directphysiological evidencefor …. nati. acad. sci. usa vol. 88, pp. 9136-9140, october 1991...
TRANSCRIPT

Proc. Nati. Acad. Sci. USAVol. 88, pp. 9136-9140, October 1991Neurobiology
Direct physiological evidence for scene segmentation bytemporal coding
(cat/visual cortex/correlation analysis/synchronization/cell assembly)
ANDREAS K. ENGEL, PETER KONIG, AND WOLF SINGERMax-Planck-Institut fur Hirnforschung, Deutschordenstrasse 46, 6000 Frankfurt 71, Federal Republic of Germany
Communicated by G. M. Edelman, July 9, 1991 (received for review May 6, 1991)
ABSTRACT Theoretical studies have suggested that scenesegmentation may be accomplished by a temporal coding mech-anism using synchronization of neuronal responses. Here wereport a direct experimental test of this hypothesis. Neuronalresponses were recorded simultaneously from two to four siteswith overlapping receptive fields in cat visual cortex. Correlationanalysis revealed that all cells synchronized their responsesirrespective of their orientation preference when they wereactivated by a single light bar. However, when stimulated withtwo superimposed light bars of different orientations, the samecells segregated into distinct assemblies according to their ori-entation preferences. Within each of these assemblies responseswere synchronized, but correlation was absent between the twoassemblies. These results are compatible with the hypothesis thatresponses to individual objects in a scene are distuished bysynchrony, whereas responses to different objects show notemporal correlation, thus allowing for the segmentation ofsuperimposed stimuli. We conclude that stimulus-specific syn-chronization of spatially distributed neuronal responses mayprovide a physiological mechanism for scene segmentation.
Since the early work of the Gestalt psychologists (1, 2)psychophysical research has established that scene segmen-tation and perceptual grouping represent important aspects ofearly vision. It is generally agreed (3) that, as a prerequisitefor object recognition, a visual scene has to be parsed, andcoherence between elementary features has to be evaluatedto permit the discrimination of different objects within thescene. The physiological correlate of this segmentation pro-cess has not yet been identified with certainty. According toa now-classic proposal, segmentation should be accom-plished by single neurons dedicated to certain feature con-stellations (4). However, such a model is not supported byexperimental evidence and leads to a "combinatorial explo-sion," because every new pattern requires an additionaldedicated unit. Therefore, alternative mechanisms have beensuggested which imply that feature constellations are repre-sented by cell assemblies rather than by the activity of singlecells (5-10). A presumed advantage of assembly coding isthat the combinatorial exhaustion of the system can beavoided, because one particular cell may participate in dif-ferent assemblies (9). In the classical model introduced byHebb (5), assemblies are defined by amplitude coding-i.e.,the concurrent elevation of the average firing frequency ofthe participating cells for several hundred milliseconds. How-ever, the Hebb mechanism does not permit the coactivationoftwo spatially superimposed assemblies in the same corticalregion (9, 11). Because all cells raise their average firingfrequencies, information about the partition into distinctassemblies is lost. This "superposition catastrophe" willresult in false conjunctions of features and render object
recognition impossible (9, 11). Therefore, it has been sug-gested that precise synchronization of neuronal discharges inthe millisecond range should be used to define functionalassemblies and to achieve scene segmentation (7-9). Accord-ing to these proposals, perceptual coherence within a seg-ment of a scene should be expressed by response synchro-nization of feature-detecting neurons. In contrast, cells re-sponding to different objects in a scene should beuncorrelated. This mechanism of assembly formation will inthe following be referred to as temporal coding. As shown bysimulation studies (11, 12), such a mechanism permits thecoexistence of several assemblies in the same network and,thus, seems capable of solving the superposition problem.
Increasing physiological evidence suggests the presence ofa temporal-coding mechanism in cortical networks that usesneuronal responses with a periodic temporal structure. Asshown in recent studies by us and others (13-17), neurons instriate and extrastriate areas of cat visual cortex tend todischarge in an oscillatory manner. Because adjacent cellswithin a single orientation column have a strong tendency tooscillate synchronously (15, 16), we have adopted the termneuronal group for the designation of such coherently activecell clusters (6, 10). Subsequent studies (14, 17-19) haveprovided evidence that spatially separate groups within onevisual area, as well as cell groups located in different areas,can synchronize their oscillatory responses. In addition, wehave recently observed response synchronization across thecerebral hemispheres (20). These results suggest that thephase-relationship of neuronal oscillations may be used todefine cortical assemblies-i.e., families of spatially distrib-uted neuronal groups that represent visual objects. In thismanner, temporal coding may be used to achieve perceptualgrouping and scene segmentation (17-23). If this holds true,the following predictions should be confirmed experimen-tally: The synchronization between spatially separate cellgroups should be stimulus dependent. Neuronal groupsshould oscillate in synchrony when they respond to the sameperceptual object, whereas cell groups responding to twounrelated stimuli should oscillate in an uncorrelated manner.In particular, it should be demonstrated that this mechanismis capable of segregating stimuli that are superimposed in thevisual field. In the present study, we used a simple experi-mental paradigm to test these predictions. Essentially, theexperiment involves simultaneous recording from severalcell groups with overlapping receptive fields. When activatedwith a single light bar, these groups will fire synchronously,even when they differ in their preferred orientation (18, 19).We have now tested whether the synchronization is affectedby using two superimposed light bars as stimuli. These"conflicting stimuli" constitute a simple visual scene inwhich two figures have to be discriminated. A preliminaryreport of the results (21) has appeared.
MATERIALS AND METHODSThe data were collected from eight anesthetized and para-lyzed adult cats. The animals were prepared and maintained
9136
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 88 (1991) 9137
as described in detail elsewhere (18, 19). Multiunit activitywas recorded from area 17 of cat visual cortex by using anarray of four or five platinum-iridium electrodes with aspacing of 400-500 ,um. Thus, we were able to monitor theactivity of coherently active cell groups within differentorientation columns (10, 15). The array was centered on therepresentation of the area centralis and was oriented alongthe anterior-posterior axis. Due to this electrode arrange-ment, the groups of cells recorded by different electrodesusually had overlapping receptive fields but differed in theirpreferred orientation. At the onset of a recording session,size and location of the receptive field, preferred orientation,and width of orientation tuning were assessed for each groupof cells with light stimuli projected manually onto a tangentscreen in front of the cat. Subsequently, the receptive fieldsof the two eyes were aligned with a combination of prisms.For correlation measurements, light bar stimuli were gener-ated with computer-controlled optical benches.The cases included in this study were selected in the
following way: Simultaneous recordings from four sites (qua-druplets) were included when the respective orientationpreferences matched pairwise but differed by 450 or morebetween these pairs. Recordings from three sites (triplets)were accepted when two sites had similar orientation pref-erences and when these differed from the third site by at least45°. Finally, we also evaluated simultaneous recordings fromtwo sites (doublets) when the respective orientation prefer-ences differed by at least 45°. Cases were only included if cellgroups with different orientation preference had sufficientlybroad tuning to be coactivated with a single stimulus ofintermediate orientation.The cells were activated with either single moving light
bars of various orientations or two bars moving simultane-ously across the receptive fields (conflicting stimuli). In thelatter case, orientations of the two light bars were adjusted tomatch orientation preferences of the cell groups (see Figs.1-3). The length ofthe bar stimuli always corresponded to theextension of the receptive fields. The bars were movedorthogonal to their orientation, and their velocity wasadjusted to elicit strong and sustained responses. For eachtrial, light bars were moved forward and backward across thereceptive fields within 10 sec, and 10 trials to the samestimulus were run in succession. For both single as well asconflicting stimuli, several blocks of 10 trials were recordedin an alternating manner to control for the consistency of theeffects observed.To record multiunit activity, electrode signals were ampli-
fied, bandpass-filtered between 1 and 3 kHz, and then fedthrough a Schmitt trigger, the threshold of which was set toat least twice the noise level. The resulting pulse train wasdigitized with a time resolution of 1 ms. As describedpreviously, peri-stimulus-time histograms as well as auto-and cross-correlation functions were computed for the re-corded spike trains (18, 19). As a control, we computedshift-predictors of all correlograms (19). The oscillatory mod-ulation of neuronal responses is reflected by a periodicauto-correlogram. A periodic cross-correlogram indicatesthe synchronization of two oscillatory responses. In bothcases, the significance of the modulation can be estimated byfitting a damped sine wave (Gabor) function to the correlo-grams (19) (see Figs. 1-3). The strength ofthe modulation canthen be quantified by using the amplitude, decay, and offsetof the respective Gabor function. To obtain a measure that isindependent of the absolute response strength, we deter-mined the relative modulation amplitude by computing theratio of the Gabor function amplitude over its offset (19).Cross-correlograms were considered as reflecting a signifi-cant synchronization when the Gabor function amplitude wassignificantly different from zero (at the 5% level), when therelative modulation amplitude exceeded a value of 0.1, and
when the Gabor function displayed at least three distinctpeaks (19) (see Figs. 1 and 2). The same criteria were appliedto auto-correlograms to quantify the periodic modulation ofthe responses. It should be noted that cross-correlogramswith only one center peak, which reflect the interaction oftwo nonoscillatory responses, can be described by a Gaborfunction with a rapid decay. Therefore, we used the samequantification procedure in cases where we studied theinfluence of conflicting stimuli on the interaction of nonos-cillating cells (see Fig. 3).
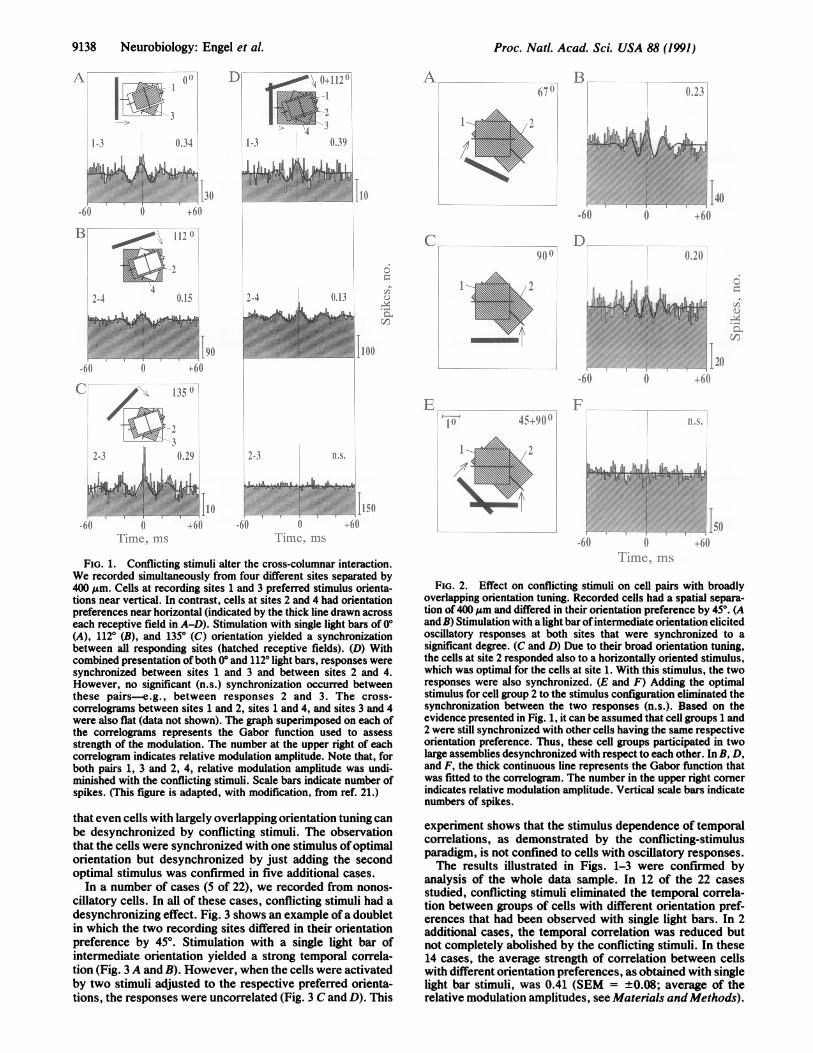
RESULTSWe studied the effect of single and conflicting stimuli on thecross-columnar interaction in 22 cases, corresponding to 4quadruplets, 7 triplets, and 11 doublets. As described previ-ously, cells in different orientation columns tend to synchro-nize their oscillatory responses when their receptive fieldsare overlapping and when they are activated with a singlelight bar (19). We have now tested whether conflicting stimulieliminate the temporal correlation between cells of differentorientation preference. The constellation of quadruplets andtriplets allowed us to investigate whether within pairs ofrecordings with matching orientation preference the corre-lation is maintained when conflicting stimuli are applied. Thisresult should be expected because in such pairs both cellgroups respond to the same component of the stimulusconfiguration.One of the quadruplets is illustrated in Fig. 1. We recorded
from four sites with overlapping receptive fields and roughlyalternating orientation preferences. Stimulation with singlelight bars of different orientations yielded a synchronizationof oscillatory responses between all sites activated by therespective orientation. Thus, cells at sites 1 and 3 respondedsynchronously to a vertical (0°) light bar (Fig. 1A), cells atsites 2 and 4 responded synchronously to a light bar of 1120orientation (Fig. 1B), and cells at sites 2 and 3 respondedsynchronously to an intermediate stimulus (Fig. 1C). Whena 00 and a 1120 light bar were presented together to form a pairof conflicting stimuli, the cells at all four recording sites weresimultaneously activated (Fig. 1D). In this case, responsesynchronization was still seen between sites 1 and 3 andbetween sites 2 and 4 and was as good as with single barstimuli. However, no correlation occurred between the twosynchronously active pairs. For instance, responses wereuncorrelated between sites 2 and 3 (Fig. 1D), although at bothsites the responses were even stronger than with a singlestimulus of intermediate orientation. Thus, the conflictingstimuli segregate the cells in two distinct assemblies accord-ing to their orientation preferences. Cells responding to thesame stimulus synchronize their responses, but they do notcorrelate with cells preferring the other stimulus. Impor-tantly, this result cannot be explained by assuming selectiveanatomical connections between cells with similar orienta-tion preferences (24) because with single stimuli a temporalcorrelation can be observed between cells preferring differentorientations (Fig. 1C).
Similar observations were made in triplets and doublets,which represent fragments of the quadruplet constellation. Adoublet of oscillatory responses is illustrated in Fig. 2. As inthe previous example, synchronized responses were evokedwith a single stimulus of orientation intermediate betweenthose preferred by the two groups of cells (Fig. 2 A and B).Because the cells exhibited a rather broad orientation tuning,they could also be activated simultaneously with a light baroriented optimally for one of the recording sites. In this case,the responses were also synchronized (Fig. 2 C and D).However, combined presentation of two light bars withorientations optimal for the two cell groups still eliminatedthe synchronization (Fig. 2 E and F). This case demonstrates
Neurobiology: Engel et al.
Proc. Natl. Acad. Sci. USA 88 (1991)
A
I-3
lit
0.34
A
'-3 01.39
I ............................
-60>i -60l
C
2.-4 ..15 2-4
9(1
611
Dt1.2(1
1.13
he*
f-
r7
I11(111211-6(1
E
2-3 '9
1 1.
2.3
-60l -61)fill} -6o 1) -601iiic. ls Ii i . IIIS
FIG. 1. Conflicting stimuli alter the cross-columnar interaction.We recorded simultaneously from four different sites separated by400 gum. Cells at recording sites 1 and 3 preferred stimulus orienta-tions near vertical. In contrast, cells at sites 2 and 4 had orientationpreferences near horizontal (indicated by the thick line drawn acrosseach receptive field in A-D). Stimulation with single light bars of 00(A), 1120 (B), and 1350 (C) orientation yielded a synchronizationbetween all responding sites (hatched receptive fields). (D) Withcombined presentation ofboth 00 and 1120 light bars, responses weresynchronized between sites 1 and 3 and between sites 2 and 4.However, no significant (n.s.) synchronization occurred betweenthese pairs-e.g., between responses 2 and 3. The cross-correlograms between sites 1 and 2, sites 1 and 4, and sites 3 and 4were also flat (data not shown). The graph superimposed on each ofthe correlograms represents the Gabor function used to assessstrength of the modulation. The number at the upper right of eachcorrelogram indicates relative modulation amplitude. Note that, forboth pairs 1, 3 and 2, 4, relative modulation amplitude was undi-minished with the conflicting stimuli. Scale bars indicate number ofspikes. (This figure is adapted, with modification, from ref. 21.)
that even cells with largely overlapping orientation tuning canbe desynchronized by conflicting stimuli. The observationthat the cells were synchronized with one stimulus of optimalorientation but desynchronized by just adding the secondoptimal stimulus was confirmed in five additional cases.
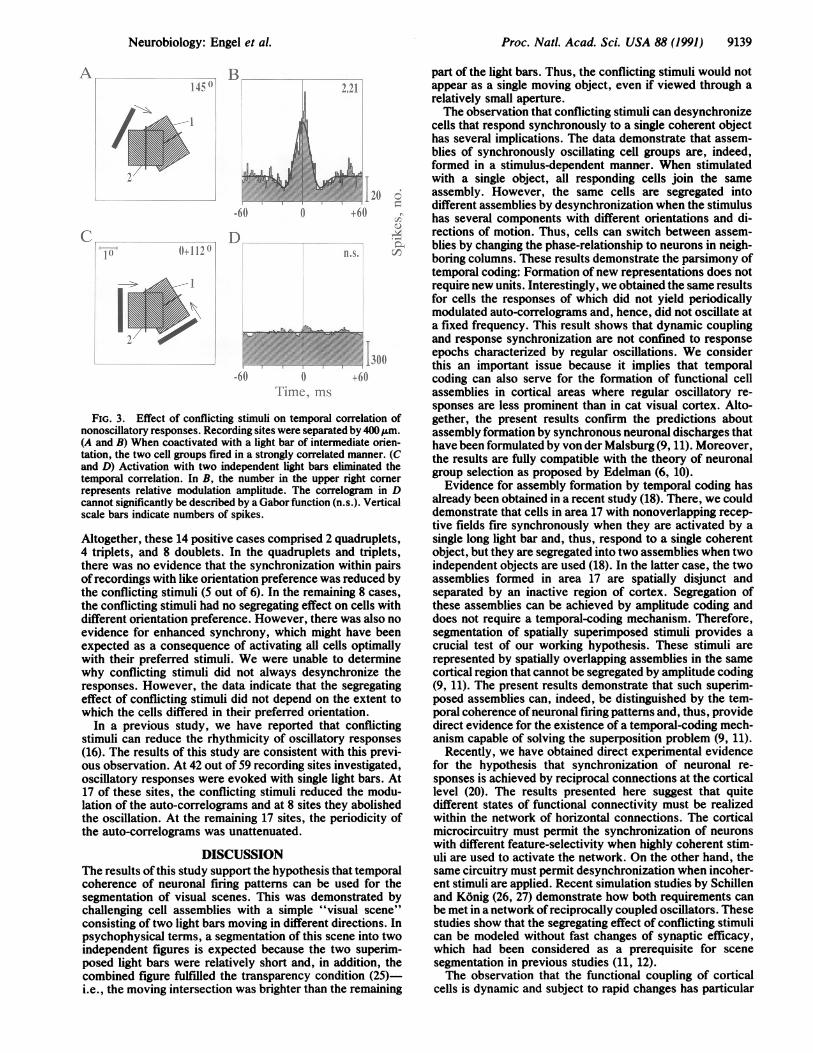
In a number of cases (5 of 22), we recorded from nonos-cillatory cells. In all of these cases, conflicting stimuli had adesynchronizing effect. Fig. 3 shows an example ofa doubletin which the two recording sites differed in their orientationpreference by 45°. Stimulation with a single light bar ofintermediate orientation yielded a strong temporal correla-tion (Fig. 3A and B). However, when the cells were activatedby two stimuli adjusted to the respective preferred orienta-tions, the responses were uncorrelated (Fig. 3 C and D). This
611 I-^ ^ .5a __i 61)^^
,.h.. l1
.60~~~~~~-60-
FIG. 2. Effect on conflicting stimuli on cell pairs with broadlyoverlapping orientation tuning. Recorded cells had a spatial separa-tion of 400 Am and differed in their orientation preference by 45°. (Aand B) Stimulation with a light bar ofintermediate orientation elicitedoscillatory responses at both sites that were synchronized to asignificant degree. (C and D) Due to their broad orientation tuning,the cells at site 2 responded also to a horizontally oriented stimulus,which was optimal for the cells at site 1. With this stimulus, the tworesponses were also synchronized. (E and F) Adding the optimalstimulus for cell group 2 to the stimulus configuration eliminated thesynchronization between the two responses (n.s.). Based on theevidence presented in Fig. 1, it can be assumed that cell groups 1 and2 were still synchronized with other cells having the same respectiveorientation preference. Thus, these cell groups participated in twolarge assemblies desynchronized with respect to each other. In B, D,and F, the thick continuous line represents the Gabor function thatwas fitted to the correlogram. The number in the upper right cornerindicates relative modulation amplitude. Vertical scale bars indicatenumbers of spikes.
experiment shows that the stimulus dependence of temporalcorrelations, as demonstrated by the conflicting-stimulusparadigm, is not confined to cells with oscillatory responses.The results illustrated in Figs. 1-3 were confimed by
analysis of the whole data sample. In 12 of the 22 casesstudied, conflicting stimuli eliminated the temporal correla-tion between groups of cells with different orientation pref-erences that had been observed with single light bars. In 2additional cases, the temporal correlation was reduced butnot completely abolished by the conflicting stimuli. In these14 cases, the average strength of correlation between cellswith different orientation preferences, as obtained with singlelight bar stimuli, was 0.41 (SEM = +0.08; average of therelative modulation amplitudes, see Materials andMethods).
9138 Neurobiology: Engel et A
D 0-112 OI
II
II4
11. S.
B ]I'll,11000-00,2
4
Proc. Natl. Acad. Sci. USA 88 (1991) 9139
A. B14 5(
.601
C(1-i 12}
l)
2 do
.............
Ti1 lII C IIs
FIG. 3. Effect of conflicting stimuli on temporal (nonoscillatory responses. Recording sites were separatf(A and B) When coactivated with a light bar of interntation, the two cell groups fired in a strongly correlateand D) Activation with two independent light bars etemporal correlation. In B, the number in the upperrepresents relative modulation amplitude. The corrncannot significantly be described by a Gabor function (Iscale bars indicate numbers of spikes.
Altogether, these 14 positive cases comprised 2 c4 triplets, and 8 doublets. In the quadrupletsthere was no evidence that the synchronizationofrecordings with like orientation preference wawthe conflicting stimuli (5 out of 6). In the remairthe conflicting stimuli had no segregating effectdifferent orientation preference. However, thereevidence for enhanced synchrony, which mighexpected as a consequence of activating all celwith their preferred stimuli. We were unable t
why conflicting stimuli did not always desyncresponses. However, the data indicate that theeffect of conflicting stimuli did not depend on t]which the cells differed in their preferred orient
In a previous study, we have reported thatstimuli can reduce the rhythmicity of oscillator(16). The results of this study are consistent witous observation. At 42 out of 59 recording sites ioscillatory responses were evoked with single li17 of these sites, the conflicting stimuli reduce(lation of the auto-correlograms and at 8 sites thtthe oscillation. At the remaining 17 sites, the pathe auto-correlograms was unattenuated.
DISCUSSIONThe results of this study support the hypothesis tlcoherence of neuronal firing patterns can be i
segmentation of visual scenes. This was demcchallenging cell assemblies with a simple "vi:consisting of two light bars moving in different dpsychophysical terms, a segmentation of this scaindependent figures is expected because the tNposed light bars were relatively short and, in acombined figure fulfilled the transparency coni.e., the moving intersection was brighter than ti
part of the light bars. Thus, the conflicting stimuli would not2.21 appear as a single moving object, even if viewed through a
relatively small aperture.The observation that conflicting stimuli can desynchronize
cells that respond synchronously to a single coherent objecthas several implications. The data demonstrate that assem-blies of synchronously oscillating cell groups are, indeed,formed in a stimulus-dependent manner. When stimulatedwith a single object, all responding cells join the same
10/)assembly. However, the same cells are segregated intodifferent assemblies by desynchronization when the stimulus
I6(J has several components with different orientations and di-/ rections of motion. Thus, cells can switch between assem-~- blies by changing the phase-relationship to neurons in neigh-
'i'> . boring columns. These results demonstrate the parsimony oftemporal coding: Formation ofnew representations does notrequire new units. Interestingly, we obtained the same resultsfor cells the responses of which did not yield periodicallymodulated auto-correlograms and, hence, did not oscillate ata fixed frequency. This result shows that dynamic coupling
0 72:'11 and response synchronization are not confined to responseepochs characterized by regular oscillations. We consider300- -----this an important issue because it implies that temporal
-6( coding can also serve for the formation of functional cellassemblies in cortical areas where regular oscillatory re-sponses are less prominent than in cat visual cortex. Alto-
correlation of gether, the present results confirm the predictions abouted by 400 jim. assembly formation by synchronous neuronal discharges thatnediate orien- have been formulated by von der Malsburg (9, 11). Moreover,4d manner. (C the results are fully compatible with the theory of neuronalliminated the group selection as proposed by Edelman (6, 10).
elogram in D Evidence for assembly formation by temporal coding hasn.s.). Vertical already been obtained in a recent study (18). There, we could
demonstrate that cells in area 17 with nonoverlapping recep-tive fields fire synchronously when they are activated by a
quadruplets, single long light bar and, thus, respond to a single coherentand triplets, object, but they are segregated into two assemblies when twowithin pairs independent objects are used (18). In the latter case, the twosreduced by assemblies formed in area 17 are spatially disjunct andfing 8 cases, separated by an inactive region of cortex. Segregation ofan cells with these assemblies can be achieved by amplitude coding andwas also no does not require a temporal-coding mechanism. Therefore,t have been segmentation of spatially superimposed stimuli provides aIs optimally crucial test of our working hypothesis. These stimuli areo determine represented by spatially overlapping assemblies in the same:hronize the cortical region that cannot be segregated by amplitude codingsegregating (9, 11). The present results demonstrate that such superim-
;he extent to posed assemblies can, indeed, be distinguished by the tem-tation. poral coherence ofneuronal firing patterns and, thus, providet conflicting direct evidence for the existence of a temporal-coding mech--y responses anism capable of solving the superposition problem (9, 11).-h this previ- Recently, we have obtained direct experimental evidenceinvestigated, for the hypothesis that synchronization of neuronal re-ight bars. At sponses is achieved by reciprocal connections at the corticald the modu- level (20). The results presented here suggest that quiteey abolished different states of functional connectivity must be realizedPeriodicity of within the network of horizontal connections. The cortical
microcircuitry must permit the synchronization of neuronswith different feature-selectivity when highly coherent stim-uli are used to activate the network. On the other hand, the
hat temporal same circuitry must permit desynchronization when incoher-used for the ent stimuli are applied. Recent simulation studies by SchillenCnstrated by and Konig (26, 27) demonstrate how both requirements cansual scene" be met in a network ofreciprocally coupled oscillators. TheseDirections. In studies show that the segregating effect of conflicting stimuliene into two can be modeled without fast changes of synaptic efficacy,wo superim- which had been considered as a prerequisite for sceneaddition, the segmentation in previous studies (11, 12).lition (25)- The observation that the functional coupling of corticalhe remaining cells is dynamic and subject to rapid changes has particular
Neurobiology: Engel et al.
An:1.I.r 11v.. A).
1" 'T ..........
-60

Proc. Natl. Acad. Sci. USA 88 (1991)
implications for the interpretation ofcross-correlograms (19).Our data demonstrate that no direct inferences can be madefrom correlation data on the underlying microcircuitry andthat, in particular, the absence of correlogram peaks does notprove a lack of anatomical connectivity. In a recent cross-correlation study of cat visual cortex, Ts'o and coworkers(24) reported preferential correlation between cells withsimilar orientation preference and concluded that only cellswith like receptive-field properties are linked via horizontalconnections. At least with respect to short-range interac-tions, our results (18, 19) are not consistent with this inter-pretation because with single light bar stimuli we readily findtemporal correlations between cells in adjacent columns withdissimilar orientation preferences. The apparent discrepancymay, in fact, be explained by the stimulus dependence oftemporal correlations as observed in this study. Ts'o et al.(24) always stimulated the cells with their preferred orienta-tion. Therefore, when recording from cells with overlappingreceptive fields and different orientation preferences, theseauthors used a configuration similar to our conflicting stimuli.Thus, they may have decreased the probability of observingcorrelated firing of cells with dissimilar orientation prefer-ences.
In conclusion, the present results provide further evidencethat cortical representations may be created in a highlydynamic manner by transient synchronization of neuronalresponses (17-23). As demonstrated here, superposition oftwo stimuli in the visual field induces the formation of twoassemblies of synchronously firing cells that are desynchro-nized with respect to each other. This result suggests theexistence ofa temporal-coding mechanism for scene segmen-tation, which involves synchronization of neurons respond-ing to the same object, but desynchronization of cells codingfor different objects of the scene.
We thank A. Herrmann and R. Ruhl-Volsing for technical assis-tance and T. B. Schillen and G. Rager for helpful comments on themanuscript.
1. Kohler, W. (1930) Gestalt Psychology (Bell & Sons, London).2. Koffka, K. (1935) Principles ofGestalt Psychology (Routledge
& Kegan Paul, London).3. Treisman, A. (1986) in Handbook of Perception and Human
Performance, eds. Boff, K., Kaufman, L. & Thomas, I. (Wiley,New York), pp. 35.1-35.70.
4. Barlow, H. B. (1972) Perception 1, 371-394.
5. Hebb, D. 0. (1949) The Organization of Behaviour (Wiley,New York).
6. Edelman, G. M. & Mountcastle, V. B. (1978) The MindfulBrain (MIT, Cambridge, MA).
7. Abeles, M. (1982) Local Cortical Circuits (Springer, Berlin).8. Crick, F. (1984) Proc. Natl. Acad. Sci. USA 81, 4586-4590.9. von der Malsburg, C. (1986) in Brain Theory, eds. Palm, G. &
Aertsen, A. (Springer, Berlin), pp. 161-176.10. Edelman, G. M. (1989) The Remembered Present (Basic
Books, New York).11. von der Malsburg, C. & Schneider, W. (1986) Biol. Cybern. 54,
29-40.12. Sporns, O., Tononi, G. & Edelman, G. M. (1991) Proc. Natl.
Acad. Sci. USA 88, 129-133.13. Gray, C. M. & Singer, W. (1987) Soc. Neurosci. Abstr. 13,
404.3.14. Eckhorn, R., Bauer, R., Jordan, W., Brosch, M., Kruse, W.,
Munk, M. & Reitboeck, H. J. (1988) Biol. Cybern. 60, 121-130.15. Gray, C. M. & Singer, W. (1989) Proc. Natd. Acad. Sci. USA
86, 1698-1702.16. Gray, C. M., Engel, A. K., Konig, P. & Singer, W. (1990) Eur.
J. Neurosci. 2, 607-619.17. Engel, A. K., Kreiter, A. K., Konig, P. & Singer, W. (1991)
Proc. NatI. Acad. Sci. USA 88, 6048-6052.18. Gray, C. M., Konig, P., Engel, A. K. & Singer, W. (1989)
Nature (London) 338, 334-337.19. Engel, A. K., Konig, P., Gray, C. M. & Singer, W. (1990) Eur.
J. Neurosci. 2, 588-606.20. Engel, A. K., Konig, P., Kreiter, A. K. & Singer, W. (1991)
Science 252, 1177-1179.21. Engel, A. K., Konig, P., Gray, C. M. & Singer, W. (1990) in
Parallel Processing in Neural Systems and Computers, eds.Eckmiller, R., Hartmann, G. & Hauske, G. (Elsevier, Amster-dam), pp. 105-108.
22. Singer, W., Gray, C. M., Engel, A. K., Konig, P., Artola, A.& Brocher, S. (1990) Cold Spring Harbor Symp. Quant. Biol.55, 939-952.
23. Engel, A. K., Konig, P., Kreiter, A. K., Gray, C. M. & Singer,W. (1991) in Nonlinear Dynamics and Neural Networks, eds.Schuster, H. G. & Singer, W. (VCH, Weinheim, F.R.G.), inpress.
24. Ts'o, D. Y., Gilbert, C. D. & Wiesel, T. N. (1986) J. Neurosci.6, 1160-1170.
25. Stoner, G. R., Albright, T. D. & Ramachandran, V. S. (1990)Nature (London) 344, 153-155.
26. Konig, P. & Schillen, T. B. (1990) in Parallel Processing inNeural Systems and Computers, eds. Eckmiller, R., Hartmann,G. & Hauske, G. (Elsevier, Amsterdam), pp. 117-120.
27. Schillen, T. B. & Konig, P. (1991) Neural Comput. 3, 167-177.
9140 Neurobiology: Engel et al.