a palaeogene record of extant lower photic zone calcareous nannoplankton
TRANSCRIPT

A PALAEOGENE RECORD OF EXTANT LOWERPHOTIC ZONE CALCAREOUS NANNOPLANKTON
by P. R. BOWN* , T. DUNKLEY JONES* , J. R. YOUNG! and R. RANDELL**Department of Earth Sciences, University College London, Gower Street, London WC1E 6BT, UK;e-mails: [email protected]; [email protected]; [email protected]!Department of Palaeontology, The Natural History Museum, London SW7 5BD, UK; e-mail: [email protected]
Typescript received 18 January 2008; accepted in revised form 28 July 2008
Abstract: In this paper, we document Palaeogene occur-
rences of the extant nannoplankton genera Gladiolithus, Al-
girosphaera and Solisphaera from claystones of the Kilwa
Group microfossil lagerstatte, coastal Tanzania. In the mod-
ern ocean, these taxa are restricted to tropical and subtropi-
cal lower photic zone habitats (100–200 m). All three genera
produce small and ⁄ or fragile coccoliths, which have not pre-
viously been found as fossils or are only found in late Qua-
ternary sediments. Their presence in the Kilwa Group
sediments demonstrates the exceptional quality of preserva-
tion and provides a minimum origination time for these eco-
logically specialized nannoplankton. Gladiolithus is present in
the oldest Palaeogene Kilwa Group material studied (late
Paleocene, c. 59 Ma), which pushes back the origination time
for lower photic zone coccolithophores by over 40 million
years. The co-occurrence of several species of Gladiolithus
alongside Algirosphaera and Solisphaera suggests that a rela-
tively diverse lower photic zone assemblage was established
by at least the Eocene. The abundant and consistent occur-
rence of specimens that are indistinguishable from the mod-
ern species Gladiolithus flabellatus has been observed in
samples of late Paleocene (Nannofossil Zone NP6) to early
Oligocene (Nannofossil Zone NP21) age and support an
open ocean palaeoenvironmental interpretation for the Kilwa
Group sediments. We describe three new species of Gladioli-
thus, G. brevis, G. ornatus and G. contus and one new combi-
nation, Algirosphaera fabaceus.
Key words: nannoplankton, lower photic zone, Palaeogene,
Gladiolithus, Algirosphaera.
Calcareous nannofossils are the preserved calciticcell-wall components of a major extant phytoplanktongroup, the coccolithophores (algal Division Haptophyta).The majority of coccolithophores live high in the marinewater column (<100 m) and their biogeography andabundance is controlled by factors such as temperature,nutrient levels, thermocline depth and water transparency(Winter et al. 1994). Vertical sampling of the photic zonereveals a tiered structure to plankton distribution (Okadaand Honjo 1973; Winter et al. 1994) and, in particular,the presence of a specialized lower photic zone (LPZ)assemblage, adapted to low light conditions and charac-terized by highly modified coccoliths and coccospheres(Young 1994; Young et al. 2003). The lower photic zonetaxa produce coccoliths that are typically small and oftenquite unlike the classic circular or elliptical liths producedby most coccolithophores and are consequently easilyoverlooked in the fossil record. However, the extant genusFlorisphaera is now frequently documented in Quaternarystudies and occasionally in older Neogene sediments, butprior to this paper, there was no unequivocal documenta-tion of LPZ nannoplankton in the pre-Neogene fossilrecord. Here we report abundant Palaeogene (late Paleo-cene – early Oligocene) occurrences of the extant LPZ
taxon Gladiolithus from the Kilwa Group of Tanzaniaalongside rare occurrences of Algirosphaera and Solisph-aera. Gladiolithus and Algirosphaera have not previouslybeen found as fossils, except in several late Quaternarystudies (e.g. Okada and Matsuoka 1996; Lars Legge et al.2006) and our records push back the minimum date oforigination of these ecologically specialized coccolitho-phores by over 57 million years. Solisphaera has onlyrecently been discovered in the modern plankton and hasno previous fossil record. The presence of these taxa inthe Palaeogene has important implications both for ourunderstanding of the evolution of modern phytoplanktonand for the documentation of pre-Neogene photic zoneenvironments.
MATERIAL AND METHODS
Our records of Gladiolithus, Algirosphaera and Solisphaeracome from the Kilwa Group (Cretaceous–Oligocene) ofcoastal Tanzania. These sediments have been studied inoutcrop and shallow drill cores around the towns of Kilwa,Lindi and Pande (Text-fig. 1), as part of the Tanzania Dril-ling Project (Pearson et al. 2004, 2006; Nicholas et al. 2006;
[Palaeontology, Vol. 52, Part 2, 2009, pp. 457–469]
ª The Palaeontological Association doi: 10.1111/j.1475-4983.2009.00853.x 457

Bown et al. 2008). The Kilwa Group comprises a kilometrethick succession of homogeneous dark clays and claystoneswith variably developed secondary lithologies of siltstones,limestones and sandstones (Nicholas et al. 2006). The sedi-ments have never been deeply buried and the claystonesare frequently uncemented and unlithified. The results wepresent here come from the Palaeogene part of the KilwaGroup, specifically the Kivinje Claystone (Paleocene –Lower Eocene), Masoko Clay (Middle Eocene) and PandeClay (Upper Eocene – Lower Oligocene) formations (Nich-olas et al. 2006). The quality of foraminifera and calcareousnannofossil preservation in these formations has led totheir designation as a Konservat-Lagerstatte for calcareousmicrofossils (Bown et al. 2008). Palaeontological andsedimentological data indicate that the succession wasdeposited at 19!S palaeolatitude in an outer shelf to upperslope position, at water depths of 300–500 m (Pearsonet al. 2004, 2006; Nicholas et al. 2006).
The calcareous nannofossils were studied using transmit-ted light and scanning electron microscopy. Light micro-scope (LM) observation was carried out on simple smearslides (Bown and Young 1998) using a Zeiss Axiophot LMat x1000 to 1250 magnification in cross polarized and phasecontrast light. Accounts of the taxonomic composition of
the Palaeogene nannofossil assemblages based on lightmicroscopy are presented in Bown (2005) and Bown andDunkley Jones (2006). Selected samples have also beenstudied using a JEOL JSM-6480LV scanning electron micro-scope (SEM), primarily viewing broken surfaces of smallrock chips (Lees et al. 2004). Other methods of SEM samplepreparation were unsuccessful because the abundant clayobscured the nannofossils. Rock surface observation wastime consuming, but yielded images of both dispersed nan-nofossils and clusters of nannofossils, in many cases exqui-sitely preserved. The clusters probably represent collapsedfaecal pellets or marine-snow aggregates.
The comparative samples of modern coccolithophoreswere collected during oceanographic research cruises byfiltration of sea-water onto cellulose nitrate or polycar-bonate microfilters. The filters were subsequently ovendried, and imaged using a Philips XL-30 FEG electronmicroscope.
RESULTS
A broad stratigraphic range of Palaeogene samples (latePaleocene – early Oligocene) has been studied as part of
TEXT -F IG . 1 . Location andstratigraphy of the Tanzania DrillingProject core sites and stratigraphicdistribution of the lower photic zonetaxa.
458 PALAEONTOLOGY, VOLUME 52

the Tanzania Drilling Project, but the abundance, diver-sity, preservation and taxonomic character of the cal-careous nannofossil assemblages are rather consistentthroughout. The nannofossils are rare to common andoften significantly diluted by clay and silt in these predom-inantly dark green to black claystone lithologies. However,preservation of the nannofossils is uniformly good andoften exceptional, with delicate structures, small coccoliths(<3.0 lm), holococcoliths and coccospheres consistentlypresent. The assemblages have high species richness values,which are generally significantly higher than have previ-ously been encountered in coeval sections (Bown et al.2008). Biostratigraphic work using light microscopy hasalready resulted in over 80 new Palaeogene species beingdescribed (Bown 2005; Bown and Dunkley Jones 2006).High diversities are conspicuous in several coccolithgroups, in particular, the rhabdoliths (Rhabdosphaeraceae:Blackites), pontosphaerids (Pontosphaeraceae: Pontosphae-ra), holococcoliths (Holodiscolithus, Daktylethra, Varolia,etc.) and pentaliths (Braarudosphaeraceae: Micrantholi-thus, Pemma). These high diversities are primarily theresult of enhanced preservation, with the exception of thepentaliths, which appear to have been more diverse inshelf environments. The quality of preservation is demon-strated not only by enhanced diversity but also by the con-servation of small and delicate coccoliths, completecoccospheres, delicate central structures and taxa that areprone to postmortem dissolution.
Gladiolithus was not identified during the initial LMstudy, but was conspicuous on rock surfaces observedusing a SEM (e.g. Pl. 2, figs 2 and 6). Abundant elongatecalcite laths within coccolith concentrations were initiallyconsidered indeterminate spicules. However, on furtherstudy, they were observed to form tubular coccoliths, andcomplete collapsed coccospheres formed of these tube-coccoliths were also found (Pl. 1, figs 5–6; Pl. 2, figs 2and 5). Comparison of these specimens with modern coc-colithophores revealed a remarkable similarity with thespecies Gladiolithus flabellatus. This includes not only thegross coccolith morphology and size but also the charac-teristic angular element terminations, element edge serra-tions, surface pitting and association with lepidolithcoccoliths, (see below and Systematic Palaeontology sec-tion for descriptions). A majority of specimens are indis-tinguishable from those of the living species Gladiolithusflabellatus (Pl. 1, figs 1–6); however, distinct specimenswith short tube-coccoliths (Pl. 2, figs 1–2, 5–6) and formswith long tube-coccoliths have also been observed (Pl. 2,figs 3–4). Gladiolithus is consistently observed in all of thePalaeogene TDP drill holes, which comprise sediments oflate Paleocene, Eocene and early Oligocene age (Nanno-fossil Zones NP6-7, 9-11, 14-17, 19-21 and 23 of Martini1971). Specimens are often as abundant or more abun-dant than other coccoliths in the assemblages. The
observation of these coccoliths in such abundance andwithin the fabric of the sediments precludes any possibil-ity of contamination.
Algirosphaera and Solisphaera specimens have beenobserved only sporadically and rarely in Eocene and Oli-gocene TDP holes. Solisphaera is found as detached blade-like processes (Pl. 3, figs 10–12), but these are small anddelicate objects and there is no previous record of themhaving survived export from the modern photic zone. Al-girosphaera has been documented in one SEM sample(Sample TDP12 ⁄ 26-2, 62 cm), but has a light microscopicrecord through the late Eocene – early Oligocene of TDPsites 11 and 12 (Nannofossil Zones 19 ⁄ 20-21).
On re-analysis of the LM slides, Gladiolithus was identi-fiable (Pl. 2, figs 8–11), but observation is difficultbecause of the combination of small size, nonuniqueshape and low birefringence in cross-polarized light. Thetube-coccoliths are delicate and tend to disintegrate intothe separate elongate elements, which are thin, narrow,calcite crystals with high-angle c axes. This means theyare virtually invisible in cross-polarized light and evenwhen abundant, they can resemble clay minerals and ⁄orindeterminate micritic particles. The lepidolith coccolithsare also prone to disintegrate into their two semicircularparts, and, in any case, are very small (1–2 lm) and atthe resolution limit for light microscopy. Nevertheless, inmany LM slides, Gladiolithus attains abundances of 100 sof elements per field of view.
LOWER PHOTIC ZONENANNOPLANKTON
Coccolithophores inhabit all marine photic zone environ-ments, but are typically most abundant around the ther-mocline in stratified water masses or near the surface inless stratified settings (Hagino et al. 2000). Vertical differ-entiation of coccolithophores within the photic zone wasfirst documented by Okada and Honjo (1973) whoobserved taxonomically-distinct assemblages between waterdepths of 100 and 200 m over a wide range of tempera-tures (10–28!C) underlying tropical and subtropical Pacificsurface-waters. This assemblage has now been observed inall ocean basins, but is highly correlated with water depthand becomes less abundant across the shelf break and rareor absent in shallow bays and inland seas (Okada 1983,1992). Initially, the LPZ assemblage was considered taxo-nomically conservative, and dominated by Florisphaeraprofunda, Gladiolithus flabellatus and Algirosphaera robusta.However, recent observations of pseudo-cryptic variabilitywithin Florisphaera (Quinn et al. 2005) together withsporadic occurrences of new and distinctive LPZ taxa,notably Syracosphaera spp., Placorhombus (Young et al.2003), Navilithus (Young and Andruleit 2006) and
BOWN ET AL . : PALAEOGENE LOWER PHOTIC ZONE NANNOPLANKTON 459

Solisphaera (Bollmann et al. 2006) suggest that this assem-blage may be more heterogeneous and diverse than haspreviously been assumed. With the exception of Florisphae-ra, all of these taxa produce small and ⁄or fragile coccolithsthat have low fossilization potential.
Florisphaera and Gladiolithus are obligate LPZ dwellersand both have highly modified coccoliths and cocco-spheres, but differ significantly in morphology and crystal-lography. Florisphaera produces just one type of nannolith(i.e. a noncoccolith-like lith), which are small, subrectan-gular plates that form globe-artichoke-shaped cocco-spheres (Pl. 2, fig. 7). Gladiolithus produces bowl-shapedcoccospheres formed from two types of coccoliths: simple,flat basal coccoliths (lepidoliths) and long, six sided, gentlyflaring tube-shaped coccoliths (tube-coccoliths) (Jordanand Chamberlain 1992; Hagino and Okada 1998; Younget al. 2003; Plate 1). The entire Florisphaera lith is a singlecalcite crystal with the c-axis lying parallel to the long axis.In contrast, the Gladiolithus tube-coccoliths comprise sixelements each of which is a separate crystal-unit with thecalcite c-axis orientated sub-parallel to the long axis (basedon light microscope observations). These differences incoccolith structure make it unlikely that Florisphaera andGladiolithus are closely related and hence the rather similarcoccosphere morphology is likely to be a result of func-tional convergence. An attractive possibility is that thelarge bowl-shaped coccospheres surrounding a small cellmay act as light concentrating devices, with the cocco-sphere refracting light onto the chloroplasts (Young 1994).If this is the case, it is likely that these taxa have notgreatly changed their ecological preferences through time.
Of these extant LPZ taxa, only Florisphaera has a well-documented fossil record, although it was largely over-looked until the work of Molfino and Mcintyre (1990)established its use as a proxy for photic zone environ-ments in late Quaternary deep-sea sediments. They
suggested that the abundance of Florisphaera increases rel-ative to other coccolithophores when the nutricline is deepand overlain by a nutrient-depleted upper photic zone.These conditions are associated with stable, stratified, oli-gotrophic surface waters, but disrupted by increased windstress and ⁄or upwelling and divergence. The abundance ofFlorisphaera can therefore be used as an extremely effectiveproxy for surface water environments and primary pro-ductivity and it is commonly strongly correlated with Mil-ankovitch (glacial-interglacial) frequency variation inother geochemical (e.g. planktonic d18O) and palaeonto-logical indices (e.g. planktonic foraminifera) (Jordan et al.1996; Beaufort et al. 1997, 2001; Gibbs et al. 2004).Indeed, the relative abundance of Florisphaera has provento be a much more robust indicator of surface-water oli-gotrophy than the abundance of surface water oligotrophicspecies, such as Umbellosphaera and Discosphaera.
The fossil record of Florisphaera is established back tothe Upper Miocene (c. 7 Ma) with questionable occur-rences in the Middle Miocene (Young 1998), although itsprecise origination time has been questioned because ofthe possible preservation biases introduced in older sedi-ments. In contrast, Gladiolithus is rarely documented inthe fossil record and never in sediments older than lateQuaternary (Okada and Matsuoka 1996; Lars Legge et al.2006). This reflects the fragility of Gladiolithus, whichrapidly disaggregates into constituent tube-coccoliths andlepidoliths postmortem and either dissolve while sinkingthrough the water column or, soon after, at the sedi-ment ⁄water interface. Hence, they may not be preservedin sea-floor sediments and have little chance of survivinginto the fossil record (Roth and Berger 1975). Moreover,the tube-coccoliths are delicate and liable to disintegrateinto the separate elongate elements that are inconspicuousin the LM. There is also the possibility that Gladiolithushas been overlooked in the fossil record because of its
EXPLANATION OF PLATE 1
Modern and Palaeogene Gladiolithus.
Fig. 1. Modern Gladiolithus coccosphere in apical view showing the bowl-shaped morphology with the outer layer of tube-coccoliths
fallen outward (AMT16 Station, North Atlantic).
Fig. 2. Modern Gladiolithus coccosphere in antapical view showing the layer of lepidoliths covering the antapical surface (HOT169
Station, Hawaii, central Pacific).
Fig. 3. Detail of antapical surface of a modern Gladiolithus coccosphere showing bases of the tube-coccoliths and lepidoliths (HOT169
Station, Hawaii, central Pacific).
Fig. 4. Tube-coccoliths from a collapsed modern Gladiolithus coccosphere. The lower two coccoliths show the outer convex pitted
surface, the others show the inner concave smooth surface (HOT169 Station, Hawaii, central Pacific).
Fig. 5. Upper Eocene Gladiolithus collapsed coccosphere showing cluster of tube-coccoliths and scattered lepidoliths on the upper right
(Sample TDP12 ⁄ 26-2, 62 cm, Zone NP19 ⁄ 20, Kilwa Group, Tanzania).
Fig. 6. Upper Paleocene Gladiolithus collapsed coccosphere with tube-coccoliths and scattered lepidoliths on the upper right (Sample
TDP16b ⁄ 12-2, 9 cm, Zone NP9, Kilwa Group, Tanzania).
Scale bars are 2 lm.
460 PALAEONTOLOGY, VOLUME 52

1
3
2
4
5 6
PLATE 1
BOWN et al., Gladiolithus

similarity to inorganic diagenetic calcite and ⁄or elongateclay minerals. However, in modern ocean sediments,Gladiolithus liths can be abundant and when present insuch numbers in fossil material, their identificationshould be relatively straightforward. The other principlerepresentatives of the modern LPZ assemblage, i.e. Al-girosphaera, Solisphaera and Syracosphaera, also producesmall and ⁄or fragile coccoliths that do not readily exportfrom the photic zone and which have rarely or never beenrecorded as fossils (Young et al. 2003; Young and Andru-leit 2006; Bollmann et al. 2006).
ORIGINATION OF LOWER PHOTICZONE NANNOPLANKTON
The Kilwa Group fossil record provides important timeconstraints on the emergence of the modern LPZ nanno-plankton. Florisphaera has a reasonably good Neogene fos-sil record because of its relatively robust liths that arelarger and thicker than those of Gladiolithus, and can con-sistently be found in reasonably well-preserved Late Mio-cene to Recent deep-sea sediments. However, Florisphaerahas not been observed in the Kilwa Group samples wherepreservation is exceptionally good and Gladiolithus isabundant. We therefore consider this to be a significantabsence, indicating it was a later addition to the LPZassemblage, appearing between the late Oligocene andmiddle Miocene, and suggesting that its documented fossilrecord is a reliable one (Young 1998). In turn, we have notobserved Gladiolithus in the upper Cretaceous of the KilwaGroup. Nannofossil preservation is good in the upper Cre-taceous (Lees 2007), but not as exceptional as the Palaeo-gene record and so, the absence of Gladiolithus may reflect
either a preservational effect or a genuine absence, inwhich case, its origination must have occurred within thePaleocene (minimum age 58 Ma). The Kilwa Group recordof Solisphaera and Algirosphaera is sporadic and rare, butthe former is present from the middle Eocene and the lat-ter from the late Eocene. The presence of Algirosphaeraand Solisphaera, alongside several varieties of Gladiolithus,suggests that a relatively diverse, taxonomically disparateand modern-looking LPZ assemblage (but without Flori-sphaera) was established by at least the Eocene.
LOWER PHOTIC ZONENANNOPLANKTON ASPALAEOENVIRONMENTALINDICATORS
The modified coccospheres of Gladiolithus are thought toreflect adaptation to life in the LPZ environment and sowe assume that it has maintained a similar ecologicalstrategy since the Palaeogene. The presence of abundantGladiolithus in the Kilwa Group therefore strongly sug-gests that this area was overlain by oligotrophic open-ocean water masses throughout this time interval. This isinformative because there has been debate surroundingthe surface-water conditions represented by the KilwaGroup sediments and, in particular, the possible effects ofvariable salinity on the stable isotope records that havebeen used to infer palaeoclimate trends (Zachos et al.2002; Pearson et al. 2002, 2007). Gladiolithus providescompelling evidence that the surface-waters were strictlyopen-ocean and, stratified, at least seasonally if not yearround. Furthermore, a LPZ community can only developbeneath totally clear water indicating the environment
EXPLANATION OF PLATE 2
Palaeogene Gladiolithus and modern Florisphaera.
Fig. 1. Holotype of Upper Paleocene Gladiolithus ornatus. Collapsed coccosphere (Zone NP9, Sample TDP16b ⁄ 12-2, 9 cm, Kilwa
Group, Tanzania).
Fig. 2. Upper Eocene Gladiolithus brevis (Sample TDP12 ⁄ 26-2, 62 cm, Zone NP19 ⁄ 20, Kilwa Group, Tanzania).
Fig. 3. Holotype of Upper Eocene Gladiolithus contus. Collapsed coccosphere (Sample TDP12 ⁄ 26-2, 62 cm, Zone NP19 ⁄ 20, KilwaGroup, Tanzania).
Fig. 4. Collapsed coccosphere of Upper Eocene Gladiolithus contus (Sample TDP12 ⁄ 26-2, 62 cm, Zone NP19 ⁄ 20, Kilwa Group,
Tanzania).
Fig. 5. Holotype of Upper Eocene Gladiolithus brevis. Collapsed coccosphere (Sample TDP12 ⁄ 26-2, 62 cm, Zone NP19 ⁄ 20, KilwaGroup, Tanzania).
Fig. 6 Upper Eocene Gladiolithus brevis (Sample TDP12 ⁄ 26-2, 62 cm, Zone NP19 ⁄ 20, Kilwa Group, Tanzania).
Fig. 7. Modern coccosphere of Florisphaera profunda var. striatus (MATER Station 69, Alboran Sea).
Figs 8–11. Light micrographs of Palaeogene Gladiolithus tube-coccolith elements. Dark background images are in cross-polarized light
and light background images in phase contrast light (figs 8–10, TDP16B ⁄ 12-1, 10 cm, Zone NP9; fig. 11, TDP12 ⁄ 26-2, 62 cm,
Zone NP19 ⁄ 20. Kilwa Group, Tanzania).
Scale bars are 1 lm.
462 PALAEONTOLOGY, VOLUME 52
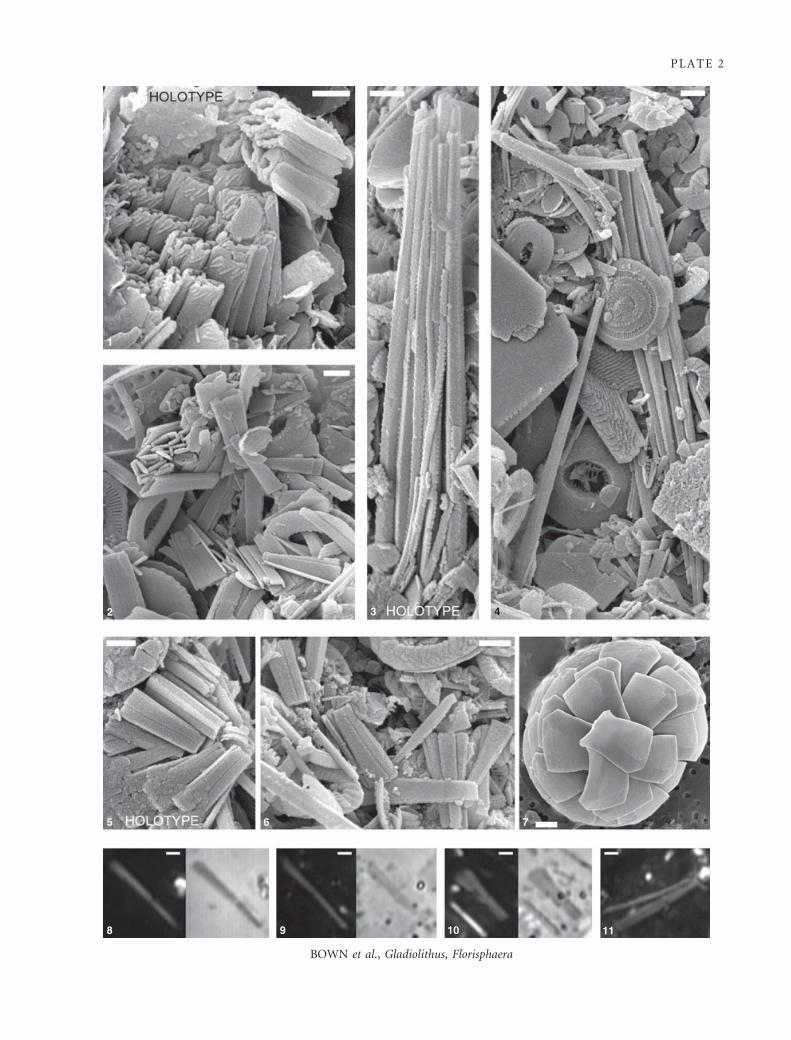
PLATE 2
BOWN et al., Gladiolithus, Florisphaera
1
2 3 4
5 6 7
8 9 10 11

was free of suspended sediment and well above the bot-tom nepheloid layer. Such a situation require depths ofat least the 300–500 m that has been inferred from theKilwa Group benthic micropalaeontological data (Pearsonet al. 2004, 2006; Nicholas et al. 2006).
To take the Gladiolithus palaeoenvironmental proxyfurther, however, we would need to generate high-resolu-tion quantitative abundance records in much the sameway as has been developed for Florisphaera in Neogeneand Quaternary research. Several late Quaternary studieshave already documented co-varying Gladiolithus andFlorisphaera abundance trends (Okada and Matsuoka1996; Lars Legge et al. 2006), but the application of Glad-iolithus in older sediments will not be straightforwardbecause of difficulties related to its preservation and iden-tification. First, Gladiolithus coccoliths routinely disaggre-gate into the constituent elements, which are small, lowbirefringence objects and inconspicuous in light micros-copy (see Pl. 2, figs 8–11). Second, these elements arerather nonunique shapes even at high magnifications inSEM and may be difficult to identify unequivocally, par-ticularly at low abundances. Finally, the presence of thesedelicate coccoliths will be dependent upon, or at the veryleast, significantly modified by, subtle variations in preser-vation and they may in fact be restricted to well preservedand young sediments (i.e. Quaternary) or older sedimentswith exceptional preservation. We have examined severallow-latitude Palaeogene sections with good nannofossilpreservation (St. Stevens Quarry, Alabama; DSDP Site242, Indian Ocean and Nanggulan, Java), but are yet tofind any evidence of Gladiolithus in SEM. Similarly, otherLPZ coccoliths such as Algirosphaera and Solisphaera arealso dissolution-prone taxa and the same preservationsensitivity will apply to their fossil records.
The presence of LPZ taxa in the Palaeogene signifi-cantly extends the stratigraphic record of the modern
assemblage and suggests that the taxonomic compositionof this specialized group may have been relatively stablethrough much of the Cenozoic. Prior to this study, therewas no unequivocal fossil record of pre-Neogene LPZnannoplankton, although following the success of Flori-sphaera as a paleoceanographic proxy in Pliocene andQuaternary studies, there has been speculation over possi-ble LPZ analogues within extinct lineages in the oldernannofossil record. Mesozoic palaeoceanographers haveproposed a number of candidates, including Schizosphae-rella in the Jurassic (Erba 2004) and Nannoconus in theCretaceous (Erba 1994; Herrle 2003; Watkins et al. 2005).These interpretations are primarily based on inverseabundance relationships with other coccoliths, a directanalogy with the younger Florisphaera studies. However,morphological considerations are also discussed with thesuggestion that these heavy liths may be LPZ adaptive(Erba 2004). The presence of heavy liths or coccospheres,however, is not a diagnostic feature of the modern LPZtaxa and also by analogy with modern distributions, theshallow-water Mesozoic epicontinental seaways in whichSchizosphaerella and Nannoconus flourished are not obvi-ous settings for the LPZ habitat. The inverse abundancerelationships associated with these large and robust taxaare also difficult to disentangle from the possible effectsof selective dissolution. We therefore await stronger evi-dence that might establish these or other taxa as MesozoicLPZ phytoplankton that are directly comparable to theCenozoic and modern examples. An alternative approachto this question is to use stable isotopes from size-sepa-rated nannofossil fractions, as demonstrated by Minolettiet al. (2001) to test the hypothesis of Aubry (1992) thatDiscoaster was a LPZ taxon. They analysed a Plioceneooze sample and were able to show that discoasters hadoxygen and carbon isotope signatures characteristic ofsurface waters rather than the LPZ.
EXPLANATION OF PLATE 3
Modern and Palaeogene Algirosphaera and Solisphaera.
Figs 1–5. Upper Eocene Algirosphaera fabaceus coccoliths from a collapsed coccosphere (Sample TDP12 ⁄ 26-2, 62 cm, Zone NP19 ⁄ 20,Kilwa Group, Tanzania).
Fig. 6. Light micrographs of Lower Oligocene Algirosphaera fabaceus coccolith – same specimen in cross polarized and phase contrast
illumination (Sample TDP11 ⁄ 15-1, 24 cm, Zone NP21, Kilwa Group, Tanzania).
Fig. 7. Modern Algirosphaera robusta coccolith (Algobank Culture Collection).
Fig. 8. Modern Algirosphaera robusta coccosphere (MATER Station 69, Alboran Sea).
Fig. 9. Modern Solisphaera blagnacensis coccosphere. Inset is an enlarged coccolith showing the blade-like distal process (MATER
Station 69, Alboran Sea).
Fig. 10. Middle Eocene sp. cf. Solisphaera (TDP13 ⁄ 20-1, 50 cm, Nannofossil Subzone NP15b, Kilwa Group, Tanzania).
Fig. 11. Middle Eocene sp. cf. Solisphaera (TDP13 ⁄ 20-1, 50 cm, Nannofossil Subzone NP15b, Kilwa Group, Tanzania).
Fig. 12. Middle Eocene sp. cf. Solisphaera (Nannofossil Subzones NP14b-15a, TDP20 ⁄ 32-1, 40 cm, Kilwa Group, Tanzania). Fig. 6. Upper
Eocene Algirosphaera sp. coccoliths (Sample TDP12 ⁄ 26-2, 62 cm, Zone NP19 ⁄ 20, Kilwa Group, Tanzania).Scale bars are 1 lm.
464 PALAEONTOLOGY, VOLUME 52

PLATE 3
BOWN et al., Algirosphaera, Solisphaera
1 2 3
4 5 6 7
8 9
10 11 12 13

SYSTEMATIC PALAEONTOLOGY
This section describes the Palaeogene LPZ taxa from the KilwaGroup of southern Tanzania. Smear slides, SEM stub-mountsand electronic images prepared for this study are stored in theDepartment of Earth Sciences, University College London, UK.The morphometric and occurrence data are based on our KilwaGroup observations. Systematic references not included in thereference section can be found in Jordan et al. (2004).
Division HAPTOPHYTA Hibberd, 1972 ex Edvardsen and
Eikrem in Edvardsen et al., 2000
Class PRYMNESIOPHYCEAE Hibberd, 1976 emend. Cavalier-
Smith in Cavalier-Smith et al., 1996
Order SYRACOSPHAERALES Hay, 1977 emend. Young et al.,
2003
Family RHABDOSPHAERACEAE Haeckel, 1894
Genus ALGIROSPHAERA Schlauder, 1945 emend. Norris,
1984
Algirosphaera fabaceus (Bown and Dunkley Jones, 2006)
comb. nov.
Plate 3, figures 1–6
Basionym. Holodiscolithus fabaceus Bown and Dunkley Jones,2006, p. 20, pl. 3, fig. 31, Journal of Nannoplankton Research, 28,17–34.
Description. Rhabdoliths with basal coccoliths comprising a rim,radial cycle and lamellar cycle from which a domed, double-lipped hood extends. The hood is parallel-sided for around halfits height, then broadens distally to dimensions similar to thatof the coccolith base and is topped by a gently curving uppersurface. The hood is formed from a three-dimensional complexof laths and in the upper part is partially covered in thin, squareand rectangular plates (Pl. 3, figs 1–4). In side view, the constit-uent laths show at least four different orientations, includingperpendicular and oblique to the coccolith base (Pl. 3, fig. 4),but it is unclear how these interconnect. Discrete sets of lathsoccur on either side of the hood, giving rise to the double-lippedappearance on the upper surface and in the light microscope(Pl. 3, fig. 6).
Dimensions. Length: 4.2 lm, width 2.7 lm, height 2.8 lm.
Remarks. The coccolith base is comparable to otherrhabdoliths, including both Palaeogene genera such asBlackites and extant forms such as Acanthoica. The hood,however, is highly distinctive and closely resembles thatof the modern coccoliths of Algirosphaera robusta (Pl. 3,figs 7–8), as described by Probert et al. (2007). Obvioussimilarities are the double-lipped overall form and theinternal construction from a mass of rod- or lath-shaped
elements. However, in A. robusta, the entire hood iscovered by a thin veneer of cover plates, whilst inA. fabaceus, these only occur around the top of thehood. Also the hood of A. fabaceus appears to be con-structed of broader, lath-like, elements than the hood ofA. robusta.
The basal coccolith and hood construction of these Pal-aeogene specimens is directly comparable to modernspecimens of Algirosphaera and supports their classifica-tion within this extant genus. Algirosphaera has beenobserved rarely in late Quaternary sediments (Okada andMatsuoka 1996) and its presence in the Kilwa Grouppushes back its origination time by over 30 my. AlthoughAlgirosphaera robusta is a characteristic taxon of the LPZ,it is not obligate, but can occur in surface waters, espe-cially at high latitudes.
Occurrence. Upper Eocene – Lower Oligocene (Nannofossil zonesNP19 ⁄ 20 ⁄ 21), TDP site 11 and 12, Pande, southern Tanzania.
Algirosphaera? sp.
Plate 3, figure 13
Description. Small rhabdolith coccoliths with hoods covered byimbricating laths. The internal structure of the hood has notbeen observed and so we cannot confirm whether it has thecomplex structure that is characteristic of the extant genus. Thegeneral appearance, however, is strongly reminiscent of Algiro-sphaera coccoliths.
Dimensions. Length: 1.5 lm, height 2.0 lm.
Occurrence. Upper Eocene (Nannofossil Zone NP19 ⁄ 20), TDPSite 12, Pande, southern Tanzania.
INCERTAE SEDIS aff. family RHABDOSPHAERACEAE Haeckel,
1894
Genus SOLISPHAERA Bollmann et al., 2006
Remarks. This extant genus is characterized by polymor-phic coccospheres that comprise simple coccoliths andcoccoliths with longitudinal blade-like processes (Pl. 3,fig. 9). These processes are constructed from small(< 0.5 lm) rhombic elements arranged in an imbricatepattern. We have observed disarticulated processes as fos-sils, with comparable shape and ultrastructure, but theyare slightly larger than the extant species and the constit-uent elements are more elongate (Pl. 3, figs 10–12). Forreasons of these differences and the lack of basal cocco-liths, our assignment to the extant genus is currentlyinformal.
466 PALAEONTOLOGY, VOLUME 52

Solisphaera sp.
Plate 3, figures 10–12
Description. Flat, trapezoid processes constructed from small,overlapping, rhombic elements. In the fossil form, the elementsslope from upper left to lower right, whilst in the living forms,they slope from lower left to upper right. This gives the impres-sion of a major difference in orientation, but may be simply aproduct of different element shape (i.e. elongation) with nochange in crystallographic orientation. The narrow end of theprocess has a row of elements with different orientation, whichmay represent where the process joined the basal coccolith. Thiscycle is not so conspicuous in published images of extant speci-mens. The fossil specimens are comparable to the living speciesSolisphaera emidasia and S. blagnacensis (Pl. 3, fig. 9).
Dimensions. Length: 3.2–4.2 lm, width 3.3–3.8 lm.
Occurrence. Middle Eocene (Nannofossil subzones NP14a–15b),TDP sites 13 (Pande) and 20 (Kilwa), southern Tanzania.
NANNOLITHS INCERTAE sedis
Genus GLADIOLITHUS Jordan and Chamberlain, 1993
Type species. Gladiolithus flabellatus (Halldal and Markali, 1955)Jordan and Chamberlain, 1993 from a plankton sample, NorthAtlantic Ocean (Gulf Stream, 38!N, 70!W).
Remarks. In the modern ocean, Gladiolithus flabellatus isa common obligate LPZ dweller. A second rare species,G. striatus, resembles G. flabellatus in overall morphology,but has striking external ornamentation and rectangularlepidoliths. Here we describe three new Palaeogene spe-cies, G. ornatus, G. contus and G. brevis, that also resem-ble G. flabellatus but which have distinct tube-coccolithlength and in one case ornamentation. Although we haveobserved abundant G. flabellatus lepidoliths, we have notbeen able to determine whether distinct lepidolith mor-phologies are associated with these new taxa.
Gladiolithus flabellatus (Halldal and Markali, 1955) Jordan
and Chamberlain, 1993
Plate 1, figures 1–6; Plate 2, figures 8–11
Description. The following description is primarily sourced fromobservations of modern coccospheres, but the coccolith andinferred coccosphere morphologies and dimensions of most ofthe Palaeogene specimens do not differ significantly from themodern specimens of G. flabellatus (compare Pl. 1, figs 1 and 5).The base of the coccosphere is formed of small (<2 lm), simple,flat coccoliths, termed lepidoliths (Pl. 1, fig. 2), which consist oftwo elements joined along a median suture. The sides of thecoccosphere are formed of long (5–8 lm) tube-shaped coccoliths
(length ⁄width ratio 4.2–7.0) with a closed base and open apex(Pl. 1, fig. 4). There are often several layers of these tube-cocco-liths and the outer layers may collapse to produce radiatingarrangements of coccoliths (Pl. 1, fig. 1), but in well-preservedspecimens, all tube-coccoliths are arranged parallel to the cocco-sphere surface (Pl. 1, fig. 2). Each tube-coccolith is formed of sixelongate elements and has a flattened hexagonal cross-section,with the elements parallel to the coccosphere surface beinglonger than those forming the sides of the tube. Two, minuteelements form the base of the tube (Pl. 2, fig. 3). The elongateelements gently taper and are narrowest at the basal end wherethey terminate bluntly and widest at the top end where theyhave triangular terminations. On modern coccospheres, thetube-coccolith elements that form the outer coccosphere surfaceare pitted, whereas those facing inwards are smoother (Pl. 1,fig. 4). The long edge contact surfaces are often serrated and twoof the elements tend to be broader than the rest.
Dimensions. Palaeogene specimens. Tube-coccolith length 4.5–9.3 lm, maximum width 2.5 lm; lepidoliths length 1.3–1.8 lm,width 0.8–0.9 lm. There is some evidence that the Paleocenespecimens are consistently smaller than those of the later Eocene.
Occurrence. Upper Paleocene – Lower Oligocene (Nannofossilzones NP6–NP23), all Palaeogene TDP sites.
Gladiolithus ornatus sp. nov.
Plate 2, figure 1
Derivation of name. Latin ornatus, meaning ornament, referringto the external striae on the tube-coccoliths.
Holotype. Plate 2, figure 1 Collapsed coccosphere.
Type locality and horizon. TDP Site 16B, Pande, southern Tanza-nia, Sample TDP16B ⁄ 12–2, 9 cm, Upper Paleocene (NannofossilZone NP9).
Diagnosis. Gladiolithus with short tube-coccoliths(length ⁄width ratio c. 3.6) that have a zone of obliqueexternal striate ornament at the base.
Dimensions. Tube-coccolith length 1.7–2.7 lm; width 0.2–0.9 lm.
Description. A single articulated coccosphere (Pl. 2, fig. 1) hasbeen observed comprising 26 tube-coccoliths and several basalelements. The complete coccosphere was at least 6.5 lm across.The tube-coccoliths are weakly flaring with blunt distal termina-tion and are closed at the base by simple plates. The striae arearound 0.4 lm in length and orientated at 40–50! to the base ofthe tube-coccolith. The extant Gladiolithus striatus also hasexternal striate ornamentation, but it is transversely orientated,perpendicular to the long axis of the elongate elements and pres-ent over the entire external surface of tube-coccoliths and lepi-doliths (Hagino and Okada 1998).
BOWN ET AL . : PALAEOGENE LOWER PHOTIC ZONE NANNOPLANKTON 467

Occurrence. Upper Paleocene (Nannofossil Zone NP9), SiteTDP16B, Pande, southern Tanzania.
Gladiolithus brevis sp. nov.
Plate 2, figures 2, 5–6
Derivation of name. Latin brevis, meaning short, referring to thesquat tube-coccoliths.
Holotype. Plate 2, figure 5. Collapsed coccosphere.
Type locality and horizon. TDP Site 12, Pande, southern Tanza-nia, Sample TDP12 ⁄ 26-2, 62 cm, Upper Eocene (NannofossilZone NP19 ⁄ 20).
Diagnosis. Gladiolithus with short, unornamented tube-coccoliths (length ⁄width ratio 2.0–4.0) that have bluntdistal terminations.
Dimensions. Tube-coccolith length 2.1–3.0 lm, maximum width0.8–1.2 lm.
Occurrence. Middle – Upper Eocene (Nannofossil zones NP15–19 ⁄ 20), TDP Site 12, Pande, southern Tanzania.
Gladiolithus contus sp. nov.
Plate 2, figures 3–4
Derivation of name. Latin contus, meaning long spear, referringto the shape of the long, narrow tube-coccoliths.
Holotype. Plate 2, figure 3 Collapsed coccosphere.
Type locality and horizon. TDP Site 12, Pande, southern Tanza-nia, Sample TDP12 ⁄ 26-2, 62 cm, Upper Eocene (NannofossilZone NP19 ⁄ 20).
Diagnosis. Gladiolithus with very long, narrow tube-coc-coliths (length ⁄width ratio 24.0–37.0).
Description. The tube-coccoliths are elongate, narrow tubes thatappear to taper at both ends. The edges of the elements displayprominent serrations.
Dimensions. Tube-coccolith length 14–20 lm, maximum width0.6–0.7 lm.
Occurrence. Upper Paleocene – Upper Eocene (Nannofossilzones NP9–19 ⁄ 20), TDP sites 12 and 14, Pande, southern Tan-zania.
Acknowledgements. Thanks to the Tanzania Drilling Project teamand also the Natural Environment Research Council, TanzaniaPetroleum Development Corporation, Tanzania Commission for
Science and Technology and UCL Graduate School that pro-vided resources and funding.
REFERENCES
AUBRY, M.-P. 1992. Late paleogene calcareous nannoplanktonevolution: a tale of climatic deterioration. 272–309. InPROTHERO, D. R. and BERGGREN, W. A. (eds). Eocene– oligocene climatic and biotic evolution. Princeton UniversityPress, 568 pp.
BEAUFORT, L., LANCELOT, Y., CAMBERLIN, P.,CAYRE, O., VINCENT, E., BASSINOT, F. and LABEY-RIE , L. 1997. Insolation cycles as a major control of Equato-rial Indian Ocean primary production. Science, 278, 1451–1454.
—— DE GARIDEL-THORON, T., MIX, A. C. andPISIAS , N. G. 2001. ENSO-like forcing on oceanic primaryproduction during the Late Pleistocene. Science, 293, 2440–2444.
BOLLMANN, J., CORTES , M. Y., KLEI JNE, A.,OSTERGAARD, J. B. and YOUNG, J. R. 2006. Solisphaeragen. nov. (Prymnesiophyceae), a new coccolithophore genusfrom the lower photic zone. Phycologia, 45, 465–477.
BOWN, P. R. 2005. Palaeogene calcareous nannofossils fromthe Kilwa and Lindi areas of coastal Tanzania (Tanzania Dril-ling Project 2003–4). Journal of Nannoplankton Research, 27,21–95.
—— and YOUNG, J. R. 1998. Techniques. 16–28. In BOWN,P. R. (ed.). Calcareous nannofossil biostratigraphy. KluwerAcademic, London, 315 pp.
—— and DUNKLEY JONES, T. 2006. New Paleogene calcar-eous nannofossil taxa from coastal Tanzania: Tanzania DrillingProject Sites 11 to 14. Journal of Nannoplankton Research, 28,17–34.
—— —— LEES, J. A., PEARSON, P. N., YOUNG, J. R.,RANDELL, R., COXALL, H. K., MIZZI , J., NICHO-LAS , C. J., KAREGA, A., S INGANO, J. and WADE, B. S.2008. A Paleogene calcareous microfossil Konservat-Lagerstattefrom the Kilwa Group of coastal Tanzania. Geological Societyof America Bulletin, 120, 3–12.
ERBA, E. 1994. Nannofossils and superplumes: the early Aptian‘nannoconid crisis’. Paleoceanography, 9, 483–501.
——2004. Calcareous nannofossils and Mesozoic oceanic anoxicevents. Marine Micropaleontology, 52, 86–106.
GIBBS , S., SHACKLETON, N. and YOUNG, J. R. 2004.Orbitally forced climate signals in mid-Pliocene nannofossilassemblages. Marine Micropaleontology, 51, 39–56.
HAGINO, K. and OKADA, H. 1998. Gladiolithus striatus sp.nov. (Prymnesiophyceae), a living coccolithophore from thelower photic zone of the Pacific Ocean. Phycologia, 37, 246–250.
—— —— and MATSUOKA, H. 2000. Spatial dynamics ofcoccolithophore assemblages in the Equatorial Western-Central Pacific Ocean. Marine Micropaleontology, 39, 53–72.
HERRLE, J. O. 2003. Reconstructing nutricline dynamics ofmid-Cretaceous oceans: evidence from calcareous nannofossilsfrom the Niveau Pacquier black shale (SE France). MarineMicropaleontology, 47, 307–321.
468 PALAEONTOLOGY, VOLUME 52

JORDAN, R. W. and CHAMBERLAIN, A. H. L. 1992.Vexillarius cancellifer gen. et sp. nov. and its possible affiniteswith other living coccolithophorids. 305–325. In HAMR-SMID, B. and YOUNG, J. R. (eds), INA 1991. Nanno-plankton research, biostratigraphy and paleobiology; Quaternarycoccoliths. HND Hodonin, Hodonin, 330 pp.
—— ZHAO, M., EGLINTON, G. and WEAVER, P. P. E.1996. Coccolith and alkenone stratigraphy and palaeoceanog-raphy at an upwelling site off NW Africa (ODP 658C) duringthe last 130,000 years. 111–130. In MOGUILEVSKY, A.and WHATLEY, R. (eds). Microfossils and oceanic environ-ments. University of Wales, Aberystwyth–Press, 434 pp.
—— YOUNG, J. R. and CROS, L. 2004. A revised classifica-tion scheme for living haptophytes. Micropaleontology, 50(supplement no. 1), 55–79.
LARS LEGGE, H., MUTTERLOSE, J. and ARZ, H. W.2006. Climatic changes in the northern Red Sea during thelast 22,000 years as recorded by calcareous nannofossils. Pale-oceanography, 21, PA1003, doi: 10.1029/2005PA001142.
LEES , J. A. 2007. New and rarely reported calcareous nannofos-sils from the Late Cretaceous of coastal Tanzania: outcropsamples and Tanzania Drilling Project Sites 5, 9 and 15. Jour-nal of Nannoplankton Research, 29, 39–65.
—— BOWN, P. R. and YOUNG, J. R. 2004. Evidence forannual records of phytoplankton productivity in the Kimme-ridge Clay Formation coccolith stone bands (Upper Jurassic,Dorset, UK). Marine Micropaleontology, 52, 29–49.
MARTINI , E. 1971. Standard Tertiary and Quaternary calcare-ous nannoplankton zonation. 739–785. In FARANACCI , A.(ed.). Proceedings of the Second Planktonic Conference Roma1970. Edizioni Tecnoscienza, Rome.
MINOLETTI , F., GARDIN, S., NICOT, E., RENARD, M.and SPEZZAFERRI , S. 2001. Mise au point d’un protocoleexperimental de separation granulometrique d’assemblages denannofossiles calcaires: applications paleoecologiques et geo-chimiques. Bulletin Societa Geologie Francais, 172, 437–446.
MOLFINO, B. and MCINTYRE, A. 1990. Precesional forcingof nutricline dynamics in the equatorial Atlantic. Science, 249,766–769.
NICHOLAS, C. J., PEARSON, P., BOWN, P. R., DUNK-LEY JONES, T., HUBER, B. T., KAREGA, A., LEES , J. A.,MCMILLAN, I. K., O ’HALLORAN, A., S INGANO, J. M.and WADE, B. S. 2006. Stratigraphy and sedimentology of theUpper Cretaceous to Paleogene Kilwa Group, southern coastalTanzania. Journal of African Earth Sciences, 45, 431–466.
OKADA, H. 1983. Modern nannofossil assemblages in sedi-ments of coastal and marginal seas along the western PacificOcean. Utrecht Micropaleontological Bulletins, 30, 171–187.
——1992. Biogeographic control on modern nannofossil assem-blages in surface sediments of Ise Bay, Mikawa Bay andKumano-Nada, off coast of central Japan. Memorie di ScienzeGeologich, 43, 431–449.
—— and HONJO, S. 1973. The distribution of oceaniccoccolithophorids in the Pacific. Deep Sea Research, 20, 355–374.
—— and MATSUOKA, M. 1996. Lower-photic nannoflora asan indicator of the late Quaternary monsoonal paleo-record inthe tropical Indian Ocean. 231–245. In MOGUILEVSKY, A.
and WHATLEY, R. (eds). Microfossils and oceanic environ-ments. University of Wales, Aberystwyth–Press, 434 pp.
PEARSON, P. N., DITCHFIELD, P. W. and SHACKL-ETON, N. J. 2002. Reply – tropical temperatures in green-house episodes. Nature, 419, 899.
—— NICHOLAS , C. J., S INGANO, J., BOWN, P. R.,COXALL, H. K., VAN DONGEN, B. E., HUBER, B. T.,KAREGA, A., LEES , J. A., MSAKY, E., PANCOST,R. D., PEARSON, M. and ROBERTS, A. P. 2004.Paleogene and Cretaceous sediment cores from the Kilwa andLindi areas of coastal Tanzania: Tanzania Drilling Project Sites1 to 5. Journal of African Earth Sciences, 39, 25–62.
————SINGANO, J. M., BOWN, P. R., COXALL, H. K.,VAN DONGEN, B. E., HUBER, B. T., KAREGA, A.,LEES , J. A., MACLEOD, K., MCMILLAN, I. K., PAN-COST, R. D., PEARSON, M. and MSAKY, E. 2006. Tan-zania Drilling Project Sites 6 to 10: Further Paleogene andCretaceous sediment cores from the Kilwa area of coastal Tan-zania. Journal of African Earth Sciences, 45, 279–317.
—— VAN DONGEN, B. E., NICHOLAS, C. J., PAN-COST, R. D., SCHOUTEN, S., S INGANO, J. M. andWADE, B. 2007. Stable warm tropical climate through theEocene epoch. Geology, 35, 211–214.
PROBERT, I., FRESNEL, J., B ILLARD, C., GEISEN, M.and YOUNG, J. R. 2007. Light and electron microscopeobservation of Algirosphaera robusta (Prymnesiophyceae).Journal of Phycology, 43, 319–332.
QUINN, P. S., CORTES , M. Y. and BOLLMANN, J. 2005.Morphological variation in the deep ocean-dwelling cocco-lithophore Florisphaera profunda (Haptophyta). European Jour-nal of Phycology, 40, 125–133.
ROTH, P. H. and BERGER, W. H. 1975. Distribution anddissolution of coccoliths in the south and central Pacific.Cushman Foundation for Foraminiferal Research, Special Publi-cation, 13, 87–113.
WATKINS , D. K., COOPER, M. J. and WILSON, P. A.2005. Calcareous nannoplankton response to late Albian oce-anic anoxic event 1d in the western north Atlantic. Paleocea-nography, 20, PA2010, doi 10.1029/2004PA001097.
WINTER, A., JORDAN, R. W. and ROTH, P. 1994. Bioge-ography of living coccolithophores in ocean waters. 161–177.In WINTER, A. and SIESSER, W. G. (eds). Coccolitho-phores, Cambridge University Press, 242 pp.
YOUNG, J. R. 1994. Functions of coccoliths. 63–82. InWINTER, A. and SIESSER, W. G., (eds). Coccolithophores.Cambridge University Press, 242 pp.
——1998. Neogene. 225–265. In BOWN, P. R. (ed.). CalcareousNannofossil Biostratigraphy. Kluwer Academic, London, 315 pp.
—— and ANDRULEIT , H. 2006. Navilithus altivelum: aremarkable new genus and species of deep photic coccolitho-phores. Journal of Micropalaeontology, 25, 141–151.
—— GEISEN, M., CROS, L., KLEI JNE, A., SPRENGEL,C., PROBERT, I. and OSTERGAARD, J. 2003. A guide toextant coccolithophore taxonomy. Journal of NannoplanktonResearch Special Issue, 1, 125 pp.
ZACHOS, J. C., ARTHUR, M. A., BRALOWER, T. J. andSPERO, H. J. 2002. Tropical temperatures in greenhouse epi-sodes. Nature, 419, 897–899.
BOWN ET AL . : PALAEOGENE LOWER PHOTIC ZONE NANNOPLANKTON 469