direct visuomotor mapping for fast visually-evoked arm movements
TRANSCRIPT

Neuropsychologia 50 (2012) 3169–3173
Contents lists available at SciVerse ScienceDirect
Neuropsychologia
0028-39
http://d
n Corr
E-m
brian.da1 Te
journal homepage: www.elsevier.com/locate/neuropsychologia
Direct visuomotor mapping for fast visually-evoked arm movements
Raymond F. Reynolds a,n, Brian L. Day b,1
a School of Sport & Exercise Sciences, College of Life & Environmental Sciences, University of Birmingham, Birmingham, B15 2TT, UKb Sobell Department of Motor Neuroscience and Movement Disorders, UCL Institute of Neurology, 33 Queen Square, London WC1N 3BG, UK
a r t i c l e i n f o
Article history:
Received 18 June 2012
Received in revised form
1 October 2012
Accepted 5 October 2012Available online 12 October 2012
Keywords:
Pointing
Motor control
Visuomotor
32/$ - see front matter & 2012 Elsevier Ltd. A
x.doi.org/10.1016/j.neuropsychologia.2012.10
esponding author. Tel.: þ44 121 414 4107; fa
ail addresses: [email protected] (R.F. R
[email protected] (B.L. Day).
l: þ44 0 20 3448 8782.
a b s t r a c t
In contrast to conventional reaction time (RT) tasks, saccadic RT’s to visual targets are very fast and
unaffected by the number of possible targets. This can be explained by the sub-cortical circuitry
underlying eye movements, which involves direct mapping between retinal input and motor output in
the superior colliculus. Here we asked if the choice-invariance established for the eyes also applies to a
special class of fast visuomotor responses of the upper limb. Using a target-pointing paradigm we
observed very fast reaction times (o150 ms) which were completely unaffected as the number of
possible target choices was increased from 1 to 4. When we introduced a condition of altered stimulus–
response mapping, RT went up and a cost of choice was observed. These results can be explained by
direct mapping between visual input and motor output, compatible with a sub-cortical pathway for
visual control of the upper limb.
& 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Many situations require fast visually-guided limb movements,for example when catching a ball or stepping over an obstacle.Human behavioral studies demonstrate that during such tasks,hand or foot trajectories can be adjusted within a remarkablyshort latency of a visual stimulus (o150 ms; Carlton, 1981;Soechting & Lacquaniti, 1983; Day & Lyon, 2000; Reynolds &Day, 2005; Reynolds & Day, 2007; Pruszynski et al., 2010; Brenner& Smeets, 1997; Prablanc & Martin, 1992). The speed of thevisuomotor pathway suggests that it may bypass the cerebralcortex and evidence from a split-brain patient supports this idea(Day & Brown, 2001). However, the notion of a sub-corticalpathway for visually-guided reaching is contentious, and thereis some evidence to the contrary (Desmurget et al., 1999;Greaet al., 2002;Pisella et al., 2000).
By contrast, the existence of a sub-cortical pathway for oculo-motor control is well established both at a behavioral and anato-mical level. Saccadic reaction times (RT) to visual stimuli aretypically within 250 ms (Fischer & Ramsperger, 1984;Saslow,1967). Furthermore, the cost of choice which affects most RT tasks(formalized as Hick’s law; Hick, 1952) does not apply; saccadic RTremains unaffected even as the number of possible target locationsincreases from 1 to 8 (Kveraga & Hughes, 2005;Kveraga, Boucher, &Hughes 2002). This violation of Hick’s law can be explained when
ll rights reserved.
.006
x: þ44 121 414 4121.
eynolds),
we consider the basic nature of the underlying circuitry. Thebrainstem circuitry for visually-guided saccades involves a directpathway from the retina to the oculomotor nuclei via the superiorcolliculus (SC). Visual afferents terminate in a retinotopic fashionwithin the SC, lying in spatial register with premotor neurons whichform a motor map encoding saccade direction and amplitude(Wurtz & Albano, 1980;Gandhi & Katnani, 2011). Thus the anato-mical arrangement involves direct mapping between sensory inputand motor output. This explains why no additional processing isrequired to map required eye movements onto additional targets,thus accounting for the violation of Hick’s law. If the relationshipbetween target and response is rendered arbitrary (e.g., during ‘anti-saccades’) then the direct anatomical mapping is no longer availableand a cost of choice becomes apparent (Kveraga et al., 2002).
If visual guidance of the upper limb is sub-served by a similarpathway then it should exhibit the same behavioral properties assaccades. Specifically we should expect to see fast RT’s and noeffect of choice, but the evidence thus far is equivocal. Studieswhich have varied target choice from 2 upwards have demon-strated minimal or no increase in RT with choice but, unlikesaccades, the transition from 1 to 2 choices imposes a clearincrease in RT (Wright, Marino, Belovsky, & Chubb, 2007;Dassonville, Lewis, Foster, & Ashe, 1999;Berryhill, Kveraga, &Hughes, 2005;Kveraga, Berryhill, & Hughes, 2006). However,these studies have employed indirect reaching paradigms inwhich a joystick or manipulandum is used. We therefore set outto determine if visually-guided pointing is genuinely imperviousto choice using a target-jump paradigm where the task involvesdirect pointing reactions with the index finger. For comparison,we also included a condition of altered stimulus–response

R.F. Reynolds, B.L. Day / Neuropsychologia 50 (2012) 3169–31733170
compatibility involving a 901 rotation. The results clearly showthat direct, but not orthogonal pointing, displays exactly the sameproperties as saccadic eye movement, i.e., fast reaction timescompletely unaffected by choice, even between 1 and 2 choices.Furthermore, analysis of pointing errors during orthogonal point-ing shows that this fast visuomotor process competes for controlof the arm with a slower, more flexible system. The resultssuggest two distinct control pathways for visual guidance of thearm; a sub-cortical pathway which is fast but inflexible, and acortical pathway which is slower but capable of dealing witharbitrary stimulus–response mappings. As the number of choicesincreases, interference between these two systems becomesapparent.
2. Methods
2.1. Subjects
We studied seven healthy right-handed volunteers with no history of
neurological disease (3 female; mean age 31). All subjects gave informed consent
to participate, with ethical approval given by the local ethics committee at the
Institute of Neurology.
2.2. Apparatus
Pointing targets consisted of 2 cm diameter circles of electroluminescent
paper (Pacel Electronics, Dorset, UK) situated on a vertically oriented white board
placed approximately 50 cm in front of the subject’s outstretched right hand
(Fig. 1). The targets could be illuminated and extinguished with precise timing by
controlling the voltage supply to the electroluminescent paper. Four targets were
situated 15 cm to the left, right, above and below a central target. The subject’s left
index finger rested on a proximity sensor (a custom-made capacitance device)
which generated a signal when the finger was lifted. This was used to control trial
timing. Trajectory of the right index finger was measured using an infra-red
Fig. 1. Apparatus and protocol. Subjects pointed with their right index finger
towards a central target. Each trial was initiated by lifting the left index finger
from a proximity sensor. In 1/3rd of trials this caused the target to jump 15 cm to
one of four possible new locations 50 ms later. Subjects were asked to make
pointing adjustments, as depicted by the arrows. These were either in the same
direction as the target jump (‘normal’), or 901 clockwise (‘orthogonal’).
marker placed at its tip. This was sampled at 400 Hz by a CODA mpx30 motion
analysis system (Charnwood Dynamics, Leicestershire, UK).
2.3. Protocol
The task required participants initially to point the index finger of their
outstretched right arm towards the illuminated central target, without any
reaching action and with both vision of hand and target available. The subject
then initiated the trial by raising their left index finger which activated the
proximity sensor. In 1/3rd of randomly selected trials, the target was made to
jump to a new location after 50 ms by simultaneously extinguishing the central
target and illuminating one of the other four targets. The subject was instructed to
attempt to point to the new location as quickly as they could. The end of the trial
was signified by a beep 2.5 s later. To reduce the likelihood of anticipatory
movements, 2/3rd of trials involved no target jump, termed ‘null’ trials. Subjects
were not given any specific gaze instructions.
We implemented two pointing manipulations. First, we varied the number of
possible target-jump locations from 1 to 4. Subjects were made aware of these
locations by placing 2 cm2 squares of coloured tape next to the relevant targets.
Second, there were two conditions of stimulus–response compatibility (SRC); the
instruction was either to follow the target directly (‘normal’) or to point 901
clockwise to the new target location (‘orthogonal’). Hence, there were a total of
eight conditions (4 choice�2 SRC). These were implemented in a blocked design.
Each block comprised 120 trials, consisting of 40 target jumps and 80 null trials.
Each subject therefore experienced 960 trials in total (2 (SRC)�4 (choice)�120).
Block order was randomised between subjects. To control for potential effects of
jump direction upon reaction time (RT), the 1, 2 and 3-choice conditions were
further sub-divided into two 60-trial blocks of differing target combinations. All
possibilities of target combinations were included across the subject group in a
randomised fashion.
2.4. Data analysis
Finger position was low-pass filtered (15 Hz) and differentiated twice to
derive acceleration. RT was defined as the point at which acceleration deviated
in any direction by two standard deviations from baseline, for at least 50 ms.
Baseline was defined as the 0.5 s period prior to target jump. Response direction
was categorised as up, down, left or right. The latency from target illumination to
the initial response (reaction time, RT) was measured in all trials in which the
finger moved. Each response was classified according to whether it was correct or
incorrect. Correct and incorrect responses were analysed separately. The incorrect
responses were subdivided into whether they occurred during null trials, in which
a response was not required, or in stimulus trials but with an initial response in
the wrong direction (termed errors). These response errors were further sub-
divided into those that were directed at the target when it was inappropriate to do
so during orthogonal pointing (termed T-error), and those that were directed
towards any other incorrect location (termed O-error). Incorrect responses in null
trials provided an estimate of the level of predictive ‘false start’ movements.
Incorrectly directed responses (errors) provided information about the difficulty of
suppressing automatic behavior.
Mean RT, and proportions of errors and false starts were analysed with
repeated-measures ANOVA. Po0.05 was considered significant.
3. Results
3.1. Reaction time
During normal pointing, RT remained constant in the face ofincreasing choice (Fig. 2A). This contrasted with orthogonalpointing, where RT systematically increased with choice. ANOVArevealed a significant interaction between choice and stimulus–response compatibility (F3,15¼13.7, po0.001). RT remained vir-tually identical for normal reaching between 1 and 2 choices(140721 ms and 139718 ms, respectively), whereas it increasedfor orthogonal reaching (170739 ms and 240735 ms).
3.2. Proportion of false starts
The tendency for subjects to produce an anticipatory pointingreaction in the absence of a target jump is termed here a falsestart. The proportion of false starts was less than 6% for normalpointing, and less than 2% for the orthogonal condition. However,false starts became slightly less likely as choice increased during
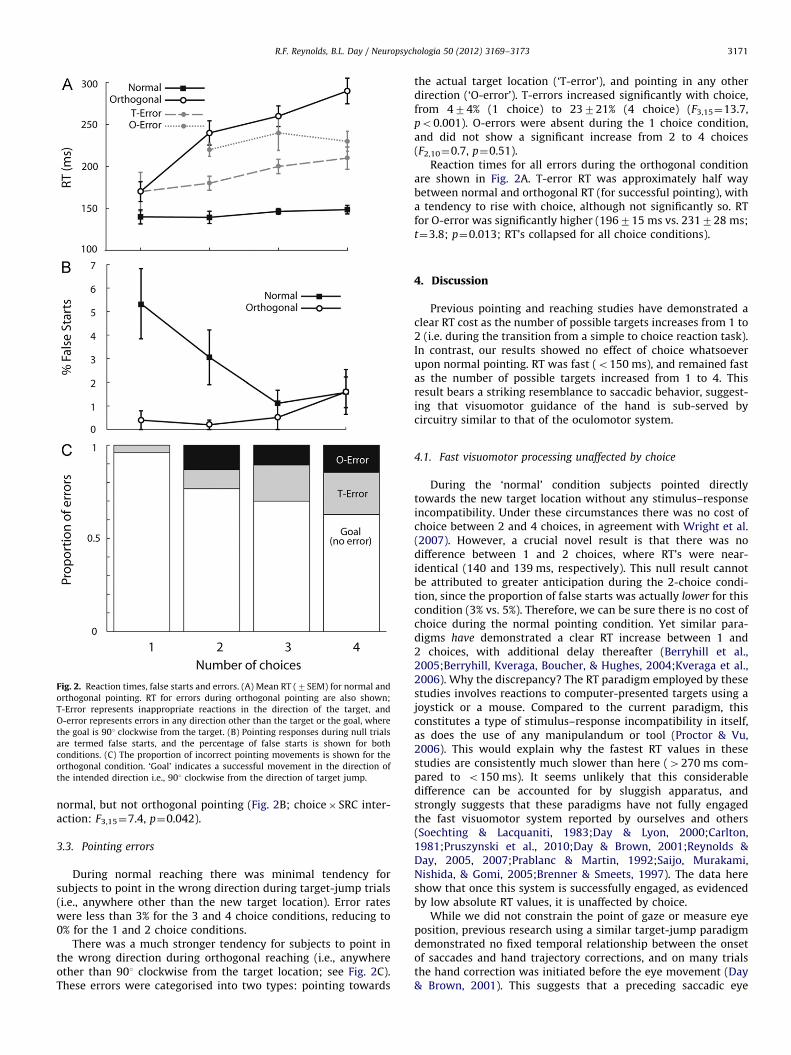
Fig. 2. Reaction times, false starts and errors. (A) Mean RT (7SEM) for normal and
orthogonal pointing. RT for errors during orthogonal pointing are also shown;
T-Error represents inappropriate reactions in the direction of the target, and
O-error represents errors in any direction other than the target or the goal, where
the goal is 901 clockwise from the target. (B) Pointing responses during null trials
are termed false starts, and the percentage of false starts is shown for both
conditions. (C) The proportion of incorrect pointing movements is shown for the
orthogonal condition. ‘Goal’ indicates a successful movement in the direction of
the intended direction i.e., 901 clockwise from the direction of target jump.
R.F. Reynolds, B.L. Day / Neuropsychologia 50 (2012) 3169–3173 3171
normal, but not orthogonal pointing (Fig. 2B; choice� SRC inter-action: F3,15¼7.4, p¼0.042).
3.3. Pointing errors
During normal reaching there was minimal tendency forsubjects to point in the wrong direction during target-jump trials(i.e., anywhere other than the new target location). Error rateswere less than 3% for the 3 and 4 choice conditions, reducing to0% for the 1 and 2 choice conditions.
There was a much stronger tendency for subjects to point inthe wrong direction during orthogonal reaching (i.e., anywhereother than 901 clockwise from the target location; see Fig. 2C).These errors were categorised into two types: pointing towards
the actual target location (‘T-error’), and pointing in any otherdirection (‘O-error’). T-errors increased significantly with choice,from 474% (1 choice) to 23721% (4 choice) (F3,15¼13.7,po0.001). O-errors were absent during the 1 choice condition,and did not show a significant increase from 2 to 4 choices(F2,10¼0.7, p¼0.51).
Reaction times for all errors during the orthogonal conditionare shown in Fig. 2A. T-error RT was approximately half waybetween normal and orthogonal RT (for successful pointing), witha tendency to rise with choice, although not significantly so. RTfor O-error was significantly higher (196715 ms vs. 231728 ms;t¼3.8; p¼0.013; RT’s collapsed for all choice conditions).
4. Discussion
Previous pointing and reaching studies have demonstrated aclear RT cost as the number of possible targets increases from 1 to2 (i.e. during the transition from a simple to choice reaction task).In contrast, our results showed no effect of choice whatsoeverupon normal pointing. RT was fast (o150 ms), and remained fastas the number of possible targets increased from 1 to 4. Thisresult bears a striking resemblance to saccadic behavior, suggest-ing that visuomotor guidance of the hand is sub-served bycircuitry similar to that of the oculomotor system.
4.1. Fast visuomotor processing unaffected by choice
During the ‘normal’ condition subjects pointed directlytowards the new target location without any stimulus–responseincompatibility. Under these circumstances there was no cost ofchoice between 2 and 4 choices, in agreement with Wright et al.(2007). However, a crucial novel result is that there was nodifference between 1 and 2 choices, where RT’s were near-identical (140 and 139 ms, respectively). This null result cannotbe attributed to greater anticipation during the 2-choice condi-tion, since the proportion of false starts was actually lower for thiscondition (3% vs. 5%). Therefore, we can be sure there is no cost ofchoice during the normal pointing condition. Yet similar para-digms have demonstrated a clear RT increase between 1 and2 choices, with additional delay thereafter (Berryhill et al.,2005;Berryhill, Kveraga, Boucher, & Hughes, 2004;Kveraga et al.,2006). Why the discrepancy? The RT paradigm employed by thesestudies involves reactions to computer-presented targets using ajoystick or a mouse. Compared to the current paradigm, thisconstitutes a type of stimulus–response incompatibility in itself,as does the use of any manipulandum or tool (Proctor & Vu,2006). This would explain why the fastest RT values in thesestudies are consistently much slower than here (4270 ms com-pared to o150 ms). It seems unlikely that this considerabledifference can be accounted for by sluggish apparatus, andstrongly suggests that these paradigms have not fully engagedthe fast visuomotor system reported by ourselves and others(Soechting & Lacquaniti, 1983;Day & Lyon, 2000;Carlton,1981;Pruszynski et al., 2010;Day & Brown, 2001;Reynolds &Day, 2005, 2007;Prablanc & Martin, 1992;Saijo, Murakami,Nishida, & Gomi, 2005;Brenner & Smeets, 1997). The data hereshow that once this system is successfully engaged, as evidencedby low absolute RT values, it is unaffected by choice.
While we did not constrain the point of gaze or measure eyeposition, previous research using a similar target-jump paradigmdemonstrated no fixed temporal relationship between the onsetof saccades and hand trajectory corrections, and on many trialsthe hand correction was initiated before the eye movement (Day& Brown, 2001). This suggests that a preceding saccadic eye

R.F. Reynolds, B.L. Day / Neuropsychologia 50 (2012) 3169–31733172
movement is not necessary for, or predictive of, visually-guidedcorrections of upper limb trajectory.
During the ‘orthogonal’ condition there was an increase in RTbetween 1 and 2 choices (65 ms), and between 2 and 4 choices(54 ms), which corresponds reasonably well to the log2 increasepredicted by Hick’s law (Hick, 1952;Hyman, 1953). Thus, alteringthe stimulus–response relationship imposed a cost of choice, aneffect previously reported for the oculomotor system whensaccades have been compared to anti-saccades (Kveraga et al.,2002). In addition to the differential effects of choice betweennormal and orthogonal pointing, the absolute RT values were alsohigher during orthogonal pointing. This was true even for the1 choice condition which is equivalent to a simple reaction task(140 and 173 ms for normal and orthogonal pointing, respec-tively). This means that even when the required movement isknown in advance, and when both stimuli and movement are ofthe same type in both conditions, a simple alteration in stimulus–response mapping produces a clear RT deficit. This constitutescompelling evidence that the slightest deviation from directpointing will disengage fast visuomotor processing, even forsimple reaction tasks. This could explain why previous researchhas failed to unequivocally demonstrate choice invariance for theupper limb.
4.2. Two competing visuomotor control systems
During orthogonal pointing subjects were instructed to point901 away from the target jump direction. However, finger trajec-tories revealed a tendency to automatically follow the targetdespite the intention. This tendency has previously beendescribed by Day and Lyon (2000) who asked subjects to eitherreach towards a jumped target, or to move in the oppositedirection (‘anti-reaching’). During the latter condition, subjectsunintentionally followed the target on many trials. Similar tointentional following reactions, these unintended reactions werefast, (RTo160 ms), whereas movements which took the handaway from the target were slower (RTc160 ms) (Also see Saijoet al., 2005 for similar finding). The results here extend thesefindings in two ways. First, we have shown that the tendency toautomatically follow a visual target increases systematically withchoice (from 4% with 1 choice, to 23% with 4 choices). Hence, asthe complexity of the task increases, the ability to suppressautomatic visual interception is impaired. Second, the RT of theseunintended following reactions lies halfway between the fastautomatic responses and the slow orthogonal responses. Thissuggests two classes of visually-guided reactions; a fast automaticsystem and a slower, more flexible system. The former system isappropriate for simple interceptive reactions, such as catching aball or swatting a fly. The latter system allows for more complexvisuomotor behavior required in situations where the visualstimulus and the required hand position are not in the samelocation. Such arbitrary stimulus–response mapping regularlyoccurs during tool usage, a common example being manipulationof a computer screen cursor with a mouse. For the flexible systemto operate successfully requires suppression of automatic inter-ception responses. However, as the stimulus–response relation-ship becomes more complex, this suppression is compromised,and there is competition between the two visuomotor systems forcontrol of the hand.
4.3. Neural substrates for visuomotor control of the hand
Given that the pointing behavior we observed is very similar tothat of saccades, this raises the possibility that manual and ocularvisuomotor pathways share similar circuitry. As discussed inthe introduction, visually-guided saccades are sub-served by a
sub-cortical pathway from the retina to the oculomotor nuclei, viathe superior colliculus (SC). Could fast visual guidance of theupper limb use a similar pathway? Behavioral data is consistentwith the idea of a sub-cortical pathway; reaching trajectoryadjustments occur even in the absence of perception of a targetjump (Goodale, Pelisson, & Prablanc, 1986), and the earliest aremuscle responses are time-locked to, and within 100 ms of, thevisual stimulus (Pruszynski et al., 2010). Neurophysiologic data insupport of a sub-cortical route also exists, with animal experi-ments implying a central role for the SC in reaching. For example,direct stimulation of the cat SC causes short-latency perturbationsof reaching movements (Courjon, Olivier, & Pelisson, 2004), andthe primate SC has been shown to contain neurons whose activityis closely related to visually-guided upper limb movement(Werner, 1993). However, the most direct evidence for a sub-cortical route comes from a split-brain patient (Day & Brown,2001). This patient exhibited a severe RT deficit during a conven-tional reaction task in which a right finger-lift movement wasdemanded in response to a right hemi-field visual stimulus. Thisis consistent with a visuomotor pathway traversing the corpuscallosum. However, when a reach adjustment was demanded inresponse to a target-jump, using the same visual stimulus and thesame hand, no such deficit was observed. This suggests that thevisuomotor pathway for online control of reaching can bypass thecerebral cortex. Despite this evidence other studies suggest acrucial role for the posterior parietal cortex (PPC). In patients withPPC damage, or in healthy subjects undergoing transcranialmagnetic stimulation of PPC, online reach adjustments are atte-nuated or abolished while normal reaching is left unaffected(Desmurget et al., 1999;Grea et al., 2002;Pisella et al., 2000).The PPC may therefore play a key role in visuomotor transforma-tions underlying reach corrections. One possible explanation forthese contradictory findings is that cortical areas, such as the PPC,may play a supervisory role over faster sub-cortical routes(Glickstein, 2003), analogous to a spinal reflex being modulatedby descending input (Schieppati, 1987). So while perturbation ofPPC activity may, in turn, perturb online control of reaching, thefastest visuomotor route may nevertheless bypass the cortex.Perhaps more direct neurophysiologic evidence will determine ifthis is the case.
What neural mechanism might underlie the switch from the fastvisuomotor system to the more flexible, slower one? Primatebrainstem recordings show that anti-saccade generation is asso-ciated with active suppression of saccade-related neurons in the SC,probably caused by descending input from cortical eye movementareas (Everling, Dorris, Klein, & Munoz, 1999). A similar process mayoccur for reaching movements. As the task becomes more complex(i.e., more choice), the ability of the cerebral cortex to successfullyinhibit automatic responses may be compromised. This wouldexplain the pattern of errors and associated RT observed here.
In summary, we have shown that fast visually-guided pointingmovements exhibit the same behavioral characteristics as sac-cades. That is, they are very fast and completely unaffected bychoice. This contrasts with the slower system which is capable ofdealing with altered stimulus–response mappings. To account forour data, we hypothesise two anatomical pathways sub-servingtwo visuomotor functions. First, there is a fast, automatic inter-ception system relying upon circuitry which has approximatelyone-to-one mapping between retinal input and motor output.This is likely to be a sub-cortical circuit possibly including thesuperior colliculus. This pathway allows for very fast interceptionresponses. Second, there is a slower, more flexible pathwayinvolving the cerebral cortex, allowing for more complex beha-viors. For any given situation the correct pathway must beselected, which becomes increasingly difficult during conditionsof ambiguous or complex stimulus–response mappings.

R.F. Reynolds, B.L. Day / Neuropsychologia 50 (2012) 3169–3173 3173
References
Berryhill, M., Kveraga, K., Boucher, L., & Hughes, H. C. (2004). Smooth pursuitunder stimulus–response uncertainty. Brain Research: Cognitive Brain Research,19, 100–102.
Berryhill, M., Kveraga, K., & Hughes, H. C. (2005). Effects of directional uncertaintyon visually-guided joystick pointing. Perceptual and Motor Skills, 100, 267–274.
Brenner, E., & Smeets, J. B. (1997). Fast responses of the human hand to changes intarget position. Journal of Motor Behavior, 29, 297–310.
Carlton, L. G. (1981). Processing visual feedback information for movementcontrol. Journal of Experimental Psychology: Human Perception and Performance,7, 1019–1030.
Courjon, J. H., Olivier, E., & Pelisson, D. (2004). Direct evidence for the contributionof the superior colliculus in the control of visually guided reaching movementsin the cat. Journal of Physiology, 556, 675–681.
Dassonville, P., Lewis, S. M., Foster, H. E., & Ashe, J. (1999). Choice and stimulus–response compatibility affect duration of response selection. Brain Research:Cognitive Brain Research, 7, 235–240.
Day, B. L., & Brown, P. (2001). Evidence for subcortical involvement in the visualcontrol of human reaching. Brain, 124, 1832–1840.
Day, B. L., & Lyon, I. N. (2000). Voluntary modification of automatic arm move-ments evoked by motion of a visual target. Experimental Brain Research, 130,159–168.
Desmurget, M., Epstein, C. M., Turner, R. S., Prablanc, C., Alexander, G. E., & Grafton,S. T. (1999). Role of the posterior parietal cortex in updating reachingmovements to a visual target. Nature Neuroscience, 2, 563–567.
Everling, S., Dorris, M. C., Klein, R. M., & Munoz, D. P. (1999). Role of primatesuperior colliculus in preparation and execution of anti-saccades and pro-saccades. Journal of Neuroscience, 19, 2740–2754.
Fischer, B., & Ramsperger, E. (1984). Human express saccades: extremely shortreaction times of goal directed eye movements. Experimental Brain Research,57, 191–195.
Gandhi, N. J., & Katnani, H. A. (2011). Motor functions of the superior colliculus.Annual Reviews in Neuroscience, 34, 205–231.
Glickstein, M. (2003). Subcortical projections of the parietal lobes. Advances inNeurology, 93, 43–55.
Goodale, M. A., Pelisson, D., & Prablanc, C. (1986). Large adjustments in visuallyguided reaching do not depend on vision of the hand or perception of targetdisplacement. Nature, 320, 748–750.
Grea, H., Pisella, L., Rossetti, Y., Desmurget, M., Tilikete, C., Grafton, S., et al. (2002).A lesion of the posterior parietal cortex disrupts on-line adjustments duringaiming movements. Neuropsychologia, 40, 2471–2480.
Hick, W. E. (1952). On the rate of gain of information. Quarterly Journal ofExperimental Psychology, 11–26.
Hyman, R. (1953). Stimulus information as a determinant of reaction time. Journal
of Experimental Psychology, 45, 188–196.Kveraga, K., Berryhill, M., & Hughes, H. C. (2006). Directional uncertainty in
visually guided pointing. Perceptual and Motor Skills, 102, 125–132.Kveraga, K., Boucher, L., & Hughes, H. C. (2002). Saccades operate in violation of
Hick’s law. Experimental Brain Research, 146, 307–314.Kveraga, K., & Hughes, H. C. (2005). Effects of stimulus–response uncertainty on
saccades to near-threshold targets. Experimental Brain Research, 162, 401–405.Pisella, L., Grea, H., Tilikete, C., Vighetto, A., Desmurget, M., Rode, G., et al. (2000).
An ‘automatic pilot’ for the hand in human posterior parietal cortex: towardreinterpreting optic ataxia. Nature Neuroscience, 3, 729–736.
Prablanc, C., & Martin, O. (1992). Automatic control during hand reaching atundetected two-dimensional target displacements. Journal of Neurophysiology,67, 455–469.
Proctor, R. W., & Vu, K. L. (2006). Stimulus–response compatibility principles: data,
theory, and application. Boca Raton, Fl; London: CRC 2006.Pruszynski, J. A., King, G. L., Boisse, L., Scott, S. H., Flanagan, J. R., & Munoz, D. P.
(2010). Stimulus-locked responses on human arm muscles reveal a rapid
neural pathway linking visual input to arm motor output. European Journal of
Neuroscience, 32, 1049–1057.Reynolds, R. F., & Day, B. L. (2005). Rapid visuomotor processes drive the leg
regardless of balance constraints. Current Biology, 15, R48–R49.Reynolds, R. F., & Day, B. L. (2007). Fast visuomotor processing made faster by
sound. Journal of Physiology, 583, 1107–1115.Saijo, N., Murakami, I., Nishida, S., & Gomi, H. (2005). Large-field visual motion
directly induces an involuntary rapid manual following response. Journal of
Neuroscience, 25, 4941–4951.Saslow, M. G. (1967). Latency for saccadic eye movement. Journal of the Optical
Society of America, 57, 1030–1033.Schieppati, M. (1987). The Hoffmann reflex: a means of assessing spinal reflex
excitability and its descending control in man. Progress in Neurobiology, 28,
345–376.Soechting, J. F., & Lacquaniti, F. (1983). Modification of trajectory of a pointing
movement in response to a change in target location. Journal of Neurophysiol-
ogy, 49, 548–564.Werner, W. (1993). Neurons in the primate superior colliculus are active before
and during arm movements to visual targets. European Journal of Neuroscience,5, 335–340.
Wright, C. E., Marino, V. F., Belovsky, S. A., & Chubb, C. (2007). Visually guided,aimed movements can be unaffected by stimulus–response uncertainty.
Experimental Brain Research, 179, 475–496.Wurtz, R. H., & Albano, J. E. (1980). Visual-motor function of the primate superior
colliculus. Annual Reviews in Neuroscience, 3, 189–226.