dna polymerase α does not mediate g0–g1 increase in yield of x-ray-induced exchange aberrations...
TRANSCRIPT

Mutation Research, 244 (1990) 111-114 111 Elsevier
MUTLET 0346
D N A polymerase ct does not mediate Go-G 1 increase in yield of X-ray- induced exchange aberrations in human peripheral blood lymphocytes*
M . A Bender a and Ruth C. M o o r e b
aMedical Department, Brookhaven National Laboratory, Upton, N Y 11973 (U.S.A.) and b The Cancer Institute, Peter MacCallum Hospital, Melbourne, Vic. 3000 (Australia)
(Accepted 21 December 1989)
Keywords: DNA polymerase ~; Blood lymphocytes, human peripheral; Phytohaemagglutinin stimulation; Butylphenyl deoxyguanosine
Summary
We report experiments to test the hypothesis that the increased yield of dicentric chromosomes observed in human peripheral blood lymphocytes treated with X-rays during the G1 phase of their first cell cycle, as compared with the yield when the cells are treated in their Go phase prior to phytohemagglutinin stimulation, is a manifestat ion of the recently-reported conversion of an inactive fo rm of DNA polymerase c~ to its active fo rm as the PHA-st imulated cells pass f rom Go into G1 (Sylvia et al., 1988). The specific polymerase o~ in- hibitor butylphenyl deoxyguanosine was used as an X-ray post-treatment. The results show that polymerase
a is not involved.
When human peripheral blood lymphocytes
(HPBL) are stimulated to divide by phytohemag- glutinin (PHA), the yield of dicentrics induced by a dose of X-rays increases by 50°7o or more as the
cells progress f rom Go to GI (Beck and Obe, 1977; Holmberg, 1976; Wolff , 1972), though the yield of
*Research carried out under the auspices of the United States Department of Energy under Contract No. DE-AC02- 76CH00016. By acceptance of this article, the publisher and/or recipient acknowledges the U.S. Government's right to retain a nonexclusive, royalty-free license in and to any copyright covering this paper.
Correspondence: Dr. M.A Bender, Medical Department, Brookhaven National Laboratory, Upton, NY 11973 (U.S.A.).
deletions does not appear to increase (Gundy and Bender, 1984). This change in dicentric yield ap- pears to explain the greater aberrat ion yield seen in irradiated H P B L f rom persons with Down syn- drome; the Down syndrome lymphocytes ap- parently are in a state more nearly of delayed G1 than in a true Go while in the peripheral circulation
(Dekaban et al., 1966; Leonard and Merz, 1983). We have shown that the frequency of aberra-
tions in HPBL treated with X-rays in Go is greatly increased by a post- treatment with a replicative D N A synthesis polymerase inhibitor, aphidicolin (Bender and Preston, 1982), and more recently that this increase is attributable to D N A pol ~, not to pol et (Bender et al., 1989). However, it has also recently been shown that the pol a in Go cells is
0165-7992/90/$ 03.50 © 1990 Elsevier Science Publishers B.V. (Biomedical Division)

112
largely, if not entirely, in an inactive form, and that this is rapidly converted to the active enzyme by a phosphatidyl inositol second messenger system when cells are stimulated to divide (Sylvia et al., 1988). It thus seemed possible that the activation of pol t~ as HPBL progress f rom Go to G1 following stimulation with an agonist such as P H A might possibly mediate the change in X-ray-induced aber- ration yields. We here report the results of ex- periments to test whether this is so.
Materials and methods
Two separate experiments were done. Whole, heparinized blood samples f rom healthy volunteers were put into culture by adding 1-ml aliquots to 10 ml of medium RPMI 1640 (Gibco), containing 15% fetal calf serum, penicillin and streptomycin, in plastic 15-ml conical centrifuge tubes.
The DNA polymerase inhibitor butylphenyl deoxyguanosine (BuPdG), the tr iphosphate of which selectively inhibits pol a (Lee et al., 1985) was used. We have shown these inhibitors to be equally effective in inhibiting replicative DNA syn- thesis in HPBL (Bender and Moore, 1988; Bender et al., 1989). Both were dissolved in dimethyl sulfoxide (DMSO) and used at a final concentra- tion of 5 x 10-5 M.
Cultures were treated, either prior to P H A stimulation or 3 or 6 h after P H A stimulation and incubation, with 200 R of X-rays from a General Electric Maxitron 250 machine (250 kVp, 30 mA, 0.5 mm C u + 1.0 mm AI added filtration) at a dose rate of approx. 85 R/min . 30 min prior to irradia- tion, 10 k of DMSO or of 5 x 10- 2 M solutions of either APC or BuPdG were added to the ap- propriate cultures. Cultures were reincubated for 2 h and then washed twice with medium and rein- cubated in medium containing PHA. Colcemid was added 2 h prior to fixation to a level of 0.1 mg/ml .
Fixations for the first experiment were done after 50 h in culture except for those treated with X-rays plus APC, which were fixed at 53 h because of earlier experience with division delay in cultures so treated in Go. However, there were too few
mitoses to allow scoring of the X-ray plus APC cultures even in spite of the extra 3 h allowed, and mitoses were in fact somewhat sparse even in the X- ray treated DMSO and BuPdG cultures. Conse- quently, for the second experiment all of the
cultures without X-ray treatment were fixed at 50 h, while the X-ray-treated cultures with either sol- vent or BuPdG were fixed at 53 h, and the X-ray plus APC cultures were fixed at 56 h. To offset the possibility of confounding the results through in- clusion of second-division metaphases at the later
fixation times, 5-bromodeoxyuridine was added to cultures in the second experiment at the time of medium replacement after washing. Thus, the fre- quency of second and later mitoses could be deter- mined and if necessary scoring could be confined to first-division mitoses in differentially stained preparations. Differential staining was done by the method of Goto et al. (1975).
Results and discussion
All of the aberration scoring for both ex- periments was done on conventionally-stained slides. Scoring of differentially stained slides of material f rom the second experiment, in which the cultures were exposed to BdrU, showed that no cultures had more than 9% second mitoses (average 3.4%) or had any third or later mitoses. Hence, any significant influence of fixation times on the results appeared impossible.
The results of the 2 Expts. were somewhat dif- ferent, and consequently we have presented the raw data for them separately in Table 1. As may be seen, the yields of chromosome-type deletions were consistently somewhat greater in the second experi- ment than in the first, though the results for the other aberration classes are quite consistent. As this difference appears unrelated to the presence or absence of a BuPdG post-treatment, we have pool- ed the data for the 2 Expts. to calculate average yields of the chromosome-aberrat ion types shown in Table 2.
It is evident that though the expected increase in dicentric yield occurs in the cells irradiated after 3 or 6 h incubation in the presence of PHA, there is
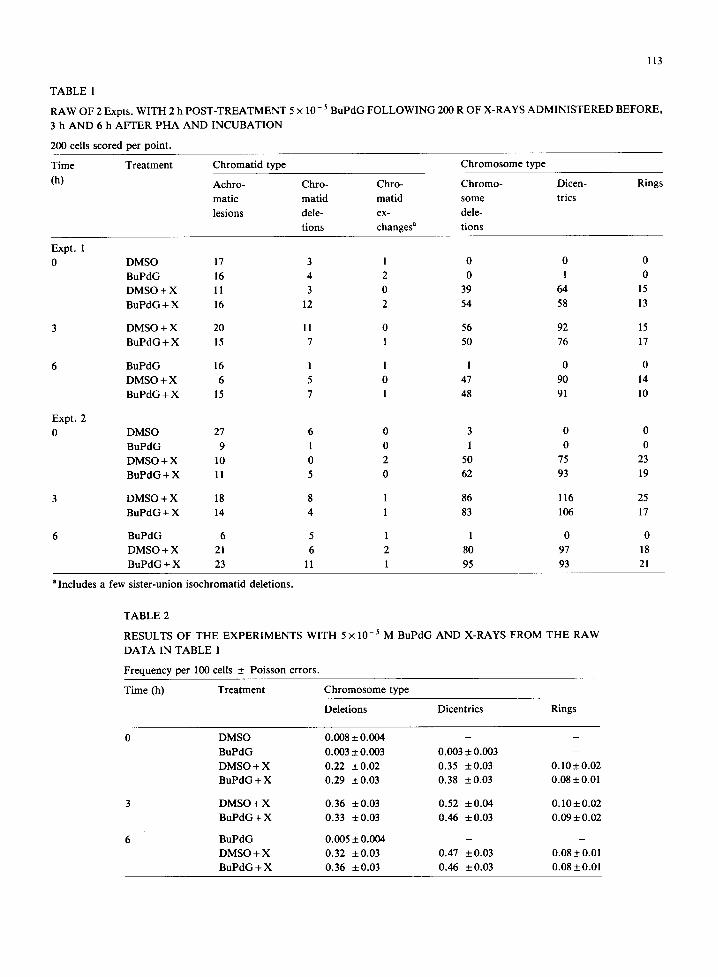
113
TABLE 1
RAW OF 2 Expts. WITH 2 h POST-TREATMENT 5 × 10 - 5 BuPdG FOLLOWING 200 R OF X-RAYS ADMINISTERED BEFORE,
3 h AND 6 h AFTER P H A AND INCUBATION
200 cells scored per point.
Time Treatment Chromat id type Chromosome type
(h) Achro- Chro- Chro- Chromo-
matic matid matid some
lesions dele- ex- dele-
tions changes a tions
Dicen- Rings
trics
Expt. 1
0
Expt. 2
0
DMSO 17 3 1 0 0 0
BuPdG 16 4 2 0 1 0
DMSO + X 11 3 0 39 64 15
BuPdG + X 16 12 2 54 58 13
DMSO + X 20 11 0 56 92 15
BuPdG + X 15 7 1 50 76 17
BuPdG 16 1 1 1 0 0
DMSO + X 6 5 0 47 90 14
BuPdG + X 15 7 1 48 91 10
DMSO 27 6 0 3 0 0
BuPdG 9 1 0 1 0 0
DMSO + X 10 0 2 50 75 23
BuPdG + X 11 5 0 62 93 19
3 DMSO + X 18 8 1 86
BuPdG 4- X 14 4 1 83
116 25
106 17
BuPdG 6 5 1 1 0 0
DMSO + X 21 6 2 80 97 18
BuPdG + X 23 11 l 95 93 21
aIncludes a few sister-union isochromatid deletions.
TABLE 2
RESULTS OF THE EXPERIMENTS WITH 5 × 10- ~ M BuPdG AND X-RAYS FROM THE RAW
DATA IN TABLE 1
Frequency per 100 cells ± Poisson errors.
Time (h) Treatment Chromosome type
Deletions Dicentrics Rings
0 DMSO 0.008 +_ 0.004 - -
BuPdG 0.003 ± 0.003 0.003 ± 0.003 -
DMSO + X 0.22 + 0.02 0.35 + 0.03 0.10 ± 0.02
B u P d G + X 0.29 _+0.03 0.38 _+0.03 0.08+_0.01
3 D M S O + X 0.36 ±0.03 0.52 ±0 .04 0 .10±0.02
B u P d G + X 0.33 +0.03 0.46 ±0.03 0 .09±0.02
6 BuPdG 0.005 ± 0.004 - -
D M S O + X 0.32 _+0.03 0.47 ±0.03 0.08±0.01
B u P d G + X 0.36 _+0.03 0.46 +0.03 0.08_+0.01

114
no effect of BuPdG post-treatment at any of the ir- radiation times.
The aberration yields we observed in our earlier experiments with BuPdG post-treatment of Go H P B L irradiated with the same dose (Bender et al., 1989) were substantially lower than in the present experiments. In the earlier experiments however, we used 1 o70 DMSO, instead of the 0.1 °70 we have generally used for such experiments and that was used for the present ones. Littlefield et al. (1988) have recently shown that l°/0 DMSO, and OH radical scavenger, substantially reduces chromo- somal aberration yields in X-irradiated Go HPBL when present during irradiation, However, the present results for cells irradiated in Go demonstrate that the DMSO reduction in aberra- tion yield did not affect our conclusion that polymerase o~ is not involved in mediating aberra- tion in these cells.
The BuPdG results shown in Table 2 seem clearly to rule out the activation of DNA polymerase ~ as the cause of the increase in dicentric yield in HPBL X-irradiated in Go or in G~. The increase is evident in cells irradiated both 3 and 6 h after P H A stimulation, and yet no difference is seen between the cells post-treated with BuPdG or with solvent. Averaging the dicentric yields for 3 and 6 h treat- ment, the increase over the Go yield is about 30°70. Though somewhat smaller than the maximum we reported earlier for the same dose (Gundy and Bender, 1984), there can be no doubt as to its statistical significance. Had we used the later treat- ment times used in the earlier work (8-12 h), it seems likely that the increase would have been even
greater. The cause of the consistently greater yield of X-
ray-induced deletions in the second experiment compared to the first is unexplained. As the blood donors for the 2 Expts. were different, possibly this represents an idiosyncratic response. However, as dicentric (and ring) yields were the same in the 2 Expts. it does not seem possible that any such dif- ference between donors could affect our conclu- sion that activation of DNA polymerase o~ is not, in fact, responsible for the increased dicentric yields in G~ as compared with Go lymphocytes.
Acknowledgement
We thank Dr. George E. Wright for kindly pro- viding the BuPdG used for these experiments.
References
Beck, B., and G. Obe (1977) Differential chromosomal radiosensitivity within the first Gl-phase of the cell cycle of early-dividing human leukocytes in vitro after stimulation with PHA, Human Genet., 35, 209-218.
Bender, M.A, and R.C. Moore (1988) Dose relationships for different effects of aphidicolin in human peripheral blood leukocytes, Mutation Res., 198,227-231.
Bender, M.A, R.C. Moore and B.E. Pyatt (1989) Role of DNA polymerase a and 6 in radiation clastogenesis, Proc. Natl. Acad. Sci. (U.S.A.), in press.
Dekaban, A.S., R. Thron and J.K. Steusing (1966) Chromo- somal aberrations in irradiated blood and blood cultures of normal subjects and of selected patients with chromosomal abnormality, Radiation Res., 27, 50-63.
Fry, M., and L.A. Loeb (1986) Animal cell DNA polymerases, CRC Press, Boca Raton, FL.
Goto, K., T. Akematsu, H. Shimazu and T. Sugiyama (1975) Simple differential Giemsa staining of sister chromatids after treatment with photosensitive dyes and exposure to light and the mechanism of staining, Chromosoma, 53,223-230.
Gundy, S., and M.A Bender (1984) Increased yields of ex- changes but not deletions in X-irradiated human peripheral lymphocytes following phytohemagglutinin stimulation, Radiation Res., 97, 519-525.
Holmberg, M. (1976) Lack of synergistic effect between X-ray and UV irradiation on the frequency of chromosome aberra- tions in PHA-stimulated human lymphocytes in the GI stage, Mutation Res., 34, 141-148.
Lee, M.Y.W.T., N.L. Toomey and G.E. Wright (1985) Dif- ferential inhibition of human placental DNA polymerase 6 and a by BuPdGTP and BuAdATP, Nucleic Acids Res., 13, 8623-8630.
Leonard, J.C., and T. Mertz (1983) The influence of cell cycle kinetics on the radiosensitivity of Down's syndrome lym- phocytes, Mutation Res., 109, 112-121.
Littlefield, L.G., E.E. Joiner, S.P. Colyer, A.M. Sayer and E.L. Fromes (1988) Modulation of radiation-induced chromosome aberrations by DMSO, on OH radical scavenger, 1. Dose-response studies in human lymphocytes exposed to 220 kV x-rays, Int. J. Radiat. Biol., 53,875-890.
Sylvia, V., G. Curtin, J. Norman, J. Stec and D. Busbee (1988) Activation of a low specific activity form of DNA polymerase tx by inositol-l,4-hiphosphate, Cell, 54, 651-659.
Wolff, S. (1972) The repair of X-ray-induced chromosome aber- rations in stimulated and unstimulated human lymphocytes, Mutation Res., 15, 435-444.
Communicated by F.H. Sobels