altered vegetative assemblage trajectories within an urban brownfield
TRANSCRIPT

lable at ScienceDirect
Environmental Pollution xxx (2011) 1e8
Contents lists avai
Environmental Pollution
journal homepage: www.elsevier .com/locate/envpol
Altered vegetative assemblage trajectories within an urban brownfield
Frank J. Gallagher a,*, Ildiko Pechmann b, Claus Holzapfel b, Jason Grabosky a
aUrban Forestry Program, Department of Ecology, Evolution and Natural Resources, Rutgers, The State University, 14 College Farm Road, New Brunswick, NJ 08901-8551, USAbDepartment of Biological Sciences, Rutgers, The State University, 195 University Avenue, Newark, NJ 07102, USA
High concentrationsof soil metals, impact the trajectory of vegetativalternate stable state.
e assemblages in an urban brownfield leading to the speculation of an
a r t i c l e i n f o
Article history:Received 13 June 2010Received in revised form2 February 2011Accepted 4 February 2011
Keywords:BrownfieldSoil metalsVegetative guildAssemblage trajectory
* Corresponding author.E-mail address: [email protected] (F.J. G
0269-7491/$ e see front matter Published by Elseviedoi:10.1016/j.envpol.2011.02.007
Please cite this article in press as: GallagherPollution (2011), doi:10.1016/j.envpol.2011.0
a b s t r a c t
Recognizing the growing importance of both structure (maintenance of biodiversity) and function(fostering natural cycles) of urban ecologies, we examine coarse scale (herbaceous, shrub and forest) betaguild trajectory in an urban brownfield. The distribution of the pioneer forest assemblage dominated byBetula populifolia Marsh. and Populus spp. could be correlated positively with total soil metal load(arsenic, cadmium, chromium, copper, lead, zinc, lead and vanadium),whereas herbaceous and shrubguilds were negatively correlated. Distinct assemblage development trajectories above and belowa critical soil metal threshold are demonstrated. In addition, we postulate that the translocation of metalsinto the plant tissue of several dominant species may provide a positive feedback loop, maintainingrelatively high concentrations of metals in the litter and soil. Therefore assembly theory, which allows forthe development of alternate stable states, may provide a better model for the establishment of resto-ration objectives on degraded urban sites.
Published by Elsevier Ltd.
1. Introduction
Many post-industrial urban landscapes present the opportunityto convert brownfields into future open green spaces. However,before such restoration initiatives can be undertaken and reasonableobjectives established, a clear understanding of urban ecology anda corresponding philosophy of restoration must be developed. Inaddition to typical competitive and facilitativebiological interactions,assemblage development on degraded sites is often subject to thefollowing three strongfilters; a) the ability of plant diaspores to reachthese isolated urban areas is reduced; b) the soil resources needed toestablish and sustain plant growth are often limited; and c) toxicsubstances within the soil often exceed the tolerance threshold ofmany plant species (Wagner, 2004). This paper focuses on the latterof these filters and the resulting implications for assemblage devel-opment and the potential for alternate stable states.
Brownfield soils often contain trace metals such as arsenic (As),cadmium (Cd), copper (Cu), zinc (Zn), lead (Pb) and others, inconcentrations above regulatory screening criteria (Dudka et al.,1996). While these elements can be adsorbed or occluded bycarbonates, organic matter, iron magnesium oxides and primary or
allagher).
r Ltd.
, F.J., et al., Altered vegetative2.007
secondary minerals their soluble fractions create an effectiveabiotic filter that “can limit the establishment, growth and/orreproduction” (Adriano, 1986; Ross, 1994), of sensitive plantspecies. Plants that respond passively to increasing soil metalconcentrations tend to allow accumulation of metals in the plantproportionate to the concentration in the external environmentuntil, some threshold is reached, after which homeostasis or failureof growth ensues (Baker, 1981). Other species exclude metal ionsfrom particular organs via active processes by sequestering metalions in metallothioneins or phytochelatins (Vesk et al., 1999;MacFarlane and Burchett, 2000). In rare cases hyperaccumulationof metal ions is achieved through highly specialized physiologicalmechanisms that concentrate certain metals.
Under optimal growing conditions, when abiotic filters do notlimit plant growth, it is probable that species interaction, specifi-cally competition, contributes significantly to vegetative assem-blage development (Lawton, 1987; Wilson and Gitay, 1995).However, the stress associated with polluted environments tendsto favor species that exhibit resistance traits over species withstrong competitive ability, relationships within these assemblagestend toward facilitation rather than competition (Wagner, 2004;Connell and Slatyer, 1977; Bertness and Callaway, 1994). There-fore under conditions of stress the corresponding vegetativeassemblages may exhibit both structural and developmentaldifferences.
assemblage trajectories within an urban brownfield, Environmental

F.J. Gallagher et al. / Environmental Pollution xxx (2011) 1e82
Ecological restrictions resulting in the presence, absence, orabundance of species can produce repeatable guild developmentalpatterns or trajectories (Wilson and Gitay, 1995). For example,Sanguer and Jetschke (2004) demonstrated that birch dominatedthe early colonization of a former uranium-mining dump. Theyargue that the early arrival of this metal-tolerant species alters thedevelopment sequence (i.e. birch e woodland vs. grass e herbassemblage) and that colonization by many other species may bedelayed or totally inhibited. Such examples provide evidence fornon-equilibrium or stochastic models of assemblage developmentwhere species transition is determined by both abiotic and bioticfilters (Hobbs and Norton, 2004).
In this paper we describe the dynamics of coarse scale (i.e. betaguild) vegetation assemblages trajectory based on a sequence ofaerial photographs of an urban brownfield known to have hetero-geneous soil metal loading (Gallagher et al., 2008a). We hypothe-sized that assemblage trajectories had been influenced, if notdirected, by soil metal contamination, which limited recruitmentand or establishment of non-tolerant species from the regionalspecies pool. We compared guild trajectories on soils with con-trasting metal loads that had been isolated from human impacts forapproximately 40 years. More specifically, we expected that betaguild trajectory would be distinctively different above and belowa previously defined critical soil metal threshold (Gallagher et al.,2008b).
In addition, it may be possible for an ecological restriction to bereinforced through a feedback loop that is capable of operating ona large spatial scale (Belyea, 2004). Such feedback systems couldresult in an alternate stable steady state that is maintained for anunusually long period of time. In the Chihuahuan Desert, derelictagricultural fields exhibit soils with slow infiltration rates. Therainfall runs off rapidly and therefore the soils do not hydrate well.This results in drier conditions that produce sparse vegetationfurther decimating the soils, which results in increased evaporationfrom the soils (Suding et al., 2004). We postulate that such a feed-back mechanism could exist in urban brownfield. Several metaltolerant plant species that exhibit passive or active uptake patternsare known to inhabit brownfields (Gallagher et al., 2008a; Frenchet al., 2006). Should the resulting detritus maintain soil metals atconcentrations that prohibit germination and/or growth of non-tolerant species the current metal induced ecological restrictioncould be reinforced.
2. Material and methods
2.1. Study area
The 251-acre (102 ha) study site is located within Liberty State Park on the westbank of Upper New York Bay at the Hudson Estuary (centered at 40�4201400 N;74�0301400W). Prior to its development by the Central Rail Road of New Jersey(CRRNJ) the site was an intertidal mud flat and salt marsh. The area was filled usingdebris from construction projects and refuse fromNewYork City and the surfacewasstabilized with cinder and ash typically used by the rail road. The industrial use ofthe site for commodity transport and storage, including coal, resulted in relativelyhigh levels of soil metals. In 1967, the CRRNJ discontinued operations leaving the siteisolated and undisturbed. Today the park consists of approximately 1100 acres, ofwhich 251 remain undeveloped, fenced with limited access, which serves as thestudy site.
2.2. Soil sampling and spatial analysis
To determine the spatial distribution of metal contamination, soil samples werecollected in triplicate from 32 sites representing the various vegetative assemblagetypes during the summer of 2005. Soil samples were collected with a hand spade orsoil borer from the depth of the greatest root concentration. Depending on thevegetation type the samples were collected between 10 and 25 cm from the surface.The sites were also examined for depth of soil above the original rail yard fill bymeasurement of visual soil layering in the cores as contamination was associatedwith rail yard activity. In general, the bottom of the root zone correlated well withthe top of the rail yard fill. The samples were examined for concentrations of arsenic
Please cite this article in press as: Gallagher, F.J., et al., Altered vegetativePollution (2011), doi:10.1016/j.envpol.2011.02.007
(As), chromium (Cr), copper (Cu), mercury (Hg), lead (Pb), vanadium (V), and zinc(Zn) using Atomic Absorption Spectroscopy (AAS). Since all the metal data werepositively skewed, 16 additional sites were sampled in 2006. In addition, to betterdefine the site border areas, five sites were added to the eastern boarder of the studyarea. Finally, at the three sites with the highest metal load, four additional sampleswere taken (one in each compass direction at 20 m). Kriged maps with least meanerror and root mean square error values were used to calculate area soil metalconcentrations. The resulting contour maps were transformed into vector data andanalyzed in ArcGIS environment. (see Gallagher et al., 2008a for details on soilsample analysis).
Individual soil datawere kriged to estimate the distributions of soil metals. Blockkriging was used to accommodate the considerable standard deviation found in thesoil metal data (Stein, 1999). Since the metal data was highly skewed it was trans-formed before performing kriging. Logarithmic (McGrath et al., 2004) and rankorder (Juang et al., 2001) transformations were used to normalize the data distri-bution and provide more stable variograms. The results were back-transformedusing the reverse function of the linear regression between the original metal dataand the ranks (Wu et al., 2006). To asses the performance of kriging on the differ-entially transformed data sets, the mean error (ME), rootmean square error (RMSE)and the coefficient of determination (r2p) were calculated and yielded acceptableresults (see Gallagher et al., 2008a for details on performance evaluation).
Since the concentration of individual soil metals did not produce significantrelationships when compared to either plant productivity or assemblage diversitythe total soil metal load (TML) was determined by calculating the summation of therank order transformation of the individual metal concentration as described byJuang et al. (2001). The spatial distribution of the data set was developed using theblock kriging method and Surfer Surface Mapping Software (Release 8.0, GoldenSoftware Inc., Colden, CO, USA), which compensates for the high standard deviation(Leenaers et al., 1990).
The resulting index provided a way to evaluate the cumulative soil metal loadusing a scale from 0e5 in 0.1 unit increments, with 5 indicating the highestcumulative soil metal concentrations. This metric was then used as a relative indexof soil contamination. A critical TML threshold value of 3.0 was used as a benchmarkfor areas of high metal load as our previous work indicated that soil metals loadabove this level had a significant sublethal impact resulting in metal-induced stresson plant productivity, long-term growth and diversity (Gallagher et al., 2008b).
2.3. Soil metal distribution
The total soil metal load (TML) was unevenly distributed (Fig. 1a) and exhibitedconsiderable local variation (Gallagher et al., 2008a). The concentration of Asexceeded the Lowest Observed Effective Concentration (LOEC, Oak Ridge, 1997) at20% and Pb at 16% of the sites. Cr exceeded the LOEC at most of the sites (80%). Theother measured soil metals exceeded the LOEC at approximately half of the sites(Table 1).
2.4. Spatial vegetation classification
The variability in substrate materials combined with the undulating topographyof the abandoned rail yard has created a patchy distribution of soils and cinders thathas influenced plant colonization over the last 40 years. A unique mosaic ofassemblage types existswithin the study site (United States ArmyCorps of Engineers,2004). The hardwood assemblage is composed primarily of Gray Birch (Betula pop-ulifolia Marsh.) (35% cover), Eastern Cottonwood (Populus deltoids Marsh.) (16%cover) and Quaking Aspen (Populus. tremuloidesMichx.) (14% cover) (Gallagher et al.,in press). Shrubs are dominated by three species of the genus Rhus; Staghorn Sumac(R. typhina L.), Smooth Sumac (R. glabra L.), and Winged Sumac (R. copallinum L.).Herb and grass assemblages are dominated by amix of several native and non-nativespecies: Goldenrod (Solidago spp.), Mugwort (Artemisia vulgaris L.), Switchgrass(Panicum virgatum L.), Chee Reedgrass (Calamagrostis epigeios L.) and Red Fescue(Festuca rubra L.) (all nomenclature after Gleason and Cronquist, 1991).
Historic true color aerial photographs from 1976, 1984, 1993, 1996, 2000, anda 1969 gray scale photograph, were scanned at 600 dpi and were geo-referenced inArcGIS using a 2002 aerial photo (released by NJDEP, spatial resolution .3 m) asreference. The rectification resulted in less than 0.6 m RSM (root square mean) errorfor each of the photographs. In delineating the assemblages, three field-verifieddigital vegetation maps were used, the first from the 2003 United States Army Corpof Engineers (USACE) Liberty State Park Environmental Assessment (USACE, 2004)based on the 2002 NJDEP aerial photo, the second from a 1996 aerial photographwhich was field verified during a park natural resource assessment (MacFarlane,1996) and the third was the 2009 field survey. In addition, an early report (TexasInstruments, 1976) was also used to verify the 1976 digital vegetation map.
The 2003 vegetation map differentiated 12 alpha guilds based on speciescomposition (Table 2). The alpha guilds were classified according to the EcologicalCommunities of New York State (Edinger et al., 2002). The identified assemblagesranged from successional northern hardwood to grasslands. Linear regressionsbetween assemblage distribution (area ratios) and total soil metal load, indicatesignificant relationships within the successional northern hardwood (r2¼ 0.62,p< 0.01) and semi-emergent marsh (r2¼ 0.49, p< 0.01) (Gallagher et al., 2008a).
assemblage trajectories within an urban brownfield, Environmental
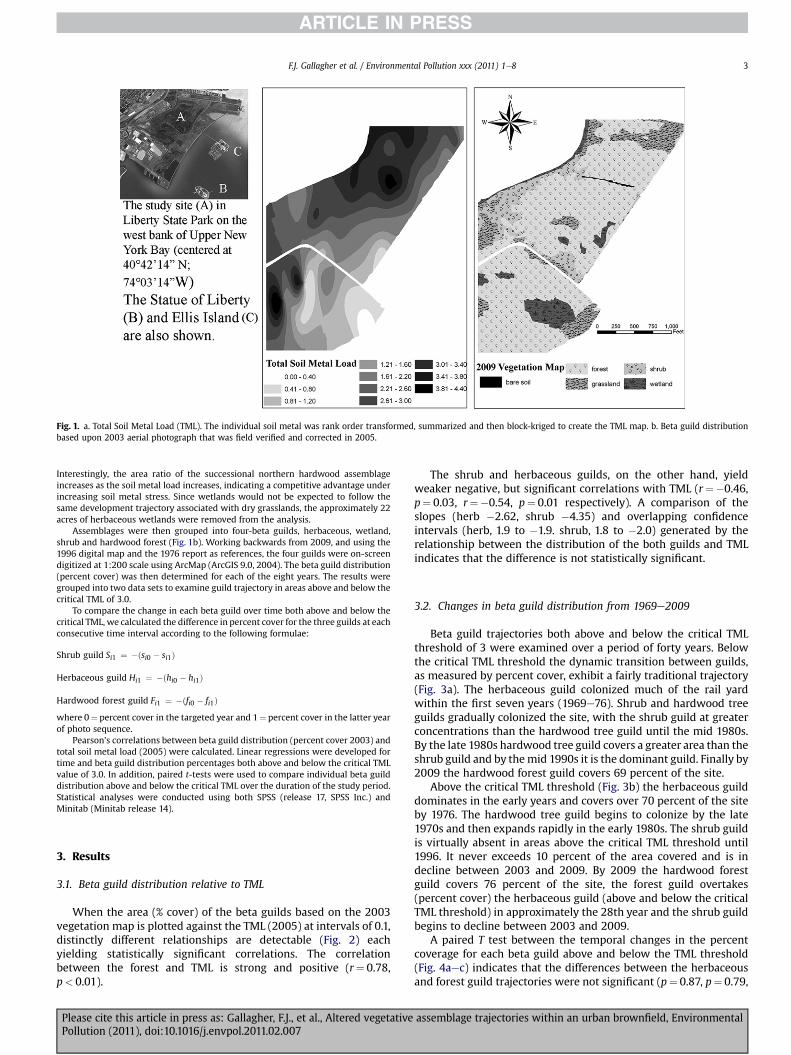
Fig. 1. a. Total Soil Metal Load (TML). The individual soil metal was rank order transformed, summarized and then block-kriged to create the TML map. b. Beta guild distributionbased upon 2003 aerial photograph that was field verified and corrected in 2005.
F.J. Gallagher et al. / Environmental Pollution xxx (2011) 1e8 3
Interestingly, the area ratio of the successional northern hardwood assemblageincreases as the soil metal load increases, indicating a competitive advantage underincreasing soil metal stress. Since wetlands would not be expected to follow thesame development trajectory associated with dry grasslands, the approximately 22acres of herbaceous wetlands were removed from the analysis.
Assemblages were then grouped into four-beta guilds, herbaceous, wetland,shrub and hardwood forest (Fig. 1b). Working backwards from 2009, and using the1996 digital map and the 1976 report as references, the four guilds were on-screendigitized at 1:200 scale using ArcMap (ArcGIS 9.0, 2004). The beta guild distribution(percent cover) was then determined for each of the eight years. The results weregrouped into two data sets to examine guild trajectory in areas above and below thecritical TML of 3.0.
To compare the change in each beta guild over time both above and below thecritical TML, we calculated the difference in percent cover for the three guilds at eachconsecutive time interval according to the following formulae:
Shrub guild Si1 ¼ �ðsi0 � si1Þ
Herbaceous guild Hi1 ¼ �ðhi0 � hi1Þ
Hardwood forest guild Fi1 ¼ �ðfi0 � fi1Þwhere 0¼ percent cover in the targeted year and 1¼ percent cover in the latter yearof photo sequence.
Pearson’s correlations between beta guild distribution (percent cover 2003) andtotal soil metal load (2005) were calculated. Linear regressions were developed fortime and beta guild distribution percentages both above and below the critical TMLvalue of 3.0. In addition, paired t-tests were used to compare individual beta guilddistribution above and below the critical TML over the duration of the study period.Statistical analyses were conducted using both SPSS (release 17, SPSS Inc.) andMinitab (Minitab release 14).
3. Results
3.1. Beta guild distribution relative to TML
When the area (% cover) of the beta guilds based on the 2003vegetation map is plotted against the TML (2005) at intervals of 0.1,distinctly different relationships are detectable (Fig. 2) eachyielding statistically significant correlations. The correlationbetween the forest and TML is strong and positive (r¼ 0.78,p< 0.01).
Please cite this article in press as: Gallagher, F.J., et al., Altered vegetativePollution (2011), doi:10.1016/j.envpol.2011.02.007
The shrub and herbaceous guilds, on the other hand, yieldweaker negative, but significant correlations with TML (r¼�0.46,p¼ 0.03, r¼�0.54, p¼ 0.01 respectively). A comparison of theslopes (herb �2.62, shrub �4.35) and overlapping confidenceintervals (herb, 1.9 to �1.9. shrub, 1.8 to �2.0) generated by therelationship between the distribution of the both guilds and TMLindicates that the difference is not statistically significant.
3.2. Changes in beta guild distribution from 1969e2009
Beta guild trajectories both above and below the critical TMLthreshold of 3 were examined over a period of forty years. Belowthe critical TML threshold the dynamic transition between guilds,as measured by percent cover, exhibit a fairly traditional trajectory(Fig. 3a). The herbaceous guild colonized much of the rail yardwithin the first seven years (1969e76). Shrub and hardwood treeguilds gradually colonized the site, with the shrub guild at greaterconcentrations than the hardwood tree guild until the mid 1980s.By the late 1980s hardwood tree guild covers a greater area than theshrub guild and by themid 1990s it is the dominant guild. Finally by2009 the hardwood forest guild covers 69 percent of the site.
Above the critical TML threshold (Fig. 3b) the herbaceous guilddominates in the early years and covers over 70 percent of the siteby 1976. The hardwood tree guild begins to colonize by the late1970s and then expands rapidly in the early 1980s. The shrub guildis virtually absent in areas above the critical TML threshold until1996. It never exceeds 10 percent of the area covered and is indecline between 2003 and 2009. By 2009 the hardwood forestguild covers 76 percent of the site, the forest guild overtakes(percent cover) the herbaceous guild (above and below the criticalTML threshold) in approximately the 28th year and the shrub guildbegins to decline between 2003 and 2009.
A paired T test between the temporal changes in the percentcoverage for each beta guild above and below the TML threshold(Fig. 4aec) indicates that the differences between the herbaceousand forest guild trajectories were not significant (p¼ 0.87, p¼ 0.79,
assemblage trajectories within an urban brownfield, Environmental

Table 1Comparison of soil metal concentrations (mg g�1 , mean� S.D.) with the Lowest Observed Effective Concentration (LOEC, OakRidge, 1997) and Maximum Acceptable ToxicConcentration (MATC, ECO-SSL, 2003). Triplicate samples were taken at one meter intervals at 35 sampling points. MDL¼Minimum Detection Limits, % above¼ area of thestudy site above ECO-SSL or LOEC, **¼No established limit.
As Cr Cu Hg Pb V Zn
MDL 0.005 <0.01 3.1 0.002 <0.01 <0.01 17.9Min <MDL 9.7� 2.5 44.0� 2.5 <MDL 86.0� 11.1 <MDL 80.0� 12.925% 16.7� 2.5 22.5� 4.8 74.0� 11.6 0.1� 0.1 185� 38.8 26.9� 15.2 93.1� 21.5Median 33.5� 6.8 38.8� 7.9 153� 27.7 0.3� 0.1 406.0� 73.6 44.0� 20.7 159.0� 48.475% 121.9� 29.8 60.2� 25.7 253.0� 58.2 0.7� 0.3 520� 181.1 76.1� 33.2 547.0� 221.8Max 977.6� 44.3 208.8� 10.4 1870.0� 315. 3.6� 6.0 4640� 1799 193.2� 112.6 6501.0� 1491% above ECO-SSL 76 ** 84 ** 88 ** **% above LOEC 20 80 48 0 16 50 44
F.J. Gallagher et al. / Environmental Pollution xxx (2011) 1e84
respectfully). The difference in the shrub guild trajectory wassignificant (p¼ 0.03).
4. Discussion
4.1. Soil metal induced stress
Under natural conditions, metals are present in soils as deriva-tives of parent geologicmaterials. Serving as amicronutrient,metalsat low concentrations are required for normal metabolic function ofboth flora and fauna. At concentrations above functional ranges, theplant can no longer maintain homeostasis and metabolic functionsare inhibited (Hillary and Wilkins, 1987). It follows then, thata species intolerant of highmetal concentrationswould be excludedfrom the assemblage on the site with high soil metal loads even ifa seed source existedwithin the regional pool. Therefore, vegetativeassemblages growing on metalliferious soil should exhibit struc-tural differences when compared to similar but uncontaminated
Table 2The dominant species within each alpha guild as classified according to EcologicalCommunities of New York State (Edinger et al., 2002). Beta guilds grouped asfollows: 1) forest, 2) shrub, 3) herb.
Northern Hardwood1 Floodplain Forest Wetland1
Populus tremuloides Michx.(Quaking Aspen)
Betula populifolia Marsh. (Grey Birch)
Populus deltoides Bartr.(Eastern Cottonwood)
Betula populifolia Marsh. (Grey Birch) Shrub Swamp Wetland2
Rubus laciniatus Willd.(Cutleaf Blackberry)
Successional Shrubland2 Rhus copallina L. (Winged Sumac)Rubus laciniatus Willd.
(Cutleaf Blackberry)Rhus typhina L. (Staghorn Sumac)
Rhus glabra L. (Smooth Sumac)Myrica pensylvanica Loisel. (Bayberry) Successional Grassland3
Festuca rubra L. (Red Fescue)Maritime Shrubland2 Panicum virgatum L. (Switchgrass)Rhus copallina L. (Winged Sumac) Calamagrostis epigeios L .
(Chee Reedgrass)Rhus glabra L. (Smooth Sumac)Rhus typhina L. (Smooth Sumac) Successional Old Field3
Calamagrostis epigeios L.(Chee Reedgrass)
Common Reed/Mugwort3 Verbascum thapsus L.(Common Mullen)
Artemisia vulgaris L. (Mugwart) Solidago canadensis L.(Canadian Goldenrod)
Phragmites australis Cav. (Common Reed)Shallow Emergent Marsh3
Common Reed-dominated Wetland3 Scirpus cyperinus L. (Wool Grass)Phragmites australis Cav. (Common Reed) Spiraea tomentosa L. (Steeplebush)Lythrum salicaria L. (Purple Loosetrife) Lythrum salicaria L.
(Purple Loosetrife)Onoclea sensibilis L. (Sensitive Fern) Phragmites australis Cav.
(Common Reed)
Please cite this article in press as: Gallagher, F.J., et al., Altered vegetativePollution (2011), doi:10.1016/j.envpol.2011.02.007
regional environments. These differences have been shown tomanifest themselves in arrested successional developments(Niering andGoodwin,1974), such as those foundonnaturallyheavymetal-rich rock outcrops (e.g. serpentine outcrops, Reynolds et al.,1997; Harrison, 1999). Kimmerer (1981) as well as Kalin and vanEverdingen (1988) demonstrated that assemblage development onmine tailings proceeded slowly, often remaining in the herbaceousstage for decades or centuries. In addition, the colonization anddistribution of plant species on mine tailings from five Pb/Zn minesin Chinawere still dominated (69.4% of total) by herbaceous speciesafter twenty years of development (Li et al., 2006).
Our study corroborates the concept that patterns of assemblagedevelopment, especially the advancement of tree species, can beimpacted if not driven by soil metal concentration. Interestingly, incontrast to the previously mentioned mine tailing studies, itappears that the hardwood assemblage at Liberty State Park hasdeveloped preferentially on the soils with increased total metalload. Sanguer and Jetschke (2004) reported similar results ona former uranium-mining disposal area (East Germany). They arguethat the early arrival of tolerant woody species alters the devel-opment sequence (i.e. birch-woodland vs. grass -herb assemblage)hence the colonization by many other species may be delayed ortotally inhibited.
This is apparently the case at Liberty State Park. An examinationof historic photos indicates that assemblage development in areaswhere the TML was high (3 or above) favored colonization of GrayBirch (B. populifolia) and Cottonwood (Populous spp.) known to bemetal tolerant species. The shrub guild dominated by three speciesof Sumac (Rhus spp.) not known to be metal tolerant does notappear until 1996 and never covers more than 10% of the area. Thestrong significant correlation between soil TML and forest distri-bution is probably the result of both metal tolerance and the
Fig. 2. Guild distribution (percent cover) of the three beta guilds, Hardwood Forest(HF) Shrub and Herbaceous (Herb) and trend lines for 2003. Pearson correlations foreach guild and total soil metal load (2005) are: hardwood forest assembly r¼ 0.78,p< 0.01, shrub assembly r¼�0.46, p¼ 0.03, herbaceous assembly r¼�0.54, p¼ 0.01.
assemblage trajectories within an urban brownfield, Environmental
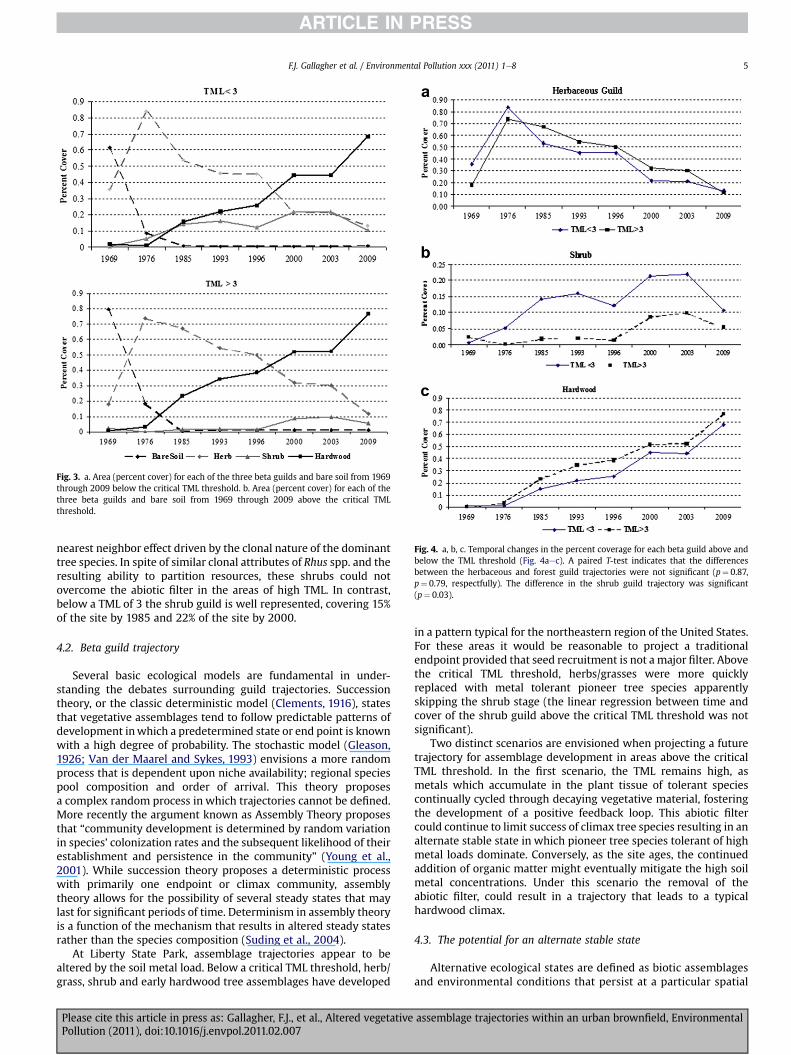
Fig. 4. a, b, c. Temporal changes in the percent coverage for each beta guild above andbelow the TML threshold (Fig. 4aec). A paired T-test indicates that the differencesbetween the herbaceous and forest guild trajectories were not significant (p¼ 0.87,p¼ 0.79, respectfully). The difference in the shrub guild trajectory was significant(p¼ 0.03).
Fig. 3. a. Area (percent cover) for each of the three beta guilds and bare soil from 1969through 2009 below the critical TML threshold. b. Area (percent cover) for each of thethree beta guilds and bare soil from 1969 through 2009 above the critical TMLthreshold.
F.J. Gallagher et al. / Environmental Pollution xxx (2011) 1e8 5
nearest neighbor effect driven by the clonal nature of the dominanttree species. In spite of similar clonal attributes of Rhus spp. and theresulting ability to partition resources, these shrubs could notovercome the abiotic filter in the areas of high TML. In contrast,below a TML of 3 the shrub guild is well represented, covering 15%of the site by 1985 and 22% of the site by 2000.
4.2. Beta guild trajectory
Several basic ecological models are fundamental in under-standing the debates surrounding guild trajectories. Successiontheory, or the classic deterministic model (Clements, 1916), statesthat vegetative assemblages tend to follow predictable patterns ofdevelopment inwhich a predetermined state or end point is knownwith a high degree of probability. The stochastic model (Gleason,1926; Van der Maarel and Sykes, 1993) envisions a more randomprocess that is dependent upon niche availability; regional speciespool composition and order of arrival. This theory proposesa complex random process in which trajectories cannot be defined.More recently the argument known as Assembly Theory proposesthat “community development is determined by random variationin species’ colonization rates and the subsequent likelihood of theirestablishment and persistence in the community” (Young et al.,2001). While succession theory proposes a deterministic processwith primarily one endpoint or climax community, assemblytheory allows for the possibility of several steady states that maylast for significant periods of time. Determinism in assembly theoryis a function of the mechanism that results in altered steady statesrather than the species composition (Suding et al., 2004).
At Liberty State Park, assemblage trajectories appear to bealtered by the soil metal load. Below a critical TML threshold, herb/grass, shrub and early hardwood tree assemblages have developed
Please cite this article in press as: Gallagher, F.J., et al., Altered vegetativePollution (2011), doi:10.1016/j.envpol.2011.02.007
in a pattern typical for the northeastern region of the United States.For these areas it would be reasonable to project a traditionalendpoint provided that seed recruitment is not a major filter. Abovethe critical TML threshold, herbs/grasses were more quicklyreplaced with metal tolerant pioneer tree species apparentlyskipping the shrub stage (the linear regression between time andcover of the shrub guild above the critical TML threshold was notsignificant).
Two distinct scenarios are envisioned when projecting a futuretrajectory for assemblage development in areas above the criticalTML threshold. In the first scenario, the TML remains high, asmetals which accumulate in the plant tissue of tolerant speciescontinually cycled through decaying vegetative material, fosteringthe development of a positive feedback loop. This abiotic filtercould continue to limit success of climax tree species resulting in analternate stable state in which pioneer tree species tolerant of highmetal loads dominate. Conversely, as the site ages, the continuedaddition of organic matter might eventually mitigate the high soilmetal concentrations. Under this scenario the removal of theabiotic filter, could result in a trajectory that leads to a typicalhardwood climax.
4.3. The potential for an alternate stable state
Alternative ecological states are defined as biotic assemblagesand environmental conditions that persist at a particular spatial
assemblage trajectories within an urban brownfield, Environmental
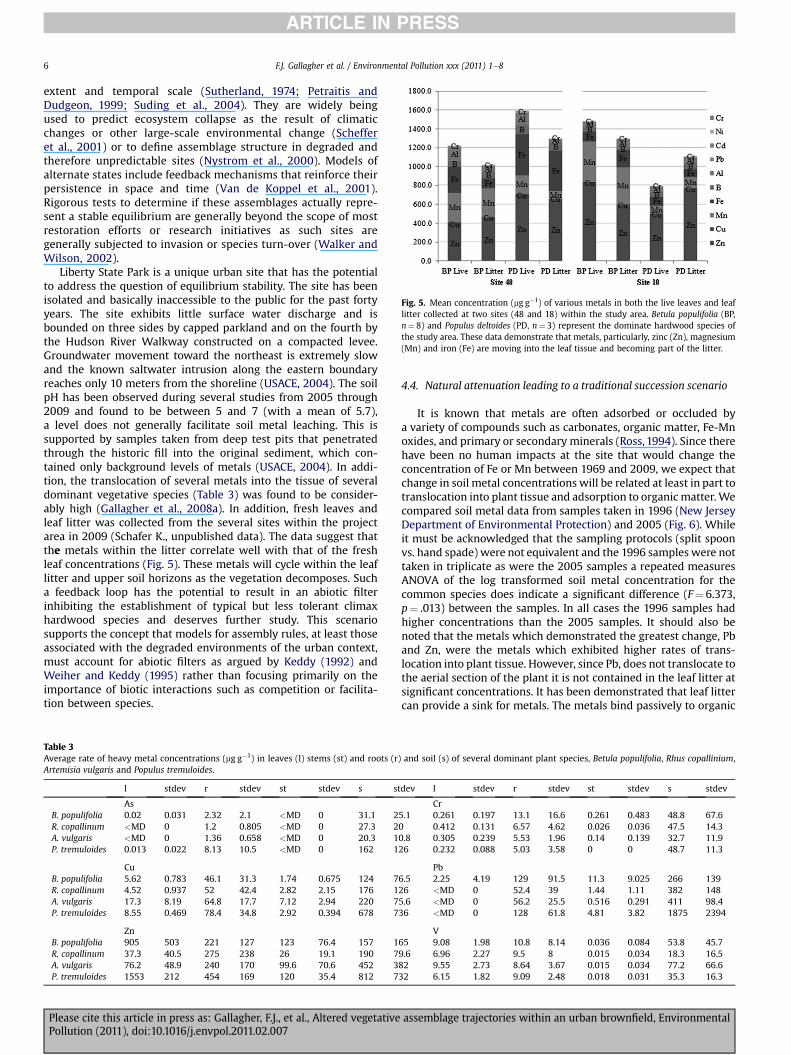
Fig. 5. Mean concentration (mg g�1) of various metals in both the live leaves and leaflitter collected at two sites (48 and 18) within the study area. Betula populifolia (BP,n¼ 8) and Populus deltoides (PD, n¼ 3) represent the dominate hardwood species ofthe study area. These data demonstrate that metals, particularly, zinc (Zn), magnesium(Mn) and iron (Fe) are moving into the leaf tissue and becoming part of the litter.
F.J. Gallagher et al. / Environmental Pollution xxx (2011) 1e86
extent and temporal scale (Sutherland, 1974; Petraitis andDudgeon, 1999; Suding et al., 2004). They are widely beingused to predict ecosystem collapse as the result of climaticchanges or other large-scale environmental change (Schefferet al., 2001) or to define assemblage structure in degraded andtherefore unpredictable sites (Nystrom et al., 2000). Models ofalternate states include feedback mechanisms that reinforce theirpersistence in space and time (Van de Koppel et al., 2001).Rigorous tests to determine if these assemblages actually repre-sent a stable equilibrium are generally beyond the scope of mostrestoration efforts or research initiatives as such sites aregenerally subjected to invasion or species turn-over (Walker andWilson, 2002).
Liberty State Park is a unique urban site that has the potentialto address the question of equilibrium stability. The site has beenisolated and basically inaccessible to the public for the past fortyyears. The site exhibits little surface water discharge and isbounded on three sides by capped parkland and on the fourth bythe Hudson River Walkway constructed on a compacted levee.Groundwater movement toward the northeast is extremely slowand the known saltwater intrusion along the eastern boundaryreaches only 10 meters from the shoreline (USACE, 2004). The soilpH has been observed during several studies from 2005 through2009 and found to be between 5 and 7 (with a mean of 5.7),a level does not generally facilitate soil metal leaching. This issupported by samples taken from deep test pits that penetratedthrough the historic fill into the original sediment, which con-tained only background levels of metals (USACE, 2004). In addi-tion, the translocation of several metals into the tissue of severaldominant vegetative species (Table 3) was found to be consider-ably high (Gallagher et al., 2008a). In addition, fresh leaves andleaf litter was collected from the several sites within the projectarea in 2009 (Schafer K., unpublished data). The data suggest thatthe metals within the litter correlate well with that of the freshleaf concentrations (Fig. 5). These metals will cycle within the leaflitter and upper soil horizons as the vegetation decomposes. Sucha feedback loop has the potential to result in an abiotic filterinhibiting the establishment of typical but less tolerant climaxhardwood species and deserves further study. This scenariosupports the concept that models for assembly rules, at least thoseassociated with the degraded environments of the urban context,must account for abiotic filters as argued by Keddy (1992) andWeiher and Keddy (1995) rather than focusing primarily on theimportance of biotic interactions such as competition or facilita-tion between species.
Table 3Average rate of heavy metal concentrations (mg g�1) in leaves (l) stems (st) and roots (rArtemisia vulgaris and Populus tremuloides.
l stdev r stdev st stdev s st
AsB. populifolia 0.02 0.031 2.32 2.1 <MD 0 31.1 2R. copallinum <MD 0 1.2 0.805 <MD 0 27.3 2A. vulgaris <MD 0 1.36 0.658 <MD 0 20.3 1P. tremuloides 0.013 0.022 8.13 10.5 <MD 0 162 1
CuB. populifolia 5.62 0.783 46.1 31.3 1.74 0.675 124 7R. copallinum 4.52 0.937 52 42.4 2.82 2.15 176 1A. vulgaris 17.3 8.19 64.8 17.7 7.12 2.94 220 7P. tremuloides 8.55 0.469 78.4 34.8 2.92 0.394 678 7
ZnB. populifolia 905 503 221 127 123 76.4 157 1R. copallinum 37.3 40.5 275 238 26 19.1 190 7A. vulgaris 76.2 48.9 240 170 99.6 70.6 452 3P. tremuloides 1553 212 454 169 120 35.4 812 7
Please cite this article in press as: Gallagher, F.J., et al., Altered vegetativePollution (2011), doi:10.1016/j.envpol.2011.02.007
4.4. Natural attenuation leading to a traditional succession scenario
It is known that metals are often adsorbed or occluded bya variety of compounds such as carbonates, organic matter, Fe-Mnoxides, and primary or secondary minerals (Ross, 1994). Since therehave been no human impacts at the site that would change theconcentration of Fe or Mn between 1969 and 2009, we expect thatchange in soil metal concentrations will be related at least in part totranslocation into plant tissue and adsorption to organic matter.Wecompared soil metal data from samples taken in 1996 (New JerseyDepartment of Environmental Protection) and 2005 (Fig. 6). Whileit must be acknowledged that the sampling protocols (split spoonvs. hand spade) were not equivalent and the 1996 samples were nottaken in triplicate as were the 2005 samples a repeated measuresANOVA of the log transformed soil metal concentration for thecommon species does indicate a significant difference (F¼ 6.373,p¼ .013) between the samples. In all cases the 1996 samples hadhigher concentrations than the 2005 samples. It should also benoted that the metals which demonstrated the greatest change, Pband Zn, were the metals which exhibited higher rates of trans-location into plant tissue. However, since Pb, does not translocate tothe aerial section of the plant it is not contained in the leaf litter atsignificant concentrations. It has been demonstrated that leaf littercan provide a sink for metals. The metals bind passively to organic
) and soil (s) of several dominant plant species, Betula populifolia, Rhus copallinium,
dev l stdev r stdev st stdev s stdev
Cr5.1 0.261 0.197 13.1 16.6 0.261 0.483 48.8 67.60 0.412 0.131 6.57 4.62 0.026 0.036 47.5 14.30.8 0.305 0.239 5.53 1.96 0.14 0.139 32.7 11.926 0.232 0.088 5.03 3.58 0 0 48.7 11.3
Pb6.5 2.25 4.19 129 91.5 11.3 9.025 266 13926 <MD 0 52.4 39 1.44 1.11 382 1485.6 <MD 0 56.2 25.5 0.516 0.291 411 98.436 <MD 0 128 61.8 4.81 3.82 1875 2394
V65 9.08 1.98 10.8 8.14 0.036 0.084 53.8 45.79.6 6.96 2.27 9.5 8 0.015 0.034 18.3 16.582 9.55 2.73 8.64 3.67 0.015 0.034 77.2 66.632 6.15 1.82 9.09 2.48 0.018 0.031 35.3 16.3
assemblage trajectories within an urban brownfield, Environmental

Fig. 6. A comparison of soil metal data (n¼ 16 for each metal species) from samplestaken in 1996 and 2005. A repeated measures ANOVA of the log transformed mean soilmetal concentration for the common species at the same sites does indicate a signifi-cant difference (F¼ 6.373, p¼ .013). In all cases the 1996 samples had higherconcentrations than the 2005 samples. Pb and Zn show the greatest differences.
F.J. Gallagher et al. / Environmental Pollution xxx (2011) 1e8 7
surfaces or actively through the physiological activity of themicrobial colonizers (Gadd, 1999; Ledin, 2000). Leaf litter can alsoact as a source when microbial activity mobilizes the metal (Gadd,1999) or through the action of deposit feeders (Weis and Weis,2004). Metal sequestration is dependent upon the rates of uptakeand retention by the various tissue types, translocation to depositfeeders and release through decomposition.
In this young terrestrial system, the continued addition oforganic material as a result of plant growth may be acting as a sink,which would reduce the pool of available soil metals. If this conceptis valid, then the strength of TML as the deterministic abiotic filtermay decrease over time leading to the establishment of specieswith lower soil metal tolerance. This suggests that the currentB. populifolia dominated hardwood assemblage may not be the endpoint for assemblage development. To better understand thepotential for litter to act as a sink that would mitigate the potentialfor metal induced plant stress the metal flux within heterogeneousmixture of soil compounds and assemblage conditions will need tobe modeled. Such a model will have to consider the complexrelationships between soil pH, soil moisture and redox potential(Bostick et al., 2001).
5. Conclusion
The restoration of degraded sites especially those associatedwith the urban context has become an area of increasing interest(Dobson et al., 1997; Young, 2000). Many of these sites lack anecological legacy and often do not respond predictably to tradi-tional management practices (Hobbs and Harris, 2001; Klotzli andGrootjans, 2001). While succession has provided the model onwhich most restoration efforts have been based, their success rateshave been varied and are somewhat disappointing (Lockwood,1997). Our study demonstrates that the anthropogenic legacy ofan urban brownfield can produce an environmental stress gradientresulting in distinct vegetative assemblages with different guildtrajectories. In addition, the eventual end point for assemblagedevelopment is uncertain as the continued presence of elevated soilmetals has the potential to lead to the development of an alterna-tive steady state.
It may therefore be more effective to define long-term restora-tion objectives with flexible endpoints that clearly articulate thepotential for the site exhibit alternate trajectories or remain in analternate steady state rather than proceed along the traditionalfixed trajectory. Specifically, managers of sites within the urbancontext should consider the role that naturally assembled urbanwildlands can play in contaminant stabilization before prescribinga traditional a species mix based upon traditional concepts ofdiversity and accepted trajectories.
Please cite this article in press as: Gallagher, F.J., et al., Altered vegetativePollution (2011), doi:10.1016/j.envpol.2011.02.007
Acknowledgements
FJG and IP contributed equally to this study. The authors wish tothank Dr Francisco Artigas (New Jersey Meadowlands Commission)for providing access to the ArcMap software and its extension, theGeostatistical Analyst and Dr. Peddrick Weis for his assistance withthe initial soils analysis. We would also like to thank both Dr. PeterSmouse and Dr. Julie Lockwood for their review and advice con-cerning the manuscript. In addition we would like to thankDr. Karina Schafer for the data concerning leaf litter metalconcentration. The New Jersey Department of EnvironmentalProtection partially funded this project.
References
Adriano, D.C., 1986. Trace Elements in the Terrestrial Environment. Springer-Verlag,New York, pp. 156.
Baker, A.J., 1981. Accumulators and excluders e strategies in the response of plantsto heavy metals. Journal of Plant Nutrition 3, 643e654.
Belyea, L.R., 2004. Beyond ecological filters: Feedback networks in the assembly andretroaction of community structure. In: Temperton, V.M., Hobbs, R.J., Nuttle, T.J.,Halle, S. (Eds.), Assembly Rules and Restoration Ecology: Bridging the GapBetween Theory and Practice. Island Press, Washington, D.C., pp. 115e131.
Bertness, M.D., Callaway, R., 1994. Positive interactions in communities. Trends inEcology and Evolution 9, 191e193.
Bostick, B.C., Hansel, C.M., Fendorf, S., 2001. Seasonal fluctuations in zinc speciationwithin a contaminated wetland. Environmental Science & Technology 35,3823e3829.
Clements, F.E., 1916. Plant Succession. Publication 242. Carnegie Institution,Washington, D.C.
Connell, J.H., Slatyer, R.O., 1977. Mechanisms of succession in natural communitiesand their role in community stability and organization. American Naturalist 111,1119e1144.
Dobson, A., Bradshaw, A.D., Baker, A.J.M., 1997. Hopes for the future: restorationecology and conservation biology. Science 227, 515e522.
Dudka, S., Ponce-Hernandez, R., Tate, G., Hutchinson, T.C., 1996. Forms of Cu, Ni andZn in soils of Sudbury, Ontario and the metal concentrations in plants. Water,Air and Soil Pollution 90, 531e542.
Edinger, G.J., Evans, D.J., Gebauer, S., Howard, T.G., Hunt, D.M., Olivero, A.M., 2002.Ecological Communities of New York State. New York Natural Heritage Program,second ed. New York State Department of Environmental Conservation, Albany,NY, pp. 21.
French, C.J., Dickinson, N.M., Putwain, P.D., 2006. Woody biomass phytoremediationof contaminated brownfield land. Environmental Pollution 141, 387e395.
Gadd, G.M., 1999. Fungal production of citric and oxalic acid: importance in metalspeciation in physiology and biochemical process. Advances in MicrobialPhysiology 41, 47e92.
Gallagher, F.J., Pechmann, I., Bogden, J.D., Grabosky, J., Weis, P., 2008a. Soil metalconcentrations and vegetative assemblage structure in an urban brownfield.Environmental Pollution 153, 351e361.
Gallagher, F.J., Pechmann, I., Bogden, J.D., Grabosky, J., Weis, P., 2008b. Soil metalconcentrations and productivity of Betula populifolia (gray birch) as measuredby field spectrometry and incremental annual growth in an abandoned urbanBrownfield in New Jersey. Environmental Pollution 156, 699e706.
Gallagher, F.J., Pechmann, I., Isaacson, B., Grabosky, J. Morphological variation in theseed of Betula populifolia Marsh: a comparison of trees from metal contami-nated soils. Urban Habitats, in press.
Gleason, H.A., 1926. The individualistic concept of plant association. Bulletin of theTorrey Botany Club 53, 7e26.
Gleason, H.A., Cronquist, A., 1991. Manual of the Vascular Plants of NortheasternUnited States and Adjacent Canada, second ed. The New York Botanical GardenPress, Bronx, NY.
Harrison, S., 1999. Local and regional diversity in a patchy landscape: native, alien,and endemic herbs on serpentine. Ecology 80, 70e80.
Hillary, J.D., Wilkins, D.A., 1987. Zinc tolerance in Betula spp., effect of externalconcentration of zinc on growth and uptake. New Phytologist 106, 517e524.
Hobbs, R.J., Harris, J.A., 2001. Restoration ecology: repairing the Earth’s ecosystemsin the new millennium. Restoration Ecology 9, 239e246.
Hobbs, J.D., Norton, D.A., 2004. In: Templeton, V.M.R.J., Hobbs, T., Nuttle, T., Halle, S.(Eds.), Assembly Rules and Restoration Ecology. Island Press, p. 77.
Juang, K.W., Lee, D.Y., Ellsworth, T.R., 2001. Using rank-order geostatistics for spatialinterpolation of highly skewed data in heavy-metal contaminated site. Journalof Environmental Quality 30, 894e903.
Kalin, M., van Everdingen, R.O., 1988. Ecological engineering: biological andgeochemical aspects, phase I. Experiments. In: Salomons, W., Förstner, U. (Eds.),Environmental Management of Solid Waste: Dredged Material and Mine Tail-ings. Springer-Verlag, Berlin, pp. 114e130.
Keddy, P.A., 1992. Assembly and response rules: two goals for predicting commu-nity ecology. Journal of Vegetative Science 3, 157e164.
assemblage trajectories within an urban brownfield, Environmental

F.J. Gallagher et al. / Environmental Pollution xxx (2011) 1e88
Kimmerer, R.W., 1981. Natural revegetation of abandoned lead and zinc mines(Wisconsin). Restoration Management, 1e20.
Klotzli, F., Grootjans, A.P., 2001. Restoration of natural and seminatural wetlandsystems in Central Europe: progress and predictability of developments.Restoration Ecology 9, 209e219.
Lawton, J.H., 1987. Are there assembly rules for successional communities? Colo-nization, succession and stability. In: Gray, A.J., Crawley, M.J., Edwards, P.J. (Eds.),Symposium 26 of the British Ecological Society. Blackwell Scientific Publica-tions, Oxford, pp. 225e244.
Ledin, M., 2000. Accumulation of metals by microorganisms-process and impor-tance for soil systems. Earth-Science Review 51, 1e31.
Leenaers, H., Okx, J.P., Burrough, P.A., 1990. Comparison of spatial predictionmethods for mapping floodplain soil pollution. Catena 15 (6), 535e550.
Li, M.S., Luo, Y.P., Su, Z.Y., 2006. Heavy metal concentrations in soils and plantaccumulation in a restored manganese mineland in Guangxi, South China.Environmental Pollution 141, 1e8.
Lockwood, J.L., 1997. An alternative to succession: assembly rules offer guide torestoration efforts. Restoration and Management Notes 15, 45e50.
MacFarlane, David M., 1996. Liberty State Park Natural Resource InventorySummary The Ecology of Liberty State Park: A Historical Perspective. Depart-ment of Environmental Protection, Trenton, NJ.
MacFarlane, G.R., Burchett, M.D., 2000. Cellular distribution of copper, lead and zincin the gray mangrove Avicennia marina (Forsk.). Vierh. Aquatic Botany 68,45e59.
McGrath, D., Zhang, Ch, Carton, O.T., 2004. Geostatistical analyses and hazardassessment on soil lead in Silvermans area, Ireland. Environmental Pollution127, 239e248.
New Jersey Department of Environmental Protection, 1995. Liberty State Park SoilStudy.
Niering, W.A., Goodwin, R.H., 1974. Creation of relatively stable shrubs withherbicides: arresting “succession” on rights- of- way and pasturelands. Ecology55, 784e795.
Nystrom, M., Folke, C., Moberg, F., 2000. Coral reef disturbance and resilience ina human dominated environment. Trends in Ecology Evolution 15, 413e417.
Petraitis, P.S., Dudgeon, S.R., 1999. Experimental evidence of the origin of alternativecommunities on rocky intertidal shores. Oikos 84, 239e245.
Reynolds, H.L., Hungate, B.A., Chapin, F.S., Dantonio, C.M., 1997. Soil heterogeneityand plant competition in an annual grassland. Ecology 78, 2076e2090.
Ross, S.M., 1994. Toxic Metals in Soil and Plant Systems. Wiley Press, New York,pp. 398.
Sanguer, H., Jetschke, G., 2004. Are assembly rules apparent in the regeneration ofa former uranium mining site? In: Temperton, V.M., Hobbs, R.J., Nuttle, T.,Jalle, S. (Eds.), Assembly Rules and Restoration Ecology: Bridging the GapBetween Theory and Practice. Island Press, Washington, D.C, p. 305.
Please cite this article in press as: Gallagher, F.J., et al., Altered vegetativePollution (2011), doi:10.1016/j.envpol.2011.02.007
Scheffer, M., Carpenter, S.J.A., Foley, C., Folke, C., Walker, B., 2001. Catastrophic shiftsin ecosystems. Nature 413, 591e596.
Stein, L., 1999. Interpolation of Spatial Data. Some Theory for Kriging. Springer, NewYork, pp. 247.
Suding, K.N., Gross, K.L., Houseman, G.R., 2004. Alternative states and positivefeedbacks in restoration ecology. Trends in Ecology and Evolution 19, 46e53.
Sutherland, J.P., 1974. Multiple stable points in natural communities. The AmericanNaturalist 108, 859e872.
Texas Instruments (TI), 1976. Liberty State Park Ecological Study. Prepared for thePort Authority of New York and New Jersey.
United States Army Corps of Engineers, 2004. Hudson-Raritan Estuary Environ-mental Restoration Study, Liberty State Park, Integrated EnvironmentalResource Inventory and Draft Feasibility Study, pp. 151.
Van der Maarel, E., Sykes, M.T., 1993. Small-scale plant species turnover in lime-stone grasslands: the carousel model and some comments on the nicheconcept. Journal of Vegetative Science 4, 179e188.
Van de Koppel, J., Herman, P.M.J., Thoolen, P., Heip, C.R., 2001. Do alternative stablestates occur in natural ecosystems? Evidence from a tidal flat. Ecology 82,3449e3461.
Vesk, P.A., Nockolds, C.E., Allaway, W.G., 1999. Metal localization in water hyacinthroots from an urban wetland. Plant, Cell and Environment 22, 149e158.
Walker, S., Wilson, J.B., 2002. Tests for nonequilibrium, instability, and stabilizingprocesses in semiarid plant communities. Ecology 83, 809e822.
Wagner, M., 2004. The roles of seed dispersal ability and seedling salt tolerance incommunity assembly of a severely degraded site. In: Temperton, V.M.,Hobbs, R.J., Nuttle, T., Jalle, S. (Eds.), Assembly Rules and Restoration Ecology:Bridging the Gap Between Theory and Practice. Island Press, Washington, DC,p. 266.
Weiher, E., Keddy, P.A., 1995. Assembly rules, null models, and trait dispersion: newquestions from old patterns. Oikos 74, 159e194.
Weis, J.S., Weis, P., 2004. Metal uptake, transport and release by wetland plants:implications for phytoremediation and restoration. Environment International30, 685e700.
Wilson, J.B., Gitay, H., 1995. Limitations to species coexistence: evidence forcompetition from field observations, using a patch model. Journal of VegetationScience 6, 369e376.
Wu, J., Norvel, W.A., Welch, R.M., 2006. Kriging on highly skewed data for DTPA-extractable soil Zn with auxiliary information for pH and organic carbon.Geoderma 134, 187e199.
Young, T.P., 2000. Restoration ecology and conservation biology. BiologicalConservation 92, 73e82.
Young, T.P., Case, J.M., Huddleston, R.T., 2001. Community succession and assembly,comparing, contrasting and combining paradigms in the context of ecologicalrestoration. Ecological Restoration 19, 5e18.
assemblage trajectories within an urban brownfield, Environmental