a new rainfrog of the pristimantis myersi group (amphibia, craugastoridae) from volcán pichincha,...
TRANSCRIPT

36 Accepted by J. Padial: 12 Feb. 2014; published: 20 Mar. 2014
ZOOTAXA
ISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2014 Magnolia Press
Zootaxa 3780 (1): 036–050
www.mapress.com/zootaxa/
Article
http://dx.doi.org/10.11646/zootaxa.3780.1.2
http://zoobank.org/urn:lsid:zoobank.org:pub:810ED246-F468-4081-A3F0-54F23A6BC895
A new rainfrog of the Pristimantis myersi Group (Amphibia, Craugastoridae)
from Volcán Pichincha, Ecuador
FERNANDO J. M. ROJAS-RUNJAIC1
, J. AMANDA DELGADO C.2
& JUAN M. GUAYASAMIN3,4
1
Museo de Historia Natural La Salle. Apartado Postal 1930, Caracas 1010-A, Venezuela. E-mail: [email protected]
2
Museo de Historia Natural, Universidad Nacional de San Antonio Abad del Cusco, Cusco, Peru. E-mail: [email protected]
3
Centro de Investigación de la Biodiversidad y Cambio Climático (BioCamb), Universidad Tecnológica Indoamérica, Av. Machala y
Sabanilla, Quito, Ecuador. E-mail: [email protected]
4
Corresponding author
Abstract
A new frog of the Pristimantis myersi Group is described from a bamboo patch within the Reserva Ecológica Verdecocha
(0°5'46.9"S, 78°36'15.3"W; 2851 m), located at northwestern flank of the Volcán Pichincha, in the vicinities of Quito, Ec-
uador. The new species is known from eight adult males, whereas the females remain unknown; it can be readily distin-
guished from all species of the P. myersi Group that inhabit the highlands of the Ecuadorian Andes by the unique
combination of the following characters: body small (adult male SVL 14.9–19.7 mm; females unknown); dorsal skin sha-
green, with a barely visible middorsal raphe, scapular and dorsolateral folds; tympanum small but well-defined; upper eye-
lid with one enlarged tubercle; males with prominent vocal slits, but without nuptial pads on thumbs; fold-like tarsal
tubercles. With this new species, the number of Pristimantis assigned to the P. myersi Group raises to 16, of which, 12 are
in Ecuador. We provide notes on morphology and color variation, advertisement call, and natural history of the new spe-
cies.
Key words: Anura, Terrarana, South America, Ecuador
Resumen
Se describe una nueva especie del grupo de Pristimantis myersi asociada a los parches de bambú en la Reserva Ecológica
Verdecocha (0°5'46.9"S, 78°36'15.3"W; 2851 m), ubicada en el flanco noroccidental del Volcán Pichincha, cerca de la ciu-
dad de Quito, Ecuador. La nueva especie es conocida sólo por ocho individuos machos adultos, en tanto que las hembras
aún se desconocen. Puede distinguirse fácilmente de las demás especies del grupo de P. myersi por la combinación única
de los siguientes caracteres: talla pequeña (machos adultos con una longitud rostro-cloacal de 14.9–19.7 mm; hembras
desconocidas); piel del dorso finamente granular, con un pliegue mediodorsal poco elevado, pliegues escapulares y dor-
solaterales presentes; tímpano pequeño pero bien definido; párpado superior con un tubérculo agrandado; macho con in-
cisuras vocales prominentes pero carentes de parches nupciales en sus pulgares; tubérculos tarsales casi formando un
pliegue interrumpido. Con la nueva especie el número de Pristimantis asignados al grupo de P. myersi asciende a 16, de
los cuales 12 están presentes en Ecuador. En este trabajo también se aportan notas sobre variación de color y morfología,
descripción del canto y notas de historia natural de la nueva especie.
Introduction
The direct-developing frog genus Pristimantis, with 469 described species (Frost 2014), is by far the most specious
group of terrestrial vertebrates in the world. This genus concentrates a substantial part of its richness in the Andes
of Colombia, Ecuador, and Peru (Lynch & Duellman 1997; Duellman & Lehr 2009; Heinicke et al. 2007; Hedges
et al. 2008). Allopatric speciation, promoted by the additive effect of the topographic and ecological complexity of
the Andes, the low dispersal abilities of terraranans, and a combination of ecological and morphological traits, has

Zootaxa 3780 (1) © 2014 Magnolia Press · 37A NEW RAINFROG OF THE PRISTIMANTIS MYERSI GROUP
been proposed as the cause of its high richness of this group in the Andes (Lynch & Duellman 1997; Gonzalez-
Voyer et al. 2011).
Despite the high number of Pristimantis known from the Ecuadorian Andes (148 spp; Centro Jambatu 2011–
2013), the continuous discovery of new species in recent years (Guayasamin & Funk 2009; Yánez-Muñoz et al.
2010; Reyes-Puig et al. 2010; Valencia et al. 2010; Arteaga-Navarro & Guayasamin 2011; Reyes-Puig & Yánez-
Muñoz 2012; Guayasamin & Arteaga 2013) demonstrates that the current richness of the genus in this region is far
from being known. Additionally, with the advent of integrative approaches in taxonomy (including molecular
techniques), it is now evident that numerous species are morphologically cryptic (Padial & De la Riva 2009; Díaz
et al. 2012; Fouquet et al. 2012; Padial et al. 2012). Thus, it is reasonable to predict that the rate of discovery and
description of new species of Pristimantis will only increase in the following years.
The Pristimantis myersi Group is a phenetic cluster of small terrestrial frogs, cloud forest and paramo dwellers,
distributed in the Andes of southern Colombia and Ecuador (Hedges et al. 2008). To date, this group comprises 15
species (Hedges et al. 2008; Guayasamin & Funk 2009; Rödder & Schimtz 2009; Yánez-Muñoz et al. 2010), 11 of
which are present in Ecuador: P. bicantus Guayasamin & Funk, 2009, P. festae (Peracca, 1904), P. floridus (Lynch
& Duellman, 1997), P. gladiator (Lynch, 1976), P. hectus (Lynch & Burrowes, 1990), P. leoni (Lynch, 1976), P.
lucidosignatus Rödder & Schimtz, 2009, P. ocreatus (Lynch, 1981), P. onorei Rödder & Schimtz, 2009, P.
pyrrhomerus (Lynch, 1976) and P. sirnigeli Yánez-Muñoz, Meza-Ramos, Cisneros-Heredia & Reyes, 2010 (Centro
Jambatu 2011–2013). Molecular phylogenies support the monophyly of the P. myersi Group, although taxon
sampling only has included up to four species (see Hedges et al., 2008; Pinto-Sánchez et al., 2012).
In this work, we described a new species of Pristimantis for this group from a cloud forest in the Volcán
Pichincha, western versant of the Ecuadorian Andes. Additionally, we provide information on the variation,
advertisement calls, and natural history of the species.
Materials and methods
Museum acronyms are those listed by Frost (2014), except when noted. Specimens were collected manually,
euthanized by immersion in Lidocaine 10%, fixed and preserved in ethanol 70%, and deposited in the collection of
amphibians of the Museo de Zoología of the Universidad Tecnológica Indoamérica, Quito (MZUTI). Generic
classification follows Hedges et al. (2008), whereas family-level taxonomy follows Pyron & Wiens (2011).
Terminology for morphological characters mostly follows Lynch & Duellman (1997) and Duellman & Lehr
(2009). Diagnosis and description follows Lynch & Duellman (1997), Duellman & Lehr (2009) and Rojas-Runjaic
et al. (2013). Gender and maturity were determined by examination of external secondary sexual characters (i.e.,
presence or absence of vocal slits) and gonads (i.e., presence of testes or ovaries) through dissection.
Measurements reported also follows Lynch & Duellman (1997), Duellman & Lehr (2009) and Rojas-Runjaic
et al. (2013), and are: Snout-Vent Length (SVL): straight length measured from tip of snout to vent; Thigh Length
(ThL): from vent opening to flexed knee; Shank Length (SL): from outer edge of flexed knee to flexed heel; Foot
Length (FL): from proximal edge of outer metatarsal tubercle to tip of Toe IV; Hand Length (HaL): from proximal
edge of palmar tubercle to tip of Finger III; Head Length (HeL): from tip of snout to posterior edge of prootic,
noted through the skin; Head Width (HW): measured across the skull at angle of jaws; Internarial Distance (InD):
measured between centers of nares; Interorbital distance (IOD): shortest distance between upper eyelids; Upper
Eyelid Width (UEW): measured transversely across widest part of upper eyelid; Eye-to-Nostril Distance (EN):
measured from the anterior edge of eye to nostril; Eye Diameter (ED): measured horizontally across eye; Eye-to-
Snout Distance (ETS): distance between the anterior corner of the eye to the tip of snout; Tympanum Diameter
(TD): measured horizontally across tympanum; Disc Width of Finger III (F3D): measured across widest part of
disk; Disc Width of Toe IV (T4D): measured across widest part of disk; Length of Finger I (F1L): measured from
outer edge of palmar tubercle to tip of disc; Length of Finger II (F2L): measured from outer edge of palmar
tubercle to tip of disc. All measurements were taken with digital caliper with precision to the nearest 0.1 mm, and
are only from adult frogs.
Comparative data was taken from Lynch (1968; 1976b; 1981), Lynch & Burrowers (1990), Lynch & Duellman
(1980; 1997), Guayasamin & Funk (2009), Rödder & Schmitz (2009), Yánez-Muñoz et al. (2010) and from
selected type and topotype specimens (see Appendix 1) housed in the following collections: MZUTI, Museo

ROJAS-RUNJAIC ET AL.38 · Zootaxa 3780 (1) © 2014 Magnolia Press
Ecuatoriano de Ciencias Naturales, Quito (MECN), Museo de Zoología of the Pontificia Universidad Católica de
Ecuador, Quito (QCAZ), Amphibian Collection of the Instituto de Investigación de los Recursos Biológicos
Alexander von Humboldt, Villa de Leyva (IAvH), Amphibian Collection of the Instituto de Ciencias Naturales,
Universidad Nacional de Colombia, Bogotá (ICN), National Museum of Natural History (USNM), and Natural
History Museum and Biodiversity Research Center of Kansas University (KU).
Calls were recorded at a distance less than 1 m of the calling males, using a Sennheiser K6-ME66
unidirectional microphone, and Olympus LS-10 Linear PCM Field Recorder. Calls were analyzed in the Sound
Analysis Sofware RAVEN (Charif et al. 2004). Call variables were obtained as described in Hutter & Guayasamin
(2012) and references therein and are: call duration, interval between calls, number of notes per call, call type
(tonal or pulsed), number of pulses per call, dominant frequency, and harmonic frequency.
Results
As stated in Hedges et al. (2008), the genus Pristimantis lacks diagnostic morphological synapomorphies.
However, we tentatively assign the new species to this genus based on its distribution in South America and overall
similarity to most members currently placed in the genus Pristimantis. We therefore restrict character states
comparisons among species to members of this group.
Pristimantis munozi sp. nov.
(Figures 1–3, Table 1)
English name: Muñoz’ Rainfrog
Spanish name: Cutín de Muñoz
Holotype. Adult male, MZUTI 1782 (field number ANF 1129), from Reserva Verdecocha, Province of Pichincha,
Ecuador (0°5'46.9"S, 78°36'15.3"W; elevation 2851 m), collected on 21 of July 2012, by F.J.M. Rojas-Runjaic and
J.A. Delgado.
Paratopotypes. Seven adult males, MZUTI 1777–1781, 1783–1784 (field numbers ANF 1124–1128, 1130–
1131 respectively), with the same collection data as the holotype.
Diagnosis. A species of the genus Pristimantis, assigned to the Pristimantis myersi species group (sensu Lynch
& Duellman 1997, and Hedges et al. 2008), by its small size (SVL < 28 mm), robust body, relative narrow head
and short snout, absence of cranial crests, tympanum differentiated, vocal slits and vomerine teeth present, limbs
short, FI shorter than FII, TV slightly longer than TIII and not extending to the proximal edge of the distal
subarticular tubercle of TIV, digital discs narrow and rounded. The new species is readily diagnosable by the
combination of the following characters: (1) body small (SVL in adult males = 14.9–19.7 mm; females unknown);
(2) dorsal skin shagreen, with a barely visible middorsal raphe, scapular and dorsolateral folds; (3) ventral skin
areolate on chest and belly, smooth to slightly areolate on throat; (4) tympanum small (1/2–2/5 of ED), well-
defined, slightly higher than long; tympanic membrane differentiated; tympanic annulus prominent anteroventrally,
posterodorsally concealed beneath skin; (5) snout acuminate in dorsal view, rounded in profile; (6) canthus
rostralis weakly concave in dorsal view, nearly angular in cross-section; (7) upper eyelid with one large and few
low tubercles; (8) choanae large, round, not concealed by palatal shelf of maxillary arch; (9) dentigerous processes
of vomers low, triangular, slightly smaller than choanae, arranged in V-shaped, posterior and medial to choanae,
each bearing one to four teeth; (10) tongue oval, longer than wide, posterior 1/4 free, posterior edge feebly notched;
(11) males with prominent vocal slits; (12) males without nuptial pads on thumbs; (13) FI shorter than FII; (14)
preaxial keels only on FII and FIII, very weak and barely visible; (15) two to four large ulnar tubercles
longitudinally aligned on the outer ventrolateral surface of forearm; (16) finger disc and pad absent on FI, pads
present and disc slightly expanded on FII to FIV; (17) calcar tubercles absent; heel with few, low tubercles; (18)
four to five fold-like tarsal tubercles aligned on the outer ventrolateral surface of tarsus; (19) inner tarsal fold short,
low and straight, extending on the distal fourth of the tarsus; (20) outer metatarsal tubercle large, round, swollen;
inner metatarsal tubercle oval, elongate, about two times the size of the outer; (21) toes without lateral fringes or

Zootaxa 3780 (1) © 2014 Magnolia Press · 39A NEW RAINFROG OF THE PRISTIMANTIS MYERSI GROUP
keels; (21) toes not webbed; (22) all toes with slightly expanded discs and digital pads; (23) TV slightly longer than
TIII; (24) throat white reticulate of black, rest of ventral surfaces pale orange finely punctuated of black.
Comparison with similar species. Pristimantis munozi has a small but well-defined tympanum and slightly
expanded discs on FII to IV, these traits differentiate P. munozi from P. scopaeus, which lacks tympanum and disc
on FII-IV. In P. munozi the tips of finger and toe discs are rounded and present an inner metatarsal tubercle two
times the size of the outer and distally not free, which differs notably from the acuminate tips of the discs and the
larger (ca. four times the size of the outer) and distally free inner metatarsal tubercle of P. hectus, P. lucidosignatus,
and P. onorei. Adult males of P. munozi have prominent vocal slits while there are absent in P. xeniolum and P.
floridus; additionally, the new species differs from P. xeniolum by having odontophores and vomerine teeth (absent
in the latter), and from P. floridus by having prominent dorsolateral folds, and discs only slightly expanded on
fingers and toes (P. floridus lacks dorsolateral folds and presents wide discs, about twice the width of the last
phalange). Prominent dorsolateral folds, weak preaxial keels on FII and III, and four to five fold-like tarsal
tubercles aligned on the outer ventrolateral surface of tarsus distinguishes to P. munozi from P. repens, which lacks
dorsolateral folds the dorsum, keels on fingers and tubercles on the tarsus. Pristimantis munozi differs from P.
ocreatus by having odontophores on vomer, preaxial keels on FII–III, pads on FII–IV, fold-like tubercles on tarsus,
and males with prominent vocal slits (P. ocreatus lacking odontophores on vomers, keels and pads on fingers,
tubercles on tarsus, and only with short vocal slits). From P. pyrrhomerus, the new species differs in having
prominent dorsolateral folds, preaxial keels on FII–III, weak discs and pads on FII–IV, fold-like tubercles on outer
ventrolateral edge of tarsus, dentigerous process of vomers low, males with prominent vocal slits and without red
color on groin and concealed parts of thighs (P. pyrrhomerus lacks dorsolateral folds and lateral keels on fingers,
discs and pads absent on FI–II, outer tarsal tubercles small and sub-conical, prominent dentigerous process, and
males whit short vocal slits and red colored groins and hidden parts of thighs). Pristimantis munozi differs from P.
myersi by having dentigerous process on vomers, white ventral color finely spotted of black, with translucent gray
groins, and by lacking papilla at tip of snout, lateral keels on toes, and by have a large tubercle on the upper eyelid
(P. myersi presents a fleshy papilla at tip of snout, lateral keels on toes, upper eyelid with numerous small tubercles,
black ventral color spotted with white, groins with red spots, and lacks dentigerous process of vomers).
Pristimantis munozi distinguished from P. festae by having the dorsal skin shagreen with middorsal raphe, scapular
and dorsolateral folds, small tympanum, snout acuminate in dorsal view, canthus nearly angular in cross-section,
enlarged tubercle on the upper eyelid, pads on FII–IV, white venter finely spotted with black, and lacking of lateral
keels on toes (P. festae have dorsal skin smooth without middorsal raphe nor dorsolateral folds, present a prominent
tympanum, snout rounded in dorsal view, canthus rounded, upper eyelid without enlarged tubercles, fingers lacking
pads, toes whit lateral keels, and black venter spotted with white and pale cream). Pristimantis gladiator differs
from the new species by lacking middorsal raphe and dorsolateral folds, and having only small tubercles on the
upper eyelid and a prominent tympanum; additionally, P. gladiator shows slightly pointed and asymmetrical discs
(discs with tip rounded in P. munozi). From P. bicantus the new species is distinguished by having prominent
dorsolateral folds, an enlarged tubercle on the upper eyelid, snout acuminate in dorsal view, prolateral keels on FII–
III, ulnar tubercles and outer tarsal tubercles present (P. bicantus usually lacks dorsolateral folds, lateral keels on
fingers, ulnar and outer tarsal tubercles, and has small tubercles on the upper eyelid, and rounded snout in dorsal
view). At the type locality, P. munozi is sympatric with P. leoni and P. sirnigeli; however, it is distinguished from
the former by having one large and rounded tubercle on the upper eyelid, preaxial keels on FII–III, toes without
lateral keels, tips of the discs on toes rounded, and fold-like outer tarsal tubercles (P. leoni with numerous enlarged
tubercles on the upper eyelid, lateral keels absent on the fingers but present on the toes, tips of the discs on toes
slightly pointed, and outer tarsal tubercles small and conical). Finally, P. munozi is distinguished from P. sirnigeli
by having a smaller body size in adult males (SVL = 14.9–17.9 mm), prominent dorsolateral folds, relatively short
fingers, and by lacking lateral keels on toes (P. sirnigeli with lateral keels on toes, relatively long and thin fingers,
and male SVL = 18.6–20.6 mm).
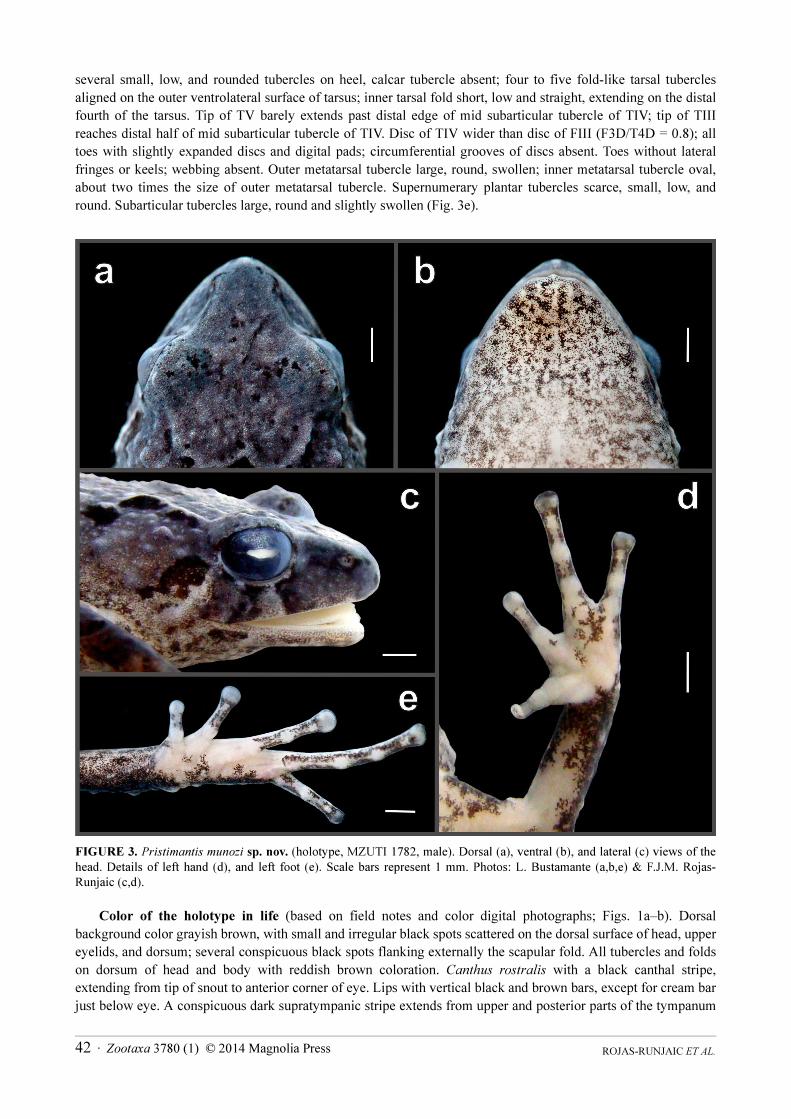
Description of the holotype. Adult male of 17.6 mm SVL. Body short and robust (Figs. 1a–b, 2a–b). Head
slightly narrower than body, slightly longer than wide (Figs. 2a–b, 3a–b); HeL 39.5% of SVL; greatest head width
between angles of jaws 38% of SVL; cranial crests absent. Snout short, acuminate in dorsal view (Fig. 3a), and
rounded in profile (Fig. 3c); larger than eye (ETS/ED = 1.5); distance eye-nostril slightly shorter than eye diameter
(EN/ED = 0.8). Canthus rostralis weakly concave in dorsal view; nearly angular in cross-section; loreal region
slightly concave, sloping outward to lip. Not protruding lips. Nares not protuberant, with dorsolateral orientation,

ROJAS-RUNJAIC ET AL.40 · Zootaxa 3780 (1) © 2014 Magnolia Press
barely visible from the front and completely observable in dorsal view. Upper eyelid wide, narrower than
interocular region (UEW/IO = 0.8), with one enlarged and rounded tubercle on each eyelid and few additional low
tubercles. Tympanum small (TD/ED = 0.4) but well-defined, slightly higher than long; tympanic membrane
differentiated; tympanic annulus prominent anteroventrally, posterodorsally concealed beneath skin, barely visible
in dorsal view. Supratympanic fold absent, with several large and conical postrictal tubercles below tympanum.
Choanae large, round, not concealed by palatal shelf of maxillary arch; dentigerous processes of vomers low,
triangular, slightly smaller than choanae, arranged in V-shape, posterior and medial to choanae, each bearing three-
to-four teeth. Tongue oval, longer than wide, posterior 1/4 free, posterior edge feebly notched. Vocal slits present
and prominent.
FIGURE 1. Pristimantis munozi sp. nov. Dorsolateral (a) and ventral (b) views of the holotype (MZUTI 1782, male) in life.
Dorsolateral (c) and ventral (d) views of the paratype (MZUTI 1781, male) in life. Photos: F.J.M. Rojas-Runjaic.
Skin of dorsum and upper eyelids shagreen; middorsal raphe present, but low and barely distinguishable;
scapular folds present, sinusoidal and prominent, ending posteriorly in a large conical tubercle; dorsolateral folds
straight and prominent, extending from posterior end of scapular folds to level of groins; no paravertebral folds
(Fig. 2a); cloacal sheath absent, small cloacal tubercles present but not conspicuous; flanks and posterior third of

Zootaxa 3780 (1) © 2014 Magnolia Press · 41A NEW RAINFROG OF THE PRISTIMANTIS MYERSI GROUP
dorsum slightly tuberculate; dorsal surface of limbs finely tuberculate. Throat smooth, chest and belly areolate;
ventral surface of limbs smooth to slightly areolate (Fig. 1b). Two-to-three large ulnar tubercles along outer
ventrolateral surface of forearm.
FIGURE 2. Pristimantis munozi sp. nov. Dorsal (a) and ventral (b) views of the holotype (MZUTI 1782, male) in preservative.
Dorsal (c) and ventral (d) views of the paratype (MZUTI 1781, male) in preservative. Scale bars represent 10 mm. Photos: L.
Bustamante.
Hand length 28.6% of SVL. Relative lengths of adpressed fingers III > IV > II > I; adpressed FI does not reach
proximal edge of disc pad of FII; nuptial pads absent; adpresed FIV reaches past distal subarticular tubercle of FIII.
Preaxial keels only present on FII and III, very weak and barely visible. Finger disc absent on FI, slightly expanded
on FII to IV; digital pad absent on FI, present in all other fingers, rounded, almost longer than wide; circumferential
groove of digital pads poorly defined laterally and incomplete proximally; distal edge of discs rounded; disc of FIII
about 1.4 times wider than distal end of adjacent phalanx. Subarticular tubercles large, longer than wide, and
slightly swollen; palmar tubercle large, slightly swollen, deeply bifid, V-shaped; thenar tubercle large, slightly
swollen, elongate; supernumerary tubercles present, relative large (between 1/3–1/2 size of subarticular tubercles)
(Fig. 3d).
Hind limbs relatively long (thigh length 50.5% SVL; shank length 56.3% SVL; foot length 50.5% SVL); heels
overlap when shanks are held perpendicular to sagittal plane; relative length of adpressed toes IV > V > III > II > I;
ROJAS-RUNJAIC ET AL.42 · Zootaxa 3780 (1) © 2014 Magnolia Press
several small, low, and rounded tubercles on heel, calcar tubercle absent; four to five fold-like tarsal tubercles
aligned on the outer ventrolateral surface of tarsus; inner tarsal fold short, low and straight, extending on the distal
fourth of the tarsus. Tip of TV barely extends past distal edge of mid subarticular tubercle of TIV; tip of TIII
reaches distal half of mid subarticular tubercle of TIV. Disc of TIV wider than disc of FIII (F3D/T4D = 0.8); all
toes with slightly expanded discs and digital pads; circumferential grooves of discs absent. Toes without lateral
fringes or keels; webbing absent. Outer metatarsal tubercle large, round, swollen; inner metatarsal tubercle oval,
about two times the size of outer metatarsal tubercle. Supernumerary plantar tubercles scarce, small, low, and
round. Subarticular tubercles large, round and slightly swollen (Fig. 3e).
FIGURE 3. Pristimantis munozi sp. nov. (holotype, MZUTI 1782, male). Dorsal (a), ventral (b), and lateral (c) views of the
head. Details of left hand (d), and left foot (e). Scale bars represent 1 mm. Photos: L. Bustamante (a,b,e) & F.J.M. Rojas-
Runjaic (c,d).
Color of the holotype in life (based on field notes and color digital photographs; Figs. 1a–b). Dorsal
background color grayish brown, with small and irregular black spots scattered on the dorsal surface of head, upper
eyelids, and dorsum; several conspicuous black spots flanking externally the scapular fold. All tubercles and folds
on dorsum of head and body with reddish brown coloration. Canthus rostralis with a black canthal stripe,
extending from tip of snout to anterior corner of eye. Lips with vertical black and brown bars, except for cream bar
just below eye. A conspicuous dark supratympanic stripe extends from upper and posterior parts of the tympanum

Zootaxa 3780 (1) © 2014 Magnolia Press · 43A NEW RAINFROG OF THE PRISTIMANTIS MYERSI GROUP
and fails to reach the shoulder; the inferior border of this stripe is also delineated of creamish white. Flank grayish
brown, slightly lighter than dorsum, with scattered small black spots and with one or two diffuse diagonal cream
stripes. Groin background translucent gray, with several small immaculate white dots. Upper arm light brown;
forearm grayish brown with two-to-three irregular blackish brown cross-bands in the distal half, more intensely
pigmented the proximal one. Hand and fingers light brown with three-to-four ill-defined pale brown cross-bands;
tips of finger discs reddish brown. Hind limbs background grayish brown, becoming paler distally (on feet), with
three to four blackish brown cross-bands on thighs, shanks, tarsi and feet; blackish cross-bands on thighs laterally
bordered with light lines; background color of the inner surface of thighs translucent gray, densely spotted of black
and with scattered small white dots; transversal rows of tubercles on shanks pigmented of reddish brown. Toes pale
orange; toe discs dorsally grayish, bordered by reddish brown coloration. Throat white, finely reticulated with
black; chest, belly, undersurface forelimbs of thighs and shanks pale orange, finely punctuated of black; tarsi
ventrally grayish brown; palms and soles pale orange; subarticular tubercles of feet bright orange. Iris golden with
fine black venation and a broad horizontal copper band; pupil ring golden.
Color of the holotype in preservative (after sixteen months, June 2013; Figs. 2a–b, 3a–e). Dorsal background
almost uniformly gray; reddish brown coloration of dorsal tubercles and folds is replaced by gray and light gray;
dorsal and lateral black spots turns more dark and conspicuous. Flanks turn grayish, lighter towards lower portion;
diagonal light bands of flanks disappear. Groins and ventral surfaces turn white, with black reticulation. Palms and
soles white, with diffuse pink tone and several small black blotches. Fingers and toes with dorsal dark gray,
transversal bars, and ventrally colored of white, densely spotted of black. Iris grayish, finely punctuated with black;
horizontal copper band turns black.
Measurements of the holotype (in mm). SVL: 17.6; ThL: 8.9; SL: 9.9; FL: 8.9; HaL: 5.0; HeL: 6.9; HW: 6.7;
InD: 1.9; IOD: 2.1; UEW: 1.7; EN: 1.7; ED: 2.2; ETS: 3.1; TD: 0.9; F3D: 0.6; T4D: 0.8; F1L: 2.6; F2L: 3.3.
Variation. Females of Pristimantis munozi are unknown; then, sexual dimorphism is not evaluated. We
mention, however, that females of all other species of Pristimantis myersi group are larger than males, and several
of them have red blotches on the groins (gender-specific coloration). Thus, we suppose that adult females of P.
munozi may be are larger than males and may have red flash coloration on the groins, as other species of the group.
Adult males range from 14.9–19.7 (16.7 ±1.6; n = 8). Variation of morphometric characters is shown in Table
1. Head length ranges from 39.1%–42.3% SVL (40.6% ± 1.1; n = 8), and maximum head width from 35.4%–
42.6% SVL (38.3% ± 2.2; n = 8). Head can be as long as wide, or slightly longer than wide (HeL/HW: 1.0 –1.1[1.1
± 0.1; n = 8]). The EN/ED ratio is somewhat variable but every eye-naris distance is smaller than eye diameter
(0.7–0.9 [0.8 ± 0.1; n = 8]). The width of the upper eyelid is narrower than the interorbital distance, and its ratio
ranges from 0.7–0.8 (0.78 ± 0.1; n = 8). The snout length also is slightly variable but always longer than eye
diameter (1.2–1.6 [1.4 ± 0.1; n = 8]). Hand length ranges from 26.4–29.7% SVL (28.2% ± 1.2; n = 8). The
tympanum is small and its ratio respect to eye has little variation (TD/ED: 0.4–0.5; n = 8). Thigh length ranges
from 49.3%–55.0% SVL (51.9% ± 2.2; n = 8); shank length from 53.6–62.1% SVL (58.0% ± 2.8; n = 8), and foot
length from 48.5%–54.4% (52.1 ± 2.0; n = 8).
In almost all specimens dorsal skin texture is shagreen with scattered tubercles of variable size, except MZUTI
1781, which have dorsal skin nearly smooth, non-tubercular, and with scattered granules on flanks and posterior
third of dorsum. Flanks and posterior third of dorsum densely tubercular in MZUTI 1777. Upper eyelid typically
with one tubercle; MZUTI 1777 has two tubercles on each upper eyelid. Middorsal raphe absent in MZUTI 1778,
but prominent in MZUTI1779. Scapular folds extending anteriorly and reaching tubercles of upper eyelids in
MZUTI 1783, nearly straight and continuous with dorsolateral folds in MZUTI 1779 and 1781. Dorsolateral folds
extending to sacral region (MZUTI 1777–1778, 1784), to groins (MZUTI 1780–1781) or inclusively converging
above the vent level (MZUTI 1779, 1783). Ulnar tubercles on forearms range from two-to-four; tarsal tubercles in
number of four or five. Throat typically smooth, but slightly areolate in some specimens (MZUTI 1777, 1780,
1784). Number of vomerine teeth somewhat variable, from one or two (MZUTI 1777–1778) to four (MZUTI
1783), but typically three-to-four (MZUTI 1779–1782).
In almost all specimens examined, the color pattern is similar to that of the holotype (Figs. 1a–b), except
specimens MZUTI 1779 and 1781 (Figs. 1c–d, 2c–d), which show striking differences. These specimens have a
dorsal background pale ocher (pale gray in preservative), extended from the tip of snout to the vent, and medially to
the scapular and dorsolateral folds; this area is bordered by a sinuous black stripe that runs from the tip of snout,
through nares, canthus, external border of upper eyelid, external flanks of the scapular an dorsolateral folds to
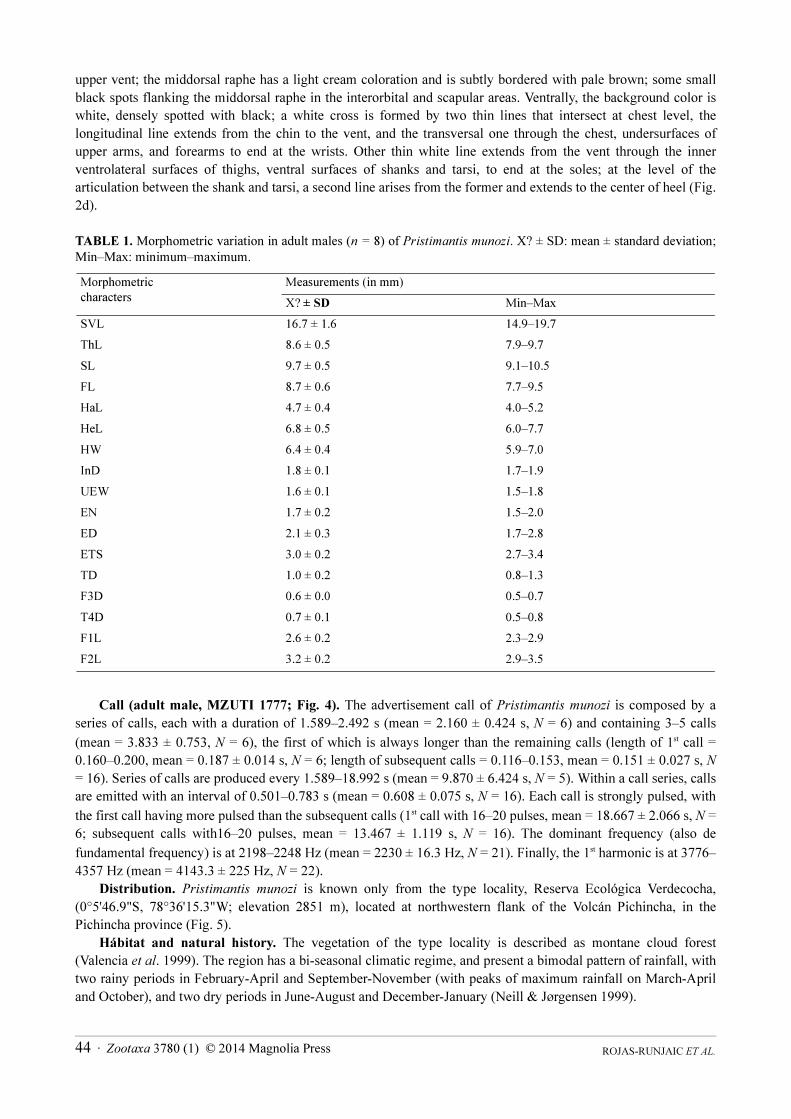
ROJAS-RUNJAIC ET AL.44 · Zootaxa 3780 (1) © 2014 Magnolia Press
upper vent; the middorsal raphe has a light cream coloration and is subtly bordered with pale brown; some small
black spots flanking the middorsal raphe in the interorbital and scapular areas. Ventrally, the background color is
white, densely spotted with black; a white cross is formed by two thin lines that intersect at chest level, the
longitudinal line extends from the chin to the vent, and the transversal one through the chest, undersurfaces of
upper arms, and forearms to end at the wrists. Other thin white line extends from the vent through the inner
ventrolateral surfaces of thighs, ventral surfaces of shanks and tarsi, to end at the soles; at the level of the
articulation between the shank and tarsi, a second line arises from the former and extends to the center of heel (Fig.
2d).
TABLE 1. Morphometric variation in adult males (n = 8) of Pristimantis munozi. X? ± SD: mean ± standard deviation;
Min–Max: minimum–maximum.
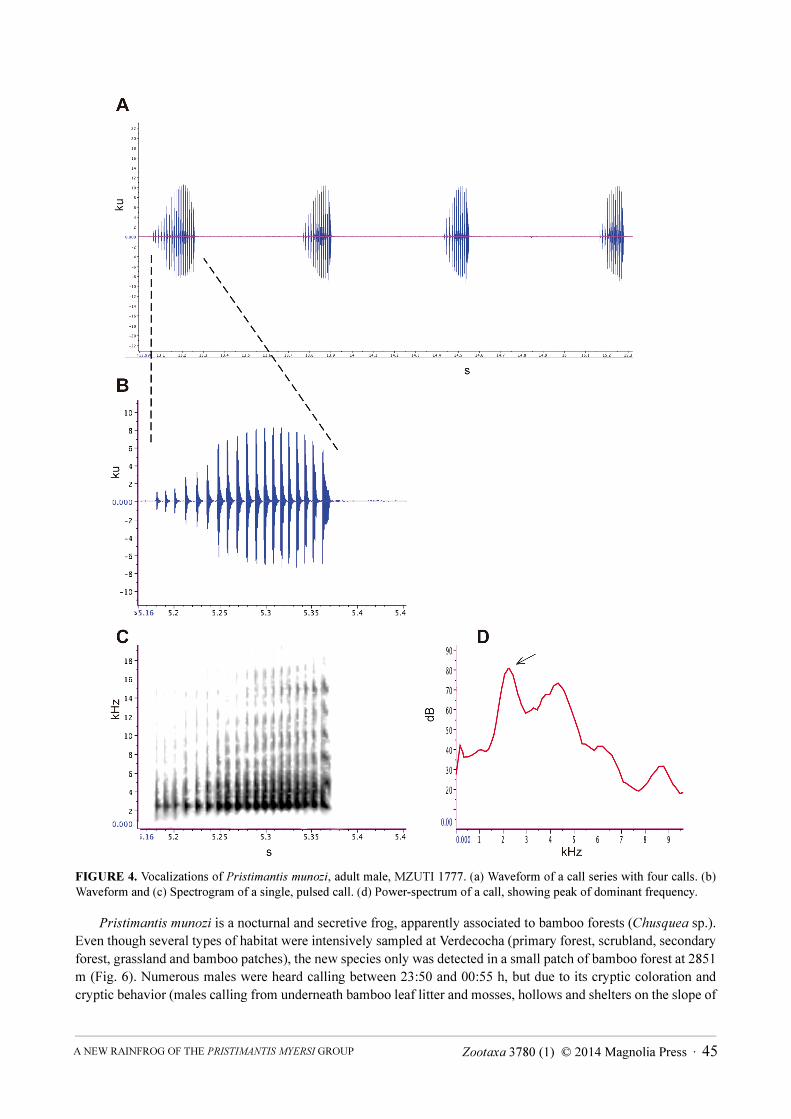
Call (adult male, MZUTI 1777; Fig. 4). The advertisement call of Pristimantis munozi is composed by a
series of calls, each with a duration of 1.589–2.492 s (mean = 2.160 ± 0.424 s, N = 6) and containing 3–5 calls
(mean = 3.833 ± 0.753, N = 6), the first of which is always longer than the remaining calls (length of 1st
call =
0.160–0.200, mean = 0.187 ± 0.014 s, N = 6; length of subsequent calls = 0.116–0.153, mean = 0.151 ± 0.027 s, N
= 16). Series of calls are produced every 1.589–18.992 s (mean = 9.870 ± 6.424 s, N = 5). Within a call series, calls
are emitted with an interval of 0.501–0.783 s (mean = 0.608 ± 0.075 s, N = 16). Each call is strongly pulsed, with
the first call having more pulsed than the subsequent calls (1st
call with 16–20 pulses, mean = 18.667 ± 2.066 s, N =
6; subsequent calls with16–20 pulses, mean = 13.467 ± 1.119 s, N = 16). The dominant frequency (also de
fundamental frequency) is at 2198–2248 Hz (mean = 2230 ± 16.3 Hz, N = 21). Finally, the 1st
harmonic is at 3776–
4357 Hz (mean = 4143.3 ± 225 Hz, N = 22).
Distribution. Pristimantis munozi is known only from the type locality, Reserva Ecológica Verdecocha,
(0°5'46.9"S, 78°36'15.3"W; elevation 2851 m), located at northwestern flank of the Volcán Pichincha, in the
Pichincha province (Fig. 5).
Hábitat and natural history. The vegetation of the type locality is described as montane cloud forest
(Valencia et al. 1999). The region has a bi-seasonal climatic regime, and present a bimodal pattern of rainfall, with
two rainy periods in February-April and September-November (with peaks of maximum rainfall on March-April
and October), and two dry periods in June-August and December-January (Neill & Jørgensen 1999).
Morphometric
characters
Measurements (in mm)
X? ± SD Min–Max
SVL 16.7 ± 1.6 14.9–19.7
ThL 8.6 ± 0.5 7.9–9.7
SL 9.7 ± 0.5 9.1–10.5
FL 8.7 ± 0.6 7.7–9.5
HaL 4.7 ± 0.4 4.0–5.2
HeL 6.8 ± 0.5 6.0–7.7
HW 6.4 ± 0.4 5.9–7.0
InD 1.8 ± 0.1 1.7–1.9
UEW 1.6 ± 0.1 1.5–1.8
EN 1.7 ± 0.2 1.5–2.0
ED 2.1 ± 0.3 1.7–2.8
ETS 3.0 ± 0.2 2.7–3.4
TD 1.0 ± 0.2 0.8–1.3
F3D 0.6 ± 0.0 0.5–0.7
T4D 0.7 ± 0.1 0.5–0.8
F1L 2.6 ± 0.2 2.3–2.9
F2L 3.2 ± 0.2 2.9–3.5
Zootaxa 3780 (1) © 2014 Magnolia Press · 45A NEW RAINFROG OF THE PRISTIMANTIS MYERSI GROUP
FIGURE 4. Vocalizations of Pristimantis munozi, adult male, MZUTI 1777. (a) Waveform of a call series with four calls. (b)
Waveform and (c) Spectrogram of a single, pulsed call. (d) Power-spectrum of a call, showing peak of dominant frequency.
Pristimantis munozi is a nocturnal and secretive frog, apparently associated to bamboo forests (Chusquea sp.).
Even though several types of habitat were intensively sampled at Verdecocha (primary forest, scrubland, secondary
forest, grassland and bamboo patches), the new species only was detected in a small patch of bamboo forest at 2851
m (Fig. 6). Numerous males were heard calling between 23:50 and 00:55 h, but due to its cryptic coloration and
cryptic behavior (males calling from underneath bamboo leaf litter and mosses, hollows and shelters on the slope of

ROJAS-RUNJAIC ET AL.46 · Zootaxa 3780 (1) © 2014 Magnolia Press
a small seasonal creek) only eight specimens were visually detected and collected. Despite the effort no females
were found. At Verdecocha, Pristimantis leoni and P. sirnigeli are sympatric with P. munozi, but none was found
syntopically with the new species at the bamboo forest, but in adjacent cloud forest dominated by relatively short
trees.
FIGURE 5. Distribution of Pristimantis munozi sp. nov. in the Ecuadorian Andes. White dot: Reserva Verdecocha (type
locality).
Etymology. The specific epithet munozi is a patronym honoring Jésus Muñoz Fuente, prominent Spanish
botanist of the Real Jardín Botánico of Spain, founder and Director of the Master’s Program in “Biodiversidad en
Áreas Tropicales y su Conservation” (MBATC) of the Universidad Internacional Menéndez Pelayo, Spain (UIMP),
and research affiliate of the Centro para la Investigación de la Biodiversidad y Cambio Climático, BioCamb, of the
Universidad Tecnológica Indoamérica, Ecuador. Through this program the Dr. Muñoz Fuente has formed over a
hundred of professionals (including the two first authors of this work), contributing significantly to the study and
conservation of the Neotropical biodiversity. We make recognition to his admirable work, support, and friendship,
naming this new species in his honor.
Acknowledgements
We thank to Ítalo Tapia (Fundación Otonga) for support provided during the fieldwork, and Carlos Montaña,
Claudia Múnera, Claudia Medina (IAvH), John D. Lynch, Jhon J. Ospina-Sarria, Teddy Angarita-Sierra, Marvin
Anganoy (ICN), Belisario Cepeda (Colección Zoológica at the Universidad de Nariño, PSO-CZ), Santiago Ron
and Diego Ortiz (QCAZ) for allowing access to the specimens and data of the Pristimantis myersi Group housed at
their corresponding institutions. Some photographs of the type series used in the illustrations were taken by Lucas
Bustamante (Tropical Herping). This study was funded by the Centro para la Investigación de la Biodiversidad y
Cambio Climático (BioCamb, Universidad Tecnológica Indoamérica), through the project “Patrones de diversidad
de los anfibios andinos del Ecuador”. Fieldwork at Reserva Ecológica Verdecocha was authorized by Mr. Jorge
Enrique Maldonado. The Ministerio del Ambiente de Ecuador provided the required research permits (No.14-
2011-IC-FAU-DPAP-MA).

Zootaxa 3780 (1) © 2014 Magnolia Press · 47A NEW RAINFROG OF THE PRISTIMANTIS MYERSI GROUP
FIGURE 6. General view of the bamboo forest at the Reserva Verdecocha, where Pristimantis munozi sp. nov. inhabits. White
arrows indicate the sites where two specimens of the type series were found while calling hidden. Photo: F.J.M. Rojas-Runjaic.

ROJAS-RUNJAIC ET AL.48 · Zootaxa 3780 (1) © 2014 Magnolia Press
References
Arteaga-Navarro, A.F. & Guayasamin, J.M. (2011) A new frog of the genus Pristimantis (Amphibia: Strabomantidae) from the
high Andes of southeastern Ecuador, discovered using morphological and molecular data. Zootaxa, 2876, 17–29.
Centro Jambatu (2011–2013) Anfibios de Ecuador. Fundación Otonga, Quito, Ecuador. Available from:
www.anfibioswebecuador.ec/anfibiosecuador.aspx (accessed 10 February 2014)
Charif, R.A., Clark, C.W. & Fristrup, K.M. (2004) Raven 1.2 user’s manual. Cornell Laboratory of Ornithology, Ithaca, New
York, 205 pp.
Díaz, L.M., Hedges, B., & Schmid, M. (2012) A new cryptic species of the genus Eleutherodactylus (Amphibia: Anura:
Eleutherodactylidae) from Cuba. Zootaxa, 3220, 44–60.
Duellman, W.E. & Lehr, E. (2009) Terrestrial-breeding frogs (Strabomantidae) in Peru. NTV Science, Münster, 382 pp.
Fouquet, A., Vences, M., Salducci, A.M., Marty, C., Blanc, M., Gilles, A. (2007) Revealing cryptic diversity using molecular
phylogenetics and phylogeography in frogs of the Scinax ruber and Rhinella margaritifera species groups. Molecular
Phylogenetics and Evolution, 43, 567–582.
http://dx.doi.org/10.1016/j.ympev.2006.12.006
Frost, D.R. (2014) Amphibian species of the World: an online reference. Version 6. American Museum of Natural History, New
York, USA. Electronic Database. Available from: http://research.amnh.org/herpetology/amphibia/index.html (accessed 10
January 2014)
Gonzalez-Voyer, A., Padial, J.M., Castroviejo-Fisher, S., De La Riva, I., & Vilá, C. (2011) Correlates of species richness in the
largest Neotropical amphibian radiation. Journal of Evolutionary Biology, 24, 931–942. http://dx.doi.org/10.1111/j.1420-9101.2011.02243.x
Guayasamin, J.M. & Arteaga, A.F. (2013). A new species of the Pristimantis orestes group (Amphibia: Strabomantidae) from
the high Andes of Ecuador, Reserva Mazar. Zootaxa, 3616 (4), 345–356.
http://dx.doi.org/10.11646/zootaxa.3616.4.3
Guayasamin, J.M. & Funk, C.W. (2009) The amphibian community at Yanayacu Biological Station, Ecuador, with a
comparison of vertical microhabitat use among Pristimantis species and the description of a new species of the
Pristimantis myersi group. Zootaxa, 2220, 41–66.
Hedges, S.B., Duellman, W.E. & Heinicke, M.P. (2008) New World direct-developing frogs (Anura: Terrarana): Molecular
phylogeny, classification, biogeography, and conservation. Zootaxa, 1737, 1–182.
Heinicke, M.P., Duellman, W.E. & Hedges, S.B. (2007) Major Caribbean and Central American frog faunas originated by
ancient oceanic dispersal. Proceedings of the National Academy of Sciences, 104, 10092–10097.
http://dx.doi.org/10.1073/pnas.0611051104
Hutter, C. & Guayasamin, J.M. (2012) A new cryptic species of glassfrog (Centrolenidae: Nymphargus) from Reserva Las
Gralarias, Ecuador. Zootaxa, 3257, 1–21.
Lynch, J.D. (1968) Systematic status of some Andean leptodactylid frogs with a description of a new species
of Eleutherodactylus. Herpetologica, 24, 289–300.
Lynch, J.D. (1976b) Three new leptodactylid frogs (genus Eleutherodactylus) from the Andean slopes of Colombia and
Ecuador. Herpetologica, 32, 310–317.
Lynch, J.D. (1981) Leptodactylid frogs of the genus Eleutherodactylus in the Andes of northern Ecuador and adjacent
Colombia. Miscellaneous Publications, Museum of Natural History, University of Kansas, Lawrence, 72, 1–46.
Lynch, J.D. & Burrowes, P.A. (1990) The frogs of the genus Eleutherodactylus (Family Leptodactylidae) at the La Planada
Reserve in southwestern Colombia with descriptions of eight new species. Occasional Papers of the Museum of Natural
History, University of Kansas, Lawrence, 136, 1–31.
Lynch, J.D. & Duellman, W.E. (1980) The Eleutherodactylus of the Amazonian slopes of the Ecuadorian Andes (Anura:
Leptodactylidae). Miscellaneous Publications, Museum of Natural History, University of Kansas, Lawrence, 69, 1–86.
Lynch, J.D. & Duellman, W.E. (1997) Frogs of the genus Eleutherodactylus (Leptodactylidae) in western Ecuador: systematics,
ecology, and biogeography. The University of Kansas Natural History Museum, Special Publications, 23, 1–236.
http://dx.doi.org/10.5962/bhl.title.16289
Neill, D.A. & Jørgensen, P.M. (1999) Climates. In: Jørgensen, P.M. & León-Yánez, S. (Eds.), Catalogue of the vascular plants
of Ecuador. Monographies in Systematic Botany from the Missouri Botanical Garden, 75, 8–13.
Padial, J.M. & De La Riva, I. (2009) Integrative taxonomy reveals cryptic Amazonian species of Pristimantis (Anura:
Strabomantidae). Zoological Journal of the Linnean Society, 155, 97–122. http://dx.doi.org/10.1111/j.1096-3642.2008.00424.x
Padial, J.M., Chaparro, J.C., Castroviejo-Fisher, S., Guayasamin, J.M., Lehr, E., Delgado, A.J., Vaira, M., Teixeira Jr, M.,
Aguayo, R. & De La Riva, I. (2012) A revision of species diversity in the Neotropical genus Oreobates (Anura:
Strabomantidae), with the description of three new species from the Amazonian slopes of the Andes. American Museum
Novitates, 3752, 1–55. http://dx.doi.org/10.1206/3752.2
Pinto-Sánchez, N.R., Ibánez, R., Madriñán, S., Sanjur, O.I., Bermingham, E. & Crawford, A. (2012) The Great American
Biotic Interchange in frogs: Multiple and early colonization of Central America by the South American genus Pristimantis
(Anura: Craugastoridae). Molecular Phylogenetics and Evolution, 62, 954–972. http://dx.doi.org/10.1016/j.ympev.2011.11.022

Zootaxa 3780 (1) © 2014 Magnolia Press · 49A NEW RAINFROG OF THE PRISTIMANTIS MYERSI GROUP
Pyron, R.A. & Wiens, J.J. (2011) A large-scale phylogeny of Amphibia including over 2800 species, and a revised
classification of extant frogs, salamanders, and caecilians. Molecular Phylogenetics and Evolution, 61, 543–583. http://dx.doi.org/10.1016/j.ympev.2011.06.012
Reyes-Puig, J.P., Yánez-Muñoz, M.H., Cisneros-Heredia, D.F. & Ramírez, S.M. (2010) Una nueva especie de rana Pristimantis
(Terrarana: Strabomantidae) de los bosques nublados de la cuenca alta del Río Pastaza, Ecuador. Avances en Ciencias e
Ingenierías, 3, B78–B82.
Reyes-Puig, J.P. & Yánez-Muñoz, M.H. (2012) Una nueva especie de Pristimantis (Anura: Craugastoridae) del corredor
ecológico Llanganates-Sangay, Andes de Ecuador. Papéis Avulsos de Zoologia, 56, 81–91.
Rödder, D. & Schimtz, A. (2009) Two new Pristimantis (Anura, Strabomantidae) belonging to the myersi group from the
Andean slopes of Ecuador. Revue Suisse de Zoologie, 116, 275–288.
Rojas-Runjaic, F.J.M., Salerno, P.E., Señaris, J.C. & Pauly, G.B. (2013) Terraranans of the Lost World: a new species of
Pristimantis (Amphibia, Craugastoridae) from Abakapá-tepui in the Chimantá massif, Venezuelan Guayana, and additions
to the knowledge of P. muchimuk. Zootaxa, 3686, 335–355.
Valencia, J., Yánez-Muñoz, M.H., Betancourt-Yépez, R., Terán-Valdez, A. & Guayasamin, J.M. (2010) Una llamativa nueva
especie de Pristimantis (Anura: Terrarana: Strabomantidae) de las estribaciones noroccidentales de los Andes de Ecuador.
Avances en Ciencias e Ingenierías, 3, B41–B45.
Valencia, R., Cerón, C., Palacios, W. & Sierra, R. (1999) Las formaciones naturales de la sierra del Ecuador. In: Sierra, R. (Ed.),
Propuesta preliminar de un sistema de clasificación de vegetación para el Ecuador continental. Proyecto INEFA/GEF-
BIRF y EcoCiencia, Quito, pp. 79–108.
Yánez-Muñoz, M.H., Meza-Ramos, P., Cisneros-Heredia, D.F., & Reyes P., J.P. (2010) Descripción de tres nuevas especies de
ranas del género Pristimantis (Anura: Terrarana: Strabomantidae) de los bosques nublados del Distrito Metropolitano de
Quito, Ecuador. Avances en Ciencias e Ingenierías, 2, 16–27.
Yánez-Muñoz, M.H., Meza-Ramos, P., Cisneros-Heredia, D.F. & Reyes, P.J.P. (2010) Descripción de tres nuevas especies de
ranas del género Pristimantis (Anura: Terrarana: Strabomantidae) de los bosques nublados del Distrito Metropolitano de
Quito, Ecuador. Avances en Ciencias e Ingenierías, 2, 16–27.
APPENDIX I. Additional specimens examined. IAvH: Instituto de Investigación de Recurso Biológicos Alexander von
Humboldt (specimens of this collection are listed under original acronym of Inderena: IND-AN); MECN: Museo
Ecuatoriano de Ciencias Naturales; MZUTI: Museo de Zoología at the Universidad Tecnológica Indoamérica; ICN:
Instituto de Ciencias Naturales, Universidad Nacional de Colombia; KU: Kansas University; QCAZ: Museo de Zoología
at the Pontificia Universidad Católica del Ecuador.
Pristimantis bicantus. Ecuador: Chimborazo: community of San Antonio de Juval (QCAZ 51559, 51561). Morona Santiago:
Chinguinda (QCAZ 24653); San Vicente, Parque Nacional Sangay (QCAZ 31988); 9 de Octubre, stream between
Cordillera Lajas and Quebrada Cugusha (QCAZ 37182); Bosque Protector Abanico, bamboo forest (QCAZ 49035). Napo:
Cosanga, Estación Científica Yanayacu (QCAZ 19030). Pastaza: Town nearest to Santa Clara, road Puyo-Tena,
community of San Rafael-Chonta Yaku, río Challuwa Yacu, Reserva Comunitaria Ankaku, buffer zone of the Parque
Nacional LLanganates (QCAZ 45803, 45805). Tungurahua: Reserva Río Zuñac (QCAZ 52464, 52466).
Pristimantis festae. Ecuador: Napo: 2 km W of Guamaní (QCAZ 1027). Pichincha: Road Otavalo-Laguna de Mojanda
(MZUTI 1807).
Pristimantis floridus. Ecuador: Pichincha: Quebrada Zapadores (QCAZ 16279, 16281–87, 16291, 16294, 16296).
Pristimantis gladiator. Ecuador: Napo: Cordillera de los Guacamayos, 10 minuts from the radio antennas, by the road to the
antennas (QCAZ 40808); Pacto Sumaco, way to volcano Sumaco, Pavayacu refuge (last refuge near to the volcano)
(QCAZ 41257, 41260, 41268, 41279); Pacto Sumaco, Parque Nacional Sumaco, La Laguna refuge (QCAZ 41305);
Cantón Quijos, Parroquia Papallacta, Locality Pap 5, Río San Pedro, direction Papallacta-Cuyuja (MZUTI 1117–1118);
Cantón Quijos, Parroquia Papallacta, Locality Pap 117, direction Papallacta-Cuyuja (MZUTI 1122–1123, 1214); Cantón
Quijos, Parroquia Papallacta, Locality Pap 9, Río Guango direction Papallacta-Cuyuja (MZUTI 1131). Pichincha: Reserva
Ecológica Cayambe-Coca, Laguna San Marcos (QCAZ 49755). Morona Santiago: Hacienda Zuleta (QCAZ 52628–
52629).
Pristimantis hectus. Colombia: Nariño: Reserva Natural La Planada, mcpo. Ricaurte (IND-AN 1947 [holotype], 1456, 1513–
1514, 1920–1924, 1926, 1928–1931, 1933–1937, 1940–1946, 1949–1957, 1959–1960 [paratypes]). Ecuador: Pichincha:
Reserva Las Gralarias (MZUTI 407, 437–438, 458, 465, 468, 517, 519, 521, 523, 562, 1463–65, 1467).
Pristimantis leoni. Ecuador: Carchi: 28 km W from Tulcán, road Tulcán-Maldonado (QCAZ 13642). Imbabura: road Otavalo-
Laguna de Mojanda, 3358 m (MZUTI 1803–1806); road Otavalo-Laguna de Mojanda, 3557 m (MZUTI 1808–1815).
Pichincha: Reserva Verdecocha (MZUTI 1785–1802, 1816–1824, 1867–1872).
Pristimantis lucidosignatus. Ecuador: Cotopaxi: Otonga (QCAZ 30266); Naranjito, Bosque Integral Otonga (QCAZ 31814,
31816, 31825).
Pristimantis myersi. Colombia: Nariño: Páramo El Tábano (ICN 2503); municipality of Cumbal, km 16–17 Chile-San Felipe,
northern slope volcano Chiles (ICN 24337–24340). Cauca: PNN Nevado del Huila, cabaña Inderena (ICN 6484, 6500);
Puracé, km 55 road Popayán-La Plata, PNN Puracé (ICN 25908–25910); laguna San Rafael, cabaña San Rafael del

ROJAS-RUNJAIC ET AL.50 · Zootaxa 3780 (1) © 2014 Magnolia Press
Inderena (ICN 33200–33204). Ecuador: Imbabura: Laguna Puruhanta (QCAZ 11677); Nueva América (QCAZ 14554–
14560). Sucumbios: El Playón (QCAZ 14561–14562).
Pristimantis ocreatus. Ecuador: Carchi: in the way to Tulcán Maldonado, 15 minuts to west of Tufiño (QCAZ 43162).
Imbabura: Lagunas de Mojanda (QCAZ 42111).
Pristimantis onorei. Ecuador: Cotopaxi: Naranjito, Bosque Integral Otonga (QCAZ 11700, 11862, 12296, 12297–12299,
12303, 12969–12971, 12974–12978, 31813, 31815, 31817–31823, 31826); Naranjito, farm of Don Tomás Granja (QCAZ
32924–32925).
Pristimantis pyrrhomerus. Ecuador: Cotopaxi: Sigchos, road Unache-Santa Rosa (MZUTI 1925–1930); 3 km E Pilaló, around
the waterfall (MZUTI 1941–1943); 1 km E Pilaló (KU 177837).
Pristimantis repens. Colombia: Nariño: Pasto, km 11 Pasto-volcano Galeras (ICN 12323 [holotype], 12324–12329
[paratypes]); Pasto, km 12–14 Pasto-volcano Galeras (ICN 12333, 12335–12339, 12341–12342, 12345, 12351
[paratypes]); Pasto km 23 (ICN 12354 [paratype]); Pasto, location El Campinero (ICN 12356–12359 [paratype]); Pasto,
carretera Panamericana, km 11 road to Río Bobo dam (ICN 12363 [paratype]).
Pristimantis scopaeus. Colombia: Tolima, Cajamarca, Amaime, Páramo de los Valles (ICN 22792 [holotype], 22789–22790,
22792, 22834 [paratypes]).
Pristimantis sirnigeli. Ecuador: Pichincha: Reserva Verdecocha (MZUTI 1825–1827).
Pristimantis xeniolum. Colombia: Cauca: Río Frio, Cerro Calima, Páramo del Duende (ICN 43891 [holotype], 43877
[paratype]).