a hairpin rna construct residing in an intron efficiently triggered rna-directed dna methylation in...
TRANSCRIPT

A hairpin RNA construct residing in an intron efficientlytriggered RNA-directed DNA methylation in tobacco
Athanasios Dalakouras1,2, Mirko Moser1, Michele Zwiebel1, Gabi Krczal1, Rudiger Hell2 and Michael Wassenegger1,2,*
1RLP AgroScience GmbH, AlPlanta-Institute for Plant Research, 67435 Neustadt, Germany, and2Heidelberg Institute for Plant Sciences, University of Heidelberg, 69120 Heidelberg, Germany
Received 23 July 2009; revised 10 August 2009; accepted 12 August 2009; published online 14 September 2009.*For correspondence (fax +49 6321 671 1313; e-mail [email protected]).
SUMMARY
So far, conventional hairpin RNA (hpRNA) constructs consisting of an inverted repeat (IR) of target promoters
directly introduced into an expression cassette have been used to mediate de novo DNA methylation.
Transcripts of such constructs resemble mRNA molecules, and are likely to be exported to the cytoplasm. The
presence of hpRNAs in the cytoplasm and the nucleus may account for the simultaneous activation of post-
transcriptional gene silencing (PTGS) and RNA-directed DNA methylation (RdDM). We hypothesized that by
retaining hpRNAs in the nucleus, efficient induction of only RdDM may be achieved. Thus, we introduced into
tobacco a transgene containing an intron into which an IR of a target promoter was inserted. The intronic
hpRNA initiated highly specific cis- and trans-methylation, but did not induce PTGS. No spreading of
methylation into sequences flanking the region of homology between the hpRNA and the target DNA was
detectable. The efficient methylation-directing activity of the intronic hpRNA may indicate a previously
unrecognized role of introns, potentially regulating gene expression at the transcriptional level.
Keywords: hairpin RNA, intron, RNA-directed DNA methylation (RdDM), RNA interference (RNAi), small RNAs.
INTRODUCTION
In plants, RNA-directed DNA methylation (RdDM) is a key
process of nuclear RNA interference (RNAi) that is primarily
associated with epigenetic gene silencing (Wassenegger,
2005; Huettel et al., 2007; Matzke et al., 2007, 2009; Wierzb-
icki et al., 2008). RdDM is induced by double-stranded RNA
(dsRNA) and, in concert with numerous proteins, leads to
de novo cytosine methylation at symmetric CpG/CpHpG and
asymmetric CpHpH sites (where H = A, T or G) (Matzke et al.,
2009).
RNA polymerase V (Pol V) is a central component of the
de novo methylation complex. The SWI/SNF-like chromatin
remodelling protein, DRD1, and the hinge domain-contain-
ing protein, DSM3, appear to recruit Pol V to chromatin,
although DRD1 does not physically interact with Pol V
(Kanno et al., 2004, 2008; Wierzbicki et al., 2009). As Pol V
does not utilize RNA templates, and is not specifically
targeted to methylated DNA, it was suggested that the
initiation of Pol V transcription occurs throughout the entire
genome. Binding of small complementary RNAs to Pol
V-produced transcripts would then lead to the sequence-
specific directing of the de novo methyltransferase(s) to the
target DNA (Pontier et al., 2005; Wierzbicki et al., 2008; ).
Indeed, in plant nuclei, 24-nucleotide (24-nt) short interfering
RNAs (siRNAs) are processed from dsRNA by Dicer-like 3
(DCL3), and are predominantly loaded onto Argonaute 4
(AGO4) (Zilberman et al., 2003; Chan et al., 2004). The 24-nt
siRNAs are generally presumed to guide the RdDM machin-
ery (Wierzbicki et al., 2009). In concert with the KOW
domain-containing transcription factor 1 (KTF1), AGO4
could either directly interact with Pol V or would associate
with Pol V via siRNA-mediated Pol V transcript binding
(El-Shami et al., 2007; He et al., 2009).
The hierarchical order of processes involved in the self-
reinforcing feedback mechanism is unclear. An SNF2
domain-containing protein (CLSY1), Pol IV, Pol V, RNA-
directed RNA polymerase 2 (RDR2), DCL3 and AGO4 were
found, although to variable extents, to be connected with the
accumulation of 24-nt siRNAs (Chan et al., 2004; Smith et al.,
2007; Mosher et al., 2008). The latter three proteins and the
NRPE1 subunit of Pol V were detectable in the nucleolus-
associated Cajal bodies, indicating that they are siRNA
processing centres (Li et al., 2006; Pontes et al., 2006). Using
RNA templates, probably generated by Pol IV, RDR2 is
producing dsRNA that is processed into secondary 24-nt
siRNAs by DCL3. (Kanno et al., 2005; Zhang et al., 2007;
Mosher et al., 2008). Yet, the substrate for Pol IV needs to be
840 ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd
The Plant Journal (2009) 60, 840–851 doi: 10.1111/j.1365-313X.2009.04003.x

identified. The currently available data appear to be incon-
sistent (Pikaard et al., 2008). There is good evidence that
Pol IV is essential for the biosynthesis of the majority of
endogenous 24-nt siRNAs. However, if 24-nt siRNAs guide
the de novo methylation machinery, the RdDM-initiating 24-
nt-siRNAs (primary 24-nt siRNAs) cannot be processed from
precursors that are produced by Pol IV-mediated transcrip-
tion of de novo methylated DNA. Thus, primary 24-nt siRNA
biosynthesis must be upstream of Pol V-dependent Pol IV
transcription. Alternatively, one may speculate that RNA
molecules other than 24-nt siRNAs are involved in the
de novo RdDM step.
So far, there is no direct evidence demonstrating that 24-
nt siRNAs guide the RdDM machinery. Daxinger and co-
workers (2009) reported that in an Arabidopsis thaliana dcl3
mutant, RdDM was impaired but not completely lost.
However, even the full inhibition of RdDM would not provide
clear evidence for a direct involvement of the 24-nt siRNAs in
guiding the RdDM machinery. One may speculate that,
instead, they are essential in an intermediate step for the
production of RdDM-guiding molecules. It is likely that the
central role of the 24-nt siRNAs is to reinforce rather than
initiate RdDM. This view is supported by the observation
that inverted repeat (IR)-mediated initiation of de novo DNA
methylation was unaffected in an A. thaliana ago4 mutant
(Zilberman et al., 2004). In ago4-1, maintenance of methyl-
ation was severely impaired, suggesting that AGO4 and
thereby 24-nt siRNAs play a vital role in the maintenance of
RdDM. However, at that time the redundant activity of AGO6
was not known (Zheng et al., 2007). Although AGO6
appeared to generally play only a minor role in RdDM, that
AGO4 was partially replaced by AGO6 in the ago4-1 line
cannot be ruled out. Notably, asymmetric DNA methylation
at a few genomic loci was reported to take place in the
absence of 24-nt siRNAs, implying a mechanism that is
independent of these RNAs (Henderson et al., 2006). How-
ever, it should be noted that diverse RdDM pathways are
likely to exist. Defined chromatin structures (hetero-/euchro-
matin), DNA arrangements (tandem repeats/IRs) and the
sequence context (CpG-rich/CpG-depleted) may require the
involvement of specific RdDM components (Fischer et al.,
2008; Luo et al., 2009).
Initiation of RdDM has been most extensively studied
using transgenic plants. Viroid and virus infections led to the
de novo methylation of transgenes sharing sequence
homology with the pathogen RNAs. Because replication of
the pathogens proceeded without the involvement of DNA,
it became evident that de novo DNA methylation was
initiated by pathogen-derived RNA molecules (Wassenegger
et al., 1994a; Jones et al., 1998). In plants, post-transcrip-
tional gene silencing (PTGS) was found to be generally
accompanied by de novo methylation of the transgene
coding region, indicating that both processes are related
(Wassenegger and Pelissier, 1998). In fact, similarly to PTGS,
RdDM is initiated by dsRNA (Mette et al., 2000). Various
processes can lead to the production of dsRNA, including
RNA virus or viroid replication and transcription of IRs.
However, to initiate RdDM, the trigger RNA needs to be
present in the nucleus.
In this study, we examined the feasibility of using an
intronic hairpin RNA (int-hpRNA) to mediate RdDM. Intron
sequences that are removed during the splicing process
were reported to remain in the nucleus (Qian et al., 1992).
Thus, we speculated that an int-hpRNA that is retained in the
nucleus could be efficiently processed into RdDM-initiating
24-nt siRNA, but not into PTGS-inducing 21-nt siRNAs.
However, only a few tobacco plant lines expressing the int-
hpRNA transgene construct produced detectable quantities
of int-hpRNA-derived 24-nt siRNAs. In addition, accumula-
tion of transgenic siRNAs of other size classes was not
detected in any lines. The absence of the 21-nt siRNAs was in
agreement with the failure of the int-hpRNA to initiate PTGS
of a sensor transgene. Despite the low accumulation of 24-nt
siRNAs the transgene containing the int-hpRNA construct
became heavily de novo methylated. Hypermethylation was
specifically found in the region that was homologous with
the int-hpRNA. Moreover, the introduction of a sensor
transgene into the int-hpRNA-expressing plants revealed
efficient trans-methylation. As was found for the trigger
transgene, only the part of the sensor transgene sharing
homology with the int-hpRNA became de novo methylated.
No spreading of methylation into flanking regions, as
previously reported, was observed (Kanno et al., 2008;
Daxinger et al., 2009). Our data show that int-hpRNA
constructs could serve as a tool to initiate efficient and
highly specific de novo DNA trans-methylation. Further-
more, they indicate that in plants, a natural intron-based
transcriptional gene silencing (TGS) mechanism may exist.
RESULTS
Generation of the GFP-int-hpRNACMPS construct and
introduction into tobacco
We hypothesized that a system predominantly enabling the
initiation of nuclear but not cytoplasmic RNAi would require
an RdDM trigger that is retained in the nucleus. As spliced
intron sequences are proposed to generally stay in the
nucleus (Qian et al., 1992), we inserted an hpRNA into an
intron. However, it was unknown whether an hpRNA would
impair the splicing process, and if int-hpRNAs could be
processed by the nuclear RNAi machinery.
The primary transgene construct, GFPint, comprised the
Solanum esculentum RNA-directed RNA polymerase 1
(SeRDR1) intron 3 inserted into the GFP5 cDNA (Haseloff
et al., 1997) (Figure S1). As an RdDM trigger, an hpRNA
targeting the promoter of a reporter gene was selected. The
hpRNA construct contained a 240-bp antisense and a 215-bp
sense fragment of the Cestrum yellow leaf curling virus
Intronic hairpin-mediated RdDM 841
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2009), 60, 840–851
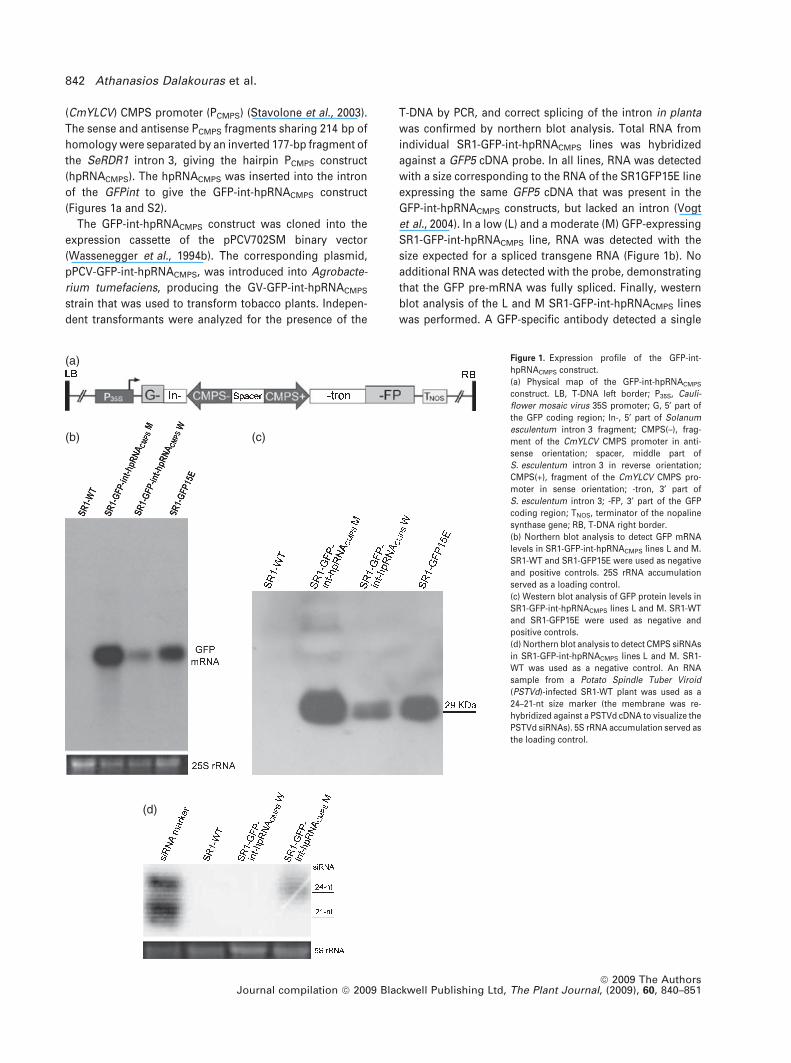
(CmYLCV) CMPS promoter (PCMPS) (Stavolone et al., 2003).
The sense and antisense PCMPS fragments sharing 214 bp of
homology were separated by an inverted 177-bp fragment of
the SeRDR1 intron 3, giving the hairpin PCMPS construct
(hpRNACMPS). The hpRNACMPS was inserted into the intron
of the GFPint to give the GFP-int-hpRNACMPS construct
(Figures 1a and S2).
The GFP-int-hpRNACMPS construct was cloned into the
expression cassette of the pPCV702SM binary vector
(Wassenegger et al., 1994b). The corresponding plasmid,
pPCV-GFP-int-hpRNACMPS, was introduced into Agrobacte-
rium tumefaciens, producing the GV-GFP-int-hpRNACMPS
strain that was used to transform tobacco plants. Indepen-
dent transformants were analyzed for the presence of the
T-DNA by PCR, and correct splicing of the intron in planta
was confirmed by northern blot analysis. Total RNA from
individual SR1-GFP-int-hpRNACMPS lines was hybridized
against a GFP5 cDNA probe. In all lines, RNA was detected
with a size corresponding to the RNA of the SR1GFP15E line
expressing the same GFP5 cDNA that was present in the
GFP-int-hpRNACMPS constructs, but lacked an intron (Vogt
et al., 2004). In a low (L) and a moderate (M) GFP-expressing
SR1-GFP-int-hpRNACMPS line, RNA was detected with the
size expected for a spliced transgene RNA (Figure 1b). No
additional RNA was detected with the probe, demonstrating
that the GFP pre-mRNA was fully spliced. Finally, western
blot analysis of the L and M SR1-GFP-int-hpRNACMPS lines
was performed. A GFP-specific antibody detected a single
(a)
(c)
(d)
(b)
Figure 1. Expression profile of the GFP-int-
hpRNACMPS construct.
(a) Physical map of the GFP-int-hpRNACMPS
construct. LB, T-DNA left border; P35S, Cauli-
flower mosaic virus 35S promoter; G, 5¢ part of
the GFP coding region; In-, 5¢ part of Solanum
esculentum intron 3 fragment; CMPS(–), frag-
ment of the CmYLCV CMPS promoter in anti-
sense orientation; spacer, middle part of
S. esculentum intron 3 in reverse orientation;
CMPS(+), fragment of the CmYLCV CMPS pro-
moter in sense orientation; -tron, 3¢ part of
S. esculentum intron 3; -FP, 3¢ part of the GFP
coding region; TNOS, terminator of the nopaline
synthase gene; RB, T-DNA right border.
(b) Northern blot analysis to detect GFP mRNA
levels in SR1-GFP-int-hpRNACMPS lines L and M.
SR1-WT and SR1-GFP15E were used as negative
and positive controls. 25S rRNA accumulation
served as a loading control.
(c) Western blot analysis of GFP protein levels in
SR1-GFP-int-hpRNACMPS lines L and M. SR1-WT
and SR1-GFP15E were used as negative and
positive controls.
(d) Northern blot analysis to detect CMPS siRNAs
in SR1-GFP-int-hpRNACMPS lines L and M. SR1-
WT was used as a negative control. An RNA
sample from a Potato Spindle Tuber Viroid
(PSTVd)-infected SR1-WT plant was used as a
24–21-nt size marker (the membrane was re-
hybridized against a PSTVd cDNA to visualize the
PSTVd siRNAs). 5S rRNA accumulation served as
the loading control.
842 Athanasios Dalakouras et al.
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2009), 60, 840–851
29-kDa protein corresponding to the GFP expressed in the
SR1GFP15E line (Figure 1c). In summary, the SeRDR1
intron 3 proved to be functional, and was efficiently and
accurately spliced, even though it contained the hpRNACMPS
fragment.
Processing of the GFP-int-hpRNACMPS into siRNAs
Total RNA from the SR1-GFP-int-hpRNACMPS L and M lines
was used for siRNA analysis. Northern blots using a PCMPS-
specific probe detected a signal in line M samples corre-
sponding to 24-nt-long siRNAs only (Figure 1d). No 21-nt
PCMPS siRNAs or siRNAs of other size classes were detect-
able. The 24-nt siRNAs were not detected in line L
(Figure 1d).
Transgene transcripts can induce the production of
siRNAs. However, expression of the GFP-int-hpRNACMPS
transgene represents a rare example in which a Pol II-
transcribed transgenic RNA is processed into 24-nt siRNAs,
but not into siRNAs of other size classes. Agroinfiltration of
SR1-GFP-int-hpRNACMPS plants using an A. tumefaciens
strain containing a ‘classical’ GFP-hairpin construct (pPCV-
GpG) produced both sizes of GFP siRNAs, indicating that
biosynthesis of 21-nt siRNA was not affected in SR1-GFP-int-
hpRNACMPS lines (Figure S3). Finally, the fact that no GFP
siRNAs were detectable in mock-infiltrated SR1-GFP-int-
hpRNACMPS lines demonstrated that GFP silencing was not
induced. This finding was in accordance with the absence
of GFP mRNA degradation products in SR1-GFP-int-
hpRNACMPS lines (Figure 1b). In summary, our data indicate
that processing of the GFP-int-hpRNACMPS into siRNAs was
exclusively catalyzed by the nuclear RNAi machinery.
Moreover, the absence of GFP siRNAs suggested that
nuclear RNAi was not associated with transitive silencing.
Notably, transgene intron-specific siRNAs, including loop-
specific siRNAs, were also absent, supporting the view that
no transitive silencing had taken place.
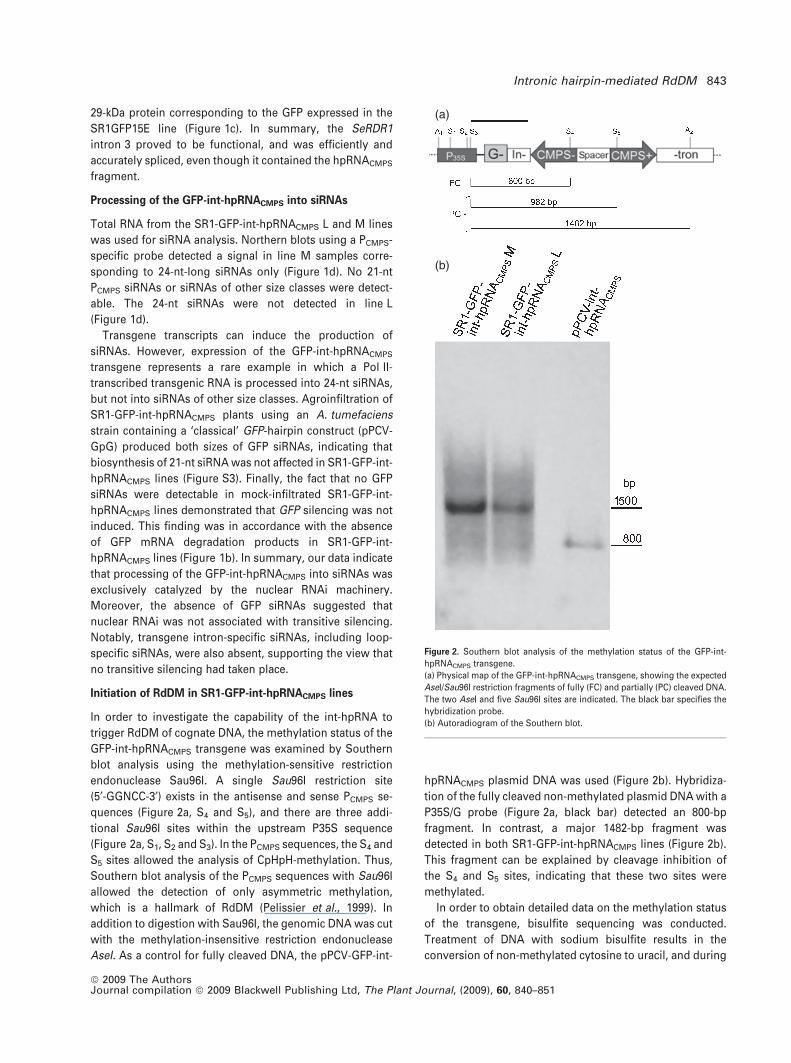
Initiation of RdDM in SR1-GFP-int-hpRNACMPS lines
In order to investigate the capability of the int-hpRNA to
trigger RdDM of cognate DNA, the methylation status of the
GFP-int-hpRNACMPS transgene was examined by Southern
blot analysis using the methylation-sensitive restriction
endonuclease Sau96I. A single Sau96I restriction site
(5¢-GGNCC-3¢) exists in the antisense and sense PCMPS se-
quences (Figure 2a, S4 and S5), and there are three addi-
tional Sau96I sites within the upstream P35S sequence
(Figure 2a, S1, S2 and S3). In the PCMPS sequences, the S4 and
S5 sites allowed the analysis of CpHpH-methylation. Thus,
Southern blot analysis of the PCMPS sequences with Sau96I
allowed the detection of only asymmetric methylation,
which is a hallmark of RdDM (Pelissier et al., 1999). In
addition to digestion with Sau96I, the genomic DNA was cut
with the methylation-insensitive restriction endonuclease
AseI. As a control for fully cleaved DNA, the pPCV-GFP-int-
hpRNACMPS plasmid DNA was used (Figure 2b). Hybridiza-
tion of the fully cleaved non-methylated plasmid DNA with a
P35S/G probe (Figure 2a, black bar) detected an 800-bp
fragment. In contrast, a major 1482-bp fragment was
detected in both SR1-GFP-int-hpRNACMPS lines (Figure 2b).
This fragment can be explained by cleavage inhibition of
the S4 and S5 sites, indicating that these two sites were
methylated.
In order to obtain detailed data on the methylation status
of the transgene, bisulfite sequencing was conducted.
Treatment of DNA with sodium bisulfite results in the
conversion of non-methylated cytosine to uracil, and during
(a)
(b)
Figure 2. Southern blot analysis of the methylation status of the GFP-int-
hpRNACMPS transgene.
(a) Physical map of the GFP-int-hpRNACMPS transgene, showing the expected
AseI/Sau96I restriction fragments of fully (FC) and partially (PC) cleaved DNA.
The two AseI and five Sau96I sites are indicated. The black bar specifies the
hybridization probe.
(b) Autoradiogram of the Southern blot.
Intronic hairpin-mediated RdDM 843
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2009), 60, 840–851

PCR amplification uracil is replaced by thymine. Thus,
sequences of PCR products from bisulfite-treated DNA
exhibit thymines for non-methylated cytosines. A mixed
plasmid/tobacco DNA sample and 2 lg of genomic DNA
pooled from the SR1-GFP-int-hpRNACMPS L and M lines
were subjected to bisulfite sequencing. Two transgene
fragments were analyzed: the 254-bp fragment (CMPS254)
containing parts of the 5¢ GFP5, the 5¢ intron and the PCMPS
antisense sequences, and the 360-bp fragment (CMPS360)
containing the 3¢ spacer, the PCMPS sense and a part of the 3¢intron sequences (Figure 3). Bisulfite sequencing is strand-
specific. The upper strand was analyzed using the primer
pairs BisF2/BisR2 for CMPS254 amplification and BisF3/BisR3
for CMPS360 amplification. Sequencing of cloned PCR prod-
ucts revealed that inserts originating from the pPCV-GFP-int-
hpRNACMPS plasmid that was not methylated were fully
converted (Figure 3, s). In contrast, sense and antisense
PCMPS sequences amplified from the plant genome-inte-
grated transgene were methylated to almost 100% (Figure 3,
). Importantly, methylation did not spread into sequences
flanking the PCMPS-specific sequences. All cytosines within
the intron, GFP5 and spacer were found to be converted in all
clones (Figure 3, h). It should be noted that in the first set of
experiments, DNA material from lines M and L was pooled
to avoid sample bias. However, methylation patterns were
identical when samples of the two lines were separately
analyzed.
To validate the bisulfite sequencing data, restriction
analysis of the PCR products obtained from amplified
bisulfite-treated DNA was also performed. If a cytosine
residue located in a restriction site of the authentic sequence
is converted, the site will be lost. Thus, PCR products
amplified from this DNA will lack the restriction site, and so
analysis of the PCR products will give an overview of the
methylation status of the tissue from which the genomic
DNA was extracted. The results of this analysis (Figure S4)
confirmed the Southern and bisulfite sequencing data,
and altogether clearly demonstrated that the GFP-int-
hpRNACMPS construct efficiently initiated RdDM. Methyla-
tion was highly specific for the sequences that shared
homology with the double-stranded region of GFP-int-
hpRNACMPS. Neither the spacer nor the intron or GFP5
sequences flanking the sense and antisense copy of the
PCMPS were targeted by RdDM.
GFP-int-hpRNACMPS efficiently triggered trans-methylation
of a sensor construct
GFP-int-hpRNACMPS induced the specific methylation of the
genome-integrated transgene region from which the hpRNA
was transcribed. To explore whether, in addition to this
cis-methylation, the GFP-int-hpRNACMPS had the capacity to
trigger trans-methylation, a ‘methylation-sensor’ construct
was introduced into tobacco. The construct comprised a 791-
bp red-shifted GFP (rsGFP) cDNA driven by the full-length
Figure 3. Bisulfite sequencing data of the GFP-int-hpRNACMPS transgene.
Physical map of the GFP-int-hpRNACMPS transgene, in which the regions analyzed by bisulfite sequencing are indicated. Abbreviations are as in Figure 1a. The 254-
bp-long (CMPS254) and the 360-bp-long (CMPS360) transgene fragments were separately analyzed. CMPS254 comprised parts of the 5¢ GFP, the 5¢ intron (underlined)
and the 5¢ CMPS(–) (bold) sequences. CMPS360 comprised parts of the spacer (lower case), the full-length CMPS(+) (bold) and the 3¢ intron (underlined) sequences.
Circles represent bisulfite sequencing data produced with pPCV-GFP-int-hpRNACMPS plasmid DNA, and squares represent those produced with SR1-GFP-int-
hpRNACMPS genomic DNA. The percentage levels of cytosine methylation are indicated by open circles and squares (0–10%), by half-filled symbols (40–60%) and by
filled symbols (90–100%). Detailed sequencing data for each clone are presented in Figure S8. The XbaI (5¢-TCTAGA-3¢) and HpyCH4IV (5¢-ACGT-3¢) restriction sites
used for restriction analysis of the bisulfite PCR product (Figure S4) are set in italic font, and are indicated by a gray and open box, respectively.
844 Athanasios Dalakouras et al.
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2009), 60, 840–851

406-bp PCMPS (PCMPS-rsGFP) (Figures 4 and S5). SR1-PCMPS-
rsGFP plant lines moderately expressing rsGFP were identi-
fied by northern blot analysis. No PCMPS siRNAs were
detectable in these lines. The SR1-PCMPS-rsGFP line 1 was
genetically crossed with the SR1-GFP-int-hpRNACMPS line M
to generate the SR1-PCMPS/int-hpRNACMPS line 1. T1 progeny
plants of this line, in which the presence of both transgenes
was verified by PCR, were used for further experiments.
In order to clarify if the presence of the GFP-int-
hpRNACMPS transgene led to the trans-methylation of the
PCMPS sequence, bisulfite sequencing was performed. A 226-
bp fragment (CMPS226) was analyzed, of which the 3¢ half
overlapped with the GFP-int-hpRNACMPS (Figure 4, upper
panel). DNA from three independent T1 plants of the
SR1-PCMPS/int-hpRNACMPS line 1 was pooled, bisulfite trea-
ted and amplified with the BisF1/BisR1 primer pair. The
pPCV800-PCMPS-rsGFP plasmid and genomic DNA from a T1
plant of the SR1-PCMPS-rsGFP line 1 served as controls. In the
controls, CMPS226 was virtually free of methylation (Fig-
ure 4, upper panel, s and h) whereas in the SR1-PCMPS/int-
hpRNACMPS progeny, the 3¢ half of CMPS226 overlapping
with GFP-int-hpRNACMPS was fully methylated (Figure 4,
upper panel, sequence in bold). Importantly, in the double
transformants, the non-overlapping part of PCMPS was not
methylated, demonstrating that the spread of methylation
into the 5¢ flanking region had not taken place. This finding
was validated by restriction analysis of PCR products from
bisulfite-treated and untreated DNA (Figure S6a).
So far, the spread of RdDM-mediated methylation along
promoter sequences was only found to proceed into
downstream sequences (Kanno et al., 2008; Daxinger et al.,
2009). Thus, we analyzed the junction between the PCMPS
and the rsGFP sequences of the PCMPS-rsGFP transgene by
bisulfite sequencing. The 354-bp CMPS354 fragment was
amplified using the BisF4/BisR4 primer pair from bisul-
fite-treated DNA of the pPCV800-PCMPS-rsGFP plasmid, the
SR1-PCMPS-rsGFP line 1 and from SR1-PCMPS/int-
hpRNACMPS T1 plants (Figure 4, lower panel). Sequencing
of individual clones containing PCR products from the
pPCV800-PCMPS-rsGFP plasmid and the SR1-PCMPS-rsGFP
line 1 revealed that the region corresponding to CMPS354
was not methylated. In contrast, the region of the CMPS354
sequence overlapping with int-hpRNACMPS was fully methy-
lated in SR1-PCMPS/int-hpRNACMPS plants. However, as was
found for cis-methylation of the GFP-int-hpRNACMPS trans-
gene, trans-methylation also appeared to decline immedi-
ately downstream of the int-hpRNACMPS/CMPS354 overlap.
Virtually no 5mC was detectable downstream of the junc-
tion between the PCMPS and the rsGFP sequence (Figure 4,
lower panel). This finding was again validated by restriction
analysis of PCR products from bisulfite-treated and
untreated DNA (Figure S6b,c).
Figure 4. Bisulfite sequencing data of the PCMPS-rsGFP sensor construct.
Physical map of the PCMPS-rsGFP transgene, in which the regions analyzed by bisulfite sequencing are indicated. The 226-bp-long (CMPS226) and the 354-bp-long
(CMPS354) transgene fragments were separately analyzed. CMPS226 comprised the middle part of the full-length CmYLCV CMPS promoter (PCMPS). CMPS354
comprised the 3¢ part of the PCMPS and the 5¢ rsGFP sequences. The bold sequences indicate regions of complementarity between int-hpRNACMPS (methylation
trigger) and PCMPS (methylation target). Circles represent bisulfite sequencing data produced with pPCV-PCMPS-rsGFP plasmid DNA, squares represent those
produced with SR1-PCMPS-rsGFP genomic DNA and triangles represent those produced with SR1-PCMPS/int-hpRNACMPS genomic DNA. The percentage levels of
cytosine methylation are indicated by open symbols (0–10%), and by filled symbols (90–100%). Detailed sequencing data for each clone are presented in Figure S8.
The guanidine of transcriptional start site (TSS) is boxed in dark gray. The BsgI (5¢-GTGCAG(N)16-3¢), Hpy188I (5¢-TCNGA-3¢) and EagI (5¢-CGGCCG-3¢) restriction sites
used for restriction analysis of the PCR products (Figure S6) are all set in italic font, and are underlined or indicated by a gray and open box, respectively.
Abbreviations are as in Figure 1a; rsGFP, red-shifted GFP.
Intronic hairpin-mediated RdDM 845
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2009), 60, 840–851
It has been proposed that the spread of 3¢ methylation
involves a nascent transcript of the target. To investigate
whether such a transcript existed, and whether it had an
effect on 3¢ spread, we mapped the transcriptional start site
(TSS) of the PCMPS-rsGFP sensor using the 5¢ RACE analysis.
The TSS was located 60-bp upstream from the end of the
overlap between the trigger and the target sequence,
demonstrating that the 3¢ region of the target was tran-
scribed (Figure 4). Nevertheless, no spreading of methyla-
tion into the 3¢ flanking region was observed.
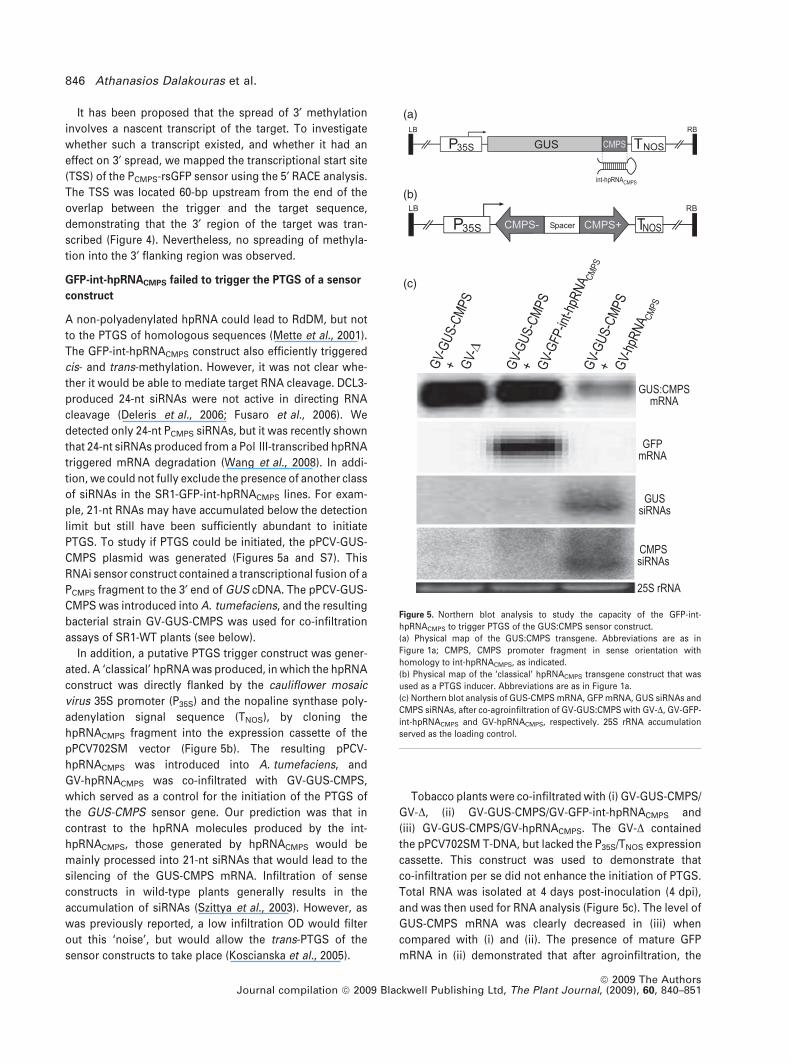
GFP-int-hpRNACMPS failed to trigger the PTGS of a sensor
construct
A non-polyadenylated hpRNA could lead to RdDM, but not
to the PTGS of homologous sequences (Mette et al., 2001).
The GFP-int-hpRNACMPS construct also efficiently triggered
cis- and trans-methylation. However, it was not clear whe-
ther it would be able to mediate target RNA cleavage. DCL3-
produced 24-nt siRNAs were not active in directing RNA
cleavage (Deleris et al., 2006; Fusaro et al., 2006). We
detected only 24-nt PCMPS siRNAs, but it was recently shown
that 24-nt siRNAs produced from a Pol III-transcribed hpRNA
triggered mRNA degradation (Wang et al., 2008). In addi-
tion, we could not fully exclude the presence of another class
of siRNAs in the SR1-GFP-int-hpRNACMPS lines. For exam-
ple, 21-nt RNAs may have accumulated below the detection
limit but still have been sufficiently abundant to initiate
PTGS. To study if PTGS could be initiated, the pPCV-GUS-
CMPS plasmid was generated (Figures 5a and S7). This
RNAi sensor construct contained a transcriptional fusion of a
PCMPS fragment to the 3¢ end of GUS cDNA. The pPCV-GUS-
CMPS was introduced into A. tumefaciens, and the resulting
bacterial strain GV-GUS-CMPS was used for co-infiltration
assays of SR1-WT plants (see below).
In addition, a putative PTGS trigger construct was gener-
ated. A ‘classical’ hpRNA was produced, in which the hpRNA
construct was directly flanked by the cauliflower mosaic
virus 35S promoter (P35S) and the nopaline synthase poly-
adenylation signal sequence (TNOS), by cloning the
hpRNACMPS fragment into the expression cassette of the
pPCV702SM vector (Figure 5b). The resulting pPCV-
hpRNACMPS was introduced into A. tumefaciens, and
GV-hpRNACMPS was co-infiltrated with GV-GUS-CMPS,
which served as a control for the initiation of the PTGS of
the GUS-CMPS sensor gene. Our prediction was that in
contrast to the hpRNA molecules produced by the int-
hpRNACMPS, those generated by hpRNACMPS would be
mainly processed into 21-nt siRNAs that would lead to the
silencing of the GUS-CMPS mRNA. Infiltration of sense
constructs in wild-type plants generally results in the
accumulation of siRNAs (Szittya et al., 2003). However, as
was previously reported, a low infiltration OD would filter
out this ‘noise’, but would allow the trans-PTGS of the
sensor constructs to take place (Koscianska et al., 2005).
Tobacco plants were co-infiltrated with (i) GV-GUS-CMPS/
GV-D, (ii) GV-GUS-CMPS/GV-GFP-int-hpRNACMPS and
(iii) GV-GUS-CMPS/GV-hpRNACMPS. The GV-D contained
the pPCV702SM T-DNA, but lacked the P35S/TNOS expression
cassette. This construct was used to demonstrate that
co-infiltration per se did not enhance the initiation of PTGS.
Total RNA was isolated at 4 days post-inoculation (4 dpi),
and was then used for RNA analysis (Figure 5c). The level of
GUS-CMPS mRNA was clearly decreased in (iii) when
compared with (i) and (ii). The presence of mature GFP
mRNA in (ii) demonstrated that after agroinfiltration, the
(a)
(b)
(c)
Figure 5. Northern blot analysis to study the capacity of the GFP-int-
hpRNACMPS to trigger PTGS of the GUS:CMPS sensor construct.
(a) Physical map of the GUS:CMPS transgene. Abbreviations are as in
Figure 1a; CMPS, CMPS promoter fragment in sense orientation with
homology to int-hpRNACMPS, as indicated.
(b) Physical map of the ‘classical’ hpRNACMPS transgene construct that was
used as a PTGS inducer. Abbreviations are as in Figure 1a.
(c) Northern blot analysis of GUS-CMPS mRNA, GFP mRNA, GUS siRNAs and
CMPS siRNAs, after co-agroinfiltration of GV-GUS:CMPS with GV-D, GV-GFP-
int-hpRNACMPS and GV-hpRNACMPS, respectively. 25S rRNA accumulation
served as the loading control.
846 Athanasios Dalakouras et al.
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2009), 60, 840–851

expression and processing of GFP-int-hpRNACMPS was not
impaired. Abundant PCMPS siRNAs were detected only when
the ‘classical’ hpRNACMPS construct, but not the GFP-int-
hpRNACMPS construct, was introduced. Accumulation of
GUS-specific secondary siRNAs was detectable only after
the co-infiltration of GV-GUS-CMPS with GV-hpRNACMPS,
but not with GV-GFP-int-hpRNACMPS. Our co-infiltration data
demonstrated that, in contrast to hpRNACMPS, GFP-int-
hpRNACMPS failed to trigger sensor mRNA degradation.
DISCUSSION
In plants, some miRNAs are known to reside within introns.
However, in contrast to our construct, they only triggered
PTGS (Voinnet, 2009). Compared with this, an intronic
transposon within FLC alleles is targeted by, and is probably
a source of, siRNAs that lead to heterochromatinization
(Mylne et al., 2006). Apart from these examples, a brief
bioinformatics-based analysis of the A. thaliana genome
provided no evidence for int-hpRNAs existing in nature.
However, so far, plants possessing larger genomes have not
been investigated. Recently, it was reported that several
thousand unique sequences exist in maize that could form
hairpins, but they were not miRNA precursors (Wang et al.,
2009). Whether such hairpins were located in introns is not
known.
Here, we demonstrate that, in tobacco, int-hpRNA trig-
gered the de novo cis- and trans-methylation of transgenes.
The efficiency of this system to trigger RdDM was remark-
able. Full methylation patterns were established already in
the primary transformants (cis-RdDM), and in the first
generations after crossing the trigger with a sensor
construct (trans-RdDM). This was in contrast to previous
findings, where more than three generations were required
(Aufsatz et al., 2002; Zilberman et al., 2004). Full methylation
patterns were homogeneous along the entire target
sequence, unlike previous reports where methylation was
most efficient near the IR center (Stam et al., 1998). Finally,
no spread of cis- and trans-methylation into 5¢ or 3¢ flanking
sequences took place, as observed previously (Daxinger
et al., 2009). After splicing, intron sequences are generally
not exported to the cytoplasm (Ernst et al., 1997; Zhou et al.,
2000). It is reasonable to assume that, as a part of the intron,
int-hpRNACMPS was also retained in the nucleus. The high
efficiency of the intronic system may thus be based on the
fact that int-hpRNACMPS was predominantly accessible to
the nuclear RNAi machinery.
Plant introns do not have absolute requirements for
branch sites and polypyrimidine tracts. An A/T-rich base
composition, a minimal size of 70-nt and the presence of
5¢-GT- and 3¢-AG- splice sites are sufficient for intron
functionality (Goodall and Filipowicz, 1991). We have ran-
domly chosen the site where the int-hpRNACMPS cDNA was
inserted into the intron. Secondary structures inside introns
could inhibit pre-mRNA splicing in dicotyledons and yeast.
Hairpin structures of only 18 or 24 bp had a strong inhibitory
effect on splicing in Nicotiana plumbaginifolia (Liu et al.,
1995). In the GFP-int-hpRNACMPS construct, the IR was
inserted 45-bp downstream of the 5¢ splice site. On the basis
of the currently available data on pre-mRNA processing, one
would have expected that the int-hpRNACMPS could interfere
with the splicing process. However, the secondary structure
of the hpRNACMPS had no obvious effect on splicing
accuracy or efficiency of the GFP-int-hpRNACMPS pre-mRNA.
Little is known about the decay of intron sequences after
splicing. Thus, it was not clear whether int-hpRNACMPS
would be accessible to the nuclear RNAi machinery, or
whether intron degradation would proceed faster than
int-hpRNACMPS processing by nuclear RNAi enzymes. The
observation that the int-hpRNACMPS of the SR1-GFP-int-
hpRNACMPS line M was processed into detectable quantities
of 24-nt siRNAs argued for the former case. However, the
concentration of int-hpRNACMPS 24-nt siRNAs in line M was
relatively low, and no int-hpRNACMPS siRNAs were found in
line L (Figure 1d). From our data, we could not conclude
whether the int-hpRNACMPS 24-nt siRNAs were primary
siRNAs directly generated from the int-hpRNACMPS by DCL3,
or whether they derived from a secondary siRNA production
pathway that may involve Pol IV and RDR2 activity.
Apart from 24-nt siRNAs, int-hpRNACMPS was apparently
not processed into 21-nt siRNAs or any other siRNA size
class. The absence of 21–22-nt siRNAs was in accordance
with the observation that the PTGS sensor was not silenced
in the presence of the GFP-int-hpRNACMPS construct. In
contrast, expression of the ‘classical’ hpRNACMPS led to the
degradation of the PTGS sensor mRNA, indicating that the
CMPS sequence was a target of RNAi (Figure 5c). In contrast
to 24-nt siRNAs that were derived from Pol II-transcribed
hpRNA, those produced from a Pol III-transcribed hpRNA
were PTGS competent (Fusaro et al., 2006; Wang et al.,
2008). This indicated that two types of 24-nt siRNAs exist
and/or that 24-nt siRNAs associate with diverse silencing
complexes, which could involve different AGO proteins.
AGO4, which is mainly associated with 24-nt siRNAs, has a
cleavage activity that is required for de novo methylation at
some loci (Qi et al., 2006). However, no cytoplasmic AGO4
activity has been reported so far. Thus, it is likely that 24-nt
siRNA-mediated PTGS involves siRNA loading onto a
different AGO protein.
The bisulfite data showed that in the SR1-GFP-int-
hpRNACMPS lines, transgene sequences sharing homology
with the double-stranded part of the hpRNACMPS displayed
essentially identical de novo methylation patterns. Accu-
mulation of different 24-nt siRNA levels, as found in these
lines, had no apparent effect on the density of target
sequence methylation. It is generally assumed that 24-nt
siRNAs guide the de novo DNA methylation machinery
(Chan et al., 2004; Wierzbicki et al., 2008; Daxinger et al.,
2009; Huang et al., 2009). Our findings raise the question of
Intronic hairpin-mediated RdDM 847
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2009), 60, 840–851

whether RNA molecules other than siRNAs direct methyl-
ation. As we observed here for line L, other reports
describe cases of RdDM in the absence of detectable
quantities of 24-nt siRNAs (Melquist and Bender, 2003;
Henderson et al., 2006; Lister et al., 2008). However, it can
not be excluded that siRNAs were produced, but to levels
that were under the detection limit. Ultra-deep sequencing
of the A. thaliana smRNAome revealed that siRNAs were
only associated with approximately a third of all genomic
cytosine methylation (Lister et al., 2008). In maize, many
siRNAs mapped to transposable elements (TEs) that were
not methylated, wereas TEs in which DNA was highly
methylated were relatively devoid of siRNAs (Wang et al.,
2009). These findings would argue against the requirement
of siRNAs for RdDM at some genomic loci. However, from
the available data it was not clear whether methylation of
regions that were not associated with siRNAs was based on
RdDM or the MET1-mediated maintenance of methylation.
Appearance of asymmetric methylation, the hallmark of
RdDM, within these regions would support the view that
siRNAs are not generally needed for the initiation of RdDM.
If 24-nt siRNAs guide the RdDM machinery, we would have
expected a denser methylation pattern in the SR1-GFP-int-
hpRNACMPS line M, accumulating 24-nt siRNAs, than in the
SR1-GFP-int-hpRNACMPS line L, where no siRNAs were
detected.
The regions of the SR1-GFP-int-hpRNACMPS lines and
SR1-PCMPS/int-hpRNACMPS progeny plants that became
methylated corresponded exclusively and precisely to
sequences sharing homology with the double-stranded
region of hpRNACMPS (Figures 3 and 4). This finding pointed
to a kind of ‘molecular ruler’ that would enable the
measurement of regions to be methylated at the nucleotide
level. An accurate ruler would be the double-stranded region
of int-hpRNACMPS or a-single-stranded RNA corresponding
to this region. The highly specific boundaries of methylation
observed here strongly argue against a function of 24-nt
siRNAs as rulers. It is unlikely that they precisely cover the
full length of the hpRNACMPS, as was also shown for a
transgenic nopaline synthase promoter fragment (Papp
et al., 2003). If the 24-nt siRNAs function as rulers, it must
be hypothesized that DCL3 processing started at the first and
stopped at the last nucleotide of the double-stranded region
of int-hpRNACMPS. In addition, accumulation of a homoge-
neous 24-nt siRNA population would help to explain the
equal distribution and density of methylation along the
targeted DNA. However, deep sequencing revealed that
thymine (T) is highly over-represented at position –1 of 24-nt
siRNA precursors, indicating that DCL3 processing occurs at
preferential sites. In addition, adenine (A) is the most
common first base of 24-nt siRNAs (Lister et al., 2008; Wang
et al., 2009). The preference for an A in the first position is in
accordance with the finding that in A. thaliana, 79% of
AGO4-associated 24-nt siRNAs exhibit a 5¢ terminal A (Mi
et al., 2008). These data indicated that 24-nt siRNAs are
unlikely to homogeneously map to the DNA that is targeted
by RdDM. Therefore, if 24-nt siRNAs were required to guide
the de novo DNA methylation machinery, one would expect
a more heterogeneous methylation pattern of the int-
hpRNACMPS target DNA. In summary, it is still unclear which
type of RNA molecules direct the de novo DNA methylation
machinery. In contrast to the general assumption, 24-nt
siRNAs may not be involved in the triggering of this process,
but in its amplification and maintenance.
So far, we have only generated a single int-hpRNA
construct. Thus, it can not be excluded that GFP-int-
hpRNACMPS-induced RdDM is a unique and artificial excep-
tion. The functionality of additional introns, RdDM triggers
and intron insertion sites must be tested to generalize our
findings. Currently, we are investigating the effect of a
nucleus-retained intronic hairpin on the induction of TGS of
transgenes and endogenous genes. In addition, we are
interested in further examining our finding of putative
selective Dicer processing of the intronic hairpin by subcel-
lular fractionation and siRNA/dsRNA detection. Finally, deep
sequencing of siRNAs and characterization of the GFP-int-
hpRNACMPS construct in a dcl2/dcl3/dcl4 triple mutant back-
ground will help to gain additional information about the
significance of 24-nt siRNAs in RdDM.
EXPERIMENTAL PROCEDURES
Plant transformation and agroinfiltration
All pPCV702SM binary vector derivatives were introduced into theA. tumefaciens strain GV3101. Nicotiana tabacum (cv. Petit HavanaSR1) plants were transformed using the leaf disc transformationmethod, as previously described (Wassenegger et al., 1994a), and/or were used for infiltration of N. tabacum leaves at an OD of 0.5, asdescribed previously (Koscianska et al., 2005).
Bisulfite analysis
Tobacco genomic DNA was isolated with the DNEasy Plant Mini Kit(Qiagen, http://www1.qiagen.com), and 2 lg of DraI-digested DNAwas bisulfite-treated with the EZ DNA Methylation Gold kit (ZYMOResearch, http://www.zymoresearch.com). The recovered materialwas amplified by PCR with AmpliTaq Gold polymerase (AppliedBiosystems, http://www.appliedbiosystems.com) under the follow-ing conditions: 10 min at 95�C, 45 amplification cycles (95�C for30 sec, 50�C for 30 sec and 72�C for 30 sec), and 72�C for 10 min.The CMPS226 was amplified with the BisF1/BisR1 primers, theCMPS254 with the BisF2/BisR2 primers and the CMPS360 with theBisF3/BisR3 primers (for primer sequences see Table S1). As apositive control for full cytosine conversion, 2 lg of SR1 wild-type(WT) DNA was mixed with approximately 40 pg of DraI-cleavedplasmid DNA, corresponding to the genome-integrated T-DNA. Theresulting PCR products were cloned into the pGEM-T Easy vector(Promega, http://www.promega.com), and 10 clones weresequenced from each experiment.
Plasmid construction
The details of all cloning strategies and the sequences of the primersused are shown in Figures S1–S7 and in Table S1, respectively. The
848 Athanasios Dalakouras et al.
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2009), 60, 840–851

pPCV-GFPint was generated by introducing a modified SeRDR1 in-tron 3 (accession number AC209589; base pairs 102578–103212) intothe GFP5 between nucleotides 375 and 376 of the cDNA (Figure S1).The intron was modified by introducing unique BglII and XbaI sites,which were subsequently used to insert the hairpin CMPS construct(hpRNACMPS). The hpRNACMPS was produced by amplifying a senseand an antisense CmYLCV CMPS promoter (PCMPS) fragment fromthe pNOV2820 plasmid (Syngenta, http://www.syngenta.com)(Figure S2). The two fragments were separated by a 120-bp frag-ment that was amplified from the SeRDR1 intron 3. The assembledhpRNACMPS was cloned into pT3T7-lac (Roche, http://www.roche.com), giving pTPCR-hpRNACMPS. From this plasmid, the hpRNACMPS
was released and introduced into pPCV-GFPint.The pPCV-GpG binary vector expressed an IR GFP5 construct
containing the second 143-bp Sau3AI fragment of the GFP5 cDNA(accession number U87973; base pairs 246–389) in sense andantisense orientation. The two fragments were separated by a 90-bpspacer that was derived from a Potato spindle tuber viroid cDNA.
The pPCV800-PCMPS-rsGFP plasmid was generated by modifyingthe pPCV702SM binary vector (Figure S5), whereby an additionalexpression cassette was introduced, comprising the full-lengthPCMPS, the red-shifted GFP (rsGFP; accession number FM883229)and the polyadenylation signal sequence of the A. tumefaciens tmrgene (pA-tmr) (accession number X00010; base pairs 1363–1755).
pPCV-GUS-CMPS was produced by fusing one of the PCMPS
fragments of the hpRNACMPS construct in sense orientation to the3¢ end of GUS cDNA of the pBI121 plasmid (Clontech, http://www.clontech.com). The GUS-CMPS transcriptional fusion productwas then introduced into the P35S/TNOS expression cassette ofpPCV702SM (Figure S7).
Northern, Southern and Western blot analysis
Total RNA was extracted from N. tabacum leaves with Tri-ReagentSolution (Ambion, http://www.ambion.com). For Northern blotanalysis, 10 lg of total RNA was separated on a 1.2% agaroseformaldehyde gel, capillary transferred onto a positively chargedmembrane BioBond Plus (Sigma-Aldrich, http://www.sigmaaldrich.com) and UV312nm-cross-linked (300 mJ cm)2). Random-primed[a-32P]dCTP-labeled PCR fragments (Random Primed DNA LabellingKit; Roche) were used as probes. The PerfectHyb Plus 1x (Sigma-Aldrich) was used for overnight hybridization at 64�C. Membraneswere washed at 64�C with buffer 1 (2x SSC, 0.1% SDS, w/v) for30 min, and with buffer 2 (0.5x SSC, 0.1% SDS, w/v) for 15 min.Membranes were exposed to FujiFilm Imaging Plates (FujiFilm,http://www.fujifilm.com) for 24 h and scanned using PharosFXPlus PhosphorImager (BioRad, http://www.bio-rad.com). For siRNAblot analysis, total RNA was extracted as above, and 20 lg wasseparated on a 20% Tris-Borate-EDTA (TBE)-acrylamide gel (Ana-med, http://www.anamed-gele.com) at 80 V for 4 h. RNA wastransferred onto positively charged nylon membranes (Ambion) byelectro-blotting at 300 mA for 1 h. Semidried membranes were UV-cross-linked as above. The appropriate PCR product for eachexperiment was labeled and purified as above. The hybridizationtemperature was 42�C, and membranes were washed once withbuffer 1 (2x SSC, 0.1% SDS, w/v) at 42�C for 30 min.
The DNEasy Plant Maxi kit (Qiagen) was used for plant DNAextraction, and Southern blot analyses were performed as previ-ously described (Vogt et al., 2004).
Western blot analysis was performed essentially as described bySambrook et al. (1989). Protein samples were separated on 9%SDS-polyacrylamide gels (Anamed) and transferred onto a polyv-inyldidene fluoride (PVDF) membrane (Roche). Membranes wereprobed with rabbit polyclonal anti-GFP (Santa-Cruz, http://
www.scbt.com). Primary antibodies were detected using a horse-radish peroxidase-labeled goat anti-rabbit IgG secondary antibody(Santa-Cruz).
5¢ RACE
The 5¢ cDNA ends were amplified using the GeneRacer Kit (Invi-trogen, http://www.invitrogen.com), following the manufacturer’sinstructions. The PCR reaction was performed using the GeneRacer5¢ primer and the gene-specific reverse primer RACE-R (Table S1).The PCR products were excised from the gel, cloned to pGEM-TEasy and sequenced.
ACKNOWLEDGEMENTS
We are grateful to Gunther Buchholz for providing the pMVA4fplasmid containing the red-shifted GFP cDNA, and to Detlef Weigeland Stefan Henz for providing the bioinformatics data on A. thali-ana. We thank Arno Putz, Milena Kitova and Marc Fullgrabe forassistance, Olivier Voinnet and Patrice Dunoyer for fruitful discus-sions, and Mike Haydon for critically reading the manuscript. Theexcellent care of plants in the glasshouse was provided by HeikoHerrmann. This work was supported by the grants of the Sixth Re-search Framework Programs of the European Union, Project LSHG-CT-2006-037900 (SIROCCO) and LSHG-CT-2004-005120 (FOSRAK).
SUPPORTING INFORMATION
Additional Supporting Information may be found in the onlineversion of this article:Figure S1. Cloning strategy for pPCV-GFPint.Figure S2. Cloning strategy for pPCV-GFP-int-hpRNACMPS.Figure S3. GV-GpG siRNAs.Figure S4. Verification of the CMPS254 and CMPS360 bisulfitesequencing data by restriction analysis of PCR products.Figure S5. Cloning strategy for pPCV800-PCMPS-rsGFP.Figure S6. Verification of the CMPS226 and CMPS354 bisulfitesequencing data by restriction analysis of PCR products.Figure S7. Cloning strategy for pPCV-GUS-CMPS.Figure S8. Detailed bisulfite sequencing data for CMPS254, CMPS360,CMPS226 and CMPS354.Table S1. Primers used for bisulfite sequencing and plasmidconstruction.Please note: Wiley-Blackwell are not responsible for the content orfunctionality of any supporting materials supplied by the authors.Any queries (other than missing material) should be directed to thecorresponding author for the article.
REFERENCES
Aufsatz, W., Mette, M.F., van der Winden, J., Matzke, A.J. and Matzke, M.
(2002) RNA-directed DNA methylation in Arabidopsis. Proc. Natl. Acad. Sci.
USA, 99(Suppl. 4), 16499–16506.
Chan, S.W., Zilberman, D., Xie, Z., Johansen, L.K., Carrington, J.C. and
Jacobsen, S.E. (2004) RNA silencing genes control de novo DNA methyl-
ation. Science, 303, 1336.
Daxinger, L., Kanno, T., Bucher, E., van der Winden, J., Naumann, U., Matzke,
A.J. and Matzke, M. (2009) A stepwise pathway for biogenesis of 24-nt
secondary siRNAs and spreading of DNA methylation. EMBO J. 28, 48–
57.
Deleris, A., Gallego-Bartolome, J., Bao, J., Kasschau, K.D., Carrington, J.C.
and Voinnet, O. (2006) Hierarchical action and inhibition of plant Dicer-like
proteins in antiviral defense. Science, 313, 68–71.
El-Shami, M., Pontier, D., Lahmy, S., Braun, L., Picart, C., Vega, D., Hakimi,
M.A., Jacobsen, S.E., Cooke, R. and Lagrange, T. (2007) Reiterated WG/
GW motifs form functionally and evolutionarily conserved ARGONA-
UTE-binding platforms in RNAi-related components. Genes Dev. 21,
2539–2544.
Intronic hairpin-mediated RdDM 849
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2009), 60, 840–851

Ernst, R.K., Bray, M., Rekosh, D. and Hammarskjold, M.L. (1997) A structured
retroviral RNA element that mediates nucleocytoplasmic export of intron-
containing RNA. Mol. Cell. Biol. 17, 135–144.
Fischer, U., Kuhlmann, M., Pecinka, M., Schmidt, R. and Mette, F. (2008) Local
DNA features affect RNA-directed transcriptional gene silencing and DNA
methylation. Plant J. 53, 1–10.
Fusaro, A.F., Matthew, L., Smith, N.A. et al. (2006) RNA interference-inducing
hairpin RNAs in plants act through the viral defence pathway. EMBO Rep. 7,
1168–1175.
Goodall, G.J. and Filipowicz, W. (1991) Different effects of intron nucleotide
composition and secondary structure on pre-mRNA splicing in monocot
and dicot plants. EMBO J. 10, 2635–2644.
Haseloff, J., Siemering, K.R., Prasher, D.C. and Hodge, S. (1997) Removal of a
cryptic intron and subcellular localization of green fluorescent protein are
required to mark transgenic Arabidopsis plants brightly. Proc. Natl. Acad.
Sci. USA, 94, 2122–2127.
He, X.J., Hsu, Y.F., Zhu, S., Wierzbicki, A.T., Pontes, O., Pikaard, C.S., Liu, H.L.,
Wang, C.S., Jin, H. and Zhu, J.K. (2009) An effector of RNA-directed DNA
methylation in arabidopsis is an ARGONAUTE 4- and RNA-binding protein.
Cell, 137, 498–508.
Henderson, I.R., Zhang, X., Lu, C., Johnson, L., Meyers, B.C., Green, P.J. and
Jacobsen, S.E. (2006) Dissecting Arabidopsis thaliana DICER function in
small RNA processing, gene silencing and DNA methylation patterning.
Nat. Genet. 38, 721–725.
Huang, L., Jones, A.M., Searle, I., Patel, K., Vogler, H., Hubner, N.C. and
Baulcombe, D.C. (2009) An atypical RNA polymerase involved in RNA
silencing shares small subunits with RNA polymerase II. Nat. Struct. Mol.
Biol. 16, 91–93.
Huettel, B., Kanno, T., Daxinger, L., Bucher, E., van der Winden, J., Matzke,
A.J. and Matzke, M. (2007) RNA-directed DNA methylation mediated by
DRD1 and Pol IVb: a versatile pathway for transcriptional gene silencing in
plants. Biochim. Biophys. Acta, 1769, 358–374.
Jones, A.L., Thomas, C.L. and Maule, A.J. (1998) De novo methylation and
co-suppression induced by a cytoplasmically replicating plant RNA virus.
EMBO J. 17, 6385–6393.
Kanno, T., Mette, M.F., Kreil, D.P., Aufsatz, W., Matzke, M. and Matzke, A.J.
(2004) Involvement of putative SNF2 chromatin remodeling protein DRD1
in RNA-directed DNA methylation. Curr. Biol. 14, 801–805.
Kanno, T., Huettel, B., Mette, M.F., Aufsatz, W., Jaligot, E., Daxinger, L., Kreil,
D.P., Matzke, M. and Matzke, A.J. (2005) Atypical RNA polymerase
subunits required for RNA-directed DNA methylation. Nat. Genet. 37, 761–
765.
Kanno, T., Bucher, E., Daxinger, L., Huettel, B., Bohmdorfer, G., Gregor, W.,
Kreil, D.P., Matzke, M. and Matzke, A.J. (2008) A structural-maintenance-
of-chromosomes hinge domain-containing protein is required for RNA-
directed DNA methylation. Nat. Genet. 40, 670–675.
Koscianska, E., Kalantidis, K., Wypijewski, K., Sadowski, J. and Tabler, M.
(2005) Analysis of RNA silencing in agroinfiltrated leaves of Nicotiana
benthamiana and Nicotiana tabacum. Plant Mol. Biol. 59, 647–661.
Li, C.F., Pontes, O., El-Shami, M., Henderson, I.R., Bernatavichute, Y.V., Chan,
S.W., Lagrange, T., Pikaard, C.S. and Jacobsen, S.E. (2006) An ARGONA-
UTE4-containing nuclear processing center colocalized with Cajal bodies in
Arabidopsis thaliana. Cell, 126, 93–106.
Lister, R., O’Malley, R.C., Tonti-Filippini, J., Gregory, B.D., Berry, C.C., Millar,
A.H. and Ecker, J.R. (2008) Highly integrated single-base resolution maps
of the epigenome in Arabidopsis. Cell, 133, 523–536.
Liu, H.X., Goodall, G.J., Kole, R. and Filipowicz, W. (1995) Effects of secondary
structure on pre-mRNA splicing: hairpins sequestering the 5¢ but not the 3¢splice site inhibit intron processing in Nicotiana plumbaginifolia. EMBO J.
14, 377–388.
Luo, C., Durgin, B., Watanabe, N. and Lam, E. (2009) Defining the functional
network of epigeentic regulators in Arabidopsis thaliana. Mol. Plant, 2, 661–
674.
Matzke, M., Kanno, T., Huettel, B., Daxinger, L. and Matzke, A.J. (2007)
Targets of RNA-directed DNA methylation. Curr. Opin. Plant Biol. 10,
512–519.
Matzke, M., Kanno, T., Daxinger, L., Huettel, B. and Matzke, A.J. (2009) RNA-
mediated chromatin-based silencing in plants. Curr. Opin. Cell Biol. 21, 1–10.
Melquist, S. and Bender, J. (2003) Transcription from an upstream promoter
controls methylation signaling from an inverted repeat of endogenous
genes in Arabidopsis. Genes Dev. 17, 2036–2047.
Mette, M.F., Aufsatz, W., van der Winden, J., Matzke, M.A. and Matzke, A.J.
(2000) Transcriptional silencing and promoter methylation triggered by
double-stranded RNA. EMBO J. 19, 5194–5201.
Mette, M.F., Matzke, A.J. and Matzke, M.A. (2001) Resistance of RNA-medi-
ated TGS to HC-Pro, a viral suppressor of PTGS, suggests alternative
pathways for dsRNA processing. Curr. Biol. 11, 1119–1123.
Mi, S., Cai, T., Hu, Y. et al. (2008) Sorting of small RNAs into Arabidopsis
argonaute complexes is directed by the 5¢ terminal nucleotide. Cell, 133,
116–127.
Mosher, R.A., Schwach, F., Studholme, D. and Baulcombe, D.C. (2008) PolIVb
influences RNA-directed DNA methylation independently of its role in
siRNA biogenesis. Proc. Natl. Acad. Sci. USA, 105, 3145–3150.
Mylne, J.S., Barrett, L., Tessadori, F., Mesnage, S., Johnson, L., Bernatavi-
chute, Y.V., Jacobsen, S.E., Fransz, P. and Dean, C. (2006) LHP1, the
Arabidopsis homologue of HETEROCHROMATIN PROTEIN1, is required
for epigenetic silencing of FLC. Proc. Natl. Acad. Sci. USA, 103, 5012–
5017.
Papp, I., Mette, F., Aufsatz, W., Daxinger, L., Schauer, S., Ray, A., van der
Winden, J., Matzke, M. and Matzke, A. (2003) Evidence for nuclear pro-
cessing of plant micro RNA and short interfering RNA precursors. Plant
Physiol. 132, 1382–1390.
Pelissier, T., Thalmeir, S., Kempe, D., Sanger, H.L. and Wassenegger, M.
(1999) Heavy de novo methylation at symmetrical and non-symmetrical
sites is a hallmark of RNA-directed DNA methylation. Nucleic Acids Res. 27,
1625–1634.
Pikaard, C.S., Haag, J.R., Ream, T. and Wierzbicki, A.T. (2008) Roles of RNA
polymerase IV in gene silencing. Trends Plant Sci. 13, 390–397.
Pontes, O., Li, C.F., Nunes, P.C., Haag, J., Ream, T., Vitins, A., Jacobsen, S.E.
and Pikaard, C.S. (2006) The Arabidopsis chromatin-modifying nuclear
siRNA pathway involves a nucleolar RNA processing center. Cell, 126, 79–
92.
Pontier, D., Yahubyan, G., Vega, D., Bulski, A., Saez-Vasquez, J., Hakimi,
M.A., Lerbs-Mache, S., Colot, V. and Lagrange, T. (2005) Reinforcement of
silencing at transposons and highly repeated sequences requires the
concerted action of two distinct RNA polymerases IV in Arabidopsis. Genes
Dev. 19, 2030–2040.
Qi, Y., He, X., Wang, X.J., Kohany, O., Jurka, J. and Hannon, G.J. (2006) Dis-
tinct catalytic and non-catalytic roles of ARGONAUTE4 in RNA-directed
DNA methylation. Nature, 443, 1008–1012.
Qian, L., Vu, M.N., Carter, M. and Wilkinson, M.F. (1992) A spliced intron
accumulates as a lariat in the nucleus of T cells. Nucleic Acids Res. 20,
5345–5350.
Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) Molecular cloning: a lab-
oratory manual, 2nd edn. New York: Cold Spring Harbor Laboratory Press.
Smith, L.M., Pontes, O., Searle, I., Yelina, N., Yousafzai, F.K., Herr, A.J., Pik-
aard, C.S. and Baulcombe, D.C. (2007) An SNF2 protein associated with
nuclear RNA silencing and the spread of a silencing signal between cells in
Arabidopsis. Plant Cell, 19, 1507–1521.
Stam, M., Viterbo, A., Mol, J. and Kooter, J. (1998) Position-dependent
methylation and transcriptional gene silencing in inverted T-DNA repeats:
implications for posttranscriptional silencing of homologous host genes in
plants. Mol. Cell. Biol. 18, 6165–6177.
Stavolone, L., Kononova, M., Pauli, S., Ragozzino, A., de Haan, P., Milligan, S.,
Lawton, K. and Hohn, T. (2003) Cestrum yellow leaf curling virus (CmYLCV)
promoter: a new strong constitutive promoter for heterologous gene
expression in a wide variety of crops. Plant Mol. Biol. 53, 663–673.
Szittya, G., Silhavy, D., Molnar, A., Havelda, Z., Lovas, A., Lakatos, L., Banfalvi,
Z. and Burgyan, J. (2003) Low temperature inhibits RNA silencing-medi-
ated defence by the control of siRNA generation. EMBO J. 22, 633–640.
Vogt, U., Pelissier, T., Putz, A., Razvi, F., Fischer, R. and Wassenegger, M.
(2004) Viroid-induced RNA silencing of GFP-viroid fusion transgenes does
not induce extensive spreading of methylation or transitive silencing. Plant
J. 38, 107–118.
Voinnet, O. (2009) Origin, biogenesis, and activity of plant microRNAs. Cell,
136, 669–687.
Wang, M.B., Helliwell, C.A., Wu, L.M., Waterhouse, P.M., Peacock, W.J. and
Dennis, E.S. (2008) Hairpin RNAs derived from RNA polymerase II and
polymerase III promoter-directed transgenes are processed differently in
plants. RNA, 14, 903–913.
Wang, X., Elling, A.A., Li, X., Li, N., Peng, Z., He, G., Sun, H., Qi, Y., Liu, X.S.
and Deng, X.W. (2009) Genome-Wide and Organ-Specific Landscapes of
850 Athanasios Dalakouras et al.
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2009), 60, 840–851

Epigenetic Modifications and Their Relationships to mRNA and Small RNA
Transcriptomes in Maize. Plant Cell, 21, 1053–1069.
Wassenegger, M. (2005) The role of the RNAi machinery in heterochromatin
formation. Cell, 122, 13–16.
Wassenegger, M. and Pelissier, T. (1998) A model for RNA-mediated gene
silencing in higher plants. Plant Mol. Biol. 37, 349–362.
Wassenegger, M., Heimes, S., Riedel, L. and Sanger, H.L. (1994a) RNA-di-
rected de novo methylation of genomic sequences in plants. Cell, 76, 567–
576.
Wassenegger, M., Heimes, S. and Sanger, H.L. (1994b) An infectious viroid
RNA replicon evolved from an in vitro-generated non-infectious viroid
deletion mutant via a complementary deletion in vivo. EMBO J. 13, 6172–
6177.
Wierzbicki, A.T., Haag, J.R. and Pikaard, C.S. (2008) Noncoding transcription
by RNA polymerase Pol IVb/Pol V mediates transcriptional silencing of
overlapping and adjacent genes. Cell, 135, 635–648.
Wierzbicki, A.T., Ream, T.S., Haag, J.R. and Pikaard, C.S. (2009) RNA poly-
merase V transcription guides ARGONAUTE4 to chromatin. Nat. Genet. 41,
630–634.
Zhang, X., Henderson, I.R., Lu, C., Green, P.J. and Jacobsen, S.E. (2007) Role
of RNA polymerase IV in plant small RNA metabolism. Proc. Natl. Acad.
Sci. USA, 104, 4536–4541.
Zheng, X., Zhu, J., Kapoor, A. and Zhu, J. (2007) Role of Arabidopsis AGO6 in
siRNA accumulation, DNA methylation and transcriptional gene silencing.
EMBO J. 26, 1691–1701.
Zhou, Z., Luo, M.J., Straesser, K., Katahira, J., Hurt, E. and Reed, R. (2000) The
protein Aly links pre-messenger-RNA splicing to nuclear export in meta-
zoans. Nature, 407, 401–405.
Zilberman, D., Cao, X. and Jacobsen, S.E. (2003) ARGONAUTE4 control of
locus-specific siRNA accumulation and DNA and histone methylation.
Science, 299, 716–719.
Zilberman, D., Cao, X., Johansen, L.K., Xie, Z., Carrington, J.C. and Jacobsen,
S.E. (2004) Role of Arabidopsis ARGONAUTE4 in RNA-directed DNA
methylation triggered by inverted repeats. Curr. Biol. 14, 1214–1220.
Intronic hairpin-mediated RdDM 851
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Publishing Ltd, The Plant Journal, (2009), 60, 840–851