acetaldehyde mediates alcohol activation of the mesolimbic dopamine system
TRANSCRIPT

Acetaldehyde mediates alcohol activation of the mesolimbicdopamine system
M. Melis,1,2 P. Enrico,3,4 A. T. Peana4 and M. Diana4
1Centre of Excellence on Neurobiology of Addiction and2B.B. Brodie Department of Neuroscience, University of Cagliari, Monserrato, Cagliari, Italy3Department of Biomedical Sciences and4G. Minardi Laboratory of Cognitive Neurosciences, Department of Drug Sciences, University of Sassari, Via Muroni 23 ⁄ A,07100 Sassari, Italy
Keywords: addiction, alcoholism, conditioned place preference, electrophysiology, microdialysis, rat
Abstract
Ethanol (EtOH), the main psychoactive ingredient of alcoholic drinks, is widely considered to be responsible for alcohol abuse andalcoholism through its positive motivational properties, which depend, at least partially, on the activation of the mesolimbicdopaminergic system. However, acetaldehyde (ACD), the first metabolite of EtOH, has been classically considered to be aversiveand useful in the pharmacological therapy of alcoholics. Here we show that EtOH-derived ACD is necessary for EtOH-induced placepreference, a pre-clinical test with high predictive validity for reward liability. We also found that ACD is essential for EtOH-increasedmicrodialysate dopamine (DA) levels in the rat nucleus accumbens and that this effect is mimicked by intra-ventral tegmental area(VTA) ACD administration. Furthermore, in vitro, ACD enhances VTA DA neuronal firing through action on two ionic currents:reduction of the A-type K+ current and activation of the hyperpolarization-activated inward current. EtOH-stimulating properties on DAneurons are prevented by pharmacological blockade of local catalase, the main metabolic step for biotransformation of EtOH intoACD in the central nervous system. These results provide in-vivo and in-vitro evidence for a key role of ACD in the motivationalproperties of EtOH and its activation of the mesolimbic DA system. Additionally, these observations suggest that ACD, by increasingVTA DA neuronal activity, would oppose its well-known peripherally originating aversive properties. Careful consideration of thesefindings could help in devising new effective pharmacological therapies aimed at reducing EtOH intake in alcoholics.
Introduction
Alcoholism is a major economic, social and health problem and thisaddictive behavior is thought to be triggered by excessive ethanol(EtOH) intake (Goodwin & Gabrielli, 1997), widely considered to bethe main psychoactive ingredient of alcoholic drinks. Further, EtOH is,after nicotine, the most abused substance in the Western world andEtOH abuse and dependence rank first of all psychiatric disorders inlifetime prevalence rates (Robins et al., 1984). As for many otherabused drugs, this compulsive behavior seems to be elicited andmaintained by the powerful reinforcing properties of the drug.However, acetaldehyde (ACD), the first metabolite of EtOH, producedperipherally by alcohol dehydrogenase (Baraona et al., 1991) andcentrally by the brain catalase ⁄ H2O2 system (Gill et al., 1996) uponEtOH ingestion, has long been thought to be aversive (Suh et al.,2006). Accordingly, ACD accumulation after administration ofdisulfiram (an aldehyde dehydrogenase blocker) still represents awidely employed pharmacological strategy to reduce EtOH drinkingin alcoholics (Suh et al., 2006).In sharp contrast with these notions, mounting evidence suggests
that ACD may share with EtOH some of its positive motivationalproperties. Indeed, ACD induces conditioned stimulus (Quertemont &De Witte, 2001) and place preference (Smith et al., 1984), and non-dependent animals self-administer ACD through an intracerebro-
ventricular route (Amit & Smith, 1985), whereas alcohol-preferringrats operate to obtain ACD directly into the ventral tegmental area(VTA) (Rodd-Henricks et al., 2002). This evidence suggests that ACDmay possess positive motivational properties per se (see Quertemontet al., 2005 for a recent review). In line with this, we recently observed(Foddai et al., 2004) that exogenous ACD stimulates VTA dopamine(DA) neurons, which play a central role in motivation, reward-relatedbehaviors and drug addiction (Girault & Greengard, 2004; Tupala &Tiihonen, 2004; Melis et al., 2005; Kienast & Heinz, 2006), and thatEtOH-stimulating properties on these cells are prevented by admin-istration of the alcohol dehydrogenase blocker 4-methyl-pyrazole(4-MP) (Foddai et al., 2004). Further, d-penicillamine, an effectiveACD-chelating agent (Nagasawa et al., 1978), selectively preventsspontaneous EtOH-drinking behavior (Font et al., 2006a), EtOH-induced place preference (Font et al., 2006b) and motor stimulation(Font et al., 2005). All of these observations suggest that ACD,formed metabolically after EtOH administration, mediates the moti-vational properties of EtOH and its activation of the mesolimbic DAsystem, although its intimate (ionic) mechanism remains unknown. Inaddition, these considerations would lead to anticipation of anincrement in dialysate DA concentrations in the nucleus accumbens(NAcc), the terminal area of VTA DA-containing neurons, although aprevious investigation reported a decrease (Ward et al., 1997).Moreover, as humans generally assume alcohol per os, its metabolisminto ACD is mainly due to gastric and hepatic alcohol dehydrogenase(Baraona et al., 1991), which provides the principal source of ACD.Therefore, in the present study we sought to investigate if
Correspondence: Dr Marco Diana, as above.E-mail: [email protected]
Received 23 May 2007, revised 13 September 2007, accepted 13 September 2007
European Journal of Neuroscience, Vol. 26, pp. 2824–2833, 2007 doi:10.1111/j.1460-9568.2007.05887.x
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd
EtOH-derived ACD would produce place preference when adminis-tered by gavage and if this effect is blocked by pharmacologicalblockade of its peripheral metabolism with 4-MP (Waller et al., 1982).We next investigated whether EtOH-formed ACD would alter DAextracellular microdialysate concentrations in the NAcc and if thiseffect is dependent on local VTA mechanisms. Therefore, weemployed whole-cell patch-clamp recordings in vitro to ascertain theionic mechanism underlying VTA DA excitation previously observedin vivo (Foddai et al., 2004). As activation of the DA system only is acommon property of both aversive (neuroleptics) and rewarding(addictive) compounds, we combined biochemical and electrophysi-ological investigations with the conditioning place preference proce-dure to provide further insights into the quality of the motivationalproperties of EtOH-derived ACD.
Materials and methods
All experiments were carried out in strict accordance with the care anduse of animals approved by the American Physiological Society, EECCouncil Directive of 24 November 1986 (86 ⁄ 609) and the EthicalCommittee of University of Sassari. All efforts were made to reducethe number of animals used.
Conditioned place preference experiments
Male Wistar rats (Harlan, Italy) weighing between 200 and 250 g wereused for these experiments. Rats were housed in groups of three tofour per cage and maintained under controlled environmentalconditions (temperature 22 ± 2 �C; humidity 60–65%; 12-h light ⁄ -dark cycle, light on at 08:00 h). All animals were given a standardlaboratory diet and tap water ad libitum. To prevent the effect of stresson behavioral experiments, subjects were habituated to the experi-mental procedures (i.e. handling and oral administration) for at least3 days. Conditioned place preference (CPP) experiments were con-ducted during the light phase of the light ⁄ dark cycle, according to themethod described by Bozarth (1990) for EtOH-induced CPP andconditions described by Quertemont & De Witte (2001) for ACD-induced CPP. The apparatus consisted of two rectangular steel boxes
(48 · 33 · 30 cm, length · width · height) separated by a guillotinedoor. Distinctive visual and tactile cues distinguished the twocompartments, i.e. the wall and floor coloring (one dark gray andthe other light gray), and the floor texture (smooth or grille). Theapparatus was placed in a sound-proof room with constant lightprovided by a 40-W lamp placed above each compartment. Eachexperiment consisted of three phases. During the first phase (day 1,pre-conditioning phase) the guillotine door was kept lifted and each ratwas placed in the center of the opening and was given access to bothcompartments of the apparatus for 30 min. The time spent by each ratin the compartments was recorded and indicated the ‘unconditionedpreference’. During the second phase (days 2–9 for ACD anddays 2–16 for EtOH, conditioning phase) the rats were administeredwith the drugs and placed for 30 min in the less preferred compart-ment. On alternate days, the rats were administered with saline andplaced in the preferred compartment. During the third phase (24 hafter the last treatment, post-conditioning phase) the guillotine doorwas removed and the time spent by each rat in the drug-pairedcompartment was recorded during 30 min (1800 s) observation. Theconditions of the post-conditioning test were identical to those ofthe pre-conditioning test. The difference between the time spent in thedrug-paired compartment in the post-conditioning test with respect tothat spent during the pre-conditioning test is a measure of the degreeof place conditioning induced by the drug (Carr et al., 1989). ACDand EtOH were dissolved in saline (0.9% NaCl) to a final volume of1 mL and administered by gavage [intragastric (i.g.)]. EtOH solutions(20% v ⁄ v) were obtained by dilution of EtOH (96%). 4-MP (Sigma-Aldrich, Italy) was dissolved in saline and administered by intraperi-toneal (i.p.) injection. All drug dilutions were freshly prepared beforeevery experiment. The gavage infusion rate was rapid (about 5 s) andgiven immediately before each conditioning session. 4-MP wasadministered at a dose of 45 mg ⁄ kg every other day (just after theconditioning session with saline to the preferred compartment,approximately 24 h before the EtOH pairing to the less preferredcompartment). Care was taken to balance the daily order of treatments(saline ⁄ saline, 4-MP ⁄ saline, saline ⁄ EtOH, 4-MP ⁄ EtOH) as well asthe daily order of exposures to each compartment. Control animalswere administered the same volume of saline (vehicle). Data areexpressed as mean ± SEM. Average side preferences were analysedby one- or two-way anova followed by Newman-Keuls test forspecific comparisons between treatment groups.
Microdialysis
Male Wistar rats weighing 290–310 g were used for these experi-ments. Microdialysis was performed using laboratory-made I-shapedprobes. Polyacrylonitrile ⁄ sodium methalyl sulfonate copolymer (innerdiameter 0.22 mm, outer diameter 0.31 mm; AN 69, Hospal, Bolo-gna, Italy) was used as the dialysis membrane. The exposed membranelength was 1.5 mm for NAcc and 1 mm for VTA. Probes wereimplanted during general anesthesia (ketamine 75 mg ⁄ kg and mede-tomidine 0.5 mg ⁄ kg, i.p.). Coordinates of the implantation (Paxinos &Watson, 1997) were AP 2.3, LM 1.1 and VD )7.8 (for the NAcc), andAP )5.3, )6.0, LM )0.9 and VD)8.0 (for the VTA) from bregma anddura, respectively. After surgery rats were administered 2.5 mL oftepid glucose solution (5%) and placed in individual perspex cageswith free access to food and water. Microdialysis experiments werecarried out 24–72 h after implantation of the probes. During theexperiments probes were perfused with a Ringer’s solution at a flowrate of 1.5 lL ⁄ min (Bee Hive System, Bioanalytical Systems, Inc.)and 15-min fractions were collected. The composition of the Ringer’ssolution was (in mm): 147 NaCl, 4.0 KCl, 2.2 CaCl2, 1.25 glucose,
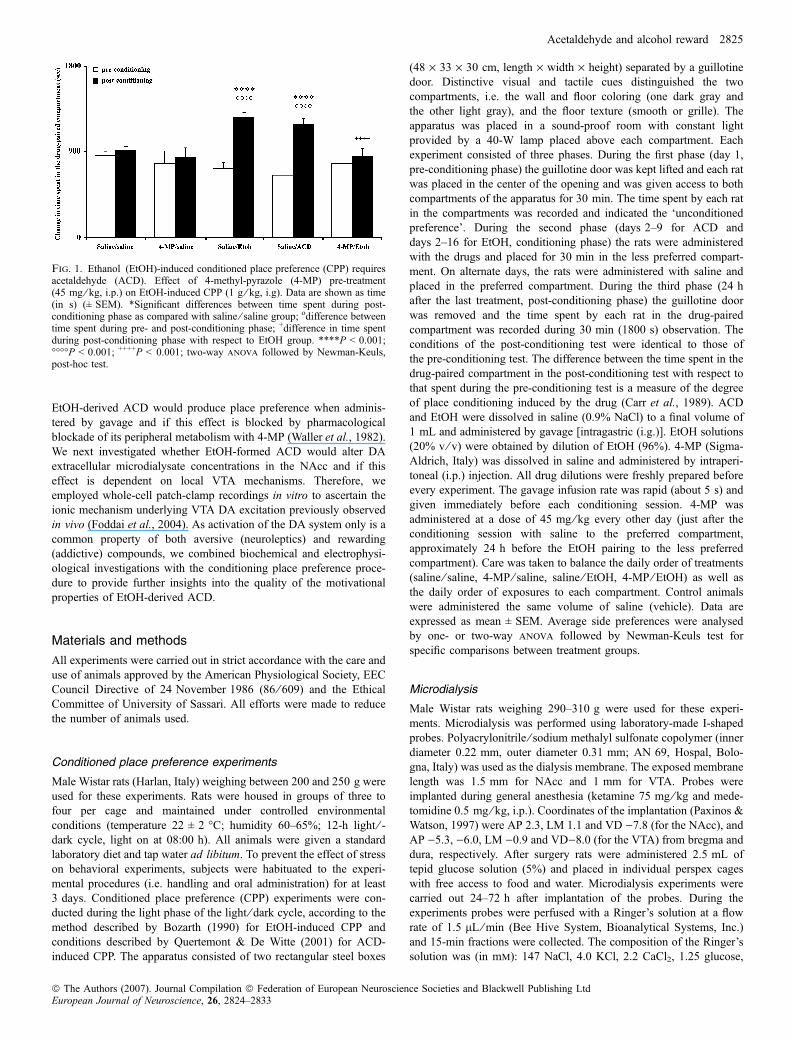
Fig. 1. Ethanol (EtOH)-induced conditioned place preference (CPP) requiresacetaldehyde (ACD). Effect of 4-methyl-pyrazole (4-MP) pre-treatment(45 mg ⁄ kg, i.p.) on EtOH-induced CPP (1 g ⁄ kg, i.g). Data are shown as time(in s) (± SEM). *Significant differences between time spent during post-conditioning phase as compared with saline ⁄ saline group; odifference betweentime spent during pre- and post-conditioning phase; +difference in time spentduring post-conditioning phase with respect to EtOH group. ****P < 0.001;����P < 0.001; ++++P < 0.001; two-way anova followed by Newman-Keuls,post-hoc test.
Acetaldehyde and alcohol reward 2825
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 26, 2824–2833

pH 7.0. For local administration in the VTA, ACD was diluted inRinger’s solution and buffered to pH 7.0 with NaOH. DA wasquantified by high-performance liquid chromatography with electro-chemical detection. A Shimazu pump (LC-10ADvp) was used inconjunction with an electrochemical detector ESA Coulochem II (firstcell 100 mV; second cell )125 mV). A reverse-phase column (length250 mm, diameter 4.7 mm; LC18, Supelco, Bellefonte, PA, USA)was used. The mobile phase consisted of 0.2 m of sodium acetateadjusted to pH 4.5 with acetic acid, 0.1 mm Na EDTA and 10%methanol (flow rate 0.8 mL ⁄ min). 4-MP was administered intraper-itoneally at a dose of 45 mg ⁄ kg, approximately 24 h before EtOHchallenge. Data are expressed as mean ± SEM. Comparisons ofvariations in DA levels within group were made by one-way repeatedmeasures anova and subjected to Dunnett’s post-hoc test whereappropriate.
Electrophysiology
The preparation of VTA slices was described previously (Melis et al.,2006). Briefly, male Sprague Dawley rats (14–21 d) were anesthetizedwith halothane and killed by decapitation. A block of tissue containingthe midbrain was rapidly dissected and sliced in the horizontal plane(300 lm) with a vibratome (Campden Instruments Ltd) in ice-cold
low-Ca2+ solution containing (in mm): 126 NaCl, 1.6 KCl, 1.2NaH2PO4, 1.2 MgCl2, 0.625 CaCl2, 18 NaHCO3 and 11 glucose.Slices (three per animal) were transferred to a holding chamber withartificial cerebrospinal fluid (37 �C) saturated with 95% O2 and 5%CO2 containing (in mm): 126 NaCl, 1.6 KCl, 1.2 NaH2PO4, 1.2MgCl2, 2.4 CaCl2, 18 NaHCO3 and 11 glucose. Slices were allowed torecover for at least 1 h before being placed, as hemislices, in therecording chamber. Cells were visualized with an upright microscopewith infrared illumination (Axioskop FS 2 plus, Zeiss), and whole-cellcurrent- and voltage-clamp recordings were made by using anAxopatch 200B amplifier (Axon Instruments, CA, USA). All record-ings were made with electrodes filled with a solution containing thefollowing (in mm): 144 KCl, 10 HEPES, 3.45 BAPTA, 1 CaCl2,2.5 Mg2ATP and 0.25 Mg2GTP (pH 7.2–7.4, 275–285 mOsm).Experiments were begun only after series resistance had stabilized(typically 15–40 MW). Series and input resistance were monitoredcontinuously on-line with a 5-mV depolarizing step (25 ms). Datawere filtered at 2 kHz, digitized at 10 kHz and collected on-line withacquisition software (pClamp 8.2, Axon Instruments). DA neuronsfrom the medial-posterior VTA were identified by the presence of alarge hyperpolarization-activated inward current (Ih) current (Fordet al., 2006) that was assayed immediately after break-in, using aseries of incremental 10-mV hyperpolarizing steps (250 ms) from a
Fig. 2. Ethanol (EtOH) increases dopamine (DA) release in the nucleus accumbens (NAcc) through its first metabolite acetaldehyde (ACD). (A) Effect of EtOHadministration on DA output in the NAcc shell of rats pre-treated with either saline (top panel) or 4-methyl-pyrazole (4-MP) (middle panel). ACD effect on DAoutput is shown in the bottom panel. (B) Effect of intra-ventral tegmental area administration of ACD on DA output in the NAcc (bar represents time ofadministration). Each point represents the mean ± SEM percent of change from baseline. Arrows indicate time of drug administration. One-way anova
(treatment · time) with repeated measures was followed by Dunnett’s test and used to determine significance. *P < 0.05 vs. baseline values. (C) Placement of theprobe illustrating local ACD administration in the ventral mesencephalon. Left: High magnification (· 100) of the ventral mesencephalon illustrating the location ofthe tip of the probe (thick arrow) in the VTA and probe track (thin arrows). Right: Low magnification (· 5) of a neutral red-stained hemislice of a rat included in thepresent study. *Red nucleus.
2826 M. Melis et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 26, 2824–2833

holding potential of )70 mV. To determine the A-type K+ current(IA), a two-step voltage protocol was used (Hille, 2001); a stepdepolarization from )30 to +50 mV activates most of the residual K+
channels but not IA, whereas depolarization from )120 to +50 mVactivates all K+ currents including IA. The difference between the twotraces represents IA. Artificial cerebrospinal fluid solution wassupplemented with lidocaine (500 lm), CdCl2 (0.4 mm) and tetrae-thylammonium (5 mm) to block Na+, Ca2+ and delayed rectifiedK+ channels, respectively. ZD72988 (ZD) was purchased from TocrisCookson (UK). ACD, EtOH, 3-aminotriazole, 4-aminopyridine(4-AP) and other chemicals were purchased from Sigma-Aldrich.For extracellular application, drugs were applied in known concen-
trations to the superfusion medium. When drugs were dissolved indimethylsulfoxide, its final concentration was < 0.01%. Averageddata from different experiments are presented as mean ± SEM.Statistical significance was assessed using one- or two-way anova
for repeated measures followed either by Dunnett’s or t-test, whereappropriate.
Results
Conditioned place preference
As expected, during the pre-conditioning phase, the spontaneous(unconditioned) preference for the two compartments was slightlyuneven (F1,90 ¼ 10.24, P < 0.05) (data from EtOH and ACD controlgroups), the rank order of preference being light-gray walls and grillefloor (985.5 ± 47.7 s), and dark-gray walls and smooth floor(819.5 ± 47.7 s) during 1800 s of observation. We then chose theless preferred compartment (based on individual choices made by rats,i.e. dark-gray walls and smooth floor) for subsequent conditioningwith rats randomly assigned to experimental groups. Two-way anova
(group · conditioning group) yielded a significant effect of condi-tioning group (F1,74 ¼ 31.13, P < 0.0000005) and group · condi-tioning group interaction (F4,74 ¼ 10.10, P < 0.000005).Rats (n ¼ 20) conditioned (15 days) with i.g. EtOH (1 g ⁄ kg) spent
more time in the less preferred compartment of the apparatus(1276.0 ± 43.1 s) with respect to the pre-conditioning phase(728.3 ± 59.5 s, P < 0.001) and with respect to the post-conditioningphase of the saline ⁄ saline group (917.1 ± 47.2 s, P < 0.001), indi-cating a shift in preference for the EtOH-paired compartment (Fig. 1).In contrast, rats (n ¼ 9) pre-treated with the alcohol dehydrogenaseinhibitor 4-MP (45 mg ⁄ kg, i.p.), per se ineffective, spent less time inthe compartment paired with 4-MP ⁄ EtOH (857.0 ± 78.1 s,P < 0.001) as compared with the group paired with saline ⁄ EtOH-induced CPP. In addition, rats (n ¼ 22) conditioned (8 days) with i.g.ACD (20 mg ⁄ kg) markedly increased their time spent in the lesspreferred compartment of the apparatus (1197.6 ± 59.3 s) as com-pared with the pre-conditioning phase (664.8 ± 62.7 s, P < 0.001)and with respect to the post-conditioning phase of the saline ⁄ salinegroup (917.1 ± 47.2 s, P < 0.001). EtOH and ACD ⁄ saline groups(n ¼ 23; 1 mL ⁄ kg, i.g., 15 and 8 days, respectively) showed nopreference as compared with the pre-conditioning phase and thus datawere pooled together (Fig. 1).
Microdialysis
Ethanol, as well as other drugs of abuse, increases DA release in therat (Imperato & Di Chiara, 1986) and human (Boileau et al., 2003)NAcc. Therefore, we investigated whether EtOH-derived ACD affectsDA release in this brain region of freely behaving rats. DA basalvalues in dialysates of the NAcc for the different experiments did notdiffer significantly and are therefore grouped together. Basal DAvalues (mean ± SEM) were 56.3 ± 3.6 fmol ⁄ 15-min sample. Asshown in Fig. 2A top panel, EtOH (1 g ⁄ kg i.g.) augmented DArelease in the NAcc shell (F7,47 ¼ 5.73, P < 0.0005, one-wayrepeated measures anova followed by Dunnett’s post-hoc test;n ¼ 6, 129.4 ± 4.9% of baseline, P < 0.05). However, its effect wasprevented by pre-treatment with 4-MP (45 mg ⁄ kg i.p.; Fig. 2A middlepanel; F7,47 ¼ 1.28, P > 0.05, one-way repeated measures anova;n ¼ 6, 100.1 ± 4.1% of baseline for 1 g ⁄ kg of EtOH), suggesting thatthe enhancing effect on DA output induced by EtOH may result fromits metabolism into ACD. Accordingly, ACD (20 mg ⁄ kg i.g) (Fig. 2Abottom panel) enhanced DA levels in the NAcc shell to a greater
Fig. 3. Electrophysiological properties of rat medial posterior ventral teg-mental area dopamine (DA) cells. (A) Left: Example of activation ofhyperpolarization-activated inward current (Ih) by hyperpolarizing voltagesteps (10-mV increments) from )70 mV (scale bar, 200 pA, 20 ms). Ih iscomputed as the difference between time points 1 and 2 (see inset). Bottomright: Histogram illustrating the Ih size distribution of all recorded neurons.(B) Top: Example of action potential (AP) waveform (left) and histogram(right) illustrating the AP width of all recorded neurons (scale bar, 5 mV, 2 ms).Bottom: Histograms illustrating the spontaneous activity (frequency, Hz)distribution of the recorded cells (left) and the effect of bath application ofDA (30 lM) on voltage membrane (right).
Acetaldehyde and alcohol reward 2827
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 26, 2824–2833

extent than EtOH and, notably, at a dose 50 times lower(F7,47 ¼ 21.39, P < 0.0001, one-way repeated measures anova
followed by Dunnett’s test; n ¼ 6, 140.7 ± 3.9% of baseline,P < 0.01). Further, we tested whether ACD perfusion, directlyadministered into the VTA, at a concentration of 75 lm (15 min)known to maintain self-administration (Rodd-Henricks et al., 2002),would increase DA output in the NAcc shell. Intra-VTA administra-tion of ACD augmented DA extracellular concentrations in the NAccshell as shown in Fig. 2B (F7,39 ¼ 26.87, P < 0.0001, one-wayrepeated measures anova followed by Dunnett’s test; n ¼ 5,137.4 ± 4.5% of baseline, P < 0.01), indicating that ACD in theVTA is a key mechanism.A histological section containing probe location within the VTA is
shown in Fig. 2C.
Electrophysiology
Whole-cell patch-clamp recordings were made from 22 DA neurons inthe rat medial posterior VTA. We chose this location within the VTAaccording to the recent work by Ford et al. (2006), which demon-
strated that NAcc-projecting DA neurons are mainly located in thisquadrant of the VTA. DA cells displayed electrophysiologicalcharacteristics that, apart from their location in the slice, facilitatedtheir identification (Grace & Onn, 1989; Melis et al., 2006). Theyshowed an Ih of 43.1 ± 3.8 pA (Fig. 3A), fired spontaneously at aregular rate of 3.0 ± 0.1 Hz (Fig. 3B), with long duration actionpotentials (measured from the action potential threshold to themaximum after-hyperpolarization period; Fig. 3B), and responded toDA (30 lm, 5 min) with a hyperpolarization (10.4 ± 0.4 mV,Fig. 3B). Under current-clamp mode, bath application of EtOH at adose (100 mm, 5 min) known to stimulate VTA DA firing (Brodieet al., 1990; Okamoto et al., 2006) induced a reversible andreproducible increase in the firing rate of these cells (69.3 ± 29%,n ¼ 6, P < 0.05, one-way anova for repeated measures; Fig. 4A–C).Because of the high levels of the enzyme catalase in the VTA(Zimatkin & Lindros, 1996; Hung & Lee, 1998), one intriguingpossibility is that EtOH might be oxidized in situ. Therefore, toinvestigate whether or not the effects of EtOH on VTA DA neuronalfrequency discharge (Brodie et al., 1990, 1999; Gessa et al., 1985;Brodie & Appel, 1998; Okamoto et al., 2006) were due to its first
Fig. 4. Ethanol (EtOH) excitation of dopamine (DA) neuronal firing rate requires its first metabolite, acetaldehyde (ACD). (A) Left: Representative traces ofspontaneous activity of DA neurons in control and after acute application of EtOH (100 mm). Right: Time graph illustrating an example of the effect of EtOH onfiring rate of DA cells. Data are normalized to the baseline. The black bar represents the time of superfusion. (B) Time graph showing that ACD is required forethanol-induced increase in firing rate of ventral tegmental area (VTA) DA cells. The effect of EtOH is abolished when ACD formation is prevented by3-aminotriazole (3-AT) (1 mm) application. All data are normalized to the respective baseline (5-min baseline). The black bars represent the time of superfusion.(C) Summary bar graph showing the effects of the first (EtOH) and second (EtOH + EtOH) application of ethanol and in the presence of 3-AT (EtOH + 3-AT) onfiring rate of VTA DA cells. Numbers above columns represent number of experiments. *P < 0.05 significantly different from both first (paired t-test) and second(two-way anova) EtOH application.
2828 M. Melis et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 26, 2824–2833
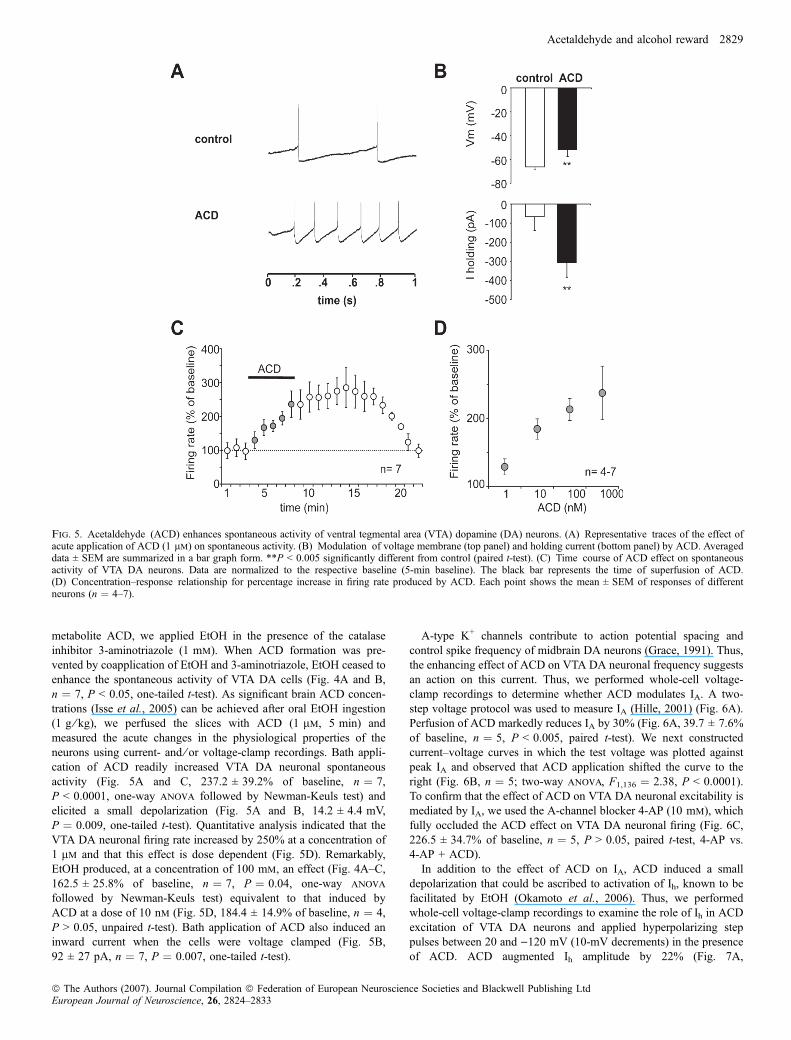
metabolite ACD, we applied EtOH in the presence of the catalaseinhibitor 3-aminotriazole (1 mm). When ACD formation was pre-vented by coapplication of EtOH and 3-aminotriazole, EtOH ceased toenhance the spontaneous activity of VTA DA cells (Fig. 4A and B,n ¼ 7, P < 0.05, one-tailed t-test). As significant brain ACD concen-trations (Isse et al., 2005) can be achieved after oral EtOH ingestion(1 g ⁄ kg), we perfused the slices with ACD (1 lm, 5 min) andmeasured the acute changes in the physiological properties of theneurons using current- and ⁄ or voltage-clamp recordings. Bath appli-cation of ACD readily increased VTA DA neuronal spontaneousactivity (Fig. 5A and C, 237.2 ± 39.2% of baseline, n ¼ 7,P < 0.0001, one-way anova followed by Newman-Keuls test) andelicited a small depolarization (Fig. 5A and B, 14.2 ± 4.4 mV,P ¼ 0.009, one-tailed t-test). Quantitative analysis indicated that theVTA DA neuronal firing rate increased by 250% at a concentration of1 lm and that this effect is dose dependent (Fig. 5D). Remarkably,EtOH produced, at a concentration of 100 mm, an effect (Fig. 4A–C,162.5 ± 25.8% of baseline, n ¼ 7, P ¼ 0.04, one-way anova
followed by Newman-Keuls test) equivalent to that induced byACD at a dose of 10 nm (Fig. 5D, 184.4 ± 14.9% of baseline, n ¼ 4,P > 0.05, unpaired t-test). Bath application of ACD also induced aninward current when the cells were voltage clamped (Fig. 5B,92 ± 27 pA, n ¼ 7, P ¼ 0.007, one-tailed t-test).
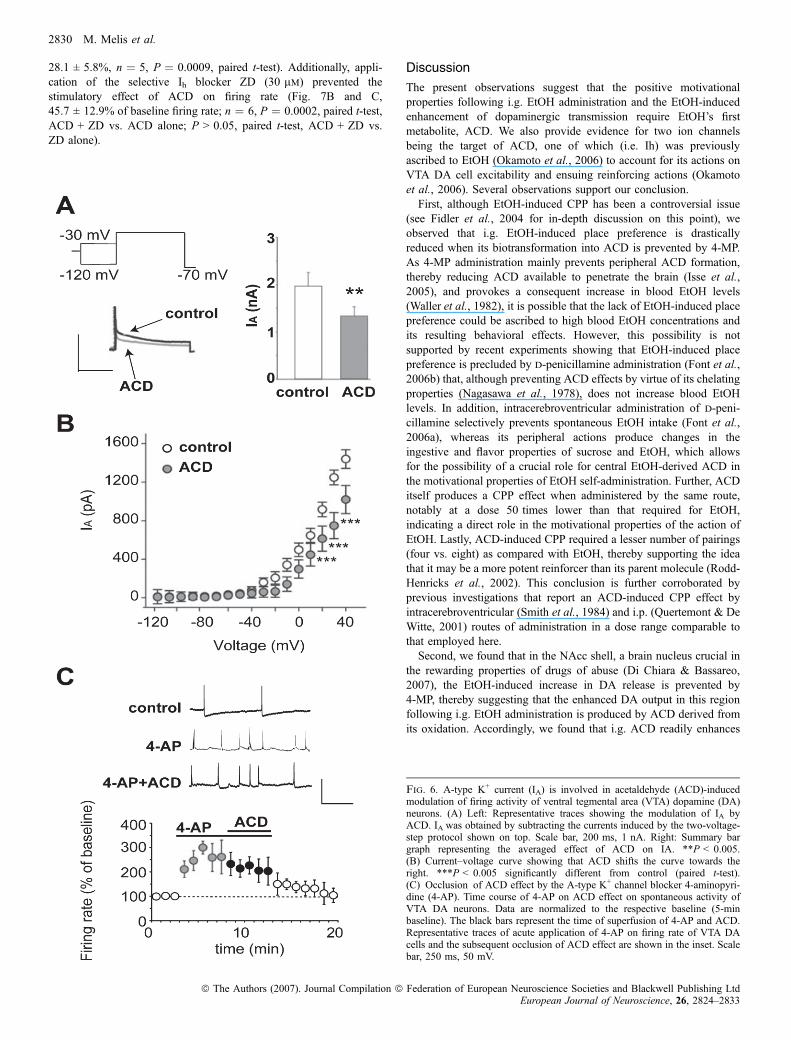
A-type K+ channels contribute to action potential spacing andcontrol spike frequency of midbrain DA neurons (Grace, 1991). Thus,the enhancing effect of ACD on VTA DA neuronal frequency suggestsan action on this current. Thus, we performed whole-cell voltage-clamp recordings to determine whether ACD modulates IA. A two-step voltage protocol was used to measure IA (Hille, 2001) (Fig. 6A).Perfusion of ACD markedly reduces IA by 30% (Fig. 6A, 39.7 ± 7.6%of baseline, n ¼ 5, P < 0.005, paired t-test). We next constructedcurrent–voltage curves in which the test voltage was plotted againstpeak IA and observed that ACD application shifted the curve to theright (Fig. 6B, n ¼ 5; two-way anova, F1,136 ¼ 2.38, P < 0.0001).To confirm that the effect of ACD on VTA DA neuronal excitability ismediated by IA, we used the A-channel blocker 4-AP (10 mm), whichfully occluded the ACD effect on VTA DA neuronal firing (Fig. 6C,226.5 ± 34.7% of baseline, n ¼ 5, P > 0.05, paired t-test, 4-AP vs.4-AP + ACD).In addition to the effect of ACD on IA, ACD induced a small
depolarization that could be ascribed to activation of Ih, known to befacilitated by EtOH (Okamoto et al., 2006). Thus, we performedwhole-cell voltage-clamp recordings to examine the role of Ih in ACDexcitation of VTA DA neurons and applied hyperpolarizing steppulses between 20 and )120 mV (10-mV decrements) in the presenceof ACD. ACD augmented Ih amplitude by 22% (Fig. 7A,
Fig. 5. Acetaldehyde (ACD) enhances spontaneous activity of ventral tegmental area (VTA) dopamine (DA) neurons. (A) Representative traces of the effect ofacute application of ACD (1 lm) on spontaneous activity. (B) Modulation of voltage membrane (top panel) and holding current (bottom panel) by ACD. Averageddata ± SEM are summarized in a bar graph form. **P < 0.005 significantly different from control (paired t-test). (C) Time course of ACD effect on spontaneousactivity of VTA DA neurons. Data are normalized to the respective baseline (5-min baseline). The black bar represents the time of superfusion of ACD.(D) Concentration–response relationship for percentage increase in firing rate produced by ACD. Each point shows the mean ± SEM of responses of differentneurons (n ¼ 4–7).
Acetaldehyde and alcohol reward 2829
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 26, 2824–2833
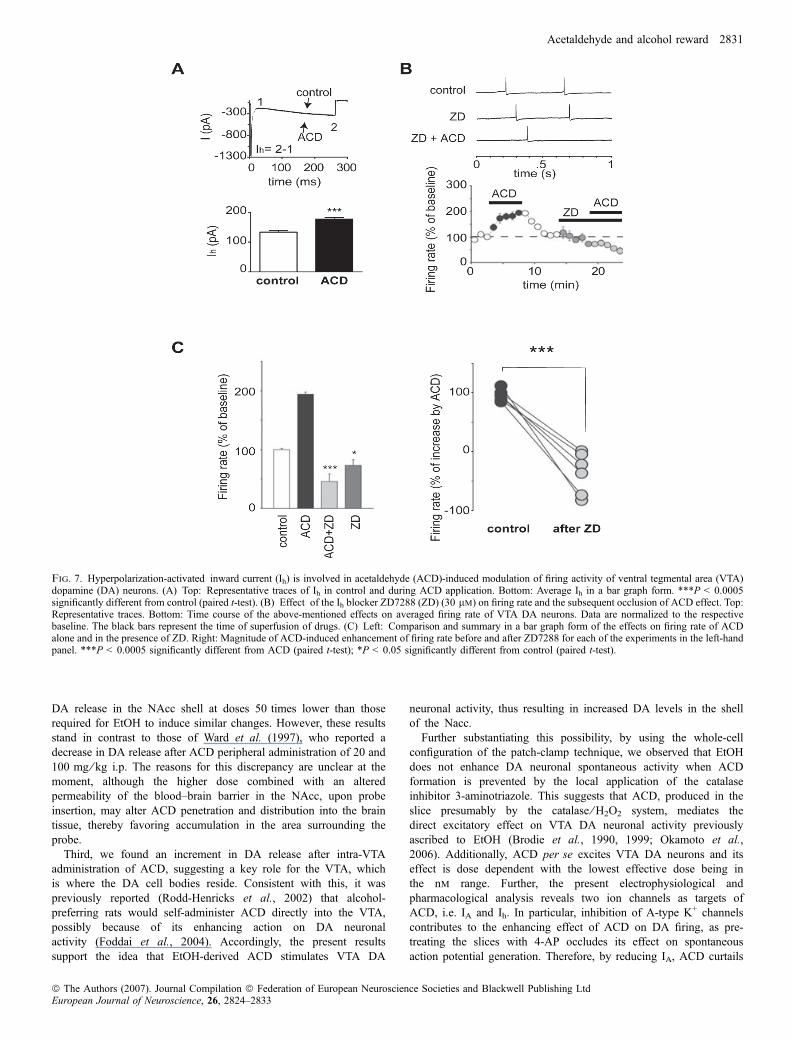
28.1 ± 5.8%, n ¼ 5, P ¼ 0.0009, paired t-test). Additionally, appli-cation of the selective Ih blocker ZD (30 lm) prevented thestimulatory effect of ACD on firing rate (Fig. 7B and C,45.7 ± 12.9% of baseline firing rate; n ¼ 6, P ¼ 0.0002, paired t-test,ACD + ZD vs. ACD alone; P > 0.05, paired t-test, ACD + ZD vs.ZD alone).
Discussion
The present observations suggest that the positive motivationalproperties following i.g. EtOH administration and the EtOH-inducedenhancement of dopaminergic transmission require EtOH’s firstmetabolite, ACD. We also provide evidence for two ion channelsbeing the target of ACD, one of which (i.e. Ih) was previouslyascribed to EtOH (Okamoto et al., 2006) to account for its actions onVTA DA cell excitability and ensuing reinforcing actions (Okamotoet al., 2006). Several observations support our conclusion.First, although EtOH-induced CPP has been a controversial issue
(see Fidler et al., 2004 for in-depth discussion on this point), weobserved that i.g. EtOH-induced place preference is drasticallyreduced when its biotransformation into ACD is prevented by 4-MP.As 4-MP administration mainly prevents peripheral ACD formation,thereby reducing ACD available to penetrate the brain (Isse et al.,2005), and provokes a consequent increase in blood EtOH levels(Waller et al., 1982), it is possible that the lack of EtOH-induced placepreference could be ascribed to high blood EtOH concentrations andits resulting behavioral effects. However, this possibility is notsupported by recent experiments showing that EtOH-induced placepreference is precluded by d-penicillamine administration (Font et al.,2006b) that, although preventing ACD effects by virtue of its chelatingproperties (Nagasawa et al., 1978), does not increase blood EtOHlevels. In addition, intracerebroventricular administration of d-peni-cillamine selectively prevents spontaneous EtOH intake (Font et al.,2006a), whereas its peripheral actions produce changes in theingestive and flavor properties of sucrose and EtOH, which allowsfor the possibility of a crucial role for central EtOH-derived ACD inthe motivational properties of EtOH self-administration. Further, ACDitself produces a CPP effect when administered by the same route,notably at a dose 50 times lower than that required for EtOH,indicating a direct role in the motivational properties of the action ofEtOH. Lastly, ACD-induced CPP required a lesser number of pairings(four vs. eight) as compared with EtOH, thereby supporting the ideathat it may be a more potent reinforcer than its parent molecule (Rodd-Henricks et al., 2002). This conclusion is further corroborated byprevious investigations that report an ACD-induced CPP effect byintracerebroventricular (Smith et al., 1984) and i.p. (Quertemont & DeWitte, 2001) routes of administration in a dose range comparable tothat employed here.Second, we found that in the NAcc shell, a brain nucleus crucial in
the rewarding properties of drugs of abuse (Di Chiara & Bassareo,2007), the EtOH-induced increase in DA release is prevented by4-MP, thereby suggesting that the enhanced DA output in this regionfollowing i.g. EtOH administration is produced by ACD derived fromits oxidation. Accordingly, we found that i.g. ACD readily enhances
Fig. 6. A-type K+ current (IA) is involved in acetaldehyde (ACD)-inducedmodulation of firing activity of ventral tegmental area (VTA) dopamine (DA)neurons. (A) Left: Representative traces showing the modulation of IA byACD. IA was obtained by subtracting the currents induced by the two-voltage-step protocol shown on top. Scale bar, 200 ms, 1 nA. Right: Summary bargraph representing the averaged effect of ACD on IA. **P < 0.005.(B) Current–voltage curve showing that ACD shifts the curve towards theright. ***P < 0.005 significantly different from control (paired t-test).(C) Occlusion of ACD effect by the A-type K+ channel blocker 4-aminopyri-dine (4-AP). Time course of 4-AP on ACD effect on spontaneous activity ofVTA DA neurons. Data are normalized to the respective baseline (5-minbaseline). The black bars represent the time of superfusion of 4-AP and ACD.Representative traces of acute application of 4-AP on firing rate of VTA DAcells and the subsequent occlusion of ACD effect are shown in the inset. Scalebar, 250 ms, 50 mV.
2830 M. Melis et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 26, 2824–2833
DA release in the NAcc shell at doses 50 times lower than thoserequired for EtOH to induce similar changes. However, these resultsstand in contrast to those of Ward et al. (1997), who reported adecrease in DA release after ACD peripheral administration of 20 and100 mg ⁄ kg i.p. The reasons for this discrepancy are unclear at themoment, although the higher dose combined with an alteredpermeability of the blood–brain barrier in the NAcc, upon probeinsertion, may alter ACD penetration and distribution into the braintissue, thereby favoring accumulation in the area surrounding theprobe.
Third, we found an increment in DA release after intra-VTAadministration of ACD, suggesting a key role for the VTA, whichis where the DA cell bodies reside. Consistent with this, it waspreviously reported (Rodd-Henricks et al., 2002) that alcohol-preferring rats would self-administer ACD directly into the VTA,possibly because of its enhancing action on DA neuronalactivity (Foddai et al., 2004). Accordingly, the present resultssupport the idea that EtOH-derived ACD stimulates VTA DA
neuronal activity, thus resulting in increased DA levels in the shellof the Nacc.Further substantiating this possibility, by using the whole-cell
configuration of the patch-clamp technique, we observed that EtOHdoes not enhance DA neuronal spontaneous activity when ACDformation is prevented by the local application of the catalaseinhibitor 3-aminotriazole. This suggests that ACD, produced in theslice presumably by the catalase ⁄ H2O2 system, mediates thedirect excitatory effect on VTA DA neuronal activity previouslyascribed to EtOH (Brodie et al., 1990, 1999; Okamoto et al.,2006). Additionally, ACD per se excites VTA DA neurons and itseffect is dose dependent with the lowest effective dose being inthe nm range. Further, the present electrophysiological andpharmacological analysis reveals two ion channels as targets ofACD, i.e. IA and Ih. In particular, inhibition of A-type K+ channelscontributes to the enhancing effect of ACD on DA firing, as pre-treating the slices with 4-AP occludes its effect on spontaneousaction potential generation. Therefore, by reducing IA, ACD curtails
Fig. 7. Hyperpolarization-activated inward current (Ih) is involved in acetaldehyde (ACD)-induced modulation of firing activity of ventral tegmental area (VTA)dopamine (DA) neurons. (A) Top: Representative traces of Ih in control and during ACD application. Bottom: Average Ih in a bar graph form. ***P < 0.0005significantly different from control (paired t-test). (B) Effect of the Ih blocker ZD7288 (ZD) (30 lm) on firing rate and the subsequent occlusion of ACD effect. Top:Representative traces. Bottom: Time course of the above-mentioned effects on averaged firing rate of VTA DA neurons. Data are normalized to the respectivebaseline. The black bars represent the time of superfusion of drugs. (C) Left: Comparison and summary in a bar graph form of the effects on firing rate of ACDalone and in the presence of ZD. Right: Magnitude of ACD-induced enhancement of firing rate before and after ZD7288 for each of the experiments in the left-handpanel. ***P < 0.0005 significantly different from ACD (paired t-test); *P < 0.05 significantly different from control (paired t-test).
Acetaldehyde and alcohol reward 2831
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 26, 2824–2833

the interspike interval and increases pacemaker spike frequency(Liss et al., 2001; Koyama & Appel, 2006b). In line withthese findings, it has recently been shown that an A-type K+
channel mediated by Kv4.3 and KChip3 subunits has a key role inpacemaker control of adjacent substantia nigra DA neurons (Lisset al., 2001). Also, ACD, by facilitating Ih, shortens and reducesthe after-hyperpolarization period, further contributing toincreased excitability (Neuhoff et al., 2002) and increased pace-maker firing. Consistently, in the present experiments, we neverobserved a shift to bursting mode but a pure increment in regularspontaneous firing after ACD, whereas in the whole animal (Foddaiet al., 2004) ACD dose dependently increased burst firing. Thisfurther suggests that, in spite of the fact that in-vivo studies haveshown that VTA DA neurons discharge less regularly and moreoften in burst mode (as compared with substantia nigra DAneurons) (Chiodo et al., 1984; Grace & Bunney, 1984a,b;Greenhoff et al., 1988), local ACD has a selective action onpacemaker-generating mechanisms, reminiscent of that described forEtOH (Gessa et al., 1985).Although we demonstrate an acute activation of VTA DA neurons
by ACD through action on these two ion channels, at this stage we canneither identify the molecular mechanisms underlying such actions norrule out the involvement of other ion channels, such as the effect onM-type currents (Koyama & Appel, 2006a; Koyama et al., 2007).Other candidates that need to be explored are persistent sodiumchannels (Grace, 1991; Catterall, 2000; Maurice et al., 2001) and low-threshold calcium channels (Kang & Kitai, 1993; Cardozo & Bean,1995; Perez-Reyes, 1999).Altogether these findings reveal a previously unexpected role of
ACD in EtOH activation of the mesolimbic DA system and itsrewarding properties. Indeed, the combination of DA neuronstimulating properties, as indexed by microdialysis and electrophys-iological findings, would not be sufficient to ascribe to ACDpositive motivational properties, as neuroleptic drugs are known toinduce similar effects although endowed with aversive properties.Thus, the behavioral experiments (CPP) are pivotal to ascribepositive motivational properties to metabolically formed ACD, afterEtOH ingestion. Further, our results challenge the assumption thatEtOH per se mediates its own rewarding properties and support thehypothesis that ACD is required for EtOH-induced euphoria. Thepresent observations support and extend previous studies reportingthat EtOH failed to stimulate locomotor activity in rats pre-treatedwith catalase inhibitors (Sanchis-Segura et al., 1999a,b; Escarabajalet al., 2000; Arizzi-LaFrance et al., 2006), thus corroborating thehypothesis that central ACD is necessary for EtOH stimulat-ing ⁄ rewarding properties.Although speculative at this time, the present observations may also
have important theoretical consequences on the therapeutic side ofalcoholism, tobacco dependence (Pianezza et al., 1998) and drugaddiction in general (Kreek et al., 2005). Indeed, these results suggestthat pharmacological blockade of EtOH metabolism would deprive itof its rewarding properties and, quite possibly, discourage individualsfrom drinking. Consistently, 4-MP has been found to be effective inreducing spontaneous alcohol intake in rodent strains selected for highalcohol preference (Waller et al., 1982) and inactivation of centralACD abolishes voluntary EtOH consumption in unselected rats (Fontet al., 2006a).In conclusion, these results indicate that the motivational and
cellular mechanisms underlying EtOH activation of the mesolimbicDA system require its metabolite ACD, which by acting on IA and Ihincreases pacemaker firing and contributes to the positive motivationalproperties of EtOH.
Acknowledgements
We thank Drs S. Perra, B. Schilstrom, S. Marinelli and M.A. Ungless for manyuseful comments, A. Lintas and S. Spiga for histology and figure preparation,and W.T. Dunn III for proof-reading this manuscript. This work was supportedby grants PRIN 2006 to M.D. (MIUR, 2006057754) and Fondazione Banco diSardegna (2006.0454) to A.T.P.
AbbreviationsACD, acetaldehyde; 4-AP, 4-aminopyridine; CPP, conditioned place prefer-ence; DA, dopamine; EtOH, ethanol; IA, A-type K+ current; i.g., intragastric;Ih, hyperpolarization-activated inward current; i.p., intraperitoneal; 4-MP, 4-methyl-pyrazole; NAcc, nucleus accumbens; VTA, ventral tegmental area; ZD,ZD7288.
References
Amit, Z. & Smith, B.R. (1985) A multi-dimensional examination of thepositive reinforcing properties of acetaldehyde. Alcohol, 2, 367–370.
Arizzi-LaFrance, M.N., Correa, M., Aragon, C.M. & Salamone, J.D. (2006)Motor stimulant effects of ethanol injected into the substantia nigra parsreticulata: importance of catalase-mediated metabolism and the role ofacetaldehyde. Neuropsychopharmacology, 31, 997–1008.
Baraona, E., Yokoyama, A., Ishii, H., Hernandez-Munoz, R., Takagi, T.,Tsuchiya,M.&Lieber, C.S. (1991) Lack of alcohol dehydrogenase isoenzymeactivities in the stomach of Japanese subjects. Life Sci., 49, 1929–1234.
Boileau, I., Assaad, J.M., Pihl, R.O., Benkelfat, C., Leyton, M., Diksic, M.,Tremblay, R.E. & Dagher, A. (2003) Alcohol promotes dopamine release inthe human nucleus accumbens. Synapse, 49, 226–231.
Bozarth, M.A. (1990) Evidence for the rewarding effects of ethanol using theconditioned place preference method. Pharmacol. Biochem. Behav., 35, 485–487.
Brodie, M.S. & Appel, S.B. (1998) The effects of ethanol on dopaminergicneurons of the ventral tegmental area studied with intracellular recording inbrain slices. Alcohol Clin. Exp. Res., 22, 236–244.
Brodie, M.S., Shefner, S.A. & Dunwiddie, T.V. (1990) Ethanol increases thefiring rate of dopamine neurons of the rat ventral tegmental area in vitro.Brain Res., 508, 65–69.
Brodie, M.S., Pesold, C. & Appel, S.B. (1999) Ethanol directly excitesdopaminergic ventral tegmental area reward neurons. Alcohol Clin. Exp.Res., 23, 1848–1852.
Cardozo, D.L. & Bean, B.P. (1995) Voltage-dependent calcium channels in ratmidbrain dopamine neurons: modulation by dopamine and GABABreceptors. J. Neurophysiol., 74, 1137–1148.
Carr, G.D., Fibiger, H.C. & Phillips, A.G. (1989) Conditioned placepreferences as a measure of drug reward. In Liebman, J. (Ed.), TheNeuropharmacological Basis of Reward. Oxford Science Publication,Clarendon Press, Oxford, pp. 264–319.
Catterall, W.A. (2000) From ionic currents to molecular mechanisms: thestructure and function of voltage-gated sodium channels. Neuron, 26, 13–25.
Chiodo, L.A., Bannon, M.J., Grace, A.A., Roth, R.H. & Bunney, B.S. (1984)Evidence for the absence of impulse-regulating somatodendritic andsynthesis-modulating nerve terminal autoreceptors on subpopulations ofmesocortical dopamine neurons. Neuroscience, 12, 1–16.
Di Chiara, G. & Bassareo, V. (2007) Reward system and addiction: whatdopamine does and doesn’t do. Curr. Opin. Pharmacol., 7, 69–76.
Escarabajal, D., Miquel, M. & Argon, C.M. (2000) A psychopharmacologicalstudy of the relationship between brain catalase activity and ethanol-inducedlocomotor activity in mice. J. Stud. Alcohol, 61, 493–498.
Fidler, T.L., Bakner, L. & Cunningham, C.L. (2004) Conditioned placeaversion induced by intragastric administration of ethanol in rats. Pharmacol.Biochem. Behav., 77, 731–743.
Foddai, M., Dosia, G., Spiga, S. & Diana, M. (2004) Acetaldehyde increasesdopaminergic neuronal activity in the VTA. Neuropsychopharmacology, 29,530–536.
Font, L., Miquel, M. & Aragon, C.M. (2005) Prevention of ethanol-inducedbehavioral stimulation by D-penicillamine: a sequestration agent foracetaldehyde. Alcohol Clin. Exp. Res., 29, 1156–1164.
Font, L., Aragon, C.M. & Miquel, M. (2006a) Voluntary ethanol consumptiondecreases after the inactivation of central acetaldehyde by d-penicillamine.Behav. Brain Res., 171, 78–86.
2832 M. Melis et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 26, 2824–2833

Font, L., Aragon, C.M. & Miquel, M. (2006b) Ethanol-induced conditionedplace preference, but not aversion, is blocked by treatment withD-penicillamine, an inactivation agent for acetaldehyde. Psychopharmacol-ogy (Berl.), 184, 56–64.
Ford, C.P., Mark, G.P. & Williams, J.T. (2006) Properties and opioid inhibitionof mesolimbic dopamine neurons vary according to target location.J. Neurosci., 26, 2788–2797.
Gessa, G.L., Muntoni, F., Collu, M., Vargiu, L. & Mereu, G. (1985) Low dosesof ethanol activate dopaminergic neurons in the ventral tegmental area. BrainRes., 348, 201–203.
Gill, K., Amit, Z. & Smith, B.R. (1996) The regulation of alcohol consumptionin rats: the role of alcohol-metabolizing enzymes ) catalase and aldehydedehydrogenase. Alcohol, 13, 347–353.
Girault, J.A. & Greengard, P. (2004) The neurobiology of dopamine signalling.Arch. Neurol., 61, 641–644.
Goodwin, D.W. & Gabrielli, W.F. Jr (1997) Alcohol: clinical aspects. InLowinson, J.H., Ruiz, P., Millman, R.B. & Langrod, J.G. (Eds), SubstanceAbuse: a Comprehensive Textbook. Williams & Wilkins, Baltimore, MD,pp. 142–148.
Grace, A.A. (1991) Regulation of spontaneous activity and oscillatory spikefiring in rat midbrain dopamine neurons recorded in vitro. Synapse, 7, 221–234.
Grace, A.A. & Bunney, B.S. (1984a) The control of firing pattern in nigraldopamine neurons: single spike firing. J. Neurosci., 4, 2866–2876.
Grace, A.A. & Bunney, B.S. (1984b) The control of firing pattern in nigraldopamine neurons: burst firing. J. Neurosci., 4, 2877–2890.
Grace, A.A. & Onn, S.P. (1989) Morphology and electrophysiologicalproperties of immunocytochemically identified rat dopamine neuronsrecorded in vitro. J. Neurosci., 9, 3463–3481.
Greenhoff, J., Ugedo, L. & Svensson, T.H. (1988) Firing patterns of midbraindopamine neurons: differences between A9 and A10 cells. Acta Physiol.Scand., 134, 127–132.
Hille, B. (2001) Ion Channels of Excitable Membranes, 3rd Edn. SinauerAssociates, Sunderland, MA.
Hung, H.C. & Lee, E.H. (1998) MPTP produces differential oxidative stressand antioxidative responses in the nigrostriatal and mesolimbic dopaminergicpathways. Free Radic. Biol. Med., 24, 76–84.
Imperato, A. & Di Chiara, G. (1986) Preferential stimulation of dopaminerelease in the nucleus accumbens of freely moving rats by ethanol.J. Pharmacol. Exp. Ther., 239, 219–228.
Isse, T., Matsuno, K., Oyama, T., Kitagawa, K. & Kawamoto, T. (2005)Aldehyde dehydrogenase 2 gene targeting mouse lacking enzyme activityshows high acetaldehyde level in blood, brain, and liver after ethanolgavages. Alcohol Clin. Exp. Res., 29, 1959–1964.
Kang, Y. & Kitai, S.T. (1993) Awhole cell patch-clamp study on the pacemakerpotential in dopaminergic neurons of rat substantia nigra compacta.Neurosci. Res., 18, 209–221.
Kienast, T. & Heinz, A. (2006) Dopamine and the diseased brain. CNS Neurol.Disord. Drug Targets, 5, 109–131.
Koyama, S. & Appel, S.B. (2006a) Characterization of M-current in ventraltegmental area dopamine neurons. J. Neurophysiol., 96, 535–543.
Koyama, S. & Appel, S.B. (2006b) A-type K+ current of dopamine andGABA neurons in the ventral tegmental area. J. Neurophysiol., 96, 544–554.
Koyama, S., Brodie, M.S. & Appel, S.B. (2007) Ethanol inhibition of m-currentand ethanol-induced direct excitation of ventral tegmental area dopamineneurons. J. Neurophysiol., 97, 1977–1985.
Kreek, M.J., Bart, G., Lilly, C., La Forge, K.S. & Nielsen, D.A. (2005)Pharmacogenetics and human molecular genetics of opiate and cocaineaddictions and their treatments. Pharmacol. Rev., 57, 1–26.
Liss, B., Franz, O., Sewing, S., Bruns, R., Neuhoff, H. & Roeper, J. (2001)Tuning pacemaker frequency of individual dopaminergic neurons by Kv4.3Land KChip3.1 transcription. EMBO J., 20, 5715–5724.
Maurice, N., Tkatch, T., Meisler, M., Sprunger, L.K. & Surmeier, D.J. (2001)D1 ⁄ D5 dopamine receptor activation differentially modulates rapidlyinactivating and persistent sodium currents in prefrontal cortex pyramidalneurons. J. Neurosci., 21, 2268–2277.
Melis, M., Spiga, S. & Diana, M. (2005) The dopamine hypothesis of drugaddiction: hypodopaminergic state. Int. Rev. Neurobiol., 63, 101–154.
Melis, M., Pillolla, G., Bisogno, T., Minassi, A., Petrosino, S., Perra, S.,Muntoni, A.L., Lutz, B., Gessa, G.L., Marsicano, G., Di Marzo, V. & Pistis,M. (2006) Protective activation of the endocannabinoid system duringischemia in dopamine neurons. Neurobiol. Dis., 24, 15–27.
Nagasawa, H.T., Goon, D.J. & De Master, E.G. (1978) 2,5,5-Trimethylthiaz-olidine-4-carboxylic acid, a D(-)-penicillamine-directed pseudometabolite ofethanol. Detoxication mechanism for acetaldehyde. J. Med. Chem., 21,1274–1279.
Neuhoff, H., Neu, A., Liss, B. & Roeper, J. (2002) I(h) channels contribute tothe different functional properties of identified dopaminergic subpopulationsin the midbrain. J. Neurosci., 22, 1290–1302.
Okamoto, T., Harnett, M.T. & Morikawa, H. (2006) Hyperpolarization-activated cation current (Ih) is an ethanol target in midbrain dopamineneurons of mice. J. Neurophysiol., 95, 619–626.
Paxinos, G. & Watson, C. (1997) The Rat Brain in Stereotaxic Coordinates, 3rdEdn. Academic Press, Sydney.
Perez-Reyes, E. (1999) Three for T: molecular analysis of the low voltage-activated calcium channel family. Cell Mol. Life Sci., 56, 660–669.
Pianezza, M.L., Sellers, E.M. & Tyndale, R.F. (1998) Nicotine metabolismdefect reduces smoking. Nature, 393, 750.
Quertemont, E. & De Witte, P. (2001) Conditioned stimulus preference afteracetaldehyde but not ethanol injections. Pharmacol. Biochem. Behav., 68,449–454.
Quertemont, E., Tambour, S. & Tirelli, E. (2005) The role of acetaldehyde inthe neurobehavioral effects of ethanol: a comprehensive review of animalstudies. Prog. Neurobiol., 75, 247–274.
Robins, L.N., Helzer, J.E., Weissman, M.M., Orvaschel, H., Gruenberg, E.,Burke, J.D. Jr & Regier, D.A. (1984) Lifetime prevalence of specificpsychiatric disorders in three sites. Arch. Gen. Psych., 41, 949–958.
Rodd-Henricks, Z.A., Melendez, R.I., Zaffaroni, A., Goldstein, A., McBride,W.J. & Li, T.K. (2002) The reinforcing effects of acetaldehyde in theposterior ventral tegmental area of alcohol-preferring rats. Pharmacol.Biochem. Behav., 72, 55–64.
Sanchis-Segura, C., Miquel, M., Correa, M. & Aragon, C.M. (1999a) Thecatalase inhibitor sodium azide reduces ethanol-induced locomotor activity.Alcohol, 19, 37–42.
Sanchis-Segura, C., Miquel, M., Correa, M. & Aragon, C.M. (1999b)Cyanamide reduces brain catalase and ethanol-induced locomotor activity:is there a functional link? Psychopharmacology (Berl.), 144, 83–89.
Smith, B.R., Amit, Z. & Splawinsky, J. (1984) Conditioned place preferenceinduced by intraventricular infusions of acetaldehyde. Alcohol, 1, 193–195.
Suh, J.J., Pettinati, H.M., Kampman, K.M. & O’Brien, C.P. (2006) The statusof disulfiram: a half of a century later. J. Clin. Psychopharmacol., 26, 290–302.
Tupala, E. & Tiihonen, J. (2004) Dopamine and alcoholism: neurobiologicalbasis of ethanol abuse. Prog. Neuropsychopharmacol. Biol. Psychiat., 28,1221–1247.
Waller, M.B., McBride, W.J., Lumeng, L. & Li, T.K. (1982) Effects ofintravenous ethanol and of 4-methylpyrazole on alcohol drinking in alcohol-preferring rats. Pharmacol. Biochem. Behav., 17, 763–768.
Ward, R.J., Colantuoni, C., Dahchour, A., Quertemont, E. & De Witte, P.(1997) Acetaldehyde-induced changes in monoamine and amino acidextracellular microdialysate content of the nucleus accumbens. Neurophar-macology, 36, 225–232.
Zimatkin, S.M. & Lindros, K.O. (1996) Distribution of catalase in rat brain:aminergic neurons as possible targets for ethanol effects. Alcohol Alcohol.,31, 167–174.
Acetaldehyde and alcohol reward 2833
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 26, 2824–2833