an archaeobotanical investigation of plant use at pre-pottery neolithic chogha golan in southwestern...
TRANSCRIPT

Eberhard-Karls-Universität Tübingen
Mathematisch-Naturwissenschaftliche Fakultät
Fachbereich Geowissenschaften
Institut für Naturwissenschaftliche Archäologie
Archäobotanik
An archaeobotanical investigation of plant
use at Pre-Pottery Neolithic Chogha Golan
in southwestern Iran
Masterarbeit zur Erlangung des akademischen Grades
Master of Science (M.Sc.)
im Studiengang
M.Sc. Naturwissenschaftliche Archäologie - Paläoanthropologie
vorgelegt von
Doğa Karakaya B.A.
Tübingen
September 2013

Tag der Abgabe: 30.09.2013
Tag der mündlichen Prüfung: 18.10.2013
Erstbetreuer: PD Dr. Simone Riehl
Zweitbetreuerin: Prof. Dr. Nicholas J. Conard, Ph.D.

Eigenständigkeitserklärung
Hiermit versichere ich, dass ich vorliegende Arbeit selbständig verfasst und nur unter Ver-
wendung der angegebenen Hilfsmittel und Quellen angefertigt habe. Die Stellen meiner Ar-
beit, die dem Wortlaut oder dem Sinn nach anderen Werken entnommen sind, habe ich in
jedem Fall unter Angabe der Quelle als Entlehnung kenntlich gemacht. Dasselbe gilt sinnge-
mäß für Tabellen und Abbildungen.
Die eingereichte Arbeit ist nicht anderweitig als Prüfungsleistung verwendet oder in deutscher
bzw. einer anderen Sprache veröffentlicht worden.
Tübingen, den 30.09.2013

TABLE of CONTENTS
ABSTRACT ............................................................................................................................................................ I
ZUSAMMENFASSUNG ..................................................................................................................................... II
ACKNOWLEDGEMENTS ................................................................................................................................ III
I. INTRODUCTION ............................................................................................................................................. 1
I.1 RESEARCH QUESTIONS ................................................................................................................................. 3
II. ARCHAEOBOTANICAL EVIDENCE AND THEORETHICAL FRAMEWORK FOR THE
ORIGINS OF AGRICULTURE .......................................................................................................................... 5
II.1 CURRENT STATE OF ARCHAEOBOTANICAL EVIDENCE IN SOUTHWEST ASIA ............................................ 5
II.2. EXPLANATORY MODELS OF THE ORIGINS OF AGRICULTURE .................................................................. 10
III. ENVIRONMENT AND ARCHAEOLOGY OF CHOGHA GOLAN ..................................................... 16
III.1 GEOLOGICAL SETTINGS .......................................................................................................................... 16
III.2 PALAEOCLIMATIC SETTINGS AND VEGETATION HISTORY IN WESTERN IRAN ...................................... 17
III.3 ARCHAEOLOGICAL RESEARCH IN CHOGHA GOLAN .............................................................................. 20
III.3.1 Site Description and Excavations ...................................................................................................... 20
III.3.2 Chronology ........................................................................................................................................ 23
IV. MATERIALS AND METHODS ................................................................................................................. 25
IV.1 METHODOLOGICAL CONSIDERATIONS ON SAMPLE SIZE, TAPHONOMY AND QUANTITATIVE
MEASUREMENTS ............................................................................................................................................... 26
IV.2 FURTHER LIMITATIONS IN THE CURRENT RESEARCH ............................................................................ 30
V. RESULTS ....................................................................................................................................................... 31
V.1 THE COMPOSITION OF POACEAE (GRASS FAMILY) ................................................................................. 32
V.1.1 Large-seeded Poaceae remains .......................................................................................................... 32
V.1.2 Small-seeded Poaceae remains ........................................................................................................... 36
V.2 THE COMPOSITION OF FABACEAE (PULSE FAMILY) ............................................................................... 37
V.2.1 Large-seeded Fabaceae remains ........................................................................................................ 38
V.2.2 Small-seeded Fabaceae remains ......................................................................................................... 39
V.3 THE COMPOSITION OF OTHER PLANT FAMILIES .................................................................................... 41
V.3.1 Anacardiceae (Sumac Family) ............................................................................................................ 41
V.3.2 Brassicaceae (Mustard Family) .......................................................................................................... 42
V.3.3 Caryophyllaceae (Pink Family) .......................................................................................................... 42
V.3.4 Malvaceae (Mallow Family) ............................................................................................................... 43
V.3.5 Chenopodiaceae/Amaranthaceae (Goosefoot/Amaranth Family) ...................................................... 43

V.3.6 Cyperaceae (Sedge Family) ................................................................................................................ 44
V.3.7 Boraginaceae (Borage Family)........................................................................................................... 44
V.3.8 Asteraceae (Sunflower Family) ........................................................................................................... 44
V.3.9 Asparagaceae ...................................................................................................................................... 44
V.3.10 Rubiaceae (Bedstraw Family) ........................................................................................................... 45
V.3.11 Papaveraceae (Poppy Family) .......................................................................................................... 45
V.4. GENERAL PATTERNS IN THE CHOGHA GOLAN ASSEMBLAGE ................................................................ 45
VI. DISCUSSION ................................................................................................................................................ 50
VI.1 VARIATIONS IN THE CHOGHA GOLAN PLANT ASSEMBLAGE .................................................................. 50
VI.2 EVALUATIONS OF PLANT MANAGEMENT STRATEGIES IN EASTERN FERTILE CRESCENT ..................... 54
VI.3.1 Wild plant-food procurement ............................................................................................................. 59
VI.2.2 Wild plant-food production ................................................................................................................ 64
VI.2.3 The prevalence of domesticated crop plants ...................................................................................... 75
VII. CONCLUDING REMARKS ...................................................................................................................... 80
VII.1 FUTURE RESEARCH AT CHOGHA GOLAN .............................................................................................. 81
BIBLIOGRAPHY ............................................................................................................................................... 83
APPENDIX 1: INVENTORY OF IDENTIFIED TAXA ................................................................................. 96
APPENDIX 2: PLATES ................................................................................................................................... 115
APPENDIX 3: MACROBOTANICAL RAW DATA .................................................................................... 123

TABLE of ILLUSTRATIONS
Figures
Fig. 1: The geographical centers of plant domestication around the world (Balter 2007).
Fig. 2: The comparative data from the eastern Fertile Crescent shown the abundance scores of
major crops, and small-seeded taxa and three nuts; modified from Charles (2007).
Fig. 3: The pollen record of Lake Zeribar (Stevens et al. 2001).
Fig. 4: Map showing the location of Chogha Golan and other PPN sites in Central Zagros
Mountains, Western Iran (Zeidi et al, 2012).
Fig. 5: The stratigraphy of Chogha Golan excavation (Zeidi pers. comm.. 2013) with
calibrated AMS dates published in Riehl et al. (2013).
Fig. 6: The chronological sequence in southwest Asia correlated with southern Levantine
sequence (Zeder 2011).
Graphs
Graph 1: The floral composition of the Chogha Golan assemblage in percentages.
Graph 2: Relative percentages of Poaceae chaff and seed remains in the plant assemblage.
Graph 3: The changes in percentages of wild barley grains and chaff remains in the floral
composition.
Graph 4: The changes in composition of small- versus large-seeded Fabaceae remains in the
plant assemblage.
Graph 5: The contribution of larger-grained pulses in the assemblage in percentages.
Graph 6: The find density analysis for the large-grained pulses.
Graph 7: The absolute percentages of small-seeded pulses throughout the occupation period.
Graph 8: The density of small-seeded pulses per one liter of soil throughout the occupation
period
Graph 9: The composition of other plant families in the assemblage
Graph 10: Correspondence Analysis plot for the compositional variations of analyzed
samples.
Graph 11: Correspondence analysis plot for the co-variational relationship between samples
and species.

I
ABSTRACT
Chogha Golan is a Pre-Pottery Neolithic tell site in Ilam province, Southwestern Iran. The site
is located on the hilly flanks of the Central Zagros Mountains, which is within the natural
distribution range of wild relatives of crop plants. In comparison to other regions of the Fertile
Crescent only few sites have been investigated in this area.
Excavations of the Tübingen Iranian Stone Age Research Project (TISARP) in 2009 and 2010
documented 11 distinct archaeological strata in the 8m of anthropogenic deposits. The
excavations focused on the systematic recovery of botanical remains by floating the majority
of all the sediments excavated. This led to the recovery of the richest record of botanical
remains available for the Pre-Pottery Neolithic in the Fertile Crescent. AMS radiocarbon
dating demonstrates that the occupation of the site lasted about 2.500 years from 11.500 to
9.000 cal BP.
The current research aims to document the most important archaeobotanical finds from
Chogha Golan throughout the occupation sequence, and provides key insights into
development of plant subsistence in the foothills of the Zagros from the end of the Younger
Dryas into the early Holocene.

II
ZUSAMMENFASSUNG
Chogha Golan ist ein Tell des präkeramischen Neolithikums in der Ilam Provinz in Südwest
Iran. Der Tell liegt an den hügeligen Flanken des zentralen Zagros Gebirges, welches sich
innerhalb der natürlichen Verbreitungsgrenze der wilden Verwandten der heutigen
Nutzpflanzen befindet. Im Vergleich mit anderen Regionen des Fruchtbaren Halbmondes sind
bis jetzt nur wenige Standorte im Zagros Gebirge untersucht worden.
Ausgrabungen des Tübingen Iran Steinzeit Forschungsprojektes (TISARP) in 2009 und 2010
dokumentierten 11 distinkte archäologische Strata in 8m anthropogenen Ablagerungen. Die
Ausgrabungen fokussierten sich auf die systematische Bergung botanischer Überresten,
indem ein Großteil der ausgegrabenen Sedimente flotiert wurde. Dadurch wurde einer der
reichsten archäobotanischen Datensätze des präkeramischen Neolithikums innerhalb des des
Fruchtbaren Halbmondes geschaffen. AMS Radiokarbondatierung zeigten das die
Besiedlungsgeschichte von Cogha Golan ungefähr 2500 Jahre, von 11000 bis 9000 v.Chr.,
andauerte.
Ziel der vorliegenden Forschungsarbeit ist die Dokumentation der archäobotanischen Funde
von Chogha Golan aus der Besiedlungszeit und Schlüsseleinblicke in die Entwicklung der
Pflanzen Subsistenz an den Ausläufern des Zagros Gebirges vom Ende der Jüngeren
Dryaszeit bis in das frühe Holozän.

III
ACKNOWLEDGEMENTS
This master thesis was written with the help of a number of people.
Foremost among them is my first adviser PD Dr. Simone Riehl. Her invaluable support and
guidance started with giving the idea about working on Chogha Golan plant materials and
continued in every step of research. I would not be at the end of this process without her
assistance, advice and support and encouragement.
I also thank to Prof Dr. Nicholas Conard for accepting to be my second adviser and also to
constitute this master program in English language. This program expanded my horizon,
changed my views to past cultures through giving me perspective of an archaeologist.
Mohsen Zeidi, the head of Chogha Golan excavations, was a strong supporter during the
process of understanding Chogha Golan by endlessly giving information about the
excavations and archaeology. I personally gained alot from his perspective of approaching the
archaeology through investigating every aspects in a very detailed manner.
A number of other people helped me alot during this period. I am especially grateful first of
all to Dr. Canan Çakırlar who first informed me about the presence of this program and later
helped me to settle down in Tübingen. I would like to thank to my rewiever Bethany
Mendenhall because she spent a substantial amount of time to correct and correct my mistakes
in English language with patience and for her helpful comments to look the thesis from a
different perspective. I am grateful to have such friends like Özgür Çizer, Hakan Mutlu,
Ahmet Aytek and Andrea Orendi who assisted me during the breaks and shared laughs among
intense working hours. Additionally, I would like to thank to my dear friend Maxi Herberich
who kindly provided me companion with her positive energy.
Lastly, I have to mention the support of my family. They were so kind not to say anything
about my prolonged studies and supported me in every conditions. Their endless love and
support was the principal reason to pursue master degree in Germany. I strongly believe that
science is a team job and this master thesis would not have come to an end without the help
and support of this people and of course all of my professors and lecturers.

1
I. INTRODUCTION
The origins of agriculture and its subsequent dispersal in Southwest Asia have been one of the
most intensively discussed topics in archaeological research. Today, scientific studies on the
emergence and dispersal of agriculture embrace a large part of the archaeobotanical literature.
From the beginning of the last century on archaeological research recognized a transitional
period between the Paleolithic and Bronze Ages. An emergent body of archaeological data on
the increasing amount of finds of agricultural implements, pottery sherds and changes in lithic
industry indicated the necessity of establishing a historical and theoretical framework (Moore
1985). The concept of “Neolithic” was the answer to this intellectual search. The period‟s
distinctive character is a set of socio-cultural and economic transformations initiated through
the changes from mobile hunting-gathering to a sedentary way of living that depends on
agriculture and stockbreeding.
The first two decades after World War II witnessed an escalating amount of archaeological
research following two important developments for our current understanding of the Neolithic
period. One was the introduction of radiocarbon dating, which allowed researchers to
establish an absolute chronology for the Neolithic period. This new method of dating reduced
the need to rely on seriation and cross-cultural trait distributions to construct chronologies
(Trigger 2006). This in turn produced the need to learn more about economic and cultural
changes during this transitional stage and entailed the investigation of plant and animal
remains. This objective required multi-disciplinary research and the cooperation of specialists
from the natural sciences to gather the information on the faunal and floral composition of
paleoenvironments. Of significant importance to the archaeology of the Neolithic period, the
excavation at Jarmo led by Robert Braidwood was the first attempt at a multi-disciplinary
approach that has become standard procedure ever since (Moore 1985).
Almost simultaneously, Kathleen Kenyon at Jericho and Robert Braidwood at Jarmo
discovered a “Pre-Pottery Neolithic” (PPN) phase that lacked pottery remains but indicated a
sedentary and semi-sedentary lifestyle. Kenyon divided this period into two subphases “A”
and “B” to identify successive cultures at Jericho. This chronological classification later

2
became a standard period term in Near Eastern archaeology for the timeframe between the
Paleolithic and Pottery Neolithic (Moore 1985; Sheratt 2007; Watkins 2008).
The introduction of water floatation methods into archaeological fieldwork for effectively
retrieving charred plant remains was another important development in the 1960s. This new
method was first applied to Near Eastern archaeology by Helbaek at Tepe Ali Kosh, Iran
(Helbaek 1969; Fuller and Colledge 2008). Floatation quickly became widespread during the
1970s and 1980s on excavations in the Near East. Following the introduction of on-site
recovery methods such as floatation, archaeobotanical studies develop into a distinct
discipline within archaeological research (Fuller and Colledge 2008) and extensively
contributed to our understanding of the origins of agriculture in Southwest Asia. The potential
of archaeobotanical investigation has proven to be valuable in detecting the changes in
subsistence economy and dietary practices in the prehistoric past.
It should be noted that the archaeological studies that directly aim to comprehend the overall
changes in the PPN period are not evenly distributed in Southwest Asia. Western Iran (also
known as central Zagros) is relatively less investigated in comparison to the numerous
research projects in the Levant, southeastern Turkey, and northern Syria. Zeidi et al. (2012)
note that the archaeological evidence for PPN occupation in western Iran is limited to the
published materials of Tepe Asiab and Sarab, Ganj Dareh (Smith 1978), Tepe Guran, Tepe
Abdul Hosein (Pullar 1990), and Tepe Ali Kosh (Hole et al. 1969). In addition to these early
sites, two newly excavated PPN sites in the central Zagros (Sheikh-e Abad and Jani) will soon
contribute to the archaeological knowledge of this region (Matthews et al. 2010).
Moreover, there are even fewer reliable and representative records for Pre-Pottery Neolithic
plant remains in western Iran. Few archaeobotanical researches in western Iran leave a large
chronological gap between the early PPNA and late PPNB occupations in eastern Fertile
Crescent. The evidence for Late Epipaleolithic and PPNA are mainly coming from a number
of sites at the northern ranges of Zagros Mountains, Hallan Çemi, Demirköy, Qermez Dere,
M‟lefaat (Savard et al. 2003, 2006) while there is no contemporary sites in central Zagros for
this timeframe. The archaeobotanical information on PPNB occupation in central Zagros is
mostly derived from late PPN sites such as Jarmo, Tepe Ali Kosh (Helbaek 1969), Tell
Magzhaliyeh, Chogha Bonut, Tepe Abdul Hossein (Hubbard 1990) that mainly represent the
plant assemblages already predominated by domesticated plants. Ganj Dareh among others

3
stands forward as a middle PPNB occupation while it bears mainly short-habited occupation
with an ambivalent evidence of plant domestication. Riehl et al. (2012, 2013) recently
published two comprehensive accounts on the results of archaeobotanical investigations at
Chogha Golan that bring into light valuable information for the simultaneous developments in
eastern Fertile Crescent. All in all, the overall picture of the establishment of the farming
economy is hampered due to the lack of sites that are inhabited for a long chronological
sequence.
The rarity of published archaeological and archaeobotanical records also obscures the overall
understanding of the evidence on the transition to a farming economy in western Iran.
Therefore, Smith (1971) indicates that the theoretical models tend to consider the origins and
dispersal of agriculture from the perspective of expansion from a single center and to
underestimate the role and probable contribution of the central Zagros region to plant and
animal domestication.
I.1 Research Questions
In the research that is the subject of this paper, analytical results from the examination of
archaeobotanical samples of a Pre-Pottery Neolithic site located in the central Zagros region,
Chogha Golan, will be described to detect any changing patterns in the subsistence economy
that would indicate an evolution towards the establishment of an agricultural food economy.
In this regard, two research questions were formulated in conjunction with the overall
research goals of the Chogha Golan project as defined by archaeologists:
1) Are there any variations in the composition of the archaeobotanical plant assemblage
throughout the occupation period?
2) How could the timing of the appearance of different plant species be related to the
development of cultivation and domestication in western Iran, in comparison to other regions
of the Fertile Crescent?
Considering the few archaeobotanical investigations of the PPN period in western Iran, this
research aims to illustrate the overall developments in the subsistence economy of Chogha

4
Golan habitation following the contributions of Riehl et al. (2012, 2013) about this particular
site.
In this respect, it documents the most important archaeobotanical finds from Chogha Golan
throughout the occupation sequence and provides key insights into the development of plant
subsistence in the foothills of the Zagros from the end of the Younger Dryas into the early
Holocene. In addition, the current state of knowledge and different explanatory models for the
origins of agriculture will be reviewed to further develop background information and to
explore the developmental route to the establishment towards the farming economy during the
PPN period.

5
II. ARCHAEOBOTANICAL EVIDENCE AND THEORETHICAL FRAMEWORK
FOR THE ORIGINS OF AGRICULTURE
II.1 Current state of archaeobotanical evidence in southwest Asia
The early pioneering efforts of de Candolle, Vavilov, and Harlan represent the cornerstone
research for our present understanding of the geographical centers of plant domestication
around the world (Abbo et al. 2010). Today, it is certain that the cultivation and
domestication1 of plants emerged independently in different regions (Figure 1) and that
chronologically this phenomenon first appeared in southwest Asia, involving a number of
wild species of grasses2, pulses, and flax at about 10.000 B.P. (Salamini et al. 2002, Diamond
2002).
The contemporary geographical distributions of wild relatives of the first crop plants intersect
in a region called the Fertile Crescent, which covers the Taurus-Zagros mountain range and
the Levant. Eight plants are considered to be those first cultivated and domesticated. This
assemblage of “founder crops”3 comprises wild progenitors of einkorn (Triticum monococcum
1 Nesbitt (2002) draws attention to the need to define concepts such as cultivation, domestication and
agriculture/farming more concretely. The term cultivation, in this text will designate “the sowing and harvesting
of wild plants in tilled soil”. The concept of domestication will be used to characterize “[…] the process in which
humans take control of the reproduction of plants and animals, and consciously or unconsciously select for
attributes favourable to human use. For cereals control of reproduction means repeated sowing and harvesting of
the same population, and the key attribute selected for is loss of the ability to disseminate seed without human
intervention”. Finally, agriculture/farming “involves the cultivation of domesticated crop plants” (Nesbitt 2002).
2 Wheat genus (Triticum) is divided into six biological species at three ploidy levels. Diploid wheats consist of T.
monococcum (genomic composition is Am
Am) and T. urartu (AA). Tetraploid wheats are T. turgidum (AABB)
and T. timopheveii (AAGG). Hexaploid wheats consists of T. aestivum (AABBDD) and T. zhukovskyi
(AAAm
Am
GG). Genomic relationship exhibits that T. monococcum, T. timopheveii and T. zhukovskyi form a
separate lineage which is not related to the principal wheat lineage, formed by T. urartu, T. turgidum, T.
aestivum (Dvorak et al. 2012).
3 Recently, some authors (Fuller et al. 2012; Asouti and Fuller 2011) presumed that more plants might have been
cultivated by early Holocene communities. Ten additional crop plants were proposed to contribute to early
subsistence of human communities together with the eight abovementioned species. These “lost” crops include

6
ssp. boeticum), emmer (Triticum turgidum ssp. dicoccoides), barley (Hordeum spontaneum),
lentil (Lens orientalis), pea (Pisum humile), chickpea (Cicer reticulatum), bitter vetch (Vicia
ervilia), and flax (Linum bienne). All these plants exhibit almost similar biological
characteristics, in that all of them are diploid (except emmer is tetraploid), annual, self-
pollinated plants. Moreover, all of these eight species are interfertile within each crop and
between the crop and its wild progenitors (Zohary and Hopf 2000; Weiss and Zohary 2011).
Figure 1: The geographical centers of plant domestication around the world (Balter 2007).
In this same respect, Zohary et al. (1969; after Flannery 1973) estimates that harvests of up to
500 to 800 kilos of grain could be gathered per hectare from wild stands of emmer wheat
today in Mount Hermon, Israel. At the same time, complementary food sources like
leguminous plants and hunted animals that were rich in protein content were also native and
abundantly found in this particular region (Miller 1984; Diamond 2002; Kislev and Bar-Yosef
1988; Harlan and Zohary 1966).
Archaeobotanical data from the Upper Paleolithic site Ohalo II, in the Levant suggests that
two important crop progenitors, wild barley and emmer were being used by hunter-gatherer
the two grained forms of Triticum monococcum or T. urartu (einkorn wheat), Secale sp. (rye), Triticum
turgidum/timopheevi (striate emmeroid tetraploid wheat), Avena sterilis (oat), Vicia faba (broad bean), Lathyrus
sativus (grass pea), Lens nigricans (black wild lentil), and Ficus carica (common fig). This issue is particularly
important to understand divergent PPN strategies for food procurement in the prehistoric record.

7
communities as early as 21.000 cal. B.P. Exceptional preservation at this particular site
exposed a rich floral assemblage with plenty of food plants as well as the ground stone
assemblage associated with food-processing activities (Kislev et al. 1989). Piperno et al.
(2004) argue that the majority of the starch grains sampled on a grinding slab indicate the
processing mainly of wild barley species.
Another aspect for the significance of the Fertile Crescent is that the climatic warming at the
onset of the Holocene coincides with the emergence of several new settlements in this region.
Unlike the scarce information on plant remains from Middle Palaeolithic (Lev et al. 2005) and
Upper Paleolithic (Kislev et al. 1989; Hillman 2000, 2001) sites, comparatively more floral
remains were recovered from PPN localities. The archaeological remains of the wild
progenitors of crop plants that were correlated in radiocarbon age and stratigraphy occur
frequently over the region (Salamini et al. 2002; Charles 2007).
The morphological analysis of seeds and chaffs shows equivalent evidence of plant
domestication during the PPNA (Nesbitt 2002). Nevertheless, many scholars agree that cereal
and pulse cultivation might have been practiced before the morphological domestication of
crop plants throughout the PPNA (Willcox 2004, 2008, 2012) or even to some extent much
earlier during the Younger Dryas climatic degradation (Hillman 2000, 2001). This
assumption, known as “predomestication cultivation”, depends on the frequent appearance of
certain plants in archaeobotanical assemblages, “arable weeds”, which today thrive only on
disturbed lands such as rocky slopes, roadsides, abandoned gardens, and tilled fields (Hillman
2000, 2001; Colledge 2002; Willcox 2012).
The key event in the domestication of cereals and pulses was the elimination of the natural
dispersal mechanism4 through human-induced selective pressures on cultivated wild plants,
mostly referred to as “domestication syndrome” in archaeobotanical literature (Nesbitt 2002).
Wild wheats and barley seeds are enclosed by thick and though glumes and located on top of
4 The adaptive traits modified through human intervention to the reproductive cycles of crop plants are classified
into two categories. The major qualitative traits in domesticated cereals comprise rachis brittleness, glume
tenacity, and free-threshing state, which together result in the elimination of the natural seed dispersal
mechanism in domesticated cereals. The additional traits, which are quantitatively inherited, are seed size, grain
yield, plant height, grain hardness, tillering capacity, seed dormancy, developmental timing, and heading date
(Peng et al. 2011; Matsuoka 2011).

8
a stalk (spike or ear) that spontaneously disarticulates between each spikelet at maturity in
order to disseminate in the natural environment. Through a single gene mutation at two major
loci controlling rachis brittleness in emmer, the ears of domesticated cereals fail to disperse
and remain intact until being harvested by humans (Peng et al. 2011; Salamini et al. 2002;
Matsuoka 2011; Nesbitt and Samuel 1996). Both wild and domesticated emmer, einkorn and
barley have though glumes and hulled seeds. The though rachis of a domesticated plant can
only be broken by a mechanical force like threshing that eventually leaves a jagged scar at the
base of each spikelet which can be detected archaeobotanically. The first attributes that are
essential for pulse domestication were also the reduction of natural dispersal mechanism (pod
dehiscent) and seed dormancy (Sonnante et al 2009, Weiss and Zohary 2011; Hillman, 1984).
Seed size enlargement of early grain crops is another criterion which is readily visible in
archaeobotanical records. This adaptive trait is considered as a “response that lead to
successful germination with increased soil disturbance and depth of burial” (Purugganan and
Fuller 2009; Fuller 2007). Further genetic modifications in cereal domestication include
glume tenacity and free-threshing state that were central on the emergence of novel
phenotypes such as Triticum aestivum (bread wheat) and Triticum durum (hard wheat). These
two species are better adapted to agricultural production. In comparison to the tetraploid
wheat, free-threshing phenotypes have broader ecological adaptations to different photoperiod
and vernalization requirements such as improved tolerance to salt, low PH, aluminium and
frost (Dubcovsky and Dvorak 2007). All in all, it is noteworthy to mention that no significant
further changes had happened in ear shattering and in average size for barley and wheats
during the past 8.000 years. This suggests that a key domestication period must have occurred
before this temporal frame in southwest Asia (Fuller et al. 2012b).
Meanwhile, molecular studies in the last decade greatly contributed to the understanding of
the origins of agriculture by investigating two central issues, namely how often and how fast
the first crop plants became morphologically domesticated One aspect of these recent research
focuses on the geographical origins of domestication in Southwest Asia, in an attempt to learn
whether the domesticated species have monophyletic or polyphyletic origins. The principal
method that molecular studies employ is to measure the genetic distance between
contemporary populations of wild relatives and domesticated crops. The molecular studies in
the first half of the last decade tended to support monophyletic origination from a single
localized area through a single domestication event. But in contrast to these early works, the

9
accumulation of more research estimating the genetic distance among cultivated and wild
accessions of cereals indicates polyphyletic origins for domestication that happened
independently over a wide area in more than one location in the Fertile Crescent (Weiss and
Zohary 2011)
How fast the adaptive traits of domestication might become fixed was also intensely debated
in archaeobotany and plant genetics during the last decade. In the early 1990s, Hillman and
Davies (1990) argued that cereal domestication might have been a rapid process that took no
more than 200 years. However, some authors argue that wild cereals could have been
cultivated for over one millennium without leading the fixation of non-shattering phenotypes
in the environment. The recognition of a time delay for the appearance of non-shattering
phenotypes in archaeobotanical records resulted in refining this model and replacing it with
one that postulates a prolonged process of domestication (Tanno and Willcox 2006). This new
paradigm reconsidered the scientific knowledge on the fixation of domestication traits to
propose a protracted model of domestication extending the timeframe by as long as 2000
years (Fuller et al. 2012).
The first unequivocal archaeobotanical evidence for cereal and pulse domestication signals to
the PPNB period as the beginning of the appearance of domesticated phenotypes in the plant
assemblages (Nesbitt 2002). There is a stepwise increase of domesticated types of emmer,
einkorn, and barley from early the PPNB on (Fuller 2007). Settlements in southeastern
Turkey, Cafer Höyük, and Çayönü, yielded the earliest definitive domesticated einkorn and
emmer; in addition, a large stock of possibly domesticated lentil was retrieved from Yiftah‟el
in the middle PPNB levels (Weiss and Zohary 2011). The middle PPNB is also characterized
by the first appearance of novel species such as bread wheat and hard wheat (Dvorak 2012;
Nesbitt 2002; Asouti and Fuller 2011). The late PPNB plant assemblage indicates widespread
occurrences of domesticated founder crops all over the Fertile Crescent (Asouti and Fuller
2012).
The developmental route to farming economy in eastern Fertile Crescent is less understood
due to the rarity of archaeobotanical research (Charles 2007; Nesbitt 2002). The data indicates
that PPNA sites at the northern ranges of Zagros display few crop progenitors and no
domesticated crops while much later PPN sites in central Zagros demonstrates that the
domesticated plants started to be represented in the plant assemblages from the end of
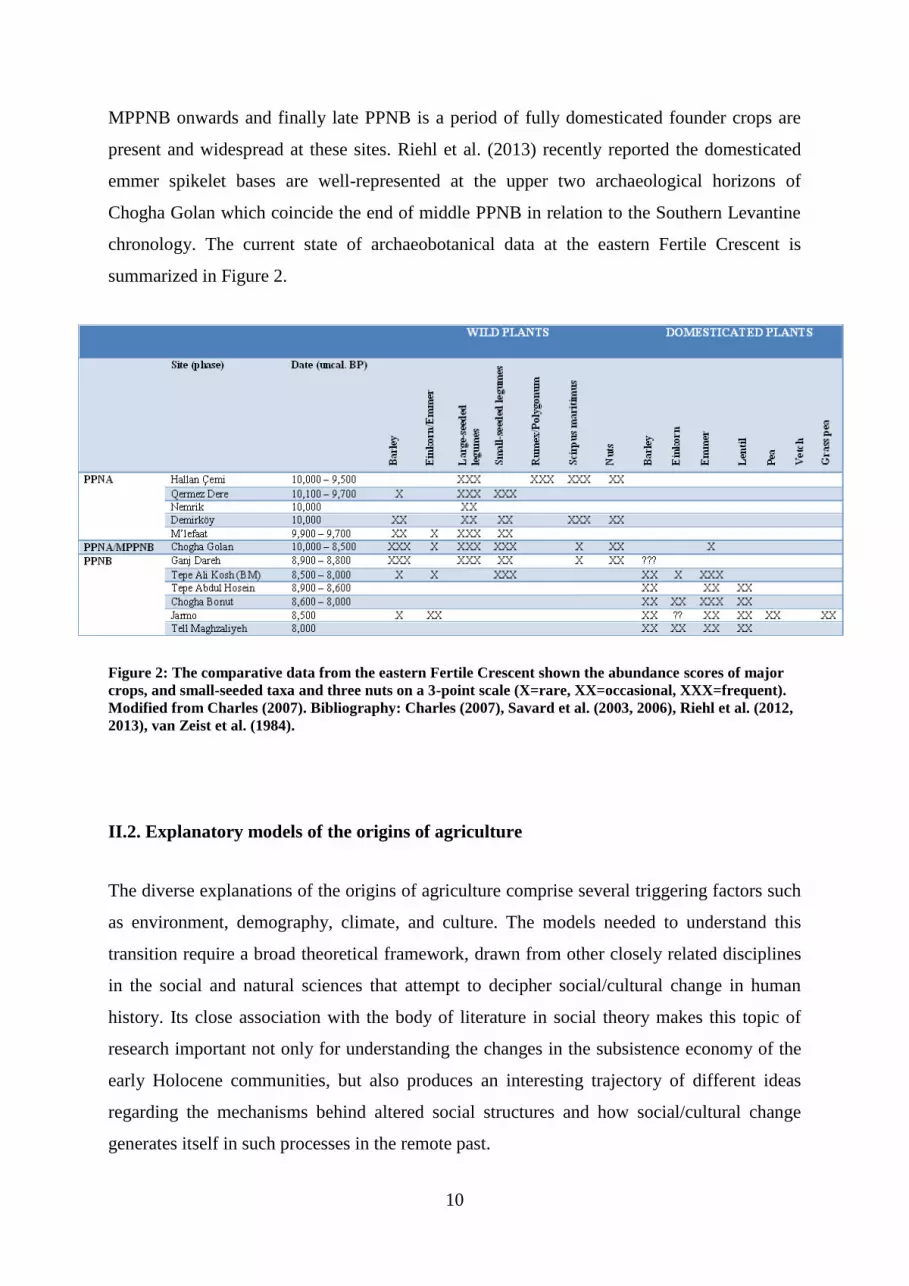
10
MPPNB onwards and finally late PPNB is a period of fully domesticated founder crops are
present and widespread at these sites. Riehl et al. (2013) recently reported the domesticated
emmer spikelet bases are well-represented at the upper two archaeological horizons of
Chogha Golan which coincide the end of middle PPNB in relation to the Southern Levantine
chronology. The current state of archaeobotanical data at the eastern Fertile Crescent is
summarized in Figure 2.
Figure 2: The comparative data from the eastern Fertile Crescent shown the abundance scores of major
crops, and small-seeded taxa and three nuts on a 3-point scale (X=rare, XX=occasional, XXX=frequent).
Modified from Charles (2007). Bibliography: Charles (2007), Savard et al. (2003, 2006), Riehl et al. (2012,
2013), van Zeist et al. (1984).
II.2. Explanatory models of the origins of agriculture
The diverse explanations of the origins of agriculture comprise several triggering factors such
as environment, demography, climate, and culture. The models needed to understand this
transition require a broad theoretical framework, drawn from other closely related disciplines
in the social and natural sciences that attempt to decipher social/cultural change in human
history. Its close association with the body of literature in social theory makes this topic of
research important not only for understanding the changes in the subsistence economy of the
early Holocene communities, but also produces an interesting trajectory of different ideas
regarding the mechanisms behind altered social structures and how social/cultural change
generates itself in such processes in the remote past.

11
Before describing the theoretical framework for this issue, it should be noted that a more
comprehensive approach to the study of prehistoric research had earlier been developed by
Childe, with his systematic application of the concept of “archaeological cultures”. This
conceptual formulation was based on defining every culture in terms of the constituent
artifacts recovered from archaeological excavations and then establishing spatiotemporal
limits empirically by means of stratigraphy (Trigger 2006). Childe framed his concept of
archaeological cultures as follows:
“We find certain types of remains – pots, implements, ornaments, burial rites, house forms
– constantly recurring together. Such a complex of associated traits we shall call a „cultural
group‟ or just a „culture‟. We assume that such a complex is the material expression of what
today we would call „a people‟” (Childe 1929, after Trigger 2006; Watkins 2008).
Moreover, this approach allowed Childe to combine the vocabulary of an emerging
archaeological taxonomy with the ethnographic classification of the anthropologist Henri
Lewis Morgan, “where pottery and polished stone axes signified the emancipation of
„Barbarism‟ from „Savagery‟” (Sheratt 2007, Childe 1958).
Starting in the second half of the last century, a number of archaeologists offered theories
principally to explain how and why agriculture emerged mainly in Southwest Asia. Smith
(2007) infers that there are “two largely disconnected scales –at the level of individual plant
and animal species to document the „what, when, and where‟ of domestication worldwide,
and at a regional or larger scale, to identify the causal „macro‟ variables (such as climate
change and population growth) that may account for „why‟ human societies first domesticated
target species”. These theoretical models for searching for macro-variables in the
domestication and the origins of agriculture can be grouped within certain broad topics such
as environmental change, demographically induced resource pressure, and changes in social
organization and ideology (Zeder 2006).
The role of environmental change or degradation in the origins of agriculture drew scholarly
attention at a very early stage in research history. Childe recognized the significance of the
Neolithic period as one of revolutionary change in the subsistence economy from food-
procurement to food production, defining it “a universal historical stage in the progress
towards modern civilization”; he was also the first to explore in detail this particular
phenomenon in the context of environmental degradation. He proposed that climatic change

12
to desiccated conditions after the Pleistocene would have led to the concentration of humans,
animals, and plants in close vicinity to water resources. This new environmental situation
promoted a degree of interaction among humans and animals and in the long term caused the
domestication of animals and plants (Childe 1951).
Childe pointed out a belt from the Atlantic to the Tigris River as the probable geographical
center of this consequential interaction. Nevertheless, it should be noted that at the time
Childe offered this model, there was reasonably good evidence for climate change at the end
of the Pleistocene in Europe, but no comparative data was present from Southwest Asia
(Bender 1975, Childe 1951).
The recent advances in paleoenvironmental reconstruction have resulted in reconsidering
climatic change as a causal factor (Zeder 2006). The well-documented Younger Dryas
climatic amelioration between around 10.700 and 9.700 cal. B.C. has been featured as having
had a role in the emergence of agriculture in Southwest Asia. Some scholars (Hillman et al.
2001, Bar-Yosef 1998) argue that the domestication of plants and animals was a response to
degraded environmental conditions and the need to maintain the already-established
subsistence economy that depended on wild grasses, legumes and hunted animals before this
particular climatic event.
The models of climatically driven environmental change as a causal factor were largely out of
favor between the 1960s and 1980s. Braidwood‟s multidisciplinary approach in the 1950s
allowed him to search for evidence of desiccation following Childe‟s propositions. However,
no major climatic change event had been detected in his field investigations in Iraq. Thus,
Braidwood offered a cultural model indicating that “farming was seen as the culmination of
ever-increasing cultural differentiation, specialization and knowledge of habitat” (Bender
1975, Braidwood 1969).
The theory of culture as an adaptive system in which the artifacts and modes of social
organization are seen as responses to changing physical and behavioral environments echoed
through the systemic approach (more specifically General Systems Theory) in archaeology.
According to this new paradigm, “culture comprises a series of interacting or articulated parts
(sub-systems) which include the effective environment – that perceived and used by human
groups under discussion – economic activities, technology, social organization, and religious

13
beliefs, all of which act like a rubber bands – pull one and all the others respond” (Bender
1975). This conceptualization of culture was important in signifying an irrevocable shift from
Childe‟s concept of archaeological cultures.
The incorporation of the environment into cultural processes was further enhanced through
ecosystem models (another interpretation of the systemic approach) of which the concept of
“broad-spectrum” economy proposed by Flannery drew considerable interest among scholars.
This mainly presupposes an intimate knowledge (pre-adaptations) of available plants and
animals, and of a variety of ecological niches to which plants can be transplanted and where
the animals can be hunted. This knowledge had already been developed by Upper Paleolithic
hunter-gatherers with well-defined seasonal migratory patterns to exploit the food resources
(Flannery 1969).
The systemic approach has resonated in the writings of the selectionist school of Neo-
Darwinian evolutionary archaeology, but with a different perspective. Dunnell, for example,
opted for biological evolutionary theory to explain cultural as well as biological variability, by
arguing that “traditional cultural evolutionism has failed to internalize such key tenets of
biological evolutionism as random variation and natural selection” (Trigger 2006). Material
culture is being interpreted here as a direct expression of human behavioral variability,
providing the basic constituent traits of a human cultural phenotype (collections of human
behaviors practiced by spatially and temporally bounded groups of people) that could be used
to reconstruct cultural lineages (Zeder 2009).
Following the same approach, Rindos assumed that domestication can be understood as
mutualistic relations of varying degrees between different biological species such as humans-
plants or humans-animals. He does not recognize the adaptation of plants and animals to
human needs as being completely different in nature from the adaptation of human beings to
the needs of plants and animals (Trigger, 2006).
Demographically induced resource pressure has been the focus of some other scholars. The
best-known example of this explanatory model was proposed by Binford with his “Marginal
Zone Hypothesis”. Population density (population packing) in his model is associated with the
carrying capacity of the area concerned. Accordingly, agricultural origins were the result of
resource pressures in an optimal area where the population increases over the carrying

14
capacity of the environment. This pressure would have been relieved through the constant
emigration of certain individuals and groups to neighboring, less favorable environments
where less preferable food resources were extensively used. The population increase in these
marginal environments eventually forced the domestication of plants and animals (Binford
1968). Another demographic stress approach that has been formulated is Cohen‟s food crisis
model, which assumes that a global food crisis following the population growth (not only in
marginal environments) at the beginning of the Holocene forced people to abandon more
nutritious hunting and gathering strategies and obliged them to tend domesticated plants and
animals (Zeder 2006).
Hodder‟s role in developing a challenging paradigm to the basic premises of processual
archaeology should also be mentioned to understand the study of cultural change in the
origins of agriculture. According to his interpretation, “material culture is not merely a
reflection of ecological adaptation or socio-political organization but also an active element in
group relations that can be used to disguise as well as reflect social relations” (Trigger 2006).
His contextual approach is based upon the idea that archaeologists need to examine all
possible lines of evidence about a culture to comprehend the significance of each part of it in
the formation of the archaeological record. Hodder brings out the dangers inherent in an
interpretation of archaeological evidence that is isolated from its broader context (Trigger,
2006).
Regarding the models concerning the problem of origins to changes in social organization and
ideology, Hayden proposed a model that denies external triggering factors as playing any
causative role in the origins of agriculture. Hayden claims that in contrast to resource
pressures, agriculture might have developed within an intra- and inter-communal competition
system in which high-prestige food items would serve to gain social advantages for some
“aggrandizers” through competitive feasting. This model predicts that domesticates were
considered as not ubiquitous dietary stables for early communities, but rather rare and
desirable exchange items (Zeder 2006, Hayden 2001, 2003, 2009).
Bender (1975, 1978) also objects to looking for external prime movers in explanations of the
origins of agriculture. She argues that the emergence of agriculture cannot be understood as a
question of changes in techno-cultural complexity but instead is a consequence of changing
social relations related to the ''commitment'' to produce more for marriage, ceremonial

15
purposes, and trade alliances among kin groups. In this case, she mentions that the evidence
of skull cults and items in circulation such as obsidian and shells from Natufian levels in the
Levant suggests continuity of settlement and a degree of organization and authority. The
increasing demands on production in relation to descent groups, exchange networks,
ceremonial institutions, and positions of authority provided a basic impulse for the
intensification of food production (Bender 1975, 1978).
Another incentive within the same framework is that of Cauvin. He proposed that
domestication is a direct consequence of a conceptual shift in human perception that dictates
that humans hold a dominant position over nature. This, when codified in religious ideology,
has had profound and irreversible effects on how humans recognize themselves in relation to
nature, freeing human communities to manipulate and transform nature through symbolic
reconstructions. Hodder similarly emphasized the role of symbols as central for the
domestication of public and private spaces (Zeder 2006, Watkins 2008).
Recently, an attempt by Asouti and Fuller (2013) to produce a site-by-site contextual analysis
of archaeobotanical evidence shares the same intellectual heritage although the authors clearly
state that their “contextual” approach is purely methodological, unlike Hodder‟s contextual
archaeology. The authors aim “to reconstruct the site-specific practices associated with plant
production, consumption, storage, and disposal and to determine how such activities might
have related to other domains of social life.” Asouti and Fuller (2013) assume the central role
of communal food consumption as a means in the formation of the early PPN plant-based
subsistence economy in the interplay of such community interactions as the negotiation and
reproduction of social identities. They further assume a balanced understanding of the
transition from foraging to farming that requires multiscalar interpretations through
contextual, micro- and macro-evolutionary perspectives.

16
III. ENVIRONMENT AND ARCHAEOLOGY OF CHOGHA GOLAN
III.1 Geological settings
The formation of the Zagros Mountains and Mesopotamia was the result of a series of tectonic
movements that caused the more mobile central plateau of Iran to move closer to the stable
massif of Arabia in the late Pliocene. The land between these two heavier formations was
compressed and folded into parallel mountain ridges or anticlines. The center of this
compressed zone collapsed and subsided further and its parallel ridges became the irregular
plains of Mesopotamia, which continue to fill with alluvial deposits from the rivers that cross
the mountainous ranges. To the east, the Zagros Mountains run in a northwest-southeast
direction. This area is characterized by parallel ridges with deep intermountain valleys and
other lowlands formed by water courses (Zohary 1973, Flannery 1969, Hole et al. 1969)
Chogha Golan, in Ilam Province, is within the folded zone of the Zagros Mountains; where
the deposition was contributed by the Upper and Lower Fars Formation of the Miocene. On
top of the Upper Fars Formation there lie the Lower Bakhtiyari beds. The transition between
these beds is characterized by the presence of red chert pebbles in the sandstone (Zeidi et al.
2012). The youngest bed formation in the Folded Zagros Zone is the Bakhtiyari
Conglomeratic Formation. This formation includes pebbles of red and yellow chert in the red
sandstone. On top of the pebble beds, cobbles of Asmari Limestone and pieces of gypsum
along with the chert became abundant. This chert is presumed to have served as raw material
for the inhabitants of Chogha Golan (Zeidi et al., 2012).
According to Zeidi et al. (2012), another probable source of raw material might have been a
group of hill formations just to the east side of the Konjam Cham River, north of the town of
Meheran. They report that “[t]he source of sediments is the Aghajari Formation and the
overlying Bakhtiyari Conglomerate of the Zagros front ranges. Raw materials of various
qualities are available in the area as pebbles, cobbles and nodules of chert, as well as pieces of
sandstone and small amount of quartzite. The density and fine quality of these lithic resources
may have attracted people to this region throughout prehistory”.

17
III.2 Palaeoclimatic settings and vegetation history in Western Iran
The climatic conditions during the Quaternary were governed by pronounced oscillations
from interglacial to glacial and interstadial to stadial times, which are documented in oxygen
isotope values (δ18
O) of deep sea sediments and ice cores. The timing of climate change
according to the Milankovitch effects is correlated through the 100.000-, 40.000-, and 23.000-
year cycles that are caused by variations in the sun-earth geometry. This phenomenon is
assumed to reflect the changes in eccentricity, obliquity, and precession of the earth‟s orbit
(Wright 1993, Kehl 2009). On the other hand, millennial-scale oscillations in δ18
O, which are
called Daasgaard-Oeschger events, have been recorded with slow cooling phases at the
beginning of a stadial, followed by fast temperature rises at the start of an interstadial. The
proposed reasons behind these fluctuations are deviations in ocean surface currents, surges of
ice sheets, variations of sunspot activity, or instabilities in the atmospheric carbon-dioxide
(CO2) system (Kehl 2009).
The regional differences of vegetation history between the western and eastern Fertile
Crescent have been posited by Stevens et al. (2001). The palaeoenvironmental conditions
during the Late Glacial Maximum (LGM) have been documented in the pollen cores of Lake
Van and Lake Urmia in the eastern Fertile Crescent. The characteristic feature of Late
Pleistocene vegetation was dry steppe dominated by Artemisia and chenopods, which
represent the semi-desert vegetation of a cold and arid climate (Stevens, 2001; Kehl, 2009).
The impact of the Younger Dryas climatic oscillations following the LGM is not pronounced
in the pollen spectra of the Lake Zeribar region, unlike the dramatic vegetation change
recorded at other pollen sites in the western Fertile Crescent. In addition to this point, Stevens
et al. (2001) report certain anomalies such as a significant increase in δO18
values at the end of
Pleistocene. The maximum inferred salinity and low lake levels between 12.600 and 12.000
cal. B.P. have also been reported through the investigation of plant macrofossil record (Kehl,
2009).
The Early Holocene in Southwest Asia was characterized by a rapid rise in temperature and
consequently in sea and lake levels. Data show the pollens of oak, pistachio, and grasses
becoming more and more pronounced in the pollen assemblages. The vegetative sequence in
the eastern Fertile Crescent, comprising the five pollen sites in the Zagros-Taurus range,

18
shows that the chenopod-Artemisia assemblage was replaced by grasses (Figure 3). Apart
from that, oak and pistachio appear in low percentages, contrary to the western parts of the
Fertile Crescent. Oak percentages in pollen sites do not reach the modern value of 40 % until
the mid-Holocene (Stevens et al. 2001).
Figure 3: The pollen record of Lake Zeribar (Stevens et al. 2001).
Today, the climate in Iran has extreme continental conditions characterized by the contrast
between cold winters and hot, dry summers common to most of Southwest Asia. The climatic
conditions are primarily governed by the pressure systems of westerly cyclones, the Siberian
High, and the Southwest Monsoon (Stevens 2001; Kehl 2009). Winters in Iran are dominated
by cold, dry air coming from the Siberian High pressure systems, while moisture-bearing low
pressure cells from the Atlantic or the Mediterranean penetrate inland to bring most of the
precipitation that falls in the winter and spring (Stevens 2001). Most of the precipitation falls
from October to April, with an average of ~350 mm per year in all Iran (Kehl 2009).
Zohary‟s (1973) phytogeographical map shows the province of Ilam situated in the Irano-
Turanian plant region, more particularly in the western Irano-Turanian subregion, comprising
the Mesopotamian province, the Irano-Anatolian province, and the East Sharo-Arabian
province of the Saharo-Arabian plant region. The Irano-Anatolian province extends farther

19
west, including a part of the Central Anatolian Plateau, the Armenian, Kurdistanian, and
Zagrosian mountain ranges, the eastern and southern slopes of the Elburz Mountains, and,
farther east, most of Afghanistan. The Irano-Anatolian province is described as floristically
highly diversified (Zohary 1973).
The prominent vegetation zone in the vicinity of Ilam province is the Kurdo-Zagrosian
steppe-forest vegetation. The dominant arboreal elements of this zone are oaks (mainly
Quercus brantii and Q. persica), pistachios (Pistacia khinjuk and P. atlantica), and also to
some extent Cerasus, Crataegus, and Pyrus. This is mainly characterized as steppe-forest in
which the trees are fairly well spaced from each other. Interspaces are vegetated by steppic
elements (Zohary 1973). The ecological characteristics of this type of forest are tolerance for
low precipitation, tolerance for low temperatures, sensitivity to snow, positive response to
increased moisture during the growing season, and inability or limited ability to disperse
under present summer-dry conditions (El-Moslimany 1986).
Zohary‟s (1973) reasons for the inclusion of the Zagros Mountains in the Irano-Turanian
phytogeographical unit instead of in the Mediterranean region are summarized in the
following arguments. Firstly, in the deciduous forest such species as (Pistacia khinjuk, P.
atlantica var. latifolia and var. kurdica, (as well as some species of Amygdalus, Cerasus,
Prunus and Acer, etc.) have no relation to Mediterranean flora. Secondly, Zohary assumes
that Kurdo-Zagrosian flora include some species that might have originated in the Zagros and
migrated westwards to the Mediterrenean zone; these species are Quercus libani and Quercus
boissieri. Lastly, he notes that Iran should be considered as an evolutionary center of
speciation (e.g., the genus Pistacia).
The precipitation rates change within close proximities to where Chogha Golan is located due
to large altitudinal variations. The annual rainfall for the town of Mehran (altitude 500
meters), which is 30 km south of the site, is ~360 mm and the mean annual temperature is
recorded as 22.7 °C with a range of 35.0 in August and 9.4 °C in January. The climatic data
from 30 km southwest of the town of Ilam (altitude 1000 meters) registers an annual rainfall
of 430 mm and a mean annual temperature of 17.0 °C with a range of 29 °C in August and 3.4
°C in January (Nevo et al. 1986).

20
III.3 Archaeological research in Chogha Golan
III.3.1 Site Description and Excavations
Chogha Golan is a Pre-Pottery Neolithic tell site situated at the outskirts of the Central Zagros
Mountains on the Amirabad plain in Ilam Province. Chogha Golan lies between the towns of
Ilam and Mehran (Figure 4). The site is located at 33° 22‟38, 50” N latitude and 46° 16‟15,
93” E longitude, at an elevation of 485 m above sea level, adjacent to the Mesopotamian
plains in western Iran (Zeidi et al. 2012).
Figure 4: Map showing the location of Chogha Golan and other PPN sites in Central Zagros Mountains,
Western Iran (Zeidi et al, 2012).
The site was excavated as part of a joint project with the Iranian Center for Archaeological
Research and the Tübingen-Iranian Stone Age Research Project (TISARP) during the
excavation seasons 2009 and 2010 (Zeidi and Conard in press). The primary goals of the
excavation of Chogha Golan were “the recovery of all classes of organic and inorganic
materials needed to reconstruct the paleoenvironmental setting, the subsistence economy and
the technology of the site‟s inhabitants” (Zeidi et al. 2012). Overall, the excavation of this

21
particular site aims to examine how the timing of early Neolithic development in Western Iran
compares to that of other regions of the Fertile Crescent (Zeidi et al. 2012). Conard et al.
(n.d.) summarize the importance of the Chogha Golan project as follows;
“Since archaeological sites dating to this period are poorly documented in western Iran, the
excavation of Chogha Golan helps to fill on important gap for reconstructing the region‟s
settlement history. This work aims to test the hypothesis that the first phase of lowland
Neolithic settlement would occur in a region adjacent to the mountains, the presumed
natural habitat of key domesticated species […] These results will also help to answer how
the timing of early Neolithic development in Western Iran compares to that of other regions
in the Fertile Crescent. Well dated materials from clear archaeological contexts are needed
to clarify whether or not the Western Zagros represents a separate region of independent
domestication of plants and animals or rather an area to which domesticates from elsewhere
were imported”.
The first excavation season in 2009 aimed to clarify the cultural and chronostratigraphic
sequence of the excavation site (Figure 5). A trench of 4 x 2 meters at the apex of the mound
was excavated to 1 meter deep. Meanwhile, one pit which had already been dug up by looters
to the depth of 5 meters revealed useful information for excavators, providing a guideline of
the stratigraphy of the site. The 7 meter profile wall of the pit was meticulously cleaned for
further analysis.
In 2010, the archaeologists devoted their efforts to a systematic examination of the
stratigraphy by excavating the trench down to the geogenic deposits. Chogha Golan preserves
a thick stratigraphic sequence that includes 8-meter-deep archaeological deposits. Eleven
archaeological horizons (AH) were identified and associated with lithic artefacts, mud brick
walls, and other material culture remains. Also, it has been reported that sediments are rich in
floral remains (Riehl et al. 2012). The zooarchaeological examinations is still awaiting to be
fully investigated but the preliminary results document that the faunal data is large in species
diversity including caprines, wild boar, gazelles, equids, large bovids, rodents, hares, reptiles,
birds, fish, mussels and freshwater crustaceans (Riehl et al. 2013).
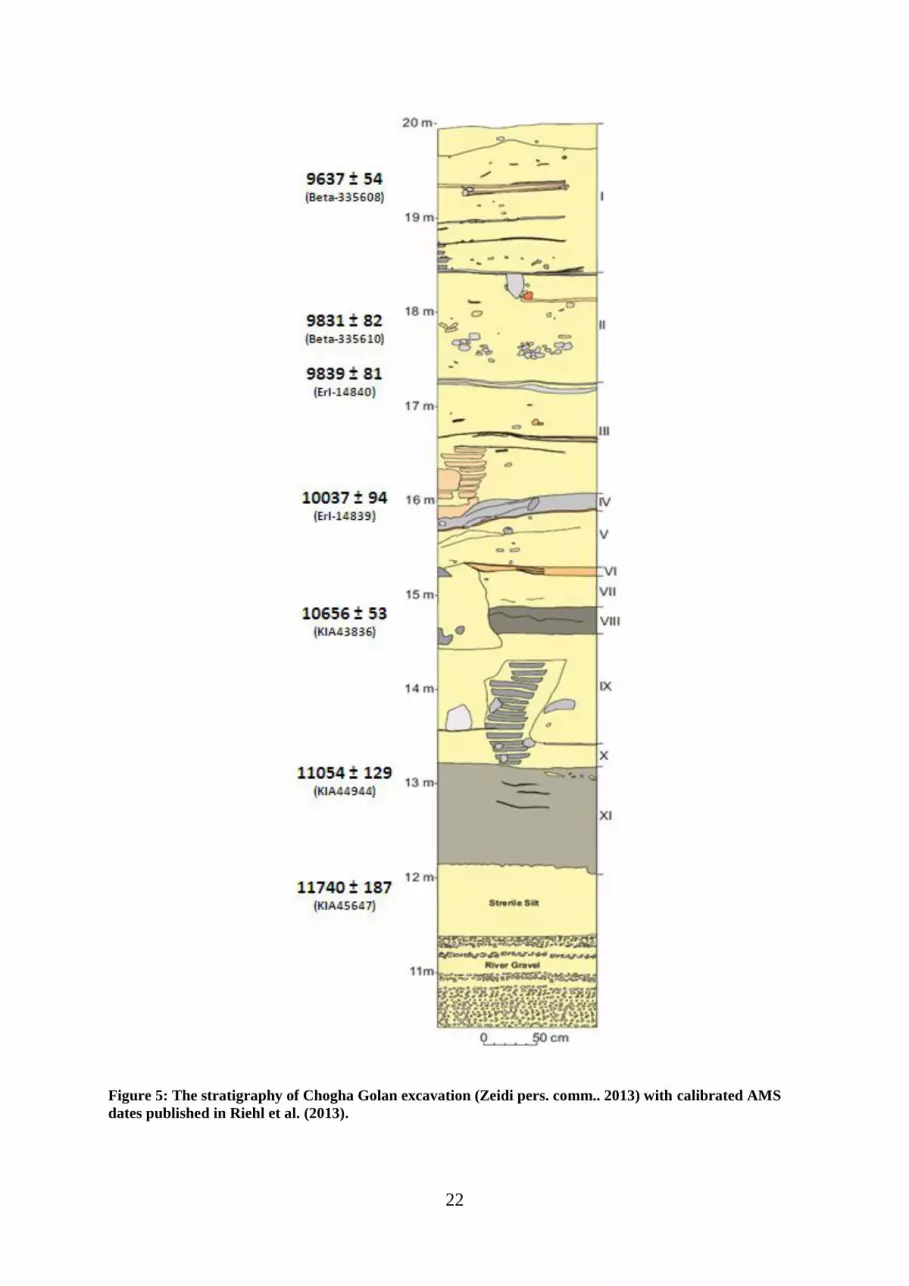
22
Figure 5: The stratigraphy of Chogha Golan excavation (Zeidi pers. comm.. 2013) with calibrated AMS
dates published in Riehl et al. (2013).

23
The majority of findings in AH I were chipped stone artefacts and bone remains. Zeidi and
Conard (in press) report that AH I consists of three sub-layers that were characterized by the
first appearance of mud brick structures (AH Ib) in addition to mortars and pestles, grinding
slabs, and stone and clay objects (AH Ib and AH Ic). Zeidi and Conard (in press) report that
AH II displays “relatively soft and light homogenous ashy silt with light brown to grey color”.
This archaeological horizon is rich with ground stone assemblage and organic remains. It also
includes abundant chipped stones, mud brick, stone structures, and plastered floors.
III.3.2 Chronology
The absence of pottery in all eleven archaeological layers together with the typology of the
lithic assemblage suggests that Chogha Golan was inhabited during the Pre-Pottery Neolithic
period (Zeidi and Conard, in press). This earlier assumption of excavators had been verified
by ten available mass spectrometry (AMS) dates which locate the occupation period of the
site between 12.000 and 9.800 calendar years before present (cal. yr. B.P.). Riehl (2013)
assumes that these dates make the site contemporary with PPNA and middle PPNB sites in
relation to the Southern Levantine chronology5
Radiocarbon dating demonstrates that regionally the start of Chogha Golan occupation
antedates any other PPN archaeological sites in western Iran that have been published so far
(Figure 6). Only Sheikh-e Adab shows evidence for earlier dates than the start of Chogha
Golan occupation (Matthews et al. 2010). According to Riehl et al. (2011), Chogha Golan is
“simultaneous with Nevali Çori in Southeast Anatolia, final Jerf el-Ahmar and Dja‟de in Syria
and Yiftahel, Jericho and Beidha in the Levant. The earliest horizons at Chogha Golan are at
least 1,000 years older than the earliest PPN layers of Ganj Dareh and roughly 1,500 years
older than Ali Kosh”.
5 On the issue of chronological timeframe among different sites in Fertile Crescent, Riehl et al. (2011) emphasize
the difficulty of interpreting the absolute dating as a reference to the cultural similarities and differences in a
relatively large geographical area like Southwest Asia. Nesbitt (2002) points out the same issue as “period terms
such as PPNA are used here [in the text] simply as a convenient shorthand for sites similar in date and do not
necessarily imply cultural similarities”.
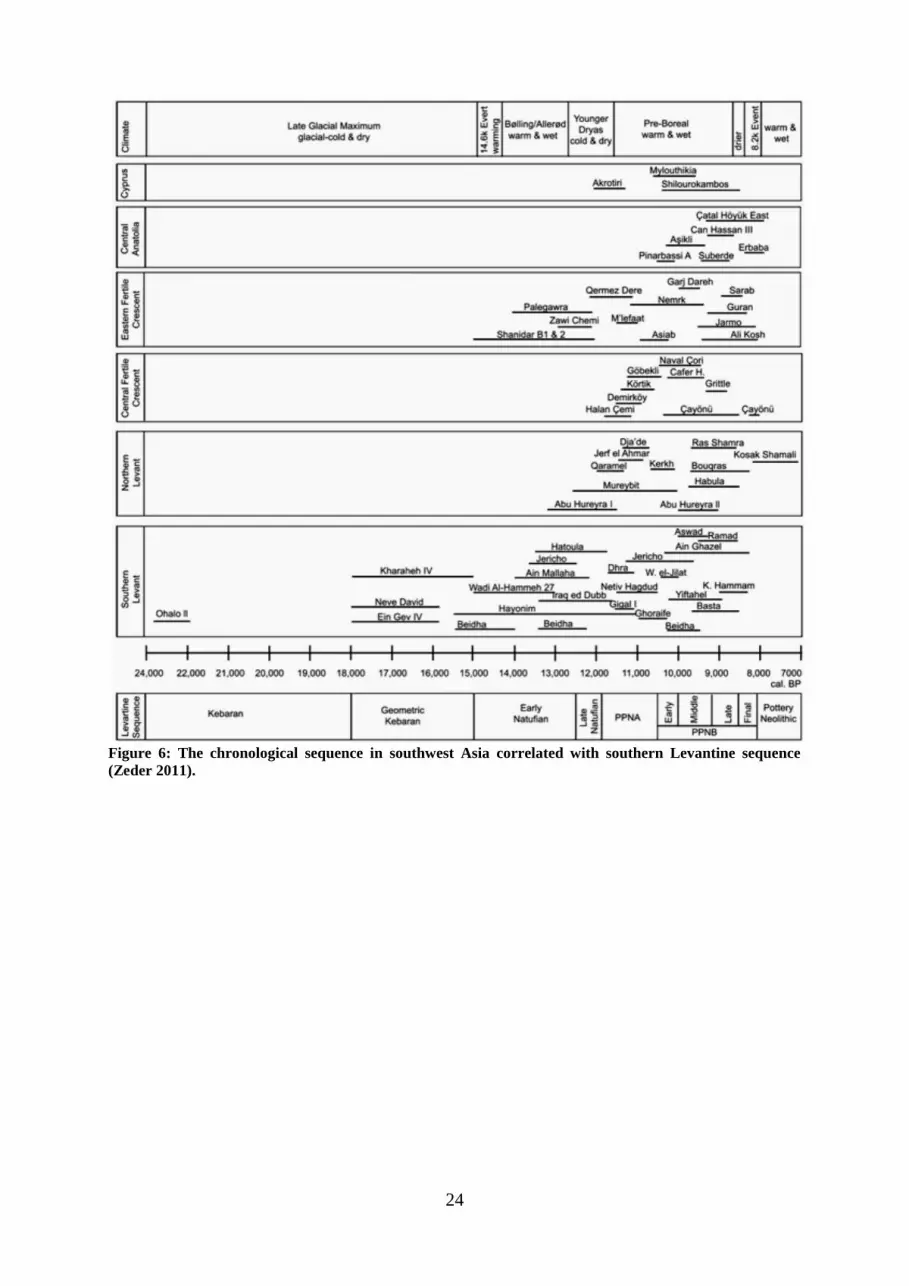
24
Figure 6: The chronological sequence in southwest Asia correlated with southern Levantine sequence
(Zeder 2011).

25
IV. MATERIALS AND METHODS
Archaeobotanical sampling was carried out by the excavators, Mohsen Zeidi and Nicholas
Conard from the University of Tübingen, through a judgmental sampling method based on the
small area of excavation sections and visible changes in the soil profile. In total 717
archaeobotanical samples had been collected by excavators with a mean sediment volume of
10 liters. The collected samples were processed at the site by bucket floatation, using sieves
with mesh sizes of 200 µm (Riehl et al. 2012; 2013). In the meantime, excavators dry-
screened all the sediment removed from the excavation unit with 2 mm mesh (Zeidi and
Conard, in press).
In later analysis, all samples from Chogha Golan were accompanied by archaeobotanical
documentation sheets, providing information including the date, the trench, coordinates, and
sediment volume. During the laboratory process, the samples were separated by dry sieving
them into different fractions (2 mm, 1 mm, 0.63 mm and 0.090 mm) to capture as many plant
remains as possible for efficient sorting. Typical remains of the 2 mm fraction are cereal
grains, large seeded pulses and charcoal. The 1 mm fraction mostly contains small-seeded
pulses and most of the other taxa/genera. Most of the small-seeded grasses were recovered
from the smaller fractions. Other types of objects such as charcoals, increments, and straw,
and awn segments are not directly relevant to the purpose of this research and have been
omitted from the analysis.
The material studied consists of 28 archaeobotanical samples, which are now housed in the
University of Tübingen. From these samples, over 25.000 seed and chaff remains have been
identified and at least categorized in 2012. The identification of plant remains was carried out
in the archaeobotanical laboratory in the Institute for Archaeological Sciences, University of
Tübingen using the comparative collections in the Laboratory of Archaeobotany at the
University of Tübingen and with reference to relevant archaeobotanical publications (e.g.
Anderberg 1994; Berggren 1969, 1981; van Zeist et al. 1984; van Zeist and Bakker-Heeres
1982, 1984a, 1984b, 1985; Nesbitt 2006; Bojnanskỳ and Fargavsová 2007) The plant remains
were identified using a Euromex brand binocular with 10-30x magnification. On the advice of
Dr. Simone Riehl, sub-sampling was conducted for the smallest fraction (0.090 mm) using a
cumulative sampling method with a rifle-type sample splitter, which divides samples in two
identical halves. Sub-samples of 1/8 or 1/4 were sorted to obtain an appropriate number of

26
seeds. In the case of „missing taxa‟, the seed, if it matched one of the available species in the
collection, was described with the extension name „type‟.
The analyzed samples were tabulated in Excel by counting the number of seeds for every
taxa/genera. In total, 61 analytical categories were identified and have been further
amalgamated for clarity, resulting in 37 categories for which the find density and percentages
were calculated using Excel. Correspondence Analysis was performed by using find densities
in CANOCO 4.5 for Windows. A cut-off level of 10 seeds was assigned to exclude the rare
taxa from the data. There were only two plant taxa of which seeds occurred sporadically in the
assemblage (Adonis sp. and Rumex/Polygonum type) and which were removed from the
analysis.
IV.1 Methodological considerations on sample size, taphonomy and quantitative
measurements
An important aspect of archaeobotanical sampling is estimating the required sample size to
recover a representative and accurate dataset (van der Veen 1982). It is generally agreed that
the archaeobotanical material found at a site represents only a small fraction of what was once
present. Furthermore, the principal way those samples are selected from the archaeological
context will possibly influence every later phase of the analysis and interpretation.
Van der Veen‟s (1982) mathematical modeling of sample size resulted in the estimation of a
minimum of 541 seeds/objects in 4 levels of investigation, namely, the site as a whole, each
occupation phase of the site, each category of feature (ditch, pit, postholes etc.) and the
individual samples. According to her formulation, the archaeobotanical material would be
representative only by including 541 seeds in every level described above.
Considering both propositions on sample size, analyzed samples in this research fulfill the
criteria of representativeness in most cases. Accepting van der Veen‟s proposition, only 12
out of 28 individual samples include more than 541 seeds/objects while the figure changes
after amalgamation of individual samples that come from the same horizons. In this level of
investigation, only samples from AH I and AH IX include a number of seeds lower than the

27
minimum amount. These archaeological horizons always demonstrate low number of objects
with low density.
It is also important to consider that the plant species represented in archaeobotanical
assemblages are heavily influenced through various taphonomic processes (i.e., pre-
sedimentary dung burning or crop processing) as well as the effects of post-sedimentary
bioturbation and differential preservation of plants under carbonization. Popper (1988) and
van der Veen (2007) both mention that the source of patterning in plant assemblage has
diverse origins ranging from the human exploitation of plants to the recording of each taxon
by the archaeobotanist.
With regard to this issue, the effects of carbonization and taphonomical processes produced a
substantial amount of over-fragmented plant remains and poorly preserved morphological
features in the Chogha Golan assemblage. van der Veen (2007) noted that the carbonization of
plants would lead to a differential preservation of some species at the expense of others (i.e.,
fruits, condiments, vegetables, and oil-rich seeds are less likely to become preserved).
Hubbard and Clapham (1992, after Fuller 2008b) divide archaeobotanical assemblages into
three distinct groups according to the relationship between context and assemblage. In the
first group, called “class A”, are the remains that were found in situ in the context from which
they were recovered (primary deposition). The context as well in this case should indicate the
signs of burning. The second group of findings (class B) represents an assemblage that comes
from an event (here a burning event) but has been re-deposited from the original context to a
secondary one (secondary deposition). The last group (class C) includes the assemblage from
diverse charring events and many different activities and is considered the most ubiquitous
find class in archaeobotany.
Van der Veen (2007) and Hillman (1984) state that the formation of carbonized plant
assemblages depends on five routes of deposition activity. Following are the routes of entry of
plant remains into an archaeological context, of which the first two represent recurrent daily
activities while the other three display rarer events in the formation of an archaeobotanical
assemblage:
“1) first and foremost, plant remains used as fuel, both intentional and „causal‟ use.
„Causal‟ use refers to the discard into a fire of fine-sieving residues of glume wheats,

28
dehusked on daily basis, as well as of nut shells, fruit stones, and similar. Intentional use
represents the deliberate use of chaff and straw of free-threshing cereals as fuel (in Roman
Egypt traded for such a purpose), and in arid and semi-arid regions the use of animal dung
(which will include chaff and straw remains as well as arable weeds and seeds of grazed
vegetation);
2) foods (especially cereal grains and pulses) accidentally burnt during food preparation
(e.g. bread baking, cooking, roasting), including parching of glume wheats where practiced;
3) stored foods and fodder destroyed by fire in accidents or in deliberate and/or hostile
fires;
4) plants destroyed during the cleaning out of grain storage pits using fire;
5) diseased or infested crop seeds that needed to be destroyed” (van der Veen 2007).
As the Chogha Golan excavations were operated for only two excavation seasons because of
limited time and funding, the archaeological contexts are poorly defined. After the first
excavation season in 2009, excavators decided to dig through the whole sequence down to the
geogenic deposits. This vertical excavation eventually limited the contextual analysis of the
plant remains in the current research. In the mean time, Riehl et al. (2013) informs that source
deposits were relatively uniform in most cases as the collected samples are coming from
“mixed accumulation of ashes from many years of fires that incorporated numerous cycles of
seasonal activities”.
All aspects related to the nature of archaeobotanical data are further complicated by the low
number of samples processed in this research, which could lead to misinterpretation of the
archaeobotanical data. Riehl (1999) stressed that enlarging the number of samples to the
widest possible range of sampled units provides the highest probability for a representative
investigation. However, during this research it was impossible to meet either goal (a large
number of samples and of sampled units) due to limited time and the excavation strategy.
Quantitative measurements are necessary methodological tools to search and describe the
patterning in the data and to distinguish the patterning defined by the research questions from
other sources of patterning. Different methods of quantification would eventually treat the
data with different degrees specificity, require different conditions and provide different
information (Popper 1988). That indicates that the best method of quantification mostly
depends on the condition of the archaeobotanical data.

29
Jones (1991) stresses two basic approaches to statistical analysis, namely pattern searching
and problem-oriented analysis. She mentions that pattern searching starts with counts of the
individual taxa identified in plant parts and uses statistical techniques “to group samples or to
identify major axes of variation on the basis of botanical composition”. On the other hand,
problem-oriented analysis starts with specific questions and applies handles the dataset to
analyze the particular research problem. During this research, pattern searching analysis was
employed mainly to characterize the spatial and temporal variations in the assemblage.
One of the methods applied to the dataset is to express the data through percentage
occurrences of plant taxa within one sample and layer. Converting absolute ratios to a
standardized measure of percentages gives an account in difference of sample size. This
analysis indicates a clear pattern in the increase and decrease of certain species taking the
different occupation periods into consideration (Riehl 1999). A disadvantage of this kind of
measurement is that an increase in one species always results in a decrease in the relative
proportions of others (Jones 1991).
Another method of quantification is to reflect the data as densities per one liter of soil
sediment. According to Jones (1991), the find density analysis “partly reflects the rate of
deposition and can therefore help to distinguish material discarded all at once from that
discarded piecemeal over a period of time and mixed with other refuse”. Riehl (1999),
however, mentions that taphonomic considerations should be taken cautiously as it is not
possible to discern the true number of seeds originally discarded in one deposition event.
A third method that is commonly used in archaeobotany but lacking in this research is to
compare the relative frequencies (ubiquity scores) of plant species through time. This analysis
shows how common a species is within the set of samples. Another important characteristic of
this measurement is that the score of one taxon does not affect the score of another thus
making it possible to evaluate each of the taxa independently. The principal reason for not
applying this measurement to the current dataset is the low total number of samples processed
and the comparatively homogeneous nature of the botanical composition of the samples.
Correspondence Analysis was carried out to distinguish what individual taxa are associated to
certain archaeological horizons in Chogha Golan. Additionally another CA plot was
performed to explore patterning in the dataset to identify similarities between samples on the

30
basis of their species compositions and identifies which species co-occur frequently. Lange
(1990; after Colledge 1999) mentions that “in graphical form the results of a Correspondence
Analysis bring out the position of each samples relative to all other samples and to all the
species, and of each species relative to all other species and to all the samples in the analysis”.
Colledge (2002) and van der Veen (2007) noted that this multivariate analysis enable the
researcher to identify similarities between samples on the basis of their species compositions
and allows looking for any meaningful grouping of samples and species from the sites and
chronological periods and phases to be investigated.
IV.2 Further limitations in the current research
Certain constraints that affect the archaeobotanical interpretation of the Chogha Golan
assemblage should be further considered in detail. For example, the degree of expertise of the
examiner is an important criterion in the identification process. It is obvious that identification
of carbonized plant materials requires well-grounded experience in the morphology of the
seeds as well as a deep understanding of plant ecology and biology. Choices in research
strategy and limited time for laboratory analysis made it impossible to assign detailed species-
level identifications; rather, most taxa were coarsely identified at the genus level or
identifications remained tentative and indeterminate. This caused the loss of valuable
palaeoecological information in this research.
There are two other major limitations, both closely related to the biological aspects of plants.
The difficulty in identifying the pulses (Fabaceae) at the species level was the result of large
intraspecies variations within this family, which are mostly attested to their nitrogen-fixing
ability through symbiotic bacteria that makes them adapt well to nitrogen-deficient soils. For
instance, the Astragalus genus in Iran comprises over 500 species, complicating the issue
further, even for botanists. Another biologically-derived aspect could be the presence of
extinct or still-undiscovered species. Some taxa in the assemblage such as the Triticoid type
and Agrostis type have no modern equivalents in identification manuals. Both types were
defined through their close morphological similarities to Triticum and Agrostis genera, but it
is uncertain that these remains truly belong to these groups.
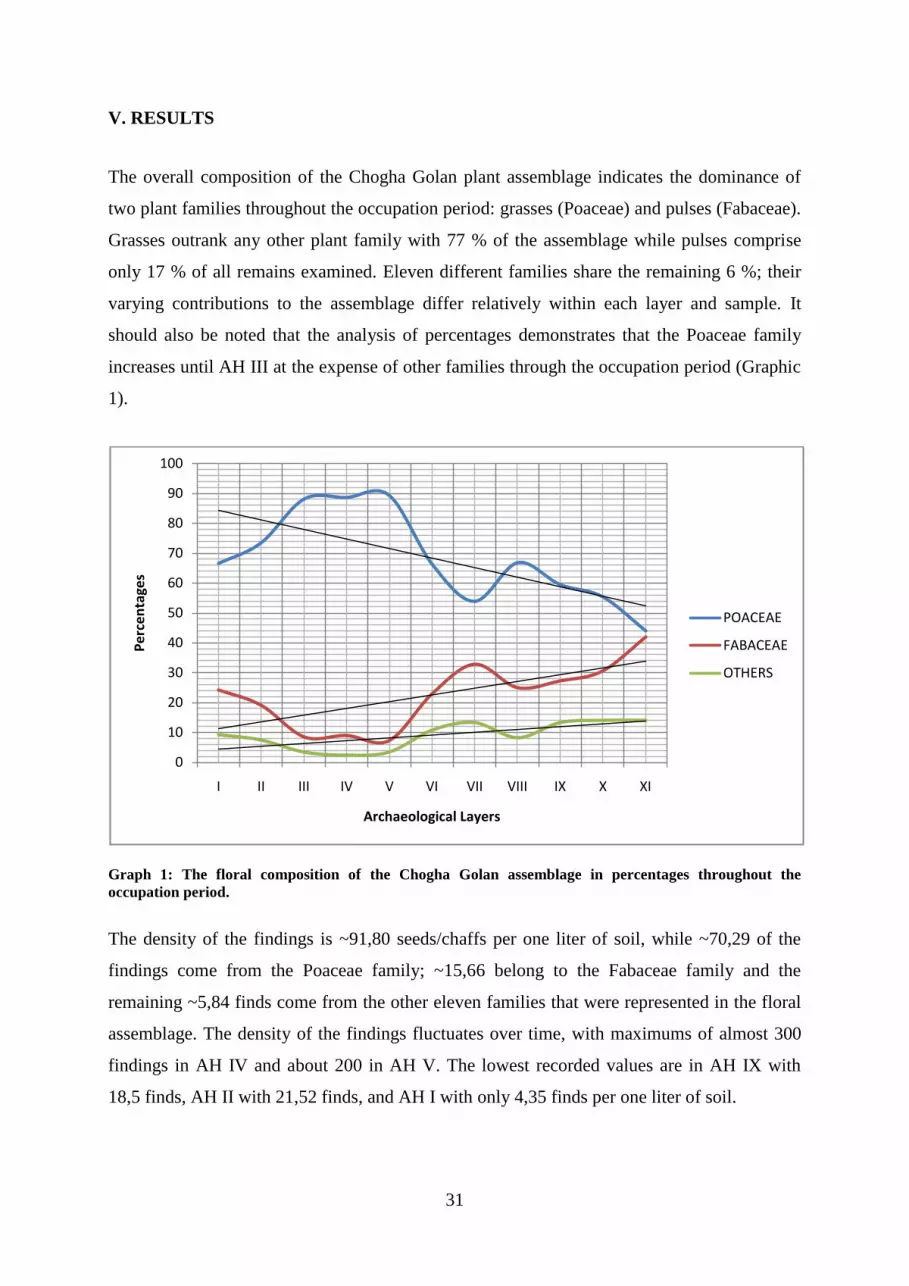
31
V. RESULTS
The overall composition of the Chogha Golan plant assemblage indicates the dominance of
two plant families throughout the occupation period: grasses (Poaceae) and pulses (Fabaceae).
Grasses outrank any other plant family with 77 % of the assemblage while pulses comprise
only 17 % of all remains examined. Eleven different families share the remaining 6 %; their
varying contributions to the assemblage differ relatively within each layer and sample. It
should also be noted that the analysis of percentages demonstrates that the Poaceae family
increases until AH III at the expense of other families through the occupation period (Graphic
1).
Graph 1: The floral composition of the Chogha Golan assemblage in percentages throughout the
occupation period.
The density of the findings is ~91,80 seeds/chaffs per one liter of soil, while ~70,29 of the
findings come from the Poaceae family; ~15,66 belong to the Fabaceae family and the
remaining ~5,84 finds come from the other eleven families that were represented in the floral
assemblage. The density of the findings fluctuates over time, with maximums of almost 300
findings in AH IV and about 200 in AH V. The lowest recorded values are in AH IX with
18,5 finds, AH II with 21,52 finds, and AH I with only 4,35 finds per one liter of soil.
0
10
20
30
40
50
60
70
80
90
100
I II III IV V VI VII VIII IX X XI
Pe
rce
nta
ges
Archaeological Layers
POACEAE
FABACEAE
OTHERS

32
V.1 The Composition of Poaceae (Grass Family)
The plant family Poaceae consists of fourteen categories in varying frequencies. In total, nine
of the fourteen categories include seed remains, which were classified into two categories as
large- and small-seeded grasses according to their sizes.
Another characteristic of the Poaceae spectrum is the high percentages of chaff remains in
general. Five categories have been documented as rachis remains in the findings belong to
Hordeum cf. spontaneum, Aegilops sp., Triticum sp., Taeniatherum sp., and “unidentified
Poaceae spikelet base type” (Graphic 2).
Graph 2: Relative percentages of Poaceae chaff and seed remains in the plant assemblage.
V.1.1 Large-seeded Poaceae remains
Hordeum cf. spontaneum (wild barley) is a well-represented species in the assemblage. It is
ubiquitous in every layer and findable in most samples except AH I. The absence of barley
grains in AH I could be associated with the small sample size, since the chaff remains of this
species do exist in this layer. In total, the absolute percentages indicate low values that range
from 0,22 % to 1,95 %. The highest values are recorded in AH VII and AH IX, although the
sample size for both those layers is relatively small. In the meantime, the find density analysis
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
I II III IV V VI VII VIII IX X XI
Pe
rce
nta
ges
Archaeological Layers
TOTAL (CHAFF)
TOTAL (SEED)
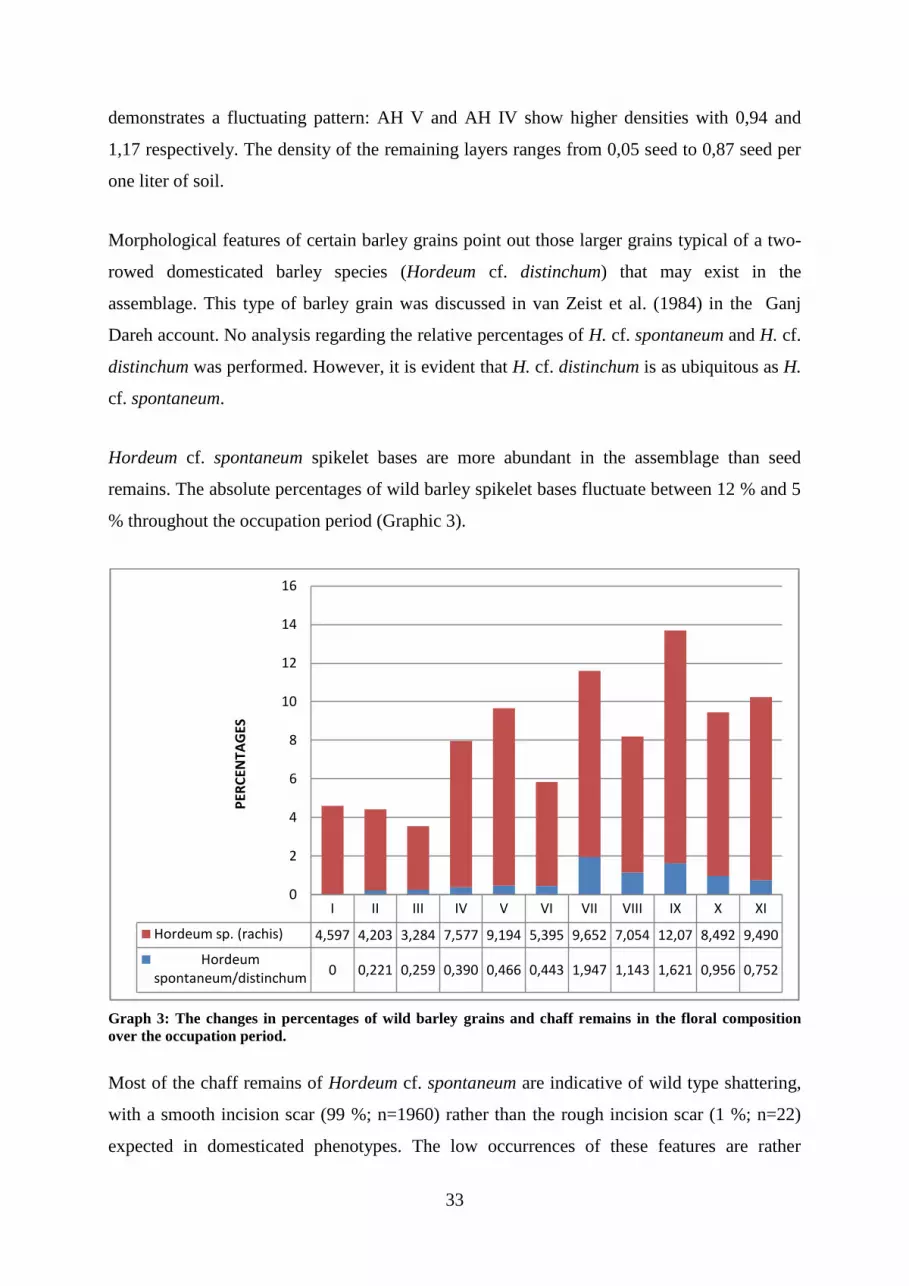
33
demonstrates a fluctuating pattern: AH V and AH IV show higher densities with 0,94 and
1,17 respectively. The density of the remaining layers ranges from 0,05 seed to 0,87 seed per
one liter of soil.
Morphological features of certain barley grains point out those larger grains typical of a two-
rowed domesticated barley species (Hordeum cf. distinchum) that may exist in the
assemblage. This type of barley grain was discussed in van Zeist et al. (1984) in the Ganj
Dareh account. No analysis regarding the relative percentages of H. cf. spontaneum and H. cf.
distinchum was performed. However, it is evident that H. cf. distinchum is as ubiquitous as H.
cf. spontaneum.
Hordeum cf. spontaneum spikelet bases are more abundant in the assemblage than seed
remains. The absolute percentages of wild barley spikelet bases fluctuate between 12 % and 5
% throughout the occupation period (Graphic 3).
Graph 3: The changes in percentages of wild barley grains and chaff remains in the floral composition
over the occupation period.
Most of the chaff remains of Hordeum cf. spontaneum are indicative of wild type shattering,
with a smooth incision scar (99 %; n=1960) rather than the rough incision scar (1 %; n=22)
expected in domesticated phenotypes. The low occurrences of these features are rather
I II III IV V VI VII VIII IX X XI
Hordeum sp. (rachis) 4,597 4,203 3,284 7,577 9,194 5,395 9,652 7,054 12,07 8,492 9,490
Hordeum spontaneum/distinchum
0 0,221 0,259 0,390 0,466 0,443 1,947 1,143 1,621 0,956 0,752
0
2
4
6
8
10
12
14
16
PER
CEN
TAG
ES

34
striking with respect to the long occupation period at Chogha Golan. No pattern is evident in
the assemblage to show an increase of non-shattering phenotypes during the succeeding
periods of occupation.
Some seed remains show a close resemblance to the wild siblings of Hordeum cf.
spontaneum. This group of wild barley remains was not successfully determined at the species
level. The absolute percentages of the findings for this group are lower than those of H. cf.
spontaneum, which has values higher than those for wild barley only in AH IV. The density
values also display the same pattern in that AH IV is significant for higher find density of this
category.
Aegilops sp. (goat grass) seeds are rarely represented in the assemblage. The absolute
percentages vary from 0,16 % to 0,76 % regarding the whole assemblage although it is absent
in the two uppermost layers. However, the chaff remains of this genus are very well
represented in the assemblage, as in the case of Hordeum cf. spontaneum.
Aegilops sp. chaff remains compose one of the most abundant categories within the large-
seeded grasses category and within the plant assemblage as a whole. The rachis remains of
this genus are highly fragmented into smaller objects, while complete remains of spikelet
bases and glume bases exist rather in low counts. The absolute percentages for these findings
are always high, the lowest being 4,50 % in AH IV and the highest 36,70 % in AH VIII. The
find density analysis, on the other hand, demonstrates the densest values in AH III with ~40
fragments per one liter of soil. In other layers, the values range from 0,90 (AH I) to 26,67
(AH XI) per one liter of soil.
Taenitherum sp. and Bromus sp. are relatively smaller grasses in comparison with Aegilops
sp. and Hordeum cf spontaneum. Both genera appear ubiquitously in most samples which
could indicate that these two plants were a permanent element of the vegetation. However, the
absolute percentages of both taxa vary between the ranges of 3 % to 1 %.
The highest percentage is recorded in AH VII for Bromus sp. with 2,79 %. Taenitherium sp. is
much better represented by its chaff remains than by its seeds. The spikelet bases of this genus
compose 0,76 % of the assemblage in AH VII. The density of Bromus sp. reaches its peak in

35
AH VII with ~1 seed per one liter of soil, while Taenitherium sp. seed and chaff get their
higher values in AH V and AH IV, respectively.
“Indeterminate large-medium seeded grasses” include all grass remains left unidentified
during the analysis of the plant remains. This category is relatively large and includes many
different types of grasses. The higher percentages were recorded in the levels below AH III,
with the exception of a suspicious decrease in AH V. The upper layers show lower
percentages than were recorded for the earlier layers. The highest densities of these finds
occur in AH IV (7,17 seeds per liter) and in AH XI (3 seeds per liter).
Only one specimen of Triticum sp. (wheat genus) seed remains was recovered from AH III.
Preliminary examinations demonstrate that this finding may belong to T. monococcum ssp.
boeticum (wild einkorn) or T. turgidum ssp. dicoccoides (wild emmer).
Another pattern in the Chogha Golan assemblage is the frequent occurrence of Triticum sp.
chaff remains. Even though these remains appear from the earlier layers on, their absolute
percentages become increasingly pronounced in the later levels of occupation. In particular,
the three uppermost layers consist of a considerable amount of Triticum sp. chaff remains.
This observation aside, further analysis of the domestication status of Triticum sp. spikelet
bases, based on the differentiation of chaff morphology, was hard to confirm due to the highly
fragmented nature of preservation.
In AH VII, the absolute percentages of Triticum sp. is suspiciously high (2,71 %) although the
neighboring layers show little or no evidence for this type of chaff remains. The percentages
gradually rise from 0,35 % in AH IV to 18,39 % in AH I. The find density on the other hand,
fluctuates between 0,03 (AH XI) and 1,90 (AH II). The uppermost layer indicates relatively
low values of density with 0,90 chaff per one liter of soil.
One specimen of Triticum sp. chaff remains needs special attention in the evaluation of the
Chogha Golan plant assemblage. This specimen, found in AH III, shows close morphological
similarities to the free-threshing type of spikelet bases. The morphological characteristics
resemble T. aestivum (bread wheat) rather than the T. durum (hard wheat) type. In addition,
the abscission scar of this particular spikelet base is smoothly broken, rather than resembling
the non-shattering free-threshing types.

36
V.1.2 Small-seeded Poaceae remains
Four categories within the Poaceae family were defined as small-seeded grasses. These
categories consist of Agrostis type, Phalaris sp., Triticoid type and “unidentified Poaceae
rachis type”. Agrostis type makes up almost half of the whole assemblage (52 %). In total,
these four categories contribute 63 % of the assemblage.
Agrostis type is a broad group of grass remains that the group mainly include several types of
seeds less than 2 mm in length. The type was defined previously in the publication by van
Zeist et al. (1984) on the Ganj Dareh plant assemblage. The most abundant finding in this
category, this type of grass was classified as “Agrostis type” in accordance with that
publication. The fossil plants in this category were concentrated in AH V, IV, and III, while
their overall contributions are much less pronounced in the remaining layers.
Phalaris sp. (canary grass) is a well-represented genus in the assemblage. The percentages of
this genus fluctuate from 8 % to 1 % throughout the occupation period, with the steady
occurrences in earlier layers altering dramatically in AH V with a rapid increase in percentage
The find density of Phalaris sp. reaches its highest values in AH V and AH IV, while the
density in other layers is generally less than 1 grain per one liter of soil. Considering its
ubiquity in the assemblage, it is obvious that Phalaris sp. is an important vegetational
element.
Triticoid type grass remains are frequently occurring in every layer, albeit in low percentages
(from 2 % to 0,3 %). This type of grass remains was also first defined by van Zeist (1984) at
Ganj Dareh, due to its close morphological similarities to the Triticum genus, although it is
comparatively much smaller in size. The density of these findings indicates that AH VI and V
differentiate with the highest values of these objects. On the other hand, the percentage
analysis displays the highest value in AH XI, with a decrease through succeeding layers. No
remains of the Triticoid type were recorded in AH II and AH I.
“Unidentified Poaceae rachis type” is a type of object that was not successfully identified in
this research. Among many other unidentified objects in the assemblage, the remains defined
under this category demonstrate high density values and relatively high percentages (from 8
% to >1 %). For this reason, this type is included pending further analysis. The highest values
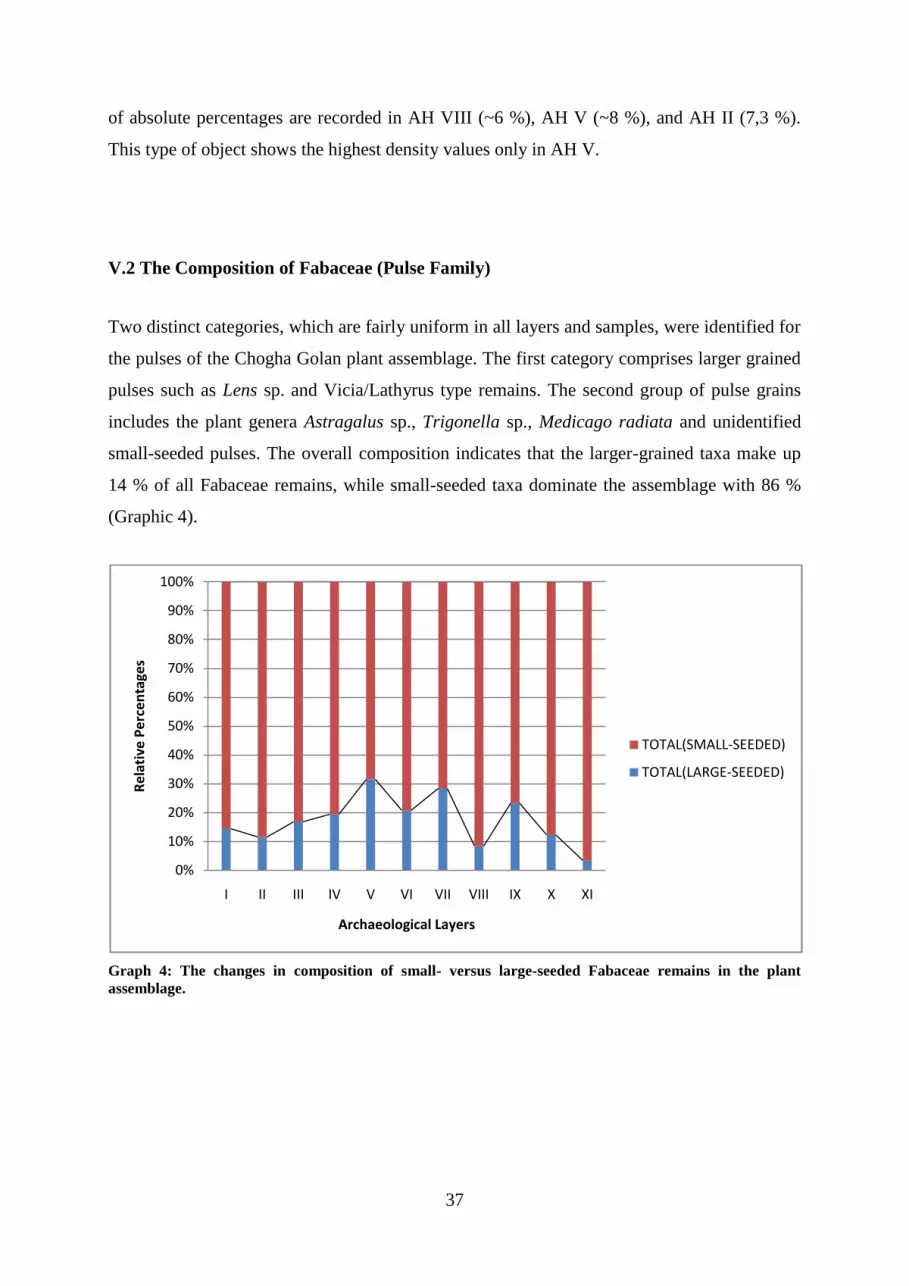
37
of absolute percentages are recorded in AH VIII (~6 %), AH V (~8 %), and AH II (7,3 %).
This type of object shows the highest density values only in AH V.
V.2 The Composition of Fabaceae (Pulse Family)
Two distinct categories, which are fairly uniform in all layers and samples, were identified for
the pulses of the Chogha Golan plant assemblage. The first category comprises larger grained
pulses such as Lens sp. and Vicia/Lathyrus type remains. The second group of pulse grains
includes the plant genera Astragalus sp., Trigonella sp., Medicago radiata and unidentified
small-seeded pulses. The overall composition indicates that the larger-grained taxa make up
14 % of all Fabaceae remains, while small-seeded taxa dominate the assemblage with 86 %
(Graphic 4).
Graph 4: The changes in composition of small- versus large-seeded Fabaceae remains in the plant
assemblage.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
I II III IV V VI VII VIII IX X XI
Re
lati
ve P
erc
en
tage
s
Archaeological Layers
TOTAL(SMALL-SEEDED)
TOTAL(LARGE-SEEDED)
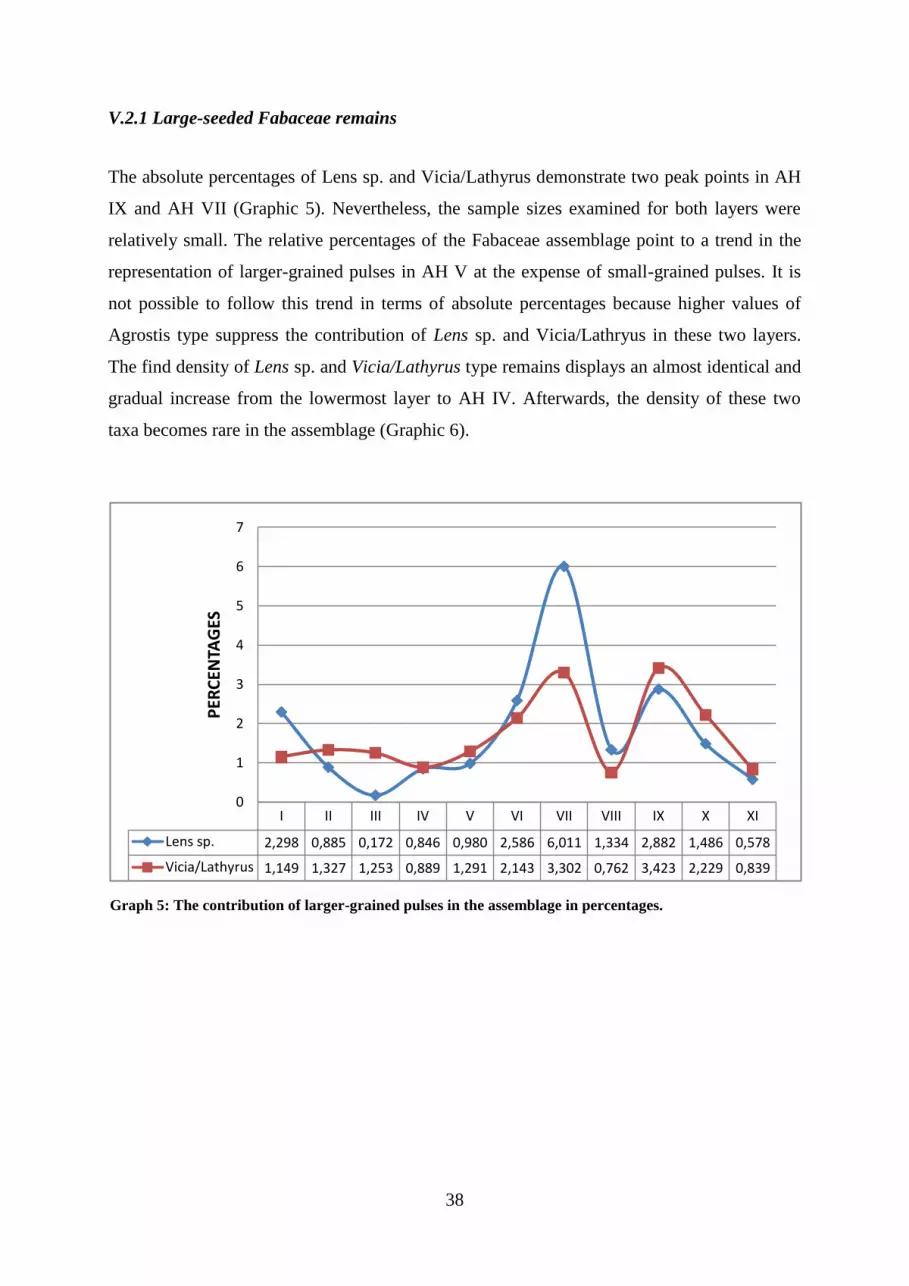
38
V.2.1 Large-seeded Fabaceae remains
The absolute percentages of Lens sp. and Vicia/Lathyrus demonstrate two peak points in AH
IX and AH VII (Graphic 5). Nevertheless, the sample sizes examined for both layers were
relatively small. The relative percentages of the Fabaceae assemblage point to a trend in the
representation of larger-grained pulses in AH V at the expense of small-grained pulses. It is
not possible to follow this trend in terms of absolute percentages because higher values of
Agrostis type suppress the contribution of Lens sp. and Vicia/Lathryus in these two layers.
The find density of Lens sp. and Vicia/Lathyrus type remains displays an almost identical and
gradual increase from the lowermost layer to AH IV. Afterwards, the density of these two
taxa becomes rare in the assemblage (Graphic 6).
I II III IV V VI VII VIII IX X XI
Lens sp. 2,298 0,885 0,172 0,846 0,980 2,586 6,011 1,334 2,882 1,486 0,578
Vicia/Lathyrus 1,149 1,327 1,253 0,889 1,291 2,143 3,302 0,762 3,423 2,229 0,839
0
1
2
3
4
5
6
7
PER
CEN
TAG
ES
Graph 5: The contribution of larger-grained pulses in the assemblage in percentages.
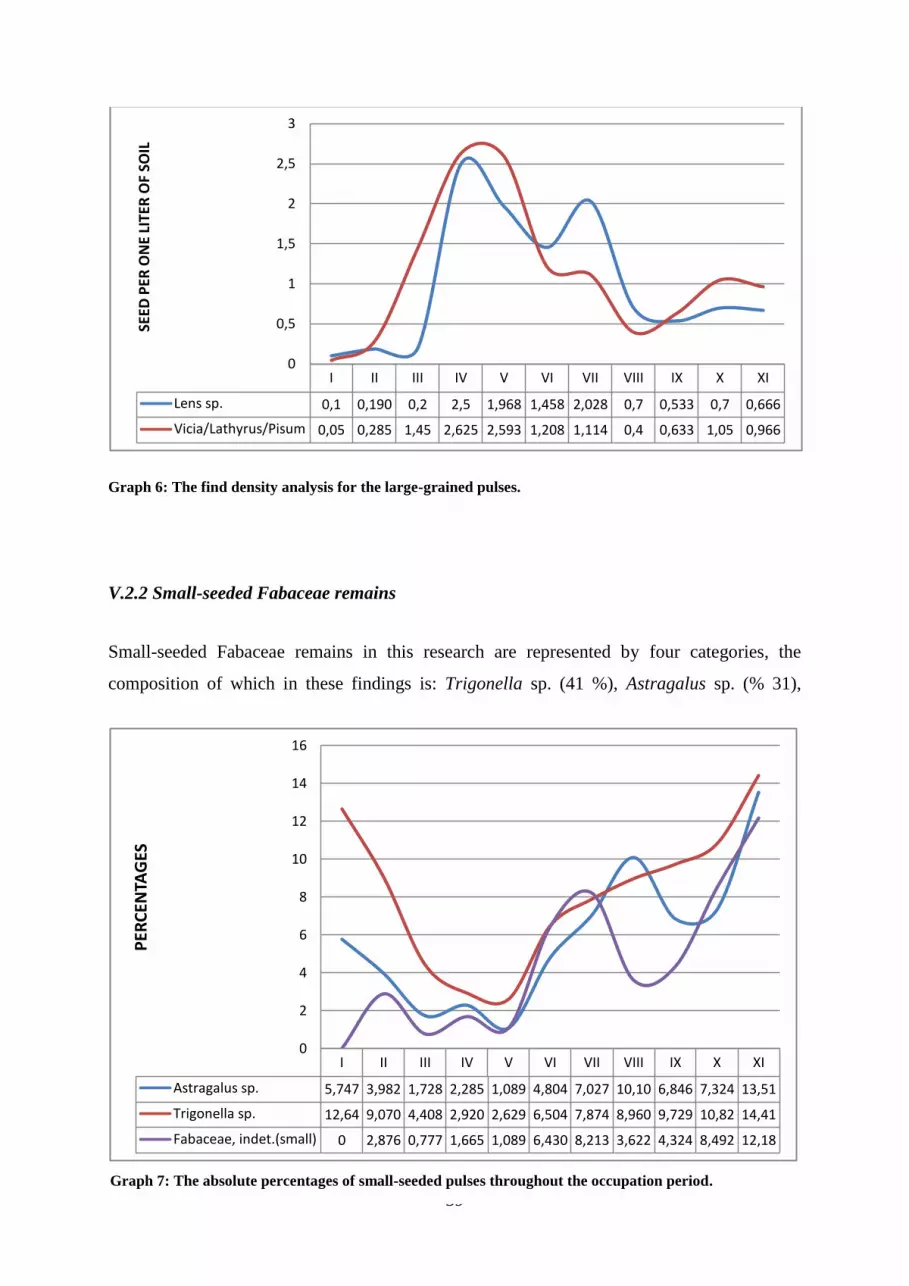
39
Graph 6: The find density analysis for the large-grained pulses.
V.2.2 Small-seeded Fabaceae remains
Small-seeded Fabaceae remains in this research are represented by four categories, the
composition of which in these findings is: Trigonella sp. (41 %), Astragalus sp. (% 31),
I II III IV V VI VII VIII IX X XI
Lens sp. 0,1 0,190 0,2 2,5 1,968 1,458 2,028 0,7 0,533 0,7 0,666
Vicia/Lathyrus/Pisum 0,05 0,285 1,45 2,625 2,593 1,208 1,114 0,4 0,633 1,05 0,966
0
0,5
1
1,5
2
2,5
3
SEED
PER
ON
E LI
TER
OF
SOIL
I II III IV V VI VII VIII IX X XI
Astragalus sp. 5,747 3,982 1,728 2,285 1,089 4,804 7,027 10,10 6,846 7,324 13,51
Trigonella sp. 12,64 9,070 4,408 2,920 2,629 6,504 7,874 8,960 9,729 10,82 14,41
Fabaceae, indet.(small) 0 2,876 0,777 1,665 1,089 6,430 8,213 3,622 4,324 8,492 12,18
0
2
4
6
8
10
12
14
16
PER
CEN
TAG
ES
Graph 7: The absolute percentages of small-seeded pulses throughout the occupation period.

40
“unidentified small-seeded Fabaceae taxa” (26 %), and Medicago radiata (2 %). Except for
Medicago radiata which occurs sporadically in the assemblage, the other three categories
display relatively similar distributions throughout the occupation period. The absolute
percentages demonstrate high values at the lower layers until AH V. Then, a sharp decrease is
visible for AH V and AH IV. The percentages increase again to a certain degree in the three
succeeding layers (Graphic 7).
It should be noted that the prevelance of Trigonella sp. is comparatively higher than any other
small-seeded Fabaceae genera from AH V on. Trigonella sp. in this research includes two
types of plant remains: Trigonella astroides and unidentified Trigonella (most probably)
objects. Unlike most of the fossil plants in the assemblage, Trigonella astroides is one of the
few taxa that were identified to the species level due to its distinct morphological
characteristics.
The find density of Trigonella sp., Astragalus sp. and unidentified small-seeded legumes
displays a constantly decreasing pattern from the lowermost layers to the uppermost layer.
The remains display the highest density in AH XI, around 16–14 seeds per one liter of soil.
These values from this particular layer are never achieved again in the remaining layers. AH
VIII differentiates from the neighboring layers with an increase in density for Astragalus sp.
and Trigonella sp., and then another rapid increase is apparent in AH IV.
I II III IV V VI VII VIII IX X XI
Astragalus sp. 0,25 0,857 2 6,75 2,187 2,708 2,371 5,3 1,266 3,45 15,56
Trigonella sp. 0,55 1,952 5,1 8,625 5,281 3,666 2,657 4,7 1,8 5,1 16,6
Medicago radiata 0,1 0,238 0,25 1,041 0,281 0,208 0,114 0,15 0 0,05 0,466
Fabaceae, indet.(small) 0 0,619 0,9 4,916 2,187 3,625 2,771 1,9 0,8 4 14,03
-2
0
2
4
6
8
10
12
14
16
18
FIN
D D
ENSI
TY
Graph 8. The density of small-seeded pulses per one liter of soil throughout the occupation period.

41
Interestingly, the density of small-seeded Fabaceae genera in AH V is not as dense as in AH
IV, even though both layers indicate an almost similar floral composition. While Trigonella
sp. has the highest density among small-seeded legumes, the last three layers show very low
density values in all three categories (Graphic 8).
V.3 The Composition of Other Plant Families
Plant families which are neither Poaceae nor Fabaceae compose 6 % of the assemblage.
Eleven plant families were identified during this research, consisting of 18 genera/categories
in total (Graphic 9). The number of genera/categories included in the data analysis does not
reflect the real picture regarding the species diversity in the Chogha Golan assemblage. Some
unidentified seed remains and some sporadically occurring genera were not included in the
actual data analysis due to low counts of these remains. Thus it should be kept in mind that
the species diversity in the Chogha Golan plant assemblage is much larger than is expressed
in this study.
V.3.1 Anacardiceae (Sumac Family)
The plant family Anacardiceae is represented by only one genus, Pistacia sp. This genus
occurs in every sample in varying percentages and it is the largest category (29 %) among
other genera that do not belong to the Poaceae and Fabaceae families.
The highest values in percentages indicate its significance in AH XI (6 % of the whole
assemblage). In the upper layers, the absolute percentages vary from 3 % to >1 %. The find
density analysis demonstrates a similar picture as the highest values are recorded in AH XI
with much lower densities in the remaining layers. Pistacia sp. is relatively abundant in AH
VIII, and AH V and IV display a sharp peak in the find density of the remains. Interestingly,
AH VII and AH VI demonstrate very low densities for this genus.
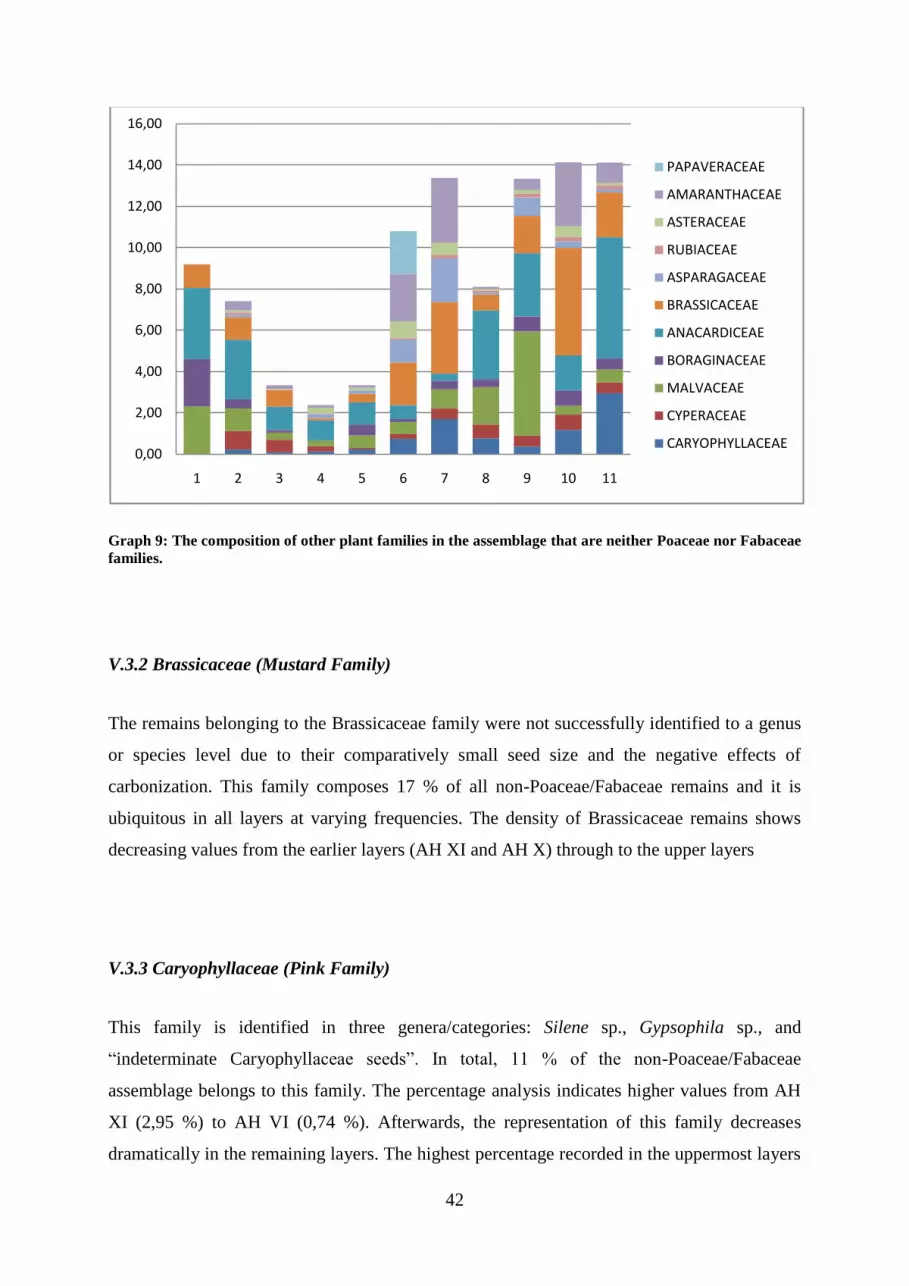
42
Graph 9: The composition of other plant families in the assemblage that are neither Poaceae nor Fabaceae
families.
V.3.2 Brassicaceae (Mustard Family)
The remains belonging to the Brassicaceae family were not successfully identified to a genus
or species level due to their comparatively small seed size and the negative effects of
carbonization. This family composes 17 % of all non-Poaceae/Fabaceae remains and it is
ubiquitous in all layers at varying frequencies. The density of Brassicaceae remains shows
decreasing values from the earlier layers (AH XI and AH X) through to the upper layers
V.3.3 Caryophyllaceae (Pink Family)
This family is identified in three genera/categories: Silene sp., Gypsophila sp., and
“indeterminate Caryophyllaceae seeds”. In total, 11 % of the non-Poaceae/Fabaceae
assemblage belongs to this family. The percentage analysis indicates higher values from AH
XI (2,95 %) to AH VI (0,74 %). Afterwards, the representation of this family decreases
dramatically in the remaining layers. The highest percentage recorded in the uppermost layers
0,00
2,00
4,00
6,00
8,00
10,00
12,00
14,00
16,00
1 2 3 4 5 6 7 8 9 10 11
PAPAVERACEAE
AMARANTHACEAE
ASTERACEAE
RUBIACEAE
ASPARAGACEAE
BRASSICACEAE
ANACARDICEAE
BORAGINACEAE
MALVACEAE
CYPERACEAE
CARYOPHYLLACEAE

43
is 0,23 % in AH V. The find density analysis displays very low values of less than 1 grain per
one liter of soil, except AH XI with 3,40 grains.
V.3.4 Malvaceae (Mallow Family)
This family is represented in the assemblage by only one genus, Malva sp. The genus exhibits
the highest percentage in AH IX, while in other layers the percentages are between 2,30 % to
0,27 % of the whole assemblage. The find density analysis records about one seed remain per
one liter of soil in AH IX, AH VII, and AH V. The remaining layers show lower densities.
Apart from that, this genus is ubiquitous in all archaeological horizons.
V.3.5 Chenopodiaceae/Amaranthaceae (Goosefoot/Amaranth Family)
The family Chenopodiaceae/Amaranthaceae is represented by four genera/categories
including Salsola sp., Atriplex sp., Suaeda sp., and “indeterminate Chenopodiceae objects”.
This family is ubiquitous in all layers except in AH I.
The range of percentages varies from 3,13 % to 0,10 % of the whole assemblage. It is evident
that there are distinct differences between AH X, AH VII, and AH VI, which have relatively
high percentages of this family, and the remaining layers that indicate lower percentages,
mostly less than 1 % of the whole assemblage. The density of these remains fluctuates from
1,45 to 0,10 seeds per one liter of soil. As seen in the percentage analysis, the particular layers
mentioned above exhibit high densities together with AH XI. Furthermore, an interesting
pattern occurs when all four genera/categories are plotted in the same chart. This shows the
density of Suaeda sp. peaking in certain layers such as AH X, AH VI, and AH IV, while it is
non-existent in the remaining layers. At the same time, Atriplex sp. and Salsola sp. are more
ubiquitous in the assemblage despite their lower densities, while “indeterminate
Chenopodiceae objects” are concentrated only in AH VII and VI.

44
V.3.6 Cyperaceae (Sedge Family)
The absolute percentages of this family are less than 1 %. It is represented by only one genus,
Scirpus sp. The percentages exhibit comparatively similar values in each layer, on average
0,50 % of the whole assemblage. Only in AH VI, AH V, and AH IV do the percentages
decrease notably. The find density analysis displays the highest values in AH XI, AH VI, and
AH III. Nevertheless, this genus is ubiquitous in all layers except AH I.
V.3.7 Boraginaceae (Borage Family)
The family Boraginaceae composes very small portion of the whole assemblage (~1 %) even
though it is ubiquitous in all horizons except AH IV. The remains of this family are most
abundant in AH XI and AH V. Among other genera like Lithospermum sp. and Arnebia sp.,
Heliotropium sp. is the most abundant genus recorded in the assemblage.
V.3.8 Asteraceae (Sunflower Family)
The only plant category identified for this family is Centaurea type. The absolute percentages
of this category are less than 1 % of the whole assemblage. The analysis exhibits the highest
percentages in AH X, VII, and VI, while the density of this plant is higher in AH VII and VI.
It is present in all layers except AH I.
V.3.9 Asparagaceae
The remains for this family are classified under the category of
Ornithagalum/Muscari/Bellavalia. Because of the difficulties in identifying these genera due
to the effects of carbonization, they are all included in the same category. The absolute
percentages are the highest in AH VII, with 2 % of the whole assemblage. Other layers show
percentages lower than 1 %. The find density for this category displays higher values from

45
AH VII to AH IV, while it is less densely represented in the remaining archaeological
horizons.
V.3.10 Rubiaceae (Bedstraw Family)
Galium sp. is the only genus identified for this family and is present in low percentages (<
0,21 %), although it is not present at all in AH V, AH III, and AH I. The find density analysis
shows the highest density of this genus in AH XI, with decreasing densities after this
lowermost layer.
V.3.11 Papaveraceae (Poppy Family)
Papaver sp. is present only in AH VI, with roughly 2 % of the whole assemblage. This genus
is represented by a density of 1,20 seeds per one liter of soil in this particular layer.
V.4. General Patterns in the Chogha Golan Assemblage
A closer look at the composition of the plant assemblage indicates that a large number of
plant genera/taxa occur rather ubiquitously throughout the occupation period. However, they
are not uniformly distributed over all layers and samples. It is evident that certain layers and
samples are more associated with certain plant taxa than others in the assemblage.
Plotting the covariational relationships between taxonomic compositions of individual
samples reveals that three patterns predominate in separatingthe location of samples in the CA
plot (Graph 10). The strongest separation appears along the first principal axis (horizontal)
and explains 42.8 % of all variations in the dataset. This shows that the first principal axis
separates the samples that include a large number of small-seeded taxa (both grasses and
legumes) from the samples with predominantly including large-seeded grasses and wheat
spikelets.

46
This situation can be illustrated by looking at some samples that come from the same
archaeological horizon but are located in negative or positive sides of the plot. For instance,
among four samples examined from AH VII, three of them (sample no. 52, 366, and 367) tend
to locate at the negative side of the first axis. These three samples are enriched with small-
seeded legumes and the remains of Chenopodiceae family, Ornithagalum/Muscari, and
Centaurea type as well as large-seeded pulses like Lens sp. and Vicia/Lathyrus type. On the
other hand, sample no. 613 dislocated from this cluster at the positive side of the axis due to a
strikingly large amount of wheat spikelets. Such a pattern is also identifiable in AH II
samples, in which samples no. 160 and 147 located at the positive side of the axis while
sample no. 137 tends to cluster more negatively. This is mainly the result of the sample
composition of no. 137, which includes a substantial amount of large- and small-seeded
pulses in contrast to low proportions in other samples. Instead samples no. 160 and 147
consist of large amounts of wheat spikelets with small amounts of large- and small-seeded
pulses.
The second principal axis (vertical) explains about 12 % of the variations in the assemblage.
The negative side of the axis is characterized by the samples that include a large amount of
small-seeded grasses, while the positive side of the axis shows the groupings of samples that
generally include the remaining taxa. For instance, all three samples from AH V indicate a
different pattern according to their location in the plot. Sample no. 327 consists of a large
amount of small-seeded grasses (Agrostis type, Phalaris sp.) and is located at the negative
side of the axis while two other samples differ from this pattern by the variations in their
composition. Sample no. 320 tends to cluster with the samples at the positive side of the
second principal axis but also at the negative side of first principal axis. This sample is
enriched by a large amount of small-seeded legumes and Brassicaceae remains in comparison
with other samples in this horizon. Yet, sample no. 536 also differs from the others at the
positive side of second axis and this time appears at the positive side of first axis. The main
reason for this separation is the enrichment of the sample by the remains of Aegilops sp. The
rest of plant species in these samples appear proportionally identical.
The second CA plot in this research (Graphic 11) has more explanatory power than the first
plot. It distinguishes 59.5 % of the cumulative variance of species data in the assemblage in
the first axis (horizontal) and 71 % in the second axis (vertical). In total, the four principal
axes explain 88.8 % of the cumulative variance. In this plot, all samples from each

47
archaeological horizon were amalgamated into one sample and additionally species data were
inserted to identify the variations layer-by-layer and species-by-species.
One feature of this plot clearly shows a separation in the first principal axis concerning
probable food plants with large seeded pulses (lentil and vetch/grass pea) and wild barley
grains and rachis remains. It is highly possible that this distinction could be related to food-
processing activities that contributed differently to the depositional history.
A second feature of the assemblage expressed in the first principal axis is the distinction
between large-seeded and small-seeded pulses. Small-seeded pulses strongly cluster with
plant taxa from the Caryophyllaceae, Chenopodiceae, and Brassicaceae families rather than
the large-seeded pulses. On the other hand, large-seeded pulses display a sort of closely
clustering with Centaurea type, Medicago radiata, Aegilops sp. grain and rachis remains,
Malva sp., Scirpus sp., and the Ornithagalum/Muscari complex. This may indicate a different
source of patterning rather than crop processing residues, or food preparations may have been
prominent for large- and small-seeded pulses.
The two lowest layers (AH XI and X) tend to be associated more with the cluster of small-
seeded pulses, pistachio, Chenopodiceae, and Caryophyllaceae remains and to locate at the
negative side of the axis. The next four horizons (AH IX, VIII, VII and VI) exhibit a more
centroid distribution on the plot, closely clustering at the positive side of the first principal
axis.
The second principal axis separates Agrostis type and Phalaris sp. from the rest of species
dataset. These two taxa are closely associated with layers AH V and IV. The final three layers
(AH III, II, and I) tend to cluster at the positive side of second principal axis in contrast to AH
V and AH IV. The large amount of Aegilops sp. finds in AH III makes this layer different
from the next two subsequent layers but if this taxon is excluded AH III tends to cluster more
closely to AH II and AH I.

48
Graph 10: Correspondence Analysis plot for the compositional variations of analyzed samples. The first
letter on the sample point denotes AH number following by sample number assigned during the
excavations. The encircled samples represents that of explained in the text.
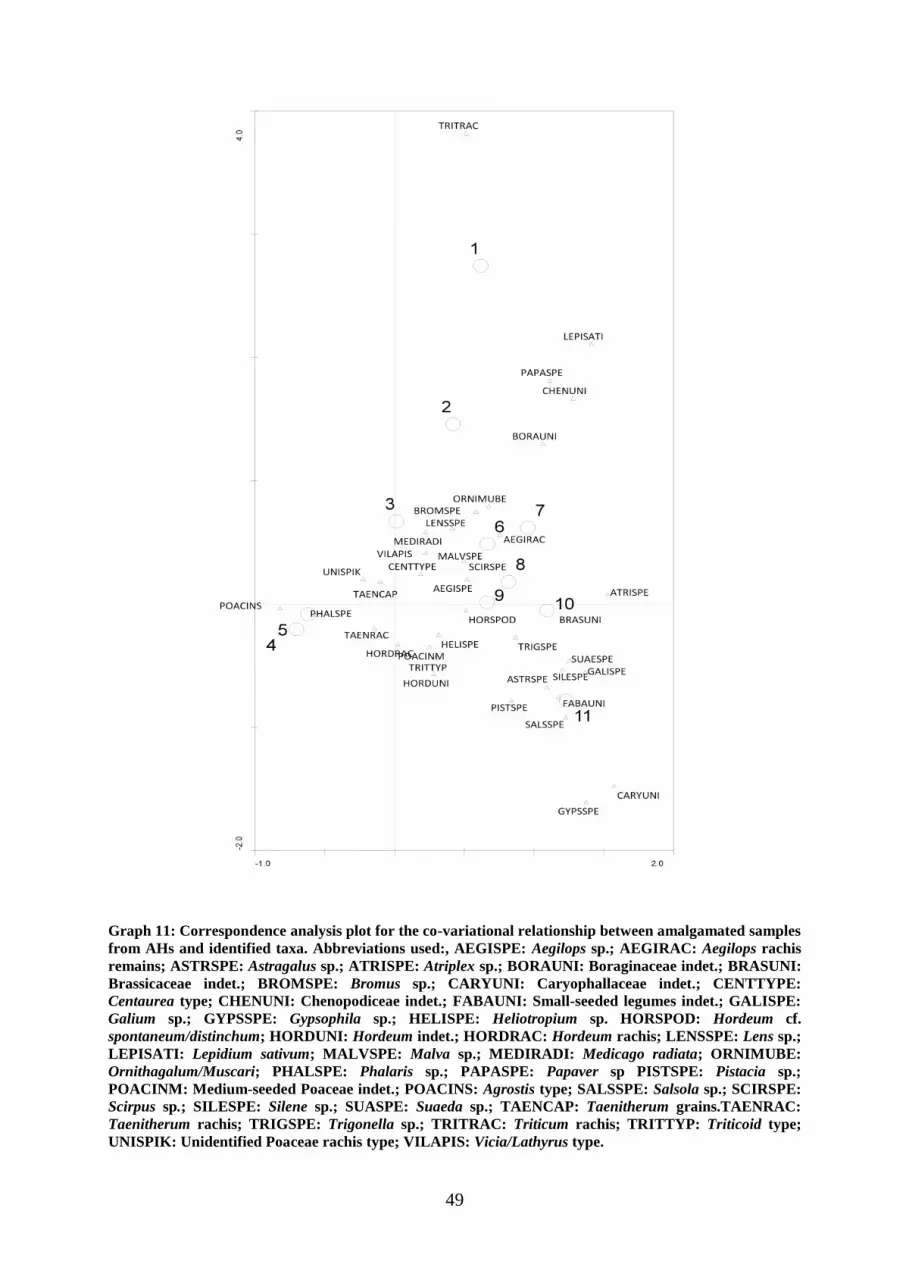
49
Graph 11: Correspondence analysis plot for the co-variational relationship between amalgamated samples
from AHs and identified taxa. Abbreviations used:, AEGISPE: Aegilops sp.; AEGIRAC: Aegilops rachis
remains; ASTRSPE: Astragalus sp.; ATRISPE: Atriplex sp.; BORAUNI: Boraginaceae indet.; BRASUNI:
Brassicaceae indet.; BROMSPE: Bromus sp.; CARYUNI: Caryophallaceae indet.; CENTTYPE:
Centaurea type; CHENUNI: Chenopodiceae indet.; FABAUNI: Small-seeded legumes indet.; GALISPE:
Galium sp.; GYPSSPE: Gypsophila sp.; HELISPE: Heliotropium sp. HORSPOD: Hordeum cf.
spontaneum/distinchum; HORDUNI: Hordeum indet.; HORDRAC: Hordeum rachis; LENSSPE: Lens sp.;
LEPISATI: Lepidium sativum; MALVSPE: Malva sp.; MEDIRADI: Medicago radiata; ORNIMUBE:
Ornithagalum/Muscari; PHALSPE: Phalaris sp.; PAPASPE: Papaver sp PISTSPE: Pistacia sp.;
POACINM: Medium-seeded Poaceae indet.; POACINS: Agrostis type; SALSSPE: Salsola sp.; SCIRSPE:
Scirpus sp.; SILESPE: Silene sp.; SUASPE: Suaeda sp.; TAENCAP: Taenitherum grains.TAENRAC:
Taenitherum rachis; TRIGSPE: Trigonella sp.; TRITRAC: Triticum rachis; TRITTYP: Triticoid type;
UNISPIK: Unidentified Poaceae rachis type; VILAPIS: Vicia/Lathyrus type.

50
VI. DISCUSSION
The next sections aim to discuss the results derived from the analysis of the Chogha Golan
plant assemblage followed by questions formulated during this research. The first section
summarizes and discusses the current state of archaeobotanical, archaeological evidence from
Chogha Golan. This is designed to answer the first research question, whether there are
variations in the composition of floral assemblage of Chogha Golan.
The subsequent sections discuss the Chogha Golan dataset from the perspective of the
similarities and dissimilarities of this site‟s floral compositions in relation to other sites in the
region. Its aim is to find out any pattern in the plant assemblages that would be connected to a
general development towards the establishment of farming economy. First the theoretical
background of changes in plant management strategies and the archaeological sites in Eastern
Fertile Crescent will be considered in detail; then the recent discussions on the fixation rate of
domesticated phenotypes and geographical origins of domestication will be reviewed to
pinpoint the temporal and spatial extent of domestication and farming economy at eastern
Fertile Crescent. ,
VI.1 Variations in the Chogha Golan plant assemblage
The composition of the lowest layer, AH XI, is characterized by the highest percentage of the
Fabaceae family with ~42 %, and the lowest recorded for Poaceae with ~44 %. Moreover, the
proportions of small-seeded pulses compose ~40 % of the dataset. In this respect, AH XI
represents an unusual figure compared with the succeeding layers. Zeidi (pers. comm. 2012)
reports regarding the mixture of knapping debris with debris from fireplaces that “this layer
seems to be an unusually rich midden deposit”. Neither bone remains nor flints have any sign
of contact with fire. In short, this layer suggests a complex pattern of accumulation including
many phases of deposition.
AH X represents a divergence from the previous layer in showing increasing values of grasses
at the expense of pulses but interestingly the contribution of other plant families is as high as
it is in AH XI. Nevertheless, the composition of plants that are neither grasses nor pulses is

51
different from the uppermost layer. This difference is characterized by the increase of the
Brassicaceae and Chenopodiceae families and the decreasing contribution of the Pistacia
genus and the Caryophillaceae family. This layer displays the first appearance of mud-brick
walls and floors plastered with red ochre. There are also indications of a fixed in situ pecked
boulder and probably mortars as well (Zeidi pers. comm. 2012).
AH IX shows an increasing trend for Poaceae and decreasing values for other plant families.
The find density declines for all organic and inorganic remains. The composition of the plant
assemblage shows that barley seeds and chaffs, lentils, Vicia/Lathyrus type, Agrostis type,
and mallow are higher in comparison with the two lower layers. This layer is defined as the
probable remnants of building debris (Zeidi pers. comm. 2012).
In AH VIII, a sudden increase becomes evident in the percentages and density of Poaceae
remains. This change can be seen chiefly in the increase of two categories in the assemblage:
the chaffs of Aegilops sp. and the unidentified Poaceae spikelet bases. Meanwhile the other
categories of grasses and pulses decrease in relation to earlier layers and only Astragalus sp.
shows a small increase in percentages at the expense of other small-seeded pulses. It is
reported that the find density of archaeological remains is high. The excavators also reported
that very thin ash lenses are apparent in this layer (Zeidi pers. comm. 2012).
Interestingly, AH VII displays a sudden decrease in Poaceae percentages, which is atypical
given the generally increasing trend of Poaceae remains in the assemblage. The remains in
this layer include a sudden occurrence of Triticum sp. chaffs in high counts. This figure could
be important because, with this exception, such high counts were not recorded in the
assemblage until AH III. However, it should be noted that the sudden occurrence of Triticum
sp. chaff remains in this particular layer may be related to the contextual variation among
sampling units. The composition of other samples coming from subsquares 0/99 and 1/99
does not contain as large an amount of Triticum chaff remains. The excavators reported that
the sample that has this high amount of wheat chaffs also includes human skeletal remains.
AH VI is characterized by the rapid increase of Aegilops sp. and Agrostis type in the
assemblage, with high density values. On the other hand, Papaver sp., which is not recorded
in any other layers and samples, is present in AH VI. The sediment in this layer is reported to

52
be very soft and silty, consisting of substantial amounts of charcoal, ash, and bone fragment
as well as river pebbles. The pisé wall from the previous horizon penetrated into AH VI.
The next two succeeding layers, AH V and AH IV, show a constant increase of the Poaceae
remains, which could be owing to the extremely high density and proportions of small-seeded
grasses (both Agrostis type and Phalaris sp.). Interestingly, the density and percentages of
Hordeum cf. spontaneum chaffs become higher than Aegilops sp. chaff remains in AH V and
AH IV. This rapid boost in the density of these findings is accompanied by the relatively
small but still apparent increase of larger-grained Fabaceae seeds in AH V at the expense of
small-grained pulse genera.
The uppermost three layers, AH III, AH II, and AH I, do indicate a decreasing pattern of
grasses in contrast to the other families. Hence, the composition of the Poaceae assemblage in
the uppermost layers is different from others in respect to the frequent occurrences of
Triticum sp. chaff remains. Among all the categories tabulated in the percentage analysis,
only Triticum sp. chaff remains show continuous increases from AH IV to AH I.
AH III shows the highest densities for Aegilops sp. chaff remains as well as the sudden
decrease of Agrostis type seed remains. This layer also differs from other layers with the
occurrence of one rachis specimen, most probably belonging to a free-threshing type, and
another grain specimen identified as wild einkorn or emmer. Archeologically, this layer
shows several thin plaster floors on top of each other. Zeidi (pers. comm. 2013) also reports
burnt surfaces, mud brick features with straw temper, and an impression of matting made of
reed that is visible on clay lumps.
In AH II and AH I, the archaeological evidence recorded several grinding and pounding
implements that were classified as hoes, mortars, grinding slabs, handstones, pestles, and
pounders. Most of the chipped stones represent debitage. Sickle blades are present but not
very numerous in the lithic assemblage; in total, 13 blades and 17 bladelets with sheen were
recovered from these layers. It was reported that traces of natural asphalt were found on one
edge of some blades. The main architectural features are chineh walls and associated stone
structures and mortars. AH I has mud brick structures; this layer shows partially burnt,
trampled floors that seem to have been renewed several times, visible as different layers that
average 2-3 cm thick.

53
The floral composition from AH II clearly indicates the rapid increase of certain plant
taxa/genera, namely, those that belong to small-seeded pulses and other wild plants, at the
expense of diminishing percentages of grass finds such as Aegilops sp. chaffs and Agrostis
type. Among the grasses, only two categories display a significant increase, Triticum sp.
chaffs and Phalaris sp. seeds, while new plant taxa like Brassica rapa type and
Rumex/Polygonum type are first recorded in this layer, although they exist in small counts.
In contrast to other layers that include plentiful archaeobotanical findings, AH I has a very
low count of remains (n=87) in two samples examined. This low amount of singular finds
raises questions as to whether the findings from AH 1 are sufficiently representative for the
accurate evaluation of this layer. However, the same spectrum of frequently-occurring taxa
found in previous layers is still present in this layer, together with the apparent high
percentage of wheat chaff remains.
Another important issue is that wheat spikelet base remains fall outside of the main clusters in
the CA plots which recognizes wheat rachis remains as an outlier. It is quite possible that the
samples from AH I might indicate a primary deposition rather than having been discarded to
this context from an original one. All in all, the distribution of plant taxa on two CA plots is
highly homogenous and does not primarily suggest a clear patterning for different formation
processes such as crop processing activities as had been argued in van der Veen (2007). The
lack of spatial data from Chogha Golan also limits the interpretation of probable causes for
various routes of entry for plant materials.
In the mean time, many herbaceous taxa had been defined as an element of steppic vegetation
cover also occurs in Chogha Golan assemblage. Leguminous genera such as Astragalus,
Trigonella, Medicago are common steppe plants (van Zeist et al. 1984). Similarly, the taxa
defined under the family Brassicaceae are also common constituents of steppe vegetation.
Furthermore, the large variations of grass taxa demonstrate that the natural vegetation cover
included those taxa of steppic associations. Helbaek observed that the earliest layer of Tepe
Ali Kosh contain mostly drought-resistant and salt-tolerant species which is also true for
Chogha Golan as the taxa coming from two plant families, namely Caryophyllaceae and
Chenopodiceae occur proportionally high in the lowermost layers. More detailed
investigations needed to be done for the ecological significance of the distribution of plant
taxa over different AHs, which remain obscure in the current research.

54
VI.2 Evaluations of plant management strategies in eastern Fertile Crescent
According to the most recent archaeobotanical and archaeological evidence, the process of
subsistence change from true hunting-gathering to farming-herding was a gradual transition.
The conceptual framework of the classification of the plant exploitation systems confers three
evolutionary stages6 of food procurement and prodaction. Harris (2007) and Zvelebil and
Dulokhanov (1991) proposed basicly similar approaches to explore the developmental route
toward resource specialization of harvesting seeds of wild grasses and other herbaceous
plants. Those modes are wild plant-food procurement (foraging as the principal plant
subsistence mode), wild plant-food production (cultivation of crop plants and foraging
strategies together) and agriculture of domesticated crops (farming as the principal mode of
subsistence).
This predictive model comprises two aspects of early human-plant relationships. First, it is the
recognition of human communities with “low-level food production” that encompasses
temporally-extended plant management strategies between a hunting-gathering subsistence
economy and a farming-herding economy. A second feature for the transition to agriculture is
the intensification of production by generating increasing amounts of labor into the managed
landscape and followed by a predominance of domesticated plants (Harris 2007; Fuller 2007;
Smith 2001, 2007a, 2007b; Zvelebil and Dulokhanov 1991).
The communities with low-level food production are considered to be neither hunter-gatherers
nor agriculturalists; they relied on low-level production of domesticates with a heavy
6 Trigger (2006) rightly puts forward some major limitations of the role of law-like generalizations in the
archaeological explanation following the remarks of Murdock (1959 in “Evolution in social organization”); “[…]
Many evolutionary generalizations may be formulated inductively as a result of detailed efforts to interpret
individual cultural sequences and then raised to a higher level of significance after their cross-cultural
applicability is noted. Because of the overlapping nature of many competing high-level theories of human
behavior, it often remains unclear which of them best accounts for such empirical generalizations. It may further
be argued that the ultimate task of evolutionary theory, and the standard by which it must be judged, is its ability
to explain what has happened in the past, as revealed through idiographic studies, rather than to construct
hypothetical schemes of development that are invariably too general to predict what actually happened in the
past.”

55
emphasis on using and management of wild resources. This assumption contrasts to Childe‟s
dualistic approach, which posited a radical and rapid shift between two static stages, with no
turning point from farming into other forms of subsistence strategies. Such “middle ground
territory” between two modes of subsistence has long been recognized in studies on the
emergence of agriculture in Mexico. The temporal-developmental distances between
domestication and the emergence of the first agricultural communities is separated by 5.500
years in Mexico characterized by the temporal gap between the domestication of squash
(Cucurbita pepo) and the beginning of the village-based communities with maize-beans-
squash farming (Flannery 1973; Smith 2001). Furthermore, Zvelebil and Dulokhanov (1996)
and Smith (2001) stress that these communities should not be seen as incipient or transitional
reference points; as once Braidwood presumed; in the evolution of agricultural life but rather
as successful and appropriate solutions to local environmental settings.
Human efforts to shape local biotic communities use different forms of niche construction or
ecosystem engineering activities such as controlled burning of vegetation or management of
wild plants by tilling, tending and sowing. Although archaeological records are scarce for
active human intervention to the landscape, this has been documented in the North American
Southwest in 550 individual locations where agave plants were transplanted by Hohokam
communities (A.D. 600 – 1350). These agave cultivation sites include archaeological features
for water manipulation, rock pile complexes around individual plants, discarded processing
tools, roasting pits and relict agave populations (Smith 2001). Smith (2007a) further reframes
the nature of niche contruction as follows “[…] all of these different activities comprised an
integrated and traditional resource management strategy of direct and sustained manipulation
of a broad array of culturally significant populations of plants and animals and their habitats
in order to maintain their abundance, productivity and diversity”.
With regard the archaeological research in the Near East, Asouti and Fuller (2012) infer a
similar developmental route through interpreting well-tuned archaeological, archaeobotanical
and chronological data in order to explore the changes in plant management strategies and the
subsequent prevalence of fully domesticated “crop packages”. In their interpretation of the
comparative dataset from southern Levantine PPN sites they cautiously note that, “full
domesticated status was assigned where non-shattering rachis remains represented the
majority of the assemblage, or they were found at late sites alongside significantly enlarged
grains”. Their regional analysis concludes that the intensification of subsistence production

56
(the appearance of fully domesticated crop packages) may have happened after the
demographic aggregation that occurred in the context of LPPNB megasites in southern
Levant. Prior to this phenomenon, what is largely seen is that “local early PPN traditions of
plant management” (Asouti and Fuller 2011) in which it is proposed (Fuller et al. 2010) to
exist a balance of differing selection pressures on cultivated and wild populations that allowed
for the coexistence of both natural shattering populations and populations under domestication
pressure in the same environment.
Both Hole (1984) and Harris (2007) recognized the very same assumption that the subsistence
strategies and habitation patterns during the PPN might have been more diversified and
permeable among each other than had been presumed on the basis of archaeological records.
Consequently, Hole (1984) argues that prior to the middle PPNB in western Iran, subsistence
economy may have depended heavily on different combined strategies like gathering and
herding or cultivating and hunting. For instance, Hole (1996) mentions that seasonal sheep-
goat transhumance could be a prominent subsistence strategy that it was herders-cultivators
who first introduce agriculture into the region and subsequently spread it through the Zagros
foothills. The harvesting profiles of faunal assemblage of Ganj Dareh also signal to active
management of caprine herds that the reexamination of goat remains show at this site
uncertain evidence of domesticated phenotypes but it appears that they were actively managed
to maintain an age-sex profile resembling to that of farmers-herders practiced (Zeder and
Hesse 2000; Zeder 1999). The archaeological records on the rapid increase of sedentism and
the number of known sites reflect that there was a convergence toward heavy dependence on
both farming and herding evidenced at Ganj Dareh Level D, the lower phases of Tepe Guran,
the Bus Mordeh phase of Tepe Ali Kosh and Çayönü (Hole 1984).
Bearing in mind Harris‟s conceptual framework and related archaeological evidence, it is
possible to identify certain similarities and dissimilarities in the development of plant
management strategies among western Iranian sites regardless of the rarity of
archaeobotanical records. Comparative evaluations of plant remains from three relatively
well-studied sites; Chogha Golan, Tepe Ali Kosh (Helbaek 1969), Ganj Dareh (van Zeist et al.
1984) were able to demonstrate that on theoretical grounds, that there are similar site-specific
trajectories in Western Iran akin to that of Harris‟ model in many aspects. This trajectory is
also basically concurrent with the early management of wild plants and the beginnings of
cereal domestication over much of the Fertile Crescent in terms of the empirical data retrieved

57
during the last decade (Willcox 2004; Fuller 2007; Purugganan and Fuller 2009; Fuller et al.
2011).
The comparative evaluation of published data for three PPN plant assemblages in western Iran
exhibit that the lowest archaeological levels always consist of high proportions of small-
seeded legumes and other wild taxa that basically may reflect local food procurement
practices, while subsequent occupation phases display a decrease in small-seeded pulses and
the increasing “visibility” of large-seeded taxa such as wild barley and wild lentil as occurs at
Chogha Golan and Ganj Dareh. This shift to heavy emphasis of large-seeded wild taxa might
have been the result of changes in plant management strategies from wild food procurement
to wild food production in the context of pre-domestication cultivation. Finally, following the
comparative data from Charles (2007), the archaeological levels that are inhabited from late
PPNB onwards such as Tepe Ali Kosh, Tell Maghzaliyeh, Jarmo, Chogha Bonut, Tepe Abdul
Hossein, are characterized by the abundance of morphologically domesticated emmer wheat
remains supplemented by domesticated barley and crop plants which signal the evidence for
the establishment of the agriculture in the region. The presumed developmental trajectory
during the Aceramic period in western Iran can offer a firm basis in order to compare the
developmental trajectories to better identify the temporal frame of probable changes in plant
management strategies.
Certain constraints of this interpretation should be taken into consideration. It is highly likely
that wild food procurement might always have been a simultaneous element in the subsistence
of PPN communities which might have been adopted along with other modes of plant
management strategies in varying arrangements in relation to local environmental settings,
seasonal changes or during the time of decrease in food resources. In the case of site occupied
for as long as Chogha Golan, it is not easy to gauge the importance of these remains by
analyzing their varying proportions. Even though a large amount of small-seeded legumes and
pistachio predominates in samples at the earlier layers, there is also sufficient evidence from
these levels that to some extent the early inhabitants of Chogha Golan simultaneously
exploited large-seeded grasses and pulses.
Contextual evidence from the earliest level of Ganj Dareh can provide helpful insights for this
issue. Although many samples contain large amounts of small-seeded legumes and pistachio
remains in the two lowest levels of occupation, wild barley must also have been utilized

58
intensively, judging by its dominance in a sample from one particular context. Accordingly,
Helbaek (1969) also recognizes two types of plant use that occurred simultaneously at Tepe
Ali Kosh that are based on both the heavy dependence on collecting wild, endemic legume
plants and the cultivation of both wild and domesticated types of wheat and barley. At Tepe
Ali Kosh the exploitation of endemic legumes appears to have been practiced more intensely
in the course of the earliest phase and again in the last phase of occupation, but as with
Chogha Golan and Ganj Dareh, it was always accompanied by large-seeded grass taxa like
emmer and/or einkorn and to lesser extent barley. Finally, the chronological significance of
these specific patterns of plant occurrences poses another problem. There is a large
chronological gap between the earliest habitation phases of Chogha Golan and Ganj Dareh
about 1.000 years and roughly 1.500 years with Tepe Ali Kosh occupation that creates
problem in incorporating the specific temporal range of plant occurrences into predefined
evolutionary stages that eventually requires well-established chronological data which was
missing in the current research.
Therefore, the simplification of the overall occurrences of certain plants into predefined
evolutionary stages should be approached cautiously and only for analytic purposes. For
instance, a wide spectrum of potential wild edibles can be found in the plant assemblages of
western Iranian sites and occasionally these may have contributed to the dietary practices.
However, the principal problem is that the presence of such wild taxa cannot be recognized as
direct evidence of either intentional harvesting or collecting from the wild unless they have
been found in large concentrations associated with primary depositional contexts (Hillman
2000, van Zeist et al. 1984). In the mean time, dung burning for fuel has been recognized as
an alternative taphonomic process in the formation of the archaeobotanical assemblage
(Miller 1984) and specifically for Western Iran sites (Charles 2007). Instead of interpreting
the plant remains only for the dietary importance, Miller (1984) and Miller and Smart (1984)
observed the fact that seeds and chaffs eaten by a range of animals can survive the passage
trough digestive system. They further proposed that accumulation of many seeds of non-
cultivated plants including hard-to-collect small-seeded legumes and so-called arable weeds,
may have resulted in dung burning as fuel and incorporated into the plant assemblage from
this route.

59
VI.3.1 Wild plant-food procurement
Contemporary hunter-gatherers such as the indigenous communities inhabited in temperate
regions of North America encounter higher levels of dietary diversity occupying resource
environments that are ecologically more varied. Harris (1977) proposes that broad spectrum
subsistence economies would possibly occur within generalized natural eco-systems where
there is a large variety of plant and animal species and populated by a small number of
individuals, especially in transitional vegetation zones such as on the margins of upland and
lowland mountainous regions or woodland and steppe. On the contrary, specialized
economies are likely to emerge where there are limited varieties of plant and animal species
but a large number of each species.
Hillman (2000) points out “the remarkably diverse diet of the Maidu of the Sacramento
Valley in California derived from a catchment that embraced open oak-dominated park-
woodland, grading below into a chaparral-grassland mosaic, the equivalent of the woodland-
steppe in Western Asia, with marshlands, rivers, and lakes below that”. Regarding his
paleoenvironmental reconstruction at Tell Abu Hureyra in 11.500 cal. B.P., Hillman (2000)
assumes a similar richness in dietary diversity resembling to that of indeginous communities.
Among 142 plant taxa in the assemblage, of these 118 were ethnographically identified plants
used as food sources among hunter-gatherers. It is proposed that such diversity in potential
foods was used to supplement the meat-based diet with plants that are rich in starch and oil,
and several of them were used as caloric staples. Adding up the probable uses of roots, green
shoots, leaves, flower buds, and other soft tissues that have little chance of being preserved
from charring, he further predicts that the total number of wild edibles consumed at Abu
Hureyra may have been over 250. This assumption would also be suitable for Chogha Golan
as considering the diversity of grasses and pulses found in the archaeobotanical assemblage.
Small seeded legumes
The small-seeded legumes comprise a large amount of plant species especially from the
genera of Trifolium, Astragalus, Medicago, Trigonella and Coronilla. Today they are a major
constituent of pasture, animal feed and green manure worldwide. These remains are usually
well-represented amongst the plant remains recovered from PPN sites. In general, seed pods
are not found intact but usually seed remains are present (Weiss and Zohary, 2011).

60
Helbaek‟s comments on endemic species gathered at Tepe Ali Kosh include various plant
species that are also present in the Chogha Golan and Ganj Dareh assemblages. The
characteristic feature of the lowest occupation phase (Bus Mordeh) is an extreme abundance
of “endemic legumes” (94,2 % of the assemblage), which consists a large number of
Trigonella species, two probable Astragalus species and certainly Medicago radiata. The
lower occupation phase of Ganj Dareh (Level E) also shows high ubiquities of small-seeded
pulses with decreasing values in the succeeding levels. Even though the small-seeded legumes
of Chogha Golan were only partially identified at the species level, the close association of
these “endemic legumes” with the lower levels of occupation of both sites needs further
consideration.
Hillman (2000) suggests that the accumulation of the small-seeded pulses at Tell Abu Hureyra
must have been the result of a deliberate activity, considering such a large number of findings.
It is worth noting that also in the Chogha Golan assemblage the find density values of small-
seeded legumes, including Astragalus sp., Trigonella sp., and the indeterminate small-seeded
legume category, follow the exact same gradient over different AHs, while Medicago radiata
differentiates from others with its sporadic occurrences throughout occupation period. This
species appears highly associated with Lens sp. and Vicia/Lathyrus type in the CA plot, which
could indicate that Medicago radiata might have charred as a result of another taphonomic
pathway (e.g. field weeds) or simply that its distribution was not as wide as other legume
species.
Butler (1995) discusses further possible routes of entry of these small-seeded leguminous
species into archaeobotanical plant assemblages. She considers these remains to be human
food resources rather than having been deposited in the form of dung, basing her argument on
the absence of domesticated flocks during the PPN, since wild species like gazelle leave their
dung in a dispersed manner that would not be time-efficient to collect. Materials used for
thatching, bedding or basketry are discounted, that is, generally not included among legume
plants. Alternatively, Hillman (2000) suggests food resources, flavorings, medicines, and dyes
as probable routes of entry for these remains. Helbaek (1969) and van Zeist et al. (1984) also
consider the probable role of small-seeded legumes as human food resources. Riehl (2013)

61
classifies some legume seeds7 from the Chogha Golan assemblage as arable weeds, which are
assumed to have been transported to the site along with the cultivated plants.
Ethnographic data suggests that there are a number of different uses for small-seeded
legumes, ranging from insect repellants, perfume, and honey plants to flavoring, coloring or
food preservatives to medicinal use and human food. For instance, the indigenous tribes in
North America use clovers (Trifolium sp.) as food resources that vary from season to season.
The seeds of clovers were rarely eaten by these communities because consuming the plant
parts as green vegetables is more common. Moreover, the seeds were eaten raw and
unprepared, when they were still in the green state8 that may represent a recent parallel to the
past gathering activities in temperate regions. Luomala (1978, after Butler 1995) reports that
the Tipai-Ipai tribe of southern California exploits two species of Trifolium genus for their
seeds which are collected into baskets using seed beaters, later on the seed are parched,
threshed and winnowed before being stored in covered pots (Butler 1995).
In the meantime, small-seeded legumes are also frequently represented in the assemblages of
Hallan Çemi, Demirköy, and Qermez Dere but in very small proportions, in contrast to sites
in western Iran. For instance, the fact that the ubiquity figures of small-seeded legumes show
high values, with about 50 % of all samples, illustrates the presence of these taxa, but their
proportions vary between 0,2 % and 7.4 % of the whole assemblage for each of these three
sites (Savard et al. 2003, 2006). This evidence demonstrates that small-seeded legumes are
7 In Riehl (2013) the classification of small-seeded legumes of Chogha Golan includes more plant taxa than
those are discussed in the present research. Additional legume taxa that are not successfully defined in this
research are Coronilla sp. and Medicago sp.
8 I personally also remember that during my childhood in Turkey in the 1980s chickpea seeds were sold together
with the whole plant in its green state, either in the market or by individual sellers from a cart on the streets. The
seeds were consumed raw, like eating nuts. I assume that this way of consuming chickpeas still continues today
in provincial towns in Turkey and that they can be easily found at farmers‟ markets. Luczaj et al. (2012) also
reports how children approach nature by collecting wild edible plants as so-called “snacks”, proposing that this
kind of collecting and consuming of wild food plants could be the true relict of once prevailing hunting-
gathering practices. The authors give examples from around Europe of children eating the immature seeds of
various Fabaceae species such as Lathyrus cicera, Vicia villosa and V. lutea as well as the flowers of Trifolium
spp. Finally, during a conversation with one of my acquaintances from western Iran, he informed me that his
family too usually ate wild cereal grains like nuts, after slightly parching them on the fire in order to easily
remove the grains from the enclosing glumes.

62
present but their contributions to the composition of the plant assemblage are not as
significant as at western Iranian sites.
Other probable wild edible plants
The highly nutritive fruits of the pistachio tree must have constituted a valued source of
vegetable fat for early inhabitants of western Iranian sites. As at Chogha Golan, the earlier
archaeological horizon of Ganj Dareh displays higher proportions while pistachio nutshells
contributed less in succeeding levels. Even though pistachio is the only arboreal fruit taxon
recorded at Chogha Golan and Tepe Ali Kosh, Ganj Dareh has wild almond nutshells, albeit
in low frequencies. Beyond this, no site shows evidence for the presence of acorns, except for
Jarmo in the late Aceramic (van Zeist et al 1984; Helbaek 1969; Charles 2007).
With regard to the dietary potential of the plant species of the Chenopodiceae family, Hillman
(2000) posits that these plants might have served as food plants during the second phase of
Abu Hureyra‟s occupation. Most plants produce their seeds in winter when other staple foods
may have been scarce. The seeds of the shrubby chenopods in particular have been intensely
collected by hunter-gatherers in North America. The Cahuilla tribe incorporated Suaeda
suffruticosa and other two species of Atriplex sp. in their diet; the use of seeds from
chenopods by the Seri and the Lakota has also been recorded ethnographically (Hillman
2000). The combined finds of seeds of various chenopods is mostly associated with earlier
levels of occupation at Chogha Golan. What is interesting is that the remains of chenopods
were not reported from either Ganj Dareh or Tepe Ali Kosh. On the other hand, no reliable
records for this group of plants exist for other sites of western Iran.
Scirpus maritimus has also had scholarly attention for its contribution as a potential food. The
nutlets of Scirpus are rich in starch and its incorporation into the human diet is evidenced by a
find of a human coprolite from Late Paleolithic Wadi Kubbaniya, Egypt (Hillman 2000). This
species never reaches high proportions at Chogha Golan, but it is a ubiquitous one and no
apparent proportional change exists in earlier and later horizons. At Ganj Dareh, the high
ubiquities of the remains of this species in earlier levels drop sharply in succeeding
occupation periods. Interestingly, the use of this species was more widely practiced at PPNA
sites at southeastern Turkey such as Hallan Çemi and Demirköy (32.1 % and 70 % of the
whole assemblages respectively) than at Iraqi and western Iranian sites. This sort of sharp

63
contrast can also be seen in the near absence of the Rumex/Polygonum complex at western
Iranian sites in contrast to the abundant finds at sites in southeastern Turkey and northern
Syria.
Aegilops sp. is widespread at sites all over the Fertile Crescent and is chiefly identified as a
weed taxon of cultivated cereal fields. Owing to its apparent mechanical difficulties in
separating the grains from its tightly enclosed glumes, the probable contribution of Aegilops
sp. type remains may have been misjudged in the literature, where it has been considered as a
noxious weed of cultivated fields. Keeping this in mind, ethnographic records from Europe
(Luczaj et al. 2012) demonstrate that Aegilops geniculata was used as a bread additive
alongside other less preferable plants during famines in 19th
century. It is highly likely that the
Aegilops species at Chogha Golan may have contributed to the human diet rather than being
merely a noxious weed infesting the disturbed grounds. The spikelet bases of this plant were
always found to be highly fragmented at Chogha Golan, which could be interpreted as the
processing residues of barley cultivation or an intentional activity directly aimed at
incorporating this plant into dietary practices. In short, one should certainly not reject the
possibility that Aegilops sp. contributed to the diet of the inhabitants of Chogha Golan.
The role of two types of small-seeded grass taxa remains obscure, that is to say, Agrostis type
and Triticoid type. Agrostis type remains frequently occur from those sites such as Chogha
Golan, Ganj Dareh, Tepe Abdul Hosein in western Iran. Ubiquity scores of this taxa display a
sudden increase after the lowermost layer at Ganj Dareh. The same pattern is also visible in
Chogha Golan assemblage that an increasing trend after two lowermost horizons and another
suspicious increase in AH V and AH IV makes up approx. 57 % and 64 % of the assemblage
composition respectively.
Riehl et al. (2013) tentatively interpret this pattern as a shift in subsistence economy. When
looking at Ganj Dareh dataset, it is evident that there is a contrast between Hordeum finds and
Agrostis type finds indicating that contextual evidence could be significant to determine the
status of this plant small-seeded taxa. Agrostis is most ubiquitous in the provenance group IV
(”other deposits” such as loose brown soil, occupation layer, room fill) while Hordeum mostly
associated the area of food preparations in the provenance group I which comprises hearts,
ovens and kilns. Additionally, this type of grasses reported from Tepe Abdul Hosein as the

64
most abundant plant finds in the site but no further evaluations has been made in relation of
these finds.
Triticoid type is also recorded in two other sites, Ganj Dareh and M‟lefaat, outside of Chogha
Golan. In any sites referred above, these remains do occur ubiquitously albeit in very low
proportions. Riehl et al. (2013) mentions this type of remains in respect to the evidence of the
already-high diversity in wild Triticum-type cereals in the local environment. Gale et al.
(2003) prefers to include this group into Triticum boeticum/Secale complex. Ganj Dareh data
indicates that this type relatively occur more such contexts like heart, kiln, oven where the
food preparations activities are centered around.
VI.2.2 Wild plant-food production
The eight founder crops of Near Eastern agriculture appear to form a well-balanced crop
package between cereals and pulses that complement each other with their nutritional and
agronomic properties. In the early Holocene, the wild grasses of the Fertile Crescent were
(and still are) highly productive, forming dense stands in their natural environments, and they
progressively served as important food sources for early Holocene communities (Miller
1984). Hillman (2001) mentions the reasons why the wild cereals and pulses were selected as
the first cultivated crops rather than other caloric staples. According to him, the emphasis on
wild cereals and pulses;
“[…] reflects the fact that, of the various staples, they were the only ones amenable to annual
cultivation on a large scale in densely sown stands, they produce a heavy yield (unlike perennial
equivalents), and were able to produce an easily stored product in a single season. Cereals also
yield secondary product: straw, that is useful as tinder, thatching and bedding, and that would one
day also prove useful as fodder for domestic herbivores”.
The nature of early human-plant interaction and the role of morphologically wild large-seeded
taxa in plant management strategies during the PPN is also widely discussed that the large-
seeded grasses and pulses were either collected from the wild as Kislev (2004) and
Ladizinsky (1987) proposed for grasses and pulses respectively or that they may have been
actively cultivated by humans without initially leading to domestication in the course of Late

65
Epipaleolithic (Hillman 2000, 2001) or PPNA (Weiss et al. 2006; Colledge 2002, Willcox
2012).
Early cultivation before the appearance of domesticated phenotypes of crop progenitors is
evident from an abrupt increase of probable arable weeds9 at the beginning of the PPN, as
well as some other markers such as indications of the heavy use of particular food plants
outside the area of natural distribution, shifts in specific patterns of wear on flint sickle-blades
and archaeological evidence of large sedentary populations that would not have been
supportable by hunting-gathering alone (Hillman 2000, 2001; Weiss et al. 2006). Willcox
(2012) suggests that the presence of seeds of presumed weeds becomes more and more
readily visible in the PPN floral assemblages that would most probably signal human-induced
vegetation changes in the environment through an intensification of cultivation10
. Some
scholars argue that the assumption of pre-domestication cultivation fits well to Harris‟ model
of the evolutionary continuum for plant management strategies as the stage of wild plant food
production (Fuller et al. 2012).
In many respects, the evolutionary relationship between human and weeds went hand-in-hand
with that between humans and cultivars. The early weeds were pioneers of secondary
succession and possessed adaptive properties such as high reproductive capacity or short life
cycles that allowed them to proliferate in disturbed grounds. A different set of adaptations was
imposed on the weed populations within an arable field through the selective pressures of
agricultural practices (Barrett 1983). For example, such selection for a weedy habit of Vicia
sativa and Lathyrus sativus in lentil cultivation enforced both plants to adopt adaptations to
9 Zohary‟s remarks on the modern distribution and origins of segetals and ruderals include valuable observations.
He argues that Near East is a center of speciation and more than 30 % of the weeds around the world are
endemic to the Near East and the largest centre of local weeds is eastern and southern fringe of Mediterranean
territory and the adjacent borderland with Irano-Turanian territory. Furthermore, according to him, there is a two
way traffic in the migration of the weeds that induce many typical Mediterranean weeds such as Trifolium,
Medicago, Vicia, Bromus, Erodium pushed eastwards into Irano-Turanian cultivated lands and many others
occurring among Eastern Mediterranean crops are Irano-Turanian by origin. It is most interesting to see that the
weeds which are associated with Irano-Turanian vegetation zone are missing in Chogha Golan assemblage.
10 A principal criticism of this assumption is that in many cases the identification of weed seeds remains at the
family or genus level. Given the species diversity of such families and genera in southwest Asia, only a few of
them are obligatory weeds (Abbo 2012). Hillman (2000) also recognized that the ecological and nutritional
characteristics of the presumed weeds are too broad to be restricted to cultivated lands as “weeds”.

66
agricultural cycles through loss of dormancy, early vigor, biomass production and
phenological adaptation (Erskine et al. 1994). In this regard, Harlan (1992; after Leonti et al.
2006) states that modern weeds presumably did not exist in their present form before
agriculture and following on this assumption, it is highly likely that the formation of
cultivated lands in the early Holocene landscape may not only have favored the first crop
plants but also consciously or unconsciously a wide array of “weedy” plants that had a broad
ecological tolerance and were well-adapted to newly emerging and relatively less-competitive
agricultural habitats.
It appears that arable weeds do constantly become the subject of intentional gathering among
traditional farmer communities11
. Many scholars have observed that wild gathered weedy
plants were widely utilized as foodstuffs and for medicinal purposes by both farmers and
hunter-gatherer communities (Leonti et al. 2006; Luczaj et al. 2012; Stepp and Moerman
2001; Abbo et al. 2012; Hillman 2000). According to their observations, the weeds of fallow
and cultivated fields constitute a large number of species gathered for their potential
pharmacological and physiological properties (e.g. toxic, antimicrobial, anti-flammatory, anti-
oxidant, appetizing, etc.) by agriculturalists and contemporary foraging communities (Leonti
et al. 2006, Steep and Moerman 2001). Therefore, Leonti et al. (2006) argue that the culinary
use of wild gathered weedy greens may have evolved together with neolithization process,
initiated through tolerating these weedy taxa in cultivated fields, because these taxa are more
fibrous than cultivated plants and contain higher concentrations of secondary compounds that
provide crucial chemicals to compensate pharmacologically active substances. This
ethnographically derived observation can give a new perspective to the role of presumed
weeds in the PPN. It is plausible that human intentionality may have played a central role in
selecting a wide array of other plants together with the early cultivars, eventually confining
both plant groups to the agricultural habitat in order to enrich dietary breadth and avoid a
heavy dependence on cultivated crop plants as well as to secure other crucial medicinal and
nutritional elements. This perspective accords with Smith‟s remarks (2001, 2007a, 2007b) on
11
Such observation can also be distinguished in the contrast between the total number of obligate weeds in the
Near East and the total number of obligate weeds without any ethnographically recorded use. Zohary (1973)
states that among nearly 1500 segetal and ruderal species in the Near East, 420 of them are obligatory weeds that
only flourish in cultivated fields. On the other hand, Willcox (2012) mentions only 19 taxa that are well-suited
for the interpretation of pre-domestication cultivation and have any textually and ethnographically defined use in
the literature.

67
the central role of human intentionality in the co-evolutionary relationships between human
and target species that eventually resulted in the intensification of human efforts to modify
local environments.
“[…] domestication quite likely occurred within integrated strategies of ecosystem engineering
based on a comprehensive storehouse of knowledge about local biotic communities that had
been acquired over hundreds, if not thousands, of years of direct experience (Smith 2007b).”
In addition to Chogha Golan, so far twelve archaeological sites have been proposed as that the
pre-domestication cultivation might have been practiced during early stages of the PPN
(Hillman 2000; Colledge 2002; Willcox 2012). Riehl et al. (2013) recently argue (and as it is
apparent in the current research) that crop progenitors such as barley, wheats, lentil and other
large-seeded pulses and also probable arable weeds12
are present in the Chogha Golan
assemblage, which signifies that such an early cultivation of morphologically wild taxa might
have been performed by the inhabitants. Wild barley and lentil are particularly important early
grain crops, considering their widespread occurrence at PPN sites all over the Fertile
Crescent. Weiss and Zohary (2011) note this aspect of these two plants by defining them as
“pioneer crops” because their cultivation seems to appear somewhat earlier than others in the
Fertile Crescent. The probable role of Vicia/Lathyrus type in the subsistence economy will
also be considered in detail.
Wild barley (Hordeum spontaneum)
Today, wild barley (Hordeum spontaneum) is massively spread over southwest Asia, where
the Fertile Crescent constitutes its primary habitat. It is considered as a part of the rich grass
cover associated with the open woodland Quercus brantii belt that occurs at elevations from
500 to 1.500 meters in the Zagros region of Iraq and Iran. It does not tolerate extreme cold
conditions while it withstands drier and warmer environments, poorer soils, and some salinity.
Bor in Rechinger (1970) stresses the weedy habit of this species as well, noting that it thrives
12
The arable weeds of Chogha Golan as identified by Riehl et al. (2013) following the list of arable weeds in
Willcox (2012) includes following taxa; Trigonella sp., Silene sp., Reseda luteola, Ornithagalum/Muscari,
Medicago radiata, Malva sp., Lithospermum sp., Heliotropium sp., Gypsophila sp., Galium sp., Fumaria sp.,
Erodium sp., Coronilla sp., Centaurea sp. and Adonis sp.

68
in “deserts, gravelly, sandy or silky soils; in waste places and invading cultivation”.13
Nevo et
al. (1986) and Harlan and Zohary (1966) state that there are several wildraces of wild barley
in the Fertile Crescent, and that robust plants with large spikes and big seeds occur in more
mesic environments like oak woodland belt, while a special wadi race has slender and smaller
forms and grows mostly in elevations from about 600 meters above sea level to 350 meters
below sea level. Another race of an intermediate type thrives in arid steppes or at the edge of
plains.
On the geographic origin of barley domestication and early research by Badr et al. (2000)
inferred that barley was only domesticated once in the Levant. However, certain
methodological problems arise from this research, relating to sampling that is flawed through
the overrepresentation of Israeli wild accessions (Abbo et al. 2001). The most recent
molecular evidence (Morrell and Klegg 2007) exhibits two domestication events for wild
barley; one occurred on the western barley wildrace in the Levant and, based on their analysis,
the other originated approximately 1.500 to 3.000 km farther east. This evidence suggests that
the western foothills of Zagros or a locale farther east in central Iran may well be a center of
barley domestication, as this eastern wildrace contributed most of the diversity in barley from
Central Asia to the Far East. Jones et al. (2008, after Fuller et al. 2011) recently distinguished
photoperiod insensitive barleys that are better adapted to northern latitudes [and] share a
genetic mutation that derives uniquely from wild barleys of the mountains of Iran and not
from post-domestication mutations. This evidence suggests that there should be at least three
source populations from domesticated barley phenotypes.
The grains of wild and domesticated barley from the sites in western Iran are frequently
occurring. A marked increase in the proportions of wild barley grains is apparent after the
lowest level at Ganj Dareh as also recorded at Chogha Golan and is simultaneous with the
decrease in ubiquities of small-seeded legumes at both sites. Wild barley is also the only
large-seeded cereal species in the assemblage of Ganj Dareh and it is also ubiquitous. Van
Zeist et al. (1984) define the two types of barley grains in their account as Hordeum
13
Interestingly, according to Nevo et al. (1986), the allozymic variations of wild barley in western Iran display a
high level of genetic diversity that varies in relation to certain factors such as altitude variations and
ecogeographical and climatic variables. The population of genetic variations of wild barley specimens collected
from both the Amirabad plain and the town of Ilam exhibits a higher genetic diversity than the mean value
recorded for most wildraces and landraces from other regions in western Iran.

69
spontaneum-type and Hordeum distichum-type. Additionally, they report no marked increase
in the proportions of the two-rowed domesticated distichum-type remains at the expense of
the two-rowed wild spontaneum-type throughout the occupation period as well as few rachis
internodes were present in the assemblage. That is why the authors refrained from drawing
conclusions about the domestication of barley in the absence of firm evidence from the
examination of rachis internodes.
Considering later sites such as Tepe Ali Kosh and Tepe Abdul Hossein, it is also hard to
evaluate the status of barley domestication because of the poor evidence of barley rachis
internodes and the lack of quantitative data. However, Helbaek (1969) reports wild barley
(Hordeum spontaneum) in quantity from the earlier level on and six-rowed hulled forms
(Hordeum vulgare), identified through their twisted grains, appear with lower counts later
during the occupation but hegives no information about the rachis internodes of his barley
findings. On the other hand, his remarks match the temporal development of plant
assemblages in that the six-rowed forms appear late in the middle of final phase (Mohammad
Jaffar) of the Tepe Ali Kosh occupation, which is close to the end of the Aceramic Neolithic
in western Iran. Hubbard (1990) notes that there are not enough rachis internodes of barley
recovered from Tepe Abdul Hossein to make a judgment. But, he argues that the barley grains
are two-rowed domesticated barley (Hordeum distichon) and that this is the only barley type
found at Tepe Abdul Hossein.
The rachis internodes of wild barley from Chogha Golan comprise a significant portion of the
composition as recorded in Graph 3 in the current research. The appearance of chaff remains
in high quantity is rather dissimilar in comparison with the other sites in western Iran such as
Tepe Ali Kosh, Ganj Dareh, and Tepe Abdul Hossein. In terms of context, chaff-rich
assemblages are usually encountered from secondary and tertiary depositional contexts such
as refuse deposits pits, ditches, hearths, dung, and fuel (van der Veen, 2007).
On the other hand, no apparent changes have been detected regarding the probable increase of
non-shattering phenotypes throughout the occupation period at Chogha Golan. Only 1 % of
all the wild barley rachis remains indicate that non-shattering phenotypes did not usually
prevail in the environment. Furthermore, Kislev (2004) documents that in nature
approximately 10 % of barley rachis internodes may show rough articulation scars owing to

70
their position on the stalk. It has become a standard criterion in archaeobotany to assume
cereal domestication over this limit.
In this aspect, the Chogha Golan assemblage shows a close resemblance to many other PPN
sites in regard to the fixation rate of domesticated phenotypes in the environment, that is to
say, the frequent appearance of brittle rachis remains of cereals in high counts rather than
non-brittle rachis is a widespread phenomenon that occurs at several early PPN sites in the
Fertile Crescent. Tanno and Willcox (2006, 2012) illustrate a slow increase of non-shattering
rachis remains of barley and emmer/einkorn recovered from six PPN sites. Fuller et al. (2011)
also show that the time frame of the domestication process should extend for as long as 1.500
years in emmer and einkorn and 2.000 years in barley through compiling a comparative
dataset from all over the Fertile Crescent. This allowed the authors to suppose that weaker
selective pressures cause a rate of phenotypic change for non-shattering types of around 0.03
– 0.04 % per year in wild populations under domestication pressure (Purugganan and Fuller
2009; cf. Abbo 2012, Hillman and Davies 1990).
It is thought that various types of harvesting methods can retard the fixation of this adaptive
trait (Hillman 2001; Hillman and Davies 1990). For instance, harvesting by sickle-reaping or
uprooting would favor those mutant phenotypes with a non-shattering trait, but harvesting by
beating into baskets – which is the type of harvesting usually recorded among contemporary
North American hunter-gatherers and also apparently a more productive way, as experimental
research shows in Hillman (1990) – would favor the types already about to shatter.
Nevertheless, Hillman (2001) argues that “even after the potentially domestication inducing
methods of harvesting were applied; several other factors might have slowed dramatically the
fixation of domestic traits. These include the effects of harvesting crops that were
incompletely ripe; wet weather during harvesting time; introgression from nearby stands of
wild cereals of the same species (then much more widespread than they are today); and
modifier genes” or, as Kislev noted (2004), gathering disarticulated dispersal units from the
ground may have prolonged this time frame considerably. Also, some other researchers have
suggested the constant gene flow into proto-domesticates through bolstering the stored
harvests with collected wild grains as the principal reason for a slow rate for the fixation of
the non-shattering rachis trait (Fuller 2007; Purugganan and Fuller 2009; Fuller et al. 2010;
Fuller et al. 2011; Asouti and Fuller 2011; Fuller et al. 2012a). White and Makarewicz (2011)
note that distinct markers on barley rachis internodes can indicate a practice of harvesting the

71
barley ears when they are still partially in the green state. The experimental harvesting of
unripened barley ears by hand showed that a considerable proportion of the barley internodes
(% 22) exhibit a domestic scar or a “ripped” scar that extends down the ventral surface as if
they were ripped off of the ear. Similar markers on barley rachis internodes also exist at
Chogha Golan.
On the other hand, the comparative data from carbonized grain measurements shows that an
increase in grain size precedes the fixation of non-shattering types in the archaeobotanical
record. Fuller et al. (2012b) discovered that the breadth of wild barley increased about 50 %
during the course of domestication. Relatively faster fixation in grain size and shape points to
a 500 – 1.000 year time frame in archaeological records, while the non-shattering trait became
fixed over about 1.000 – 2.000 years (Willcox 2004, 2006; Fuller 2007; Purugganan and
Fuller 2009; Fuller et al. 2011). However, both Willcox (2004) and some other scholars
(Weiss and Zohary 2011; Abbo et al. 2012) approach this evidence cautiously because such
an increase would impose some other factors such as a plastic response to improved soil
conditions, the participation of different varieties, or simply a founder effect.
Wild lentil (Lens culinaris)
Lentil is a characteristic companion of a cereal-based diet in Southwest Asia. What makes it
so appreciated by the Near Eastern farmers is its protein content (about 25 %), which
constitutes an important meat substitute along with other pulses. In nature, wild lentil grows
primarily on shallow stony soils and on gravelly hillsides in open habitats. It also enters
disturbed localities such as stony patches near orchards and cultivated fields of cereals,
especially barley. It usually forms small scattered colonies in the wild, but some populations
are rich, comprising hundreds of plants together. The species occurs frequently side-by-side
with annual vetches, clovers, medics, and grass peas in their primary habitats (Weiss and
Zohary 2011; Abbo et al. 2009; Sonnante et al. 2009).
The first signs of domestication for lentil and pulses in general are the retention of the seed in
the pod (pod indehiscence) and the loss of germination regulation (seed dormancy). The
examination of chloroplast DNA restriction patterns indicates that lentil might have originated
from the regions around southern Turkey and northern Syria, but this evidence is not definite.
Ladizinsky (1987) suggests that wild lentil might have undergone a stage of intensive

72
collection by humans through which a strong selective pressure on the plant allowed it to
mutate and lose germination regulation (seed dormancy) before the beginning of its
cultivation. Ladizinsky‟s assumption has been challenged by Zohary (1989), whose
experimental research on wild lentil cultivation demonstrated that wild lentil reacts rapidly to
the improved soil conditions in cultivated fields, boosting the productivity of individuals up to
70 seeds, as opposed to an average of about 10 seeds in natural conditions. On the other hand,
even under present-day traditional farming practices in the Near East, farmers tend to harvest
domesticated pulse crops after most pods have attained full maturity but before the crop has
completely dried up in order to avoid the potential seed losses from dehiscent pods (Abbo et
al. 2009).
This plant displays widespread occurrences in most of the PPN sites and is always
encountered together with emmer, einkorn, or barley. Among the richest sites are Tell Aswad,
Tell Abu Hureyra, Jericho, and Çayönü. A large hoard of carbonized lentils (ca. 1.400.000
seeds) that was contaminated by a typical weed of lentil cultivation (Galium tricornutum) was
retrieved from the MPPNB site of Yiftah‟el. This evidence indicates that lentil was already
under cultivation in northern Israel during this period. The available archaeobotanical
information indicates that wild lentils must have been a valuable food resource for PPN
communities (Weiss and Zohary 2011).
In western Iran, one striking feature of the Tepe Ali Kosh assemblage is the rare occurrences
of lentil throughout all occupation phases; Helbaek notes that these few lentil finds must be a
small-seeded race of wild lentils endemic to the Zagros Mountains. The presence of lentils is
firmly evidenced in the Ganj Dareh account, albeit in low quantity. Van Zeist et al. (1984)
observed that the size of lentils corresponds with that of wild Lens orientalis as well.
M‟lefaat, Qermez Dere and Nemrik also show evidence of wild lentil in the assemblages
(Charles 2007; Savard et al. 2006). In Neolithic Tepe Sabz, which was settled on the Deh
Luran Plain after the abandonment of Tepe Ali Kosh, the size of lentils is reported to be as
large as that of domesticated lentils, 4.2 mm in diameter.
The rates of lentil size increase are equivalent to those of wheats and barley. According to
Fuller et al. (2011, 2012b), a time frame of 2.000 years is proposed to capture a high rate of
directional change, comparable to the rates of barley and einkorn, following which a slower
rate toward an increasing lentil seed size continued for the next 2.000 years over the course of

73
the Neolithic period. Over the entire domestication period, the total amount of seed size
change for lentils is over 50 % (Fuller et al. 2012b). The seed size measurements of Chogha
Golan lentils (n = 57) display varying sizes between a range of 2.76 and 1,95 mm that
corresponds to Helbaek‟s and van Zeist et al.‟s remarks on a small-seeded race. The results
display an interesting progression: the lowest layer contains relatively large-sized grains, after
which the size suddenly drops in the next layer and then enlarges constantly up to AH III,
when it reaches values similar to those of AH XI. The next two layers contain so few lentils
that no measurements were taken.
Vicia/Lathyrus type (vetch/grass pea)
The recognition of the lost crops or “false starts” opens a valuable discussion of the possibility
that the collection of the classical eight founder crops may be only a relic of a large number of
taxa that were genuine crops in early cultivation in certain locations at certain points in time
(Fuller et al. 2011; cf. Abbo et al. 2013). Apart from Vicia ervilia (bitter vetch), which is
within the classical eight, three of these lost crops may be present in the Chogha Golan
assemblage: Vicia peregrina (rambling vetch), Vicia faba (broad bean), and Lathyrus sativus
(grass pea). The assemblage may also contain cultivated Vicia species like Vicia sativa
(common vetch). On the other hand, Zohary and Hopf (2000) argue that “since there are no
reliable diagnostic traits by which wild and cultivated forms of bitter vetch cannot be
distinguished from each other”. This situation is also prevalent for grass pea finds so that it is
difficult to determine whether they represent cultivated forms or collection from the wild.
In recent times, pulses like Vicia ervilia, V. sativa, and Lathyrus sativus appear as tolerated
weeds in traditional farming systems (Abbo et al. 2009) and they are always associated with
fodder and famine food, but it is also well-known that they can be consumed as food after
detoxification (Melamed et al. 2008; Valamoti et al 2011). There are three members of the
genus Vicia that are characteristic of Mediterranean grain agriculture. Vicia faba (of which the
wild form is still unknown) is considered as one of the principal crops in southwest Asia. The
origin and natural distribution of Vicia sativa is also obscure. Wherever this plant is present, it
often occurs in low quantity in comparison to other pulse remains; it also mimics the seed of
lentils, making it difficult to separate the seeds of both genera from each other. Erskine et al.
(1994) argue that the simplest hypothesis for the domestication of this species could be that it
was selected as a secondary crop from weeds of the primary crop lentil. Vicia ervilia is

74
capable of colonizing abandoned fields and roadsides. This plant is always accompanied by
annual vetches, annual chickpeas, peas, and lentils in natural habitats. Sporadic occurrences of
this species have been reported from several PPN sites, but its widespread cultivation was
centered in Anatolia and the Balkans and is heavily associated with Neolithic and Bronze Age
contexts. Lev et al. (2005) report that up to the present day the seeds of Vicia ervilia have
been used to treat skin diseases and burns, coughing, leprosy, and hemorrhoids.
The finds of the genera Vicia sp. and Lathyrus sp. are reported at the Middle Paleolithic
Kebara Cave (65 – 48 ka B.P.; Lev et al. 2005) and from numerous sites during the Aceramic
Neolithic. Both genera were found at PPN Tell Abu Hureyra, Yiftah‟el, and Nevali Çori.
Helbaek (1969) does not report any occurrences of large-seeded pulses such as Vicia sp. or
Lathyrus sp. in his findings from Tepe Ali Kosh, but Vicia sp. is firmly evidenced in the Ganj
Dareh account, and there is also evidence for the presence of both genera and Vicia ervilia
from M‟lefaat (Savard et al. 2003).
Experimental research by Valamoti et al. (2011) indicates important results for the Chogha
Golan remains. The authors argue that the processing of Vicia to render it edible by removing
the toxic substance from the seeds can be detected in the archaeobotanical record. By
investigating the probable processing stages of Vicia ervilia and Lathyrus sativus, their
experiment shows that specimens boiled prior to processing have a clear depression on the
inner cotyledon surface that was not observed in specimens that were not treated by water or
in specimens that were soaked. They concluded that the seeds would have split into separate
cotyledons either prior to charring, through grinding or pounding, or perhaps after charring,
due to soaking or more probably boiling.
Most specimens in the Chogha Golan data are also separated into cotyledons but no
depression was observed on the inner cotyledon surfaces, which are usually found flat. In the
experimental study by Valamoti et al. (2011), specimens that had been untreated or soaked
(but not boiled) also demonstrated this flat surface pattern. Moreover, many specimens of
large-seeded pulse taxa recovered from AH III are extremely appressed on both dorsal
surfaces, giving the impression that they were somehow subject to a mechanical force like
grinding or pounding.

75
VI.2.3 The prevalence of domesticated crop plants
A clear pattern of change over Fertile Crescent PPN sites is that domesticated plants prevail in
the plant assemblages from the end of MPPNB onwards; they are well-documented at a
number of sites together with larger settlement sizes and presumably an increase in
population. Although today domesticated emmer wheat is a relic crop and its area of
production is sporadic, it was the principal wheat of southwest Asian agriculture in the
Neolithic and early Bronze Ages (Nesbitt 2002; Weiss and Zohary 2011). As Hillman (1984b)
noted during his ethnographic studies in southeastern Turkey in the 1970s, farmers even then
still favored the use of domesticated hulled emmer wheat for making cracked wheat (bulgur)
food.
Domesticated emmer (Triticum turgidum subsp. dicoccum)
In natural habitats, wild emmer is an element of the herbaceous flora typical of the Near
Eastern oak-pistachio park-forest belt. There are two distinct races of wild emmer that are
geographically, morphologically, and genetically separated from each other: a western
Palestine race and a Middle Eastern Turkish-Iraqi race. The western emmer populations are
more interconnected and intermixed within each other, so they do not indicate a clear
population structure. On the other hand, the eastern race, which consists of all accessions from
Turkey, Iraq, and Iran, has populations that are genetically more separated from each other
and so harbors more genetic diversity. The wild lines from this race are proposed as better
candidates for domesticated emmer because they might have better adapted to the climatic
conditions in mountainous regions in central Anatolia, the Caucasus, Central Asia, and
Europe than the heat- and drought-tolerant lines coming from the southern Levant. Moreover,
most wild emmer from the southern Levant has the disadvantage of having an extremely thick
glume, which makes it hard to process the grain (Abbo 2009; Özkan et al. 2005, 2010; Weiss
and Zohary 2011).
The genetic studies considering the geographic origin of the domestication of emmer and
einkorn have pointed to the Karacadağ and Kartal regions, which today are located
respectively in the Diyarbakır and Gaziantep provinces of southeastern Turkey (Özkan et al.
2005, 2010; Heun et al. 1997). This molecular evidence for signaling a single geographic
origin for emmer and einkorn is opposed by some scholars They mainly argue that the

76
molecular studies based on estimating the genetic distance of today‟s wild and domesticated
plant population is inefficient in determining multiple domestication events because the
combination of hybridization among populations, migration, and genetic drift might affect the
shape of phylogenetic trees, which would eventually result in the reduction of the allelic
variation among populations and lead to false assumptions of a single origination event
(Purugganan and Fuller 2009; Fuller et al. 2010; Fuller et al. 2011; Allaby et al. 2008, Brown
et al. 2009).
The earliest unequivocal finds of domesticated emmer appear during EPPNB Çayönü and
Cafer Höyük, with hundreds of spikelet bases showing rough abscission scars. The evidence
of emmer domestication represents region-wide occurrences all over the Fertile Crescent after
the middle PPNB, while prior to this period wild emmer appeared only at southern Levantine
sites and not in northern Syria or southwestern Turkey, where einkorn was present alongside
rye and barley (Willcox 2005; Weiss and Zohary 2011).
Later phases of the Aceramic period in western Iran are characterized by widespread
occurrences of domesticated plants at several sites, including Chogha Golan. Riehl et al.
(2013) found firm evidence that domesticated phenotypes of emmer wheat were being used
during the AH II and AH I periods by the inhabitants of Chogha Golan; that finding is also
associated with the probable weed taxa in the assemblage. This association of domesticated
types and later levels of the occupation is also consistent with the results of the current
research. The proportional contribution of emmer spikelet bases – although defining the status
of their domestication was out of the scope of this research – becomes more pronounced at
later AHs of the occupation.
Ganj Dareh has no or equivocal evidence of the use of any domesticated crop plants. This
situation could be attributed to its brief occupation period, which was less than 200 years
according to Zeder and Hesse (2000). AMS dates show that the site was occupied around
10.000 years ago, that could be possible that the site was abandoned before the earliest
appearance of domesticated emmer at Chogha Golan in AH II. The final phase of Tepe Ali
Kosh (Mohammad Jaffar) indicates the presence of domesticated einkorn/emmer spikelet
bases together with the grains of two-rowed hulled barley. One interesting phenomenon is that
during this last phase of occupation it appears that the collection of wild edible plants of the
steppic environment, especially small-seeded legumes, increased again on a huge scale, in

77
contrast to the contribution of cultivated plants, which declines from 55 % to 4 % of the
composition. Additionally, during the last phase of the occupation, wild cereals completely
disappeared from the assemblage. Furthermore, all other sites in Western Iran that postdate or
were inhabited during the final phases of Chogha Golan occupation (Tell Maghzaliyeh,
Jarmo, Chogha Bonut, Tepe Abdul Hosein) demonstrate the presence of domesticated plants
with a wide crop spectrum. Charles (2007) notes that the predominant domesticated crops of
this and subsequent phases are emmer and barley, while lentil and einkorn appear less
abundantly in these sites. In addition, there is also evidence for the presence of peas, grass
peas, and/or vetches. The persistence of wild forms of these latter species in plant
assemblages from the late Aceramic of western Iran were also noted by Charles (2007).
Free-threshing wheats
Singular finds of a free-threshing type spikelet base with brittle-rachised abscission scar (Plate
7G) was found in layer AH III of Chogha Golan (and also two other specimens in AH XI, as
Riehl et al. (2013) noted). This find represents an unusual figure comparing the lack of
evidence on free-threshing type findings in western Iran during the Aceramic Neolithic. The
earliest firm evidence for free-threshing wheat spikelet bases on the basis of a reliable rachis
criterion comes from the MPPNB onwards in low quantities at the sites of Tell Aswad, Can
Hasan III, Tell Bouqras, Çatalhöyük, and Tell Ramad (Weiss and Zohary 2011).
Genetic studies and crossing experiments indicate that the progenitors of Triticum aestivum
(AABBDD) are the tetraploid Triticum turgidum (AABB) and a diploid wild grass Aegilops
tauschii ssp. strangulata (DD). At the same time, to date no wild form of Triticum aestivum
has ever been found in the nature, strengthening the theory of natural hybridization among the
two species in cultivated fields (Matsuoka, 2011).
The role of the D-genome donor plant, Ae. tauschii subsp. strangulata, is central to this
speciation event. The contemporary natural distribution of this species is restricted to
Transcaucasia and southwest fringes of Caspian Sea in northern Iran. This strengthens the
theory that agriculture and tetraploid wheats radiated to this region to allow hybridization with
Ae. tauschii. However, the earliest archaeobotanical evidence of Neolithic farming in the
Caspian region suggests that this might have happened during the Neolithic Period, while
hexaploid free-threshing wheats from the Neolithic have already been identified at several

78
sites in the Fertile Crescent (Nesbitt and Samuel 1996). Considering this discrepancy, Riehl et
al. (2013) and Nesbitt and Samuel (1996) raise the question whether the past distribution of
this species was more widespread than today, allowing for much earlier hybridization in a
different region.
Two major qualitative trait loci (Tg and Q) contribute to the modifications of rachis fragility
and glume tenacity in the T. turgidum lineage, which further gave rise to free-threshing
tetraploid ssp. durum (hard wheat) and hexaploid T. aestivum (bread wheat). The Tg
(tenacious glume) locus controls glume toughness while the Q locus supports the formation of
square-headed ears with good threshing capacity (Peng et al. 2011; Salamini et al. 2002;
Matsuoka 2011). The origin of the dominant Q allele is ambivalent in the sense that it might
have first appeared in hexaploid wheats, which later introgressed into tetraploid wheats
through hybridization. But it is also probable that the recessive q to dominant Q mutation
might have appeared only once in tetraploid wheats and was inherited by hexaploid wheats.
Therefore, on a genetic basis, a genotypic change from qqTgTg to QQtgtg is considered
essential to the emergence of the free-threshing wheat species. In other words, because Tg
conceals the action of Q allele, the QQTgTg genotype results in a non-free threshing (hulled)
phenotype (Matsuoka 2011).
In the case of Triticum and Aegilops hybridization, Matsuoka et al. (2008; 2011) state that
there are at least 57 natural hybrids known in botanical sources but no natural occurrence of a
Triticum turgidum and Ae. tauschii hybrid has ever been found in nature. Furthermore,
crossing experiments show that hybridization of T. turgidum ssp. dicoccoides with Ae.
tauschii results in fragile-rachised hulled hexaploid wheat. The only known cases of this type
of wheat are the wild variety of Triticum macha var. megrelicum and another brittle-rachised
hulled hexaploid wheat in Tibet (Nesbitt and Samuel 1996; Cao et al. 1997). The progenies in
the cross of Tibetian semi-wild wheat with T. aestivum spp. spelta indicates that three genes
regulating rachis fragility interact to result in three different types of this trait such as a semi-
wild wheat type with shattering rachis, a spelta type, and the tough rachis of common wheat
(Cao et al. 1997).
In view of the molecular evidence and crossing experiments, it is highly probable that the
hybridization of two genera may have occurred at Chogha Golan. In particular, the
appearance of Aegilops and Triticum specimens in layers AH XI and AH III of the occupation

79
is rather striking insofar as the highest find density values of Aegilops remains were recorded
in these two AHs. It is nearly impossible to understand the true nature of what species might
have been involved in the emergence of these types of findings, but given the recorded cases
of hybridization between the Aegilops and Triticum genera in the wild and the species
diversity of Triticum complex at Chogha Golan shown in the current research, it is highly
likely that these singular spikelet base finds may represent a hulled hexaploid type resembling
weedy wheats such as Triticum macha var. megrelicum or the Tibetian semi-wild wheat.

80
VII. CONCLUDING REMARKS
On the basis of existing knowledge, Chogha Golan is a good candidate to fill the
developmental-temporal distance between the Upper Paleolithic and middle PPNB periods in
western Iran. The particular changes in the composition of the Chogha Golan plant
assemblage signal the strong likelihood of simultaneous plant management strategies over
central Zagros sites similar to those over much of the Fertile Crescent.
Although archaeobotanical studies of western Iran are only now emerging and there are few
comparable well-studied sites, certain similarities are evident among assemblages. In the
course of this research, the changes in the Chogha Golan plant assemblage are defined as
three systems of plant exploitation that are also congruent with the theoretical assumption of a
stepwise evolutionary continuum towards the intensification of human intervention in the
managed landscape and, eventually, the establishment of village-based agricultural
communities.
The first stage of this trajectory comprises wild plant-food procurement, which is represented
by the suspicious abundance of small-seeded taxa, especially legumes in western Iran sites.
Even though the earliest phases of Chogha Golan are separated by more than 1.000 years
from the earliest phases of Tepe Ali Kosh and Ganj Dareh, the heavy emphasis of small-
seeded taxa during earlier occupations is quite striking. This pattern favoring small-seeded
taxa comprising Scirpus maritimus, Rumex/Polygonum, and various other species is also
prevalent in PPNA sites like Hallan Çemi and Demirköy.
A second stage in the developmental route to a farming economy is considered to be wild
plant-food production that focused on the cultivation of large-seeded grasses and pulses, and
is characterized by a decreasing dependence on gathering wild plants for food and an
increasing input of human labor per unit area of exploited land. Following the argument of
Riehl et al. (2013) on the appearance of crop progenitors and arable weeds in the Chogha
Golan assemblage that signal pre-domestication cultivation, it is proposed that this stage of
plant exploitation is represented through a conspicious decrease of small-seeded pulses and
other wild taxa while there is an apparent increase in the representations of large-seeded taxa.
This pattern is also evident in the Ganj Dareh assemblage although there is a long
chronological gap between two sites. It is most probable that this changing trend represents a

81
change in subsistence strategy from a heavy emphasis on wild food procurement to wild food
production in the Chogha Golan and Ganj Dareh assemblages.
A final stage of plant management strategies is proposed to be the establishment of a farming
economy in western Iran that is evident with the widespread appearance of domesticated crop
plants. This phenomenon is stressed more after the end of the MPPNB or, specifically, at the
final phase of the PPN in western Iran. The sites that are inhabited within this temporal frame
show well-developed crop plant assemblages as do many of the sites in the Fertile Crescent.
Additionally, singular free-threshing type spikelet bases are among most interesting finds in
the Chogha Golan assemblage that illustrate the high species diversity of the Triticum
complex and the genetic potential for speciation at the Zagros foothills.
In conclusion, the floral remains examined during this research clearly coincide with an
expected regional trajectory towards the establishment of a farming economy regarding the
developmental timing of PPN settlements and the appearance of wild and domesticated crop
plants over much of Fertile Crescent.
VII.1 Future Research at Chogha Golan
Continuing research at Chogha Golan will help to clarify the role and nature of neolithization
processes in the central Zagros region. In particular, Chogha Golan offers a wide array of
research topics for bioarchaeological disciplines. In addition to the recent contributions of
Riehl (2011, 2013) and this current study, more research in archaeobotany, zooarchaeology,
micromorphology, and archaeology is being conducted by the scholars and students of the
University of Tübingen. These new studies on Chogha Golan will make positive additions to
our current understanding of the early history of farming societies in this region.
The current research did not undertake a ubiquity analysis due to small sample size.
However, the application of more quantitative measures to the Chogha Golan data, with more
analyzed samples, could bring out valuable information on the variations of the floral remains
throughout the occupation period. For instance, as Ganj Dareh data is readily available for

82
researchers in the publication of van Zeist et al. (1984), a comparison of the Ganj Dareh and
Chogha Golan data could display very interesting results.
The formation process of the plant assemblage was out of the scope of this current research.
Among different routes of entry for plant remains at Chogha Golan, intentional/deliberate
activities like gathering wild plants for nutritional or pharmacological purposes was
investigated through ethnographic data. However, other principal routes of formation such as
crop processing activities or dung burning as fuel should also be taken into consideration
regarding the evidence posited in Riehl et al. (2013) for the pre-domestication cultivation at
the site.
The necessity of contextual evidence is widely echoed in archaeobotanical literature. The
increaseof horizontal excavations in prehistoric archaeology now represents a prerequisite for
developing a better comprehension of intra-site contextual variations within different phases.
For this reason, resuming excavations at Chogha Golan would help to clarify contextual
evidence for various plant use practices as well as to pinpoint the area of crop processing or
perhaps communal consumption of cultivated plants.
The lack of archaeological research in the eastern part of the Fertile Crescent belt also retards
the probable contribution of this region to our understanding of the development of farming
communities. The contribution of Riehl et al. (2011, 2013) at Chogha Golan clearly
demonstrated that even though studies on the origins of agriculture started early in the last
century, we are still at the very beginnings for understanding this major transformation. More
regional and site-specific investigations will shed light on new perspectives. In this respect,
archaeological research in western Iran is enhanced by the potential to reshape our
understanding of the origins of agriculture and will help to develop models better suited to
clarify the true nature of neolithization in Southwest Asia.

83
BIBLIOGRAPHY
Abbo, S., Lev-Yadun, S., Ladizinsky, G., 2001. Tracing the wild genetic stocks of crop plants.
Genome, 44, 309–310.
Abbo, S., Saranga, Y., Peleg, Z., Kerem, Z., Lev-Yadun, S., Gopher, A., 2009. Reconsidering
domestication of legumes versus cereals in the ancient Near East. The Quarterly Review
of Biology, 84, 29–50.
Abbo, S., Lev-Yadun, S., Gopher, A., 2010. Agricultural origins: centers and noncenters: a
Near Eastern reappraisal. Critical Reviews in Plant Sciences, 29, 317–328.
Abbo, S., Lev-Yadun, S., Gopher, A., 2012. Plant domestication and crop evolution in the
Near East: on events and processes. Critical Reviews in Plant Sciences, 31, 241–257.
Abbo, S., Lev-Yadun, S., Heun, M., Gopher, A., 2013. On the “lost”crops of the neolithic
Near East. Journal of Experimental Botany, 64, 815–822.
Allaby, R.G., Fuller, D.Q., Brown, T.A., 2008. The genetic expectations of a protracted model
for the origins of domesticated crops. Proceedings of the National Academy of Sciences,
105, 13982–13986.
Anderberg, A.L., 1994. Atlas of seeds and small fruits of Northwest-European plant species
with morphological descriptions. Part 4, Resedacea-Umbelliferae. Swedish Museum of
Natural History, Risbergs Tryckeri, Stockholm.
Asouti, E., 2006. Beyond the Pre-pottery Neolithic B interaction sphere. Journal of World
Prehistory, 20, 87–126.
Asouti, E., Fuller, D.Q., 2012. From foraging to farming in the southern Levant: the
development of Epipalaeolithic and Pre-pottery Neolithic plant management strategies.
Vegetation History and Archaeobotany, 21 (2), 149–162.
Asouti, E., Fuller, D.Q., 2013. A Contextual Approach to the Emergence of Agriculture in
Southwest Asia. Current Anthropology, 54, 299–345.

84
Badr, A., Müller, K., Schäfer-Pregl, R., El Rabey, H., Effgen, S., Ibrahim, H.H., Pozzi, C.,
Rohde, W., Salamini, F., 2000. On the origin and domestication history of barley
(Hordeum vulgare). Molecular Biology and Evolution, 17 (4), 499–510.
Balter, M., 2007. Seeking agriculture‟s ancient roots. Science, 316, 1830–1835.
Barrett, S.H., 1983. Crop mimicry in weeds. Economic Botany, 37, 255–282.
Bar-Yosef, O., 1998. On the nature of transitions: the Middle to Upper Palaeolithic and the
Neolithic Revolution. Cambridge Archaeological Journal, 8, 141–163.
Bender, B., 1975. Farming in prehistory: from hunter-gatherer to food-producer. John Baker,
London.
Bender, B., 1978. Gatherer-hunter to farmer: a social perspective. World Archaeology, 10,
204–222.
Berggren, G. 1969. Atlas of Seeds and small fruits of Northwest-European plant species with
morphological descriptions. Part 2, Cyperaceae. Swedish Natural Science Research
Council, Berlingska Boktryckeriet, Stockholm.
Berggren, G. 1981. Atlas of Seeds and small fruits of Northwest-European plant species with
morphological descriptions. Part 3, Salicaceae-Cruciferae. Swedish Museum of
Natural History, Berlings, Stockholm.
Binford, L.R., 1968. Post-Pleistocene adaptations. In: Prehistoric agriculture. pp. 22–49.
Struever S., Ed. 1971. The Natural History Press, New York.
Bojnanskỳ, V., Fargavsová, A., 2007. Atlas of seeds and fruits of Central and East-European
flora: the Carpathian Mountains region. Springer, Dordrecht.
Braidwood, R.J., 1969. The earliest village communities of southwestern Asia reconsidered.
In: Prehistoric agriculture. pp. 236–251. Struever S., Ed. 1971. The Natural History
Press, New York.
Brown, T.A., Jones, M.K., Powell, W., Allaby, R.G., 2009. The complex origins of
domesticated crops in the Fertile Crescent. Trends in Ecology & Evolution, 24, 103–
109.

85
Butler, A., 1995. The small-seeded legumes: An enigmatic prehistoric resource. Acta
Palaeobotanica, 35, 105–116.
Butler, A., 1996. Trifolieae and related seeds from archaeological contexts: problems in
identification. Vegetation History and Archaeobotany 5, 157–167.
Cao, W., Scoles, G.J., Hucl, P., 1997. The genetics of rachis fragility and glume tenacity in
semi-wild wheat. Euphytica, 94, 119–124.
Charles, M., 2007. East of eden? A consideration of Neolithic crop spectra in the eastern
Fertile Crescent and beyond. In: The origins and spread of domestic plants in southwest
Asia and Europe. pp. 53–74. Colledge S. and Connolly J., Eds. Left Coast Press,
California.
Childe, V.G., 1951. The Neolithic Revolution. In: Prehistoric agriculture. pp. 15–21.
Struever S., Ed. 1971. The Natural History Press, New York.
Childe, V.G., 1958. The prehistory of European society. Penguin Books, Great Britain.
Colledge, S., 1999. Identifying pre-domestication cultivation using multivariate analysis. In:
The origins of agriculture and crop domestication: proceedings of the Harlan
Symposium, 10 – 14 May 1997. pp. 121-132. Damania A.B., Valkoun J., Willcox G.,
Qualset C.O, Eds. ICARDA, Aleppo, Syria.
Colledge, S., 2002. Identifying pre-domestication cultivation in the archaeobotanical record
using multivariate analysis presenting the case for quantification. In: The dawn of
farming in the Near East. pp. 141–152. Cappers, R.T.J. and Bottema, S., Eds. Ex
Oriente, Berlin.
Colledge, S., Conolly, J., 2010. Reassessing the evidence for the cultivation of wild crops
during the Younger Dryas at Tell Abu Hureyra, Syria. Environmental Archaeology, 15,
124–138.
Conard, N.J., Riehl, S., Zeidi, M., (n.d.) Revisiting neolithization in the Zagros foothills;
excavations at Chogha Golan, a PPN site in Ilam Province. s.n., s.l.
Davis, P. H., 1965-2000. Flora of Turkey and the East Aegean Islands. Vol. 1-11. Edinburgh
University Press, Edinburgh.

86
Diamond, J., 2002. Evolution, consequences and future of plant and animal domestication.
Nature, 418, 700–707.
Dubcovsky, J., Dvorak, J., 2007. Genome plasticity a key factor in the success of polyploid
wheat under domestication. Science, 316, 1862–1866.
Dvorak, J., Deal, K.R., Luo, M.-C., You, F.M., von Borstel, K., Dehghani, H., 2012. The
origin of spelt and free-threshing hexaploid wheat. Journal of Heredity, 103, 426–441.
El-Moslimany, A.P., 1986. Ecology and late-Quaternary history of the Kurdo-Zagrosian oak
forest near Lake Zeribar, western Iran. Vegetation, 68, 55–63.
El-Moslimany, A.P., 1990. Ecological significance of common nonarboreal pollen: examples
from drylands of the Middle East. Review of Palaeobotany and Palynology, 64, 343–
350.
Erskine, W., Smartt, J., Muehlbauer, F.J., 1994. Mimicry of lentil and the domestication of
common vetch and grass pea. Economic Botany, 48, 326–332.
Flannery, K.V., 1969. Origins and ecological effects of early domestication in Iran and the
Near East. In: Prehistoric agriculture. pp. 51–100. Struever S., Ed. 1971. The Natural
History Press, New York.
Flannery, K.V., 1973. The origins of agriculture. Annual Review of Anthropology, 2, 271–310.
Fuller, D.Q., 2007. Contrasting patterns in crop domestication and domestication rates: recent
archaeobotanical insights from the Old World. Annals of Botany, 100, 903–924.
Fuller, D.Q., Colledge, S., 2008a. Recent lessons from Near Eastern archaeobotany: wild
cereal use, pre-domestication cultivation and tracing multiple origins and dispersals.
Praghara, 18, 105–134.
Fuller, D.Q., Stevens, C., McClatchie, M., 2008b. Routine activities, tertiary refuse and labor
organisation: social inference from everyday archaeobotany. Ancient Plants and People
– Contemporary Trends in Archaeobotany.

87
Fuller, D.Q., Allaby, R.G., Stevens, C., 2010. Domestication as innovation: the entanglement
of techniques, technology and chance in the domestication of cereal crops. World
Archaeology, 42, 13–28.
Fuller, D.Q., Willcox, G., Allaby, R.G., 2011. Cultivation and domestication had multiple
origins: arguments against the core area hypothesis for the origins of agriculture in the
Near East. World Archaeology, 43, 628–652.
Fuller, Dorian Q., Willcox, G., Allaby, R.G., 2012a. Early agricultural pathways: moving
outside the “core area” hypothesis in Southwest Asia. Journal of Experimental Botany,
63, 617–633.
Fuller, D. Q., Asouti, E., Purugganan, M.D., 2012b. Cultivation as slow evolutionary
entanglement: comparative data on rate and sequence of domestication. Vegetation
History and Archaeobotany, 21 (2), 131–145
Harlan, J.R., Zohary, D., 1966. Distribution of wild wheats and barley. Science, 153, 1074.
Harris, D.R., 1977. Alternative pathways toward agriculture. In: Origins of agriculture. pp.
179–243. Reed C.A., Ed., Mouton Publishers, Paris.
Harris, D.R., 2007. Agriculture, cultivation and domestication: exploring the conceptual
framework of early food production. In: Rethinking agriculture: archaeological and
ethnoarchaeological perspectives. pp. 16–35. Denham T., Iriarte, J., and Vyydaghs, L.,
Eds., Left Coast Press, Walnut Creek, CA.
Hayden, B., 2001. The dynamics of wealth and poverty in the transegalitarian societies of
Southeast Asia. Antiquity, 75, 571–581.
Hayden, B., 2003. Were luxury foods the first domesticates? Ethnoarchaeological
perspectives from Southeast Asia. World Archaeology, 34, 458–469.
Hayden, B., 2009. The proof is in the pudding. Current Anthropology, 50, 597–601.
Helbaek, H., 1969. Plant collecting, dry-farming, and irrigation agriculture in prehistoric Deh
Luran. In: Prehistory and human ecology of the Deh Luran Plain. pp. 383-425. Hole F.,
Flannery K.V., Neely J.A., Eds. Ann Arbor: University of Michigan,

88
Helbaek, H., 1969. Plant collecting, dry-farming, and irrigation agriculture in prehistoric Deh
Luran. In: Prehistory and human ecology of the Deh Luran Plain. Hole, F., Flannery,
K.V., Neely, J.A., Ed. University of Michigan. Ann Arbor.
Heun, M., Schäfer-Pregl, R., Klawan, D., Castagna, R., Accerbi, M., Borghi, B., Salamini, F.,
1997. Site of einkorn wheat domestication identified by DNA fingerprinting. Science,
278, 1312–1314.
Hillman, G.C, 1984. Interpretation of archaeological plant remains: the application of
ethnographic models from Turkey. In: Plants and Ancient Man: Studie sin
Paleoethnobotany. pp. 1-41. Van Zeist, W., and Casparie, W.A., Eds. A.A. Balkerna,
Rotterdam.
Hillman, G.C., 1984. Traditional husbandry and processing of archaic cereals in modern
times: Part I, the glume wheats. Bulletin of Sumerian Agriculture, 1, 114-152.
Hillman, G.C., Davies, M.S., 1990. Measured domestication rates in wild wheats and barley
under primitive cultivation, and their archaeological implications. Journal of World
Prehistory, 4, 157–222.
Hillman, G.C., 2000. In: Village on the Euphrates: from foraging to farming at Abu Hureyra.
Moore, A.M.T., Hillman, G.C., Legge, A.J., Eds. Oxford University Press, New York.
Hillman, G.C, Hedges, R., Moore, A., Colledge, S., Pettitt, P., 2001. New evidence of
Lateglacial cereal cultivation at Abu Hureyra on the Euphrates. The Holocene, 11, 383–
393.
Hodder, I., 2001. Review feature: The birth of the Gods and the origins of agriculture by
Jacques Cauvin. Cambridge Archaeological Journal. 11 (1), 105–121.
Hole, F., 1984. A reassessment of the Neolithic revolution. Paléorient, 10 (2), 49–60.
Hole, F., 1996. The context of caprine domestication in the Zagros region. In: The origins and
spread of agriculture and pastoralism in Eurasia. pp. 263–281. Harris, D., Ed.
Smithsonian Institution Press, Washington, DC.

89
Hole, F., Flannery, K.V., Neely, J.A., 1969. Prehistory and human ecology of the Deh Luran
Plain. In: Prehistoric agriculture. pp. 252 - 311. Struever, S., Ed. 1971. The Natural
History Press, New York.
Hubbard, R., 1990. Archaeobotany of Abdul Hosein: The carbonised seeds from Tepe Abdul
Hosein, results of preliminary analyses. In: Tepe Abdul Hosein: A Neolithic site in
western Iran: excavations 1978. pp. 217-226. Pullar J., Ed. B.A.R., Oxford.
Jones, G., 1991. Numerical analysis in archaeobotany. In: Progress in old world
palaeoethnobotany. pp. 63–80. Van Zeist, W., Wasylikowa, K., and Behre, K.-E., Eds.
A.A. Balkema, Rotterdam.
Kehl, M., 2009. Quaternary climate change in Iran – the state of knowledge. Erdkunde, 1–17.
Kislev, M.E., Bar-Yosef, O., 1988. The legumes: The earliest domesticated plants in the Near
East? Current Anthropology, 29, 175–179.
Kislev, M.E., Nadel, D., Carmi, I., 1992. Epipalaeolithic (19,000 BP) cereal and fruit diet at
Ohalo II, Sea of Galilee, Israel. Review of Palaeobotany and Palynology, 73, 161–166.
Kislev, M.E., Weiss, E., Hartmann, A., 2004. Impetus for sowing and the beginning of
agriculture: ground collecting of wild cereals. Proceedings of the National Academy of
Sciences, 101, 2692–2695.
Ladizinsky, G., 1987. Pulse domestication before cultivation. Economic Botany, 41, 60–65.
Leonti, M., Nebel, S., Rivera, D., Heinrich, M., 2006. Wild gathered food plants in the
European Mediterranean: a comparative analysis. Economic Botany, 60, 130–142.
Lev, E., Kislev, M.E., Bar-Yosef, O., 2005. Mousterian vegetal food in Kebara cave, Mt.
Carmel. Journal of Archaeological Science, 32, 475–484.
Luczaj, L., Pieroni, A., Tardío, J., Pardo-de-Santayana, M., Sõukand, R., Svanberg, I., Kalle,
R., 2012. Wild food plant use in 21st century Europe: the disappearance of old
traditions and the search for new cuisines involving wild edibles. Acta Societatis
Botanicorum Poloniae, 81, 359–370.

90
Matthews, R., Mohammadifar, Y., Matthews, W., Motarjem, A., 2010. Investigating the early
Neolithic of western Iran: the Central Zagros Archaeological Project. [online] Available
at: http://antiquity.ac.uk/projgall/matthews323/ [Accessed 02.08.2013]
Matsuoka, Y., 2011. Evolution of polyploid Triticum wheats under cultivation: the role of
domestication, natural hybridization and allopolyploid speciation in their diversification.
Plant and Cell Physiology, 52, 750–764.
Melamed, Y., Plitmann, U., Kislev, M.E., 2008. Vicia peregrina: an edible early Neolithic
legume. Vegetation History and Archaeobotany, 17 (1, suppl.), 29–34.
Miller, N.F., 1992. The origins of plant cultivation in the Near East. pp. 39–58. In: The
origins of agriculture: an international perspective. Cowan, C.W., and Watson, P.J.,
Eds. Smithsonian Institution Press, Washington, DC.
Miller, N.F., 1997. The macrobotanical evidence for vegetation in the Near East, C. 18 000/16
000 BC 4 000 BC. Paléorient, 23 (2), 197–207.
Miller, N.F., Smart, T.L., 1984. International burning of dung as fuel: a mechanism for the
incorporation of charred seeds into the archeological record. Journal of Ethnobiology, 4,
15–28.
Moore, A.M.T., 1985. The development of Neolithic societies in the Near East. Advances in
World Archaeology, 4, 1–69.
Morrell, P.L., Clegg, M.T., 2007. Genetic evidence for a second domestication of barley
(Hordeum vulgare) east of the Fertile Crescent. Proceedings of the National Academy of
Sciences, 104, 3289–3294.
Nesbitt, M., and D. Samuel, 1996. From staple crop to extinction? The archaeology and
history of the hulled wheats. pp. 41–10. In: Hulled wheats. Proceedings of the 1st
International Workshop on Hulled Wheats. Padulosi S., Hammer K., and Heller J., Eds.
Castelvecchio Pascoli, Italy.
Nesbitt, M., 2002. When and where did domesticated cereals first occur in southwest Asia?
pp. 113–132. In: The dawn of farming in the Near East. Cappers, R.T.J., and Bottema,
S., Eds. Ex Oriente, Berlin.

91
Nesbitt, M., 2006. Identification guide for Near Eastern grass seeds. Archetype Publications,
London.
Nevo, E., Zohary, D., Belles, A., Kaplan, D., Storch, N.,1986. Genetic diversity and
environmental associations of wild barley, Hordeum spontaneum, in Iran. Plant
Systematics and Evolution, 153, 141–164.
Özkan, H., Brandolini, A., Pozzi, C., Effgen, S., Wunder, J., Salamini, F., 2005. A
reconsideration of the domestication geography of tetraploid wheats. Theoretical and
Applied Genetics, 110, 1052–1060.
Özkan, H., Willcox, G., Graner, A., Salamini, F., Kilian, B., 2010. Geographic distribution
and domestication of wild emmer wheat (Triticum dicoccoides). Genetic Resources and
Crop Evolution, 58, 11–53.
Peng, J., Sun, D., Nevo, E., 2011. Wild emmer wheat, Triticum dicoccoides, occupies a
pivotal position in wheat domestication process (Invited review article). Australian
Journal of Crop Science, 5 (9), 1127–1143.
Piperno, D.R., Weiss, E., Holst, I., Nadel, D., 2004. Processing of wild cereal grains in the
Upper Palaeolithic revealed by starch grain analysis. Nature, 430, 670–673.
Popper, V.S., 1988. Selecting quantitative measurements in paleoethnobotany. In: Current
paleoethnobotany: analytical methods and cultural interpretations of archaeological
plant remains pp. 53–71. Hastorf, C.A., and Popper, V.S., Eds. University of Chicago
Press, Chicago.
Pullar, J., 1990. Tepe Abdul Hosein: a Neolithic site in Western Iran: excavations 1978.
B.A.R., Oxford.
Purugganan, M.D., Fuller, D.Q., 2009. The nature of selection during plant domestication.
Nature, 457, 843–848.
Rechinger, K.H., 1963 – 2012. Flora Iranica. Akademische Drück –u. Verlagsanstalt, Graz.
Riehl, S., 1999. Bronze Age environment and economy in the Troad: the archaeobotany of
Kumtepe and Troy. Dissertation. BioArchaeologica 2, Mo-Vince-Verlag, Tübingen.

92
Riehl, S., Benz, M., Conard, N.J., Darabi, H., Deckers, K., Nashli, H.F., Zeidi-Kulehparcheh,
M., 2012. Plant use in three Pre-Pottery Neolithic sites of the northern and eastern
Fertile Crescent: a preliminary report. Vegetation History and Archaeobotany, 21 (2),
95–106.
Riehl, S., Zeidi, M., Conard, N.J., 2013. Emergence of agriculture in the foothills of the
Zagros Mountains of Iran. Science, 341, 65–67.
Salamini, F., Özkan, H., Brandolini, A., Schäfer-Pregl, R., Martin, W., 2002. Genetics and
geography of wild cereal domestication in the Near East. Nature Reviews Genetics, 3,
429–441.
Savard, M., Nesbitt, M., Gale, R., 2003. Archaeobotanical evidence for early Neolithic diet
and subsistence at M‟lefaat (Iraq). Paléorient, 29 (1), 93–106.
Savard, M., Nesbitt, M., Jones, M.K., 2006. The role of wild grasses in subsistence and
sedentism: new evidence from the northern Fertile Crescent. World Archaeology, 38,
179–196.
Sherratt, A., 2007. Diverse origins: regional contributions to the genesis of farming. In: The
origins and spread of domestic plants in southwest Asia and Europe. pp. 1–20. Colledge
S. and Connolly J., Eds. Left Coast Press, California.
Smith, B.D., 2001. Low-level food production. Journal of Archaeological Research, 9, 1–43.
Smith, B.D., 2007a. Niche construction and the behavioral context of plant and animal
domestication. Evolutionary Anthropology: Issues, News, and Reviews, 16, 188–199.
Smith, B.D., 2007b. The ultimate ecosystem engineers. Science 315, 1797.
Smith, P.E., 1971. Iran. 9000-4000 BC: The Neolithic. Expedition, 13, 9.
Smith, P.E., 1978. An interim report on Ganj Dareh Tepe, Iran. American Journal of
Archaeology, 82 (4), 537–540.
Sonnante, G., Hammer, K., Pignone, D., 2009. From the cradle of agriculture a handful of
lentils: history of domestication. Rendiconti Lincei, 20, 21–37.

93
Stepp, J.R., Moerman, D.E., 2001. The importance of weeds in ethnopharmacology. Journal
of Ethnopharmacology, 75, 19–23.
Stevens, L.R., Wright Jr, H.E., Ito, E., 2001. Proposed changes in seasonality of climate
during the Lateglacial and Holocene at Lake Zeribar, Iran. The Holocene, 11, 747–755.
Tanno, K., Willcox, G., 2006. How fast was wild wheat domesticated? Science, 311, 1886–
1886.
Tanno, K., Willcox, G., 2012. Distinguishing wild and domestic wheat and barley spikelets
from early Holocene sites in the Near East. Vegetation History and Archaeobotany, 21
(2), 107–115.
Trigger, B.G., 2006. A history of archaeological thought. 12th ed. Cambridge University
Press, Cambridge.
Valamoti, S.M., Moniaki, A., Karathanou, A., 2011. An investigation of processing and
consumption of pulses among prehistoric societies: archaeobotanical, experimental and
ethnographic evidence from Greece. Vegetation History and Archaeobotany, 20 (5),
381–396.
Van der Veen, M., Fieller, N., 1982. Sampling seeds. Journal of Archaeological Science, 9,
287–298.
Van der Veen, M., 2007. Formation processes of desiccated and carbonized plant remains–the
identification of routine practice. Journal of Archaeological Science, 34, 968–990.
Van Zeist W., Bakker-Heeres, J.A.H., 1982. Archaeobotanical studies in the Levant 1.
Neolithic sites in the Damascus Basin: Aswad, Ghoraife, Ramad. Palaeohistoria, 24,
165-256.
Van Zeist W., Smith, P.E.L., Palfenier-Vegter, R.M., Suwijn, M., Casparie, W.A., 1984. An
archaeobotanical study of Ganj Dareh Tepe, Iran. Palaeohistoria, 26, 201-224.
Van Zeist W., Bakker-Heeres, J.A.H., 1984a. Archaeobotanical studies in the Levant 2.
Neolithic and Halaf levels at Ras Shamra. Palaeohistoria, 26, 151-170.

94
Van Zeist W., Bakker-Heeres, J.A.H., 1984b. Archaeobotanical studies in the Levant 3. Late-
Paleolithic Mureybit. Palaeohistoria, 26, 171-199.
Van Zeist W., Bakker-Heeres, J.A.H., 1985. Archaeobotanical studies in the Levant 4. Bronze
age sites on the North Syrian Euphrates. Palaeohistoria Acta et Communicationes
Instituti Bio-Archaeologici Universitatis Groninganae, 27, 165-256.
Watkins, T., 2008. Supra-regional networks in the Neolithic of Southwest Asia. Journal of
World Prehistory, 21, 139–171.
Weiss, E., Kislev, M.E., Hartmann, A., 2006. Autonomous cultivation before domestication.
Science, 312, 1608.
Weiss, E., Zohary, D., 2011. The Neolithic southwest Asian founder crops. Current
Anthropology, 52, 237–S254.
White, C.E., Makarewicz, C.A., 2012. Harvesting practices and early Neolithic barley
cultivation at el-Hemmeh, Jordan. Vegetation History and Archaeobotany, 21 (2), 85–
94.
Willcox, G., 2004. Measuring grain size and identifying Near Eastern cereal domestication:
evidence from the Euphrates valley. Journal of Archaeological Science, 31, 145–150.
Willcox, G., 2005. The distribution, natural habitats and availability of wild cereals in relation
to their domestication in the Near East: multiple events, multiple centres. Vegetation
History and Archaeobotany, 14 (4), 534–541.
Willcox, G., 2012. Searching for the origins of arable weeds in the Near East. Vegetation
History and Archaeobotany, 21 (2), 163–167.
Willcox, G., Fornite, S., Herveux, L., 2008. Early Holocene cultivation before domestication
in northern Syria. Vegetation History and Archaeobotany, 17 (3), 313–325.
Wright, H.E., 1993. Environmental determinism in Near Eastern prehistory. Current
Anthropology, 34, 458–469.
Zeder, M.A., 1999. Animal domestication in the Zagros: a review of past and current
research. Paléorient, 25 (2), 11–25.

95
Zeder, M.A., Hesse, B., 2000. The initial domestication of goats (Capra hircus) in the Zagros
Mountains 10,000 years ago. Science, 287, 2254–2257.
Zeder, M.A., 2006. Central questions in the domestication of plants and animals. Evolutionary
Anthropology: Issues, News, and Reviews, 15, 105–117.
Zeder, M.A., 2009. The Neolithic macro-(r) evolution: macroevolutionary theory and the
study of culture change. Journal of Archaeological Research, 17, 1–63.
Zeder, M.A., 2011. The origins of agriculture in the Near East. Current Anthropology, 52,
221–235.
Zeidi M., Conard N.J., (in press). Preliminary report on the chipped lithic assemblage from
Chogha Golan, a PPN site in the foothills of the Zagros Mountains, Ilam Province, Iran.
Ex Oriente Series.
Zeidi M., Riehl S., Napierala H., Conard NJ. 2012. Chogha Golan: a PPN site in the foothills
of the Zagros Mountains, Ilam Province, Iran (Report on the first season of excavation
in 2009). In: Proceedings of the 7th International Congress on the Archaeology of the
Ancient Near East. pp. 259-275. Matthews, R., Curtis, J., Eds. Harrassowitz Verlag,
Weisbaden.
Zohary, D., 1989. Pulse domestication and cereal domestication: how different are they?
Economic Botany, 43, 31–34.
Zohary, D., Hopf, M., 2000. Domestication of plants in the Old World: The origin and spread
of cultivated plant in West Asia, Europe and the Nile Valley. 3rd ed. Oxford University
Press, Oxford.
Zohary, M., 1973. Geobotanical foundations of the Middle East. Fischer Stuttgart.
Zvelebil, M., Dolukhanov, P., 1991. The transition to farming in eastern and northern Europe.
Journal of World Prehistory, 5, 233–278.

96
APPENDIX 1: INVENTORY OF IDENTIFIED TAXA
The contents of this inventory comprise morphological and ecological descriptions of
identified taxa as well as length of the finds. For some taxa, the information of economic
significance has also been added.
CHENOPODICEAE/AMARANTHACEAE/ (Goosefoot/Amaranth family)
Atriplex sp. (orache)
Plate: 1-A, 1-B
Seed length: 1.34 mm
ID criteria: One seeded nutlet, enclosed within two appressed bracteoles which are fused to
nutlet at the margins. Due to charring only impressions of bracteoles attached to the seed can
be recognized in most cases. Exocarp is not recognizable as well. Seeds have a circular,
elliptic shape in outline with a slightly projected radicular tip. Bracteoles are diamond-shaped
in outline and the surface is distinctly nerved.
Ecology: Hedge in Rechinger (1997) notes that in SW Asia the species are distinctly segetal,
halophytic, and semi-desertic; many species are important grazing plants. Most thrives into
salty marshes and cultivated lands. 21 species were described in Iran and adjacent regions.
Hedge in Rechinger (1997) informs that Atriplex hastata records from Iran are certainly
misidentifications of A. patula and A. microantha. A. hastata is a European species
Salsola sp. (saltwort)
Plate: 1-C
Seed length: 2.10 mm
ID criteria: The seed is distinctly helix-shaped. No pseudocarp remains detected.

97
Ecology: Small or dwarf shrubs, annuals and rarely small trees. 48 species described in Flora
Iranica. The genus often associated with semi-desertic plant communities although a few of
the species penetrating into humid areas along coast lines and on ruderal and other disturbed
habitats (Freitag and Rilke in Rechinger 1997). In Davies (1967), it is noted that mostly
thrived into “sandy, salt-rich soils near the sea or scattered and partly naturalized in sandy,
salty places inland.”
Suaeda sp. (sea blite)
Plate: 1-D
Seed length: 0.88 mm
ID criteria: The shape of seed is reniform or elliptic with usually projected tip of radical. The
margin of the seed is transversely obovate-rhombic. The surface is glossy.
Ecology: Annuals, undershrubs or shrubs. Sandy, salty places (Davies, 1967). 16 species
defined in Flora Iranica (Akhani and Podlech in Rechinger 1997).
Indeterminate Chenopodiceae objects
ID criteria: The presence of distinct tip of radicle and the peripheral embryo surrounding the
albumen like a ring is attributed to the amaranth family.
ANACARDICEAE (sumac family)
Pistacia sp. (pistachio)
Plate: 1-E
Seed length: 4.36 mm

98
ID criteria: The pericarp of pistachio nuts was mostly found fragmented. Only one specimen
recovered intact which is oblong in outline with an opening on top.
Ecology: The arboreal species is widely distributed over Mediterranean and further inland to
Afghanistan, Pakistan and Central Asia. Following Zohary and Hopf‟s (2000) description,
Pistacia vera ranks among the most drought resistant fruit trees in west Asia. The cultivated
form, Pistacia vera is “dioecious, wind pollinated tree and fruit-bearing female clones have
been traditionally planted intermixed with male individuals”. P. khinjuk is an Irano-Turanian
element thrives rocky slopes of gorges at the altitude between 1000 – 1800 m. P. atlantica is a
deciduoud tree up to 7 m thrives into dry hillsides, cemeteries, edge of fields, roadsides. Often
occurring as a relic of destroyed forest. According to Yaltırık in Davies (1967), “[I]ts survival
in many localities may be explained by its economic use: its seeds are (or have been) used for
tanning and for soup-making, and its gum is applied as an antiseptic to wounds.”
ASPARAGACEAE (asparagus family)
Ornithagalum/Muscari/Bellevalia type
Plate: 2-F
Seed length: 2.46 mm
ID criteria: The seeds of this group of plants are almost globular or obovate with mostly
distinct suture on the ventral side. There is a circular opening visible in most of objects
accrossing cylindrically from the tip to the base. The surface is always smooth and no
recognizable testa exists.
Ecology: Bulbous perennials, mostly found on a wide range of different habitats such as
rocky slopes, hillsides and meadows.

99
ASTERECEAE (sunflower family)
.
Centaurea type (knapweed)
Plate: 2-A, 2-B
Seed Length: Small type: 1.77 mm; Big type: 2.87 mm.
ID Criteria: The usually fragmented nature of these objects and distinctly large species
variability in the sunflower family hampers species or genus level of identification. Following
the Ganj Dareh account of van Zeist et al. (1984), all objects of this family were attributed to
the category of Centaurea type. Some specimens which has an intact pappus rim show
similarities with the description of small Centaurea type in van Zeist et al. (1984). The shape
is obovate in outline, truncated at the apex and tapering and hooked at the base, where the
hilum should have been located. While another specimen has 3 layered-rim and a larger fruit
size. This diversity would indicate that there is more than one plant species may have
involved in Chogha Golan.
Ecology: Thrives into different habitats such as steppic vegetation, fallow fields, stony slopes
and sandy dunes.
BORAGINACEAE (borage family)
According to van Zeist and Bakker-Heeres (???), the finds of boragineous fruits pose some
problems. They note that “[…] On burning, the nutlets do not turn black, but they acquire a
whitish to grey colour which is due to the silica skeleton. Because of the silica skeleton also
of non-carbonized fruits the wall may remain preserved in archaeological deposits”.
Considering this remark, some Chogha Golan boragineous fruits show sign of carbonization
and some others not. As no intrusion reported from both archaeologists and in the papers of
Riehl (2011, 2013), these remains can mainly be considered as a part of archaeological
deposition rather than being modern intrusions.

100
Arnebia sp.
Plate: 2-C
Seed Length: ~2.97 mm
ID criteria: The nutlets of this boraginous plant are trigonous, have a broad triangular base
and widely spaced, a large verrucae. The apex is rather obtuse. The triangular base is bordered
by a distinct collar. The ventral keel extends over the upper 2/3 of the fruit. The surface is
densely covered with wart-like projections.
Ecology: Annual and perennial herbs; rootstock is yielding purple dye (Edmonson in Davies
1978). Riedl in Rechinger (1967) described 19 species of this genus from Iran and adjacent
regions.
Heliotropium sp. (turnsole)
Plate: 2D
Seed Length: 0.90 mm – 1.12 mm
ID criteria: The nutlets of this plant are slightly compressed on both sides of the keel. The
characteristic attribute is the protruding hilum, that is easily recognizable on CG objects.
Ecology: Annuals and perennials, 74 species of this genus described by Riedl in Rechinger
(1967) in Iran and adjacent regions. Riedl in Davies (1978) described this taxa thrives broad
range of habitats such as gravelly and rocky slopes, streambeds, vineyards, fields, roadsides.
Mostly ruderal.

101
Lithospermum sp. (groomwell)
Plate: 2E
Seed length: ~1.61 mm
ID Criteria: The prominent humps on sides, small rounded triangular base and rather pointed
apex are the main characteristics of this species. The surface is densely covered by wart-like
projections.
Ecology: The species thrives in cultivated fields as well as steppe vegetations.
BRASSICACEAE (mustard family)
The remains of this family are not successfully identified in most cases to a genus or species
level due to comparatively small seed size and the degrading effects of carbonization. Most
identification remains indeterminate for these reasons. In addition, some objects were found
intact to the fruit pods.
Allysum type
ID criteria: Obovate-elliptic shape in outline with a narrow tip of radicle, which is shorter
than the cotylenodary one. No membranous wing is still present due to charring.
Ecology: Annuals, biennials and perennials. An element of steppic habitats; many species
thrives into disturbed grounds.
cf. Arabis sp.
Plate: 3A
Seed Length: 0.92 mm

102
ID Criteria: The cotylenodary part of the seed is twice as wide as the radicular one. The tip
of radicle is longer than the tip of cotyledons. The shape is oblong-elliptic and flattened in
outline.
Ecology: Cullen in Davies (1965) defines most species thrives into cliff, stony, rocky places,
limestone rocks, fields and cultivated lands.
Brassica type
Plate: 3B
Seed length: 1.12 mm
ID criteria: The seed is spheroidal and the cotyledons are conduplicately located to the
radicle. Due to charring, the cotyledons slightly get apart from the hilum and make humps on
both sides exposing the hilum and the radicle. The surface is papillose.
Ecology: Annual, biennial or perennial herbs. Many species of Brassica genus have been
cultivated as food plants nowadays and others are widespread segetals thrive in dry rocky
slopes, steppe and cultivated fields. 7 species described in Flora Iranica (Hedge in Rechinger
1968).
Lepidium cf. sativum (garden cress)
Plate: 3C
Seed length: 1.49 mm – 1.01 mm
ID criteria: The seeds are obovate to semiobovate. The tip of radicle and cotyledons severely
deformed due to carbonization. Therefore no comparison between two elements is possible.
Testa is in all case partially lost; exposing the cotyledons.
Ecology: Annual. Hedge in Rechinger (1968) notes that ssp. sativum is a cultivated form of
this species while ssp. spinescens is xerophytic plant. Both subspecies occurs in waste places
and cultivated fields.

103
CARYOPHYLLACEAE (pink family)
Many objects remain indeterminate because of densely damaged outer surface. It should be
noted that more species/genus of similar morphological attributes may have been involved in
the assemblage.
Gypsophila sp.
Plate: 4A
Seed length: 1.25 mm
ID criteria: The seeds are almost circular in outline, with a short but projected radicle-tip.
The surface is papillose, presents the concentric rows of radially elongated verrucae.
Ecology: Huber-Morath in Davies (1967) mentions that the main distribution area of this
genus comprises the arid parts of Anatolia and adjacent areas. Rechinger (1988) classifies 41
species of this genus in Iran and adjacent areas.
Silene sp. (catchfly)
Plate: 4B
Seed length: 1.36 mm
ID criteria: The shape is kidney-shaped/reniform in outline, compressed together around the
hilum. Concentric rows of radially elongated wart-like projections on side faces and dorsal
face. Both features are heavily influenced through charring; especially dorsal surface is
completely missing in most cases.
Ecology: Wide range of ecological adaptations. Some species (Silene alba) are weeds
growing in refuse sites, field margins and such disturbed habitats. Some (silene vulgaris)
occurs in impoverished meadows, stony areas and forest margins.

104
CYPERACEAE (sedge family)
Scirpus cf. maritimus (sea club-rush)
Plate: 4C
Seed length: 1.95 mm
ID criteria: The nutlets are obovate-trigonous in outline, tapering towards the base. The
ventral side is flat, while the dorsal side is roof-shaped with a rounded median ridge. Single
fruit contents had also been found. The surface is smooth.
Ecology: Davies (1967) informs that the species occurs in “freshwater and saline marshes
with Typha, Juncus and Phragmites, stagnant swamps, water meadows with Orchis palustris,
by streams and rivers, dried up river beds, soda lakes, thermal springs, saline and alluvial
flats, beaches, edge of irrigation ditches and ricefields.” In Flora Iranica, Kukkonen in
Rechinger (1998) described the habitat of this species as “in salt marshes, in shallow water
brooks, rivers and lakes.”
FABACEAE (pulse family)
Lens sp. (lentil)
Plate: 4D
Seed length: varies between 1,93 mm and 2,76 mm; mean=2,17 mm; n=57
ID criteria: Seeds are lenticular in outline, slightly compressed at both sides with sharp or
blunt margins. The surface is smooth. The size class of Chogha Golan lentils corresponds to
the small-seeded race of wild lentil (Lens orientalis) rather than domestic lentils (Lens
culinaris) with larger sizes.
Ecology: The natural distribution area of the wild lentil comprises the Zagros Mountains in
Iran at elevations between 700 – 1700 m. (van Zeist et al. 1984). ABBO! On the other hand,

105
Chrtkova-Zertova in Rechinger (1979) reports only 3 species in Iran which are Lens cyanea,
Lens culinaris and Lens orientalis.
Vicia/Lathyrus type (vetch/grass pea)
Plate: 4E, 4F
Seed length: ~2.70 mm
ID criteria: Vicia/Lathyrus type seeds show large variations in shape; some objects are
almost spherical to biconvex types, while others are rounded-cubical. Hilum in most cases is
missing; which is considered an important attribute to distinguish various types of species in
these genera (Butler 1996). van Zeist and Bakker-Heeres (1982) mention the difficulty to
make a distinction between lenticular vetches and lentil seeds and also between small peas
and large, spherical vetch seeds. Most probably, several different vetches, grass pea and pea
species may have involved under this category. Pisum sp. (pea) is an important early crop
plant in Near Eastern agriculture but its identification was also problematic for the same
purposes. For that reason it was added up into this category.
Ecology: 51 Vicia species had been described in Iran and adjacent regions by Chrtkova-
Zertova in Rechinger (1979). Apart from that the genus Lathyrus has 25 species identified by
the same author. The genus Pisum is represented by only two species in Iran; P. sativum and
P. formosum. The latter species is a boreal form discovered in Mount Tuchal (2.400 mt.),
Tehran. The former P. sativum is divided into two subspecies subsp. sativum and subsp.
elatius.
Astragalus sp. (milk vetch)
Plate: 5A
Seed length: 1.66 mm
ID criteria: Typical specimens attributed to this genus are laterally compressed obliquely
quandrangular in outline (van Zeist and Bakker-Heeres, 1982). Anderberg (1994) and van
Zeist and Bakker-Heeres (1982) note that the seeds of Astragalus genus are variable in shape.

106
Hilum is usually sunken in the hilar notch that is mostly missing; the divergence of radicular
lobe is variable among specimens in the assemblage.
Ecology: The genus Astragalus is extremely rich in species diversity. Parsa (1948, after van
Zeist et al 1984) states that there are more than 550 species in Iran, that the majority of which
are dwarf shrubs.
Medicago radiata (ray-podded medick)
Plate: 5B
Seed length: 1.23 mm
ID criteria: Laterally compressed seeds, ovate to almost circular in outline. A characteristic
surface pattern is the irregular longitudinal wrinkles.
Ecology: Heyn in Davies (1970) mentions that the species appears in steppe and desert-steppe
vegetations.
Trigonella sp. (trigonel)
Plate: 5C, 5D
Seed length: varying from 2.21 to 1.68 mm
ID criteria: The seeds consist of two types of remains, which look morphologically similar to
each other. Trigonella astroides type remains are cylindrical in outline with transversely
irregular wrinkles on the surface. It is characterized by the truncated upper and lower ends. In
some specimens upper end is rounded. Apart from this, some seeds defined underthe category
of Trigonella sp., have also cylindrical shape in outline, while not truncated at both ends and
lacks wrinkles but finely punctuate on the surface.
These identification criteria should be taken seriously because of phenotypic similarities of
various pulse genera such as Melilotus, Trifolium and Medicago.

107
Ecology: Trigonella astroides appears to be an Irano-Turanian element, thrives in steppe
vegetations as well as cultivated fields (van Zeist and Bakker-Heeres, ???). Other Trigonella
species are also associated with steppic vegetations.
Indeterminate small-seeded pulses
Plate: 5E
Seed length: 1.73 mm
ID criteria: This category is composed of heavily carbonized or fragmented objects as well as
the remains that are not successfully attributed to one of the identified genera or any other
genera. This is mostly the result of the phenotypic plasticity of this plant family.
MALVACEAE (mallow family)
Malva sp. (mallow)
Plate: 5F
Seed length: 1.32 mm
ID criteria: The seed is reniform, has a deep hilar notch. The length of radicular lobe and
cotylenodary lobe is equal or the radicular lobe is slightly longer in some specimens. The seed
is the thinnest at the ventral side. The surface is smooth, mostly dorsally damaged by
carbonization. Mericarps of Malva sp. had been also recovered, which demonstrates radiaaly
favulariate surface patterns. This feature resembles that of M. nicaeensis or M. parviflora
species.
Ecology: Various mallow species are common in disturbed habitats such as fields, roadsides
or waste places. Riedl in Rechinger (1976) records 13 species of this genus in Iran and
adjacent regions.

108
POACEAE (grass family)
Aegilops sp. (goat-face grass)
Plate: 6A, 6B, 6C, 6D
Seed Length: 5.05 mm
ID criteria: The robust caryopsis is oblong-ovate in outline, that the longest breadth on the
dorsal side is usually seen above the proximal end. Parallel sided specimens are also present
and show smaller grain sizes than other specimens. The striations on the dorsal face are
recognizable. Most Aegilops objects are laterally grooved, while for some specimens, it is
already lost due to carbonization or fragmentation. Without exception, all objects display
wide ventral groove with often flattened ventral flanks. In cross-section, it is dorsally
compressed ventrally flattened. For too few specimens it is dorsally domed.
The rachis remains of this genus are highly fragmented into smaller objects, while complete
remains of spikelet bases and glume bases exist rather in low counts. All intact spikelets show
attributes of barrel-type disarticulation. Nesbitt (2006) notes that these type of disarticulation
are found in the modern species from the section Vertebrata (Ae. crassa, Ae. juvenalis, Ae.
tauschii, Ae. vavilovii, Ae. ventricosa) and Ae. cylindrica. Except Ae. vavilovii and Ae.
ventricosa, other 4 species appears in the inventory of Flora Iranica (Bor in Rechinger, 1970).
The glume tips are generally missing and no venation patterns on glumes is easily
recognizable. The attachment scars at the base triangular and sometimes quadrate.
Ecology: Thrives into stony slopes, dry grasslands and weedy places.
Hordeum cf. spontaneum (wild barley)
Plate: 6E, 6F, 6G, 6H, 6I, 6J
Seed Length: 6.18 mm
ID criteria: The caryopses of this species are narrowly elliptic/spindle shaped that the breadth
becomes wider above the scutellum. Dorsally compressed through a keel crossing 2/3 of the

109
grain and angular margins. These two dorsal features are not always present. Mostly laterally
and ventrally grooved.
Another type of objects defined as Hordeum distinchum type show close morphological
similarities with domesticated two-row barley species. The shape is almost circular in cross-
section, that the ventral and lateral grooves are not pronounced at all. This type of barley
grains had been also discussed in van Zeist et al. (1984) in Ganj Dareh account. Addiitionally,
some seeds remains show close resemblance to wild siblings of Hordeum spontaneum. Those
grouped under the category of “Hordeum sp.” in the dataset also includes the fragmented
Hordeum caryopses that has no clear identification attributes.
Ecology: Today, wild barley (Hordeum spontaneum) is massively spread over southwest Asia
where Fertile Crescent constitutes its primary habitat. It is considered as a part of the rich
grass cover associated with open-woodland Quercus brantii belt occurs at elevations from
500 to 1500 meters in the Zagros region of Iraq and Iran. It does not tolerate extreme cold
conditions while withstands drier and warmer environments, poorer soils and some salinity.
Bor in Rechinger (1970) stresses the weedy habit of this species as well, noting that it thrives
in “deserts, gravelly, sandy or silky soils; in waste places and invading cultivation”
Economy: Two domesticated types of barley serve as principal crop plants for world
economy. Most barley cultivars are of the “hulled” variety which is mainly used for different
purposes such as animal fodder or the production of malt for brewing. The other form of
barley cultivar is “naked” barley principally makes use of human food because of its ease of
processing and edibility. This type is mostly prevalent staple food in the high altitude areas of
Nepal and Tibet, whereas its cultivation is very limited in southwest Asia and Europe (Lister
and Jones 2012).
Triticum sp. (wheats)
Plate: 7A, 7B, 7C, 7D, 7G
Seed length: 4.95 mm
ID criteria: Only one specimen of Triticum sp. (wheat genus) seed remains is recovered from
AH III. Preliminary examinations demonstrate that this finding may belong to T. monococcum
ssp. boeticum (wild einkorn) and/or T. turgidum ssp. dicoccoides (wild emmer). Several

110
specimens of spikelet bases had been recovered. No further analysis to attribute the
domestication status had been performed but Riehl et al. (2013) clearly identified wild and
domesticated emmer in the findings.
One specimen of Triticum sp. that is found in AH III shows close similarities to the free-
threshing type of spikelet bases. The morphological characteristics resemble T. aestivum
(bread wheat) rather than T. durum (hard wheat) type. In addition, the abscission scar of this
particular spikelet base is smoothly broken, not resembling the non-shattering domesticated
types (Plate 7G).
Ecology: Zohary and Hopf (2000) notes that wild emmer grow “as common annual
components in the herbaceous cover of oak park-forest belt and related steppe-like herbaceous
plant formations. They are confined to basaltic and hard limestone bedrocks and completely
absent on marls and chalks. In rocky places which have not been severely overgrazed,
dicocoides wheat often grows in large stands […] Whereas dicocoides wheat occurs alone in
Syrian-Palestine area, it grows sympatrically with a second wild tetraploid wheat T.
timopheevi subsp. araraticum in the northeastern part of its distribution area.”
Economy: Today, bread wheat is one of the most important crops for world economy among
other domesticated cereals such as rice, maize. It accounts for about 95 % of global wheat
production. Additionally, about 20 % of the total human food calories are provided by this
cultivar (Peng et al. 2011).
Bromus sp. (bromegrass)
Seed length: 3.67 mm
ID criteria: Dorsally compressed in cross-section. Distinct ventral grooves, always flattened.
Many objects are somehow quadrate in cross section, while others have wide and deep ventral
furrows. V-shaped scutellum is not recognizable due to fragmented nature of these remains.
Ecology: A large genus including about 150 species worldwide. 44 of them are reported from
Iran and adjacent regions (Bor in Rechinger 1970). Such large species diversity hampers
taxonomic identifications even in botany (Nesbitt 2006). Stebbins (1981; after Nesbitt 2006)
suggests that the annual species of Bromus may have evolved rapidly from perennial species

111
in section Phigma, through adaption to agriculture, particularly grazing by animals. The
annual species are weeds of disturbed ground and arable fields.
Taeniatherum caput-medusae (medusahead)
Plate: 7E, 7F
Seed length: ~6.16 mm
ID criteria: Narrow and elongated caryopses. Apex rounded. Dorsally compressed. The
ventral grooves shallow and wide or deep and narrow. Lateral groves are distinct features in
every specimen. The spikelet bases mostly found with glumes still attached.
Ecology: Both subspecies of this plant (ssp. asper and ssp. crinitum) are reported in Flora
Iranica (Bor in Rechinger, 1970). Thrives into sandy places in the mountains or in sandy, dry
places.
Indeterminate large-seeded grasses
Plate: 7H
Seed length: 3.87 - 3.27 mm
ID criteria: This group of caryopses includes all grass remains left out unidentified during the
identification process. This category is relatively large including many different types of
caryopses. Morphological attributes of one type of these remains display similarities with the
identification made by van Zeist and Baaker-Heeres in Bronze Age Selenkahiye and Hadidi as
“Gramineae type B”. This type of caryopses has slender shape in outline, almost parallel
lateral sides and it has a circular shape in cross section. A ventral groove generally is not
present.

112
Agrostis type
Plate: 8A
Seed length: 1.07 – 0.99 mm
ID criteria: This type of remains has no modern equivalent described in the literature.
Following the description of van Zeist et al. (1984), it is defined as Agrostis type due to close
morphological similarities with Agrostis genus. It is important to note that more than one
species and genus may have involved in the assemblage.
The shape of the caryopses is mostly ellipsoid in outline; dorsally compressed, attenuate
towards apex but always rounded at the apex. Circular-ellipsoid in cross-section. The hilum is
missing in most specimens. van Zeist et al.‟s (1984) description show sub-basal linear hilum.
The effects of carbonization are mostly seen on the ventral side that the caryopsis puffed up
open.
Ecology: Possibly weed of disturbed habitats.
Triticoid type
Plate: 8B
Seed length: 2.36 mm – 2.72 mm
ID criteria: No modern equivalent exists for this type of remains. van Zeist et al. (1984)
described this sort of caryopses while there are also recognizable morphological variations
and size differences among the remains. The authors also mention that although it is named
after the resemblance of Triticum genus, it is also highly likely that this type of plants belongs
to another genus.
One pattern appears to be common. The caryopses with flat ventral surface are dorsally and
laterally curved at the apex while other specimens with deep ventral furrow are somehow
dorsally compressed and slender in outline. Striations which are caused by stiff glumes are
visible in many objects. The carbonization affects most the lateral surfaces of the remains.
Ecology: Possibly weeds of disturbed habitats.

113
Phalaris sp. (canary grass)
Plate: 8C
Seed length: 1.50 mm
ID criteria: Laterally densely compressed caryopses with longitudinally curved ventral and
dorsal sides. Relatively large radicle shield extending over 1/4 to 1/3 of the dorsal side. Apical
and basal ends rounded; some specimens have slender morphology with less laterally
compressed outline and pointed apex. More than one species must have involved in the
assemblage.
Ecology: Usually dry weedy places. P. arundinaceae prefers moist soils. Five species are
described in Flora Iranica (Bor in Rechinger 1970). Riehl (1999) informs that the grains of
Phalaris minor are very nutritious, but more suitable as animal fodder than human nutrition
because of the difficulty to harvest.
Unidentified Poaceae rachis type
One group of rachis remains appears to be the third numerous rachis finds in the assemblage
after Hordeum and Aegilops chaffs. “Unidentified Poaceae rachis type” comprises one type of
object that is not successfully identified in this research. No relevant information had been
found in archaeobotanical literature about this type of remains. Given relatively small sizes of
this type of chaff remains, it can possibly belong to small-seeded taxa of grasses.
Plate: 8D
ID criteria: Nesbitt (2006) notes that “the fertile spikelet of Phalaris paradoxa is subtended
by deformed or reduced sterile spikelets, which sometimes survive charring”. Interestingly,
the percentages of these unidentified objects follow more or less the same trend of Phalaris
finds in the assemblage. However, ne any other reliable criterion is present to identify these
remains.

114
RUBIACEAE (bedstraw family)
Galium sp. (cleavers)
Plate: 8E
Seed length: ~0.79 in diameter.
ID criteria: The seed is more or less spherical in outline with a round concavity on the ventral
side indicating the position of hilum. On the surface there are slight humps in some
specimens, others are smooth.
Ecology: Galium is a large genus, mainly described as “typical weeds” in the
archaeobotanical literature (Riehl 1999).
PAPAVERACEAE (poppy family)
Papaver sp. (poppy)
Plate: 8F
Seed length: 0.66 mm
ID criteria: The shape is broadly reniform in outline with distinct reticulate surface structure.
Dorsal side convex, ventral side concave. Upper and lower lobes are usually equally long.
Ecology: Annual, biennial and perennial herbs. A genus with 30 species over Iran and
adjacent regions (Cullen in Rechinger, 1966). Papaver somniferum is a cultivated species
grown for medicinal, culinary and narcotic purposes (Bojnansky and Fargasova 2007). Other
species are usually described as weeds in cultivated fields and waste places.
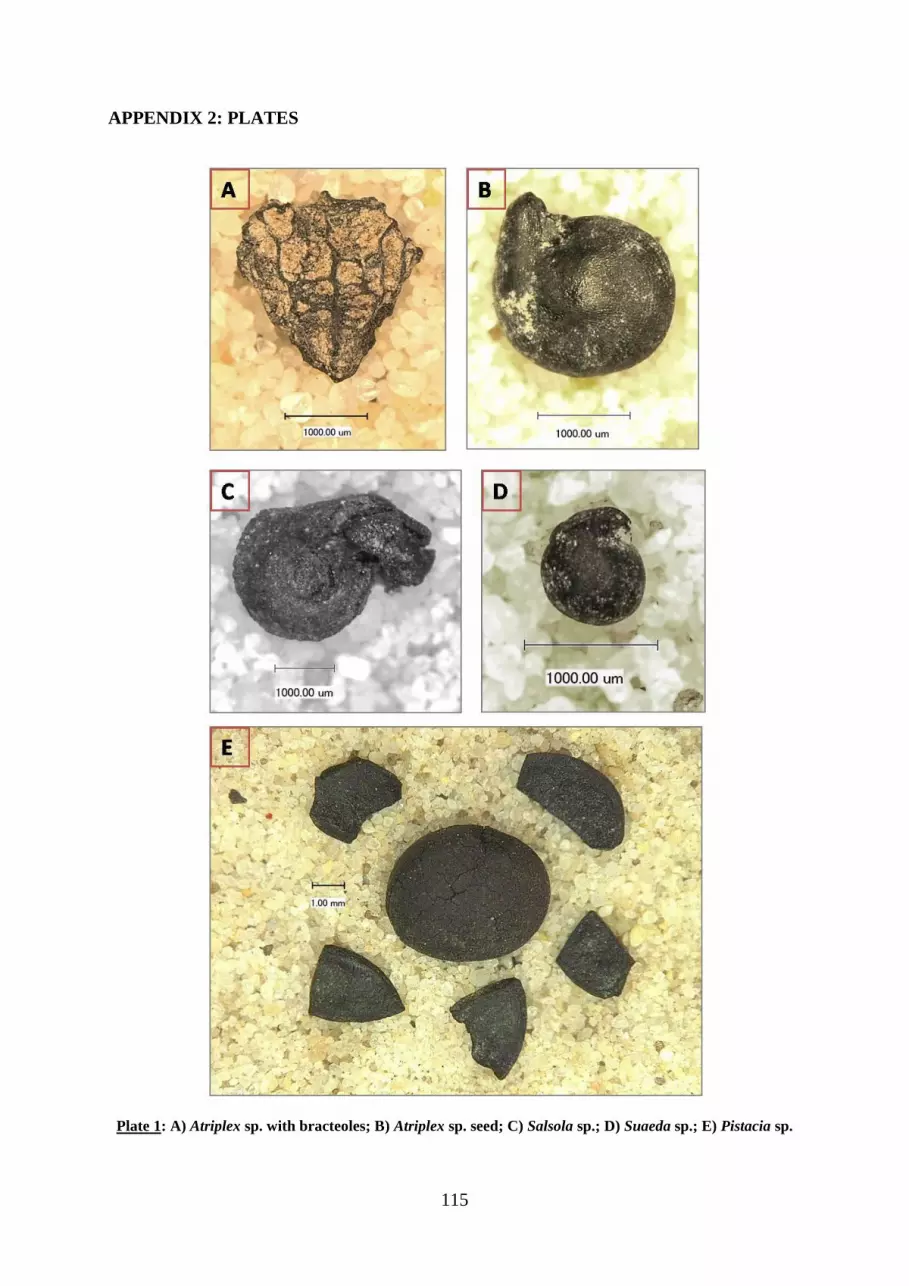
115
APPENDIX 2: PLATES
Plate 1: A) Atriplex sp. with bracteoles; B) Atriplex sp. seed; C) Salsola sp.; D) Suaeda sp.; E) Pistacia sp.

116
Plate 2 A) Centaurea type three-rimed specimens; B) Centaurea type small-headed specimen; C) Arnebia
sp. D) Heliotropium sp.; E) Lithospermum sp. F) Ornithagalum/Muscari.

117
Plate 3 A) cf. Arabis sp.; B) Brassica type C) Lepidium sativum.

118
Plate 4 A) Gypsophila sp.; B) Silene sp., C) Scirpus cf. maritimus; D) Lens sp.; E) Vicia/Lathyrus type; F)
Large-seeeded pulse specimens.

119
Plate 5 A) Astragalus sp.; B) Medicago radiata; C) Trigonella sp.; D) Trigonella astroides; E) Indeterminate
small-seeded pulses; F) Malva sp.

120
Plate 6 A – C) Aegilops sp. grain; D) Aegilops rachis; E – F) Hordeum cf. spontaneum; G) Hordeum
rachis internodes, smooth scar; H – I) Hordeum distinchum type; J) Hordeum rachis internodes; rough
scar.

121
Plate 7 A - C) Triticum sp. grain; D) Triticum spikelet base; E) Taenitherium cf. caput-medusae; F)
Taenitherium cf. caput-medusae rachis remain; G) Hexaploid type spikelet base; H) Indeterminate
medium-seeded grasses.

122
Plates 8 A) Agrostis type; B) Triticoid type; C) Phalaris type; D) Unidentified Poaceae rachis type; E)
Galium sp.; F) Papaver sp.

123
APPENDIX 3: MACROBOTANICAL RAW DATA
Cultural Layer I I IIb IIb IIb Trench 1/99 1/0 1/1 1/1 1/99 Botanical Sample No 106 168 160 137 147 Finding Code BFb BFd BFd BFc BFd Exact Provenance (x, y, z) 75, 75, 18.65 75, 25, 18.71 75, 25, 17.90 25, 25, 18.07 75, 60, 17.99 Sediment Volume (ml) 10000 10000 14000 12000
Excavator MZ MZ MZ MZ MZ
Plant Name Absolute counts
I-106 I-168 II-160 II-137 II-147
Hordeum spontaneum C.Koch 1
Hordeum distichum-type 1
Hordeum sp.
Hordeum fragments
Hordeum sp. (smooth incision) 2 10 10
Hordeum sp. (rough incision)
Hordeum sp. (lower part of spikelet) 3 1 3 13
Aegilops/Hordeum
Aegilops sp.
Aegilops sp. (spikelet base) 4 7
Aegilops sp. (glume base) 6 12 23 68 123
Unidentified Poaceae grain type
Unidentified Poaceae spikelet base type 2 4 4 58
Phalaris sp. 2 5 10 17 27
Triticoid type 1 1 1
Hulled Triticum type
Triticum type free-threshing grain
Triticum type spikelet bases 4 1 7 2 7
Triticum type glume bases 6 5 12 4 48
Free-threshing wheat spikelet base
Taeniatherum caput-medusae 1
Taeniatherum sp. (spikelet bases)
Bromus sp.
Poaceae, indet. (large, medium) 1 1 5
Poaceae, indet. (small) 4 6 26 66 91
Poaceae, indet. (grain fragments) 5
Lens sp. 2 1 4 3
Vicia/Lathyrus/Pisum 1 1 9 2
Astragalus sp. 2 3 10 17 9
Trigonella astroides 2 4 30 6
Trigonella sp. 5 4 15 27
Medicago sp.
Medicago radiata 1 1 8 2
Fabaceae, indet.(medium)
Fabaceae, indet.(small) 3 13 10
Silene sp. 1
Gypsophila sp.
Caryophyllaceae, indet. 1
Scirpus/Bolbochenius sp. 5 3
Malva sp. 2 3 2 5
Heliotropium sp. 1 1 1
Lithospermum sp. 1 1
Arnebia sp.
Boraginaceae, indet. 1
Pistacia sp. 1 2 3 13 10
Brassica type 2
Brassicaceae, indet. 1 4 4
Alyssum type
cf. Arabis
cf. Lepidium sativum
cf. Lepidium
Ornithogalum/Muscari/Bellevalia 1
Galium sp. 1
Centaurea type 1
Salsola sp.
Atriplex sp.
Suaeda sp.
Beta sp.
Chenopodiceae, indet. 4
Rumex/Polygonum 4
Papaver sp.
Adonis sp.
TOTAL 43 44 118 314 476
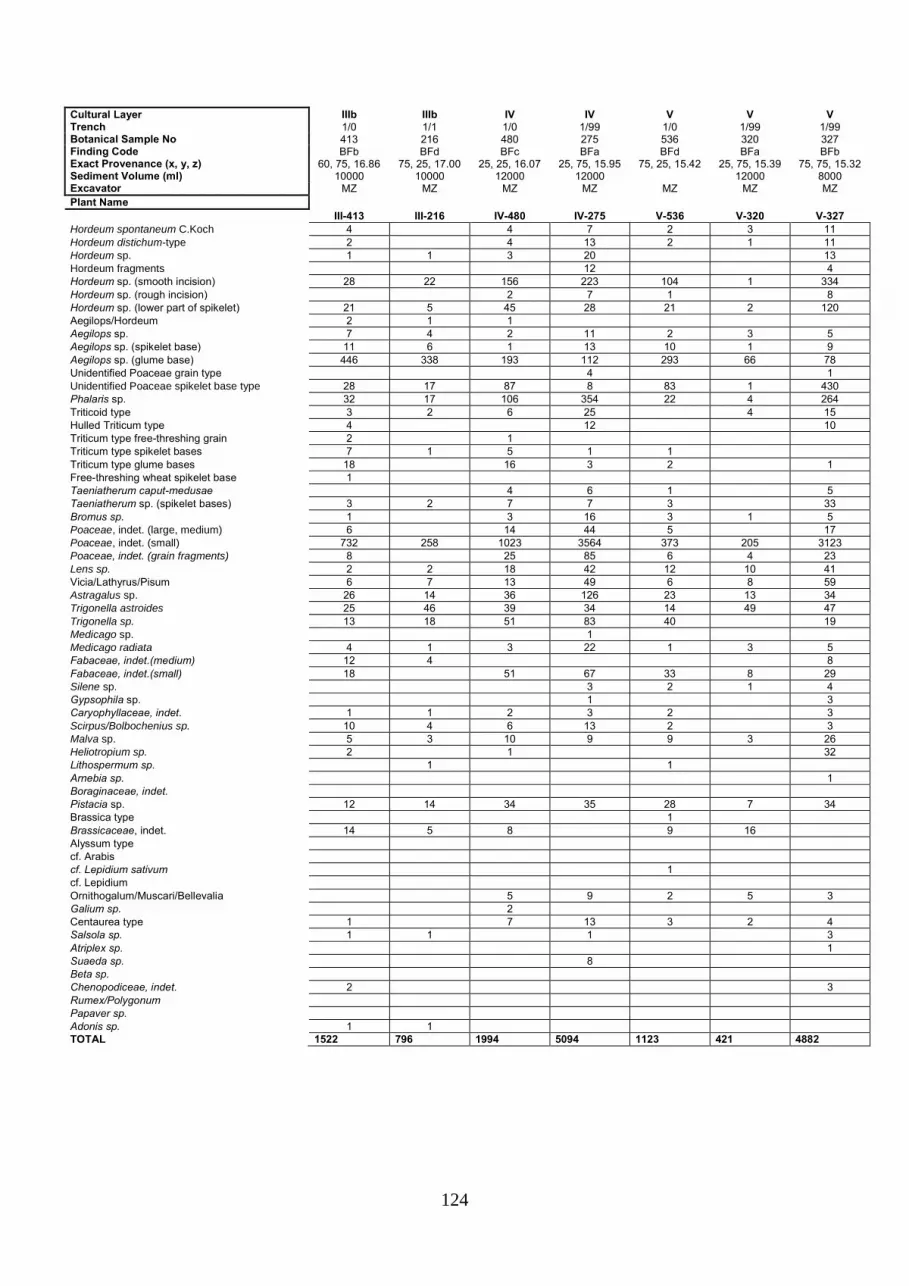
124
Cultural Layer IIIb IIIb IV IV V V V Trench 1/0 1/1 1/0 1/99 1/0 1/99 1/99 Botanical Sample No 413 216 480 275 536 320 327 Finding Code BFb BFd BFc BFa BFd BFa BFb Exact Provenance (x, y, z) 60, 75, 16.86 75, 25, 17.00 25, 25, 16.07 25, 75, 15.95 75, 25, 15.42 25, 75, 15.39 75, 75, 15.32 Sediment Volume (ml) 10000 10000 12000 12000
12000 8000
Excavator MZ MZ MZ MZ MZ MZ MZ
Plant Name
III-413 III-216 IV-480 IV-275 V-536 V-320 V-327
Hordeum spontaneum C.Koch 4 4 7 2 3 11
Hordeum distichum-type 2 4 13 2 1 11
Hordeum sp. 1 1 3 20 13
Hordeum fragments 12 4
Hordeum sp. (smooth incision) 28 22 156 223 104 1 334
Hordeum sp. (rough incision) 2 7 1 8
Hordeum sp. (lower part of spikelet) 21 5 45 28 21 2 120
Aegilops/Hordeum 2 1 1
Aegilops sp. 7 4 2 11 2 3 5
Aegilops sp. (spikelet base) 11 6 1 13 10 1 9
Aegilops sp. (glume base) 446 338 193 112 293 66 78
Unidentified Poaceae grain type 4 1
Unidentified Poaceae spikelet base type 28 17 87 8 83 1 430
Phalaris sp. 32 17 106 354 22 4 264
Triticoid type 3 2 6 25 4 15
Hulled Triticum type 4 12 10
Triticum type free-threshing grain 2 1
Triticum type spikelet bases 7 1 5 1 1
Triticum type glume bases 18 16 3 2 1
Free-threshing wheat spikelet base 1
Taeniatherum caput-medusae 4 6 1 5
Taeniatherum sp. (spikelet bases) 3 2 7 7 3 33
Bromus sp. 1 3 16 3 1 5
Poaceae, indet. (large, medium) 6 14 44 5 17
Poaceae, indet. (small) 732 258 1023 3564 373 205 3123
Poaceae, indet. (grain fragments) 8 25 85 6 4 23
Lens sp. 2 2 18 42 12 10 41
Vicia/Lathyrus/Pisum 6 7 13 49 6 8 59
Astragalus sp. 26 14 36 126 23 13 34
Trigonella astroides 25 46 39 34 14 49 47
Trigonella sp. 13 18 51 83 40 19
Medicago sp. 1
Medicago radiata 4 1 3 22 1 3 5
Fabaceae, indet.(medium) 12 4 8
Fabaceae, indet.(small) 18 51 67 33 8 29
Silene sp. 3 2 1 4
Gypsophila sp. 1 3
Caryophyllaceae, indet. 1 1 2 3 2 3
Scirpus/Bolbochenius sp. 10 4 6 13 2 3
Malva sp. 5 3 10 9 9 3 26
Heliotropium sp. 2 1 32
Lithospermum sp. 1 1
Arnebia sp. 1
Boraginaceae, indet.
Pistacia sp. 12 14 34 35 28 7 34
Brassica type 1
Brassicaceae, indet. 14 5 8 9 16
Alyssum type
cf. Arabis
cf. Lepidium sativum 1
cf. Lepidium
Ornithogalum/Muscari/Bellevalia 5 9 2 5 3
Galium sp. 2
Centaurea type 1 7 13 3 2 4
Salsola sp. 1 1 1 3
Atriplex sp. 1
Suaeda sp. 8
Beta sp.
Chenopodiceae, indet. 2 3
Rumex/Polygonum
Papaver sp.
Adonis sp. 1 1
TOTAL 1522 796 1994 5094 1123 421 4882

125
Cultural Layer VI VI VII VII VII VII Trench 1/99 1/0 0/99 1/99 1/99 1/0 Botanical Sample No 342 555 52 366 367 613 Finding Code BFa BFc BFb BFa BFa pLa Exact Provenance (x, y, z) 25, 75, 14.13 25, 25, 14.99 75, 55, 15.00 25, 75, 14.74 25, 75, 14.72 45, 68, 14.49 Sediment Volume (ml) 12000 12000 5000 10000 10000 10000 Excavator MZ MZ MZ MZ MZ MZ
Plant Name
VI-342 VI-555 VII-52 VII-366 VII-367 VII-613
Hordeum spontaneum C.Koch 3 1 8 4 5 3
Hordeum distichum-type 2 3
Hordeum sp. 4 2 4 2
Hordeum fragments 2
Hordeum sp. (smooth incision) 28 25 30 15 19 15
Hordeum sp. (rough incision)
Hordeum sp. (lower part of spikelet) 16 4 32 3
Aegilops/Hordeum
Aegilops sp. 3 4 3 2 3 1
Aegilops sp. (spikelet base) 9 2 2 1 2 5
Aegilops sp. (glume base) 362 69 49 74 69 52
Unidentified Poaceae grain type
Unidentified Poaceae spikelet base type 63 5 4 2 2
Phalaris sp. 8 4 4 3 6
Triticoid type 3 4 3 1 2
Hulled Triticum type 1
Triticum type free-threshing grain
Triticum type spikelet bases 2 5
Triticum type glume bases 1 1 24
Free-threshing wheat spikelet base
Taeniatherum caput-medusae 1 1 1 1
Taeniatherum sp. (spikelet bases) 1 7 2
Bromus sp. 2 2 3 18 12
Poaceae, indet. (large, medium) 2 13 10 5 14
Poaceae, indet. (small) 173 78 64 16
Poaceae, indet. (grain fragments) 6 8 4
Lens sp. 4 31 11 32 20 8
Vicia/Lathyrus/Pisum 14 13 8 11 12 5
Astragalus sp. 31 34 8 28 27 20
Trigonella astroides 41 29 13 12 11 29
Trigonella sp. 12 6 5 11 5 7
Medicago sp.
Medicago radiata 3 2 2 2
Fabaceae, indet.(medium) 2 3
Fabaceae, indet.(small) 67 20 13 31 38 15
Silene sp. 4 1 4 3
Gypsophila sp. 1
Caryophyllaceae, indet. 3 2 3 7 2
Scirpus/Bolbochenius sp. 2 1 1 1 4
Malva sp. 4 4 3 3 4 1
Heliotropium sp. 1 2
Lithospermum sp. 1
Arnebia sp. 1
Boraginaceae, indet. 1 1
Pistacia sp. 8 1 1 3
Brassica type
Brassicaceae, indet. 16 12 11 2 14
Alyssum type 1
cf. Arabis
cf. Lepidium sativum 13
cf. Lepidium
Ornithogalum/Muscari/Bellevalia 7 8 12 8 5
Galium sp. 1 1 1
Centaurea type 5 6 3 1 3
Salsola sp. 2 1
Atriplex sp. 5 4 3
Suaeda sp. 8 1 1
Beta sp.
Chenopodiceae, indet. 16 4 13 10
Rumex/Polygonum
Papaver sp. 28
Adonis sp. 1 2
TOTAL 944 409 326 314 288 256

126
Cultural Layer VIII VIII IX IX IX Trench 1/0 1/99 1/99 1/0 1/0 Botanical Sample No 601 390 403 625 636 Finding Code BFc BFa BFb BFc BFd Exact Provenance (x, y, z) 25, 25, 14.37 25, 75, 14.39 75, 75, 14.17 25, 25, 14.08 75, 25, 13.95 Sediment Volume (ml) 8000 12000 10000 10000 10000 Excavator MZ MZ MZ MZ MZ
Plant Name
VIII-601 VIII-390 IX-403 IX-625 IX-636
Hordeum spontaneum C.Koch 2 6 1
Hordeum distichum-type 3 7 2
Hordeum sp. 6 1
Hordeum fragments
Hordeum sp. (smooth incision) 9 52 13 5 29
Hordeum sp. (rough incision) 2 1
Hordeum sp. (lower part of spikelet) 4 7 18 1
Aegilops/Hordeum 1 1
Aegilops sp. 2 1 1
Aegilops sp. (spikelet base) 2 9 7 2
Aegilops sp. (glume base) 166 208 51 28 45
Unidentified Poaceae grain type 1
Unidentified Poaceae spikelet base type 5 61 1 1 1
Phalaris sp. 3 7 7 1 2
Triticoid type 3 1 1
Hulled Triticum type 2 1
Triticum type free-threshing grain
Triticum type spikelet bases
Triticum type glume bases
Free-threshing wheat spikelet base
Taeniatherum caput-medusae 1
Taeniatherum sp. (spikelet bases) 3
Bromus sp. 3 1 1
Poaceae, indet. (large, medium) 5 4 3 2
Poaceae, indet. (small) 4 113 80 7
Poaceae, indet. (grain fragments) 7 6
Lens sp. 8 6 2 5 9
Vicia/Lathyrus/Pisum 2 6 10 3 5
Astragalus sp. 12 94 14 3 21
Trigonella astroides 51 18 6 5 20
Trigonella sp. 10 15 23
Medicago sp.
Medicago radiata 1 2
Fabaceae, indet.(medium) 1
Fabaceae, indet.(small) 9 29 12 2 10
Silene sp. 3
Gypsophila sp. 1
Caryophyllaceae, indet. 4 1 1
Scirpus/Bolbochenius sp. 1 6 2 1
Malva sp. 4 15 6 2 20
Heliotropium sp. 3 3
Lithospermum sp. 1
Arnebia sp.
Boraginaceae, indet. 1
Pistacia sp. 4 31 7 1 9
Brassica type
Brassicaceae, indet. 8 8 1 1
Alyssum type
cf. Arabis
cf. Lepidium sativum
cf. Lepidium
Ornithogalum/Muscari/Bellevalia 1 2 3
Galium sp. 1 1
Centaurea type 1 1
Salsola sp. 1
Atriplex sp. 1
Suaeda sp.
Beta sp.
Chenopodiceae, indet. 1 1
Rumex/Polygonum
Papaver sp.
Adonis sp.
TOTAL 306 743 289 66 200
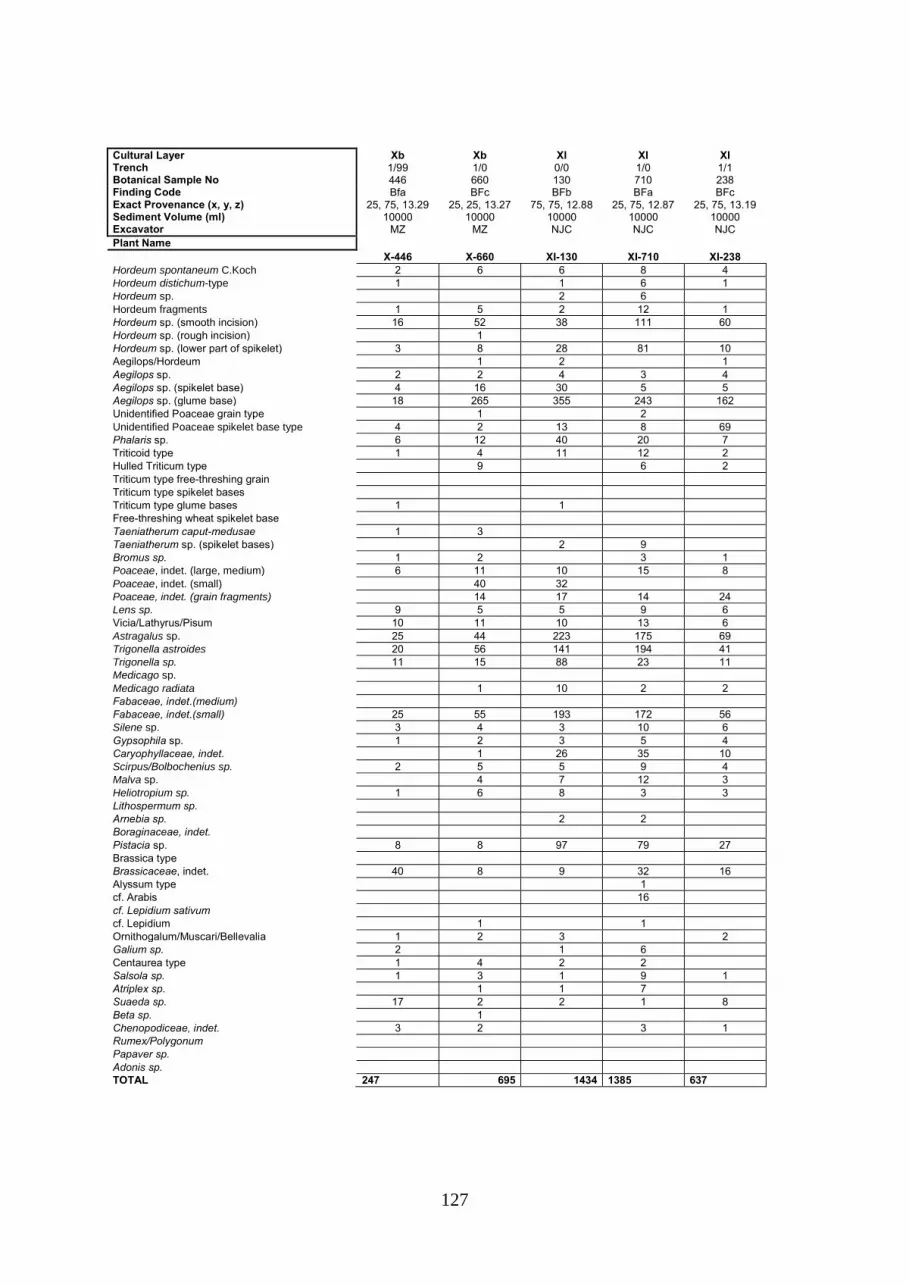
127
Cultural Layer Xb Xb XI XI XI Trench 1/99 1/0 0/0 1/0 1/1 Botanical Sample No 446 660 130 710 238 Finding Code Bfa BFc BFb BFa BFc Exact Provenance (x, y, z) 25, 75, 13.29 25, 25, 13.27 75, 75, 12.88 25, 75, 12.87 25, 75, 13.19 Sediment Volume (ml) 10000 10000 10000 10000 10000 Excavator MZ MZ NJC NJC NJC
Plant Name
X-446 X-660 XI-130 XI-710 XI-238
Hordeum spontaneum C.Koch 2 6 6 8 4
Hordeum distichum-type 1 1 6 1
Hordeum sp. 2 6
Hordeum fragments 1 5 2 12 1
Hordeum sp. (smooth incision) 16 52 38 111 60
Hordeum sp. (rough incision) 1
Hordeum sp. (lower part of spikelet) 3 8 28 81 10
Aegilops/Hordeum 1 2 1
Aegilops sp. 2 2 4 3 4
Aegilops sp. (spikelet base) 4 16 30 5 5
Aegilops sp. (glume base) 18 265 355 243 162
Unidentified Poaceae grain type 1 2
Unidentified Poaceae spikelet base type 4 2 13 8 69
Phalaris sp. 6 12 40 20 7
Triticoid type 1 4 11 12 2
Hulled Triticum type 9 6 2
Triticum type free-threshing grain
Triticum type spikelet bases
Triticum type glume bases 1 1
Free-threshing wheat spikelet base
Taeniatherum caput-medusae 1 3
Taeniatherum sp. (spikelet bases) 2 9
Bromus sp. 1 2 3 1
Poaceae, indet. (large, medium) 6 11 10 15 8
Poaceae, indet. (small) 40 32
Poaceae, indet. (grain fragments) 14 17 14 24
Lens sp. 9 5 5 9 6
Vicia/Lathyrus/Pisum 10 11 10 13 6
Astragalus sp. 25 44 223 175 69
Trigonella astroides 20 56 141 194 41
Trigonella sp. 11 15 88 23 11
Medicago sp.
Medicago radiata 1 10 2 2
Fabaceae, indet.(medium)
Fabaceae, indet.(small) 25 55 193 172 56
Silene sp. 3 4 3 10 6
Gypsophila sp. 1 2 3 5 4
Caryophyllaceae, indet. 1 26 35 10
Scirpus/Bolbochenius sp. 2 5 5 9 4
Malva sp. 4 7 12 3
Heliotropium sp. 1 6 8 3 3
Lithospermum sp.
Arnebia sp. 2 2
Boraginaceae, indet.
Pistacia sp. 8 8 97 79 27
Brassica type
Brassicaceae, indet. 40 8 9 32 16
Alyssum type 1
cf. Arabis 16
cf. Lepidium sativum
cf. Lepidium 1 1
Ornithogalum/Muscari/Bellevalia 1 2 3 2
Galium sp. 2 1 6
Centaurea type 1 4 2 2
Salsola sp. 1 3 1 9 1
Atriplex sp. 1 1 7
Suaeda sp. 17 2 2 1 8
Beta sp. 1
Chenopodiceae, indet. 3 2 3 1
Rumex/Polygonum
Papaver sp.
Adonis sp.
TOTAL 247 695 1434 1385 637