apolipoprotein e isoform-specific regulation of dendritic spine morphology in apolipoprotein e...
TRANSCRIPT

APOLIPOPROTEIN E ISOFORM-SPECIFIC REGULATION OFDENDRITIC SPINE MORPHOLOGY IN APOLIPOPROTEIN ETRANSGENIC MICE AND ALZHEIMER’S DISEASE PATIENTS
Y. JI,a Y. GONG,d W. GAN,d T. BEACH,e
D. M. HOLTZMANf AND T. WISNIEWSKIa,b,c*aDepartment of Neurology, New York University School of Medicine,New York, NY 10016, USAbDepartment of Psychiatry, New York University School of Medicine,New York, NY 10016, USAcDepartment of Pathology, New York University School of Medicine,New York, NY 10016, USAdDepartment of Physiology and Neuroscience, Skirball Institute, NewYork University School of Medicine, New York, NY 10016, USAeSun Health Research Institute, Sun City, AZ 85351, USAfThe Departments of Neurology, Molecular Biology and Pharmacology,Center for the Study of Nervous System Injury, Washington UniversitySchool of Medicine, 660 South Euclid Avenue, Box 8111, St. Louis,MO 63110, USA
Abstract—Dendritic spines are postsynaptic sites of excita-tory input in the mammalian nervous system. Apolipoprotein(apo) E participates in the transport of plasma lipids and inthe redistribution of lipids among cells. A role for apoE isimplicated in regeneration of synaptic circuitry after neuralinjury. The apoE4 allele is a major risk factor for late-onsetfamilial and sporadic Alzheimer’s disease (AD) and is asso-ciated with a poor outcome after brain injury. ApoE isoformsare suggested to have differential effects on neuronal repairmechanisms. In vitro studies have demonstrated the neuro-trophic properties of apoE3 on neurite outgrowth. We haveinvestigated the influence of apoE genotype on neuronal celldendritic spine density in mice and in human postmortemtissue. In order to compare the morphology of neurons de-veloping under different apoE conditions, gene gun labelingstudies of dendritic spines of dentate gyrus (DG) granulecells of the hippocampus were carried out in wild-type (WT),human apoE3, human apoE4 expressing transgenic mice andapoE knockout (KO) mice; the same dendritic spine parame-ters were also assessed in human postmortem DG from in-dividuals with and without the apoE4 gene. Quantitative anal-ysis of dendritic spine length, morphology, and number wascarried out on these mice at 3 weeks, 1 and 2 years of age.Human apoE3 and WT mice had a higher density of dendriticspines than human E4 and apoE KO mice in the 1 and 2 yearage groups (P<0.0001), while at 3 weeks there were no dif-ferences between the groups. These age dependent differ-ences in the effects of apoE isoforms on neuronal integritymay relate to the increased risk of dementia in aged individ-uals with the apoE4 allele. Significantly in human brain,
apoE4 dose correlated inversely with dendritic spine densityof DG neurons cell in the hippocampus of both AD(P�0.0008) and aged normal controls (P�0.0015). Our find-ings provide one potential explanation for the increased cog-nitive decline seen in aged and AD patients expressingapoE4. © 2003 IBRO. Published by Elsevier Ltd. All rightsreserved.
Key words: Alzheimer’s disease, apolipoprotein E, trans-genic mice, dendrite spine, dentate gyrus, neuronal plastic-ity.
Many types of neurons have dendrites with specializedsynaptic contact sites, called dendritic spines (Bonhoefferand Yeste, 2002; van Rossum, 1999). Dendritic spines arethe loci of synaptic interactions among central neurons.Their susceptibility to changes in afferent stimulationmakes dendritic spines the prime candidates for serving asthe long-term morphological substrates of neuronal plas-ticity. In the hippocampus, which has been associated withlearning and memory, spines may be essential for theinduction, specificity, and endurance of long-term potenti-ation (Bonhoeffer and Yeste, 2002; Harris and Kater,1994). Dendrite spine formation is the most characteristicfeature in structural plasticity of mature neuronal syn-apses.
Alzheimer’s disease (AD) is an age-dependent neuro-degenerative disorder that is characterized by a progres-sive cognitive decline and by characteristic morphologicalCNS alterations, including deposition of amyloid � (A�)peptides in parenchymal plaques and cerebral blood ves-sels; intraneuronal formation of neurofibrillary tangles; andloss of neuronal subpopulations, synaptophysin-immuno-reactive presynaptic terminals, and cholinergic fibers(Terry, 1998; Wegiel et al., 2003). One major hallmark ofAD is the massive loss of synapses that occurs at an earlyclinical stage of the disease, particularly in the molecularlayer of the dentate gyrus (DG) of the hippocampal forma-tion (Masliah et al., 1991, 1992; Sze et al., 1997) and, to alesser degree, in other hippocampal and neocortical re-gions (Honer et al., 1992; Lippa et al., 1992; Masliah et al.,1991). Synaptic loss has been correlated with the cognitiveimpairment of AD by several authors (DeKosky and Scheff,1990; Lippa et al., 1992; Terry et al., 1991). Previousstudies have also demonstrated a negative correlationbetween synaptic size and synaptic density in the hip-pocampus, as well as in other brain regions in AD patientsand normal aged individuals (DeKosky and Scheff, 1990;Scheff et al., 1996).
*Correspondence to: T. Wisniewski, New York University MedicalCenter, Department of Neurology, Millhauser Laboratory, HN419, 550First Avenue, New York, NY 10016, USA. Tel: �1-212-263-7993; fax:�1-212-263-7528.E-mail address: [email protected] (T. Wisniewski).Abbreviations: A�, amyloid �; AD, Alzheimer’s disease; ANOVA, anal-ysis of variance; apoE, apolipoprotein E; DG, dentate gyrus; KO,knock-out; WT, wild-type.
Neuroscience 122 (2003) 305–315
0306-4522/03$30.00�0.00 © 2003 IBRO. Published by Elsevier Ltd. All rights reserved.doi:10.1016/j.neuroscience.2003.08.007
305

Apolipoprotein E (apoE), a 34-kDa protein involved incholesterol uptake and transport (Mahley, 1988), is ex-pressed throughout the brain, and is produced mainly inastrocytes and microglia (Mahley et al., 1996). ApoE isimportant for synaptic maintenance during aging (Masliahet al., 1997) and it may protect the CNS from a variety ofchallenges (De Chaves et al., 1997; Handelmann et al.,1992; Poirier et al., 1995). The association of allele four ofapoE (apoE4) as a genetic risk factor for AD has been wellestablished, with patients expressing the E4 allele havingan earlier age at onset and having a greater amyloidburden (Corder et al., 1993; Sanan et al., 1994; Wisniewskiand Frangione, 1992).
Previous in vitro studies have shown that exogenousapoE3 can stimulate neurite outgrowth, while exogenousapoE4, decreased outgrowth (Nathan et al., 1994). Similarobservations of isoform-specific effects of apoE3 andapoE4 have been described in a murine neuroblastomacell line (Neuro-2a; Bellosta et al., 1995). Furthermore,embryonic hippocampal neurons from WT mice grown inthe presence of astrocytes derived from apoE3 transgenicmice showed significantly greater neurite outgrowth thanthose grown in the presence of astrocytes from apoE4transgenic mice (Sun et al., 1998). Subsequent studiesemploying hippocampal slices harvested from transgenicanimals, with human apoE gene insertions, revealed im-paired sprouting in slices expressing human apoE4 (Teteret al., 1999). A further study using human apoE transgenicmice demonstrated that after a global ischemic insult,apoE4 expressing mice displayed greater neuronal celldeath than E3 mice (Horsburgh et al., 2000). On the otherhand, another study showed no effect of apoE isotypeexpression on neuronal cell death due to excitotoxic andapoptotic agents in vitro or following neonatal hypoxicischemia in vivo (Lendon et al., 2000).
Using a newly developed plasma membrane labelingtechnique (Gan et al., 2000), we examined detailed struc-tural changes in dendritic spines in the DG of human apoEtransgenic mice and human brains. We show apoE iso-form-specific effects on dendritic spine density in vivo inboth mouse and human postmortem tissue.
EXPERIMENTAL PROCEDURES
Animals
C57BL/6 male mice used for these experiments were wild-type(WT), and murine apoE�/� (knock-out, KO). In addition, humanapoE3�/� and apoE4�/� on a murine apoE�/� background, wereused (Holtzman et al., 1999; Holtzman et al., 2000a; Sun et al.,1998). The expression level of apoE in the transgenic mouselines, and in the WT mice was equivalent in the brain as deter-mined by Western blot analysis, whereas immunoreactivity wasnot detected in apoE KO mice (Sun et al., 1998). Mice weremaintained in a temperature- and humidity-controlled environmentand kept under a 12-h light/dark cycle with access to food andsterile water ad libitum. Surgical and animal care procedures wereperformed with strict adherence to the Guide for the Care and useof Laboratory Animals (National Institutes of Health Publicationnumber 86–23; Bethesda, MD, USA). The minimum number ofanimals was used and care was taken to minimize any suffering.
Quantitative analysis of dendritic spine length, morphology,and number was carried out at 3 weeks, 12 and 24 months of age.Mice were deeply anesthetized with a mixture of 80 mg/kg ket-amine and 5 mg/kg xylazine delivered intraperitoneally for eachgroup, and then perfused with 4% paraformaldehyde. Brains wereremoved and postfixed in 4% paraformaldehyde for 1 h and then10 �m sections were cut on a vibratome.
Human brains and apoE isotyping
Two-hundred micrometer thick fresh sections including the DG ofhippocampus tissue was obtained from the Sun Health ResearchInstitute (Sun City, AZ, USA) and the New York University Alzhei-mer’s Disease Center (New York, NY, USA). Twenty-four ADcases (mean age�S.E., 84.07�2.24 years; postmortem delay1–3 h), and 10 cases without any AD pathological changes(88.19�3.05 years; postmortem delay 1–3 h) were examined. ADcases were diagnosed clinically with dementia and fulfilled Na-tional Institute on Aging-Reagan criteria for AD at autopsy andwere matched for clinical stage of disease (Gearing et al., 1995;The National Institute on Aging and The Reagan Institute WorkingGroup on Diagnositic Criteria for the Neuropathological Assess-ment of Alzheimer’s Disease, 1997). Control subjects had noclinical history of dementia and no significant neuropathologicalchanges indicative of AD or other clinically relevant disease (Table1). Subjects that by clinical history had no indication of dementia,but on histological examination of the hippocampus were found tohave more than rare amyloid plaques, were excluded from thestudy group.
Genomic DNA was isolated by treating 15–25 mg of frozenbrain tissue with proteinase K. DNA was extracted by phenol/chloroform/isoamyl alcohol treatment, recovered by ethanol pre-cipitation, dried, and resuspended in buffer. A 227 bp sequence ofthe apoE gene was amplified by PCR using primers and reactionconditions as described (Wenham et al., 1991). Genotyping wasperformed after digestion with the restriction enzyme CfoI andseparation of fragments on a 12% polyacrylamide gel (Wenham etal., 1991).
Western blot
Western blot analysis of neocortical tissue extracts was performedto assess the expression of apoE in the mice in each group, usingmethods as described (Permanne et al., 1997; Sun et al., 1998).Three mice per genotype and per age group were analyzed and50 �g of protein were loaded per lane.
Labeling of neurons
Neuronal structures were labeled with the DiOlistic technique aspreviously described (Gan et al., 2000; Grutzendler et al., 2003).Dye-coated particles were delivered to the brain slices using acommercially available biolistic device, “gene gun” (BioRad, Her-cules, CA, USA; Helios Gene Gun System catalog no. 165–2431).Dye-coated particles were delivered at a helium gas pressure of180 pounds per square inch (psi) with a 5 mm distance betweenthe gene gun and the preparation. Brain slices were shot on bothsides to ensure sufficient labeling.
Table 1. Age, post-mortem delay and apoE genotypes of the patientsstudied
Age in years(�S.D.)
Postmortemdelay�h
ApoE genotypes
E3.3 E3.4 E4.4
Controls 89.60�5.62 2.52�0.58 6 4 0AD 83.08�7.85 2.63�0.60 9 12 3
Y. Ji et al. / Neuroscience 122 (2003) 305–315306

Slices were then kept in 0.1 M PBS solution for 2–3 h at roomtemperature or 4 °C overnight, to allow complete dye diffusion.Tissues were then post-fixed in 4% paraformaldehyde for 1 h atroom temperature and washed with 0.1 M PBS.
Amyloid staining
In human tissue, amyloid plaques were stained with Thioflavin Sand Campbell-Switzer techniques using standard methods(Wisniewski et al., 1995). In addition, to assess the amyloid bur-den in AD and control patient tissue, sections that were sequentialto sections used for neuronal labeling were immunostained withmonoclonal anti-A� antibody 6E10 as previously described (Sig-urdsson et al., 2001). Briefly, sections were incubated in 6E10 at1:1000 dilution, followed by an anti-mouse IgG secondary anti-body at a 1:2000 dilution (Vector Elite staining kit; Vector labora-tories, Burlingame, CA, USA). The sections were reacted in 3,3�-diaminobenzidine tetrahydrochloride with or without nickel ammo-nium sulfate (Mallinckrodt, Paris, KY, USA) intensification.
Imaging of neuronal structures
Images of labeled neurons were acquired by a Zeiss LSM 510confocal attached to an upright Zeiss Axioplan two microscope. Asingle low-power scan (10�) was made of each filled neuron,followed by using a 60�/1.30 oil-immersion DIC Plan-Neuofluarobjective. Neuronal structures labeled with different lipophilic dyesare sequentially scanned using one of the three appropriate exci-tation lasers (argon 458, 488, 514 nm for DiO and Thioflavin S,HeNe1 543 nm for DiI, HeNe2 633 nm for DiD) combined with theappropriate emission filter (LP 560 for DiI, LP 505 for DiO). Stacksof images at 0.5 �m steps are acquired to generate three-dimen-sional data sets of imaged neurons. The total number of opticalsections taken during the 60� high-power scan to complete thethree dimensional reconstruction of the cell, depended on thelength of the dendrites and the depth that it traveled through thesection. Quantification of spine density, dendritic diameter, andspine number were quantified with Metamorph software (Univer-sal Imaging Corporation, PA, USA). Confocal microscopy andanalysis of neuronal morphology was performed blind, and thecode was not broken until the data collection was complete. In themouse tissue, a subset of filled DG neurons from each animal waschosen for confocal microscopy based on the quality of the fill andlocalization in the DG (six to 10 cells per animal; total 20–44 cellsper group in five mice). Only granule neurons in the DG thatsatisfied the following criteria were included: (I) complete impreg-nation (including all dendrites), not obscured by other neurons orartifacts; (II) clear image. Cells used for counting which met boththese criteria were picked using systemic random sampling, withoverlay of an x–y grid (Hyman et al., 1998; Wegiel et al., 1998).For each neuron, all of the dendrites and their primary brancheswere traced to their natural or artificial ends in order to quantifytheir number and length. The length of more distal dendritebranches was not counted. The length of 10 (�2, S.D.) dendriteswas calculated per cell. Spine density was calculated by dividingthe total number of spines counted on all identified dendrites bythe total length of the dendrites and was expressed as the numberof spines per 10 �m of dendrite.
For human brain, these neurons were located within the DG ofthe hippocampus and were required to have no breaks in stainingalong its dendrites. In addition, counted cells were at least 200 �maway from any amyloid plaques seen by Thioflavin S staining. The
Three Week Old Mice
WT E3 E4 KO
750
1000
1250
1500
A
n=16 n=20 n=12 n=18
To
tal
Den
dri
te S
pin
es/
DG
neu
ron
12 Month Old Mice
WT E3 E4 KO
750
1000
1250
1500
n=31 n=17 n=44 n=24To
tal
Den
dri
te S
pin
es/
DG
neu
ron
24 Month Old Mice
WT E3 E4 KO
750
1000
1250
1500
C
n=17 n=14 n=26 n=15
To
tal
Den
dri
te s
pin
es/
DG
Neu
ron
B
Fig. 1. Total dendritic spine numbers per neuron in the DG of mice. A)Shows there were no significant differences in the total number ofdendritic spines in the four mouse groups at 3 weeks of age. B) Showsthe reduced total number of spines at 1 year of age in the transgenicmice expressing apoE4 and apoE KO mice compared with WT andapoE3 expressing mice (P�0.0001, one-way ANOVA for genotypeeffect). Post hoc Newman-Keuls multiple comparison tests show bothapoE4 and KO mice to be significantly different from both WT andapoE3 mice (P�0.001). C) Shows the reduced total number of spinesat 2 years of age in the transgenic mice expressing apoE4 and apoEKO mice compared with WT and apoE3 expressing mice (P�0.0001,one-way ANOVA, genotype effect). Post hoc Newman-Keuls multiple
comparison tests show apoE4 mice to be significantly different fromboth WT and apoE3 expressing mice (P�0.001). KO mice are alsosignificantly different from both WT and apoE3 mice (P�0.01). Num-bers below the bars indicate the total number of cells examined in eachgroup of mice.
Y. Ji et al. / Neuroscience 122 (2003) 305–315 307

number of dendritic spines within a 50-�m radius ring progres-sively more distal from the soma was counted in human brainsamples. Only spines that were distinct from the dendritic branchwere counted. Spine density was calculated by dividing the num-ber of spines on a segment by the length of the segment and wasexpressed as the number of spines per 10 �m of dendrite.
Amyloid staining
Immunohistochemistry of the amyloid burden in human AD tissuesections was quantitated with a Bioquant image analysis systememploying unbiased sampling. All procedures were performed byan individual blind to the experimental conditions of the study. Theamyloid burden was determined in the DG. The area of the gridwas 800�800 �m2 and the amyloid load was measured in 10frames randomly chosen per AD patient sample (each640�480 �m2). The A� burden is defined as the percentage ofarea in the measurement field occupied by reaction product.
Statistical analysis
Data were analyzed with analysis of variance (ANOVA), followedby Newman-Keuls multiple comparison tests or t-tests when ap-plicable (GraphPad Prism Software, version 4.0). The length ofeach dendrite was traced and the number of dendritic spinescounted along each dendrite using a tag point technique. Nocorrection for hidden spines was made, but while counting thespines in the reconstructed images, position and verification ofspines were aided by the images in the single plane, to assure thatall seen spines were counted.
RESULTS
Total dendritic spine number per neuron in mice
Confocal microscopic imaging of gene gun labeling neu-rons provided excellent visualization of dendritic spines aspreviously reported (Gan et al., 2000; Grutzendler et al.,2003; see Fig. 4). A total of 20–43 granule cells wereincluded in the final analysis per group (five to 10 cells peranimal, four to five mice per group). All of the neuronsresided in the DG of the hippocampus. At 3 weeks of ageno difference was evident among the four groups of mice(Fig. 1A). However, by 12 months of age a striking differ-ence was noted in apoE4 and apoE KO mice versus WT(Fig. 1B) and apoE3 mice (P�0.0001, one-way ANOVA;genotype effect). Post hoc Newman-Keuls multiple com-parison tests showed both apoE4 and KO mice to besignificantly different from both WT and apoE3 mice(P�0.001). At 24 months a similar significant differencewas observed between both the apoE4 and KO mice ver-sus the WT and apoE3 mice (P0.0035, one-way ANOVA;genotype effect; Fig. 1C).
Dendritic length in mice
No significant difference in dendritic length was noted be-tween the four groups at 3 weeks of age (Fig. 2A). At both12 and 24 months of age some significant differences werenoted (Fig. 2B and C). At 12 months of age one-way
Three weeks mice
WT E3 E4 KO
300
400
500
n=16 n=20 n=12 n=18
Den
dri
te L
en
gth
(µ
m)
WT E3 E4 KO
300
400
500
n=30 n=17 n=40 n=23
Den
dri
te L
en
gth
( µm
)
A
B12 Month Old Mice
24 Month Old Mice
WT E3 E4 KO
300
400
500
n=17 n=14 n=26 n=15
Den
dri
te L
en
gth
( µm
)
C
Fig. 2. Dendritic length in the DG of mice. A) Shows there were nosignificant differences in the length of dendrites in the four mousegroups at 3 weeks of age. B) Shows the reduced length of dendrites at1 year of age in the transgenic mice expressing apoE4 and apoE KOmice (P0.0234, one-way ANOVA for genotype effect). Post hocNewman-Keuls multiple comparison tests show the apoE KO group tobe significantly different from the apoE3 group (P�0.05). The othergroups did not differ significantly by post hoc tests. C) Shows thereduced length of dendrites at 2 years of age in the transgenic miceexpressing apoE4 or apoE KO mice compared with WT or apoE3expressing mice (P0.0035, one-way ANOVA, genotype effect). Posthoc Newman-Keuls multiple comparison tests show the apoE4 mice todiffer from the apoE3 and WT mice significantly (P�0.01 and P�0.05
espectively). The KO group did not differ significantly from the othergroups. Numbers below the bars indicate the total number of cellsexamined in each group of mice.
Y. Ji et al. / Neuroscience 122 (2003) 305–315308
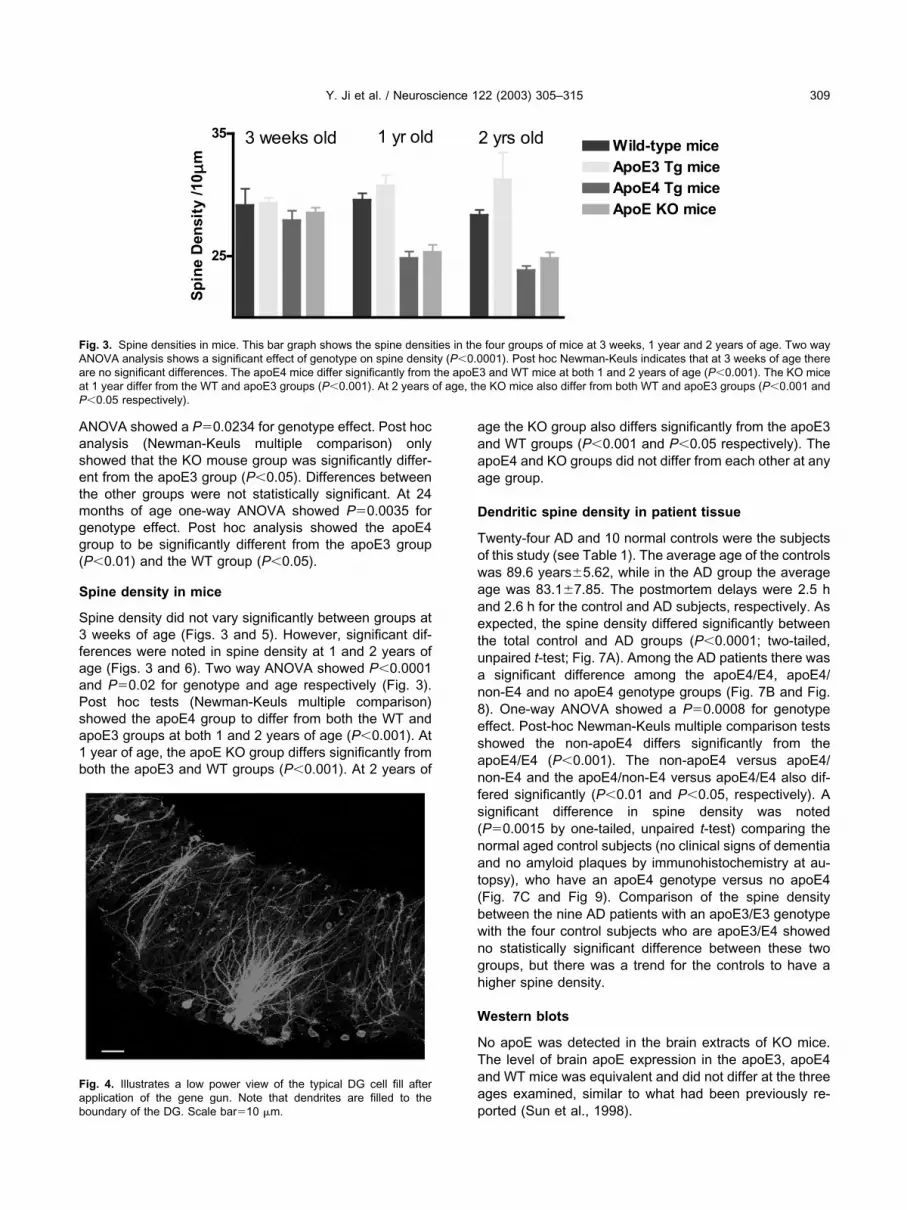
ANOVA showed a P0.0234 for genotype effect. Post hocanalysis (Newman-Keuls multiple comparison) onlyshowed that the KO mouse group was significantly differ-ent from the apoE3 group (P�0.05). Differences betweenthe other groups were not statistically significant. At 24months of age one-way ANOVA showed P0.0035 forgenotype effect. Post hoc analysis showed the apoE4group to be significantly different from the apoE3 group(P�0.01) and the WT group (P�0.05).
Spine density in mice
Spine density did not vary significantly between groups at3 weeks of age (Figs. 3 and 5). However, significant dif-ferences were noted in spine density at 1 and 2 years ofage (Figs. 3 and 6). Two way ANOVA showed P�0.0001and P0.02 for genotype and age respectively (Fig. 3).Post hoc tests (Newman-Keuls multiple comparison)showed the apoE4 group to differ from both the WT andapoE3 groups at both 1 and 2 years of age (P�0.001). At1 year of age, the apoE KO group differs significantly fromboth the apoE3 and WT groups (P�0.001). At 2 years of
age the KO group also differs significantly from the apoE3and WT groups (P�0.001 and P�0.05 respectively). TheapoE4 and KO groups did not differ from each other at anyage group.
Dendritic spine density in patient tissue
Twenty-four AD and 10 normal controls were the subjectsof this study (see Table 1). The average age of the controlswas 89.6 years�5.62, while in the AD group the averageage was 83.1�7.85. The postmortem delays were 2.5 hand 2.6 h for the control and AD subjects, respectively. Asexpected, the spine density differed significantly betweenthe total control and AD groups (P�0.0001; two-tailed,unpaired t-test; Fig. 7A). Among the AD patients there wasa significant difference among the apoE4/E4, apoE4/non-E4 and no apoE4 genotype groups (Fig. 7B and Fig.8). One-way ANOVA showed a P0.0008 for genotypeeffect. Post-hoc Newman-Keuls multiple comparison testsshowed the non-apoE4 differs significantly from theapoE4/E4 (P�0.001). The non-apoE4 versus apoE4/non-E4 and the apoE4/non-E4 versus apoE4/E4 also dif-fered significantly (P�0.01 and P�0.05, respectively). Asignificant difference in spine density was noted(P0.0015 by one-tailed, unpaired t-test) comparing thenormal aged control subjects (no clinical signs of dementiaand no amyloid plaques by immunohistochemistry at au-topsy), who have an apoE4 genotype versus no apoE4(Fig. 7C and Fig 9). Comparison of the spine densitybetween the nine AD patients with an apoE3/E3 genotypewith the four control subjects who are apoE3/E4 showedno statistically significant difference between these twogroups, but there was a trend for the controls to have ahigher spine density.
Western blots
No apoE was detected in the brain extracts of KO mice.The level of brain apoE expression in the apoE3, apoE4and WT mice was equivalent and did not differ at the threeages examined, similar to what had been previously re-ported (Sun et al., 1998).
Fig. 4. Illustrates a low power view of the typical DG cell fill afterapplication of the gene gun. Note that dendrites are filled to theboundary of the DG. Scale bar10 �m.
25
35
ApoE3 Tg mice
Wild-type mice
ApoE4 Tg mice
ApoE KO mice
3 weeks old 1 yr old 2 yrs old
Sp
ine D
en
sit
y /
10µ
m
Fig. 3. Spine densities in mice. This bar graph shows the spine densities in the four groups of mice at 3 weeks, 1 year and 2 years of age. Two wayANOVA analysis shows a significant effect of genotype on spine density (P�0.0001). Post hoc Newman-Keuls indicates that at 3 weeks of age thereare no significant differences. The apoE4 mice differ significantly from the apoE3 and WT mice at both 1 and 2 years of age (P�0.001). The KO miceat 1 year differ from the WT and apoE3 groups (P�0.001). At 2 years of age, the KO mice also differ from both WT and apoE3 groups (P�0.001 andP�0.05 respectively).
Y. Ji et al. / Neuroscience 122 (2003) 305–315 309

Amyloid staining
In control human tissue samples no amyloid plaques weredetected by 6E10 immunohistochemistry in the hippocam-pus. The amyloid burden in the DG, as expected, washigher among AD patients who were apoE3/4 or E4/4(5.2�0.6%) versus apoE3/3 (4.1�0.4%, P0.0003).
DISCUSSION
Our studies demonstrate the importance of apoE isotypeexpression for age-related changes in dendritic spine den-sity, both in animal models and in human tissue. Theinheritance of the apoE4 allele is well recognized to be themajor identified, genetic risk factor for the development oflate-onset AD (Laws et al., 2003; Wisniewski and Fran-gione, 1996). AD is the most common cause of dementiaand is characterized by the accumulation of A� in the formof senile plaques and congophilic angiopathy, neurofibrill-lary tangle formation and extensive synaptic loss. TheapoE protein exists in three major isoforms (apoE2, E3 andE4), which differ by single amino acid substitutions atresidues 112 and 158 (Mahley, 1988). In Caucasian pop-ulations the risk of developing AD rises greatly with in-creasing apoE4 dose. The odds ratio for apoE genotypes 2/ 4, 3/ 4, 4/ 4 are 2.6, 3.3 and 14.9 respectively
(Farrer et al., 1997). The mechanism by which apoE playsa role in AD pathogenesis is not clear and multiple path-ways may be involved. However, abundant evidence sug-gests that apoE4 can act as a “pathological chaperone”(Wisniewski and Frangione, 1992), promoting A� peptideaggregation and deposition into amyloid plaques. It hasbeen shown that under certain in vitro conditions apoE canpromote a �-sheet conformation in A� peptides (Ma et al.,1994; Sanan et al., 1994; Soto et al., 1996; Wisniewski etal., 1994). Studies using apoE knockout mice that werecrossed with AD models, APPV717F and APPsw transgenicmice, have shown that the presence of apoE is needed foramyloid plaque formation, consistent with this hypothesis(Bales et al., 1997; Holtzman et al., 2000b).
ApoE has also been shown, in some studies, to becritical for neuronal plasticity (Poirer et al., 1995). ApoE isthe major lipid transporter in the CNS (Mahley, 1988). Anincrease in apoE synthesis is associated with neuronalgrowth and repair following injury in both the peripheraland CNS (Boyles et al., 1989; Poirier et al., 1995). ApoEexpression is also increased in association with AD relatedinjury (Yamagata et al., 2001; Zarrow and Victoroff, 1998).ApoE’s role in neuronal plasticity also shows significantisoform specificity. Following entorhinal cortical injury thecompensatory sprouting and reactive synaptogenesis
Fig. 5. Shows the typical appearance of spines density in WT (A), apoE3 (B), apoE4 (C) and apoE KO (D) mice at 3 weeks of age. No apparentdifferences in spine density are noted. Scale bar10 �m.
Y. Ji et al. / Neuroscience 122 (2003) 305–315310

have been shown to be impaired in mice expressing hu-man apoE4 compared with apoE3 expressing mice (Poireret al., 1995; White et al., 2001). A similar negative effect ofapoE4 expression has been reported following excitotoxicinduced neurodegeneration (Buttini et al., 2000) and fol-lowing global ischemia in adult mice (Horsburgh et al.,2000). However, no apoE isotype expression effect wasseen following hypoxic ischemia in neonatal mice (Lendonet al., 2000). Tissue culture studies have also shown areduced function of apoE4. Neurons in vitro receivingapoE3 containing lipoprotein particles show increasedneurite outgrowth and branching compared with those re-ceiving apoE4 lipoparticles (Nathan et al., 1995; Postumaet al., 1998). These findings in mouse and cell culturemodels have been linked to the reported association of aworse outcome among apoE4 expressing individuals afterhead injury (Jang et al., 1996; Nicoll et al., 1995; Teasdaleet al., 1997), intracerebral hemorrhage (Alberts et al.,1995) and following cardiac surgery (Tardiff et al., 1997).Indeed, studies have shown that normal asymptomaticmiddle-aged individuals expressing the apoE4 allele havepreclinical memory impairment (Caselli et al., 2001; Floryet al., 2000) and normal aged individuals may have a morerapid memory decline (Deary et al., 2002; Dik et al., 2000).Such mild cognitive impairment has also been noted in
aged (but not young) apoE4 expressing transgenic micecompared with apoE3 expressing mice (Hartman et al.,2001; Raber et al., 2000).
The hippocampus and the perforant pathway of thehippocampal formation play an important role in learningand memory function. Synapse loss is an important andearly change in patients with clinical dementia due to AD.This change correlates well with the dementia that char-acterizes this disorder (Masliah et al., 1992; Terry, 1994)and has a predilection to affect the hippocampus and theperforant pathway (Wegiel et al., 2003). We examined thedendritic spines of neurons of the DG, which are part of theperforant pathway, using a novel gene gun technique.These dendritic spines are the sites of synapses and aredynamic structures critical for neuronal plasticity (Bonhoef-fer and Yeste, 2002). Our results show that in apoE4 andapoE KO mice, the total number, dendritic length and spinedensity are significantly reduced at both 1 and 2 years ofage with no differences being seen at 3 weeks of age(Figs. 1–3). The mice expressing apoE4 did not differ frommice where apoE was knocked out, suggesting a relativeloss of function of apoE4 versus apoE3 or murine apoE inthe maintenance of dendritic spines in the face of normalage induced stress. These results are in keeping with the
Fig. 6. Shows the typical appearance of spines density in WT (A), apoE3 (B), apoE4 (C) and apoE KO (D) mice at 1 year of age. A reduced numberof spines is evident in the apoE4 and apoE KO groups compared with the apoE3 and WT mice. Scale bar10 mm.
Y. Ji et al. / Neuroscience 122 (2003) 305–315 311

past reported isotype differences between the functions ofapoE3 and E4 in neuronal plasticity.
Significantly we found similar differences in humanpostmortem tissue. In the human AD tissue a clear apoE4dosage effect was seen with apoE4 homozygous individ-uals having the lowest spine density compared with pa-tients with no apoE4. A possible confounding variable inthe analysis of spine density in the AD patients is the
increased amyloid burden among our patients with anapoE4 allele, as has been previously reported among ADpatients at equivalent clinical stages of disease (Rebeck etal., 1993; Schmechel et al., 1993). This was minimized inour study in that spines were counted in neurons that wereat least 200 �m away from any amyloid plaques. However,among the control patients, where apoE4 expression wasalso associated with reduced spine density, amyloid
AD CTL
0
5
10
15
20
25
Sp
ine D
en
sit
y /
10
µm
E4/E4 E4/0 Non-E4
0
10
20
Sp
ine D
en
sit
y /
10µ
m
E4 Non-E4
0
5
10
15
20
25
Sp
ine D
en
sit
y /
10
µm
A
B
C
Fig. 7. Spine density in the DG using human tissue. A) Shows a bargraph comparing the spine densities in control versus all AD patients(P�0.0001, unpaired, two-tailed t-test). B) Shows a bar graph com-paring the spine densities in AD patients with differing apoE genotype(P0.0008, one-way ANOVA for genotype effect). Post hoc Neuman-Keuls tests show the non-apoE4 differs significantly from theapoE4/E4 (P�0.001). The non-apoE4 versus apoE4/non-E4 and theapoE4/non-E4 versus apoE4/E4 also differed significantly (P�0.01and P�0.05, respectively). C) Shows a bar graph comparing the spinedensity in control patients expressing the apoE allele and those that donot (P0.0015, unpaired, one-tailed t-test).
Fig. 8. Shows the typical spine density in AD patients with apoE3/E3(A), apoE3/E4 (B) and apoE4/E4 (C) genotypes. A much reducedspine density is evident in the apoE4/E4 group, with the apoE3/E4group having intermediate spine density. Scale bar10 �m.
Y. Ji et al. / Neuroscience 122 (2003) 305–315312

plaques could not be a confounding variable, since by theselection criteria, these individuals had no amyloid plaquesin the hippocampus by histology. The effect of expressingan apoE4 gene in older life appears to be detrimental fordendritic spine density in the DG. Comparison of the spinedensity between the nine AD patients with an apoE3/E3genotype with the four control subjects who are apoE3/E4showed no statistically significant difference betweenthese two groups, although the AD group had a trend forlower density. This suggests that the loss of dendriticspines in the DG of aged controls expressing apoE4 ap-proaches that seen with AD patients with extensive amy-loid deposition at that site, who do not express an apoE4.
Our findings of reduced dendritic spine density in theDG of aged mice and humans expressing apoE4, using anovel gene gun technique, provides one potential contrib-uting factor for the cognitive deficits that have been re-ported in animals and humans with this genotype. Thediffering negative roles apoE4 plays in both normal agingand in AD suggests that two potential therapeutic ap-proaches are agents that make apoE4 behave moreapoE2 or apoE3-like as well as compounds that block theinteraction between apoE and A� peptides that promotes
amyloid deposition. Such compounds are currently beingdeveloped.
Acknowledgements—This work was supported by NIH grantsAG15408, AG17617 and AG13956.
REFERENCES
Alberts MJ, Graffagnino C, McCleeny C, DeLong D, Strittmatter W,Saunders A, Roses AD (1995) ApoE genotype and survival fromintracerebral haemorrhage. Lancet 346:575.
Bales KR, Verina T, Dodel RC, Du YS, Altstiel L, Bender M, Hyslop P,Johnstone EM, Little SP, Cummins DJ, Piccardo P, Ghetti B, PaulSM (1997) Lack of apolipoprotein E dramatically reduces amyloid�-peptide deposition. Nat Genet 17:263–264.
Bellosta S, Nathan BP, Orth M, Dong LM, Mahley RW, Pitas RE (1995)Stable expression and secretion of apolipoproteins E3 and E4 inmouse neuroblastoma cells produces differential effects on neuriteoutgrowth. J Biol Chem 270:27063–27071.
Bonhoeffer T, Yeste R (2002) Spine motility: phenomenology, mech-anisms and function. Neuron 35:1019–1027.
Boyles JK, Zoellner CD, Anderson LJ, Kosik LM, Pitas RE, WeisgraberKH, Hui DY, Mahley RW, Gebicke-Haerter PJ, Ignatias MJ, ShooterEM (1989) A role for apolipoprotein E, apoprotein A-1, low densitylipoprotein receptors in cholesterol transport during regenerationand remyelination of the sciatic nerve. J Clin Invest 83:1015–1031.
Buttini M, Akeefe H, Lin C, Mahley RW, Pitas RE, Wyss-Coray T,Mucke L (2000) Dominant negative effects of apolipoprotein E4revealed in transgenic models of neurodegenerative disease. Neu-roscience 97:207–210.
Caselli RJ, Osborne D, Reiman EM, Hentz JG, Barbieri CJ, SaundersAM, Hardy J, Graff-Radford NR, Hall GR, Alexander GE (2001)Preclinical cognitive decline in late middle-aged asymptomatic apo-lipoprotein E-e4/4 homozygotes: a replication study. J Neurol Sci189:93–98.
Corder EH, Saunders AM, Strittmatter WJ, Schmechel DE, GaskellPC, Small GW, Roses AD, Haines JL, Pericak-Vance MA (1993)Gene dose of apolipoprotein E type 4 allele and the risk of Alzhei-mer’s disease in late onset families. Science 261:921–923.
De Chaves EI, Rusinol AE, Vance DE, Campenot RB, Vance JE(1997) Role of lipoproteins in the delivery of lipids to axons duringaxonal regeneration. J Biol Chem 272:30766–30773.
Deary IJ, Whiteman MC, Pattie A, Starr JM, Hayward C, Wright AF,Carothers A, Whalley LJ (2002) Cognitive change and the APOEepsilon 4 allele. Nature 418:932.
DeKosky ST, Scheff SW (1990) Synapse loss in frontal cortex biopsiesin Alzheimer’s disease: correlation with cognitive severity. AnnNeurol 27:457–464.
Dik MG, Jonker C, Bouter LM, Geerlings MI, van Kamp GJ, Deeg DJ(2000) APOE-epsilon 4 is associated with memory decline in con-gnitively impaired elderly. Neurology 54:1492–1497.
Farrer LA, Cupples LA, Haines JL, Hyman B, Kukull WA, Mayeux R,Myers RH, Pericak-Vance MA (1997) Effects of age, sex andethnicity on the association between apolipoprotein E genotype andAlzheimer disease: a meta-analysis: APOE and Alzheimer diseasemeta analysis consortium. JAMA 278:1349–1356.
Flory JD, Manuck SB, Ferrell RE, Ryan CM, Muldoon MF (2000)Memory performance and the apolipoprotein E polymorphism in acommunity sample of middle-aged adults. Am J Med Genet 96:707–711.
Gan WB, Grutzendler J, Wong WT, Wong RO, Lichtman JW (2000)Multicolor “DiOlistic” labeling of the nervous system using lipophilicdye combinations. Neuron 27:219–225.
Gearing M, Mirra SS, Hedreen JC, Sumi SM, Hansen LA, Heyman A(1995) The Consortium to Establish a Registry for Alzheimer’sDisease (CERAD): X. Neuropathology confirmation of the clinicaldiagnosis of Alzheimer’s disease. Neurology 45 (Pt 1):461–466.
Grutzendler J, Tsai J, Gan WB (2003) Rapid labeling of neuronal
Fig. 9. Shows the typical spine density aged patients with apoE3/E3(A) and apoE3/E4 (B) genotypes. A reduced spine density is evident inB. Scale bar10 �m.
Y. Ji et al. / Neuroscience 122 (2003) 305–315 313

populations by ballistic delivery of fluorescent dyes. Methods 30:79–85.
Handelmann GE, Boyles JK, Weisgraber KH, Mahley RW, Pitas RE(1992) Effects of apolipoprotein E, beta-very low density lipopro-teins, and cholesterol on the extension of neurites by rabbit dorsalroot ganglion neurons in vitro. J Lipid Res 33:1677–1688.
Harris KM, Kater SB (1994) Dendritic spines: cellular specializationsimparting both stability and flexibility to synaptic function. Ann RevNeurosci 17:341–371.
Hartman RE, Wozniak DF, Nardi A, Olney JW, Sartorius L, HoltzmanDM (2001) Behaviioral phenotyping of GFAP-apoE3 and apoE4transgenic mice: apoE4 mice show profound working memory im-pairments in the absence of Alzheimer’s like neuropathology. ExpNeurol 179:326–344.
Holtzman DM, Bales KR, Tenkova T, Fagan AM, Parsadanian M,Sartorius LJ, Mackey B, Olney J, McKeel D, Wozniak D, Paul SM(2000a) Apolipoprotein E isoform-dependent amyloid depositionand neuritic degeneration in a mouse model of Alzheimer’s dis-ease. Proc Natl Acad Sci USA 97:2892–2897.
Holtzman DM, Bales KR, Wu S, Bhat P, Parsadanian M, Fagan AM,Chang LK, Sun Y, Paul SM (1999) Expression of human apoli-poprotein E reduces amyloid-beta deposition in a mouse model ofAlzheimer’s disease. J Clin Invest 103:R15–R21.
Holtzman DM, Fagan AM, Mackey B, Tenkova T, Sartorius L, PaulSM, Bales KR, Hsiao Ashe K, Irizarry MC, Hyman BT (2000b)Apolipoprotein E facilitates neuritic and cerebrovascular plaqueformation in an Alzheimer’s disease model. Ann Neurol 47:739–747.
Honer WG, Dickson DW, Gleeson J, Davies P (1992) Regional syn-aptic pathology in Alzheimer’s disease. Neurobiol Aging 13:375–382.
Horsburgh K, McCulloch J, Nilsen M, Roses AD, Nicoll JA (2000)Increased neuronal damage and apoE immunoreactivity in humanapolipoprotein E, E4 isoform-specific, transgenic mice after globalcerebral ischemia. Eur J Neurosci 12:4309–4317.
Hyman BT, Gomez-Isla T, Irizarry MC (1998) Stereology: a practicalprimer for neuropathology. J Neuropathol Exp Neurol 57:305–310.
Jang M, Maestre G, Tsai W, Jui X, Feng L, Chung W, Churi M,Schofield P, Stern Y, Tycko B, Mayeux R (1996) Effects of age,ethnicity and head injury on the association between apoE geno-types and Alzheimer’s disease. Anna NY Acad Sci 802:6–15.
Laws SM, Hone E, Gandy S, Martins RN (2003) Expanding the asso-ciation between the APOE gene and the risk of Alzheimer’sdisease: possible roles for APOE promoter polymorphisms andalterations in APOE transcription. J Neurochem 84:1215–1236.
Lendon CL, Han BH, Salimi K, Fagan AM, Behrens MI, Muller MC,Holtzman DM (2000) No effect of apolipoprotein E on neuronal celldeath due to excitotoxic and apoptotic agents in vitro and neonatalhypoxia ischaemia in vivo. Eur J Neurosci 12:2235–2242.
Lippa CF, Hamos JE, Pulaski-Salo D, DeGennaro LJ, Drachman D(1992) Alzheimer’s disease and aging: effects on perforant pathwayperikarya and synapses. Neurobiol Aging 13:405–411.
Ma J, Yee A, Brewer HB Jr, Das S, Potter H (1994) Amyloid-associ-ated proteins alpha 1-antichymotrypsin and apolipoprotein E pro-mote assembly of Alzheimer beta-protein into filaments. Nature372:92–94.
Mahley RW (1988) Apolipoprotein E: cholesterol transport protein withexpanding role in cell biology. Science 240:622–630.
Mahley RW, Nathan BP, Pitas RE (1996) Apolipoprotein E: structure,function, and possible roles in Alzheimer’s disease. Ann NY AcadSci 777:139–145.
Masliah E, Ellisman M, Carragher B, Mallory M, Young S, Hansen L,DeTeresa R, Terry RD (1992) Three-dimensional analysis of therelationship between synaptic pathology and neuropil threads inAlzheimer disease. J Neuropathol Exp Neurol 51:404–414.
Masliah E, Samuel W, Veinbergs I, Mallory M, Mante M, Saitoh T(1997) Neurodegeneration and cognitive impairment in apoE-defi-
cient mice is ameliorated by infusion of recombinant apoE. BrainRes 751:307–314.
Masliah E, Terry RD, Alford M, DeTeresa R, Hansen LA (1991) Cor-tical and subcortical patterns of synaptophysin like immunoreactiv-ity in Alzheimer’s disease. Am J Pathol 138:235–246.
Nathan BP, Bellosta S, Sanan DA, Weisgraber KH, Mahley RW, PitasRE (1994) Differential effects of apolipoproteins E3 and E4 onneuronal growth in vitro. Science 264:850–852.
Nathan BP, Chang KC, Bellosta S, Brisch E, Ge N, Mahley RW, PitasRE (1995) The inhibitory effect of apolipoprotein E4 on neuriteoutgrowth is associated with microtubule depolymerization. J BiolChem 270:19791–19799.
Nicoll JA, Roberts GW, Graham DI (1995) Apolipoprotein E epsilon 4allele is associated with deposition of amyloid beta-protein followinghead injury. Nat Med 1:135–137.
Permanne B, Perez C, Soto C, Frangione B, Wisniewski T (1997)Detection of apolipoprotein E dimeric soluble amyloid � complexesin Alzheimer’s disease brain supernatants. Biochem Biophys ResCommun 240:715–720.
Poirer J, Minnich A, Davignon J (1995) Apolipoprotein E, synapticplasticity and Alzheimer’s disease. Ann Med 27:663–670.
Postuma RB, Martins RN, Cappai R, Beyreuther K, Masters CL,Strickland DK, Mok SS, Small DH (1998) Effects of the amyloidprotein precursor of Alzheimer’s disease and other ligands of theLDL receptor related protein on neurite outgrowth from sympatheticneurons in culture. FEBS Lett 428:13–16.
Raber J, Wong D, Gui-Qui Y, Buttini M, Mahley RW, Pitas RE, MuckeL (2000) Apolipoprotein E and cognitive performance. Nature 404:352–353.
Rebeck GW, Reiter JS, Strickland DK, Hyman BT (1993) Apolipopro-tein E in sporadic Alzheimer’s disease: allelic variation and receptorinteractions. Neuron 11:575–580.
Sanan DA, Weisgraber KH, Russell SJ, Mahley RW, Huang D, Saun-ders A, Schmechel D, Wisniewski T, Frangione B, Roses AD,Strittmatter WJ (1994) Apolipoprotein E associates with beta amy-loid peptide of Alzheimer’s disease to form novel monofibrils: iso-form apoE4 associates more efficiently than apoE3. J Clin Invest94:860–869.
Scheff SW, Sparks DL, Price DA (1996) Quantitative assessment ofsynaptic density in the outer molecular layer of the hippocampaldentate gyrus in Alzheimer’s disease. Dementia 7:226–232.
Schmechel D, Saunders A, Strittmatter W, Crain BJ, Hulette C, JooSH, Pericak-Vance M, Goldgaber D, Roses A (1993) Increasedamyloid beta-peptide deposition in cerebral cortex as a conse-quence of apolipoprotein E genotype in late-onset Alzheimer’s dis-ease. Proc Natl Acad Sci USA 90:9649–9653.
Sigurdsson EM, Scholtzova H, Mehta P, Frangione B, Wisniewski T(2001) Immunization with a nontoxic/nonfibrillar amyloid-� homol-ogous peptide reduces Alzheimer’s disease associated pathologyin transgenic mice. Am J Pathol 159:439–447.
Soto C, Golabek AA, Wisniewski T, Castano EM (1996) Alzheimer’ssoluble �-amyloid is conformationally modified by apolipoproteinsin vitro. Neuroreport 7:721–725.
Sun Y, Wu S, Bu G, Onifade MK, Patel SN, LaDu MJ, Fagan AM,Holtzman DM (1998) Glial fibrillary acidic protein-apolipoprotein E(apoE) transgenic mice: astrocyte-specific expression and differingbiological effects of astrocyte-secreted apoE3 and apoE4 lipopro-teins. J Neurosci 18:3261–3272.
Sze CI, Troncoso JC, Kawas CH, Mouton P, Price DL, Martin LJ(1997) Loss of the presynaptic vesicle protein synaptophysin inhippocampus correlates with cognitive decline in Alzheimer dis-ease. J Neuropathol Exp Neurol 56:933–944.
Tardiff B, Newman M, Saunders A, Strittmatter W, Blumenthal JA,White WD, Croughwell ND, Davis RD, Roses AD, Reves JG (1997)Preliminary report of a genetic basis for cognitive decline aftercardiac operations: the Neurological Outcome Research Group ofthe Duke Heart Center. Ann Thorac Surg 64:715–720.
Teasdale GM, Nicoll JAR, Murray G, Fiddes M (1997) Association of
Y. Ji et al. / Neuroscience 122 (2003) 305–315314

apolipoprotein E polymorphism with outcome after head injury.Lancet 350:1069–1071.
Terry RD (1994) Neuropathological changes in Alzheimer disease.Prog Brain Res 101:383–390.
Terry RD (1998) The cytoskeleton in Alzheimer’s disease. J NeuralTransm 53:141–145.
Terry RD, Masliah E, Salmon DP, Butters N, DeTeresa R, Hill R,Hansen LA, Katzman R (1991) Physical basis of cognitive alter-ations in Alzheimer’s disease: synapse loss is the major correlate ofcognitive impairment. Ann Neurol 30:572–580.
Teter B, Xu PT, Gilbert JR, Roses AD, Galasko D, Cole GM (1999)Human apolipoprotein E isoform-specific differences in neuronalsprouting in organotypic hippocampal slice culture. J Neurochem73:2613–2616.
The National Institute on Aging and The Reagan Institute WorkingGroup on Diagnositic Criteria for the Neuropathological Assess-ment of Alzheimer’s Disease (1997) Consensus recommendationsfor the postmortem diagnosis of Alzheimer’s disease. NeurobiolAging 18:S1–S2.
van Rossum D (1999) Cytoskeletal dynamics in dendritic spines: directmodulation by glutamate receptors? Trends Neurosci 22:290–295.
Wegiel J, Wisniewski HM, Kuchna I, Tarnawski M, Badmajew E,Popovitch E, Kulczycki J, Dowjat WK, Wisniewski T (1998) Cell-type specific enhancement of amyloid-� deposition in a novel pre-senilin-1 mutation (P117L). J Neuropathol Exp Neurol 57:831–838.
Wegiel J, Wisniewski T, Reisberg B, Silverman W (2003) Alzheimerdementia neuropathology. In: Dementia; presentations, differential
diagnosis and nosology (Emery OB, Oxman TE, eds), pp 89–120.Baltimore: Johns Hopkins University Press.
Wenham PR, Price WH, Blandell G (1991) Apolipoprotein E genotyp-ing by one-stage PCR. Lancet 337:1158–1159.
White F, Nicoll JAR, Roses A, Horsburgh K (2001) Impaired neuronalplasticity in transgenic mice expressing human apolipoprotein E4compared to E3 in a model of entorhinal cortex lesion. NeurobiolDis 8:611–625.
Wisniewski T, Castano EM, Golabek AA, Vogel T, Frangione B (1994)Acceleration of Alzheimer’s fibril formation by apolipoprotein E invitro. Am J Pathol 145:1030–1035.
Wisniewski T, Frangione B (1992) Apolipoprotein E: a pathologicalchaperone protein in patients with cerebral and systemic amyloid.Neurosci Lett 135:235–238.
Wisniewski T, Frangione B (1996) Apolipoprotein E, amyloidosis andAlzheimer’s disease. Dementia 10:171–183.
Wisniewski T, Golabek AA, Kida E, Wisniewski KE, Frangione B(1995) Conformational mimicry in Alzheimer’s disease; role of apo-lipoproteins in amyloidogenesis. Am J Pathol 147:238–244.
Yamagata K, Urakami K, Ikeda K, Ji Y, Adachi Y, Arai H, Sasaki H,Sato K, Nakashima K (2001) High expression of apolipoprotein EmRNA in the brains with sporadic Alzheimer’s disease. DementGeriatr Cogn Disord 12:57–62.
Zarrow C, Victoroff J (1998) Increased apolipoprotein E mRNA in thehippocampus in Alzheimer disease and in rats after entorhinalcortex lesioning. Exp Neurol 149:79–86.
(Accepted 6 August 2003)
Y. Ji et al. / Neuroscience 122 (2003) 305–315 315