1α,25-dihydroxy-3-epi-vitamin d3, a natural metabolite of 1α,25-dihydroxy vitamin d3: production...
TRANSCRIPT

1a,25-Dihydroxy-3-epi-vitamin D3, a natural metabolite of1a,25-dihydroxy vitamin D3: production and biological activity
studies in pulmonary alveolar type II cellsq
Virender K. Rehan,a,* John S. Torday,a Sara Peleg,b Lynn Gennaro,c Paul Vouros,c
James Padbury,d D. Sunita Rao,d and G. Satyanarayana Reddyd
a Department of Pediatrics, Harbor-UCLA Research and Education Institute, Torrance, CA, USAb Department of Endocrine Neoplasia and Hormonal Disorders, The University of Texas, M.D. Anderson Cancer Center, Houston, TX 77030, USA
c Northeastern University, Boston, MA, USAd Department of Pediatrics, Women and Infants Hospital of Rhode Island, Brown University School of Medicine, 101 Dudley Street,
Providence, RI 02905, USA
Received 20 December 2001; received in revised form 26 March 2002; accepted 26 March 2002
Abstract
Pulmonary alveolar type II cells have been shown to be a possible target for the secosteroid hormone, 1a,25-dihydroxyvitamin D3[1a,25ðOHÞ2D3], during perinatal transition. At present, there is great interest to isolate and identify the metabolites of 1a,25ðOHÞ2D3produced in its target tissues and to determine the contribution of each individual metabolite of 1a,25ðOHÞ2D3 to the final expressionof the pleiotropic actions attributed to 1a,25ðOHÞ2D3. Of all the known metabolites of 1a,25ðOHÞ2D3, 1a,25ðOHÞ2-3-epi-D3 hasgained most attention as it is produced only in specific tissues and possesses significant activity in tissues in which it is produced.
Furthermore, in vivo studies indicate that this metabolite when compared to 1a,25ðOHÞ2D3 is less calcemic. Therefore, we performedthe present study to identify production of 1a,25ðOHÞ2-3-epi-D3 in alveolar type II cells, and to evaluate its effect on surfactantsynthesis.We incubatedNCI-H441 cells, an alveolar type II cell line, with 1a,25ðOHÞ2D3 and demonstrated that these cells metabolize1a,25ðOHÞ2D3 to various previously well-characterized polar metabolites, and to a less polar metabolite which was unequivocallyidentified as 1a,25ðOHÞ2-3-epi-D3 by GC/MS and HPLC analysis. Further, biological activity studies in H441 cells indicated that1a,25ðOHÞ2-3-epi-D3 possesses significant activity in terms of its ability: (i) to increase surfactant phospholipid synthesis, (ii) to inducesurfactant SP-B mRNA gene expression, and (iii) to increase surfactant SP-B protein synthesis. However, the activity of 1a,25ðOHÞ2-3-epi-D3 when compared to 1a,25ðOHÞ2D3 in generating VDR-mediated transcriptional activity in ROS 17/2.8 cells transfected withhuman osteocalcin VDRE/growth hormone gene construct, was significantly reduced. The high metabolic stability of 1a,25ðOHÞ2-3-epi-D3, as previously proposed by us, may be a possible explanation for the high in vitro activity in spite of the reduced VDR-mediated
transcriptional activity. In summary, we report for the first time the pathways of 1a,25ðOHÞ2D3 metabolism in pulmonary alveolartype II cells and indicate that 1a,25ðOHÞ2-3-epi-D3, a natural intermediary metabolite of 1a,25ðOHÞ2D3 possesses significant activityin stimulating surfactant synthesis in alveolar type II cells. � 2002 Elsevier Science (USA). All rights reserved.
Keywords: 1a,25ðOHÞ2D3; 1a,25ðOHÞ2-3-epi-D3; C-24 oxidation pathway; C-3 epimerization pathway; Metabolism; H441 cells
1. Introduction
The secosteroid hormone 1a,25-dihydroxyvitamin D3½1a,25ðOHÞ2D3� has been suggested to play an impor-tant role in lung development and function based on thefollowing lines of evidence: (1) the presence of receptorsfor 1a,25ðOHÞ2D3 in fetal rat lung during late intra-uterine life (days 19–21 of gestation) [1], at a time when
Molecular Genetics and Metabolism 76 (2002) 46–56
www.academicpress.com
q Abbreviations used: 1a,25ðOHÞ2D3, 1a,25-Dihydroxyvitamin D3;1a,25ðOHÞ2-3-epi-D3, 1a,25-dihydroxy-3-epi-vitamin D3; 1a; 24ðRÞ25ðOHÞ3D3, 1a,24(R)25-trihydroxyvitamin D3; 1a,25ðOHÞ2-24-oxo-D3,1a,25-dihydroxy-24-oxo-vitamin D3; 1a,23(S),25ðOHÞ3-24-oxo-D3,1a,23(S),25-trihydroxy-24-oxo-vitamin D3; DSPC, disaturated phos-phatidylcholine; GC/MS, gas chromatography/mass spectroscopy;
HPLC, high performance liquid chromatography; SP-B, surfactant
protein-B; VDR, vitamin D receptor.* Corresponding author. Fax: 310-222-3887.
E-mail address: [email protected] (V.K. Rehan).
1096-7192/02/$ - see front matter � 2002 Elsevier Science (USA). All rights reserved.PII: S1096 -7192 (02 )00022-7

two major events take place in the lung: (i) the appear-ance of differentiated alveolar type II cells, and (ii) theonset of surfactant synthesis and release [2,3]. (2) Usinga monoclonal antibody directed against the 1a,25ðOHÞ2D3 receptor, it was shown that the latter islocalized to alveolar type II cells, [4] and 1a,25ðOHÞ2D3is involved in the differentiation and maturation of al-veolar type II cells [5,6]. (3) 1a,25ðOHÞ2D3 stimulatesphospholipid biosynthesis and surfactant release fromthese fetal rat alveolar type II cells in vitro [7,8].Thus, alveolar type II cells, which are responsible forsurfactant biosynthesis, not only possess the receptorsfor 1a,25ðOHÞ2D3, but also 1a,25ðOHÞ2D3 is also in-volved in their differentiation and maturation and instimulating surfactant synthesis and release from thesecells.It is now well established that 1a,25ðOHÞ2D3 is
metabolized by its target tissues into several naturalmetabolites via modifications of both the side chain andthe A-ring. The C-24 oxidation pathway is initiated byhydroxylation at C-24. This pathway is the main sidechain modification pathway and leads to the formationof the end product, calcitroic acid [9–12]. The C-23 andC-26 oxidation pathways are initiated by hydroxylationsat C-23 and C-26, respectively. These minor side chainmodification pathways together lead to the formation ofthe end product, 1a,25ðOHÞ2D3-lactone [11,13]. The C-3epimerization pathway is initiated by epimerization ofthe hydroxyl group at C-3. This newly discovered A-ringmodification pathway leads to the conversion of1a,25ðOHÞ2D3 into 1a,25-dihydroxy-3-epi-vitamin-D3½1a,25ðOHÞ2-3-epi-D3� [14–16]. The pathways of1a,25ðOHÞ2D3 metabolism via modifications of both theside chain and the A-ring are described in detail in ourrecent paper [17].There is increasing evidence to show that some of
these natural metabolites possess significant biologicalactivities [18]. Out of all the natural metabolites of1a,25ðOHÞ2D3; 1a,25ðOHÞ2-3-epi-D3 has gained spe-cial significance as it is produced only in specific tis-sues and possesses significant biological activities inthe tissues in which it is produced. For example,1a,25ðOHÞ2-3-epi-D3 produced in keratinocytes isalmost equipotent to 1a,25ðOHÞ2D3 in inhibitingproliferation of these cells [19,20]. Also, 1a,25ðOHÞ2-3-epi-D3 produced in bovine parathyroid cells is equi-potent to 1a,25ðOHÞ2 D3 in suppressing parathyroidhormone secretion in these cells [21]. At present, thebiological role of 1a,25ðOHÞ2-3-epi-D3 in alveolar typeII cells is not known. Therefore, we performed thepresent study first to identify the production of1a,25ðOHÞ2-3-epi-D3 in alveolar type II cells, and thento evaluate its effects on these surfactant producingcells: (i) to stimulate surfactant phospholipid synthesis,(ii) to induce surfactant SP-B mRNA gene expression,(iii) to increase surfactant SP-B protein synthesis, and
(iv) to induce growth hormone production in rat os-teosarcoma cells (ROS 17/2.8) transfected with anosteocalcin VDRE/growth hormone gene reporterconstruct. We wish to report in the present paper, adetailed description of the metabolic pathways of1a,25ðOHÞ2D3 and the actions of 1a,25ðOHÞ2D3 and1a,25ðOHÞ2-3-epi-D3 in alveolar type II cells.
2. Materials and methods
2.1. Materials
The human pulmonary adenocarcinoma-derived cellline, NCI-H441 (H441), was obtained from the Ameri-can Type Culture Collection (ATCC, Rockville, MD).Streptomycin, pencillin, and RPMI 1640 medium wereobtained from Life Technologies (Gaithersburg, MD).Fetal bovine serum was purchased from Hyclone (Lo-gan, UT). All solvents were obtained from Burdick andJackson Laboratories, Muskegan, MI.
2.2. Vitamin D compounds
Crystalline 1a,25ðOHÞ2D3 and 1a; 25ðOHÞ2D3-3-epi-D3 were synthesized at Hoffmann-La-Roche (Nutley,NJ, USA). All the known natural metabolites of1a,25ðOHÞ2D3, which include 1a,24(R)25-trihydroxyvi-tamin D3½1a,24(R)25ðOHÞ3D3�, 1a,25-dihydroxy-24-oxo-vitamin D3 ½1a,25ðOHÞ2-24-oxo-D3�, and 1a,23(S), 25-trihydroxy-24-oxo-vitamin D3½1a;23(S),25ðOHÞ3-24-oxo-D3� were biologically synthesized in the rat kidneyperfusion system as previously described [9,22].
2.3. High performance liquid chromatography (HPLC)
HPLC analysis of the lipid extracts from the cellsand media was performed with a Waters SystemController (Millennium 32) equipped with a photo-diode array detector (Model PDA 996) to monitor theultraviolet (UV) absorbing material at 265 nm (Wa-ters Associates, Milford, MA). The vitamin D com-pounds were isolated and purified using bothstraight and reverse phase HPLC systems. Analysis bystraight phase HPLC involved the use of a Zorbax-SIL column (9� 250mm) (Dupont, Wilmington, DE,USA), eluted with three different solvent mixtures at aflow rate of 2ml/min. The solvent mixtures used wereas follows: 10% isopropanol in hexane (HPLC system-I), 6% isopropanol in hexane (HPLC system-II), and4% isopropanol in methylene chloride (HPLC system-III). Analysis by reverse phase HPLC involved the useof a Zorbax-ODS column ð4:5� 250mm) (Dupont,Wilmington, DE, USA) eluted with 25% water inmethanol at a flow rate of 1ml/min (HPLC system-IV).
V.K. Rehan et al. / Molecular Genetics and Metabolism 76 (2002) 46–56 47

2.4. Gas chromatography/mass spectrometry (GC/MS)
GC/MS analysis was performed using a Hewlett-Packard GC-MSD system which is equipped with a5890 series II chromatograph, a 5971 mass-selectivedetector and a 7673 GC autosampler (Hewlett-Packard,Wilmington, DE). The vitamin D compounds weretrimethylsilylated in 30 ll of a 1:1 mixture of acetonitrileand Power SIL-Prep (Alltech Associates, Deerfield, IL,USA) and incubated at 70 �C for 15min. The trimeth-ylsilyl ether derivatives were analyzed in quadruplicate.Samples were analyzed on an HP-5 capillary columnð30m� 0:25mm� 0:25lmÞ with a temperature pro-gram ranging from 150 to 300 �C (10 �C/min ramp).Full-scan electron impact spectra (m=z 50–650) wereacquired for each experiment.
2.5. Study of the metabolism of 1a,25ðOHÞ2D3 in H441
H441 cells were maintained in RPMI 1640 mediumsupplemented with 10% fetal bovine serum (FBS) andantibiotics [penicillin (100 IU/ml) and streptomycin(100 lg/ml). Cell culture medium was changed every 3–4days. The cells were subcultured when approximately80% confluent and were not subcultured beyond fivepassages. For the metabolism studies, 3� 106 cells wereseeded in T150 tissue culture flasks and grown to con-fluence. The incubations were carried out at 37 �C in ahumidified atmosphere under 5% CO2.Confluent H441 cells were incubated with 1 lM
concentration of 1a,25ðOHÞ2D3 in 50ml of media con-taining 10% FBS. The incubations were stopped after 24and 48 h with 10ml of methanol, and the lipids fromboth cells and media were extracted for HPLC analysisusing the extraction procedure described earlier [9].Prior to the lipid extraction, the cells and media werespiked with 5 lg of 25OHD3, which was used as an in-ternal standard. The recovery of the internal standardwas used to assess the extraction efficiencies of the var-ious lipid-soluble vitamin D metabolites.Control incubations without cells, containing only
media and the vitamin D compounds, were performed.The control studies indicated that the vitamin D com-pounds did not undergo any chemical change orbreakdown either during the 24 and 48 h incubationperiods or during the extraction procedure (data notshown).
2.6. Determination of 3H-saturated phosphatidylcholine(PC)
Incorporation of [methyl-3H]-choline chloride (NENDupont) into saturated PC was studied in monolayers ofH441 cells using a modification of previously describedprocedures [23]. Briefly, subconfluent cultures of H441in 6-well plates were incubated with graded concentra-
tions of 1a,25ðOHÞ2D3, 1a,25ðOHÞ2-3-epi-D3, or vehiclein serum-less medium containing 0.1% bovine serumalbumin (BSA) for 24 h at 37 �C. During the last 4 h, thewells were incubated with fresh medium ½1a,25ðOHÞ2D3; 1a,25ðOHÞ2-3-epi-D3 or vehicle] and 1 lCi/ml of[methyl-3H]-choline chloride. After incubation, cellswere washed once with fresh medium, recovered bygentle trypsinization and counted in a particle counter(Hyaleah, FL). Cellular lipids were extracted withchloroform and methanol (2:1). The organic phase wasdried under a stream of nitrogen at 60 �C, resuspendedin 0.5ml of carbon tetrachloride containing 3.5mg ofosmium tetroxide and left at room temperature for15min. The reaction mixture was redried under nitrogenand resuspended in 70 ll of chloroform/methanol (9:1,vol/vol). The lipid extracts were transferred to silica gelplates (Kodak, Rochester, NY) and developed in achloroform/methanol/water (65:25:4) solvent system.Pure dipalmitoyl phosphatidylcholine was used as thechromatographic standard. The developed plates werestained with bromothymol blue, blotted, and vacuum-dried for 5min at 90 �C. Chromatogram spots corre-sponding to the migration of saturated PC were scrapedfrom the plates and counted by liquid scintillationspectrometry. The amounts of [methyl-3H]-cholinechloride incorporated into saturated PC were expressedas disintegrations per min (dpm) per mg protein.
2.7. Reverse transcriptase-polymerase chain reaction(RT-PCR)
Total cellular RNA was isolated using a modificationof the single-step method of Chomczynski and Sacchi[24]. Briefly, cells were lysed directly by vortexing in 2mllysis solution [2M guanidinium thiocyanate, 12.5mMsodium citrate (pH 7.0), 0.25% sarcosyl, 50mM 2-mer-captoethanol, and 50% (vol/vol) water–saturated phe-nol]. Chloroform–isoamyl alcohol (49:1, vol/vol) wasadded to each sample and the mixture was vortexed for10 s, and cooled on ice for 15min. After centrifugation at10,000g for 20min at 4 �C, RNA in the upper aqueousphase was precipitated in ethanol at )20 �C. The RNAwas pelleted by sedimentation at 10,000g for 20min at4 �C, solubilized in 1ml lysis solution, extracted in phe-nol/chloroform/isoamyl alcohol and precipitated inethanol. The RNA pellets were resuspended in 100llDEPC-treated water and quantitated by absorbance at260 nm. The quality of RNA was assessed on a 1% aga-rose gel after ethidium bromide staining. Twenty-five lgof total RNA was reverse transcribed with the randomhexamers and deoxyribonucleotides by superscript re-verse transcriptase enzyme (Promega). Five lg of reversetranscript was amplified in 30 cycles of PCR with humanSP-B primers (50-CATCGACTACTTCCAGAACCAGAC-30 and 30-GCAGATGCCGCCCGCCACCAGAGG-30). Co-amplification with glyceraldehyde-3-phosphate
48 V.K. Rehan et al. / Molecular Genetics and Metabolism 76 (2002) 46–56

dehydrogenase (GAPDH) cDNA was used as an internalstandard. The reactions were run according to the stan-dard protocols at 42 �C for 75min and terminated byheating at 95 �C for 5min. PCR was initiated by TaqDNA polymerase and allowed to proceed for 30 cycleswith an annealing temperature at 50 �C. Five lg ofreaction products was separated on a 1% agarose gel andintensities of the PCR product bands were quantitativelyanalyzed (Eagle Eye, Stratagene).
2.8. Western blot analysis
Protein extraction and Western blot analysis forsurfactant proteins were performed according to modi-fications of methods described previously [25]. Briefly,cells were homogenized in 10mM tris (hydroxymethyl)aminomethane (Tris, pH 7.5), 0.25M sucrose, 1mMEDTA, 5mM benzamidine, 2mM phenylmethylsulfonylfluoride, and 10lg=ml each of pepstatin A, aprotinin,and leupeptin, and centrifuged at 140� for 10min(4 �C). Equal amounts of the protein from the super-natant were dissolved in electrophoresis sample bufferand were subjected to sodium dodecyl sulfate–poly-acrylamide (4–12% gradient) gel electrophoresis fol-lowed by electrophoretic transfer to a nitrocellulosemembrane, as previously described [26]. The membranewas blocked with 5% milk in Tris–buffered saline andwas incubated with rabbit anti-human SP-B antibody(1:10,000) overnight at 4 �C. The surfactant protein Bantibody used was rabbit antihuman (Chemicon,Temecula, CA). Subsequently, the membrane was wa-shed with Tris–buffered saline containing 0.1% Tween 20and incubated with a 1:150,000 dilution of horseradishperoxidase-conjugated goat-antirabbit immunoglobulinG (Calbiochem, La Jolla, CA) for 2 h at room temper-ature, washed again, and developed with a chemilumi-nescent substrate [enhanced chemiluminescence (ECL);Amersham, Arlington Heights, IL] following the man-ufacturer’s protocol. The density of the SP-B bands wasquantified using a scanning densitometer (Eagle Eye,Stratagene).
2.9. Transcription assay
To test the vitamin D receptor-mediated transcrip-tional activities of the 19-nor vitamin D3 compounds,ROS 17/2.8 cells were plated in 35-mm dishes at a den-sity of 3� 105 cells/dish in Dulbecco’s modified essentialmedium (DMEM) and 10% FCS. Forty-eight hours la-ter, the cells were transfected with plasmid ð2lg=dishÞcontaining the vitamin D response element (VDRE)from the human osteocalcin gene (GGTGACTCACCG-GGTGAACGGGGGCATT) [27]. This response ele-ment was inserted upstream of a thymidine kinasepromoter/growth hormone fusion gene. All transfec-tions were performed by the DEAE dextran method
[28], and the cells were treated for 1min with 10% fetalcalf serum (FCS). Samples of medium harvested for themeasurements of growth hormone were collected 24 hafter ligand treatment. Growth hormone production bythe reporter gene was measured by a radioimmunoassay(RIA) as described by the manufacturer (Nichols Insti-tute, San Juan Capistrano, CA, USA).
2.10. Statistics
Values were calculated as mean� standard error(SEM) from duplicate or triplicate samples from at leastthree different experiments. Significance levels were de-termined by Student’s t test.
3. Results
3.1. Metabolism of 1a,25ðOHÞ2D3 in human lung cells,H441
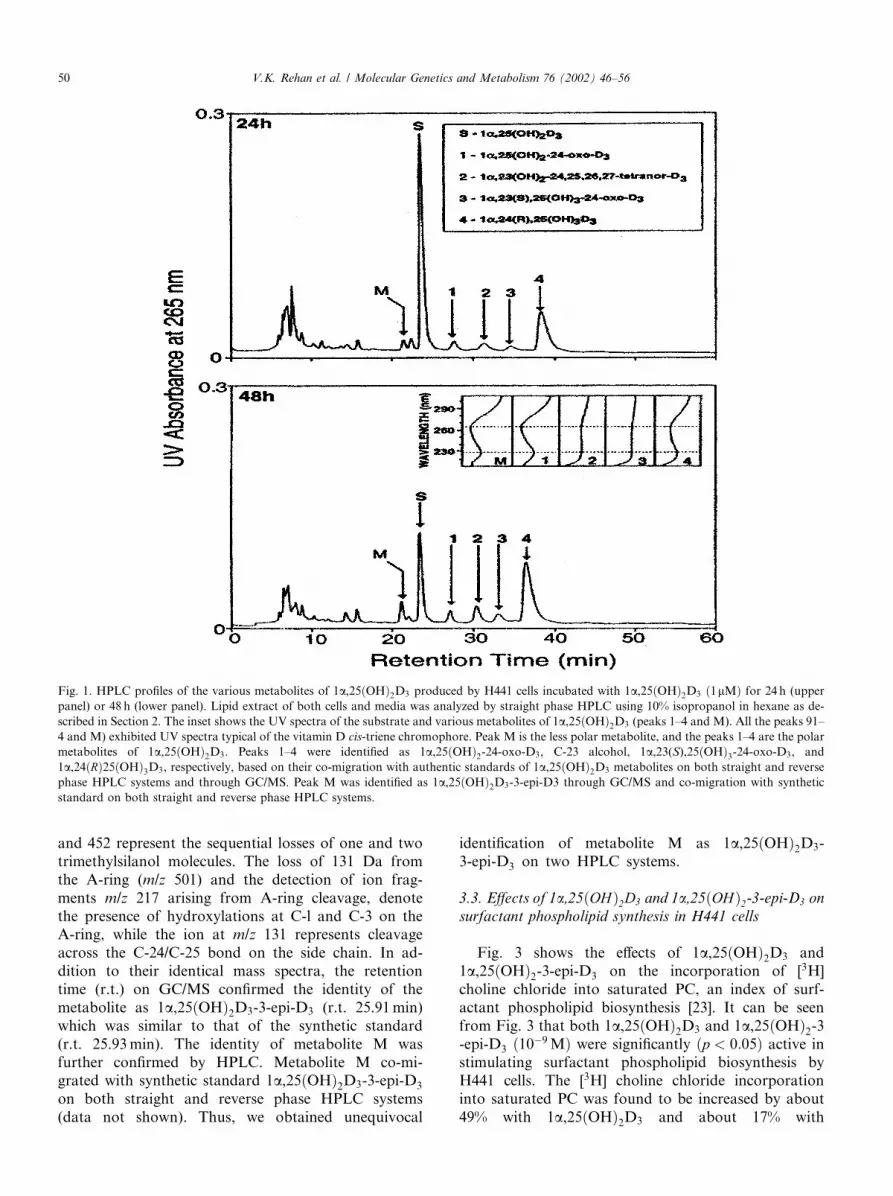
Fig. 1 shows the HPLC profiles obtained when con-fluent H441 cells were incubated with 1lM 1a,25ðOHÞ2D3 for 24 (panel A) and 48 h (panel B). It can beseen that 1a,25ðOHÞ2D3 was metabolized by H441 cellsinto several polar metabolites (peaks 1–4), all of whichexhibited the UV absorption spectra with absorbancemaxima at 265 nm and absorbance minima at 228 nm,indicating the presence of an intact 5,6-cis-triene chro-mophore, a characteristic of the D vitamins (Fig. 1, in-set). Peaks 1–4 are identified through their co-migrationwith the corresponding synthetic standards on differentHPLC systems described in Section 2. Peak 1 wasidentified as 1a,25ðOHÞ2-24-oxo-D3, peak 2 as C-23 al-cohol, peak 3 as 1a; 23ðSÞ; 25ðOHÞ3-24-oxo-D3, andpeak 4 as 1a; 24ðRÞ25ðOHÞ3D3, i.e., the metabolitesderived through the C-24 oxidation pathway. Theidentity of each polar metabolite was further confirmedby GC/MS (data not shown).In addition to these aforementioned polar metabo-
lites produced through the C-24 oxidation pathway (Fig.1, peaks 1–4), H441 cells also produced a novel lesspolar metabolite (peak M), which possesses the char-acteristic UV chromophore of vitamin D (kmin228,kmax265) (Fig. 1, inset). Metabolite M was identified as1a,25ðOHÞ2D3-3-epi-D3, as described below.
3.2. Identification of metabolite M as 1a,25ðOHÞ2D3-3-epi-D3 by GC/MS and HPLC
Metabolite M was purified using four differentHPLC systems, as described in Section 2. Fig. 2 showsthe identical mass spectral characteristics of the trim-ethylsilylated 1a,25ðOHÞ2D3-3-epi-D3 synthetic stan-dard and those of metabolite M. The molecular ionwas detected at m/z 632. The fragments at m/z 542
V.K. Rehan et al. / Molecular Genetics and Metabolism 76 (2002) 46–56 49
and 452 represent the sequential losses of one and twotrimethylsilanol molecules. The loss of 131 Da fromthe A-ring (m/z 501) and the detection of ion frag-ments m/z 217 arising from A-ring cleavage, denotethe presence of hydroxylations at C-l and C-3 on theA-ring, while the ion at m/z 131 represents cleavageacross the C-24/C-25 bond on the side chain. In ad-dition to their identical mass spectra, the retentiontime (r.t.) on GC/MS confirmed the identity of themetabolite as 1a,25ðOHÞ2D3-3-epi-D3 (r.t. 25.91min)which was similar to that of the synthetic standard(r.t. 25.93min). The identity of metabolite M wasfurther confirmed by HPLC. Metabolite M co-mi-grated with synthetic standard 1a,25ðOHÞ2D3-3-epi-D3on both straight and reverse phase HPLC systems(data not shown). Thus, we obtained unequivocal
identification of metabolite M as 1a,25ðOHÞ2D3-3-epi-D3 on two HPLC systems.
3.3. Effects of 1a,25ðOHÞ2D3 and 1a,25ðOHÞ2-3-epi-D3 onsurfactant phospholipid synthesis in H441 cells
Fig. 3 shows the effects of 1a,25ðOHÞ2D3 and1a,25ðOHÞ2-3-epi-D3 on the incorporation of [3H]choline chloride into saturated PC, an index of surf-actant phospholipid biosynthesis [23]. It can be seenfrom Fig. 3 that both 1a,25ðOHÞ2D3 and 1a,25ðOHÞ2-3-epi-D3 ð10�9MÞ were significantly ðp < 0:05Þ active instimulating surfactant phospholipid biosynthesis byH441 cells. The [3H] choline chloride incorporationinto saturated PC was found to be increased by about49% with 1a,25ðOHÞ2D3 and about 17% with
Fig. 1. HPLC profiles of the various metabolites of 1a,25ðOHÞ2D3 produced by H441 cells incubated with 1a,25ðOHÞ2D3 ð1lMÞ for 24 h (upperpanel) or 48 h (lower panel). Lipid extract of both cells and media was analyzed by straight phase HPLC using 10% isopropanol in hexane as de-
scribed in Section 2. The inset shows the UV spectra of the substrate and various metabolites of 1a,25ðOHÞ2D3 (peaks 1–4 and M). All the peaks 91–4 and M) exhibited UV spectra typical of the vitamin D cis-triene chromophore. Peak M is the less polar metabolite, and the peaks 1–4 are the polar
metabolites of 1a,25ðOHÞ2D3. Peaks 1–4 were identified as 1a,25ðOHÞ2-24-oxo-D3, C-23 alcohol, 1a,23(S),25ðOHÞ3-24-oxo-D3, and1a,24ðRÞ25ðOHÞ3D3, respectively, based on their co-migration with authentic standards of 1a,25ðOHÞ2D3 metabolites on both straight and reversephase HPLC systems and through GC/MS. Peak M was identified as 1a,25ðOHÞ2D3-3-epi-D3 through GC/MS and co-migration with syntheticstandard on both straight and reverse phase HPLC systems.
50 V.K. Rehan et al. / Molecular Genetics and Metabolism 76 (2002) 46–56

1a,25ðOHÞ2 -3-epi-D3, respectively. The ED50 for both1; 25 ðOHÞ2 D3 and 3-epi-1,25ðOHÞ2D3 (for surfactantphosp- holipid synthesis) in the dose range between10�6 and 10�9 M were calculated, and were 16:8� 9and 17:8� 8 nM (mean� SD), respectively (data notshown).
3.4. Effects of 1a,25ðOHÞ2D3 and 1a,25ðOHÞ2-3-epi-D3 onthe expression of surfactant SP-B mRNA in H441 cells
H441 cells were cultured for 2 days with either1a,25ðOHÞ2D3 or 1a,25ðOHÞ2-3-epi-D3 ð10�9MÞ andtheir effects on SP-B mRNA gene expression were de-termined by semi-quantitative RT-PCR. Fig. 4 shows theeffects of 1a,25ðOHÞ2D3 and 1a,25ðOHÞ2-3-epi-D3 onthe stimulation of SP-B mRNA gene expression in H441
cells. As shown in Fig. 4, panel A, the PCR productsappeared as a single band, 2.0 kb, indicating the presenceof SP-B mRNA. Densitometric analysis (Fig. 4, panel B)revealed that both 1a,25ðOHÞ2D3 and 1a,25ðOHÞ2-3-epi-D3 significantly ðp < 0:05Þ increased SP-B mRNAexpression when compared to the control. The SP-BmRNA levels were increased by about 30% with1a,25ðOHÞ2D3, and about 20% with 1a,25ðOHÞ2-3-epi-D3, respectively.
3.5. Effects of 1a,25ðOHÞ2D3 and 1a,25ðOHÞ2-3-epi-D3 onsurfactant SP-B protein synthesis in H441 cells
H441 cells were cultured for 2 days with either1a,25ðOHÞ2D3 or 1a,25ðOHÞ2-3-epi-D3 (10�9 M) andSP-B protein content was analyzed by Western blot.
Fig. 2. Mass spectra of trimethylsilylate derivates of the synthetic standard of 1a,25ðOHÞ2D3-3-epi-D3 and of metabolite M produced in H441
cells. Upper panel: metabolite M, retention time 25.91min; Lower panel: synthetic standard of 1a,25ðOHÞ2D3-3-epi-D3, retention time25.93min.
V.K. Rehan et al. / Molecular Genetics and Metabolism 76 (2002) 46–56 51

Fig. 5 shows the effects of 1a,25ðOHÞ2D3 and 1a,25ðOHÞ2-3-epi-D3 on the stimulation of SP-B proteinsynthesis in H441 cells. Immunoblot in Fig. 5, panel A,shows the SP-B protein detected as an 8.7 kD band.Densitometric analysis (Fig. 5, panel B) of the immu-noblots revealed that both 1a,25ðOHÞ2D3 and1a,25ðOHÞ2-3-epi-D3 significantly ðp < 0:05Þ increasedSP-B protein content when compared to the control.The SP-B protein content was found to be increased byabout 25% with 1a,25ðOHÞ2D3, and about 15% with1a,25ðOHÞ2 -3-epi-D3, respectively.
3.6. Transcriptional activities of 1a,25ðOHÞ2D3 and1a,25ðOHÞ2-3-epi-D3 in ROS 17/2.8 cells
To determine vitamin D receptor (VDR)-mediatedtranscriptional activities, we assessed the abilities of1a,25ðOHÞ2D3 and 1a,25ðOHÞ2-3-epi-D3 to inducegrowth hormone production in ROS 17/2.8 cells trans-fected with an osteocalcin VDRE/growth hormone geneconstruct. Fig. 6 shows that the VDR-mediated tran-scriptional activity through the osteocalcin VDRE wasdose-dependent for both 1a,25ðOHÞ2D3 and1a,25ðOHÞ2-3-epi-D3. The C-3 epimer, 1a,25ðOHÞ2-3-epi-D3 had very little transcriptional activity at 1 nM.Detectable transcriptional activity was observed at10 nM. Among both these vitamin D3 compounds,½1a,5ðOHÞ2D3� had higher transcriptional activity at thevarious concentrations studied.
4. Discussion
The association of vitamin D deficiency and respi-ratory disorders in infants [29], the abnormal alveolarand connective tissue development in pups born tovitamin D deficient mothers [30], and the detection ofa vitamin D-dependent calbindin in human fetal lung[31] suggest that the lung may be a target tissue for1a,25ðOHÞ2D3, especially during fetal life. Most im-portantly, the presence of 1a,25ðOHÞ2D3 receptors inalveolar type II cells of the lung and the involvementof 1a,25ðOHÞ2D3 in the maturation and differentiationof these cells and surfactant synthesis and release fromthese cells indicated that the alveolar type II cells maybe major targets for 1a,25ðOHÞ2D3. Therefore, in thepresent study we examined the metabolism of 1a,25ðOHÞ2D3, and the effects of its metabolite 1a,25ðOHÞ2-3-epi-D3, on the alveolar type II cells. But it is im-portant to emphasize that since no specific pulmonaryabnormalities have been reported in VDR null mutantmice and in patients that lack VDR, it is likely thatvitamin D is not absolutely essential in pulmonarydevelopment and function. Since transition from liquidbreathing fetus to air breathing neonate is such a vitalcomponent for the initiation of extrauterine life, it islikely that there is a great deal of redundancy in themechanisms involved and vitamin D is only one of themany molecules that affect pulmonary alveolar type IIcell maturation.
Fig. 3. Effects of 1a,25ðOHÞ2D3 and 1a,25ðOHÞ2D3-3-epi-D3 on surfactant phospholipid synthesis in H441 cells. The H441 cells were treated for 24 hwith either 1a,25ðOHÞ2D3 or 1a,25ðOHÞ2D3-3-epi-D3 ð10�9MÞ. During the last 4 h the cells were also treated with [3H] choline chloride. Lipids wereextracted from the cells and were transferred to silica gel plates and developed in chloroform/methanol/water (65:25:4) solvent system. The amount of
[3H] choline chloride incorporated into saturated PC was measured by counting the radioactivity in the chromatogram spots corresponding to the
migration of saturated PC. Data are expressed as dpm/mg protein. Each value represents the means�SE of three experiments with duplicate dishes.
52 V.K. Rehan et al. / Molecular Genetics and Metabolism 76 (2002) 46–56

In the present study, we report for the first time themetabolism of 1a,25ðOHÞ2D3 in alveolar type II cellsinto the novel A-ring modified metabolite, 1a,25ðOHÞ2-3-epi-D3, derived via the C-3 epimerization pathwayalong with several previously well-characterized side-chain modified metabolites, namely 1a,24ðRÞ25ðOHÞ3D3, 1a,25ðOHÞ2-24-oxo-D3, 1a,23ðSÞ; 25ðOHÞ3-24-oxo-D3, and C-23 alcohol derived via the C-24 oxidationpathway. The identity of 1a,25ðOHÞ2-3-epi-D3 was un-equivocally established using the techniques of GC/MSand HPLC analysis. The epimer 1a,25ðOHÞ2-3-epi-D3 isformed as a result of a change in the orientation of C-3hydroxy group from b to a. The possible mechanismsresponsible for the C-3 epimerization of 1a,25ðOHÞ2D3are discussed in our previous paper [16].Out of all the natural metabolites of 1a,25ðOHÞ2D3,
1a,25ðOHÞ2-3-epi-D3 has gained special significance, asits production is target tissue specific. For example,1a,25ðOHÞ2-3-epi-D3 is produced in human keratino-cytes [14–16], bovine parathyroid cells [21], rat osteo-
sarcoma cells [32], human colon carcinoma cells [33], ratvascular smooth muscle cells [34], but not in perfused ratkidney [32], and human promyelocytic leukemia cells[17]. The present study indicates for the first time thatalveolar type II cells also possess the ability to convert1a,25ðOHÞ2D3 into 1a,25ðOHÞ2-3-epi-D3. Not only is1a,25ðOHÞ2-3-epi-D3 produced in specific tissues, but itis also found to possess significant in vitro biologicalactivities in the tissues in which it is produced. For ex-ample, keratinocytes produce 1a,25ðOHÞ2-3-epi-D3which is almost equipotent to 1a,25ðOHÞ2D3 in inhib-iting keratinocyte proliferation [19,20]. In another study,bovine parathyroid cells produce 1a,25ðOHÞ2-3-epi-D3which was found to be almost equipotent to 1a,25ðOHÞ2D3 in suppressing parathyroid hormone secretion inbovine parathyroid cells [21]. Further, in the presentstudy we show for the first time that alveolar type II cellsproduce 1a,25ðOHÞ2-3-epi-D3 which, like its parent,1a,25ðOHÞ2D3, was significantly active in stimulatingsurfactant synthesis in alveolar type II cells. 1a,25ðOHÞ2
Fig. 4. Effects of 1a,25ðOHÞ2D3 and 1a,25ðOHÞ2D3-3-epi-D3 on the expression of SP-B mRNA in H441 cells. The H441 cells were treated for 48 hwith either 1a,25ðOHÞ2D3 or 1a,25ðOHÞ2D3-3-epi-D3 ð10�9MÞ. About 20lg of RNA extracted from cells was subjected to electrophoresis andsubsequently analyzed by Northern blot. Panel A: Immunoblot. SP-B mRNA, 2 kb. Panel B: Densitometric analysis of SP-B mRNA/GADPH
transcripts ratio. Each value represents the means�SE of three experiments with duplicate dishes.
V.K. Rehan et al. / Molecular Genetics and Metabolism 76 (2002) 46–56 53

-3-epi-D3 stimulates surfactant phospholipid lipid syn-thesis (assessed by [3H] incorporation into saturatedPC), increases surfactant SP-B mRNA gene expression(assessed by RT-PCR), and increases surfactant SP-Bprotein synthesis (assessed by Western blot). Bothsurfactant phospholipid and surfactant SP-B protein arecritical components of pulmonary surfactant. Thus, ourpresent study indicates that, in addition to 1a,25ðOHÞ2D3, its natural metabolite 1a,25ðOHÞ2- 3-epi-D3also may play a potentially important role in lung de-velopment and function.The biological activities of 1a,25ðOHÞ2D3 and its
analogs are primarily vitamin D receptor mediated.Therefore, in the present study we measured the abilityof 1a,25ðOHÞ2-3-epi-D3 to generate VDR-mediatedtranscription. Our results indicate that the VDR-medi-ated transcriptional activity of 1a,25ðOHÞ2-3-epi-D3 ininducing growth hormone production in ROS 17/2.8cells transfected with an osteocalcin VDRE/growthhormone gene construct is significantly reduced whencompared to its parent compound at the various con-
centrations tested (0.0001–1000 nM). Also, a previousstudy measured the transcriptional activities of 3-epimetabolites of 1a,25ðOHÞ2D3 and its analogs, namely,1a,25ðOHÞ2-16-ene-D3, 1a,25ðOHÞ2-16-ene-23-yne-hexafluro-D3 using a single concentration (10 nM). The re-sults indicated that the transcriptional activities of 3-epimetabolites of various vitamin D compounds were sig-nificantly reduced when compared to their parent com-pounds [35]. These findings suggest that the ‘3-epi’modification significantly reduces VDR-mediated tran-scription. Furthermore, the reduced transcriptional ac-tivity of 1a,25ðOHÞ2-3-epi-D3 can be explained based onits binding affinity to the VDR which is only 24% of thatof 1a,25ðOHÞ2D3 [19]. Nonetheless, the high metabolicstability of 1a,25ðOHÞ2-3-epi-D3 [21], as proposed by usearlier, may be an explanation for the high in vitro ac-tivity in spite of reduced binding to the VDR [19] andreduced VDR-mediated transcriptionat activity. Thesimilar ED50s of 3-epi-1; 25ðOHÞ2D3 in comparison tothat of 1; 25ðOHÞ2D3, despite its lower VDR affinity,again emphasize that a greater metabolic stability might
Fig. 5. Effects of 1a,25ðOHÞ2D3 and 1a,25ðOHÞ2D3-3-epi-D3 on the expression of SP-B protein in H441 cells. The cells were treated for 48 h witheither 1a,25ðOHÞ2D3 or 1a,25ðOHÞ2D3-3-epi-D3 ð10�9MÞ. About 25 lg of protein extracted from the cells was subjected to SDS–PAGE and
subsequently analyzed by Western blot. Panel A: Immonoblot. SP-B protein, 42 kD. Panel B: Densitometric analysis of the protein band. Data are
expressed as percentage of control cells not treated with the vitamin D3 compounds. Each value represents the means� SE of three experiments withduplicate dishes.
54 V.K. Rehan et al. / Molecular Genetics and Metabolism 76 (2002) 46–56

be contributing to its significant functional activity.However, it is important to note that since ROS 17/2.8 cells do not metabolize vitamin D compounds effi-ciently, it is unlikely that the metabolic stability of3-epi-1; 25ðOHÞ2D3 will increase its activity relative to1,25ðOHÞ2D3 in these cells, but is likely to do so in cellswith active metabolic pathways such as pulmonary al-veolar type II cells.EB1213, a synthetic vitamin D analog has been
shown to possess significant activity in terms of syn-thesis and release of DSPC by alveolar type II cellswithout generating significant in vivo calcemic activity[8]. Our present study indicates for the first time that1a,25ðOHÞ2-3-epi-D3, a natural metabolite of 1a,25ðOHÞ2D3 (which was previously shown to be lesscalcemic; [19] produced by the alveolar type II cells, alsopossessed significant activity in its ability to stimulatesurfactant synthesis in these cells. This finding suggeststhat the ability of 1a,25ðOHÞ2D3 to stimulate surfactantsynthesis was maintained even after its metabolism into1a,25ðOHÞ2-3-epi-D3. Further, our results also suggestthat 1a,25ðOHÞ2-3-epi-D3, the noncalcemic natural in-termediary metabolite of 1a,25ðOHÞ2D3 may have sig-nificant therapeutic value.In summary, we report that 1a,25ðOHÞ2D3 is me-
tabolized in pulmonary alveolar type II cells into severalintermediary metabolites derived via both the C-24 ox-idation and the C-3 epimerization pathways. 1a,25ðOHÞ2-3-epi-D3, a natural intermediary metabolite of1a,25ðOHÞ2D3 derived via the C-3 epimerization path-way was found to retain significant biological activity ofits parent in terms of its ability to stimulate surfactant
phospholipid and protein synthesis in alveolar type IIcells.
Acknowledgments
This work was supported by research grants fromNational Institutes of Health to Dr. G.S. Reddy(DK52488) and Dr. S. Peleg (DK50583). We also ac-knowledge the expert technical assistance of Mei-LingSiu-Caldera in performing HPLC.
References
[1] M. Nguyen, H. Guillozo, M. Garabedian, S. Balsan, Lung as a
possible additional target organ for vitamin D during fetal life in
the rat, Biol. Neonate 52 (1987) 232–240.
[2] F.B. Askin, C. Kuhn, The cellular origin of pulmonary surfactant,
Lab. Invest. 25 (1971) 260.
[3] M.C. Williams, Development of the alveolar structure of the fetal
rat in late gestation, Fed. Proc. 36 (1977) 2653.
[4] T.M. Nguyen, H. Guillozo, L. Martin, M.E. Dufour, C. Tordet,
J.W. Pike, M. Garabedian, 1,25-Dihydroxyvitamin D3 receptors
in rat lung during the perinatal period: regulation and immuno-
histochemical localization, Endocrinology 127 (1990) 1755–1762.
[5] L. Marin, M.E. Dufour, T.M. Nguyen, C. Tordet, M. Garabe-
dian, Maturational changes induced by 1a,25-dihydroxyvitaminD3 in type II cells from fetal rat lung explants, Am. J. Physiol. 265
(9) (1993) L45–L52.
[6] J.D. Edelson, S. Chan, D. Jassal, M. Post, A.K. Tanswell,
Vitamin D stimulates DNA synthesis in alveolar type II cells,
Biochim. Biophys. Acta 1211 (1994) 159–166.
[7] L. Marin, M.E. Dufour, C. Tordet, M. Nguyen, 1; 25ðOHÞ2D3stimulates phospholipid biosynthesis and surfactant release in
fetal rat lung explants, Biol. Neonate 57 (1990) 257–260.
[8] T.M. Nguyen, H. Guillozo, L. Martin, C. Tordet, S. Koite, M.
Garabedian, Evidence for a vitamin D paracrine system regulating
maturation of developing rat lung epithelium, Am. J. Physiol. 271
(15) (1996) L392–L399.
[9] G.S. Reddy, K-Y. Tserng, Calcitroic acid end product of renal
metabolism of 1,25-dihydroxyvitamin D3 through C-24 oxidation
pathway, Biochemistry 28 (1989) 1763–1769.
[10] G. Makin, D. Lohnes, V. Byford, R. Ray, G. Jones, Target cell
metabolism of 1a,25-dihydroxyvitamin D3 to calcitroic acid,Biochem. J. 262 (1989) 173–180.
[11] R. Bouillon, W.H. Okamura, A.W. Norman, Structure–function
relationships in the vitamin D endocrine system, Endocrine Rev.
16 (2) (1995) 202–257.
[12] A.J. Brown, A. Dusso, E. Slatopolsky, Vitamin D, Am. J. Physiol.
277 (2 Pt 2) (1999) F157–F175.
[13] S. Ishizuka, A.W. Norman, Metabolic pathways from 1a,25-dihydroxyvitamin D3 to 1a,25-dihydroxyvitamin D3-26,23-lac-tone, J. Biol. Chem. 262 (1987) 7165–7170.
[14] G.S. Reddy, K.R. Muralidharan, W.H. Okamura, K.-Y. Tserng,
J.A. McLane, Metabolism of 1a,25-dihydroxyvitamin D3 and oneof its A-ring diastereomer 1a,25-dihydroxy-3-epi-vitamin D3 inneonatal human keratinocytes, in: A.W. Norman, R. Bouillon, M.
Thomasset (Eds.), Vitamin D a pluripotent steroid hormone:
structural studies, molecular endocrinology and clinical applica-
tions, Walter de Gruyter, New York, 1994, pp. 172–173.
[15] G.S. Reddy, M.-L. Siu-Caldera, I. Schuster, N. Astecker, K.-Y.
Tserng, K.R. Muralidharan, W.H. Okamura, J.A. McLane, M.R.
Uskokovic, Target tissue specific metabolism of 1a,25-di-
Fig. 6. Transcriptional activity of 1a,25ðOHÞ2D3 and 1a,25ðOHÞ2D3-3-epi-D3 in ROS 17/2.8 cells. The cells were transfected by the DEAE-
dextran method with the fusion gene containing the osteocalcin VDRE
attached to the thymidine kinase/growth hormone reporter gene. Im-
mediately after transfection, the cells were treated with either
1a,25ðOHÞ2D3 or 1a,25ðOHÞ2D3-3-epi-D3 for 24 h in media contain-ing 10% FCS. Culture medium was assayed for growth hormone
production by radioimmunoassay (Nichols Institute). Each point of
the dose–response curve is the mean of duplicate transfections and the
results shown are representative of four transfection experiments.
V.K. Rehan et al. / Molecular Genetics and Metabolism 76 (2002) 46–56 55

hydroxyvitamin D3 through A-ring modification, in: A.W. Nor-
man, R. Bouillon, M. Thomasset (Eds.), Vitamin D, Chemistry,
Biology and Clinical Applications of the Steroid Hormone,
University of California, Printing and Reprographics, Riverside,
CA, USA, 1997, pp. 139–146.
[16] G.S. Reddy, K.R. Muralidharan, W.H. Okamura, K.-Y. Tserng,
J.A. McLane, Metabolism of 1a,25-dihydroxyvitamin D3, and oneof its C-3 epimer, 1a,25-dihydroxy-3-epi-vitamin D3 in neonatalhuman kertinocytes, Steroids 66 (3–5) (2001) 441–450.
[17] D. Sunita Rao, M.J. Campbell, H.P. Koeffler, S. Ishizuka, M.R.
Uskokovic, P. Spagnuolo, G.S. Reddy, Metabolism of 1a,25-dihydroxyvitamin D3 in human promyelocytic leukemic [HL-60]
cells: in vitro biological activities of the natural metabolites of
1a,25-dihydroxyvitamin D3 produced in HL-60 cells, Steroids 66(3–5) (2001) 423–432.
[18] G.S. Reddy, D. Sunita Rao, M.-L. Siu-Caldera, Natural metab-
olites of 1a,25-dihydroxyvitamin D3 and its analogs, in: A.W.Norman, R. Bouillon, M. Thomasset (Eds.), Vitamin D Endo-
crine System, Structural, Biological, Genetic and Clinical Aspects,
University of California, Printing and Reprographics, Riverside,
CA, USA, 2000, pp. 139–146.
[19] A.W. Norman, R. Bouillon, M.C. Farach-Carson, J.E. Bishop,
L.X. Zhou, I. Nemere, J. Zhao, K.R. Muralidharan, W.H.
Okamura, Demonstration that 1b,25-dihydroxyvitamin D3 is anantagonist of the nongenomic but not genomic biological
responses and biological profile of the three A-ring diastereomers
of 1a,25-dihydroxyvitamin D3, J. Biol. Chem. 268 (1993) 20022–20030.
[20] I. Schuster, N. Astecker, H. Egger, G. Herzig, G.S. Reddy, J.
Schmid, G. Vorisek, Vitamin D-metabolism in human keratino-
cytes and biological role of products, in: A.W. Norman, R.
Bouillon, M. Thomasset (Eds.), Vitamin D, Chemistry, Biology
and Clinical Applications of the Steroid Hormone, University of
California, Printing and Reprographics, Riverside, CA, USA,
1997, pp. 551–558.
[21] A.J. Brown, C. Ritter, E. Slatopolsky, K.R. Muralidharan, W.H.
Okamura, G.S. Reddy, 1a,25-Dihydroxy-3-epi-vitamin D3 a
natural metabolite of 1a,25-dihydroxyvitamin D3, is a potentsuppressor of parathyroid hormone secretion, J. Cell. Biochem. 73
(1999) 106–113.
[22] G.S. Reddy, K.-Y. Tserng, B.R. Thomas, R. Dayal, A.W.
Norman, Isolation and identification of l,23-dihydroxy-24,25,
26,27-tetranorvitamin D3 a new metabolite of 1,25-dihydroxyvi-
tamin D3 produced in rat kidney, Biochemistry 26 (1987) 324–331.
[23] J.S. Torday, J. Hua, R. Slavin, Metabolism and fate of fetal rat
lung fibroblast neutral lipids, Biochim. Biophys. Acta 1254 (1995)
198–206.
[24] P. Chomczynski, N. Sacchi, Single-step method of RNA isolation
by acid guanidinium thiocynate-phenol chloroform extraction,
Anal. Biochem. 162 (1987) 156–159.
[25] O. Ayad, H.R. Wong, Nitric oxide decreases surfactant protein A
gene expression in H441 cells, Crit. Care Med. 26 (1998) 1277–
1282.
[26] J.C. Clark, S.E. Wert, C.J. Bachurski, M.T. Stahlman, B.R.
Stripp, T.E. Weaver, J.A. Whitsett, Targeted disruption of the
surfactant protein G gene disrupts surfactant homeostasis, causing
respiratory failure in newborn mice, Proc. Natl. Acad. Sci. 92
(1995) 7794–7798.
[27] K. Ozono, J. Liao, S.A. Kerner, R.A. Scott, J.W. Pike, The
vitamin D-responsive element in the human osteocalcin gene.
Association with a nuclear proto-oncogene enhancer, J. Biol.
Chem. 265 (35) (1990) 21881–21888.
[28] M.A. Lopata, D.W. Cleveland, B. Sollner-Webb, High level
transient expression of a chloramphenicol acetyl transferase gene
by DEAE-dextran mediated DNA transfection coupled with a
dimethyl sulfoxide or glycerol shock treatment, Nucl. Acids Res.
12 (14) (1984) 5707–5717.
[29] J.F.T. Glasgow, P.S. Thomas, Rachitic respiratory distress in
small preterm infants, Arch. Dis. Child. 52 (1977) 262–273.
[30] C. Gaultier, A. Harf, N. Balmain, P. Cuisinier-Gleizes, H.
Mathieu, Lung mechanics in rachitic rats, Am. Rev. Respir. Dis.
130 (1984) 1100–1108.
[31] P.K. Dudeja, E.S. Foster, T.A. Brasitus, Modulation of rat
colonic brush-border membrane Naþ–Hþ exchange by dexameth-
asone role of lipid fluidity, Biochim. Biophys. Acta 905 (1987)
485–493.
[32] M-L. Siu-Caldera, H. Sekimoto, A. Weiskopf, P. Vouros, K.R.
Muralidharan, W.H. Okamura, J. Bishop, A.W. Norman, M.R.
Uskokovic, I. Schuster, G.S. Reddy, Production of 1a,25-dihydroxy-3-epi-vitamin D3 in two rat osteosarcoma cell lines
(UMR 106 and ROS 17/2.80: existence of the C-3 epimeri-
zation pathway in ROS 17/2.8 cells in which the C-24
oxidation pathway is not expressed, Bone 24 (5) (1999) 457–
463.
[33] M.G. Bischof, M.-L. Siu-Caldera, A. Weiskopf, P. Vouros, H.S.
Cross, M. Peterlik, G.S. Reddy, Differentiation-related pathways
of 1a,25-dihydroxycholecalciferol metabolism in human colon
adenocarcinoma-derived Caco-2 cells: production of 1a,25-dihydroxy-3-epi-cholecalciferol, Exp. Cell Res. 241 (1998) 194–
201.
[34] M.-L. Siu-Caldera, H. Sekimoto, A.S. Brem, R.B. Bina, G.S.
Reddy, Metabolism of 1a,25-dihydroxyvitamin D3 into 1a,25-dihydroxy-3-epi-vitamin D3 in rat osteosarcoma cells (UMR 106
and ROS 17/2.8) and rat vascular smooth muscle cells, J. Bone
Min. Res. 23 (Suppl) (1998) S426.
[35] K. Nakagawa, Y. Sowa, M. Kurobe, K. Ozono, M.-L. Siu-
Caldera, G.S. Reddy, M.R. Uskokovic, T. Okano, Differential
activities of 1a,25-dihydroxy-16-ene-vitamin D3 analogs and their3-epimers on human promyelocytic leukemia (HL-60) cell differ-
entiation and apoptosis, Steroids 66 (2001) 327–337.
56 V.K. Rehan et al. / Molecular Genetics and Metabolism 76 (2002) 46–56