a note on modeling mechano-chemical transduction with an application to a skin receptor
TRANSCRIPT

J. Math. Biol. (1994) 32:275-285 Journa l ai r
mathematical 61olo9y
© Springer-Verlag 1994
A note on modeling mechano-chemica! transduction with an application to a skin receptor
Jonathan Bell i, Mark H. Holmes 2 1 Department of Mathematics, SUNY at Buffalo, Buffalo, NY 14214, USA 2 Department of Mathematical Sciences, Rensselaer Polytechnic Institute, Troy, NY 12180, USA
Received 13 January 1993; received in revised form 16 July 1993
Abstract. Strain-sensitive (also called stretch-sensitive) ionic channels are thought to be present in various mechanoreceptors. The gating of these channels is precipitated by mechanical strains, as opposed to the usual activation processes of changes in membrane potential or other ligands. Below we present a class of models for the strain-activated mechanism, compare our approach to one of Sachs and Lecar (1991), and apply the gating mechanism to a model of a specific mechanoreceptor, namely a Pacinian corpuscle neurite model. The simulation experiment suggests the activation energy of the channel depends linearly, rather than nonlinearly, on the (hoop) strain in the receptor membrane.
Key words: Transduction - Mechanoreceptors - Pacinian corpuscle - Stretch- sensitive ionic channel
Introduction
Mechanoreceptors in the body range from those associated with hearing, balance, and sense of touch, to those that measure variables such as blood pressure, filling of the bladder and intestines, etc. (Sachs 1986; Morris 1990). In primary mechanore- ceptors such as the Pacinian corpuscle, the stimuli is first received and then filtered by the external structure of the receptor. When this transformed mechanical signal reaches the receptor membrane it changes the strain in the membrane. In the case of the strain-activated (SA) channels that are thought to be present in the Pacinian corpuscle, cochlea, and labyrinth, this opens ionic channels, causing a transduction current, which in turn initiates a neural response. The bridge between the mechan- ical and electrical components of the mechanoreceptor is the gating mechanism. There is both direct and indirect evidence that associated with mechanoreceptors is an ionic channel whose activation is precipitated by mechanical strains, as opposed to the usual activation processes of changes in membrane potential or concentra- tion changes of other ionic species (Guharay and Sachs 1986; Morris 1990). These

276 J. Bell, M. H. Holmes
strain-activated (or, stretch-activated) channels have been studied intensively only recently, but a large number have been identified (Sachs 1986; Morris 1990). They fall within the family of strain-sensitive (SS) channels which also includes strain- inactivated (SI) channels. Such channels have been implicated in auto-regulation of blood flow (Lansman et al. 1987), various osmoregulators and as control of cell di- vision (Methfessel et al. 1986; Yang and Sachs 1987), and even sensitivity to weak magnetic fields (Frankel 1984).
Below we present a model for the strain-activated channel. We start, like Sachs and Lecar (1991), with an assumed form for the energy of a two-state deformable channel. However, we use a different mechanical paradigm which leads to a dif- ferent model formulation. We then derive an analytical expression for the depen- dence of activation energy on strain. We use this formulation for activation en- ergy in a model for a specific mechanoreceptor, namely the Pacinian corpuscle. This will be done to help suggest an experiment that will help in determining how the channel depends on strain, which is something that has been an open question in the area (Morris 1990).
A strain-activated ion channel gating model
A number of SA ionic channels are now known and their properties vary somewhat from species to species. Generally, one or more rate of opening parameters are strain dependent, but all rates associated with channel closing appear to be independent of strain (at least for strain magnitudes within normal physiological range). Typically the channels are not very ion specific, but some are K + selective. Often times they have more than two conformational states. For simplicity of modeling, and because there is no evidence otherwise in the mechanoreceptors we are concerned with, it is assumed that the channel has only two states (one open state, one closed state), and it is stretch-activated (SA). Thus, the ionic channel responsible for the transduction current has one opening rate, ~, which is considered strain and voltage dependent, and one closing rate, /~, which is assumed to be only voltage dependent. It is also assumed that the energy of opening depends on crossing a single energy barrier. In our mathematical representation below we consider that it is sufficient to allow only a single axis of deformation, but at the membrane level the deformation must be three dimensional. One possible scenario for this deformation is given in Sachs (1986).
To deyelop the description of the channel dynamics we rely on Eyring's absolute reaction rate theory (Hille 1984) which represents the opening rate ~ as
ot = ~o T e x p ( - AE/~c T ), (1)
where ~0 is a scale constant, x is Boltzmann's constant, and T is absolute tem- perature. To use this formulation we assume that the energy of activation A E of the transducer channel is the Helmholtz free energy AA and it has two compo- nents, namely a component due to the voltage across the filopodial membrane and a component due to the strain in the receptor membrane.
One way of looking at the problem is to consider the channel as a forced nonlin- ear spring with a potential energy function ~b(x) as pictured in Fig. 1. This particular form for the energy was proposed by Sachs and Lecar (1991) for a deformable two state channel with a single axis of deformation. Sachs and Lecar describe a mech- anical paradigm for their channel model in terms of two, independent, harmonic
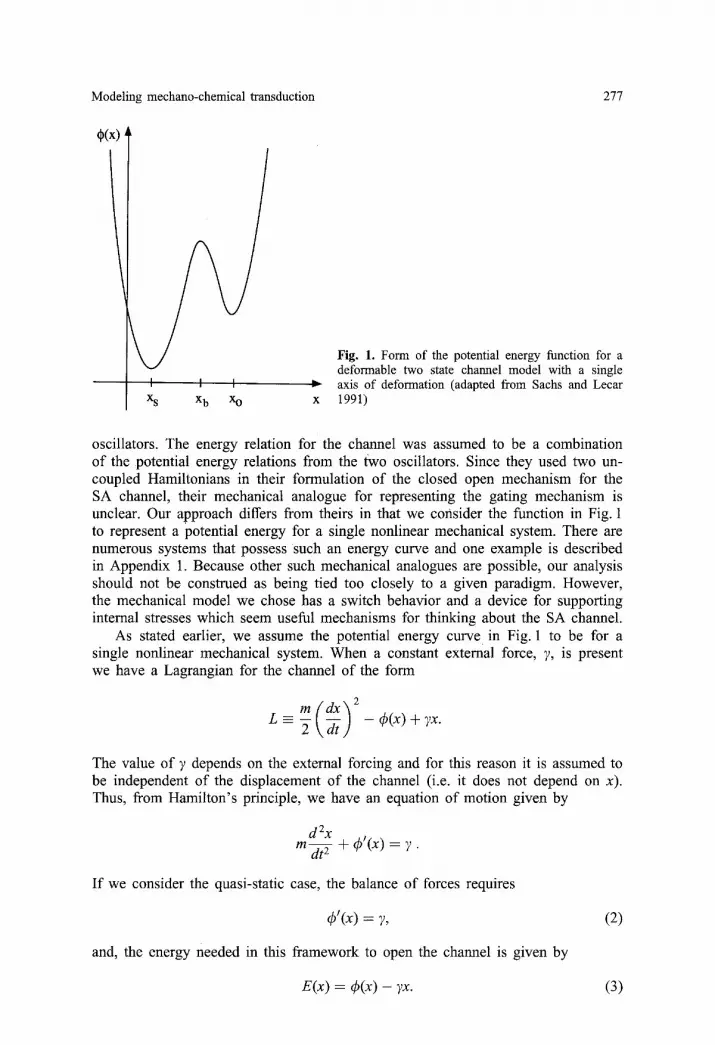
Modeling mechano-chemical transduction 277
~(x)
I I I X s X b XO
Fig. 1. Form of the potential energy function for a deformable two state channel model with a single axis of deformation (adapted from Sachs and Lecar 1991)
oscillators. The energy relation for the channel was assumed to be a combination of the potential energy relations from the two oscillators. Since they used two un- coupled Hamiltonians in their formulation of the closed open mechanism for the SA channel, their mechanical analogue for representing the gating mechanism is unclear. Our approach differs from theirs in that we consider the function in Fig. 1 to represent a potential energy for a single nonlinear mechanical system. There are numerous systems that possess such an energy curve and one example is described in Appendix 1. Because other such mechanical analogues are possible, our analysis should not be construed as being tied too closely to a given paradigm. However, the mechanical model we chose has a switch behavior and a device for supporting internal stresses which seem useful mechanisms for thinking about the SA channel.
As stated earlier, we assume the potential energy curve in Fig. 1 to be for a single nonlinear mechanical system. When a constant external force, 7, is present we have a Lagrangian for the channel of the form
m ( d x ) 2 L = ~ \ dt j - (a(X) + TX"
The value of 7 depends on the external forcing and for this reason it is assumed to be independent of the displacement of the channel (i.e. it does not depend on x). Thus, from Hamilton's principle, we have an equation of motion given by
d 2X m~t 5- + q~'(x) = 7.
I f we consider the quasi-static case, the balance of forces requires
qS'(x) = 7, (2)
and, the energy needed in this framework to open the channel is given by
E(x) = 49(x) - 7x. (3)

278 J. Bell, M. H. Holmes
To find the transition energy associated with going from the closed to the open state let xj, f o r j = s, b, be the equilibrium points as indicated in Fig. 1. Assuming a small forcing, we can expand x about each point as follows
x ~ xj +x j17 +xj2? 2 + . . . . (4)
Substituting this into (2), expanding ~b'(x) using Taylor 's theorem, and assuming the function q~; is smooth, yields
1 2 ; t i t , , 2 4)j -~- Xj14)~I ~J -}- (Xj2 4)j' -I - "~ Xjl q) j )'Y -1- . . . . ~,
where 4)~ = 4)'(xj), etc. From this it follows that 4)~ = 0 and Xjl = 1/4)j'. Inserting these values into (3) gives the following expansion for the energy
1 2 ~ 4)j-xj -
Therefore, for a small forcing, the Helmholtz free energy has the form
AA = AAns + AAstrain, (5)
where AAns is the deformation energy due to non-strain induced forces. In our case we consider the transmembrane voltage case so that AAstrain = -,~w, where w = w(t) is the voltage. Here the constant 2 is positive and is associated with the difference in dipole moments of the two channel conformations. Also,
AAstrain = Ab -- As
l ( ~ t 1 )Y 2" ~ 4 ) b - 4 ) s - ( x b - - 4 ) s - 7 7 / (6)
Note that, based on the assumed form of the energy curve in Fig. 1, 4)b > 4)s, xb > x~, and 4)~' < 0 < 4)'J. Thus the coefficients of 7 and 7 2 in (6) are negative. Also note that the above derivation can be used to find the transition energy AA c-° for going from the closed to open state. The result is
1(1 1) 2 AA c-° ~ - 2 w + 4)0 - 4)b -- (Xo -- xb)y -- ~ ~o' 4)~' " (7)
Similar expressions to (6) and (7) can be obtained even if the external forcing is an analytic function of x. To demonstrate, note that in this case the energy in (3) takes the form
E(x ) = 4 ) ( x ) - 2G(s )ds .
As before 7 is constant but now we are allowing for a dependence on x through the function G(x). In this context 7 is the magnitude of the forcing function and we can therefore take 0 < G ( x ) < 1. The expansions in (4) still apply and substituting them into the force balance equation 4)'(x) = 2G(x) yields 4)j = 0 and xjl = Gj/4)j I where Gj = G(xj). From this we get the following expansion for the energy
f0 xj 2

Modeling mechano-chemical transduction 279
u i ( x ) =
Thus, their energy functions where
Thus, in place of (6) we obtain
AAstrain ~ 4 b - - Os - - 7 G(s)ds - ~
A similar expression is found in place of (7). Another way to derive an analytical expression for AAstrain has been considered
by Sachs and Lecar (1991), who treat the SA channel as composed of two Hookian springs (with constants K o , K s ) . Using statistical mechanical arguments, and subject- ing the channel to a constant force F, their total energy is p 2 / 2 m + ui + u ° , where p is the momentum, u ° is a constant, and ui is the potential energy given by
K : ( x - - F x , i = o , s . x i ) 2 for
of the open and shut states have the form A i = A ° - q9 i
and
A ~ = ~ m + ~- x - x i - ~ i + u ° ,
F 2
(Pi = F x i + - - 2 K i "
They interpret the first term, A °, in the expression for Ai as the Hamiltonian for an unperturbed oscillator, while the second term is the contribution of the external force. From this they arrive at the following for the Helmholtz flee energy
( 1 1 ) F 2 (8) AAstrain = A°o - A°s - (Xo - x s ) F - I£o K s "
This expression is qualitatively similar to (6) if we identify F with the constant 7- The principle difference between (6) and (8), besides the differences in derivations, is that (6) represents the energy barrier in going from the closed to open state, whereas (8) is the difference between the energy levels for the closed and open states. As a consequence the 72 term in (6) is always negative, whereas the corresponding term in (8) can be both positive or negative. If it is assumed that the linear terms in these expressions are much larger than the quadratic components, an assumption which is investigated below, then this discrepancy is a second order effect and is difficult to resolve by present experimental means.
The expressions in (6) and (8) for the energy of activation differ from others that have been proposed. Guharay and Sachs (1984) only considered the energy stored in the channel and did not include the component associated with the difference between equilibrium positions. Thus, their energy function did not contain the O ( F )
term in (8). Corey and HudsPeth (1983) on the other hand obtained a linear function of F because they assumed that a single spring constant can be used for the channel (hence, K o = K s in (8)). However, they cannot account for the fact that the forward rate is strain sensitive while the reverse rate is not. A linear function was also used by Bell and Holmes (1986) in describing the channels in the auditory hair cell.
The expression for the activation energy can be simplified if it is assumed that the elastic forcing is small enough that the 0(7 2 ) term in (6) is negligible. More precisely, it is assumed that the term

280 J. Bell, M. H. Holmes
-k-/7 7 respectively qb-~b ,
is small compared to 2(xb --x~). In this case
A A = - 2 w + C~b - - (gs - - (Xb - - X s ) 7 . (9)
At this point it is unclear how to experimentally test this assumption, but such an experiment will be discussed in the next section. Also, using a similar assumption for the transition from closed to open we have, from (7), that
A A c - ° = - 2 w + (ao - q~b - (Xo - x b ) ~ . (10)
In the case of a SA channel, this result would imply that Xo and xb are relatively closely spaced (much more so than xs and Xo) .
In the application of our formulation of the Helmholtz free energy given in the next section, ~ is proportional to the hoop strain (represented by zoo), so the exponent for the rate constant ~ given in (1) is, from (5) and (6),
A A - - a l e o o - a 2 ( ~ o o ) 2 + a 3 w ( t ) - a4, (11)
xT
where the a~s are positive constants. This represents a generalization of the form derived by Bell and Holmes (1986) for the transduction channels in the auditory hair cell where it was assumed that the activation energy is a linear function of strain. As will be shown below the linearized version is most likely the dependence for the PC as well.
Applying the gating model to a receptor model of the Pacinian corpuscle
One of the more intensively studied somatosensory mechanoreceptors from a phys- iological viewpoint is the Pacinian corpuscle (PC). This interest is partly due to its size and its accessibility, and partly due to the fact that it is regarded as the prototypical non-ciliated mechanoreceptor. In somatosensory terms it is the receptor associated with the P-channel, which is one of the four somatosensory pathways in the skin (Bolanowski et al. 1988). At the present time the PCs associated with the skin are thought to be responsible for the tranduction of relatively high frequency mechanical vibrations. For details on the anatomy and physiology of PCs, see Bell et al. (1993).
One of the standard experimental protocols used to study the response char- acteristics of the PC involves a periodic forcing. It is found from these tests that the PC displays what is known as phase locking. In these experiments the fre- quency of the stimulus is fixed and the amplitude of the forcing function is ini- tially set so small that there is no spike activity from the neurite. The amplitude is then increased in small increments and the neural response is recorded. It is found that once the amplitude is large enough the neural response jumps to one spike/ cycle (see Fig. 2). The firing rate will continue at this level until the amplitude reaches another threshold, after which the rate jumps to two spikes/cycle. Thus the rate locks to a specific integer value. The location and length of these ampli- tude steps depends on the fixed stimulus frequency, the temperature, and numerous other conditions. This behaviour is also observed in other preparations, which do

Modeling mechano-chemical transduction 281
not posses a mechanical filter like the capsule (Nemoto et al. 1975; Guttman et al. 1980). Moreover, the PC shows phase locking with or without the capsule. Thus, it appears that this response characteristic is due to the neural component of the PC and not to the mechanics of the capsule. It is this experiment that will be used to investigate the dependence of the activation energy on the stimulus forcing.
Although there is no direct evidence for SA channels in PCs, the evidence is strong enough to assume that there are (Bell et al. 1993). Our model for the PC has been developed elsewhere (Holmes and Bell 1990; Bell and Holmes 1992) so the description given here is brief. In essence it consists of a mechanical capsule that is coupled through a SA channel model to a model for the nerve ending, or neurite. The activation o f the channel is from the hoop strain in the receptor membrane and this is determined using the capsule model (see (11)). However, because the capsule component is not needed here we only consider the channel and neurite portions (the model is summarized in Appendix 2). The input in this case is an imposed sinusoidal hoop strain, as it would be for a decapsulated corpusle. The model's output is a sequence of action potentials generated by the neurite, and the firing rates are compared to similar experimental results conducted on PCs.
The PC model displays phase locking and this can be used to examine the dependence of the activation of the transduction channel on the forcing (see Fig. 2). In particular, phase locking can be used to investigate whether or not the free energy of activation is a linear or quadratic function of strain. To do this two versions of the model are considered. In the first version the energy of activation is taken to be a linear function of hoop strain, so,
A A
K T - - aaeoo + a 3 w ( t ) - a 4 • (12)
For version two, a strictly quadratic dependence is used, in which case
A A
roT - - a2(eoO) 2 q- a 3 w ( t ) - - a4 . (13)
Figure 3 shows the results from computing the intensity characteristics for the two versions of the model using parameters that are typical for the PC. What is im- mediately evident from these results is that the quadratic dependence given in (13) is not capable of producing a plateau at 1 spike/cycle. It is relatively easy to find a principle contributor to this behavior. During a cycle the strain in the receptor membrane is positive for half the cycle and negative the other half. Because the hoop strain is squared in (13) there is no way to distinquish betwen positive and negative strain. Consequently the stimulus reaches a point twice in the cycle where it is large enough to produce an action potential. For the linear version, however, this occurs only once and hence 1 spike/cycle is produced. For a similar reason (13) has difficulty remaining locked to three spikes/cycle or, for that matter, any odd integer number of spikes per cycle.
From this we are lead to hypothesize that the linear term in the energy depen- dence on strain, (11), dominates the quadratic term, at least over normal physiologi- cal range of stimuli for the PC. A definitive answer will have to wait until sensitive enough experimental techniques can be devised but this does appear to provide a test which will help resolve the strain dependence on the channel.

282 J. Bell, M. H. Holmes
IO00
Z I00 tO W g l
n,, IJd O.
I,=.1 v
a. i o tJl
t~l I -
n,.,
Z
¢ 1 .0 i ,
I I ' I I I D' El
c
E N S I T Y C H A R A C T E R I S T I I C - 7 / 3 / 7 9 - 1
l 2 5 0 H z
0 . / 4 0 ~ I I l I I I I I I J 1 - 5 0 - 2 0 - t O 0 I 0 2 0
D I S P L A C E M E N T IN d B RE I/~rn P E A K 3 0
Fig. 2. Neural firing rate obtained from a Pacinian corpuscle subjected to a periodic forcing. Note the plateau in the firing rate corresponding to 1 spike/cycle. The driving frequency is 250 Hz (from Bolanowski and Zwis- locki 1984)
5 .0
m
3.0 r ~
2 .0
1.0 :r.
0 .0 0 .0
4 . 0 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . : . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . r - . - - ~ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
t I
t !
...................................... i ............. r - - " .................. i ................... " t - - - - - - - - i - i
L_ i _ ' i . . . . . . . . . . . T . . . . . . . . . . . . . . 4 - - - i
, i / ..... I[' . Linear Model ] .....
. . . . . Quadratic Model ]
T 5 .0 10 .0
N o r m a l i z e d A m p l i t u d e
15.0
Fig. 3. Firing rate as a function of ampli- tude using an imposed periodic stimulus. Shown is the response using the linear de- pendence on strain in (12) and the quadratic dependence in (13). The driving frequency is 250Hz. The amplitudes have been nor- malized by the threshold value necessary to produce a response of one spike/cycle, and the parameter values are typical of those for a PC (see Bell and Holmes 1992)
Appendix 1
The starting point in the derivation o f the dependence of the activation energy on strain is the energy curve shown in Fig. 1. It is sometimes helpful to have a simple mechanical paradigm that has the same characteristics as the system under study. There are numerous systems with energy curves like the one shown in Fig. 1 and an example is summarized in Fig. 4. It consists o f a mass that is restricted to move on a circle with radius r (one can think o f it as a rigid rod, free to rotate frictionless at its base, with a mass m at the upper end). Acting in the circumferential direction is a spring which exerts a force F1. There is a second spring rthat exerts a fo rce / ' 2 on the mass (note F2 is the component o f the restoring force o f the spring in the circumferential direction).To keep the formulation simple we will assume each spring is linear, so, the model contains a geometric rather than a material nonlinearity. In

Modeling mechano-chemical transduction 283
Fig. 4. One mechanical paradigm involving a two spring system tied to a gate which leads to a potential energy function representation as depicted in Fig. 1
this case, for the cireumferential spring we have that F1 = - k l r ( O - 00) where kl is the spring constant and 00 is the angle when the spring is unloaded. For the second spring we have F2 = - k z ( d - do ) f (O) where f (O) = ~sin(O). Here k2 is the spring constant, d is the length of the spring, E is the vertical height o f the spring (see Fig. 4), and do is the length of the spring when unloaded. It is not hard to show that
d = v / r 2 - 2r~cos(0) ÷ E ? .
Assuming, without loss of generality that r = 1, the equation of motion is
m O 't = FI (0 ) + F2(0) = - k l ( O - 0o ) - k z ( d - d o g ( O ) .
A Lagrangian for this system is
where
m(a0) 2 Ls = 2 k at / - (~(0),
(a,(O) = lk1(02 - 20o0) + l k2 (d2 - 2dod) .
It is not hard to show that q~s has the form shown in Fig. 1 for certain ranges of the parameters. For example, it is obtained if one takes, 00 = - ~ , kl = 1, k2 = 10, # = 2, and do = d(~/4).
Although a gating function must be present in a strain-activated channel, there is no physiological evidence a s to the nature of any particular gating mechanism. Consequently, it is easy to think of possible gates that have the required properties. For example, we might take the paradigm of a twisted protein structure as pictured in Fig 2(E), Chapt. 14 in (Hille 1984). In this case the imposed membrane strain untwists the protein from its rest potential (the minimum energy configuration). The channel, being a complicated structure, has one (or more charged) subunits affected mechanically by the untwisting. The relative position (and charge distribution) during this changing conformational process can determine the energy barrier between states as pictured in our Fig. 1 given that variable x represents the relative bending angle of the channel.

284 J. Bell, M. H. Holmes
Appendix 2
The model of the receptor membrane and strain-sensitive gating mechanisms relies on the classical principles in electrophysiology, the circuit model of Bolanowski and Zwislocki (1984), and the work on dendritic spine modeling by Baer and Rinzel (1990). The receptor membrane is modeled as a passive circular cable with active filopodia distributed along its outer surface. The filopodia are considered to be 'in- dividual units connected electrically to the neurite via a stem resistance, R/, but they are assumed to be disconnected from each other. The filopodia are considered as being distributed along the neurite with density N = N(x) which represents the number of filopodia per unit length. The formulation of the model allows this dis- tribution to be nonuniform along the neurite (i.e. there may be more filopodia in the cleft region), and of different morphofunctional types (i.e. directional selectivity). The number and distribution of filopodia in the model is given by their number density per characteristic length, n(x) = AN(x). From current balance considerations
~t) 2 ~2v W -- V 1 Ow w - v (A.2)
+ I - RI ,
where v(x, t) and w(x, t) represent the membrane potential at time t in the neurite and filopodial head (at location x), respectively. Equation (A.1) is the standard cable equation for a passive dendritic cable, with time constant z and length constant 2, with the last term representing an applied current source (Rall 1976). This source is the contribution of adjacent filopodia with density N = n/2, and input resistance 2R0. The parameters z, 2,Ru are parameters are determined from the anatomy or physiology using the following formulas
: R m f m ; ,~2 : Rmd /4R i ; Rf = 4Rilf/7~d~; Ro : Rm/Tz,~d,
w h e r e Rm is the membrane resistance, R i is the cytoplasmic resistivity, and Cm is the dendritic membrane capacitance. The filopodium membrane capacitance, CU, is calculated from xAf/dfm, where Z is the permittivity, and Af, dim are the averaged filopodial membrane surfacearea and membrane thickness, respectively. The other parameters are diameter of the neurite, d, length of the filopodium, lf, and diameter of filopodium, df. The current voltage relation I in (A.2) is
I = g r m 3 h ( w - v r ) + gAn4(w - VA) + gL(W -- VL), (A.3)
where vr, vA, vL are, respectively, the transducer, non-transducer, and leakage reversal potentials. Also gr, gA, gL are maximal conductance parameters for the transducer, non-transducer, and leakage currents, respectively. For lack of specific experimental information, values for vr and vA are taken from the Hodgkin-Huxley (HH) theory. Similarly, gr, gA, gL are versions of the HH conductances scaled so that the neurite model supports 1000 Hz spiking behavior. The leakage potential VL is calculated so that I = 0 at rest potential. The variables m,h,n are, respectively, activation and inactivation variables for the transducer channel, and the activation variable for the non-transducer "A" type channel. These satisfy first order kinetic equations (see Bell and Holmes 1992).

Modeling mechano-chemical transduction 285
Acknowledgement. We wish to thank Dr. Fred Sachs for consultation on various aspects of SA channel experiments, including the energy approach presented here. The work was partially sup- ported by NSF grant DM8801502 (J.B) and the Whitaker Fotmdation (M.H.H.).
References
Baer, S. M., Rinzel, J.: Propagation of dendritic spikes mediated by excitable spines: a continuum theory. J. Neurophys. 65, 874-890 (1991)
Bell, J., Holmes, M. H.: A nonlinear model for transduction in hair cells. Hearing Res. 21, 97-108 (1986)
Bell, J., Holmes, M. H.: Model of the dynamics of receptor potential in a mechanoreceptor. Math. Biosci. 110, 139-174 (1992)
Bell, J., Bolanowski, S. J., Holmes, M. H.: The structure and function of Pacinian corpuscles: a review. Prog Neurobiol. (in press)
Bolanowski, S. J., Gescheider, G. A., Verrillo, R. T., Checkosky, C. M.: Four channels mediate the mechanical aspects of touch. J. Acoust. Soc. Am. 84, 1680-1694 (1988)
Bolanowski, S. J., Zwisloeki, J. J.: Intensity and frequency characteristics of Pacinian corpuscles. I. Action potentials. J. Neurophysiol. 51, 793-811 (1984)
Corey, D. R., Hudspeth, A. J.: Kinetics of the receptor current in bullfrog saccular hair cells. J. Neurosci. 3, 962-976 (1983)
Frankel, R. B.: Magnetic guidance of organisms. Ann. Rev. Biophys. Bioeng. 13, 85-95(1984) Guharay, F., Sachs, F.: Stretch-activated single ion channel currents in tissue-cultured embryonic
chick skeletal muscle. J. Physiol. 352, 685-701 (1984) Guttman, R., Feldman, L., Jakobsson, E.: Frequency entrainment of squid axon membrane.
J. Membr. Biol. 56, 9-18 (1980) Hille, B.: Ionic Channels of Excitable Membranes. Sunderland, MA: Sinauer (1984) Holmes, M. H., Bell, J.: A model of a sensory mechanoreceptor derived from homogenization.
SIAM J. Appl. Math. 50, 147-166 (1990) Lansman, J. B., Hallam, J., Rink, T. J.: Single stretch-activated ion channels in vascular endothelial
cells as mechanotransducers? Nature 325, 811-813 (1987) Methfessel, C., et al.: Patch clamp measurements on Xenopus laevis oocytes: currents- through end
ogenous channels and implanted acetylcholine receptor and sodium channels. Pflugers Arch. 407, 577-590 (1986)
Morris, C. E.: Mechanosensitive ion channels. J. Memb. Biol. 113, 93-107 (1990) Nemoto, I., et al.: Behavior of solutions of the Hodgkin-Huxley equations and its relation to
properties of mechanoreceptors. Biophys. J. 15, 469-479 (1975) Rail, W.: In: Handbook of Physiology, the Nervous System, vol. 1. Washington, DC: Am. Physiol.
Soc. (1976) Sachs, F.: Biophysics of mechanoreception. Memb. Biochem. 6, 173-193 (1986) Sachs, F., Lecar, H.: Stochastic models for mechanical transduction. Biophys J. 59, 1143-1145
(1991) Yang, X. C., Sachs, F.: Stretch-activated channels in several tissues. Biophys. J. 51, 252a (1987)