
les populations des cinq espèces. La technique du "Non Metric Multidimentional Scaling" a été
présentée en détail au sein du chapitre précédent (c. f. partie "matériel et méthodes"). Dans ce cas précis,
les calculs ont été basés sur une matrice de distances n × n, calculée à partir de la matrice d'origine de n
lignes (observations = groupes de poissons) et p colonnes (variables = catégories de proies ingérées) en
utilisant la distance de Bray-Curtis. Les relations trophiques entre individus des cinq espèces mises en
évidence par l'ordination des groupes de poissons seront détaillées pour les trois tranches
bathymétriques étudiées (0-20 m, 30-50 m et 70-100 m) afin d'affiner le schéma global observé. Leur
analyse, ainsi que celle des indices de recouvrement des niches alimentaires permettront de tirer des
conclusions sur le partage des ressources alimentaires entre ces espèces et leur compétition éventuelle
au large du Rhône.
II/III.3 - PARTAGE DES RESSOURCES ALIMENTAIRES ENTRE ESPECES
II/III.3.1 - COMPETITION POTENTIELLE POUR LES RESSOURCES ALIMENTAIRES
Les largeurs de niches alimentaires (B) obtenues pour l'ensemble des groupes de poissons
individualisés (espèce / stade de vie / profondeur / saison) fluctuent entre 1,2 à 18,7, la moitié des
valeurs obtenues se situant entre 3,5 et 12 (Tableau II/III.1). Compte-tenu de ces observations, les
largeurs de niches alimentaires seront considérées comme faibles lorsqu'elles sont inférieures à 5,
comme moyennes entre 5 et 11 et comme élevées au delà de 11.
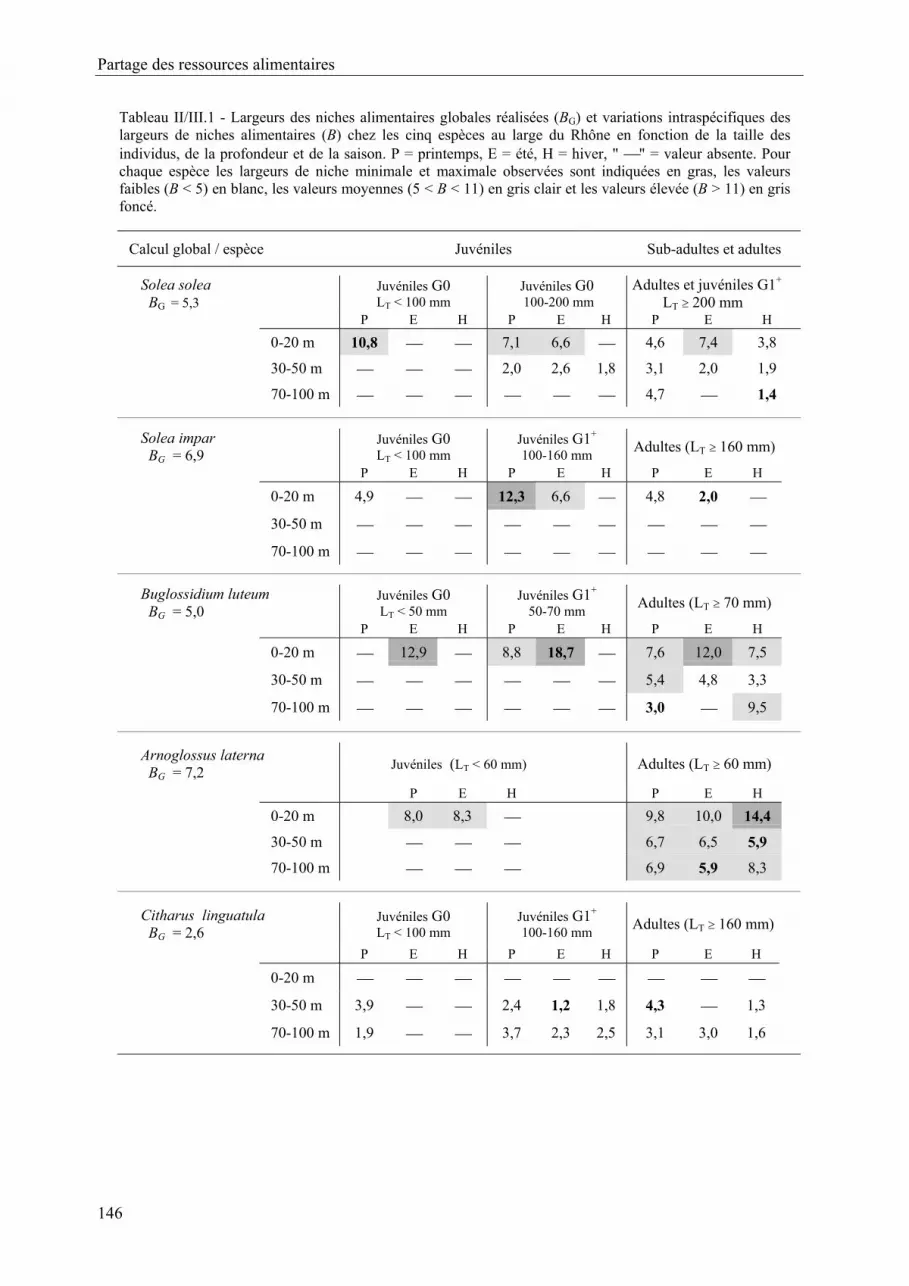
Partage des ressources alimentaires
146
Tableau II/III.1 - Largeurs des niches alimentaires globales réalisées (BG) et variations intraspécifiques deslargeurs de niches alimentaires (B) chez les cinq espèces au large du Rhône en fonction de la taille desindividus, de la profondeur et de la saison. P = printemps, E = été, H = hiver, " " = valeur absente. Pourchaque espèce les largeurs de niche minimale et maximale observées sont indiquées en gras, les valeursfaibles (B < 5) en blanc, les valeurs moyennes (5 < B < 11) en gris clair et les valeurs élevée (B > 11) en grisfoncé.
Calcul global / espèce Juvéniles Sub-adultes et adultes
Solea solea BG = 5,3
Juvéniles G0LT < 100 mm
Juvéniles G0 100-200 mm
Adultes et juvéniles G1+
LT ≥ 200 mmP E H P E H P E H
0-20 m 10,8 7,1 6,6 4,6 7,4 3,8
30-50 m 2,0 2,6 1,8 3,1 2,0 1,9
70-100 m 4,7 1,4
Solea impar BG = 6,9
Juvéniles G0LT < 100 mm
Juvéniles G1+
100-160 mm Adultes (LT ≥ 160 mm)
P E H P E H P E H
0-20 m 4,9 12,3 6,6 4,8 2,0
30-50 m
70-100 m
Buglossidium luteum BG = 5,0
Juvéniles G0LT < 50 mm
Juvéniles G1+
50-70 mm Adultes (LT ≥ 70 mm)
P E H P E H P E H
0-20 m 12,9 8,8 18,7 7,6 12,0 7,5
30-50 m 5,4 4,8 3,3
70-100 m 3,0 9,5
Arnoglossus laterna BG = 7,2 Juvéniles (LT < 60 mm) Adultes (LT ≥ 60 mm)
P E H P E H
0-20 m 8,0 8,3 9,8 10,0 14,4
30-50 m 6,7 6,5 5,9
70-100 m 6,9 5,9 8,3
Citharus linguatula BG = 2,6
Juvéniles G0LT < 100 mm
Juvéniles G1+
100-160 mm Adultes (LT ≥ 160 mm)
P E H P E H P E H
0-20 m
30-50 m 3,9 2,4 1,2 1,8 4,3 1,3
70-100 m 1,9 3,7 2,3 2,5 3,1 3,0 1,6

Partage des ressources alimentaires
147
Au large du Rhône, les largeurs de niche alimentaire globale réalisée des cinq espèces sont
comprises entre 2,6 et 7,2 (Tableau II/III.1). Le régime alimentaire de chaque espèce est donc peu ou
moyennement diversifié. La largeur de la niche alimentaire globale ne semble liée ni à la taille de
l'espèce ni à sa répartition bathymétrique. Elle est maximale (BG = 7,2) pour A. laterna qui présente,
malgré sa petite taille, le régime alimentaire le plus diversifié, et minimale pour C. linguatula (BG =
2,6). Parmi les trois soléidés, la largeur de niche alimentaire globale est maximale chez S. impar (BG =
6,9), exclusivement répartie sur les fonds de 0-20 m de profondeur, et réduite chez S. solea et B. luteum,
pourtant tous deux distribués entre 0 et 100 m de profondeur. Ces deux dernières espèces présentent des
largeurs de niche globales voisines (BG ≈ 5) malgré des tailles maximales très différentes. Les largeurs
de niches réalisées observées relèvent donc plus de capacités adaptatives et de preferenda alimentaires
propres à chaque espèce que d'une modification des ressources disponibles à chaque profondeur ou de
différences de gamme de taille des proies capturables par les poissons. Ce constat est renforcé par
l'étude des variabilités intra-spécifiques de la largeur de niche alimentaire, qui reflètent la capacité
d'adaptation des individus des cinq espèces suivant leur taille et l'environnement fréquenté (Tableau
II/III.1). Elles passent du simple au double entre les deux espèces de petite taille, A. laterna (± 8,5) et B.
luteum (± 15,7), et du simple au triple entre celles de grande taille, C. linguatula (± 3,2), S. solea (±
9,5), S. impar (± 10,3), sans aucun lien apparent avec la distribution bathymétrique.
On distingue, chez les cinq espèces, deux principaux cas de figure : (1) celui de C. linguatula
qui, avec une largeur de niche faible et peu variable, présente une capacité d'adaptation réduite et est
donc particulièrement "vulnérable" vis-à-vis d'une compétition potentielle pour les proies qu'elle ingère,
(2) celui d'A. laterna, B. luteum, S. solea et S. impar qui, avec des largeurs de niche alimentaires
moyennes mais variables, semblent plus capables de s'adapter à une éventuelle modification de la nature
et/ou de l'abondance des ressources disponibles dans le milieu.
De fait, les indices globaux de recouvrement théorique des niches alimentaires les plus faibles
concernent tous C. linguatula, pour qui les valeurs de TT ne dépassent jamais 0,36 (Tableau II/III.2).
Cette espèce possède donc un régime alimentaire très différent de ceux des quatre autres poissons plats
étudiés. Elle est, de ce fait, peu concernée par une éventuelle compétition interspécifique pour les
ressources alimentaires. Les indices de recouvrement globaux théoriques des quatre autres espèces sont
plus élevés (TT = 0,41 - 0,64). La compétition potentielle pour les ressources alimentaires concerne
surtout S. solea et B. luteum. Les similitudes de composition du régime alimentaire sont maximales
entre ces deux espèces, avec un indice de recouvrement alimentaire global théorique (TT = 0,64)
dépassant la valeur seuil de 0,6. La compétition pour les ressources alimentaires est également
potentiellement importante entre B. luteum et A. laterna, avec une valeur de TT élevée (0,51) bien que
non significative entre ces deux espèces.

Partage des ressources alimentaires
148
Tableau II/III.2. Indices de recouvrement globaux théoriques des niches alimentaires (TT) calculés aularge du Rhône pour les cinq espèces prises deux à deux.
TTSoleaImpar
Buglossidiumluteum
Arnoglossuslaterna
Citharuslinguatula
Solea solea
Solea impar
Buglossidium luteum
Arnoglossus laterna
0,42 0,64
0,41
0,46
0,34
0,51
0,15
0,14
0,17
0,36
II/III.3.2 - PARTAGE EFFECTIF DES RESSOURCES ALIMENTAIRES
Au large du Rhône, l'intensité des interactions trophiques entre les cinq espèces étudiées est
maximale à 0-20 m de profondeur (Fig. II/III.3). Dans le plan proposé par l'ordination des groupes de
poissons (pour chaque espèce : stade de vie / saison / profondeur) en fonction de leur alimentation, la
disposition des points obtenus pour les espèces étudiées est plus compacte à 0-20 m qu'à 30-50 m ou 70-
100 m, ce qui indique des similarités de régimes alimentaires plus fortes sur les petits fonds sableux,
qu'au niveau des vases terrigènes plus profondes.
Figure II/III.3 - Ordination des différents groupes de poissons individualisés pour les cinq espècesétudiées en fonction de leur alimentation au large du Rhône : N.M.S. (solution à 2 dimensions) réaliséesur les poids moyens (en mg) des proies ingérées par les individus de chaque espèce groupés par stade devie benthique / saison / profondeur. Les différences de couleurs indiquent les profondeurs (voir légende).
AlAl
AlAlAl
Al
Al
AlAl
AlAl
Bl
Bl
Bl
Bl
Bl
BlBl
Bl
Bl
Bl
ClCl
Cl
Cl
Cl
ClCl
Cl
Cl
Cl
Cl
Cl
Cl
Si
SiSi
Si
SiSs
Ss
SsSs
SsSs
Ss SsSs
Ss
Ss
Ss
Ss
Ss
-2
0
2
-2 0 2
Axe 1
Axe
2
Stress = 0,131
70 - 100 m
30 - 50 m
0 - 20 m
Tranchebathymétrique
juvéniles G0(LT < 100 mm)
sub-adultes etadultes
juvéniles G1+ et juvéniles G0 de L T > 100 mm
Stade de viebenthique

Partage des ressources alimentaires
149
Ce phénomène est accru par les différences de densité de poissons existant entre ces deux types
de fonds (c. f. Partie II - Chapitre I). Le nombre de groupes de poissons présents ainsi que leur densité
sont nettement plus élevés à 0-20 m qu'entre 30 et 100 m de profondeur, où l'importance potentielle des
interactions trophique entre espèces est réduite du fait de l'absence de S. impar et des plus petits
juvéniles de S. solea, B. luteum et A. laterna.
II/III.3.2.1 - PARTAGE DES RESSOURCES ALIMENTAIRES SUR LES PETITS FONDS SABLEUX (0-
20 m)
Situation à 0-20 m au printemps
Au printemps, les stades de vie benthiques de S. solea, S. impar, B. luteum et A. laterna sont
tous présents à 0-20 m à l'exception des plus petits juvéniles de B. luteum. Les indices de recouvrement
des niches alimentaires observés varient de 0,13 à 0,82 (Tableau II/III.3). Seuls les adultes de S. impar
ont un régime alimentaire relativement distinct de celui des autres groupes de poissons avec des indices
de recouvrement de niches intra et interspécifiques inférieurs à 0,52. Pour les autres groupes en
présence, le recouvrement des niches alimentaires est significatif avec 1 à 6 autre(s) groupe(s) ce qui
porte à 20 le nombre de recouvrements supérieurs à 0,6 pour cette saison. La compétition alimentaire
potentielle est surtout interspécifique et concerne principalement les soléidés. Elle est maximale pour B.
luteum (juvéniles G1+ et adultes) chez qui on observe des recouvrements significatifs avec les juvéniles
de S. solea (T = 0,63 - 0,73) et ceux de S. impar (T = 0,64 - 0,69). La compétition intra-spécifique
potentielle est également élevée à cette saison avec un recouvrement de niche alimentaire maximal (T =
0,82) entre adultes et juvéniles de B. luteum et une valeur de T élevée (0,67) entre adultes et juvéniles de
S. solea.
Tableau II/III.3 - Indices de recouvrement des niches alimentaires (T) observés au printemps à 0-20 m deprofondeur, pour les stades de vie benthique des espèces présentes, pris deux à deux. S s = S. solea ; S i = S.impar ; B l = B. luteum ; A l = A. laterna ; C l = C. linguatula. Juv. = juvéniles ; G0 = juvéniles de l'année ; G1+ = juvénilesde plus d'un an ; Ad. = adultes ; LT = longueur totale en mm.
T S s G0LT < 100 mm
S s G0LT ≥ 100 mm
S sAd.
S iG0
S iG1+
S iAd.
B lG1+
B lAd.
A lJuv.
S s G0LT ≥ 100 mm 0,52S s Ad. 0,38 0,67
S i G0 0,66 0,65 0,55
S i G1+ 0,77 0,43 0,22 0,51
S i Ad. 0,52 0,23 0,13 0,26 0,49
B l G1+ 0,73 0,63 0,42 0,64 0,69 0,32
B l Ad. 0,72 0,75 0,54 0,67 0,68 0,41 0,82
A l Juv. 0,51 0,45 0,26 0,49 0,65 0,20 0,61 0,50
A l Ad 0,62 0,67 0,55 0,74 0,48 0,24 0,60 0,70 0,44

Partage des ressources alimentaires
150
La compétition alimentaire potentielle porte essentiellement sur les polychètes, consommés de
façon importante (Cp = 16 - 65%) par les 9 groupes de poissons impliqués et, à moindre échelle, sur les
bivalves (Cp = 19 - 32% pour 4 groupes), les amphipodes (Cp = 12 - 19% pour 5 groupes) et les
cumacés (Cp = 12 - 23% pour 4 groupes) (Tableau IV).
Tableau II/III.4 - Pourcentages pondéraux (Cp) des différentes catégories de proies ingérées au printemps à 0-20 mpar les 10 groupes de poissons présents à cette profondeur et à cette saison. S s = S. solea ; S i = S. impar ; B l = B.luteum ; A l = A. laterna ; C l = C. linguatula. Juv. = juvéniles ; G0 = juvéniles de l'année ; G1+ = juvéniles de plus d'un an ;Ad. = adultes ; LT = longueur totale en mm; "" = proie absente pour ce groupe de poissons.
S s G0LT < 100 mm
S s G0LT ≥ 100 mm
S sAd.
S iG0
S iG1+
S iAd.
B lG1+
B lAd.
A l Juv.
A l Ad.
Polychètes 31,8% 51,1% 50,4% 65,6% 16,4% 3,6% 38,0% 47,1% 21,5% 51,9%
Bivalves 32,1% 8,9% 2,7% 7,8% 28,4% 68,7% 19,6% 23,1% 7,2% 3,9%
Amphipodes 9,2% 16,1% 1,3% 6,4% 18,8% 5,9% 19,2% 18,8% 15,8% 12,4%
Isopodes 11,5%
Macroures 0,5%
Mysidacés 6,7% 6,5% 6,4% 14,0% 2,8% 8,2%
Brachyoures 1,1%
Anomoures 1,2%
Leptostracés 3,4%
Cumacés 13,1% 0,2% 13,7% 23,1% 1,9% 12,0% 2,8% 47,3% 5,4%
Copépodes 2,1% 1,0% 11,2% 2,8% 4,5% 0,6%
Crust. Indét. 1,3% 3,0% 0,3% 4,3%
Ophiures 1,8% 12,4% 44,1% 1,8% 4,8% 2,6%
Phoronidiens 4,0% 11,9%
Situation à 0-20 m en été
En été, période où, parmi les plus petits juvéniles, seuls les G0 de B. luteum et d'A. laterna sont
présents à 0-20 m, neuf groupes de poissons sont représentés dans les captures. Les indices de
recouvrement des niches varient de 0,07 à 0,79 (Tableau II/III.5). La composition du régime alimentaire
diffère de façon importante d'une espèce et d'un groupe à l'autre, et la compétition alimentaire
potentielle est réduite entre les 4 espèces. Le seul indice de recouvrement significatif des niches
alimentaires (T = 0,79) est observé entre juvéniles (G0 vs G1+) de B. luteum. Il est lié à une ingestion
importante de polychètes, de bivalves, de gastéropodes, de cumacés et de copépodes. Ces 5 catégories
de proies constituent chacune 10 à 22 % en poids du total des proies ingérées par ces deux groupes
d'individus.

Partage des ressources alimentaires
151
Tableau II/III.5 - Indices de recouvrement des niches alimentaires (T) observés en été et à 0-20 m deprofondeur pour les stades de vie benthique des espèces présentes, pris deux à deux. S s = S. solea ; S i = S.impar ; B l = B. luteum ; A l = A. laterna ; C l = C. linguatula. Juv. = juvéniles ; G0 = juvéniles de l'année ; G1+ =juvéniles de plus d'un an ; Ad. = adultes ; LT = longueur totale en mm.
T S s G0LT ≥ 100 mm
S sAd.
S iG1+
S iAd.
B lG0
B lG1+
B lAd.
A lJuv.
S s Ad. 0,44
S i G1+ 0,22 0,40
S i Ad. 0,26 0,49 0,42
B l G0 0,44 0,40 0,49 0,21
B l G1+ 0,44 0,53 0,48 0,28 0,79
B l Ad. 0,56 0,26 0,26 0,15 0,44 0,42
A l Juv. 0,17 0,14 0,50 0,08 0,36 0,31 0,31
A l Ad 0,55 0,17 0,20 0,07 0,39 0,30 0,42 0,38
Situation à 0-20 m en hiver
En hiver, période où seuls les adultes de S. solea, B. luteum et A. laterna sont abondamment
présents à 0-20 m, le recouvrement des niches alimentaires varie de 0,41 à 0,74 et n'est significatif
qu'entre adultes de S. solea et B. luteum (Tableau II/III.6). La compétition alimentaire potentielle entre
ces deux groupes de poissons porte alors essentiellement sur les polychètes (Cp > 51%) et les bivalves
(Cp > 18%).
Tableau II/III.6 - Indices de recouvrement des niches alimentaires (T)observés en hiver à 0-20 m de profondeur pour les stades de vie benthiquedes espèces présentes, pris deux à deux. S s = S. solea ; B l = B. luteum ; A l= A. laterna. Ad. = adultes.
TS sAd.
B lAd.
B l Ad. 0,74
A l Ad 0,41 0,53
Conclusion sur le partage des ressources à 0-20 m
Sur les petits fonds sableux compris entre 0 et 20 m de profondeur, la compétition alimentaire
est maximale au printemps. Elle est très réduite en été grâce à des différences accrues de régimes
alimentaires d'un groupe à l'autre, et en hiver par une diminution importante du nombre de groupes de
poissons présents. Elle concerne principalement les soléidés avec un recouvrement interspécifique
souvent significatif entre les niches alimentaires des différents groupes de S. solea, S. impar et B.
luteum. On observe de plus, chez B. luteum, un recouvrement intra-spécifique significatif et important
(T > 0,79) au printemps et en été.

Partage des ressources alimentaires
152
Tableau II/III.7 - Poids moyens (mg) des proies ingérées à 0-20 m par deux groupes de poissons ou plus. Pourchaque groupe de poissons, seules sont indiquées les catégories de proies représentant plus de 20% en poids dupoids total des proies ingérées, avec en gras, les proies préférentielles. S s = S. solea ; S i = S. impar ; B l = B.luteum ; A l = A. laterna ; C l = C. linguatula. Juv. = juvéniles ; G0 = juvéniles de l'année ; G1+ = juvéniles de plusd'un an ; Ad. = adultes ; LT = longueur totale en mm.
Polychètes Bivalves Gastéropodes Amphipodes Macroures Cumacés Copépodes
S s G0 LT < 100 mm 0,97 0,99 0,08
S s G0 LT ≥ 100 mm 1,34 1,59 17,64 0,60
S s Ad. 4,66 9,12 30,86
S i G0 0,52 0,06
S i G1+ 1,06 1,40 0,13 0,11
S i Ad. 1,73 3,65 6,33 0,24 0,23
B l G0 0,84 1,00 1,58 0,48 0,26 0,05
B l G1+ 0,69 0,72 5,33 0,25 0,41 0,13
B l Ad. 2,73 1,35 1,35
A l Juv. 0,33 0,11 0,07 0,66 0,04
A l Ad 2,56 1,20 0,67 0,11
Figure II/III.4 - Positionnement des groupes alimentaires présents à 0-20 m dans le plan obtenu parordination des espèces de poissons en fonction de leur alimentation (N.M.S. réalisée sur les individusgroupés par stade de vie benthique / saison / profondeur, solution à 2 dimensions). Les différences decouleur indiquent les saisons : printemps (vert), été (jaune), hiver (bleu).
Bl
Al
Al
Al
Bl
Bl
Bl
Bl
SiSi Si
SiSi Ss
SsSs Ss
SsSs
Al
Bl
Al
-2
0
2
-2 0 2
Axe 1
Axe
2
Stress = 0,131 été
printemps
hiver
Saison
juvéniles G0(LT < 100 mm)
adultes
juvéniles G1+ et G0 de LT > 100 mm
Stade de vie benthique

Partage des ressources alimentaires
153
A 0-20 m, la compétition alimentaire potentielle porte essentiellement sur les polychètes, les
bivalves, les amphipodes et les cumacés. Or, pour ces quatre taxons, la taille des individus ingérés
diffère entre les groupes de poissons présents à cette profondeur (Tableau II/III.7). Les poids moyens
des proies ingérées ne sont similaires que pour les adultes d'A. laterna et de B. luteum, d'une part, et
pour les juvéniles G1+ de S. impar et les plus petits G0 (LT < 100 mm) de S. solea, de l'autre. Ces
différences de taille des proies ingérées réduisent de façon considérable le recouvrement effectif des
niches alimentaires pour la majorité des groupes de poissons présents à 0-20 m et résultent en une
séparation nette des alimentations de S. solea, S. impar et B. luteum à cette profondeur (Fig. II/III.4).
Les régimes alimentaires d'A. laterna et de B. luteum sont plus semblables, mais les variations
saisonnières d'abondances de ces deux espèces et les fluctuations temporelles des proies ingérées par
leurs différents stades de vie benthiques réduisent la compétition interspécifique entre elles.
Les variations saisonnières de la composition des régimes alimentaires et des groupes de
poissons présents à 0-20 m réduisent donc de façon importante la compétition effective entre S. solea, S.
impar, B. luteum et A. laterna à cette profondeur. Les indices de recouvrements globaux effectifs
(Tableau II/III.8), calculés uniquement à partir de l'alimentation des individus présents simultanément à
0-20 m et tenant compte, dans chaque cas, des effectifs correspondants, fournissent une estimation plus
précise du partage réel des ressources alimentaires entre populations des quatre espèces au niveau des
petits fonds sableux côtiers. Les valeurs trouvées varient de 0,10 à 0,22. Elles sont maximales pour les
soléidés (surtout pour B. luteum et S. solea) mais restent très largement en dessous de la valeur seuil de
0,6. Il n'y a donc pas de possibilité de compétition importante pour les ressources alimentaires entre les
différents stades de vie benthique de S. solea, S. impar, B. luteum et A. laterna présents sur ces fonds.
La compétition alimentaire potentielle y est limitée aux juvéniles de S. solea (G0, LT < 100 mm) et de S.
impar (G1+) et aux adultes de B. luteum et d'A. laterna au printemps, les différences du poids des proies
ingérées réduisant les interactions trophiques entre individus des autres groupes présents à cette
profondeur.
Tableau II/III.8 - Indices de recouvrements globaux effectifs des nichesalimentaires (TE) des populations des espèces présentes à 0-20 m deprofondeur, prises deux à deux.
TE 0 – 20 m S. solea S. impar B. luteum
S. impar 0,20
B. luteum 0,22 0,21
A. laterna 0,15 0,10 0,17

Partage des ressources alimentaires
154
II/III.3.2.2 - PARTAGE DES RESSOURCES ALIMENTAIRES SUR LES VASES TERRIGENES DE
HAUT NIVEAU (30 - 50 m)
Situation à 30-50 m au printemps
Au printemps, sur les fonds vaseux entre 30 et 50 m, la compétition alimentaire potentielle est
surtout interspécifique, avec des recouvrements de niche alimentaire significatifs entre adultes de B.
luteum et juvéniles de S. solea d'une part (T = 0,68), et entre adultes de S. solea et d'A. laterna (T = 0,82)
de l'autre (Tableau II/III.9). La compétition intraspécifique ne concerne véritablement que C. linguatula,
avec un recouvrement de niche significatif entre juvéniles G0, juvéniles G1+ et adultes de l'espèce (T =
0,63-0,78). Le recouvrement des niches est également élevé (bien que non significatif) entre adultes de
S. solea et de B. luteum (T = 0,59) et entre juvéniles et adultes de S. solea (T = 0,58). Le partage des
ressources alimentaires porte essentiellement sur les polychètes, consommés de façon importante (Cp >
45%) par les individus de S. solea, B. luteum et A. laterna, les poissons (Cp > 34% pour tous les
individus de C. linguatula) et les crevettes (Cp > 10% pour les juvéniles G1+ et adultes de C.
linguatula).
Tableau II/III.9 - Indices de recouvrement des niches alimentaires (T) observés au printempsà 30-50 m de profondeur pour les stades de vie benthique des espèces présentes, pris deux àdeux. "+" = valeur inférieure à 0,01. S s = S. solea ; B l = B. luteum ; A l = A. laterna ; C l = C.linguatula. G0 = juvéniles de l'année ; G1+ = juvéniles de plus d'un an ; Ad. = adultes ; LT = longueurtotale en mm.
TS s G0
LT ≥ 100 mmS sAd.
C lG0
C lG1+
C lAd.
B lAd.
S s Ad. 0,58
C l G0 0,04 0,15
C l G1+ 0,04 0,11 0,78
C l Ad. 0,04 0,41 0,63 0,53
B l Ad. 0,68 0,59 + + +
A l Ad 0,51 0,82 0,17 0,12 0,36 0,52
Situation à 30-50 m en été
En été, le recouvrement des niches alimentaires varie de 0,006 à 0,67 entre les groupes de
poissons présents à 30-50 m de profondeur (Tableau II/III.10). La composition du régime alimentaire
diffère de façon importante d'un groupe à l'autre et la compétition alimentaire potentielle est réduite
entre les 4 espèces. Le seul recouvrement significatif (T = 0,67) est observé entre les niches alimentaires
des juvéniles et adultes de S. solea. Il est lié à une consommation très importante de polychètes par ces
deux groupes d'individus pour qui cette catégorie de proies constitue 56 à 91% en poids du total des
proies ingérées.

Partage des ressources alimentaires
155
Tableau II/III.10 - Indices de recouvrement des niches alimentaires (T) observésen été pour les stades de vie benthique des espèces présentes à 30-50 m deprofondeur, pris deux à deux. "+" = valeur inférieure à 0,01. S s = S. solea ; B l = B.luteum ; A l = A. laterna ; C l = C. linguatula. G0 = juvéniles de l'année ; G1+ = juvénilesde plus d'un an ; Ad. = adultes ; LT = longueur totale en mm.
TS s G0
LT ≥ 100 mmS sAd.
C lG1+
B lAd.
S s Ad. 0,67
C l G1+ + +
B l Ad. 0,29 0,02 0,01
A l Ad. 0,20 0,25 0,02 0,42
Situation à 30-50 m en hiver
En hiver, le recouvrement des niches alimentaires varie de 0,001 à 0,88 avec quatre valeurs
significatives (Tableau II/III.11). La compétition alimentaire interspécifique potentielle ne concerne que
B. luteum et S. solea, pour qui le recouvrement des niches alimentaire est, à cette saison, significatif et
élevé quel que soit le stade de vie benthique considéré (T > 0,80). La compétition intraspécifique
potentielle concerne S. solea et C. linguatula avec, pour chacune de ces deux espèces, un recouvrement
de niche élevé et significatif entre juvéniles et adultes (T > 0,86). La compétition alimentaire porte
essentiellement sur les polychètes, consommés de façon très importante (Cp > 78% pour tous les
individus de S. solea et de B. luteum) et les poissons (Cp > 86% pour tous les individus de C.
linguatula).
Tableau II/III.11 - Indices de recouvrement des niches alimentaires (T) observés en hiver à 30-50 m deprofondeur entre les stades de vie benthique des espèces présentes, pris deux à deux. "+" = valeurinférieure à 0,01. S s = S. solea ; B l = B. luteum ; A l = A. laterna ; C l = C. linguatula. G0 = juvéniles de l'année ;G1+ = juvéniles de plus d'un an ; Ad. = adultes ; LT = longueur totale en mm.
T 30-50 mS s G0
LT ≥ 100 mmS sAd.
C lG1+
C lAd.
B lAd.
S s Ad. 0,88
C l G1+ + +
C l Ad. + + 0,86
B l Ad. 0,85 0,80 0,07 +
A l Ad. 0,05 0,08 0,06 0,04 0,17

Partage des ressources alimentaires
156
Conclusion sur le partage des ressources à 30-50 m
Sur les vases terrigènes comprises entre 30 et 50 m de profondeur, la compétition alimentaire
potentielle est minimale en été, sous le double effet d'une réduction du nombre de groupes de poissons
présents et d'une augmentation des différences entre régimes alimentaires. Elle est maximale en hiver
tant sur le plan interspécifique, entre soléidés, que sur le plan intraspécifique, pour S. solea et C.
linguatula. A cette profondeur, le partage des ressources alimentaires porte essentiellement sur les
polychètes et les poissons. Or, pour ces deux taxons, de grandes différences de taille existent entre les
proies ingérées par les juvéniles et les adultes des quatre espèces étudiées (Tableau II/III.12). Ce
phénomène réduit le recouvrement réel des niches alimentaires des groupes de poissons présents à 30-
50 m et résulte en une nette séparation des alimentations des quatre espèces à cette profondeur (Fig.
II/III.5). Bien qu'A. laterna et B. luteum présentent, à 30-50 m, les alimentations les plus similaires, les
fluctuations saisonnières de leur alimentation réduisent la compétition interspécifique effective entre
leurs populations.
Tableau II/III.12 - Poids moyen (mg) des proies ingérées à 30-50 m par deux groupes de poissonsou plus. Pour chaque groupe de poissons, seules sont indiquées les catégories de proiesreprésentant plus de 20% en poids du poids total des proies ingérées, avec en gras, les proiespréférentielles. S s = S. solea ; B l = B. luteum ; A l = A. laterna ; C l = C. linguatula. G0 =juvéniles de l'année ; G1+ = juvéniles de plus d'un an ; Ad. = adultes ; LT = longueur totale en mm.
Polychètes Bivalves Macroures Mysidacés Poissons
S s G0 LT ≥ 100 1,52 3,92 0,60
S s Ad. 2,78
C l G0 0,37 6,06
C l G1+ 6,95 1,69 38,22
C l Ad. 48,72 1,14 195,81
B l Ad. 0,76 2,28
A l Ad 4,22 4,15 0,20
Pour conclure, à 30-50 m de profondeur, les variations saisonnières de la composition des
régimes alimentaires et le nombre réduit de groupes de poissons présents à chaque saison limitent de
façon importante la compétition effective globale entre S. solea, B. luteum, A. laterna et C. linguatula.
Les indices de recouvrements globaux effectifs calculés pour les populations des quatre espèces varient
de 0,003 à 0,36 et sont très largement en dessous de la valeur seuil de 0,6 (Tableau II/III.13). De plus,
pour tous les groupes de poissons présents à cette profondeur, des différences de poids des proies
réduisent les interactions trophiques entre individus de tailles différentes. L'ensemble de ces facteurs fait
qu'il n'y a pas de compétition réelle possible pour les ressources alimentaires entre les différents stades
de vie benthique de S. solea, B. luteum, A. laterna et C. linguatula présents sur ces fonds.

Partage des ressources alimentaires
157
Figure II/III.5 - Positionnement des groupes alimentaires présents à 30-50 m dans le plan obtenu par ordination desespèces en fonction de leur alimentation (N.M.S. réalisée sur les individus groupés par stade de vie benthique /saison / profondeur, solution à 2 dimensions). Les couleurs indiquent les saisons : printemps (vert), été (jaune), hiver(bleu).
Tableau II/III.13 - Indices de recouvrement globaux effectifs des nichesalimentaires (TE) observés entre les populations des espèces présentes à 30-50m de profondeur, prises deux à deux. "+" = valeur inférieure à 0,01.
TE 30 – 50 m S. solea B. luteum A. laterna
B. luteum 0,36
A. laterna 0,18 0,09
C. linguatula 0,03 + 0,06
Al
Al
Al
BlCl
Cl
Cl
Cl
Cl
Cl
SsSs
Ss
SsSs
Ss
Bl
-2
0
2
-2 0 2
Axe 1
Axe 2
Stress = 0,131
été
printemps
hiver
Saison
juvéniles G0(LT < 100 mm)
adultes
juvéniles G1+ et G0 de LT > 100 mm
Stade de vie benthique

Partage des ressources alimentaires
158
II/III.3.2.3 - PARTAGE DES RESSOURCES ALIMENTAIRES SUR LES VASES TERRIGENES
PROFONDES ( 70-100 m)
Situation à 70-100 m au printemps
Au printemps, le recouvrement des niches alimentaires des poissons plats présents entre 70 et
100 m de profondeur varie de 0,001 à 0,74 avec quatre valeurs supérieures à 0,6 (Tableau II/III.14). La
compétition alimentaire potentielle est essentiellement interspécifique, entre adultes de B. luteum et S.
solea d'une part (T = 0,63), et adultes de C. linguatula et d'A. laterna de l'autre (T = 0,60). La
compétition intraspécifique potentielle ne concerne que C. linguatula, avec un recouvrement de niche
significatif entre juvéniles G0 et juvéniles G1+ (T = 0,74) et entre G1+ et adultes (T = 0,66). Le partage
des ressources alimentaires porte essentiellement sur les poissons (Cp > 40% pour tous les individus de
C. linguatula), les macroures (Cp > 56% pour les adultes de C. linguatula et d'A. laterna) et les
polychètes (Cp > 62% pour les adultes de S. solea et de B. luteum).
Situation à 70-100 m en été
En été, la composition du régime alimentaire diffère de façon importante entre les groupes
présents à 70-100 m. Aucun des indices de recouvrement des niches alimentaires calculés n'est
significatif (Tableau II/III.15). Il n'y a donc pas de réelle compétition possible pour les ressources
alimentaires à cette 70-100 m en été.
Situation à 70-100 m en hiver
En hiver, le recouvrement des niches alimentaire varie de 0,002 à 0,80 (Tableau II/III.16). Il n'y
a pas, à cette saison, de compétition interspécifique pour les ressources alimentaires à 70-100 m. La
seule valeur de recouvrement significative est observée entre juvéniles G1+ et adultes de C. linguatula et
provient d 'une consommation importante de poissons par ces deux groupes (Cp > 80%).
Conclusion sur le partage des ressources à 70-100 m
Entre 70 et 100 m, la compétition potentielle pour les ressources alimentaires entre S. solea, B.
luteum, A. laterna et C. linguatula est minimale en été et maximale au printemps. Elle concerne surtout
C. linguatula pour qui le recouvrement intraspécifique des niches alimentaires est significatif en hiver et
au printemps quel que soit le stade de vie benthique. Le partage des ressources alimentaires porte
essentiellement, à cette profondeur, sur les poissons, les macroures et les polychètes. Les différences de
taille des proies consommées par les juvéniles et les adultes des quatre espèces (Tableau II/III.17)
réduisent le recouvrement réel des niches alimentaires des groupes de poissons présents à 70-100 m et
augmente encore leur séparation (Fig. II/III.6).

Partage des ressources alimentaires
159
Tableau II/III.14 - Indices de recouvrement des niches alimentaires (T) observésau printemps pour les stades de vie benthique des espèces présentes à 70-100 mde profondeur, pris deux à deux. "+" = valeur inférieure à 0,01. S s = S. solea ; B l =B. luteum ; A l = A. laterna ; C l = C. linguatula. G0 = juvéniles de l'année ; G1+ =juvéniles de plus d'un an ; Ad. = adultes.
T 70-100 m
S sAd.
C lG0
C lG1+
C lAd.
B lAd.
C l G0 +C l G1+ 0,03 0,74C l Ad. + 0,43 0,66B l Ad. 0,63 + + +A l Ad 0,08 0,16 0,33 0,60 0,11
Tableau II/III.15 - Indices de recouvrement des niches alimentaires (T)observés en été à 70-100 m de profondeur entre les stades de viebenthique des espèces présentes, pris deux à deux. A l = A. laterna ; C l =C. linguatula. G1+ = juvéniles de plus d'un an ; Ad. = adultes.
T 70-100 m
C lG1+
C lAd.
C l Ad. 0,27A l Ad. 0,25 0,46
Tableau II/III.16 - Indices de recouvrement des niches alimentaires (T)observés en hiver à 70-100 m de profondeur entre les stades de viebenthique des espèces présentes, pris deux à deux. "+" = valeur inférieure à0,01. S s = S. solea ; B l = B. luteum ; A l = A. laterna ; C l = C. linguatula.. G1+ =juvéniles de plus d'un an ; Ad. = adultes.
T 70-100 mS sAd.
C lG1+
C lAd.
B lAd.
C l G1+ +C l Ad. 0,04 0,80B l Ad. 0,20 0,09 0,08A l Ad 0,05 0,09 0,08 0,55
Tableau II/III.17 - Poids moyen (mg) des proies ingérées à 70-100 m par deux groupes depoissons ou plus. Pour chaque groupe de poissons, seules sont indiquées les catégories deproies représentant plus de 20% en poids du poids total des proies ingérées, avec en gras,les proies préférentielles. S s = S. solea ; B l = B. luteum ; A l = A. laterna ; C l = C. linguatula. G0= juvéniles de l'année ; G1+ = juvéniles de plus d'un an ; Ad. = adultes.
Polychètes Macroures Mysidacés Poissons
S s Ad. 5,30
C l G0 0,22 5,46
C l G1+ 10,00 0,69 102,73
C l Ad. 38,06 0,63 166,06
B l Ad. 0,40 7,48 1,94
A l Ad 8,85 1,40 0,08

Partage des ressources alimentaires
160
Figure II/III.6 - Positionnement des groupes alimentaires présents à 70-100 m dans le plan obtenu parordination des espèces en fonction de leur alimentation (N.M.S. réalisée sur les individus groupés par stade devie benthique / saison / profondeur, solution à 2 dimensions). Les couleurs indiquent les saisons : printemps (vert),été (jaune), hiver (bleu).
Pour conclure, à 70-100 m de profondeur, les variations saisonnières de la composition des
régimes alimentaires et le nombre réduit de groupes de poissons présents à chaque saison évitent toute
compétition effective entre S. solea, B. luteum, A. laterna et C. linguatula. Les indices de recouvrements
globaux effectifs calculés pour les populations des quatre espèces à 70-100 m varient de 0,001 à 0,19 et
sont très largement en dessous de la valeur seuil de 0,6 fixée par Keast (1978) pour une compétition
alimentaire significative (Tableau II/III.18).
Tableau II/III.18 - Indices de recouvrement globaux effectifs des niches alimentaires(TE) observés pour les populations des cinq espèces présentes à 70-100 m deprofondeur, prises deux à deux. "+" = valeur inférieure à 0,01.
TE 70-100 m S. solea S. impar B. luteum
S. impar 0,15
B. luteum 0,02 0,06
A. laterna + + 0,19
Al Al
Al
Bl
Bl
Cl
Cl Cl
Cl
Cl Cl
Cl
Ss
Ss
-2
0
2
-2 0 2
Axe 1
Axe 2
Stress = 0,131
été
printemps
hiver
Saison
juvéniles G0(LT < 100 mm)
adultes
juvéniles G1+ et G0 de LT > 100 mm
Stade de vie benthique

Partage des ressources alimentaires
161
II/III.4 - CONCLUSION SUR LE PARTAGE DES RESSOURCES ALIMENTAIRES ENTRE
POISSONS PLATS AU LARGE DU RHONE
Au large du Rhône, la compétition pour les ressources alimentaires entre les cinq espèces de
poissons plats étudiées est réduite, non seulement par des modifications spatio-temporelles de
répartition de leurs individus mais également par des variations saisonnières de l'alimentation des
différents stades de vie benthique. Quelle que soit la saison, le recouvrement des niches alimentaires est
plus intense sur les petits fonds sableux côtiers (0-20 m) que sur les vases terrigènes plus profondes (30-
100 m). Pour toutes les tranches bathymétriques, les interactions d'ordre trophique sont maximales au
printemps, période où le nombre de groupes de poissons présents à chaque profondeur est le plus élevé
et les alimentations les plus similaires. Quelle que soit la saison et/ou la profondeur, l'alimentation de C.
linguatula est totalement distincte de celle des quatre autres poissons plats étudiés. De même, en dépit
de valeurs de recouvrements de niche souvent élevées entre individus de B. luteum, S. impar et S. solea,
les similitudes alimentaires entre ces trois espèces de soléidés sont réduites par les différences
importantes de poids des proies ingérées. Les ressemblances alimentaires sont maximales à chaque
profondeur entre B. luteum et A. laterna, pour lesquelles le poids des proies ingérées diffère peu (Fig.
II/III.7). Cependant, les variations spatio-temporelles de leurs alimentations respectives réduisent la
compétition effective entre les classes de taille présentes à une saison donnée dans la même tranche
bathymétrique.
Figure II/III.7 - Positionnement des groupes alimentaires présents au large du Rhône dans le plan obtenu parordination des cinq espèces en fonction de leur alimentation (N.M.S. réalisée sur les individus groupés par stade devie benthique / saison / profondeur, solution à 2 dimensions). Les différences de couleurs indiquent les profondeurs : 0-20m (bleu-vert), 30-50 m (bleu ciel), 70-100 m (bleu foncé).
Al
Al
Al
AlAl
Al
Al
AlAl
AlAl
Bl
Bl
Bl
Bl
BlBl
Bl
Bl
Bl
Bl
Cl Cl
Cl
Cl
Cl
ClCl
Cl
Cl
ClCl
Cl
Cl
Si
SiSi
SiSi
Ss
Ss
SsSs
SsSs
Ss Ss SsSs
SsSs
Ss
Ss
-2
0
2
-2 0 2Axe 1
Axe
2
70 - 100 m
30 - 50 m
0 - 20 m
Tranchebathymétrique
juvéniles G0 (LT < 100 mm)
adultes
juvéniles G0 et G1+
de LT ≥ 100 mm
Stade de viebenthique
Stress = 0,131

Partage des ressources alimentaires
162
Les interactions trophiques directes entre individus des cinq espèces étudiées sont donc réduites
au large du Rhône. Les recouvrements effectifs globaux sont très faibles et varient de 0,001 à 0,12
(Tableau II/III.19). La seule compétition pour les ressources alimentaires envisageable est limitée aux
juvéniles de S. solea (G0 : LT < 100 mm) et de S. impar (G1+) et aux adultes de B. luteum et A. laterna à
0-20 m. Or, les différences de rythme d'alimentation réduisent les interactions entre ces différents
groupes de poissons, dans la mesure où S. solea et B. luteum se nourrissent surtout de nuit, tandis qu'A.
laterna et (vraisemblablement) S. impar s'alimentent essentiellement de jour. A ceci s'ajoutent des
différences spécifiques des proies consommées, notamment en ce qui concerne les familles de
polychètes.
Tableau II/III.19 - Indices de recouvrement globaux effectifs des niches alimentaires (TE)observés sur l'ensemble de la zone d'étude (0-100 m) entre les populations des cinq espèces,prises deux à deux. "+" = valeur inférieure à 0,01 ; "─" = valeur absente.
TE S. impar B. luteum A. laterna C. linguatula
S. solea 0,12 0,10 0,05 +
S. impar 0,04 0,02 —
B. luteum 0,04 +
A. laterna 0,03
Ainsi, des modifications saisonnières de régime alimentaire ainsi que des différences de
répartition spatio-temporelle, de rythme d'alimentation et de taille des proies ingérées réduisent les
interactions trophiques entres les cinq espèces de poissons plats étudiées qui se partagent les ressources
alimentaires disponibles au large du Rhône sans qu'il y ait de réelle compétition entre elles. Ce résultat
sera discuté ultérieurement en termes de stratégies de vie des poissons (c. f. Partie IV).

PARTIE III
RESEAUX TROPHIQUES ET SUIVI DE LA MATIERE ORGANIQUE PARTICULAIRE TERRIGENE


Réseaux trophiques et suivi de la MOP terrigène
163
RESEAUX TROPHIQUES ET SUIVI DE LA MATIERE
ORGANIQUE PARTICULAIRE TERRIGENE
III.1 - INTRODUCTION
Dans les écosystèmes marins côtiers situés à proximité de l'embouchure des fleuves, l'étude des
flux de matière organique dans les réseaux trophiques benthiques se heurte à un certain nombre de
difficultés. Dans ce type d'écosystèmes, les sources potentielles de matière organique à la base des
chaînes trophiques sont multiples et variables (Mann, 1982, 1988). Elles se différencient non seulement
par leur provenance (production in situ ou apports allochtones) mais également par leur nature
(organismes vivants ou formes détritiques) et sont caractérisées par une importante variabilité spatiale et
temporelle (blooms phytoplanctoniques saisonniers, apports continentaux variables entre les périodes de
crues et d'étiage). Le caractère détritique de la matière organique et la variabilité des apports rendent
particulièrement difficile la détermination de son origine et l'estimation de la contribution des
différentes sources primaires à la production secondaire. De plus, différentes sources de matière
organique constituant la base du réseau trophique (phytoplancton, détritus, bactéries) sont étroitement
associées en un "pool particulaire" indifférencié (Knox, 1986). Il est alors difficile de classer les
consommateurs benthiques en catégories trophiques simples (i.e., herbivores, carnivores, détritivores)
dans la mesure où peu d'organismes se nourrissent sélectivement à partir d'un composant unique du pool
particulaire. La plupart des consommateurs primaires benthiques sont omnivores et consomment, selon
leurs disponibilités dans le milieu, un mélange variable de microalgues, détritus, bactéries, protozoaires,
champignons, mais aussi de petites espèces méiobenthiques ou de formes larvaires (Massé, 1972a ;
Haines et Montague, 1979 ; Kuipers et al., 1981). Dans ces conditions, la connaissance des régimes
alimentaires se révèle difficile car aucune méthode d'étude classique ne s'avère parfaitement adaptée
pour résoudre une telle complexité et déterminer l'origine première de la matière organique consommée.
Dans ce domaine, l'analyse des contenus stomacaux présente de nombreuses limites (Pinnegar et
Polunin, 2000). Elle reflète souvent un état instantané et donc partiel de l'alimentation d'une population.
De plus, l'assimilation différentielle des aliments ingérés peut biaiser l'image fournie par l'étude des
contenus stomacaux (Gearing et al., 1984 ; Stoner et Zimmerman, 1988). Les sources de nourriture les
plus rapidement digérées deviennent vite difficilement identifiables et leur importance alimentaire
risque d'être sous estimée. Au contraire, certaines catégories de proies sont ingérées mais peu ou pas
assimilées et leur contribution dans l'alimentation des organismes est alors surestimée.

Réseaux trophiques et suivi de la MOP terrigène
164
Nombre de ces difficultés peuvent être levées par l'utilisation du traçage isotopique. L'analyse
des compositions tissulaires en isotopes stables fournit un enregistrement intégré et continu des sources
de nourriture utilisées par les organismes pour leur croissance (Dufour et Gerdeaux, 2001). Elle est de
ce fait moins sensible aux biais temporels que l'étude des contenus stomacaux et prend en compte
l'ensemble des sources de nourriture assimilées par l'organisme, en contournant le biais lié à la digestion
différentielle (Pinnegar et Polunin, 2000). C'est pourquoi l'analyse des rapports isotopiques de la matière
organique s'est peu à peu imposée pour identifier les sources nutritives des organismes (Van Dover et
al., 1992 ; Riera et Richard, 1996 ; Hsieh et al., 2000 ; Chong et al., 2001) et caractériser les transferts
de carbone entre les différents niveaux trophiques (Peterson et al., 1985 ; Thomas et Cahoon, 1993 ;
Polunin et al., 2001 ; Takai et al., 2002). Depuis les premières études dans les années 70 (Parker et
Calder, 1970 ; Haines, 1976), la méthode a été de plus en plus utilisée pour tracer l'origine de la matière
ingérée par les animaux marins, du fait de l'existence de signatures isotopiques souvent très différentes
entre les sources potentielles de matière organique à la base des réseaux trophiques océaniques (Owens,
1987 ; Peterson et Fry, 1987 ; Ostrom et Fry, 1993). Dans les écosystèmes marins côtiers sous influence
continentale, l'utilisation des isotopes stables s'est révélée particulièrement concluante (Riera, 1995 ;
Deegan et Garritt, 1997 ; Riera et Richard, 1997 ; Lee, 2000). Cependant, peu d'études ont été réalisées
en Méditerranée (Pinnegar, 2000 ; Pinnegar et Polunin, 2000 ; Polunin et al., 2001) où les travaux
concernant les substrats meubles sont rares (Dauby et Mosora, 1988 ; Dauby, 1989 ; Camusso et al.,
1998 ; Camusso et al., 1999).
Le but de ce chapitre est de déterminer, à l'aide des isotopes stables du carbone et de l'azote, les
voies éventuelles d'incorporation de la MOP terrigène dans les réseaux trophiques au large du Rhône et
de décrire son cheminement potentiel jusqu'aux populations de poissons démersaux étudiées. Pour cela,
les signatures isotopiques des juvéniles et des adultes de S. solea, S. impar, B. luteum, A. laterna et C.
linguatula ont été établies et comparées à celles de leurs proies afin de déterminer les composantes
majeures de leurs réseaux trophiques respectifs aux différentes profondeurs étudiées (0-20 m, 30-50 m
et 70-100 m). Les flux de matière organique d'origine terrestre et d'origine marine dans les réseaux
trophiques benthiques ont également été étudiés à ces différentes profondeurs afin de déterminer
l'impact potentiel des apports du Rhône en MOP terrigène sur les stades de vie benthique de chaque
espèce de poisson.

Réseaux trophiques et suivi de la MOP terrigène
165
III.2 - MATERIEL ET METHODES
III.2.1 - L'OUTIL "ISOTOPES STABLES"
Au cours des vingt dernières années, l'étude de la composition en isotopes stables de différents
éléments (carbone, azote, oxygène, soufre…) s'est peu à peu imposée comme le principal "outil"
méthodologique pour tracer les flux de matière organique dans les réseaux trophiques aquatiques, qu'ils
soient lacustres (Hesslein et al., 1991 ; Cabana et Rasmussen, 1994 ; Kiriluk et al., 1995 ; Vander
Zanden et al., 1999 ; Vander Zanden et Rasmussen, 2001) ou marins (Mc Connaughey et Mc Roy, 1979
; Fry et al., 1984 ; Polunin et al., 2001 ; Takai et al., 2002). Cette approche est essentiellement basée sur
le fait que la composition isotopique d'un organisme est similaire ou varie de façon définie par rapport à
celle de sa nourriture (De Niro et Epstein, 1978, 1981 ; Minagawa et Wada, 1984). L'analyse des
signatures isotopiques des composantes d'un réseau trophique permet donc de décrire non seulement les
relations trophiques entre les organismes mais également les transferts de matière le long des chaînes
alimentaires.
Isotopes stables : définition, propriétés et méthode de mesure
Les isotopes sont des éléments chimiques possédant le même nombre d'électrons et de protons,
mais un nombre différent de neutrons, ce qui résulte en un nombre de masse particulier à chaque
isotope. La plupart des éléments ont plusieurs isotopes stables (hydrogène : 1 H, D ; carbone : 12 C, 13 C ;
azote : 14 N, 15 N ; oxygène : 16 O, 17 O, 18 O ; soufre : 32 S, 33 S, 34 S, 36 S ; etc.). Dans la nature, un des
isotopes est toujours nettement majoritaire par rapport aux autres (Tableau III.1) et la composition
isotopique des éléments n'est pas absolument stable.
Tableau III.1 - Abondances naturelles moyennes des isotopes stables du carbone et de l'azote(d'après Riera, 1995).
Elément Isotope majoritaire (% du total) Isotope minoritaire (% du total)
Azote 14N (99,63 %) 15N (0,37 %)
Carbone 12C (98,89 %) 13C (1,11 %)
De par les légères variations de masse qui existent entre elles, les formes isotopiques d'un même
élément présentent des propriétés physico-chimiques différentes (densité, volume molaire, température
de vaporisation et de condensation, conditions de précipitation, pression de vapeur, viscosité). De
même, la vitesse des réactions et les constantes d'équilibre sont influencées par la composition
isotopique. Ainsi, au cours des réactions physiques, chimiques ou biologiques, il se produit un
fractionnement isotopique qui résulte en des différences de composition isotopique entre réactifs et
produits formés (Owens, 1987). Les variations d'abondances isotopiques naturelles résultent de ce

Réseaux trophiques et suivi de la MOP terrigène
166
phénomène de fractionnement. Elles sont très faibles et ne dépassent que rarement quelques millièmes.
Leur mesure nécessite donc l'utilisation d'appareils très précis et un mode de notation approprié.
Les compositions isotopiques sont mesurées à l'aide de spectromètres de masse à source gazeuse
de grande précision analytique et exprimées par la notation conventionnelle "delta" (δ) : pour un
élément X, le rapport (R) "isotope le moins abondant/isotope le plus abondant" (qui correspond en
général au rapport "isotope lourd/isotope léger") dans l'échantillon est comparé à la mesure du même
rapport pour une référence internationale dont la valeur est fixée à 0. La référence internationale pour le
carbone, le PDB, est un rostre de Belemnite fossile provenant de la Pee Dee Formation aux Etats-Unis.
Pour l'azote, le N2 atmosphérique est utilisé. Les variations mesurées étant généralement très faibles, les
valeurs sont multipliées par mille et la signature isotopique est exprimé en ‰. On a donc, pour chaque
élément X :
δX (‰) = [(R échantillon / R référence) -1] × 1000
Par convention, l'isotope le moins abondant (isotope lourd) est utilisé pour la notation δX (δ13C,
δ15N, δ34S, etc). Le signe (positif ou négatif) de delta dépend essentiellement des abondances relatives
des isotopes lourd et léger de l'élément considéré dans l'échantillon.
Signatures isotopiques des différentes sources de matière organique
Les principales sources de matière organique à la base des réseaux trophiques (producteurs
primaires) possèdent des signatures isotopique différentes (Tableau III.2) car elles dépendent non
seulement de la composition isotopique des éléments minéraux nutritifs originels, mais également des
cycles biochimiques utilisés au cours de la synthèse de la matière organique.
Tableau III.2 - Compositions isotopiques (δ en ‰) typiques en carbone et en azote des principauxproducteurs primaires en milieu terrestre et aquatique. D'après la revue réalisée par Ostrom et Fry (1993).
δ13 C δ 15N δ 34S
Plantes en C3 (terrestres) -30 à -23 -7 à -6 -7 à 14
Plantes en C3 (marais) -29 à -23 3 à 5 6
Plantes en C4 (marais) -15 à -12 1 à 8 -10 à 6
Phytoplancton marin -24 à -18 -2 à 12 18 à 19
Phytoplancton estuarien -30 à -15 2 à 19 18 à 20
Macrophytes benthiques -27 à -10 -1 à 10 15 à 20
Phanérogames marines -16 à -4 0 à 6 -13 à 15
Mangroves -29 à -25 6 à 7 5 à 14

Réseaux trophiques et suivi de la MOP terrigène
167
Parmi les éléments les plus utilisés pour le traçage isotopique (C, N, S, O), le carbone est
particulièrement efficace pour la caractérisation de l'origine (terrestre/marine, benthique/pélagique) de la
matière organique. Le δ13C varie de façon importante entre les différents producteurs primaires. En
effet, le carbone fixé par les organismes autotrophes est directement dérivé du pool de carbone
inorganique présent dans leur environnement immédiat. Les plantes terrestres utilisent le CO2
atmosphérique alors que les producteurs primaires aquatiques (phytoplancton, algues, phanérogames
marines) effectuent leur photosynthèse à partir du carbone inorganique dissous dans l'eau (CID). Or, le
fractionnement isotopique complexe qui se produit lors de la dissolution du CO2 dans l'eau résulte en
des différences de signature entre le CO2 atmosphérique (-7‰), et le CID présent dans les eaux douces
(-15‰) ou en mer (0‰) (Faure, 1986 ; Peterson et Fry, 1987). Ce phénomène explique, en partie, les
différences de δ13C observées entre les producteurs primaires de ces trois milieux. Cependant, le
caractère plus ou moins fractionnant des enzymes utilisées au cours de la synthèse de la matière
organique peut également engendrer de fortes variations de la composition isotopique. Dans le cas de la
photosynthèse, deux types principaux sont classiquement considérés en fonction de l'enzyme utilisée
pour la fixation initiale du carbone : (1) celui qui utilise la ribulose-1,5-biphosphate (RuBP), appelé
cycle de Calvin ou cycle C3, qui passe par la formation de molécules à 3 atomes de carbone, et (2) celui
qui utilise la phosphoénolpyruvate (PEP), appelé cycle de Hatch-Slack, ou cycle C4, qui produit des
molécules à 4 atomes de carbone. Ces deux types photosynthétiques ne conduisent pas à des
fractionnements isotopiques égaux. Ainsi, en milieu terrestre, à partir du même CO2 atmosphérique (-7
‰), les plantes en C3 (l'essentiel des végétaux terrestres et aquatiques des régions tempérées) ont un
δ13C moyen de -28‰ alors que les végétaux en C4 (xérophytes des biotopes secs ou salés,
légumineuses, Agaves, Crassulacées…) ont un δ13C moyen de -12‰ (Smith et Epstein, 1971 ; Peterson
et Fry, 1987 ; Boutton, 1991).
Par la combinaison de ces deux mécanismes, les producteurs primaires des milieux terrestres,
dulçaquicoles, océaniques ou des marais maritimes ont le plus souvent des signatures isotopiques en
carbone clairement distinctes, dans une gamme de variation assez large (entre -40 et -5‰). La
connaissance du δ13C de ces organismes permet donc d'estimer leurs contributions respectives à la
matière organique particulaire (MOP) des milieux côtiers. Dans les systèmes estuariens, les δ13C de la
MOP attribuée aux apports terrestres sont généralement compris entre -31 et -26‰ (Simenstad et
Wissmar, 1985 ; Riera et Richard, 1996 ; Camusso et al., 1999 ; Hsieh et al., 2000). A l'opposé, le
phytoplancton marin montre des δ13C variant essentiellement de -22 à -19‰ (Fontugne et Duplessy,
1981). Ainsi, les δ13C des diverses sources de MOP présentes dans les écosystèmes côtiers ont été
largement utilisés pour examiner divers aspects du cycle du carbone dans ces milieux. Ils ont
notamment permis de déterminer la source de MOP dominante dans les matières en suspension ou dans
les sédiments de plusieurs systèmes marins et estuariens (Gearing et al., 1984 ; Fichez et al., 1993 ;
Kerhervé et al., 2001).

Réseaux trophiques et suivi de la MOP terrigène
168
Relation entre les compositions isotopiques d'un consommateur et de sa nourriture
Une fois fixée dans la matière organique, la composition isotopique ne subit que de faibles
variations lors des processus de dégradation et des transferts dans les chaînes alimentaires (Ostrom et
Fry, 1993). C'est sur cette relation étroite qu'est basée l'utilisation des abondances naturelles en isotopes
stables pour l'étude des relations trophiques dans les milieux naturels. Il existe cependant, quel que soit
l'élément, une légère différence de composition isotopique entre un animal et sa nourriture, le
consommateur étant très généralement enrichi en isotope lourd par rapport à sa nourriture.
L'enrichissement est faible pour le carbone, avec des différences de δ13C entre un organisme et sa source
de nourriture généralement de l'ordre de 1‰ et ne dépassant jamais 2‰ (De Niro et Epstein, 1978 ;
Peterson et Fry, 1987 ; Pinnegar et Polunin, 1999 ; Vander Zanden et Rasmussen, 2001). Du fait de
cette forte conservation de la composition isotopique en carbone de la proie au prédateur, la
connaissance des δ13C d'un animal et des sources potentielles de nourriture permet de déterminer
rapidement l'origine principale de la matière organique ingérée par cet organisme au cours de sa
croissance. Pour l'azote, l'enrichissement en isotope lourd est plus important. Il varie suivant les
organismes mais le δ15N d'un animal montre en moyenne un enrichissement de 3,4‰ par rapport à sa
nourriture (De Niro et Epstein, 1981 ; Minagawa et Wada, 1984 ; Vander Zanden et Rasmussen, 2001).
Lorsque ce fractionnement constant est pris en compte, les signatures isotopiques en azote sont
également de bons indicateurs de l'origine de la matière organique.
L'utilisation couplée des isotopes du carbone et de l'azote permet un suivi efficace du devenir de
la matière organique dans les réseaux trophiques. En effet, certaines sources de matière organique ou
certaines catégories de proies ne peuvent être différenciées sur la base d'un seul isotope. De même, le
mélange, dans certaines proportions, de deux sources (proies) isotopiquement distinctes peut présenter
un rapport isotopique équivalent à celui d'une troisième source (proie), ce qui empêche l'identification
précise de cette dernière. L'utilisation simultanée de deux traceurs isotopiques (ici le carbone et l'azote)
est susceptible de lever ce type d'ambiguïté grâce à une séparation bidimensionnelle de chaque source
de matière organique. De plus, la représentation graphique des signatures isotopiques (δ15N versus δ13C)
des différents composants d'un même réseau trophique (i.e. basé sur une seule et même source de
matière organique) montre, du fait de l'enrichissement trophique en isotope lourd de la proie au
prédateur, une relation linéaire de pente positive (Fig. III.1). Ce continuum trophique est vérifié dans de
nombreux réseaux trophiques terrestres et marins, quels que soient la latitude, le type de substrat, ou la
profondeur (Dickson, 1987 ; Harrigan et al., 1989 ; Wada et al. 1987b) et devrait permettre de séparer
rapidement, au large du Rhône, les organismes benthiques dépendant principalement des sources de
production primaire marine de ceux dépendant surtout des apports en MOP terrestre (Fig. III.2).

Réseaux trophiques et suivi de la MOP terrigène
169
Figure III.1 – Représentation d'un réseau trophique théorique basé sur l'utilisation d'une sourcede matière organique unique. P1 = producteur primaire, C1 = consommateurs primaires (herbivores),C2 = consommateurs secondaires (carnivores).
Figure III.2 – Représentation d'un réseau trophique théorique basé sur l'utilisation de deuxsources de matière organique distinctes A (en blanc) et B (en noir). P1 = producteur primaire, C1 =consommateurs primaires (herbivores), C2 = consommateurs secondaires (carnivores). Les points grisreprésentent les organismes (C1, C2) dépendant des deux sources de matière organique.
δ15N
δ13C
0
P1
(source de matièreorganique unique)
C1
C2
+3,4‰+1‰
δ15N
δ 13C
0
P1
C1
C2
+1‰+3,4‰
RéseauTrophique Aseulement
Réseautrophique Bseulement
A
B

Réseaux trophiques et suivi de la MOP terrigène
170
Comme l'enrichissement en isotope lourd se produit à chaque saut vers un niveau trophique
supérieur, l'étude des compositions isotopiques en azote fournissent également une base solide pour
calculer le niveau trophique des organismes et établir la structure des chaînes alimentaires dans un
écosystème (Vander Zanden et al., 1997). Ainsi, l'utilisation conjointe des isotopes de l'azote et du
carbone devrait permettre, grâce aux avantages respectifs présentés par ces deux éléments, une
description précise de la structure des réseaux trophiques au large du Rhône et un suivi efficace du
devenir des différentes sources de matière organique (marine et terrestre) le long des chaînes trophiques
benthiques.
III.2.2 - PREPARATION DES ECHANTILLONS
Les échantillons de poisson (muscle blanc natatoire dorsal), de proies benthiques (animaux
entiers), de zooplancton, de sédiment superficiel, de végétaux (phytoplancton marin, débris de
macrophytes marines et de végétaux terrestres) et de seston (MOP rhodanienne et de l'eau de la zone
d'étude) récoltés ont été préparés en vue des analyses isotopiques. Pour cela, les échantillons conservés
au congélateur (-30°C), ont été lyophilisés puis réduits en poudre fine (< 60 µm) à l'aide d'un mortier en
agate et d'un tamis en acier, avant d'être stockés dans un dessiccateur. Afin d'éviter toute contamination
des échantillons au cours de la dissection et du broyage, le matériel utilisé a été passé aux ultrasons
(tamis) ou lavé à l'acide chlorhydrique 1% (mortier, pilon, scalpel, pinces) puis rincé à l'eau distillée et
séché en étuve entre chaque individu. En ce qui concerne les invertébrés benthiques, les individus de
chaque catégorie de proie ont été lyophilisés groupés par saison et par profondeur après avoir été, dans
la mesure du possible, disséqués sous la loupe binoculaire afin d'éliminer leur tube digestif et un
maximum de tissus durs (coquilles pour les mollusques, exosquelette pour les crustacés).
Les carbonates inorganiques ont un δ13C particulier (0‰) et peuvent affecter de façon
importante la signature isotopique en carbone de certains échantillons. Leur élimination est donc
conseillée pour l'analyse de cet élément (De Niro et Epstein, 1978). Cependant, les méthodes
d'acidification employées dans ce but modifient de façon significative le δ15N des échantillons (Pinnegar
et Polunin, 1999). Pour cette raison, les échantillons obtenus pour le sédiment, les végétaux, le seston, le
zooplancton et les invertébrés benthiques ont été divisés, chacun, en deux sous-échantillons dont un seul
a été acidifié (HCl 1M), rincé à l'eau distillée, re-séché et re-broyé. L'autre a été conservé intact pour les
analyses de δ15N. En ce qui concerne les poissons, l'utilisation de muscle blanc a permis d'éviter l'étape
d'acidification des échantillons. En effet, celui-ci, très pauvre en carbonates et en lipides (Pinnegar et
Polunin, 1999), ne nécessite aucune préparation particulière en vue des analyses isotopiques. De plus, il
présente, parmi tous les tissus mous analysables chez les poissons, la plus faible variabilité
interindividuelle en δ13C et δ15N ce qui réduit considérablement les risques d'erreurs d'interprétation des
signatures liés à la taille parfois réduite des effectifs analysés.

Réseaux trophiques et suivi de la MOP terrigène
171
III.2.3 - ANALYSES ISOTOPIQUES
Les analyses isotopiques en δ13C et δ15N ont été effectuées lors de deux séjours (janvier 2000 et
janvier 2001) au sein du département "Marine Sciences and Coastal Management" de l'Université de
Newcastle-upon-Tyne (U.K.). Les compositions isotopiques des différents échantillons ont été
déterminées par spectrométrie de masse isotopique en flot continu à combustion assistée par un
analyseur élémentaire (EA- IRMS) (Barrie et Prosser, 1996). Compte tenu des propriétés de l'appareil
utilisé (Europa Scientific "ANCA-SL/PDZ 20-20" system), la masse utile de matériel sec à analyser
dans chaque cas (tissus animal, végétaux, sédiment) a été déterminée afin de disposer d'un minimum de
150 µg d'azote et 200 µg de carbone par échantillon, ce qui garantit une précision analytique de 0,06‰.
Les masses de matière sèche nécessaires ainsi déterminées (1 mg pour les poissons et les invertébrés et
10 mg pour les filtres de seston et le sédiment) ont été pesées au microgramme près et placées dans des
capsules en étain.
L'analyse des compositions isotopiques par le système utilisé comporte plusieurs étapes (Fig.
III.3). L'analyseur élémentaire réalise tout d'abord la combustion des capsules dans un four en quartz
maintenu à 1000°C. Une quantité définie d'oxygène est introduite en même temps que l'échantillon et
sert de catalyseur pour la combustion qui se produit, de ce fait, de façon quasi instantanée à une
température avoisinant les 1600°C au niveau de l'échantillon. A la sortie du four, les gaz issus de la
combustion sont entraînés par un flux d'hélium au-dessus de différents réactifs (Cr2O3, CuO, Ag) afin
d'oxyder les hydrocarbones et éliminer les halides (chlorures, sulfures) puis pénètrent dans un deuxième
four à 600°C où sont réduits les oxydes d'azote. L'eau produite précédemment est piégée, puis une
colonne chromatographique permet la séparation du CO2 et du N2 qui, maintenus à une température
constante de 48°C, sont envoyés successivement dans le spectromètre. A ce niveau, le gaz introduit est
ionisé par bombardement électronique et accéléré sous plusieurs kilovolts vers l'analyseur de masse. Les
faisceaux ioniques y sont séparés par des champs magnétiques en fonction de leur rapport masse/charge.
Les deux faisceaux d'ions correspondant aux deux isotopes sont ensuite collectés par des détecteurs
ioniques. Les impulsions électriques qu'ils engendrent sont amplifiées pour donner le signal de mesure,
traité ensuite par un système informatique.
Les ratios isotopiques en carbone et en azote fournis par le système sont exprimés, selon la
norme internationale, en δ (‰) :
δX (‰) = [(R échantillon / R référence) -1] × 1000
où X correspond soit à 15N soit à 13C et R représente le ratio isotopique 15N / 14N ou 13C / 12C.

Réseaux trophiques et suivi de la MOP terrigène
172
Figure III.3- Schéma descriptif du système utilisé pour les analyses isotopiques. Le système est constitué d'unanalyseur élémentaire automatisé Europa Scientific ANCA-SL (Automated Nitrogen Carbon Analysis unit forSolids and Liquids), relié à un spectromètre de masse isotopique à flot continu (Europa Scientific PDZ 20-20). Lesnuméros indiquent les différentes étapes du traitement des échantillons. 0 = positionnement initial des échantillons,1 = combustion (capsule → gaz), 2 = réduction et purification des gaz, 3 = élimination de l'eau, 4 = séparation duN2 et du CO2, 5 = ionisation et accélération des particules, 6 = séparation des faisceaux d'ions et détection .
Lors du calibrage du système, des échantillons de farine de blé et de muscle blanc de morue
(Gadus morhua L.) de signatures homogènes connues (δ13C = -26,64 et -16,45‰ et δ15N = 3,01 et
16,45‰) ont été utilisés comme références internes, respectivement pour les échantillons de végétaux,
de seston et de sédiment et pour ceux de tissus animaux (poissons et invertébrés). Afin d'assurer une
précision maximale des mesures réalisées et re-calibrer régulièrement le système d'analyse en flot
continu, deux échantillons de référence (farine ou morue suivant les analyses) ont été passés tout les 5
échantillons. La précision expérimentale (basée sur l'erreur standard des mesures obtenues pour ces
échantillons de référence) obtenue au cours de cette étude est de 0,14‰ pour le δ13C et de 0,11‰ pour
le δ15N. Elle englobe les variances liées aux précisions respectives de la balance utilisée pour les pesées
et de l'analyseur isotopique, ainsi que, pour le δ13C, les fluctuations de signatures liées à l'acidification
des échantillons.
III.2.4 - EXPLOITATION DES DONNEES
Sources de matière organique
Les productions macro et micro-phytobenthiques étant très réduites sur la zone (Bodoy et
Plante-Cuny, 1980), le phytoplancton marin et la matière organique particulaire (MOP) en suspension
dans l'eau du fleuve constituent les deux principales sources potentielles de matière organique au large
du Rhône. Divers débris végétaux d'origine terrestre (feuilles et branches apportées par le Rhône) ou
marine (fragments de macrophytes benthiques transportées par les courants) sont également susceptibles
de contribuer à la constitution des pools de matière organique de la zone. Cette participation étant
dépendante soit du débit du fleuve, soit de la direction et de l'intensité des courants marins, elle reste

Réseaux trophiques et suivi de la MOP terrigène
173
sporadique et potentiellement minime comparée à celles de la MOP rhodanienne et du phytoplancton.
Néanmoins, afin de déterminer au mieux l'origine de la matière organique au large du fleuve, les
signatures isotopiques des fragments de végétaux terrestres et de macrophytes marines (Chlorophycées)
récoltés ponctuellement sur les fonds échantillonnés (surtout à 0-20 m) ont été déterminées et
confrontées à celles du phytoplancton marin, de la MOP rhodanienne et de la MOP de l'eau de surface
et du sédiment superficiel de la zone d'étude. Les différences de signature isotopique (δ13C, δ15N)
observées entre ces différentes catégories (MOP rhodanienne, végétaux terrestres, phytoplancton marin,
fragments de macrophytes benthiques, MOP de l'eau de surface et sédiment superficiel) ont été testées
par une MANOVA à un facteur (catégorie). La réalisation, pour chaque élément (C, N), d'une ANOVA
à un facteur (catégorie) complétée par un test à posteriori de Newman-Keuls a permis de préciser le
schéma obtenu.
Une tentative d'estimation des contributions respectives des deux principales sources d'origine
terrestre et marine (MOP rhodanienne et phytoplancton marin) dans la composition de la matière
organique présente dans l'eau de surface et dans le sédiment superficiel de la zone échantillonnée a été
réalisée à l'aide de modèles de mélange (Dunton et Shell, 1987 ; Harrigan et al., 1989). Les signatures
en carbone de ces deux sources de matière organique étant très différentes, un système d'équations de
mélange impliquant leurs δ13C et celui obtenu soit pour la MOP de l'eau de surface, soit pour le
sédiment superficiel de la zone, ont été utilisées :
(1) δ13C(X) = ∑ F i × δ13C(i)
(2) ∑ F i = 1
où X désigne soit le sédiment superficiel soit l'eau de surface récoltés pour chaque tranchebathymétrique et Fi la contribution de la source i à la composition de X.
En ce qui concerne la MOP de l'eau de surface, filtrée à 250 µm comme la MOP rhodanienne et
le phytoplancton marin et ne contenant donc que des particules ou microorganismes de même taille que
ceux de ces deux sources de matière organique, un seul système d'équations (basé sur les δ13C de la
MOP rhodanienne et du phytoplancton) a été résolu. Pour la matière organique présente dans le
sédiment superficiel, le problème est plus complexe. En effet, celle-ci correspond à un mélange
indifférencié de matériel détritique (pelotes fécales, restes d'organismes planctoniques sédimentés,
matériel d'origine rhodanienne, pseudo-fèces et fèces du benthos…) et d'organismes benthiques (micro
et méiofaune non extraites car difficilement repérables). Les proportions de ces différents types de
matériel sont difficilement estimables mais correspondent essentiellement soit aux deux sources de
matière organique étudiées ("producteurs primaires"), soit à des organismes s'en nourrissant directement
("consommateurs primaires": zooplancton, bactéries hétérotrophes, méiofaune). Pour cette raison,
quatre systèmes d'équations ont été résolus pour le sédiment. Le premier fait intervenir les δ13C du
phytoplancton et de la MOP rhodanienne; le second a été établi à partir de ces deux signatures

Réseaux trophiques et suivi de la MOP terrigène
174
incrémentées du +1‰ correspondant à l'enrichissement type en carbone lié au passage à un échelon
trophique supérieur (De Niro et Epstein, 1978); les deux derniers correspondent aux deux autres
combinaisons possibles de ces quatre signatures. Ce procédé a permis d'obtenir une estimation
approximative des contributions des sources de matière organique terrestre et marine à la composition
du sédiment superficiel au large du Rhône.
Afin de préciser l'importance des crues du Rhône dans la constitution des pools de matière
organique présents dans l'eau de surface et dans le sédiment superficiel à chaque profondeur, les
signatures isotopiques de la MOP rhodanienne et du phytoplancton marin ont été déterminées en
période de crue (printemps-hiver) et en période d'étiage (fin de l'été). Les différences de δ13C entre ces
deux sources de matière organique, le sédiment superficiel et l'eau de surface à chaque période ont été
testées statistiquement par des ANOVA à deux facteurs croisés fixes (période × catégorie) tout d'abord
de façon globale, puis en séparant les δ13C obtenus pour les deux compartiments à chaque profondeur
étudiée. Dans chaque cas, des test de Newman-Keuls ont été réalisés a posteriori afin de déterminer les
catégories responsables des différences observées à chaque période. De même que pour l'ensemble de la
zone d'étude, les contributions approximatives des sources terrestres et marines à la composition de la
MOP de l'eau de surface et du sédiment superficiel ont été estimées à chaque profondeur étudiée afin de
déceler d'éventuelles différences bathymétriques de la quantité de matière organique terrigène
disponible à la base des réseaux trophiques benthiques.
MOP terrigène et réseaux trophiques benthiques
Les signatures globales des principaux échelons des réseaux trophiques benthiques (sources
primaires de matière organique, proies benthiques et poissons) ont été déterminées et les différences de
δ13C et de δ15N entre ces trois catégories ont été testées par une MANOVA à deux facteurs croisés fixes
(catégorie × profondeur). Des tests à posteriori de Newman-Keuls ont permis de déterminer les
catégories et les profondeurs responsables des différences observées pour chaque élément (C ou N). Les
enrichissements moyens en δ13C et en δ15N d'une catégorie à l'autre ont été calculés pour chaque tranche
de profondeur afin de déceler d'éventuelles différences d'incorporation et de transfert de la MOP
terrestre au sein des réseaux trophiques benthiques suivant la profondeur. Le graphe δ13C vs δ15N obtenu
pour l'ensemble de la zone d'étude a permis de décrire l'intensité de l'exploitation des sources de matière
organique d'origine marine et d'origine terrestre par différents invertébrés benthiques et de déterminer
les enrichissements trophiques moyens en δ13C et en δ15N au sein des communautés benthiques au large
du Rhône. Les graphes δ13C vs δ15N ont ensuite été réalisés pour chaque profondeur afin d'étudier les
différences bathymétriques de l'incorporation de la MOP terrestre dans les réseaux trophiques des cinq
poissons.

Réseaux trophiques et suivi de la MOP terrigène
175
Transfert de la MOP jusqu'aux poissons
Afin de déceler l'incorporation éventuelle de matériel organique dérivé des sources de MOP
terrigène dans la chair des poissons, la signature isotopique moyenne globale et les δ13C et δ15N moyens
spécifiques des cinq espèces au large du Rhône ont été déterminés. Pour cela, un nombre égal (10
individus) de juvéniles G0 et d'adultes a été utilisé pour chaque poisson afin de supprimer le biais
éventuel lié à la taille différente des effectifs analysés pour chaque stade de vie et chaque espèce. La
normalité et l'homoscédaticité des données étant vérifiées, tant pour le δ13C que pour le δ15N des
poissons, l'existence potentielle de différences de signature entre les cinq espèces a été testée par une
MANOVA à un facteur (espèce). Des tests de Newman-Keuls ont été réalisés a posteriori afin de
déterminer les espèces responsables des différences de signature en carbone et en azote observées.
Pour chaque espèce, l'influence de la croissance et de la profondeur sur la signature isotopique
des individus a également été étudiée en détail. Pour cela, l'existence ou non d'une corrélation linéaire
entre la taille et le δ15N ou le δ13C des individus a été déterminée. Les différences de δ13C et de δ15N
observées entre juvéniles G0 et adultes de chaque espèce ont également été testées par des tests de
Student et les distributions statistiques du δ13C et du δ15N obtenues pour chaque stade de vie benthique
ont été illustrées par des boites de Tukey (c. f. Partie II, Chapitre II). Pour les stades de vie capturés sur
plusieurs tranches bathymétriques (adultes chez S. solea, B. luteum et A. laterna ; juvéniles et adultes
chez C. linguatula), l'effet de la profondeur sur le δ13C et le δ15N des poissons a été testé par des
ANOVA à un facteur, pour S. solea, B. luteum et A. laterna, ou à deux facteurs fixes croisés (stade ×
profondeur), pour C. linguatula. Des tests de Newman-Keuls ont été réalisés a posteriori afin de
déterminer les profondeurs responsables des différences de signature en carbone et en azote observées et
les distributions obtenues pour chaque tranche bathymétrique ont été illustrées par des boites de Tukey.
Réalisé à différents niveaux de précision (5 espèces groupées, espèces séparées, juvéniles et
adultes de chaque poisson), le calcul (1) du niveau trophique moyen théorique attendu sur la base d'une
alimentation exclusivement d'origine marine (phytoplancton) et (2) de la signature isotopique en
carbone correspondante, a permis, par simple comparaison des δ13C théoriques calculés et des δ13C
réellement observés, de déceler la présence ou non de matériel d'origine terrestre dans la chair des
poissons. Le niveau trophique (NT) d'un organisme représente le nombre de transferts trophiques
d'énergie (pondérés suivant leurs importances énergétiques relatives) existant entre cet organisme et les
sources d'énergie présentes sous forme inorganique dans le milieu (Vander Zanden et al., 1997 ; Vander
Zanden et Rasmussen, 1999). Son calcul permet de positionner les organismes au sein des réseaux
trophiques, composantes majeures des écosystèmes aquatiques car englobant l'ensemble des relations
d'ordre alimentaire existant entre les producteurs primaires et les consommateurs présents dans le milieu
et certains autres éléments, comme les détritus. Par définition, les producteurs primaires (végétaux,
bactéries autotrophes) et les détritus ont des niveaux trophiques égaux à 1. Le niveau trophique des

Réseaux trophiques et suivi de la MOP terrigène
176
consommateurs primaires stricts (herbivores) est de 2, celui des consommateurs secondaires stricts de 3,
etc. Cependant, la majorité des organismes hétérotrophes ayant une alimentation de type omnivore et
s'alimentant souvent à partir de proies de différents NT, les niveaux trophiques obtenus dans la nature
correspondent en général à des valeurs décimales. Une des approches quantitatives de l'utilisation des
isotopes stables est le calcul du niveau trophique des organismes en fonction de leur δ15N et de celui,
soit des producteurs primaires (Kline et Pauly, 1998), soit des consommateurs primaires du milieu
(Vander Zanden et Rasmussen, 2001). Etant donné les différences de fractionnement isotopique
alimentaire existant généralement entre herbivores et carnivores, l'utilisation des consommateurs
primaires comme organismes de référence dans les équations reliant le δ15N et la position trophique des
animaux augmente considérablement la précision du calcul (Vander Zanden et Rasmussen, 2001). Dans
la présente étude, le niveau trophique des poissons a donc été calculé grâce à la formule suivante :
(1) NT = [δ15N poisson – (δ15N phytopK + ∆δ15NPI-CI)] / ∆δ15NC + 2
où NT correspond au niveau trophique moyen théorique attendu sur la base d'une alimentationexclusivement d'origine marine (phytopK = phytoplancton) et ∆δ15NPI-CI et ∆δ15NC désignent lesenrichissements moyens en azote obtenus respectivement entre producteurs primaires et consommateursprimaires et pour les carnivores de niveau trophique supérieur de la zone d'étude.
L'estimation du δ13C moyen théorique attendu sur la base d'une alimentation exclusivement
d'origine marine a ensuite été calculé à l'aide de la formule :
(2) δ13Cth. = δ13C phytopK +[ (NT – 1) × ∆δ13C]
où NT correspond au niveau trophique théorique obtenu sur la base d'une alimentation exclusivementd'origine marine (phytopK = phytoplancton) et ∆δ13C désigne l'enrichissement moyen en carbone d'unéchelon trophique à l'autre estimé pour la zone d'étude.
Afin d'expliquer les différences de signature observées suivant le stade et/ou la profondeur, les
signatures isotopiques des adultes et des juvéniles des cinq espèces ont été confrontées aux données
obtenues par l'examen de leurs contenus stomacaux. Afin de comparer les résultats issus des deux
méthodes, les signatures isotopiques moyennes obtenues pour les juvéniles et les adultes de chaque
espèce ont été comparées avec les signatures isotopiques théoriques correspondantes calculées, sur la
base des régimes alimentaires observés, à l'aide de la formule :
δXth. = ∆δX+∑ δXi × Cpi
où δXth. désigne la signature théorique en élément X (carbone ou azote) attendue, ∆δX l'enrichissementmoyen en élément X d'un échelon trophique à l'autre (pour les carnivores de la zone d'étude), δXi lasignature isotopique de la proie i et Cpi son importance pondérale dans l'alimentation du groupe depoisson considéré.
Ce procédé a permis d'estimer la capacité des relations proies-prédateur révélées par l'étude de
l'alimentation des poissons à expliquer ou non la signature isotopique obtenue dans chaque cas. Une fois
validées par cette étape, les données alimentaires ont servi à reconstruire les réseaux trophiques de

Réseaux trophiques et suivi de la MOP terrigène
177
chaque espèce et, grâce à l'élaboration des graphes δ13C vs δ15N à chaque profondeur, de décrire les
transferts de matière organique d'origine terrestre et d'origine marine jusqu'aux populations des cinq
poissons étudiés.
III.3 - SOURCES ET COMPOSITION DE LA MATIERE ORGANIQUE AU LARGE DU
RHONE
Situation globale au large de l'embouchure du Rhône
Au large du Rhône, les signatures isotopiques obtenues pour les quatre principales sources
potentielles de matière organique individualisées (phytoplancton marin, MOP rhodanienne, débris de
végétaux terrestres véhiculés par le fleuve et fragments de macrophytes marines amenés par les
courants) sont distinctes. Elles varient de -27,72 à -22,36 ‰ pour le carbone et de 1,16 à 4,25 pour
l'azote (Fig. III.4).
Figure III.4 – Signatures isotopiques (moyennes et erreurs-types) obtenues pour les quatreprincipales sources potentielles de matière organique ainsi que pour la MOP de l'eau de surface etle sédiment superficiel au large du Rhône.
Par rapport à ces différentes sources, la matière organique particulaire disponible à la base des
réseaux trophiques benthiques au large du Rhône présente une signature isotopique intermédiaire qui
varie peu entre la MOP de l'eau de surface et le sédiment (Tableau III.3). Les différences de signature
isotopique observées d'une source à l'autre sont significatives (MANOVA RRao (10, 142) = 23,61 ; P <
0,001) aussi bien pour le δ13C (ANOVA : F(5, 72) = 30,60 ; P < 0,001) que pour le δ15N (ANOVA : F(5, 72)
végétauxterrestres
sédiment
phytoplancton
MOP Rhône
macrophytes marines(Chlorophycées)
MOP eau
-29
-28
-27
-26
-25
-24
-23
-22
-210 1 2 3 4 5 6
δ 15 N
δ 13 C

Réseaux trophiques et suivi de la MOP terrigène
178
= 18,47 ; P < 0,001). A l'exception des macrophytes (Chlorophycées) et du phytoplancton marins d'une
part, et de la MOP de l'eau de surface et du sédiment de l'autre, la signature en carbone diffère de façon
significative (P < 0,01 ; Annexe 34) d'une catégorie à l'autre. L'utilisation du δ13C permet donc de
différencier la majorité des sources potentielles de matière organique au large du Rhône et fournit
notamment une séparation nette entre sources d'origine terrestre (δ13C < -26‰) et marine (δ13C > -
23‰). L'utilisation complémentaire de l'azote permet une individualisation totale des sources de matière
organique du fait des différences de δ15N significatives (P < 0,05; Annexe 35) existant entre le
phytoplancton marin, les macrophytes marines, les végétaux terrestres et le groupe formé par la MOP
rhodanienne, le sédiment et la MOP de l'eau de surface.
Tableau III.3 – signatures en carbone (δ13C) et en azote (δ15N) des sources potentielles de matière organiquerecensées au large du Rhône et de la matière organique présente dans l'eau de surface et le sédimentsuperficiel de la zone d'étude. MOP = matière organique particulaire ; N =Effectif analysé.
δ13C(moy. ± erreur-type)
δ15N(moy. ± erreur-type)
N
Sources d'origine terrestreMOP rhodanienne -26,11 ± 0,23 3,47 ± 0,42 9Végétaux terrestres (débris) -27,72 ± 0,23 1,16 ± 0,18 4
Sources d'origine marinePhytoplancton (Frioul) -22,36 ± 0,24 2,33 ± 0,11 9Macrophytes (débris de Chlorophycées) -22,56 ± 0,04 4,25 ± 0,27 3
Matière organique au large du RhôneMOP eau de surface -24,38 ± 0,29 3,03 ± 0,23 27Sédiment superficiel -24,70 ± 0,14 3,81 ± 0,11 26
Les signatures en carbone obtenues pour la MOP de l'eau de surface et le sédiment sont proches
de celle de la MOP rhodanienne et nettement inférieures à celles des producteurs primaires marins. Ceci
implique une importante contribution des apports rhodaniens en MOP terrigène à la constitution des
pools de matière organique au large du Rhône, tant au niveau de l'eau de surface que du sédiment. Sur la
base des signatures en carbone observées, la contribution de la MOP rhodanienne à la constitution du
pool de matière organique présent dans l'eau de surface peut être estimée à 56,5%, contre 43,5% pour le
phytoplancton marin. Au niveau du sédiment superficiel, les contributions estimées de la MOP terrigène
et du phytoplancton marin sont respectivement de 62,7- 89,3 % et de 10,7-37,3 % (Tableau III.4). Plus
de la moitié (≥ 56%) de la matière organique disponible au large du fleuve pour les consommateurs
primaires marins, qu'ils soient benthiques ou planctoniques, est donc d'origine terrestre.

Réseaux trophiques et suivi de la MOP terrigène
179
Tableau III.4 – Estimations des contributions (%) respectives de la MOP terrigène et du phytoplancton marinà la constitution du pool de matière organique du sédiment au large du Rhône. Résultats obtenus pour leséquations de mélange réalisées à partir de 4 combinaisons de δ13C associées à ces deux sources.
Signatures utilisées δ13C %
(1) Phytoplancton -22,36 37,3%MOP rhodanienne -26,11 62,7%
(2) C1 marin = phytoplancton + 1‰ -21,36 29,5%MOP rhodanienne -26,11 70,5%
(3) Phytoplancton -22,36 14,5%C1 MOP = MOP rhodanienne + 1‰ -25,11 85,5%
(4) C1 marin = phytoplancton + 1‰ -21,36 10,7%C1 MOP = MOP rhodanienne + 1‰ -25,11 89,3%
Variabilité spatio-temporelle des apports terrestre et marins
A l'embouchure du Rhône, la composition de la matière organique (δ13C) diffère non seulement
suivant le compartiment considéré (ANOVA : P < 0,001; Annexe 36) mais également suivant la période
de l'année (ANOVA : P < 0,001; Annexe 36) (Tableau III.5). Quelle que soit la période considérée, la
différence de δ13C entre le phytoplancton marin et la MOP rhodanienne est élevée (> 3,6 ‰) et
significative (P < 0,001; Annexe 36). La signature isotopique en carbone de la MOP rhodanienne diffère
pourtant de façon importante (P < 0,001) entre les crues (printemps-hiver) et les périodes d'étiage (été).
Ces modifications saisonnières concernent également le phytoplancton marin dont le δ13C diffère de
façon significative (P < 0,01) entre la saison chaude (été) et la saison froide (printemps-hiver). Les
modifications de δ13C des deux sources de matière organique se répercutent sur les signatures en
carbone des pools de matière organique au large de l'embouchure, mais les différences temporelles
observées ne sont significatives que pour la MOP de l'eau de surface (P < 0,05), le sédiment ayant un
δ13C beaucoup plus stable. En période de crue, deux groupes de signatures en carbone distinctes (P <
0,001) se forment, opposant le phytoplancton marin d'une part, et la MOP rhodanienne, la MOP de l'eau
de surface et le sédiment superficiel de la zone d'étude de l'autre. Par contre, en période d'étiage, les
δ13C de la MOP rhodanienne, du sédiment superficiel et de la MOP de l'eau de surface diffèrent (P <
0,01) et la signature en carbone de la MOP de l'eau de surface correspond à celle obtenue pour le
phytoplancton marin à cette période. Ainsi, les contributions respectives des sources terrestres et
marines de matière organique diffèrent suivant le pool de matière organique considéré. La composition
du pool présent dans l'eau de surface réagit très vite aux modifications des apports des deux sources et
est dominée tantôt par la matière organique d'origine terrestre (périodes de crues), tantôt par le
phytoplancton marin (périodes d'étiage). Au contraire, la composition de la matière organique dans le
sédiment est relativement stable d'une saison à l'autre et reste très proche, quelle que soit la période, de
celle observée en période de crues pour la MOP rhodanienne.

Réseaux trophiques et suivi de la MOP terrigène
180
Tableau III.5 – Signatures en carbone (δ13C : moyenne ± erreur-type) des deux principales sources,terrestre et marine, de matière organique au large du Rhône et de la matière organique présente dansl'eau de surface et le sédiment superficiel de la zone d'étude, en période de crues (décembre, mars) et enpériode d'étiage (septembre). MOP = matière organique particulaire, N = effectif analysé.
Période de crues(printemps-hiver)
Période d'étiage(été)
Principales sources de matière organique
MOP rhodanienne -25,52 ± 0,24(N = 6)
-26,62 ± 0,12(N = 3)
Phytoplancton (Frioul) -22,03 ± 0,22(N = 6)
-23,01 ± 0,37(N = 3)
Matière organique au large du Rhône
MOP eau de surface -24,59 ± 0,22(N = 18)
-23,42 ± 0,37(N = 9)
Sédiment superficiel -24,45 ± 0,11(N = 18)
-25,13 ± 0,17(N = 9)
A ces modifications de la composition de la matière organique suivant le compartiment et la
période de l'année considérés, s'ajoutent des différences liées à la localisation des zones étudiées par
rapport à l'embouchure du fleuve. En effet, les signatures en carbone des pools de matière organique de
l'eau de surface et du sédiment superficiel varient suivant la tranche bathymétrique considérée (ANOVA
: P < 0,001; Annexe 37) (Tableau III.6). La signature en carbone de la MOP en suspension dans l'eau de
surface diffère de façon significative (P < 0,001) d'une tranche bathymétrique à l'autre. Elle est
minimale au-dessus des fonds de 30-50 m et maximale au-dessus de ceux de 70-100 m. Pour ce qui est
du sédiment, le δ13C varie également selon la profondeur avec une signature significativement moins
négative à 70-100 m qu'à 0-20 m et 30-50 m (P < 0,001). Les signatures des deux compartiments (eau
de surface, sédiment) sont similaires à 30-50 m mais diffèrent de façon significative (P < 0,001) à 0-20
m et à 70-100 m de profondeur.
Tableau III.6 – signatures en carbone (δ13C moyen ± erreur-type, en ‰) de la matière organiqueprésente dans l'eau de surface et le sédiment superficiel de la zone d'étude à chaque tranchebathymétrique étudiée (0-20 m, 30-50 m, 70-100 m). MOP = matière organique particulaire.
MOP eau de surface (effectif analysé)
Sédiment superficiel (effectif analysé)
0-20 m -24,52 ± 0,41(N = 9)
-24,91 ± 0,21(N = 8)
30-50 m-25,38 ± 0,16
(N = 9)-24,94 ± 0,14
(N = 9)
70-100 m -23,31 ± 0,25(N = 9)
-24,24 ± 0,14(N = 9)

Réseaux trophiques et suivi de la MOP terrigène
181
Quelle que soit la tranche bathymétrique considérée, les signatures moyennes obtenues pour
l'eau de surface et le sédiment sont significativement différentes de celles obtenues pour la MOP
rhodanienne (-26,11 ± 0,23‰) et le phytoplancton marin (-22,36 ± 0,24‰). Les estimations des
contributions respectives des deux sources de matière organique (terrestre et marine) dans la
composition de la matière organique présente dans l'eau de surface et le sédiment de chaque tranche
bathymétrique sont résumées dans le Tableau III.7. A 0-20 m et surtout à 30-50 m, l'essentiel de la
matière organique disponible à la base des réseaux trophique est d'origine terrestre, avec de 68 à 95% du
pool de matière organique sédimentaire issu des apports rhodaniens en MOP terrigène.
Tableau III.7 – Estimations des contributions (%) respectives de la MOP terrigène et duphytoplancton marin à la constitution des pools de matière organique de l'eau de surface et dusédiment à 0-20 m, 30-50 m et 70-100 m de profondeur au large du Rhône.
% MOP rhodanienne % Phytoplancton
MOP de l'eau de surface0-20 m 57,6 % 42,4 %30-50 m 80,5 % 19,5 %70-100 m 25,3 % 74,7 %
Sédiment superficiel0-20 m 68,0 – 94,7 % 5,3 - 32,0 %30-50 m 68,8 – 95,5 % 4,5 - 31,2 %70-100 m 50,1 – 76,8 % 23,2 - 49,9 %
La part de matière organique d'origine rhodanienne dans la MOP de l'eau de surface est
également élevée pour ces deux tranches bathymétriques mais, au-dessus des fonds de 0-20 m,
l'influence marine est non négligeable (42%) alors qu'au-dessus de ceux de 30-50 m la contribution de la
MOP rhodanienne atteint plus de 80%. A 70-100 m, le schéma est différent. Le phytoplancton marin
domine clairement (75%) dans la MOP de l'eau de surface et la contribution de la MOP terrestre à la
constitution du sédiment ne dépasserait pas les 77% (contre 68% au minimum pour les deux autres
tranches bathymétriques). Ainsi, l'influence rhodanienne est maximale au niveau des fonds de 30-50 m,
importante à 0-20 m, et plus réduite à 70-100 m où seul le pool de matière organique présent au niveau
du sédiment contiendrait une fraction importante de matériel d'origine terrestre. Le sédiment, qui
présente le δ13C le plus négatif et le moins variable, semble donc constituer le principal réservoir de
matière organique d'origine terrestre au large du Rhône.

Réseaux trophiques et suivi de la MOP terrigène
182
Cette observation est confirmée par l'étude des variations temporelles de signature isotopique
des deux sources de matière organique (terrestre et marine) ainsi que de la MOP de l'eau de surface et
du sédiment récoltés à chaque profondeur (Fig. III.5). Les variations spatio-temporelles de signature
isotopique observées sont plus importantes pour la MOP de l'eau de surface que pour le sédiment. Le
δ13C de la première fluctue entre celui du phytoplancton et celui de la MOP rhodanienne alors que celui
du second, plus stable, est toujours distinct du δ13C du phytoplancton mais approche la signature du
Rhône à certaines saisons et profondeurs.
Figure III.5 – Moyennes et erreurs-types des signatures isotopiques (δ13C versus δ15N) obtenues enpériode de crues (C) et en périodes d'étiage (E) pour les 2 principales sources de matière organiqued'origine terrestre (MOP rhodanienne) et marine (phytoplancton) et pour l'eau de surface et le sédimentsuperficiel de la zone d'étude à chaque profondeur échantillonnée (0-20 m = 10 m, 30-50 m = 30 m, 70-100 m = 90 m). MOP = matière organique particulaire.
En période de crue, les signatures en carbone de la MOP de l'eau de surface et du sédiment
diffèrent de façon significative de celle du phytoplancton (P < 0,001) mais sont similaires à celle de la
MOP rhodanienne, excepté celle de la MOP de l'eau de surface à 70-100 m (P < 0,01) (Fig. III.6).
L'influence rhodanienne s'étend sur l'ensemble de la zone d'étude avec un gradient progressif de
signature de moins en moins terrestre depuis l'eau de surface et le sédiment à 0-20 m et 30-50 m, de
δ13C très négatifs, jusqu'à l'eau de surface à 70-100 m.
C
30 m E 10 m E
E
C30 m C
10 m C30 m C
E
90 m C
10 m E
90 m E
30 m E 10 m C
90 m E
90 m C
-27
-26
-25
-24
-23
-22
-211,5 2,0 2,5 3,0 3,5 4,0 4,5 5,0 5,5 6,0
δ 15 N
δ 13 C
Phytoplancton
MOP eau surface
Sédiment
MOP rhodanienne

Réseaux trophiques et suivi de la MOP terrigène
183
Figure III.6 - Signatures en carbone (δ13C moyen ± erreur-type, en ‰) observées en période de crue pour les 2principales sources de matière organique d'origine terrestre (MOP Rhône) et marine (phytoplancton) et pour l'eaude surface et le sédiment superficiel de la zone d'étude à chaque tranche bathymétrique échantillonnée (0-20 m,30-50 m, 70-100 m). MOP = matière organique particulaire; Sed. = sédiment. Les barres horizontales indiquent lesmoyennes non-statistiquement différentes (P > 0,05).
En période d'étiage, les signatures de la MOP de l'eau de surface et du sédiment diffèrent de
façon significative de celle de la MOP rhodanienne (P < 0,01) et, mis à part pour la MOP de l'eau de
surface à 0-20 m et 70-100 m, de celle du phytoplancton marin (P < 0,01) (Fig. III.7). L'influence
rhodanienne est donc réduite à cette période et n'est sensible qu'au niveau du sédiment de l'ensemble de
la zone d'étude et de l'eau de surface à 30-50 m.
Figure III.7 - Signatures en carbone (δ13C moyen ± erreur-type, en ‰) observées en période d'étiage pour les 2principales sources de matière organique d'origine terrestre (MOP Rhône) et marine (phytoplancton) et pour l'eaude surface et le sédiment superficiel de la zone d'étude à tranche bathymétrique échantillonnée (0-20 m, 30-50 m,70-100 m). MOP = matière organique particulaire; Sed. = sédiment. Les barres horizontales indiquent les moyennes non-statistiquement différentes (P > 0,05).
En ce qui concerne l'azote, le δ15N de la MOP de l'eau de surface est moins variable que celui du
sédiment et reste proche, aux deux saisons, de celui du phytoplancton (Tableau III.8). Il est maximal
pendant l'étiage, ce qui reflète en partie l'augmentation de δ15N de la MOP rhodanienne à cette saison.
Cependant, en période de crue, les signatures en azote observées pour la MOP de l'eau de surface sont
supérieures à celles des deux sources potentielles, indiquant la présence de matériel associé à des
organismes hétérotrophes dans les prélèvements. Ce matériel, de taille réduite (< 250 µm), est
probablement également présent dans les échantillons récoltés en période d'étiage et contribue à
l'enrichissement en δ15N observé pour la MOP de l'eau de surface à cette période (notamment à 70-100
m où l'enrichissement en δ15N par rapport au phytoplancton marin est de 0,5‰).
Eau 30-50 m MOP Rhône Eau 0-20 m Sed. 30-50 m Sed. 0-20 m Sed. 70-100 m Eau 70-100 m Phytoplancton (-25,64 ± 0,10) (-25,52 ± 0,24) (-25,34 ± 0,06) (-24,72 ± 0,13) (-24,54 ± 0,16) (-24,12 ± 0,19) (-23,71 ± 0,23) (-22,03 ± 0,22)
MOP Rhône Sed. 0-20 m Sed. 30-50 m Eau 30-50 m Sed. 70-100 m Eau 70-100 m Eau 0-20 m Phytoplancton (-26,62 ± 0,12) (-25,52 ± 0,10) (-25,39 ± 0,09) (-24,87 ± 0,07) (-24,47 ± 0,09) (-22,49 ± 0,11) (-22,89 ± 0,06) (-23,01 ± 0,37)

Réseaux trophiques et suivi de la MOP terrigène
184
Tableau III.8 - Signatures en azote (δ15N moyen ± erreur-type, en ‰) observées en période de crue et en périoded'étiage pour les 2 principales sources de matière organique, d'origine terrestre (MOP Rhône) et marine(phytoplancton), et pour l'eau de surface et le sédiment superficiel de la zone d'étude à chaque tranchebathymétrique échantillonnée (0-20 m, 30-50 m, 70-100 m). MOP = matière organique particulaire; Sed. = sédiment.
Période de crues Période d'étiage
MOP rhodanienne 2,66 ± 0,19 5,07 ± 0,16
Phytoplancton (Frioul) 2,32 ± 0,11 2,37 ± 0,04
MOP eau de surface0-20 m 3,06 ± 0,13 3,03 ± 0,2130-50 m 2,95 ± 0,20 3,29 ± 0,0370-100 m 3,13 ± 0,21 3,48 ± 0,07
Sédiment superficiel0-20 m 3,61 ± 0,13 4,35 ± 0,0230-50 m 4,04 ± 0,23 4,15 ± 0,0170-100 m 3,76 ± 0,18 3,08 ± 0,02
Le δ15N du sédiment est plus élevé que celui de la MOP de l'eau de surface et plus variable,
avec des δ15N maximaux à 0-20 m et 30-50 m de profondeur en période d'étiage. La signature en azote
observée oscille entre celle de la MOP de l'eau de surface et celle de la MOP rhodanienne en période
d'étiage. Ce δ15N élevé peut s'expliquer en partie par la présence non négligeable d'organismes
hétérotrophes associés à la matière organique d'origine terrestre et/ou marine dans le sédiment,
notamment en périodes de crues, mais résulte vraisemblablement surtout de la contribution élevée de la
MOP rhodanienne à la constitution du pool de matière organique du sédiment, qui intègre la signature
de cette source aux deux périodes.
III.4 - MOP TERRIGENE ET RESEAUX TROPHIQUES BENTHIQUES
Les contributions différentes des sources marine et terrestre à la constitution des pools de
matière organique de l'eau de surface et du sédiment superficiel selon la profondeur sont susceptibles
d'entraîner des différences d'incorporation de la MOP rhodanienne au sein des réseaux trophiques
présents à 0-20 m, 30-50 m et 70-100 m. La MANOVA réalisée sur les δ13C et les δ15N obtenus pour
l'ensemble des composantes des réseaux trophiques benthiques au large de l'embouchure indique
l'existence de différences significatives de signature isotopique non seulement entre les grandes
catégories analysées (producteurs primaires, invertébrés benthiques, poissons) (R Rao (4, 494) = 144,74; P <
0,001), mais également suivant la profondeur (R Rao (4, 494) = 9,11; P < 0,001). La signature isotopique en
carbone varie de façon significative (ANOVA: F(2, 248) = 230,29; P < 0,001; Annexe 38) suivant la
catégorie considérée (Tableau III.9). Le δ13C augmente significativement entre les producteurs
primaires, les invertébrés benthiques et les poissons, tant pour l'ensemble de la zone d'étude (P < 0,001)
qu'à chacune des trois profondeurs étudiées (P < 0,001).

Réseaux trophiques et suivi de la MOP terrigène
185
Tableau III.9 – Signatures en carbone (δ13C, ‰) des principales composantes des réseaux trophiquesbenthiques obtenues à chaque profondeur (0-20 m, 30-50 m et 70-100 m) au large du Rhône. N =effectif analysé.
ProfondeurCatégorie 0-20 m 30-50 m 70-100 m
Global(0-100 m)
Producteurs primaires(eau de surface, sédiment, macrophytes
marines, végétaux terrestres)
-25,19 ± 0,43 (N = 14)
-25,46 ± 0,22 (N = 14)
-23,68 ± 0,21 (N = 14)
-24,78 ± 0,21 (N = 42)
Invertébrés benthiques(polychètes, mollusques, crustacés,
échinodermes…)
-20,37 ± 0,37 (N = 36)
-22,31 ± 0,33 (N = 20)
-20,29 ± 0,83 (N = 16)
-20,89 ± 0,23 (N = 72)
Poissons(juvéniles et adultes des 5 espèces)
-18,11 ± 0,16 (N = 72)
-19,49 ± 0,28 (N = 35)
-18,20 ± 0,26 (N = 36)
-18,47 ± 0,13 (N = 143)
Global(toutes catégories confondues)
-19,59 ± 0,25 (N = 122)
-21,52 ± 0,33 (N = 69)
-19,87 ± 0,31 (N = 66)
L'élévation du δ13C moyen d'une catégorie à l'autre varie, suivant la profondeur, de +3,2 ‰ (30-
50 m) à +4,8‰ (0-20 m) entre les producteurs primaires et les invertébrés benthiques et de +2,1‰ (70-
100 m) à +2,8‰ (30-50 m) entre ces derniers et les poissons, autour de valeurs moyennes respectives
de +3,9‰ et +2,4‰. La signature en carbone varie également suivant la profondeur (ANOVA: F (2, 248)
= 16,60; P < 0,001; Annexe 38), avec un δ13C global significativement plus négatif à 30-50 m qu'aux
deux autres tranches bathymétriques (P < 0,001; Annexe 38). Ce phénomène résulte principalement des
modifications de δ13C des organismes benthiques hétérotrophes. En effet, les signatures en carbone des
invertébrés et des poissons sont significativement plus négatives à 30-50 m qu'aux deux autres
profondeurs (P < 0,001 et P < 0,01 respectivement; Annexe 38) alors que le δ13C des producteurs
primaires est similaire à 0-20 m et 30-50 m et ne se modifie de façon significative (P < 0,05) qu'à 70-
100 m. La signature en azote ne diffère pas suivant la tranche bathymétrique considérée (ANOVA: H (2,
248) = 0,71; P > 0,48; Annexe 39) mais varie de façon significative (ANOVA: H (2, 248) = 314,53; P <
0,001; Annexe 39) d'une catégorie à l'autre (Tableau III.10). Le δ15N augmente de façon significative
entre les producteurs primaires, les invertébrés benthiques et les poissons (P < 0,001; annexe 39) avec
un enrichissement moyen de +3,4‰ entre les deux premiers groupes et de +3,6‰ entre les deux
derniers (Tableau III.10). Ce phénomène se vérifie pour chaque tranche bathymétrique étudiée, avec un
enrichissement en δ15N toujours significatif mais variant selon la profondeur, de +3,0‰ (70-100 m) à
+3,6‰ (0-20 m et 30-50 m) entre producteurs primaires et invertébrés, et de + 3,4‰ (0-20 m) à + 4,0‰
(70-100 m) entre invertébrés et poissons.

Réseaux trophiques et suivi de la MOP terrigène
186
Tableau III.10 – Signatures en azote (δ15N, ‰) des principales composantes des réseaux trophiquesbenthiques obtenues à chaque profondeur (0-20 m, 30-50 m et 70-100 m) au large du Rhône. N =effectif analysé.
ProfondeurCatégorie 0-20 m 30-50 m 70-100 m
Global (0-100 m)
Producteurs primaires(eau de surface, sédiment, macrophytes
marines, végétaux terrestres)
3,25 ± 0,37(N = 14)
3,59 ± 0,49(N = 14)
3,62 ± 0,54(N = 14)
3,49 ± 0,27 (N = 42)
Invertébrés benthiques(polychètes, mollusques, crustacés,
échinodermes…)
6,83 ± 0,33(N = 36)
7,15 ± 0,27(N = 20)
6,61 ± 0,35(N = 16)
6,87 ± 0,20 (N = 72)
Poissons(juvéniles et adultes des cinq espèces)
10,21 ± 0,14(N = 72)
10,62 ± 0,17(N = 35)
10,64 ± 0,19(N = 36)
10,42 ± 0,10 (N = 143)
Global(toutes catégories confondues)
6,74 ± 0,55(N = 158)
7,02 ± 0,29(N = 84)
6,98 ± 0,34(N = 86)
Ainsi, l'incorporation de la matière organique d'origine terrestre et son transfert le long des
chaînes trophiques aboutissant aux poissons au large du Rhône diffère selon la tranche bathymétrique
considérée. Les différences de δ13C observées entre les consommateurs primaires et les sources de
matière organique disponibles à chaque profondeur suggèrent des modifications bathymétriques du
degré d'exploitation de la MOP terrigène par les organismes benthiques. De même, les fluctuations
bathymétriques de l'enrichissement en δ13C et en δ15N d'une catégorie à l'autre traduisent l'existence,
selon la profondeur, de différentes voies de transfert de la MOP rhodanienne. Ces deux phénomènes
(incorporation et transfert de la MOP terrestre au sein des réseaux) seront détaillés au sein des deux sous
chapitres suivants afin de préciser le schéma observé pour les communautés benthiques présentes au
large du Rhône.
III.4.1 - INCORPORATION DE LA MOP TERRIGENE DANS LES RESEAUX TROPHIQUES ?
Les signatures isotopiques obtenues à 0-20 m, 30-50 m et 70-100 m au large du Rhône pour
l'ensemble des composantes des réseaux trophiques des cinq poissons plats étudiés sont réunies dans
l'Annexe 40. Pour la zone d'étude prise dans son ensemble, deux grands types de réseaux trophiques
benthiques s'individualisent, respectivement basés, l'un sur l'exploitation principale des sources de
matière organique d'origine marine (essentiellement le phytoplancton), et l'autre majoritairement sur
celle des sources terrestres (MOP rhodanienne, sédiment, seston de la zone d'étude) (Fig. III.8). La
MOP d'origine rhodanienne participe donc en partie à la production des écosystèmes benthiques côtiers
et intègre les réseaux trophiques des poissons étudiés. Cependant, parmi les proies benthiques analysées,
la majorité dépend essentiellement de la production primaire in situ. Seuls les polychètes dépositivores
et carnivores ont des signatures isotopiques compatibles avec une assimilation importante de matériel
organique d'origine terrestre. Compte tenu des signatures isotopiques observées au sein de chacun des

Réseaux trophiques et suivi de la MOP terrigène
187
deux réseaux et des connaissances disponibles sur la biologie des organismes impliqués,
l'enrichissement moyen en azote peut être estimé à 2,6±0,05‰ entre producteurs primaires et
consommateurs primaires et à 3,2±0,1‰ entre organismes carnivores de niveau trophique supérieur.
L'enrichissement moyen en carbone d'un niveau trophique à l'autre est de 1,8±0,2‰.
Figure III.8 - Exploitation des sources de matière organique d'origine marine et terrestre par les différentescomposantes des réseaux trophiques benthiques aboutissant aux cinq espèces de poissons plats étudiées au largedu Rhône. Les erreurs-types associées aux signatures moyennes (δ13C et δ15N) de chaque catégorie sontindiquées en gris. MOP = matière organique particulaire, brach. = brachyoure, pol = polychète, S = suspensivore, DS =dépositivore de surface, DsS = dépositivore de sub-surface, C = carnivore, pK = plancton.
Fonds Sableux entre 0 et 20 m de profondeur
Sur les petits fonds sableux côtiers situés entre 0 et 20 m de profondeur à proximité de
l'embouchure du Rhône, la MOP d'origine terrestre, surtout présente au niveau du sédiment, participe à
la production des écosystèmes benthiques et intègre les réseaux trophiques aboutissant aux juvéniles et
aux adultes des cinq poissons étudiés. En effet, à cette profondeur, les deux grands types de réseaux
trophiques benthiques individualisés précédemment (terrestre/marin) se retrouvent de façon très nette
(Fig. III.9). La majorité les proies benthiques analysées dépend essentiellement de la production
primaire in situ, et seuls les polychètes dépositivores ont des signatures isotopiques compatibles avec
une assimilation préférentielle de matériel organique d'origine terrestre. Ces derniers semblent capables
d'exploiter directement la matière organique rhodanienne sédimentée, et ce de façon sélective, leurs
signatures en carbone étant égales ou inférieures à celle du sédiment qu'ils ingèrent (Annexe 40). Les
Réseau trophique reposantessentiellement sur :
le phytoplancton marin
la MOP terrigène
végétauxterrestres
MOP Rhône
sédimentMOP eau
phytopKpol C
pol S
pol DsSpol DS
gobiidae
cumacé copépode
mysidacé
brach.
ophiure
bivalvejuvénile
zoopK
AlpheusCrangon
amphipode
holothurie
anomouregastéropode
macrophytes marines(Chlorophycées)
siphon bivave
-28
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-170 1 2 3 4 5 6 7 8 9 10 11
δ15
Ν
δ13 C

Réseaux trophiques et suivi de la MOP terrigène
188
polychètes carnivores et, à moindre échelle, les juvéniles de bivalves, les amphipodes et les brachyoures
semblent dépendre partiellement, à 0-20 m, des sources de matière organique terrestre. Les autres proies
(dont les polychètes suspensivores) ont des signatures clairement marines. Parmi elles, les macroures du
genre Crangon, les anomoures et les bivalves adultes présentent des signatures isotopiques particulières,
différentes de celles attendues sur la base d'une ingestion de matériel exclusivement dérivé du
phytoplancton marin. Leurs δ13C suggèrent l'exploitation d'une source de carbone enrichie en 13C (et
peut être également en 15N) par rapport aux sources de matière organique analysées.
Figure III.9 - Exploitation des sources de matière organique d'origine marine et terrestre par les différentescomposantes des réseaux trophiques benthiques aboutissant aux cinq espèces de poissons plats à 0-20 m deprofondeur au large du Rhône. Les erreurs-types associées aux signatures moyennes (δ13C et δ15N) de chaquecatégorie sont indiquées en gris. MOP = matière organique particulaire, pol = polychète, S = suspensivore, DS =dépositivore de surface, DsS = dépositivore de sub-surface, C = carnivore, pK = plancton.
Fonds vaseux entre 30 et 50 m de profondeur
A 30-50 m de profondeur, où la matière organique d'origine terrestre domine à la fois au niveau
du sédiment et dans l'eau de surface, les proies benthiques se répartissent moins clairement au sein des
deux types de réseaux trophiques benthiques (terrestre/marin) individualisés pour la zone d'étude dans
son ensemble (Fig. III.10). La majorité des proies benthiques analysées dépend toujours de la
production primaire in situ. Cependant, un bon nombre d'invertébrés benthiques présentent un δ13C très
négatif, signe d'une exploitation partielle des apports terrigènes pour leur croissance (Annexe 40). A
cette profondeur, la matière organique particulaire d'origine rhodanienne participe donc fortement à la
macrophytes marines
mysidacé
anomoure
ophiure
zoopK
Crangon
siphonbivalve
cumacé amphipode
gastéropode
brachyoure
gobiidae
copépode
bivalve juvénile
pol S
pol DsS
pol C
pol DS
phytopK
MOP eau
sédiment
MOP Rhône
végétauxterrestres-29
-28
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-17
-161 2 3 4 5 6 7 8 9 10 11
δ15Ν
δ13C
Réseau trophique reposantessentiellement sur :
le phytoplancton marin
la MOP terrigène

Réseaux trophiques et suivi de la MOP terrigène
189
production des écosystèmes benthiques. L'ensemble des polychètes présents (y compris les
suspensivores) montre des signatures isotopiques correspondant à une assimilation préférentielle de
matériel organique d'origine terrestre. L'utilisation de la MOP terrigène se fait de façon directe (via le
sédiment et la MOP en suspension dans l'eau ingérés) pour les polychètes dépositivores et
suspensivores. Elle est indirecte pour les polychètes carnivores qui semblent se nourrir essentiellement à
partir de ces derniers. A cette profondeur, les bivalves, les brachyoures et les macroures du genre
Crangon semblent également dépendre de façon non négligeable des sources de matière organique
terrestre. Les autres proies (mysidacés, amphipodes, holothuries, gastéropodes…) ont des signatures
typiquement marines.
Figure III.10 - Exploitation des sources de matière organique d'origine marine et terrestre par les différentescomposantes des réseaux trophiques benthiques aboutissant aux aux cinq espèces de poissons plats à 30-50 m deprofondeur au large du Rhône. Les erreurs-types associées aux signatures moyennes (δ13C et δ15N) de chaquecatégorie sont indiquées en gris. MOP = matière organique particulaire, pol = polychète, S = suspensivore, DS =dépositivore de surface, DsS = dépositivore de sub-surface, C = carnivore, pK = plancton.
Fonds vaseux entre 70 et 100 m de profondeur
A 70-100 m, où la matière organique terrigène est essentiellement présente au niveau du
sédiment, le réseau trophique basé sur la matière organique terrigène est très réduit (Fig. III.11). A cette
profondeur, la quasi totalité des proies benthiques analysées dépend de la production primaire in situ. La
matière organique particulaire d'origine rhodanienne participe donc peu à la production des écosystèmes
Réseau trophique reposantessentiellement sur :
le phytoplancton marin
la MOP terrigène
végétauxterrestres MOP Rhône
sédimentMOP eau
phytopK
pol DsS
pol DS
pol Cbrachyoure
gastéropode
holothurie
cumacé
pol S
Alpheus mysidacé
Crangon
amphipode
gobiidae
copépode
bivalve
-28
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-17
-160 1 2 3 4 5 6 7 8 9 10 11
δ15 Ν
δ13 C
zoopK

Réseaux trophiques et suivi de la MOP terrigène
190
benthiques des fonds vaseux. Elle intègre malgré tout les réseaux trophiques des poissons étudiés, par le
biais des polychètes dépositivores de sub-surface (Annexe 40). Les polychètes carnivores et, à moindre
échelle, les polychètes dépositivores de surface et les macroures du genre Alpheus semblent également
dépendre en partie des sources de matière organique d'origine terrestre. Les autres proies, dont les
polychètes suspensivores, les brachyoures, les macroures du genre Crangon et les bivalves, ont des
signatures clairement marines.
Figure III.11 - Exploitation des sources de matière organique d'origine marine et terrestre par les différentescomposantes des réseaux trophiques benthiques aboutissant aux aux cinq espèces de poissons plats à 70-100 mde profondeur au large du Rhône. Les erreurs-types associées aux signatures moyennes (δ13C et δ15N) de chaquecatégorie sont indiquées en gris. MOP = matière organique particulaire, pol = polychète, S = suspensivore, DS =dépositivore de surface, DsS = dépositivore de sub-surface, C = carnivore, pK = plancton.
III.4.2 - TRANSFERT DE LA MOP TERRIGENE JUSQU'AUX POISSONS ?
Les signatures isotopiques individuelles obtenues pour les cinq poissons plats au large du Rhône
fluctuent entre -22,08 et -16,13 ‰ pour le carbone et entre 7,94 et 13,39 ‰ pour l'azote. La signature
moyenne en azote des cinq espèces est de 10,15±0,12‰, ce qui, sur la base d'une alimentation
strictement dépendante des sources de matière organique marines (phytoplancton) et d'un
enrichissement moyen en azote constant de +3,2‰ par échelon trophique à partir des consommateurs
primaires, correspondrait à un niveau trophique moyen de 3,66. Compte tenu du δ13C obtenu pour le
zooplancton marin de la zone (-21,36‰) et de l'enrichissement moyen en carbone par échelon trophique
bivalve
brachyoure
pol S
copépode
amphipode Crangon
holothurie
Alpheuspol C
mysidacé
gobiidae
gastéropode
cumacé
pol Ds
pol DsS
phytopK
MOP eau
sédiment
MOP Rhône
végétauxterrestres
-28
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-170 1 2 3 4 5 6 7 8 9 10 11
δ15Ν
δ13 C
zoopK
Réseau trophique reposantessentiellement sur :
le phytoplancton marin
la MOP terrigène

Réseaux trophiques et suivi de la MOP terrigène
191
(+1,8‰), le δ13C attendu dans ce cas est de -17,57‰. Or le δ13C moyen obtenu pour les cinq espèces est
de -18,49±0,36‰. L'obtention de cette signature globale, inférieure à celle calculée, suggère une
participation non négligeable de la MOP terrigène à la constitution générale des stocks des cinq espèces
au large du Rhône. Cependant, d'importantes différences interspécifiques existent (Fig. III.12). La
signature isotopique varie de façon significative suivant l'espèce considérée (MANOVA : RRao (8,188) =
7,48 ; P < 0,001) aussi bien pour le carbone (F(4,95) = 10,15 ; P < 0,001), que pour l'azote (F (4,95) = 5,16 ;
P < 0,001). La sole commune Solea solea, présente un δ13C très négatif et significativement différent (P
< 0,01; Annexe 41) de celui des autres espèces. Pour l'azote, le δ15N ne diffère de façon significative (P
< 0,01; Annexe 41) qu'entre Arnoglossus laterna (pour qui il est minimal) et les deux soles, Solea solea
et S. impar (chez qui il est maximum). Buglossidium luteum et Citharus linguatula présentent des δ15N
intermédiaires. Compte tenu de ces différences, l'exploitation des apports rhodaniens en MOP terrigène
est susceptible de varier fortement d'une espèce à l'autre.
Figure III.12 – Signatures isotopiques moyennes des cinq espèces de poissons plats étudiées aularge du Rhône : Graphe δ13C vs δ15N obtenu à partir des signatures de 10 juvéniles G0 et 10adultes de chaque espèce. Les erreurs-types associées aux signatures moyennes (δ13C et δ15N) dechaque poisson sont indiquées en gris.
S. impar
A. laterna
B. luteum
S. solea
C. linguatula
-21
-20
-19
-18
-17
-168 9 10 11 12
δ 15 N
δ13C

Réseaux trophiques et suivi de la MOP terrigène
192
III.4.2.1 - SOLEA SOLEA
Signature isotopique globale
Les δ13C obtenus pour les 80 juvéniles et adultes de S. solea analysés (LT = 30-371 mm)
fluctuent entre -22,08 et -16,57 ‰ autour d'une moyenne de -19,95±0,23‰. Cette signature isotopique
en carbone indique l'incorporation, dans la chair des individus, de matériel organique provenant d'une
source de δ13C plus négatif que le phytoplancton marin. En effet, les δ15N obtenus pour cette espèce
varient de 9,18 à 13,39 ‰ avec une valeur moyenne de 10,80±0,13‰. Sur la base d'une alimentation
d'origine exclusivement marine, le niveau trophique global moyen de l'espèce serait de 3,83 et la
signature en carbone correspondante de -17,26‰. Compte tenu des signatures en carbone des sources
potentielles de matière organique au large du Rhône, la différence observée entre le δ13C attendu et celui
observé ne peut s'expliquer que par une forte incorporation de matériel d'origine terrestre dans la chair
des individus de l'espèce. Il est cependant possible, du fait des variations ontogéniques et
bathymétriques de l'alimentation observées pour S. solea sur la zone d'étude (c. f. Partie II, Chapitre II),
que ce transfert de MOP terrigène soit plus important pour l'un des stades de vie benthique ou à une
profondeur donnée.
Taille des individus, signature isotopique et transfert de la MOP terrigène
La signature isotopique varie en fonction de la taille des individus. En ce qui concerne l’azote,
le δ15N est positivement corrélé (P < 0,001) avec la taille des individus, indiquant une augmentation du
niveau trophique au cours de la croissance (Fig. III.13).
Figure III.13 - Régression linéaire significative (r = 0,622, r2 = 0,387 ; P < 0,001) obtenue entre le δ15N(en ‰) et la taille (LT, en mm) chez Solea solea au large du Rhône. ----- = intervalle de confiance (95%).
δ15N
(‰)
LT (mm)
8,5
9,0
9,5
10,0
10,5
11,0
11,5
12,0
12,5
13,0
13,5
14,0
0 50 100 150 200 250 300 350 400
δ15N = 0,0075 LT + 9,2539
+

Réseaux trophiques et suivi de la MOP terrigène
193
La signature en azote augmente de façon significative (Test de Student : T (34, 21, 15) = 4,532 ; P <
0,001) entre les plus petits juvéniles (30-90 mm de LT) et les adultes (LT ≥ 262 mm) de l'espèce (Fig.
III.14). Le δ15N moyen passe de 10,07±0,14‰ chez les premiers à 11,06±0,15‰ chez les derniers, ce
qui correspond à une augmentation de niveau trophique de +0,31.
Figure III.14 – Représentation en boites de Tukey des δ15N (‰) observés chez les plus petits juvéniles(G0 : 30-90 mm de LT) et les adultes (LT ≥ 262 mm) de Solea solea au large du Rhône. NT = niveautrophique calculé à partir du δ15N moyen.
En ce qui concerne le carbone, aucune relation significative n’a été mise en évidence entre le
δ13C et la taille des individus étudiés (régression linéaire : r = 0,223 ; r2 = 0, 0497 ; P > 0,303). Les
modifications de δ13C entre les plus petits juvéniles (δ13C moy. = -20,03±0,33 ‰) et les adultes (δ13C moy. =
-19,56±0,38 ‰) de l'espèce ne sont pas significatives (Test de Student : T (34, 21, 15) = 1,091; P > 0,283).
Compte tenu des signatures obtenues, l'influence de la MOP rhodanienne concerne l'ensemble
des stades de vie benthique de S. solea mais est plus forte chez les adultes (Tableau III.11).
Tableau III.11 – Signatures isotopiques moyennes en azote et en carbone observées et niveau trophique et δ13Cthéoriques calculés dans le cas d'une alimentation d'origine exclusivement "marine" pour les juvéniles G0 (LT< 100 mm) et les adultes de S. solea au large du Rhône.
δ15N observéNiveau trophique
théorique δ13C théorique δ13C observé δ13C obs. - δ13C théo.
Juvéniles G0 10,07 3,25 -17,67 -20,03 - 2,36
Adultes 11,06 3,56 -17,11 -19,56 - 2,45
Max non-aberrantMin non-aberrant
75%25%
Médiane
9,0
9,5
10,0
10,5
11,0
11,5
12,0
12,5
Juvéniles G0 LT < 100 mm Adultes ( NT = 3,25 ) ( NT = 3,56 )
δ15N
(‰)

Réseaux trophiques et suivi de la MOP terrigène
194
Ceci s'explique par l'importance particulièrement élevée des polychètes dans le régime
alimentaire de S. solea. En effet, au large du Rhône, les polychètes dépositivores constituent une part
importante de l'alimentation des juvéniles et l'essentiel de celle des adultes de l'espèce (Fig. III.15).
Figure III.15 – Composition pondérale (Cp) du régime alimentaire des juvéniles G0 (a) et des adultes (b)de S. solea au large du Rhône.
L'alimentation décrite pour chacun des deux stades de vie benthique explique totalement la
signature en carbone observée. Les δ13C théoriques calculés sur la base des signatures des proies
ingérées pondérées par leur importance alimentaire, pour chacun des deux stades, sont très proches des
signatures obtenues, surtout pour les juvéniles (Tableau III.12).
Tableau III.12 – Signatures isotopiques théoriques en azote et en carbone calculées en fonction del'alimentation des individus et δ13C et δ15N moyens observés pour les juvéniles G0 et les adultes de S. solea aularge du Rhône. (pour les calculs, voir chapitre matériel et méthodes).
Juvéniles G0 Adultesδ15N δ13C δ15N δ13C
Proie théorique (calcul alimentation) 6,10 -21,82 7,05 -21,83
Signature théorique attendue 9,30 -20,02 10,25 -20,03
Signature isotopique observée 10,07±0,14 -20,03±0,33 11,06±0,15 -19,56±0,38
Par contre, pour les deux stades de vie, les signatures en azote attendues sont légèrement
inférieures à celles obtenues. Il est possible que les signatures isotopiques en azote des phoronidiens et
des siponcles, non disponibles pour la présente étude, modifient la signature en azote des juvéniles et
des adultes de l'espèce ou que la taille des proies analysées soit inférieure à celle des proies ingérées
dans le milieu. Mais l'obtention d'un écart en δ15N entre juvéniles et adultes similaire entre les valeurs
calculées et les valeurs mesurées (+1‰ environ) suggère plutôt l'existence d'un enrichissement
trophique global en azote supérieur à 3,2‰ chez S. solea. Le positionnement des juvéniles et des adultes
de S. solea par rapport à leurs proies potentielles sur la zone d'étude explique les signatures observées
(b) Adultes (LT > 260 mm)
polychètes 75,3% (surtout DsS)
anomoures 0,6%
brachyoures1,3%
macroures3,3%
bivalves5,4%
ophiures3,1%
gastéropodes2,4%
holothuries5,7%
siponcles2,2%
(a) Juvéniles G0 (LT < 100 mm)
amphipodes11,0%
mysidacés5,5%
cumacés17,7%
ophiures1,5%
bivalves26,6%
polychètes 34,5%(surtout DS)
phoronidiens3,3%

Réseaux trophiques et suivi de la MOP terrigène
195
Ss J
gastéropode
anomoure
holothurie
amphipodeCrangon
pol C
Alpheus
ophiure
brach.
pol DsS
Ss A
cumacépol S
mysidacé
phytopK
MOP eau
sédiment
MOP Rhône
pol DS
-28
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-17
-160 1 2 3 4 5 6 7 8 9 10 11 12
δ 15 Ν
δ 13 C
siphonbivalve
bivalve juvénile
pour ces deux stades de vie benthique et permet de décrire le transfert de la matière organique d'origine
terrestre vers cette espèce (Fig. III.16).
Figure III.16 - Réseaux trophiques des juvéniles G0 et des adultes de S. solea au large du Rhône. MOP =matière organique particulaire, brach. = brachyoure, pol = polychète, S = suspensivore, DS = dépositivore de surface,DsS = dépositivore de sub-surface, C = carnivore. Les flèches indiquent les principales voies de transfert de matièreorganique déterminées par l'étude des contenus stomacaux des poissons (proies dont Cp > 10%).
Les juvéniles et les adultes de S. solea se situent en bout du réseau trophique basé
essentiellement sur la MOP terrigène, ce qui confirme l'importance primordiale des polychètes
dépositivores ingérés pour la croissance et la condition des individus de l'espèce. La faible augmentation
de niveau trophique (∆δ15N = +1‰) entre juvéniles et adultes est essentiellement liée à la
consommation préférentielle de polychètes dépositivores de sub-surface (de faible δ15N) par les adultes,
qui contrebalance, chez ces derniers, l'augmentation potentielle en δ15N liée à l'ingestion de proies plus
volumineuses et de niveau trophique supérieur (macroures, siphons de bivalves adultes, gastéropodes,
etc).
Signature isotopique et profondeur de capture des individus
La profondeur n’a pas d’influence significative sur le δ15N des individus de S. solea récoltés au large
du Rhône (ANOVA : F(2, 18) = 0,082 ; P > 0,921). En revanche, une différence significative (ANOVA :
F(2, 18) = 4,428 ; P < 0,05) du δ13C avec la profondeur a été mise en évidence (Fig. III.17). A taille égale,
les soles adultes capturées entre 30 et 50 m présentent un δ13C plus négatif (P < 0,05 ; Annexe 42) que
celles récoltées dans des zones moins profondes (0-20 m) ou plus profondes (70-100 m).

Réseaux trophiques et suivi de la MOP terrigène
196
Figure III.17 - Représentation en boites de Tukey des δ13C obtenus pour les individus de Solea solea deLT > 200 mm présents au large du Rhône, en fonction de la profondeur de récolte des poissons. Lesvaleurs entre parenthèses correspondent aux δ13C moyens ± erreur-types obtenus à chaque profondeur.
Ces différences bathymétriques de signatures en carbone s'expliquent par une double
modification, selon la profondeur, de l'alimentation des adultes de l'espèce et de l'exploitation des
sources terrestres par les invertébrés benthiques (Fig. III.18). Au niveau des petits fonds sableux entre 0
et 20 m de profondeur (Fig. III.18-a), l'importance pondérale des polychètes dans l'alimentation des
adultes de l'espèce est minimale (Cp = 38%). Le régime alimentaire s'enrichit d'un nombre non
négligeable de proies de signature marine (bivalves, gastéropodes, ophiures) dont l'ingestion provoque
la hausse de δ13C observée pour les soles adultes à cette profondeur. Au niveau des vases plus
profondes, l'alimentation des adultes est globalement similaire, avec une large dominance de polychètes
(Cp > 73%). Les différences de signatures observées entre les soles capturées à 30-50 m et à 70-100 m
résultent de modifications bathymétriques du transfert des sources de matière organique marine et
terrestre au sein du réseau trophique des soles adultes. En effet, à 30-50 m, l'ensemble des polychètes
présente une signature nettement "terrestre" (Fig. III.18-b) alors qu'à 70-100 m, seuls les dépositivores
de sub-surface dépendent des apports rhodaniens en MOP terrigène (Fig. III.18-c). La variabilité
bathymétrique des signatures en carbone des polychètes dépositivores de surface et carnivores est donc
le principal facteur responsable des différences de δ13C observées pour les soles adultes entre les fonds
vaseux de 30-50 m et ceux de 70-100 m de profondeur.
δ13C
(‰)
Max non-aberrantMin non-aberrant75%25%Médiane
-22,5
-22,0
-21,5
-21,0
-20,5
-20,0
-19,5
-19,0
-18,5
-18,0
-17,5
-17,0
-16,5
-16,0
-15,5
0-20 m 30-50 m 70-100 m ( -18,57±0,51‰) ( -20,99±0,36‰) ( -19,23±0,67 ‰)

Réseaux trophiques et suivi de la MOP terrigène
197
(a) 0-20 m de profondeur
(b) 30-50 m de profondeur
(c) 70-100 m de profondeur
Figure III.18- Composition pondérale du régime alimentaire (Cp), réseau trophique et transfert de la matière chezles adultes de S. solea ( LT ≥ 260 mm) à 0-20 m (a), 30-50 m (b) et 70-100 m (c) de profondeur au large du Rhône.Les flèches indiquent les transferts alimentaires de matière organique.
polychète 73,5%(dont 50% DsS, 26% DS, 22% C et 2% S)
holothurie9,2%
siponcle5,1%
poisson (gobiidae)0,1%
bivalve5,0%
brachyoure0,2%
macroure6,0%
gastéropode0,8%
brachyoure0,7%
anomoure1,3%
amphipode0,5%
poisson (gobiidae)0,4%
polychète 37,8% (dont 41% DS, 36% DsS, 16% C et 6% S)
bivalve23,6%
ophiure20,0%
gastéropode13,9%
macroure0,8%
cumacé0,5%
polychète 88,3%(dont 81% DsS, 10% DS, 8% C et 1% S)
holothurie6,2%
poisson (gobiidae)0,2%
brachyoure1,6%
anomoure0,9%mysidacé
0,1%
amphipode0,1%
macroure1,2%
bivalve1,2%
MOP Rhône
sédiment
MOP eau
phytopK
pol DsS
gastéropode
gobiidae
pol SAlpheus
holothurie
Crangon
pol C
pol DS
brachyoure
bivalve
Ss A
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-172 3 4 5 6 7 8 9 10 11 12
δ 15 Ν
δ 13C
pol DS
bivalve
brachyoure
gobiidae
amphipode
Ss A crangon
pol DsS
Alpheus mysidacé holothurie
anomoure
pol SphytopK
MOP eau
sédiment
MOP Rhône
pol C
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-17
-162 3 4 5 6 7 8 9 10 11 12
δ 15 Ν
δ 13C
pol DsS
MOP Rhône
sédiment
MOP eau
phytopK
pol S
bivalve juvénile
Ss A
gobiidae
brachyoure
gastéropode
amphipode
cumacé
siphonbivalve
macroureophiure
anomoure
pol C
pol DS
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-17
-162 3 4 5 6 7 8 9 10 11 12
δ 15
Ν
δ 13
C

Réseaux trophiques et suivi de la MOP terrigène
198
III.4.2.2 - SOLEA IMPAR
Les signatures isotopiques en carbone et en azote de la sole adriatique, S. impar, ont été
déterminées à partir d'échantillons de muscle blanc provenant de 10 juvéniles G0 (LT = 29-72 mm) et de
14 adultes (LT = 162-212 mm). Les signatures individuelles obtenues fluctuent entre -20,53‰ et -
16,13‰ pour le δ13C et entre 10,18‰ et 12,09‰ pour le δ15N, autour de moyennes respectives
de -17,69±0,23‰ et de 10,93±0,11‰. Sur la base d'une ingestion de matériel organique exclusivement
d'origine marine, le niveau trophique moyen global estimé pour l'espèce est de 3,88 et la signature en
carbone correspondante de -17,19‰, ce qui est très proche du δ13C moyen obtenu pour l'espèce.
L'influence de la MOP terrestre sur la croissance et le développement des individus de l'espèce au large
du Rhône est donc très réduite. Cependant, compte tenu des modifications ontogéniques du régime
alimentaire observées chez S. impar au large du Rhône (c. f. Partie II, Chapitre II), il est possible que
l'importance du transfert de la MOP terrigène diffère entre les différents stades de vie benthique de
l'espèce et qu'il intervienne de façon non négligeable dans la croissance pondérale soit des juvéniles, soit
des adultes de l'espèce.
Taille des individus, signature isotopique et transfert de la MOP terrigène
La signature isotopique varie en fonction de la taille des individus. En ce qui concerne l’azote,
le δ15N augmente significativement avec la taille des individus, indiquant une légère élévation du niveau
trophique au cours de la croissance (Fig. III.19).
Figure III.19 - Régression linéaire significative (r = 0,458, r2 = 0,210 ; P < 0,05) obtenue entre le δ15N (en ‰)et la taille (LT, en mm) chez Solea impar au large du Rhône (0-20 m). ----- = intervalle de confiance (95%).
LT (mm)
10,0
10,2
10,4
10,6
10,8
11,0
11,2
11,4
11,6
11,8
12,0
12,2
12,4
12,6
0 50 100 150 200 250
δ15N = 0,0039 LT + 10,4235
δ15N
(‰)

Réseaux trophiques et suivi de la MOP terrigène
199
Compte tenu de la forte variabilité du δ15N à chaque taille, la signature en azote ne diffère pas
de façon significative (Test de Student : T (22) = 1,834 ; P > 0,080) entre les juvéniles G0 (29-72 mm de
LT) et les adultes (LT ≥ 162 mm) de l'espèce (Figure III.20). Le δ15N moyen passe néanmoins de
10,69±0,12‰ chez les premiers à 11,10±0,17‰ chez les seconds, ce qui correspond à une augmentation
moyenne du niveau trophique de +0,13.
Figure III.20 – Représentation en boites de Tukey des δ15N (‰) observés chez les plus petits juvéniles(G0 : 29-72 mm de LT) et les adultes (LT ≥ 162 mm) de Solea impar au large du Rhône (0-20 m). NT =niveau trophique calculé à partir du δ15N moyen.
Une relation significative a également été mise en évidence entre le δ13C et la taille des
individus de S. impar (Fig. III.21).
Figure III.21 - Régression linéaire significative (r = 0,804, r2 = 0,646 ; P < 0,001) obtenue entre le δ13C (en ‰)et la taille (LT, en mm) chez Solea impar au large du Rhône (0-20 m). ----- = intervalle de confiance (95%).
Max non-aberrantMin non-aberrant75%25%Médiane
10,0
10,5
11,0
11,5
12,0
12,5
Juvéniles G0 Adultes
δ15N
(‰)
( NT = 3,44 ) ( NT = 3,57 )
LT (mm)
-20,5
-20,0
-19,5
-19,0
-18,5
-18,0
-17,5
-17,0
-16,5
-16,0
-15,5
0 40 80 120 160 200 240
δ13C
(‰)
δ13C = 0,0136 LT - 19,465

Réseaux trophiques et suivi de la MOP terrigène
200
Les modifications de δ13C entre les plus petits juvéniles et les adultes de l'espèce sont
significatives (Test de Student : T (22) = 5,927; P < 0,001) (Fig. III.22). Le δ13C augmente en moyenne
de +1,75‰ entre ces deux stades de vie benthique qui, pourtant, fréquentent les même fonds (0-20 m de
profondeur).
Figure III.22 – Représentation en boites de Tukey des δ13C (‰) observés chez les plus petits juvéniles(G0 : 29-72 mm de LT) et les adultes (LT ≥ 162 mm) de Solea impar au large du Rhône (0-20 m). Lesvaleurs entre parenthèses correspondent aux δ13C moyens ± erreur-types obtenus à chaque profondeur.
Compte tenu des signatures obtenues, l'influence de la MOP rhodanienne est uniquement
perceptible chez les plus petits juvéniles de S. impar, la signature isotopique des adultes étant clairement
"marine" (Tableau III.13).
Tableau III.13 – Signatures isotopiques moyennes en azote et en carbone observées, et niveau trophique et δ13Cthéoriques calculés dans le cas d'une alimentation d'origine exclusivement "marine" pour les juvéniles G0 et lesadultes de S. impar au large du Rhône.
δ15N observéNiveau trophique
théorique δ13C théorique δ13C observé δ13C obs. - δ13C théo.
Juvéniles G0 10,69 3,44 -17,32 -18,71±0,15 - 1,39
Adultes 11,10 3,57 -17,09 -16,96±0,22 + 0,13 (n.s.)
Ceci s'explique par une modification de la composition du régime alimentaire de S. impar entre
ces deux stades de vie benthique. En effet, l'alimentation des juvéniles G0 de l'espèce est dominée par
les polychètes, alors que les bivalves constituent l'essentiel du régime alimentaire des adultes (Fig.
III.23).
Max non-aberrantMin non-aberrant75%25%Médiane
-19,5
-19,0
-18,5
-18,0
-17,5
-17,0
-16,5
-16,0
-15,5
Juvéniles G0 Adultes
( -18,71±0,15‰ ) ( -16,96±0,22‰ )
δ13C
(‰)

Réseaux trophiques et suivi de la MOP terrigène
201
Figure III.23 – Composition pondérale (Cp) du régime alimentaire des juvéniles G0 (a) et desadultes (b) de S. impar au large du Rhône.
L'alimentation décrite pour chacun des deux stades de vie benthique explique relativement bien
la signature en carbone observée. Les δ13C théoriques obtenus sur la base des signatures des proies
ingérées pondérées par leur importance pour chacun des deux stades sont proches des signatures
obtenues, surtout pour les juvéniles (Tableau III.14).
Tableau III.14 – Signatures isotopiques théoriques en azote et en carbone calculées en fonction del'alimentation des individus et δ13C et δ15N moyens observés pour les juvéniles G0 et les adultes de S. impar aularge du Rhône. (pour les calculs, voir chapitre matériel et méthodes).
Juvéniles G0 Adultesδ15N δ13C δ15N δ13C
Proie théorique (calcul alimentation) 6,76 -20,25 7,82 -19,45
Signature théorique attendue 9,96 -18,85 11,02 -17,65
Signature isotopique observée 10,69±0,12 -18,71±0,15 11,10±0,17 -16,96±0,22
La signature en azote observée pour les adultes est très proche de celle calculée à partir du δ15N
des proies ingérées. Par contre, le δ15N calculé pour les juvéniles est inférieur de 1‰ à celui réellement
obtenu. Il est possible que la signature isotopique en azote des Spionidés ingérés par ces individus soit
légèrement supérieure à celle retenue pour la présente étude (calculée à partir de celles des polychètes
suspensivores et dépositivores) ou que la taille de certaines proies analysées soit inférieure à celle des
organismes ingérés dans le milieu.
En dépit des légères différences de signature observées, le positionnement des juvéniles et des
adultes de S. impar par rapport à leurs proies potentielles sur la zone d'étude explique les signatures
observées pour ces deux stades de vie benthique et permet de décrire le transfert de la matière organique
d'origine terrestre vers cette espèce au large du Rhône (Fig. III.24).
(a) Juvéniles G0 (LT < 100 mm)
Polychètes(spionidés) 65,6%
bivalves7,8%
amphipodes6,4%
mysidacés6,5%
cumacés13,7%
(b) Adultes (LT > 160 mm)
bivalves78,4%
cumacés2,2%
amphipodes4,2%
copépodes0,9%
gastéropodes1,3%
mysidacés8,3%
ophiures1,8%
polychètes 2,9%

Réseaux trophiques et suivi de la MOP terrigène
202
Figure III.24 - Réseaux trophiques des juvéniles G0 (Si J) et des adultes (Si A) de S. impar au large duRhône. MOP = matière organique particulaire, pol = polychète, S = suspensivore, DS = dépositivore de surface, C =carnivore. Les flèches indiquent les principales voies de transfert de matière organique déterminées par l'étude descontenus stomacaux des poissons (proies dont Cp > 10%).
Les juvéniles G0 et les adultes de l'espèce se situent en bout du réseau trophique basé sur la
matière organique d'origine marine. Les proies de signature "marine" ingérées ont donc, quel que soit le
stade de vie considéré, une importance prépondérante pour la croissance et la condition des individus de
l'espèce. Les juvéniles G0 ont néanmoins une signature beaucoup plus négative que les adultes et se
situent en marge du réseau trophique "marin". Ceci confirme l'importance des polychètes ingérés
(espèces suspensivores et/ou dépositivores de surface de la famille des Spionidae) dans leur
alimentation et indique une légère influence des apports en MOP terrigène (via les espèces
dépositivores) sur la croissance des individus de S. impar au cours des premiers mois de vie benthique.
pol C
MOP Rhône
sédiment
MOP eau
phytopKcumacé
copépode
Si J
bivalve juvénile
bivalvesiphon
mysidacé
amphipode
gastéropodeophiure
Si A
pol DS
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-17
-161 2 3 4 5 6 7 8 9 10 11 12
δ 15 Ν
δ 13 C
pol S

Réseaux trophiques et suivi de la MOP terrigène
203
III.4.2.3 - BUGLOSSIDIUM LUTEUM
Pour la solenette, B. luteum, les signatures isotopiques en carbone obtenues pour l'ensemble des
individus analysés (10 juvéniles G0 de 30-45 mm et 21 adultes de 94-130 mm de LT) fluctuent entre -
21,66‰ et -17,10‰, autour d'une moyenne de -18,55±0,31‰. Cette signature isotopique indique
l'incorporation, dans la chair des individus, de matériel organique provenant en partie d'une source de
δ13C plus négatif que le phytoplancton marin. En effet, les signatures en azote obtenues pour l'espèce au
large du Rhône varient de 8,15 à 11,76‰ autour d'une moyenne de 10,33±0,21‰. Sur la base d'une
assimilation exclusive de matériel organique d'origine marine, le niveau trophique moyen global estimé
pour l'espèce serait de 3,69 et la signature en carbone correspondante de -17,52‰. De même que pour S.
solea, la différence observée entre le δ13C attendu et celui observé ne peut s'expliquer que par une
incorporation non négligeable de matériel d'origine terrestre dans la chair des individus de l'espèce. Il
est cependant possible, du fait des variations ontogéniques et bathymétriques de l'alimentation observées
pour B. luteum sur la zone d'étude (c. f. partie II, Chapitre II), que ce transfert de MOP terrigène diffère
selon le stade de vie benthique ou la profondeur considérés.
Taille des individus, signature isotopique et transfert de la MOP terrigène
La signature isotopique varie en fonction de la taille des individus. En ce qui concerne l’azote,
le δ15N est positivement corrélé (P < 0,001) avec la taille des individus, indiquant une augmentation du
niveau trophique au cours de la croissance (Fig. III.25).
Figure III.25 - Régression linéaire significative (r = 0,458, r2 = 0,210 ; P < 0,05) obtenue entre le δ15N (en ‰)et la taille (LT, en mm) chez Buglossidium luteum au large du Rhône. ----- = intervalle de confiance (95%).
LT (mm)
7,0
7,5
8,0
8,5
9,0
9,5
10,0
10,5
11,0
11,5
12,0
12,5
0 10 20 30 40 50 60 70 80 90 100 110 120 130 140 150
δ15N = 0,0312 LT + 7,6455
δ15N
(‰)

Réseaux trophiques et suivi de la MOP terrigène
204
La signature en azote diffère de façon significative (Test de Student : T (22) = 12,544 ; P < 0,001)
entre les juvéniles G0 (30-45 mm de LT) et les adultes (LT ≥ 94 mm) de l'espèce (Fig. III.26). Le δ15N
moyen passe de 8,77±0,18‰ chez les juvéniles à 11,07±0,09‰ chez les adultes, ce qui correspond à une
augmentation de niveau trophique moyenne de +0,82.
Figure III.26 – Représentation en boites de Tukey des δ15N (‰) observés chez les plus petits juvéniles(G0 : 30-45 mm de LT) et les adultes (LT ≥ 94 mm) de Buglossidium luteum au large du Rhône. NT =niveau trophique calculé à partir du δ15N moyen.
En ce qui concerne le carbone, une relation significative (P < 0,01) a également été mise en
évidence entre le δ13C et la taille des individus (Fig. III.27).
Figure III.27 - Régression linéaire significative (r = 0,485, r2 = 0,235 ; P < 0,01) obtenue entre le δ13C (en ‰)et la taille (LT, en mm) chez Buglossidium luteum au large du Rhône. ----- = intervalle de confiance (95%).
Max non-aberrantMin non-aberrant75%25%MédianeAberrants
7,5
8,0
8,5
9,0
9,5
10,0
10,5
11,0
11,5
12,0
12,5
13,0
Juvéniles G0 Adultes
δ15N
(‰)
(NT =3,20) (NT = 3,92)
δ13C
(‰)
LT (mm)
-22,5
-22,0
-21,5
-21,0
-20,5
-20,0
-19,5
-19,0
-18,5
-18,0
-17,5
-17,0
-16,5
-16,0
-15,5
-15,0
0 10 20 30 40 50 60 70 80 90 100 110 120 130 140
δ13C = - 0,0251 LT - 16,392

Réseaux trophiques et suivi de la MOP terrigène
205
Les modifications de δ13C entre les plus petits juvéniles et les adultes de l'espèce sont
significatives (Test de Student : T (22) = 5,927; P < 0,001). Le δ13C, relativement stable et élevé chez les
juvéniles G0 de B. luteum, présents uniquement à 0-20 m, diminue en moyenne de 1,52‰ et devient très
variable chez les adultes de l'espèce répartis entre 0 et 100 m de profondeur (Fig. III.28).
Figure III.28 – Représentation en boites de Tukey des δ13C (‰) observés chez les plus petits juvéniles(G0 : 30-45 mm de LT) et les adultes (LT ≥ 94 mm) de Buglossidium luteum au large du Rhône (0-100 m).Les valeurs entre parenthèses correspondent aux δ13C moyens ± erreur-types obtenus à chaqueprofondeur.
Compte tenu des signatures obtenues, l'influence de la MOP rhodanienne n'est perceptible que
chez les adultes de B. luteum, les juvéniles G0 de l'espèce ayant une signature clairement "marine"
(Tableau III.15). Ceci s'explique par la modification de la composition du régime alimentaire de B.
luteum entre ces deux stades de vie benthique. En effet, l'alimentation des adultes de l'espèce est
dominée par les polychètes dépositivores, alors que les juvéniles G0 présentent une alimentation plus
diversifiée à base de crustacés, de mollusques et de polychètes, (Fig. III.29).
Tableau III.15 – Signatures isotopiques moyennes en azote et en carbone observées et niveau trophique et δ13Cthéoriques calculés dans le cas d'une alimentation d'origine exclusivement "marine" pour les juvéniles G0 et lesadultes de B. luteum au large du Rhône.
δ15N observéNiveau trophique
théorique δ13C théorique δ13C observé δ13C obs. - δ13C théo.
Juvéniles G0 8,77 3,20 -18,40 -17,52±0,15 + 0,88
Adultes 11,07 3,92 -17,11 -19,04±0,22 - 1,93
Max non-aberrantMin non-aberrant75%25%Médiane
-22,0
-21,5
-21,0
-20,5
-20,0
-19,5
-19,0
-18,5
-18,0
-17,5
-17,0
-16,5
Juvéniles G0 Adultes
δ13C
(‰)
( -17,52±0,15‰ ) ( -19,04±0,22 ‰)

Réseaux trophiques et suivi de la MOP terrigène
206
Figure III.29 – Composition pondérale (Cp) du régime alimentaire des juvéniles G0 (a) et desadultes (b) de B. luteum au large du Rhône.
Pour B. luteum, l'alimentation explique relativement bien la signature en carbone observée,
surtout chez les adultes. Pour ces derniers, les δ15N et δ13C théoriques obtenus sur la base des signatures
des proies ingérées pondérées par leur importance alimentaire sont très proches des signatures obtenues
par analyse de la chair des poissons (Tableau III.16).
Par contre, le δ15N et le δ13C calculés pour les juvéniles diffèrent respectivement de -2‰ et de
+1,5‰ à ceux obtenus par l'analyse isotopique de la chair des poissons. Il est possible que les signatures
globales des proies utilisées pour le calcul des signatures théoriques soient différentes de celles des
organismes ingérés dans le milieu. La petite taille des juvéniles de B. luteum réduit notamment la taille
de leurs proies et les polychètes carnivores et les gastéropodes ingérés par ce groupe de poissons sont
essentiellement des individus encore au stade juvénile pour lesquels la signature isotopique exacte n'a
pu être déterminée. De même les signatures utilisées pour les copépodes et les cumacés sont des
signatures globales susceptibles de différer de celles des espèces de ces deux catégories ingérées à 0-20
m par les jeunes solenettes. Dans tous les cas, la signature obtenue pour les juvéniles de l'espèce indique
l'ingestion importante d'un matériel ayant une signature en carbone élevée, de l'ordre de celles obtenues
à 0-20 m pour les macroures, les polychètes suspensivores, les ophiures ou les siphons de bivalve
analysés. Parmi ces proies, les polychètes suspensivores et les macroures présentent les δ15N les plus
compatibles avec la signature en azote attendue.
Tableau III.16– Signatures isotopiques théoriques en azote et en carbone calculées en fonction de l'alimentationdes individus et δ13C et δ15N moyens observés pour les juvéniles G0 et les adultes de B. luteum au large duRhône. (pour les calculs, voir chapitre matériel et méthodes).
Juvéniles G0 Adultesδ15N δ13C δ15N δ13C
Proie théorique (calcul alimentation) 6,63 -20,86 7,55 -21,29
Signature théorique attendue 9,83 -19,06 10,75 -19,49
Signature isotopique observée 8,77±0,18 -17,52±0,15 11,07±0,09 -19,04±0,22
(a) Juvéniles G0 (LT < 50 mm)
copépodes
16,4% polychètes C21,9%
bivalves10,4%
gastéropodes
16,4%
amphipodes17,4%
cumacés17,4%
(b) Adultes (LT > 100 mm)
bivalves14,8%
mysidacés4,2%
amphipodes8,3%
copépodes1,2%
cumacés1,3%
polychètes 56,6%(surtout C +DsS)
gastéropodes
0,8%
macroures9,8%
brachyoures2,4%

Réseaux trophiques et suivi de la MOP terrigène
207
Malgré la légère différence de signature observée pour les juvéniles par rapport à celle attendue,
le positionnement des juvéniles et des adultes de B. luteum par rapport à leurs proies potentielles sur la
zone d'étude explique de façon acceptable les signatures observées pour ces deux stades de vie
benthique et permet de décrire le transfert de la matière organique d'origine terrestre vers cette espèce au
large du Rhône (Fig. III.30).
Figure III.30 - Réseaux trophiques des juvéniles G0 (Bl J) et des adultes (Bl A) de B. luteum au large duRhône. MOP = matière organique particulaire, brach. = brachyoure, pol = polychète, S = suspensivore, DS =dépositivore de surface, DsS = dépositivore de sub-surface, C = carnivore. Les flèches indiquent les principales voiesde transfert de matière organique déterminées par l'étude des contenus stomacaux des poissons (proies dont Cp > 10%).
Les juvéniles G0 et les adultes de l'espèce se situent respectivement en bout du réseau trophique
basé sur la matière organique d'origine marine et en bout de celui impliquant la MOP terrigène. Ceci
confirme l'importance des polychètes ingérés dans l'alimentation des adultes de B. luteum et indique une
influence non négligeable des apports en MOP terrigène sur la condition des reproducteurs de l'espèce
alors que les proies de signature "marine" ont une importance prépondérante pour la croissance et la
condition des juvéniles.
pol DsS
MOP Rhône
sédiment
MOP eau
phytopKcumacé
copépode
mysidacé
brach.
bivalve
Bl J
pol C
macroure
amphipode
Bl Agastéropode
pol DS
-28
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-17
-160 1 2 3 4 5 6 7 8 9 10 11 12
δ 15 Ν
δ 13 C

Réseaux trophiques et suivi de la MOP terrigène
208
Variation de la signature isotopique en fonction de la profondeur
A taille égale, la profondeur n’a pas d’influence significative sur le δ15N des individus de B.
luteum au large du Rhône (ANOVA : F(2, 18) = 0,253 ; P > 0,779). En revanche, une différence
significative (ANOVA : F (2, 18) = 7,863 ; P < 0,01) du δ13C a été mise en évidence avec la profondeur
(Fig. III.31). Le δ13C des adultes de B. luteum capturés entre 30 et 50 m est significativement plus
négatif (P < 0,01; Annexe 43) que celui de ceux récoltés dans des zones moins profondes (0-20 m) ou
plus profondes (70-100 m). Il diffère également de façon significative (P < 0,001) du δ13C des juvéniles,
ces derniers, uniquement capturés à 0-20 m, présentant une signature en carbone similaire à celle des
adultes à 0-20 m et 70-100 m. Ainsi, la différence globale de δ13C observée entre juvéniles et adultes de
B. luteum au large du Rhône est essentiellement liée à la fréquentation des fonds de 30-50 m par ces
derniers.
Figure III.31 - Représentation en boites de Tukey des δ13C obtenus pour les individus de B. luteum de LT
≥ 94 mm présents au large du Rhône, en fonction de la profondeur de récolte des poissons. Les valeursentre parenthèses correspondent aux δ13C moyens ± erreur-types obtenus à chaque profondeur.
Les différences bathymétriques de signatures en carbone s'expliquent par une double
modification, selon la profondeur, de l'alimentation des adultes de l'espèce et de l'exploitation des
sources terrestres par les invertébrés benthiques (Fig. III.32). Au niveau des petits fonds sableux entre 0
et 20 m de profondeur (Fig. III.32-a), les polychètes représentent une part non négligeable de
l'alimentation des adultes de l'espèce (Cp = 46%) mais leur régime alimentaire comprend également de
nombreuses proies de signature "marine" (bivalves, amphipodes, mysidacés, etc) dont l'ingestion
provoque la hausse de δ13C et la forte variabilité inter-individuelle observée à cette profondeur.
δ13C
(‰)
Max non-aberrantMin non-aberrant75%25%Médiane
-22,5
-22,0
-21,5
-21,0
-20,5
-20,0
-19,5
-19,0
-18,5
-18,0
-17,5
-17,0
-16,5
-16,0
-15,5
0-20 m 30-50 m 70-100 m ( -18,47±0,61‰ ) ( -20,71±0,44‰ ) ( -17,99±0,52‰ )

Réseaux trophiques et suivi de la MOP terrigène
209
(a) 0-20 m de profondeur
(b) 30-50 m de profondeur
(c) 70-100 m de profondeur
Figure III.32- Composition pondérale du régime alimentaire (Cp), réseau trophique et transfert de la matière chezles adultes de B. luteum ( LT ≥ 94 mm) à 0-20 m (a), 30-50 m (b) et 70-100 m (c) de profondeur au large duRhône. Les flèches indiquent les transferts alimentaires de matière organique.
pol DS
MOP Rhône
sédimentMOP eau
phytopKbivalvejuvénile
copépode
gobiidae
brachyoure
amphipodecumacé
bivalvesiphon
macroure
pol S
ophiureBl A
mysidacé
pol C
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-17
-161 2 3 4 5 6 7 8 9 10 11 12
δ 15Ν
δ13 C
brachyoures2,9%
amphipodes11,8%
poisson (gobiidae)6,8%
polychètes 46,0%(dont 57% C, 33% DS et 10% S)
bivalves17,2%
ophiures0,9%
macroures4,7%
copépodes2,2% cumacés
3,2%
mysidacés3,1%
leptostracés1,2%
bivalves8,3%
gastéropodes1,0%
isopodes0,8%
macroures2,2%
amphipodes8,1%
mysidacés4,5% cumacés
2,6%
brachyoures2,6%
polychètes 69,9% (dont 52% DsS, 38% C, 8% DS et 2% S)
amphipodes23,5%
macroures25,8%
mysidacés20,1%
cumacés9,2%
copépodes3,3% polychètes
18,1% (dont 72% C, et 28% DS)
mysidacé
MOP Rhône
sédiment
MOP eau
phytopK
Bl A
cumacé
macroure
amphipode
copépode
pol DS
polC
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-171 2 3 4 5 6 7 8 9 10 11 12
δ 15
Ν
δ13 C
Bl A
MOP Rhône
sédimentMES eau
phytopKcrabe
Gastéropode
cumacé
pol DS
mysidacé
crevette
pol C
amphipode
pol DsS
bivalve
pol S
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-171 2 3 4 5 6 7 8 9 10 11 12
δ15Ν
δ13C
macrourebrachyoure
MOP

Réseaux trophiques et suivi de la MOP terrigène
210
Au niveau des vases plus profondes, l'alimentation des adultes est moins diversifiée qu'à 0-20
m. Elle diffère selon la profondeur, entraînant des différences de signatures isotopiques entre les
poissons capturés à 30-50 m et à 70-100 m. Ce phénomène est accentué par des modifications
bathymétriques du transfert des sources de matière organique marine et terrestre au sein du réseau
trophique des solenettes. En effet, à 30-50 m, les polychètes dominent dans le régime alimentaire des
adultes (Cp = 70%), alors qu'à 70-100 m, ils représentent moins de 20% en poids du total des proies
ingérées. On observe également, pour cette catégorie de proies, une modification, selon la profondeur,
des groupes éthologiques consommés : les polychètes ingérés sont essentiellement des dépositivores de
sub-surface et des carnivores à 30-50 m et des carnivores et des dépositivores de surface à 70-100 m.
Or, à 30-50 m, l'ensemble des polychètes présente une signature nettement terrestre (Fig. III.32-b) alors
qu'à 70-100 m, seuls les dépositivores de sub-surface (non ingérés à cette profondeur) dépendent des
apports rhodaniens en MOP terrigène (Fig. III.32-c). Ce phénomène explique les différences de δ13C
moyen observées et la forte homogénéité des signatures obtenues pour chacune des deux tranches
bathymétriques. Sur les fonds vaseux de 30-50 m, les solenettes adultes tirent la quasi totalité de leur
alimentation du réseau trophique basé sur la MOP rhodanienne, alors qu'à 70-100 m de profondeur,
l'ensemble des proies ingérées dépend de l'exploitation des sources de production marine.
III.4.2.4 - ARNOGLOSSUS LATERNA
Les signatures isotopiques en carbone et en azote de la population d'A. laterna au large du
Rhône ont été déterminées à partir d'échantillons de muscle blanc provenant de 12 juvéniles G0 (LT =
32-49 mm) et de 16 adultes (LT = 97-136 mm) de l'espèce. Les signatures individuelles obtenues
fluctuent entre -21,21 et -16,56‰ pour le δ13C et entre 7,94 et 10,77‰ pour le δ15N, autour de
moyennes respectives de -18,53 ± 0,25‰ et de 9,42±0,18‰. Sur la base d'une alimentation d'origine
exclusivement marine, le niveau trophique global moyen de l'espèce serait de 3,40 et la signature en
carbone correspondante de -18,03‰, ce qui est très proche du δ13C moyen observé. L'influence de la
MOP terrestre sur la croissance et le développement des individus de l'espèce au large du Rhône est
donc très réduite. Cependant, compte tenu des modifications ontogénétiques et bathymétriques du
régime alimentaire d'A. laterna au large du Rhône (c. f. partie II, Chapitre II), il est possible que le
transfert de la MOP terrigène soit non négligeable pour l'un des stades de vie benthique et/ou à une
profondeur donnée.
Taille des individus, signature isotopique et transfert de la MOP terrigène
La signature isotopique varie en fonction de la taille des individus. En ce qui concerne l’azote,
le δ15N est positivement corrélé (P < 0,001) avec la taille des individus, indiquant une augmentation du
niveau trophique au cours de la croissance (Fig. III.33).

Réseaux trophiques et suivi de la MOP terrigène
211
Figure III.33 - Régression linéaire significative (r = 0,820, r2 = 0,672 ; P < 0,001) obtenue entre le δ15N (en ‰)et la taille (LT, en mm) chez Arnoglossus laterna au large du Rhône. ----- = intervalle de confiance (95%).
La signature en azote augmente de façon significative (Test de Student : T (26, 16, 12) = 7,440 ; P <
0,001) entre les plus petits juvéniles (32-49 mm de LT) et les adultes (LT ≥ 97 mm) de l'espèce (Fig.
III.34). Le δ15N moyen passe de 8,52±0,16‰ chez les premiers à 10,10±0,14‰ chez les derniers, ce qui
correspond à une augmentation de niveau trophique de +0,50.
Figure III.34 – Représentation en boites de Tukey des δ15N (‰) observés chez les plus petits juvéniles(G0 : 32-49 mm de LT) et les adultes (LT ≥ 97 mm) d'A. laterna au large du Rhône. NT = niveautrophique calculé à partir du δ15N moyen.
Max non-aberrantMin non-aberrant75%25%MédianeAberrants
7,5
8,0
8,5
9,0
9,5
10,0
10,5
11,0
Juvéniles G0 Adultes
δ15N
( NT = 3,12) ( NT = 3,62 )
LT (mm)
7,5
8,0
8,5
9,0
9,5
10,0
10,5
11,0
0 10 20 30 40 50 60 70 80 90 100 110 120 130 140 150
δ15N
(‰)
δ15N = 0,0233 LT + 7, 5385

Réseaux trophiques et suivi de la MOP terrigène
212
En ce qui concerne le carbone, aucune relation significative n’a été mise en évidence entre le
δ13C et la taille des individus étudiés (régression linéaire : r = 0,125, r2 = 0,016 ; P > 0,264). Les
modifications de δ13C entre les plus petits juvéniles (δ13C moy. = -18,93 ± 0,46‰) et les adultes (δ13C moy.
= -18,23 ± 0,26‰) de l'espèce ne sont pas significatives (Test de Student : T (26, 16, 12) = 1,423; P >
0,167).
Compte tenu des signatures obtenues, l'influence de la MOP rhodanienne est réduite quel que
soit le stade de vie d'A. laterna, mais légèrement plus marquée chez les adultes de l'espèce (Tableau
III.17). En effet, les signatures en carbone observées sont plus négatives que celles attendues sur la base
d'une alimentation exclusivement marine mais restent cependant proches de ces dernières.
Tableau III.17 – Signatures isotopiques moyennes en azote et en carbone observées et niveau trophique et δ13Cthéoriques calculés dans le cas d'une alimentation d'origine exclusivement "marine" pour les juvéniles G0 et lesadultes d'A. laterna au large du Rhône.
δ15N observéNiveau trophique
théorique δ13C théorique δ13C observé δ13C obs. -δ13C théo.
Juvéniles G0 8,52 3,12 -18,54 -18,93±0,46 - 0,39
Adultes 10,10 3,62 -17,65 -18,23±0,26 - 0,58
Ceci s'explique par l'étude des régimes alimentaires de ces deux stades de vie benthiques.
L'importance des polychètes dans l'alimentation des juvéniles et des adultes est réduite (Cp ≤ 26%) et
les individus ingérés appartiennent surtout à des espèces suspensivores et carnivores (Fig. III.35).
Figure III.35 – Composition pondérale (Cp) du régime alimentaire des juvéniles G0 (a) et desadultes (b) d'A. laterna au large du Rhône. C = carnivores, DS = dépositivores de surface, DsS =dépositivores de sub-surface, S = suspensivores.
(a) Juvéniles G0 (LT ≤ 50 mm)
cumacés47,3%
amphipodes15,8%
bivalves7,2%
polychètes 21,5% (dont 60% C, 30% S, et 10% DS)
copépodes4,5%
leptostracés3,4%
(b) Adultes (LT ≥ 97 mm)
anomoures0,9%
brachyoures11,4%
mysidacés10,4%
cumacés0,6%
copépodes0,2%
poissons(gobiidae)
0,2% phoronidiens3,8%
macroures39,1%
amphipodes5,7%
bivalves1,7%
polychètes 26,0%(dont 52% S, 29% C,18% DS et 1% DsS)

Réseaux trophiques et suivi de la MOP terrigène
213
L'alimentation décrite pour chacun des deux stades de vie explique bien la signature en carbone
observée. Les δ13C et δ15N théoriques obtenus sur la base des signatures des proies ingérées pondérées
par leur importance alimentaire pour chacun des deux stades sont très proches des signatures obtenues
par analyse de la chair des poissons (Tableau III.18).
Tableau III.18 – Signatures isotopiques théoriques en azote et en carbone calculées en fonction del'alimentation des individus et δ13C et δ15N moyens observés pour les juvéniles G0 et les adultes d'A. laterna aularge du Rhône. (pour les calculs, voir chapitre matériel et méthodes).
Juvéniles G0 Adultesδ15N δ13C δ15N δ13C
Proie théorique (calcul alimentation) 5,40 -21,31 7,04 -20,51
Signature théorique attendue 8,60 -19,51 10,24 -18,71
Signature isotopique observée 8,52±0,16 -18,93±0,46 10,10±0,14 -18,23±0,26
La signature en carbone observée pour les deux stades est cependant légèrement moins négative
que celle attendue dans chaque cas. Il est possible que ce phénomène provienne des signatures des
phoronidiens et des leptostracés, ingérés en petites quantités respectivement par les adultes et les
juvéniles de l'espèce mais dont les δ13C et le δ15N n'ont pu être déterminés au cours de la présente étude.
Dans ce cas, ces deux catégories de proies présenteraient une signature clairement marine avec un δ13C
particulièrement peu négatif.
Le positionnement des juvéniles et des adultes d'A. laterna par rapport à leurs proies potentielles
explique les signatures observées pour ces deux stades de vie benthique et permet de décrire le transfert
de la matière organique d'origine terrestre vers cette espèce au large du Rhône (Fig. III.36). Les
juvéniles G0 et les adultes de l'espèce se situent en bout du réseau trophique basé sur la matière
organique d'origine marine. Les proies de signature "marine" ingérées ont donc, quel que soit le stade de
vie considéré, une importance prépondérante pour la croissance et la condition des individus de l'espèce.
Les adultes ont néanmoins une signature en carbone proche de celle des juvéniles (malgré un niveau
trophique supérieur) et se situent légèrement en marge du réseau trophique "marin", ce qui suggère
l'existence, pour ce stade de vie benthique, d'une incorporation supérieure de matériel d'origine terrestre
dans la chair des poissons.

Réseaux trophiques et suivi de la MOP terrigène
214
Figure III.36 - Réseaux trophiques des juvéniles G0 (Al J) et des adultes (Al A) d'A. laterna au large duRhône. MOP = matière organique particulaire, brach. = brachyoure, pol = polychètes. DS = dépositivore de surface, C= carnivore. Les flèches indiquent les principales voies de transfert de matière organique déterminées par l'étude descontenus stomacaux des poissons.
Variation de la signature isotopique en fonction de la profondeur
Chez A. laterna, la signature en azote se modifie de façon significative (ANOVA : F (2, 13) =
4,528 ; P < 0,05) avec la profondeur (Fig. III.37). Chez les adultes de l'espèce (seuls présents aux trois
tranches bathymétriques prospectées), le δ15N diminue de façon progressive avec la profondeur et les
individus capturés à 70-100 m présentent un δ15N significativement plus faible (P < 0,05 ; Annexe 44)
que ceux récoltés dans des zones moins profondes (0-20 m, 30-50 m). Le δ15N des adultes reste
cependant, quelle que soit la profondeur, significativement supérieur (P < 0,001) à celui des juvéniles,
uniquement capturés à 0-20 m.
Une modification significative (ANOVA : F(2, 13) = 7,200 ; P < 0,001) du δ13C avec la
profondeur a également été mise en évidence (Fig. III.38). Chez les adultes de l'espèce, le δ13C des
individus capturés à 0-20 m diffère de façon significative (P < 0,05 ; Annexe 45) de celui des individus
récoltés dans des zones plus profondes (30-50 m, 70-100 m). Il est également significativement différent
(P < 0,01) de celui des juvéniles dont la signature en carbone à 0-20 m est par contre similaire à celle
des adultes capturés à 30-50 m et 70-100 m de profondeur (P > 0,263).
Al J
anomoure
pol DS
amphipode
macroure
pol S
Al A
bivalve
brach.
pol C
copépodecumacé
phytopK
MOP eau
sédiment
MOP Rhône
mysidacé
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-171 2 3 4 5 6 7 8 9 10 11
δ 15 Ν
δ13C

Réseaux trophiques et suivi de la MOP terrigène
215
Figure III.37 – Représentation en boites de Tukey des δ15N (‰) observés chez les adultes (LT ≥ 97 mm)d'A. laterna présents au large du Rhône, en fonction de la profondeur de récolte des poissons. Les valeursentre parenthèses correspondent aux δ15N moyens ± erreur-types obtenus à chaque profondeur.
Figure III.38 - Représentation en boites de Tukey des δ13C obtenus pour les individus d'A. laterna de LT ≥97 mm présents au large du Rhône, en fonction de la profondeur de récolte des poissons. Les valeursentre parenthèses correspondent aux δ13C moyens ± erreur-types obtenus à chaque profondeur.
δ13C
(‰)
( -16,75±0,47‰ ) ( -19,20±0,16‰ ) ( -18,24±0,25‰ )
Max non-aberrantMin non-aberrant75%25%Médiane-20,0
-19,5
-19,0
-18,5
-18,0
-17,5
-17,0
-16,5
-16,0
0-20 m 30-50 m 70-100 m
Max non-aberrantMin non-aberrant75%25%Médiane9,5
10,0
10,5
11,0
0-20 m 30-50 m 70-100 m
( 10,49±0,66‰ ) ( 10,25±0,18‰ ) ( 9,67±0,12‰ )
δ15N
(‰)

Réseaux trophiques et suivi de la MOP terrigène
216
Ces différences bathymétriques de signatures en carbone s'expliquent par une double
modification, selon la profondeur, de l'alimentation des adultes de l'espèce et de l'exploitation des
sources "terrestres" par les invertébrés benthiques (Fig. III.39). Entre 0 et 20 m et à 30-50 m de
profondeur, les polychètes (surtout des suspensivores) tiennent une place importante (Cp > 28%) dans
l'alimentation des adultes de l'espèce (Fig. III.39 a et b). Cependant, l'exploitation des sources de
matière organique par les espèces ingérées diffère d'une tranche bathymétrique à l'autre. Les polychètes
suspensivores, majoritairement ingérés aux deux profondeurs, dépendent essentiellement du matériel
organique dérivé des sources de production marines à 0-20 m, alors qu'à 30-50 m, ils exploitent
majoritairement la MOP terrigène apportée par le Rhône. De plus, à 30-50 m de profondeur, l'essentiel
de l'alimentation des adultes d'A. laterna est constituée de macroures et de brachyoures. Or, à cette
profondeur, ces deux catégories de proies dépendent en partie des apports rhodaniens en MOP terrestre
pour leur croissance. Il en résulte une signature en carbone significativement plus négative à 30-50 m,
où la MOP terrigène influence de façon non négligeable la croissance pondérale des adultes de l'espèce,
qu'à 0-20 m où la quasi totalité de la matière organique exploitée par les arnoglosses est issue de la
production marine in situ. A 70-100 m, les polychètes ont une importance réduite (Cp < 7%) dans
l'alimentation (Fig. III.39-c). De plus, les individus ingérés (essentiellement des carnivores) dépendent
essentiellement, à cette profondeur, des sources de production marines. La signature moins négative
obtenue pour les adultes de l'espèce à 70-100 m s'explique par leur exploitation exclusive, à cette
profondeur, de proies appartenant au réseau trophique marin.

Réseaux trophiques et suivi de la MOP terrigène
217
(a) 0-20 m de profondeur
(b) 30-50 m de profondeur
(c) 70-100 m de profondeur
Figure III.39 - Composition pondérale du régime alimentaire (Cp), réseau trophique et transfert de la matière chezles adultes d'A. laterna ( LT ≥ 97 mm) à 0-20 m (a), 30-50 m (b) et 70-100 m (c) de profondeur au large du Rhône.Les flèches indiquent les transferts alimentaires de matière organique.
polychètes 28,8% (dont 62% S, 14% DS et 24%C)
bivalves 1,2%
amphipodes2,9%
macroures43,9%
poissons (gobiidae)0,1%
cumacés0,4%
mysidacés2,9%
brachyoures12,4%
anomoures1,3%
crust. ind.6,0%
crust. ind.7,9%
brachyoures5,3%
mysidacés3,7%
cumacés2,5%
poissons (gobiidae)
0,5%
phoronidiens16,1%
macroures10,6%
amphipodes18,0%
bivalves4,4%
polychètes 30,8% (dont 52% S, 27% DS, 18% C et 2% DsS)
Polychètes 6,2% (C > DS > S)
bivalves0,1%
amphipodes 3,4%
macroures41,7%
poissons(gobiidae)
0,5%copépodes0,8%
cumacés0,2%
mysidacés33,2%
brachyoures11,1%
crust. ind.2,7%
phoronidiens ?
pol DS
MOP Rhône
sédimentMOP eau
phytopK
brachyoure
Al A
cumacé pol C
mysidacé
macroure
anomoure
amphipode
gobiidae
bivalve pol S
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-17
-162 3 4 5 6 7 8 9 10 11 12
δ15 Ν
δ 13 C
pol C
mysidacé
gobiidae
cumacé
macroure
pol S
amphipode brachyoure
Al A
bivalve phytopK
MOP eau
sédiment
MOP Rhône
pol Ds
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-17
-162 3 4 5 6 7 8 9 10 11 12
δ15 Ν
δ13 C
pol S
MOP Rhône
sédiment
MOP eau
phytopK
cumacé
Al A
mysidacé
pol DS
pol C
macroure
amphipode
copépode brachyoure
bivalve
gobiidae
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-172 3 4 5 6 7 8 9 10 11 12
δ15
Ν
δ13 C

Réseaux trophiques et suivi de la MOP terrigène
218
III.4.2.5 - CITHARUS LINGUATULA
Pour la cithare, C. linguatula, les δ13C obtenus pour l'ensemble des individus analysés (12
juvéniles G0 de 63-84 mm et 12 adultes de 192-230 mm de LT) fluctuent entre -18,07‰ et -16,86‰
autour d'une moyenne de -17,62 ± 0,06‰. Cette signature, très marine, indique que la contribution de la
MOP terrigène à la croissance des individus de l'espèce au large du Rhône est négligeable. En effet, les
δ15N obtenus pour C. linguatula varient de 8,53‰ à 12,19‰ avec une valeur moyenne de 10,30±0,28‰.
Sur la base d'une alimentation d'origine exclusivement marine, le niveau trophique global moyen de
l'espèce serait de 3,68 et la signature en carbone correspondante de -17,54 ‰, ce qui est très proche du
δ13C moyen obtenu pour l'espèce. L'influence de la MOP terrestre sur la croissance et le développement
des individus de C. linguatula au large du Rhône est donc négligeable.
Taille des individus, signature isotopique et transfert de la MOP terrigène
La signature isotopique varie en fonction de la taille des individus. En ce qui concerne l’azote,
le δ15N augmente significativement avec la taille des individus (Fig. III.40), indiquant une augmentation
du niveau trophique au cours de la croissance.
Figure III.40 - Régression linéaire significative (r = 0,979, r2 = 0,958 ; P < 0,001) obtenue entre le δ15N (en‰) et la taille (LT, en mm) chez Citharus linguatula au large du Rhône. ----- = intervalle de confiance (95%).
Le δ15N augmente de façon significative (Test de Student : T(22) = 19,225 ; P < 0,001) entre les
plus petits juvéniles (63-84 mm de LT) et les adultes (LT ≥ 192 mm) de l'espèce (Fig. III.41). Il passe en
moyenne de 9,13±0,08‰ chez les juvéniles à 11,56±0,11‰ chez les adultes, ce qui correspond à une
augmentation de niveau trophique de +0,76.
LT (mm)
7,0
7,5
8,0
8,5
9,0
9,5
10,0
10,5
11,0
11,5
12,0
12,5
0 50 100 150 200 250
δ15N = 0,0187 LT + 7,6727
δ15N
(‰)

Réseaux trophiques et suivi de la MOP terrigène
219
Figure III.41 – Représentation en boites de Tukey des δ15N (‰) observés chez les plus petits juvéniles(G0 : 63-84 mm de LT) et les adultes (LT ≥ 192 mm) de C. linguatula au large du Rhône. NT = niveautrophique calculé à partir du δ15N moyen.
En ce qui concerne le carbone, aucune relation significative n’a été mise en évidence entre le
δ13C et la taille des individus étudiés (régression linéaire : r = 0,175, r2 = 0,031 ; P > 0,761). Les
modifications de δ13C entre les plus petits juvéniles (δ13C moy. = -17,60 ± 0,08 ‰) et les adultes (δ13C moy.
= -17,64 ± 0,08 ‰) de l'espèce ne sont pas significatives (Test de Student : T(22) = -0,289; P > 0,776).
Compte tenu des signatures obtenues, l'influence de la MOP rhodanienne est uniquement
perceptible chez les adultes de l'espèce, les juvéniles G0 ayant une signature clairement marine (Tableau
III.19).
Tableau III.19– Signatures isotopiques moyennes en azote et en carbone observées et niveau trophique et δ13Cthéoriques calculés dans le cas d'une alimentation d'origine exclusivement "marine" pour les juvéniles G0 et lesadultes de C. linguatula au large du Rhône.
δ15N observéNiveau trophique
théorique δ13C théorique δ13C observé δ13C obs. - δ13C théo.
Juvéniles G0 9,13 3,31 -18,20 -17,60±0,08 + 0,60
Adultes 11,56 4,07 -16,83 -17,64±0,08 - 0,81
Ceci s'explique par des modifications ontogéniques de l'alimentation chez C. linguatula. Bien
que les catégories de proies ingérées varient peu d'un stade de vie à l'autre, les mysidacés, abondamment
consommés par les juvéniles de l'espèce, sont en grande partie remplacés, chez les adultes, par des
macroures auxquelles s'ajoutent quelques polychètes dépositivores de sub-surface (Fig. III.42).
Max non-aberrantMin non-aberrant75%25%Médiane
8,0
8,5
9,0
9,5
10,0
10,5
11,0
11,5
12,0
12,5
Juvéniles G0 Adultes
δ15N
(‰)
( NT = 3,31) ( NT = 4,07 )

Réseaux trophiques et suivi de la MOP terrigène
220
Figure III.42 – Composition pondérale (Cp) du régime alimentaire des juvéniles G0 (a) et des adultes(b) de C. linguatula au large du Rhône.
Ces changements de composition du régime alimentaire s'accompagnent en outre d'une
modification de la taille des individus ingérés pour plusieurs catégories de proies, notamment les
poissons, les mysidacés et les macroures. La signature isotopique étant susceptible de varier fortement
(surtout pour l'azote) entre les individus de ces catégories de proies ingérés par les juvéniles et les
adultes, les signature moyennes par catégorie n'ont pas été utilisées. Elles ont été remplacées, dans
chaque cas, par les signatures des individus de tailles minimale et maximale capturés entre 30 et 100 m
de profondeur.
A condition de tenir compte des variations de la taille des proies ingérées, l'alimentation décrite
pour chacun des deux stades de vie explique relativement bien la signature en carbone observée. Les
δ13C théoriques obtenus sur la base des signatures des proies ingérées pondérées par leur importance
alimentaire pour chacun des deux stades sont proches des signatures obtenues, surtout pour les juvéniles
(Tableau III.20).
Tableau III.20 – Signatures isotopiques théoriques en azote et en carbone calculées en fonction del'alimentation des individus et δ13C et δ15N moyens observés pour les juvéniles G0 et les adultes de C.linguatula au large du Rhône. (pour les calculs, voir chapitre matériel et méthodes).
Juvéniles G0 Adultesδ15N δ13C δ15N δ13C
Proie théorique (calcul alimentation) 7,26 -19,76 9,19 -19,03
Signature théorique attendue 10,46 -17,96 12,39 -17,23
Signature isotopique observée 8,98±0,08 -17,60±0,08 11,63±0,11 -17,64±0,08
(a) Juvéniles G0 (LT< 100 mm )
poisson (gobiidae)64,8%
macroures5,4%
mysidacés29,8%
(b) Adultes (LT > 160 mm )
mysidacés9,2%
poisson (gobiidae)73,5%
macroures16,1%
polychètes1,1%

Réseaux trophiques et suivi de la MOP terrigène
221
Par contre, les δ15N calculés pour les juvéniles et les adultes de l'espèce sont supérieurs à ceux
obtenus par analyse isotopique de la chair des poissons. Il est probable que ce phénomène reflète le fait
que la signature isotopique en azote des poissons, des mysidacés et des macroures ingérés diffère des
valeurs extrêmes retenues pour la présente étude.
Le positionnement des juvéniles et des adultes de C. linguatula par rapport à leurs proies
potentielles sur la zone d'étude explique les signatures observées pour ces deux stades de vie benthique
et permet de décrire les modalités du transfert de la matière organique vers cette espèce au large du
Rhône (Fig. III.43).
Figure III.43- Réseaux trophiques des juvéniles G0 (Cl J) et des adultes (Cl A) de C. linguatula au large duRhône (30-100 m de profondeur). Les flèches indiquent les voies de transfert de matière organique déterminées parl'étude des contenus stomacaux des poissons. MOP = matière organique particulaire, pol = polychètes, DS =dépositivore de surface, DsS = carnivore, "min." et "max." désignent respectivement les individus de taille minimale etmaximale observés pour chaque catégorie de proie.
Les juvéniles G0 et les adultes de l'espèce se situent en bout du réseau trophique basé sur la
matière organique d'origine marine. Les proies de signature "marine" ingérées ont donc, quel que soit le
stade de vie considéré, une importance prépondérante pour la croissance et la condition des individus de
l'espèce. Les adultes ont néanmoins une signature légèrement plus "terrestre" que les juvéniles. En effet,
ils présentent le même δ13C que ces derniers malgré leur δ15N supérieur. Ceci s'explique essentiellement
par l'ingestion, chez les adultes de C. linguatula, de fortes quantités de grands macroures (Crangonidae
et Alpheidae adultes) de δ13C inférieur et dépendant vraisemblablement en partie des sources de MOP
terrestre pour leur croissance. Le transfert de matériel organique d'origine terrestre vers les adultes de
l'espèce est en outre complété par l'ingestion de polychètes dépositivores.
gobiidae min.
Cl J
Alpheus
pol DsS
pol DS
gobiidaemax
mysidacé min.
Crangonmax.
Crangonmin.
Cl A
phytopK
MOP eau
sédiment
MOP Rhône
mysidacémax.
-28
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-17
-16
1 2 3 4 5 6 7 8 9 10 11 12
δ15Ν
δ13C

Réseaux trophiques et suivi de la MOP terrigène
222
Variation de la signature isotopique en fonction de la profondeur
La profondeur n’a d'influence significative ni sur le δ15N (ANOVA : F (1, 22) = 0,660 ; P > 0,425)
ni sur le δ13C des individus de C. linguatula (ANOVA : F (1, 22) = 0,0645 ; P > 0,064) récoltés au large
du Rhône. Le δ13C des cithares reste le même quelle que soit la profondeur considérée (P > 0,454;
Annexe 45), ce qui confirme les résultats obtenus par l'étude des contenus stomacaux de l'espèce (pour
chaque stade de vie benthique, la composition de l'alimentation reste la même aux deux profondeurs).
Le δ15N global des individus est identique aux deux profondeurs de capture de l'espèce mais la signature
en azote diffère de façon significative entre juvéniles et adultes tant à 30-50 m (P < 0,001 ; Annexe 46)
qu'à 70-100 m (P < 0,001 ; Annexe 46) (Fig. III.44).
Figure III.44 - Représentation en boites de Tukey des δ15N obtenus pour les juvéniles et les adultes de C.linguatula au large du Rhône, en fonction de la profondeur de récolte des poissons.
Le schéma global de transfert de la matière organique d'origine marine et d'origine terrestre vers
les juvéniles et les adultes de l'espèce est donc le même à 30-50 m qu'à 70-100 m. Les modifications de
la signature isotopique des proies selon la profondeur, liées à leurs différences bathymétriques
d'exploitation des deux principales sources de matière organique, n'influencent pas de façon
significative la signature des poissons du fait de l'importance réduite dans l'alimentation des deux
stades, des proies de signature variable (macroures, polychètes dépositivores de surface).
Max non-aberrantMin non-aberrant75%25%Médiane
8,0
8,5
9,0
9,5
10,0
10,5
11,0
11,5
12,0
12,5
30-50 m 70-100 m 30-50 m 70-100 m
δ15N
(‰)
juvéniles G0 adultes

Réseaux trophiques et suivi de la MOP terrigène
223
III.5 - DISCUSSION
III.5.1 - SOURCES POTENTIELLES ET ORIGINE DE LA MATIERE ORGANIQUE
L'utilisation des isotopes stables pour caractériser l'origine de la matière organique dans les
écosystèmes est d'autant plus efficace que les sources potentielles sont peu nombreuses et de signature
isotopiques distinctes (Peterson et Fry, 1987 ; Dufour et Gerdeaux, 2001). Au large de l'embouchure du
Rhône, ces deux conditions sont respectées. On dénombre au maximum cinq sources potentielles de
matière organique, dont deux sources autochtones marines (producteurs primaires planctoniques et
microphytobenthos) et trois sources allochtones, dont deux terrestres (MOP rhodanienne et débris de
végétaux terrestres charriés par le fleuve) et une marine (fragments de macrophytes marines amenés par
les courants). Bien que la signature isotopique du microphytobenthos n'ait pu être déterminée pour la
zone d'étude, les δ13C reportés dans la littérature pour cette catégorie de producteurs primaires (δ13C = -
18,5 à -11,9‰) (Riera et Richard, 1996, 1997 ; Kiyashko et al., 1998 ; Herman et al., 2000 ; Riera et al.,
2000 ; Riera et al., 2002 ; Takai et al., 2002) sont très différents de ceux obtenus pour les quatre autres
sources potentielles (δ13C < -22,1 ‰). Pour ces dernières, les signatures en carbone et en azote obtenues
sont en accord avec les données de la littérature (Ostrom et Fry, 1993) et permettent une
individualisation totale de chaque source par rapport aux trois autres. Cependant, en ce qui concerne les
fragments de macrophytes marines, le problème de la représentativité de la signature isotopique obtenue
se pose, dans la mesure où seuls des débris de Chlorophycées ont pu être analysés. Compte tenu du fait
que l'un des plus vastes herbiers à Posidonia oceanica de la région Marseillaise est situé à moins de 6
km de la zone d'étude et que le golfe de Fos abrite de vastes herbiers à Nanozostera noltii et Zostera
marina, des fragments de phanérogames marines sont également susceptibles d'être transportés de façon
ponctuelle sur les fonds échantillonnés. Les δ13C des phanérogames marines varient cependant
généralement de -19 à -3‰ avec des moyennes de -14‰ pour Posidonia oceanica et de -10‰ pour
Zostera marina (Nichols et al., 1985 ; Cooper et De Niro, 1989 ; Bricout et al., 1990 ; Fourqurean et al.,
1997) et sont très éloignés de ceux des autres sources principales de matière organique (MOP
rhodanienne, phytoplancton marin et végétaux terrestres), ce qui devrait permettre de déceler toute
contribution importante de ce groupe de producteurs primaires à la constitution des pools de matière
organique de la zone d'étude.
Pour les signatures isotopiques obtenues au large du Rhône, le carbone est celui des deux
éléments analysés qui permet la meilleure séparation des sources de matière organique individualisées.
La gamme de variation des δ13C obtenus (± 5,4‰) est plus importante que celle observée pour les δ15N
(± 3,1‰) et, surtout, l'étude des δ13C permet une séparation des sources selon leur origine, terrestre
(δ13C ≤ -25,5 ‰) ou marine (δ13C ≥ -23,0 ‰), contrairement à celle des δ15N. Ces résultats confirment
la faible efficacité des δ15N pour caractériser les sources de matière organique dans les écosystèmes
marins côtiers sous influence continentale (Couch, 1989). Dans ce type de milieux, leur pouvoir

Réseaux trophiques et suivi de la MOP terrigène
224
discriminant est réduit par rapport à celui des δ13C dont l'utilisation suffit généralement pour séparer les
sources de matière organique selon leur origine, marine ou continentale (Riera, 1995 ; Camusso et al.,
1999 ; Lee, 2000). Cependant, seule l'utilisation combinée des deux éléments (carbone et azote) permet
une individualisation totale des sources potentielles de matière organique au large du Rhône, notamment
dans le cas du phytoplancton marin et des débris de macrophytes marines (Chlorophycées), dont les
δ13C sont identiques mais les δ15N très différents. Ceci confirme l'intérêt, déjà souligné par plusieurs
auteurs (Peterson et al., 1985 ; Thomas et Cahoon, 1993 ; Riera, 1995), d'une approche pluri-
élémentaire lorsqu'il s'agit de déterminer l'origine exacte de la matière organique incorporée dans les
réseaux trophiques benthiques côtiers.
Contribution des différentes sources à la constitution des pools de matière organique
Les signatures isotopiques globales moyennes de la MOP de l'eau de surface au large du Rhône
et du sédiment de la zone d'étude sont similaires, surtout en ce qui concerne le carbone, avec des δ13C
très proches et situés entre ceux des sources de matière organique d'origine marine et ceux des sources
terrestres (Fig. III.45). Ceci suggère des origines analogues pour les pools de matière organique de l'eau
de surface et du sédiment et une participation non négligeable des apports provenant des sources
terrestres à leurs constitutions respectives.
Figure III.45 - Signatures isotopiques (δ13C et δ15N, en ‰) des sources potentielles et des pools dematière organique présents au niveau de l'eau de surface et du sédiment superficiel au large duRhône.
végétaux terrestres
sédiment
phytoplancton
MOP Rhône
macrophytes marines(Chlorophycées)
diatomées benthiques(Riera et al ., 1999)
MOP eau
-28
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-17
-16
-15
-14
0 1 2 3 4 5 6
δ 15 N
δ 13 C
Posidonia oceanica(Pinnegar et Polunin, 2000)
-13
SOURCES MARINES
SOURCES TERRESTRES

Réseaux trophiques et suivi de la MOP terrigène
225
Au large du Rhône, les macrophytes benthiques sont quasiment absentes et la production
primaire microphytobenthique ne concerne que les fonds sableux situés au-dessus de 20 m de
profondeur où elle reste très réduite (Bodoy et Plante-Cuny, 1980). La contribution potentielle des
micro et macrophytes benthiques locales à la constitution des pools de carbone de l'eau et du sédiment
est donc négligeable. L'essentiel de la matière organique disponible à la base des réseaux trophiques
benthiques provient de la sédimentation du matériel présent au niveau de la colonne d'eau, qu'il s'agisse
de MOP terrestre, de plancton marin ou de débris de macrophytes terrestres ou marines. Le δ13C des
phanérogames marines, très éloigné des signatures globales moyennes obtenues pour le sédiment
superficiel (Fig. III.45) et les consommateurs primaires (δ13C = -23,1 à -21,1‰) de la zone d'étude, va
dans le sens d'une participation négligeable de ce groupe de producteurs primaires au fonctionnement
les écosystèmes côtiers au large du Rhône. De même, bien que la signature globale des fragments de
Chlorophycées recueillis sur la zone d'étude n'aille pas à l'encontre de leur participation à la constitution
de la matière organique du sédiment superficiel, cette source d'apports dépend fortement de la direction
et de l'intensité des courants marins, ce qui la rend très sporadique. La biomasse de matière organique
qu'elle représente est ainsi, dans le temps, globalement négligeable par rapport à celles de la production
planctonique marine autochtone et des apports rhodaniens allochtones.
Les signatures très proches obtenues pour la MOP de l'eau de surface et le sédiment superficiel
semblent indiquer que la majeure partie du sédiment provient de la sédimentation de la matière en
suspension dans l'eau sus-jacente. Cependant, si l'augmentation de δ15N entre MOP de l'eau de surface
et sédiment superficiel s'explique par les modifications de signature en azote associées à l'assimilation
trophique et à la décomposition de la matière organique détritique par les organismes endogés
(Minagawa et Wada, 1984 ; Owens, 1987), le sédiment présente un δ13C légèrement inférieur à celui de
l'eau de surface qui implique une incorporation plus importante de matériel d'origine terrestre à son
niveau. L'origine de ce phénomène réside probablement dans le devenir particulier des débris végétaux
charriés par le fleuve. Par leur taille importante, ceux-ci sédimentent rapidement au large de
l'embouchure et leur contribution au pool de carbone organique présent dans la colonne d'eau est
négligeable alors qu'ils sont localement présents en quantités importantes au niveau du fond (Massé et
al., 1977). Si on se réfère à leur signature isotopique (δ13C = -28,4‰ ; δ15N. = 1,2‰), ces derniers
semblent ne contribuer que faiblement (du moins de façon directe) à la constitution du pool de matière
organique du sédiment de la zone d'étude (δ13C = -24,7‰; δ15N = 3,8‰). Etant donné leur fort taux en
cellulose et en lignine (Bouloubassi et al., 1997), ces fragments végétaux constituent un pool de
matériel réfractaire difficilement assimilable par les organismes hétérotrophes marins (Temore et al.,
1982). Ils ne sont directement exploitables que par les microorganismes benthiques (bactéries,
champignons) et la matière organique qu'ils contiennent n'est disponible pour la méiofaune et la
macrofaune benthique qu'après de longues périodes (plusieurs mois) de décomposition par l'activité
microbienne associée. Leur contribution à la constitution du pool de matière organique sédimentaire est

Réseaux trophiques et suivi de la MOP terrigène
226
donc différée et se fait de manière indirecte par le biais du compartiment microbien. Du fait des
augmentations de δ13C et, surtout, de δ15N associées à l'assimilation trophique et à la décomposition
bactérienne de la matière organique (De Niro et Epstein, 1978 ; Minagawa et Wada, 1984 ; Faure,
1986), la signature isotopique attendue pour le matériel végétal terrestre après ce type de médiation est
proche de celle de la MOP rhodanienne. Compte tenu de l'ensemble de ces considérations, les signatures
isotopiques du phytoplancton marin (δ13C = -22,4‰ et δ15N = 2,3‰) et de la MOP rhodanienne (δ13C =
-26,1‰ ; δ15N = 3,5‰) peuvent être respectivement retenues comme valeurs de références pour la
matière organique d'origine marine et pour le matériel d'origine terrestre disponibles à la base des
réseaux trophiques benthiques. Les apports de ces deux sources constituent, en biomasse, l'essentiel des
entrées de matière organique dans les écosystèmes à l'embouchure du Rhône (Cauwet et al., 1990) et
leurs signatures représentent celles de la matière de chaque origine directement assimilable par les
organismes hétérotrophes marins, qu'ils soient benthiques ou planctoniques. Les δ13C observés pour le
phytoplancton marin au large (-23,0 à -21,3‰) se situent dans la gamme de signatures communément
admises pour ce type de producteurs primaires (Dauby et Mosora, 1988 ; Riera et Richard, 1997 ;
Kiyashko et al., 1998 ; Herman et al., 2000 ; Takai et al., 2002). De même, les signatures en carbone
observées pour la MOP rhodanienne (-27,0 à -24,4‰) sont en accord avec celles relevées pour d'autres
fleuves (Incze et al., 1982 ; Lee, 2000 ; Riera et al., 2000).
Les apports des ces deux sources de matière organique varient de façon importante au cours de
l'année, tant du point de vue quantitatif que du point de vue qualitatif. L'essentiel du carbone et de
l'azote organiques apporté par le Rhône est déversé en mer en hiver et au printemps, au cours des
épisodes de crues pendant lesquels le flux de carbone organique particulaire peut dépasser 500 kg C s-1,
contre 0,3 kg C s-1 en période d'étiage (Cauwet, 1996). A ceci s'ajoutent des modifications saisonnières
de la nature du matériel d'origine continentale apporté par le fleuve. En effet, la signature isotopique de
la MOP rhodanienne varie au cours de l'année, avec notamment une diminution sensible du δ13C
pendant l'étiage (été) par rapport aux périodes de crues du fleuve (hiver-printemps). Une évolution
saisonnière semblable a déjà été observée pour d'autres fleuves européens, où elle s'explique par une
modification de la composition de la MOP fluviale, avec une proportion supérieure de phytoplancton
d'eau douce en été (Riera, 1995 ; Hellings et al., 1999). Pour le Rhône, les trois principaux pôles qui
contribuent à la matière organique particulaire sont les détritus arrachés aux sols par l'érosion, les
organismes et débris issus de la production aquatique et les rejets liés à l'activité humaine. Si on admet
que le troisième pôle est, en terme de carbone et d'azote, intermittent, localisé et globalement
négligeable (Cauwet, 1996), on peut considérer qu'il y a deux pôles principaux. La signature en carbone
obtenue pour la MOP rhodanienne (-26,1‰), proche de celle des débris végétaux terrestres récoltés au
large de l'embouchure (-27,7‰) est éloignée des signatures admises pour le phytoplancton d'eau douce
(δ13C = -32 à -44‰) (Mook et Tan, 1991). Ceci plaide en faveur d'une dominance de la matière
détritique d'origine terrestre dans la composition globale du pool de carbone organique véhiculé par le

Réseaux trophiques et suivi de la MOP terrigène
227
fleuve. Ce résultat est confirmé par le rapport C/N élevé (10,3) obtenu en moyenne pour la MOP
rhodanienne. En effet, les rapports C/N sont inférieurs à 6 pour le phytoplancton et la matière organique
animale, supérieurs à 15 dans les détritus issus des macrophytes et peuvent dépasser 40 pour les acides
humiques et certains sédiments anciens (Pourriot et Meybeck, 1995). Dans le cas du Rhône, la valeur
moyenne trouvée, en accord avec les C/N déjà reportés pour le fleuve par Cauwet (1996), dépasse la
valeur moyenne des fleuves et rivières mondiaux (8,5) et indique une forte contribution de matériel
organique détritique d'origine terrestre (Meybeck, 1982). La composition de la MOP rhodanienne se
modifie cependant fortement entre périodes de crues et d'étiage (Tableau III.21). En période de crues,
lorsque l'apport de particules liées à l'érosion des sols est prépondérant, le rapport C/N est maximal et le
δ13C très proche des valeurs rencontrées pour l'humus terrestre (-25,8‰) (Dauby, 1989). En période
d'étiage, lorsque la production primaire aquatique du Rhône est forte (Cauwet, 1996), la proportion de
phytoplancton d'eau douce dans la MOP fluviale est plus importante, ce qui se traduit par un δ13C et un
rapport C/N plus faibles. Il est possible que l'augmentation du δ15N de la MOP rhodanienne à cette
période reflète l'existence d'une signature isotopique en azote élevée (δ15N > 6‰) pour le phytoplancton
du Rhône, hypothèse compatible avec les données disponibles sur les δ15N du phytoplancton d'eau
douce qui couvrent une large gamme de valeurs, entre 1 et 9‰ (Minagawa et Wada, 1984 ; Owens,
1987). Il est également possible que cette augmentation de δ15N traduise un vieillissement plus
important de la matière organique détritique charriée par le fleuve à cette période, les détritus arrachés
au sol lors des pluies de fin d'été ayant largement eu le temps, pendant la saison sèche, d'être dégradés
sur place avant d'être emportés par ruissellement.
Tableau III.21 – Signatures isotopiques (carbone et azote) et rapports C/Nmoyens (± erreur-type) des eaux rhodaniennes en période de crue et en périoded'étiage.
δ13C (‰) δ15N (‰) Corg. / Ntotal
Crue
Etiage
Total
-25,7 ± 0,3
-26,6 ± 0,02
-26,1 ± 0,2
2,7 ± 0,2
5,1 ± 0,2
3,5 ± 0,4
12,5 ± 0,1
7,1 ± 0,2
10,3 ± 0,2
Tout comme les apports rhodaniens, les apports liés à la production primaire marine varient
selon la saison considérée. Dans le golfe du Lion, la production primaire est maximale d'avril à juin,
période où elle dépasse 1,6 mg m-3 h-1, contre un maximum de 1 mg m-3 h-1 le reste du temps (Conan et
al., 1998). Au large du Rhône, ce schéma d'évolution saisonnière est respecté bien que les apports
fluviaux en nutriments maintiennent un taux de production primaire élevé toute l'année, avec des pics de
production primaire d'importances secondaires en fin d'hiver et en été (Blanc et al., 1969 ; Lefèvre et
al., 1997). A ces modifications quantitatives des apports en biomasse d'origine marine se surimposent

Réseaux trophiques et suivi de la MOP terrigène
228
des modifications d'ordre qualitatif. En effet, si le δ15N et le C/N du phytoplancton marin restent stables
quelle que soit la saison, on observe une modification significative de la signature en carbone entre la
période printemps-hiver et l'été, avec un δ13C plus négatif pendant la saison estivale (Tableau III.22).
Cette évolution saisonnière ne traduit pas forcément une modification fondamentale de la composition
de la MES des eaux marines. En effet, le fractionnement isotopique du carbone par le phytoplancton
marin dépend essentiellement de la température de l'eau de surface (Fontugne et Duplessy, 1981) qui, en
Méditerranée, varie beaucoup entre le début du printemps et l'été du fait de l'établissement de la
thermocline estivale (Younes, 2000). Ce phénomène pourrait expliquer les différences saisonnières de
δ13C observées au sein des communautés phytoplanctoniques marines au large du Rhône. Celles-ci sont
pourtant dominées à toutes les saisons par les diatomées et les dinoflagellés (Lefèvre et al., 1997) et
présentent une signature en azote et un C/N très stables. Cependant, la composition spécifique des
producteurs primaires en présence influence également le fractionnement isotopique (Fontugne et
Duplessy, 1981 ; Fry et Wainright, 1991) et il est possible que les différences de δ13C observées
reflètent de légères modifications saisonnières de la composition spécifique des assemblages
phytoplanctoniques présents au large du Rhône.
Tableau III.22 – Signatures isotopiques (carbone et azote) et rapports C/Nmoyens (± erreurs-types) des eaux marines au large (Frioul) en période de crue(printemps-hiver) et en période d'étiage (été).
δ13C (‰) δ15N (‰) Corg. / Ntotal
Crue
Etiage
Total
-22,0 ± 0,2
-23,0 ± 0,4
-22,4 ± 0,2
2,3 ± 0,1
2,4 ± 0,04
2,3 ± 0,1
6,6 ± 0,2
6,3 ± 0,2
6,5 ± 0,2
Origine des pools de matière organique au large du Rhône
Les signatures isotopiques de l'eau et du sédiment de la zone d'étude suggèrent une participation
majeure des apports en matériel d'origine terrestre à la constitution des pools de carbone organique de
ces deux compartiments. Les contributions estimées sur la base des δ13C obtenus indiquent que plus de
56% de la matière organique disponible pour les consommateurs primaires au large du Rhône serait
d'origine terrestre. La validité de ces calculs dépend essentiellement de l'efficacité du δ13C à différencier
le matériel allochtone (apports du fleuve) de la matière organique d'origine marine au large du Rhône.
En ce qui concerne l'eau de surface, le δ13C global obtenu correspond à un mélange entre les signatures
du phytoplancton, des bactéries et du matériel détritique autochtones d'une part, et celles du
phytoplancton et du matériel détritique allochtone (apporté par le Rhône), de l'autre. Si la signature en
carbone globale des deux composantes terrigènes de ce mélange peut être estimée par le δ13C de la MOP
rhodanienne (-26,1‰), celle des composantes autochtones ne correspond pas forcément à celle obtenue

Réseaux trophiques et suivi de la MOP terrigène
229
pour la MOP strictement marine (-22,4‰). En effet, la signature des microorganismes marins
(phytoplancton, bactéries autotrophes et hétérotrophes) dépend de celle du carbone exploité par ces
organismes pour leur croissance. Ceux-ci peuvent utiliser aussi bien le CID (carbone inorganique
dissous) et le COD (carbone organique dissous) d'origine marine ou terrestre. Certains travaux en zones
estuariennes ont montré que la signature en carbone des producteurs primaires et des bactéries
hétérotrophes marins est susceptible de varier fortement selon l'origine (terrestre ou marine) du CID ou
du COD assimilé, avec des δ13C très négatifs (δ13C = -26,3‰) en cas d'exploitation du carbone dissous
d'origine continentale (Bouillon et al., 2000 ; Rolff et Elmgren, 2000). Compte tenu de l'importance
élevée des apports du Rhône en matériel dissous par rapport aux concentrations relevées en
Méditerranée, il est fort probable que ce type de phénomène se produise au large de l'embouchure du
fleuve, où la majorité du COD et du CID disponibles sont d'origine terrestre (Cauwet et al., 1997). Ceci
n'a cependant pas une grande incidence sur les conclusions des équations de mélange utilisées pour
estimer la part de matière organique d'origine "terrestre" et d'origine "marine" dans la matière en
suspension dans l'eau de surface de la zone d'étude dans la mesure où, selon qu'ils utilisent le carbone
dissous terrestre ou marin, la signature en carbone des microorganismes marins du panache va osciller
entre la signature de la MOP rhodanienne et celle du phytoplancton marin. Cependant, l'efficacité du
δ13C à différencier le matériel allochtone (apports du fleuve) de la matière organique d'origine marine
dans l'eau de surface récoltée au large du Rhône peut être estimée par l'étude de différents paramètres
(T°, S‰, C/N) mesurés sur la zone d'étude. Les δ13C obtenus pour la MOP de l'eau de surface sont en
accord avec les températures et les salinités relevées au large du Rhône en période de crues (printemps-
hiver) et d'étiage (été) (Tableau III.23). Les signatures en carbone les plus négatives, toujours associées
aux salinités les plus faibles, sont associées aux températures les plus basses au printemps-hiver et aux
températures les plus élevées en été, suivant ainsi l'évolution thermique des eaux rhodaniennes, plus
froides en hiver et plus chaudes en été que les eaux marines du golfe du Lion (Younes, 2000).
Tableau III.23 – Signatures en carbone, température et salinité des eaux rhodaniennes et marinescomparées à celles de l'eau de surface à 0-20 m, 30-50 m et 70-100 m de profondeur au large du Rhône,en période de crue et en période d'étiage.
δ13C (‰) T (°C) Salinité (ppt)
Crue Etiage Crue Etiage Crue Etiage
Rhône -25,52 -26,62 11,7 20,2 0,0 0,0Mer (Frioul) -22,03 -23,01 15,1 16, 3 38,0 37,8
Zone d'étude0-20 m (10 m) -25,34 -22,89 13,4 18,2 6,47 37,430-50 m (30 m) -25,64 -24,87 12,7 19,3 5,72 35,970-100 m (90 m) -23,71 -22,49 14,8 17, 9 29,80 37,1

Réseaux trophiques et suivi de la MOP terrigène
230
Il semblerait donc que le δ13C soit un bon indicateur de l'origine (terrestre/marine) des pools de
matière organique présents dans l'eau de surface au large du Rhône et que, sur l'ensemble de la zone
d'étude, la signature en carbone obtenue pour la MOP de l'eau de surface résulte plutôt d'un mélange de
MOP détritique d'origine terrestre et de phytoplancton marin que d'une production primaire in situ
essentiellement basée sur l'exploitation du carbone dissous d'origine continentale. Ceci est confirmé par
le rapport C/N élevé (7,8) de ce compartiment qui indique une présence non négligeable de matériel
détritique.
Pour le sédiment, la situation est encore plus complexe que pour l'eau de surface car de
nombreux facteurs peuvent contribuer à modifier la signature isotopique de la matière sédimentée avant
ou après son arrivée sur le fond. En effet, si la présence d'organismes hétérotrophes (bactéries,
champignons, méiofaune, restes de zooplancton) au sein des échantillons de sédiment analysés a été
prise en compte lors du calcul des contributions des sources terrestres et marines à sa composition,
d'autres facteurs sont susceptibles de modifier le δ13C de la matière organique, dont le principal est le
passage dans le tractus digestif des organismes planctoniques et/ou benthiques sans assimilation.
Plusieurs études portant sur la signature en carbone des pelotes fécales du zooplancton, ont démontré
l'existence d'une différence significative de δ13C entre le matériel assimilé (muscle) et le matériel rejeté
(fèces) par certains copépodes (Klein Breteler et al., 2002) et mysidacés (Gorokhova et Hansson, 1999)
planctoniques. La signature en carbone des pelotes fécales est, selon l'espèce et la source de nourriture
considérées, tantôt enrichie, tantôt appauvrie en 13C par rapport au matériel ingéré, ce qui introduit une
source de variation supplémentaire dans la signature isotopique de la MOP marine qui sédimente. De
même, la faune benthique est susceptible de modifier le signal des sources de matière organique au
niveau du sédiment par le biais de l'ingestion et de la défécation. Ceci a été démontré pour plusieurs
espèces de polychètes dépositivores (Thomas et Blair, 2002). La signature en carbone des fèces diffère
de celle de la nourriture ingérée et dépend essentiellement de l'espèce considérée. L'ensemble de ces
facteurs peut entraîner une modification conséquente de la signature de la matière organique des deux
origines (marine et terrestre) avant et après son incorporation au sein du sédiment. Les résultats des
équations de mélange obtenus sur la base des δ13C du phytoplancton marin, de la MOP rhodanienne et
du sédiment doivent donc être considérés avec précaution. Selon ces calculs, la contribution minimale
de la MOP rhodanienne à la constitution du pool de carbone organique du sédiment superficiel serait de
62% et les apports terrestres pourraient constituer jusqu'à 89% du pool de matière organique présent
dans ce compartiment. Ces valeurs sont en accord avec les fourchettes déjà obtenues sur la zone par
Bouloubassi et al. (1997), grâce à l'utilisation de différents marqueurs moléculaires de la matière
organique terrestre. Selon ces auteurs, la fraction du sédiment d'origine planctonique ne dépasserait pas
30 % au large du Rhône et, au niveau du prodelta du fleuve, le pourcentage de carbone d'origine
terrestre serait supérieur à 50%. Si l'on considère le fait que 30 à 45 % du sédiment apporté par le Rhône
est déposé au niveau du prodelta (Zuo et al., 1997), ce qui représente un taux d'accumulation de matériel

Réseaux trophiques et suivi de la MOP terrigène
231
terrestre au niveau du sédiment de 1,4. 106 t an-1 (Bouloubassi et al., 1997), ces estimations sont tout à
fait plausibles. Il semblerait donc que la modification de la signature du sédiment liée à l'activité de la
faune benthique soit réduite sur la zone d'étude ou que les déviations de δ13C associées à différents
phénomènes (défécation, décomposition bactérienne, assimilation) s'annulent mutuellement.
Variations spatio-temporelles des apports terrestres et marins au niveau de la zone d'étude
Les modifications saisonnières de la signature isotopique de la MOP rhodanienne et du
phytoplancton marin se répercutent sur celles de la MOP de l'eau de surface et du sédiment de la zone
d'étude qui varient suivant la saison, mais également suivant la profondeur. Ceci suggère l'existence
d'une double variabilité spatio-temporelle, quantitative et qualitative, des apports de matière organique
marine et continentale au niveau des écosystèmes benthiques situés au large du Rhône.
L'analyse des variations de la signature isotopique de la MOP de l'eau de surface permet de
reconstituer en partie la dynamique spatio-temporelle des apports de chaque source au large du fleuve.
En effet, la composition de la matière en suspension dans les eaux de surface à l'embouchure du Rhône
peut se modifier complètement en l'espace de quelques heures sous la double influence des apports du
fleuve et des courants (Younes, 2000). Les signatures isotopiques obtenues à chaque saison pour la
MOP de l'eau de surface représentent donc autant d'instantanés de la position du panache du fleuve au-
dessus de la zone étudiée. De fait, les signatures isotopiques en carbone et les rapports C/N observés
pour ce compartiment indiquent que la composition du pool présent dans l'eau de surface réagit
fortement aux modifications des apports des deux sources (Tableau III.24). La MOP présente dans l'eau
de surface sur la zone d'étude est ainsi dominée tantôt par la matière organique d'origine continentale,
tantôt par le phytoplancton marin.
Tableau III.24 – Signatures isotopiques en carbone et rapports C/N des eaux rhodaniennes etmarines comparés à ceux de la MOP de l'eau de surface et du sédiment superficiel à 0-20 m, 30-50 m et 70-100 m de profondeur au large du Rhône, en période de crue et en période d'étiage.
δ13C (‰) Corg. / Ntotal
Crue Etiage Total Crue Etiage Total
MOP Rhône -25,5 -26,6 -26,1 12,5 7,1 10,3phytoplancton marin -22,0 -23,0 -22,4 6,6 6,3 6,5
MOP eau de surface -24,6 -23,4 -24,4 8,1 6,5 7,8
Sédiment superficiel0-20 m (10 m) -24,5 -25,5 -24,9 11,9 12,3 12,230-50 m (30 m) -24,7 -25,4 -24,9 11,0 10,6 10,970-100 m (90 m) -24,1 -24,5 -24,2 10,9 10,1 10,7

Réseaux trophiques et suivi de la MOP terrigène
232
En période de crue, l'influence de la MOP rhodanienne domine. Elle s'étend sur l'ensemble de la
zone d'étude, avec un maximum pour l'eau de surface à 30-50 m (δ13C = -25,6‰), une situation
intermédiaire à 0-20 m (δ13C = -25,3‰) et un minimum pour l'eau de surface à 70-100 m (δ13C = -
23,7‰). En période d'étiage, elle est réduite et sensible uniquement au niveau de l'eau de surface à 30-
50 m (δ13C = -24,9‰). Ainsi, l'importance des apports en matériel d'origine terrestre diffère selon la
tranche bathymétrique considérée. Elle est maximale à 30-50 m, profondeur où la MOP de l'eau de
surface comprend, quelle que soit la saison, une part non négligeable de MOP terrigène. Elle est réduite
à 70-100m, où l'influence des apports continentaux n'est sensible que pendant les crues, et intermédiaire
à 0-20 m, profondeur où les apports terrestres prédominent en période de crue mais sont réduits pendant
l'étiage. Ces observations sont en accord avec les données disponibles sur l'extension du panache
rhodanien à proximité de l'embouchure du fleuve. Dans la majorité des cas (> 60% du temps), le
panache est initialement dévié vers l'Est de l'embouchure et s'étend au niveau de la zone d'étude au-
dessus des fonds de 20 à 70 m de profondeur avant d'être (ou non) dévié vers l'Ouest plus au large
(Demarcq, 1985). En cas de forts vents de Sud-Est, il est poussé à la côte et recouvre les fonds de la
zone d'étude entre 0 et 50 m de profondeur (Denis et al., 2001). Les épisodes de Sud-Est étant
essentiellement observés en fin d'hiver et au début du printemps, l'influence des apports rhodaniens est
surtout sensible à 0-20 m en périodes de crues du fleuve. De même, la déviation vers l'Ouest du panache
rhodanien par le courant liguro-provençal n'est contrebalancée par la force du courant Nord/Ouest-
Sud/Est de l'écoulement rhodanien qu'en cas de très fort débit du fleuve (Broche et al., 1998), ce qui
explique l'influence réduite des apports rhodaniens sur la MOP de l'eau de surface à 70-100 m et ce,
même en période de crues du fleuve.
Les variations spatio-temporelles de la position du panache du fleuve au-dessus de la zone
étudiée ont un impact primordial sur la constitution du sédiment à chaque profondeur. Du fait de la
hauteur de matériel échantillonnée (2-4 cm), les signatures obtenues pour le sédiment superficiel
fournissent une image intégrée des apports en matière organique au niveau du fond sur une échelle de
temps annuelle, voire pluriannuelle selon le taux de sédimentation. Ceci est confirmé par les rapports
C/N obtenus pour le sédiment superficiel au large de l'embouchure qui varient de 10,1 à 12,3. En effet,
le rapport C/N de la matière sédimentée varie en général de 8 à 20 en milieu marin et augmente
rapidement avec la profondeur et l'ancienneté des sédiments (Berner, 1971). Le matériel sédimentaire
échantillonné ici correspond donc à un sédiment relativement récent mais déjà en cours de
vieillissement. Parce qu'elle intègre les variations de signatures liées aux dépôts de sédiment antérieurs à
la période échantillonnée, la composition de la matière organique dans le sédiment est relativement
stable d'une saison à l'autre et très proche, quelle que soit la période et/ou la profondeur, de celle
observée en période de crues pour la MOP rhodanienne et l'eau de surface (Tableau III.24). Ceci
confirme l'importance primordiale des apports en MOP organique d'origine continentale associés aux
crues rhodaniennes dans la formation et le maintien des structures sédimentaires du prodelta du fleuve.

Réseaux trophiques et suivi de la MOP terrigène
233
L'essentiel des apports solides du Rhône est déchargé en mer pendant les courtes périodes de crues
(Cauwet, 1996) et sédimente au niveau de son prodelta où le taux d'accumulation atteint par endroit 300
à 500 kg m-2 a-1 (Bouloubassi et al., 1997). Si l'on compare ce chiffre à celui de la production primaire
marine au large du fleuve, estimée à 0,159 kg m-2 a-1 (Tusseau-Vuillemain, 1998), on comprend
aisément la dominance de la signature terrestre dans le pool de matière organique des sédiments de la
zone d'étude. L'utilisation des équations de mélange basées sur les δ13C du sédiment et des sources
marines et terrestres indiquent que l'incorporation de la MOP rhodanienne au sein des sédiments est
maximale (68-95%) à 0-50 m de profondeur et réduite plus profondément. Ceci confirme les données
préliminaires déjà disponibles sur la zone, selon lesquelles l'essentiel de la MOP rhodanienne serait
déposé au niveau du prodelta du fleuve au-dessus de l'isobathe 70 m, avec pour résultat des
pourcentages de carbone d'origine terrestre dans le sédiment de 55-80% à 50 m de profondeur et de 30-
65% à 90 m (Bouloubassi et al., 1997). Cette dernière valeur est inférieure au pourcentage de carbone
d'origine terrestre (50-77%) calculé pour le sédiment à 70-100 m au cours de la présente étude. Il est
possible que l'estimation obtenue à cette profondeur soit biaisée par les modifications du δ13C du
sédiment liées à l'activité biologique de la méiofaune endogée, ce qui expliquerait l'image différente
obtenue pour la MOP de l'eau de surface à cette profondeur, qui est dominée par le phytoplancton
marin. Cependant, dans la zone côtière étudiée, le forçage des vents en surface et les remontées d'eaux
profondes qui lui sont associées ainsi que l'écoulement en profondeur des eaux chargées en matières en
suspension provenant du Rhône sont à l'origine de structures benthiques éphémères de type néphéloïde
(Naudin et al., 1992 ; Naudin et Cauwet, 1997) qui s'écoulent en profondeur et enrichissent l'ensemble
du plateau continental en matière organique d'origine côtière (Chassefiere, 1990). L'enrichissement
ponctuel en matériel terrigène du sédiment des fonds de 70-100 m de profondeur par le dépôt massif de
matériel associé à ce type de structures expliquerait en grande partie le schéma observé. Cette hypothèse
est d'autant plus probable que la présence du néphéloïde benthique rhodanien, qui prend sa source dès le
chenal fluviatile et entraîne avec lui une charge importante de particules d'origine terrestre, a déjà été
signalée au niveau de la zone d'étude sur les fonds de 50 à 90 m de profondeur (Naudin et al., 1992).
III.5.2 - RESEAUX TROPHIQUES BENTHIQUES
L'utilisation des isotopes stable pour déceler l'incorporation éventuelle de matière organique
d'origine continentale dans les réseaux trophiques marins benthiques situés au large de l'embouchure des
fleuves a été validée dans plusieurs zones estuariennes (Incze et al., 1982 ; Simenstad et Wissmar, 1985
; Riera, 1995 ; Riera et Richard, 1996, 1997). Cependant, les études basées comme ici sur l'analyse
simultanée d'au moins deux éléments, seules véritablement fiables dans ce type de système à sources de
matière organique multiples, sont rares (Camusso et al., 1998 ; Camusso et al., 1999 ; Lee, 2000) et le
plus souvent limitées à un groupe d'invertébrés (Kiyashko et al., 1998 ; Riera et al., 2002) voire à une
seule espèce (Riera et al., 2000). Le présent travail, couvrant, à plusieurs profondeurs, l'ensemble de la

Réseaux trophiques et suivi de la MOP terrigène
234
chaîne trophique depuis les producteurs primaires jusqu'aux poissons, constitue donc à l'heure actuelle
l'une des études les plus détaillées du transfert de la MOP terrigène au sein des écosystèmes marins
benthiques sous influence continentale. L'utilisation des isotopes stables du carbone et de l'azote a
permis de reconstituer les réseaux trophiques benthiques au large du Rhône et de démontrer que la MOP
d'origine terrestre est exploitée, à différentes profondeurs, par des organismes de position trophique et
de modes d'alimentation différents.
Reconstitution des réseaux trophiques
L'utilisation de l'outil isotopique pour la reconstruction des réseaux trophiques aquatiques a été
validée pour de nombreux systèmes, qu'ils soient lacustres (Hesslein et al., 1991 ; Kiriluk et al., 1995 ;
Vander Zanden et al., 1999 ; Harvey et Kitchell, 2000) ou marins (Jennings et al., 1997 ; Paterson et
Whitfield, 1997 ; Kaehler et al., 2000 ; Sheaves et Molony, 2000). Dans chaque cas, la fiabilité de la
méthode repose sur la capacité des signatures isotopiques obtenues à décrire les relations trophiques
entre les organismes présents dans le(s) milieu(x) étudié(s). Cette fiabilité est rarement remise en
question bien que l'application consciencieuse de la méthode implique un certain nombre de précautions
d'emploi dont le non respect constitue une source d'erreur importante dans la reconstruction des réseaux
trophiques (Vander Zanden et Rasmussen, 2001). La capacité de l'outil isotopique à décrire de façon
fiable la structure des réseaux trophiques étudiés repose essentiellement sur deux postulats : (1)
l'enrichissement isotopique de la proie au prédateur est connu dans le milieu considéré et, (2) les δ15N
obtenus pour les organismes impliqués permettent de déterminer leurs niveaux trophiques respectifs et
de les positionner le long des chaînes alimentaires. En ce qui concerne le premier de ces deux points,
peu d'études (Harrigan et al., 1989 ; Kaehler et al., 2000) prennent en compte la variabilité
interspécifique du fractionnement isotopique, pourtant soulignée par plusieurs auteurs (De Niro et
Epstein, 1978, 1981 ; Minagawa et Wada, 1984 ; Vander Zanden et Rasmussen, 2001). La plupart des
travaux réalisés, même récents, repose sur l'utilisation d'un enrichissement trophique moyen de 0-1‰
pour le carbone et de 2-3‰ pour l'azote (Jennings et al., 1997 ; Harvey et Kitchell, 2000 ; Pinnegar et
Polunin, 2000 ; Sheaves et Molony, 2000 ; Chong et al., 2001). Ces valeurs correspondent aux
moyennes obtenues par divers auteurs dans les années 80 sur la base d'un nombre élevé d'organismes
issus d'écosystèmes variés, aquatiques ou non (De Niro et Epstein, 1978, 1981 ; Minagawa et Wada,
1984 ; Peterson et Fry, 1987). Malgré l'importance des écart-types qui leur sont associés (>1‰),
l'utilisation de ces moyennes en tant que valeurs absolues de référence pour les enrichissement
trophiques en δ13C et en δ15N dans les écosystèmes s'est peu à peu généralisée et est rarement remise en
cause. Or plusieurs études, réalisées sur le terrain et en laboratoire, ont souligné les limites de l'emploi
systématique de ces valeurs moyennes pour la description des réseaux trophiques aquatiques (Paterson
et Whitfield, 1997 ; Vander Zanden et Rasmussen, 1999, 2001). En effet, le fractionnement isotopique
varie de façon importante suivant l'éthologie alimentaire des organismes, avec un enrichissement

Réseaux trophiques et suivi de la MOP terrigène
235
trophique en δ15N beaucoup plus variable et nettement inférieur chez les herbivores (∆ δ15N = 2,5±2,5
‰) que chez les carnivores (∆ δ15N = 3,2±0,4 ‰) (Vander Zanden et Rasmussen, 1999, 2001). Ce
phénomène peut entraîner des erreurs non négligeables dans l'estimation du niveau trophique des
prédateurs lorsque ces différences ne sont pas prises en compte et qu'un enrichissement isotopique
constant est utilisé pour l'ensemble des transferts trophiques impliqués (Vander Zanden et Rasmussen,
2001). De même, la sur- ou la sous-estimation des fractionnements isotopiques en carbone peut
conduire à des erreurs importantes quant à la nature des composantes du régime alimentaire de chaque
consommateur (Paterson et Whitfield, 1997). Au cours de la présente étude, le calcul des
enrichissements en δ15N et en δ13C particuliers aux communautés benthiques échantillonnées a permis
de limiter le biais lié à ce type d'erreur. Les enrichissements moyens en δ15N estimés pour les herbivores
(+2,6‰) et les carnivores (+3,2‰) capturés au large du Rhône sont en accord avec les valeurs utilisées
dans la littérature bien que légèrement différents (De Niro et Epstein, 1981 ; Minagawa et Wada, 1984 ;
Vander Zanden et Rasmussen, 2001). L'enrichissement trophique moyen en δ13C obtenu (+1,8‰) se
situe dans la gamme des valeurs proposées par Vander Zanden et al. (2001) pour les réseaux trophiques
aquatiques mais est relativement élevé par rapport à la valeur de 0-1‰ généralement admise pour cet
élément (De Niro et Epstein, 1978). L'utilisation des enrichissements estimés pour la zone d'étude en
lieu et place des valeurs communément utilisées a ainsi permis une meilleure description des réseaux
trophiques étudiés.
Pour ce qui est de l'estimation des positions trophiques, l'étude des δ15N permet le plus souvent
à elle seule de positionner les organismes entre eux et d'obtenir ainsi la structure des réseaux trophiques
(Vander Zanden et Rasmussen, 1996 ; Jennings et al., 1997 ; Vander Zanden et al., 1997). Cependant,
lorsque les sources potentielles de matière organique sont multiples et de signatures en azote différentes,
seule une approche pluri-élémentaire permet de décrire de façon précise la structure des réseaux
trophiques (Harrigan et al., 1989). L'estimation fine du niveau trophique des organismes nécessite alors
une étude préalable de(s) source(s) de matière organique exploitée(s) dans chaque cas, ceci afin de
préciser quelle signature en azote utiliser comme référence pour les calculs (Lee, 2000). La prise en
compte de ce facteur de variation n'est pas systématique, même dans les études récentes (Sauriau et
Kang, 2000 ; Griffin et Valiela, 2001 ; Nisbet et al., 2002). Ceci peut pourtant conduire à des erreurs
non négligeables dans l'estimation du niveau trophique des organismes. Au large du Rhône, où les δ15N
des deux principales sources de matière organique (phytoplancton marin et MOP rhodanienne) diffèrent
de plus de 1‰, le niveau trophique estimé pour les cinq poissons plats étudiés varie ainsi d'environ 0,35
suivant le δ15N de référence utilisé (Tableau III.25). De plus, l'erreur commise en ne choisissant pas la
source de matière organique adéquate pour le calcul des niveaux trophiques peut aboutir à des
conclusions erronées quant aux stratégies alimentaires des espèces.

Réseaux trophiques et suivi de la MOP terrigène
236
Ainsi, sur la base du δ15N du phytoplancton marin, B. luteum présente un niveau trophique
inférieur à celui de S. solea et proche de celui de C. linguatula, alors qu'au large du Rhône cette espèce
s'alimente en fait à un niveau trophique similaire à celui de la sole et nettement inférieur à celui de la
cithare. Cet exemple montre toute l'importance d'une étude basée sur l'analyse des rapports isotopiques
de deux éléments (ou plus) pour la description des réseaux trophiques benthiques à l'interface mer-
continent et la nécessité de déterminer l'origine de la matière organique ingérée par les organismes pour
l'estimation de leur niveau trophique.
Tableau III.25– Niveaux trophiques moyens (NT) des poissons obtenus en utilisant le δ15N, soit duphytoplancton marin, soit de la MOP rhodanienne, soit d'un mélange équilibré des deux sources, commesignature de référence des producteurs primaires dans les calculs (c. f. chapitre matériel et méthodes). Danschaque cas, les valeurs en gras indiquent le niveau trophique le plus probable au large du Rhône.
NT phytoplancton MOP rhodanienne mixte
Solea solea 3,83 3,48 3,66Solea impar 3,88 * 3,52 3,70Buglossidium luteum 3,69 3,33 3,51Arnoglossus laterna 3,40 3,05 3,23Citharus linguatula 3,68 3,32 3,50
* Calcul ne tenant pas compte de la part de matière organique issue de la production microphytobenthiquesusceptible d'être transférée vers les adultes de l'espèce via les bivalves adultes (pour les détails, voir plus loin)
En ce qui concerne la structure des réseaux trophiques proposée au large du Rhône, l'obtention
d'une large gamme de signatures isotopiques pour chaque "échelon trophique" (surtout pour l'azote) ne
remet pas en question le schéma proposé. En effet, ce phénomène s'explique par l'interaction de
différents facteurs, dont le principal est la variabilité interspécifique du fractionnement isotopique. Chez
les organismes hétérotrophes, l'enrichissement trophique en azote dépend essentiellement du prédateur
(De Niro et Epstein, 1978). Le fractionnement isotopique est très variable chez les herbivores (Vander
Zanden et Rasmussen, 2001), ce qui explique en partie la large gamme de signatures obtenue pour cet
échelon au sein de chacun des réseaux trophiques étudiés. Chez les carnivores, l'enrichissement
trophique est plus stable (Vander Zanden et Rasmussen, 2001) mais, étant donné la diversité supérieure
des proies potentiellement ingérées, la variabilité des signatures isotopiques attendues augmente (du fait
du nombre supérieur de signatures obtenues par combinaisons de celles des proies). Ce phénomène est
accentué par le caractère omnivore de l'alimentation de certains invertébrés étudiés (Tableau III.26).
Dans l'ensemble, le niveau trophique estimé pour chaque taxon est en accord avec les données
disponibles sur son alimentation. Cependant, certaines divergences existent, notamment en ce qui
concerne les bivalves.

Réseaux trophiques et suivi de la MOP terrigène
237
Tableau III.26 – Gamme de δ15N obtenue et niveaux trophiques (NT) estimés pour les organismes hétérotrophesrécoltés au large du Rhône : comparaison avec les données connues sur le régime alimentaire et le niveau trophiquedes animaux. "MOP" = matière organique particulaire; "C I", "C II", "C III"… = consommateur primaire, secondaire,tertiaire (etc).
δ15N (‰) NT estimé Alimentation (référence) : position trophique
Polychètes suspensivores 6,1 - 7,2 C I - C II MOP, phyto et zooplancton (4) : C I – C IIPolychètes dépositivores 5,8 - 7,0 C I Sédiment et microorganismes associés (4) : C I
Polychètes carnivores 8,6 - 9,2 C II Méiofaune endogée et épigée (4) : C II
Copépodes 4,8 C I MOP, phytoplancton (8) : C I
Mysidacés (< 2 cm) 8,7 - 9,6 C I - C II phyto et zooplancton, MOP, méiofaune (3) :C I - C II
Cumacés 4,2 - 4,4 C I MOP, phytoplancton (8) : C I
Amphipodes (< 5 mm) 4,7 - 5,5 C I MOP, phytoplancton (8) : C I
Crangonidae (< 2 cm) 5,4 - 8,3 C I - C II Méiofaune (8) : C II
Alpheidae (< 2 cm) 8,3 - 8,4 C II Méiofaune (8) : C II
Anomoures (< 2 cm) 7,3 C II ? Micro et macroalgues, méiofaune, cadavres (2) : C I -C IIIBrachyoures (< 2 cm) 6,2 - 7,0 C I - CII Omnivores (9) : C I - C II
Bivalves 5,4 - 7,9 C I - C II ? Microalgues marines et MOP (9) : C I
Gastéropodes (< 2 cm) 9,7 - 9,8 C II (limite CIII) Méiofaune, larves, cadavres (8) : > C II
Ophiures 7,8 C II Méiofaune, larves (8) : C II
Holothuries (Thyone spp.) 6,4 C II (limite CI) phyto et zooplancton, MOP (7) : C I - CII
Poissons (gobiidés < 4 cm) 7,6 - 9,6 C II Méiofaune (1) (6) : C II
Solea solea 10,1 - 11,1 C III NT moyen de l'espèce = 3,28±0,44 (5)
Solea impar 10,7 - 11,1 C III NT moyen de l'espèce = 3,23±0,41 (5)
Buglossidium luteum 8,8 - 11,1 CII - C III NT moyen de l'espèce = 3,39±0,43 (5)
Arnoglossus laterna 8,5 - 10,1 CII - C III NT moyen de l'espèce = 3,59±0,54 (5)
Citharus linguatula 9,1 - 11,6 CII - C IV NT moyen de l'espèce = 3,97±0,65 (5)
Références bibliographiques : (1) Aarnio et Bonsdorff (1993), (2) Barnes (1965), (3) Dauby (1995), (4) Fauchald et Jumars (1979), (5) Froese
et Pauly (2000), (6) Hamerlynck et Cattrijsse (1994), (7) Massin (1982), (8) Pérès (1980), (9) Riera et al. (1999).
La différence importante de δ15N observée entre les juvéniles et les adultes de cette catégorie de
proie est surprenante, les bivalves étant largement décrits comme des organismes filtreurs herbivores
(Riera et Richard, 1996 ; Sauriau et Kang, 2000 ; Raikow et Hamilton, 2001) et souvent cités comme
consommateurs primaires de référence (Cabana et Rasmussen, 1996 ; Vander Zanden et al., 1997).
Cependant, ce résultat rejoint celui de Minagawa et al. (1984) qui expliquent le changement de niveau
trophique observé chez les moules de Mer de Chine au cours des premiers mois de développement par
une modification du régime alimentaire avec la croissance. Les bivalves adultes étant capables d'ingérer
des particules de taille importante (jusqu'à 110 µm) (Newel et al., 1989), il est possible que cette

Réseaux trophiques et suivi de la MOP terrigène
238
modification du δ15N soit liée à l'ingestion, par les adultes analysés, de petits organismes hétérotrophes
planctoniques (Dupuy et al., 1999). Cependant, une ingestion importante de diatomées benthiques (de
δ15N et de δ13C nettement supérieurs à ceux du phytoplancton) peut également produire le type de
signature observée chez les bivalves adultes. Au large du Rhône, la production phytobenthique
contribue très faiblement à la production primaire totale (Bodoy et Plante-Cuny, 1980). Il est cependant
possible que les diatomées benthiques constituent une source de nourriture non négligeable pour
certains organismes capables de les sélectionner préférentiellement, surtout à 0-20 m où la production
microphytobenthique est maximale. Etant donné la profondeur de capture des bivalves adultes analysés
(0-20 m), cette hypothèse est vraisemblable, d'autant plus qu'une ingestion sélective de diatomées
benthiques a été démontrée chez les huîtres et les coques en Baie de Marennes-Oléron (Riera et Richard,
1996 ; Sauriau et Kang, 2000), où ces producteurs primaires représentent une part importante du régime
alimentaire des bivalves. Etant donné la signature en azote des diatomées benthiques (δ15N = 4,5‰), le
δ15N obtenu pour les adultes de bivalve refléterait l'ingestion d'un mélange de microphytes marines
dominé par ces dernières, ce qui est compatible avec la signature en carbone observée. L'ingestion d'une
quantité conséquente de microphytes benthiques expliquerait également les signatures isotopiques
particulières (δ13C élevés) obtenues pour les anomoures et les macroures Crangonidae à 0-20 m de
profondeur. Cette hypothèse a déjà été émise par Ansell et al. (1999) pour les individus de Crangon
crangon et Pagurus bernhardus capturés sur les fonds sableux à l'Ouest de l'Ecosse, où les quantités
élevées de grains de sable rencontrées dans les tubes digestifs de ces deux espèces aboutiraient à une
assimilation non négligeable des microorganismes benthiques épigés qui leur sont associés. La
production microphytobenthique représente donc potentiellement une part non négligeable de la matière
organique d'origine marine qui intègre les réseaux trophiques benthiques à 0-20 m de profondeur au
large du Rhône. Les fluctuations de δ15N et de δ13C liées à ce phénomène ainsi qu'à la variabilité
interspécifique du fractionnement isotopique et à l'omnivorie, expliquent les larges gammes de
signatures isotopiques obtenues par "échelon trophique" et justifient les structures de réseaux trophiques
proposées à chaque profondeur étudiée.
Incorporation de la MOP d'origine continentale au sein des réseaux trophiques
Consommateurs primaires
L'incorporation de la MOP apportée par le fleuve au sein des réseaux trophiques benthiques
concerne essentiellement les consommateurs primaires. Ceux-ci sont généralement considérés comme
étant des "omnivores opportunistes" (Haines et Montague, 1979) se nourrissant à partir d'un mélange de
microphytes benthiques, de détritus, de bactéries, de protozoaires, de champignons, mais aussi de petites
espèces méiobenthiques ou de formes larvaires (Kuipers et al., 1981), selon la disponibilité de ces
différentes catégories. Au large du Rhône, la majorité des consommateurs primaires échantillonnés

Réseaux trophiques et suivi de la MOP terrigène
239
semble cependant capable d'assimiler (si ce n'est d'ingérer) de façon sélective certaines des composantes
de la matière organique particulaire disponible dans le milieu. Ce phénomène est particulièrement
marqué chez les organismes filtreurs et détritivores (bivalves, copépodes, cumacés, amphipodes, etc).
Ceux-ci présentent une signature clairement "marine" (δ13C > -22‰), incompatible avec une
assimilation indifférenciée de la MOP disponible à proximité du fond, dans laquelle le matériel dérivé
des organismes marins autotrophes représente une fraction négligeable de la charge particulaire totale
(Naudin et al., 1992 ; Naudin et Cauwet, 1997 ; Denis, 1999). L'exploitation préférentielle des
microphytes marines (diatomées, phytoplancton) aux dépends des autres particules sestoniques
(bactéries, matière détritique, zooplancton) est largement documentée chez les bivalves (Newel et al.,
1989 ; Riera et Richard, 1996 ; Charles et al., 1999 ; Sauriau et Kang, 2000 ; Raikow et Hamilton, 2001)
où elle fait appel à différents mécanismes de "tri sélectif" intervenant pendant la filtration, la digestion
et l'assimilation des particules (Shumway et al., 1985). Des phénomènes similaires sont susceptibles de
se produire chez les copépodes harpacticoïdes, les cumacés et les amphipodes échantillonnés. En effet,
ces invertébrés méiobenthiques sont décrits comme des "epistrate-feeders" essentiellement herbivores,
se nourrissant à partir de phytoplancton, de diatomées et de bactéries présentes au niveau du fond
(Pérès, 1980), mais seraient capables d'une ingestion sélective des proies disponibles dans le milieu
(Buffan-Dubau et al., 1996 ; Dittel et al., 2000). En ce qui concerne les polychètes, l'ingestion des
particules semble moins sélective. Suivant la profondeur et la composition de l'eau de surface, les
espèces suspensivores se positionnent, tantôt en CI - CII du réseau trophique phytoplanctonique, tantôt
en CI de celui basé sur la MOP terrigène, ce qui est tout à fait compatible avec les données disponibles
sur leur régime alimentaire, constitué d'un mélange variable de particules sestoniques incluant
phytoplancton, zooplancton et matière détritique (Fauchald et Jumars, 1979). De même, les polychètes
dépositivores de surface et de sub-surface présentent une signature isotopique très proche de celle du
sédiment superficiel à partir duquel ils se nourrissent (Fauchald et Jumars, 1979). Ils semblent
cependant capables, à chaque profondeur, d'assimiler de façon sélective le matériel organique le plus
frais disponible dans leur environnement. La signature isotopique des polychètes dépositivores de
surface (qui se nourrissent dans les premiers cm du sédiment) reflète ainsi les modifications de
composition de la MOP sédimentée dominée, selon la profondeur, soit par la MOP rhodanienne, soit par
le phytoplancton marin. Cette assimilation préférentielle de matériel "frais" a déjà été reportée pour les
polychètes dépositivores pour qui les microorganismes présents en surface ou à l'intérieur du sédiment
constituent la principale source de matière organique (Fauchald et Jumars, 1979). Il est d'ailleurs
possible, pour cette raison, que les débris végétaux terrestres participent également de façon non
négligeable à l'alimentation de ces organismes via les bactéries hétérotrophes qui leurs sont associées.
Ceci expliquerait la signature en carbone relativement stable et toujours très négative obtenue pour les
espèces dépositivores de sub-surface qui se nourrissent plus profond dans le sédiment et dont
l'environnement est moins soumis aux modifications qualitatives de la MOP sédimentée.

Réseaux trophiques et suivi de la MOP terrigène
240
Du fait des phénomènes de sélection et des différences de preferenda alimentaires observés chez
les consommateurs primaires présents au large du Rhône, le degré d'exploitation de la matière organique
d'origine terrestre diffère suivant le groupe d'invertébrés considéré, ce qui a d'importantes conséquences
sur le degré d'incorporation de la MOP terrigène au sein des réseaux trophiques benthiques. Dès le
premier échelon trophique, deux grands groupes trophiques distincts s'individualisent, l'un basé sur
l'exploitation principale des sources de production phytoplanctoniques et rassemblant la majorité des
consommateurs primaires étudiés, l'autre dépendant majoritairement des sources de MOP terrestres
(MOP rhodanienne, sédiment et/ou seston de la zone d'étude) et incluant essentiellement les polychètes.
Les biomasses associées à ces deux groupes d'invertébrés et leurs participations respectives à
l'alimentation des prédateurs benthiques vont déterminer en grande partie l'importance de l'incorporation
de la MOP terrigène au sein des réseaux trophiques benthiques à chaque profondeur.
Omnivorie
Les consommateurs primaires "stricts" ne semblent cependant pas être les seuls organismes
impliqués dans l'incorporation de la matière organique au sein des réseaux trophiques benthiques situés
au large du Rhône. En effet, certains invertébrés, essentiellement décrits comme carnivores, présentent
des signatures isotopiques en carbone et en azote suggérant une ingestion opportuniste importante de
matériel planctonique, microphytobenthique ou de MOP détritique d'origine terrestre. C'est le cas
notamment des juvéniles de brachyoures et de macroures, qui changent de position trophique (CI ou
CII) et semblent capables d'assimiler de façon directe des quantités non négligeables de microphytes
marines ou de matière organique terrigène selon la profondeur. Cette observation confirme le caractère
omnivore de l'alimentation de ces deux groupes d'invertébrés, déjà suggéré par plusieurs auteurs (Ansell
et al., 1999 ; Riera et al., 1999). En effet, bien qu'en général décrits comme essentiellement carnivores
(Barnes, 1965 ; Mascaro et Seed, 2000 ; Oh et al., 2001), de nombreux brachyoures et la majorité des
Crangonidae présentent une forte adaptabilité trophique et peuvent, dans certains habitats, consommer
une part non négligeable de matériel végétal ou bactérien (Ansell et al., 1999). Parmi les invertébrés
benthiques échantillonnés, d'autres "carnivores" semblent également capables d'exploiter directement la
matière organique disponible à la base des écosystèmes benthiques (Tableau III.26). C'est le cas des
anomoures et des mysidacés, pour lesquels une ingestion non sélective de matériel particulaire vivant
(diatomées benthiques, phytoplancton, bactéries) ou détritique a été démontrée (Dauby, 1995 ; Ansell et
al., 1999). En l'absence de signatures isotopiques détaillées à chaque profondeur pour ces deux groupes
d'invertébrés, il est impossible de conclure sur leur participation effective à l'incorporation de la MOP
d'origine terrestre dans les réseaux trophiques au large du Rhône. Cependant, les signatures globales
obtenues plaident en faveur d'une exploitation majoritaire de matériel d'origine marine par ces
organismes.

Réseaux trophiques et suivi de la MOP terrigène
241
Adaptabilité trophique
Le degré d'exploitation de la MOP terrestre par les organismes benthiques diffère selon la
tranche bathymétrique considérée. Ce phénomène est directement lié aux modifications de la quantité de
matière organique d'origine terrestre présente à chaque profondeur dans la matière en suspension dans
l'eau ou au niveau du sédiment superficiel. L'incorporation de la MOP d'origine terrestre et son transfert
dans les réseaux trophiques benthiques sont ainsi maximums à 30-50 m, profondeur où les apports
terrigènes fournissent plus de 50% du carbone organique disponible au sein de ces deux compartiments
(Bouloubassi et al., 1997 ; présente étude). L'adaptabilité trophique diffère cependant selon les
organismes considérés et seuls certains groupes d'invertébrés benthiques semblent capables d'exploiter,
que ce soit de façon directe ou non, le surplus de matière organique lié aux apports accrus du Rhône à
cette profondeur. C'est le cas des juvéniles de bivalve et de brachyoures, des macroures (Crangonidae et
Alpheidae) et des polychètes dépositivores de surface, suspensivores et carnivores dont la signature
isotopique, minimale à 30-50 m et maximale à 70-100 m, reflète les modifications bathymétriques des
apports en MOP terrestre au large de l'embouchure. Des variations similaires de la signature isotopique
en fonction de l'origine (terrestre/marine) de la matière organique disponible dans le milieu ont été
observées chez divers invertébrés benthiques, qu'ils soient herbivores, carnivores ou omnivores
(Simenstad et Wissmar, 1985 ; Hsieh et al., 2000 ; Lee, 2000). Chez les mollusques, de nombreuses
études en zone estuarienne ont montré une diminution significative du δ13C avec celle de la salinité
(Incze et al., 1982 ; Riera, 1995 ; Riera et Richard, 1996, 1997 ; Hsieh et al., 2000). Les huîtres
répondent aux variations de signature isotopique de la MOP en suspension avec un délai de 1 à 2 mois
et semblent capables d'exploiter directement la MOP terrestre détritique apportée par les fleuves (Riera
et Richard, 1997). Pour ce qui est des crustacés, les études sont plus rares. Cependant, quelques travaux
démontrent la capacité de ce groupe d'organismes à tirer parti des apports fluviaux en MOP terrestre
(Haines et Montague, 1979 ; Schlacher et Wooldridge, 1996 ; Paterson et Whitfield, 1997 ; Chong et al.,
2001). Les macroures du genre Penaeus répondent ainsi rapidement aux variations de la signature
isotopique de la MOP disponible à la base des réseaux alimentaires et semblent capables d'exploiter la
MOP terrigène apportée par les fleuves, non seulement de façon indirecte (médiation par le biais de
petits invertébrés benthiques) mais également de façon directe (Riera et al., 2000). De même, les
signatures isotopiques obtenues pour certains brachyoures détritivores en zone estuarienne indiquent
une incorporation importante de matériel d'origine continentale dans la chair de ces organismes
provenant d'un mélange de MOP fluviale et de feuilles de végétaux terrestres (Lee, 2000). En ce qui
concerne les polychètes, les études isotopiques sont rares. Certains travaux ont cependant montré une
incorporation de matériel terrigène chez les polychètes dépositivores de surface et de sub-surface
(Kichuki et Wada, 1996) et une diminution significative du δ13C avec celle de la salinité chez Nereis
diversicolor (Nithart, 1995, 2000).

Réseaux trophiques et suivi de la MOP terrigène
242
Les résultats obtenus au large du Rhône pour l'ensemble de ces groupes d'invertébrés
complètent ceux des études préexistantes et confirment la capacité de nombreux organismes benthiques
marins à tirer parti de la MOP terrestre apportée par les fleuves. Certains des organismes benthiques
récoltés (holothuries, amphipodes, gastéropodes) semblent par contre peu ou pas influencés par les
apports terrestres, ce qui a d'importantes implications quant à leur biologie et leur compétitivité sur ces
fonds. Les différences interspécifiques et bathymétriques de signature isotopique mises en évidence au
sein du macrobenthos (et les différents degrés d'exploitation de la MOP rhodanienne qu'elles
impliquent) ont également d'importantes conséquences quant à l'influence potentielle des apports
rhodaniens en matière organique sur les populations de poissons plats présentes au large du delta du
fleuve. Ainsi, la connaissance précise des modalités de l'exploitation de la matière organique au sein des
communautés d'invertébrés benthiques constitue une étape préliminaire fondamentale dans la
compréhension du cheminement de la MOP terrigène depuis son arrivée dans la colonne d'eau jusqu'à
son intégration dans la chair des poissons démersaux présents au large du fleuve.
III.5.3 - POISSONS PLATS : ALIMENTATION, SIGNATURE ISOTOPIQUE ET EXPLOITATION DE
LA MOP TERRIGENE
Les études isotopiques de l'incorporation et du transfert de la MOP continentale au sein des
communautés marines se limitent généralement à l'analyse des premiers échelons de la chaîne trophique
et les travaux incluant les poissons sont peu nombreux (Deegan et Garritt, 1997 ; Kwak et Zedler, 1997 ;
Paterson et Whitfield, 1997). Ces études concluent le plus souvent à une influence réduite des apports
terrestres sur les communautés marines étudiées (Deegan et Garritt, 1997 ; Kwak et Zedler, 1997 ;
Camusso et al., 1999) et peu d'entre elles mettent en évidence un impact conséquent du matériel
terrigène allochtone sur les poissons échantillonnés (Lee, 2000 ; Sheaves et Molony, 2000). Au large du
Rhône, les signatures isotopiques obtenues indiquent une incorporation de matériel organique d'origine
terrestre dans la chair de chacune des cinq espèces de poissons plats étudiées. Cependant celle-ci varie
fortement d'une espèce à l'autre et n'est primordiale que pour S. solea dont le δ13C très négatif (-
19,95‰) indique une exploitation majoritaire de matériel d'origine terrestre. La détection de l'influence
des apports en MOP terrigène sur la croissance des quatre autres poissons n'a pu être obtenue que grâce
à l'étude détaillée des réseaux trophiques benthiques. Celle-ci a été facilitée par le nombre réduit de
sources de matière organique en présence au large du Rhône (2 à 3 suivant les profondeurs), inférieur à
celui habituellement recensé en zone estuarienne (Mann, 1982 ; Riera, 1995, 1998 ; Lee, 2000). Il est
donc possible que, dans certains des autres écosystèmes étudiés (Deegan et Garritt, 1997 ; Kwak et
Zedler, 1997), la multitude des sources potentielles de matière organique présentes à la base des réseaux
trophiques analysés brouille l'information fournie par l'utilisation des isotopes et que l'influence des
apports fluviaux sur certains poissons passe inaperçue. Il semblerait néanmoins, compte tenu des

Réseaux trophiques et suivi de la MOP terrigène
243
résultats de la présente étude et des données disponibles dans la littérature (Lee, 2000 ; Sheaves et
Molony, 2000), que le transfert important de MOP terrigène jusqu'aux poissons marins côtiers ou
estuariens soit un phénomène limité à un mode de vie (benthique) et un régime alimentaire (proies
dépositivores) particuliers. Parmi l'ensemble des facteurs susceptibles de déterminer l'intensité du
transfert de la MOP terrigène jusqu'aux poissons (mode de vie, habitat, alimentation), la composition du
régime alimentaire semble primordiale. En effet, des différences importantes de régime alimentaire
expliquent en général les différences de signatures isotopiques observées, que ce soit d'une espèce de
poisson à l'autre (Lee, 2000 ; Sheaves et Molony, 2000) ou entre habitats différents abritant la même
espèce (Dauby et Mosora, 1988 ; Camusso et al., 1998 ; Camusso et al., 1999). Les individus de S.
solea capturés au large du Pô (se nourrissant essentiellement à partir d'amphipodes) présentent ainsi, à
δ15N équivalent et pour une signature de MOP terrestre proche (δ13C = -27‰), une signature beaucoup
plus marine (δ13C = -18,5‰) que celle observée ici (Camusso et al., 1998 ; Camusso et al., 1999). De
même, au large du Rhône, les différences interspécifiques et bathymétriques de la nature des proies
ingérées expliquent les différences de signatures isotopiques observées d'un groupe de poissons à l'autre.
Analyse isotopique et étude des contenus stomacaux : deux approches complémentaires
Etant donné la complexité du régime alimentaire des cinq espèces étudiées, les modalités
précises du transfert de la matière organique de chaque origine (terrestre/marine) vers leurs populations
n'ont pu être élucidées, dans chaque cas, que grâce à l'étude combinée des contenus stomacaux et des
signatures isotopiques des poissons. La complémentarité des deux approches a déjà été soulignée par
plusieurs études (Harrigan et al., 1989 ; Pinnegar et Polunin, 2000). Cependant, ces travaux utilisent en
général des données alimentaires issues de la littérature (Pinnegar et Polunin, 2000) ou ne concernent
qu'une seule espèce (Harrigan et al., 1989). Le présent travail, basé sur l'analyse des compositions
isotopique musculaires et des contenus stomacaux des même poissons et confrontant les signatures
isotopiques et les régimes alimentaires détaillés des juvéniles et des adultes de cinq espèces, permet une
estimation fiable des apports respectifs des deux méthodes à l'étude de l'alimentation des poissons.
L'analyse des contenus stomacaux permet d'obtenir un degré de détail quant à la nature des proies
consommées non décelable par l'utilisation des isotopes. Ce phénomène, déjà souligné par Pinnegar et
Polunin (2000), est particulièrement bien illustré au large du Rhône où, par exemple, seule l'étude des
contenus stomacaux a permis de différencier l'alimentation des juvéniles de B. luteum (polychètes,
mollusques, crustacés) et de C. linguatula (poissons, mysidacés) qui présentent des signatures
isotopiques voisines. Cette méthode empirique, bien qu'extrêmement chronophage (du fait du nombre
important d'individus à analyser afin d'obtenir une image globale fiable de l'alimentation), se révèle
donc indispensable lorsque la détermination précise des proies ingérées est nécessaire. Elle ne procure
cependant aucune indication quant au degré d'assimilation des proies ingérées. Les analyses isotopiques,
qui fournissent directement une image intégrée de ce qui est utilisé par le poisson pour sa croissance,

Réseaux trophiques et suivi de la MOP terrigène
244
sont donc plus informatives d'un point de vue purement "énergétique" (à condition, bien sûr, que les
signatures des proies soient connues et suffisamment distinctes pour que leurs contributions respectives
à l'alimentation des poissons soient estimées). Elles permettent de plus, lorsque les différentes sources
de nourriture sont prises en compte, de déterminer la position trophique précise des poissons dans le
milieu étudié. Leur utilisation constitue donc un "plus" dans l'étude des flux de matière et la
reconstitution des réseaux trophiques. Cependant, les analyses de la chair des poissons, bien que plus
rapides à réaliser et nécessitant un nombre réduit d'individus, sont encore très coûteuses et ne permettent
pas, seules, de décrire précisément les relations proies-prédateurs lorsque les proies potentielles sont
multiples. L'étude de l'alimentation des poissons par les méthodes conventionnelles est alors nécessaire
pour limiter le nombre de proies à analyser à celles consommées en majorité par chaque groupe de
prédateurs et permettre une utilisation "éclairée" de la méthode. Dans ce cas, la détermination fiable des
sources de matière organique est possible et les résultats obtenus permettent de dégager les stratégies
alimentaires des espèces étudiées.
Signature isotopique en azote et niveau trophique des poissons
Pour les cinq poissons plats étudiés, le δ15N augmente de façon progressive depuis les espèces
de taille réduite (A. laterna, B. luteum) vers celles de taille plus importante (soles). De même, au sein de
chaque espèce, le δ15N augmente de façon significative entre le stade juvénile et le stade adulte. Ce
phénomène, largement décrit chez les poissons, est en général attribué à une élévation intra et
interspécifique du niveau trophique avec la taille des individus (Hesslein et al., 1991 ; Ponsard et
Averbuch, 1999 ; Griffin et Valiela, 2001). Au large du Rhône, ce schéma global semble parfois
respecté, mais plusieurs observations suggèrent l'existence de particularités inhérentes aux espèces en
présence. Le δ15N moyen obtenu pour la sole commune, S. solea, est réduit par rapport à celui attendu
étant donné sa taille adulte élevée. De même, l'augmentation du δ15N entre juvéniles et adultes varie
d'une espèce à l'autre et ce, sans aucun lien avec l'augmentation de taille correspondante (Fig. III.46).
L'élévation du δ15N pendant la croissance est ainsi maximale chez les deux poissons de petite taille (A.
laterna et B. luteum) et cinq fois plus forte chez C. linguatula que chez S. impar en dépit des tailles
similaires des adultes de ces deux dernières espèces.

Réseaux trophiques et suivi de la MOP terrigène
245
Figure III.46 – Droites de régression δ15N = f (LT) obtenues pour les cinq poissons étudiés. Dans chaquecas, la longueur de la droite indique l'intervalle de taille couvert par les individus de l'espèce au large duRhône.
L'évolution des δ15N au large du Rhône ne reflète pas forcément celle des niveaux trophiques.
Le calcul du NT moyen des juvéniles et des adultes des cinq espèces étudiées sur la base des δ15N de
leurs sources principales de carbone montre, pour certaines d'entre elles, d'importantes différences entre
l'évolution des NT calculés et celle des δ15N au cours de la croissance (Tableau III.27). Ainsi, les
augmentations de δ15N observées entre juvéniles et adultes des cinq espèces reflètent souvent plus un
changement de chaîne alimentaire et de source de matière organique qu'une réelle augmentation du
niveau trophique des poissons. Les implications de ces différences interspécifiques d'évolution des
niveaux trophiques seront discutées pour les cinq espèces en termes de stratégies alimentaires dans la
Partie IV. Il est cependant important de préciser ici que l'évolution du NT calculé dépend fortement, tout
comme celle du δ15N, de l'espèce considérée mais que l'augmentation de niveau trophique au cours de la
croissance n'est pas systématique. On constate également que, dans la mesure où la signature de la
source principale de matière organique est utilisée de façon systématique pour le rétro-calcul des
niveaux trophiques, les valeurs fournies par l'utilisation de la formule proposée sont très proches de
celles habituellement indiquées pour chaque espèce. Ceci confirme la validité de la formule utilisée (c. f.
"matériel et méthodes") et la bonne représentativité des enrichissements trophiques moyens estimés
pour les herbivores et les carnivores au large du Rhône.
0 50 100 150 200 250 300 350
δ 15 N
LT (mm)7
8
9
10
11
12
13 C. linguatula
B. luteum
A. laterna
S. solea
S. impar

Réseaux trophiques et suivi de la MOP terrigène
246
Tableau III.27 – δ15N et niveaux trophiques moyens (NT) obtenus pour les juvéniles et les adultes des cinqpoissons au large du Rhône. Dans chaque cas, le δ15N de la source principale de matière organique indiquée aservi de signature de référence pour les producteurs primaires dans les calculs (cf. chapitre matériel etméthodes). En cas de mélange de deux sources, la moyenne des deux signatures a été utilisée. Pour chaqueespèce, les NT fournis sur le site de Fishbase (Froese et Pauly, 2000) sont indiqués à titre comparatif.
δ15N (‰) Principale source de MOP NT calculéNT de l'espèce
(Fishbase)
S. solea juvéniles adultes
10,111,1
MOP rhodanienneMOP rhodanienne
3,483,56 3,28 ± 0,44
S. impar juvéniles adultes
10,711,1
Phytoplancton/ DiatoméesDiatomées
3,473,25 3,23 ± 0,41
B. luteum juvéniles adultes
8,811,7
PhytoplanctonMOP rhodanienne
3,213,54 3,39 ± 0,43
A. laterna juvéniles adultes
8,510,1
PhytoplanctonPhytoplancton
3,123,62 3,59 ± 0,54
C. linguatula juvéniles Adultes
9,111,6
PhytoplanctonPhytoplancton
3,314,07 3,97 ± 0,65
Signature isotopique en carbone et alimentation des poissons
La signature en carbone varie, tout comme la composition du régime alimentaire, selon l'espèce,
le stade de vie benthique et la profondeur considérés. Il est également probable que, du fait des
modifications saisonnières de l'alimentation des cinq espèces, le transfert de la MOP rhodanienne vers
les poissons dépende également de la saison. Cependant, les modifications saisonnières de l'alimentation
sont réduites au large du Rhône et nettement moins marquées que celles liées à la profondeur. Les
variations saisonnières induites de signature du matériel ingéré sont donc faibles et il est peu probable
que ces différences soient décelables au niveau du muscle blanc des individus analysés du fait de la
faiblesse du renouvellement musculaire chez les poissons (0,1 - 0,2 % par jour) (Hesslein et al., 1993).
Les modifications de l'alimentation observées selon la profondeur et entre les deux stades de vie
benthique justifient les modifications ontogéniques et bathymétriques de la signature en carbone
obtenues pour chaque espèce au large du Rhône. En ce qui concerne les modifications ontogéniques, on
observe un net parallélisme entre l'évolution de la signature en carbone et celle du régime alimentaire au
cours de la croissance des individus de chaque espèce (Fig. III.47). Pour S. solea, A. laterna et C.
linguatula, chez qui la signature en carbone ne varie pas de façon significative entre juvéniles et adultes,
la composition du régime alimentaire diffère peu d'un stade de vie à l'autre. Par contre, pour S. impar et
B. luteum, chez qui le δ13C se modifie de façon significative d'un stade à l'autre, les proies consommées
diffèrent de façon importante entre juvéniles et adultes. Les modifications de la signature en carbone
d'un groupe de poissons à l'autre sont directement liée à la quantité de polychètes ingérée. Les deux
espèces ayant les signatures en carbone les plus négatives, S. solea (-20,0‰) et B. luteum (-18,6‰),
sont celles qui consomment le plus de polychètes (respectivement 71 et 47% du poids total des proies
ingérées).

Réseaux trophiques et suivi de la MOP terrigène
247
Figure III.47 – Composition pondérale du régime alimentaire des juvéniles G0 (J) et des adultes (A) des cinqpoissons plats étudiés au large du Rhône. Pour chaque espèce, la flèche indique s'il y a modificationsignificative (*) ou non (ns) de la signature en carbone (δ13C, en ‰) entre les deux stades de vie benthique.
De même, chez B. luteum et S. impar, le stade de vie benthique présentant la signature la plus
négative est celui qui ingère le plus de polychètes. Cependant, l'éthologie des polychètes consommés
joue également et la signature est d'autant plus négative que la proportion de dépositivores ingérés est
plus élevée. A condition de tenir compte des modifications de signatures des proies à chaque profondeur
de capture des poissons et de l'importance des différents polychètes (suspensivores, carnivores, et
dépositivores de surface ou de sub-surface) dans leur alimentation, le calcul de la signature en carbone
moyenne attendue pour chaque stade de vie benthique en fonction de son alimentation fournit une bonne
approximation de sa signature isotopique réelle. Pour les cinq espèces, les différences entre δ13C
théoriques et δ13C observés sont légères et peuvent être raisonnablement imputées à un manque de
précision des signatures utilisées pour les proies benthiques (signature manquantes pour certaines proies
à certaines profondeurs, taille des individus analysés différente de celle des individus ingérés). Ce
résultat permet de valider l'enrichissement trophique en δ13C de +1,8‰ proposé au large du Rhône. Il
indique également une absence d'assimilation différentielle des proies ingérées par les juvéniles et les
adultes de poissons étudiés, le matériel organique non réfractaire présent dans les estomacs (i.e. débris
végétaux et sédiment exclus) reflétant totalement la composition de la chair des individus analysés.
En ce qui concerne la profondeur, le calcul des δ13C et des δ15N théoriques n'a pas pu être
réalisé, la signature de certaines proies n'ayant pas été déterminée précisément pour chacune des trois
tranches bathymétriques. Cependant, le positionnement des adultes de S. solea, B. luteum et A. laterna
����������
��������������������������
���������������������������������������
������������������������������������������
������������������������������������������
�������������
����������������������������
����������������������������
����������������������������������������������������������������������������������������������������������������
�������������������������������������������������������
�������������
���������������������������������������
����������������������������
0%
25%
50%
75%
100%
J A J A J A J A J A
Cp
polychète bivalve gastéropode amphipode macroure mysidacé brachyoure anomoure
leptostracé������������ cumacé copépode ophiurepoisson siponcle holothurie phoronidien
ns * * ns ns
S. solea S. impar B. luteum A. laterna C. linguatula

Réseaux trophiques et suivi de la MOP terrigène
248
par rapport à leurs proies potentielles sur les graphes δ13C vs δ15N obtenus pour chaque profondeur
explique en grande partie les différences bathymétriques de signature en carbone observées pour ces
groupes de poissons. Pour les trois espèces, le δ13C des adultes est minimum à 30-50 m, profondeur où
l'alimentation comprend le plus de polychètes et seule tranche bathymétrique où certaines des autres
proies ingérées (brachyoures, macroures) exploitent de façon non négligeable du matériel d'origine
terrestre. Ainsi, le transfert accru de la matière organique d'origine terrigène vers les adultes de S. solea,
B. luteum et A. laterna à 30-50 m de profondeur résulte non seulement de la forte adaptabilité trophique
de ces trois espèces mais également de celle des invertébrés benthiques qui leur servent de proies. Ce
résultat est compatible avec les données disponibles pour l'ensemble des espèces impliquées∗ et sera
discuté en termes de stratégies de vie des poissons au sein de la Partie IV.
III.6 - CONCLUSION
Au large du Rhône, la matière organique terrigène véhiculée par le fleuve participe à la
production des différentes communautés d'invertébrés benthiques situées entre 0 et 100 m de profondeur
et intègre, à chaque profondeur, les réseaux trophiques aboutissant aux cinq poissons plats étudiés.
L'importance de cette incorporation varie cependant suivant la profondeur considérée, avec une
exploitation de la MOP terrigène par les communautés benthiques maximale à 30-50 m, profondeur où
les apports rhodaniens sont les plus constants et les plus élevés. Parmi l'ensemble des proies benthiques
échantillonnées, d'importantes différences de sensibilité aux apports rhodaniens ont été mises en
évidence. Les polychètes dépositivores de surface et de sub-surface sont les seuls organismes
benthiques capables d'exploiter majoritairement la matière organique apportée par le fleuve pour leur
croissance. A ce titre, ils constituent le principal vecteur de transfert potentiel de la MOP terrigène vers
les poissons. Du fait des différences de régimes alimentaires existant entre les cinq poissons plats
étudiés, l'influence potentielle des apports du fleuve en matière organique d'origine terrestre diffère
d'une espèce à l'autre. Sur le graphe général δ13C = f(δ15N) obtenu au large du Rhône, trois grands
groupes de poissons peuvent être séparés en fonction de leur alimentation et de leurs signatures
isotopiques (Fig. III.48). Le premier (1), regroupant les adultes de B. luteum et l'ensemble des stades de
vie benthiques de S. solea, dépend essentiellement des sources de matière organique terrestres pour sa
croissance. Le second (2), formé des juvéniles de S. impar, des adultes de C. linguatula et de l'ensemble
des stades de vie benthique d'A. laterna, ingère une majorité de proies issues du réseau trophique
"marin" mais consomme également des quantités non négligeables de proies de signature "terrestre".
L'influence potentielle des apports rhodaniens sur les poissons qu'il englobe est donc réduite. Enfin, le
dernier groupe (3), incluant les adultes de S. impar et les juvéniles de B. luteum et de C. linguatula,
dépend essentiellement des sources de production marines.
∗ pour les proies : se référer plus haut ; pour les poissons : voir chapitre "Partage des ressources alimentaires"

Réseaux trophiques et suivi de la MOP terrigène
249
Figure III.48 – Schéma récapitulatif de la structure des réseaux trophiques benthiques au large du Rhône. CI =consommateur primaire, CII = consommateur secondaire. Les numéros indiquent les trois groupes de poissonsindividualisés suivant leur alimentation : 1 = poissons consommant majoritairement CI et/ou CII "terrestres"; 2= poissons consommant surtout CI et/ou CII "marins" + des quantités moindres de CI et/ou CII "terrestres"; 3 =poissons consommant presque exclusivement CI et/ou CII "marins".
L'étude combinée de l'alimentation des poissons et de la composition isotopique (carbone et
azote) de leur chair permet donc non seulement de reconstituer le cheminement de la matière organique
le long des réseaux trophiques benthiques, mais également d'estimer l'influence potentielle des apports
rhodaniens en MOP terrigène sur chacune des cinq espèces étudiées. Celle-ci est maximale chez la sole
commune S. solea, pour qui on observe une incorporation massive de matériel d'origine terrestre dans la
chair de l'ensemble des stades de vie benthique. Elle est réduite pour B. luteum et A. laterna, où elle
concerne, soit un seul des deux stades de vie benthique (adultes de B. luteum), soit les deux mais de
façon moindre (A. laterna). Enfin, elle est minimale chez S. impar et C. linguatula chez qui un seul des
deux stades de vie benthique (respectivement les juvéniles et les adultes) est légèrement influencé,
l'autre dépendant exclusivement des sources de production marines. Les implications biologiques et
démographiques de ces différences interspécifiques d'exploitation de la MOP terrigène seront discutées
au sein de la Partie IV afin de préciser l'impact potentiel des crues du Rhône sur les populations des cinq
espèces étudiées.
1
2
3
C I
C II
C II
C I
Réseau trophique:
" marin" (surtout phytoplancton)
" terrestre (MOP rhodanienne)
Cl A Cl JBl J
Bl A
Al A
MOPRhône
sédiment
MOPeau
phytopK pol C
pol S
pol DsS.
pol DS
gobiidae
cumacé
copépode
mysidacé
brachyoure
ophiure
bivalve juv.
Si A
Alpheus
Crangon
amphipode
holothurieanomoure
gast.
Ss JSs A
Si J
Al J
-28
-27
-26
-25
-24
-23
-22
-21
-20
-19
-18
-17
-161 2 3 4 5 6 7 8 9 10 11 12
δ15N
δ13C
bivalve ad.

Réseaux trophiques et suivi de la MOP terrigène
250

PARTIE IV
DISCUSSION SUR L'INFLUENCE DES APPORTS RHODANIENS ET CONCLUSIONS GENERALES


Discussion et conclusions générales
251
DISCUSSION SUR
L'INFLUENCE DES APPORTS RHODANIENS
ET CONCLUSIONS GENERALES
Cette étude contribue en partie à élucider la question du devenir de la matière organique
déversée par le Rhône en Méditerranée, dans la mesure où elle a permis de démontrer que la MOP
terrigène apportée par le fleuve participe de façon non négligeable à la production des communautés
macrobenthiques situées au large de son embouchure. Cependant, le principal apport de ce travail réside
dans la mise en évidence d'un transfert trophique de MOP rhodanienne vers les stades de vie benthique
des principales espèces de poissons plats capturées au large du fleuve (Solea solea, Solea impar,
Buglossidium luteum, Arnoglossus laterna et Citharus linguatula). D'après les signatures isotopiques
obtenues, l'influence potentielle des apports du Rhône serait maximale pour S. solea, plus réduite chez
B. luteum et A. laterna et minimale chez S. impar et C. linguatula.
Il est cependant possible que ce schéma se modifie d'une année sur l'autre en fonction des fluctuations
du débit du Rhône. En effet, l'intensité de l'incorporation de la MOP terrigène et les modalités de son
transfert au sein des réseaux trophiques benthiques dépendent directement de la quantité de matière
organique disponible à la base des réseaux trophiques, comme en témoignent les différences
bathymétriques mises en évidence au cours de ce travail (c. f. Partie III). L'échantillonnage réalisé,
effectué de mars 2000 à juin 2001, survient à la fin d'une période de débits annuels rhodaniens modérés,
initiée en 1996 (Fig. IV.1). L'année 2001, avec un débit annuel moyen supérieur à la moyenne des 20
dernières années et plus de 65 jours de crues au printemps (mars - avril) peut être considérée comme
une année de débit relativement élevé pour le Rhône. Cependant, l'entrée de la MOP rhodanienne dans
les réseaux trophiques benthiques se fait essentiellement via les polychètes dépositivores, qui
constituent le principal vecteur de transfert de matériel organique d'origine continentale vers les
poissons (c. f. Partie III). Or, le délai de réponse de leurs populations aux crues du fleuve varie de
quelques mois à plusieurs années (2-3 ans) suivant l'éthologie alimentaire des espèces (Salen-Picard et
al., 2002). L'influence des crues printanières de 2001 sur la composition des communautés n'a donc pu
être décelée au cours de cette étude. Les résultats obtenus correspondent ainsi à une situation de débit
modéré du Rhône et reflètent l'influence "moyenne" des apports rhodaniens sur les communautés
benthiques et les populations de poissons plats.

Discussion et conclusions générales
252
500 600 700 800 900
1000 1100
1200 1300 1400 1500 1600 1700 1800 1900 2000 2100 2200 2300
1980
1981
1982
0 10 20 30 40 50 60 70 80 90 100 110 120 130 140 150 160 170 180 190 200
1983
1984
1985
1986
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
Débit annuel moyen (m 3 s -1 )
Nombre annuel
de jours de crue
(débit ≥ 3000 m3 s-1)
Figure IV.1 – Débits moyens annuels (courbe : moyenne en noir ± erreur-type en gris) et nombres annuelsde jours de crues (histogrammes) du Rhône pour la période 1980-2001, calculés d'après les observations àBeaucaire de la Compagnie Nationale du Rhône. La droite en pointillés indique le débit annuel moyen(Dmoy.= 1770 ± 83 m3 s-1) obtenu sur l'ensemble de la période 1980-2001.
En cas de modification importante du débit du Rhône, différents facteurs sont susceptibles
d'interagir et de modifier le schéma observé. La répartition spatio-temporelle des poissons, la
composition et la variabilité de leur régime alimentaire et le partage intra et interspécifique des
ressources peuvent agir sur le transfert de la matière terrigène, dans la mesure où des différences
d'exploitation de la MOP terrestre ont été mises en évidence suivant les catégories de proies et la tranche
bathymétrique considérées (c. f. Partie III). Pour cette raison, les implications des différences de
stratégie de vie des poissons seront discutées, afin d'estimer l'intensité potentielle maximale du transfert
de MOP terrigène vers les juvéniles et les adultes de chaque espèce. Des hypothèses sur l'impact
éventuel des crues du Rhône sur le cycle de vie et la dynamique de population des cinq espèces seront
ensuite émises en envisageant essentiellement les effets positifs des crues sur la biologie des espèces
mais en évoquant également les effets négatifs potentiels liés à la pollution rhodanienne.

Discussion et conclusions générales
253
IV.1 - STRATEGIES DE VIE BENTHIQUE ET IMPACT POTENTIEL DES APPORTS
RHODANIENS EN MOP TERRIGENE SUR LES CINQ ESPECES
Au large du Rhône, l'alimentation de S. solea, S. impar, B. luteum, A. laterna et C. linguatula
est basée sur l'ingestion des trois mêmes groupes zoologiques (polychètes, mollusques, crustacés).
Cependant, la compétition alimentaire entre espèces est réduite, non seulement grâce à des différences
de rythmes d'activité, de preferenda alimentaires et de taille des proies ingérées, mais également par des
modifications spatio-temporelles de la densité des individus et de la composition de leur régime
alimentaire. Les implications de ces différences en termes de stratégies de vie et de sensibilité des
poissons aux crues du Rhône sont discutées ci-dessous.
IV.1.1 - STRATEGIES DE VIES ET PARTAGE DES RESSOURCES ALIMENTAIRES
Périodes d'alimentation
L'existence de différences de périodes d'alimentation est considérée comme un moyen efficace
de réduire le partage interspécifique des ressources alimentaires disponibles dans le milieu (Carter et al.,
1991). Parmi les espèces de poissons étudiées, deux (S. solea et B. luteum) s'alimentent principalement
de nuit et trois (A. laterna, C. linguatula et, semble-t-il, S. impar) de jour. Ceci permet de réduire en
permanence la compétition alimentaire directe entre ces poissons, d'autant plus que les pics alimentaires
sont décalés dans le temps entre espèces nocturnes d'une part et entre espèces diurnes de l'autre.
L'intensité de la nutrition varie également suivant la saison, mais de façon relativement similaire pour
les cinq espèces. La prise de nourriture est maximale en été, importante au printemps et fortement
réduite en hiver, sauf chez A. laterna, pour qui elle est minimale au printemps. L'intensité de la nutrition
intervient donc peu dans la réduction de la compétition alimentaire et est plutôt à relier aux
modifications saisonnières des conditions environnementales. Chez les poissons, la disponibilité des
proies dans le milieu et les conditions abiotiques déterminent souvent la durée de la période végétative
(i.e. la période où la population peut maintenir un métabolisme élevé et augmenter sa biomasse) au
cours de laquelle se fait l'essentiel de la prise de nourriture (Nikolskii, 1969). L'alimentation des cinq
poissons plats étudiés est ainsi maximale en été et au printemps, saisons caractérisées par de fortes
biomasses de proies benthiques (Massé, 1972b ; Salen-Picard et al., in press), et des températures
élevées entre 0 et 20 m de profondeur (17 – 25°C de mai à septembre) (Younes, 2000). De même, la
réduction hivernale de l'alimentation est à relier à la baisse de métabolisme associée à des températures
plus faibles et à la diminution d'abondance des proies dans le milieu. Cependant, si la période et la durée
de l'alimentation dépendent essentiellement, chez les juvéniles, de la disponibilité de la nourriture et des
conditions abiotiques, avec en général une alternance de périodes d'alimentation et de pauses hivernales
jusqu'à l'établissement de la maturité sexuelle, elles dépendent également, chez les adultes, de la période
de reproduction (Nikolskii, 1969). Chez ces derniers, la prise de nourriture est généralement importante

Discussion et conclusions générales
254
pendant la phase de gamétogenèse et réduite, voire inexistante, pendant la période de frai. Ce
phénomène, et la dominance d'adultes dans la population d'A. laterna, expliquent la stratégie annuelle
d'alimentation maximale de cette espèce, peu sensible aux baisses de température hivernales (Tito de
Morais et Bodiou, 1984) et à reproduction surtout printanière (Giovanardi et Piccinetti, 1984). Le
décalage de la période d'alimentation d'A. laterna (été-hiver) par rapport à celles des autres espèces
(printemps-automne) présente des avantages potentiels. Il permet aux juvéniles de l'espèce de
poursuivre leur croissance pendant la période hivernale (et ainsi, non seulement de réduire plus vite leur
vulnérabilité vis-à-vis de prédateurs potentiels, mais également d'atteindre plus rapidement la taille de
première maturité sexuelle) et à ses adultes d'élaborer leurs réserves énergétiques en vue de la
reproduction pendant une période de compétition minimale pour les ressources alimentaires. Ceci
pourrait expliquer en partie la dominance numérique d'A. laterna sur les quatre autres espèces.
Preferenda alimentaires
La nature des organismes ingérés par chaque poisson dépend en grande partie de ses capacités
de détection et de capture des proies et de la morphologie de son appareil digestif (De Groot, 1971). Les
différences de morphologie et d'équipement sensoriel existant entre les soleidés d'une part, et les
bothidés et les citharidés de l'autre, expliquent les différences de preferenda alimentaire observées. Les
soleidés sont morphologiquement adaptés à la capture de proies épigées ou endogées vulnérables et peu
mobiles, comme les polychètes et les bivalves qui constituent généralement l'essentiel de leur régime
alimentaire (De Groot, 1971). De fait, l'alimentation est dominée par des polychètes chez S. solea et B.
luteum, et par des mollusques bivalves chez S. impar. Par contre, les bothidés et les citharidés sont
décrit comme de redoutables chasseurs à vue, capables de capturer et d'ingérer des proies plus
volumineuses et plus mobiles (De Groot, 1971). Ceci explique la dominance des crustacés (mysidacés,
amphipodes, macroures du genre Crangon) dans l'alimentation d'A. laterna et celle des poissons et des
crustacés (mysidacés, macroures du genre Alpheus) dans le régime alimentaire de C. linguatula.
Au delà de ces différences qualitatives, les alimentations de S. solea, S. impar, B. luteum, A.
laterna et C. linguatula au large du Rhône diffèrent fortement du point de vue de la taille des proies
consommées. Pour chaque taille de prédateur, il existe une taille de proie optimale, correspondant à un
équilibre entre sa capacité à capturer les proies les plus volumineuses et la possibilité qu'il a de se
procurer suffisamment de proies de taille réduite, pour satisfaire ses besoins physiologiques (Steven,
1930). L'augmentation du poids moyen des proies ingérées entre les espèces de petite taille (Pm = 0,2-
0,4 mg chez B. luteum et A. laterna) et celles de taille supérieure (Pm = 0,6-2,5 mg chez S. solea, S.
impar et C. linguatula) se vérifie au large du Rhône. Cependant, le poids moyen des proies varie du
simple au double entre B. luteum (0,2 mg) et A. laterna (0,4 mg) et est multiplié par 4 entre S. impar
(0,6 mg) et C. linguatula (2,5 mg). Ce phénomène s'explique par l'existence de différences

Discussion et conclusions générales
255
interspécifiques de la taille de l'ouverture buccale chez les Pleuronectiformes (De Groot, 1971). D'une
façon générale, les soléidés présentent, à taille égale, une ouverture de la bouche nettement inférieure à
celle des bothidés et des citharidés, ce qui, combiné avec les différences interspécifiques de taille du
corps, explique les modifications de taille des proies consommées.
Chez chacune des espèces étudiées, l'alimentation subit des modifications quantitatives et
qualitatives au cours de la croissance. Ce phénomène, largement documenté chez les poissons (Steven,
1930 ; Nikolskii, 1969 ; Gerking, 1994) résulte principalement de l'existence de besoins alimentaires
différents entre juvéniles (petite taille, énergie exclusivement dédiée à la croissance) et adultes (taille
supérieure, énergie essentiellement allouée à la reproduction) (Nikolskii, 1969). Cependant, chez les
poissons plats, les modifications ontogéniques de l'alimentation correspondent également à une
adaptation progressive à la vie benthique (Steven, 1930 ; Sorbe, 1981). La consommation importante de
copépodes harpacticoïdes et de cumacés, observée au large du Rhône chez les juvéniles G0 de S. solea,
S. impar, B. luteum et A. laterna, se retrouve chez les juvéniles de nombreux poissons démersaux et
correspond à un vestige de planctonophagie (Marchand et Masson, 1989). La fréquence de capture de
ces proies diminue avec la croissance des individus (Rogers et Jinadasa, 1989) qui diversifient peu à peu
leur alimentation en recherchant des invertébrés benthiques de plus grande taille. Nos résultats
correspondent à ce schéma, mais les modifications observées dépendent de l'espèce considérée. Deux
stratégies principales se dégagent suivant le système de sélection des proies. La première, observée chez
C. linguatula, consiste en une simple augmentation de la taille des individus ingérés sans modification
de la composition du régime alimentaire (poissons, mysidacés et macroures quel que soit le stade de vie
benthique). La seconde, observée chez les autres espèces, combine une modification de la nature, de la
taille et du nombre des proies ingérées. Ces modifications sont maximales chez S. impar et B. luteum,
pour qui les catégories de proies consommées diffèrent de façon importante entre juvéniles et adultes.
Niveau trophique
Puisque la taille du prédateur détermine la gamme de taille des proies consommées (Cohen et
al., 1993), on peut s'attendre à une augmentation du niveau trophique avec la taille des poissons.
Cependant, malgré des tailles adultes et des poids moyens de proies ingérées différents, les poissons
étudiés présentent des niveaux trophiques moyens relativement similaires au large du Rhône (Tableau
IV.). De plus, l'augmentation du niveau trophique des proies ingérées entre juvéniles et adultes d'une
même espèce n'est pas systématique. S. solea et S. impar présentent ainsi un niveau trophique similaire
quel que soit le stade de vie benthique, du fait de régimes alimentaires particuliers des adultes
(polychètes dépositivores chez S. solea et siphons de bivalves chez S. impar).

Discussion et conclusions générales
256
Tableau IV.1 – Poids moyen des proies ingérées et niveau trophique calculé pour les juvéniles, les adultes etl'ensemble des individus de chaque espèce au large du Rhône. En cas de modification ontogénique sensible de l'unde ces deux paramètres, la différence observée est indiquée sous la flèche d'évolution correspondante.
Poids moyen des proies (en mg) Juvéniles Adultes Moyenne
Niveau trophique calculé Juvéniles Adultes Moyenne
S. solea 4,9
1,0 1,9 3,5 3,6 3,5
S. impar 2,5
0,4 0,6 3,5 3,3 3,4
B. luteum 1,2
0,4 0,2
3,5
3,2 3,4
A. laterna 1,3
0,1 0,4
3,6
3,1 3,4
C. linguatula
54,1
2,5
1,4
4,1
3,7
3,3
Répartition spatio-temporelle
Chez les poissons plats, l'habitat des juvéniles est généralement réduit à la zone la plus côtière
de l'aire de répartition totale de la population et les individus présentent une distribution bathymétrique
de plus en plus profonde au cours de leur croissance (Gibson, 1994). Cependant, plusieurs espèces de
Pleuronectiformes échappent à cette règle (Marinaro et al., 1983 ; Amara et al., 1998 ; Garcia-
Rodriguez et Eesteban, 2000 ; Bolle et al., 2001). Au large du Rhône, S. impar et C. linguatula
effectuent ainsi la totalité de leur vie benthique dans une même tranche bathymétrique (respectivement
0-20 m et 30-100 m de profondeur) alors que, chez S. solea, B. luteum et A. laterna, le déroulement de
la phase de vie benthique est plus traditionnel. Pour ces dernières, le recrutement et l'essentiel de la
croissance se font entre 0 et 20 m de profondeur, les adultes se répartissant ensuite de 10 à 100 m de
profondeur, avec un maximum d'abondance entre 30 et 50 m. Cette modification ontogénique de
répartition bathymétrique présente l'avantage de limiter les interactions entre juvéniles et adultes d'une
même espèce, diminuant ainsi la compétition alimentaire intraspécifique. Sur le plan interspécifique, la
compétition pour l'espace est maximale chez S. impar (dont l'aire de répartition bathymétrique est
également exploitée par l'ensemble des stades de vie benthique de S. solea, B. luteum et A. laterna) et
minimale chez C. linguatula (pour qui le recrutement et la croissance se réalisent sur des fonds
uniquement fréquentés par les adultes de S. solea, B. luteum et A. laterna).
+ 3,9
+ 2,1
+ 0,8
+ 1,2
+ 0,3
+ 0,5
+ 0,8+ 52,7

Discussion et conclusions générales
257
Au large du Rhône, les migrations saisonnières des espèces étudiées sont peu marquées et les
déplacements bathymétriques des adultes de S. solea, B. luteum et A. laterna suffisamment réduits pour
permettre l'acquisition de signatures isotopiques caractéristiques des profondeurs de capture (δ13C plus
négatif à 30-50 m, c. f. Partie III). Ce résultat est assez surprenant, du moins en ce qui concerne S. solea.
Au large des côtes atlantiques françaises, les adultes de sole effectuent d'importantes migrations
saisonnières entre leurs zones d'alimentation côtières et leurs aires de ponte au large (Koutsikopoulos et
al., 1995 ; Amara et al., 1998). Les différences de comportement observées entre les adultes de S. solea
en face du Rhône et au large des côtes Atlantiques peuvent s'expliquer par les différences de pente du
talus continental entre ces deux régions. En Méditerranée comme en Atlantique, les soles adultes se
reproduisent au dessus des fonds de 30-70 m de profondeur (Campillo et al., 1989 ; Koutsikopoulos et
al., 1995). Cependant, ces derniers sont situés à plusieurs dizaines de km des zones d'alimentation
côtières en Atlantique, alors qu'au large du Rhône, les fonds de 10 m et ceux de 90 m ne sont distants
que de 8 km. Il est donc possible pour les adultes de l'espèce présents à 0-20 m et 70-100 m au large du
Rhône de demeurer à profondeur constante la majorité du temps et de ne rejoindre les zones de pontes
que de façon ponctuelle, au moment du frai. Cette explication est d'autant plus probable que les proies
principales de S. solea (polychètes) sont présentes tout au long de l'année en grandes quantités au large
du Rhône et ce, quelle que soit la tranche bathymétrique considérée (Massé, 1972b ; Salen-Picard et al.,
in press). Les conditions trophiques de la zone d'étude, aussi favorables pour les adultes de l'espèce à 0-
20 m qu'à 30-50 m ou à 70-100 m, ne rendent pas obligatoire une migration alimentaire vers les zones
les plus côtières pendant la saison chaude.
Les cinq poissons plats étudiés présentent des stratégies différentes en ce qui concerne la
profondeur d'installation benthique de leurs juvéniles. Solea solea, S. impar, B. luteum et A. laterna
recrutent à 0-20 m, et C. linguatula à 30-100 m. En s'installant sur les fonds vaseux de plus de 30 m de
profondeur, les jeunes recrues de C. linguatula évitent toute compétition alimentaire importante avec les
autres espèces étudiées puisque seuls les adultes de S. solea, B. luteum et A. laterna (qui consomment
des proies plus grandes) sont présents à 30-100 m. Par contre, l'installation benthique des juvéniles des
quatre autres espèces sur les fonds de 0-20 m de profondeur au printemps et en été est susceptible
d'entraîner une compétition alimentaire importante entre ces individus, qui présentent des régimes
alimentaires relativement similaires (c. f. Partie II chapitre I). Au sein des assemblages de poissons, des
différences de périodes de recrutement et de taux de croissance sont en général développées par les
espèces pour limiter la compétition alimentaire au moment de l'installation benthique des post-larves
(Nikolskii, 1969). C'est le cas au large du Rhône pour S. solea, S. impar, B. luteum et A. laterna dont les
périodes de recrutement se succèdent, d'avril à mai pour S. solea, de mai à juin pour S. impar et A.
laterna et de juin à juillet pour B. luteum. De plus, la métamorphose et l'installation benthique
surviennent à des tailles différentes suivant les espèces. La métamorphose commence à environ 8 mm
de LT chez S. solea et B. luteum, contre 30 mm de LT chez S. impar et A. laterna (Giovanardi et

Discussion et conclusions générales
258
Piccinetti, 1984 ; Shéhata, 1984 ; Amara et Lagardère, 1995). Les importantes différences
interspécifiques de taille de la bouche qui résultent de ce phénomène réduisent également, au cours des
premiers mois de vie benthique, la compétition alimentaire potentielle entre les juvéniles des cinq
espèces ayant les régimes alimentaires les plus proches.
Adaptabilité trophique et modifications spatio-temporelles de l'alimentation
Les régimes alimentaires observés traduisent l'existence d'importantes différences de stratégies
alimentaires entre les cinq espèces étudiées. Citharus linguatula présente la stratégie la plus distincte,
avec une alimentation très spécialisée, centrée sur l'ingestion d'un nombre réduit de proies
nectobenthiques (crustacés, poissons) et une adaptabilité trophique faible, comme en témoigne la
stabilité de la composition de son régime alimentaire, tant au large du Rhône que sur l'ensemble de son
aire de répartition (Jardas, 1984 ; Belghyti et al., 1993 ; Redon et al., 1994 ; Belghyti et al., 1995).
Inversement, avec des gammes de proies relativement larges et une composition variable de leur
alimentation, S. solea, B. luteum, A. laterna et S. impar sont des prédateurs "généralistes" (Gerking,
1994), capables de modifier leur alimentation en fonction des proies disponibles dans le milieu. Au
large du Rhône, l'adaptabilité trophique de ces quatre espèces se traduit par une ingestion accrue de
polychètes et de bivalves, proies qui représentent à elles seules plus de 60% de la biomasse et de la
densité des invertébrés macrobenthiques présents dans le milieu (Abou-Hamdan, 2001). Leur
importance particulière dans l'alimentation de S. solea, S. impar et B. luteum au large du Rhône
confirme le caractère "opportuniste" de l'alimentation de ces espèces, déjà décrite par plusieurs auteurs
(Marinaro et Bouabib, 1983 ; Nottage et Perkins, 1983 ; Molinero et Flos, 1992 ; Ansel et al., 1999).
Les modifications de l'alimentation avec la profondeur ont essentiellement été observées chez S.
solea, B. luteum et A. laterna. Pour ces trois espèces, le régime alimentaire est plus diversifié à 0-20 m,
avec une ingestion accrue de bivalves, d'amphipodes et de cumacés. Les polychètes sont surtout
consommés à 30-50 m, où ils forment l'essentiel de l'alimentation des adultes de S. solea, B. luteum et A.
laterna, alors que les macroures sont principalement ingérés à 70-100 m. Ces modifications
bathymétriques de l'alimentation reflètent en grande partie celles de la composition des communautés
macrobenthiques au large du fleuve (Bodoy et Plante-Cuny, 1980 ; Salen-Picard, 1982) et résultent
vraisemblablement plus du caractère "opportuniste" de l'alimentation de S. solea, B. luteum et A. laterna
que d'une modification bathymétrique des besoins alimentaires de ces espèces.
La composition des régimes alimentaires des cinq poissons étudiés varie d'une saison à l'autre
mais de façon plus ou moins intense suivant les espèces. Les variations observées résultent en partie des
fluctuations saisonnières d'abondance des proies dans le milieu. C'est le cas pour les bivalves, plus
abondants au moment du recrutement de leurs juvéniles au printemps et en été (Massé, 1971 ; Massé et
Guérin, 1976) et alors consommés en quantité par S. impar, B. luteum et S. solea. De même, la

Discussion et conclusions générales
259
consommation d'amphipodes est maximale en été, saison où leurs juvéniles sont plus abondants (Massé,
1971, 1972a) et l'ingestion occasionnelle, par les trois soléidés, de juvéniles d'ophiures (au printemps) et
de gastéropodes (en été) semble liée aux périodes d'abondance maximale de ces proies (Massé, 1972a).
Cependant, les variations saisonnières du macrobenthos n'expliquent qu'en partie celles de l'alimentation
des poissons. La composition du régime alimentaire d'une espèce peut être influencée par sa propre
dynamique de population (Molinero et Flos, 1992). Ainsi, la consommation accrue de polychètes et de
cumacés par S. impar au printemps et de cumacés par B. luteum en été résulte essentiellement d'une
augmentation des abondances relatives de leurs juvéniles à ces saisons. L'ingestion de certaines
catégories de proies pendant la saison précédant la reproduction peut également correspondre à la
satisfaction de besoins alimentaires spécifiques des adultes en vue de la gamétogenèse (Nikolskii, 1969 ;
Molinero et Flos, 1992). Au large du Rhône, les macroures sont abondamment consommés par les
adultes d'A. laterna en hiver et au printemps, et par ceux de C. linguatula au printemps et en été. De
telles modifications saisonnières des préférences alimentaires ont déjà été observées pour C. linguatula
en Atlantique, où les femelles de l'espèce ingèrent des quantités maximales de macroures pendant les
quelques mois précédant directement la ponte (Belghyti et al., 1993). Enfin, les modifications
saisonnières de l'alimentation des espèces à forte adaptabilité trophique peuvent être liées au passage
d'un régime alimentaire à un autre en fonction de l'intensité de prédation totale exercée sur certaines
catégories de proies. Au large du Rhône, ce phénomène est relativement marqué pour B. luteum et A.
laterna, qui présentent des ressemblances alimentaires maximales du point de vue des proies ingérées et
de leur taille, mais pour qui l'alimentation diffère à chaque saison donnée entre individus présents à une
même profondeur (c. f. Partie II, Chapitre II et III).
Conclusion sur la réduction de la compétition alimentaire
Au large du Rhône, la compétition pour les ressources alimentaires entre les cinq espèces de
poissons plats étudiées est réduite, non seulement par des modifications spatio-temporelles de
répartition de leurs individus, mais également par des variations saisonnières de l'alimentation des
différents stades de vie benthique. Quelle que soit la saison, le recouvrement des niches alimentaires est
plus intense sur les petits fonds sableux côtiers (0-20 m) qu'au niveau des vases terrigènes plus
profondes (30-100 m). Pour toutes les tranches bathymétriques, les interactions d'ordre trophique sont
maximales au printemps, période où le nombre de groupes de poissons présents à chaque profondeur est
le plus élevé et les alimentations les plus similaires. Ce type d'évolution spatio-temporelle des
recouvrements des niches alimentaires, avec un maximum concernant les profondeurs et les saisons
d'abondance maximale des proies a déjà été observé dans de nombreux milieux (Thorman, 1982 ;
Macdonald et Green, 1986 ; Piet et al., 1999 ; Amara et al., 2001). Il n'implique pas forcément
l'existence d'une compétition alimentaire significative. En effet, si la ressource est présente dans le
milieu en quantité suffisante pour subvenir aux besoins de l'ensemble des espèces en présence, le

Discussion et conclusions générales
260
recouvrement des niches alimentaires, même élevé, n'entraîne pas de compétition (Thorman et
Wiederholm, 1986). Il est ainsi possible que l'abondance élevée des proies benthiques observée au
printemps et en été à 0-20 m (Massé, 1971 ; Abou-Hamdan, 2001) suffise à couvrir la totalité des
besoins alimentaires des prédateurs présents sur ces fonds à cette période, comme cela a déjà été
suggéré pour différentes zones sableuses côtières (Evans, 1983 ; Tito de Morais et Bodiou, 1984 ;
Thorman et Wiederholm, 1986 ; Rogers, 1994b).
En dépit de valeurs de recouvrement des niches alimentaires souvent élevées entre individus de
B. luteum, S. impar et S. solea, les similitudes alimentaires entre ces trois espèces de soléidés sont
réduites par les importantes différences de taille des proies ingérées existant entre leurs individus. Ce
phénomène, déjà observé entre S. solea et B. luteum (Rogers et Jinadasa, 1989 ; Darnaude, 1999 ;
Darnaude et al., 2001), résulte principalement de différences de taille de l'ouverture buccale, dues à
l'effet combiné (1) des différences de LT existant au moment de l'installation benthique entre S. impar et
les deux autres espèces (voir plus haut) et (2) des vitesses de croissance supérieures des deux soles par
rapport à B. luteum. Les interactions trophiques entre populations des cinq poissons sont donc faibles au
large du Rhône. La compétition potentielle pour les ressources alimentaires y est limitée, au printemps
et exclusivement à 0-20 m, aux juvéniles de S. solea et de S. impar et aux adultes de B. luteum et A.
laterna. Or, des différences de rythme alimentaire réduisent fortement les interactions directes entre ces
individus, S. solea et B. luteum s'alimentant essentiellement de nuit et A. laterna et S. impar, de jour. A
ceci s'ajoutent des différences au niveau de la nature des proies ingérées, qui diffère entre les juvéniles
de S. solea et de S. impar d'une part et entre les adultes de B. luteum et d'A. laterna de l'autre. Les cinq
espèces de poissons étudiées ont donc globalement réussi le partage des ressources alimentaires entre
leurs différents stades de vie benthiques. La compétition alimentaire entre leurs populations est réduite
et lorsqu'elle survient, elle ne concerne jamais l'ensemble des stades de vie benthique d'une espèce et se
limite à une saison et une profondeur données. De plus, dans ce cas, des différences de nature et de taille
des proies ingérées réduisent les interactions trophiques entre les groupes impliqués.
IV.1.2 - STRATEGIES DE VIE ET INTENSITE DU TRANSFERT DE MOP TERRIGENE
Au large du Rhône, les juvéniles et les adultes des cinq espèces étudiées ont adopté des
stratégies de vie différentes (Tableau IV.2). Celles-ci seront résumées ci-dessous afin de discuter leurs
influences respectives sur la sensibilité des poissons aux crues du Rhône avant de conclure sur l'intensité
maximale potentielle du transfert de la MOP terrigène vers chaque espèce au cours de sa phase de vie
benthique.

Discussion et conclusions générales
261
Com
pétition avec les autresespèces pour l'ingestion des
proies principales?
Adaptabilité trophique
Répartition bathym
étrique(m
aximum
d'abondance)
Niveau trophique
Preferenda alimentaires
Périodes d'alimentation
maxim
ale :
Nycthém
érale
Saisonnière
Oui
(réduite)
moyenne
0-20 m
3,5
Cu
+ Pol. (D)
Nuit
P
Juvéniles
Oui
(réduite)
élevée
0-20 m30-50 m70-100 m
3,6
Pol. (D)
Nuit
E
Adultes
Solea solea
Oui
(réduite)
moyenne
0-20 m
3,5
Pol. (D/S)
+ Cu
Jour ?
P
Juvéniles
non
moyenne
0-20 m
3,3 B
Jour ?
P - E
Adultes
Solea impar
Oui
(réduite)
moyenne
0-20 m
3,2
Co + C
u
Nuit
E
Juvéniles
Oui
(réduite)
élevée
0-20 m30-50 m70-100 m
3,5
Pol. (C+D
)+ A
Nuit
P - E
Adultes
Buglossidium
l
Oui
(réduite)
moyenne
0-20 m
3,1
A + C
u
Jour
P
Juvéniles
Oui
(réduite)
élevée
0-20 m30-50 m70-100 m
3,6
My + M
a
Jour
E - H
Adultes
Arnoglossus laterna
Oui
(réduite)
réduite
30-50 m70-100 m
3,3
My + Po
Jour
P-E
Juvéniles
Oui
(réduite)
réduite
30-50 m70-100 m
4,1
My + Po
Jour
P-E
Adultes
Citharus linguatula
Tableau IV.2 - Stratégies de vie adoptées par les juvéniles et les adultes des cinq espèces étudiées au large du R
hône. P = printemps, E = été, H
= hiver, A =
amphipodes, B
= bivalves, Co = copépodes, C
u = cumacés, M
a = macroures, M
y = mysidacés, Po = poissons, Pol. = polychètes (C
= carnivores, D =
dépositivores, S = suspensivores).

Discussion et conclusions générales
262
Preferenda alimentaires
L'existence de preferenda alimentaires distincts pour les cinq espèces étudiées conditionne
fortement leurs sensibilités respectives vis-à-vis des apports rhodaniens en MOP terrigène et détermine
en grande partie l'intensité de leurs réponses potentielles aux crues du fleuve. En effet, les différences de
signature isotopique en carbone (et donc du degré d'incorporation de matériel d'origine terrestre) des
poissons sont directement liées à la composition de leurs régimes alimentaires, du fait des différences
d'exploitation des sources de production terrestre et marine existant entre catégories de proies. Parmi ces
dernières, seuls les polychètes dépositivores et carnivores sont capables d'exploiter majoritairement la
matière organique apportée par le fleuve. Les autres proies benthiques dépendent essentiellement de la
production primaire marine, même si certains des organismes analysés (polychètes suspensivores,
bivalves, amphipodes, brachyoures, macroures du genre Crangon) semblent également capables
d'exploiter en partie les apports d'origine terrestre. Suite à une forte crue du fleuve, on peut s'attendre, au
niveau du fond, à une augmentation sélective des abondances des proies capables d'exploiter la MOP
terrigène et donc à une modification de la composition des communautés. Celle-ci se fera en partie en
faveur des bivalves et de certains crustacés (amphipodes, brachyoures, macroures du genre Crangon),
mais surtout en faveur des polychètes, comme cela a été démontré au large du Rhône (Salen-Picard et
al., 1997 ; Massé, 2000). Parmi les espèces de poissons plats étudiées, S. solea et B. luteum, toutes deux
consommatrices de grandes quantités de polychètes dépositivores et/ou carnivores, seront donc les plus
favorisées par l'augmentation des ressources alimentaires benthiques. Solea impar et A. laterna sont
également susceptibles de réagir fortement, dans l'hypothèse où l'augmentation des apports de MOP
terrestre sur le fond se traduise par un accroissement sensible de l'abondance des polychètes
suspensivores, des bivalves, des amphipodes et des Crangonidae qu'elles consomment
préférentiellement. Compte-tenu de son alimentation, l'impact des crues du fleuve sur C. linguatula est
par contre potentiellement négligeable, puisqu'aucune de ses proies préférentielles ne semble capable
d'exploiter de façon importante la MOP terrigène. Cependant, d'autres facteurs que les preferenda
alimentaires peuvent modifier le schéma attendu.
Niveau trophique
Le niveau trophique des proies ingérées peut intervenir en partie dans la détermination de
l'intensité du transfert de matériel d'origine terrestre vers chaque groupe de poissons. En effet, si
l'exploitation de MOP terrigène a une influence directe sur la dynamique de population des
consommateurs primaires capables de l'exploiter, la quantité de biomasse animale associée à cette
exploitation initiale va diminuer au fur et à mesure que l'on s'élève dans la chaîne alimentaire du fait des
déperditions d'énergie associées au passage d'un échelon trophique à l'autre (Pimm, 1982). La
consommation majoritaire de consommateurs primaires sensibles aux apports rhodaniens en MOP
terrigène (polychètes dépositivores, bivalves, amphipodes) est donc potentiellement plus avantageuse

Discussion et conclusions générales
263
pour les poissons que celle de consommateurs secondaires (macroures, polychètes carnivores). Ainsi,
même si le degré d'exploitation de MOP terrigène des proies principales de C. linguatula augmentait
suite à une crue du fleuve, le niveau trophique plus élevé de ces dernières limiterait le transfert potentiel
de matériel terrestre vers ce poisson par rapport aux quatre autres espèces étudiées.
Répartition spatio-temporelle
Au large du Rhône, la matière organique terrigène véhiculée par le fleuve participe à la
production des différentes communautés d'invertébrés benthiques situées entre 0 et 100 m de profondeur
et intègre, à chaque profondeur, les réseaux trophiques aboutissant aux cinq poissons plats étudiés (c. f.
Partie III). L'importance de cette incorporation varie cependant suivant la tranche bathymétrique
considérée, avec une exploitation de la MOP terrigène par les communautés benthiques maximale à 30-
50 m, profondeur où les apports rhodaniens sont les plus constants et les plus élevés. La répartition
spatio-temporelle des cinq espèces conditionne donc en partie l'intensité du transfert de MOP terrigène
vers leurs individus, dans la mesure où le temps passé par chaque groupe de poisson à s'alimenter à 30-
50 m est susceptible d'accroître l'incorporation de matière organique terrigène dans ses tissus. La
répartition spatiale très côtière des juvéniles de S. solea, B. luteum et A. laterna, ainsi que de l'ensemble
des stades de vie de S. impar, empêche ces individus de profiter de l'augmentation des ressources
benthiques attendue à 30-50 m de profondeur. Les adultes de S. solea, B. luteum et A. laterna et
l'ensemble des stades de vie de C. linguatula (surtout les adultes) ont par contre cette opportunité. En
cas d'augmentation du débit du fleuve, la répartition bathymétrique des espèces devrait conduire à un
accroissement des différences de transfert de MOP terrigène entre ces deux groupes de poissons.
Adaptabilité trophique et modifications spatio-temporelles de l'alimentation
Le degré d'adaptabilité trophique des poissons joue un rôle clé dans la détermination de
l'intensité du transfert de MOP terrigène vers les cinq espèces étudiées, dans la mesure où les preferenda
alimentaires observés pour S. solea, S. impar, B. luteum et A. laterna au large du fleuve résultent en
grande partie du caractère "opportuniste " de leur alimentation. Suite à une forte crue du Rhône, les
groupes de poissons consommateurs de polychètes (i. e. tous les groupes étudiés à l'exception des
juvéniles de C. linguatula) vont être plus ou moins capables de modifier leur alimentation de façon à
exploiter les pics successifs d'abondance de ces invertébrés dans le milieu. On peut ainsi s'attendre,
compte-tenu des différences d'adaptabilité trophique et de composition des régimes alimentaires mises
en évidence au cours de cette étude, à une augmentation de l'ingestion de polychètes importante chez les
adultes de S. solea, B. luteum et A. laterna, modérée chez les juvéniles de S. solea, B. luteum et A.
laterna et l'ensemble des stades de vie benthique de S. impar, et très faible chez les adultes de C.
linguatula. Des phénomènes similaires sont également susceptibles de se produire pour l'ensemble des

Discussion et conclusions générales
264
autres catégories de proies benthiques capables d'exploiter en partie les apports rhodaniens en MOP
terrigène (bivalves, amphipodes, brachyoures, etc).
Conclusion sur l'importance potentielle du transfert de MOP terrigène vers les stades de vie
benthiques des cinq espèces
Par ses preferenda alimentaires, sa répartition bathymétrique, son niveau trophique et
l'adaptabilité alimentaire de ses individus, S. solea reste, quelles que soient les conditions de débit du
fleuve, l'espèce étudiée la plus susceptible de profiter des apports rhodaniens en MOP d'origine
continentale. Le transfert de matériel terrigène vers ses juvéniles et ses adultes au large du Rhône, déjà
important en période de débit modéré, est susceptible de s'accroître encore suite à une période de fortes
crues.
Buglossidium luteum et, à un degré moindre, A. laterna sont également susceptibles de réagir
fortement aux crues du Rhône. En effet, le transfert de matière organique terrigène, élevé chez les
adultes de B. luteum et moyen chez ses juvéniles ainsi que chez l'ensemble des stades de vie benthiques
d'A. laterna, est susceptible de s'accroître en cas d'augmentation des apports rhodaniens du fait de la
forte adaptabilité trophique de ces espèces. Dans les deux cas, il est potentiellement maximal chez les
adultes, seuls présents à 30-50 m de profondeur.
Du point de vue composition du régime alimentaire et adaptabilité trophique, S. impar est
comparable à B. luteum et A. laterna. Cependant, la répartition bathymétrique de cette espèce, plus
côtière (0-20 m), ne lui permet pas d'accéder en totalité à la biomasse engendrée à l'embouchure du
fleuve par l'exploitation de la MOP terrigène par les différentes proies benthiques. L'incorporation de
matériel d'origine terrestre au sein des réseaux trophiques benthiques étant réduite à 0-20 m de
profondeur (surtout par rapport à la situation observée à 30-50 m), le transfert de MOP terrigène vers S.
impar est peu important en période de débit modéré du fleuve, où il concerne essentiellement le stade
juvénile. Cependant, dans l'hypothèse d'une augmentation de l'abondance des bivalves et des polychètes
dépositivores ou suspensivores à 0-20 m suite à une forte crue du Rhône, le transfert de matériel
terrigène vers les juvéniles et les adultes de cette espèce est susceptible d'augmenter de façon non
négligeable et d'être comparable à celui observé pour B. luteum et A. laterna en période de débit
modéré.
Par contre, il est peu probable qu'une augmentation de l'intensité des crues du Rhône ne se
traduise par un effet plus marqué des apports en MOP terrestre sur les juvéniles et adultes de C.
linguatula. En raison de son régime alimentaire particulier (juvéniles de poissons et mysidacés), cette
espèce est la moins sensible aux apports du fleuve, ceci malgré sa présence sur les fonds de 30-50 m. De
plus, sa très faible adaptabilité trophique et son niveau trophique élevé rendent peu probable une

Discussion et conclusions générales
265
augmentation de l'intensité du transfert de matériel terrigène vers ses individus en cas de modification
du milieu suite à une forte crue du fleuve. Si elle survient, celle-ci sera vraisemblablement limitée aux
adultes, seuls individus de l'espèce pour lesquels l'ingestion de polychètes, bien que ponctuelle, ait été
observée.
Pour conclure, on peut donc, en cas de crues du Rhône, s'attendre à un transfert de MOP
terrigène :
- maximal chez les adultes de B. luteum et l'ensemble des stades de vie benthique de S. solea,
- élevé chez les juvéniles de B. luteum et l'ensemble des stades de vie benthique d'A. laterna,
- moyen chez l'ensemble des stades de vie benthique de S. impar
- réduit chez les adultes de C. linguatula,
- et négligeable chez les juvéniles de cette dernière espèce.
Ces différences de sensibilité ont d'importantes implications quant aux délais et au délai et à
l'intensité des réponses des populations des cinq espèces étudiées aux crues du Rhône.
IV.2 - CONSEQUENCES POTENTIELLES DES CRUES DU FLEUVE SUR LE CYCLE DE
VIE ET LA DYNAMIQUE DE POPULATION DES POISSONS PLATS
La matière organique particulaire ne représente qu'une partie des apports du Rhône à la
Méditerranée. Ainsi, l'influence positive des apports en MOP terrigène sur la croissance et la condition
des stades de vie benthique des poissons, suggérée au cours du présent travail, ne constitue pas le seul
impact potentiel des crues du fleuve. Les apports en matière organique dissoute et en sels nutritifs
peuvent également avoir une influence dans la mesure où ils conditionnent l'intensité de la production
primaire au large du Rhône (Tusseau-Vuillemain, 1998). Ils déterminent ainsi potentiellement la
quantité de nourriture disponible pour les poissons au cours de leur phase de vie larvaire et ont
également une influence sur la quantité de nourriture présente au niveau du fond. L'impact des polluants
véhiculés par le fleuve sur la biologie des poissons est également susceptible de contrebalancer
l'influence positive de l'enrichissement du milieu associé aux crues, dans la mesure où ils sont, pour la
plupart, essentiellement déversés pendant ces périodes de fort débit (Dorten et al., 1991 ; Elbaz-
Poulichet et al., 1996). Les impacts potentiels de ces deux autres types d'apports (matériel dissous et
polluants) seront donc discutés et comparés à ceux en MOP terrigène. Des hypothèses concernant les
mécanismes et les délais de réponse des cinq espèces aux crues du fleuve seront ensuite émises afin de
prédire, dans la mesure du possible, les réactions potentielles de leurs populations à une augmentation
conséquente du débit du Rhône.

Discussion et conclusions générales
266
IV.2.1 - APPORTS RHODANIENS EN MATERIEL DISSOUS
Le Rhône fournit l'essentiel du carbone organique dissous et des sels nutritifs présents dans le
golfe du Lion (Coste et Raimbault, 1993). Ces apports enrichissent l’ensemble du golfe (Cauwet, 1996)
et ont un impact primordial sur la production primaire planctonique (Denis et al., 2001). Celle-ci est
beaucoup plus intense à la côte qu'au large (Tusseau-Vuillemain, 1998) et atteint des valeurs maximales
à l'intérieur du panache du fleuve, avec des biomasses de phytoplancton et de zooplancton élevées toute
l'année et maximales au printemps (Blanc et al., 1969 ; Blanc et Leveau, 1971 ; Gaudy et al., 1996 ;
Lefèvre et al., 1997). Cependant, contrairement aux apports en matériel particulaire, les apports dissous
ne suivent que modérément les fluctuations du débit liquide du Rhône (Cauwet, 1996). La production
primaire planctonique du golfe du Lion varie ainsi peu d'une année sur l'autre, et semble relativement
indépendante des fluctuations interannuelles du débit du fleuve (Lefèvre et al., 1997).
Apports dissous et particulaire du Rhône et quantité de nourriture disponible pour les poissons
Les apports du Rhône en matériel dissous et en matériel particulaire diffèrent par leur
importance, leur variabilité et leurs capacité de dispersion en mer (Cauwet et al., 1990). Ceci a
d'importantes conséquences quant à la nature de la matière terrigène disponible pour les organismes
marins, qu'ils soient planctoniques ou benthiques, et résulte en des différences d'impact potentiel de ces
deux types d'apports sur les stades de vie pélagiques et benthiques des poissons.
Phase de vie larvaire
Chez les poissons plats, la disponibilité en nourriture constitue, avec la température et la
prédation, les trois principaux facteurs affectant la mortalité des larves (Miller, 1994). L'alimentation a
une influence directe sur le taux de survie des individus pendant leurs premières semaines de vie. Chez
S. solea, de mauvaises conditions alimentaires pendant les jours suivant immédiatement l'ouverture de la
bouche entraînent une augmentation de la mortalité au cours de la phase larvaire (Lagardère, 1989).
Cependant, les variations du taux de survie des larves, et par conséquent du recrutement, sont en général
essentiellement déterminées par la taille des individus et la durée de la période de vie planctonique
(Miller et al., 1988 ; Pepin, 1991). Une durée prolongée de la phase larvaire accroît le temps de
vulnérabilité à la prédation (Amara et al., 1993) et, lorsque la mortalité dépend de la taille, les variations
de ce facteur peuvent avoir pour effet de modifier les risques de prédation, non seulement au cours de la
vie planctonique (Miller et al., 1988), mais également pendant la période qui suit l'installation benthique
(Van der Veer et Bergman, 1987). Or, la quantité de nourriture disponible dans le plancton, par son
impact direct sur la vitesse de croissance des larves (Amara et al., 1993), détermine non seulement leur
taille mais également la durée de leur vie planctonique puisque la métamorphose commence, le plus
souvent, à une taille constante pour chaque espèce (Amara et Lagardère, 1995). Ainsi, par le biais des

Discussion et conclusions générales
267
fortes productions planctoniques qu'ils engendrent, les apports du Rhône ont un impact bénéfique
important sur le succès de l'installation benthique et donc du recrutement des cinq espèces de poissons
plats étudiées.
Les eaux côtières au large du delta du fleuve abritent un nombre élevé de larves des cinq
espèces étudiées (Le Direac'h-Boursier, 1990), qui profitent des accroissements de productions primaire
et secondaire planctoniques à l'intérieur comme à l'extérieur du panache. L'augmentation de la quantité
de nourriture dont elles bénéficient est essentiellement liée aux apports dissous du Rhône. En effet, bien
que bon nombre de bactéries hétérotrophes marines (Omnes, 1996 ; Rolff et Elmgren, 2000) et une
partie des organismes zooplanctoniques (Bouilllon et al., 2000 ; Rolff et Elmgren, 2000) soient capables
d'exploiter la MOP terrigène en suspension dans les eaux du panache, la production phytoplanctonique
et son exploitation consécutive par le zooplancton constituent la principale source de biomasse
disponible dans le plancton de la zone côtière (Blanc et al., 1969). Pour cette raison et compte-tenu de la
faible variabilité interannuelle de la productivité planctonique du golfe du Lion (Lefèvre et al., 1997),
on peut s'attendre à ce que la quantité de nourriture disponible pour les larves de poissons soit
relativement constante d'une année sur l'autre.
Il est par contre possible que l'influence des apports rhodaniens en matériel dissous sur la phase
larvaire des poissons diffère d'une espèce à l'autre. Chez les poissons plats, la durée de la phase de vie
planctonique varie non seulement selon les espèces mais également suivant les conditions du milieu
(Amara et al., 1993 ; Amara et al., 1998). Les espèces de poissons plats étudiées, passent toutes un à
deux mois dans le plancton avant de s'installer sur le fond (Giovanardi et Piccinetti, 1984 ; Shéhata,
1984 ; Sabatés, 1988 ; Amara et Lagardère, 1995). Cependant, les périodes de reproduction de ces
espèces diffèrent, avec des conséquences différentes sur la quantité de nourriture disponible pour leurs
larves. Citharus linguatula est ainsi la seule espèce à avoir une reproduction automnale. Ce décalage de
la période de reproduction par rapport à celles généralement observées chez les poissons plats des zones
tempérées (fin d'hiver - été) peut être considérée comme une adaptation visant à augmenter le taux de
survie des larves et des juvéniles par une diminution de la mortalité par prédation et du partage des
ressources alimentaires au cours des premiers mois de vie (Nikolskii, 1969). De fait, au large du Rhône,
C. linguatula évite toute compétition avec les autres espèces au cours de sa phase de vie planctonique.
Cependant, la présence de ses larves dans le plancton à l'automne et en début d'hiver peut présenter
certains désavantages. Du fait des températures plus faibles des eaux de surface entre novembre et mars
(Younes, 2000), le taux de croissance des larves est potentiellement moins élevé à cette période qu'au
printemps ou en été. Mais, surtout, la production planctonique des eaux côtières du golfe du Lion en fin
d'automne est nettement inférieure à celle observée au printemps (Blanc et Leveau, 1971). Ceci explique
peut-être la répartition spatiale des populations de C. linguatula, dont les densités sont maximales au
large du Rhône (Campillo et al., 1989). En effet, les fortes productions phytoplanctonique et bactérienne

Discussion et conclusions générales
268
liées aux apports du fleuve aboutissent, même en automne, à des biomasses de zooplancton à son
embouchure jusqu’à 10 fois supérieures à celles relevées dans le golfe de Marseille (Blanc et al., 1969).
La zone de dilution rhodanienne constitue donc, en fin d'automne, une zone privilégiée pour la
croissance des larves planctoniques par rapport au reste du golfe. Pour les quatre autres espèces
étudiées, les périodes de reproduction s'étalent entre la fin de l'hiver (S. solea, A. laterna) et l'été (B.
luteum, S. impar) mais se chevauchent au printemps (avril-mai), période où la production planctonique
est maximale au large du Rhône (Blanc et Leveau, 1971). Ainsi la localisation printanière de la phase de
vie larvaire de ces espèces permet à leurs juvéniles de profiter de la période d'abondance maximale des
proies planctoniques. Pendant leur période de présence simultanée au sein du plancton, les individus de
S. solea, A. laterna, B. luteum et S. impar sont en compétition pour l'exploitation des ressources
alimentaires. Cependant, si les larves de poissons se nourrissent toutes dans le plancton par une capture
active de petits organismes zooplanctoniques (Last, 1980), des phénomènes de sélection des proies
(Fonds 1979) et l'existence de différences importantes de régime alimentaire des larves suivant la taille
des individus (Sanchez-Velasco, 1998) sont susceptibles de réduire fortement la compétition alimentaire
potentielle entre ces espèces. De plus, si les larves sont trop peu nombreuses pour affecter l'abondance
de leurs proies, comme cela a été suggéré par plusieurs études portant sur le nombre de proies
disponibles en zone côtière et les rations alimentaires des larves de poissons plats (Jenkins, 1987 ;
Sabatés, 1988), le partage intra et interspécifique des ressources n'entraînera pas de compétition
significative pour la nourriture.
Phase de vie benthique
Les modifications temporelles des communautés benthiques ont souvent été reliées aux
modifications de quantité de nourriture disponible associées aux fluctuations de la production
phytoplanctonique (Beukema, 1991 ; Josefson et al., 1993 ; Frid et al., 1996 ; Josefson et Conley,
1997). Les apports du Rhône en matériel dissous influencent la quantité de nourriture disponible pour
les poissons au niveau du fond, dans la mesure où bon nombre d'invertébrés benthiques dépendent
majoritairement des sources de production phytoplanctoniques pour leur croissance. Cependant, les
conséquences de ces apports sur l'abondance et la composition des communautés benthiques au large de
l'embouchure sont totalement différentes de celles des apports en matière organique particulaire. En
effet, les apports dissous du Rhône ne suivant que modérément les fluctuations du débit liquide, la
production primaire planctonique varie peu d'une année sur l'autre. Ainsi, seules les proies benthiques
exploitant la MOP terrigène apportée par le Rhône sont susceptibles de réagir fortement à la suite d'une
augmentation sensible du débit, et donc des apports, du fleuve.

Discussion et conclusions générales
269
IV.2.2 - POLLUTION ASSOCIEE
Même si l'on tient compte d'une réelle amélioration au cours des dix dernières années, le Rhône
reste une source non négligeable d'apport de polluants au milieu marin (Tableau IV.3). En ce qui
concerne les métaux lourds, le niveau de contamination du fleuve est relativement faible par
comparaison avec d'autres grands cours d'eau mondiaux (Lascombe, 1996). Le Rhône déverse
cependant chaque année en Méditerranée près de 300×103 t d'Al, 200×103 t de Fe, 4500 t de Mg, 450 t
de Pb, 320 t de Cu, 310 t de Ni, 60 t de Co, 10 t de Cd et 4 t de Hg (Dorten et al., 1991 ; Guieu et al.,
1991). Pour la majorité de ces contaminants, l'essentiel des apports rhodaniens se fait sous forme
particulaire (Lascombe, 1996). Ceci entraîne d'importantes variations des concentrations en métaux
lourds des eaux du panache selon le débit du fleuve (Dorten et al., 1991) et résulte en un dépôt final de
ces polluants dans les sédiments, avec une accumulation maximale au niveau du prodelta rhodanien
(Nolting, 1989 ; Fernex et al., 2001). Le fleuve constitue également une source d'apports en
radionucléides émetteurs α (Pu, Am, Cm, U) et β/γ (tritium, 106Ru-Rh, 90Sr-Y, 125Sb, 137Cs, 134Cs, 60Co)
(Charmasson et al., 1998). Ces éléments présentent des propriétés physico-chimiques différentes ayant
d'importantes conséquences sur leur devenir en milieu marin (Charmasson, 1998). Ainsi, le 106Ru,
élément prépondérant des apports rhodaniens, est essentiellement transporté sous forme dissoute et n'est
retrouvé qu'épisodiquement dans les sédiments au large de l'embouchure. Le césium (134Cs et 137Cs),
beaucoup moins rejeté mais présentant une forte affinité pour la phase particulaire en eaux douces, est
essentiellement déversé en périodes de fortes crues et s'accumule au niveau du prodelta (Radakovitch et
al., 1999).
Tableau IV.3 – Liste des principaux polluants apportés par le Rhône en Méditerranée et effetsrecensés sur les organismes marins.
Type de polluant Impact connu sur les organismes marins(Pérez et al., 2000)
Métaux lourds (Al, Cd, Co, Cu, Cr,Fe, Mn, Ni, Pb, Hg)
Cytotoxique, génotoxique, problèmesimmunologiques et morphologiques
Radionucléides émetteurs et β/γGénotoxique, tératogène
Composés organochlorés(PCB, HCH, DDT)
Cytotoxique, génotoxique, problèmesimmunologiques
Hydrocarbures (PAH, NAH)Cytotoxique, génotoxique, problèmes
immunologiques, physiologiques,morphologiques
Détergents Cytotoxique, problèmes physiologiques

Discussion et conclusions générales
270
En ce qui concerne la pollution organique, le Rhône constitue une source non négligeable
d'hydrocarbures (NAH et PAH) qui se concentrent dans les sédiments de son prodelta (Lipiatou et
Saliot, 1991). Les composés organochlorés les plus souvent signalés (PCB, isomères HCH, dérivés de
DDT, chlorobiphényles, hexachlorobenzène…) sont également présents dans le fleuve, mais à très
faibles concentrations (Lascombe, 1996). En revanche, la contamination par les détergents associés à la
pollution domestique, bien qu'inférieure à celle de certains autres grands fleuves européens, est non
négligeable (Maldonaldo et al., 1999).
Malgré de récents développements des recherches sur la distribution des contaminants dans les
eaux côtières et les sédiments au large de l'embouchure du Rhône (Dachs et al., 1997 ; Radakovitch et
al., 1999 ; Eyrolle et Charmasson, 2001 ; Fernex et al., 2001), peu d'études concernent leur impact sur
les organismes marins (Bocquene et al., 1993 ; Burgeot et al., 1994 ; Charmasson, 1998 ; Charmasson
et al., 1999). De plus, la majorité de ces travaux concerne les effets des PCB et des PAH (pour lesquels
le complexe industriel de Fos-sur-Mer constitue une source de contamination potentiellement aussi
importante que celle du Rhône) ou sont limitées à quelques espèces bioindicatrices. Il est probable que
la pollution rhodanienne ait un effet, même réduit, sur les cinq espèces de poissons plats étudiées. En
effet, des phénomènes de bioaccumulation et des impacts négatifs sur la biologie des poissons plats ont
été démontrés pour bon nombre de contaminants présents dans le Rhône (voir Pérez et al., 2000 pour
une synthèse) bien que, dans chaque cas, l'intensité de la réponse dépende essentiellement de la
concentration des polluants dans le milieu et de leurs interactions. Il est également probable que des
différences de taux de contamination existent entre ces espèces à cause de leurs différences de taille et
de régime alimentaire, comme cela a été démontré pour d'autres zones estuariennes (Camusso et al.,
1998 ; Camusso et al., 1999 ; Arcos et al., 2002). Cependant, en l'absence de données plus précises sur
les taux de contamination des poissons ou de leurs proies benthiques au large du Rhône, il est difficile
de discuter de l'impact potentiel des apports du Rhône en polluants sur leurs populations, compte-tenu
de l'extrême variété des contaminants impliqués.
IV.2.3 - BIOLOGIE ET DELAIS DE REPONSE DES ESPECES
Les apports du Rhône sont susceptibles d'intervenir en favorisant la croissance et la condition
des individus des cinq espèces non seulement pendant leur courte période de vie larvaire mais
également au cours de leurs années de vie benthique, soit au stade juvénile, soit au stade adulte. Les
conséquences des modifications du débit du fleuve sur la dynamique de population de chaque espèce
vont ainsi varier suivant le nombre de stades de vie impliqués et l'importance de l'exploitation de la
MOP terrigène par chacun de ces stades, elle même étroitement liée à la répartition spatio-temporelle, la
composition du régime alimentaire et les relations intra et interspécifiques des individus au large de
l'embouchure.

Discussion et conclusions générales
271
Biologie des poissons plats et impact potentiel des apports en MOP rhodanienne sur le cycle de vie
des cinq espèces
En fonction des connaissance disponibles sur la biologie des poissons plats et les facteurs
conditionnant le cycle de vie et le succès du recrutement chez les poissons, il est possible d'émettre un
certain nombre d'hypothèses concernant les conséquence potentielles des crues du Rhône sur la
dynamique de population des cinq espèces étudiées. Chez les poissons, l'importance des cohortes est
considérée comme étant principalement déterminée entre la fin de la vie larvaire et les premiers mois de
vie benthique, période où survient la plus forte mortalité (Cushing, 1982). De faibles modulations de la
mortalité naturelle à ce stade peuvent entraîner d'importantes différences d'abondance annuelle des
juvéniles (Bailey et Spring, 1992 ; Leggett et DeBlois, 1994 ; Bailey et al., 1995). Les facteurs
susceptibles de favoriser la survie des individus pendant cette période critique de leur cycle de vie vont
donc avoir une influence importante sur la dynamique de population des espèces. Parmi ces facteurs,
l'abondance des ressources alimentaires sur le fond joue un rôle prépondérant chez les poissons plats.
Dans les zones de nurseries, la quantité de proies disponibles conditionne pour l'essentiel le taux de
croissance des juvéniles (Van der Veer et Witte, 1993) et son effet sur la survie des individus semble
plus important que la température, la nature du sédiment ou encore des phénomènes de régulation
densité-dépendants (Fonds et al., 1992 ; Rogers, 1992, 1994a). Ainsi, pour chacune des espèces
étudiées, une augmentation de l'abondance des proies consommées par les juvéniles devrait favoriser la
croissance et la survie de ces derniers et augmenter ainsi de façon directe le nombre d'individus
atteignant la taille adulte et rejoignant le stock de poissons exploité. Dans ce cas de figure, on peut
s'attendre à une augmentation des captures des espèces concernées au cours des années suivant la crue,
avec un délai de réponse variant d'une espèce à l'autre selon l'âge d'entrée des poissons dans la pêcherie
et les stades juvéniles (G0, G1 et/ou G2) concernés par l'augmentation de nourriture dans le milieu.
Cependant, l'influence des crues sur la croissance et la survie des larves et des juvéniles n'est
pas le seul facteur susceptible d'avoir une influence sur la dynamique de population des poissons plats.
Les variations interannuelles du succès de la reproduction ont ainsi également un impact important, dans
la mesure où elles conditionnent le nombre d'œufs émis dans le milieu (Miller, 1994) et agissent par ce
biais sur le succès du recrutement. L'alimentation joue un rôle prépondérant dans la détermination de ce
succès puisqu'elle influence la croissance et la condition des reproducteurs (Bromley, 2000 ; Bromley et
al., 2000). Chez S. solea, la fécondité relative des femelles augmente avec la taille (Horwood, 1993).
Elle est cependant très variable d'un individu à l'autre, tout comme chez la plie pour qui la fécondité en
milieu naturel varie de 40-60% pour des femelles de tailles similaires (Horwood et Greer Walker, 1989).
Ces variations ont été reliées, par des expériences de nutrition en laboratoire, à des différences de
régimes et de rations alimentaires. Une augmentation significative du poids absolu et relatif des ovaires,
du nombre d'ovocytes et de la taille des œufs s'observe pour les femelles les mieux nourries et, au bout
d'un an d'expérience, la fécondité relative des plies soumises à de fortes rations est approximativement

Discussion et conclusions générales
272
60% plus élevée que celles des plies soumises à des rations faibles (Horwood et Greer Walker, 1989).
Ainsi, puisque la fécondité relative est plus importante chez les poissons plus grands et mieux nourris,
des quantités supérieures d'œufs de meilleure qualité sont susceptibles d'être pondues par le stock de
géniteurs lorsque la croissance et la condition des individus est favorisée par une augmentation des
ressources alimentaires. Il a également été démontré que la condition des géniteurs influence aussi la
qualité des œufs émis (Rijnsdorp et al., 1991) et détermine en partie la taille et l'âge des larves à la
métamorphose (Chambers et Legett, 1987 in Amara et Lagardère, 1995). Il semblerait donc que la
condition et la taille des géniteurs, intimement liée à leur alimentation, aient un impact direct sur le
recrutement. Une telle relation a été démontrée chez Gadus morhua, avec les individus plus âgés et plus
grands produisant la plus grande partie des œufs et le plus fort nombre de recrues avec des taux de
survie plus importants (Cardinale et Arrhenius, 2000). Dans le cas de figure d'un impact positif des
crues sur les sub-adultes et les adultes des cinq espèces étudiées au large du Rhône, on peut donc
s'attendre à une optimisation du succès reproducteur de ces individus et, ainsi, à une augmentation des
captures des espèces concernées plusieurs années après celle de la crue et variant principalement d'une
espèce à l'autre suivant l'âge d'entrée des poissons dans la pêcherie.
Délais de réponse des proies benthiques et synergie potentielle des effets des crues sur les juvéniles
et adultes de poisson
D'après les résultats obtenus au cours de cette étude, l'accroissement des apports continentaux
en matière organique devrait se traduire par une augmentation de la biomasse et de la densité des proies
benthiques essentiellement en faveur des polychètes (dépositivores de surface et de sub-surface, mais
aussi suspensivores et carnivores) et, à moindre échelle des bivalves, des amphipodes, des brachyoures
et des macroures Crangonidae. Les délais de réponses des populations de ces différents organismes sont
susceptible de différer fortement du fait de leurs répartitions au niveau du fond, leurs durées de cycle de
vie et leurs niveaux trophiques différents. Ainsi, à la suite d'un épisode de crues intense, comme celui de
1994, l'augmentation des densités de polychètes au large du Rhône se poursuit sur plusieurs années
(environ 4 ans), du fait de la succession dans le temps de pics d'espèces ayant des modes de vie et des
éthologies alimentaires différents (Salen-Picard et Arlhac, 2002). Les espèces dépositivores de surface
et suspensivores de petite taille (de niveau trophique inférieur, s'alimentant à la surface ou dans les
premiers cm du sédiment et à durées de vie courtes) réagissent en premier aux apports du fleuve, alors
que la réponse des polychètes carnivores (de niveau trophique supérieur) et dépositivores de sub-surface
de grande taille (à durée de vie plus longue et se nourrissant plus profond dans le sédiment), est différée
de plusieurs années (Fig. IV.2). Un phénomène similaire de décalage dans le temps est susceptible de se
produire pour les autres catégories de proies consommées, les bivalves et les amphipodes étant
potentiellement plus rapidement influencés que les brachyoures et les Crangonidae, de niveau trophique
supérieur.

Discussion et conclusions générales
273
Figure IV.2 – délais de réponse des différents groupes de polychètes échantillonnés au cours de laprésente étude aux crues du Rhône (d'après Salen-Picard et Arlhac, 2002 et Salen-Picard et al., 2002).
Ce phénomène a d'importantes conséquences potentielles quant aux délais de réponse des
poissons aux crues du fleuve. En ce qui concerne les polychètes par exemple, les poissons
consommateurs d'espèces dépositivores de surface seront les premiers à bénéficier de l'augmentation de
biomasse macrobenthique associée aux crues, alors que l'influence potentielle de cette dernière sur les
poissons consommateurs de polychètes carnivores sera beaucoup plus tardive. Pour cette raison, les
effets favorables des crues sur la biologie des juvéniles et des adultes de poissons sont susceptibles
d'agir en synergie chez certaines espèces étudiées. En effet, dans la mesure où l'alimentation diffère
entre ces deux stades de vie benthique, il est possible que l'augmentation différée du stock de
reproducteurs induite par la réaction aux crues des juvéniles coïncide avec l'augmentation de nourriture
disponible dans le milieu pour les adultes de l'espèce et maximise leur succès reproducteur plusieurs
années après la crue, différant d'autant la réponse maximale aux crues du stock exploité.
Impact des crues sur Solea solea et délais de réponse de sa population au large du Rhône
Les hypothèses émises ci-dessus quant aux conséquences potentielles des crues du Rhône sur le
cycle de vie et la dynamique de population des espèces de poissons plats étudiées sont étayée par un
certain nombre de résultats concernant les pêcheries de sole commune dans le golfe du Lion. En effet,
la confrontation des données disponibles sur les fluctuations temporelles du débit du Rhône, des
densités de polychètes et des captures de S. solea au large du fleuve avec les données obtenues au cours
Dépositivores de surfaceSuspensivores Dépositivores de sub-surface
Carnivores
103 ind.m-2
0
1
2
3
4
5
6
7
1993 1994 1995 1997 19981996Années
Crues de 1994

Discussion et conclusions générales
274
du présent travail a permis d'expliquer le délai de réponse de cette espèce aux crues du Rhône par une
synergie d'effets positifs impliquant l'ensemble des stades de vie de l'espèce et découlant directement de
la composition de l'alimentation de ses juvéniles et ses adultes (c.f. Salen-Picard et al. 2001) (Fig. IV.3).
Au large du fleuve, les jeunes soles consomment essentiellement des polychètes dépositivores de
surface opportunistes qui réagissent rapidement aux crues, alors que les sub-adultes et les adultes se
nourrissent principalement à partir de polychètes dépositivores de sub-surface de plus grande taille dont
l’abondance augmente après un délai de 2 à 3 ans. Ainsi, l’augmentation de la quantité de proies
présentes dans le milieu après de fortes crues va favoriser successivement l'ensemble des stades du cycle
de vie de la sole. L’année même de la crue, les larves de S. solea vont bénéficier de l’augmentation
printanière de la productivité planctonique associée aux apports rhodaniens en sels nutritifs, et ses
juvéniles (surtout les G0 qui s'installent sur le fond mais également les G1+ déjà présents) des pics
d'abondance des polychètes opportunistes qu'ils consomment préférentiellement. L'optimisation de la
croissance et de la survie de l'ensemble de ces individus doit se traduire par une augmentation du succès
du recrutement de ces deux cohortes et donc une augmentation de biomasse au niveau du stock de soles
exploité, ce qui est effectivement observé 1 à 2 ans après la crue. Deux à trois ans après la crue,
l’augmentation d'abondance des polychètes à plus longue durée de vie va favoriser la croissance et la
condition des adultes, maximisant ainsi leur succès de reproduction et augmentant les taux de
recrutement des années ultérieures (de 3 à 4 ans après la crue). La sole se reproduisant à partir de 3 ans
en Méditerranée, l'augmentation différée du stock de géniteurs induite par la réaction aux crues des
juvéniles coïncide en partie avec l'augmentation de nourriture disponible dans le milieu pour les adultes.
Ainsi, on peut s'attendre, pour S. solea, à un succès reproducteur (et donc, à un succès du recrutement
benthique consécutif) maximal deux à trois ans après la crue. Ceci explique tout à fait la dynamique
temporelle de réponse aux crues du Rhône observé pour le stock de sole commune exploité au large du
Rhône (Fig. IV.4), avec une réponse maximale au bout de cinq ans (Salen-Picard et al., 2002) qui
s'explique par l'âge modal (2 ans) de capture des individus de l'espèce pour les pêcheries du golfe du
Lion (Benouada, 1985 ; Campillo et Bigot, 1992).

Discussion et conclusions générales
275
-200
0
200
400
600
800
1000
-1800
-1400
-1000
-600
-200
200
600
1000
1400
1970 1975 1980 1985 1990 1995
Débit du RhôneflCaptures de sole
Années
Déb
it an
nuel
moy
en (m
3 s-1)
Débarquem
ents annuels (t a-1)
(a)
0 1 2 3 4 5 6 7 8
*** *0,6
0,2
0,4
0,8
1,0
R
Délai (ans)
(b)
Figure IV.3 – Influence des apports rhodaniens sur les écosystèmes côtiers et sur le cycle de la solecommune, Solea solea. MOP = matière organique particulaire. D'après Salen-Picard et al. (2002).
Figure IV.4 – Relation entre le débit du Rhône et les débarquements de sole (Solea solea). (a) courbes des écartscumulés à la moyenne obtenues pour le débit annuel moyen du Rhône et les débarquements annuels moyens desole à Martigues (golfe du Lion). (b) Evolution du coefficient de corrélation de Spearman (R) en fonction dudécalage entre ces deux paramètres. "*" = P < 0,05 ;"**" = P < 0,01 ; "***" = P < 0,001. D'après Salen-Picard etal. (2002).
adultes
oeufs������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������
juvéniles
larves
Apportsdu Rhône
Sels nutritifs
M.O.P
+
Plancton
Polychètes
Croissance+ survie
Croissance + reproduction
Nombre + taille

Discussion et conclusions générales
276
Hypothèses sur les réactions des populations des quatre autres espèces de poisson étudiées
Seule la connaissance des délais de réponse de S. solea et de ses proies aux crues du Rhône a
permis d'expliquer les relations observées entre le débit du fleuve et les pêcheries du golfe du Lion. Les
informations disponibles pour les quatre autres espèces étudiées au cours de ce travail sont nettement
moins nombreuses, voire inexistantes pour ce qui est des stocks exploités. Cependant, le schéma obtenu
pour S. solea peut être utilisé afin d'émettre certaines hypothèses quant aux importances et aux délais
des réponses de ces espèces.
Solea impar
Chez S. impar, l'alimentation se modifie nettement au cours de la croissance. Les juvéniles G0
(LT < 100 mm) consomment essentiellement de petits polychètes dépositivores de surface susceptibles
de réagir dans les premiers mois suivant la crue (2-6 mois) alors que les juvéniles G1+ (100-160 mm) et
les adultes (LT > 160 mm) ingèrent respectivement une majorité d'amphipodes et de bivalves. Ces deux
dernières catégories de proies étant susceptibles de réagir également aux apports du Rhône en MOP
terrigène, on peut envisager un effet positif des crues du Rhône sur l'ensemble du cycle de vie de S.
impar. Cependant, compte-tenu de la répartition bathymétrique très côtière de l'espèce (0-20 m), ses
proies benthiques ne vont profiter que modérément des apports du fleuve en matière particulaire. La
réponse aux crues devrait donc essentiellement impliquer les juvéniles, seuls individus consommant de
fortes quantités de proies dépendantes de la MOP terrigène pour leur croissance (polychètes
dépositivores de surface).
Ainsi, l’année même de la crue, les juvéniles G0 de S. impar vont bénéficier des pics d'abondance
des polychètes opportunistes qu'ils consomment préférentiellement. Ceci devrait se traduire par une
optimisation de la croissance et de la survie de l'ensemble de ces individus et donc une augmentation du
succès du recrutement de cette cohorte. La taille commune de S. impar dans les captures étant de 12-24
cm (Fisher et al., 1987) et les individus de l'espèce acquérant leur première maturité sexuelle au cours
de leur deuxième année de vie (Shéhata, 1984), une augmentation des prises commerciales pourrait être
sensible dès l'année suivant la crue et se poursuivre sur deux ans au moins, par un impact indirect
associé à la reproduction de la cohorte de juvéniles ayant bénéficié directement de la crue.
Buglossidium luteum
L'alimentation de B. luteum au large du Rhône comprend une part non négligeable de
polychètes, tant au stade juvénile qu'au stade adulte. Cependant, la nature des familles ingérées se
modifie au cours de la croissance. Les juvéniles G0 (LT < 50 mm) consomment exclusivement des
espèces carnivores alors que les juvéniles G1+ (50-70 mm) et les adultes (LT > 70 mm) ingèrent
également d'importantes quantités d'espèces dépositivores de petite taille. Ainsi, chez cette espèce,

Discussion et conclusions générales
277
l'ensemble des stades de vie benthique est susceptible d'être favorisé mais avec des délais différents.
L'année de la crue, l'augmentation de nourriture au large du fleuve va favoriser la croissance et la
condition des juvéniles G1+ et des adultes. Cependant, l'effet des apports en MOP terrigène sur la
quantité de nourriture disponible pour l'espèce sera également sensible plusieurs années après la crue au
moment de l'augmentation de densité des populations de carnivores. Bien que la réponse de ces derniers
soit potentiellement réduite par rapport à celle des consommateurs primaires que constituent les espèces
dépositivores, ce deuxième pic d'abondance des proies dans le milieu est susceptible d'avoir un impact
non négligeable sur B. luteum dans la mesure où il concerne l'ensemble des stades de vie benthique de
l'espèce. La réponse de la population de B. luteum aux crues du Rhône est donc relativement complexe
et difficile à prédire compte-tenu des incertitudes concernant les délais et l'intensité de réponse des
proies benthiques de l'espèce aux crues. Cependant, la taille commune de capture de B. luteum étant de
8-13 cm (Fisher et al., 1987) et ses individus se reproduisant dès l'âge de 2 ans (Shéhata, 1984), on peut
s'attendre à un impact des crues sensible dès la deuxième année suivant la crue et étalé sur plusieurs
années par une synergie de réponses différées des proies et des juvéniles favorisés.
Arnoglossus laterna
Chez A. laterna, l'alimentation des juvéniles (LT < 60 mm) est largement dominée par les
amphipodes alors que les sub-adultes et adultes (LT ≥ 60 mm) ingèrent essentiellement des macroures
Crangonidae et des mysidacés. Des quantités moindres de polychètes sont ingérées par tous les stades
de vie benthiques, avec une consommation maximale chez les adultes (à 30-50 m de profondeur) et un
remplacement progressif des espèces carnivores par des espèces suspensivores au cours de la croissance.
La majorité des proies consommées par l'espèce (polychètes suspensivores et carnivores, amphipodes et
Crangonidae) étant susceptibles de réagir aux apports rhodaniens en MOP terrigène, on peut envisager
un effet positif des crues du Rhône sensible sur l'ensemble des stades de vie benthique d'A. laterna.
Cependant, compte-tenu de la répartition bathymétrique plus côtière des juvéniles de l'espèce, leurs
proies ne vont profiter que modérément des apports du fleuve en matière particulaire et la réponse aux
crues devrait essentiellement impliquer les adultes (les polychètes suspensivores dépendent
essentiellement, à 30-50 m de profondeur, de la MOP terrigène pour leur croissance). Comme pour B.
luteum, la réponse de la population d'A. laterna aux crues du fleuve va faire intervenir plusieurs pics
successifs de proies benthiques dont les intensités et les délais respectifs sont difficiles à prédire.
Cependant, on peut supposer que l'augmentation de nourriture au large du fleuve l'année de la crue va
favoriser la croissance et la condition des adultes, maximisant ainsi la reproduction de ces derniers et le
recrutement benthique de l'année suivante. Arnoglossus laterna étant essentiellement capturé à une taille
de 8-15 cm (Fisher et al., 1987), on peut s'attendre à un impact des crues sensible dès la deuxième année
suivant la crue. L'impact direct de la crue sur les stades juvéniles de l'espèce intervenant plus
tardivement, du fait des délais de réponses des polychètes carnivores, il est susceptible de concerner la

Discussion et conclusions générales
278
cohorte issue de la reproduction maximisée des adultes et d'ainsi accroître encore l'importance de cette
dernière. La première maturité sexuelle des individus d'A. laterna survenant à un âge de 1 à 2 ans dans
le golfe du Lion (Shéhata, 1984), l'impact de la reproduction de cette cohorte est susceptible de
maintenir l'augmentation du stock de l'espèce sur plusieurs années après la crue.
Citharus linguatula
L'alimentation de C. linguatula au large du Rhône est très stable et essentiellement basée sur
l'ingestion de poissons et de mysidacés. L'influence potentielle des apports rhodaniens en MOP
terrigène sur l'abondance de ces proies benthiques étant très réduite, il est peu probable que les crues du
fleuve aient un impact sensible sur la dynamique de population et la taille du stock de cette espèce, si ce
n'est par une augmentation de la production primaire à l'embouchure pendant la période de vie larvaire
de ses individus dans le cas de crues automnales exceptionnelles.
IV.3 - CONCLUSION GENERALE ET PERSPECTIVES
Par sa simplicité, le système rencontré au large du Rhône, avec uniquement deux principales
sources de matière organique de signatures isotopiques bien distinctes, a permis un suivi optimal du
devenir de la matière organique particulaire (MOP) terrigène au sein des écosystèmes marins côtiers.
Celle-ci, essentiellement présente au sein du sédiment, intègre les réseaux trophiques benthiques par le
biais de divers consommateurs primaires parmi lesquels les polychètes dépositivores tiennent une place
prépondérante. Elle est ensuite transférée jusqu'aux poissons soit par ingestion directe de ces organismes
soit par le biais de consommateurs secondaires s'en nourrissant.
Cette étude confirme l'efficacité de l'utilisation des isotopes stables du carbone et de l'azote pour
le suivi du transfert trophique de matière organique au sein des écosystèmes marins côtiers. Elle
souligne néanmoins l'existence de plusieurs exigences concernant leur utilisation dans le cas de chaînes
trophiques complexes et de variabilité spatiale importante du milieu. Le point principal soulevé
concerne la détermination précise des proies ingérées par les carnivores. Ce travail a permis de
confirmer que cette étape, souvent négligée lors des études portant sur les réseaux trophiques des
poissons, est en fait indispensable pour tirer un maximum d'informations des signatures isotopiques
obtenues. En effet, les variations spatio-temporelles et ontogéniques de l'alimentation des poissons et
des signatures isotopiques de leurs proies benthiques peuvent conduire à de graves erreurs
d'interprétation des signatures obtenues en cas d'étude basée sur des informations approximatives.
L'analyse combinée des contenus stomacaux des poissons (et plus généralement des carnivores) et des
compositions en isotopes stables (δ13C et δ15N) de la chair des organismes constitue par contre un outil
très puissant pour l'étude du devenir de la matière organique terrigène au sein des communautés marines
côtières sous influence continentale.

Discussion et conclusions générales
279
Cette étude a permis d'estimer l'intensité du transfert trophique de MOP terrigène vers les
juvéniles et les adultes des cinq espèces de poissons plats étudiées et d'émettre des hypothèses
concernant les réponses potentielles de leurs populations à une augmentation des apports du fleuve. Les
apports dissous du Rhône ne suivant que modérément les fluctuations du débit liquide, leur influence
positive sur la production primaire est relativement constante d'une année sur l'autre et intervient peu
dans les variations interannuelles d'abondance des poissons plats au large du fleuve. Ainsi, la réponse de
chaque population à une augmentation de débit du Rhône va essentiellement dépendre de l'intensité du
transfert de MOP terrigène vers les stades de vie benthique (juvéniles et/ou adultes) de l'espèce
concernée. Celle-ci dépend du régime alimentaire des poissons et notamment des quantités de
polychètes ingérées. Elle diffère fortement entre Solea solea, Solea impar, Buglossidium luteum,
Arnoglossus laterna et Citharus linguatula, du fait de l'existence d'importantes différences de
preferenda alimentaires entre ces cinq poissons plats. Elle varie également, pour chaque espèce, suivant
le stade de vie benthique (juvénile/adulte) et la tranche bathymétrique considérés, en raison de
modifications ontogéniques et spatio-temporelles de l'alimentation des poissons et de différences
bathymétriques du degré d'exploitation de la MOP terrestre par les communautés. Compte-tenu des
preferenda alimentaires, de la répartition spatio-temporelle et de l'adaptabilité trophique des poissons
étudiés, on peut s'attendre, en cas d'augmentation des apports du Rhône au niveau du fond, à un transfert
de MOP terrigène maximal chez l'ensemble des stades de vie benthique de S. solea et les adultes de B.
luteum, élevé chez les juvéniles de B. luteum et l'ensemble des stades de vie benthique d'A. laterna,
moyen chez l'ensemble des stades de vie benthique de S. impar et négligeable chez les adultes et les
juvéniles de C. linguatula. Les différences de sensibilité entre juvéniles et adultes de chaque espèce ont
d'importantes implications quant aux délais de réponse de leurs populations aux crues du Rhône. Les
réponses attendues diffèrent ainsi d'une espèce à l'autre en fonction du nombre de stades de vie
benthiques concernés et des délais de réponse des proies benthiques impliquées. Au vu des résultats
obtenus, on peut s'attendre à une réponse importante et durable des populations de S. solea, B. luteum,
A. laterna et S. impar mais à un impact négligeable des crues sur la population de C. linguatula.
Cependant, plusieurs zones d'ombre subsistent à ce niveau, notamment en ce qui concerne les
délais de réponse aux crues des organismes et l'effet de la pollution rhodanienne sur leur biologie. De
plus, les hypothèses émises sur la dynamique de population des poissons plats étudiés, bien qu'étayées
par de nombreux travaux préexistants, doivent être vérifiées au large du Rhône. Les recherches futures
devront ainsi s'attacher en priorité à l'étude de l'évolution de divers paramètres biologiques (croissance,
condition, fécondité, succès du recrutement, etc) des cinq espèces après une forte crue du fleuve, en
comparaison avec des périodes de débit faible ou moyen. L'impact potentiel de la pollution rhodanienne
sur les communautés benthiques à l'embouchure devra également être étudié afin de déterminer son
influence potentielle sur la dynamique de population des organismes macrobenthiques et des poissons.

Discussion et conclusions générales
280
Pour cela, la connaissance des réseaux trophiques benthiques apportée par ce travail de thèse sera d'une
aide précieuse pour le choix des organismes à analyser et la compréhension des concentrations
obtenues. Enfin, la connaissance des réseaux trophiques individualisés devra être affinée (notamment en
ce qui concerne les espèces de proies ingérées et leurs réactions potentielles au crues), avec pour
objectif ultime une modélisation du transfert de la matière organique terrigène au sein des écosystèmes
benthiques à l'embouchure du Rhône. Cette modélisation pourrait permettre, non seulement de prédire
les fluctuations interannuelles des stocks de poissons plats au large du fleuve, mais également d'estimer
le transfert de carbone d'origine rhodanienne vers les niveaux trophiques supérieurs et son "piégeage"
dans le compartiment benthique.

REFERENCES
BIBLIOGRAPHIQUES


281
BIBLIOGRAPHIE(Au total, 361 références bibliographiques)
Aarnio, K. et Bonsdorff, E. (1993). Seasonal variation in abundance and diet of the sand gobyPomatoschistus minutus (Pallas) in a northern Baltic archipelago. Ophelia, 37 (1): 19-30.
Abou-Hamdan, H. (2001). Gradient bathymétrique et influence des apports du Rhône sur la biodiversité,la densité et la biomasse du macrobenthos du plateau continental méditerranéen. Rapport deDEA, Univ. Aix-Marseille II. 32 p.
Allam, S. M. (1995). Feeding habits of soles (Solea vulgaris, Solea aegyptiaca, Solea impar and Soleakleini) from Abukir Bay, Southeastern Mediterranean Sea, Egypt. Bull. Nat. Inst. Oceanogr.Fish., A.R.E., 21 (2): 477-500.
Amara, R., Laffargue, P., Dewarumez, J. M., Maryniak, C., Lagardère, F. et Luczac, C. (2001). Feedingecology and growth of 0-group flatfish (sole, dab and plaice) on a nursery ground (SouthernBight of the North Sea). J. Fish Biol., 58: 788-803.
Amara, R. et Lagardère, F. (1995). Taille et âge au début de la métamorphose chez la sole (Solea solea(L.)) du golfe de Gascogne. ICES J. mar. Sci., 52 (2): 247-256.
Amara, R., Lagardère, F. et Désaunay, Y. (1993). Seasonal distribution and duration of the planktonicstage of Dover sole, Solea solea, larvae in the Bay of Biscay: an hypothesis. J. Fish Biol., 43(suppl. a): 17-30.
Amara, R., Poulard, J. C., Lagardère, F. et Désaunay, Y. (1998). Comparison between the life cycles oftwo Soleidae, the common sole, Solea solea, and the thickback sole, Michrochirus variegatus,in the Bay of Biscay (France). Envir. Biol. fishes, 53: 193-209.
Anderson, M. J. (2000). NPMANOVA: a fortran computer program for non-parametric multivariateanalysis of variance (for any two-factor ANOVA design) using permutation tests., Departmentof Statistics, University of Auckland: 16 p.
Anderson, M. J. (2001). Permutation tests for univariate or multivariate analysis of variance andregression. Can. J. Fish. aquat. Sci., 58: 636-639.
Anderson, M. J. et Legendre, P. (1999). An empirical comparison of permutation methods for test ofpartial regression coefficients in a linear model. J. Statist. Comput. Simul., 62: 271-303.
Ansel, A. D., Harvey, R. et Günther, C.-P. (1999). Recovery from siphon damage in Donax vittatus (DaCosta) (Bivalvia Donacidae). J. Moll. Stud., 65: 223-232.
Ansell, A. D., Comely, C. A. et Robb, L. (1999). Distribution, movements and diet of macrocrustaceanson a Scottish sandy beach with particular reference to predation on juvenile fishes. Mar. Ecol.Prog. ser., 176: 115-130.
Arcos, J. M., Ruiz, X., Bearhop, S. et Furness, R. W. (2002). Mercury levels in seabirds and their fishprey at the Ebro Delta (NW Mediterranean): the role of trawler discards as a source ofcontamination. Mar. Ecol. Prog. ser., 232: 281-290.
Ascensio, E., Bordreuil, C., Frasse, M., Orieux, A. et Roux, D. (1977). Une approche des conditionsmétéorologiques sur le golfe du Lion. Ann. Inst. Océanogr. Paris, 53: 155-169.
Avsar, D. (1994). Diel diet and feeding behavior of scaldfish (Arnoglossus laterna Walbaum, 1792) inthe Bay of Mersin. Acta adriat., 34 (1-2): 89-101.
Bailey, K. M., Canino, M. F., Napp, J. M., Spring, S. M. et Brown, A. L. (1995). Contrasting years ofprey levels, feeding conditions and mortality of larval walleye pollock Theragra chalcogrammain the western Gulf of Alaska. Mar. Ecol. Prog. ser., 119: 11-23.
Bailey, K. M. et Spring, S. M. (1992). Comparison of larval, age-0 juvenile and age-2 recruit abundanceindices of walleye pollock, Theragra chalcogramma, in the western Gulf of Alaska. ICES J.mar. Sci., 49: 297-304.

282
Bakun, A. et Agostini, V. N. (2001). Seasonal patterns of wind-induced upwelling/downwelling in theMediterranean Sea. Sci. mar., 65 (3): 243-257.
Barnes, R. D. (1965). Invertebrate zoology. W. B. Saunders Compagny, Philadelphia / London: 632 p.Barrie, A. et Prosser, S. J. (1996). Automated analysis of light element stable isotopes by isotope ratio
mass spectrometry. In: Boutton, T. W. et Yamasaki, S. (ed.), Mass Spectrometry of Soils,Marcel Dekker Inc, New-York, pp 1-46.
Batty, R. S. et Hoyt, R. D. (1995). The role of sense organs in the feeding behaviour of juvenile soleand plaice. J. Fish Biol., 47: 931-939.
Bekers, J.-M., Brasseur, P. et Nihoul, J. C. J. (1997). Circulation of the western Mediterranean: fromglobal to regional scales. Deep-sea Res.II, 44 (3-4): 531-549.
Belghyti, D., Aguesse, P. et Gabrion, C. (1993). Feeding ethology of Citharus linguatula andDicologlossa cuneata of the Moroccan Atlantic area. Vie Milieu, 43 (2-3): 95-108.
Belghyti, D., El Kharrim, K., Idelhaj, A., Menioui, M., Bouchereau, J.-L., Sorbe, J.-C. et Ahami, A.(1995). Inventaire zoologique par analyse des contenus stomacaux de deux espèces de poissonspleuronectiformes du littoral à Casablanca et Mehdia (Maroc). Bullet. Inst. Sci. Rabat, 19: 103-110.
Benouada, H. (1985). Exploitation halieutique partagée (Dorade, Loup, Muges, Sars et Soles)Interaction entre pêcheries marines et lagunaires du quartier de Sète. Thèse : Univ. MontpellierII. 218 p + annexes.
Berner, R. A. (1971). Principles of chemical sedimentology. Franck Press: 231 p.Beukema, J. J. (1991). Changes in composition of bottom fauna of a tidal-flat area during a period of
eutrophication. Mar. Biol., 111: 293-301.Beyst, B., Cattrijsse, A. et Mees, J. (1999). Feeding ecology of juvenile flatfishes of the surf zone of a
sandy beach. J. Fish Biol., 55: 1171-1186.Bianchi, G., Gislason, H., Graham, K., Hill, L., Jin, X., Koranteng, K., Manickchand-Heileman, S.,
Payá, I., Sainsbury, K., Sanchez, F. et Zwanenburg, K. (2000). Impact of fishing on sizecomposition and diversity of demersal fish communities. ICES J. mar. Sci., 57: 558-571.
Blanc, F. et Leveau, M. (1971). Plancton et eutrophie : aire d'épandage rhodanienne et golfe de Fos(traitement mathématique des données). Thèse : Univ. Aix-Marseille II. 681 p. + annexes.
Blanc, F., Leveau, M. et Szekielda, K. H. (1969). Effets eutrophiques au débouché d'un grand fleuve(Grand Rhône). Mar. Biol., 3: 233-242.
Blanc, J. J. (1974). La sédimentation sur le précontinent de Provence et ses modalités. Comparaisonavec d'autres secteurs de la Méditerranée. Bull. B.R.G.M. 2è série. Section IV, 3: 206-211.
Blanchard, F. (2001). Une approche de la dynamique des peuplements de poissons démersaux exploités: analyse comparée de la diversité spécifique dans le golfe de Gascogne (océan Atlantique) etdans le golfe du Lion (mer Méditerranée). Aquat. Living Resour., 14: 29-40.
Blanchard, F. et Boucher, J. (2001). Temporal variability of total biomass in harvested communities ofdemersal fishes. Fisheries Research, 49: 283-293.
Blondel, J. (1995). Biogéographie. Approche écologique et évolutive. Masson, Paris: 297 p.Bocquene, G., Galgani, F., Burgeot, T., Le Dean, L. et Truquet, P. (1993). Acetylcholinesterase levels in
marine organisms along French coasts. Mar. Pollut. Bull., 26 (2): 101-106.Bodoy, A. et Plante-Cuny, M.-R. (1980). Evaluation simultanée des biomasses et productions primaires
phytoplanctonique et microphytobenthique en milieu côtier. C.R. Acad. Sci. Paris, 290 (D):667-670.
Bolle, L. J., Rijnsdorp, A. D. et Van der Veer, H. W. (2001). Recruitment variability in dab (Limandalimanda) in the southeastern North Sea. J. Sea Res., 45 (3-4): 255-270.

283
Borsa, P. et Quignard, J.-P. (2001). Systematics of the Atlantic-Mediterranean soles Pegusa impar, P.lascaris, Solea aegyptiaca, S. senegalensis ans S. solea (Pleuronectiformes: Soleidae). Can. J.Zool., 79: 2297-2302.
Bouilllon, S., Chandra Mohan, P., Sreenivas, N. et Dehairs, F. (2000). Sources of suspended organicmatter and selective feeding by zooplankton in an estuarine mangrove ecosystem as traced bystable isotopes. Mar. Ecol. Prog. ser., 208: 79-92.
Bouloubassi, I., Lipiatou, E., Saliot, A., Tolosa, I., Bayona, J. M. et Albaiges, J. (1997). Carbon sourcesand cycle in the western Mediterranean - the use of molecular markers to determine the originof organic matter. Deep-sea Res. II, 44 (3-4): 781-799.
Bourquard, C. (1985). Structure et mécanismes de mise en place, de maintien et d'évolution despeuplements ichthyiques lagunaires du Golfe du Lion. Thèse de 3ème cycle, Univ. Montpellier II.337 p.
Boutton, T. W. (1991). Stable carbon isotope ratios of natural materials: II. Atmospheric, terrestrial,marine, and freshwater environments. In: Coleman, D. C. et Fry, B. (ed.), Carbon isotopestechniques, Academic Press, San Diego, pp 173-185.
Braber, L. et De Groot, S. J. (1973). The food of five flatfish species (Pleuronectiformes) in the northernNorth Sea. Neth. J. sea res., 6: 163-172.
Bricout, J., Knoepffler-Peguy, M., Pergent, G. et Boudouresque, C.-F. (1990). Le rapport 13C/12C chezquelques végétaux marins de Méditerranée. Posidonia Newsletter, 3 (2): 19-21.
Broche, P., Devenon, J.-L., Forget, P., Maistre, J.-C., Naudin, J.-J. et Cauwet, G. (1998). Experimentalstudy of the Rhone plume. Part I: physics and dynamics. Oceanol. Acta, 21 (6): 725-735.
Bromley, P. J. (2000). Growth, sexual maturation and spawning in central North Sea plaice(Pleuronectes platessa L.), and the generation of maturity ogives from commercial catch data. J.Sea Res., 44: 27-43.
Bromley, P. J., Ravier, C. et Witthames, P. R. (2000). The influence of feeding regime on sexualmaturation, fecundity and atresia in first-time spawning turbot. J. Fish Biol., 56: 264-278.
Buffan-Dubau, E., De Wit, R. et Castel, J. (1996). Feeding selectivity of the harpacticoid copepodCanuella perplexa in benthic muddy environments demonstrated by HPLC analyses of chlorinand carotenoid pigments. Mar. Ecol. Prog. ser., 137 (1-3): 71-82.
Burgeot, T., Bocquene, G., Pingray, G., Godefroy, D., Legrand, J., Dimeet, J., Marco, F., Vincent, F.,Henocque, Y., Oger Jeanneret, H. et Galgani, F. (1994). Monitoring biological effects ofcontamination in marine fish along french coast by measurement of ethoxyresorufin-O-deethylase activity. Ecotoxicol. Envir. Saf., 29: 131-147.
Cabana, G. et Rasmussen, J. B. (1994). Modelling food chain structure and contaminantbioaccumulation using stable nitrogen isotopes. Nature, 372: 255-257.
Cabana, G. et Rasmussen, J. B. (1996). Comparing aquatic food chains using nitrogen isotopes. Proc.Nat. Acad. Sci. U.S.A., 93: 10844-10847.
Caddy, J. F. (1993). Towards a comparative evaluation of human impacts on fishery ecosystems ofenclosed and semi-enclosed seas. Rev. Fish Sci., 1: 57-95.
Caddy, J. F. (2000). Marine catchment basin effects versus impacts of fisheries on semi-enclosed seas.ICES J. mar. Sci., 57: 628-640.
Campillo, A., Aldebert, Y., Bigot, J.-L. et Liorzou, B. (1989). Données sur la distribution desprincipales espèces commerciales du Golfe du Lion. Rapports internes de la Direction desRessources Vivantes de l'IFREMER (DRV-89.041-RH/Sète): 175 p.
Campillo, A. et Bigot, J. L. (1992). Les pêcheries françaises de Méditerranée. Synthèse desconnaissances. IFREMER-CEE (92/1211625/TF): 206 p.
Camusso, M., Balestrini, R., Martinotti, W. et Arpini, M. (1999). Spatial variations in trace metal andstable isotope content of autochtonous organisms and sediments in the river Po. J. Aqua.Ecosyst. Health Manag., 2: 93-53.

284
Camusso, M., Martinotti, W., Balestrini, R. et Guzzy, L. (1998). C and N stable isotopes and tracemetals in selected organisms from the river Po delta. Chemosphere, 37 (14-15): 2911-2920.
Cao, Y., Bark, A. W. et Williams, W. P. (1996). Measuring the response of macroinvertebratecommunities to water pollution: a comparison of multivariate approaches, biotic and diversityindices. Hydrobiologia, 341: 1-19.
Cardinale, M. et Arrhenius, F. (2000). The relationship between stock and recruitment: are theassumptions valid? Mar. Ecol. Prog. ser., 196: 305-309.
Carter, C. G., Grove, D. J. et Carter, D. M. (1991). Trophic resource partitioning between twocoexisting flatfish species of the north coast of Anglesey, North Wales. Neth. J. sea res., 27(3/4): 325-335.
Castel, J. et Lasserre, P. (1982). Régulation biologique du meiobenthos d'un écosystème lagunaire parun alevinage expérimental en soles (Solea vulgaris). Oceanologica acta, Actes SymposiumInternational sur les lagune côtières SCOR/IABO/UNESCO, Bordeaux, 8-14 septembre1981: 243-251.
Cauwet, G. (1996). Apports du Rhône en carbone et azote dissous et particulaire de juin 1994 à juin1995. Colloque Potam'mes. 7es Rencontres de l'Agence Regionale pour l'EnvironnementProvence-Alpes-Côte d'Azur, Digne-les-Bains (ed.): 263-271.
Cauwet, G., Gadel, F., De Souza Sierra, M. M., Donard, O. et Ewald, M. (1990). Contribution of theRhône River to organic carbon inputs to the northwestern Mediterranean Sea. Cont. shelf Res.,10 (9-11): 1025-1037.
Cauwet, G., Miller, A., Brasse, S., Fengler, G., Mantoura, R. F. C. et Sptzy, A. (1997). Dissolved andparticulate organic carbon in the western Mediterranean Sea. Deep-sea Res.II, 44 (3-4): 769-779.
Charles, F., Amouroux, J. M. et Gremare, A. (1999). Comparative study of the utilization of bacteriaand microalguae by the suspension-feeding bivalve: Callista chione. J. mar. Biol. Assoc. U.K.,79 (4): 577-584.
Charmasson, S. (1998). Cycle du combustible nucléaire et milieu marin. Devenir des effluentsrhodaniens en Méditerranée et des déchets immergés en Atlantique Nord-Est. Thèse : Univ.Aix-Marseille II. 365 p. + annexes.
Charmasson, S., Barker, E., Calmet, D., Pruchon, A.-S. et Thébault, H. (1999). Long-term variations ofman-made radionuclide concentrations in a bio-indicator Mytilus galloprovincialis from theFrench Mediterranean coast. Sci. total Environ., 237/238: 93-103.
Charmasson, S., Radakovitch, O., Arnaud, M., Bouisset, P. et Pruchon, A.-S. (1998). Long-core profilesof 137Cs, 134Cs, 60Co and 210Pb in sediment near the Rhône River (Northwestern MediterraneanSea). Estuaries, 21 (3): 367-378.
Chassefiere, B. (1990). Mass-physical properties of surficial sediments on the Rhône continentalmargin: implications for the nepheloid benthic layer. Cont. shelf Res., 10: 857-867.
Chong, V. C., Low, C. B. et Ichikawa, T. (2001). Contribution of mangrove detritus to juvenile prawnnutrition: a dual stable isotope study in a Malaysian mangrove forest. Mar. Biol., 138: 77-86.
Clarke, K. R. (1993). Non-parametric multivariate analyses of changes in community structure. Austr. J.Ecol., 18: 117-143.
Cohen, J. E., Pimm, S. L., Yodzis, P. et Saldana, J. (1993). Body size of animal predators and animalprey in food webs. J. anim. Ecol., 62 (67-78):
Conan, P., Pujo-Pay, M., Raimbault, P. et Leveau, M. (1998). Variabilité hydrologique et biologique duGolfe du Lion. II. Productivité sur le bord interne du courant. Oceanol. Acta, 21 (6): 767-781.
Cooper, L. W. et De Niro, M. J. (1989). Stable carbon isotope variability in the sea grass Posidoniaoceanica: evidence for light intensity effect. Mar. Ecol. Prog. Ser., 50: 225-229.
Coste, B. et Raimbault, P. (1993). Recent data on the nutrient input into the Mediterranean Sea by theRhone River. Water pollut. Res. Rep., 30: 47-51.

285
Couch, C. A. (1989). Carbon and nitrogen stable isotopes of meiobenthos and their food resources.Estuar. coast. Shelf Sci., 28: 433-441.
Cruzado, A. et Velasquez, Z. R. (1990). Nutrients and phytoplankton in the Gulf of Lions, northwesternMediterranean. Cont. shelf Res., 10 (9-11): 931-942.
Cushing, D. H. (1982). Climate and fisheries. Academic Press, New-York: 373 p.Cushing, D. H. (1995). The long-term relationship between zooplankton and fish. ICES J. mar. Sci., 52:
611-626.Dachs, J., Bayona, J. M. et Albaiges, J. (1997). Spatial distribution, vertical profiles and budget of
organochlorine compounds in Western Mediterranean seawater. Mar. Chem., 57: 313-324.Darnaude, A. M. (1999). Relations proies-prédateurs et compétition alimentaire chez deux Soléidés du
Golfe de Fos : Solea vulgaris Quensel et Buglossidium luteum (Risso). Rapport de DEA, Univ.Aix-Marseille II. 40 p.
Darnaude, A. M., Harmelin-Vivien, M. L. et Salen-Picard, C. (2001). Food partitioning among flatfish(Pisces: Pleuronectiforms) juveniles in a mediterranean coastal shallow sandy area. J. mar. Biol.Assoc. U.K, 81: 119-127.
Dauby, P. (1989). The stable carbon isotope ratios in benthic food webs of the Gulf of Calvi, Corsica.Cont. shelf Res., 9 (2): 181-195.
Dauby, P. (1995). A δ13C study of the feeding habits in four mediterranean Leptomysis species(Crustacea: Mysidacea). P.S.Z.N.I.: Mar. Ecol., 16 (2): 93-102.
Dauby, P. et Mosora, F. (1988). Analyse à l'aide des isotopes stables du carbone des sources de nutritiondes détritivores et suspensivores benthiques. Bull. Soc. Roy. Liège, 57 (4-5): 241-248.
Day, J. W., Hall, C. S. A., M., K. et Yanez-Arancibia, A. (1989). Estuarine Ecology. John Wiley &Sons, New-York: 558 p.
De Groot, S. J. (1969). Digestive system and sensorial factors in relation to the feeding behaviour offlatfish (Peuronectiformes). J. Cons. int. Explor. Mer, 32 (3): 385-395.
De Groot, S. J. (1971). On the interrelationship between morphology of the alimentary tract, food andfeeding behaviour in flatfishes (Pisces: Pleuronectiformes). Neth. J. sea res., 5 (2): 121-196.
De Niro, M. J. et Epstein, S. (1978). Influence of diet on the distribution of carbon isotopes in animals.Geochim. Cosmochim. Acta, 42: 495-506.
De Niro, M. J. et Epstein, S. (1981). Influence of diet on the distribution of nitrogen isotopes in animals.Geochim. Cosmochim. Acta, 45: 341-351.
Deegan, L. A. et Garritt, R. H. (1997). Evidence for spatial variability in estuarine food webs. Mar.Ecol. Prog. ser., 147: 31-47.
Demarcq, H. (1985). Applications de la télédétection infra-rouge et visible en océanographie: étude dela zone rhodanienne, observations des zones de production dans le Golfe du Lion et estimationde l'éclairement solaire global en Méditerranée occidentale. Thèse 3e cycle, Univ. Aix-MarseilleII. 226 p.
Deniel, C. (1981). Les poissons plats (Téléostéens, Pleuronectiformes) en baie de Douarnenez.Reproduction, croissance et migration des Bothidae, Scophthalmidae, Pleuronectidae etSoleidae. Thèse 3e cycle, Univ. de Bretagne Occidentale. 476 p.
Deniel, C. (1983). La reproduction des poissons plats (Téléostéens - Pleuronectiformes) en baie deDouarnenez. 1. Cycles sexuels et fécondité des arnoglosses Arnoglossus thori, A. laterna, Aimpérialis (Bothidae). Cah. Biol. Mar., 24: 231-252.
Deniel, C. (1984). Relations entre l'activité reproductrice et la croissance chez les poissons plats de laBaie de Douarnenez. Cybium, 8 (1): 83-93.
Deniel, C., Le Blanc, C. et Rodriguez, A. (1989). Comparative study of sexual cycles, oogenesis andspawning of two Soleidae, Solea lascaris (Risso, 1810) and Solea impar (Bennett, 1831), on thewestern coast of Brittany. J. Fish Biol., 35 (1): 49-58.

286
Denis, L. (1999). Dynamique des flux d'oxygène et de sels nutritifs à l'interface eau-sédiment sur lamarge continentale du Golfe du Lion (Méditerranée Nord-Occidentale). Thèse : Univ. Aix-Marseille II. 193 p. + annexes.
Denis, L., Grenz, C., Alliot, E. et Rodier, M. (2001). Temporal variability in dissolved inorganicnitrogen fluxes at the sediment-water interface and related annual budget on a continental shelf(NW Mediterranean). Oceanol. Acta, 24 (1): 85-97.
Dittel, A. I., Epifanio, C. E., Schwalm, S. M., Fantle, M. S. et Fogel, M. L. (2000). Carbon and nitrogensources for juvenile blue crabs Callinectes sapidus in coastal wetlands. Mar. Ecol. Prog. ser.,194: 103-112.
Dorten, W., Elbaz-Poulichet, F., Mart, L. et Martin, J.-M. (1991). Reassessment of the river input oftrace metals into the Mediterranean Sea. Ambio, 20: 2-6.
Dufour, E. et Gerdeaux, D. (2001). Apport des isotopes stables (13C/12C, 15N/14N, 18O/16O, 36S/34S,87Sr/86Sr) aux études écologiques sur les poissons. Cybium, 25 (4): 369-382.
Dunton, C., Le Gall, S., Hartmann, H. J. et Bréret, M. (1999). Retention of ciliates and flagellates by theoyster Crassostrea gigas in French Atlantic coastal ponds: protists as a trophic link betweenbacterioplankton and benthic suspension-feeders. Mar. Ecol.Prog. Ser., 177: 165-175.
Dupuy, K. H. et Shell, D. M. (1987). Dependance of consumers on macroalgal (Laminaria solidungula)carbon in an artic kelp community: δ13C evidence. Mar. Biol., 93: 615-625.
Durrieu de Madron, X., Abassi, A., Heussner, S., Monaco, A., Aloisi, J. C., Radakovitch, O., Giresse,P., Buscail, R. et Kerhervé, P. (2000). Particulate matter and organic carbon budgets for theGulf of Lions (NW Mediterranean). Oceanol. Acta, 23 (6): 717-730.
Elbaz-Poulichet, F., Garnier, J.-M., Guan, D. M., Martin, J.-M. et Thomas, A. J. (1996). Theconservative behaviour of trace metals (Cd, Cu, Ni and Pb) and As in the surface plume ofstratified estuaries: example of the Rhone River (France). Estuar. coast. shelf Sci., 42: 289-310.
Evans, S. (1983). Production, predation and food niche segregation in a marine shallow soft-bottomcommunity. Mar. Ecol. Prog. ser., 10: 147-157.
Eyrolle, F. et Charmasson, S. (2001). Distribution of organic carbon, selected stable elements andartificial radionuclides among dissolved, colloidal and particulate phases in the Rhône River(France): Preliminary results. J. environ. Radioact., 55: 145-155.
Farrugio, H. et Le Corre, G. (1987). Etude pour une gestion optimale des pêcheries démersalesinteractives dans le Golfe du Lion. Rapport IFREMER (DRV-87-015/RH/Sète): 180 p.
Fauchald, K. et Jumars, P. A. (1979). The diet of worms : a study of polychaete feeding guilds.Oceanogr. Mar. Biol. Ann. Rev., 17: 193-284.
Faure, G. (1986). Principles of Isotope geology. 2nd Edition. J. Wiley, New-York: 589 p.Febvre-Chevalier, C. (1969). Etude bionomique des substrats meubles dragables du Golfe de Fos.
Téthys, 1 (2): 421-476.Fernex, F. E., Migon, C. et Chisholm, J. R. M. (2001). Entrapment of pollutants in Mediterranean
sediments and biogeochemical indicators of their impact. Hydrobiologia, 450: 31-46.Fichez, R., Dennis, P., Fontaine, M. F. et Jickells, T. D. (1993). Isotopic and biochemical composition
of particulate organic matter in a shallow water estuary (Great Ouse, North Sea, England). Mar.Chem., 43: 263-276.
Fisher, W., Schneider, M. et Bauchot, M.-L. (1987). Fiches FAO d'identification des espèces pour lesbesoins de la pêche. Méditerranée et Mer Noire (Zone de pêche 37). Révision 1, 2 (vertébrés).FAO, Rome: 769 (761-1530) p.
Fonds, M. (1975). The influence of temperature and salinity on growth of young sole Solea solea L..10th European Symposium on Mar. Biol., Ostend, Belgium (1975). Persoone, G. et Jaspers, E.(ed.), Univers Press, Wetteren: 109-125.

287
Fonds, M. (1979). Laboratory Observations on the Influence of Temperature and salinity onDevelopment of the Eggs and Growth of the Larvae of Solea solea. Mar. Ecol. Prog. ser., 1: 91-99.
Fonds, M., Cronie, R., Vethaak, A. D. et Van der Puyl, P. (1992). Metabolism, food consumption andgrowth of plaice (Pleuronectes platessa) and flounder (Platichthys flesus) in relation to fish sizeand temperature. Neth. J. sea res., 29 (1-3): 127-143.
Fonds, M. et Saksena, V. P. (1977). The daily food intake of young soles (Solea solea L.) in relation totheir size and the water temperature. 3rd Meeting of the ICES Working Group on Mariculture,Brest, France (ed.), Actes de Colloques CNEXO: 51-58.
Fontugne, M. R. et Duplessy, J. C. (1981). Organic carbon isotopic fractionation by marine plankton inthe temperature range -1 to 31°C. Oceanol. Acta, 4 (1): 85-90.
Fourqurean, J. W., Moore, T. O., Fry, B. et Hollibaugh, J. T. (1997). Spatial and temporal variation inC:N:P ratios, δ15N, and δ13C of eelgrass Zostera marina as indicators of ecosystem processes,Tomales Bay, California, USA. Mar. Ecol. Prog. ser., 157: 147-157.
Frid, C. L. J., Buchanan, J. B. et Garwood, P. R. (1996). Variability and stability in benthos: twenty-twoyears of monitoring off Northumberland. ICES J. mar. Sci., 53: 978-980.
Froese, R. et Pauly, D. (2000). Fishbase 2000. Concepts, design and data sources. 4CD-ROMs (updateson http://www.fishbase.org). International Centre for Living Aquatic Resource Management,Los Banos, Philippines.
Fry, B., Anderson, R. K., Entzeroth, L., Bird, J. L. et Parker, P. L. (1984). 13C enrichment and oceanicfood web structure in the northern gulf of Mexico. Contrib. mar. Sci., 27: 49-63.
Fry, B. et Wainright, S. C. (1991). Diatom sources of 13C-rich carbon in marine food webs. Mar. Ecol.Prog. ser., 76 (2): 149-157.
Gaertner, J. C. (1997). Organisation des assemblages démersaux dans le golfe du Lion : structuresspatiales et stabilité temporelle. Thèse : Univ. Aix-Marseille II. 139 p. + annexes.
Gaertner, J. C. (1999). Spatial structure and habitat associations of demersal assemblages in the Gulf ofLions : a multicompartmental approach. Mar. Biol., 135: 199-208.
Gaertner, J. C. (2000). Seasonal organization patterns of demersal assemblages in the Gulf of Lions(north-western Mediterranean sea). J. mar. Biol. Assoc. U.K., 80: 777-783.
Garcia-Rodriguez, M. et Eesteban, A. (2000). Contribution to the knowledge of Citharus linguatula(Linnaeus, 1758) (Osteicthyes: Heterosomata) in the Iberian Mediterranean. Conférence :Ressources halieutiques en Méditerranée, Pisa (Italy), (ed.), Actes de Colloques IFREMER.Plouzané (France): 131-140.
Gaudy, R., Bianchi, M., Pagano, M. et Soto, Y. (1996). Cross frontal variability in hydrological andbiological structures observed in a river plume area (Rhone mouth, NW Mediterranean Sea).Hydrobiologia, 324 (2): 131-140.
Gautier, Y. (1957). Recherches sur les biocénoses benthiques des côtes de Camargue et du Golfe deFos. Rec. Trav. Stn mar. Endoume, 22: 55-64.
Gearing, G. N., Gearing, P. L., Rudnick, D. T., Requejo, A. G. et Hutchins, M. J. (1984). Isotopevariability of organic carbon in a phytoplancton-based temperate estuary. Geochim. Cosmochim.Acta, 48: 1089-1098.
Gerking, S. D. (1994). Feeding Ecology of Fish. Academic Press, San Diego, California: 416 p.Gibson, R. N. (1994). Impact of habitat quality and quantity on the recruitment of juvenile flatfishes.
Neth. J. sea res., 32 (2): 191-206.Gibson, R. N. et Ezzi, I. A. (1980). The biology of the scaldfish Arnoglossus laterna (Walbaum) on the
west coast of Scotland. J. Fish Biol., 17: 565-575.Gibson, R. N. et Ezzi, I. A. (1987). Feeding relationships of a demersal fish assemblage on the west
coast of Scotland. J. Fish Biol., 31: 55-69.

288
Giovanardi, O. et Piccinetti, C. (1981). Biology and fishery of the yellow sole, Solea lutea (Risso) 1810,in the Adriatic Sea. FAO Fish. Rep., 253: 101-113.
Giovanardi, O. et Piccinetti, C. (1984). Biology and fishery aspects of the scaldfish Arnoglossus laterna(Walbaum) 1792, in the Adriatic Sea. FAO Fish. Rep., 290: 161-166.
Gorokhova, E. et Hansson, S. (1999). An experimental study on variations in stable carbon and nitrogenisotope fractionation during growth of Mysis mixta and Neomysis integer. Can. J. Fish. aquat.Sci., 56: 2203-2210.
Greig-Smith, P. (1983). Quantitative Plant Ecology. Third edition. Blackwell Scientific Publications,Oxford: 359 p.
Griffin, M. P. A. et Valiela, I. (2001). δ15N isotope studies of life history and trophic position ofFundulus heteroclitus and Menidia menidia. Mar. Ecol. Prog. ser., 214: 299-305.
Guieu, C., Martin, J.-M., Thomas, A. J. et Elbaz-Poulichet, F. (1991). Atmospheric versus river inputsof metals to the Gulf of Lions: total concentrations, partitioning and fluxes. Mar. Pollut. Bull.,22 (4): 176-183.
Haines, E. B. (1976). Stable carbon isotope ratios in the biota, soils, and tidal water of a Georgia saltmarsh. Estuar. coast. mar. Sci., 4: 609-616.
Haines, E. B. et Montague, C. l. (1979). Food sources of estuarine invertebrates analysed using 13C/12Cratios. Ecology, 60: 48-56.
Hamerlynck, O. et Cattrijsse, A. (1994). The food of Pomatoschistus minutus (Pisces, Gobiidae) inBelgian coastal waters, and a comparison with the food of its potential competitor P. lozanoi. J.Fish Biol., 44: 753-771.
Harmelin-Vivien, M. L. (1979). Ichtyofaune des récifs coralliens de Tuléar (Madagascar) : écologie etrelations trophiques. Thèse d'Etat, Univ. Aix-Marseille II. 151 p.
Harmelin-Vivien, M. L. (1981). Description d'un petit chalut à perche pour récolter la faune vagile desherbiers de Posidonies. Rapp. Comm. int. Mer Medit., 27 (5): 199-200.
Harmelin-Vivien, M. L., Kaim-Malka, R. A., Ledoyer, M. et Jacob-Abraham, S. S. (1989). Foodpartitioning among scorpaenid fishes in Mediterranean seagrass beds. J. Fish Biol., 34: 715-734.
Harrigan, P., Zieman, J. C. et Macko, S. A. (1989). The base of nutritional support for the gray snapper(Lutjanus griseus): an evaluation based on a combined stomach content and stable isotopeanalysis. Bull. mar. Sci., 44: 65-77.
Harvey, C. J. et Kitchell, J. F. (2000). A spatial isotope evaluation of the structure and spatialheterogeneity of a Lake Superior food web. Can. J. Fish. aquat. Sci., 57: 1395-1403.
Hellings, L., Dehairs, F., Tackx, M., Keppens, E. et Baeyens, W. (1999). Origin and fate of organiccarbon in the freshwater part of the Scheldt Estuary as traced by stable carbon isotopecomposition. Biogeochemistry, 47: 167-186.
Herman, P. J. M., Middelburg, J. J., Widdows, J., Lucas, C. H. et Heip, C. H. R. (2000). Stable isotopesas trophic tracers: combining field sampling and manipulative labelling of food resources formacrobenthos. Mar. Ecol. Prog. ser., 204 (79-92):
Hesslein, R. H., Capel, M. J., Fox, D. E. et Hallard, K. A. (1991). Stable isotopes of Sulfur, Carbon, andNitrogen as indicators of trophic level and fish migration in the lower Mackenzie River basin,Canada. Can. J. Fish. aquat. Sci., 48: 2258-2265.
Hesslein, R. H., Hallard, K. A. et Ramlal, P. (1993). Replacement of Sulfur, Carbon, and Nitrogen inTissue of Growing Broad Whitefish (Coregonus nasus) in Response to a Change in Diet Tracedby δ34S, δ13C, and δ15N. Can. J. Fish. aquat. Sci, 50: 2071-2076.
Horwood, J. W. (1993). The Bristol Channel Sole (Solea solea (L.)) : A Fisheries Case Study. Adv. mar.Biol., 29: 215-367.
Horwood, J. W. et Greer Walker, M. (1989). The effects of feeding level on the fecundity of plaice(Pleuronectes platessa). J. mar. Biol. Assoc. U.K., 69 (81-92):

289
Hsieh, H.-L., Kao, W.-Y., Chen, C.-P. et Liu, P.-J. (2000). Detrital flows through the feeding pathwayof the oyster (Crassostrea gigas) in a tropical lagoon: δ13C signals. Mar. Biol., 136: 677-684.
Hureau, J. C. (1970). Biologie comparée de quelques poissons antarctiques (Nototheniidae). Bull. Inst.Océanogr. Monaco, 68 (1391): 244.
Hyslop, E. J. (1980). Stomach content analysis : a review of methods and their application. J. Fish Biol.,17: 411-430.
Incze, L. S., Mayer, L. M., Evelyn, B. S. et Macko, S. A. (1982). Carbon inputs to Bivalve Mollusks: acomparison of two estuaries. Can. J. Fish. aquat. Sci., 39: 1348-1352.
Jardas, I. (1984). Citharus macrolepidotus (Bloch, 1787) (Pisces, Pleuronectiformes) III - Nourishmentand the length-weight relationship in the Adriatic Sea. Ichthyologia, 16 (1-2): 1-4.
Jenkins, G. P. (1987). Comparative diets, prey selection, and predatory impact of co-occuring larvae oftwo flounder species. J. Exp. Mar. Biol. and Ecol., 110: 147-170.
Jennings, S., Renones, O., Morales-Nin, B., Polunin, N. V. C., Moranta, J. et Coll, J. (1997). Spatialvariation in the 15N and 13C stable isotope composition of plants, invertebrates and fishes onMediterranean reefs: implications for the study of trophic pathways. Mar. Ecol. Prog. ser., 146:109-116.
Josefson, A. B. et Conley, D. J. (1997). Benthic response to a pelagic front. Mar. Ecol. Prog. ser., 147:49-62.
Josefson, A. B., Jensen, J. N. et Ærtebjerg, G. (1993). The benthos community structure anomaly in thelate 1970s and early 1980s - a result of major food pulse? J. exp. mar. Biol. Ecol., 172: 31-45.
Kaehler, S., Pakhamov, E. A. et McQuaid, C. D. (2000). Trophic structure of the marine food web at thePrince Edward Islands (Southern Ocean) determined by δ13C and δ15N analysis. Mar. Ecol.Prog. ser., 208: 13-20.
Keast, A. (1978). Trophic and spatial interrelationships in the fish species of an Ontario temperate lake.Envir. Biol. fishes., 3: 7-31.
Kenkel, N. C. et Orloci, L. (1986). Applying metric and nonmetric multidimensional scaling to someecological studies: some new results. Ecology, 67: 919-928.
Kerhervé, P., Minagawa, M., Heussner, S. et Monaco, A. (2001). Stable isotopes (13C/12C and 15N/14N)in settling organic matter of the northwestern Mediterranean Sea: biogeochemical implications.Oceanol. Acta, 24 (Suppl.): S77-S85.
Kerr, S. R. et Ryder, R. A. (1992). Effects of cultural eutrophication on coastal marine fisheries: acomparative approach. In: Vollenweider, R. A., Marchetti, R. et Viviani, R. (ed.), Marinecoastal eutrophication: the response of marine transitional systems to human impact: problemsand perspectives for restoration, Elsevier Science Publishers B. V., Amsterdam, pp 599-614.
Kichuki, E. et Wada, E. (1996). Carbon and nitrogen stable isotope ratios of deposit-feeding polychaetesfrom the Nanakita River estuary, Japan. Hydrobiologia, 321 (1): 69-75.
Kirchman, D., Soto, Y., Van Wambeck, F. et Bianchi, M. (1989). Bacterial production in the RhôneRiver plume: effect of mixing on relationships among microbial assemblages. Mar. Ecol. Prog.ser., 53: 267-275.
Kiriluk, R. M., Servos, M. R., Whittle, D. M., Cabana, G. et Rasmussen, J. B. (1995). Using ratios ofstable nitrogen and carbon isotopes to characterize the biomagnification of DDE, mirex, andPCB in a Lake Ontario pelagic food web. Can. J. Fish. aquat. Sci., 52: 2660-2674.
Kiyashko, S. I., Kharlamenko, V. I. et Imbs, A. B. (1998). Stable isotope ratios and fatty acids as foodsource markers of deposit feeding invertebrates. Russian J. mar. Biol., 24 (3): 170-174.
Klein Breteler, W. C. M., Grice, K., Schouten, S., Kloosterhuis, H. T. et Sinninghe Damsté, J. S. (2002).Stable carbon isotope fractionantion in the marine copepod Temora longicornis: unexpectedlylow δ13C value of faecal pellets. Mar. Ecol. Prog. ser., 240: 195-204.

290
Kline, T. C. J. et Pauly, D. (1998). Cross-validation of trophic level estimates from a mass balancemodel of Prince William Sound using 15N/14N data. In: Funk, F., Quinn, T. J., Heifetz, J.,Ianelli, J. N., Powers, J. E., Sullivan, P. J. et Zhang, C. I. (ed.), Proceedings of the InternationalSymposium on Fishery Stock Assessment Models, Alaska Sea Grant College Report, pp 98-101.
Knox, G. A. (1986). Estuarine ecosystems: a systems approach. Vol. II. CRC Press, Boca Raton,Florida: 230 p.
Koutsikopoulos, C., Dorel, D. et Desaunay, Y. (1995). Movement of sole (Solea solea) in the Bay ofBiscay: coastal environment and spawning migration. J. mar. Biol. Assoc. U.K., 75: 109-126.
Kruskal, J. B. (1964). Multi-dimensional scaling by optimizing goodness of fit to a non-parametrichypothesis. Psychometrica, 29: 1-27.
Krüuk, H. (1963). Diurnal periodicity in the activity of the common sole, Solea vulgaris Quensel. Neth.J. sea res., 2 (1): 1-28.
Kuipers, B. R., De Wilde, P. A. W. J. et Creutzberg, F. (1981). Energy flow in a tidal flat ecosystem.Mar. Ecol. Prog. ser., 5: 215-221.
Kwak, T. J. et Zedler, J. B. (1997). Food web analysis of southern California coastal wetlands usingmultiple stable isotopes. Oecologia, 110: 262-277.
Labropoulou, M. et Markakis, G. (1998). Morphological-dietary relationships within two assemblagesof marine demersal fishes. Envir. Biol. fishes, 51 (3): pp 309.
Lagardère, F. (1989). Influence of feeding conditions and temperature on the growth rate and otolith-increment deposition of larval Dover sole (Solea solea (L.)). Rapports et Procès-verbaux desréunions de la Commission internationale pour l'Exploration scientifique de la MerMéditerranée, 191: 390-399.
Lagardère, J. P. (1987). Feeding ecology and daily food consumption of common sole, Solea vulgarisQuensel, juveniles on the French Atlantic coast. J. Fish Biol., 30: 91-104.
Largier, J. L. (1993). Estuarine fronts: how important are they ? Estuaries, 16: 1-11.Lascombe, C. (1996). Suivi des apports polluants du Rhône à la Méditerranée. Phase expérimentale :
rappel des objectifs et premiers résultats. Colloque Potam'mes. 7es Rencontres de l'AgenceRegionale pour l'Environnement Provence-Alpes-Côte d'Azur, Digne-les-Bains (ed.): 281-282.
Lasserre, G. et Lasserre, P. (1979). Croissance, production et demande d'énergie d'une population desoles (Solea vulgaris) introduite dans une lagune aménagée (Bassin d'Arcachon). Publ. Sci.Tech. CNEXO : Actes Colloq., 7: 689-712.
Last, J. M. (1980). The food of twenty species of pleuronectiform larvae in the west-central North Sea.Fish. Res. Tech. Rep., 60: 44 p.
Le Direac'h-Boursier, L. (1990). Recrutement et comportement larvaire : ichtyoplancton du golfe de Fos- Etude expérimentale du déterminisme des migrations des larves et juvéniles de sole Soleavulgaris Q. Thèse 3e cycle, Univ. Aix-Marseille II. 314 p.
Lê, K. L. (1983). Les phases initiales du cycle biologique (oeufs, larves et jeunes) de Solea vulgarisQuensel et Solea senegalensis Kaup (Poissons, Pleuronectiformes, Soleidae). Relations avec lebassin de Marennes-Oléron, écosystème estuarien a vocation aquicole. Thèse 3e cycle, Univ.Aix-Marseille II. 136 p.
Lee, S. Y. (2000). Carbon dynamics of Deep Bay, eastern Pearl River estuary, China. II: Trophicrelationship based on carbon and nitrogen stable isotopes. Mar. Ecol. Prog. ser., 205: 1-10.
Lefèvre, D., Minas, H. J., Minas, M., Robinson, C., Williams, P. J. L. B. et Woodward, E. M. S. (1997).Review of gross community production, primary production, net community production anddark community respiration in the Gulf of Lions. Deep-sea Res. II, 44 (3-4): 801-832.
Legendre, P. et Gallagher, E. D. (2001). Ecological meaningful transformations for ordination of speciesdata. Oecologia, 129 (2): 271-280.

291
Legendre, P. et Legendre, L. (1998). Numerical ecology, 2nd English edition. Elsevier Science B.V.,Amsterdam: p. 853.
Leggett, W. C. et DeBlois, E. (1994). Recruitment in marine fishes: is it regulated by starvation andpredation in the eggs and larval stages? Neth. J. sea res., 32: 119-134.
Letourneur, Y., Darnaude, A. M., Salen-Picard, C. et Harmelin-Vivien, M. L. (2001). Spatial andtemporal variation of fish assemblages in a shallow Mediterranean soft-bottom area (Gulf ofFos, France). Oceanol. Acta, 24 (3): 273-285.
Linke, T. E., Platell, M. E. et Potter, I. C. (2001). Factors influencing the partitioning of food resourcesamong six fish species in a large embayment with juxtaposing bare sand and seagrass habitats.J. Exp. Mar. Biol. Ecol., 266: 193-217.
Lipiatou, E. et Saliot, A. (1991). Hydrocarbon contamination of the Rhone Delta and WesternMediterranean. Mar. Pollut. Bull., 22 (6): 297-304.
Lloret, J., Lleonart, J., Solé, I. et Fromentin, J. M. (2001). Fluctuations of landings and environmentalconditions in the north-western Mediterranean Sea. Fish. Oceanogr., 10 (1): 33-50.
Lochet, F. et Leveau, M. (1990). Transfers between a eutrophic ecosystem, the river Rhône, and anoligotrophic ecosystem, the north-western Mediterranean Sea. Hydrobiologia, 207: 95-103.
Ludwig, J. A. et Reynolds, J. F. (1988). Statistical ecology: a primer on methods and computing. Wiley& Sons, New-York: p. 337.
Macdonald, J. S. et Green, R. H. (1983). Redundancy of variables Used to Describe Importance of Preyin Fish Diets. Can. J. Fish. aquat. Sci., 40: 635-637.
Macdonald, J. S. et Green, R. H. (1986). Food resource utilization by five species of benthic feedingfish in Passamaquoddy Bay, New Brunswick. Can. J. Fish. aquat. Sci., 43: 1534-1546.
Macpherson, E. et Duarte, A. P. (1991). Bathymetric trends in demersal fish size : is there a generalrelationship? Mar. Ecol. Prog. ser., 71: 103-112.
Maldonaldo, C., Dachs, J. et Batyona, J. M. (1999). Trialkylamines and Coprostanol as Tracers ofUrban Pollution in Waters from Enclosed Seas: The Mediterranean an Black Sea. Environ. Sci.Technol., 33: 3290-3296.
Mann, K. H. (1982). Ecology of coastal waters. A systems approach., 8. Blackwell ScientificPublications, Oxford: 322 p.
Mann, K. H. (1988). Production and use of detritus in various freshwater, estuarine and coastal marineecosystems. Limnol. Oceanogr, 33: 910-930.
Marchand, J. et Masson, G. (1989). Process of estuarine colonization by 0-group sole ( Solea solea):hydrological conditions, behaviour, and feeding activity in the Vilaine estuary. Rapports etProcès-verbaux des réunions de la Commission internationale pour l'Exploration scientifiquede la Mer Méditerranée, 191: 287-295.
Marinaro, J. Y. (1991). Croissance de la sole de sable Solea (Pegusa) theophilus (Poissons, Soléidés)sur la côte du Roussillon (France). Vie Milieu, 41 (2-3): 141-151.
Marinaro, J. Y., Baumgartner, N., Lencioni, R. et Barrois, J. M. (1978). Contribution à l'étude des oeufset larves pélagiques de poissons méditerranéens. VII - Premiers stades de la limande Citharusmacrolepidotus (Bloch, 1787). Pelagos, 5: 89-101.
Marinaro, J. Y. et Bouabib, M. (1983). Ecologie alimentaire de deux soles d'Algérie,Pegusa nasuta(Pallas) et P. lascaris (Risso) (Téléostéens, soléidés). Rapp. Comm. int. Mer Medit., 28 (5): 73-75.
Marinaro, J. Y., Chebab, L. et Chalabi, A. (1983). Croissance de Pegusa nasuta (Pallas) sur les côtesd'Algérie (Téléostéen, soléidé). Rapp. Comm. int. Mer Medit., 28 (5): 65-67.
Mascaro, M. et Seed, R. (2000). Foraging behavior of Carcinus maenas (L.): comparisons of size-selective predation on four species of bivalve prey. J. Shellfish Res., 19 (1): 283-291.

292
Massé, H. (1970). La suceuse hydraulique, bilan de quatre années d'emploi, sa manipulation, sesavantages et inconvénients. Peuplements benthiques. Téthys, 2 (2): 547-556.
Massé, H. (1971). Contribution à l'étude quantitative et dynamique de la macrofaune de peuplementsdes sables fins infralittoraux des côtes de Provence. Thèse : Univ. Aix-Marseille II. 310 p.
Massé, H. (1972a). Contribution à l'étude de la macrofaune de peuplements des sables fins infralittorauxdes côtes de Provence - VI : Données sur la biologie des espèces. Téthys, 4 (1): 63-84.
Massé, H. (1972b). Contribution à l'étude de la macrofaune de peuplements des sables fins infralittorauxdes côtes de Provence - VII: discussion, comparaison, et interprétation des donnéesquantitatives. Téthys, 4 (2): 379-422.
Massé, H. (1998). Conséquences à long terme de travaux d'aménagements littoraux sur la macrofaunedes sables fins de deux stations de la baie du Prado (Méditerranée Nord-Occidentale - Golfe deMarseille). Vie Milieu, 48: 79-87.
Massé, H. (2000). Long-term changes in sand-bottom macrofauna along the coast of Provence(northwest Mediterranean Sea). Oceanol. Acta, 23 (2): 229-242.
Massé, H. et Guérin, J.-P. (1976). Etude expérimentale sur le recrutement des espèces de la macrofaunebenthique des substrats meubles. 2- Données sur les cycles biologiques des polychètes et desbivalves. Téthys, 8 (3): 283-294.
Massé, H., Plante, R. et Reys, J.-P. (1977). Etude comparative de l'efficacité de deux bennes et d'unesuceuse en fonction de la nature du fond. Biology of benthic organisms. 11th EuropeanSymposium on Mar. Biol., Galway : 539-568.
Massin, C. (1982). Food and feeding mechanisms: holothuroidea. In: Jangoux, M. et Lawrence, J. M.(ed.), Echinoderm Nutrition, A. A. Balkema Publishers, Rotterdam, pp 43-55.
Maurin, C. (1968). Ecologie ichthyologique des fonds chalutables atlantiques (de la baie ibéro-marocaine à la Mauritanie) et de la Méditerranée occidentale. Rev. Trav. Inst. pêches mar., 32(1): 5-147.
Mc Ardle, B. H. et Anderson, M. J. (2001). Fitting multivariate models to community data : a commenton distance-based redundancy analysis. Ecology, 82 (1): 290-297.
Mc Arthur, R. H. (1972). Geographical Ecology. Harper & Row, New-York: 269 p.Mc Connaughey, T. et Mc Roy, C. P. (1979). Food-web structure and the fractionation of carbon
isotopes in the Bering Sea. Mar. Biol., 53: 257-262.Mendez de Elguezabal, E. (1978). Contribution à l'étude biologique et écologique des populations de
Soles, Solea vulgaris du Golfe du Lion. Thèse 3e cycle, Univ. Montpellier II. 115 p.Meybeck, M. (1982). Carbon, nitrogen and phosphorus transport by world rivers. Amer. J. Sci., 282:
401-450.Miller, J. M. (1994). An overview of the second flatfish symposium: recuitment in flatfish. Neth. J. sea
res., 32 (2): 103-106.Miller, T. J., Crowder, L. B., Rice, J. A. et Marschall, E. A. (1988). Larval size and recruitment
mechanisms in fishes: toward a conceptual framework. Can. J. Fish. aquat. Sci., 45: 1657-1670.Millner, R. S. et Whiting, C. L. (1996). Long term changes in growth and population abundance of sole
in the North Sea from 1940 to the present. ICES J. mar. Sci., 53: 1185-1195.Millot, C. (1990). The Gulf of Lions' hydrodynamics. Cont. shelf Res., 10: 885-894.Minagawa, M. et Wada, E. (1984). Stepwise enrichment of 15N along food chains: Further evidence and
the relation between δ15N and animal age. Geochi. et Coasochim. Acta, 48: 1135-1140.
Molinero, A. et Flos, R. (1992a). Influence of sex and age on the feeding habits of the common soleSolea solea. Mar. Biol., 111: 493-501..
Molinero, A. et Flos, R. (1992b). Influence of season on the feeding habits of the common sole Soleasolea. Mar. Biol., 113: 499-507

293
Molinero, A., Garcia, E. et Flos, R. (1991). Comparative study of feeding habits for two species of sole:Solea vulgaris and Solea senegalensis. Spec. Publ. Eu. Aquacult. Soc., 14: 219-220
.
Mook, W. G. et Tan, T. C. (1991). Stable carbon isotopes in rivers and estuaries. In: Degens, E. T.,Kempe, S. et Richey, J. E. (ed.), Biogeochemistry of Major world Rivers., SCOPE, John Wiley& Sons Ltd, pp 245-264.
Moutin, T., Raimbault, P., Golterman, H. L. et Coste, B. (1998). The input of nutrients by the RhôneRiver into the Mediterranean Sea: recent observations and comparison with earlier data.Hydrobiologia, 373/374: 237-246.
Naudin, J. J. et Cauwet, G. (1997). Transfer mechanisms and biogeochemical implications in the bottomnepheloid layer. A case study of the coastal zone off the Rhone River (France). Deep-sea Res.II, 44 (3-4): 551-575.
Naudin, J. J., Cauwet, G., Leveau, M., Lochet, F., Pauc, H., Romano, J. C. et Sempéré, R. (1992). Lenéphéloïde benthique au large du Rhône. Transfert particulaires à l'interface continent-océan.Oceanol. Acta, 15 (6): 621-638.
Newel, R. C., Shumway, S. E., Cucci, T. L. et Selvin, R. (1989). The effects of natural seston particlesize and type on feeding rate, feeding selectivity and food resource avalability for the musselMythilus edulis Linnaeus, 1758 at bottom culture sites in Maine. J. Shellfish Res., 8 (1): 187-196.
Nichols, J. H. (1976). Soleidae of the Eastern North Atlantic. 150/151. 10 p.Nichols, P. D., Klumpp, D. W. et Johns, R. B. (1985). A study of food chains in seagrass communities.
III. Stable carbon isotope ratios. Austr. J. mar. Freshw. Res., 36: 683-690.Nielsen, E. et Richardson, K. (1996). Can changes in the fisheries yield in the Kattegat (1950-1992) be
linked to changes in primary production? ICES J. mar. Sci., 53: 988-994.Nikolskii, G. V. (1969). Fish Population Dynamics. Oliver & Boyd, Edinburgh: 323 p.Nisbet, I. C. T., Montoya, J. P., Burger, J. et Hatch, J. J. (2002). Use of stable isotope to investigate
individual differences in diets and mercury exposures among common terns Sterna hirundo inbreeding and wintering grounds. Mar. Ecol. Prog. ser., 242: 267-274.
Nithart, M. (1995). Role of two Polychaete species (Nereis diversicolor, Scoloplos armiger) inprocessing Organic Matter and Nutrient Cycling in a norfolk SaltMarsh (U.K.). Phd, Universityof East Anglia. 136 p.
Nithart, M. (2000). Comparison of stable carbon and nitrogen isotope signatures of the polychaeteNereis diversicolor from different estuarine sites. J. mar. Biol. Assoc. U.K., 80 (4): 763-765
.
Nolting, R. F. (1989). Dissolved and particulate trace metals in sediments of the Gulf of Lions. In:Martin, J.-M. et Barth, H. (ed.), Water pollut. Res. Rep. 13, EROS-2000 (European River OceanSystem). First workshop on the north-west Mediterranean Sea, pp 314-340.
Nottage, A. S. et Perkins, E. J. (1983). The biology of the solenette, Buglossidium luteum (Risso), in theSolway Firth. J. Fish Biol., 22 (1): 21-27.
Oh, C.-W., Hartnoll, R. G. et Nash, R. D. M. (2001). Feeding ecology of the common shrimp Crangoncrangon in Port Erin Bay, Isle of Man, Irish Sea. Mar. Ecol. Prog. ser., 214: 211-223.
Ohman, M. C. et Rajasuriya, A. (1998). Relationships between habitat structure and fish communitieson coral and sandstone reefs. Envir. Biol. fishes, 53 (1): 19-31.
Omnes, P. (1996). Mise en évidence de la voie de réduction dissimilatrice du nitrate en ammonium.Application in situ aux systèmes particulaires et sédimentaires en Méditerranée. Thèse 3e cycle,Univ. Aix-Marseille II. 199 p.
Ostrom, P. H. et Fry, B. (1993). Sources and cycling of organic matter within modern and prehistoricfood webs. In: Engel, M. H. et Macko, S. A. (ed.), Organic Geochemistry, Plenum Press, NewYork, pp 785-798.
Owens, N. J. P. (1987). Natural variations in 15N in the marine environment. Adv. mar. Biol., 24: 389-451.

294
Parker, P. L. et Calder, J. A. (1970). Stable carbon isotope ratio variations in biological systems. In:Hood, D. W. (ed.), Organic matter in natural waters, Institute of marine Science, Alaska, pp107-127.
Paterson, A. W. et Whitfield, A. K. (1997). A stable carbon isotope study of the food-web in afreswater-deprived south African estuary, with particular emphasis on the ichthyofauna. Estuar.Coast. Shelf Sci., 45: 705-715.
Pepin, P. (1991). The effect of temperature and size on development and mortality rates of the pelagicearly life history stages of marine fishes. Can. J. Fish. aquat. Sci., 48: 503-518.
Pérès, J. M. (1980). Rapports trophiques entre le micro- et le méiobenthos et le macrobenthos.Production primaire et secondaire. Colloque franco-soviétique. Marseille, 1979, (1980). Pérès,J. M. et C., E. (ed.), CNEXO (Actes de Colloques): 187-194.
Pérez, T., Sartoretto, S., Soltan, D., Capo, S., Fourt, M., Dutrieux, E., Vacelet, J., Harmelin, J. G. etRebouillon, P. (2000). Etude bibliographique sur les bioindicateurs de l'état du milieu marin.Système d'évaluation de la qualité des milieux littoraux - volet biologique., Document Agencede l'Eau, fasc. 1-4: 642 p.
Peterson, B. J. et Fry, B. (1987). Stable isotopes in ecosystem studies. Annu. Rev. Ecol. Syst., 18: 293-320.
Peterson, B. J., Howarth, R. W. et Garritt, R. H. (1985). Multiple stable isotopes used to trace the flowof organic matter in estuarine food webs. Science, 227: 1361-1363.
Picard, C. (1971). Les peuplements de vase au large du golfe de Fos. Téthys, 3 (3): 569-618.Picard, J. (1965). Recherches qualitatives sur les biocénoses marines des substrats meubles dragables de
la région marseillaise. Rec. Trav. Stn mar. Endoume, 52 (36): 1-160.Pielou, E. C. (1969). An Introduction to Mathematical Ecology. Wiley Inter-science, New-York: 286 p.Piet, G. J., Pet, J. S., Guruge, W. A. H. P., Vijverberg, J. et Van Densen, W. L. T. (1999). Resource
partitioning along three niche dimensions in a size-structured tropical fish assemblage. Can. J.Fish. aquat. Sci., 56: 1241-1254.
Pimm, S. L. (1982). Food webs. Chapman and Hall Ltd, London: 219 p.Pinazo, C., Marsaleix, P., Millet, B., Estournel, C., Kondrachoff, V. et Véhil, R. (2001). Phytoplankton
variability in summer in the Northwestern Mediterranean: Modelling of the wind and freshwaterimpacts. J. coastal Res., 17 (1): 146-161.
Pinnegar, J. K. (2000). Planktivorous Fishes: Links Between the Mediterranean Littoral and Pelagic.PhD, Univ. of Newcastle upon Tyne. 208 p.
Pinnegar, J. K. et Polunin, N. V. C. (1999). Differential fractionation of δ13C and δ15N among fishtissues: implications for the study of trophic interactions. Funct. Ecol., 13: 225-231.
Pinnegar, J. K. et Polunin, N. V. C. (2000). Contributions of stable-isotope data to elucidating foodwebs of Mediterranean rocky littoral fishes. Oecologia, 122: 399-409.
Poll, M. (1947). Poissons marins. Faune de Belgique. Musée Royal Hist. Nat. Belgique: 452 p.Polunin, N. V. C., Morales-Nin, B., Pawsey, W. E., Cartes, J. E., Pinnegar, J. K. et Moranta, J. (2001).
Feeding relationships in Mediterranean bathyal assemblages elucidated by stable nitrogen andcarbon isotope data. Mar. Ecol. Prog. ser., 220: 13-23.
Ponsard, S. et Averbuch, P. (1999). Should growing and adult animals fed on the same diet showdifferent delta 15N values ? Rap. Comm. Mass spectrom., 13 (13): 1305-1310.
Pont, D. (1997). Les débits solides du Rhône à proximité de son embouchure: données récentes (1994-1995). Rev. Géogr. Lyon, 72: 23-33.
Pourriot, R. et Meybeck, M. (1995). Limnologie générale. Masson, Paris: 956 p.

295
Probst, J. L. (1989). Hydroclimatic fluctuations of some European rivers since 1800. In: Petts, G. E.(ed.), Historical Change of Large Alluvial Rivers: Western Europe, John Wiley & Sons Ltd,New York, pp 41-55.
Quéro, J.-C. et Vayne, J.-J. (1997). Les poissons de mer des pêches françaises. Delachaux et Niestlé,Paris: 304 p.
Radakovitch, O., Charmasson, S., Arnaud, M. et Bouisset, P. (1999). 210Pb and Caesium Accumulationin the Rhone Delta Sediments. Estuar. coast. mar. Sci., 48 (1): 77-92.
Raikow, D. F. et Hamilton, S. K. (2001). Bivalve diets in a midwestern U.S. stream: a stable isotopeenrichment study. Limnol. Oceanogr., 46 (3): 514-522.
Randall, P. J. et Myers, A. A. (2001). Effects of resource matrix, gut region analysed and sampled sizeon diet statistics in co-existing species of flatfish. J. marine Biol. Assoc. U.K., 81: 1041-1048.
Redon, M. J., Morte, M. S. et Sanz-Brau, A. (1994). Feeding habits of the spotted flounder Citharuslinguatula off the eastern coast of Spain. Mar. Biol., 120 (2): 197-201.
Reys, J. P. (1960). Etude de la nourriture de quelques poissons démersaux du golfe du Lion. Rec. Trav.Stn mar. Endoume, 33 (20): 463-530.
Riera, P. (1995). Origine et devenir de la matière organique dans un écosystème estuarien à vocationconchylicole. Utilisation du traçage isotopique naturel. Thèse : Univ. Bordeaux I. 214 p.
Riera, P. (1998). δ15N of organic matter sources and benthic invertebrates along an estuarine gradient inMarennes-Oléron Bay (France): implications for the study of trophic structure. Mar. Ecol. Prog.ser., 166: 143-150.
Riera, P., Montagna, P. A., Kalke, R. D. et Richard, P. (2000). Utilization of estuarine organic matterduring growth and migration by juvenile brown shrimp Penaeus aztecus in a South Texasestuary. Mar. Ecol. Prog. ser., 199: 205-216.
Riera, P. et Richard, P. (1996). Isotopic determination of food sources of Crassostrea gigas along atrophic gradient in the estuarine bay of Marennes-Oléron. Estuar. coast. shelf Sci., 42: 347-360.
Riera, P. et Richard, P. (1997). Temporal variation of δ13C in particulate organic matter and oysterCrassostrea gigas in Marennes-Oléron Bay (France): effect of freshwater inflow. Mar. Ecol.Prog. ser., 147: 105-115.
Riera, P., Stal, L. J. et Nieuwenhuize, J. (2002). δ13C versus δ15N of co-occuring molluscs within acommunity dominated by the Crassostrea gigas and Crepidula fornicata (Oosterschelde, TheNetherlands). Mar. Ecol. Prog. ser., 240: 291-295.
Riera, P., Stal, L. J., Nieuwenhuize, J., Richard, P., Blanchard, G. et Gentil, F. (1999). Determination offood sources for benthic invertebrates in a salt marsh (Aiguillon Bay, France) by carbon andnitrogen stable isotopes: importance of locally produced sources. Mar. Ecol. Prog. ser., 187:301-307.
Rijnsdorp, A. D., Daan, N., Van Beek, F. A. et Heessen, H. J. L. (1991). Reproductive variability inNorth Sea plaice, sole and cod. J. Cons. – Cons. perm. int. Explor. mer, 47: 352-375.
Robertson, A. I. (1980). The structure and organization of an eelgrass fish fauna. Oecologia, 47: 76-82.Rodriguez, A. (1996). Diet of two Soleidae, Solea lascaris and Solea impar, on the western coast of
Brittany. Cybium, 20 (3): 261-277.
Rogers, S. I. (1991). The occurrence of young scaldfish Arnoglossus laterna (Walbaum, 1792) in theeastern Irish Sea. J. Fish Biol., 39 (6): 889-891.
Rogers, S. I. (1992). Environmental factors affecting the distribution of sole (Solea solea (L.)) within anursery area. Neth. J. sea res., 29 (1-3): 153-161.
Rogers, S. I. (1994a). Population density and growth rate of juvenile sole Solea solea (L.). Neth. J. seares., 32 (3/4): 353-360.

296
Rogers, S. I. (1994b). Species composition and production of sole, Solea solea L., in a flatfish nurseryground on the North Wales coast, U.K. An International Symposium. Sea Ranching of Cod andOther Marine Fish Species, Danielssen, D. S., Howell, B. R. et Mocksness, E. (ed.), Aquacut.Fish Manage.: 161-177.
Rogers, S. I. et Jinadasa, J. (1989). Prey partitioning among juvenile flatfish with particular reference tothe Soleidae (Abstract). In: Blaxter, J. H. S., Gamble, J. C. et Westernhagen, H. v. (ed.), TheEarly Life History of Fish., 191. Third Ices Symposium, Bergen, pp pp 466.
Rolff, C. et Elmgren, R. (2000). Use of riverine organic matter in plankton food webs of the Baltic Sea.Mar. Ecol. Prog. ser., 197: 81-101.
Ross, S. T. (1986). Ressource partitioning in fish assemblages : a review of field studies. Copeia, 2:352-388.
Sabatés, A. (1988). Larval development and spawning of Citharus lingatula (Linnaeus, 1758) in thewestern Mediterranean. J. Plankton Res., 10 (6): 1131-1140.
Sale, P. F. (1977). Maintenance of high diversity in coral reef fish communities. Am. Nat., 111: 337-359.
Salen-Picard, C. (1982). Contribution à l'étude dynamique des peuplements marins de substratsmeubles: les peuplements benthiques circalittoraux soumis à l'envasement dans la régionprovençale. Thèse d'Etat, Univ. Aix-Marseille II. 262 p.
Salen-Picard, C. et Arlhac, D. (2002). Long-term changes in a Mediterranean benthic community:relationships between the polychaete assemblages and hydrological variations of the Rhôneriver. Estuaries, 25 (6): 1121-1130.
Salen-Picard, C., Arlhac, D. et Alliot, E. (in press). Response of a Mediterranean soft bottomcommunity to short term (1993-1996) hydrological changes in the Rhône river. Mar. Environ.Res.
Salen-Picard, C., Bellan-Santini, D., Bellan, G., Arlhac, D. et Marquet, R. (1997). Changements à longterme dans une communauté benthique d'un golfe méditerranéen (golfe de Fos). Oceanol. Acta,20: 299-310.
Salen-Picard, C., Darnaude, A. M., Arlhac, D. et Harmelin-Vivien, M. L. (2002). Fluctuations ofmacrobenthic populations: a link between climate variability, run-off and coastal fisheries.Oecologia, 133 (380-388):
Sanchez-Velasco, L. (1998). Diet composition and feeding habits of fish larvae of two co-occuringspecies (Pisces: Callionymidae and Bothidae) in the north-western Mediterranean. ICES J. mar.Sci., 55: 299-308.
Sartor, P., Sbrana, M., Ungaro, N., Marano, C., Piccinetti, C. et Piccinetti Manfrin, G. (2002).Distribution and abundance of Citharus linguatula, Lepidorhombus boscii, and Solea vulgaris(Osteichthyes: Pleuronectiformes) in the Mediterranean Sea. Sci. mar., 66 (Suppl. 2): 83-102.
Sauriau, P.-G. et Kang, C.-K. (2000). Stable isotope evidence of benthic microalgae-based growth andsecondary production in the suspension feeder Cerastoderma edule (Mollusca, Bivalvia) in theMarennes-Oléron Bay. Hydrobiologia, 440: 317-329.
Schlacher, T. A. et Wooldridge, T. H. (1996). Origin and trophic importance of detritus - Evidence fromstable isotopes in the benthos of a small, temperate estuary. Oecologia, 106 (3): 382-388.
Schoener, T. W. (1970). Non-synchronous spatial overlap of lizards in patchy habits. Ecology, 51: 408-418.
Schoener, T. W. (1974). Ressource partitioning in ecological communities. Science, 185: 27-39.Sempéré, R., Charrière, B., Van Wambeke, F. et Cauwet, G. (2000). Carbon inputs of the Rhône River
to the Mediterranean Sea: Biogeochemical implications. Glob. Biogeochem. cycles, 14 (2): 669-681.
Sheaves, M. et Molony, B. (2000). Short-circuit in the mangrove food chain. Mar. Ecol. Prog. ser., 199:97-109.

297
Shéhata, S. (1984). Contribution à la connaissance des Soléidés (Poissons, Téléostéens) du golfe duLion. Systématique - Ecobiologie. Thèse 3e cycle, Univ. Montpellier II. 311 p.
Shumway, S. E., Cucci, T. L., Newel, R. C. et Yentsch, C. M. (1985). Particle selection, ingestion, andabsorption in filter-feeding bivalves. J. Exp. Mar. Biol. Ecol., 91 (1-2): 77-92.
Simenstad, C. A. et Wissmar, R. C. (1985). δ13C evidence of the origins and fates of organic carbon inestuarine and nearshore food webs. Mar. Ecol. Prog. ser., 22: 141-152.
Smith, B. N. et Epstein, S. (1971). Two categories of 13/12C ratios for higher plants. Pl. Physiol., 47:380-384.
Solbrig, O. T. et Solbrig, D. L. (1979). Introduction to Population Biology and Evolution. Addison-Wesley Reading, MA: 368 p.
Sorbe, J. C. (1972). Ecologie et éthologie alimentaire de l'ichtyofaune chalutable du plateau continentalSud Gascogne. Thèse 3e cycle, Univ. Aix-Marseille II. 125 p.
Sorbe, J. C. (1981). Rôle du benthos dans le régime alimentaire des poissons démersaux du secteur SudGascogne. Kiel. Meeresforsch., 5: 479-489.
Steven, G. A. (1930). Bottom fauna and the food of fishes. J. mar. Biol. Assoc. U.K., 16 (3): 677-700.
Stoner, A. W. et Zimmerman, R. J. (1988). Food pathways associated with penaeid shrimps in amangrove-fringed estuary. Fish. Bull., 86: 543-551.
Sutcliffe, W. H. J., Drinkwater, K. et Muir, B. S. (1976). Correlations of Fish Catch and EnvironmentalFactors in the Gulf of Maine. J. Fish. Res. Board Can., 34: 19-30.
Takai, N., Mishima, Y., Yorozu, A. et Hoshika, A. (2002). Carbon sources for demersal fish in thewestern Seto Inland Sea, Japan, examined by δ13C and δ15N analyses. Limnol. Oceanogr, 47 (3):730-741.
Temore, K. R., Cammen, L., Findlay, S. E. G. et Phillips, N. (1982). Perspectives of research ondetritus: do factors controlling the availability of detritus to macroconsumers depend on itssource? J. Mar. Res., 40: 473-490.
Thomas, C. J. et Blair, N. E. (2002). Transport and digestive alteration of uniformly 13C-labeled diatomsin mudflat sediments. J. Mar. Res., 60: 517-535.
Thomas, C. J. et Cahoon, L. B. (1993). Stable isotope analyses differentiate between different trophicpathways supporting rocky-reef fishes. Mar. Ecol. Prog. ser., 95: 19-24.
Thorman, S. (1982). Niche dynamics and resource partitioning in a fish guild inhabiting a shallowestuary on the Swedish West Coast. Oikos, 39: 32-39.
Thorman, S. et Wiederholm, A.-M. (1983). Seasonal occurence and food resource use of an assemblageof nearshore fish species in the Bothnian Sea, Sweden. Mar. Ecol. Prog. ser., 10: 223-229.
Thorman, S. et Wiederholm, A.-M. (1986). Food, habitat and time niches in a coastal fish speciesassemblage in a brackish water bay in the Bothnian Sea, Sweden. J. exp. mar. Biol. Ecol., 95:67-86.
Tirasin, E. M. et Jorgensen, T. (1999). An evaluation of the precision of diet description. Mar. Ecol.Prog. ser., 182: 243-252.
Tito de Morais, L. (1983). Etude de la nutrition et de ses aspects énergétiques chez des formes juvénilesde Pleuronectiformes. Thèse 3e cycle., Univ. Paris VI. 171 p.
Tito de Morais, L. (1984). Rythmes et rations alimentaires des juvéniles de deux espèces dePleuronectiformes de Méditerranée occidentale. Vie Milieu, 34 (2/3): 141-147.
Tito de Morais, L. (1986). Gastric evacuation of juveniles of two flatfish from a Western MediterraneanBay. Mar. Ecol., 7 (3): 255-264.
Tito de Morais, L. et Bodiou, J. Y. (1984). Predation on meiofauna by juvenile fish in a WesternMediterranean flatfish nursery ground. Mar. Biol., 82 (209-215):
Todd, R. A. (1907). Second report on the food of fishes (North Sea 1904-1905). Rep. North Sea Fish.Invest. Comm., South Area, 2 (1): 49-163.

298
Touzani, A. et Giresse, P. (2002). The Rhône River prodelta: short-term (100-103 year) sedimentationpatterns and human impact. J. coastal Res., 18 (1): 102-117.
Tusseau-Vuillemain, M.-H. (1998). Modeling nitrate fluxes in an open coastal environment (Gulf ofLions): transport versus biogeochemical processes. J. Geophys. Res., 103 (C4): 7693-7708.
Van der Veer, H. W. et Bergman, M. J. N. (1987). Predation by crustaceans on a newly settled 0-groupplaice Pleuronectes platessa L.) population in the western Wadden Sea. Mar. Ecol. Prog. ser.,35: 203-215.
Van der Veer, H. W. et Witte, J. I. J. (1993). The "maximum growth/optimal food condition"hypothesis: a test for 0-group plaice Pleuronectes platessa in the Dutch Wadden Sea. Mar. Ecol.Prog. ser., 101: 81-90.
Van Dover, C. L., Grassle, J. F., Fry, B., Garritt, R. H. et Srtarczak, V. R. (1992). Stable isotopeevidence for entry of sewage-derived organic material into a deep-sea food web. Nature, 360:153-156.
Vander Zanden, M. J., Cabana, G. et Rasmussen, J. B. (1997). Comparing trophic position of freshwaterfish calculated using stable nitrogen isotope ratios (δ15N) and literature dietary data. Can. J.Fish. aqaut. Sci., 54: 1142-1158.
Vander Zanden, M. J., Casselman, J. M. et Rasmussen, J. B. (1999). Stable isotope evidence for thefood web consequences of species invasion in lakes. Nature, 401: 464-467.
Vander Zanden, M. J. et Rasmussen, J. B. (1996). A trophic position model of pelagic food webs:impact on contaminant bioaccumulation in lake trout. Ecological Monographs, 66 (4): 451-477.
Vander Zanden, M. J. et Rasmussen, J. B. (1999). Primary consumer δ13C and δ15N and the trophicposition of aquatic consumers. Ecology, 80 (4): 1395-1404.
Vander Zanden, M. J. et Rasmussen, J. B. (2001). Variation in δ15N and δ13C trophic fractionation:Implications for aquatic food web studies. Limnol. Oceanogr., 46 (8): 2061-2066.
Vassilopoulou, V. et Papaconstantinou, C. (1994). Age, growth and mortality of the spotted flounder(Citharus linguatula Linnaeus, 1758) in the Agean Sea. Sci. mar., 58 (3): 261-267.
Vivian, H. (1989). Hydrological changes of the Rhône River. In: Petts, G. E. (ed.), Historical Change ofLarge Alluvial Rivers: Western Europe, John Wiley & Sons Ltd, New York, pp 57-77.
Wallace, R. K. (1981). An assessment of diet-overlap indexes. Trans. am. Fish. Soc., 110: 72-76.Warwick, R. M. et Clarke, K. R. (1991). A comparison of some methods for analysing changes in
benthic communities. J. mar. Biol. Assoc. U.K, 71: 225-244.
Warwick, R. M. et Clarke, K. R. (1993). Comparing the severity of disturbance: a meta-analysis ofmarine benthic communities. Mar. Ecol. Prog. ser., 92: 221-231.
Wheeler, A. C. (1969). Fishes of the British Isles and North West Europe. Mc Millan, London: 529 p.
Wolff, W. J. (1983). Estuarine benthos. In: Ketchum, B. H. (ed.), Estuaries and enclosed seas, Elsevier,Amsterdam, pp 151-182.
Woodhead, P. M. J. (1964). Changes in the behaviour of the sole, Solea vulgaris, during cold winters,and the relation between the winter catch and sea temperature. Helgol. Wiss. Meeresunters, 10:328-342.
Yáñez, E., Garcia, M. et Barbieri, M. A. (1998). Pelagic fish stocks and environmental changes in thesouth-east Pacific. In: Durand, M., Curry, P., Mendelssohn, R., Roy, C., Bakun, A. et Pauly, D.(ed.), Global versus local changes in upwelling systems, ORSTOM Editions, Paris, pp 275-291.
Younes, W. A. N. (2000). Observation à haute définition spatiale de la variabilité temporelle àdifférentes échelles d'un hydrosystème estuarien, le Rhône, et de son influence sur les eauxcôtières. Thèse : Univ. Aix-Marseille II. 427 p.
Zar, J. H. (1999). Biostatistical Analysis - Fourth Edition. Prentice-Hall, New Jersey: 663 p.Zuo, Z., Eisma, D., Gieles, R. et Beks, J. (1997). Accumulation rates and sediment deposition in the
northwestern Mediterranean. Deep-sea Res. II, 44 (3-4): 597-609.

LISTE DES FIGURES ET
DES TABLEAUX


299
LISTE DES FIGURES(Au total, 145 figures)
PARTIE I
Figure I.1 – Situation géographique de la zone retenue pour cette étude.Figure I.2 – Directions des courants de surface dans le Golfe du Lion et forme et étendue du panache rhodanien:principaux cas de figure observés au printemps-été (a) et en automne-hiver (b). D'après Bekers et al. (1997),modifié. Figure I.3 – Adulte de Solea solea. D'après Fisher et al. (1987).Figure I.4 – Répartition géographique de Solea solea. D'après Deniel (1981), modifié.Figure I.5 – Adulte de Solea impar. D'après Fisher et al. (1987).Figure I.6 – Répartition géographique de Solea impar. D'après Shéhata (1984), modifié.Figure I.7 – Adulte de Buglossidium luteum. D'après Fisher et al. (1987)Figure I.8 – Répartition géographique de Buglossidium luteum. D'après Deniel (1981), modifié.Figure I.9 – Adulte d'Arnoglossus laterna. D'après Deniel (1981).Figure I.10 – Répartition géographique d'Arnoglossus laterna. D'après Deniel (1981), modifié.Figure I.11 – Adulte de Citharus linguatula. D'après Fisher et al. (1987).Figure I.12 – Répartition géographique de Citharus linguatula. D'après Deniel (1981), modifié.Figure I.13 – Localisation (en rouge) des prélèvements de poissons effectués à l'embouchure en 2000-2001.♦ = traits de senne de plage ; chalutages.Figure I.14 – Schéma du chalut à panneaux utilisé.Figure I.15 – pêche réalisée sur 30 m de fond au chalut à panneaux.Figure I.16 – Schéma du petit gangui utilisé.Figure I.17 – Juvéniles récoltés au petit gangui sur 5-10 m de profondeur.Figure I.18 – Senne de plage utilisée. Schéma du constructeur.Figure I.19 – Juvéniles de soles récoltés à la senne de plage sur 0-2 m de profondeur (photo : M. Harmelin).Figure I.20 – Localisation des prélèvements d'eau de surface et de sédiment superficiel réalisés en 2000-2001.Les prélèvements de sédiment n'ont été réalisés que sur R1.Figure I.21 – Carottier multitubes utilisé : mise à bord après prélèvement sur le fond de 4 carottes de sédiment.Figure I.22 – Carottier-benne Flucha utilisé : mise à bord après récolte du sédiment (photo : C. Salen).Figure I.23 – Remontée de la spatangue utilisée. Les premiers cm superficiels du sédiment ont été écrémés grâce àla fente située sur la partie métallique puis piégés dans la poche qui prolonge l'engin.Figure I.24 – Localisation des prélèvements de faune benthique réalisés à l'embouchure de juin 2000 à septembre2001.Figure I.25 – Spatangue utilisée pour la récolte des proies benthiques sur 10-100 m de fond.Figure I.26 – Drague Waren utilisée pour la récolte des crustacés nectobenthiques sur 30-100 m de fond (photo C.Marschal).Figure I.27 – Utilisation de la suceuse pneumatique en plongée pour la récolte des proies benthiques et descrustacés nectobenthiques sur 0-20 m de fond (photo R. Graille).
PARTIE IICHAPITRE I
Figure II/I.1 – Fluctuations spatio-temporelles des abondances relatives des 5 espèces étudiées au large du Rhône.Les effectifs par espèce sont exprimés en pourcentages relatifs du nombre total d'individus des cinq espècescapturés à chaque saison pour chaque profondeur.

300
Figure II/I.2 – Fluctuations spatio-temporelles de la taille des individus de S. solea récoltés au large du Rhône.Les effectifs par classe de taille sont exprimés en fréquences relatives (F%) du nombre total d'individus capturés àchaque saison et chaque profondeur.Figure II/I.3 – Fluctuations spatio-temporelles de la taille des individus de S impar récoltés au large du Rhône.Les effectifs par classe de taille sont exprimés en fréquences relatives (F%) du nombre total d'individus capturés àchaque saison et chaque profondeur.Figure II/I.4 – Fluctuations spatio-temporelles de la taille des individus de B. luteum récoltés au large du Rhône.Les effectifs par classe de taille sont exprimés en fréquences relatives (F%) du nombre total d'individus capturés àchaque saison et chaque profondeur.Figure II/I.5 – Fluctuations spatio-temporelles de la taille des individus d'A. laterna récoltés au large du Rhône.Les effectifs par classe de taille sont exprimés en fréquences relatives (F%) du nombre total d'individus capturés àchaque saison et chaque profondeur.Figure II/I.6 – Fluctuations spatio-temporelles de la taille des individus de C. linguatula récoltés au large duRhône. Les effectifs par classe de taille sont exprimés en fréquences relatives (F%) du nombre total d'individuscapturés à chaque saison et chaque profondeur.
CHAPITRE II
Figure II/II.1 - Variations nycthémérales de l'indice de réplétion stomacal moyen (IR) chez S. solea (juin 2000 ;heure locale = heure GMT +1). CS = coucher du soleil ; LS = lever du soleil.Figure II/II.2 - Représentation en Boites de Tukey des nombres (a) et des poids totaux (b) de proies par estomacchez S. solea au large du Rhône.Figure: II/II.3 - Ordination (NMS, solution à 2 dimensions) des différentes classes de taille (LT, mm) de S. soleaen fonction de leur régime alimentaire (Poids moyen des proies par estomac).Figure II/II.4 - Classification par groupement agglomératif des classes de taille de S. solea en fonction de leurrégime alimentaire (poids moyens des proies par estomac). La flèche verticale rouge indique le pourcentaged'information (65 %) retenu comme valeur seuil pour l'établissement des groupes alimentaires (A, B, B' et C).Figure II/II.5 - Représentation en Boites de Tukey des nombres de proies (Np, ind. est.-1) rencontrés dans lesestomacs des individus de chaque groupe de taille (LT, mm) chez S. solea au large du Rhône.Figure II/II.6 - Relation linéaire entre la taille (LT, mm) des individus de S. solea et le poids moyen (Pm, mg) desproies ingérées (r = 0,629; r2 = 0,395; P < 0,001).Figure II/II.7 - Variations de la composition pondérale (Cp) du régime alimentaire de Solea solea au large duRhône en fonction de la taille des individus (LT, mm).Figure II/II.8 - Représentation en Boites de Tukey des indices de réplétion stomacale (IR) observés aux troissaisons étudiées chez S. solea au large du Rhône. CV = coefficient de vacuité (% d'estomacs vides).Figure II/II.9 - Variations de l'alimentation (pourcentages pondéraux des proies ingérées) de S. solea en fonctionde la saison et de la taille des individus (longueur totale LT, en mm). N = nombre d'estomacs pleins analysé pourchaque groupe de poissons. Dans chaque cas, les proies surlignées en gris sont rencontrées dans plus de 20% desestomacs et/ou représentent plus de 10% du poids total des proies ingérées.Figure II/II.10 - Variations de l'alimentation (pourcentages pondéraux des proies ingérées) de S. solea en fonctionde la profondeur et de la taille des individus (longueur totale LT, en mm). N = nombre d'estomacs pleins analysépour chaque groupe de poissons. Dans chaque cas, les proies surlignées en gris sont rencontrées dans plus de 20%des estomacs et/ou représentent plus de 10% du poids total des proies ingérées.Figure II/II.11 - Variations de l'alimentation de S. solea en fonction de la profondeur (0-20 m, 30-50 m, 70-100m), de la saison (P = printemps, E = été, H = hiver), et de la taille (LT, mm) des individus. Ordination (N.M.S.,solution à 2 dimensions) réalisée sur les poids moyens des proies par estomac.Figure II/II.12 - Variations nycthémérales de l'indice de réplétion stomacal moyen (IR) chez S. impar (juin 2000;heure locale = heure GMT + 1). CS = coucher du soleil; LS = lever du soleil.Figure II/II.13 - Représentation en Boites de Tukey des nombres (a) et des poids totaux (b) des proies parestomac chez S. impar au large du Rhône.Figure II/II.14 - Ordination (NMS, solution à 2 dimensions) des différentes classes de taille (LT, mm) de S. imparen fonction de leur régime alimentaire (Poids moyen des proies par estomac).Figure II/II.5 - Classification par groupement agglomératif des classes de taille de S. impar en fonction de leurrégime alimentaire (poids moyens des proies par estomac). La flèche verticale rouge indique le pourcentaged'information (65%) retenu comme valeur seuil pour l'établissement des groupes alimentaires (A,B,C et D).

301
Figure II/II.16 - Représentation en Boites de Tukey des nombres de proies (Np, ind. est.-1) rencontrés dans lesestomacs des individus de chaque groupe de taille (LT, mm) chez S. impar au large du Rhône.Figure II/II.17 - Relation linéaire entre la taille (LT, mm) des individus de S. impar et le poids moyen (Pm, mg)des proies ingérées (r = 0,602; r2 = 0,362; P < 0,001).Figure II/II.18 - Variations de la composition pondérale (Cp) du régime alimentaire de S. impar au large duRhône en fonction de la taille des individus (LT, mm).Figure II/II.19 - Représentation en Boites de Tukey des indices de réplétion stomacale (IR) observés à chacunedes trois saisons étudiées chez les individus de S. impar au large du Rhône. CV = coefficient de vacuité (%d'estomacs vides).Figure II/II.20 - Variations de l'alimentation (pourcentages pondéraux des proies ingérées) de S. impar enfonction de la saison et de la taille des individus (longueur totale LT, en mm). N = nombre d'estomacs pleinsanalysé pour chaque groupe de poissons. Dans chaque cas, les proies surlignées en gris sont rencontrées dans plusde 20% des estomacs et/ou représentent plus de 10% du poids total des proies ingérées.Figure II/II.21 - Variations de l'alimentation de S. impar en fonction de la saison (P = printemps, E = été, H =hiver), et de la taille (LT, mm) des individus à 0-20 m de profondeur. Ordination (N.M.S., solution à 2 dimensions)réalisée sur les poids moyens des proies par estomac.Figure II/II.22 - Variations nycthémérales de l'indice de réplétion stomacal moyen (IR) chez B. luteum (juin 2000;heure locale = heure GMT + 1). CS = coucher du soleil; LS = lever du soleil.Figure II/II.23 - Représentation en Boites de Tukey des nombres (a) et des poids totaux (b) des proies parestomac chez B. luteum au large du Rhône.Figure II/II.24 - Ordination (NMS, solution à 2 dimensions) des différentes classes de taille (LT, mm) de B.luteum en fonction de leur régime alimentaire (Poids moyen des proies par estomac).Figure II/II.25 - Classification par groupement agglomératif des classes de taille de B. luteum en fonction de leurrégime alimentaire (poids moyens des proies par estomac). La flèche verticale rouge indique le pourcentaged'information (65 %) retenu comme valeur seuil pour l'établissement des groupes alimentaires (A, B, C et C').Figure II/II.26 - Relation linéaire entre la taille (LT, mm) des individus de B. luteum et le poids moyen (Pm, mg)des proies ingérées (r = 0,410; r2 = 0,168; P < 0,001).Figure II/II.27 - Variations de la composition pondérale (Cp) du régime alimentaire de B. luteum au large duRhône en fonction de la taille des individus (LT, mm).Figure II/II.28 - Représentation en Boites de Tukey des indices de réplétion stomacale (IR) observés aux troissaisons étudiées chez B. luteum au large du Rhône. CV = coefficient de vacuité (% d'estomacs vides).Figure II/II.29 - Variations de l'alimentation (pourcentages pondéraux des proies ingérées) de B. luteum enfonction de la saison et de la taille des individus (longueur totale LT, en mm). N = nombre d'estomacs pleinsanalysé pour chaque groupe de poissons. Dans chaque cas, les proies surlignées en gris sont rencontrées dans plusde 20% des estomacs et/ou représentent plus de 10% du poids total des proies ingérées.Figure II/II.30 - Variations de l'alimentation (pourcentages pondéraux des proies ingérées) de B. luteum enfonction de la profondeur et de la taille des individus (longueur totale LT, en mm). N = nombre d'estomacs pleinsanalysé pour chaque groupe de poissons. Dans chaque cas, les proies surlignées en gris sont rencontrées dans plusde 20% des estomacs et/ou représentent plus de 10% du poids total des proies ingérées.Figure II/II.31 -Variations de l'alimentation de B. luteum en fonction de la profondeur (0-20 m, 30-50 m, 70-100m), de la saison (P = printemps, E = été, H = hiver), et de la taille (LT, mm) des individus. Ordination (N.M.S.,solution à 2 dimensions) réalisée sur les poids moyens des proies par estomac.Figure II/II/32 - Variations nycthémérales de l'indice de réplétion stomacal moyen (IR) chez A. laterna (juin2000; heure locale = heure GMT +1). CS = coucher du soleil; LS = lever du soleil.Figure II/II.33 - Représentation en Boites de Tukey des nombres (a) et des poids totaux (b) des proies parestomac rencontrés chez A. laterna au large du Rhône.Figure II/II.34 - Ordination (NMS, solution à 2 dimensions) des différentes classes de taille (LT, mm) d'A. laternaen fonction de leur régime alimentaire (Poids moyen des proies par estomac).Figure II/II.35 - Classification par groupement agglomératif des différentes classes de taille d'A. laterna enfonction de leur régime alimentaire (poids moyens des proies par estomac). La flèche verticale rouge indique lepourcentage d'information (65 %) retenu comme valeur seuil pour l'établissement des groupes alimentaires (G. A,B et C).Figure II/II.36 - Représentation en Boites de Tukey des nombres de proies (Np, ind. est.-1) rencontrés dans lesestomacs des individus de chaque groupe de taille (LT, mm) chez A. laterna au large du Rhône.

302
Figure II/II.37 - Relation linéaire entre la taille (LT, mm) des individus d'A. laterna et le poids moyen (Pm, mg)des proies ingérées (r = 0,402; r2 = 0,161; P < 0,001).Figure II/II.38 - Variations de la composition pondérale (Cp) du régime alimentaire d'Arnoglossus laterna aularge du Rhône en fonction de la taille des individus (LT, mm).Figure II/II.39 - Représentation en Boites de Tukey des indices de réplétion stomacale (IR) observés à chaquesaison chez A. laterna au large du Rhône. CV = coefficient de vacuité (% d'estomacs vides).Figure II/II.40 - Variations de l'alimentation (pourcentages pondéraux des proies ingérées) d'A. laterna enfonction de la saison, de la profondeurr et de la taille des individus (longueur totale LT, en mm). N = nombred'estomacs pleins analysé pour chaque groupe de poissons. Dans chaque cas, les proies surlignées en gris sontrencontrées dans plus de 20% des estomacs et/ou représentent plus de 10% du poids total des proies ingérées.Figure II/II.41 - Variations de l'alimentation de A. laterna en fonction de la profondeur (0-20 m, 30-50 m, 70-100m), de la saison (P = printemps, E = été, H = hiver), et de la taille (LT, mm) des individus. Ordination (N.M.S.,solution à 2 dimensions) réalisée sur les poids moyen des proies par estomac.Figure II/II.42 - Variations nycthémérales de l'indice de réplétion stomacal moyen (IR) chez C. linguatula (juin2000; heure locale = heure GMT + 1). CS = coucher du soleil; LS = lever du soleil.Figure II/II.43 - Représentation en Boites de Tukey des nombres (a) et des poids totaux (b) des proies parestomac rencontrés chez C. linguatula au large du Rhône.Figure II/II.44 - Ordination (N.M.S., solution à 2 dimensions) des différentes classes de taille (LT, mm) de C.linguatula en fonction de leur régime alimentaire (poids moyen des proies par estomac).Figure II/II.45 - Classification par groupement agglomératif (Cluster Analysis) des différentes classes de taille deC. linguatula en fonction de leur régime alimentaire (poids moyens des différentes proies ingérées par estomac).La flèche verticale rouge indique le pourcentage d'information (65 %) retenu comme valeur seuil pourl'établissement des groupes alimentaires (A, B et C).Figure II/II.46 - Variations de la composition pondérale (Cp) du régime alimentaire de C. linguatula au large duRhône en fonction de la taille des individus (LT, mm).Figure II/II.47 - Relation linéaire entre la taille (LT, mm) des individus de C. linguatula et le poids moyen (Pm,mg) des proies ingérées (r = 0,438; r2 = 0,192; P < 0,001).Figure II/II.48 - Représentation en Boites de Tukey des nombres de proies (Np, ind. est.-1) rencontrés dans lesestomacs des individus de chaque groupe de taille (LT, mm) chez C. linguatula au large du Rhône.Figure II/II.49 - Représentation en Boites de Tukey des indices de réplétion stomacale (IR) observés à chacunedes trois saisons étudiées chez C. linguatula au large du Rhône. CV = coefficient de vacuité (% d'estomacs vides).Figure II/II.50 - Variations de l'alimentation (pourcentages pondéraux des proies ingérées) de C. linguatula enfonction de la saison et de la taille des individus (longueur totale LT, en mm). N = nombre d'estomacs pleinsanalysé pour chaque groupe de poissons. Dans chaque cas, les proies surlignées en gris sont rencontrées dans plusde 20% des estomacs et/ou représentent plus de 10% du poids total des proies ingérées.Figure II/II.51 - Variations de l'alimentation (pourcentages pondéraux des proies ingérées) de C. linguatula enfonction de la profondeur et de la taille des individus (longueur totale LT, en mm). N = nombre d'estomacs pleinsanalysé pour chaque groupe de poissons. Dans chaque cas, les proies surlignées en gris sont rencontrées dans plusde 20% des estomacs et/ou représentent plus de 10% du poids total des proies ingérées.Figure II/II.52 - Variations de l'alimentation de C. linguatula en fonction de la profondeur (30-50 m, 70-100 m),de la saison (P = printemps, E = été, H = hiver), et de la taille (LT, mm) des individus. Ordination (N.M.S.,solution à 2 dimensions) réalisée sur les poids moyen des proies par estomac.Figure II/II.53 - Rythmes nycthéméraux d'alimentation observés chez les cinq espèces étudiées au large du Rhône(juin 2000 ; heure locale = heure GMT + 1). AL = A. laterna, BL = B. luteum, CL = C. linguatula, SI = S. impar,SS = S. solea.
CHAPITRE III
Figure II/III.1 - Modèle illustrant le concept de la niche écologique : position d'un hypervolume-niche dans unespace à trois dimensions et projection de cet hypervolume sur les plans définis par ces dimensions (d'aprèsPianka, 1974 in Blondel, 1995)Figure II/III.2 - Projection de la niche de cinq espèces types sur deux dimensions de l'hypervolume-niche. Lesespèces dont les niches se chevauchent sur une dimension (la niche de l'espèce 3 est totalement incluse dans cellede l'espèce 4 sur la dimension I) peuvent être isolées par une deuxième dimension. (D'après Pianka, 1974 in

303
Blondel, 1995). Dans le cas présent, si les dimensions I et II correspondent respectivement à l'alimentation etl'habitat (profondeur), le cas de S. impar et C. linguatula est assimilable à celui des espèces 4 et 3 ou 4 et 2.Figure II/III.3 - Ordination des différents groupes de poissons individualisés pour les cinq espèces étudiées enfonction de leur alimentation au large du Rhône : N.M.S. (solution à 2 dimensions) réalisée sur les poids moyens(en mg) des proies ingérées par les individus de chaque espèce groupés par stade de vie benthique / saison /profondeur. Les différences de couleurs indiquent les profondeurs (voir légende).Figure II/III.4 - Positionnement des groupes alimentaires présents à 0-20 m dans le plan obtenu par ordination desespèces de poissons en fonction de leur alimentation (N.M.S. réalisée sur les individus groupés par stade de viebenthique / saison / profondeur, solution à 2 dimensions). Les différences de couleur indiquent les saisons :printemps (vert), été (jaune), hiver (bleu).Figure II/III.5 - Positionnement des groupes alimentaires présents à 30-50 m dans le plan obtenu par ordinationdes espèces en fonction de leur alimentation (N.M.S. réalisée sur les individus groupés par stade de vie benthique /saison / profondeur, solution à 2 dimensions). Les couleurs indiquent les saisons : printemps (vert), été (jaune),hiver (bleu).Figure II/III.6 - Positionnement des groupes alimentaires présents à 70-100 m dans le plan obtenu par ordinationdes espèces en fonction de leur alimentation (N.M.S. réalisée sur les individus groupés par stade de vie benthique /saison / profondeur, solution à 2 dimensions). Les couleurs indiquent les saisons : printemps (vert), été (jaune),hiver (bleu).Figure II/III.7 - Positionnement des groupes alimentaires présents au large du Rhône dans le plan obtenu parordination des cinq espèces en fonction de leur alimentation (N.M.S. réalisée sur les individus groupés par stade devie benthique / saison / profondeur, solution à 2 dimensions). Les différences de couleurs indiquent lesprofondeurs : 0-20 m (bleu-vert), 30-50 m (bleu ciel), 70-100 m (bleu foncé).
PARTIE IIIFigure III.1 – Représentation d'un réseau trophique théorique basé sur l'utilisation d'une source de matièreorganique unique. P1 = producteur primaire, C1 = consommateurs primaires (herbivores), C2 = consommateurssecondaires (carnivores).Figure III.2 – Représentation d'un réseau trophique théorique basé sur l'utilisation de deux sources de matièreorganique distinctes A (en blanc) et B (en noir). P1 = producteur primaire, C1 = consommateurs primaires(herbivores), C2 = consommateurs secondaires (carnivores). Les points gris représentent les organismes (C1, C2)dépendant des deux sources de matière organique.Figure III.3 - Schéma descriptif du système utilisé pour les analyses isotopiques. Le système est constitué d'unanalyseur élémentaire automatisé Europa Scientific ANCA-SL (Automated Nitrogen Carbon Analysis unit forSolids and Liquids), relié à un spectromètre de masse isotopique à flot continu (Europa Scientific PDZ 20-20). Lesnuméros indiquent les différentes étapes du traitement des échantillons. 0 = positionnement initial des échantillons,1 = combustion (capsule → gaz), 2 = réduction et purification des gaz, 3 = élimination de l'eau, 4 = séparation duN2 et du CO2, 5 = ionisation et accélération des particules, 6 = séparation des faisceaux d'ions et détection .Figure III.4 – Signatures isotopiques (moyennes et erreurs-types) obtenues pour les quatre principales sourcespotentielles de matière organique ainsi que pour la MOP de l'eau de surface et le sédiment superficiel au large duRhône.Figure III.5 – Moyennes et erreurs-types des signatures isotopiques (δ13C = f (δ15N)) obtenues en période de crues(C) et en périodes d'étiage (E) pour les 2 principales sources de matière organique d'origine terrestre (MOPrhodanienne) et marine (phytoplancton) et pour l'eau de surface et le sédiment superficiel de la zone d'étude àchaque profondeur échantillonnée (0-20 m = 10 m, 30-50 m = 30 m, 70-100 m = 90 m). MOP = matière organiqueparticulaire; Sed. = sédiment.Figure III.6 - Signatures en carbone (δ13C moyen ± erreur-type, en ‰) observées en période de crue pour les 2principales sources de matière organique d'origine terrestre (MOP Rhône) et marine (phytoplancton) et pour l'eaude surface et le sédiment superficiel de la zone d'étude à chaque tranche bathymétrique échantillonnée (0-20 m,30-50 m, 70-100 m). MOP = matière organique particulaire; Sed. = sédiment. Les barres horizontales indiquent lesmoyennes non-statistiquement différentes (P > 0,05).Figure III.7 - Signatures en carbone (δ13C moyen ± erreur-type, en ‰) observées en période d'étiage pour les 2principales sources de matière organique d'origine terrestre (MOP Rhône) et marine (phytoplancton) et pour l'eaude surface et le sédiment superficiel de la zone d'étude à tranche bathymétrique échantillonnée (0-20 m, 30-50 m,

304
70-100 m). MOP = matière organique particulaire; Sed. = sédiment. Les barres horizontales indiquent lesmoyennes non-statistiquement différentes (P > 0,05).Figure III.8 - Exploitation des sources de matière organique d'origine marine et terrestre par les différentescomposantes des réseaux trophiques benthiques aboutissant aux cinq espèces de poissons plats étudiées au largedu Rhône. Les erreurs-types associées aux signatures moyennes (δ13C et δ15N) de chaque catégorie sont indiquéesen gris. MOP = matière organique particulaire, brach. = brachyoure, pol = polychète, S = suspensivore, DS =dépositivore de surface, DsS = dépositivore de sub-surface, C = carnivore, pK = plancton.Figure III.9 - Exploitation des sources de matière organique d'origine marine et terrestre par les différentescomposantes des réseaux trophiques benthiques aboutissant aux cinq espèces de poissons plats à 0-20 m deprofondeur au large du Rhône. Les erreurs-types associées aux signatures moyennes (δ13C et δ15N) de chaquecatégorie sont indiquées en gris. MOP = matière organique particulaire, pol = polychète, S = suspensivore, DS =dépositivore de surface, DsS = dépositivore de sub-surface, C = carnivore, pK = plancton.Figure III.10 - Exploitation des sources de matière organique d'origine marine et terrestre par les différentescomposantes des réseaux trophiques benthiques aboutissant aux aux cinq espèces de poissons plats à 30-50 m deprofondeur au large du Rhône. Les erreurs-types associées aux signatures moyennes (δ13C et δ15N) de chaquecatégorie sont indiquées en gris. MOP = matière organique particulaire, pol = polychète, S = suspensivore, DS =dépositivore de surface, DsS = dépositivore de sub-surface, C = carnivore, pK = plancton.Figure III.11 - Exploitation des sources de matière organique d'origine marine et terrestre par les différentescomposantes des réseaux trophiques benthiques aboutissant aux aux cinq espèces de poissons plats à 70-100 m deprofondeur au large du Rhône. Les erreurs-types associées aux signatures moyennes (δ13C et δ15N) de chaquecatégorie sont indiquées en gris. MOP = matière organique particulaire, pol = polychète, S = suspensivore, DS =dépositivore de surface, DsS = dépositivore de sub-surface, C = carnivore, pK = plancton.Figure III.12 – Signatures isotopiques moyennes des cinq espèces de poissons plats étudiées au large du Rhône :Graphe δ13C vs δ15N obtenu à partir des signatures de 10 juvéniles G0 et 10 adultes de chaque espèce. Les erreurs-types associées aux signatures moyennes (δ13C et δ15N) de chaque poisson sont indiquées en gris.Figure III.13 - Régression linéaire significative (r = 0,622, r2 = 0,387 ; P < 0,001) obtenue entre le δ15N (en ‰) etla taille (LT, en mm) chez Solea solea au large du Rhône.Figure III.14 – Représentation en boites de Tukey des δ15N (‰) observés chez les plus petits juvéniles (G0 : 30-90 mm de LT) et les adultes (LT ≥ 262 mm) de Solea solea au large du Rhône. NT = niveau trophique calculé àpartir du δ15N moyen.Figure III.15 – Composition pondérale (Cp) du régime alimentaire des juvéniles G0 (a) et des adultes (b) de S.solea au large du Rhône.Figure III.16 - Réseaux trophiques des juvéniles G0 et des adultes de S. solea au large du Rhône. MOP = matièreorganique particulaire, brach. = brachyoure, pol = polychète, S = suspensivore, DS = dépositivore de surface, DsS= dépositivore de sub-surface, C = carnivore. Les flèches indiquent les principales voies de transfert de matièreorganique déterminées par l'étude des contenus stomacaux des poissons (proies dont Cp > 10%).Figure III.17 - Représentation en boites de Tukey des δ13C obtenus pour les individus de Solea solea de LT >200mm présents au large du Rhône, en fonction de la profondeur de récolte des poissons. Les valeurs entreparenthèses correspondent aux δ13C moyens ± erreur-types obtenus à chaque profondeur.Figure III.18 - Composition pondérale du régime alimentaire (Cp), réseau trophique et transfert de la matière chezles adultes de S. solea ( LT ≥ 260 mm) à 0-20 m (a), 30-50 m (b) et 70-100 m (c) de profondeur au large du Rhône.Les flèches indiquent les transferts alimentaires de matière organique.Figure III.19 - Régression linéaire significative (r = 0,458, r2 = 0,210 ; P < 0,05) obtenue entre le δ15N (en ‰) etla taille (LT, en mm) chez Solea impar au large du Rhône (0-20 m).Figure III.20 – Représentation en boites de Tukey des δ15N (‰) observés chez les plus petits juvéniles (G0 : 29-72 mm de LT) et les adultes (LT ≥ 162 mm) de Solea impar au large du Rhône (0-20 m). NT = niveau trophiquecalculé à partir du δ15N moyen.Figure III.21 - Régression linéaire significative (r = 0,804, r2 = 0,646 ; P < 0,001) obtenue entre le δ13C (en ‰) etla taille (LT, en mm) chez Solea impar au large du Rhône (0-20 m).Figure III.22 – Représentation en boites de Tukey des δ13C (‰) observés chez les plus petits juvéniles (G0 : 29-72 mm de LT) et les adultes (LT ≥ 162 mm) de Solea impar au large du Rhône (0-20 m). Les valeurs entreparenthèses correspondent aux δ13C moyens ± erreur-types obtenus à chaque profondeur.Figure III.23 – Composition pondérale (Cp) du régime alimentaire des juvéniles G0 (a) et des adultes (b) de S.impar au large du Rhône.

305
Figure III.24 - Réseaux trophiques des juvéniles G0 (Si J) et des adultes (Si A) de S. impar au large du Rhône.MOP = matière organique particulaire, pol = polychète, S = suspensivore, DS = dépositivore de surface, C =carnivore. Les flèches indiquent les principales voies de transfert de matière organique déterminées par l'étude descontenus stomacaux des poissons (proies dont Cp > 10%).Figure III.25 - Régression linéaire significative (r = 0,458, r2 = 0,210 ; P < 0,05) obtenue entre le δ15N (en ‰) etla taille (LT, en mm) chez Buglossidium luteum au large du Rhône.Figure III.26 – Représentation en boites de Tukey des δ15N (‰) observés chez les plus petits juvéniles (G0 : 30-45 mm de LT) et les adultes (LT ≥ 94 mm) de Buglossidium luteum au large du Rhône. NT = niveau trophiquecalculé à partir du δ15N moyen.Figure III.27 - Régression linéaire significative (r = 0,485, r2 = 0,235 ; P < 0,01) obtenue entre le δ13C (en ‰) etla taille (LT, en mm) chez Buglossidium luteum au large du Rhône.Figure III.28 – Représentation en boites de Tukey des δ13C (‰) observés chez les plus petits juvéniles (G0 : 30-45 mm de LT) et les adultes (LT ≥ 94 mm) de Buglossidium luteum au large du Rhône (0-100 m). Les valeurs entreparenthèses correspondent aux δ13C moyens ± erreur-types obtenus à chaque profondeur.Figure III.29 – Composition pondérale (Cp) du régime alimentaire des juvéniles G0 (a) et des adultes (b) de B.luteum au large du Rhône.Figure III.30 - Réseaux trophiques des juvéniles G0 (Bl J) et des adultes (Bl A) de B. luteum au large du Rhône.MOP = matière organique particulaire, brach. = brachyoure, pol = polychète, S = suspensivore, DS = dépositivorede surface, DsS = dépositivore de sub-surface, C = carnivore. Les flèches indiquent les principales voies detransfert de matière organique déterminées par l'étude des contenus stomacaux des poissons (proies dont Cp >10%).Figure III.31 - Représentation en boites de Tukey des δ13C obtenus pour les individus de B. luteum de LT ≥ 94mm présents au large du Rhône, en fonction de la profondeur de récolte des poissons. Les valeurs entreparenthèses correspondent aux δ13C moyens ± erreur-types obtenus à chaque profondeur.Figure III.32- Composition pondérale du régime alimentaire (Cp), réseau trophique et transfert de la matière chezles adultes de B. luteum ( LT ≥ 94 mm) à 0-20 m (a), 30-50 m (b) et 70-100 m (c) de profondeur au large duRhône. Les flèches indiquent les transferts alimentaires de matière organique.Figure III.33 - Régression linéaire significative (r = 0,820, r2 = 0,672 ; P < 0,001) obtenue entre le δ15N (en ‰) etla taille (LT, en mm) chez Arnoglossus laterna au large du Rhône.Figure III.34 – Représentation en boites de Tukey des δ15N (‰) observés chez les plus petits juvéniles (G0 : 32-49 mm de LT) et les adultes (LT ≥ 97 mm) d'A. laterna au large du Rhône. NT = niveau trophique calculé à partirdu δ15N moyen.Figure III.35 – Composition pondérale (Cp) du régime alimentaire des juvéniles G0 (a) et des adultes (b) de A.laterna au large du Rhône. C = carnivores, DS = dépositivores de surface, DsS = dépositivores de sub-surface, S =suspensivores.Figure III.36 - Réseaux trophiques des juvéniles G0 (Al J) et des adultes (Al A) d'A. laterna au large du Rhône.MOP = matière organique particulaire, brach. = brachyoure, pol = polychètes. DS = dépositivore de surface, C =carnivore.Les flèches indiquent les principales voies de transfert de matière organique déterminées par l'étude descontenus stomacaux des poissons.Figure III.37 – Représentation en boites de Tukey des δ15N (‰) observés chez les adultes (LT ≥ 97 mm) d'A.laterna présents au large du Rhône, en fonction de la profondeur de récolte des poissons. Les valeurs entreparenthèses correspondent aux δ15N moyens ± erreur-types obtenus à chaque profondeur.Figure III.38 - Représentation en boites de Tukey des δ13C obtenus pour les individus d'A. laterna de LT ≥ 97 mmprésents au large du Rhône, en fonction de la profondeur de récolte des poissons. Les valeurs entre parenthèsescorrespondent aux δ13C moyens ± erreur-types obtenus à chaque profondeur.Figure III.39 - Composition pondérale du régime alimentaire (Cp), réseau trophique et transfert de la matière chezles adultes d' A. laterna ( LT ≥ 97 mm) à 0-20 m (a), 30-50 m (b) et 70-100 m (c) de profondeur au large du Rhône.Les flèches indiquent les transferts alimentaires de matière organique.Figure III.40 - Régression linéaire significative (r = 0,979, r2 = 0,958 ; P < 0,001) obtenue entre le δ15N (en ‰) etla taille (LT, en mm) chez Citharus linguatula au large du Rhône.Figure III.41 – Représentation en boites de Tukey des δ15N (‰) observés chez les plus petits juvéniles (G0 : 63-84 mm de LT) et les adultes (LT ≥ 192 mm) de C. linguatula au large du Rhône. NT = niveau trophique calculé àpartir du δ15N moyen.

306
Figure III.42 – Composition pondérale (Cp) du régime alimentaire des juvéniles G0 (a) et des adultes (b) de C.linguatula au large du Rhône.Figure III.43 - Réseaux trophiques des juvéniles G0 (Cl J) et des adultes (Cl A) de C. linguatula au large duRhône (30-100 m de profondeur). Les flèches indiquent les voies de transfert de matière organique déterminéespar l'étude des contenus stomacaux des poissons. MOP = matière organique particulaire, pol = polychètes, DS =dépositivore de surface, DsS = carnivore, "min." et "max." désignent respectivement les individus de tailleminimale et maximale observés pour chaque catégorie de proie.Figure III.44 - Représentation en boites de Tukey des δ15N obtenus pour les juvéniles et les adultes de C.linguatula au large du Rhône, en fonction de la profondeur de récolte des poissons.Figure III.45 - Signatures isotopiques (δ13C et δ15N, en ‰) des sources potentielles et des pools de matièreorganique présents au niveau de l'eau de surface et du sédiment superficiel au large du Rhône.Figure III.46 – Droites de régression δ15N = f (LT) obtenues pour les cinq poissons étudiés. Dans chaque cas, lalongueur de la droite indique l'intervalle de taille couvert par les individus de l'espèce au large du Rhône.Figure III.47 – Composition pondérale du régime alimentaire des juvéniles G0 (J) et des adultes (A) des cinqpoissons plats au large du Rhône. Pour chaque espèce, la flèche indique s'il y a modification significative (*) ounon (ns) de la signature en carbone (δ13C, en ‰) entre les deux stades de vie benthique.Figure III.48 – Schéma récapitulatif de la structure des réseaux trophiques benthiques au large du Rhône. CI =consommateur primaire, CII = consommateur secondaire. Les numéros indiquent les trois groupes de poissonsindividualisés suivant leur alimentation : 1 = poissons consommant majoritairement CI et/ou CII "terrestres"; 2 =poissons consommant surtout CI et/ou CII "marins" + des quantités moindres de CI et/ou CII "terrestres"; 3 =poissons consommant presque exclusivement CI et/ou CII "marins".
PARTIE IV
Figure IV.1 – Débits moyens annuels (courbe : moyenne en noir ± erreur-type en gris) et nombres annuels dejours de crues (histogrammes) du Rhône pour la période 1980-2001, calculés d'après les observations à Beaucairede la Compagnie Nationale du Rhône. La droite en pointillés indique le débit annuel moyen (Dmoy.= 1770 ± 83 m3
s-1) obtenu sur l'ensemble de la période 1980-2001.Figure IV.2 – délais de réponse des différents groupes de polychètes échantillonnés au cours de la présente étudeaux crues du Rhône (d'après Salen-Picard et Arlhac, 2002 et Salen-Picard et al., 2002).Figure IV.3 – Influence des apports rhodaniens sur les écosystèmes côtiers et sur le cycle de la sole commune,Solea solea. MOP = matière organique particulaire. D'après Salen-Picard et al. (2001).Figure IV.4 – Relation entre le débit du Rhône et les débarquements de sole (Solea solea). (a) courbes des écartscumulés à la moyenne obtenues pour le débit annuel moyen du Rhône et les débarquements annuels moyens desole à Martigues (Golfe du Lion). (b) Evolution du coefficient de corrélation de Spearman (R) en fonction dudécalage entre ces deux paramètres. "*" = P < 0,05 ;"**" = P < 0,01 ; "***" = P < 0,001. D'après Salen-Picard etal. (2001).

307
LISTE DES TABLEAUX(Au total, 76 tableaux)
PARTIE I
Tableau I.1 – Principales caractéristiques biologiques et écologiques des cinq poissons choisis pour cette étude.P = polychètes, M = mollusques, C = crustacés, Po = poissons.Tableau I.2 – Récapitulatif des principaux types de missions réalisées entre mars 2000 et novembre 2001. "*" =la mission SOLMAR 4 (novembre 2000) a été interrompue à cause d'une importante quantité de filets calés par lespêcheurs sur la zone d'échantillonnage. Non complets, les quelques échantillons récoltés au cours de cettemission ont finalement été écartés des résultats.Tableau I.3 – Récapitulatif des principales missions d'échantillonnage des poissons.Tableau I.4 – Missions d'échantillonnage de l'eau de surface (dans le Rhône, en mer et à l'embouchure) et dusédiment supérieur (à l'embouchure). Les crues du fleuve sont surlignées en gris.Tableau I.5 – Récapitulatif des principales missions d'échantillonnage des communautés d'invertébrésbenthiquesTableau I.6 – Utilisation et nombre des prélèvements de chaque type (traits de chaluts, de senne de plage, despatangue…) effectués à chaque saison et à chacune des différentes stations échantillonnées.
PARTIE IICHAPITRE I
Tableau II/I.1 – Données bibliographiques utilisées pour l'établissement des groupes de taille (LT, mm)correspondant aux trois "stades" de vie benthique (juvéniles G0, juvéniles G1+, adultes) étudiés chez S. solea, S.impar, B. luteum, A. laterna et C. linguatula au large du Rhône.Tableau II/I.2 – Densités moyennes globales (moyenne ± erreur-type) obtenues sur l'ensemble de la zoned'étude (0-100 m de profondeur) pour les cinq espèces étudiées. Données SOLMAR (84 traits de chalut àpanneaux). Les barres horizontales grises indiquent les densités moyennes non statistiquement différentes (P ≥0,05).Tableau II/I.3 – Fluctuations bathymétriques des densités (D moy. ± erreur-type, en ind. ha-1) des cinq espècesde poissons plats étudiées au large du Rhône. Les barres horizontales grises indiquent les densités moyennes nonstatistiquement différentes (P ≥ 0,05). "" = espèce absente à cette profondeur. N = nombre de traits de chalutsà panneaux réalisés à chaque profondeur.Tableau II/I.4 – Fluctuations spatio-temporelles au large du Rhône de la densité totale (DT, en ind. ha-1) des cinqespèces groupées.Tableau II/I.5 – Pourcentages des différents stades de vie benthique retenus (juvéniles G0, juvéniles G1+, sub-adultes et adultes) dans les captures des cinq espèces de poissons plats étudiées au large du Rhône. Pour chaqueespèce, le pourcentage le plus élevé est indiqué en gras.Tableau II/I.6 – Tailles moyennes (LT en mm) et erreurs-types des individus des cinq espèces à chaqueprofondeur étudiée au large du Rhône. Les barres horizontales grises indiquent les tailles moyennes nonstatistiquement différentes (P ≥ 0,05). "" = espèce absente à cette profondeur.Tableau II/I.7 – Répartition bathymétrique des différents stades de vie benthique (juvéniles G0, juvéniles G1+ etadultes) des cinq espèces étudiées au large du Rhône. "" = stade absent à cette profondeur pour l'espèceconcernée. Dans chaque cas, le pourcentage maximal d'individus obtenu pour chaque stade est indiqué en gras.
CHAPITRE II
Tableau II/II.1 - Régime alimentaire global de S. solea au large du Rhône. F = fréquence ; N = nombre deproies ; Cn = pourcentage numérique ; P = poids sec des proies (mg) ; Cp = pourcentage pondéral ; Q =coefficient alimentaire (Q = Cn × Cp). "+" = valeur inférieure à 0,01.

308
Tableau II/II.2 - Variations de l'alimentation de S. solea en fonction de la taille. Q = coefficient alimentaire. "+"= valeur inférieure à 0,01 ; "" = valeur absente. LT = longueur totale des individus (en mm).Tableau II/II.3 - Régime alimentaire global de S. impar au large du Rhône. F = fréquence ; N = nombre deproies ; Cn = pourcentage numérique ; P = poids sec des proies (mg) ; Cp = pourcentage pondéral ; Q =coefficient alimentaire des proies (Q = Cn × Cp).Tableau II/II.4 - Variations de l'alimentation de S. impar en fonction de la taille. Q = coefficient alimentaire desproies (Q = Cn × Cp). "" = valeur absente. LT = longueur totale (en mm).Tableau II/II.5 - Régime alimentaire global de B. luteum au large du Rhône. F = fréquence ; N = nombre deproies ; Cn = pourcentage numérique ; P = poids sec des proies (mg) ; Cp = pourcentage pondéral ; Q =coefficient alimentaire des proies (Q = Cn × Cp). "+" = valeur inférieure à 0,01.Tableau II/II.6 - Variations de l'alimentation de B. luteum en fonction de la taille. Q = coefficient alimentaire."+" = valeur inférieure à 0,01 ; "" = valeur absente. LT = longueur totale (en mm).Tableau II/II.7 - Régime alimentaire global d'A. laterna au large du Rhône. F = fréquence ; N = nombre deproies ; Cn = pourcentage numérique ; P = poids sec des proies (mg) ; Cp = pourcentage pondéral ; Q =coefficient alimentaire des proies (Q = Cn × Cp). "+" = valeur inférieure à 0,01.Tableau II/II.8 - Variations de l'alimentation d'A. laterna en fonction de la taille. Q = coefficient alimentaire."+" = valeur inférieure à 0,01 ; "" = valeur absente. LT = longueur totale des individus (en mm).Tableau II/II.9 - Régime alimentaire global de C. linguatula au large du Rhône. F = fréquence ; N = nombre deproies ; Cn = pourcentage numérique ; P = poids sec des proies (mg) ; Cp = pourcentage pondéral ; Q =coefficient alimentaire des proies (Q = Cn × Cp). "+" = valeur inférieure à 0,01.Tableau II/II.10 - Variations de l'alimentation de C. linguatula en fonction de la taille. Q = coefficientalimentaire. "+" = valeur inférieure à 0,01 ; "" = valeur absente. LT = longueur totale des individus (en mm).Tableau II/II.11 - Stratégies alimentaires des cinq espèces au large du Rhône. Np = nombre total desproies/estomac ; Pm proies = poids individuel moyen des proies ; med. = médiane.Tableau II/II.12 - Modifications ontogéniques de l'alimentation des cinq espèces au large du Rhône. Pm proies= poids moyen des proies ingérées ; Pp = poids total des proies/estomac ; Np = nombre total des proies/estomac; med. = médiane. A = amphipodes ; B = bivalves ; Co = copépodes ; Cu = cumacés ; Ma = macroures ; My =mysidacés ; P = polychètes ; Ph = phoronidiens ; Po = poissons. DSS = dépositivores de sub-surface ; DS =dépositivores de surface ; C = carnivores ; S = suspensivores.Tableau II/II.13 - Principales proies (Cp ≥ 10%) des cinq espèces aux différentes profondeurs étudiées au largedu Rhône. A = amphipodes ; B = bivalves ; Br = brachyoures ; Co = copépodes ; Cu = cumacés ; Ma =macroures ; My = mysidacés ; P = polychètes ; Po = poissons. Les proies préférentielles sont indiquées en graspour chaque espèce.Tableau II/II.14 - Principales proies (Cp ≥ 10%) observées à chaque saison pour les cinq espèces étudiées aularge du Rhône. A = amphipodes ; B = bivalves ; Br = brachyoures; Cu = cumacés ; Ma = macroures ; My =mysidacés ; P = polychètes ; Po = poissons. Les proies préférentielles sont indiquées en gras pour chaque espèce.
CHAPITRE III
Tableau II/III.1 - Largeurs des niches alimentaires globales réalisées (BG) et variations intraspécifiques deslargeurs de niches alimentaires (B) chez les cinq espèces au large du Rhône en fonction de la taille des individus,de la profondeur et de la saison. P = printemps, E = été, H = hiver, " " = valeur absente. Pour chaque espèceles largeurs de niche minimale et maximale observées sont indiquées en gras, les valeurs faibles (B < 5) en blanc,les valeurs moyennes (5 < B < 11) en gris clair et les valeurs élevée (B > 11) en gris foncé.Tableau II/III.2 - Indices de recouvrement globaux théoriques des niches alimentaires (TT) calculés au large duRhône pour les cinq espèces prises deux à deux.Tableau II/III.3 - Indices de recouvrement des niches alimentaires (T) observés au printemps à 0-20 m deprofondeur, pour les stades de vie benthique des espèces présentes, pris deux à deux. S s = S. solea ; S i = S.impar ; B l = B. luteum ; A l = A. laterna ; C l = C. linguatula. G0 = juvéniles de l'année ; G1+ = juvéniles de plusd'un an ; Ad. = adultes ; LT = longueur totale en mm.Tableau II/III.4 - Pourcentages pondéraux (Cp) des différentes catégories de proies ingérées au printemps à 0-20 m par les 10 groupes de poissons présents à cette profondeur et à cette saison. S s = S. solea ; S i = S. impar ;B l = B. luteum ; A l = A. laterna ; C l = C. linguatula. G0 = juvéniles de l'année ; G1+ = juvéniles de plus d'un an; Ad. = adultes ; LT = longueur totale en mm; "" = proie absente pour ce groupe de poissons.

309
Tableau II/III.5 - Indices de recouvrement des niches alimentaires (T) observés en été et à 0-20 m deprofondeur pour les stades de vie benthique des espèces présentes, pris deux à deux. S s = S. solea ; S i = S.impar ; B l = B. luteum ; A l = A. laterna ; C l = C. linguatula. G0 = juvéniles de l'année ; G1+ = juvéniles de plusd'un an ; Ad. = adultes ; LT = longueur totale en mm.Tableau II/III.6 - Indices de recouvrement des niches alimentaires (T) observés en hiver à 0-20 m de profondeurpour les stades de vie benthique des espèces présentes, pris deux à deux. S s = S. solea ; B l = B. luteum ; A l =A. laterna. Ad. = adultes.Tableau II/III.7 - Poids moyens (mg) des proies ingérées à 0-20 m par deux groupes de poissons ou plus. Pourchaque groupe de poissons, seules sont indiquées les catégories de proies représentant plus de 20% en poids dupoids total des proies ingérées, avec en gras, les proies préférentielles. S s = S. solea ; S i = S. impar ; B l = B.luteum ; A l = A. laterna ; C l = C. linguatula. G0 = juvéniles de l'année ; G1+ = juvéniles de plus d'un an ; Ad. =adultes ; LT = longueur totale en mm.Tableau II/III.8 - Indices de recouvrements globaux effectifs des niches alimentaires (TE) des populations desespèces présentes à 0-20 m de profondeur, prises deux à deux.Tableau II/III.9 - Indices de recouvrement des niches alimentaires (T) observés au printemps à 30-50 m deprofondeur pour les stades de vie benthique des espèces présentes, pris deux à deux. "+" = valeur inférieure à0,01. S s = S. solea ; B l = B. luteum ; A l = A. laterna ; C l = C. linguatula. G0 = juvéniles de l'année ; G1+ =juvéniles de plus d'un an ; Ad. = adultes ; LT = longueur totale en mm.Tableau II/III.10 - Indices de recouvrement des niches alimentaires (T) observés en été pour les stades de viebenthique des espèces présentes à 30-50 m de profondeur, pris deux à deux. "+" = valeur inférieure à 0,01. S s =S. solea ; B l = B. luteum ; A l = A. laterna ; C l = C. linguatula. G0 = juvéniles de l'année ; G1+ = juvéniles deplus d'un an ; Ad. = adultes ; LT = longueur totale en mm.Tableau II/III.11 - Indices de recouvrement des niches alimentaires (T) observés en hiver à 30-50 m deprofondeur entre les stades de vie benthique des espèces présentes, pris deux à deux. "+" = valeur inférieure à0,01. S s = S. solea ; B l = B. luteum ; A l = A. laterna ; C l = C. linguatula. G0 = juvéniles de l'année ; G1+ =juvéniles de plus d'un an ; Ad. = adultes ; LT = longueur totale en mm.Tableau II/III.12 - Poids moyen (mg) des proies ingérées à 30-50 m par deux groupes de poissons ou plus. Pourchaque groupe de poissons, seules sont indiquées les catégories de proies représentant plus de 20% en poids dupoids total des proies ingérées, avec en gras, les proies préférentielles. S s = S. solea ; B l = B. luteum ; A l = A.laterna ; C l = C. linguatula. G0 = juvéniles de l'année ; G1+ = juvéniles de plus d'un an ; Ad. = adultes ; LT =longueur totale en mm.Tableau II/III.13 - Indices de recouvrement globaux effectifs des niches alimentaires (TE) observés entre lespopulations des espèces présentes à 30-50 m de profondeur, prises deux à deux. "+" = valeur inférieure à 0,01.Tableau II/III.14 - Indices de recouvrement des niches alimentaires (T) observés au printemps pour les stadesde vie benthique des espèces présentes à 70-100 m de profondeur, pris deux à deux. "+" = valeur inférieure à0,01. S s = S. solea ; B l = B. luteum ; A l = A. laterna ; C l = C. linguatula. G0 = juvéniles de l'année ; G1+ =juvéniles de plus d'un an ; Ad. = adultes.Tableau II/III.15 - Indices de recouvrement des niches alimentaires (T) observés en été à 70-100 m deprofondeur entre les stades de vie benthique des espèces présentes, pris deux à deux. A l = A. laterna ; C l = C.linguatula. G1+ = juvéniles de plus d'un an ; Ad. = adultes.Tableau II/III.16 - Indices de recouvrement des niches alimentaires (T) observés en hiver à 70-100 m deprofondeur entre les stades de vie benthique des espèces présentes, pris deux à deux. "+" = valeur inférieure à0,01. S s = S. solea ; B l = B. luteum ; A l = A. laterna ; C l = C. linguatula.. G1+ = juvéniles de plus d'un an ; Ad.= adultes.Tableau II/III.17 - Poids moyen (mg) des proies ingérées à 70-100 m par deux groupes de poissons ou plus.Pour chaque groupe de poissons, seules sont indiquées les catégories de proies représentant plus de 20% en poidsdu poids total des proies ingérées, avec en gras, les proies préférentielles. S s = S. solea ; B l = B. luteum ; A l =A. laterna ; C l = C. linguatula. G0 = juvéniles de l'année ; G1+ = juvéniles de plus d'un an ; Ad. = adultes.Tableau II/III.18 - Indices de recouvrement globaux effectifs des niches alimentaires (TE) observés pour lespopulations des cinq espèces présentes à 70-100 m de profondeur, prises deux à deux. "+" = valeur inférieure à0,01.Tableau II/III.19 - Indices de recouvrement globaux effectifs des niches alimentaires (TE) observés surl'ensemble de la zone d'étude (0-100 m) entre les populations des cinq espèces, prises deux à deux. "+" = valeurinférieure à 0,01 ; "─" = valeur absente.

310
PARTIE III
Tableau III.1 - Abondances naturelles moyennes des isotopes stables du carbone et de l'azote (d'après Riera,1995).Tableau III.2 - Compositions isotopiques (δ en ‰) typiques en carbone et en azote des principaux producteursprimaires en milieu terrestre et aquatique. D'après la revue réalisée par Ostrom et Fry (1993).Tableau III.3 – signatures en carbone (δ13C) et en azote (δ15N) des sources potentielles de matière organiquerecensées au large du Rhône et de la matière organique présente dans l'eau de surface et le sédiment superficielde la zone d'étude. MOP = matière organique particulaire.Tableau III.4 – Estimations globales des contributions (%) respectives de la MOP terrigène et du phytoplanctonmarin à la constitution du pool de matière organique du sédiment au large du Rhône. Résultats obtenus pour leséquations de mélange réalisées à partir de quatre combinaisons de signatures en carbone associées à ces deuxsources.Tableau III.5 – Signatures en carbone (δ13C : moyenne ± erreur-type) des deux principales source, terrestre etmarine, de matière organique au large du Rhône et de la matière organique présente dans l'eau de surface et lesédiment superficiel de la zone d'étude, en période de crues (décembre, mars) et en période d'étiage (septembre).MOP = matière organique particulaire, N = effectif analysé.Tableau III.6 – signatures en carbone (δ13C moyen ± erreur-type, en ‰) de la matière organique présente dansl'eau de surface et le sédiment superficiel de la zone d'étude à chaque tranche bathymétrique étudiée (0-20 m, 30-50 m, 70-100 m). MOP = matière organique particulaire.Tableau III.7 – Estimations globales des contributions (%) respectives de la MOP terrigène et du phytoplanctonmarin à la constitution des pools de matière organique de l'eau de surface et du sédiment à 0-20 m, 30-50 m et70-100 m de profondeur au large du Rhône.Tableau III.8 - Signatures en azote (δ15N moyen ± erreur-type, en ‰) observées en période de crue et enpériode d'étiage pour les 2 principales sources de matière organique, d'origine terrestre (MOP Rhône) et marine(phytoplancton), et pour l'eau de surface et le sédiment superficiel de la zone d'étude à chaque tranchebathymétrique échantillonnée (0-20 m, 30-50 m, 70-100 m). MOP = matière organique particulaire; Sed. =sédiment.Tableau III.9 – Signatures en carbone (δ13C, ‰) des principales composantes des réseaux trophiques benthiquesobtenues à chaque profondeur (0-20 m, 30-50 m et 70-100 m) au large du Rhône. N = effectif analysé.Tableau III.10 – Signatures en azote (δ15N, ‰) des principales composantes des réseaux trophiques benthiquesobtenues à chaque profondeur (0-20 m, 30-50 m et 70-100 m) au large du Rhône. N = effectif analysé.Tableau III.11 – Signatures isotopiques moyennes en azote et en carbone observées, et niveau trophique et δ13Cthéoriques calculés dans le cas d'une alimentation d'origine exclusivement "marine" pour les juvéniles G0 (LT <100 mm)et les adultes de S. solea au large du Rhône.Tableau III.12 – Signatures isotopiques théoriques en azote et en carbone calculées en fonction de l'alimentationdes individus et δ13C et δ15N moyens observés pour les juvéniles G0 et les adultes de S. solea au large du Rhône.(pour les calculs, voir chapitre matériel et méthodes)Tableau III.13 – Signatures isotopiques moyennes en azote et en carbone observées, et niveau trophique et δ13Cthéoriques calculés dans le cas d'une alimentation d'origine exclusivement "marine" pour les juvéniles G0 et lesadultes de S. impar au large du Rhône.Tableau III.14 – Signatures isotopiques théoriques en azote et en carbone calculées en fonction de l'alimentationdes individus et δ13C et δ15N moyens observés pour les juvéniles G0 et les adultes de S. impar au large duRhône. (pour les calculs, voir chapitre matériel et méthodes)Tableau III.15 – Signatures isotopiques moyennes en azote et en carbone observées, et niveau trophique et δ13Cthéoriques calculés dans le cas d'une alimentation d'origine exclusivement "marine" pour les juvéniles G0 et lesadultes de B. luteum au large du Rhône.Tableau III.16 – Signatures isotopiques théoriques en azote et en carbone calculées en fonction de l'alimentationdes individus et δ13C et δ15N moyens observés pour les juvéniles G0 et les adultes de B. luteum au large duRhône. (pour les calculs, voir chapitre matériel et méthodes).Tableau III.17 – Signatures isotopiques moyennes en azote et en carbone observées, et niveau trophique et δ13Cthéoriques calculés dans le cas d'une alimentation d'origine exclusivement "marine" pour les juvéniles G0 et lesadultes d'A. laterna au large du Rhône.

311
Tableau III.18 – Signatures isotopiques théoriques en azote et en carbone calculées en fonction de l'alimentationdes individus et δ13C et δ15N moyens observés pour les juvéniles G0 et les adultes d'A. laterna au large duRhône. (pour les calculs, voir chapitre matériel et méthodes)Tableau III.19 – Signatures isotopiques moyennes en azote et en carbone observées, et niveau trophique et δ13Cthéoriques calculés dans le cas d'une alimentation d'origine exclusivement "marine" pour les juvéniles G0 et lesadultes de C. linguatula au large du Rhône.Tableau III.20 – Signatures isotopiques théoriques en azote et en carbone calculées en fonction de l'alimentationdes individus et δ13C et δ15N moyens observés pour les juvéniles G0 et les adultes de C. linguatula au large duRhône. (pour les calculs, voir chapitre matériel et méthodes)Tableau III.21 – Signatures isotopiques (carbone et azote) et rapports C/N moyens (± erreur-type) des eauxrhodaniennes en période de crue et en période d'étiage.Tableau III.22 – Signatures isotopiques (carbone et azote) et rapports C/N moyens (± erreurs-types) des eauxmarines au large (Frioul) en période de crue (printemps-hiver) et en période d'étiage (été).Tableau III.23 – Signatures en carbone, température et salinité des eaux rhodaniennes et marines comparées àcelles de l'eau de surface à 0-20 m, 30-50 m et 70-100 m de profondeur au large du Rhône, en période de crue eten période d'étiage.Tableau III.24 – Signatures isotopiques en carbone et rapports C/N des eaux rhodaniennes et marines comparésà ceux de la MOP de l'eau de surface et du sédiment superficiel à 0-20 m, 30-50 m et 70-100 m de profondeur aularge du Rhône, en période de crue et en période d'étiage.Tableau III.25 – Niveaux trophiques moyens (NT) des poissons obtenus en utilisant le δ15N, soit duphytoplancton marin, soit de la MOP rhodanienne, soit d'un mélange équilibré des deux sources, commesignature de référence des producteurs primaires dans les calculs (cf. chapitre matériel et méthodes). Danschaque cas, les valeurs en gras indiquent le niveau trophique le plus probable au large du Rhône.Tableau III.26 – Gamme de δ15N obtenue et niveaux trophiques (NT) estimés pour les organismes hétérotrophesrécoltés au large du Rhône : comparaison avec les données connues sur le régime alimentaire et le niveautrophique des animaux. "MOP" = matière organique particulaire; "C I", "C II", "C III"… = consommateurprimaire, secondaire, tertiaire (etc).Tableau III.27 – δ15N et niveaux trophiques moyens (NT) obtenus pour les juvéniles et les adultes des cinqpoissons au large du Rhône. Dans chaque cas, le δ15N de la source principale de matière organique indiquée aservi de signature de référence pour les producteurs primaires dans les calculs (cf. chapitre matériel etméthodes). En cas de mélange de deux sources, la moyenne des deux signatures a été utilisée. Pour chaqueespèce, les Pour chaque espèce, les NT fournis sur le site de Fishbase (Froese et Pauly, 2000) sont indiqués à titrecomparatif.
PARTIE IV
Tableau IV.1 – Poids moyen des proies ingérées et niveau trophique calculé pour les juvéniles, les adultes etl'ensemble des individus de chaque espèce au large du Rhône. En cas de modification ontogénique sensible del'un de ces deux paramètres, la différence observée est indiquée sous la flèche d'évolution correspondante.Tableau IV.2 - Stratégies de vie adoptées par les juvéniles et les adultes des cinq espèces étudiées au large duRhône. P = printemps, E = été, H = hiver, A = amphipodes, B = bivalves, Co = copépodes, Cu = cumacés, Ma =macroures, My = mysidacés, Po = poissons, Pol. = polychètes (C = carnivores, D = dépositivores, S =suspensivores).Tableau IV.3 – Liste des principaux polluants apportés par le Rhône en Méditerranée et effets recensés sur lesorganismes marins.

312

ANNEXES


Annexe 1 – Relations d'allométrie reliant la longueur standard (LS, en mm) et le poids (P, en g) des individus àleur longueur totale (LT, en mm) obtenues pour Solea solea, Solea impar, Buglossidium luteum, Arnoglossuslaterna et Citharus linguatula au large du Rhône. r = coefficient de corrélation ; P = probabilité associée; N =effectif utilisé pour l'établissement de la relation.
Relationlongueur totale - longueur standard
Relationpoids - longueur totale N
S. soleaLT = 0,878 × LS - 1,646(r = 0,999 ; P < 0,001)
P = 6.10-6 × LT 3,069
(r = 0,997 ; P < 0,001) 514
S. impar LT = 1,136 × LS + 2,158(r = 0,999 ; P < 0,001)
P = 3.10-6 × LT 3,228
(r = 0,998 ; P < 0,01) 138
B. luteumLT = 1,190 × LS + 0,646(r = 0,996 ; P < 0,001)
P = 6.10-6 × LT 3,114
(r = 0,987 ; P < 0,001) 231
A. laterna LT = 1,206 × LS + 0,356(r = 0,996 ; P < 0,001)
P = 2.10-5 × LT 2,801
(r = 0,961 ; P < 0,001) 1922
C. linguatulaLT = 0,817 × LS - 0,849(r = 0,999 ; P < 0,001)
P = 3.10-6 × LT 3,186
(r = 0,993 ; P < 0,001) 282
313

Annexe 2 – Modifications spatio-temporelles de la densité totale des 5 espèces de poissons plats étudiées aularge du Rhône.
(a) MANOVA réalisée sur les effectifs totaux des 5 espèces récoltées à chaque saison (hiver, printemps, été) pour chaqueprofondeur (0-20 m, 30-50 m, 70-100 m).
(b) Résultats des tests de comparaisons a posteriori par saison (H = hiver, P = printemps, E = été), par profondeur (P1 =0-20 m, P2 = 30-50 m, P3 = 70-100 m), et par saison pour chaque profondeur.
(a)
STATISTICA. MANOVA GENERALESynthèse de tous les EffetsPlan: 1-SAISON, 2-PROF
dl Mc dl Mc
Effet Effet Effet Erreur Erreur F niveau P
1 2 ,152109 63 ,014147 10,75218 ,000096
2 2 ,143375 63 ,014147 10,13477 ,000153
1 × 2 4 ,081626 63 ,014147 5,76994 ,000507
(b)
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocEFFET PRINC.:PROFONDEUR
PROF P1 P2 P3
P1 ,000380 ,000141P2 ,000380 ,350680P3 ,000141 ,350680
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocEFFET PRINC.:SAISON
SAISON P H E
P ,000134 ,163655H ,000134 ,001083E ,163655 ,001083
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocINTERACTION: PROFONDEUR × SAISON
P1 × H P1 × P P1 × E P2 × H P2 × P P2 × E P3 × H P3 × P P3 × E
P1 × H ,000128 ,040424 ,530922 ,505050 ,504129 ,778872 ,716847 ,686166
P1 × P ,000128 ,003563 ,000172 ,000142 ,000222 ,000140 ,000186 ,000161
P1 × E ,040424 ,000563 ,255195 ,152964 ,173607 ,023807 ,156351 ,191676
P2 × H ,530922 ,000172 ,255195 ,684880 ,907086 ,528521 ,982253 ,976810
P2 × P ,505050 ,000142 ,152964 ,684880 ,815845 ,609287 ,935265 ,900245
P2 × E ,504129 ,000222 ,173607 ,907086 ,815845 ,404736 ,624519 ,796831
P3 × H ,778872 ,000140 ,023807 ,528521 ,609287 ,404736 ,639372 ,636343
P3 × P ,716847 ,000186 ,156351 ,982253 ,935265 ,624519 ,639372 ,880250
P3 × E ,686166 ,000161 ,191676 ,976810 ,900245 ,796831 ,636343 ,880250
314

Annexe 3 - Modifications spatio-temporelles des abondances relatives des 5 espèces étudiées au large duRhône.(a) MANOVA non paramétrique réalisée sur les effectifs des 5 espèces récoltés pour 6 traits de chalut (3 traits de jour et 3
de nuit) à chaque saison (hiver, printemps, été) pour chaque profondeur (0-20 m, 30-50 m, 70-100 m).
(b) Résultats des tests de comparaisons a posteriori par saison (H = hiver, P = printemps, E = été), par profondeur (P1 = 0-20 m, P2 = 30-50 m, P3 = 70-100 m) et par saison pour chaque profondeur.
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- Two-way crossed (orthogonal) ANOVA Factor 1 is saison with 3 levels Factor 2 is prof with 3 levels Factors 1 and 2 are fixed The no. of replicates = 6 The no. of variables = 5
--- Results --- Non-parametric Multivariate Analysis of Variance
Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- saison 2 8596.8150 4298.4075 18.3633 0.0002 >1.0E+10 Res prof 2 4577.8872 2288.9436 9.7786 0.0002 >1.0E+10 Res s x p 4 1314.8701 328.7175 1.4043 0.2064 >1.0E+10 Res Resid 45 10533.4071 234.0757 Total 53 25022.9794 -----------------------------------------------------------------------------
Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of residuals(reduced model) No. of permutations used = 4999
315

(b)
--- Results --- Pair-wise a posteriori comparisons
Tests among groups in factor 1 (saison)
Possible Average dissimilarities Groups t P No.perm. within/between groups ---------------------------------------------- ( H, P) 3.2749 0.0004 0.4538E+10 H P E ( H, E) 4.4314 0.0002 0.4538E+10 H 30.482 ( P, E) 2.5292 0.0012 0.4538E+10 P 32.705 18.051 ---------------------------------------------- E 37.371 20.498 17.740 No. of permutations used = 4999
Tests among groups in factor 2 (profondeur)
Possible Average dissimilarities Groups t P No.perm. within/between groups ----------------------------------------------- ( P1, P2) 2.9405 0.0018 0.4538E+10 P1 P2 P3 ( P1, P3) 1.9237 0.0374 0.4538E+10 P1 24.857 ( P2, P3) 2.1876 0.0084 0.4538E+10 P2 31.014 25.840 ----------------------------------------------- P3 27.196 28.275 24.236 No. of permutations used = 4999
Tests among groups in factor 1 (saison) within level 1 of factor 2 (P1)
Possible Average dissimilarities Groups t P No.perm. within/between groups --------------------------------------------- ( H, P) 5.3450 0.0016 462.0 H P E ( H, E) 6.0610 0.0028 462.0 H 14.176 ( P, E) 2.3549 0.0168 462.0 P 31.056 10.034 --------------------------------------------- E 41.422 16.755 15.166 No. of permutations used = 4999
Tests among groups in factor 1(saison) within level 2 of factor 2 (P2)
Possible Average dissimilarities Groups t P No.perm. within/between groups -------------------------------------------- ( H, P) 1.3858 0.1738 462.0 H P E ( H, E) 1.8411 0.0720 462.0 H 29.096 ( P, E) 1.4066 0.1646 462.0 P 28.468 23.089 -------------------------------------------- E 29.077 22.732 18.717 No. of permutations used = 4999
Tests among groups in factor 1 (saison) within level 3 of factor 2 (P3)
Possible Average dissimilarities Groups t P No.perm. within/between groups -------------------------------------------- ( H, P) 2.4303 0.0290 462.0 H P E ( H, E) 2.6680 0.0112 462.0 H 29.732 ( P, E) 1.3809 0.2098 462.0 P 31.545 11.610 -------------------------------------------- E 34.414 13.998 13.970 No. of permutations used = 4999
Note: The pair-wise tests have not been corrected for multiple comparisons.
316

Ann
exe
4 -
Com
posi
tion
du r
égim
e al
imen
taire
des
indi
vidu
s co
nstit
utifs
des
4 g
roup
es d
e ta
ille
(A, B
, B',
C)
indi
vidu
alis
és p
our
Sole
a so
lea
au
larg
e du
Rhô
ne p
aror
dina
tion
et c
lass
ifica
tion
des c
lass
es d
e ta
ille
(LT,
en m
m) d
e l'e
spèc
e à
parti
r de
l'alim
enta
tion
(poi
ds d
es p
roie
s ing
érée
s) d
e le
urs i
ndiv
idus
. F =
fréq
uenc
e de
s pro
ies,
Cn
= po
urce
ntag
e nu
mér
ique
, C
p =
pour
cent
age
pond
éral
, Q =
coe
ffic
ient
alim
enta
ire.
A :
LT <
100
mm
B :
100-
160
mm
B' :
160
-180
mm
B :
180-
200
mm
C :
LT >
200
mm
FC
pC
nQ
FC
pC
nQ
FC
pC
nQ
FC
pC
nQ
FC
pC
nQ
Olig
ochè
tes
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
0,00
0,0%
0,1%
0,0
Pol
ychè
tes
0,52
34,5
%14
,2%
489,
90,
7670
,3%
71,8
%50
48,1
0,72
44,0
%93
,5%
4113
,81,
0078
,0%
98,9
%77
08,7
0,86
75,3
%95
,0%
7154
,0
Sip
oncl
es--
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
-0,
012,
2%0,
0%0,
1
Cop
épod
es--
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
-0,
020,
0%0,
1%0,
0
Lep
tost
racé
s--
---
----
---
---
----
-0,
020,
4%0,
1%0,
0--
---
----
---
---
----
-0,
021,
2%0,
0%0,
1--
---
----
---
---
----
-
Mys
idac
és0,
035,
5%0,
3%1,
6--
---
----
---
---
----
---
---
----
---
---
----
-0,
081,
4%0,
0%0,
10,
020,
0%0,
1%0,
0
Cum
acés
0,58
17,7
%77
,7%
1374
,40,
020,
5%0,
4%0,
2--
---
----
---
---
----
-0,
150,
2%0,
0%0,
00,
050,
1%0,
1%0,
0
Iso
pode
s--
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
-0,
020,
9%0,
0%0,
00,
020,
0%0,
0%0,
0
Am
phip
odes
0,12
11,0
%2,
0%22
,20,
272,
1%7,
4%15
,40,
342,
2%0,
3%0,
60,
080,
6%0,
1%0,
10,
100,
1%0,
4%0,
0
Mac
rour
es--
---
----
---
---
----
-0,
021,
6%0,
2%0,
3--
---
----
---
---
----
---
---
----
---
---
----
-0,
033,
3%0,
1%0,
4
Ano
mou
res
----
---
---
----
---
---
----
---
---
----
---
---
0,06
0,4%
0,3%
0,1
0,08
4,3%
0,1%
0,6
0,02
0,5%
0,1%
0,0
Bra
chyo
ures
----
---
---
----
---
---
0,01
0,4%
0,1%
0,0
----
---
---
----
---
---
----
---
---
----
---
---
0,10
1,3%
0,6%
0,8
Cru
stac
é in
dét.
----
---
---
----
---
---
0,00
0,5%
0,2%
0,1
----
---
---
----
---
---
----
---
---
----
---
---
0,01
0,1%
0,0%
0,0
Biv
alve
s0,
3926
,6%
4,9%
130,
90,
6423
,7%
19,4
%46
0,1
0,69
8,2%
2,8%
22,6
0,62
8,8%
0,4%
4,0
0,47
5,4%
2,3%
12,5
Gas
téro
pode
s--
---
----
---
---
----
-0,
020,
4%0,
3%0,
10,
0937
,1%
1,0%
37,3
----
---
---
----
---
---
0,07
2,4%
0,3%
0,6
Mol
lusq
ue in
det.
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
0,01
0,2%
0,0%
0,0
Oph
iure
s0,
031,
5%0,
3%0,
4--
---
----
---
---
----
-0,
034,
6%1,
3%5,
80,
232,
5%0,
1%0,
20,
043,
1%0,
4%1,
2
Hol
othu
ries
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
0,04
5,7%
0,3%
1,8
Pho
roni
dien
s0,
003,
3%0,
6%1,
9--
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
-
Poi
sson
s--
---
----
---
---
----
---
---
----
---
---
----
-0,
063,
4%1,
0%3,
40,
152,
1%0,
1%0,
30,
050,
2%0,
1%0,
0
Pro
ies i
ndet
.0,
010,
1%0,
0%0,
0--
---
----
---
---
----
-0,
030,
2%0,
1%0,
00,
010,
1%0,
0%0,
00,
010,
1%0,
0%0,
0
317

Annexe 5 - Modifications du nombre de proies par estomac en fonction de la taille des individus chez Soleasolea au large du Rhône.
.
ANOVA non paramétrique de Kruskal-Wallis :
Groupe Somme des rangs effectifLT < 100 mm 4032,5 33100-200 mm 22994,5 177LT > 200 mm 17823 97
H (2, N = 307) = 29,63978 P = 0,000
Test de Dunn (test non paramétrique de comparaison des échantillons a posteriori) :
Effectif total = 307 GroupesLT < 100 mm 100-200 mm LT > 200 mm
Somme des rangs 4032,5 22994,5 17823effectif 33 177 97
Rang moyen 122,197 129,912 183,742
Matrice des SE : LT < 100 mm 100-200 mm100-200 mm 16,83
LT > 200 mm 17,89 11,21
Matrice des Q : LT < 100 mm 100-200 mm100-200 mm 0,46LT > 200 mm 3,44 4,80
Q théorique (0,05; k=3) : 2,394Q théorique (0,01; k=3) : 2,936 Rejet H0 si Q > Q théoriqueQ théorique (0,001; k=3) : 3,588
Matrice des résultats : LT < 100 mm 100-200 mm100-200 mm nsLT > 200 mm * * * * *
318

Annexe 6 - Modifications saisonnières de l'intensité de la nutrition chez Solea solea au large du Rhône.Variations de l'indice de réplétion stomacal (IR) en fonction de la saison.
ANOVA non paramétrique de Kruskal-Wallis :
Groupe Somme des rangs effectifPrintemps 21370 118
Eté 48915 226Hiver 13970 88
H (2, N = 432) = 7,7266 P = 0,0210
Test de Dunn (test non paramétrique de comparaison des échantillons a posteriori) :
Effectif total = 432 GroupesPrintemps Eté Hiver
Somme des rangs 21370 48915 13970effectif 118 226 88
Rang moyen 181,102 216,438 158,750
Matrice des SE : Printemps Eté
Eté 14,18Hiver 17,59 15,69
Matrice des Q : Printemps EtéEté 2,49
Hiver 1,27 3,68
Q théorique (0,05; k=3) : 2,394Q théorique (0,01; k=3) : 2,936 Rejet H0 si Q > Q théoriqueQ théorique (0,001; k=3) : 3,588
Matrice des résultats : Printemps EtéEté *
Hiver ns * * *
319

Annexe 7 - Modifications saisonnières de l'alimentation des juvéniles et adultes de Solea solea de plus de 100mm de LT au large du Rhône.
(a) MANOVA non paramétrique à deux facteurs (stade de vie benthique × saison) réalisée sur les contenus stomacaux(Poids des proies ingérées en mg) des individus de S. solea présents aux trois saisons étudiées (hiver, printemps, été).
(b) Résultats des tests de comparaisons a posteriori par saison (H = hiver, P = printemps, E = été) et par groupe (G2 = 100-200 mm, G3 = LT > 200 mm).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- Two-way crossed (orthogonal) ANOVA Factor 1 is groupe with 2 levels Factor 2 is saison with 3 levels Factors 1 and 2 are fixed The no. of replicates = 10 The no. of variables = 13
--- Results --- Non-parametric Multivariate Analysis of Variance
Possible Denom. Source df SS MS F P No.perm. MS ---------------------------------------------------------------------------- groupe 1 7376.1982 7376.1982 3.0891 0.0050 >1.0E+10 Res saison 2 11369.8242 5684.9121 2.3808 0.0104 >1.0E+10 Res gr x sa 2 5873.7448 2936.8724 1.2300 0.2566 >1.0E+10 Res Resid. 54 128940.6659 2387.7901 Total 59 153560.4330
Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of residuals(reduced model) No. of permutations used = 4999
(b)
--- Results ---Pair-wise a posteriori comparisons
Tests among groups in factor 2 (saison)
Possible Groups t P No.perm. ------------------------------------------------- ( P, E) 1.8877 0.0040 >1.0E+10 ( P, H) 1.5698 0.0128 >1.0E+10 ( E, H) 0.9123 0.5402 >1.0E+10 ------------------------------------------------- No. of permutations used = 4999
Average dissimilarities within/between groups
P E H P 64.448 E 71.701 68.143
320

H 67.789 65.447 63.822
Tests among groups in factor 1 (Groupe) within level 1 of factor 2 (Printemps)
Possible Groups t P No.perm. ------------------------------------------------ ( G2, G3) 1.7499 0.0044 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Tests among groups in factor 1 (Groupe) within level 2 of factor 2 (Eté)
Possible Groups t P No.perm. ------------------------------------------------ ( G2, G3) 0.9947 0.4050 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Tests among groups in factor 1 (Groupe) within level 3 of factor 2 (Hiver)
Possible Groups t P No.perm. ------------------------------------------------ ( G2, G3) 1.3116 0.0994 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Tests among groups in factor 2 (saison) within level 1 of factor 1 (G2)
Possible Groups t P No.perm. ------------------------------------------------ ( P, E) 2.1211 0.0054 >1.0E+10 ( P, H) 1.9690 0.0056 >1.0E+10 ( E, H) 1.2675 0.1590 >1.0E+10------------------------------------------------- No. of permutations used = 4999
Average dissimilarities within/between groups
G2 G3 G2 48.775 G3 68.617 70.855
Average dissimilarities within/between groups
G2 G3 G2 69.746 G3 68.635 65.447
Average dissimilarities within/between groups
G2 G3 G2 50.296 G3 65.809 72.932
Average dissimilarities within/between groups
P E H P 48.775
E 73.085 69.746H 59.464 61.893 50.296
321

Tests among groups in factor 2 (saison) within level 2 of factor 1 (G3)
Possible Groups t P No.perm. ------------------------------------------------ ( P, E) 0.9689 0.4762 0.9238E+05 ( P, H) 0.8821 0.6710 0.9238E+05 ( E, H) 0.8649 0.6004 0.9238E+05------------------------------------------------- No. of permutations used = 4999
Note: The pair-wise tests have not been corrected for multiple comparisons.
Average dissimilarities within/between groups
P E H P 70.855
E 67.431 65.447H 71.260 67.720 72.932
322

Annexe 8 - Modifications de l'alimentation printanière en fonction de la taille des individus chez Solea solea aularge du Rhône.(a) MANOVA non paramétrique à un facteur (stade de vie benthique) réalisée sur les contenus stomacaux (Poids des proies
ingérées en mg) des individus de S. solea présents au printemps.
(b) Résultats des tests de comparaisons a posteriori (G1 = LT < 100 mm, G2 = 100-200 mm, G3 = LT > 200 mm).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- One-way ANOVA Factor 1 is groupe with 3 levels The no. of replicates = 10 The no. of variables = 13
--- Results --- Non-parametric Multivariate Analysis of Variance
Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- group 2 15059.3986 7529.6993 3.7525 0.0004 >1.0E+10 Res Resid 27 54177.6031 2006.5779 Total 29 69237.0017 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of raw data No. of permutations used = 4999
(b)
--- Results ---Pair-wise a posteriori comparisons
Tests among groups in factor 1 (groupe)
Possible Groups t P No.perm. ------------------------------------------------ ( G1, G2) 2.4015 0.0010 0.9238E+05 ( G1, G3) 1.7125 0.0072 0.9238E+05 ( G2, G3) 1.7797 0.0036 0.9238E+05 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G1 G2 G3G1 52.029G2 65.456 48.775G3 70.155 69.445 71.769
Note: The pair-wise tests have not been corrected for multiple comparisons.
323

Annexe 9 - Modifications bathymétriques de l'alimentation des juvéniles de Solea solea de 100-200 mm de LTau large du Rhône. MANOVA non paramétrique à un facteur (profondeur : P1 = 0-20 m, P2 = 30-50 m) réalisée sur les contenusstomacaux (Poids des proies ingérées en mg) des individus de S. solea de 100-200 mm de LT.
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- One-way ANOVA Factor 1 is prof. with 2 levels (P1, P2) The no. of replicates = 13 The no. of variables = 13
--- Results --- Non-parametric Multivariate Analysis of Variance
Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- prof. 1 6121.5431 6121.5431 3.1006 0.0204 0.520E+07 Res Resid. 24 47382.9904 1974.2913 Total 25 53504.5335 -----------------------------------------------------------------------------
Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of raw data No. of permutations used = 4999
324

Annexe 10 - Modifications bathymétriques de l'alimentation des juvéniles et adultes de Solea solea de LT > 200mm au large du Rhône.(a) MANOVA non paramétrique à un facteur (profondeur) réalisée sur les contenus stomacaux (Poids des proies ingérées
en mg) des individus de S. solea de LT > 200 mm.
(b) Résultats des tests de comparaisons a posteriori (P1 = 0-20 m, P2 = 30-50 m, P3 = 70-100 m).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- One-way ANOVA Factor 1 is prof with 3 levels The no. of replicates = 13 The no. of variables = 13
--- Results --- Non-parametric Multivariate Analysis of Variance
Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- prof 2 9993.9869 4996.9934 2.1037 0.0188 >1.0E+10 Res Resid 36 85512.6378 2375.3510 Total 38 95506.6247 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of raw data No. of permutations used = 4999
(b)
--- Results ---Pair-wise a posteriori comparisons
Tests among groups in factor 1 (profondeur)
Possible Groups t P No.perm. ------------------------------------------------ ( P1, P2) 1.8191 0.0076 0.5200E+07 ( P1, P3) 1.5113 0.0364 0.5200E+07 ( P2, P3) 0.8206 0.6938 0.5200E+07 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P1 P2 P3P1 68.239P2 68.955 57.019P3 71.210 59.401 64.060
Note: The pair-wise tests have not been corrected for multiple comparisons.
325

Annexe 11 - Modifications de l'alimentation à 0-20 m en fonction de la taille des individus chez Solea solea aularge du Rhône .(a) MANOVA non paramétrique à un facteur (stade de vie benthique) réalisée sur les contenus stomacaux (Poids des proies
ingérées en mg) des individus de S. solea présents à 0-20 m.
(b) Résultats des tests de comparaisons a posteriori (G1 = LT < 100 mm, G2 = 100-200 mm, G3 = LT > 200 mm).
(a)Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
Input file:SsG1G2G3prof1.txt
--- Experimental Design --- One-way ANOVA Factor 1 is groupe with 3 levels The no. of replicates = 13 The no. of variables = 13
--- Results --- Non-parametric Multivariate Analysis of Variance
Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- groupe 2 16409.9560 8204.9780 3.7380 0.0004 >1.0E+10 Res Resid. 36 79021.3385 2195.0372 Total 38 95431.2944 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of raw data No. of permutations used = 4999
(b)
--- Results --- Pair-wise a posteriori comparisons
Tests among groups in factor 1 (groupe)
Possible Groups t P No.perm. ------------------------------------------------ ( G1, G2) 2.6205 0.0002 0.5200E+07 ( G1, G3) 2.0428 0.0016 0.5200E+07 ( G2, G3) 0.8866 0.5652 0.5200E+07 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G1 G2 G3 G1 53.860 G2 72.497 61.253 G3 71.400 64.203 68.929
Note: The pair-wise tests have not been corrected for multiple comparisons.
326

Annexe 12 - Modifications de l'alimentation à 30-50 m en fonction de la taille des individus chez Solea solea aularge du Rhône. MANOVA non paramétrique à un facteur (stade de vie benthique) réalisée sur les contenus stomacaux (Poids desproies ingérées en mg) des individus de S. solea présents à 30-50 m (G2 = 100-200 mm, G3 = LT > 200 mm).
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- One-way ANOVA Factor 1 is groupe with 2 levels The no. of replicates = 13 The no. of variables = 13
--- Results --- Non-parametric Multivariate Analysis of Variance
Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- groupe 1 5748.8690 5748.8690 3.0990 0.0160 0.520E+07 Res Resid. 24 44522.1170 1855.0882 Total 25 50270.9860 -----------------------------------------------------------------------------
Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of raw data No. of permutations used = 4999
327

Annexe 13 - Familles de polychètes ingérées par Solea solea au large du Rhône à chaque profondeurétudiée.
0 - 20 m 30 - 50 m 70 - 100 m
Carnivores
Polynoidae 9 6 0 Phyllodocidae 5 5 3 Nereidae 4 4 0 Nephtydae 8 5 0 Glyceridae 1 9 3 Eunicidae 2 2 0 Onuphidae 27 0 0 Lumbrineridae 1 19 5
Total 57 50 11
Dépositivores de surface
Paraonidae 21 5 0 Spionidae 4 38 8 Cossuridae 1 2 0 Cirratulidae 1 3 0 Magelonidae 244 1 0
Total 271 49 8
Dépositivores de sub-surface
Capitellidae 30 1432 0 Maldanidae 0 0 0 Sternaspidae 27 12 6
Total 57 1444 6
Suspensivores
Ampharetidae 1 11 1 Terebellidae 7 1 0 Sabellidae 2 0 0 Owenidae 0 0 0
Total 10 12 1
328

Annexe 14 – Modifications du nombre de proies par estomac en fonction de la taille des individus chez Soleaimpar au large du Rhône.
(a) ANOVA réalisée sur les nombres de proies par estomac des 4 groupes de taille individualisés par la NMS chez S.impar.
(b) Résultats des tests de comparaisons a posteriori (G1 = LT < 100 mm, G2 = 100-140m, G3 = 140-160m, G4 = LT >160 mm).
(a)
STATISTICA. MANOVA GENERALESynthèse de tous les EffetsPlan: 1-GROUPE
dl Mc dl Mc
Effet Effet Effet Erreur Erreur F niveau P
1 3 ,802021 97 ,190044 4,220193 ,007872
(b)
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocEFFET PRINC.:GROUPE
GROUPE G1 G2 G3 G4
G1 ,050548 ,016195 ,945520G2 ,050548 ,402846 ,106125G3 ,016195 ,402846 ,024602G4 ,945520 ,106125 ,024602
329

Annexe 15 - Modifications saisonnières de l'intensité de la nutrition chez Solea impar au large du Rhône.Variations de l'indice de réplétion stomacal (IR) en fonction de la saison.
ANOVA non paramétrique de Kruskal-Wallis :
Groupe Somme des rangs effectifPrintemps 3985,000 71
Eté 864,000 17Hiver 302,000 13
H (2, N = 101) = 8,609129 P = 0,0135
Test de Dunn (test non paramétrique de comparaison des échantillons a posteriori) :
Effectif total = 101 GroupesPrintemps Eté Hiver
Somme des rangs 3985 864 302effectif 71 17 13
Rang moyen 56,127 50,824 23,231
Matrice des SE : Hiver Printemps
Printemps 8,84Eté 10,80 7,91
Matrice des Q : Hiver Printemps
Printemps 3,72Eté 2,56 0,67
Q théorique (0,05; k=3) : 2,394Q théorique (0,01; k=3) : 2,936 Rejet H0 si Q > Q théoriqueQ théorique (0,001; k=3) : 3,588
Matrice des résultats : Hiver PrintempsPrintemps * * *
Eté * ns
330

Annexe 16 - Modifications saisonnières de l'alimentation des juvéniles et adultes de Solea impar de plus de 100mm de LT au large du Rhône .
(a) MANOVA non paramétrique à deux facteurs (stade de vie benthique × saison) réalisée sur les contenusstomacaux (Poids des proies ingérées en mg) des individus de S. solea présents aux deux saisons d'alimentationde l'espèce (printemps, été).
(b) Résultats des tests de comparaisons a posteriori par saison (E = été, P = printemps) et par groupe (G2 = 100-160mm, G3 = LT > 160 mm).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- Two-way crossed (orthogonal) ANOVA Factor 1 is groupe with 2 levels Factor 2 is saison with 2 levels Factors 1 and 2 are fixed The no. of replicates = 10 The no. of variables = 9
--- Results --- Non-parametric Multivariate Analysis of Variance
Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- group 1 11861.8957 11861.8957 8.2213 0.0002 >1.0E+10 Res saiso 1 3555.7284 3555.7284 2.4644 0.0488 >1.0E+10 Res grxsa 1 2955.0926 2955.0926 2.0481 0.0974 >1.0E+10 Res Resid 36 51941.9891 1442.8330 Total 39 70314.7058 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of residuals(reduced model) No. of permutations used = 4999
(b)
--- Results ---Pair-wise a posteriori comparisons
Tests among groups in factor 1 (Groupe) within level 1 of factor 2 (Eté)
Possible Groups t P No.perm. ------------------------------------------------ ( G2, G3) 2.5710 0.0016 0.9238E+05 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G2 G3 G2 21.705 G3 57.016 56.641
331

Tests among groups in factor 1 (Groupe) within level 2 of factor 2 (Printemps)
Possible Groups t P No.perm. ------------------------------------------------ ( G2, G3) 2.0470 0.0070 0.9238E+05 ------------------------------------------------ No. of permutations used = 4999
Tests among groups in factor 2 (saison) within level 1 of factor 1 (G2)
Possible Groups t P No.perm. ------------------------------------------------ ( E, P) 2.0886 0.0034 0.9238E+05 ------------------------------------------------ No. of permutations used = 4999
Tests among groups in factor 2 (saison) within level 2 of factor 1 (G3)
Possible Groups t P No.perm. ------------------------------------------------ ( E, P) 0.9648 0.4392 0.9238E+05 ------------------------------------------------ No. of permutations used = 4999
Note: The pair-wise tests have not been corrected for multiple comparisons.
Average dissimilarities within/between groups
G2 G3 G2 58.849 G3 63.751 48.973
Average dissimilarities within/between groups
E P E 21.705 P 53.147 58.849
Average dissimilarities within/between groups
E P E 56.641 P 52.115 48.973
332

Annexe 17 - Modifications de l'alimentation printanière en fonction de la taille des individus chez Solea imparau large du Rhône .
(a) MANOVA non paramétrique à un facteur (stade de vie benthique) réalisée sur les contenus stomacaux (Poids desproies ingérées en mg) des individus de S. impar présents au printemps.
(b) Résultats des tests de comparaisons a posteriori (G1 = LT < 100 mm, G2 = 100-160 mm, G3 = LT > 160 mm).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- One-way ANOVA Factor 1 is groupe with 3 levels The no. of replicates = 10 The no. of variables = 9
--- Results --- Non-parametric Multivariate Analysis of Variance Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- group 2 26552.7254 13276.3627 9.8374 0.0002 >1.0E+10 Res Resid 27 36438.7282 1349.5825 Total 29 62991.4536 -----------------------------------------------------------------------------
Data were transformed to log10(x+1)No standardizationAnalysis based on Bray-Curtis dissimilaritiesPermutation of raw dataNo. of permutations used = 4999
(b)
--- Results --- Pair-wise a posteriori comparisons
Tests among groups in factor 1 (groupe)
Possible Groups t P No.perm. ------------------------------------------------ ( G1, G2) 3.1145 0.0008 0.9238E+05 ( G1, G3) 4.3266 0.0002 0.9238E+05 ( G2, G3) 1.9738 0.0074 0.9238E+05 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G1 G2 G3 G1 34.614 G2 64.293 55.069 G3 76.986 61.385 48.973
Note: The pair-wise tests have not been corrected for multiple comparisons.
333

Ann
exe
18 -
Com
posi
tion
du r
égim
e al
imen
taire
des
indi
vidu
s de
s 4
grou
pes
de ta
ille
(A, B
, C, C
') in
divi
dual
isés
pou
r Bu
glos
sidi
um lu
teum
au
larg
e du
Rhô
ne p
ar o
rdin
atio
n et
cla
ssifi
catio
n de
s cl
asse
s de
taill
e (L
T, en
mm
) de
l'esp
èce
à pa
rtir d
e l'a
limen
tatio
n (p
oids
des
pro
ies
ingé
rées
) de
leur
s in
divi
dus.
F =
fréq
uenc
e de
s pr
oies
, Cn
= po
urce
ntag
e nu
mér
ique
, C
p =
pour
cent
age
pond
éral
, Q =
coe
ffic
ient
alim
enta
ire. "
+" =
val
eur
infé
rieur
e à
0,01
.
A :
LT <
50
mm
B :
50-7
0 m
mC
: 70
-110
mm
C' :
LT >
110
mm
FC
pC
nQ
FC
pC
nQ
FC
pC
nQ
FC
pC
nQ
Pol
ychè
tes
0,28
21,9
%5,
1%11
1,5
0,53
21,4
%14
,2%
303,
00,
6845
,2%
30,3
%13
70,1
0,72
56,6
%61
,7%
3495
,6
Cop
épod
es0,
6716
,4%
70,4
%11
57,6
0,41
12,8
%39
,6%
508,
1+
1,7%
2,9%
5,0
+1,
2%0,
5%0,
6
Lep
tost
racé
s--
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
-+
1,3%
0,5%
0,7
Mys
idac
és--
---
----
---
---
----
-0,
065,
1%0,
9%4,
90,
036,
8%3,
2%21
,80,
044,
2%1,
5%6,
4
Cum
acés
0,44
17,4
%13
,3%
231,
20,
3514
,5%
10,4
%15
0,6
0,04
6,4%
14,5
%92
,9--
---
----
---
---
----
-
Iso
pode
s--
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
-0,
040,
7%0,
5%0,
3
Am
phip
odes
0,44
17,4
%7,
1%12
4,5
0,65
12,0
%21
,7%
259,
40,
2014
,6%
41,3
%60
4,0
0,20
8,3%
25,0
%20
7,5
Cre
vette
s--
---
----
---
---
----
---
---
----
---
---
----
-+
7,1%
0,6%
4,6
0,08
9,8%
1,5%
14,9
Bra
chyo
ures
----
---
---
----
---
---
----
---
---
----
---
---
+2,
2%0,
6%1,
40,
042,
4%1,
0%2,
5
Biv
alve
s0,
1710
,4%
2,0%
21,2
0,53
17,8
%11
,3%
202,
00,
079,
5%5,
8%55
,00,
1514
,8%
7,2%
105,
8
Gas
téro
pode
s0,
1116
,4%
2,0%
33,6
0,06
11,0
%0,
9%10
,4--
---
----
---
---
----
-0,
040,
8%0,
5%0,
4
Oph
iure
s--
---
----
---
---
----
-0,
065,
3%0,
9%5,
0+
0,8%
0,3%
0,2
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
----
---
---
Poi
sson
s--
---
----
---
---
----
---
---
----
---
---
----
-0,
015,
8%0,
3%1,
9--
---
----
---
---
----
-
334

Annexe 19 - Modifications saisonnières de l'intensité de la nutrition chez Buglossidium luteum au large duRhône. Variations de l'indice de réplétion stomacal (IR) en fonction de la saison.
ANOVA non paramétrique de Kruskal-Wallis :
Groupe Somme des rangs effectifPrintemps 4224,500 61
Eté 2258,500 36Hiver 1020,000 25
H (2, N = 122) = 12,31268 P = 0,0021
Test de Dunn (test non paramétrique de comparaison des échantillons a posteriori) :
Effectif total = 122 GroupesPrintemps Eté Hiver
Somme des rangs 4224,5 2258,5 1020effectif 61 36 25
Rang moyen 69,254 62,736 40,800
Matrice des SE : Hiver Printemps
Printemps 8,40Eté 9,21 7,43
Matrice des Q : Hiver Printemps
Printemps 3,40Eté 2,38 0,88
Q théorique (0,05; k=3) : 2,394Q théorique (0,01; k=3) : 2,936 Rejet H0 si Q > Q théoriqueQ théorique (0,001; k=3) : 3,588
Matrice des résultats : Hiver PrintempsPrintemps * * *
Eté * ns
335

Annexe 20 - Modifications saisonnières de l'alimentation des juvéniles et adultes de Buglossidium luteum deplus de 50 mm de LT au large du Rhône.
(a) MANOVA non paramétrique à deux facteurs (stade de vie benthique × saison) réalisée sur les contenusstomacaux (Poids des proies ingérées en mg) des individus de B. luteum présents à au moins deux saison(printemps, été).
(b) Résultats des tests de comparaisons a posteriori par saison (P = printemps, E = été) et par groupe (G2 = 50-70mm, G3 = LT > 70 mm).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- Two-way crossed (orthogonal) ANOVA Factor 1 is groupe with 2 levels Factor 2 is saison with 2 levels Factors 1 and 2 are fixed The no. of replicates = 12 The no. of variables = 13
--- Results --- Non-parametric Multivariate Analysis of Variance
Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- group 1 8674.1922 8674.1922 4.8420 0.0054 >1.0E+10 Res saiso 1 4869.4875 4869.4875 2.7182 0.0514 >1.0E+10 Res grxsa 1 2196.1771 2196.1771 1.2259 0.3076 >1.0E+10 Res Resid 44 78822.9483 1791.4306 Total 47 94562.8051 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of residuals(reduced model) No. of permutations used = 4999
336

(b) --- Results --- Pair-wise a posteriori comparisons
Tests among groups in factor 1 (groupe) within level 1 of factor 2 (Printemps)
Possible Groups t P No.perm. ------------------------------------------------ ( G2, G3) 1.1817 0.2584 0.1352E+07 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G2 G3 G2 43.266 G3 47.829 48.988
Tests among groups in factor 1 (groupe) within level 2 of factor 2 (Eté)
Possible Groups t P No.perm. ------------------------------------------------ ( G2, G3) 2.0297 0.0080 0.1352E+07 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G2 G3 G2 65.230 G3 70.315 55.527
Tests among groups in factor 2 (saison) within level 1 of factor 1 (G2)
Possible Groups t P No.perm. ------------------------------------------------ ( P, E) 1.5028 0.1170 0.1352E+07 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P E P 43.266 E 58.311 65.230
Tests among groups in factor 2 (saison) within level 2 of factor 1 (G3)
Possible Groups t P No.perm. ------------------------------------------------ ( P, E) 1.2922 0.2044 0.1352E+07 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P E P 48.988 E 56.358 55.527
Note: The pair-wise tests have not been corrected for multiple comparisons
337

Annexe 21 - Modifications saisonnières de l'alimentation des adultes de Buglossidium luteum (LT > 70 mm) aularge du Rhône.
(a) MANOVA non paramétrique à un facteur (saison) réalisée sur les contenus stomacaux (Poids des proies ingéréesen mg) des individus de B. luteum de LT > 70 mm.
(b) Résultats des tests de comparaisons a posteriori (H = hiver, P = printemps, E = été).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- One-way ANOVA Factor 1 is saison with 3 levels The no. of replicates = 12 The no. of variables = 13
--- Results --- Non-parametric Multivariate Analysis of Variance Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- saison 2 4410.3147 2205.1574 1.1563 0.3430 >1.0E+10 Res Resid. 33 62934.4286 1907.1039 Total 35 67344.7433 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of raw data No. of permutations used = 4999
(b)
--- Results --- Pair-wise a posteriori comparisons
Tests among groups in factor 1 (saison)
Possible Groups t P No.perm. ------------------------------------------------ ( P, E) 0.6394 0.7164 0.1352E+07 ( P, H) 1.0869 0.3276 0.1352E+07 ( E, H) 1.3691 0.1392 0.1352E+07 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P E H P 49.385 E 56.602 63.750 H 52.667 61.574 53.585
Note: The pair-wise tests have not been corrected for multiple comparisons.
338

Annexe 22 - Modifications de l'alimentation estivale en fonction de la taille des individus chez Buglossidiumluteum au large du Rhône .
(a) MANOVA non paramétrique à un facteur (stade de vie benthique) réalisée sur les contenus stomacaux (Poids desproies ingérées en mg) des individus de B. luteum présents en été.
(b) Résultats des tests de comparaisons a posteriori (G1 = LT < 50 mm, G2 = 50-70 mm, G3 = LT > 70 mm).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- One-way ANOVA Factor 1 is groupe with 3 levels The no. of replicates = 12 The no. of variables = 13
--- Results --- Non-parametric Multivariate Analysis of Variance Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- groupe 2 18798.1877 9399.0939 4.0920 0.0006 >1.0E+10 Res Resid. 33 75799.9438 2296.9680 Total 35 94598.1315 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of raw data No. of permutations used = 4999
(b)
--- Results --- Pair-wise a posteriori comparisons
Tests among groups in factor 1 (groupe)
Possible Groups t P No.perm. ------------------------------------------------ ( G1, G2) 1.6993 0.0358 0.1352E+07 ( G1, G3) 2.6028 0.0002 0.1352E+07 ( G2, G3) 1.6481 0.0462 0.1352E+07 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G1 G2 G3 G1 65.728 G2 71.300 65.097 G3 78.120 66.479 56.500
Note: The pair-wise tests have not been corrected for multiple comparisons.
339

Annexe 23 - Modifications bathymétriques de l'alimentation des juvéniles et adultes de Buglossidium luteum deLT > 70 mm au large du Rhône.
(a) MANOVA non paramétrique à un facteur (profondeur) réalisée sur les contenus stomacaux (Poids des proiesingérées en mg) des individus de B. luteum de LT > 70 mm.
(b) Résultats des tests de comparaisons a posteriori (P1 = 0-20 m, P2 = 30-50 m, P3 = 70-100 m).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- One-way ANOVA Factor 1 is profondeur with 3 levels The no. of replicates = 28 The no. of variables = 13
--- Results --- Non-parametric Multivariate Analysis of Variance
Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- prof. 2 9844.1215 4922.0608 2.4758 0.0252 >1.0E+10 Res Resid. 81 161034.8945 1988.0851 Total 83 170879.0160 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of raw data No. of permutations used = 4999
(b)
--- Results --- Pair-wise a posteriori comparisons
Tests among groups in factor 1 (profondeur)
Possible Groups t P No.perm. ------------------------------------------------ ( P1, P2) 1.3702 0.1330 >1.0E+10 ( P1, P3) 1.6241 0.0460 >1.0E+10 ( P2, P3) 1.7244 0.0304 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P1 P2 P3 P1 60.802 P2 60.198 55.994 P3 58.844 55.729 51.246
Note: The pair-wise tests have not been corrected for multiple comparisons.
340

Annexe 24 - Modifications de l'alimentation à 0-20 m en fonction de la taille des individus chez Buglossidiumluteum au large du Rhône.
(a) MANOVA non paramétrique à un facteur (stade de vie benthique) réalisée sur les contenus stomacaux (Poids desproies ingérées en mg) des individus de B. luteum présents à 0-20 m.
(b) Résultats des tests de comparaisons a posteriori (G1 = LT < 50 mm, G2 = 50-70 mm, G3 = LT > 70 mm).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
Input file:BlG1G2G3P1.txt
--- Experimental Design --- One-way ANOVA Factor 1 is groupe with 3 levels The no. of replicates = 17 The no. of variables = 13
--- Results --- Non-parametric Multivariate Analysis of Variance
Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- groupe 2 18209.3178 9104.6589 4.2519 0.0004 >1.0E+10 Res Resid. 48 102782.9072 2141.3106 Total 50 120992.2250 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of raw data No. of permutations used = 4999
(b)
--- Results --- Pair-wise a posteriori comparisons
Tests among groups in factor 1 (groupe) Possible Groups t P No.perm. ------------------------------------------------ ( G1, G2) 1.6859 0.0368 0.1167E+10 ( G1, G3) 2.5937 0.0002 0.1167E+10 ( G2, G3) 1.7590 0.0382 0.1167E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G1 G2 G3 G1 69.471 G2 68.660 58.223 G3 72.861 58.828 52.270
Note: The pair-wise tests have not been corrected for multiple comparisons.
341

Annexe 25 - Familles de polychètes ingérées par Buglossidium luteum au large du Rhône à chaque profondeur étudiée.
0 - 20 m 30 - 50 m 70 - 100 m
Carnivores
Phyllodocidae 0 4 0 Nephthydae 11 5 6 Glyceridae 2 3 1 Eunicidae 0 0 0 Lumbrineridae 18 19 8
Total 31 31 15
Dépositivores de surface
Spionidae 5 2 2
Total 5 2 2
Dépositivores de sub-surface
Capitellidae 2 148 0
Total 2 148 0
Suspensivores
Ampharetidae 6 6 10
Total 6 6 10
342

Annexe 26 - Modifications saisonnières de l'intensité de la nutrition chez Arnoglossus laterna au large duRhône. Variations de l'indice de réplétion stomacal (IR) en fonction de la saison.
ANOVA non paramétrique de Kruskal-Wallis :
Groupe Somme des rangs effectifPrintemps 85864,0 290
Eté 54598,5 135Hiver 112653,5 286
H (2, N = 711) = 44,24509 P = 0,0000
Test de Dunn (test non paramétrique de comparaison des échantillons a posteriori) :
Effectif total = 711 GroupesPrintemps Eté Hiver
Somme des rangs 85864 54598,5 112653,5effectif 290 135 286
Rang moyen 296,083 404,433 393,893
Matrice des SE : Hiver Printemps
Printemps 17,12Eté 21,45 21,40
Matrice des Q : Hiver PrintempsPrintemps 5,7
Eté 0,49 5,06
Q théorique (0,05; k=3) : 2,394Q théorique (0,01; k=3) : 2,936 Rejet H0 si Q > Q théoriqueQ théorique (0,001; k=3) : 3,588
Matrice des résultats : Hiver PrintempsPrintemps * * *
Eté ns * * *
343

Annexe 27 - Modifications spatio-temporelles de l'alimentation des adultes d'Arnoglossus laterna (LT > 60 mm)au large du Rhône.
(a) MANOVA non paramétrique à deux facteurs (profondeur × saison) réalisée sur les contenus stomacaux (Poidsdes proies ingérées en mg) des individus d'A. laterna de LT > 60 mm.
(b) Résultats des tests de comparaisons a posteriori pour chaque profondeur (P1 = 0-20 m, P2 = 30-50 m, P3 = 70-100 m) et chaque saison (P = printemps, E = été, H = hiver).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- Two-way crossed (orthogonal) ANOVA Factor 1 is profondeur with 3 levels Factor 2 is saison with 3 levels Factors 1 and 2 are fixed The no. of replicates = 10 The no. of variables = 13
--- Results --- Non-parametric Multivariate Analysis of Variance Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- prof. 2 55662.0091 27831.0045 13.9852 0.0002 >1.0E+10 Res sais. 2 11841.8590 5920.9295 2.9753 0.0044 >1.0E+10 Res pr x sa 4 24720.0860 6180.0215 3.1055 0.0004 >1.0E+10 Res Resid 81 161192.5036 1990.0309 Total 89 253416.4576 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of residuals(reduced model) No. of permutations used =
(b)
--- Results --- Pair-wise a posteriori comparisons
Tests among groups in factor 1 (profondeur)
Possible Groups t P No.perm. ------------------------------------------------ ( P1, P2) 2.6601 0.0002 >1.0E+10 ( P1, P3) 4.9991 0.0002 >1.0E+10 ( P2, P3) 2.7699 0.0002 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P1 P2 P3 P1 54.051 P2 71.010 73.468 P3 79.853 75.140 57.498
344

Tests among groups in factor 2 (saison)
Possible Groups t P No.perm. ------------------------------------------------ ( P, E) 1.8246 0.0152 >1.0E+10 ( P, H) 1.7892 0.0210 >1.0E+10 ( E, H) 1.8090 0.0168 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P E H P 64.219 E 72.013 72.505 H 72.343 73.046 72.064
Tests among groups in factor 1 (prof.) within level 1 of factor 2 (Printemps)
Possible Groups t P No.perm. ------------------------------------------------ ( P1, P2) 1.8973 0.0352 >1.0E+10 ( P1, P3) 2.7868 0.0006 >1.0E+10 ( P2, P3) 2.0778 0.0046 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P1 P2 P3 P1 48.962 P2 61.435 59.473 P3 77.919 73.854 60.771
Tests among groups in factor 1 (prof.) within level 2 of factor 2 (Eté)
Possible Groups t P No.perm. ------------------------------------------------ ( P1, P2) 2.4804 0.0008 0.9238E+05 ( P1, P3) 2.5826 0.0002 0.9238E+05 ( P2, P3) 1.6061 0.0772 0.9238E+05 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P1 P2 P3 P1 56.463 P2 80.249 68.807 P3 77.733 72.429 63.592
Tests among groups in factor 1 (prof.) within level 3 of factor 2 (hiver)
Possible Groups t P No.perm. ------------------------------------------------ ( P1, P2) 2.3007 0.0006 0.9238E+05 ( P1, P3) 5.3599 0.0002 0.9238E+05 ( P2, P3) 3.3291 0.0002 0.9238E+05 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P1 P2 P3 P1 54.325 P2 78.922 72.328 P3 87.815 80.886 19.698
345

Tests among groups in factor 2 (saison) within level 1 of factor 1 (P1)
Possible Groups t P No.perm. ------------------------------------------------ ( P, E) 1.1621 0.2700 >1.0E+10 ( P, H) 0.9621 0.4856 >1.0E+10 ( E, H) 1.1996 0.2420 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P E H P 48.962 E 54.096 56.463 H 52.769 56.369 54.325
Tests among groups in factor 2 (saison) within level 2 of factor 1 (P2)
Possible Groups t P No.perm. ------------------------------------------------ ( P, E) 1.8246 0.0152 0.9238E+05 ( P, E) 2.2328 0.0010 0.9238E+05 ( E, H) 1.7881 0.0237 0.9238E+05 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P E H P 59.473 E 73.107 68.807 H 81.089 79.116 72.328
Tests among groups in factor 2 (saison) within level 3 of factor 1 (P3)
Possible Groups t P No.perm. ------------------------------------------------ ( P, E) 3.2062 0.0002 0.9238E+05 ( P, H) 0.4050 0.8984 0.9238E+05 ( E, H) 2.2735 0.0130 0.9238E+05 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P E H P 60.771 E 69.744 19.698 H 60.643 54.902 63.592
Note: The pair-wise tests have not been corrected for multiple comparisons.
346

Annexe 28 - Modifications de l'alimentation suivant la saison et la taille des individus chez Arnoglossus laternaà 0-20 m au large du Rhône.
(a) MANOVA non paramétrique à deux facteurs (stade de vie benthique × saison) réalisée sur les contenusstomacaux (Poids des proies ingérées en mg) des individus d'A. laterna capturés à 0-20 m de profondeur.
(b) Résultats des tests de comparaisons a posteriori pour chaque stade (G1 = LT < 50 mm, G2 = LT > 60 mm) etchaque saison (P = printemps, E = été).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- Two-way crossed (orthogonal) ANOVA Factor 1 is groupe with 2 levels Factor 2 is saison with 2 levels Factors 1 and 2 are fixed The no. of replicates = 19 The no. of variables = 13
--- Results --- Non-parametric Multivariate Analysis of Variance Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- groupe 1 25030.2154 25030.2154 13.4180 0.0002 >1.0E+10 Res saison 1 16295.4371 16295.4371 8.7355 0.0002 >1.0E+10 Res gr x sa 1 11637.4786 11637.4786 6.2385 0.0004 >1.0E+10 Res Resid. 72 134310.2903 1865.4207 Total 75 187273.4214 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of residuals(reduced model) No. of permutations used = 4999
347

(b)
--- Results --- Pair-wise a posteriori comparisons
Tests among groups in factor 1 within level 1 of factor 2 (Printemps)
Possible Groups t P No.perm. ------------------------------------------------ ( G1, G2) 3.9896 0.0002 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G1 G2G1 51.764G2 75.820 57.026
Tests among groups in factor 1 (Groupe) within level 2 of factor 2 (Eté)
Possible Groups t P No.perm. ------------------------------------------------ ( G1, G2) 2.0691 0.0004 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G1 G2 G1 49.680 G2 63.332 65.087
Tests among groups in factor 2 (saison) within level 1 of factor 1 (G1)
Possible Groups t P No.perm. ------------------------------------------------ ( P, E) 4.0433 0.0002 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P E P 51.764 E 69.496 49.680
Tests among groups in factor 2 (saison) within level 2 of factor 1 (G2)
Possible Groups t P No.perm. ------------------------------------------------ ( P, E) 1.2179 0.2130 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P E P 57.026 E 63.376 65.087
Note: The pair-wise tests have not been corrected for multiple comparisons.
348
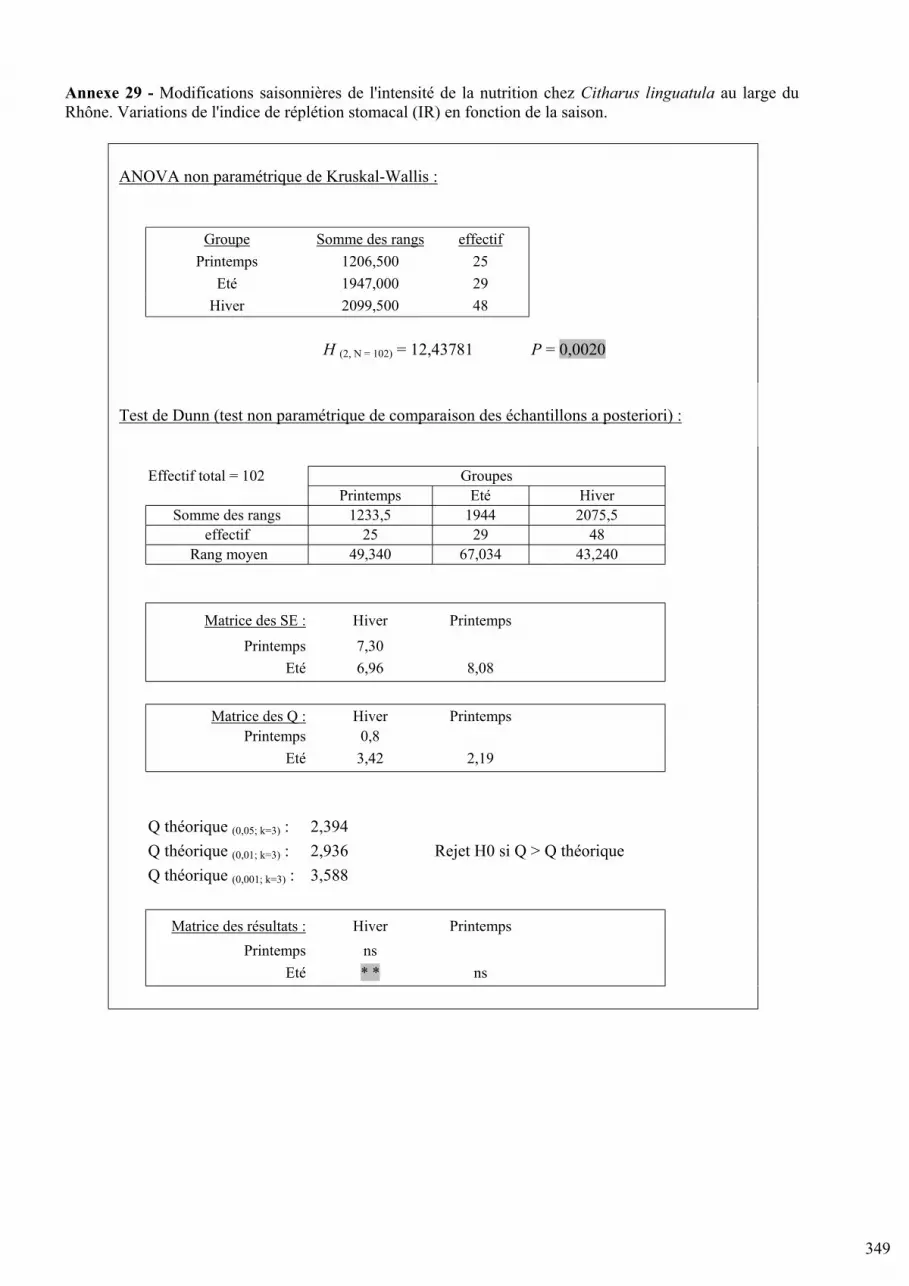
Annexe 29 - Modifications saisonnières de l'intensité de la nutrition chez Citharus linguatula au large duRhône. Variations de l'indice de réplétion stomacal (IR) en fonction de la saison.
ANOVA non paramétrique de Kruskal-Wallis :
Groupe Somme des rangs effectifPrintemps 1206,500 25
Eté 1947,000 29Hiver 2099,500 48
H (2, N = 102) = 12,43781 P = 0,0020
Test de Dunn (test non paramétrique de comparaison des échantillons a posteriori) :
Effectif total = 102 GroupesPrintemps Eté Hiver
Somme des rangs 1233,5 1944 2075,5effectif 25 29 48
Rang moyen 49,340 67,034 43,240
Matrice des SE : Hiver Printemps
Printemps 7,30Eté 6,96 8,08
Matrice des Q : Hiver PrintempsPrintemps 0,8
Eté 3,42 2,19
Q théorique (0,05; k=3) : 2,394Q théorique (0,01; k=3) : 2,936 Rejet H0 si Q > Q théoriqueQ théorique (0,001; k=3) : 3,588
Matrice des résultats : Hiver PrintempsPrintemps ns
Eté * * ns
349

Annexe 30 - Modifications de l'alimentation des individus de Citharus linguatula de plus de 100 mm de LTsuivant la saison et la taille au large du Rhône.
(a) MANOVA non paramétrique à deux facteurs (stade de vie benthique × saison) réalisée sur les contenusstomacaux (Poids des proies ingérées en mg) des individus de C. linguatula de plus de 100 mm de LT.
(b) Résultats des tests de comparaisons a posteriori pour chaque stade (G2 = 100-160 m, G2 = LT > 160 mm) etchaque saison (P = printemps, E = été, H = hiver).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- Two-way crossed (orthogonal) ANOVA Factor 1 is groupe with 2 levels Factor 2 is saison with 3 levels Factors 1 and 2 are fixed The no. of replicates = 11 The no. of variables = 5
--- Results --- Non-parametric Multivariate Analysis of Variance Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- groupe 1 13143.9806 13143.9806 8.6875 0.0006 >1.0E+10 Res saison 2 22501.7844 11250.8922 7.4363 0.0004 >1.0E+10 Res gr x sa 2 37416.8921 18708.4460 12.3653 0.0002 >1.0E+10 Res Resid. 60 90778.4595 1512.9743 Total 65 163841.1166 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of residuals(reduced model) No. of permutations used = 4999
350

(b)
--- Results --- Pair-wise a posteriori comparisons
Tests among groups in factor 2 (saison) Possible Groups t P No.perm. ------------------------------------------------ ( P, E) 1.9817 0.0260 >1.0E+10 ( P, H) 2.4793 0.0048 >1.0E+10 ( E, H) 2.1900 0.0226 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P E H P 59.100 E 59.833 49.851 H 69.107 61.585 58.004
Tests among groups in factor 1 (groupe) within level 1 of factor 2 (Printemps)
Possible Groups t P No.perm. ------------------------------------------------ ( G2, G3) 2.3240 0.0112 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G2 G3 G2 57.326 G3 65.553 46.680
Tests among groups in factor 1 (groupe) within level 2 of factor 2 (Eté)
Possible Groups t P No.perm. ------------------------------------------------ ( G2, G3) 2.4374 0.0154 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G2 G3 G2 56.739 G3 56.397 28.562
Tests among groups in factor 1 (groupe) within level 3 of factor 2 (Hiver)
Possible Groups t P No.perm. ------------------------------------------------ ( G2, G3) 5.2968 0.0002 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G2 G3 G2 35.498 G3 81.298 29.262
351

Tests among groups in factor 2 (saison) within level 1 of factor 1 (G2)
Possible Groups t P No.perm. ------------------------------------------------ ( P, E) 1.2510 0.2156 >1.0E+10 ( P, H) 1.1193 0.2992 >1.0E+10 ( E, H) 1.6672 0.0804 >1.0E+10 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P E H P 57.326 E 58.871 56.739 H 47.499 51.966 35.498
Tests among groups in factor 2 (saison) within level 2 of factor 1 (G3)
Possible Groups t P No.perm. ------------------------------------------------ ( P, E) 3.6192 0.0006 0.3527E+06 ( P, H) 4.1990 0.0002 0.3527E+06 ( E, H) 6.8804 0.0002 0.3527E+06 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P E H P 46.680 E 59.839 28.562 H 74.374 92.429 29.262
Note: The pair-wise tests have not been corrected for multiple comparisons.
352

Annexe 31 - Modifications de l'alimentation printanière en fonction de la taille des individus chez Citharuslinguatula au large du Rhône.
(a) MANOVA non paramétrique à un facteur (stade de vie benthique) réalisée sur les contenus stomacaux (Poids desproies ingérées en mg) des individus de C. linguatula présents au printemps.
(b) Résultats des tests de comparaisons a posteriori (G1 = LT < 100 mm, G2 = 100-160 mm, G3 = LT > 160 mm).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- One-way ANOVA Factor 1 is groupe with 3 levels The no. of replicates = 11 The no. of variables = 5
--- Results --- Non-parametric Multivariate Analysis of Variance Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- groupe 2 19830.7705 9915.3852 5.6031 0.0012 >1.0E+10 Res Resid. 30 53088.5422 1769.6181 Total 32 72919.3126 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of raw data No. of permutations used = 4999
(b)
--- Results --- Pair-wise a posteriori comparisons
Tests among groups in factor 1 (groupe)
Possible Groups t P No.perm. ------------------------------------------------ ( G1, G2) 2.1900 0.0260 0.3527E+06 ( G1, G3) 3.3619 0.0024 0.3527E+06 ( G2, G3) 2.2970 0.0124 0.3527E+06 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G1 G2 G3 G1 43.127 G2 62.386 57.485 G3 73.035 65.230 46.739
Note: The pair-wise tests have not been corrected for multiple comparisons.
353

Annexe 32 - Modifications bathymétriques de l'alimentation des juvéniles et adultes de Citharus linguatula deplus de 100 mm de LT au large du Rhône.
(a) MANOVA non paramétrique à deux facteurs (stade de vie benthique × profondeur) réalisée sur les contenusstomacaux (Poids des proies ingérées en mg) des individus de C. linguatula de plus de 100 mm de LT au large duRhône.
(b) Résultats des tests de comparaisons a posteriori par saison (P2 = 30-50 m, P3 = 70-100 m) et par groupe (G2 =100-160 mm, G3 = LT > 160 mm).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
Input file:C1G2G3Prof.txt
--- Experimental Design --- Two-way crossed (orthogonal) ANOVA Factor 1 is groupe with 2 levels Factor 2 is prof with 2 levels Factors 1 and 2 are fixed The no. of replicates = 13 The no. of variables = 5
--- Results --- Non-parametric Multivariate Analysis of Variance Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- groupe 1 12741.6399 12741.6399 5.2747 0.0094 >1.0E+10 Res prof. 1 5609.9373 5609.9373 2.3224 0.1046 >1.0E+10 Res Gr x pr 1 906.1689 906.1689 0.3751 0.6918 >1.0E+10 Res Resid 48 115949.1905 2415.6081 Total 51 135206.9366 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of residuals(reduced model) No. of permutations used = 4999
354

(b)
--- Results --- Pair-wise a posteriori comparisons
Tests among groups in factor 1 (groupe) within level 1 of factor 2 (P2)
Possible Groups t P No.perm. ------------------------------------------------ ( G2, G3) 1.9116 0.0376 0.5200E+07 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G2 G3G2 61.914G3 67.037 53.728
Tests among groups in factor 1 (groupe) within level 2 of factor 2 (P3)
Possible Groups t P No.perm. ------------------------------------------------ ( G2, G3) 1.3977 0.1524 0.5200E+07 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G2 G3 G2 52.793 G3 62.642 64.170
Tests among groups in factor 2 (profondeur) within level 1 of factor 1 (G1)
Possible Groups t P No.perm. ------------------------------------------------ ( P2, P3) 0.6255 0.6714 0.5200E+07 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P2 P3 P2 61.914 P3 56.085 52.793
Tests among groups in factor 2 (prof.) within level 2 of factor 1 (G2)
Possible Groups t P No.perm. ------------------------------------------------ ( P2, P3) 1.4893 0.1244 0.52000E+07 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
P2 P3 P2 53.728 P3 64.130 64.170
Note: The pair-wise tests have not been corrected for multiple comparisons.
355

Annexe 33 - Modifications de l'alimentation à 70-100 m en fonction de la taille des individus chez Citharuslinguatula au large du Rhône.
(a) MANOVA non paramétrique à un facteur (stade de vie benthique) réalisée sur les contenus stomacaux (Poids desproies ingérées en mg) des individus de C. linguatula présents à 70-100 m.
(b) Résultats des tests de comparaisons a posteriori (G1 = LT < 100 mm, G2 = 100-160 mm, G3 = LT > 160 mm).
(a)
Program NP-MANOVA by M.J. Anderson Centre for Research on Ecological Impacts of Coastal Cities University of Sydney (1999)
--- Experimental Design --- One-way ANOVA Factor 1 is groupe with 3 levels The no. of replicates = 10 The no. of variables = 5
--- Results --- Non-parametric Multivariate Analysis of Variance Possible Denom. Source df SS MS F P No.perm. MS ----------------------------------------------------------------------------- groupe 2 17451.4573 8725.7286 5.0461 0.0034 >1.0E+10 Res Resid. 27 46688.9015 1729.2186 Total 29 64140.3588 ----------------------------------------------------------------------------- Data were transformed to log10(x+1) No standardization Analysis based on Bray-Curtis dissimilarities Permutation of residuals(reduced model) No. of permutations used = 4999
(b)
--- Results --- Pair-wise a posteriori comparisons
Tests among groups in factor 1 (groupe)
Possible Groups t P No.perm. ------------------------------------------------ ( G1, G2) 1.7811 0.0416 0.9238E+05 ( G1, G3) 3.5026 0.0004 0.9238E+05 ( G2, G3) 1.3666 0.1706 0.9238E+05 ------------------------------------------------ No. of permutations used = 4999
Average dissimilarities within/between groups
G1 G2 G3G1 28.240G2 48.564 58.395G3 72.975 64.138 57.147
Note: The pair-wise tests have not been corrected for multiple comparisons.
356

Annexe 34 - Différences de signature isotopique en carbone (δ13C) entre les quatre principales sourcespotentielles de matière organique individualisées au large du Rhône et la matière organique particulaire (MOP)de l'eau de surface et le sédiment superficiel de la zone d'étude.
(a) ANOVA réalisée sur les δ13C de ces différentes catégories.(b) Résultats des tests de comparaisons a posteriori. R1 = zone d'étude, PhytopK = phytoplancton marin (Frioul),
Algue = débris de macrophyte marine (Chlorophycées), Vég. = débris de végétaux terrestres, Séd. = sédiment.
(a)
STATISTICA. MANOVA GENERALESynthèse de tous les EffetsPlan: 1- catégorie
dl Mc dl Mc
Effet Effet Effet Erreur Erreur F niveau P
1 5 23,03708 72 ,752792 30,60218 ,000000
(b)
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocEFFET PRINC.:catégorie
Echantillon MOP Rhône MOP eau R1 PhytopK Séd. R1 Vég. Rhône Algue
MOP Rhône ,005300 ,000125 ,007524 ,000607 ,000150
MOP eau R1 ,005300 ,000173 ,635635 ,000150 ,000216
PhytopK ,000125 ,000173 ,000168 ,000128 ,673345
Séd. R1 ,007524 ,635635 ,000168 ,000111 ,000161
Vég. Rhône ,000607 ,000150 ,000128 ,000111 ,000125
Algue ,000150 ,000216 ,673345 ,000161 ,000125
357

Annexe 35 - Différences de signature isotopique en carbone (δ15N) entre les quatre principales sourcespotentielles de matière organique individualisées au large du Rhône et la matière organique particulaire (MOP)de l'eau de surface et le sédiment superficiel de la zone d'étude.
(a) ANOVA réalisée sur les δ15N de ces différentes catégories.(b) Résultats des tests de comparaisons a posteriori. R1 = zone d'étude, PhytopK = phytoplancton marin (Frioul),
Algue = débris de macrophyte marine (Chlorophycées), Vég. = débris de végétaux terrestres, Séd. = sédiment.
(a)
STATISTICA. MANOVA GENERALESynthèse de tous les EffetsPlan: 1-ECHANTILLON
dl Mc dl Mc
Effet Effet Effet Erreur Erreur F niveau P
1 5 7,381503 72 ,399433 18,47997 ,000000
(b)
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocEFFET PRINC.:ECHANTILLON
Echantillon MOP Rhône MOP eau R1 PhytopK Séd. R1 Vég. Rhône Algue
MOP Rhône ,666472 ,004045 ,290630 ,000150 ,064439
MOP eau R1 ,666472 ,005203 ,298137 ,000111 ,040219
PhytopK ,004045 ,005203 ,000352 ,001304 ,000127
Séd. R1 ,290630 ,298137 ,000352 ,000125 ,226423
Vég. Rhône ,000150 ,000111 ,001304 ,000125 ,000128
Algue ,064439 ,040219 ,000127 ,226423 ,000128
358

Annexe 36 - Différences de signature isotopique en carbone (δ13C) entre les deux principales sources de matièreorganique individualisées au large du Rhône (phytoplancton marin et MOP rhodanienne) et la MOP de l'eau desurface et le sédiment superficiel de la zone d'étude (R1) en périodes de crues du fleuve et pendant l'étiage.
(a) ANOVA à deux facteurs (catégorie × débit) réalisée sur les δ13C de ces différentes catégories en périodes decrues du fleuve et pendant l'étiage.
(b) Résultats des tests de comparaisons a posteriori. MOP = matière organique particulaire, phytopK = phytoplanctonmarin (Frioul), R1 = zone d'étude, Sed. = sédiment, C = crue, E = étiage.
(a)
STATISTICA. MANOVA GENERALESynthèse de tous les EffetsPlan: 1- Catégorie 2- Débit
dl Mc dl Mc
Effet Effet Effet Erreur Erreur F niveau P
1 3 17,24967 63 ,544980 31,65191 ,000000
2 1 1,86519 63 ,544980 3,42249 ,069005
1 × 2 3 7,23329 63 ,544980 13,27257 ,000001
(b)
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocEFFET PRINC.: catégorie
Echantillon MOP Rhône MOP eau R1 Séd. R1 PhytopK
MOP Rhône ,000115 ,000311 ,000153
MOP eau R1 ,000115 ,028948 ,000108
Séd. R1 ,000311 ,028948 ,000115
PhytopK ,000153 ,000108 ,000115
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocEFFET PRINC.: Débit
Débit Crue Etiage
Crue ,037064
Etiage ,037064
359

(b) suite
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocINTERACTION : 1 × 2
Echantillon MOPRhône
MOPRhône
MOP eauR1
MOP eauR1 Séd. R1 Séd. R1 PhytopK PhytopK
× Débit × C × E × C × E × C × E × C × E
MOP Rhône × C ,001779 ,737419 ,000865 ,291823 ,847931 ,000130 ,000161
MOP Rhône × E ,001779 ,000972 ,000131 ,000160 ,002699 ,000127 ,000130
MOP eau R1 × C ,737419 ,000972 ,002784 ,304333 ,582365 ,000131 ,000359
MOP eau R1 × E ,000865 ,000131 ,002784 ,017942 ,001002 ,005312 ,343088
Séd. R1 × C ,291823 ,000160 ,304333 ,017942 ,258015 ,000155 ,003537
Séd. R1 × E ,847931 ,002699 ,582365 ,001002 ,258015 ,000131 ,000173
PhytopK × C ,000130 ,000127 ,000131 ,005312 ,000155 ,000131 ,025274
PhytopK × E ,000161 ,000130 ,000359 ,343088 ,003537 ,000173 ,025274
360

Annexe 37 - Différences de signature isotopique en carbone (δ13C) entre les deux principales sources de matièreorganique individualisées au large du Rhône (phytoplancton marin et MOP rhodanienne) et la MOP de l'eau desurface et le sédiment superficiel de chaque tranche bathymétrique échantillonnée (10 m = 0-20 m, 30 m = 30-50 m, 90 m = 70-100 m) pour la zone d'étude (R1).
(a) ANOVA à deux facteurs (catégorie × débit) réalisée sur les δ13C de ces différentes catégories en périodes decrues du fleuve et pendant l'étiage.
(b) Résultats des tests de comparaisons a posteriori. MOP = matière organique particulaire, phytopK =phytoplancton, R1 = zone d'étude, Séd. = sédiment.
(a)
STATISTICA. MANOVA GENERALESynthèse de tous les EffetsPlan: 1 - catégorie, 2 - débit
dl Mc dl Mc
Effet Effet Effet Erreur Erreur F niveau P
1 7 10,45644 55 ,156802 66,68555 ,000000
2 1 ,00039 55 ,156802 ,00248 ,960466
1 × 2 7 3,57401 55 ,156802 22,79310 ,000000
(b)
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocEFFET PRINC.: catégorie
Echantillon MOPRhône
MOP eauR1 10m
MOP eauR1 30m
MOP eauR1 90m
Séd. R110m
Séd. R130m
Séd. R190m PhytopK
Eau Rhône ,000138 ,001015 ,000136 ,000250 ,000198 ,000130 ,000133
MOP eau R1 10m ,000138 ,000130 ,000114 ,000143 ,000186 ,326642 ,000121
MOP eau R1 30m ,001015 ,000130 ,000138 ,465531 ,301273 ,000182 ,000136
MOP eau R1 90m ,000136 ,000114 ,000138 ,000161 ,000130 ,000121 ,003160
Séd. R1 10m ,000250 ,000143 ,465531 ,000161 ,886138 ,000371 ,000130
Séd. R1 30m ,000198 ,000186 ,301273 ,000130 ,886138 ,000577 ,000138
Séd. R1 90m ,000130 ,326642 ,000182 ,000121 ,000371 ,000577 ,000161
PhytopK ,000133 ,000121 ,000136 ,003160 ,000130 ,000138 ,000161
361

Annexe 38 - Différences de signature isotopique en carbone (δ13C) en fonction de l'échelon trophique(producteurs primaires, invertébrés benthiques et poissons) et de la profondeur au large du Rhône.
(a) ANOVA à deux facteurs (catégorie × profondeur) réalisée sur les δ13C obtenus pour les producteurs primaires(PI), les invertébrés benthiques et les poissons (5 espèces étudiées) à chaque profondeur (P1 = 0-20 m, P2 = 30-50m, P3 = 70-100 m).
(b) Résultats des tests de comparaisons a posteriori réalisés pour chaque facteur et pour les deux facteurs croisés.
(a)
STATISTICA. MANOVA GENERALESynthèse de tous les EffetsPlan: 1- catégorie, 2- profondeur
dl Mc dl Mc
Effet Effet Effet Erreur Erreur F niveau P
1 2 ,237065 248 ,001029 230,2971 ,000000
2 2 ,017089 248 ,001029 16,6012 ,000000
1 × 2 4 ,001729 248 ,001029 1,6800 ,155157
(b)
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocEFFET PRINC.:CATEGORIE
Catégorie PI Invertébrés Poissons
PI ,000009 ,000022
Invertébrés ,000009 ,000009
Poissons ,000022 ,000009
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocEFFET PRINC.: PROFONDEUR
Profondeur P1 P2 P3
P1 ,000010 ,133019
P2 ,000010 ,000022
P3 ,133019 ,000022
362

(b) suite
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocINTERACTION : 1 × 2
Catégorie PI PI PI Invert Invert Invert Poiss Poiss Poiss
× Profondeur × P1 × P2 × P3 × P1 × P2 × P3 × P1 × P2 × P3
PI × P1 ,614320 ,010579 ,000017 ,000022 ,000008 ,000032 ,000020 ,000026
PI × P2 ,614320 ,006260 ,000020 ,000008 ,000017 ,000010 ,000026 ,000032
PI × P3 ,010579 ,006260 ,000008 ,009791 ,000022 ,000026 ,000017 ,000020
Invert × P1 ,000017 ,000020 ,000008 ,000227 ,974752 ,000016 ,076484 ,000032
Invert × P2 ,000022 ,000008 ,009791 ,000227 ,000089 ,000020 ,000008 ,000017
Invert × P3 ,000008 ,000017 ,000022 ,974752 ,000089 ,000029 ,168515 ,000026
Poiss × P1 ,000032 ,000010 ,000026 ,000016 ,000020 ,000029 ,006606 ,845328
Poiss × P2 ,000026 ,000026 ,000017 ,076484 ,000008 ,168515 ,006606 ,004404
Poiss × P3 ,000026 ,000032 ,000020 ,000032 ,000017 ,000026 ,845328 ,004404
363

Annexe 39 - Différences de signature isotopique en carbone (δ15N) en fonction de l'échelon trophique(producteurs primaires, invertébrés benthiques et poissons) et de la profondeur au large du Rhône.
(a) ANOVA à deux facteurs (catégorie × profondeur) réalisée sur les δ15N obtenus pour les producteurs primaires(PI), les invertébrés benthiques et les poissons (5 espèces étudiées) à chaque profondeur (P1 = 0-20 m, P2 = 30-50m, P3 = 70-100 m).
(b) Résultats des tests de comparaisons a posteriori réalisés pour chaque facteur et pour les deux facteurs croisés.
(a)
STATISTICA. MANOVA GENERALESynthèse de tous les EffetsPlan: 1-CATEGORIE, 2-PROFONDEUR
dl Mc dl Mc
Effet Effet Effet Erreur Erreur F niveau P
1 2 3,233336 248 ,010280 314,5313 ,000000
2 2 ,007394 248 ,010280 ,7192 ,488143
1 × 2 4 ,001795 248 ,010280 ,1746 ,951284
(b)
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocEFFET PRINC.:CATEGORIE
Catégorie PI Invertébrés Poissons
PI ,000009 ,000021
Invertébrés ,000009 ,000009
Poissons ,000021 ,000009
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocINTERACTION : 1 × 2
Catégorie × PI PI PI Invert Invert Invert Poiss Poiss Poiss
Profondeur × P1 × P2 × P3 × P1 × P2 × P3 × P1 × P2 × P3
PI × P1 ,554232 ,792394 ,000017 ,000020 ,000008 ,000026 ,000032 ,000010
PI × P2 ,554232 ,953338 ,000008 ,000017 ,000022 ,000020 ,000026 ,000032
PI × P3 ,792394 ,953338 ,000022 ,000008 ,000009 ,000017 ,000020 ,000026
Invert × P1 ,000017 ,000008 ,000022 ,383692 ,886788 ,000022 ,000008 ,000017
Invert × P2 ,000020 ,000017 ,000008 ,383692 ,568294 ,000015 ,000023 ,000010
Invert × P3 ,000008 ,000022 ,000009 ,886788 ,568294 ,000008 ,000017 ,000020
Poiss × P1 ,000026 ,000020 ,000017 ,000022 ,000015 ,000008 ,596905 ,851009
Poiss × P2 ,000032 ,000026 ,000020 ,000008 ,000023 ,000017 ,596905 ,990227
Poiss × P3 ,000010 ,000032 ,000026 ,000017 ,000010 ,000020 ,851009 ,990227
364

Ann
exe
40 –
Sig
natu
res i
soto
piqu
es (δ
13C
et δ
15N
: m
oyen
ne ±
err
eur-
type
) obt
enue
s pou
r les
con
stitu
ant p
oten
tiels
des
rése
aux
troph
ique
s des
cin
q po
isso
ns p
lats
à 0
-20
m, 3
0-50
m e
t70
-100
m a
u la
rge
du R
hône
. "
" =
caté
gorie
pré
sent
e m
ais c
ompo
sitio
n is
otop
ique
non
ana
lysé
e po
ur c
ette
pro
fond
eur p
ar m
anqu
e de
mat
érie
l.
0-20
m30
-50
m70
-100
mG
loba
lδ13
Cδ15
Nδ13
Cδ15
Nδ13
Cδ15
Nδ13
Cδ15
NN
éch
antil
lons
Sour
ces (
P I)
MO
P R
hône
-26,
11±0
,23
3,47
±0,4
2N
= 9
phyt
opla
ncto
n m
arin
-22,
36±0
,24
2,33
±0,1
1N
= 9
Sédi
men
t sup
erfic
iel
-24,
86±0
,25
3,79
±0,1
8-2
5,00
±0,1
94,
09±0
,17
-24,
25±0
,18
3,56
±0,1
6-2
4,70
±0,1
43,
81±0
,11
N =
27
MO
P ea
u de
surf
ace
-24,
47±0
,51
3,05
±0,3
1-2
5,37
±0,2
22,
96±0
,51
-23,
31±0
,35
3,10
±0,4
3-2
4,38
±0,2
93,
03±0
,23
N =
26
Vég
étau
x te
rres
tres
-28,
40±0
,08
1,71
±0,0
1-2
7,19
±0,1
00,
81±0
,06
-27,
56±0
,05
0,96
±0,0
3-2
7,72
±0,2
31,
16±0
,18
N =
6M
acro
phyt
es m
arin
es (C
hlor
ophy
cées
)-2
2,56
±0,0
04,
25±0
,27
-22,
56±0
,04
4,25
±0,2
7N
= 3
Zoop
lanc
ton
-21,
29±0
,06
4,73±0
,01
N =
3
Proi
es b
enth
ique
s
Poly
chèt
es-2
2,67
±0,2
17,
18±0
,50
N =
24
susp
ensi
vore
s-1
9,21
±0,0
27,
24±0
,02
-23,
76±0
,09
6,43
±0,0
5-2
0,39
±0,0
36,
10±0
,05
-21,
12±0
,86
6,59
±0,2
2N
= 6
carn
ivor
es-2
1,94
±0,0
48,
55±0
,04
-22,
05±0
,02
9,12
±0,0
0-2
0,32
±0,0
69,
17±0
,01
-21,
43±0
,35
8,95
±0,1
3N
= 6
dépo
sitiv
ores
de
surf
ace
-24,
94±0
,00
5,84
±0,0
1-2
4,36
±0,0
26,
09±0
,00
-21,
82±0
,02
6,40
±0,0
2-2
3,71
±0,6
16,
11±0
,10
N =
6dé
posi
tivor
es d
e su
b-su
rfac
e-2
4,52
±0,2
96,
85±0
,10
-23,
26±0
,04
7,06
±0,0
3-2
3,65
±0,1
27,
01±0
,01
-23,
81±0
,25
6,97
±0,0
5N
= 6
Cop
épod
es
-2
1,49
±0,0
64,
79±0
,03
N =
3M
ysid
acés
-19,
53±0
,35
9,15
±0,5
0N
= 3
Cum
acés
-21,
43±0
,07
4,30
±0,1
5N
= 3
Am
phip
odes
-21,
45±0
,10
5,51
±0,1
0-2
0,59
±0,5
85,
20±0
,20
-20,
25±0
,12
4,75
±0,1
3-2
0,90
±0,2
45,
17±0
,16
N =
6
Mac
rour
es (C
reve
ttes
)-1
9,86
±0,1
87,
68±0
,90
N =
10
Alph
eus s
p.
-1
9,97
±0,0
78,
35±0
,00
-20,
22±0
,10
8,33
±0,0
0-2
0,07
±0,0
88,
34±0
,00
N =
4C
rang
on sp
.-1
8,44
±0,0
95,
43±0
,05
-20,
76±0
,06
8,27
±0,5
0-1
9,60
±0,6
76,
85±0
,85
-19,
60±0
,67
6,85
±0,8
5N
= 6
Ano
mou
res (
Pagu
res)
-17,
47±0
,05
7,25
±0,0
6
-1
7,47
±0,0
57,
25±0
,06
N =
3B
rach
your
es (C
rabe
s)-2
1,45
±0,0
96,
23±0
,05
-21,
89±0
,01
6,99
±0,0
1-2
0,94
±0,9
66,
18±0
,80
-21,
33±0
,36
6,37
±0,3
0N
= 6
Biv
alve
s-2
1,30
±0,5
26,
09±0
,40
N =
9ju
véni
les
-22,
21±0
,13
5,42
±0,0
0-2
2,85
±0,0
15,
29±0
,00
-20,
87±0
,01
5,76
±0,0
4-2
1,98
±0,3
75,
49±0
,09
N =
6Si
phon
s d'ad
ulte
s-1
9,27
±0,0
77,
88±0
,05
-19,
27±0
,07
7,8
8 ±0,
05 N
= 3
Gas
téro
pode
s-1
8,75
±0,2
19,
75±0
,02
-18,
29±0
,03
9,79
±0,0
3
-1
8,56
±0,1
69,
77±0
,02
N =
6
Oph
iure
s-1
9,02
±0,1
57,
85±0
,31
-19,
02±0
,15
7,85
±0,3
1N
= 3
Hol
othu
ries
-19,
02±0
,02
6,39
±0,2
7N
= 3
Pois
sons
(gob
iidés
)
-1
9,07
±0,2
68,
28±0
,75
N =
3
365

Annexe 41 - Différences de signature isotopique en carbone (δ13C) et en azote (δ15N) entre les cinq espècesétudiées (Solea solea, Solea impar, Buglossidium luteum, Arnoglossus laterna et Citharus linguatula).
(a) ANOVA à un facteur (espèce) et résultats des tests de comparaisons a posteriori réalisés sur les δ13C des poissonsétudiés.
(b) ANOVA à un facteur (espèce) et résultats des tests de comparaisons a posteriori réalisés sur les δ15N des poissonsétudiés.
(a)
STATISTICA. MANOVA GENERALE – DELTA C
Variabledépendante
MCEffet
MCErreur
F (dl 1,2)4,95 niveau P
DELTA C 17,92344 1,765998 10,14919 ,000001
STATISTICA. MANOVA GENERALE – DELTA CTest Newman-KeulsProbas des Tests Post HocEFFET PRINC.: ESPECE
Espèce S. solea B. luteum A. laterna S. impar C. linguatula
S. solea ,001852 ,000777 ,000142 ,000119
B. luteum ,001852 ,983806 ,064073 ,103572
A. laterna ,000777 ,983806 ,145858 ,166541
S. impar ,000142 ,064073 ,145858 ,851994
C. linguatula ,000119 ,103572 ,166541 ,851994
(b)
STATISTICA. MANOVA GENERALE – DELTA N
Variabledépendante
MCEffet
MCErreur
F (dl 1,2)4,95 niveau P
DELTA_N 5,96526 1,154977 5,16483 ,000828
STATISTICA. MANOVA GENERALE – DELTA NTest Newman-KeulsProbas des Tests Post HocEFFET PRINC.: ESPECE
Espèce S. solea B. luteum A. laterna S. impar C. linguatula
S. solea ,160883 ,006307 ,445799 ,230474
B. luteum ,160883 ,134861 ,050673 ,525459
A. laterna ,006307 ,134861 ,000863 ,086212
S. impar ,445799 ,050673 ,000863 ,124611
C. linguatula ,230474 ,525459 ,086212 ,124611
366

Annexe 42 - Modifications de la signature isotopique en carbone (δ13C) en fonction de la taille et de laprofondeur chez les individus de Solea solea au large du Rhône.
(a) ANOVA à un facteur (profondeur) réalisée sur les δ13C obtenus pour les adultes de S. solea capturés à chaqueprofondeur (P1 = 0-20 m, P2 = 30-50 m, P3 = 70-100 m).
(b) Résultats des tests de comparaisons a posteriori des δ13C obtenus pour les individus de S. solea de chaque stade(juvénile/adulte) pour chaque profondeur (P1 = 0-20 m, P2 = 30-50 m, P3 = 70-100 m).
(a)
STATISTICA. MANOVA GENERALESynthèse de tous les effetsPlan: 1- PROFONDEUR
dl Mc dl Mc
Effet Effet Effet Erreur Erreur F niveau P
Profondeur 2 9,953016 18 2,247572 4,428341 ,027289
(b)
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocINTERACTION: 1 × 2
STADE × Adulte Adulte Adulte Juvénile Juvénile Juvénile
Profondeur × P1 × P2 × P3 × P1 × P2 × P3
Adulte × P1 ,020447 ,369318 ,121455 -- --
Adulte × P2 ,020447 ,090096 ,296585 -- --
Adulte × P3 ,369318 ,090096 ,271583 -- --
Juvénile × P1 ,121455 ,296585 ,271583 -- --
Juvénile × P2 -- -- -- -- --
Juvénile × P3 -- -- -- -- --
367

Annexe 43 - Modifications de la signature isotopique en carbone (δ13C) en fonction de la taille et de laprofondeur chez les individus de Buglossidium luteum au large du Rhône.
(a) ANOVA à un facteur (profondeur) réalisée sur les δ13C obtenus pour les adultes de S. solea capturés à chaqueprofondeur (P1 = 0-20 m, P2 = 30-50 m, P3 = 70-100 m).
(b) Résultats des tests de comparaisons a posteriori des δ13C obtenus pour les individus de B. luteum de chaque stade(juvénile/adulte) pour chaque profondeur (P1 = 0-20 m, P2 = 30-50 m, P3 = 70-100 m).
(a)
STATISTICA. MANOVA GENERALESynthèse de tous les effetsPlan: 1-PROFONDEUR
dl Mc dl Mc
Effet Effet Effet Erreur Erreur F niveau P
1 2 15,03882 18 1,912686 7,862671 ,003514
(b)
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocINTERACTION: 1 × 2
STADE × Adulte Adulte Adulte Juvénile Juvénile Juvénile
Profondeur × P1 × P2 × P3 × P1 × P2 × P3
Adulte × P1 ,002692 ,488847 ,350599 -- --
Adulte × P2 ,002692 ,001265 ,000478 -- --
Adulte × P3 ,488847 ,001265 ,485712 -- --
Juvénile × P1 ,350599 ,000478 ,485712 -- --
Juvénile × P2 -- -- -- -- --
Juvénile × P3 -- -- -- -- --
368

Annexe 44 - Modifications de la signature isotopique en carbone (δ15N) en fonction de la taille et de laprofondeur chez les individus d'Arnoglossu laterna au large du Rhône.
(a) ANOVA à un facteur (profondeur) réalisée sur les δ15N obtenus pour les adultes d'A. laterna capturés à chaqueprofondeur (P1 = 0-20 m, P2 = 30-50 m, P3 = 70-100 m).
(b) Résultats des tests de comparaisons a posteriori des δ15N obtenus pour les individus d'A. laterna de chaque stade(juvénile/adulte) pour chaque profondeur (P1 = 0-20 m, P2 = 30-50 m, P3 = 70-100 m).
(a)
STATISTICA. MANOVA GENERALESynthèse de tous les effetsPlan: 1-PROFONDEUR
dl Mc dl Mc
Effet Effet Effet Erreur Erreur F niveau P
1 2 ,914483 13 ,201948 4,528310 ,032183
(b)
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocINTERACTION: 1 × 2
STADE × Adulte Adulte Adulte Juvénile Juvénile Juvénile
Profondeur × P1 × P2 × P3 × P1 × P2 × P3
Adulte × P1 ,428308 ,026853 ,000171 -- --
Adulte × P2 ,428308 ,059444 ,000137 -- --
Adulte × P3 ,026853 ,059444 ,000733 -- --
Juvénile × P1 ,000171 ,000137 ,000733 -- --
Juvénile × P2 -- -- -- -- --
Juvénile × P3 -- -- -- -- --
369

Annexe 45 - Modifications de la signature isotopique en carbone (δ13C) en fonction de la taille et de laprofondeur chez les individus d'Arnoglossus laterna au large du Rhône.
(a) ANOVA à un facteur (profondeur) réalisée sur les δ13C obtenus pour les adultes d'A. laterna capturés à chaqueprofondeur (P1 = 0-20 m, P2 = 30-50 m, P3 = 70-100 m).
(b) Résultats des tests de comparaisons a posteriori des δ13C obtenus pour les individus d'A. laterna de chaque stade(juvénile/adulte) pour chaque profondeur (P1 = 0-20 m, P2 = 30-50 m, P3 = 70-100 m).
(a)
STATISTICA. MANOVA GENERALESynthèse de tous les effetsPlan: 1-PROF
dl Mc dl Mc
Effet Effet Effet Erreur Erreur F niveau P
1 2 7,199952 13 ,110532 65,13882 ,000000
(b)
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocINTERACTION: 1 × 2
STADE × Adulte Adulte Adulte Juvénile Juvénile Juvénile
Profondeur × P1 × P2 × P3 × P1 × P2 × P3
Adulte × P1 ,003602 ,026222 ,004949 -- --
Adulte × P2 ,003602 ,293307 ,700490 -- --
Adulte × P3 ,026222 ,293307 ,263709 -- --
Juvénile × P1 ,004949 ,700490 ,263709 -- --
Juvénile × P2 -- -- -- -- --
Juvénile × P3 -- -- -- -- --
370

Annexe 46 - Modifications de la signature isotopique en carbone (δ13C) en fonction de la taille et de laprofondeur chez les individus de Citharus linguatula au large du Rhône.
(a) ANOVA à un facteur (profondeur) réalisée sur les δ13C obtenus pour les adultes de C. linguatula capturés àchaque profondeur (P2 = 30-50 m, P3 = 70-100 m).
(b) Résultats des tests de comparaisons a posteriori des δ13C obtenus pour les individus de C. linguatula de chaquestade (juvénile/adulte) pour chaque profondeur (P2 = 30-50 m, P3 = 70-100 m).
(a)
STATISTICA. MANOVA GENERALESynthèse de tous les effetsPlan: 1-PROFONDEUR
dl Mc dl Mc
Effet Effet Effet Erreur Erreur F niveau P
1 2 ,129719 22 2,012045 ,064471 ,801922
(b)
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocINTERACTION: 1 × 2
STADE × Adulte Adulte Juvénile Juvénile
Profondeur × P2 × P3 × P2 × P3
Adulte × P2 ,899492 ,877993 ,453967
Adulte × P3 ,899492 ,723035 ,651489
Juvénile × P2 ,877993 ,723035 ,602781
Juvénile × P3 ,453967 ,651489 ,602781
371

Annexe 47 - Modifications de la signature isotopique en carbone (δ15N) en fonction de la taille et de laprofondeur chez les individus de Citharus linguatula au large du Rhône.
(a) ANOVA à un facteur (profondeur) réalisée sur les δ15N obtenus pour les adultes de C. linguatula capturés àchaque profondeur (P2 = 30-50 m, P3 = 70-100 m).
(b) Résultats des tests de comparaisons a posteriori des δ15N obtenus pour les individus de C. linguatula de chaquestade (juvénile/adulte) pour chaque profondeur (P2 = 30-50 m, P3 = 70-100 m).
(a)
STATISTICA. MANOVA GENERALESynthèse de tous les effetsPlan: 1-PROFONDEUR
dl Mc dl Mc
Effet Effet Effet Erreur Erreur F niveau P
1 1 ,051810 22 ,078475 ,660215 ,425190
(b)
STATISTICA. MANOVA GENERALETest Newman-KeulsProbas des Tests Post HocINTERACTION: 1 × 2
STADE × Adulte Adulte Juvénile Juvénile
Profondeur × P2 × P3 × P2 × P3
Adulte × P2 ,163192 ,000145 ,000151
Adulte × P3 ,163192 ,000175 ,000145
Juvénile × P2 ,000145 ,000175 ,946544
Juvénile × P3 ,000151 ,000145 ,946544
372

LISTE DES PUBLICATIONS,
COMMUNICATIONS ORALES
ET POSTERS DIRECTEMENT
ISSUS DE CE TRAVAIL


373
PUBLICATIONS, COMMUNICATIONS ORALESET POSTERS DIRECTEMENT ISSUS DE CE TRAVAIL
(cette liste est non exhaustive)
PUBLICATIONS Darnaude A. M. (sous presse). Résumé de thèse. Apports fluviaux en zone côtière et réseaux
trophiques marins benthiques : transfert de matière organique particulaire terrigène jusqu'auxpoissons plats au large du Rhône. Cybium.
Darnaude A. M., Salen-Picard C., Polunin N.V.C., and M.L. Harmelin-Vivien (soumise àOecologia). Transfer of terrestrial organic matter into marine coastal food webs throughdepositivorous polychaetes elucidated by stable isotopes off the Rhone River delta (Gulf of Lions,NW Mediterranean).
En préparation: Darnaude A. M. Apports fluviaux en zone côtière et réseaux trophiques marinsbenthiques : transfert de matière organique particulaire terrigène jusqu'aux poissons plats au largedu Rhône.
En préparation: Darnaude, A. M., Salen-Picard, C. and M. L. Harmelin-Vivien. Spatial variationsin terrestrial particulate organic matter use by marine coastal benthic communities off the RhoneRiver delta (Gulf of Lions, NW Mediterranean).
En préparation: Darnaude, A. M.. Incorporation of material from terrestrial origin in the flesh offive marine flatfish species living off the Rhone River delta (Gulf of Lions, NW Mediterranean):intra- and inter-specific differences related to fish diet.
COMMUNICATIONS ORALES Darnaude A.M., Harmelin-Vivien M., Salen-Picard C., Polunin N.V.C. (2001). "Variation de la
signature en isotopes stables (δ13C et δ15N) chez les poissons plats au large du delta du Rhône".3èmes Journées de l’Ecologie Fonctionnelle, La Londe Les Maures - mars 2001.
Salen-Picard C., Darnaude A.M. and Harmelin-Vivien M. (2001). "Fluctuations of macrobenthicpopulations: a link between climatic variability, runoff and fisheries". European Marine BiologySymposium. Minorque, Espagne - septembre 2001.
Harmelin-Vivien M. L., Darnaude A.M., Salen-Picard C. (2001). "Du climat aux pêcheriesdémersales: le rôle des apports terrigènes dans les réseaux trophiques côtiers". Réunion PNEC-GLOBEC. Paris - décembre 2001.
Darnaude A.M. (2002). "Du climat aux pêcheries démersales: le rôle des apports terrigènes dansles réseaux trophiques côtiers". 5ème Congrès de l'Ecole Doctorale Sciences de l'Environnement(E.D.S.E. 2002). Marseille - mars 2002.
Darnaude A. M., Polunin N.V.C., Salen-Picard C., Harmelin-Vivien M.L. (2002). Impact ofRhone River terrestrial particulate organic matter on five NW Mediterranean marine flatfish. FifthInternational Symposium on Flatfish Ecology. Ile de Man (U.K.) - novembre 2002.
Darnaude A. M., Salen-Picard C., Harmelin-Vivien M.L. (2003). Stable isotope evidence ofterrestrial organic matter incorporation into coastal marine food webs: impact of Rhone Riverinputs on five NW Mediterranean marine flatfish species". EGS-AGU-EUG Joint Assembly 2003.Nice - avril 2003.
POSTERS Darnaude A.M. (2000). "Crues du Rhône et pêcheries de poissons plats : une histoire qui dure…
Mais par quel miracle ?". 5èmes Doctoriales. La Baume-les-Aix, France - mai 2001. Darnaude A.M., Harmelin-Vivien M.L. (2001). "Apports terrigènes et eutrophisation des zones
côtières : impact sur les populations de poissons démersaux au large du Rhône". ColloqueQuadriennal de Restitution du CIRMED. Banyuls-sur-Mer, France - janvier 2002.



Apports fluviaux en zone côtière et réseaux trophiques marins benthiques : transfert de matièreorganique particulaire terrigène jusqu'aux poissons plats au large du Rhône
La matière organique apportée par les fleuves participe fortement au fonctionnement des écosystèmes côtiers. Le Rhône,est la principale source de matière organique continentale en Méditerranée. Ses apports en matériel dissous favorisent lesproductions planctonique et benthique au large de son delta. Cependant, les fluctuations interannuelles de son débit serépercutent principalement sur ses apports en matière organique particulaire (MOP). Celle-ci sédimente au niveau du prodelta dufleuve où son incorporation dans les réseaux trophiques benthiques intervient pour une grande part dans les fluctuationsinterannuelles du macrobenthos. En se répercutant le long des chaînes trophiques, ce phénomène est susceptible d'entraîner, àterme, des variations temporelles non négligeables des populations de prédateurs benthiques, dont notamment les poissonsdémersaux. Ce travail a permis d'estimer et d'expliquer les réactions potentielles des cinq principales espèces de poissons platscapturées au large du fleuve (Solea solea, S. impar, Buglossidium luteum, Arnoglossus laterna, Citharus linguatula) à uneaugmentation de son débit. Dans un premier temps, l'étude de la répartition spatio-temporelle et de l’alimentation des cinqespèces a permis de décrire leurs stratégies de vie et le partage des ressources entre leurs individus. Dans un deuxième temps, lesuivi (grâce aux isotopes stables : δ13C et δ15N) du devenir du carbone d’origine terrestre au sein des réseaux trophiquesbenthiques a permis de reconstituer les modalités du transfert alimentaire de matériel terrigène vers les poissons, de déterminerl'influence des stratégies de vie des espèces sur leurs sensibilités respectives aux apports du fleuve et d'émettre des hypothèsesquant à l'impact éventuel de ces derniers sur leurs populations. Au large du Rhône, la MOP terrigène est essentiellement présenteau sein du sédiment, avec un maximum d'accumulation à 30-50 m de profondeur. Elle intègre les réseaux trophiques benthiquespar le biais de divers consommateurs primaires parmi lesquels les polychètes dépositivores tiennent une place prépondérante.Elle est ensuite transférée jusqu'aux poissons, soit par ingestion directe de ces organismes, soit par le biais de consommateurssecondaires s'en nourrissant. L'intensité du transfert dépend ainsi du régime alimentaire des poissons mais également de latranche bathymétrique considérée, avec un maximum à 30-50 m lié à un degré d'exploitation maximal de la MOP terrigène parles communautés macrobenthiques à cette profondeur. Elle diffère fortement entre S. solea, S. impar, B. luteum, A. laterna et C.linguatula, du fait de l'existence d'importantes différences de preferenda alimentaires et de répartition bathymétrique entre cescinq poissons plats. Compte-tenu des régimes alimentaires, de la répartition spatio-temporelle et de l'adaptabilité trophique desjuvéniles et des adultes des cinq espèces, on peut s'attendre, en cas d'augmentation des apports du Rhône au niveau du fond, à unimpact négligeable sur la population de C. linguatula mais à une réponse durable des populations des quatre autres poissons platsétudiés. Celle-ci devrait être importante chez B. luteum, A. laterna et S. impar et maximale chez la sole commune, S. solea, avecles conséquences que cela implique pour l'exploitation des stocks de cette espèce à haute valeur commerciale.
Mots-clefs : zones marines côtières, apports terrigènes, poissons plats, réseaux trophiques benthiques, isotopes stables,alimentation, pêcheries, Méditerranée NO.
River inputs to coastal areas and marine benthic food webs: trophic transfer of particulate organicmatter from terrestrial origin up to five flatfish species living off the Rhone River delta.
Rivers are known to enhance the productivity of marine coastal areas through the input of nutrients and organic matter ofterrestrial origin. The Rhone River is the main supplier of terrigenous matter to the Mediterranean basin. Its inputs in dissolvedorganic matter and nutrients influence both pelagic and benthic productions off its delta. However, the year-to-year fluctuationsof the river discharge mainly result in variations of terrigenous particulate organic matter (POM) fluxes. The POM brought bythe river is mainly deposited in the prodelta where its incorporation in marine benthic food webs plays a major role indetermining the macrobenthic communities fluctuations. This phenomenon could lead to important temporal variations in thepopulation size of some benthic top predators as demersal fishes. This work was carried out in order to estimate and explain thepotential responses of the five main flatfish species living off the Rhone delta (Solea solea, S. impar, Buglossidium luteum,Arnoglossus laterna and Citharus linguatula) to an increase in river discharge. The spatio-temporal distribution and the feedingof juvenile and adult fish were first studied. This provided a description of the five species life strategies off the Rhone Riverdelta and allowed to estimate the intensity of resource partitioning between their individuals in this area. Nitrogen (δ15N) andcarbon (δ13C) stable isotopes were then used to follow the trophic transfer of terrigenous POM along benthic food webs. Thisallowed to estimate the intensity of the terrigenous POM transfer up to each fish species, to determine the role played by fish lifestrategy in this respect and to build hypotheses about fish populations responses to an increase in river inputs. Off the RhoneRiver, terrigenous POM is mainly trapped in the sediment compartment, with a maximum between 30 and 50 m depth. It entersmarine benthic food webs via various primary consumers among which depositivorous polychaetes play a major role. It is thentransferred up to fishes by the ingestion of these prey organisms or other benthic consumers that feed upon them. Thus, transferintensity depends not only on fish diet but also on depth range, with a maximum between 30 and 50 m due to a maximal POMuse by the benthic community. It differs greatly between S. solea, S. impar, B. luteum, A. laterna and C. linguatula due todifferences both in food preferences and in bathymetric distribution between these flatfish species. According to fish speciesdiets, spatio-temporal distributions and trophic adaptabilities, one can predict a negligible effect of Rhone River floods on C.linguatula population, but a significant and long-lasting impact on the four other species populations. The response intensityshould be important in B. luteum, S. impar and A. laterna and maximum in the common sole, S. solea, with strong consequenceson the local fishery of this high value fish.
Keywords: coastal marine areas, terrestrial inputs, flatfishes, benthic food webs, stable isotopes, feeding, fisheries, NWMediterranean