1513058960transpirationetext.pdf - e-pg pathshala
TRANSCRIPT

Input Template for Content Writers
(e-Text and Learn More)
1. Details of Module and its Structure

Library Science Management of Library and Information Network
Network
Module Detail
Subject Name <BOTANY>
Paper Name <Plant Physiology II>
Module Name/Title <Transpiration>
Module Id <>
Pre-requisites Basic knowledge about transpiration, its effect on plant water relations
Objectives To make the students aware of the physical and physiological mechanisms that regulates transpirational water loss.
Keywords Evaporational water loss, stomata, transcriptional regulation, water transport, Cuticle, daytime and night time transpiration, Water use efficiency (WUE)
Development Team
Structure of Module / Syllabus of a module (Define Topic / Sub-topic of module )
<Transpiration> <Sub-topic Name1>, <Sub-topic Name2>
Role Name Affiliation
Subject Coordinator <Dr. Sujata Bhargava> Savitribai Phule Pune University
Paper Coordinator <Dr. Sujata Bhargava>
Content Writer/Author (CW) <Dr. Dhiraj Naik > Janakidevi Bajaj College of Science, Wardha
Content Reviewer (CR) Dr. Sujata Bhargava
Language Editor (LE) Dr. A.N. Latey Savitribai Phule Pune University

Library Science Management of Library and Information Network
Network
TABLE OF CONTENTS (for textual content) 1. Introduction 2. Process of Transpiration 3. Transpiration as physical process 4. Modes of transpirational water loss 5. Factors affecting rates of transpiration 6. Transpiration and water use efficiency 7. Genetic variation and gene responsible for regulating transpiration and water use efficiency Introduction
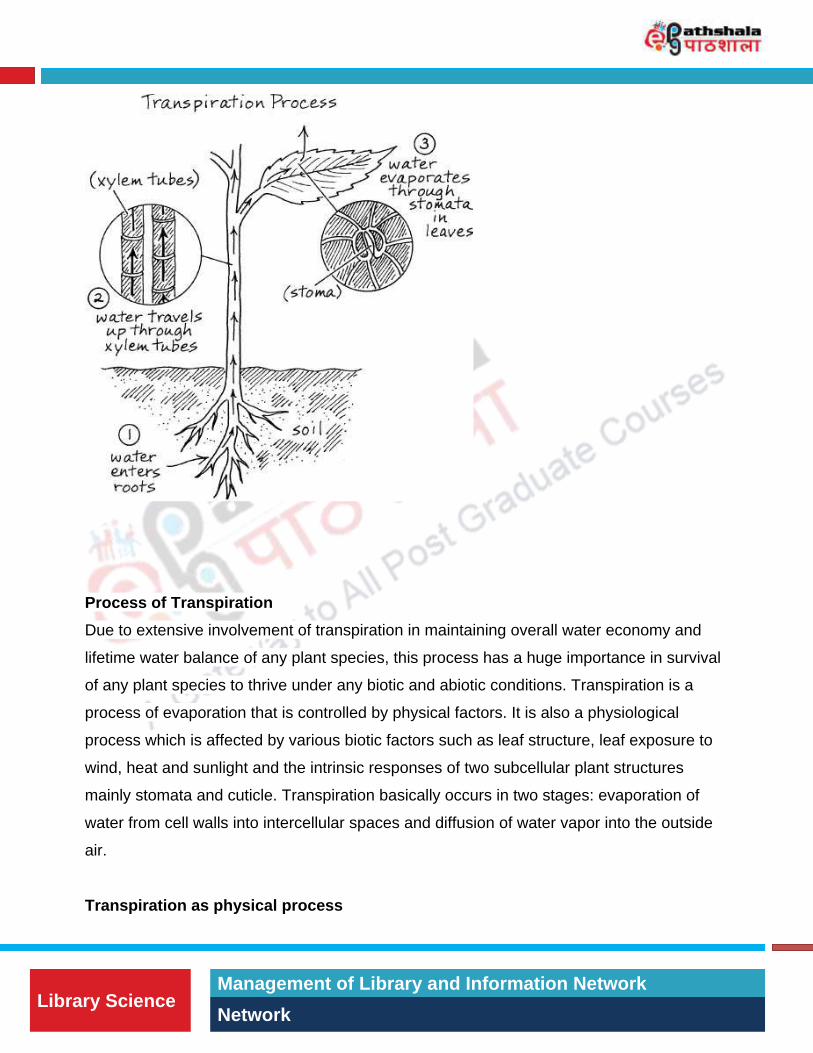
Transpiration is the dynamic process of evaporation of water molecules from the leaf
surface in plants. In worldwide scenario, transpiration process accounts for three quarters
of water loss due to vaporization from global land surface. As plants don’t have permeable
membranes for CO2 and impermeable to water, transpiration is an inevitable process
during day time when photosynthetic activity of leaves are maximum. The water molecules
through evaporational loss are replaced by additional absorption of water by soil leading to
a continuous column of water in the plant’s xylem (Figure 1). The process
of transpiration provides some benefit to the plant with evaporative cooling, nutrients,
carbon dioxide entry and water. Rates of transpiration depend on the water
potential gradient from the soil to the atmosphere and the resistances to its movement
through the plant. Water enters the root and travels through the cortex and endodermal
layers of cells to reach the xylem where water ascends to the leaf where, if not used in the
plant, evaporates. If water loss is greater than water uptake, air bubbles can form in the
xylem. Plants reduce water loss by closing their stomata, developing thick cuticles, or by
possessing leaf hairs to increase the boundary layer. Stomata are quick to respond to
environmental cues to protect the plant from losing too much water, but still allowing in
enough carbon dioxide to drive photosynthesis.
Figure 1: Diagrammatic representation of transpiration process (adapted from
www.education.com)
Library Science Management of Library and Information Network
Network
Process of Transpiration
Due to extensive involvement of transpiration in maintaining overall water economy and
lifetime water balance of any plant species, this process has a huge importance in survival
of any plant species to thrive under any biotic and abiotic conditions. Transpiration is a
process of evaporation that is controlled by physical factors. It is also a physiological
process which is affected by various biotic factors such as leaf structure, leaf exposure to
wind, heat and sunlight and the intrinsic responses of two subcellular plant structures
mainly stomata and cuticle. Transpiration basically occurs in two stages: evaporation of
water from cell walls into intercellular spaces and diffusion of water vapor into the outside
air.
Transpiration as physical process

Library Science Management of Library and Information Network
Network
Transpirational water loss is mainly driven by evaporation. The evaporation rate of water
vapor from any leaf surface depends on
(a) the energy supply available to vaporize the water molecules,
(b) the vapor concentration gradients that constitutes a driving force for water vapor
movement and
(c) the resistance in the diffusion pathway which is created by vapor concentration
gradients across the different tissues inside the leaf.
(a) Energy requirement
The solar radiations mostly serve as primary source of energy for evaporation. To certain
extent, wind velocity also serves as another source of water vapor evaporation on leaf
surfaces. During day time, most of water vapor escapes through the stomata, some water
molecules passes through the epidermis of leaves and its cuticular covering and some
water molecules escapes from the bark, stem or branches of any plant species.
Evaporation can be described by simple equation as described in equation (1):
E = Cwater - Cair Equation (1)
rair
where E is evaporation in kg m-2 s-1; Cwater and Cair are the concentration of water vapor at
the water surface and in the bulk air, respectively in kg m-3 and rair is boundary layer
resistance encountered by diffusing water molecules.
Because transpiration is considerably controlled by leaf boundary layer resistance, another
term or equation is need to explain it further as described in equation (2).
T = Cleaf-Cair Equation (2)
rleaf +rair
In this equation, T is transpiration, Cleaf is water vapor concentration at evaporating surface
within leaf, rleaf is an additional leaf resistance to diffusion in the leaf.
This equation calculates rates of transpiration (in kg m2 s-1) which is proportional to the
difference ( C) between the evaporating surfaces in the leaf and the bulk air outside the

Library Science Management of Library and Information Network
Network
leaf (Cleaf- Cair) divided by the sum of resistances to diffusion (rleaf + rair) (in s m-1). The
conceptual network of total resistance during the transpiration process is shown in figure 2.
Figure 2. Diagram showing resistances to the diffusion of water vapor from a leaf. The
rate of transpiration is proportional to steepness of the gradients in water vapor
concentration Cleaf to Cair and is inversely proportional to the resistances. Adapted from
Pallardy S.G. 2010 Physiology of Woody Plants 3 rd Edition Academic Press.
(b) the vapor concentration gradients
The water vapor concentration gradients is difference the water vapor concentration at the
evaporating surfaces and the vapor concentration outside the leaf surface surrounding the
bulk air (as described in equation 2). The vapor concentration at the evaporating surfaces
which is in close proximity of leaf surface is influenced by the temperature and water
potential at the surfaces. In well-watered conditions of plants, cell on the surfaces are
turgid and therefore water potential at the surfaces is zero. In such case, vapor
concentration is considered as saturating vapor concentration at that temperature. As
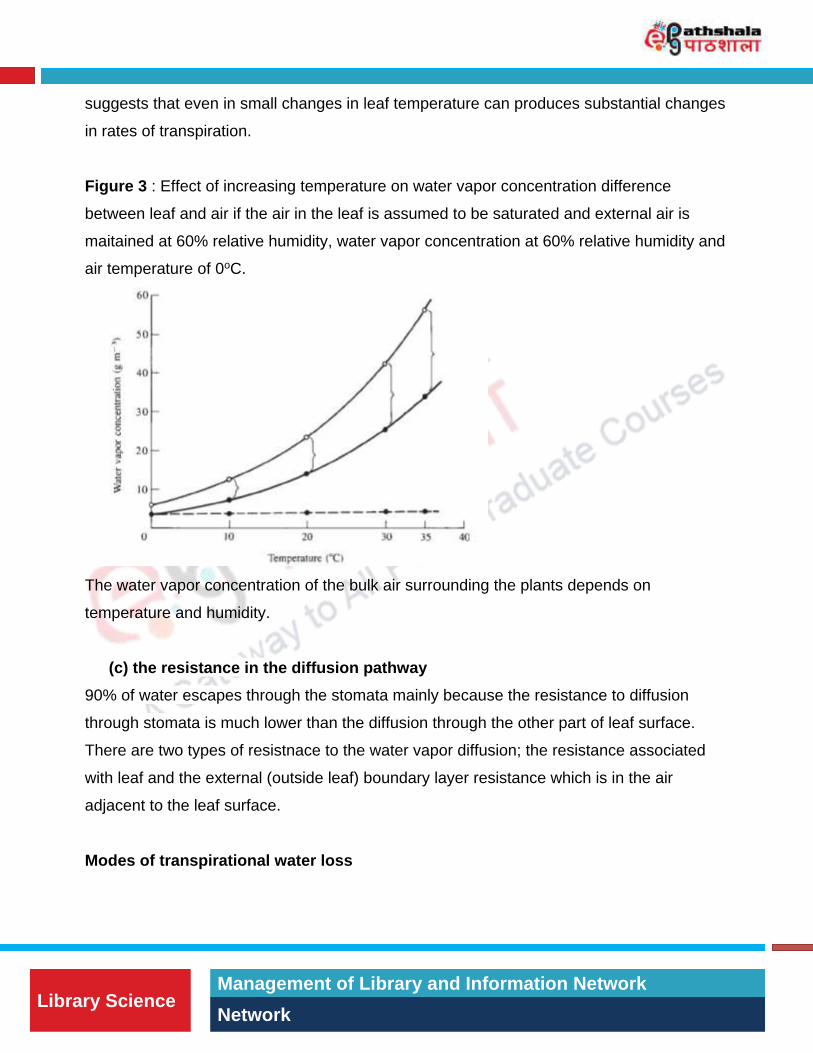
shown in figure 3, the effect of temperature increased saturating water vapor concentration
on the surface. When temperature increases from 10oC to 30oC, there is a strong increase
in saturating vapor concentration and a distinct difference between Cleaf and Cair. This
Library Science Management of Library and Information Network
Network
suggests that even in small changes in leaf temperature can produces substantial changes
in rates of transpiration.
Figure 3 : Effect of increasing temperature on water vapor concentration difference
between leaf and air if the air in the leaf is assumed to be saturated and external air is
maitained at 60% relative humidity, water vapor concentration at 60% relative humidity and
air temperature of 0oC.
The water vapor concentration of the bulk air surrounding the plants depends on
temperature and humidity.
(c) the resistance in the diffusion pathway
90% of water escapes through the stomata mainly because the resistance to diffusion
through stomata is much lower than the diffusion through the other part of leaf surface.
There are two types of resistnace to the water vapor diffusion; the resistance associated
with leaf and the external (outside leaf) boundary layer resistance which is in the air
adjacent to the leaf surface.
Modes of transpirational water loss

Library Science Management of Library and Information Network
Network
Transpiration occurs during daytime as well as in night time. Majority of transpirational
water loss occurs during day time. Stomata and cuticle plays an important role in
regulating rates of transpiration.
Stomatal transpiration: Stomatal transpiration during daytime begins with distinct
stomatal opening that results in increase in stomatal conductance leading to water loss.
Stomatal transpiration increases steadily with increase in stomatal conductance assuming
two physiological conditions that should follow simultaneously, the first one is vapor
pressure deficit is constant and the second one is nearly zero boundary layer resistance.
Water use efficiency can be improved by reducing water use through the mechanism that
regulates stomatal conductance to water vapor. This regulation involves increase in
stomatal number or density, stomatal aperture and boundary layer resistance.
Stomatal number (Density): Stomatal density i.e. number of stomata per unit area of leaf
influences water loss, as the stomatal pore area represents the principal diffusion pathway
for the water loss due to vapor pressure deficit between leaf and atmosphere. Reduced
stomatal number is correlated with reduced transpiration. Although no correlation was
observed between stomatal density and WUE. Stomatal density was negative correlated
with photosynthesis in forest tree poplar genotypes. This study suggests that WUE
increases with decreased stomatal density only to the minimum number so that they are
sufficient enough to capture CO2 efficiently. A variety of abiotic factors affects stomatal
density in plants. High light intensities and lower CO2 concentration increases stomatal
density. Increased salinity, high temperature and UV-B exposure decreases stomatal
density in plants. Drought stress invariably affects stomatal density. No distinct relationship
between drought stress and stomatal density was observed. In wild relatives of wheat,
Leymus chinensis, severe drought decreases stomatal density, but increase stomatal
densities during moderate drought. Increased stomatal density was observed with
decreased water availability in perennial grass Pseudoroegneria. These studies suggest
no clear mechanism between stomatal density and drought stress.
Stomatal aperture: Stomatal opening and closing is controlled by cascade of signaling
factors. The important stages in cascade signaling are increased in cytosolic Ca2+ ion
concentration surrounding the guard cells, which activates H+-ATPase. The activated
Library Science Management of Library and Information Network
Network
ATPase changes membrane polarization. The hyperpoplarized membrane assist K+
uptake into guard cell that results in a lowering of osmotic potential, increased uptake of
water in guard cells and ending into increased turgor pressure inside guard cells. The
ultimate increased in guard cell volume leads to stomatal opening. The reversal of this
biochemical process leads to stomatal closure (Table 1). The stomatal opening process is
controlled by various factors such as increased light intensity, lower CO2 concentration
and higher humidity, whereas stomatal closure is promoted by high CO2 concentration, low
relative humidity and high temperature and higher ABA concentration.
Table 1: Agents causing increased cytoplasmic concentrations of Ca2+ associated with
stomatal movements
Stomatal closure Stomatal opening
ABA Auxin
Elevated CO2 concentration Fusicoccin
Oxidative stress
External Ca2+ ions
Low external K+ ion concentrations
Cuticular transpiration: Apart from stomatal transpiration, cuticle is another important
tissue that loses water molecules from the outer surface layer of leaves presumably due to
higher vapor pressure deficit. The surface of plant tissues consists of cuticle which is made
up of cutin and lipid molecules. Cutin is chain of polymer matrix, whereas cuticular wax is
made of lipid molecules. Cuticular transpiration varies between plants of different growth
habit and from different habitats. The variation in cuticular transpiration depends upon
cuticular structure, thickness and on the amount of cuticular wax and lipid compositions.
The most common method of measuring cuticular transpiration is to estimate water loss
during dark conditions an d dry down curves d detached leaves and stems. The relative
water loss from leaf surface depends upon water permeability of cuticule which is directly
proportional to cuticule thickness. The Cuticle permeability is estimated by the rate of
chlorophyll extraction in an organic solvent, e.g., 80% ethanol. Although the amount of
water loss due to cuticular transpiration is not clear due to precise measurement limitation.

Library Science Management of Library and Information Network
Network
Recent advances in genetic improvement studies in drought tolerance showed important
findings. Quantitative trait loci (QTLs) for variation in epicuticular wax production in Rice
collocates with shoot and root drought tolerance. Several mutant studies showed that
altered cuticular traits such as increase and decreased in cuticular wax amount results in
significant transpirational water loss. However over-expression lines for altered wax
composition in Arabidopsis and Medicago showed weaker transpirational water loss,
suggestion no possible effect of altered cuticular traits on transpiration.
Nighttime transpiration: Transpiration during night time occurs in almost all C3 and C4
species. This type of transpiration results into 5-30 % diurnal water loss in all plant
species. Nightime transpiration results from incomplete stomatal closure. Few studies
showed strong correlationship between night time transpiration and nitrogen acquisition.
In these studies, WUE was correlated with nitrogen availability. This suggests that
lowering down night time transpiration rate to reduce water may not increase WUE if night
time transpiration is required for necessary nitrogen uptake. During drought stress,
substantial water loss would results in night time transpiration. Similar to day time drought
conditions, water loss at night increases with increased in vapor pressure deficit. This
suggests that night time transpiration results due to incomplete stomatal closure or
significant cuticular water loss.
Factors affecting transpiration:
Transpirational process is mainly controlled by physical factors, but inspite that, there are
many other biological factors which could influence rates of transpiration.
Leaf Area: Transpirational water loss in predominantly depend upon total leaf area of
plants. Plants with large leaf area usually transpire with higher rates as compared to plants
with smaller leaf areas. Some woody shrubs such as poplar, willow and black walnut shed
leaves when subjected to drought to reduce transpirational water loss. Many tree species
and crop plants show curling and rolling of wilted leaves during mid-day when transpiration
rate is maximum. Curling and rolling of leaves reduces the exposed surface to sunlight
and increases resistance to diffusion of water vapor.

Library Science Management of Library and Information Network
Network
Root: Shoot ratio: The ratio of root to shoots or more precisely, the ratio of water and
nutrient absorbing surface from the soil to transpiring surface from leaves is of greater
importance than the leaf area alone. This ratio is important if water absorption lags behind
the transpiration, then water deficit at leaf level develops which in turn closes the stomata
and reduces the transpiration. Plants with extensive, multibranched root system transpire
efficiently under limited water availabilities and survive better under drought stress than
those with shallow and sparsed root system, which indicates that root: shoot ratio is
important for plants to withstand transpiration failure during water stress.
Leaf size and shape: The leaf shape and leaf size significantly affects rates of
transpiration per unit leaf area. Thinner and dissected leaves or compound leaves tend to
be more cooler than the broad leafed plants due to thinner boundary layer permits more
rapid transfer of heat. Thinner and dissected leaves would have narrower effective leaf
widths, consequently thinner boundary layer and lowe leaf air resistance. Such leaf type
also would have capacity to avoid heat injuries due to high solar irradiance during mid-day
when leaves are more exposed to sunlight.
Leaf orientation and leaf surface: Leaf orientation in most plant species is angled in
such a way that their orientation gain maximum photosynthesis as well as minimum
transpirational water loss. Most of the leaves on any plant system grow in either more or
less perpendicular to brightest light that strikes on their surfaces. During drought stress,
many plant species show drooping and rolling characteristics in order to decrease mid-day
leaf temperature and reducing transpirational water loss.
Transpiration and water use efficiency:
Plants are always in greater demand of water supply for biochemical reactions and cell
expansion, which is necessary for growth and development (Taiz and Zeiger, 2006).
However, up to 95- 97% of water absorbed by plants is transpired and released as water
vapor to the atmosphere through stomata (Taiz and Zeiger, 2006; Gosney and Mickelbart,
unpublished). Transpirational rate is critical to the maintenance of plant water status,
proper leaf temperatures, and essential mineral uptake from soils. Stomata plays dual role,
mainly involved in CO2 uptake during day time as well as in released of water molecules
through transpiration. Stomatal pores are also responsible for carbon dioxide (CO2) uptake
Library Science Management of Library and Information Network
Network
that is necessary for carbon assimilation into biomass and yield. Therefore, the
relationship between transpiration and CO2 uptake is referred to as the transpiration ratio
(water transpired/CO2 assimilated) or water use efficiency (WUE: CO2 assimilated/water
transpired) (Farquhar and Sharkey, 1982; Yoo et al., 2009).
Plant WUE can be improved by increasing carbon assimilation relative to transpiration or
reducing transpiration relative to carbon assimilation (Chaves et al., 2004, Chaearle et al.,
2005). Data from numerous studies suggest that phenotypic traits that result in a slight
reduction in transpiration rate can increase WUE because water loss is reduced slightly,
long term water loss is reduced greatly, and CO2 uptake is not altered leading to increase
in biomass and yield. It may be feasible to improve WUE in plants by decreasing daytime
or nighttime transpiration.
Genetic variation and gene responsible for regulating transpiration and water use
efficiency:
With advent of new technologies in modern genetics, various research studies in crop
plants have shown that there is wide genetic variation for water use efficiency and for
transpiration ratios among genotypes within the species. It is now feasible to link
phenotypic traits with the help of genetic determinants which are involved in transpirational
water loss and crop yield. With the help of QTL mapping, marker assisted breeding,
various gene regulators have been identified which are potent target for water use
efficiency(Table 2).
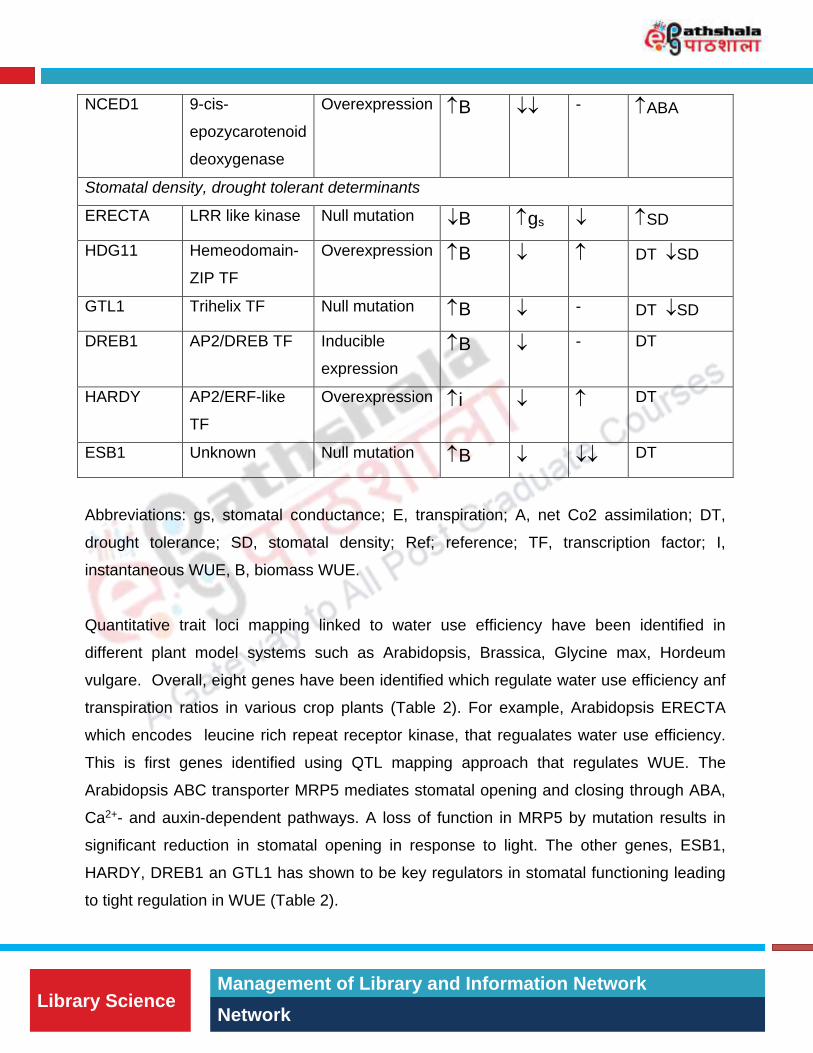
Table 2: Genes regulating WUE under well-watered conditions. Genes that are linked to
WUE and related physiological traits such as transpiration (E) and net CO2 assimilation
(A). Adapted from Yoo et al.,(2009) Regulation of Transpiration to Improve Crop Water
Use. Critical Reviews in Plant Sciences 28(6):410-431.
Gene Function Modification WUE E A Phenotypes
Stomatal opening/closing, ABA biosynthesis/signaling
MRP5 ABC transporter Null mutation i - DT
ABP9 bZIP TF Overexpression i ABA
Library Science Management of Library and Information Network
Network
NCED1 9-cis-
epozycarotenoid
deoxygenase
Overexpression B - ABA
Stomatal density, drought tolerant determinants
ERECTA LRR like kinase Null mutation B gs SD
HDG11 Hemeodomain-
ZIP TF
Overexpression B DT SD
GTL1 Trihelix TF Null mutation B - DT SD
DREB1 AP2/DREB TF Inducible
expression
B - DT
HARDY AP2/ERF-like
TF
Overexpression i DT
ESB1 Unknown Null mutation B DT
Abbreviations: gs, stomatal conductance; E, transpiration; A, net Co2 assimilation; DT,
drought tolerance; SD, stomatal density; Ref; reference; TF, transcription factor; I,
instantaneous WUE, B, biomass WUE.
Quantitative trait loci mapping linked to water use efficiency have been identified in
different plant model systems such as Arabidopsis, Brassica, Glycine max, Hordeum
vulgare. Overall, eight genes have been identified which regulate water use efficiency anf
transpiration ratios in various crop plants (Table 2). For example, Arabidopsis ERECTA
which encodes leucine rich repeat receptor kinase, that regualates water use efficiency.
This is first genes identified using QTL mapping approach that regulates WUE. The
Arabidopsis ABC transporter MRP5 mediates stomatal opening and closing through ABA,
Ca2+- and auxin-dependent pathways. A loss of function in MRP5 by mutation results in
significant reduction in stomatal opening in response to light. The other genes, ESB1,
HARDY, DREB1 an GTL1 has shown to be key regulators in stomatal functioning leading
to tight regulation in WUE (Table 2).
Library Science Management of Library and Information Network
Network
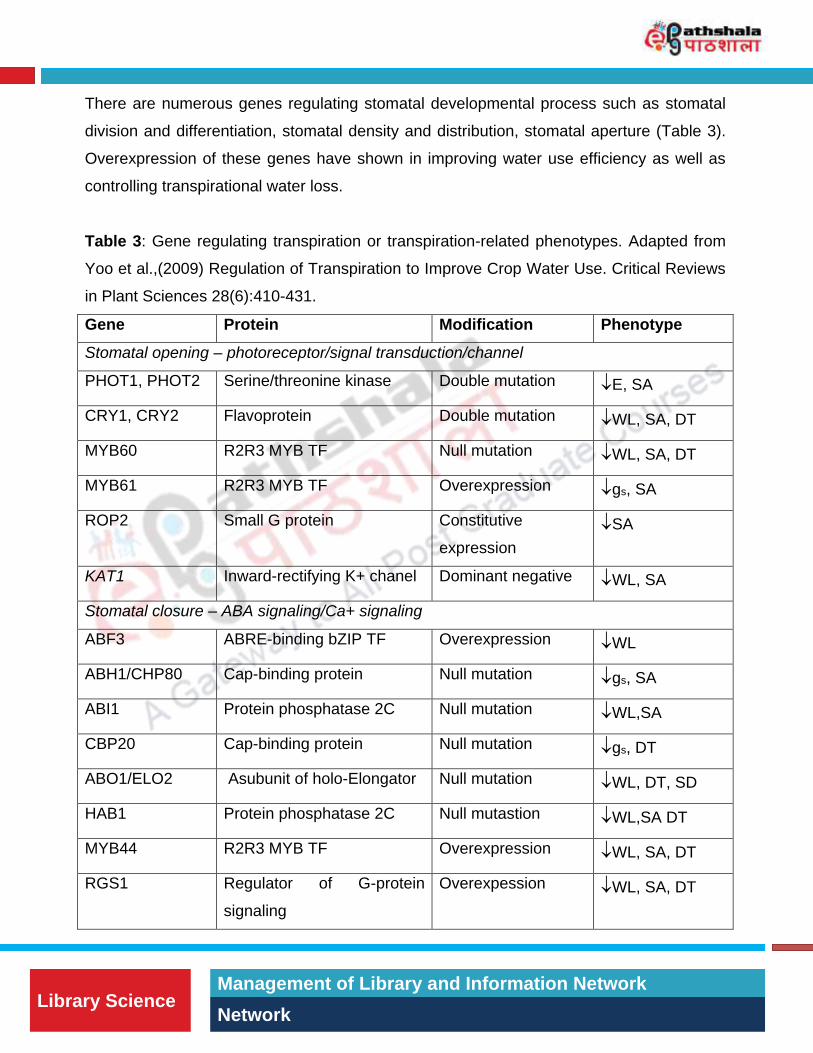
There are numerous genes regulating stomatal developmental process such as stomatal
division and differentiation, stomatal density and distribution, stomatal aperture (Table 3).
Overexpression of these genes have shown in improving water use efficiency as well as
controlling transpirational water loss.
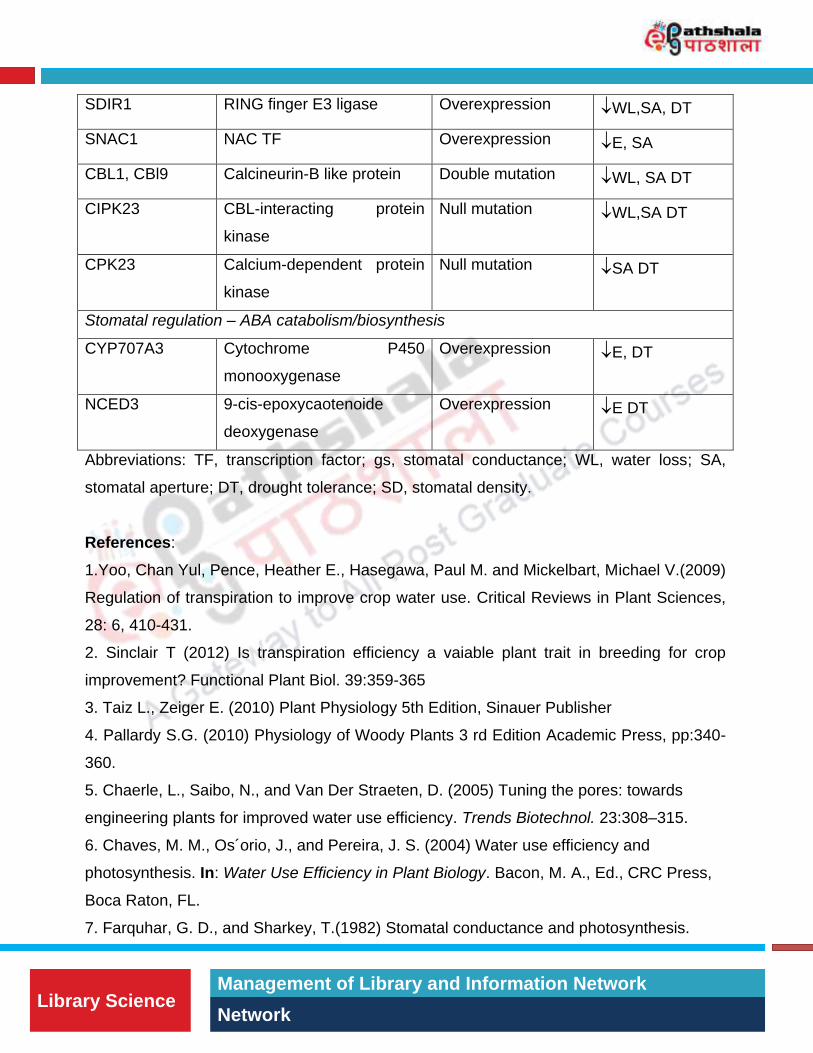
Table 3: Gene regulating transpiration or transpiration-related phenotypes. Adapted from
Yoo et al.,(2009) Regulation of Transpiration to Improve Crop Water Use. Critical Reviews
in Plant Sciences 28(6):410-431.
Gene Protein Modification Phenotype
Stomatal opening – photoreceptor/signal transduction/channel
PHOT1, PHOT2 Serine/threonine kinase Double mutation E, SA
CRY1, CRY2 Flavoprotein Double mutation WL, SA, DT
MYB60 R2R3 MYB TF Null mutation WL, SA, DT
MYB61 R2R3 MYB TF Overexpression gs, SA
ROP2 Small G protein Constitutive
expression
SA
KAT1 Inward-rectifying K+ chanel Dominant negative WL, SA
Stomatal closure – ABA signaling/Ca+ signaling
ABF3 ABRE-binding bZIP TF Overexpression WL
ABH1/CHP80 Cap-binding protein Null mutation gs, SA
ABI1 Protein phosphatase 2C Null mutation WL,SA
CBP20 Cap-binding protein Null mutation gs, DT
ABO1/ELO2 Asubunit of holo-Elongator Null mutation WL, DT, SD
HAB1 Protein phosphatase 2C Null mutastion WL,SA DT
MYB44 R2R3 MYB TF Overexpression WL, SA, DT
RGS1 Regulator of G-protein
signaling
Overexpession WL, SA, DT
Library Science Management of Library and Information Network
Network
SDIR1 RING finger E3 ligase Overexpression WL,SA, DT
SNAC1 NAC TF Overexpression E, SA
CBL1, CBl9 Calcineurin-B like protein Double mutation WL, SA DT
CIPK23 CBL-interacting protein
kinase
Null mutation WL,SA DT
CPK23 Calcium-dependent protein
kinase
Null mutation SA DT
Stomatal regulation – ABA catabolism/biosynthesis
CYP707A3 Cytochrome P450
monooxygenase
Overexpression E, DT
NCED3 9-cis-epoxycaotenoide
deoxygenase
Overexpression E DT
Abbreviations: TF, transcription factor; gs, stomatal conductance; WL, water loss; SA,
stomatal aperture; DT, drought tolerance; SD, stomatal density.
References:
1.Yoo, Chan Yul, Pence, Heather E., Hasegawa, Paul M. and Mickelbart, Michael V.(2009)
Regulation of transpiration to improve crop water use. Critical Reviews in Plant Sciences,
28: 6, 410-431.
2. Sinclair T (2012) Is transpiration efficiency a vaiable plant trait in breeding for crop
improvement? Functional Plant Biol. 39:359-365
3. Taiz L., Zeiger E. (2010) Plant Physiology 5th Edition, Sinauer Publisher
4. Pallardy S.G. (2010) Physiology of Woody Plants 3 rd Edition Academic Press, pp:340-
360.
5. Chaerle, L., Saibo, N., and Van Der Straeten, D. (2005) Tuning the pores: towards
engineering plants for improved water use efficiency. Trends Biotechnol. 23:308–315.
6. Chaves, M. M., Os´orio, J., and Pereira, J. S. (2004) Water use efficiency and
photosynthesis. In: Water Use Efficiency in Plant Biology. Bacon, M. A., Ed., CRC Press,
Boca Raton, FL.
7. Farquhar, G. D., and Sharkey, T.(1982) Stomatal conductance and photosynthesis.

Library Science Management of Library and Information Network
Network
Ann. Rev. Plant Physiol. 33: 317–345.
8. Gates,D.M. (1968) Transpiration and leaf temperature. Annu. Rev. Plant Physiol.
19: 211–238.
9. Fan, L. M., Zhao, Z., and Assmann, S. M. (2004) Guard cells: a dynamic signaling
model. Curr. Opin. Plant Biol. 7: 537–546.
10. Jarvis, P. G., and McNaughton, K. G. (1986) Stomatal control of transpiration:scaling
up from leaf to region. Adv. Ecol. Res. 15: 1–49.
Nobel, P. S. (1999) Physicochemical & Environmental Plant Physiology. 2nd ed., Academic
Press, San Diego, CA.
.