distribution of the selma translocon in secondary plastids of red algal origin and predicted...
TRANSCRIPT

Distribution of the SELMA translocon in secondary plastids of red algal origin 1
and predicted uncoupling of ubiquitin-dependent translocation from degradation 2
Simone Stork*1, Daniel Moog*1, Jude M. Przyborski2, Ilka Wilhelmi1’, Stefan Zauner1 3
and Uwe G. Maier#1,3 4
1Laboratory for Cell Biology, Philipps-University Marburg, Karl-von-Frisch Str. 8, D-5
35032 Marburg, Germany. 6
2Laboratory for Parasitology, Philipps-University Marburg, Karl-von-Frisch Str. 8, D-7
35032 Marburg, Germany 8
3LOEWE-Zentrum für Synthetische Mikrobiologie (SynMikro), Hans-Meerwein-Straße, 9
D-35032 Marburg, Germany 10
*These authors contributed equally. 11
‘Present address: Institute for Molecular Tumor Biology and Cancer Gene Therapy, 12
Philipps-University Marburg, Emil-Mannkopff-Str. 2, D-35032 Marburg, Germany. 13
#E-mail: [email protected] 14
Running title: SELMA - uncoupling translocation from degradation 15
Copyright © 2012, American Society for Microbiology. All Rights Reserved.Eukaryotic Cell doi:10.1128/EC.00183-12 EC Accepts, published online ahead of print on 5 October 2012

2
Abstract 16
Protein import into complex plastids of red algal origin is a multistep process including 17
translocons of different evolutionary origin. The symbiont-derived ERAD-like machinery 18
(SELMA), shown to be of red algal origin, is proposed to be the transport system for 19
preprotein import across the periplastidal membrane of heterokontophytes, 20
haptophytes, cryptophytes and apicomplexans. In contrast to the canonical endoplasmic 21
reticulum-associated degradation (ERAD) system, SELMA translocation is suggested to 22
be uncoupled from proteasomal degradation. We investigated the distribution of known 23
and newly identified SELMA components in organisms with complex plastids of red 24
algal origin by intensive data mining, thereby defining a set of core components present 25
in all examined organisms. These include putative pore-forming components, a 26
ubiquitylation machinery, as well as a Cdc48 complex. Furthermore, the set of known 27
20S proteasomal components in the periplastidal compartment (PPC) of diatoms was 28
expanded. These newly identified putative SELMA components as well as proteasomal 29
subunits were in vivo-localized as PPC proteins in the diatom Phaeodactylum 30
tricornutum. The presented data allows us to speculate about the specific features of 31
SELMA translocation in contrast to the canonical ERAD system, especially the 32
uncoupling of translocation from degradation. 33

3
Introduction 34
Organelles like plastids, including those of secondary origin, almost completely rely on 35
protein import from the host cytosol (46, 65). The structure of complex plastids, 36
surrounded by three or four membranes required, in contrast to primary plastids, the 37
evolution of several additional protein transport mechanisms. Complex plastids arose 38
through secondary endosymbiosis, a process which describes the engulfment of a 39
former free-living eukaryotic alga into a eukaryotic host cell (32-33). During evolution, 40
the symbiont was subsequently reduced in terms of compartmentalization and genome 41
size to an organelle strictly dependent on the host cell (16, 32). Different types of 42
secondary plastids exist in a very broad range of algae and protists, which can be 43
distinguished based on their evolutionary origin (e.g. a red or green alga derived 44
symbiont), as well as on the amount of cellular reduction inside the host cell. Our 45
understanding of the evolution of organisms harboring a secondary plastid of red algal 46
origin has changed in the last few years. According to the chromalveolate hypothesis, 47
six major lineages were grouped together to be of monophyletic origin: cryptophytes, 48
haptophytes, heterokontophytes, peridinin-containing dinoflagellates, apicomplexans 49
and the non-plastid containing ciliates, as well as several smaller lineages related to 50
some chromalveolate members (15, 41). However, recent phylogenetic analyses have 51
given rise to extended theories about the evolution of the lineages with a red algal 52
endosymbiont, including serial endosymbiotic events with secondary, as well as tertiary, 53
endosymbioses (21-22, 26-27, 56, 61, 71, 75). 54
It has been shown that the lineages with an endosymbiont of red algal origin share 55
common plastid protein import mechanisms despite remarkable differences resulting 56
from specific features in plastid ultrastructure (10, 37, 65). Import into complex plastids 57
starts co-translationally at the endoplasmatic reticulum (ER) membrane where nascent 58

4
precursor proteins are synthesized into the ER lumen. This transport step requires a 59
canonical N-terminal signal peptide (SP). In heterokontophytes, cryptophytes and 60
haptophytes, the outermost plastid membrane, termed chloroplast ER (cER) membrane, 61
is continuous with the endomembrane system of the host cell; therefore, the Sec61 62
mediated import already represents transport across the first membrane of complex 63
plastids. In contrast, the plastids of apicomplexans and peridinin-containing 64
dinoflagellates are not connected to the endomembrane system. Thus, after import into 65
the ER lumen, proteins are likely to be transported to the plastid via vesicle transport 66
mechanisms directly from the ER or via the Golgi apparatus (47, 57). 67
After the preprotein has entered the cER lumen, the SP is thought to be cleaved off and 68
a transit peptide-like sequence (TPL) is exposed at the new N-terminus. The TPL is 69
required for further transport into the periplastidal compartment (PPC), which resembles 70
the naturally reduced cytoplasm of the endosymbiont, and further into the stroma of the 71
plastid. Such transit peptide-like sequences thereby fulfill an additional function in 72
contrast to transit peptides (TP) in primary plastids. Detailed characterization of the TPL 73
revealed a difference between stroma- and PPC-localized proteins. Stromal proteins 74
possess an aromatic (Phe, Tyr, Trp) or bulky (Leu) amino acid at the +1 position of their 75
TPL, in contrast to PPC proteins (4, 31, 34, 42). However, the observed AXA-FAP motif 76
at the transition between SP and TPL of stromal proteins is not as well conserved in 77
haptophytes, apicomplexans and dinoflagellates as it is the case for heterokontophytes 78
and cryptophytes (58). Furthermore, some membrane proteins of apicomplexan plastids 79
(apicoplasts) seem to carry intrinsic targeting signals instead of a bipartite targeting 80
signal (BTS) consisting of SP and TPL (1, 20). 81
For transport across the second outermost membrane, the periplastidal membrane 82
(PPM), a translocon model was proposed to consist of a recycled ER-associated 83

5
degradation (ERAD) machinery of symbiont origin (67). Support for this model came 84
from the detection of symbiont-specific ERAD components encoded on the 85
nucleomorph of the cryptophyte Guillardia theta, this being the remnant nucleus of the 86
former red algal endosymbiont in the cryptophytes PPC (23). The canonical ERAD 87
removes aberrant or misfolded luminal (ERAD-L) and membrane (ERAD-M and ERAD-88
C) ER proteins and tags them after retro-translocation in the cytosol with poly-ubiquitin 89
moieties for subsequent proteasomal degradation (7, 40, 66). However, in the symbiont-90
specific ERAD-like pathway (SELMA) the retro-translocation machinery of ERAD-L is 91
postulated to be maintained, and possesses the capacity to transport proteins from an 92
ER luminal compartment into a cytoplasmic compartment, the PPC. This process is 93
supposed to be uncoupled from degradation. SELMA is conserved in all secondary 94
evolved organisms with a red algal endosymbiont, for which genomic data are available 95
(26, 67-68). Proteins of the derlin family are still controversially discussed elements of 96
the ERAD-specific translocon. In the diatom Phaeodactylum tricornutum, two symbiont-97
localized derlins (PtsDer1-1/PtsDer1-2) are expressed which form hetero- as well as 98
homo-oligomers and show interaction with transit peptide-like sequences of PPC-99
localized proteins (38). These components are indeed involved in the transport of 100
proteins into the plastid as indicated by a conditional knock-down mutant of the 101
Toxoplasma gondii sDer1 protein which showed impairment in plastid protein import (2). 102
The translocation process is predicted to be dependent on ubiquitylation, further 103
supported by the presence of a set of ubiquitylation enzymes (39, 67). Additional factors 104
proposed to be involved in SELMA are a symbiont-specific Cdc48 AAA-ATPase with its 105
co-factor Ufd1 and adaptor proteins (55, 67). Following translocation, the precursor 106
proteins are likely to undergo de-ubiquitylation and are either passed on to the 107
translocon in the third outermost membrane or folded in the PPC (13, 39, 55). Although 108

6
a residual set of 20S proteasomal components was identified in the PPC of diatoms, 109
there is currently no link between SELMA and proteasomal degradation (55). 110
Having passed through the PPC, transport across the innermost plastid membranes 111
seems to be comparable to primary plastids with a translocon at the inner membrane of 112
chloroplasts (TIC) and a recently identified Omp85 protein which belongs to the family 113
of Toc75 proteins, the core components of the translocon at the outer membrane of 114
chloroplasts (TOC) (1, 10, 13, 73). 115
Here, we present an update on the SELMA translocation model in organisms with a red 116
algal endosymbiont with focus on five heterokontophytes and apicomplexan parasites. 117
In particular we mined the genomes of organisms that carry secondary plastids, 118
including recently published full genome sequences, for SELMA proteins. With this 119
collected data set one would expect to define the degree of factor conservation and 120
identify main components of the SELMA system which evolved to function in protein 121
transport at a plastid membrane. Our results are compared to the respective host ERAD 122
system as well as to red algal ERAD components, from which SELMA originated. 123
Additionally, four new PPC-localized proteins similar to factors involved in ERAD could 124
be identified in the diatom P. tricornutum. We also extended the set of core proteasomal 125
components in the PPC of heterokontophytes and discuss their putative function in 126
relation to SELMA. 127
128
Materials and Methods 129
Bioinformatic Analysis 130

7
Protein sequences of ERAD and SELMA as well as proteasomal components were 131
collected from published data or retrieved via blastp and tblastn searches. As queries, 132
sequences from the Saccharomyces cerevisiae ERAD system and the P. tricornutum 133
SELMA system were used to search the genomic databases for Phaeodactylum 134
tricornutum v2.0 (12), Thalassiosira pseudonana (5), Fragilariopsis cylindrus 135
(http://genome.jgi-psf.org/Fracy1/Fracy1.home.html), Aureococcus anophagefferens 136
(30), Emiliania huxleyi CCMP1516 main genome assembly v1.0 (http://genome.jgi-137
psf.org/Emihu1/Emihu1.home.html) and Guillardia theta CCMP2712 v1.0 138
(http://genome.jgi-psf.org/Guith1/Guith1.home.html). Sequences from Ectocarpus 139
siliculosus (19) and Babesia bovis were searched at the National Center for 140
Biotechnology Information (NCBI) server (http://www.ncbi.nlm.nih.gov/guide/). 141
Apicomplexan sequences were retrieved from the Plasmodium Genomics Resource 142
Version 9.0 (6) for Plasmodium, the Toxoplasma Genomics Resource v7.2 (29) for T. 143
gondi and Neospora caninum, TparvaDB Version 1.0 (74) and the National Center for 144
Biotechnology Information (NCBI) server (http://www.ncbi.nlm.nih.gov/guide/) for 145
Theileria parva and the Cryptosporidium Genomics Resource v4.6 (36) for 146
Cryptosporidium parvum. ERAD sequences for red algae were either retrieved from the 147
genome projects of Cyanidioschyzon merolae (53) and Galdieria sulphuraria (Michigan 148
State University Galdieria Database [http://genomics.msu.edu/galdieria/about.html]), or 149
by local Blast (blast-2.2.10-ia32-win32) using expressed sequence tags (EST) of 150
Porphyridium cruentum and partial genome data from Calliarthron tuberculosum 151
(http://dbdata.rutgers.edu/data/plantae/) generated by Chan and colleagues (17). 152
In general a minimal e-value of 1e-04 was set as threshold for the identification of 153
ERAD/SELMA components on the protein level. However, in cases of weak query 154
sequence significance, also matches with a lower e-value were inspected. Additionally, 155

8
criteria like domain structure and composition similarity (NCBI Conserved Domain 156
search) were applied for identification of relevant proteins (51). For proteasomal 157
components, all S. cerevisiae 20S protein sequences were used as queries to collect a 158
data set of putative proteasomal components which were then classified according to 159
the NCBI Conserved Domain Database (51) which differs from the S. cerevisiae 160
nomenclature (detailed information on different classifications can be found in (60)). 161
All gene models of the identified proteins were aligned to genomic and EST sequences, 162
if available. Thereby, missing N- and C-termini were identified by searching for putative 163
start and stop codons in frame, respectively. If possible, intron borders of the gene 164
models were checked to be in agreement with EST data. The protein sequences were 165
additionally examined for N-terminal targeting sequences to discriminate symbiont 166
proteins from host factors. PPC directed proteins are characterized by the presence of a 167
SP and a TPL. The SignalP 3.0 Server (24) was used for the prediction of a SP with a 168
cutoff of >0.5 by the HMM algorithm. Then, the sequences were analyzed with the 169
TargetP 1.1 Server (25) with default settings to define the SP as a secretory signal 170
sequence and exclude mitochondrial targeting. In general, the TPL of PPC (symbiont) 171
proteins cannot be predicted accurately with available tools. For this reason, besides 172
performing the prediction with the TargetP 1.1 Server (25) using signal peptide-173
truncated sequences in “plant” mode, the criteria defined in (55) were applied. In some 174
cases, a protein model was identified with high similarity to a known symbiont protein of 175
the diatom P. tricornutum or the apicomplexan parasite P. falciparum but without SP 176
prediction. This can be caused by an incorrect gene model prediction due to the lack of 177
EST data or the presence of several putative start codons. Therefore, these proteins 178
were assigned as symbiont but marked to lack a signal peptide prediction. 179

9
Analyses of transmembrane spanning regions were performed with TOPCONS (9), 180
domain and coiled-coil prediction was done using SMART (45). Protein sequence 181
alignments were performed with GENEDOC Software (version 2.6.002 182
[http://www.psc.edu/biomed/genedoc]). 183
Plasmid Construction and Transfection of P. tricornutum 184
The predicted PPC proteins were cloned and transfected into the diatom P. tricornutum. 185
The sequences of genes containing introns or without EST support were amplified from 186
cDNA, the rest from gDNA, cloned in front of egfp into P. tricornutum transfection 187
vectors. ptsubx, ptspng1, ptsubq were cloned into the nitrate-inducible pPha-NR vector 188
(GenBank: JN180663), ptsnpl4, ptsβ1, ptsα3, pthβ7 and pthrpn10 into the light-inducible 189
pPha-T1 vector (GenBank: AF219942). For further information about sequences of in 190
vivo-localized proteins as well as primer sequences see supplemental file 1. Biolistic 191
transfection into P. tricornutum cells was performed as described previously (67, 77). 192
Positive transformants were cultured under standard conditions as described before (3) 193
with 1.5 mM NH4+ in permanent cultivation. Protein expression under control of the 194
nitrate reductase promoter (pPha-NR vector) was induced by cultivation on 0.9 mM 195
NO3- for two days. 196
Fluorescence Microscopy 197
P. tricornutum transformants were fixed with 4 % paraformaldehyde/ 0,0075 % 198
glutaraldehyde in 1x PBS buffer and analyzed with a confocal laser scanning 199
microscope Leica TCS SP2 using a HCX PL APO 40×/1.25 − 0.75 Oil CS objective. 200
Fluorescence of eGFP and chlorophyll was excited with an Argon laser at 488 nm and 201
detected with two photomultiplier tubes at a bandwidth of 500–520 nm and 625–720 nm 202
for eGFP and chlorophyll fluorescence, respectively. 203

10
204
Results 205
1. Identification of ERAD and SELMA components in red algae and organisms 206
with a red algal endosymbiont 207
In order to identify new ERAD and SELMA components, all available genomic 208
sequences of red algae and organisms with a red algal endosymbiont were screened 209
via BLAST search with queries from the best studied ERAD system of Saccharomyces 210
cerevisiae (40, 66). The recently published genomes of heterokontophytes (the diatom 211
Fragilariopsis cylindrus, the brown alga Ectocarpus siliculosus, the harmful alga 212
Aureococcus anophagefferens), the nuclear genome of the cryptophyte Guillardia theta 213
and the apicomplexan Neospora caninum were included in these analyses. Because 214
SELMA was shown to be phylogenetically derived from the ERAD system of the red 215
algal endosymbiont (26), we also included sequences from the red algae 216
Cyanidioschyzon merolae, Porphyridium cruentum, Calliarthron tuberculosum and 217
Galdieria sulphuraria in our analyses (see Material and Methods for detailed description 218
on used genome data). In contrast to the other chromalveolate groups, dinoflagellate 219
plastids have only three surrounding membranes and very little is known about the 220
mechanisms that transport proteins across these membranes (10, 65). Due to the 221
paucity of genomic data for these organisms, we have not included peridinin-containing 222
dinoflagellates in this study. 223
We identified genes for conserved ERAD components in all investigated red algal 224
genomes (Table 1). However, due to incomplete data for Porphyridium cruentum (EST 225
data) and especially Calliarthron tuberculosum (partial genome data), only a subset of 226
ERAD factors could be identified. The collected data set for red algae implicates that the 227

11
progenitor from which the SELMA machinery originated was capable of ERAD-L via the 228
Hrd1 complex as well as ERAD-C via the Doa10 complex in the ER membrane. 229
Additionally, all proteins required for ubiquitylation and efficient proteasomal substrate 230
delivery after ERAD retro-translocation are present in red algae. 231
In organisms with a red algal endosymbiont, the SELMA system exists in parallel with 232
the host ERAD machinery. The discrimination between proteins of both systems is 233
based on the targeting signal of the PPC localized SELMA proteins in contrast to the 234
mostly cytosolic ERAD components (see Material and Methods). Identification of a 235
SELMA protein is more reliable if a respective host protein with the same putative 236
function can be found. Therefore, a detailed analysis of the host ERAD system of the 237
investigated organisms was included and almost all ERAD proteins known from S. 238
cerevisiae could be identified in the genomes (Table 1, for detailed information see 239
supplemental file 2, Table S1). All organisms encode for the ER membrane proteins 240
Sec61α, Hrd1, the derlin proteins and, with the exception of apicomplexans, also for 241
Doa10. In addition, a cytosolic ubiquitylation machinery, the Cdc48 complex with its co-242
factors (Npl4 and Ufd1) and all proteasomal substrate delivery factors (Rad23, Dsk2, 243
Png1) were identified. 244
Our inspection of the SELMA system in secondary evolved algae and apicomplexan 245
parasites showed a high degree of conserved components for this putative protein 246
translocation machinery (Table 1). However, failures in the identification of certain 247
proteins can have different reasons. If a protein is present in most of the organisms of 248
one group but lacking in one specific organism, this is likely caused by incomplete 249
genome sequencing and assembly, or by incorrect protein model prediction (e. g. for 250
Aureococcus anophagefferens). In contrast, a protein not identified in a whole group of 251
organisms may have been lost completely during evolution. Haptophytes and 252

12
cryptophytes are represented only by one organism, hindering a final conclusion about 253
the presence or absence of specific proteins but allowing considerations of whole 254
protein complexes. The analysis of the newly available genome sequence of the 255
cryptophyte G. theta shows that the partially nucleomorph-encoded SELMA system is 256
supplemented with nucleus encoded factors (Table 1). 257
Interestingly, from the three ER membrane protein classes, Sec61α, derlin proteins and 258
the ubiquitin ligase Hrd1, which are discussed as putative ERAD channel proteins, only 259
the derlin proteins are found in the complex plastids of these organisms as membrane 260
proteins with several transmembrane domains, with the exception of a nucleomorph-261
encoded Hrd1 in cryptophytes (see below). In respect to derlins, two symbiontic 262
representatives are present in heterokontophytes, haptophytes and cryptophytes, as is 263
the case for yeast (ScDer1p and ScDfm1p). Ubiquitylation requires a cascade of three 264
enzymes starting with a ubiquitin-activating enzyme (Uba1) which is present in all 265
organisms. At least one symbiont ubiquitin conjugating enzyme (sUbc) can also be 266
found, but not all putative PPC-targeted sUbc proteins can be assigned to the same S. 267
cerevisiae Ubc protein. While heterokontophytes share a sUbc similar to ScUbc6p and 268
at least one other sUbc protein, apicomplexans seem to encode only for one sUbc 269
protein with the highest similarity to ScUbc4p. The ubiquitin ligase sHrd1 of 270
heterokontophytes differs in protein structure from the symbiont ubiquitin ligase of 271
cryptophytes. Several transmembrane domains are predicted for the GtsHrd1 protein. 272
Therefore, it more resembles the yeast ScHrd1p structure than the heterokontophyte E3 273
ligase which contains only one predicted transmembrane domain. 274
The symbiont Cdc48-complex together with sUfd1 can be found in all organisms 275
investigated and we were now able to identify a sNpl4 protein in the diatom P. 276
tricornutum which is conserved among heterokontophytes. The same is the case for 277

13
three other newly identified putative symbiont proteins, sUBX, sUbq and sPng1. These 278
share similarity to ERAD factors and are present in addition to the host version in the 279
diatom P. tricornutum and other heterokontophytes. None of these proteins is 280
conserved in apicomplexans (Table 1). 281
2. Newly identified putative SELMA components: the sCdc48 co-factor sNpl4, a 282
UBX domain-containing protein, a symbiont ubiquilin-like protein and a peptide 283
N-glycanase 284
Among the newly identified putative SELMA components are two Cdc48 binding 285
proteins, the UBX domain-containing protein sUBX (symbiont UBX) and the Ufd1 co-286
factor sNpl4, as well as proteins with sequence similarity to the de-glycosylation enzyme 287
ScPng1p (sPng1) and the poly-ubiquitin binding protein ScDsk2p (sUbq). Not all four 288
proteins are predicted to have a TPL in the diatom P. tricornutum. In such a case, a 289
signal peptide on usually cytosolic proteins is indicative for a PPC localization, but 290
remained to be verified in localization experiments. Therefore, PtsNpl4, PtsUBX, 291
PtsUbq and PtsPng1 were expressed as eGFP fusions in P. tricornutum and their 292
localization was examined in vivo. All constructs showed the typical fluorescence 293
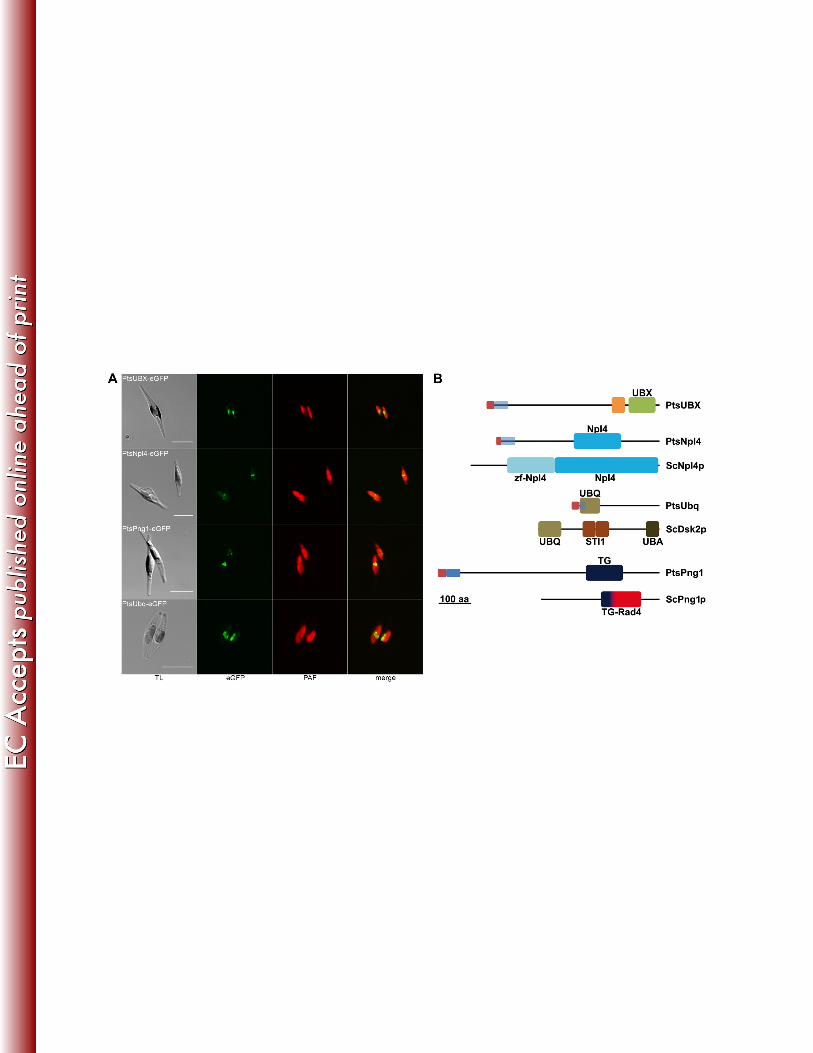
pattern of PPC-localized proteins in the middle of the two plastid lobes (Fig. 1A). 294
A comparison of the protein sequences to well known ERAD components of S. 295
cerevisiae and their domain composition points to their putative function (Fig. 1B). The 296
identified symbiont UBX domain-containing protein, PtsUBX, shares sequence similarity 297
to other proteins only in its UBX domain which has been shown to be a general Cdc48 298
binding module (63). Preceding the UBX domain, the protein harbors a coiled-coil 299
region for homo- or heterotypic protein interaction (52). The second identified Cdc48 300
binding protein, PtsNpl4, now completes the sCdc48 complex, together with the 301

14
previously described co-factor sUfd1 (67). In comparison to its yeast ERAD counterpart 302
the protein lacks the N-terminal Npl4 zinc finger domain. The symbiont ubiquilin-like 303
protein PtsUbq shares the N-terminal UBQ domain known from mammalian ubiquilins 304
and ScDsk2p (28, 69) but lacks the C-terminal UBA domain for poly-ubiquitin binding as 305
well as the internal STI1 domains. PtsPng1 as a symbiont de-glycosylation enzyme was 306
initially annotated as the host protein (67) despite the presence of a weakly predicted 307
signal peptide which can be explained by a missing second copy of this protein at that 308
time. In this current study an additional Png1 protein was identified in its place, lacking 309
an N-terminal targeting sequence and leading to reevaluation of the former prediction. 310
PtsPng1 has a transglutaminase/protease-like domain with the conserved catalytic 311
residues of cytoplasmic PNGase (data not shown) (70). 312
3. The symbiont ubiquitin in diatoms lacks the conserved lysine residues Lys48 313
and Lys63 314
The SELMA model proposes ubiquitylation of the precursor proteins during transport 315
and subsequent removal of the ubiquitin moiety via a PPC-specific de-ubiquitinating 316
enzyme (38, 67). The PPC-targeted ubiquitin of P. tricornutum (PtsUbi) was shown to 317
lack the specific lysine residue (Lys48) as the most prominent linker for poly-318
ubiquitylation involved in degradation (67). With the identification of symbiont ubiquitins 319
from T. pseudonana and F. cylindrus, this feature becomes even more apparent, as the 320
Lys48 is also absent in these diatom sequences (Fig. 2). The TpsUbi protein model (ID: 321
1539) can hardly be recognized and has to be modified according to the available EST 322
sequences in order to obtain the full sequence (see supplemental file 1). In the genome 323
of the cryptophyte Guillardia theta, a di-ubiquitin protein sequence with a signal peptide 324
prediction could be identified with the less conserved first ubiquitin domain sharing the 325
diatom lysine mutation. In contrast, sUbi sequences from the haptophyte E. huxleyi and 326

15
the apicomplexans B. bovis and T. parva still contain lysine 48. The previously identified 327
symbiont ubiquitin of P. falciparum shows only weak conservation with the symbiont 328
ubiquitins from diatoms (68). Unfortunately, it was not possible to detect a symbiont 329
ubiquitin in the other newly investigated organisms. 330
Interestingly, the position Lys63, usually used for poly-ubiquitin linkages related to 331
modifications of protein function, is also no longer present in all symbiont ubiquitins 332
except the second domain of GtsUbi. 333
4. Identification and localization of proteasomal components in the PPC of 334
heterokontophytes and cryptophytes 335
We previously reported on the presence of relict 20S proteasomal components in the P. 336
tricornutum PPC (55). Here, we expand the model of a symbiont core proteasome by 337
extensive in silico analyses. 338
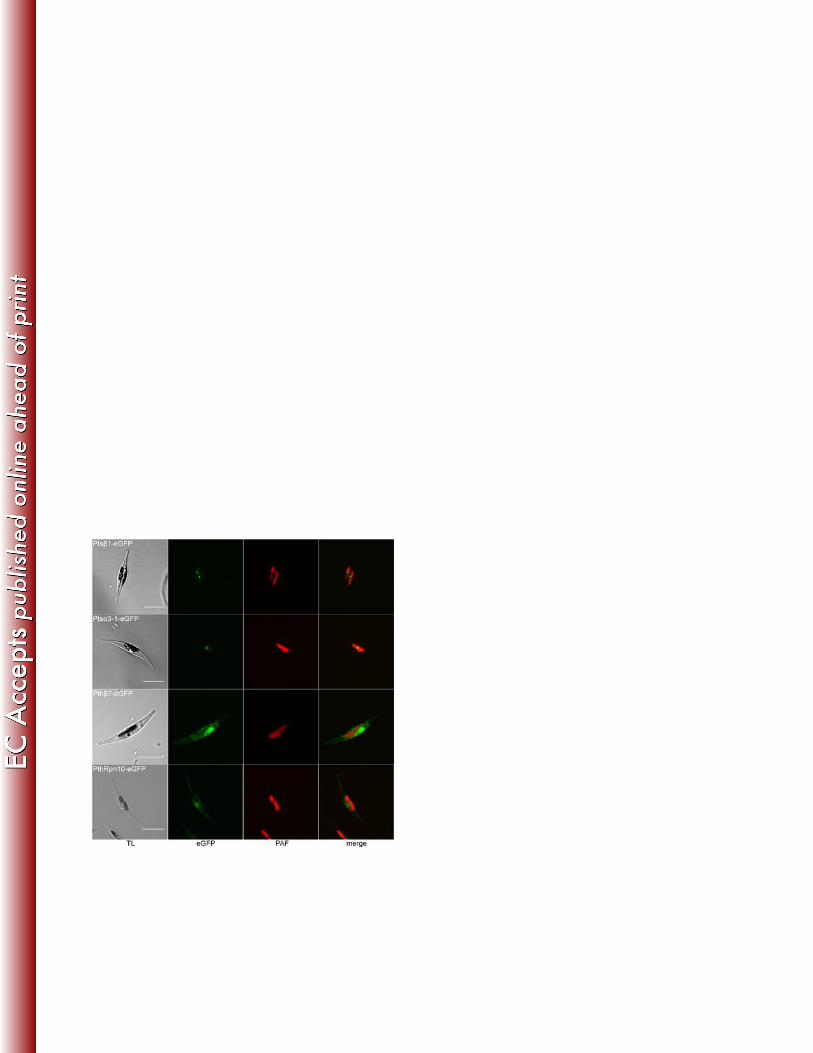
The cryptophyte Guillardia theta, which is still able to synthesize proteins in the PPC, 339
encodes for an almost complete set of PPC-localized proteasomal degradation 340
components on its nucleomorph genome (23). This is in contrast to other organisms 341
with a red algal endosymbiont and amongst them, only in heterokontophytes could 342
residual 20S subunits be identified. Importantly, we did not detect a symbiont 19S 343
regulatory particle in the PPC of heterokontophytes although it is present in 344
cryptophytes. We could not identify a complete set of 20S subunits, including 7 alpha 345
and 7 beta subunits, for any of the organisms studied (Table 2). Instead, the putative 346
20S core particle seems to vary in subunit composition, with the exception of conserved 347
sα2, sα3 and sβ6. The putative catalytically active subunits β6 and β7 could be 348
identified in almost all heterokontophytes in a PPC-directed version; a second β5 gene 349
is only detectable as a gene fusion with a 5'-3' exonuclease, also lacking a signal 350

16
peptide. In A. anophagefferens and E. siliculosus in addition to a symbiont sβ5 with 351
signal peptide prediction, several putative symbiont subunits exist but an exact defining 352
of the gene model is difficult. Therefore, a classification into host or symbiont protein 353
cannot yet conclusively be determined. 354
In addition to the already reported symbiont 20S proteasomal subunits Ptsβ2, Ptsβ6, 355
Ptsβ7, Ptsα7-1 and Ptsα7-2 (55), we successfully localized two additional subunits in 356
the PPC of P. tricornutum. Both Ptsα3-1 and Ptsβ1 showed the typical PPC 357
fluorescence pattern (see Fig. 3). As a comparison, two subunits of the host 358
proteasome were also localized, Pthβ7 and PthRpn10, which resulted in a different 359
fluorescence pattern outside of the plastid. 360
361
Discussion 362
For protein transport across the periplastidal membrane of complex plastids of red algal 363
origin, an ERAD derived mechanism (SELMA) was proposed as the protein 364
translocation machinery (67). The SELMA model, originally based on our findings in 365
cryptophytes, was shown to be conserved in organisms with a red algal endosymbiont 366
(2, 26, 67-68) and SELMA components can be identified in all available genomes in 367
addition to the host ERAD machinery (Table 1). However, the exact mechanism of this 368
transport step, the pore-forming proteins and the minimal required components remain 369
an open question. The proposed SELMA components show often a minimized structure, 370
as domains, known from ERAD proteins of other organisms, are missing. Thus, the 371
SELMA complex should indicate a minimized version of the retro-translocation activity 372
of ERAD in general. Different extents of ERAD to SELMA reduction can be found in the 373
investigated organisms according to the amount of reduction of the former 374

17
endosymbiont. The cryptophyte G. theta represents the most extended set of SELMA 375
and proteasomal components resulting most probably from its transcriptionally and 376
translationally active nucleomorph in the PPC. Apicomplexans instead have the 377
smallest set of identified SELMA components and seem to lack a symbiont proteasome. 378
So far, all investigated organisms share the following as SELMA components in the 379
PPC: derlins as membrane and putative channel proteins, a ubiquitylation machinery 380
and a Cdc48 complex. It remains to be determined if the recently identified, conserved 381
PPC protein PPP1 provides a new crucial function for protein transport (64). All other 382
identified proteins with functions related to SELMA or the proteasome in 383
heterokontophytes might represent lineage specific adaptations (Fig. 4). 384
Of all ER membrane proteins that are candidates for a translocation channel in ERAD 385
(66), only the derlin proteins could be identified as SELMA components. Apart from that, 386
the Sec61 channel and the ubiquitin ligase Hrd1 are in discussion as potentially being 387
capable of fulfilling this function. On the one hand, we could not identify additional 388
symbiont Sec61 subunits, on the other hand, the diatom symbiont E3 ubiquitin ligase is 389
predicted to have only one transmembrane domain, and is therefore most likely not 390
capable of homotypic channel formation. However, the derlin proteins and the ubiquitin 391
ligase might form a membrane complex which connects translocation to ubiquitylation. 392
The presence of the ubiquitylation enzymes sUba, sUbc and ubiquitin itself in almost all 393
investigated organisms including apicomplexans suggests the presence of a ubiquitin-394
dependent mechanism in the PPC. Once the preprotein is ubiquitylated in the PPC, it 395
can be recognized by the Cdc48 complex (76). We could identify at least one symbiont-396
specific sCdc48 protein in all organisms investigated. The Cdc48-ATPase has been 397
shown to be a central component of ERAD, acting specifically in concert with its co-398
factors Ufd1 and Npl4 (54, 76). Although Cdc48 is known to have various cellular 399

18
functions, the identification and localization of a symbiont Npl4 protein of P. tricornutum 400
presented here now define the sCdc48-sUfd1-sNpl4 complex as a SELMA component. 401
However, other functions unrelated to protein transport together with so far unidentified 402
co-factors cannot be excluded. The new PPC-localized protein PtsUBX, most likely a 403
sCdc48 binding protein due to its UBX domain, might also be involved in SELMA 404
translocation akin to the case for UBX proteins in ERAD. These proteins can direct the 405
Cdc48-ATPase to a specific protein complex, in the context of ERAD to ubiquitin ligases 406
at the ER membrane (50, 62-63). 407
After translocation is completed, ERAD substrates are recognized by a set of cytosolic 408
proteins and processed for degradation by the proteasome (59, 76). The presence of a 409
relict symbiont proteasome in heterokontophytes (Table 2) raises the question of a 410
functional link between the ERAD derived SELMA machinery and degradation in the 411
PPC of these organisms. Several features of both machineries in the PPC argue 412
against such a connection. On the one hand, the PPC of apicomplexan parasites and 413
haptophytes lacks symbiont 20S subunits and therefore harbors a SELMA system 414
which seems to be completely independent of a proteasomal function. On the other 415
hand, proteasomal substrates are not only delivered by the ERAD system but can also 416
be degraded independently of ubiquitylation (8). The 20S core particle was shown to 417
have basal proteolytic activity towards unstructured or oxidized proteins (60). It is also 418
implicated in maturation and specific cleavage of various proteins, which gain access to 419
the proteolytic chamber through an interaction with N-termini of the α-subunits (8, 49). 420
In the PPC of heterokontophytes, we identified proteasomal subunits of the 20S core 421
particle including proteolytic active subunits. It was not possible to detect a complete set 422
of 20S subunits in any of the heterokontophyte species. Either the remaining α- and β-423
subunits in the genomes are too divergent to be recognized or the putative reduced 20S 424

19
particle in the PPC can vary in subunit composition, replacing some subunits by other 425
ones. 426
In addition, the recognition and unfolding of ubiquitylated proteasomal substrates is 427
mediated by the 19S regulatory particle of the proteasome which was not identified in a 428
PPC targeted version in heterokontophytes. However, the two newly identified PPC 429
proteins sUbq and sPng1 are counterparts to ScDsk2p and ScPng1p which are known 430
from ERAD to function between retro-translocation and degradation (43). The symbiont 431
ubiquilin-like protein PtsUbq lacks the C-terminal ubiquitin-associated (UBA) domain for 432
poly-ubiquitin binding present in ScDsk2p and mammalian ubiquilins for recognition of 433
proteasomal substrates (28, 48). In addition, the ubiquitin-like domain (UBQ) at the N-434
terminus of ScDsk2p was shown to bind proteasomal components of the 19S regulatory 435
particle as well as to ScUfd2p (35, 69), both not present in a symbiont version in the 436
PPC. Most likely, both proteins had to adapt to new functions. PtsPng1, the PPC-437
localized peptide N-glycanase, might either be involved in the maturation of PPC-438
localized proteins or may be required for efficient removal of glycan moieties of plastid 439
precursor proteins added in the ER lumen before transport across the third outermost 440
plastid membrane. In contrast to heterokontophytes, haptophytes and cryptophytes, 441
apicomplexans encode neither a host nor a symbiont Png1 protein. This is likely due to 442
a reduction of N-glycosylation capacities in these organisms, especially for apicoplast 443
proteins (14). 444
Another important feature of SELMA is the symbiont ubiquitin which shows alterations 445
at specific lysine residues. In heterokontophytes, the PPC-localized ubiquitin (sUbi) 446
does not possess the conserved lysine residues Lys48 and Lys63, while haptophyte 447
and apicomplexan (except P. falciparum) sUbi sequences still contain Lys48 but show 448
mutations at Lys63. Lys48 was shown to represent the most prominent position for poly-449

20
ubiquitylation leading to proteasomal degradation (72). Although recent work suggests a 450
more complex interplay between different ubiquitin linkages on various lysine residues 451
also in degradation (44), loss of Lys48 in the symbiont ubiquitins of heterokontophytes 452
might be an evolutionary adaptation required for the establishment of the symbiont 453
ERAD as a preprotein translocation system. This is supported by the finding that only 454
organisms with a symbiont proteasome (Table 2) show this ubiquitin Lys48 modification. 455
An exception is the cryptophyte G. theta with two ubiquitin domains in the predicted 456
GtsUbi sequence, one overall conserved and another having Lys mutations at both 457
positions. One might speculate about a separation of SELMA and proteasomal 458
degradation in the cryptophytes PPC based on different ubiquitins, which might be 459
caused by the different morphology, as cryptophytes – in contrast to all other organisms 460
with a secondary red algal symbiont- still synthesize proteins in the PPC. Ubiquitin 461
Lys63 is implicated in ubiquitylation processes related to functional modifications of 462
target proteins (72). Remarkably, loss of Lys63 in all organisms with a secondary plastid 463
of red algal origin leads to reduced ubiquitylation possibilities in the PPC in contrast to 464
the manifold mechanisms regulated by ubiquitylation in the host cytosol. Thus, it 465
remains to be determined if the symbiont ubiquitins can be used for both mono- and 466
polyubiquitylation on the remaining lysine residues. 467
The SELMA translocation machinery (Fig. 4) provides an interesting view into 468
evolutionary rearrangements and modifications of already existing mechanisms. During 469
the establishment of a red alga as an organelle, the symbiont ER-associated 470
degradation machinery was split into a translocation complex on the one hand and a 471
presumed degradation machinery on the other one. The former now represents the 472
second step of protein import into complex plastids across the periplastidal membrane, 473
whereas the latter one might be required for protein homeostasis in the PPC of only 474

21
certain groups of organisms with a red algal endosymbiont. Such modularization of the 475
well conserved ERAD translocation not only gave rise to SELMA but also was shown to 476
be the principle mechanism of the peroxisomal importomer, again a ubiquitin-dependent 477
translocation independent of proteasomal degradation (11). 478
479
Acknowledgements 480
We are supported by the Deutsche Forschungsgemeinschaft (Collaborative Research 481
Centre 593 for S.S., I.W., S.Z. and U.-G.M.; SFB TR1 for J.M.P.). D.M. is a fellow of the 482
International Max Planck Research School for Environmental, Cellular and Molecular 483
Microbiology (IMPRS-MIC). 484
485
References 486
1. Agrawal S, Striepen B. 2010. More membranes, more proteins: complex protein 487
import mechanisms into secondary plastids. Protist 161:672-687. 488
2. Agrawal S, van Dooren GG, Beatty WL, Striepen B. 2009. Genetic evidence 489
that an endosymbiont-derived endoplasmic reticulum-associated protein 490
degradation (ERAD) system functions in import of apicoplast proteins. J. Biol. 491
Chem. 284:33683-33691. 492
3. Apt KE, Grossman A, Kroth-Pancic P. 1996. Stable nuclear transformation of 493
the diatom Phaeodactylum tricornutum. Mol. Gen. Genet. 252:572-579. 494
4. Apt KE, Zaslavkaia L, Lippmeier JC, Lang M, Kilian O, Wetherbee R, 495
Grossman AR, Kroth PG. 2002. In vivo characterization of diatom multipartite 496
plastid targeting signals. J. Cell Sci. 115:4061-4069. 497
5. Armbrust EV, Berges JA, Bowler C, Green BR. 2004. The genome of the 498
diatom Thalassiosira pseudonana: ecology, evolution, and metabolism. Science 499
306:79-86. 500
6. Aurrecoechea C, Brestelli J, Brunk BP, Dommer J, Fischer S, Gajria B, Gao 501
X, Gingle A, Grant G, Harb OS, Heiges M, Innamorato F, Iodice J, Kissinger 502
JC, Kraemer E, Li W, Miller JA, Nayak V, Pennington C, Pinney DF, Roos 503
DS, Ross C, Stoeckert CJ, Treatman C, Wang H. 2009. PlasmoDB: a 504

22
functional genomic database for malaria parasites. Nucleic Acids Res. 37:D539-505
D543. 506
7. Bagola K, Mehnert M, Jarosch E, Sommer T. 2011. Protein dislocation from 507
the ER. Biochim. Biophys. Acta 1808:925-936. 508
8. Baugh JM, Viktorova EG, Pilipenko EV. 2009. Proteasomes can degrade a 509
significant proportion of cellular proteins independent of ubiquitination. J. Mol. 510
Biol. 386:814-827. 511
9. Bernsel A, Viklund H, Hennerdal A, Elofsson A. 2009. TOPCONS: consensus 512
prediction of membrane protein topology. Nucleic Acids Res. 37:W465-468. 513
10. Bolte K, Bullmann L, Hempel F, Bozarth A, Zauner S, Maier U-G. 2009. 514
Protein targeting into secondary plastids. J. Eukaryot. Microbiol. 56:9-15. 515
11. Bolte K, Gruenheit N, Felsner G, Sommer MS, Maier UG, Hempel F. 2011. 516
Making new out of old: recycling and modification of an ancient protein 517
translocation system during eukaryotic evolution. Mechanistic comparison and 518
phylogenetic analysis of ERAD, SELMA and the peroxisomal importomer. 519
Bioessays 33:368-376. 520
12. Bowler C, Allen AE, Badger JH, Grimwood J, Jabbari K, Kuo A, Maheswari 521
U, Martens C, Maumus F, Otillar RP, Rayko E, Salamov A, Vandepoele K, 522
Beszteri B, Gruber A, Heijde M, Katinka M, Mock T, Valentin K, Verret F, 523
Berges JA, Brownlee C, Cadoret J-P, Chiovitti A, Choi CJ, Coesel S, De 524
Martino A, Detter JC, Durkin C, Falciatore A, Fournet J, Haruta M, Huysman 525
MJJ, Jenkins BD, Jiroutova K, Jorgensen RE, Joubert Y, Kaplan A, Kroger 526
N, Kroth PG, La Roche J, Lindquist E, Lommer M, Martin-Jezequel V, Lopez 527
PJ, Lucas S, Mangogna M, McGinnis K, Medlin LK, Montsant A, Secq M-PO-528
L, Napoli C, Obornik M, Parker MS, Petit J-L, Porcel BM, Poulsen N, 529
Robison M, Rychlewski L, Rynearson TA, Schmutz J, Shapiro H, Siaut M, 530
Stanley M, Sussman MR, Taylor AR, Vardi A, von Dassow P, Vyverman W, 531
Willis A, Wyrwicz LS, Rokhsar DS, Weissenbach J, Armbrust EV, Green BR, 532
Van de Peer Y, Grigoriev IV. 2008. The Phaeodactylum genome reveals the 533
evolutionary history of diatom genomes. Nature 456:239-244. 534
13. Bullmann L, Haarmann R, Mirus O, Bredemeier R, Hempel F, Maier UG, 535
Schleiff E. 2010. Filling the gap, evolutionarily conserved Omp85 in plastids of 536
chromalveolates. J. Biol. Chem. 285:6848-6856. 537
14. Bushkin GG, Ratner DM, Cui J, Banerjee S, Duraisingh MT, Jennings CV, 538
Dvorin JD, Gubbels M-J, Robertson SD, Steffen M, O'Keefe BR, Robbins 539 PW, Samuelson J. 2010. Suggestive evidence for Darwinian selection against 540
asparagine-linked glycans of Plasmodium falciparum and Toxoplasma gondii. 541
Eukaryot. Cell 9:228-241. 542
15. Cavalier-Smith T. 1999. Principles of protein and lipid targeting in secondary 543
symbiogenesis: euglenoid, dinoflagellate, and sporozoan plastid origins and the 544
eukaryote family tree. J. Eukaryot. Microbiol. 46:347-366. 545
16. Cavalier-Smith T. 2000. Membrane heredity and early chloroplast evolution. 546
Trends Plant Sci. 5:174-182. 547

23
17. Chan CX, Yang EC, Banerjee T, Yoon HS, Martone PT, Estevez JM, 548
Bhattacharya D. 2011. Red and green algal monophyly and extensive gene 549
sharing found in a rich repertoire of red algal genes. Curr. Biol. 21:328-333. 550
18. Chung D-WD, Ponts N, Prudhomme J, Rodrigues EM, Le Roch KG. 2012. 551
Characterization of the ubiquitylating components of the human malaria 552
parasite’s protein degradation pathway. PLoS ONE 7:e43477. 553
doi:10.1371/journal.pone.0043477. 554
19. Cock JM, Sterck L, Rouzé P, Scornet D, Allen AE, Amoutzias G, Anthouard 555
V, Artiguenave F, Aury J-M, Badger JH, Beszteri B, Billiau K, Bonnet E, 556
Bothwell JH, Bowler C, Boyen C, Brownlee C, Carrano CJ, Charrier B, Cho 557
GY, Coelho SM, Collén J, Corre E, Da Silva C, Delage L, Delaroque N, 558
Dittami SM, Doulbeau S, Elias M, Farnham G, Gachon CMM, Gschloessl B, 559
Heesch S, Jabbari K, Jubin C, Kawai H, Kimura K, Kloareg B, Küpper FC, 560
Lang D, Le Bail A, Leblanc C, Lerouge P, Lohr M, Lopez PJ, Martens C, 561
Maumus F, Michel G, Miranda-Saavedra D, Morales J, Moreau H, Motomura 562
T, Nagasato C, Napoli CA, Nelson DR, Nyvall-Collén P, Peters AF, Pommier 563
C, Potin P, Poulain J, Quesneville H, Read B, Rensing SA, Ritter A, 564
Rousvoal S, Samanta M, Samson G, Schroeder DC, Ségurens B, 565
Strittmatter M, Tonon T, Tregear JW, Valentin K, von Dassow P, Yamagishi 566
T, Van de Peer Y, Wincker P. 2010. The Ectocarpus genome and the 567
independent evolution of multicellularity in brown algae. Nature 465:617-621. 568
20. Derocher AE, Karnataki A, Vaney P, Parsons M. 2012. Apicoplast targeting of 569
a Toxoplasma gondii transmembrane protein requires a cytosolic tyrosine-based 570
motif. Traffic 13:694-704. 571
21. Deschamps P, Moreira D. 2012. Re-evaluating the Green Contribution to 572
Diatom Genomes. Genome Biol. Evol. 4:683-688. doi:10.1093/gbe/evs053. 573
22. Dorrell RG, Smith AG. 2011. Do red and green make brown?: perspectives on 574
plastid acquisitions within chromalveolates. Eukaryot. Cell 10:856-868. 575
23. Douglas S, Zauner S, Fraunholz M, Beaton M, Penny S, Deng LT, Wu X, 576
Reith M, Cavalier-Smith T, Maier UG. 2001. The highly reduced genome of an 577
enslaved algal nucleus. Nature 410:1091-1096. 578
24. Dyrløv Bendtsen J, Nielsen H, von Heijne G, Brunak S. 2004. Improved 579
prediction of signal peptides: SignalP 3.0. J. Mol. Biol. 340:783-795. 580
25. Emanuelsson O, Nielsen H, Brunak S, von Heijne G. 2000. Predicting 581
subcellular localization of proteins based on their N-terminal amino acid 582
sequence. J. Mol. Biol. 300:1005-1016. 583
26. Felsner G, Sommer MS, Gruenheit N, Hempel F, Moog D, Zauner S, Martin 584
W, Maier UG. 2011. ERAD components in organisms with complex red plastids 585
suggest recruitment of a preexisting protein transport pathway for the periplastid 586
membrane. Genome Biol. Evol. 3:140-150. doi:10.1093/gbe/evq074. 587
27. Frommolt R, Werner S, Paulsen H, Goss R, Wilhelm C, Zauner S, Maier UG, 588
Grossman AR, Bhattacharya D, Lohr M. 2008. Ancient recruitment by 589

24
chromists of green algal genes encoding enzymes for carotenoid biosynthesis. 590
Mol. Biol. Evol. 25:2653-2667. 591
28. Funakoshi M, Sasaki T, Nishimoto T, Kobayashi H. 2002. Budding yeast 592
Dsk2p is a polyubiquitin-binding protein that can interact with the proteasome. 593
Proc. Natl. Acad. Sci. U. S. A. 99:745-750. 594
29. Gajria B, Bahl A, Brestelli J, Dommer J, Fischer S, Gao X, Heiges M, Iodice 595
J, Kissinger JC, Mackey AJ, Pinney DF, Roos DS, Stoeckert CJ, Wang H, 596
Brunk BP. 2008. ToxoDB: an integrated Toxoplasma gondii database resource. 597
Nucleic Acids Res. 36:D553-D556. 598
30. Gobler CJ, Berry DL, Dyhrman ST, Wilhelm SW, Salamov A, Lobanov AV, 599
Zhang Y, Collier JL, Wurch LL, Kustka AB, Dill BD, Shah M, VerBerkmoes 600
NC, Kuo A, Terry A, Pangilinan J, Lindquist EA, Lucas S, Paulsen IT, 601
Hattenrath-Lehmann TK, Talmage SC, Walker EA, Koch F, Burson AM, 602
Marcoval MA, Tang Y-Z, Lecleir GR, Coyne KJ, Berg GM, Bertrand EM, Saito 603
MA, Gladyshev VN, Grigoriev IV. 2011. Niche of harmful alga Aureococcus 604
anophagefferens revealed through ecogenomics. Proc. Natl. Acad. Sci. U. S. A. 605
108:4352-4357. 606
31. Gould SB, Sommer MS, Kroth PG, Gile GH, Keeling PJ, Maier UG. 2006. 607
Nucleus-to-nucleus gene transfer and protein retargeting into a remnant 608
cytoplasm of cryptophytes and diatoms. Mol. Biol. Evol. 23:2413-2422. 609
32. Gould SB, Waller RF, McFadden GI. 2008. Plastid evolution. Annu. Rev. Plant 610
Biol. 59:491-517. 611
33. Green BR. 2011. After the primary endosymbiosis: an update on the 612
chromalveolate hypothesis and the origins of algae with Chl c. Photosynth. Res. 613
107:103-115. 614
34. Gruber A, Vugrinec S, Hempel F, Gould SB, Maier U-G, Kroth PG. 2007. 615
Protein targeting into complex diatom plastids: functional characterisation of a 616
specific targeting motif. Plant Mol. Biol. 64:519-530. 617
35. Hänzelmann P, Stingele J, Hofmann K, Schindelin H, Raasi S. 2010. The 618
yeast E4 ubiquitin ligase Ufd2 interacts with the ubiquitin-like domains of Rad23 619
and Dsk2 via a novel and distinct ubiquitin-like binding domain. J. Biol. Chem. 620
285:20390-20398. 621
36. Heiges M, Wang H, Robinson E, Aurrecoechea C, Gao X, Kaluskar N, 622
Rhodes P, Wang S, He C-Z, Su Y, Miller J, Kraemer E, Kissinger JC. 2005. 623
CryptoDB: a Cryptosporidium bioinformatics resource update. Nucleic Acids Res. 624
34:D419-D422. 625
37. Hempel F, Bozarth A, Sommer MS, Zauner S, Przyborski JM, Maier U-G. 626
2007. Transport of nuclear-encoded proteins into secondarily evolved plastids. 627
Biol. Chem. 388:899-906. 628
38. Hempel F, Bullmann L, Lau J, Zauner S, Maier UG. 2009. ERAD-derived 629
preprotein transport across the second outermost plastid membrane of diatoms. 630
Mol. Biol. Evol. 26:1781-1790. 631

25
39. Hempel F, Felsner G, Maier UG. 2010. New mechanistic insights into pre-632
protein transport across the second outermost plastid membrane of diatoms. Mol. 633
Microbiol. 76:793-801. 634
40. Hoseki J, Ushioda R, Nagata K. 2010. Mechanism and components of 635
endoplasmic reticulum-associated degradation. J. Biochem. 147:19-25. 636
41. Keeling PJ. 2009. Chromalveolates and the evolution of plastids by secondary 637
endosymbiosis. J. Eukaryot. Microbiol. 56:1-8. 638
42. Kilian O, Kroth PG. 2005. Identification and characterization of a new conserved 639
motif within the presequence of proteins targeted into complex diatom plastids. 640
Plant J. 41:175-183. 641
43. Kim I, Ahn J, Liu C, Tanabe K, Apodaca J, Suzuki T, Rao H. 2006. The Png1–642
Rad23 complex regulates glycoprotein turnover. J. Cell Biol. 172:211-219. 643
44. Kravtsova-Ivantsiv Y, Ciechanover A. 2012. Non-canonical ubiquitin-based 644
signals for proteasomal degradation. J. Cell Sci. 125:539-548. 645
45. Letunic I, Doerks T, Bork P. 2012. SMART 7: recent updates to the protein 646
domain annotation resource. Nucleic Acids Res. 40:D302-D305. 647
46. Li H-m, Chiu C-C. 2010. Protein transport into chloroplasts. Annu. Rev. Plant 648
Biol. 61:157-180. 649
47. Lim L, Kalanon M, McFadden GI. 2009. New proteins in the apicoplast 650
membranes: time to rethink apicoplast protein targeting. Trends Parasitol. 651
25:197-200. 652
48. Lim PJ, Danner R, Liang J, Doong H, Harman C, Srinivasan D, Rothenberg 653
C, Wang H, Ye Y, Fang S, Monteiro MJ. 2009. Ubiquilin and p97/VCP bind 654
erasin, forming a complex involved in ERAD. J. Cell Biol. 187:201-217. 655
49. Lin Y-C, Chen H-M, Chou I-M, Chen A-N, Chen C-P, Young G-H, Lin C-T, 656
Cheng C-H, Chang S-C, Juang R-H. 2012. Plastidial starch phosphorylase in 657
sweet potato roots is proteolytically modified by protein-protein interaction with 658
the 20S proteasome. PLoS ONE 7:e35336. doi:10.1371/journal.pone.0035336. 659
50. Madsen L, Kriegenburg F, Vala A, Best D, Prag S, Hofmann K, Seeger M, 660
Adams IR, Hartmann-Petersen R. 2011. The tissue-specific Rep8/UBXD6 661
tethers p97 to the endoplasmic reticulum membrane for degradation of misfolded 662
proteins. PLoS ONE 6:e25061. doi:10.1371/journal.pone.0025061. 663
51. Marchler-Bauer A, Lu S, Anderson JB, Chitsaz F, Derbyshire MK, DeWeese-664
Scott C, Fong JH, Geer LY, Geer RC, Gonzales NR, Gwadz M, Hurwitz DI, 665
Jackson JD, Ke Z, Lanczycki CJ, Lu F, Marchler GH, Mullokandov M, 666
Omelchenko MV, Robertson CL, Song JS, Thanki N, Yamashita RA, Zhang 667
D, Zhang N, Zheng C, Bryant SH. 2011. CDD: a Conserved Domain Database 668
for the functional annotation of proteins. Nucleic Acids Res. 39:D225-D229. 669
52. Mason JM, Arndt KM. 2004. Coiled Coil Domains: Stability, Specificity, and 670
Biological Implications. ChemBioChem 5:170-176. 671

26
53. Matsuzaki M, Misumi O, Shin-I T, Maruyama S, Takahara M, Miyagishima S-672
Y, Mori T, Nishida K, Yagisawa F, Nishida K, Yoshida Y, Nishimura Y, Nakao 673
S, Kobayashi T, Momoyama Y, Higashiyama T, Minoda A, Sano M, Nomoto 674
H, Oishi K, Hayashi H, Ohta F, Nishizaka S, Haga S, Miura S, Morishita T, 675
Kabeya Y, Terasawa K, Suzuki Y, Ishii Y, Asakawa S, Takano H, Ohta N, 676
Kuroiwa H, Tanaka K, Shimizu N, Sugano S, Sato N, Nozaki H, Ogasawara 677
N, Kohara Y, Kuroiwa T. 2004. Genome sequence of the ultrasmall unicellular 678
red alga Cyanidioschyzon merolae 10D. Nature 428:653-657. 679
54. Meyer H, Bug M, Bremer S. 2012. Emerging functions of the VCP/p97 AAA-680
ATPase in the ubiquitin system. Nat. Cell Biol. 14:117-123. 681
55. Moog D, Stork S, Zauner S, Maier U-G. 2011. In silico and in vivo investigations 682
of proteins of a minimized eukaryotic cytoplasm. Genome Biol. Evol. 3:375-382. 683
doi:10.1093/gbe/evr031. 684
56. Moustafa A, Beszteri B, Maier UG, Bowler C, Valentin K, Bhattacharya D. 685
2009. Genomic footprints of a cryptic plastid endosymbiosis in diatoms. Science 686
324:1724-1726. 687
57. Nassoury N, Cappadocia M, Morse D. 2003. Plastid ultrastructure defines the 688
protein import pathway in dinoflagellates. J. Cell Sci. 116:2867-2874. 689
58. Patron NJ, Waller RF. 2007. Transit peptide diversity and divergence: a global 690
analysis of plastid targeting signals. Bioessays 29:1048-1058. 691
59. Richly H, Rape M, Braun S, Rumpf S, Hoege C, S. 2005. A series of ubiquitin 692
binding factors connects CDC48/p97 to substrate multiubiquitylation and 693
proteasomal targeting. Cell 120:73-84. 694
60. Saeki Y, Tanaka K. 2012. Assembly and function of the proteasome. Methods 695
Mol. Biol. 832:315-337. 696
61. Sanchez-Puerta MV, Delwiche CF. 2008. A hypothesis for plastid evolution in 697
chromalveolates. J. Phycol. 44:1097-1107. 698
62. Schuberth C, Buchberger A. 2005. Membrane-bound Ubx2 recruits Cdc48 to 699
ubiquitin ligases and their substrates to ensure efficient ER-associated protein 700
degradation. Nat. Cell Biol. 7:999-1006. 701
63. Schuberth C, Buchberger A. 2008. UBX domain proteins: major regulators of 702
the AAA ATPase Cdc48/p97. Cell. Mol. Life Sci. 65:2360-2371. 703
64. Sheiner L, Demerly JL, Poulsen N, Beatty WL, Lucas O, Behnke MS, White 704
MW, Striepen B. 2011. A systematic screen to discover and analyze apicoplast 705
proteins identifies a conserved and essential protein import factor. PLoS Path. 706
7:e1002392. doi:10.1371/journal.ppat.1002392. 707
65. Sheiner L, Striepen B. 2012. Protein sorting in complex plastids. Biochim. 708
Biophys. Acta. doi:10.1016/j.bbamcr.2012.05.030. 709
66. Smith MH, Ploegh HL, Weissman JS. 2011. Road to ruin: targeting proteins for 710
degradation in the endoplasmic reticulum. Science 334:1086-1090. 711

27
67. Sommer MS, Gould SB, Lehmann P, Gruber A, Przyborski JM, Maier U-G. 712
2007. Der1-mediated preprotein import into the periplastid compartment of 713
chromalveolates? Mol. Biol. Evol. 24:918-928. 714
68. Spork S, Hiss JA, Mandel K, Sommer M, Kooij TWA, Chu T, Schneider G, 715
Maier UG, Przyborski JM. 2009. An unusual ERAD-like complex is targeted to 716
the apicoplast of Plasmodium falciparum. Eukaryot. Cell 8:1134-1145. 717
69. Su V, Lau AF. 2009. Ubiquitin-like and ubiquitin-associated domain proteins: 718
significance in proteasomal degradation. Cell. Mol. Life Sci. 66:2819-2833. 719
70. Suzuki T, Park H, Lennarz WJ. 2002. Cytoplasmic peptide:N-glycanase 720
(PNGase) in eukaryotic cells: occurrence, primary structure, and potential 721
functions. FASEB J. 16:635-641. 722
71. Teich R, Zauner S, Baurain D, Brinkmann H, Petersen J. 2007. Origin and 723
distribution of Calvin cycle fructose and sedoheptulose bisphosphatases in 724
plantae and complex algae: a single secondary origin of complex red plastids 725
and subsequent propagation via tertiary endosymbioses. Protist 158:263-276. 726
72. Trempe J-F. 2011. Reading the ubiquitin postal code. Curr. Opin. Struct. Biol. 727
21:792-801. 728
73. van Dooren GG, Tomova C, Agrawal S, Humbel BM, Striepen B. 2008. 729
Toxoplasma gondii Tic20 is essential for apicoplast protein import. Proc. Natl. 730
Acad. Sci. U. S. A. 105:13574-13579. 731
74. Visendi P, Ng'ang'a W, Bulimo W, Bishop R, Ochanda J, de Villiers EP. 732
2011. TparvaDB: a database to support Theileria parva vaccine development. 733
Database (Oxford) 2011:bar015. 734
75. Woehle C, Dagan T, Martin WF, Gould SB. 2011. Red and problematic green 735
phylogenetic signals among thousands of nuclear genes from the photosynthetic 736
and apicomplexa-related Chromera velia. Genome Biol. Evol. 3:1220-1230. 737
doi:10.1093/gbe/evr100. 738
76. Wolf DH, Stolz A. 2012. The Cdc48 machine in endoplasmic reticulum 739
associated protein degradation. Biochim. Biophys. Acta 1823:117-124. 740
77. Zaslavskaia LA, Lippmeier JC, Kroth PG, Grossman AR, Apt KE. 2000. 741
Transformation of the diatom Phaeodactylum tricornutum (Bacillariophyceae) 742
with a variety of selectable marker and reporter genes. J. Phycol. 36:379-386. 743
744
745

28
Figure legends 746
747
Figure 1 748
In vivo-localization and domain organization of new SELMA proteins of the diatom P. 749
tricornutum. A: In vivo-localizations of PtsUBX, PtsNpl4, PtsPng1 and PtsUbq as eGFP 750
fusion proteins in P. tricornutum show the characteristic PPC fluorescence in the middle 751
of the two plastid lobes (scale: 10 µm, TL: transmission light, PAF: plastid 752
autofluorescence). B: Structural overview of the domain composition of PtsNpl4, PtsUbq 753
and PtsPng1 in comparison to the respective S. cerevisiae ERAD protein (ScNpl4p, 754
ScDsk2p and ScPng1p). Whereas PtsNpl4 and PtsPng1 share the conserved domains 755
of their yeast counterparts, PtsUbq lacks the UBA domain for poly-ubiquitin binding. 756
PtsUBX cannot be assigned to a specific ERAD protein. (red: signal peptide, blue: TPL 757
predicted, light blue: no TPL predicted, UBX: domain present in ubiquitin-regulatory 758
proteins, orange: coiled coil region, UBQ: ubiquitin homologues, STI1: heat shock 759
chaperonin-binding motif, UBA: ubiquitin associated domain; TG: 760
transglutaminase/protease-like homologues; Rad4: Rad4 transglutaminase-like 761
domain). 762
Figure 2 763
Alignment of the ubiquitin domain of hUbi and sUbi sequences with the S. cerevisiae 764
and C. merolae ubiquitins. The host ubiquitin sequences are derived from polyubiquitins 765
consisting of multiple ubiquitin domains of the same sequence. Protein sequences of 766
sUbi from P. tricornutum, F. cylindrus and T. pseudonana share the same lysine 767
mutations at positions 48 and 63. E. huxleyi, T. parva and B. bovis only show an altered 768
Lys63 position. Both ubiquitin domains of the F. cylindrus symbiont di-ubiquitin have 769

29
identical protein sequences, in contrast, the di-ubiquitin of G. theta is depicted as two 770
independent ubiquitin domains (GtsUbi_155024_1/2) (* There is no polyubiquitin in C. 771
merolae, therefore a ubiquitin-ribosomal fusion protein was included in the alignment. 772
For detailed information on protein sequences see supplemental file 1). 773
Figure 3 774
In vivo-localization of new symbiont 20S proteasomal components in the PPC of the 775
diatom P. tricornutum. eGFP fusion proteins Ptsβ1 and Ptsα3-1 show fluorescence in 776
the middle of the two plastid lobes indicating a PPC localization in contrast to the host 777
proteins Pthβ7 and PthRpn10 which localize in areas around the plastid (scale: 10 µm; 778
TL: transmission light; PAF: plastid autofluorescence). 779
Figure 4 780
Schematic model of protein import into the PPC across the SELMA translocation 781
complex in diatoms. Upon recognition of a SELMA substrate in the cER lumen, 782
precursor proteins are translocated across the membrane complex composed of sDer1-783
1/-2 and the ubiquitin ligase ptE3p. As soon as they reach the PPC, the precursors are 784
ubiquitylated via a PPC-located ubiquitylation machinery. This leads to a recruitment of 785
the Cdc48 complex with its co-factors sUfd1 and sNpl4 and translocation is completed. 786
The ubiquitin moiety is most likely removed prior of protein maturation for PPC resident 787
proteins or further transport for plastid proteins. The identified 20S proteasomal 788
components function according to current knowledge independent of ubiquitylation. 789
(cER: chloroplast ER; PPC: periplastidal compartment; IMS: intermembrane space; 790
modified after (39), orange indicates proteins identified in this study) 791

30
TABLE 1 Overview of all identified host ERAD and symbiont SELMA components in organisms with secondary plastids of red algal origin compared to ERAD proteins of four red algal species.
Protein complex/
function
Protein name host (symbiont)
Red algae Heterokontophytes Haptophytes Cryptophytes Apicomplexans
C
. m
ero
lae
P. c
rue
ntu
m
C.
tub
erc
ulo
su
m
G.
su
lph
ura
ria
P.
tric
orn
utu
m
T.
pse
ud
on
an
a
F.
cyli
nd
rus
E.
sil
ico
los
us
A.
an
op
ha
geff
ere
ns
E.
hu
xle
yi
G.
the
ta
P.
falc
ipa
rum
T.
go
nd
ii
B. b
ovis
N. c
an
inu
m
T.
parv
a
C. p
arv
um
H S H S H S H S H S H S H S H S H S H S H S H S H
ER translocon Sec61 X X X X X(67)
Xa X X
a X X X
(67) X
(67) X X X X X
Derlin proteins Dfm1/ hDer1-1
(sDer1-1)
X X X X X(38, 67)
X(38, 67)
X X X X X X X X X(26)
X(26)
X(67)
X(67)
X(67-68)
X(67-68)
X(2, 68)
X(2, 68)
X(68)
X X X X(68)
Der1/hDer1-2
(sDer1-2)
X X X X(38, 67)
X(38, 67)
Xa X X (X) X X X X X
(26) X
(26) X X X
(67-68) X
(67-68) X
(2, 68) X
(68) X
(68) X X
(68) X
(68)
Ubiquitylation Hrd1/Der3 (ptE3P) X X X X X(67)
X(39)
X X X X X X X X X(67)
X(18, 67-68)
X(65)
X(68)
X X(68)
X(68)
Hrd3p X X X X X X Xa,(67)
X(65)
X(68)
X
Uba1 (sUba1) X X X X X(67)
X(67)
X X X X X (X) X (X) Xa,(26)
X(26)
Xa X X
(18, 67-68) X
(67-68) X
(65) X
(64) X X X X
(68)
Ubc (sUbc) Xa X
a X
a X
a X
a, (67) X
a X
a X
a X
a X
a X
a X
a X
a X
a X X
a X
a,(31) X
a(18) X
(68) X
a X
(2, 68) X
a X
(68) X
a X X
a X
(68) X
a
Doa10 X X X X X X X Xa X X
polyubiquitin (sUbi) X(67)
X(67)
X X X X X Xa X X X
a X X
(67-68) X
(68) X X
(68) X
(68) X X
(68) X X
(68)
Cdc48 complex Cdc48 (sCdc48-1) X X X X(67)
X(67)
X X X X X X X X X(26)
X(26)
X X(67)
X(67-68)
X(67-68)
X(2, 68)
X(2, 68)
X(68)
X X X X(68)
X X(68)
(sCdc48-2) X(55)
X X X X X X
Ufd1 (sUfd1) X X X X(67)
X(67)
X X X X Xa X X X X
(26) X
(26) X
a X
(67) X
(67-68) X
(67-68) X
(2, 68) X
(2, 68) X
(68) X
(68) X X X
(68) X
(68) X
(68)
Npl4 (sNpl4) X X X X(67)
X X X X X X X X X X(67-68)
X(68)
X(68)
X(68)
X(68)
(sUBX) X (X) (X) X
(sPUB) X(55)
X X X X
Processing Png1 (sPng1) X X X X(67)
X X X X X X X X X X Xa X
Dsk2 (sUbq) X X X X X X X X X X X X X X X X X X X X
Rad23 X X X X(67)
X X Xa X X X X X X
(67) X X X X X
Ufd2 X X X X X(67)
X X X X X Xa X
(67) X X X X X
(ptDUP) X(39)
(X) X X X X
Unknown (PPP1) X(64)
X(64)
X(55, 64)
X(64)
X(64)
X(64)
X(64)
X(64)
X(64)
X(64)
X(64)
X(64)
X(64)
Chaperones Hsp70 (sHsp70) X X X X(67)
X (31)
X X X X X X X X X X X X X X X X X
Hsp40 X X X X X X X X X X X X
(sDPC) X(52)
X X X X

31
Proteins similar to ScUsa1p, ScCue1p and ScUbx4p could not be identified in host or symbiont version and are therefore not included in the table. Ubc proteins are further assigned to the most similar S. cerevisiae Ubc enzyme in supplemental file 2 (Table S1). X: detected; (X): symbiont gene detected by homology but without targeting sequence;
a: more than one gene detected. Numbers in
brackets refer to respective literature. Protein identifiers can be found in supplemental file 2 (Table S1).
792
793

32
TABLE 2: Overview of host and symbiont 20S proteasomal components of heterokontophytes and cryptophytes compared to those from red algae.
Red algae Heterokontophytes
Crypto- phytes
Protein name
C.
mero
lae
P.
cru
en
tum
C.
tub
erc
ulo
su
m
G.
su
lph
ura
ria
P.
tric
orn
utu
m
T.
ps
eu
do
na
na
F.
cyli
nd
rus
E.
sil
ico
los
us
A.
an
op
ha
ge
ffe
ren
s
G.
the
ta
H S H S H S H S H S H S
α1 X X
X X (55)
X
X
X
X
X X
α2 X X
X X(55)
X(55)
X X X X
X X X X X
α3 X X X X X(55)
Xa X X
a X X X X X
X X
α4 X X X X X(55)
X
X
X X X
X X
α5 X X
X X(55)
X
X
X X X
X X
α6 X X X X X(55)
X
X
X X X X X X
α7 X X X X X(55)
X(55)
X
X X X
X
X X
alpha general
Xa,(55)
X X
Xa
X
β1 X X
X X
(55) X X X X X X
a
X
a
X X
β2 X X
X X(55)
X X X X X X
Xa
X X
β3 X X
X X(55)
X(55)
X X X X Xa
X X X X
β4 X X
X X(55)
X
X
X
X
X
β5 X X X X Xa,(55)
Xa
X
a
X X X X X X
β6 X X
X X(55)
X(55)
X X X X X X X
X X
β7 X X X X X(55)
X(55)
X
X (X) X
X
X X
X: detected; a: more than one gene detected; (X): symbiont gene detected by homology but without targeting
sequence. Numbers in brackets refer to respective literature. Protein identifiers can be found in supplemental file 2 (Table S2).
794