an anatomical study of the mechanical interactions of flexor digitorum superficialis and profundus...
TRANSCRIPT

A N A N A T O M I C A L S T U D Y OF THE M E C H A N I C A L I N T E R A C T I O N S OF F L E X O R D I G I T O R U M S U P E R F I C I A L I S A N D P R O F U N D U S A N D THE F L E X O R T E N D O N S H E A T H IN
Z O N E 2
E. T. WALBEEHM and D. A. McGROUTHER
From the Department of Plastic and Reconstructive Surgery, Rayne Institute, University College of London and Middlesex Medical Schools, London, UK :
The mechanical interactions of tendon loading and motion between FDS and FDP tendons and the distal edge of the A2-pulley (DEA2) were investigated in eadaveric hands.
The FDP tendon showed a two-stranded contrarotational spiral of tendon fibres, their course parallel to the FDS fibres. On loading, the FDP tendon changed cross-sectional shape and the two slips of FDS moved closer together, applying lateral force on the FDP tendon.
The formula DEA2-MCP/proximal phalanx length was calculated as 0.64___0.052 mm/mm. Excursions of bifurca and chiasma were measured relative to the DEA2. In full extension the bifurca was well proximal to the DEA2, the chiasma always distal. In flexion, the chiasma approximated to DEA2 level. Therefore, an additional anteroposterior force should exist at DEA2 level between FDS and the DEA2 on the FDP-tendon. The changes in tendon shape and the lateral- and anteroposterior forces produce a "compressional" mechanism on FDP by FDS which may alter frictional resistance. This mechanism is compared with the Chiropteran tendon locking mechanism. Journal of Hand Surgery (British and European Volume, 1995) 20B: 3." 269-280
Despite intensive research activity, Sterling Bunnell's "No Man's Land", zone 2 in flexor tendon injuries, is still associated with a high failure rate after tendon repair compared to the other zones of the hand (Manske and Lester, 1982; Strickland, 1989 a and b; Boulas and Strickland, 1993; Tang, 1994).
Morphologically, this area is considered to show little anatomical variation. This would suggest that it there- fore has a specific function (Shrewsbury and Kuczinsky, 1974). The functional anatomy, however, remains incompletely understood, particularly in the region of the decussation of FDS.
A review of historical anatomy showed beautifully illustrated and very accurate descriptions of the entire flexor mechanism by Leonardo da Vinci in 1510 (O'Malley and Saunders, 1952; Shrewsbury and Kuczinsky, 1974), Albinus (1734; 1747), Weitbrecht (1742) and Camper (1760), whose name is attached to the dorsal decussation of FDS (Figs 1 and 2). Their drawings illustrated in meticulous detail the flexor tendon sheath with the pulleys and the course of the fibres of both flexor tendons. The old masters, however, could not, or at least did not, explain the functional implications and often remained purely descriptive. Da Vinci, as engineer, was intrigued by the biomechanics involved in a perforator/perforans system. More recently a number of authors, Martin (1958), Field (1971), Hueston and Wilson (1972, 1973), Wilson and Hueston (1973), Shrewsbury and Kuczinsky (1974), and Enna and Dyer (1976) revisited tendon anatomy in very thorough morphological studies.
Flexor tendon sheath (FTS) (Figs 1, 2 and 3)
The FTS has been recently redescribed by a number of authors (Simmons and de la Caffini6re, 1981; Idler, 1985; Doyle, i988; Lin et al, 1989), although most of the detail was apparent in the works of Albinus. The sheath is generally accepted to consist of the sheath and the pulleys. The most important pulleys, biomechan- ically, are the A2 and A4 pulleys (Simmons and de la Caffini6re, 1981; Doyle, 1988).
Our study has focused on the A2 pulley, which is situated entirely on the proximal phalanx (PP), mainly the more proximal part. This pulley is the strongest of the pulleys, with increasing thickness from proximal to distal. The A2 is strongly attached to the PP with a fibro-osseous junction, in particular the distal edge of the A2 pulley (DEA2).
The general view on the function of the pulley system is that it prevents bowstringing of the two tendons when the finger is actively flexed. When imagining a finger in the fully flexed position, mostly used for powergrip (Backhouse, 1968; Long, 1968; Simmons and de la Caffini~re, 1981), it is obvious that forces on the DEA2 are potentially considerable.
269
Flexor digitorum superficialis and profundus
To understand how anatomical details contribute to manual mechanisms we have studied the intratendinous architecture and its functional implications in tendon loading, the positions of the bifurca and Camper's

270 THE JOURNAL OF HAND SURGERY VOL. 20B No. 3 JUNE 1995
Fig 1
b
(a and b) Copies of original drawings of a dissection performed by Bernard Siegfried Albinus (1734), drawn by Jan Wandelaar. Reproduced by the kind permission of the Trustee of the Wellcome Trust from the Historia Musculorwn Hominis by B. S. Albinus (1734).
chiasma in relation to the DEA2 in different fingerposi- tions (Brand et al, 1981; McGrouther and Ahmed, 1981; Brand, 1985) and the involved tendon and pulley surfaces.
MATERIAL AND M E T H O D S
Six fixed and five unfixed cadaveric hands were dissected. One hand dissected in detail to develop the experimental method, and a so called "blue hand" (a hand perfused with indian ink, so that the bloodvessels are visible) were excluded from the statistical analysis. Eight were right hands and three were left. The fixed hands were obtained from cadavers which were fixed with preser- vation fluid (20 L) containing 39% to 41% formaldehyde solution (4 L), 80% phenol solution (2 L), 99% methanol solution (2 L), Teepol detergent (50 ml) and 16 L tap- water. To soften the tissues in order to give suppler hands and fingers, enabling us to do the measurements with more ease, we used the technique of Blaney and Johnson (1989). The hands were soaked in concentrated
Comfort (fabric softener) diluted in tapwater (30 ml/5 1 water) for 7 days.
The unfixed hands were obtained from fresh cadavers and had been frozen at the time of dissection for periods varying between 2 weeks and 9 months.
For accurate dissection and measurements an operating microscope (Zeiss) was used with 10 x to 40 x magnification. Measurements were taken with a Vernier gauge with an accuracy of 0.1 mm and a calliper. The MP, PIP and the DIP joints were marked with a small intraarticular hypodermic needle.
The fingers were dissected by removal of the palmar skin distal to the proximal palmar crease. The flexor tendon sheath was exposed and the C1 and C2 pulleys removed. The annular pulleys were left intact, with the exception of a small window created in the A2 pulley in order to be able to perform the measurements accurately. The distal edge of the A2 pulley (DEA2) was marked to serve as a point of reference for measurements of tendon dimensions and motion. The distance of DEA2 to MP joint was recorded. Other constant points on the

FDS, FDP AND THE FLEXOR SHEATH 271
a
Fig 2
Fig 3
Copies of original drawings of a dissection performed by Petrus Camper (1760), whose name is attached to the dorsal chiasma tendinosum of FDS. Reproduced by the kind permission of the Royal Society of Medicine from the Demonstra t ionum Anatomico- pathologicarum, liber prirnus by P. Camper (1760).
t e n d o n s we re m a r k e d w i t h w a t e r - i n s o l u b l e m a r k e r s : t h e a p e x o f t h e b i f u r c a t i o n o f t h e F D S ( V a ) , a p o i n t o n F D S 1 c m d i s t a l t o Va, a n d t h e p r o x i m a l s t a r t o f t h e g r o o v e o r a r a n d o m p o i n t o n t h e F D P .
To o b t a i n t h e m e a s u r e m e n t s t h e j o i n t s w e r e f ixed in d i f f e r e n t a n g l e s ( T a b l e 1) b y m e a n s o f K - w i r e s w h i c h we re p l a c e d i n t r a a r t i c u l a r l y . T h e m e a s u r e m e n t s w e r e m a d e in t h e d i f f e r e n t p o s i t i o n s in o r d e r to b e ab l e to d r a w c o n c l u s i o n s o n t h e m o v e m e n t o f b o t h t e n d o n s
Table l - -Angles of NIP, PIP and DIP (degrees) in which measurements were performed
Finger-positions ° Flexion ° Flexion ° Flexion in M P in P I P in D I P
Picture of the inside of the flexor tendon sheath showing the different pulleys (this is clearly visible in this so called "blue hand", where the pulleys are less vascularized and therefore colour less). Top needles show the C pulleys, bottom needles show the A Pulleys. The left hand side of the picture is proximal.
00/00/00 0 o 0 o o o 00/00/90 0 ° 0 ° 90 ° 00/90/00 0 ° 90 ° 0 ° 00/90/90 0 ° 90 ° 90 ° 90/90/90 90 ° 90 ° 90 ° 45/45/45 45 ° 45 ° 45 °
272
relative to one another and to the A2 pulley. The 45/45/45 configuration was included as representation of the pinch-grip.
Sufficient force was applied to the tendons proximal to the flexor retinaculum to take up the slack. To obtain more individual freedom of the involved tendons a flexor synovectomy was performed in the carpal tunnel, leaving the flexor retinaculum intact. In order to perform the measurements of Camper's chiasma, FDP was cut at DIP level and withdrawn in a proximal direction and the A3 and A4 pulleys were severed at the fibro-osseous junction.
In all, 34 fingers were dissected and measured. Two tendon pairs within their intact flexor tendon sheath were excised (and excluded from the measurements) and fixed in Karnovsky fixative. They were prepared for Scanning Electron Microscopy (SEM) to demonstrate the inner surface of the A2 pulley, particularly the DEA2, the volar surface of the FDP tendon, which is in contact with the DEA2 and the inner surface of the two FDS tendon slips, the inner surfaces of the two FDS tendon slips, the volar surface of Camper's chi- asma, which is in contact with the dorsal surface of the FDP tendon, and the dorsal surface of the FDP tendon.
One FDP tendon was obtained during operation on a patient who sustained a ring avulsion injury to his ring finger of his dominant left hand. The tendon was intact at its insertion and at the level of the injury and avulsed at the musculo-tendinous junction. It was cut by means of a microtome and used for cross-sectional area measurements. The slices were drawn on paper by means of a camera lucida. These were cut out and weighed relative to one another.
RESULTS
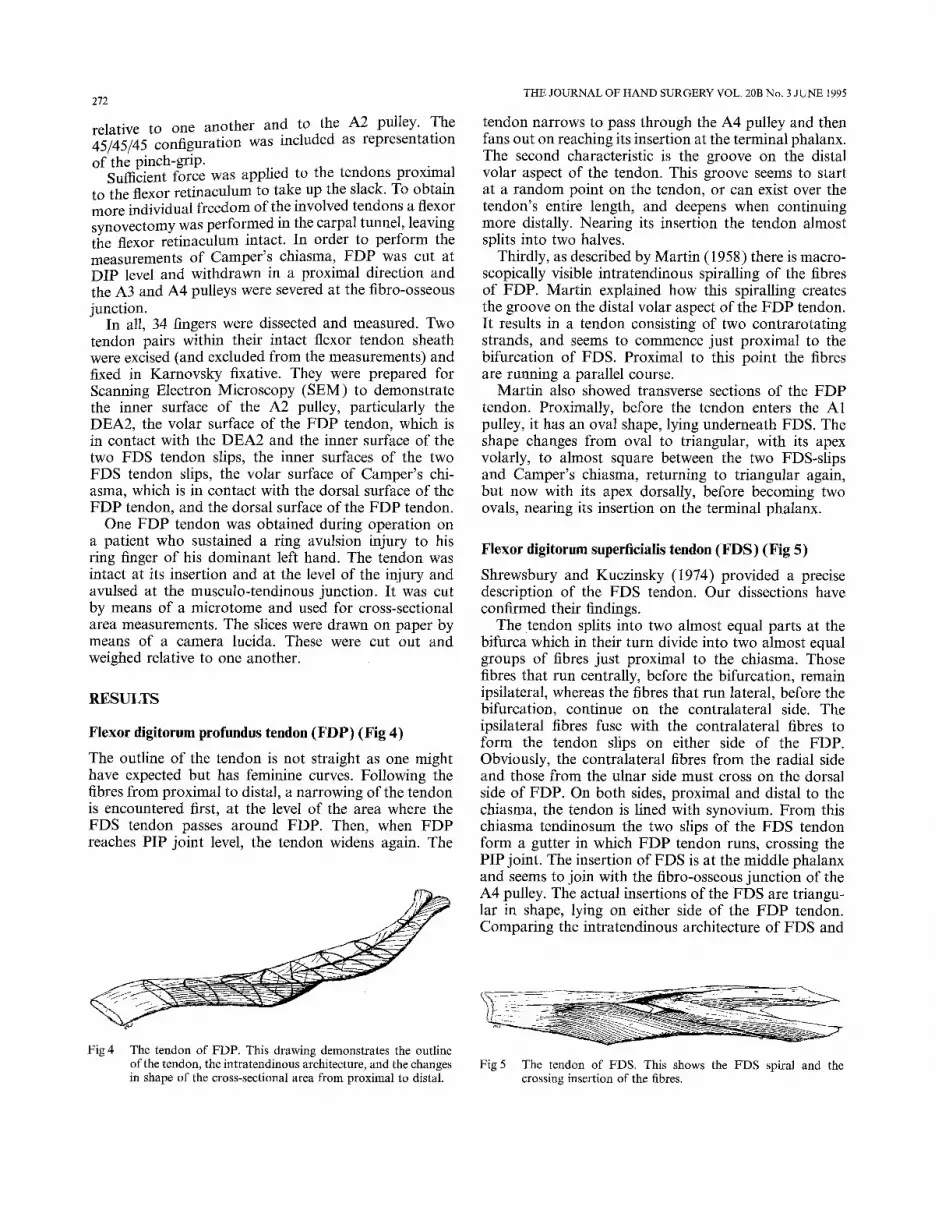
Flexor digitorum profundus tendon (FDP) (Fig 4)
The outline of the tendon is not straight as one might have expected but has feminine curves. Following the fibres from proximal to distal, a narrowing of the tendon is encountered first, at the level of the area where the FDS tendon passes around FDP. Then, when F D P reaches PIP joint level, the tendon widens again. The
Fig 4 The tendon of FDP. This drawing demonstrates the outline of the tendon, the intratendinous architecture, and the changes in shape of the cross-sectional area from proximal to distal.
THE JOURNAL OF HAND SURGERY VOL. 20B No. 3 JUNE 1995
tendon narrows to pass through the A4 pulley and then fans out on reaching its insertion at the terminal phalanx. The second characteristic is the groove on the distal volar aspect of the tendon. This groove seems to start at a random point on the tendon, or can exist over the tendon's entire length, and deepens when continuing more distally. Nearing its insertion the tendon almost splits into two halves.
Thirdly, as described by Martin (1958) there is macro- scopically visible intratendinous spiralling of the fibres of FDP. Martin explained how this spiralling creates the groove on the distal volar aspect of the F DP tendon. It results in a tendon consisting of two contrarotating strands, and seems to commence just proximal to the bifurcation of FDS. Proximal to this point the fibres are running a parallel course.
Martin also showed transverse sections of the FDP tendon. Proximally, before the tendon enters the A1 pulley, it has an oval shape, lying underneath FDS. The shape changes from oval to triangular, with its apex volarly, to almost square between the two FDS-slips and Camper's chiasma, returning to triangular again, but now with its apex dorsally, before becoming two ovals, nearing its insertion on the terminal phalanx.
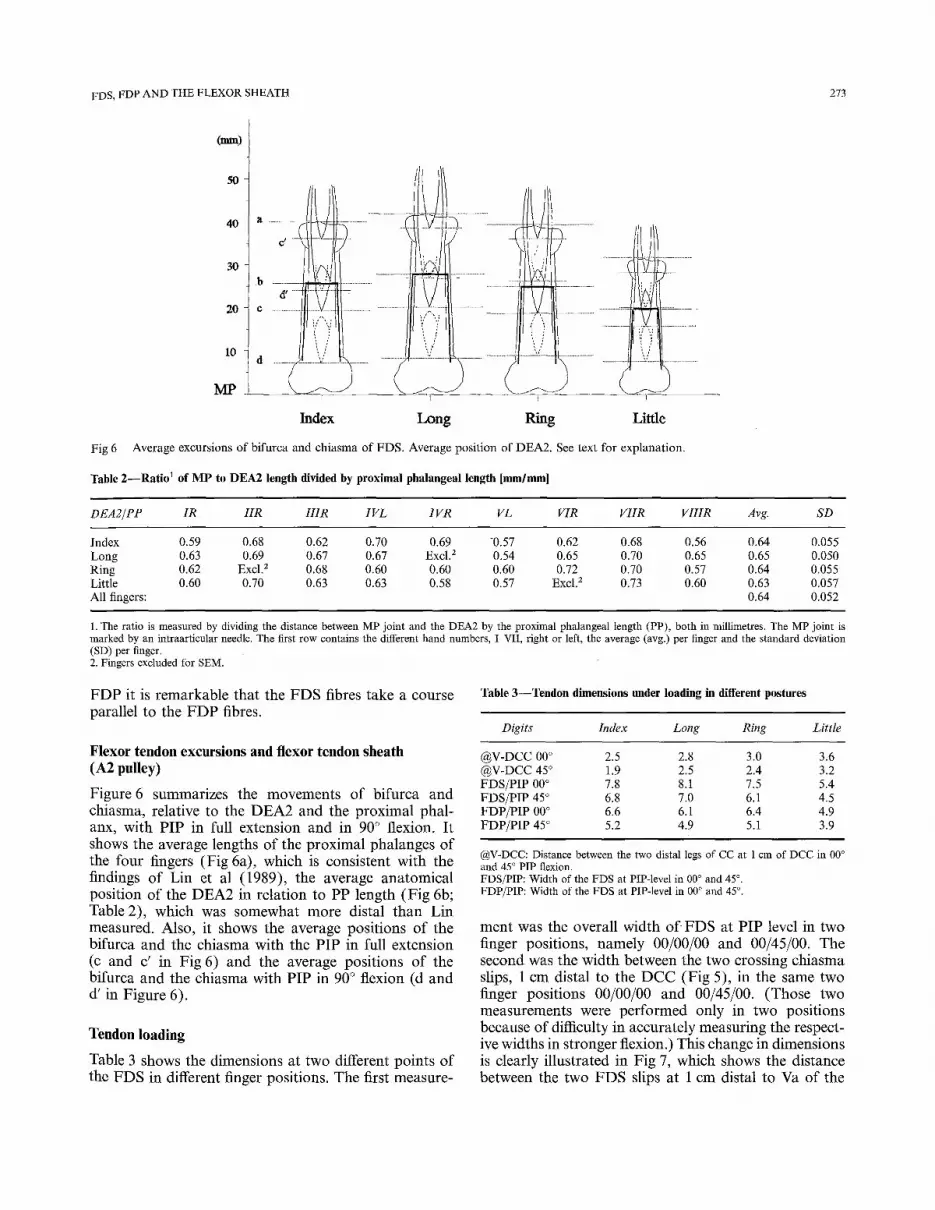
Flexor digitorum superfieialis tendon (FDS) (Fig 5)
Shrewsbury and Kuczinsky (1974) provided a precise description of the FDS tendon. Our dissections have confirmed their findings.
The tendon splits into two almost equal parts at the bifurca which in their turn divide into two almost equal groups of fibres just proximal to the chiasma. Those fibres that run centrally, before the bifurcation, remain ipsilateral, whereas the fibres that run lateral, before the bifurcation, continue on the contralateral side. The ipsilateral fibres fuse with the contralateral fibres to form the tendon slips on either side of the FDP. Obviously, the contralateral fibres from the radial side and those from the ulnar side must cross on the dorsal side of FDP. On both sides, proximal and distal to the chiasma, the tendon is lined with synovium. From this chiasma tendinosum the two slips of the FDS tendon form a gutter in which FDP tendon runs, crossing the PIP joint. The insertion of FDS is at the middle phalanx and seems to join with the fibro-osseous junction of the A4 pulley. The actual insertions of the FDS are triangu- lar in shape, lying on either side of the F D P tendon. Comparing the intratendinous architecture of FDS and
Fig 5 The tendon of FDS. This shows the FDS spiral and the crossing insertion of the fibres.
FDS, FDP AND THE FLEXOR SHEATH 273
(~ )
5O
40
30
20
10
MP
iili ! ill .... i
i:tl k
I
ti ,
t~ .................
I ............... i r
y
A; Index Long Ring Little
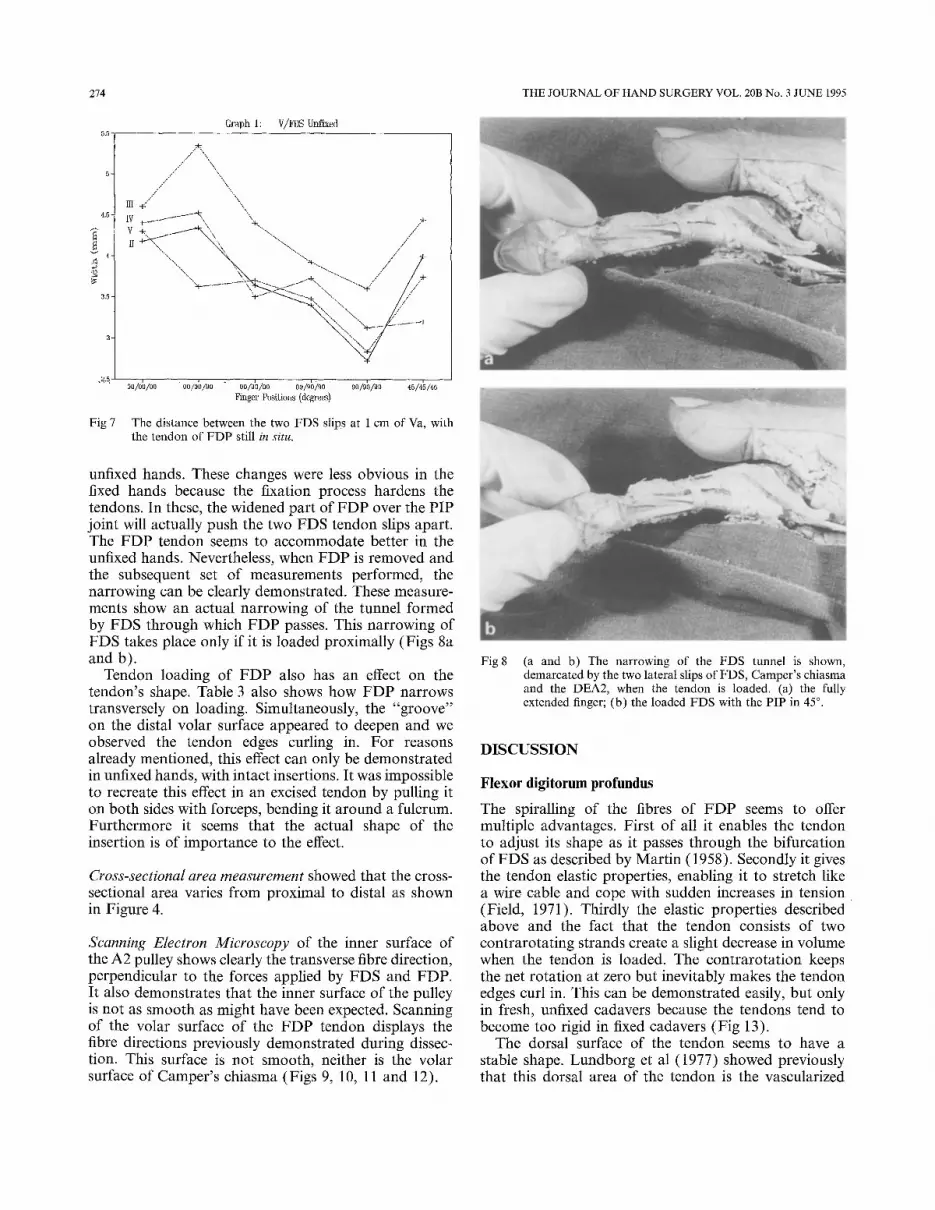
Fig 6 Average excursions of bifurca and chiasma of FDS. Average position of DEA2. See text for explanation.
Table 2--Ratio ~ of M P to DEA2 length divided by proximal phalangeal length [mm/mml
DEA2/PP IR IIR IIIR IV L IVR VL VIR VIIR VIIIR Avg. SD
Index 0.59 0.68 0.62 0.70 0.69 -0.57 0.62 0.68 0.56 0.64 0.055 Long 0.63 0.69 0.67 0.67 Excl. z 0.54 0.65 0.70 0.65 0.65 0.050 Ring 0.62 Excl, z 0.68 0.60 0.60 0.60 0.72 0.70 0.57 0.64 0.055 Little 0.60 0.70 0.63 0.63 0.58 0.57 Excl. 2 0.73 0.60 0.63 0.057 All fingers: 0.64 0.052
1. The ratio is measured by dividing the distance between MP joint and the DEA2 by the proximal phalangeal length (PP), both in millimetres. The MP joint is marked by an intraarticular needle. The first row contains the different hand numbers, I-VII, right or left, the average (avg.) per finger and the standard deviation (SD) per finger. 2. Fingers excluded for SEM.
FDP it is remarkable that the FDS fibres take a course parallel to the FDP fibres.
Flexor tendon excursions and flexor tendon sheath (A2 pulley)
Figure 6 summarizes the movements of bifurca and chiasma, relative to the DEA2 and the proximal phal- anx, with PIP in full extension and in 90 ° flexion. It shows the average lengths of the proximal phalanges of the four fingers (Fig 6a), which is consistent with the findings of Lin et al (1989), the average anatomical position of the DEA2 in relation to PP length (Fig 6b; Table 2), which was somewhat more distal than Lin measured. Also, it shows the average positions of the bifurca and the chiasma with the PIP in full extension (c and c' in Fig 6) and the average positions of the bifurca and the chiasma with PIP in 90 ° flexion (d and d' in Figure 6).
Tendon loading
Table 3 shows the dimensions at two different points of the FDS in different finger positions. The first measure-
Table 3--Tendon dimensions under loading in different postures
Digits Index Long Ring Little
@V-DCC 00 ° 2.5 2.8 3.0 3.6 @V-DCC 45 ° 1.9 2.5 2.4 3.2 FDS/PIP 00 ° 7.8 8.1 7.5 5.4 FDS/PIP 45 ° 6.8 7.0 6.1 4.5 FDP/PIP 00 ° 6.6 6.1 6.4 4.9 FDP/PIP 45 ° 5.2 4.9 5.1 3.9
@V-DCC: Distance between the two distal legs of CC at 1 cm of DCC in 00 ° and 45 ° PIP flexion. FDS/PIP: Width of the FDS at PIP-level in 00 ° and 45 °. FDP/PIP: Width of the FDS at PIP-level in 00 ° and 45 °.
ment was the overall width of FDS at PIP level in two finger positions, namely 00/00/00 and 00/45/00. The second was the width between the two crossing chiasma slips, 1 cm distal to the DCC (Fig 5), in the same two finger positions 00/00/00 and 00/45/00. (Those two measurements were performed only in two positions because of difficulty in accurately measuring the respect- ive widths in stronger flexion.) This change in dimensions is clearly illustrated in Fig 7, which shows the distance between the two FDS slips at 1 cm distal to Va of the
274 THE JOURNAL OF HAND SURGERY VOL. 20B No. 3 JUNE 1995
Graph 1: V/FDS UlffLxed
4 . 5 ¸
3 , 5 -
Fig 7
/ / / / / / " / \'\'\.\\,
lII~" \ IV+.._ ...... -, "~, .,+ V +, ',, .... -, /
",,. 2L.~ . . . . % " . . . . . . . . . . . . "
÷ ............... . ........ <5 / , , ..... ~- /
oo/o~)/oo oo/o~/9o oo/o'o/oo o~/9~)/9o 90/9h/~o ~5/4~/46 Fhlger PosiLions (degrees)
The distance between the two FDS slips at 1 cm of Va, with the tendon of FDP still in situ.
unfixed hands. These changes were less obvious in the fixed hands because the fixation process hardens the tendons. In these, the widened part of FDP over the PIP joint will actually push the two FDS tendon slips apart. The FDP tendon seems to accommodate better in the unfixed hands. Nevertheless, when FDP is removed and the subsequent set of measurements performed, the narrowing can be clearly demonstrated. These measure- ments show an actual narrowing of the tunnel formed by FDS through which FDP passes. This narrowing of FDS takes place only if it is loaded proximally (Figs 8a and b).
Tendon loading of FDP also has an effect on the tendon's shape. Table 3 also shows how FDP narrows transversely on loading. Simultaneously, the "groove" on the distal volar surface appeared to deepen and we observed the tendon edges curling in. For reasons already mentioned, this effect can only be demonstrated in unfixed hands, with intact insertions. It was impossible to recreate this effect in an excised tendon by pulling it on both sides with forceps, bending it around a fulcrum. Furthermore it seems that the actual shape of the insertion is of importance to the effect.
Cross-sectional area measurement showed that the cross- sectional area varies from proximal to distal as shown in Figure 4.
Scanning Electron Microscopy of the inner surface of the A2 pulley shows clearly the transverse fibre direction, perpendicular to the forces applied by FDS and FDP. It also demonstrates that the inner surface of the pulley is not as smooth as might have been expected. Scanning of the volar surface of the FDP tendon displays the fibre directions previously demonstrated during dissec- tion. This surface is not smooth, neither is the volar surface of Camper's chiasma (Figs 9, 10, 11 and 12).
Fig 8 (a and b) The narrowing of the FDS tunnel is shown, demarcated by the two lateral slips of FDS, Camper's chiasma and the DEA2, when the tendon is loaded. (a) the fully extended finger; (b) the loaded FDS with the PIP in 45 °.
DISCUSSION
Flexor digitorum profundus
The spiralling of the fibres of FDP seems to offer multiple advantages. First of all it enables the tendon to adjust its shape as it passes through the bifurcation of FDS as described by Martin (1958). Secondly it gives the tendon elastic properties, enabling it to stretch like a wire cable and cope with sudden increases in tension (Field, 1971). Thirdly the elastic properties described above and the fact that the tendon consists of two contrarotating strands create a slight decrease in volume when the tendon is loaded. The contrarotation keeps the net rotation at zero but inevitably makes the tendon edges curl in. This can be demonstrated easily, but only in fresh, unfixed cadavers because the tendons tend to become too rigid in fixed cadavers (Fig 13).
The dorsal surface of the tendon seems to have a stable shape. Lundborg et al (1977) showed previously that this dorsal area of the tendon is the vascularized

FDS, FDP AND THE FLEXOR SHEATH 275
Fig 9 SEM performed on the inner surface of the A2 pulley. The fibres shown are perpendicular to the direction of movement of the tendons.
part and that the volar surface is almost devoid of vascularization for nearly two-thirds of its cross- sectional surface, and is only vascularized by sparse vertical vessel loops. The spiral arrangement of fibres might be part of the regulatory mechanism for nutrition by diffusion of this area (Manske and Lester, 1982), by simply loading and unloading the tendon.
Another advantage is that the fibre direction on the dorsal surface in the curled tendon resembles the surface of a modern rain-t~cre. When the tendon tenses and is pulled against the DEA2 with great force, synovial fluid between the two surfaces may be released to the sides by the fibre pattern, thereby creating more friction. Additionally, the SEM showed the fibre direction of the inside of the A2 pulley. The angles between the direction of the fibres of FDP and those of the A2 clearly demonstrate a preferential direction for friction on the contact areas: in flexion movements the friction will be less. The moment the system becomes static, e.g. in the sustained or power grip, the direction of the friction turns 180 ° , and will increase because of the preference of the system.
Furthermore, because the fibres spiral in this way, the FDP tendon also creates two different surface planes, one, running from the palmar aspect proximally to the dorsal side of the tendon distally, and the other distal to the bifurcation of the FDS, also coming from the palmar side and crossing to the dorsal side (Fig 4).
Flexor digitorum superficialis
There have been various concepts of the function of the bifurcation of the FDS and of the interactions between the two tendons in the fingers. One of the oldest recorded ideas is that the bifurca forms a pulley-like sling to constrain the profundus tendon to the proximal phalanx (Weitbrecht, 1742; Wood Jones, 1946). Figure 6, how- ever, clearly shows that in the flexed position (PIP 90 ° ) the apex of the bifurca is well under the A2 pulley, almost at the level of the MP joint, and will not be able to keep the tendon of FDP closely applied to the proximal phalanx. The sling function seems mainly due to the A2 pulley (more specifically the DEA2).
Field (1971) and Rank et al (1973) suggested that

276 THE JOURNAL OF HAND SURGERY VOL. 20B No. 3 JUNE 1995
Fig 10 SEM performed on the volar surface of the tendon of FDP. It shows the direction of the fibres, as was confirmed by dissection.
the intratendinous architecture of FDS deals with its force-loading in such a way that the two slips and the chiasma form a perfect tunnel through which FDP glides frictionlessly. This seems to be the most widely-held belief amongst hand surgeons. However, when the FDS is loaded, the two slips inevitably come closer together. Shrewsbury and Kuczinsky (1974) made the unproven proposition that there actually may be some other function, in contradiction to the Field hypothesis. They noted that there may be some actual "gripping", a function they accredited to the bifurcation of the FDS.
Camper (1760) described the decussation of the two contralateral slips of the FDS on the dorsal side of the FDP. So this crossing, and not the bifurcation as some authors have described, is the actual chiasma tendino- sum of Camper. (Albinus had previously described it in 1734; Figs 1 and 2.) In the article of Shrewsbury and Kuczinsky (1974), the two dorsal contralateral slips of CC were drawn as lying on top of each other but they added later that the slips sometimes interweave during the crossing. In this they form a woven mat. Furthermore, looking at this architectural construction, the FDS decussation seems to resemble a so-called
"Chinese finger trap", which is wide when put over a finger, but immediately tightens on the "trapped" finger if pulled from the distal end: the trap has to be pushed off from proximally. The FDS-trap does not grip the FDP to a degree that prevents motion. The precise benefits of this mechanism are not clear, although a role in tendon nutrition and the circulation of synovial fluid are possibilities. Increased frictional resistance between the tendons and sheath may be an important normal functional mechanism. Further research of those fric- tional interactions seems important and could be essen- tial for diseases like rheumatoid arthritis (Neurath and Stofft, 1992).
Findings by Arbuckle and McGrouther (1995) suggest that there is a limiting mechanism of FDP excursion towards the end of the arc of digital motion which may be due to a relative increase in friction. Following repair a "gripping" tendency of the FDP by FDS might be an important factor in disruption (Rank et al, 1973). Tendon nutrition may also be impaired, not only by the repair itself, but also by dysfunction of the tendon compression mechanism.
The concept of a tendon compression mechanism can

FDS, FDP AND THE FLEXOR SHEATH 277
Fig 11 Detail from Figure 10.
be considered from the point of view of comparative anatomy (Sonntag, 1924; Lewis, 1989).
Tendon locking mechanisms (TLM) in nature
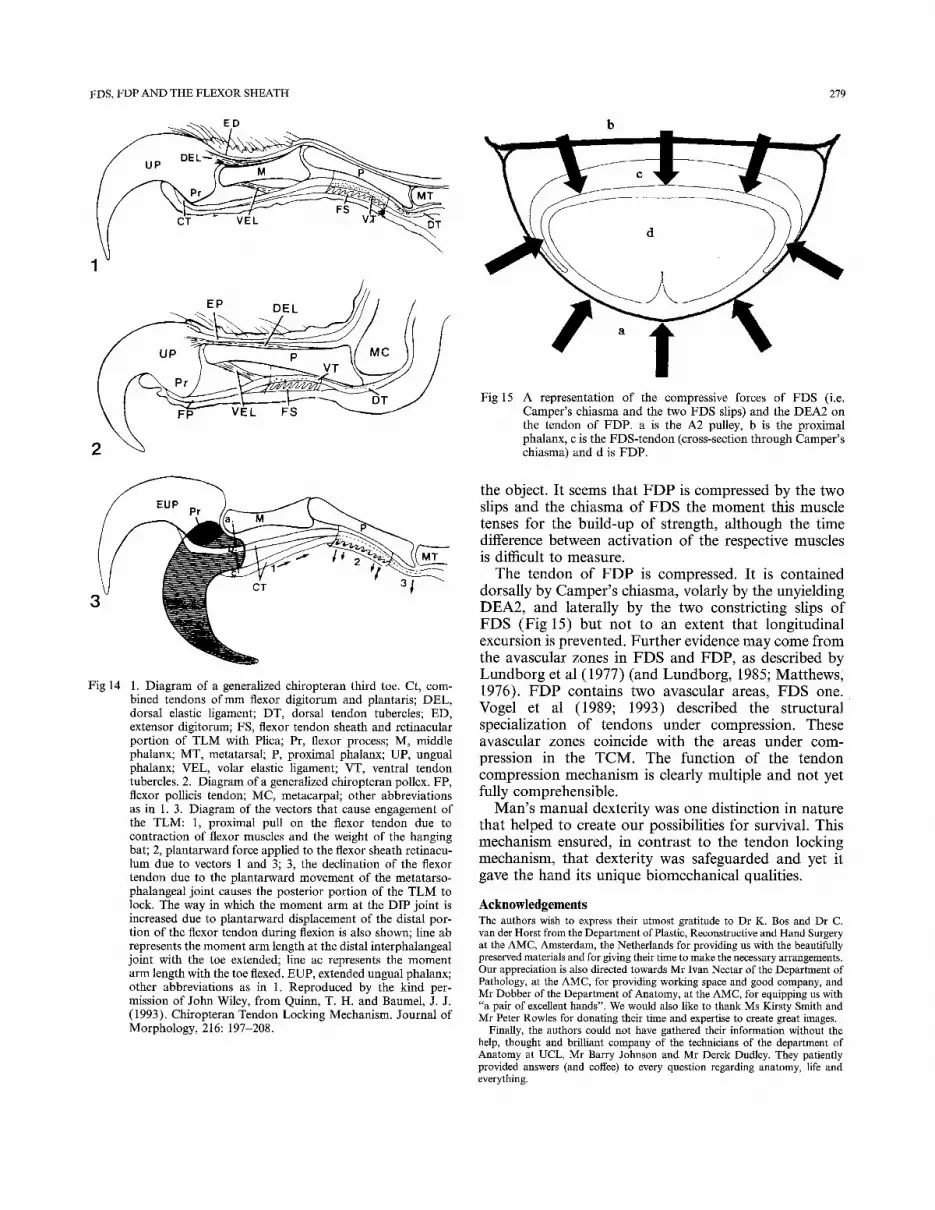
Quinn and Baumel described, in two different species, namely the bird in 1990 and the bat in 1993, a tendon locking mechanism (TLM). This mechanism is capable of sustained flexion of the digits which is independent of continuous muscular contraction. The anatomy of the bat's foot is of mammalian configuration (Fig 14). Most bats have digits consisting of three phalanges and a potlex (or hallux) which has only two. The bat has a conjoint tendon (CT), instead of FDS and FDP, con- sisting of the flexor digitorum and plantaris muscles for digital flexion, and an extensor tendon. Both tendons are inserted into the terminal phalanx called ungual phalanx (UP; better known as the claw). The CT runs through a flexor tendon sheath (FTS) which contains an unyielding fibro-osseous part, situated on the proxi- mal phalanx (PP), which has striking similarities to the human A2 pulley (for clarity this part will be referred
to as the A2 pulley). It is different in that there are two elastic ligaments, the dorsal elastic ligament (DEL) and the volar elastic ligament (VEL). In bats the TLM consists of the A2 pulley of the FTS and a tuberculated area on the corresponding plantar side of the conjoint tendon. The A2 pulley has a series of macroscopically visible transverse plicae on it's inner surface. The TLM in bats is activated by flexing the toe. As in man, as the tendon flexes, it is drawn into the FTS away from the bone. The direction of the plicae will lock the CT to the A2 pulley and the flexor musculature can relax while the digit will remain flexed. Unlocking is achieved by an upward shift of the bodyweight, passive recoil of the dorsal and volar elastic ligaments, and proximal traction on the extensor tendon. A form of TLM is also found in birds (Quinn and Baumel, 1990)
The functions of the TLM relate to swimming (form- ing a cupshaped paddle), perching, grasping and hanging upside down during hibernation without loss of meta- bolic energy. The existence of two analogous solutions to similar problems in totally different species is an example of evolutionary convergence.

278 THE JOURNAL OF HAND SURGERY VOL. 20B No. 3 JUNE 1995
Fig 12 SEM of the volar surface of Camper's chiasma.
c
DISTAL
c
Proximal Fig 13 Tendon loading effect on FDP. (a) Unloaded FDP tendon.
(b) Loaded FDP tendon. (c) Stable vascularized area.
Analogy with the human hand
In the human hand there is a difference between forces in the unloaded finger and the forces during sustained and power grip. In the unloaded finger FDP seems to be the more dominant muscle, and activity of FDS varies between subjects and even within one hand (Long, 1968; Backhouse, 1968; Long et al, 1970; Simmons and de la Caffini~re, 1981; Valentin 1981). In sustained grip and power grip the forces are distributed equally over
the long finger flexors. This grip is mostly used with the PIP joint flexed, often to 90 ° or more (hooked fist). Whether or not the MP joint is flexed (closed fist) is of no importance. McGrouther and Ahmed (1981) found no excursion of the two tendons relative to each other or to the A2 pulley during flexion of the MP joint. Our measurements show two important findings in both hook grip and closed fist. The position of the bifurca with the PIP in 90 ° is proximal, under the A2-pulley, nearing the MP joint. Camper's chiasm is at the level of the distal edge of the A2 pulley. Also, the two slips of the FDS come closer together during active flexion or stabilization of the PIP joint, i.e. during tendon loading in order to move or stabilize it. With the PIP joint, but not necessarily the DIP joint, in flexion, FDP will exert force during tendon loading on the distal edge of the A2 pulley and vice versa. The position of Camper's chiasma is exactly dorsal to the DEA2. If FDS is loaded it will apply force on the dorsal surface of the tendon of FDP, pushing its volar surface more firmly against the non-smooth interior of the pulley. The force between the tendon and the A2 pulley may be augmented by external force from the object gripped. In power grip, FDP, together with the intrinsics, adapts the finger to
FDS, FDP AND THE FLEXOR SHEATH 279
L P~~ MT \
I
EP DEL
2
Fig 14 1. Diagram of a generalized chiropteran third toe. Ct, com- bined tendons o f m m flexor digitorum and plantaris; DEL, dorsal elastic ligament; DT, dorsal tendon tubercles; ED, extensor digitorum; FS, flexor tendon sheath and retinacular portion of T L M with Plica; Pr, flexor process; M, middle phalanx; MT, metatarsal; P, proximal phalanx; UP, ungual phalanx; VEL, volar elastic ligament; VT, ventral tendon tubercles. 2. Diagram of a generalized chiropteran pollex. FP, flexor pollicis tendon; MC, metacarpal; other abbreviations as in 1. 3. Diagram of the vectors that cause engagement of the TLM: 1, proximal pull on the flexor tendon due to contraction of flexor muscles and the weight of the hanging bat; 2, plantarward force applied to the flexor sheath retinacu- lum due to vectors 1 and 3; 3, the declination of the flexor tendon due to the plantarward movement of the metatarso- phalangeal joint causes the posterior portion of the T L M to lock. The way in which the momen t arm at the DIP joint is increased due to plantarward displacement of the distal por- tion of the flexor tendon during flexion is also shown; line ab represents the moment arm length at the distal interphalangeal joint with the toe extended; line ac represents the momen t arm length with the toe flexed. EUP, extended ungual phalanx; other abbreviations as in 1. Reproduced by the kind per- mission of John Wiley, f rom Quinn, T. H. and Baumel, J. J. (1993). Chiropteran Tendon Locking Mechanism. Journal of Morphology, 216: 197-208.
Fig 15
II A representation of the compressive forces of FDS (i.e. Camper ' s chiasma and the two FDS slips) and the DEA2 on the tendon of FDP. a is the A2 pulley, b is the proximal phalanx, c is the FDS-tendon (cross-section through Camper 's chiasma) and d is FDP.
the object. It seems that FDP is compressed by the two slips and the chiasma of FDS the moment this muscle tenses for the build-up of strength, although the time difference between activation of the respective muscles is difficult to measure.
The tendon of FDP is compressed. It is contained dorsally by Camper's chiasma, volarly by the unyielding DEA2, and laterally by the two constricting slips of FDS (Fig 15) but not to an extent that longitudinal excursion is prevented. Further evidence may come from the avascular zones in FDS and FDP, as described by Lundborg et al (1977) (and Lundborg, 1985; Matthews; 1976). FDP contains two avascular areas, FDS one. Vogel et al (1989; 1993) described the structural specialization of tendons under compression. These avascular zones coincide with the areas under com- pression in the TCM. The function of the tendon compression mechanism is clearly multiple and not yet fully comprehensible.
Man's manual dexterity was one distinction in nature that helped to create our possibilities for survival. This mechanism ensured, in contrast to the tendon locking mechanism, that dexterity was safeguarded and yet it gave the hand its unique biomechanical qualities.
Acknowledgements The authors wish to express their utmost gratitude to Dr K. Bos and Dr C. van der Horst from the Department of Plastic, Reconstructive and Hand Surgery at the AMC, Amsterdam, the Netherlands for providing us with the beautifully preserved materials and for giving their time to make the necessary arrangements. Our appreciation is also directed towards Mr Ivan Nectar of the Department of Pathology, at the AMC, for providing working space and good company, and Mr Dobber of the Department of Anatomy, at the AMC, for equipping us with "a pair of excellent hands". We would also like to thank Ms Kirsty Smith and Mr Peter Rowles for donating their time and expertise to create great images.
Finally, the authors could not have gathered their information without the help, thought and brilliant company of the technicians of the department of Anatomy at UCL, Mr Barry Johnson and Mr Derek Dudley. They patiently provided answers (and coffee) to every question regarding anatomy, life and everything.

280 THE JOURNAL OF HAND SURGERY VOL. 20B No. 3 JUNE 1995
A P P E N D I X
List of Abbreviations CC C a m p e r ' s ch ia sm CSA Cross-sec t ional a rea CT Con jo in t t endon D C C Dis ta l po in t o f C a m p e r ' s eh iasm D E A 2 The d is ta l edge of the A2-Pul ley D I P Dis ta l i n t e rpha langea l j o in t F D P Flexor d ig i t o rum p ro fundus F D S Flexor d i g i t o r u m superficial is FTS F lexor t endon shea th M P M e t a c a r p o p h a l a n g e a l j o in t P C C P r o x i m a l po in t o f C a m p e r ' s ch iasm P IP P r o x i m a l i n t e rpha langea l j o in t PP P r o x i m a l p h a l a n x T C M Tendon compress ion mechan i sm T L M Tendon lock ing m e c h a n i s m S E M Scanning e lect ron mic roscopy U P U n g u a l p h a l a n x Va A p e x of the " V " of the FDS-b i furca
References ALBINUS, B. S. (1734). Historia Musculormn Hominis. Leidae Batavorum. ALBINUS, B. S. (1747). Tabulae Sceleti et Musculorum Corporis Humain.
Leidae Batavorum. ARBUCKLE, J. D. and McGROUTHER, D. A. Measurement of the arc of
digital flexion and joint movement ranges. Accepted for publication in the Journal of Hand Surgery (British and European volume).
BACKHOUSE, K. M. (1968). The mechanics of normal digital control in the hand and an analysis of the ulnar drift of rheumatoid arthritis. Annals of the Royal College of Surgeons, 43: 3: 154-173.
BLANEY, S. P. A. and JOHNSON, B. (1989). Technique for reconstituting fixed cadaveric tissue. Anatomical Record, 224: 4:550 551.
BOULAS, H. J. and STRICKLAND, J. W. (1993). Strength and functional recovery following repair of flexor digitorum superficialis in zone 2. Journal of Hand Surgery, 18B: 1: 22-25.
BRAND, P. W. Clinical Mechanics of the Hand. St Louis C. V. Mosby, 1985. BRAND, P. W., BEACH, R. B. and THOMPSON, D. E. (1981). Relative
tension and potential excursion of muscles in the forearm and hand. Journal of Hand Surgery, 6: 3: 209-219.
CAMPER, P. (1760). Demonstrationum Anatomico-pathologicarum. Liber Primus. Amstelaedami.
DOYLE, J. R. (1988). Anatomy of the flexor tendon sheath and pulley system: A current review. Journal of Hand Surgery, 13A: 4: 349-350.
ENNA, C. D. and DYER, R. F. (1976). Tendon plasticity--a property appli- cable to reconstructive surgery of the hand. The Hand, 8: 2:118-124.
FIELD, P. L. (1971). Tendon fibre arrangement and blood supply. Australian and New Zealand Journal of Surgery, 40: 3: 298-302.
HUESTON, J. F. and WILSON, W. F. (1972). The aetiology of trigger finger: Explained on the basis of intratendinous architecture. The Hand, 4: 3: 257-260.
HUESTON, J. F. and WILSON, W. F. (1973). Flexor fibre protrusion through the digital fibrous flexor sheath. The Hand, 5: 1: 39-42.
IDLER, R. S. (1985). Anatomy and biomechanics of the digital flexor tendons. Hand Clinics, 1: 1:3 11.
LEWIS, O. J. Functional Morphology o f the Evolving Hand and Foot. New York, Oxford University Press, 1989.
LIN, G. T., AMADIO, P. C., AN, K. N. and COONEY W. P. (1989). Functional anatomy of the human digital flexor pulley system. Journal of Hand Surgery, 14A: 6: 949-956.
LONG, C. (1968). Intrinsic-extrinsic muscle control of the fingers:
Electromyographic studies. Journal of Bone and Joim Surgery, 50A: 5: 973-984.
LONG, C., CONRAD, P. W., HALL, E. A. and FURLER, S. L. (1970). Intrinsic-extrinsic muscle control of the hand in power grip and precision handling: An electro myographic study. Journal of Bone and Joint Surgery, 52A: 5: 853-867.
LUNDBORG, G. Flexor Tendon Nutrition and Repair. In: Jackson I. T. (Ed.): Recent Advances in Plastic Surgery, 1985, 51-64.
LUNDBORG, G., MYRHAGE, R. and RYDEVIK, B. (1977). The vasculariz- ation of human flexor tendons within the digital synovial sheath region- structural and functional aspects. Journal of Hand Surgery, 2: 6: 417-427.
MANSKE, P. R. and LESTER, P. A. (1982). Nutrient pathways of flexor ten- dons in primates. Journal of Hand Surgery, 7: 5: 436-444.
MARTIN, B. F. (1958). The tendons of flexor digitorum profundus. Journal of Anatomy, 92: 602-608.
MATTHEWS, P. (1976). The fate of isolated segments of flexor tendons within the digital sheath: A study in synovial nutrition. British Journal of Plastic Surgery, 29: 3: 216-224.
McGROUTHER, D. A. and AHMED, M. R. (1981). Flexor tendon excursions in "no-man's land". The Hand, 13: 2: 129-141.
NEURATH, M. F. and STOFFT, E. (1992). Zur Ultrastruktur der langen Beuge- und Strecksehnen der hand bei der rheumatischen Tenosynovialitis. Handchirnrgie, Mikrochirurgie und Plastische Chirurgie, 24: 3:159 164.
O'MALLEY, C. D. and SAUNDERS, J. B. de c.m Leonardo da Vinci on the Human Body. New York, Henry Schuman, 1952.
QUINN, T. H. and BAUMEL, J. J. (1990). The digital tendon locking mechan- ism of the avian foot (Aves). Zoomorphology, 109: 281-293.
QUINN, T. H. and BAUMEL, J. J. (1993). Chiropteran tendon locking mechan- ism. Journal of Morphology, 216: 197-208.
RANK, B. K., WAKEFIELD, A. R. and HUESTON, J. T. The Surgery of Repair as Applied to Hand Injuries. 4th Edn, Edinburgh, Livingstone, 1973, 34-39; 47 49.
SHREWSBURY, M. M. and KUCZYNSKI, K. (1974). Flexor digitorum super- ficialis tendon in the fingers of the human hand. The Hand, 6: 2: 121-133.
SIMMONS, B. P. and DE LA CAFFINIERE, J. Y. Physiology o f Flexion of the Fingers. In: Tubiana, R. (Ed.): The Hand, Philadelphia, W.B. Saunders, 1981, Vol 1,377-388.
SONNTAG, C. F. The Morphology and Evolution of the Apes and Man. London, Bale Sons and DanieIssou, 1924.
STRICKLAND, J. W. (1989a). Flexor tendon surgery. Part 1: Primary flexor tendon repair. Journal of Hand Surgery, 14B: 3:261 272.
STRICKLAND, J. W. (1989b). Flexor tendon surgery. Part 2: Free tendon grafts and tenolysis. Journal of Hand Surgery, 14B: 4:368 382.
TANG, J. B. (1994). Flexor tendon repair in zone 2C. Journal of Hand Surgery, 19B: 1:72 75.
VALENTIN, P. Extrinsic Muscles of the Hand and Wrist. In: Tubiana, R. (Ed.): The Hand. Philadelphia, W,B. Sannders, 1981, Vol 1,237-243.
VOGEL, K. G. and KOOB, T. J. (1989). Structural specialization in tendons under compression. International Review of Cytology, 115: 267-293.
VOGEL, K. G., O R D t G , A., POGANY, G. and OL,~H, J. (1993). Proteoglycans in the compressed region of human tibialis posterior tendon and in ligaments. Journal of Orthopaedic Research, 11: 68-77.
WEITBRECHT, J. (1742). Syudesmohigy. WILSON, W. F. and HUESTON, J. T. (1973). The intratendinous architecture
of the tendons of the flexor digitorum profundus and flexor pollicis longus. The Hand, 5: 1: 33-38.
WOOD JONES, F. The Tendons and Tendon Sheaths. In: The Principles of Anatomy as Seen in tile Hand, 2nd Edn. London, Balli6re, Tindall and Cox, 1946, 283 299.
Accepted: 1 November 1994 Efik Walbeehm, Department of Plastic and Reconstructive Surgery, LrHR Dijkzigt/Sophia, Dr Molewaterplein 40, 3015 GD Rotterdam, The Netherlands.
© 1995 The British Society for Surgery of the Hand