distinctive molecular composition of human gingival interdental papilla
TRANSCRIPT

Distinctive Molecular Composition ofHuman Gingival Interdental PapillaAndrea Csiszar,* Colin Wiebe,* Hannu Larjava,* and Lari Hakkinen*
Background: Gingiva is composed of attached and mar-ginal (free) gingiva and interdental papilla. Increasing estheticdemands in dentistry have created a need to restore all partsof the gingiva. However, the interdental papilla has limited re-generation potential compared to other parts of the gingiva. Italso is more susceptible to gingival overgrowth, suggestingthat it has distinct cellular and molecular properties from otherparts of the gingiva. Very little is known about the possible dif-ferences in the molecular composition of different parts of thegingiva.
Methods: We compared the expression of a set of key mol-ecules in interdental papilla and marginal gingiva from sevenhealthy subjects by immunohistochemical staining.
Results: In the interdental papilla, immunoreactivity forintegrin avb6 and cytokeratin 19 in the oral epithelium wassignificantly higher than in marginal gingiva. Expression oftype I procollagen, extra domain A (EDA) and extra domain B(EDB) fibronectin isoforms, tenascin-C, transforming growthfactor-b (TGF-b), connective tissue growth factor (CTGF),and the signaling molecule son-of-sevenless (SOS)-1 alsowere increased in the interdental papilla. The expression ofsmall leucine-rich proteoglycans decorin, biglycan, fibro-modulin, and lumican in the interdental papilla was partiallydifferent from the marginal gingiva.
Conclusions: Molecular composition of the interdentalpapilla is distinct from marginal gingiva. Increased expressionof molecules normally induced in wound healing (avb6 in-tegrin, fibronectin-EDB and -EDA, tenascin-C, type I procolla-gen, TGF-b, CTGF, and SOS-1) suggests that the cells inthe interdental papilla are in an activated state and/or inher-ently display a specific phenotype resembling wound healing.J Periodontol 2007;78:304-314.
KEY WORDS
Extracellular matrix; gingiva; growth factors; integrins.
Human gingiva is composed ofdistinct anatomical units, namelyattached gingiva, which directly
binds gingiva to the underlying bone,and marginal (free) gingiva, a narrowband of tissue that does not bind to ahard tissue and surrounds the toothlike a collar. In a vertical direction, themarginal gingiva spans from the ce-mento-enamel junction to the free gin-gival groove. The part of the gingiva thatoccupies the interdental space is calledthe interdental papilla.1,2 Anatomically,the interdental papilla is the part of thegingiva that extends from the interden-tal alveolar bone to the contact point ofadjacent teeth. It is demarcated fromthe mesial and distal sides by sulcularand/or junctional epithelium, whereas thefacial and oral sides are covered by akeratinized gingival oral epithelium. Atposterior teeth, the buccal and lingualpapilla are joined by a valley-like de-pression called the col. The interdentalpapilla forms when the teeth erupt, andits shape is determined by the contactpoints of adjacent teeth, the widths ofproximal tooth surfaces, and the shapeof the cemento-enamel junction.1,3 In-terdental papilla is exposed easily toagents that cause inflammation throughthe junctional/sulcular epithelia; even ina clinically healthy papilla, some degreeof local inflammation is a common histo-logic finding.
Increasing esthetic demands in dentis-try have created a need to maintain andrestore the different parts of the gingivalcomplex, in particular the interdental
* Faculty of Dentistry, Department of Oral Biological and Medical Sciences, Laboratory ofPeriodontal Biology, University of British Columbia, Vancouver, BC.
doi: 10.1902/jop.2007.060165
Volume 78 • Number 2
304

papilla. Loss of the interdental papilla leads to estheticconcerns, affects speech, and may promote food im-paction leading to periodontal problems. However,when the interdental papilla is lost as the result of peri-odontal disease, surgery, or trauma, its capacity to re-generate is limited compared to marginal or attachedgingiva. In addition, normal marginal gingiva, includ-ing junctional and sulcular epithelium, forms arounddental implants placed in edentulous areas, whereasthe regeneration of interdental papilla remains limited.Drug-induced and hereditary gingival overgrowth ap-pears to manifest first at the interdental papilla; fromhere the overgrowth spreads to cover the other partsof the gingiva,2 suggesting that the interdental papillais predisposed to gingival overgrowth.
The limited regenerative capacity of the interdentalpapilla seems to depend on anatomical factors, boneloss, plaque, and inflammation in the interdentalarea.3-9 It also is possible that the interdental papillahas unique functional characteristics that are due todistinct cellular or molecular properties. It is welldocumented, even in the marginal gingiva, that differ-ent anatomical tissue units have specific characteris-tics. For example, marginal gingiva contains threedifferent epithelia, namely junctional, sulcular, andoral gingival epithelium. These epithelia have differ-ent cell turnover rates and can be keratinized ornon-keratinized.10,11 In addition, the expression ofcertain cytokeratins within the gingival epithelia de-pends on the anatomical location of the epithelium,i.e., whether the epithelium is exposed to the oralenvironment directly (oral gingival epithelium) orwhether it opposes tooth (sulcular epithelium andjunctional epithelium).10 Much less is known aboutmolecular differences in the connective tissue in differ-ent anatomical regions of the gingiva. It is reasonableto assume that these differences exist because theconnective tissue phenotype is modulated by the ep-ithelium and vice versa.12,13 Additionally, fibroblastscultured from different anatomical locations of gingivadisplay inherently distinct phenotypical properties.14
To characterize some of the cellular and molecularcharacteristics of the interdental papilla, we comparedthe expression of a set of molecules in healthy humanmarginal gingiva and interdental papilla.
MATERIALS AND METHODS
Tissue SamplesBiopsies of marginal gingiva and interdental papillawere obtained from seven healthy subjects (fourmales and three females) between 25 and 65 yearsof age during crown-lengthening surgery. All biopsiescontained facial/buccal marginal gingiva and inter-dental papilla. All subjects were medically healthy,did not have periodontal disease in the surgical sex-tant, were non-smokers, and were not taking any
medications. No clinical signs of inflammation werepresent at the biopsy site. Informed consent was ob-tained from all subjects, and all procedures were in ac-cordance with regulations set forth by the ResearchEthics Board, University of British Columbia. Patientswere enrolled in the study from 2005 to 2006.
After harvesting, biopsies were immersed in phys-iological saline for a maximum of 1 hour, embedded inoptimal cutting temperature compound,† and imme-diately frozen in liquid nitrogen. Tissue block sam-ples were stored at -86�C until frozen sections wereprepared. Serial bucco-lingual (marginal gingiva) orbucco-lingual and mesio-distal (interdental papilla)cryostat sections (5 mm) were prepared and placedonto 3-aminopropyltriethoxysilane–coated slides,‡
air dried, and stored at -86�C until further use.
ImmunostainingThe following primary antibodies were used for immu-nostaining: anti-cytokeratin19, anti–pan-transforminggrowth factor-b (recognizes transforming growth fac-tor [TGF]-b1, -2, and -3), anti-son-of-sevenless-1,§
anti-integrin avb6,15 anti-procollagen type I,i anti-decorin,16 anti-biglycan,17 anti-fibromodulin,18 anti-lumican,19 anti-fibronectin extra domain A (EDA),¶ anti-fibronectin extra domain B (EDB),20 anti-tenascin-C,#
and anti-connective tissue growth factor.**For immunostaining, frozen tissue samples were
thawed briefly at room temperature prior to fixationwith -20�C acetone for 5 minutes. After washing withphosphate buffered saline (PBS) containing 1 mg/mlbovine serum albumin (BSA) and 0.01% Triton-X100, the sections were incubated with the appropriatenormal blocking serum†† at room temperature for30 minutes to block non-specific binding sites. Pri-mary antibody diluted in PBS/BSA/Triton X-100 wasadded, and the samples were left in a humidifiedchamber at 4�C. After 18 hours of incubation, the sec-tions were washed with PBS/BSA/Triton X-100 andincubated with an appropriate biotinylated secondaryantibody‡‡ for 1 hour. They were then washed withPBS/BSA/Triton X-100, incubated with reagent§§
for 30 minutes, washed again, and reacted with re-agentii until suitable color development was obtained.Reaction times for each antibody were standardizedfor all samples. Sections were left to air dry and weremounted using a solution.¶¶ Negative control stainings
† Tissue-Tek, Miles, Elkhart, IA.‡ Fluka Chemie, Buchs, Switzerland.§ A53B/A2, Santa Cruz Biotechnology, Santa Cruz, CA.i Chemicon International, Temecula, CA.¶ 1ST-9, Accurate Chemical & Scientific, Westbury, NY.# Sigma BioSciences, St. Louis, MO.** FibroGen, San Francisco, CA.†† Vectastain Kit, Vector Laboratories, Burlingame, CA.‡‡ Vector Laboratories.§§ Vectastain ABC, Vector Laboratories.ii VIP, Vector Laboratories.¶¶ Vecta-Mount, Vector Laboratories.
J Periodontol • February 2007 Csiszar, Wiebe, Larjava, Hakkinen
305

were performed by omitting the primary antibody. Nocolor reaction was observed in the negative controlsamples. Representative sections were examinedand photographed using a digital camera## attachedto a microscope using 10·, 20·, and 40· objec-tives.*** Sections from two to four different areas ofthe tissue sample from the marginal gingiva and inter-dental papilla from five to seven subjects were used forimmunostaining for each molecule of interest. Thus, atleast 10 sections in total for each molecule in marginalgingiva and interdental papilla were used for the anal-yses. Relative staining intensity was assessed by twoindependent calibrated examiners for each moleculeat the marginal gingiva and interdental papilla in eachsample and scored as follows: 0 = no immunoreactiv-ity; 1 = weak, but visible staining intensity; 2 = moder-ate staining intensity; and 3 = strong staining intensity.Statistical significance between the mean scores inmarginal gingiva and interdental papilla was deter-mined by using Student t test. Every tenth slide fromeach subject was stained with hematoxylin and eosin(H&E) for gross morphological characterization andanalysis of the presence or absence of inflammatorycell infiltrate. Only tissue areas with no inflammatorycell infiltrate were used for immunohistologic analysis.To analyze the collagen fibril organization, the H&E-stained sections were photographed using a fluores-cence microscope equipped with a rhodamine filter.†††
RESULTS
Histologic CharacterizationUnder histologic examination of the H&E-stainedsections, the oral epithelium in the marginal gingivashowed keratinization, whereas the interdental papillashowed areas of parakeratinized and non-keratinizedepithelium (data not shown). The parakeratinizedareas corresponded to the oral epithelium of the in-terdental papilla, whereas the non-keratinized areasrepresented junctional/sulcular epithelium. The oralepithelium in the interdental papilla appeared slightlythicker and rete ridges were longer than in the margi-nal gingiva (Figs. 1A and 1B). The relative density offibroblasts, identified based on their typical spindle-shaped morphology, was similar in both tissue loca-tions, whereas the relative number of blood vesselswas higher in the interdental papilla. No inflammationwas noted in the marginal gingiva samples. However,tissue sections from the interdental papilla containedsome localized areas with mild to moderate inflamma-tion. Only tissue areas with no inflammatory cell infil-trates were used for immunohistochemical analyses.
Immunohistochemical Characterization ofthe EpitheliumSeveral statistically significant differences in the ex-pression of the target molecules were noted between
Figure 1.Histologic characterization and comparison of CK19, avb6 integrin,fibromodulin, and lumican expression in marginal gingiva (A, C, E, G,and I) and interdental papilla (B, D, F, H, and J). Arrows in C showisolated cells at the tips of rete ridges expressing CK19. Arrows andthe inset in F show tips of rete ridges that demonstrate the strongestimmunoreactivity for avb6 integrin. Arrows in H show strongimmunoreactivity at the tips of rete ridges. Inset in H shows highermagnification of strong immunostaining that associates with collagenfibrils. Arrows in J show strong immunoreactivity at the tips of reteridges. Inset in J shows higher magnification of strong immunostainingthat associates with collagen fibrils. Representative samples from atleast 10 sections from five to seven subjects are shown. E = oralepithelium; BC = basal cells; SS = stratum spinosum; SG = stratumgranulosum; SECT = subepithelial connective tissue; DCT = deepconnective tissue. Bar = 100 mm. (H&E, A and B; CK19immunostaining, C and D; integrin avb6 immunostaining, E and F;fibromodulin immunostaining, G and H; lumican immunostaining,I and J.)
## Coolpix 995, Nikon, Richmond, BC.*** Eclipse TS 100, Nikon.††† Zeiss Axiolab E light microscope, Carl Zeiss, Jena, Germany.
Molecular Composition of Interdental Papilla Volume 78 • Number 2
306

oral epithelia of the marginal gingiva and interdentalpapilla (Table 1). With the exception of the occasionalbasal cell, cytokeratin 19 (CK19) was never ex-pressed in the oral epithelium of marginal gingiva(Fig. 1C). In contrast, CK19 was expressed stronglyin the epithelium of the interdental papilla. In thenon-keratinized sulcular/junctional epithelium, CK19localized throughout the epithelium, whereas in theoral epithelium, where epithelium was parakerati-nized, CK19 showed strong expression in the basalcells only (Fig. 1D). No expression of avb6 integrinwas noted in the marginal gingiva (Fig. 1E), whereasthe epithelium in the interdental papilla stained posi-tively (Fig. 1F). Strongest expression of avb6 integrinwas localized at the tips of rete ridges of the interdentalpapilla oral epithelium (Fig. 1F). In the marginal gin-giva, biglycan immunoreactivity localized most abun-dantly to the basal epithelial cells of oral epithelium;however, weak immunoreactivity also was noted inthe stratum spinosum. In contrast, biglycan immuno-reactivity was moderate to strong throughout the oralepithelium in the interdental papilla (Table 1). Fibro-modulin showed weak immunoreactivity in the basalepithelial cells and in the immediate two to four supra-basal cell layers of the oral epithelium of the marginalgingiva (Fig. 1G), whereas in the interdental papilla itwas expressed strongly by the basal oral epithelialcells (Fig. 1H). Lumican showed moderate immuno-reactivity at the basal epithelial cells and weak stain-ing at the stratum spinosum in the marginal gingiva(Fig. 1I). In contrast, staining intensity for lumicanwas moderate to strong in the interdental papilla epi-thelium. The basal cells, especially at the tips of reteridges, showed strong immunostaining for lumican(Fig. 1J). TGF-b showed two different patterns of im-munostaining in the marginal gingiva. In some areas,there was relatively weak immunoreactivity at thebasal epithelial cells, whereas the other oral epitheliallayers were negative (Fig. 2A). In other areas of oralepithelium, basal cells showed moderate to strong im-munoreactivity for TGF-b at the cell membrane facingthe basement membrane. In these areas, the cells inthe stratum spinosum and granulosum also showedvariable, but weaker, immunoreactivity for TGF-b
(Fig. 2B). A similar staining pattern for TGF-b alsowas noted in the oral epithelium of the interdental pa-pilla (Figs. 2C and 2D). The relative staining intensityof TGF-b appeared stronger in the interdental papillathan in the marginal gingiva, but the difference did notreach statistical significance (Figs. 2A through 2D;Table 1). Moderate immunostaining intensity for con-nective tissue growth factor (CTGF) was localized tothe basal cells in the oral epithelium of the marginalgingiva. Furthermore, the two to four immediatesuprabasal cell layers showed weak immunostainingfor CTGF (Figs. 2G and 2I). CTGF localized to the
basal epithelial cells also in the interdental papilla oralepithelium (Figs. 2H and 2K); however, in contrast tothe marginal gingiva, the suprabasal cells, especiallyat the stratum granulosum, showed strong immunore-activity for CTGF (Fig. 2H). In the marginal gingiva,
Figure 2.Immunohistochemical staining of TGF-b (A through F), CTGF (Gthrough L), and SOS-1 (M through R) in marginal gingiva (A, B, E,G, I, J, M, O, and P) and interdental papilla (C, D, F, H, K, L, N, Q,and R). Short arrows indicate blood vessels in E, F, J, L, P, and R. Longarrows indicate tips of rete ridges showing strong immunoreactivity forSOS-1 in N. E = oral epithelium; BC = basal cells; SS = stratumspinosum; SG = stratum granulosum; CT = connective tissue.Representative samples from at least 10 sections from five to sevensubjects are shown. Bar = 100 mm.
J Periodontol • February 2007 Csiszar, Wiebe, Larjava, Hakkinen
307

son-of-sevenless (SOS)-1 localized most abundantlyat the basal oral epithelial cells, whereas the supra-basal cell layers were almost negative (Figs. 2M and2O). In contrast, SOS-1 showed significantly strongerimmunostaining intensity throughout the oral epithe-lium of the interdental papilla, with the strongest ex-pression in the basal cells, especially at the tips ofrete ridges (Figs. 2N and 2Q). No immunoreactivityfor procollagen, decorin, fibronectin-EDA, fibronec-tin-EDB, or tenascin-C was noted in the epitheliumof the marginal gingiva or interdental papilla (Table 1).
Immunohistochemical Characterization of theConnective Tissue Extracellular MatrixIn the marginal gingiva, connective tissue was com-posed of thick collagen fiber bundles that displayedthe typical basket-weave organization. In contrast,collagen fibers often were organized into parallel bun-dles that appeared thinner in the interdental papilla(Figs. 3A and 3B, respectively). Healthy marginal gin-giva and papilla showed several statistically signifi-cant differences in the expression of the connective
tissue extracellular matrix molecules (Table 2). Inmarginal gingiva and interdental papilla, procollagentype I, the precursor of type I collagen, showed thestrongest immunoreactivity at the basement mem-brane zone and in the connective tissue immediatelybeneath the basement membrane (Figs. 3C and 3D).However, only a weak immunoreactivity of procolla-gen localized in the deep connective tissue of themarginal gingiva (Fig. 3C), whereas it showed signif-icantly stronger staining in the deep connective tissueof the interdental papilla (Fig. 3D). In marginal gingivaand interdental papilla, decorin localized to collagenfiber bundles and showed the most intense stainingin the subepithelial connective tissue (Table 2 anddata not shown). In contrast to the marginal gingiva,thinner isolated collagen bundles that showed decorinstaining were present in the papilla (data not shown).Biglycan also associated with collagen fiber bundles,but it showed stronger immunoreactivity at the deepconnective tissue in the interdental papilla than inthe marginal gingiva (Table 2 and data not shown).Fibromodulin showed weak to moderate staining
Table 1.
Comparison of Relative Immunostaining Intensity in the Oral Epithelium of the MarginalGingiva and Interdental Papilla
Marginal Gingiva Interdental Papilla
Antibody SG SS BC SG SS BC
CK19 0 0 0 1.0 – 0.4 1.8 – 0.2* 2.6 – 0.2*
Integrin avb6 0 0 0 0.8 – 0.3 1.6 – 0.4† 2.6 – 0.2*
Procollagen type I 0 0 0 0 0 0
Decorin 0 0 0 0 0 0
Biglycan 0 0.6 – 0.2 2.0 – 0.0 1.4 – 0.2‡ 1.8 – 0.2‡ 2.4 – 0.2
Fibromodulin 0 0.2 – 0.2 0.8 – 0.2 0.2 – 0.2 1.0 – 0.4 2.4 – 0.2‡
Lumican 0 1.2 – 0.2 2.2 – 0.4 1.8 – 0.3‡ 2.2 – 0.4‡ 3.0 – 0.0‡
Fibronectin-EDA 0 0 0 0 0 0
Fibronectin-EDB 0 0 0 0 0 0
Tenascin-C 0 0 0 0 0 0
TGF-b 0.8 – 0.4 1.0 – 0.3 1.0 – 0.4 1.2 – 0.6 1.6 – 0.4 2.2 – 0.4
CTGF 0 0.8 – 0.2 2.2 – 0.2 2.0 – 0.3‡ 1.4 – 0.2 2.0 – 0.0
SOS-1 0.2 – 0.2 0.8 – 0.2 2.0 – 0.0 1.2 – 0.2‡ 1.8 – 0.2‡ 2.8 – 0.2‡
At least 10 sections from the marginal gingiva and interdental papilla samples from five to seven subjects were used for the analyses of the relative stainingintensity (0 = no immunoreactivity; 1 = weak, but visible staining intensity; 2 = moderate staining intensity; and 3 = strong staining intensity) of the givenmolecule in different tissue locations. Results show mean – SEM of parallel samples. Statistical analysis was performed by comparing scores of eachmolecule at the same tissue location in marginal gingiva and interdental papilla by using the Student t test.SG = stratum granulosum; SS = stratum spinosum; BC = basal cell.* P <0.001.† P <0.05.‡ P <0.005.
Molecular Composition of Interdental Papilla Volume 78 • Number 2
308

intensity throughout the connective tissue whereit associated with collagen fiber bundles (Figs. 1Gand 1H). However, in the deep connective tissue ofthe interdental papilla, fibromodulin showed strongimmunoreactivity on the thin collagen fiber bundles,whereas the staining intensity was weaker in the inter-fibrillar extracellular matrix (Fig. 1H, inset). Like theother small leucine-rich proteoglycans, lumican alsolocalized on the collagen fiber bundles. The strongestexpression of lumican was noted in the deep connec-
tive tissue (Fig. 1I); however, in the interdental papilla,immunostaining intensity was significantly weakerthan in the marginal gingiva (Fig. 1J). In the interden-tal papilla, the strongest immunoreactivity localizedto areas where thin collagen fiber bundles were abun-dant (Fig. 1J, inset). Fibronectin-EDA showed weakto moderate immunoreactivity at the basement mem-brane zone and at localized areas of the deep connec-tive tissue of the marginal gingiva (Fig. 3E). In theinterdental papilla, immunostaining intensity for fi-bronectin-EDA was variable, but was significantlystronger throughout the extracellular matrix than inthe marginal gingiva. Like in the marginal gingiva,the strongest immunoreactivity colocalized with thincollagen fiber bundles (Figs. 3E and 3F). Expressionof fibronectin-EDB was absent or very weak in themarginal gingiva (Fig. 3G). However, in the interden-tal papilla, it localized to the basement membranezone (Fig. 3H) and to areas of the extracellular matrixin the subepithelial and deep connective tissue (Fig.3H, inset). In marginal gingiva and interdental papilla,tenascin-C showed strong immunostaining intensityat the basement membrane zone and weaker stainingin a narrow area of connective tissue immediatelybeneath the epithelium (Figs. 3I and 3J). Unlike inthe marginal gingiva, tenascin-C also was expressedstrongly in localized areas of the deep connective tis-sue of the interdental papilla (Fig. 3J). In these loca-tions, tenascin-C immunoreactivity was associatedwith thin collagen fiber bundles.
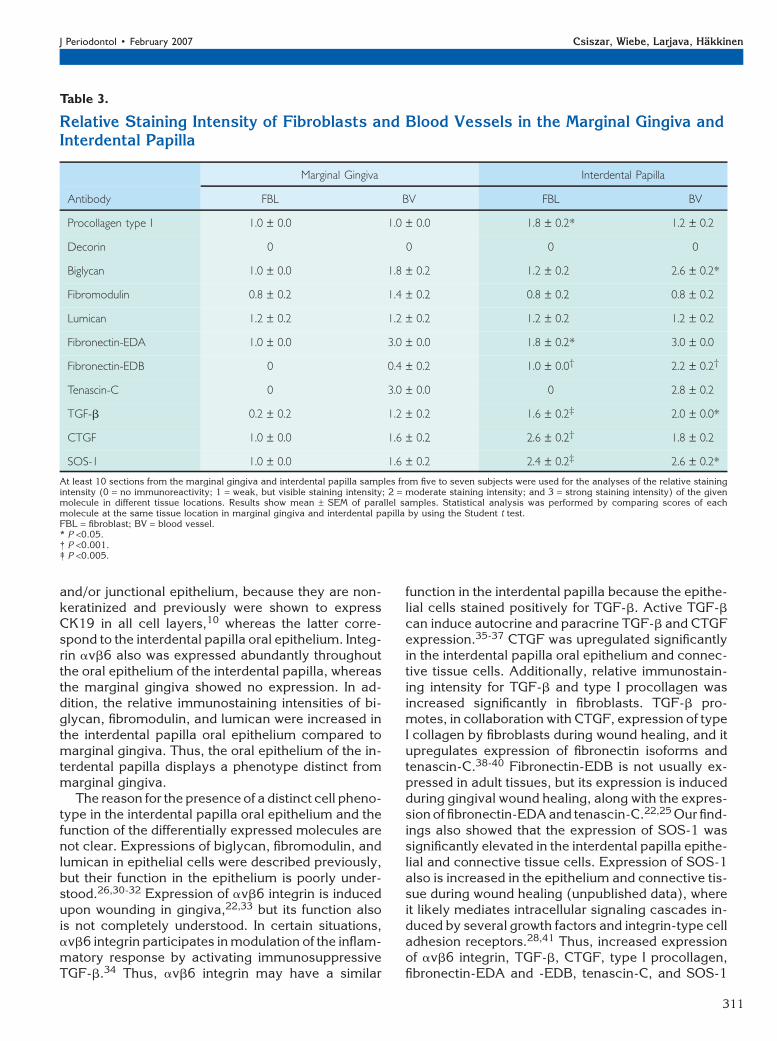
Localization of the Target Molecules inConnective Tissue CellsSeveral statistically significant differences were notedbetween the immunostaining intensity of connectivetissue cells in the marginal gingiva and interdentalpapilla (Table 3). Procollagen type I localized to fibro-blasts and blood vessels in marginal gingiva and inter-dental papilla, but the staining intensity of fibroblastswas significantly stronger in the interdental papilla(Table 3, Figs. 3C and 3D, respectively). No cell-associated staining was noted for decorin by usingthe antibody that recognizes the mature form of themolecule (Table 3). Biglycan showed moderate im-munoreactivity in the blood vessels of the marginalgingiva, whereas fibroblasts showed weaker staining.In the interdental papilla, the staining intensity ofbiglycan in blood vessels was significantly strongerthan in the marginal gingiva (Table 3). Fibromodulinand lumican showed weak immunoreactivity in fibro-blasts and blood vessels in marginal gingiva and in-terdental papilla (Table 3). In the marginal gingiva,blood vessels showed weak immunoreactivity forTGF-b, whereas fibroblasts were mostly negative(Table 3, Fig. 2E). However, blood vessels and fibro-blasts showed significantly stronger immunostaining
Figure 3.Collagen organization (A and B) and immunohistochemicallocalization of procollagen type I (C and D), fibronectin-EDA(FN-EDA) (E and F), fibronectin-EDB (FN-EDB) (G and H), andtenascin-C (I and J) in marginal gingiva (A, C, E, G and I) andinterdental papilla (B, D, F, H and J). A and B: H&E stained sectionsviewed under fluorescent light using a microscope equipped with arhodamine filter (H&E/FL). G and H: Insets show higher magnificationfrom connective tissue. E = oral epithelium; CT = connective tissue;SECT = subepithelial connective tissue; DCT = deep connective tissue.Short arrows indicate blood vessels. Representative samples from ‡10sections from five to seven subjects are shown. Bar = 100 mm.
J Periodontol • February 2007 Csiszar, Wiebe, Larjava, Hakkinen
309

intensity in the interdental papilla than in the marginalgingiva (Table 3, Fig. 2F). In marginal gingiva and in-terdental papilla, CTGF (Table 3, Figs. 2J and 2L) andSOS-1 (Table 3, Figs. 2P and 2R) localized to fibro-blasts and blood vessels, but the staining intensitywas significantly stronger in the interdental papilla.Fibronectin-EDA showed strong immunostaining ofthe blood vessels in marginal gingiva and interdentalpapilla (Figs. 3E and 3F, respectively), but fibroblastsin the interdental papilla showed significantly strongerstaining intensity (Table 3). In the marginal gingiva,fibronectin-EDB was not associated with any fibro-blasts and only occasionally localized weakly to someblood vessels (Fig. 3G). In contrast, blood vessels andfibroblasts in the interdental papilla stained positivelyfor fibronectin-EDB (Table 3, Fig. 3H). In marginalgingiva and interdental papilla, tenascin-C was local-ized to blood vessels, but not to fibroblasts (Table 3,Figs. 3I and 3J).
DISCUSSION
The distinct functional features of the interdentalpapilla suggest that it possesses specific molecularcharacteristics compared to other parts of the gingiva.In the present study, we assessed the expressionof a set of key molecules in the oral epithelium andconnective tissue of human facial/buccal marginalgingiva and interdental papilla. We studied the ex-pression of molecules that are not normally expressed
in gingiva (avb6 integrin and fibronectin-EDB), mol-ecules that are upregulated during wound healingand regulate tissue development and regeneration(avb6 integrin, fibronectin-EDA and -EDB, tenas-cin-C, TGF-b, and CTGF), and molecules that playa role in the organization and homeostasis of the ex-tracellular matrix (type I procollagen and the smallleucine-rich proteoglycans decorin, biglycan, fibro-modulin, and lumican).21-28 We also analyzed theexpression of SOS-1, an intracellular molecule thatregulates key signaling cascades initiated from growthfactor receptors and integrin-type extracellular ma-trix receptors.28 A mutation that apparently resultsin the constitutive activation of SOS-1 is linked to he-reditary gingival fibromatosis.29 We also immunolo-calized CK19 because it was shown to be expressedonly in the non-keratinized sulcular and junctionalepithelium.10
Several statistically significant differences werefound in the relative expression of the studied mole-cules between the marginal gingiva and interdentalpapilla (Table 4). The epithelium in the interdentalpapilla stained positively for CK19, whereas the mar-ginal gingiva was negative except for isolated cells atthe tips of some rete ridges. In areas of non-keratinizedinterdental papilla epithelium, CK19 was expressed inall epithelial cell layers, whereas in areas with para-keratinized epithelium, only the basal epithelial cellswere positive. The former locations represent sulcular
Table 2.
Comparison of Relative Immunostaining Intensity in the Connective Tissue ExtracellularMatrix in the Marginal Gingiva and Interdental Papilla
Marginal Gingiva Interdental Papilla
Antibody BMZ SECT DCT BMZ SECT DCT
Procollagen type I 2.0 – 0.0 1.2 – 0.2 0.8 – 0.2 2.4 – 0.2 2.2 – 0.3 2.0 – 0.2*
Decorin 0 3.0 – 0.0 1.8 – 0.2 0 2.8 – 0.4 2.6 – 0.2
Biglycan 0 1.0 – 0.0 1.6 – 0.2 0 1.4 – 0.2 2.4 – 0.2*
Fibromodulin 0.4 – 0.4 1.6 – 0.2 2.2 – 0.3 0.4 – 0.4 1.4 – 0.4 1.8 – 0.2
Lumican 0 1.8 – 0.2 2.2 – 0.2 0 0.8 – 0.2† 1.0 – 0.0†
Fibronectin-EDA 2.2 – 0.2 0.8 – 0.2 1.4 – 0.2 2.4 – 0.2 2.0 – 0.3* 2.2 – 0.2*
Fibronectin-EDB 0.4 – 0.2 0 0 2.6 – 0.2‡ 1.6 – 0.2† 2.0 – 0.0‡
Tenascin-C 3.0 – 0.0 1.4 – 0.2 0.2 – 0.2 2.8 – 0.2 2.0 – 0.0 1.8 – 0.2‡
At least 10 sections from the marginal gingiva and interdental papilla samples from five to seven subjects were used for the analyses of the relative stainingintensity (0 = no immunoreactivity; 1 = weak, but visible staining intensity; 2 = moderate staining intensity; and 3 = strong staining intensity) of the givenmolecule in different tissue locations. Results show mean – SEM of parallel samples. Statistical analysis was performed by comparing scores of eachmolecule at the same tissue location in marginal gingiva and interdental papilla by using the Student t test.BMZ = basement membrane zone; SECT = subepithelial connective tissue; DCT = deep connective tissue.* P <0.05.† P <0.005.‡ P <0.001.
Molecular Composition of Interdental Papilla Volume 78 • Number 2
310
and/or junctional epithelium, because they are non-keratinized and previously were shown to expressCK19 in all cell layers,10 whereas the latter corre-spond to the interdental papilla oral epithelium. Integ-rin avb6 also was expressed abundantly throughoutthe oral epithelium of the interdental papilla, whereasthe marginal gingiva showed no expression. In ad-dition, the relative immunostaining intensities of bi-glycan, fibromodulin, and lumican were increased inthe interdental papilla oral epithelium compared tomarginal gingiva. Thus, the oral epithelium of the in-terdental papilla displays a phenotype distinct frommarginal gingiva.
The reason for the presence of a distinct cell pheno-type in the interdental papilla oral epithelium and thefunction of the differentially expressed molecules arenot clear. Expressions of biglycan, fibromodulin, andlumican in epithelial cells were described previously,but their function in the epithelium is poorly under-stood.26,30-32 Expression of avb6 integrin is inducedupon wounding in gingiva,22,33 but its function alsois not completely understood. In certain situations,avb6 integrin participates in modulation of the inflam-matory response by activating immunosuppressiveTGF-b.34 Thus, avb6 integrin may have a similar
function in the interdental papilla because the epithe-lial cells stained positively for TGF-b. Active TGF-b
can induce autocrine and paracrine TGF-b and CTGFexpression.35-37 CTGF was upregulated significantlyin the interdental papilla oral epithelium and connec-tive tissue cells. Additionally, relative immunostain-ing intensity for TGF-b and type I procollagen wasincreased significantly in fibroblasts. TGF-b pro-motes, in collaboration with CTGF, expression of typeI collagen by fibroblasts during wound healing, and itupregulates expression of fibronectin isoforms andtenascin-C.38-40 Fibronectin-EDB is not usually ex-pressed in adult tissues, but its expression is inducedduring gingival wound healing, along with the expres-sion of fibronectin-EDA and tenascin-C.22,25 Our find-ings also showed that the expression of SOS-1 wassignificantly elevated in the interdental papilla epithe-lial and connective tissue cells. Expression of SOS-1also is increased in the epithelium and connective tis-sue during wound healing (unpublished data), whereit likely mediates intracellular signaling cascades in-duced by several growth factors and integrin-type celladhesion receptors.28,41 Thus, increased expressionof avb6 integrin, TGF-b, CTGF, type I procollagen,fibronectin-EDA and -EDB, tenascin-C, and SOS-1
Table 3.
Relative Staining Intensity of Fibroblasts and Blood Vessels in the Marginal Gingiva andInterdental Papilla
Marginal Gingiva Interdental Papilla
Antibody FBL BV FBL BV
Procollagen type I 1.0 – 0.0 1.0 – 0.0 1.8 – 0.2* 1.2 – 0.2
Decorin 0 0 0 0
Biglycan 1.0 – 0.0 1.8 – 0.2 1.2 – 0.2 2.6 – 0.2*
Fibromodulin 0.8 – 0.2 1.4 – 0.2 0.8 – 0.2 0.8 – 0.2
Lumican 1.2 – 0.2 1.2 – 0.2 1.2 – 0.2 1.2 – 0.2
Fibronectin-EDA 1.0 – 0.0 3.0 – 0.0 1.8 – 0.2* 3.0 – 0.0
Fibronectin-EDB 0 0.4 – 0.2 1.0 – 0.0† 2.2 – 0.2†
Tenascin-C 0 3.0 – 0.0 0 2.8 – 0.2
TGF-b 0.2 – 0.2 1.2 – 0.2 1.6 – 0.2‡ 2.0 – 0.0*
CTGF 1.0 – 0.0 1.6 – 0.2 2.6 – 0.2† 1.8 – 0.2
SOS-1 1.0 – 0.0 1.6 – 0.2 2.4 – 0.2‡ 2.6 – 0.2*
At least 10 sections from the marginal gingiva and interdental papilla samples from five to seven subjects were used for the analyses of the relative stainingintensity (0 = no immunoreactivity; 1 = weak, but visible staining intensity; 2 = moderate staining intensity; and 3 = strong staining intensity) of the givenmolecule in different tissue locations. Results show mean – SEM of parallel samples. Statistical analysis was performed by comparing scores of eachmolecule at the same tissue location in marginal gingiva and interdental papilla by using the Student t test.FBL = fibroblast; BV = blood vessel.* P <0.05.† P <0.001.‡ P <0.005.
J Periodontol • February 2007 Csiszar, Wiebe, Larjava, Hakkinen
311

suggests that the cells in the interdental papilla are inan activated state similar to wound healing.
Interdental papilla is exposed easily to inflamma-tion-causing agents from the oral cavity. Care wastaken in the present study to evaluate only those areasthat did not show inflammatory cell infiltrate in the vi-cinity. Inflammation upregulates expression of CK19in epithelial cells.42 Because CK19 was expressed inall regions of the interdental papilla epithelium, wecannot completely rule out the possibility that the ef-fects of mild inflammation in some areas of the tissuemay have affected other areas further away from theinflammatory infiltrate. However, it is unlikely that theexpression of avb6 integrin in the interdental papilla isa result of inflammation, but rather reflects a specificphenotype of this epithelium. This is supported bythe finding that expression of avb6 integrin still is up-regulated in gingival wounds when inflammation is re-duced.22 Furthermore, periodontal pocket epitheliumdoes not express avb6 integrin.43
In the marginal gingiva, collagen was organizedinto the typical basket-weave pattern of relatively
thick collagen fiber bundles. In contrast, in the inter-dental papilla, collagen organization was less homog-enous and included localized parallel thin collagenfiber bundles that were stained strongly with anti-bodies against decorin, biglycan, fibromodulin, andlumican. These small leucine-rich proteoglycans col-laborate in regulating collagen fibrillogenesis in peri-odontal tissues, and their major function seems to beto limit the lateral growth of collagen fibrils.27,44,45
Thus, abundant expression of these molecules mayresult in formation of the thin collagen fibers in theinterdental papilla. Apart from the localized collagenfibril bundles, the relative staining for lumican wassignificantly weaker in the extracellular matrix of theinterdental papilla than in the marginal gingiva. Inaddition, the staining intensity of biglycan in bloodvessels was higher in the interdental papilla than inthe marginal gingiva. Thus, expression of the smallleucine-rich proteoglycans is regulated differently inthe interdental papilla and marginal gingiva. In addi-tion to regulating collagen fibrillogenesis, at leastdecorin, biglycan, and fibromodulin bind TGF-b, ren-dering it inactive.46 These proteoglycans also canserve as signaling molecules that regulate cell func-tions.44,45 Thus, differential expression of the smallleucine-rich proteoglycans in the interdental papillamay result in cellular functions different from the mar-ginal gingiva. We showed previously that the proformof decorin (prodecorin) that is secreted by cells local-izes to fibroblasts and blood vessels in marginal gin-giva.26 Lack of decorin staining in fibroblasts andblood vessels can be explained by the fact that the an-tibody used in the study recognizes only the mature,extracellular matrix–associated form of the molecule.
We previously analyzed the expression avb6 in-tegrin and tenascin-C,22 type I procollagen, decorin,biglycan, fibromodulin, and lumican,21,26 and EDAand EDB fibronectin25 in human attached palatal gin-giva where the expression and localization of thesemolecules are similar to the marginal gingiva, asshown in the present study. The only difference isstrong immunostaining intensity of fibromodulinin the basal epithelial cells of the attached gingivacompared to marginal gingiva. Furthermore, our un-published data showed similar expression and locali-zation of CK19, TGF-b, CTGF, and SOS-1 in humanattached palatal gingiva and marginal gingiva. Thus,the findings in the present study show that the molec-ular composition in the interdental papilla, and hence,the cell phenotype, differ from the marginal gingivaand attached palatal gingiva, suggesting that humangingiva contains functionally different anatomicalareas. Notably, the expressions of molecules thatare involved in wound healing are upregulated inthe interdental papilla. It is possible that this specificphenotype is inherent to the cells in this anatomical
Table 4.
Summary of Expression of the StudiedMolecules in the Interdental Papilla Relativeto the Marginal Gingiva
Relative Expression
Antibody
Oral
Epithelium ECM FBL BV
CK19 [ 0 0 0
avb6 integrin [ 0 0 0
Procollagen type I 0 [ [ –
Decorin 0 – 0 0
Biglycan [ [ – [
Fibromodulin [ – – –
Lumican [ Y – –
Fibronectin-EDA 0 [ [ –
Fibronectin-EDB 0 [ [ [
Tenascin-C 0 [ 0 –
TGF-b – 0 [ [
CTGF [ 0 [ –
SOS-1 [ 0 [ [
At least 10 sections from the marginal gingiva and interdental papillasamples from five subjects were used for the analyses of expression of eachmolecule. Arrows show changes that are statistically significant.ECM = extracellular matrix; FBL = fibroblasts; BV = blood vessels. [ =upregulated; Y = downregulated; – = no change; 0 = not expressed.
Molecular Composition of Interdental Papilla Volume 78 • Number 2
312

location or that the cells are activated because of thepresence of mild localized inflammation, even in clin-ically healthy tissue. Drug-induced or hereditary gin-gival overgrowth usually starts at the interdentalpapilla.28 It is tempting to speculate that the specificactivated phenotype of the cells in the interdental pa-pilla may predispose the tissue to gingival overgrowthin the presence of additional activating factors, i.e.,certain drugs or hereditary gene mutations. Anatom-ical factors, including the extent of bone loss, likelyplay an important role in the limited regeneration abil-ity of the interdental papilla.3 However, the lack of re-generation also may depend on the inability of the cellspresent in the surrounding marginal gingiva to reca-pitulate the specific phenotypic properties of the inter-dental papilla cells. Thus, surgical techniques that canpreserve the existing papilla as a source of specificcells may be needed to be able to regenerate the inter-dental papilla. As more detailed information becomesavailable about the cellular and molecular character-istics of the interdental papilla, targeted moleculartherapies also can be developed that promote growthand differentiation of appropriate cells. Our findingsalso emphasize the importance of recording the exactgingival tissue biopsy location for molecular studies tobe able to make meaningful conclusions. Based onour results, the especially strong expression of CK19,avb6 integrin, and fibronectin-EDB can be used asmolecular markers for interdental papilla to differen-tiate it from the marginal gingiva.
ACKNOWLEDGMENTS
The authors thank Dr. Larry Fisher, National Institutes ofHealth/National Institute of Dental and Craniofacial Re-search, Bethesda, MD; Dr. Tom Krusius, Finnish RedCross Blood Transfusion Service, Helsinki, Finland; Dr.Anna Plaas, Department of Internal Medicine, Universityof South Florida, Tampa, FL; Dr. P.J. Roughley, ShrinersHospital for Children, McGill University, Montreal, Que-bec City; Dr. Dean Sheppard, Lung Biology Center, Uni-versity of California, San Francisco, San Francisco, CA;andDr.LucianoZardi,LaboratoryofCellBiology, IstitutoNazionale per la Ricerca sul Cancro, Genoa, Italy, forproviding antibodies for the study. We also thank Mr.Cristian Sperantia, Faculty of Dentistry, University ofBritish Columbia, Vancouver, BC, for expert technicalassistance.Thestudywassupportedby theCanadian In-stitutes for Health Research, Ottawa, Ontario.
REFERENCES1. Berkovitz BKB, Holland GR, Moxham BJ. Oral Anat-
omy, Histology and Embryology. New York: MosbyInternational Limited; 2002:1-7.
2. Lindhe J, Karring T, Lang NP. Clinical Periodontologyand Implant Dentistry, 4th ed. Copenhagen: BlackwellPublishing; 2003:1-49.
3. Tarnow DP, Magner AW, Fletcher P. The effect of thedistance from the contact point to the crest of bone onthe presence or absence of the interproximal dentalpapilla. J Periodontol 1992;63:995-996.
4. Weber HP, Cochran DL. The soft tissue response toosseointegrated dental implants. J Prosthet Dent 1998;79:79-89.
5. Choquet V, Hermans M, Adriaenssens P, DaelemansP, Tarnow DP, Malevez C. Clinical and radiographicevaluation of the papilla level adjacent to single-toothdental implants. A retrospective study in the maxillaryanterior region. J Periodontol 2001;72:1364-1371.
6. Shimono M, Ishikawa T, Enokiya Y, et al. Biologicalcharacteristics of the junctional epithelium. J ElectronMicrosc (Tokyo) 2003;52:627-639.
7. Tarnow D, Elian N, Fletcher P, et al. Vertical distancefrom the crest of bone to the height of the interprox-imal papilla between adjacent implants. J Periodontol2003;74:1785-1788.
8. Schropp L, Isidor F, Kostopoulos L, Wenzel A. Inter-proximal papilla levels following early versus delayedplacement of single-tooth implants: A controlled clin-ical trial. Int J Oral Maxillofac Implants 2005;20:753-761.
9. Zetu L, Wang HL. Management of inter-dental/inter-implant papilla. J Clin Periodontol 2005;32:831-839.
10. Dale BA, Salonen J, Jones AH. New approaches andconcepts in the study of differentiation of oral epithe-lia. Crit Rev Oral Biol Med 1990;1:167-190.
11. Winning TA, Townsend GC. Oral mucosal embryologyand histology. Clin Dermatol 2000;18:499-511.
12. Schweizer J, Winter H, Hill MW, Mackenzie IC. Thekeratin polypeptide patterns in heterotypically recom-bined epithelia of skin and mucosa of adult mouse.Differentiation 1984;26:144-153.
13. Lim IJ, Phan TT, Bay BH, et al. Fibroblasts coculturedwith keloid keratinocytes: Normal fibroblasts secretecollagen in a keloid-like manner. Am J Physiol CellPhysiol 2002;283:C212-C222.
14. Phipps RP, Borrello MA, Blieden TM. Fibroblast heter-ogeneity in the periodontium and other tissues. J Peri-odontal Res 1997;32:159-165.
15. Huang X, Wu J, Spong S, Sheppard D. The integrinalphavbeta6 is critical for keratinocyte migration onboth its known ligand, fibronectin, and on vitronectin.J Cell Sci 1998;111:2189-2195.
16. Krusius T, Ruoslahti E. Primary structure of an extra-cellular matrix proteoglycan core protein deducedfrom cloned cDNA. Proc Natl Acad Sci USA 1986;83:7683-7687.
17. Fisher LW, Lindner W, Young MF, Termine JD. Syn-thetic peptide antisera: Their production and use in thecloning of matrix proteins. Connect Tissue Res 1989;21:43-48.
18. Plaas AH, Wong-Palms S. Biosynthetic mechanismsfor the addition of polylactosamine to chondrocytefibromodulin. J Biol Chem 1993;268:26634-26644.
19. Grover J, Chen XN, Korenberg JR, Roughley PJ. Thehuman lumican gene. Organization, chromosomal lo-cation, and expression in articular cartilage. J Biol Chem1995;270:21942-21949.
20. Kaczmarek J, Castellani P, Nicolo G, Spina B, AllemanniG, Zardi L. Distribution of oncofetal fibronectin iso-forms in normal, hyperplastic and neoplastic humanbreast tissues. Int J Cancer 1994;59:11-16.
21. Hakkinen L, Oksala O, Salo T, Rahemtulla F, LarjavaH. Immunohistochemical localization of proteoglycans
J Periodontol • February 2007 Csiszar, Wiebe, Larjava, Hakkinen
313

in human periodontium. J Histochem Cytochem 1993;41:1689-1699.
22. Hakkinen L, Hildebrand HC, Berndt A, Kosmehl H,Larjava H. Immunolocalization of tenascin-C, alpha9integrin subunit, and alphavbeta6 integrin during woundhealing in human oral mucosa. J Histochem Cytochem2000;48:985-998.
23. Hakkinen L, Strassburger S, Kahari VM, et al. A role fordecorin in the structural organization of periodontalligament. Lab Invest 2000;80:1869-1880.
24. Hakkinen L, Uitto VJ, Larjava H. Cell biology of gingivalwound healing. Periodontol 2000 2000;24:127-152.
25. Larjava H, Koivisto L, Hakkinen L. Keratinocyte in-teractions with fibronectin during wound healing. In:Heino J, Kahari V-M, eds. Cell Invasion. Georgetown,TX: Landes Biosciences; 2002:42-64.
26. Alimohamad H, Habijanac T, Larjava H, Hakkinen L.Colocalization of the collagen-binding proteoglycansdecorin, biglycan, fibromodulin and lumican with dif-ferent cells in human gingiva. J Periodontal Res 2005;40:73-86.
27. Matheson S, Larjava H, Hakkinen L. Distinctive local-ization and function for lumican, fibromodulin anddecorin to regulate collagen fibril organization in peri-odontal tissues. J Periodontal Res 2005;40:312-324.
28. Hakkinen L, Csiszar A. Hereditary gingival fibromato-sis: Characteristics and novel putative pathogenicmechanisms. J Dent Res 2007;86:25-34.
29. Hart TC, Zhang Y, Gorry MC, et al. A mutation in theSOS1 gene causes hereditary gingival fibromatosistype 1. Am J Hum Genet 2002;70:943-954.
30. Bianco P, Fisher LW, Young MF, Termine JD, RobeyPG. Expression and localization of the two smallproteoglycans biglycan and decorin in developinghuman skeletal and non-skeletal tissues. J HistochemCytochem 1990;38:1549-1563.
31. Saika S, Shiraishi A, Liu CY, et al. Role of lumican inthe corneal epithelium during wound healing. J BiolChem 2000;275:2607-2612.
32. Schaefer L, Grone HJ, Raslik I, et al. Small proteogly-cans of normal adult human kidney: Distinct expres-sion patterns of decorin, biglycan, fibromodulin, andlumican. Kidney Int 2000;58:1557-1568.
33. Haapasalmi K, Zhang K, Tonnesen M, et al. Keratino-cytes in human wounds express alphavbeta6 integrin.J Invest Dermatol 1996;106:42-48.
34. Sheppard D. Integrin-mediated activation of latenttransforming growth factor beta. Cancer MetastasisRev 2005;24:395-402.
35. Leask A, Abraham DJ. TGF-beta signaling and thefibrotic response. FASEB J 2004;18:816-827.
36. Leivonen SK, Hakkinen L, Liu D, Kahari VM. Smad3and extracellular signal-regulated kinase 1/2 coordi-
nately mediate transforming growth factor-beta-in-duced expression of connective tissue growth factorin human fibroblasts. J Invest Dermatol 2005;124:1162-1169.
37. Phanish MK, Wahab NA, Hendry BM, DockrellME. TGF-beta1-induced connective tissue growthfactor (CCN2) expression in human renal proximaltubule epithelial cells requires Ras/MEK/ERK andSmad signalling. Nephron Exp Nephrol 2005;100:e156-e165.
38. Frazier K, Williams S, Kothapalli D, Klapper H,Grotendorst GR. Stimulation of fibroblast cell growth,matrix production, and granulation tissue formationby connective tissue growth factor. J Invest Dermatol1996;107:404-411.
39. Pearson CA, Pearson D, Shibahara S, Hofsteenge J,Chiquet-Ehrismann R. Tenascin: cDNA cloningand induction by TGF-beta. EMBO J 1988;7:2977-2982.
40. Linnala A, Kinnula V, Laitinen LA, Lehto VP, Virtanen I.Transforming growth factor-beta regulates the expres-sion of fibronectin and tenascin in BEAS 2B humanbronchial epithelial cells. Am J Respir Cell Mol Biol1995;13:578-585.
41. Nimnual A, Bar-Sagi D. The two hats of SOS. Sci STKE2002;2002:PE36.
42. Ouhayoun JP, Goffaux JC, Sawaf MH, Shabana AH,Collin C, Forest N. Changes in cytokeratin expressionin gingiva during inflammation. J Periodontal Res 1990;25:283-292.
43. Haapasalmi K, Makela M, Oksala O, et al. Expressionof epithelial adhesion proteins and integrins in chronicinflammation. Am J Pathol 1995;147:193-206.
44. Hocking AM, Shinomura T, McQuillan DJ. Leucine-rich repeat glycoproteins of the extracellular matrix.Matrix Biol 1998;17:1-19.
45. Ameye L, Young MF. Mice deficient in small leucine-rich proteoglycans: Novel in vivo models for osteopo-rosis, osteoarthritis, Ehlers-Danlos syndrome, musculardystrophy, and corneal diseases. Glycobiology 2002;12:107R-116R.
46. Hildebrand A, Romaris M, Rasmussen LM, et al. Inter-action of the small interstitial proteoglycans biglycan,decorin and fibromodulin with transforming growthfactor beta. Biochem J 1994;302:527-534.
Correspondence: Dr. Lari Hakkinen, Faculty of Dentistry,Department of Oral Biological and Medical Sciences,Laboratory of Periodontal Biology, University of BritishColumbia, 2199 Wesbrook Mall, Vancouver, BC V6T 1Z3.Fax: 604/822-3562; email: [email protected].
Accepted for publication September 3, 2006.
Molecular Composition of Interdental Papilla Volume 78 • Number 2
314