* metabolismo de proteínas y aminoácidos: biosíntesis de urea
TRANSCRIPT

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
Tema 14. Panorama global del metabolismo de los aminoácidos y ciclo dela urea.
Introducción
Los aminoácidos son un grupo de moléculas de granimportancia cuya estructura química común es R-CH(NH3
+)COO-.Aunque los aminoácidos primarios comparten la facultad depolimerizar el uno con el otro para formar proteínas yaunque algunos de ellos pueden interconvertirse mediantereacciones metabólicas, cada uno de ellos es un compuestoúnico con un metabolismo y unas funciones biológicasindividuales.
Los aminoácidos se encuentran fundamentalmenteformando parte de las proteínas, las cuales representan lamayor parte del peso seco del cuerpo humano. Muchos de losaminoácidos que se incorporan a las proteínas humanasproceden de las proteínas de la dieta a través de suabsorción por el intestino. Sin embargo, dado que lacomposición de las proteínas de la ingesta no secorresponde exactamente con las necesidades del individuo,algunos de los aminoácidos de las proteínas humanas sesintetizan a partir de otros constituyentes de la dieta; elcarbono, hidrógeno y oxígeno pueden derivar de otrosaminoácidos o de precursores glucídicos o lipídicos, peroel nitrógeno procede casi exclusivamente de los aminoácidosde la ingesta.
Los aminoácidos no esenciales son aquellos compuestosnecesarios para las proteínas que pueden ser producidos porel metabolismo humano. Los aminoácidos esenciales no puedenser sintetizados por el cuerpo humano porque sus célulascarecen de los enzimas biosintéticos necesarios.
Requerimiento dietético de aminoácidos
Esenciales No esenciales
143

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
Argininaa
Fenilalaninab
HistidinaIsoleucinaLeucinaLisina
Metioninac
TreoninaTriptófano
Valina
AlaninaAspartatoCisteínaGlutamatoGlicinaProlinaSerina
Tirosina
a La arginina se sintetiza en los tejidos de los mamíferos, pero a unavelocidad insuficiente para satisfacer las necesidades del aminoácidodurante el crecimiento.b Se necesitan grandes cantidades de fenilalanina para formar tirosina,si es que ésta no se suministra adecuadamente a través de la dieta.c La metionina se necesita en grandes cantidades para producircisteína, si es que ésta no se suministra adecuadamente a través de ladieta
Los aminoácidos esenciales y no esenciales son sólouna pequeña parte de los que se encuentran en lanaturaleza. Estos dos grupos incluyen solamente aquellosaminoácidos que deben estar presentes para la síntesis decualquier proteína. Además, se sintetizan dos derivadospara su incorporación a las proteínas, la glutamina apartir del ácido glutámico y la asparagina a partir delácido aspártico, lo que eleva la lista de monómeros de lasproteínas a 20. El resto de los aminoácidos, que se hallansólo en determinadas proteínas, son derivados que seobtienen mediante las denominadas modificacionespostraducción de una o más de las 20 unidades estructuralesprimarias, una vez que se hallan incorporadas en lascadenas polipeptídicas.
La cantidad de nitrógeno en cada individuo se hallaregulada a niveles prácticamente constantes, exceptodurante el crecimiento en que la cantidad debe aumentar enproporción al mismo. No existe ninguna forma de almacenarlas reservas de nitrógeno; sólo una pequeña parte de estenitrógeno se halla en forma de aminoácidos libres u otroscompuestos que pueden utilizarse para la síntesis deaminoácidos. En consecuencia, deben ingerirsefrecuentemente las cantidades adecuadas de aminoácidos.
144

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
Los aminoácidos son sustratos de multitud de procesosbiosintéticos esenciales. Sin embargo, las dietas normalescontienen un exceso de aminoácidos respecto a lasnecesidades de síntesis proteica o de otros constituyentescelulares, con lo que la mayor parte de este exceso sedegrada a productos que, o bien se oxidan para obtenerenergía, o se almacenan en forma de grasa o glucógeno. Seacul fuere la situación, el nitrógeno se libera comoamoniaco. Parte del amoniaco se reutiliza en la síntesis deaminoácidos, parte se destina a otras reaccionesbiosintéticas, algo se excreta a través de la orina, perola mayor parte se convierte en urea, la cual se excreta porlos riñones.
La síntesis de urea tiene lugar principalmente en elhígado, lugar donde también se lleva a cabo la mayor partede la biosíntesis de los aminoácidos no esenciales y unagran parte de la degradación de todos los aminoácidos.
Destinos metabólicos del grupo amino:Reacciones generalesde los aminoácidos.
145

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
1. Transaminación: Entre las reacciones de transferenciade grupos amino, las transaminaciones son especialmenteimportantes. Ellas son catalizadas por las transaminasas(que están implicadas tanto en las rutas catabólicas yanabólicas del metabolismo de aminoácidos). Durante latransaminación, el grupo amino de un aminoácido setransfiere a un 2-oxoácido (-cetoácido). Se forma un 2-oxoácido (-cetoácido) del aminoácido original y unaminoácido del 2-oxoácido original.
Con pocas excepciones, el primer paso de la degradaciónde los L-aminoácidos consiste en la eliminación del grupo -amino para producir el correspondiente -cetoácido. Estamodificación, conocida por transaminación, está catalizadapor las enzimas denominadas aminotransferasas otransaminasas. En estas reacciones de transaminación elgrupo -amino de un aminoácido es transferido al átomo decarbono del -cetoglutarato. No hay desaminación neta sinouna transferencia del grupo -amino desde un aminoácidohasta un -cetoácido. El objetivo de las reacciones detransaminación es recoger los grupos aminos de muchosaminoácidos diferentes en forma de uno solo, el glutamato,que los canalizará hacia rutas biosintéticas o hacia víasque generan productos nitrogenados de excreción.
Las células contienen varias aminotransferasas, muchasde ellas específicas para el -cetoglutarato como aceptor degrupos aminos. Sin embargo difieren en su especificidad parael otro sustrato, el aminoácido que cede el grupo amino, yen función del cual se denominan. Las reacciones catalizadaspor las aminotransferasas son reversibles. Las másimportantes son la aspartato aminotransferasa y la alaninaaminotransferasa.
146

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
La determinación de la actividad de ciertos enzimas ensuero se utiliza habitualmente como parámetro para eldiagnóstico de determinadas patologías. La alaninaaminotransferasa (también denominada glutamato-piruvatotransaminasa o GPT) y la aspartato aminotransferasa(denominada también glutamato-oxalacetato transaminasa oGOT) son muy abundantes en corazón e hígado por lo quedespués de un infarto de miocardio o una hepatitisinfecciosa (u otras lesiones de cualquier órgano) lasaminotransferasas de las células lesionadas pasan altorrente circulatorio pudiéndose determinar suconcentración en el suero (S). Los valores de SGPT y SGOTproporcionan información sobre la gravedad y el estado dela lesión. En el caso de infarto de miocardio, los valoresdel enzima creatina quinasa (SCK) son también muyimportantes ya que es específico del músculo cardíaco.
2. Desaminación oxidativa:Cuando el grupo amino de unaminoácido se libera como amoníaco, recibe el nombre dedesaminación. De particular importancia es ladesaminación oxidativa. En esta reacción, el grupo aminoes liberado en forma de amoníaco y se genera un 2-oxoácido. Un ejemplo de desaminación oxidativa es lacatalizada por el enzima glutamato deshidrogenasa.
Glutamato + NAD+(P) -cetoglutarato + amoníaco +NAD(P)H + H+
147
Reacciones catalizadas por los enzimas alanina
transaminasa y aspartato transaminasa
O
P iru v a to
+
G lu ta m a to
C O O -
CC H 3
O
C O O -
C HH 3 N+
C H 2C O O -
C O O -
C HH 3 N+
C H 3
C O O -
C HH 3 N+
C H 2C H 2C O O -
C O O -
CC H 2C H 2C O O -
+
A la n in a -C e to g lu ta ra to
A la n in atra n s a m in a s a
A s p a rta to
OC O O -
CC H 2C H 2C O O -
-C e to g lu ta ra to
+O
O x a la c e ta to
+
G lu ta m a to
C O O -
CC H 2
C O O -
C HH 3 N+
C H 2C H 2C O O -
A s p a rta totra n s a m in a s a C O O -

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
El glutamato generado en el citosol de las célulashepáticas por las reacciones de transaminación estransportado hasta la matriz mitocondrial donde por acciónde la glutamato deshidrogenasa sufre desaminación oxidativa,liberándose iones amonio. Este enzima únicamente seencuentra en la matriz mitocondrial y es capaz de utilizarcomo aceptores de los equivalentes de reducción NAD+ y NADP+.Sus activadores alostéricos son el GDP y el ADP y losefectores alostéricos negativos son el GTP (generado en elciclo de Krebs) y el ATP. Si un hepatocito necesitacombustible para el ciclo de Krebs aumenta la actividad dela glutamato deshidrogenasa, suministrando -cetoglutaratopara el ciclo de Krebs y liberando NH4
+ para la excreción.Por el contrario, es inhibida si se acumula GTP comoconsecuencia de una alta actividad del ciclo de Krebs. Porlo tanto, la disminución de la carga energética (disminuciónde GTP y ATP) acelera la oxidación de los aminoácidos.
Transporte de NH4+ a través de la sangre
Los iones amonio que se generan durante el metabolismocelular o bacteriano mayoritariamente no circulan librementedisueltos en la sangre sino que lo hacen en forma deaminoácidos y que son glutamina y alanina.
Glutamina: El amonio, derivado principalmente de ladesaminación de los grupos -amino de los aminoácidos es muy
148
FIG. 4 Reacción catalizada por el enzima glutamato deshidrogenasa

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
tóxico para todos los animales, especialmente para elsistema nervioso. En la mayoría de los animales el excesoNH4
+ producido en los diferentes tejidos es convertido englutamina antes de ser transportado, a través de la sangre,hasta el riñón, intestino (mucosa intestinal) y en menormedida al hígado. El glutamato, que es tan importante parael metabolismo de los grupos aminos intracelulares, essustituido por glutamina para dicha función de transporte.Ello se debe a que la glutamina es un compuesto neutro y notóxico, que puede atravesar fácilmente las membranascelulares, lo cual no podría hacer el glutamato porque poseeuna carga neta negativa. En muchos tejidos, incluído elcerebro, el amonio se combina con el glutamato y generaglutamina, reacción catalizada por la glutamina sintetasamitocondrial. Este proceso necesita ATP y transcurre en dosetapas. El enzima se denomina específicamente sintetasa, enlugar de sintasa, porque la reacción está acoplada con lahidrólisis del ATP. Ambas enzimas pertenecen a la claseligasas pero las sintasas no requieren ATP.
Como ocurre con el grupo amino del glutamato, elnitrógeno amídico de la glutamina se libera en forma deiones amonio, en las mitocondrias de los tejidos que captanglutamina (riñón, intestino, y en menor medida hígado). Lareacción, catalizada por el enzima glutaminasa, convierte laglutamina en glutamato y NH4
+.
149
Síntesis de glutamina catalizada por la glutamina sintetasa
C O O -
C HH 3 N +
C H 2C H 2C
P i
ON H 2
L - G l u t a m i n a
G l u t a m i n as i n t e t a s a
OO P
O
O -
O -
C O O -
C HH 3 N +
C H 2C H 2C
A D PA T P
G l u t a m a t o
C O O -
C HH 3 N +
C H 2C H 2C O O -
N H 4+
G l u t a m i n as i n t e t a s a
- G l u t a m i l f o s f a t o
Liberación del grupo amido de la glutamina por acción de la glutaminasa
C O O -
C HH 3 N +
C H 2C H 2CON H 2
L -G lu ta m in aG lu ta m a to
C O O -
C HH 3 N +
C H 2C H 2C O O -
N H 4+
G lu ta m in a s a
H 2 OU r e a

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
Alanina: Desempeña un papel especial en el transporte deamonio hasta el hígado en una forma no tóxica yeléctricamente neutra, mediante el ciclo de la glucosa-alanina. En el músculo y en algunos otros tejidos quedegradan aminoácidos como combustible metabólico, los gruposaminos se canalizan hacia el glutamato mediantetransaminación. El glutamato puede convertirse en glutaminapara su transporte a los tejidos capaces de captarla(fundamentalmente riñón e intestino), o puede transferir sugrupo amino al piruvato, producto fácilmente asequible de laglucólisis muscular, por acción del enzima alaninaaminotrasferasa. La alanina es neutra a pH fisiológico, porlo que pasa a la sangre y finalmente al hígado. En elhígado, el exceso de nitrógeno es descargado en lasmitocondrias de los hepatocitos en forma de NH4
+. Para ello,se invierten las reacciones que tuvieron lugar en elmúsculo. En el citosol, la alanina aminotransferasa hepáticatransfiere el grupo amino de la alanina al -cetoglutaratopara formar glutamato. Parte del glutamato formado setransporta a las mitocondrias donde, por acción de laglutamato deshidrogenasa, se libera NH4
+ que es utilizadopara la síntesis de urea. Alternativamente, en el citosoltambién puede transferirse el grupo amino del glutamato aloxalacetato para formar aspartato, catalizado por laaspartato aminotransferasa. El aspartato es el otro dador denitrógeno para la formación de urea.
Transporte del amoníaco al hígado para la síntesis de urea
150

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
La utilización de la alanina para transportar amoniodesde los músculos esqueléticos muy activos al hígado es unejemplo de la economía intrínseca de los organismos vivos.Los músculos esqueléticos sometidos a contracción vigorosaoperan en anaerobiosis y producen, no sólo amonio a partirde la degradación de las proteínas sino grandes cantidadesde piruvato a partir de la glucólisis. Los dos productos vanal hígado: el amonio para ser convertido en urea que seráexcretada y el piruvato para formar glucosa y volver a losmúsculos. Mediante el ciclo se resuelven dos problemas,transportar los átomos de carbono del piruvato y los ionesamonio desde el músculo al hígado en forma de alanina. En elhígado, la alanina forma piruvato que entra en lagluconeogénesis y libera NH4
+ para la síntesis de urea.
Excreción del NH4+
Aunque el amonio es un participante universal en lasíntesis y degradación de los aminoácidos, su acumulación enconcentraciones anormales tiene consecuencias tóxicas. Porlo tanto, las células con un catabolismo de aminoácidos muyactivo deben ser capaces de realizar la desactivación tóxicay/o excreción del amonio con la misma rapidez con que éstese genera. Existen varias estrategias para la eliminacióndel amonio y en base a las mismas, los animales puedenclasificarse en:
151

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
Amonotélicos: Excretan amonio y entre ellos están la mayoríade los animales acuáticos que, al ser capaces de captar yexpulsar agua continuamente, el amonio se elimina a travésdel líquido expulsado. En el agua del hábitat el amonio sediluye.
Uricotélicos: Excretan el amonio en forma de ácido úrico yentre ellos se encuentran aves, reptiles e insectos. Elácido úrico es bastante insoluble de modo que se excreta sinuna pérdida importante de agua que haga variar la presiónosmótica. La biosíntesis de ácido úrico tiene lugar a travésde la ruta de la biosíntesis de nucleótidos de purina.
Ureotélicos: Excretan el amonio en forma de urea y en estegrupo se encuentran la mayor parte de los mamíferos yanfibios adultos. La urea es muy soluble en agua, pero alcarecer de cargas su acumulación no afecta al pH como ocurrecon el NH4
+.
Ciclo de la urea
En los organismos ureotélicos, como los seres humanos,aproximadamente el 80% del nitrógeno total excretado estápresente en forma de urea. La síntesis de urea se realiza enel denominado ciclo de la urea, mediante un conjunto deenzimas que actúan coordinadamente. Aunque muchas de esasenzimas suelen estar presentes en la mayor parte de lostejidos de los mamíferos, el ciclo funciona únicamente en elhígado. En la producción de urea a partir de NH4
+ intervienencinco reacciones enzimáticas, dos en la matriz mitocondrialy tres en el citosol.
152

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
Reacciones que transcurren en la matriz mitocondrial:
a) Síntesis de carbamilfosfato. El primer grupo amino que entra enel ciclo de la urea proviene del NH4
+ presente en elinterior de las mitocondrias. Este a su vez procede de lasrutas ya descritas así como de la oxidación de aminoácidospor las bacterias del tracto intestinal y que llega alhígado a través de la vena porta. El NH4
+ se combina con CO2
(en forma de HCO3-) procedente de la respiración
mitocondrial, produciendo carbamil fosfato. La reacción estácatalizada por la carbamil fosfato sintetasa I. La hidrólisis dedos moléculas de ATP asegura que el proceso de síntesis seairreversible. El carbamil fosfato es un dador activado degrupos carbamilo.
b) Síntesis de citrulina. El carbamil fosfato cede su grupocarbamilo a la ornitina para formar citrulina, reaccióncatalizada por la ornitina transcarbamilasa. La citrulina,mediante un transportador específico presente en la membranamitocondrial interna, es enviada al citosol.
Reacciones que transcurren en el citosol:
a) Síntesis de argininosuccinato. La citrulina y el aspartato(procedente de la matriz mitocondrial y generado portransaminación) se condensan para formar argininosuccinato,proceso favorecido por la hidrólisis del ATP en AMP y PPi,
153

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
y posterior hidrólisis del pirofosfato. Está catalizado porel enzima argininosuccinato sintetasa. El segundo grupo amino quese introduce en el ciclo de la urea lo hace en forma deaspartato.
b) Rotura de argininosuccinato. Por acción del enzimaargininosuccinato liasa el argininosuccinato es escindido enarginina, que es el aminoácido precursor de la urea, yfumarato que entra a formar parte de los intermediarios delciclo de Krebs.
c) Hidrólisis de arginina. Se genera ornitina y urea, procesocatalizado por el enzima arginasa. Este enzima es elresponsable de la naturaleza cíclica de la ruta de labisosíntesis de la urea. Prácticamente todos los organismossintetizan arginina a partir de ornitina, mediante lasreacciones mostradas. Sin embargo, únicamente los organismosureotélicos contienen arginasa. El destino de la ornitina esvolver otra vez a la matriz mitocondrial para su utilizaciónen un nuevo ciclo.
154

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
Ciertos organismos presentan más de una forma deexcreción del amonio durante su ciclo vital. Un caso muypeculiar es el del renacuajo puesto que antes de sufrir lametamorfosis es de vida acuática y amonotélico mientras quesu forma adulta, la rana, es de vida terrestre y ureotélica.Ello se debe a que durante el proceso de metamorfosis seinduce la síntesis del enzima arginasa.
Eliminación de la urea
La urea abandona el hígado y pasa al sistemacirculatorio a través del cual llega a los riñones donde es
155

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
filtrada para su excreción. La determinación de laconcentración de urea en sangre es un indicador clínico dela función renal ya que la filtración y eliminación de urease ven afectados cuando hay una actividad renal deficiente.Existen excepciones en el caso de animales que hibernan yaque durante el período de hibernación no orinan y la urea,presente en la vejiga urinaria, se reabsorbe y vuelve a lostejidos donde aporta grupos aminos para la biosíntesis deaminoácidos.
Regulación del ciclo de la urea
La actividad del ciclo de la urea va a estarcondicionada por la composición de la dieta. Supongamos lasdos situaciones metabólicas siguientes: por un lado la de unindividuo alimentado con una dieta constituida esencialmentepor proteínas y, por otro, la de un organismo sometido ainanición severa. En ambos casos los aminoácidos (esqueletoshidrocarbonados) serán utilizados como principal fuente deenergía y se producirá abundante urea a partir de los gruposaminos excedentes. Los enzimas del ciclo y lacarbamilfosfato sintetasa I van a estar regulados a dosniveles.
Concentración de los enzimas: los enzimas del ciclo de laurea (incluido carbamilfosfato sintetasa I) sonsintetizadas a una velocidad superior cuando se ingiere unadieta rica en proteínas que cuando se consume una dietaequilibrada (abundan glúcidos y lípidos). Lo mismo esaplicable cuando se trata de inanición ya que las proteínasmusculares van a actuar como principal fuente de energíametabólica. Y al contrario, cuando no se consumen proteínasla velocidad de síntesis disminuye. Se trata de un mecanismode regulación que funciona a largo plazo.
Regulación alostérica: es ejercida sobre el enzimacarbamilfostato sintetasa I. Su activador alostérico es elN-acetilglutamato que, a su vez, se sintetiza a partir deacetil-CoA y glutamato por acción del enzima N-acetilglutamato sintetasa mitocondrial. El enzima N-acetilglutamato sintetasa es activada por arginina,intermediario del ciclo de la urea, que se acumula cuando la
156

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
síntesis de urea va muy lenta en comparación con laproducción de amonio a partir del catabolismo deaminoácidos. Este mecanismo de regulación es el primero quese ejerce para intentar controlar el flujo de nitrógeno através del ciclo y por lo tanto se trata de una regulación acorto plazo.
Balance químico total del ciclo de la urea
NH3 + CO2 + 2 ATP carbamoilfosfato + 2 ADP + 2PiCarbamoilfosfato + ornitina citrulina + PiCitrulina + ATP + Aspartato arginosuccinato + AMP + PPi
Arginosuccinato arginina + fumarato Arginina urea + ornitina
2 NH3 + CO2 + 3 ATP urea + 2 ADP + AMP + PPi + 2 Pi
Destinos metabólicos del esqueleto hidrocarbonado
Una vez se ha separado el grupo NH4+, el catabolismo del
esqueleto hidrocarbonado resultante va a generar dos tiposde moléculas: aquellas cuyo destino final es la oxidación enel ciclo de Krebs (acetil-CoA o acetacetil-CoA) o moléculasde naturaleza anfibólica (algunos intermediarios del ciclode Krebs) capaces de generar glucosa. Aunque cada uno delos veinte aminoácidos tiene su propia ruta para ser
157
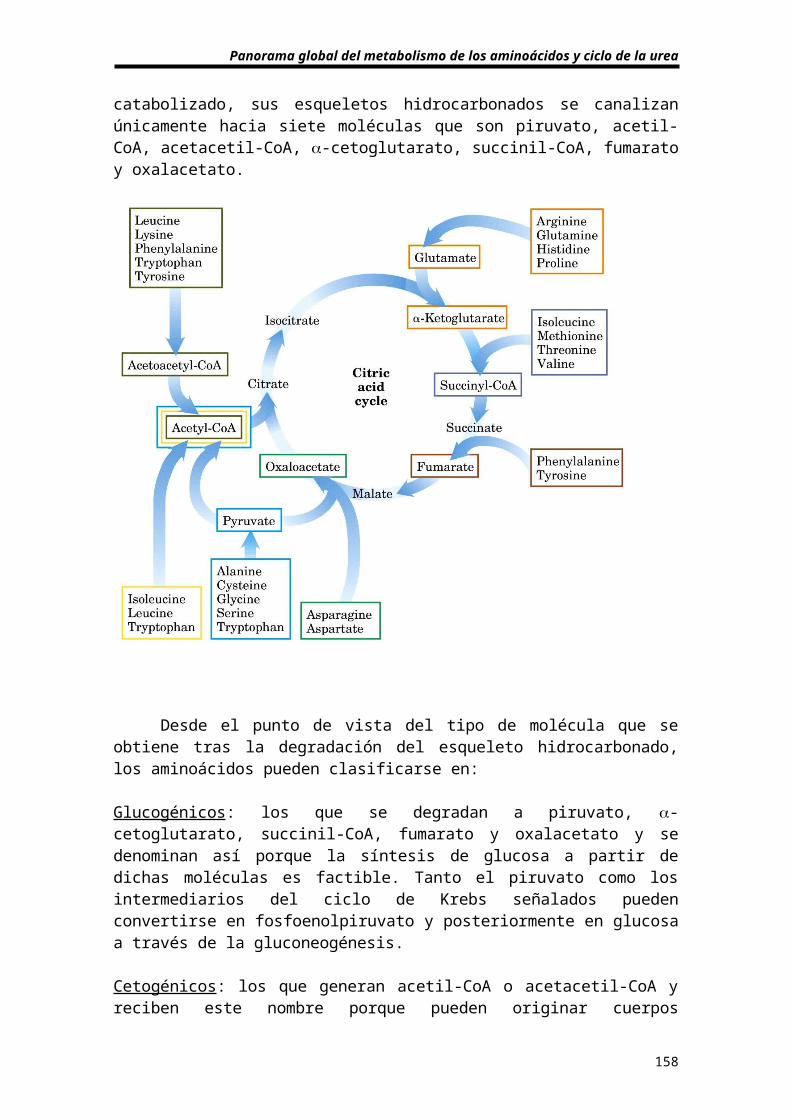
Panorama global del metabolismo de los aminoácidos y ciclo de la urea
catabolizado, sus esqueletos hidrocarbonados se canalizanúnicamente hacia siete moléculas que son piruvato, acetil-CoA, acetacetil-CoA, -cetoglutarato, succinil-CoA, fumaratoy oxalacetato.
Desde el punto de vista del tipo de molécula que seobtiene tras la degradación del esqueleto hidrocarbonado,los aminoácidos pueden clasificarse en:
Glucogénicos: los que se degradan a piruvato, -cetoglutarato, succinil-CoA, fumarato y oxalacetato y sedenominan así porque la síntesis de glucosa a partir dedichas moléculas es factible. Tanto el piruvato como losintermediarios del ciclo de Krebs señalados puedenconvertirse en fosfoenolpiruvato y posteriormente en glucosaa través de la gluconeogénesis.
Cetogénicos: los que generan acetil-CoA o acetacetil-CoA yreciben este nombre porque pueden originar cuerpos
158

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
cetónicos. Puesto que los mamíferos carecen del sistemaenzimático adecuado, estos compuestos nunca podrán serutilizados como precursores para la biosíntesis de glucosa.
De los veinte aminoácidos universales, catorce sonpuramente glucogénicos y dos puramente cetogénicos (leucinay lisina). Los cuatro restantes (isoleucina, fenilalanina,triptófano y tirosina) son glucogénicos y cetogénicossimultaneamente ya que una parte del esqueletohidrocarbonado origina precursores para la biosíntesis de laglucosa (piruvato o intermediarios del ciclo de Krebs) y laotra parte acetil-CoA o acetacetil-CoA.
Glucogénicos
Cetogénicos
Alanina Arginina Asparragina Aspartato Cisteína Glutamina Glutamato Glicina Histidina Isoleucina
Metionina Fenilalanina Prolina Serina
Isoleucina Leucina Lisina
Fenilalanina
Triptófano Tirosina
159
Clasificación de los aminoácidos enfunción
del destino del esqueleto

Panorama global del metabolismo de los aminoácidos y ciclo de la urea
Treonina Triptófano Tirosina Valina
160