diplomarbeit effektorfunktionen von dendritischen zellen ...¤t_vmf/institut_immunologie...i die...
TRANSCRIPT

I
Universität Leipzig
Fakultät für Biowissenschaften, Pharmazie und Psychologie
Diplomarbeit
Effektorfunktionen von dendritischen Zellen nach
Stimulation mit inaktiviertem Parapoxvirus ovis
vorgelegt von
Franziska Herden
geb. am 15.März 1983 in Leipzig
Studentin im Diplomstudiengang Biochemie
Leipzig, den 19. März 2008

I
Die vorliegende Arbeit wurde vom 24.09.2007 bis zum 19.03.2008 am Institut für
Immunologie der Veterinärmedizinischen Fakultät der Universität Leipzig angefertigt.
Betreut wurde die Arbeit von Herrn Professor Dr. med. vet. G. Alber, Institut für
Immunologie der Veterinärmedizinischen Fakultät der Universität Leipzig, und Frau
Professor Dr. rer. nat. A. Beck-Sickinger, Institut für Biochemie der Fakultät für
Biowissenschaften, Pharmazie und Psychologie der Universität Leipzig.

ABKÜRZUNGSVERZEICHNIS
II
Verzeichnis verwendeter Abkürzungen
Auf die Auflistung deutscher Termini wird verzichtet, wenn sie dem gängigen
Sprachgebrauch entsprechen. Physikalische und chemische Symbole werden ohne
Erläuterung verwendet, die Einbuchstaben-Schreibweise der Aminosäuren und der Purin- und
Pyrimidinbasen der Nukleinsäuren entsprechen den IUPAC-Regeln und werden nicht
gesondert aufgeführt.
Abkürzung Erklärung AK Antikörper APC Allophycoerythrin aqua bidest. Deionisiertes und durch Umkehrosmose destilliertes Wasser CD cluster of differentiation cDC conventional dendritic cell, konventionelle dendritische Zelle CpG-ODN CpG Oligodesoxynukleotide (als TLR9-Ligand verwendet) DCs dendritische Zellen def defizient DAI DNA-dependent activator of IFN-regulatory factors,
DNA-abhängiger Aktivator von IFN-regulatorischen Faktoren DMEM Dulbecco´s modified Eagle medium DMSO Dimethylsulfoxid DNA Desoxyribonukleinsäure ds doppelsträngig EDTA Ethylendiamintetraessigsäure EM Elektronenmikroskopie EMA Ethidiummonoazidbromid FACS fluorescence activated cell sorting Fcγ-Rezeptor fragment crystallizable gamma receptor FITC Fluoresceinisothiocyanat FKS Fötales Kälberserum FL Fluoreszenz Flt3L fms-like tyrosin kinase 3 ligand, fms-ähnlicher Tyrosinkinase-3-Ligand GM-CSF Granulozyten-Makrophagen-Koloniestimulierender Faktor HEPES 2-[4-(2-Hydrodyethyl)-1-piperazinyl]-ethansulfonsäure Ham Hamster HRP horse radish peroxidase HSV Herpes-Simplex-Virus iPPVO chemisch inaktiviertes Parapoxvirus ovis hu human Ig Immunglobulin IRF Interferon-regulatorischer Faktor (en) IFN Interferon(e) IL Interleukin(e) LPS Lipopolysaccharid (als TLR4-Ligand verwendet) M-CSF Makrophagen-Koloniestimulierender Factor MDA5 melanoma-differentiation-associated gene 5 MHC major histocompatibility complex, Haupthistokompatibilitätskomplex

ABKÜRZUNGSVERZEICHNIS
III
MOI multiplicity of infection Ms Maus mu murin MyD88 myeloid differentiation primary response gene 88 NK cell(s) natürliche Killerzelle(n) NO Stickstoffmonoxid NF-κB nukleärer Faktor-kappa B PPVO Parapoxvirus ovis Pam3CSK4 N-Palmitoyl-S-[2,3-bis(palmitoyloxy)-(2RS)-propyl]-Cys-Ser-(Lys)4 x
3HCl (als TLR2-Ligand verwendet) PAMP pathogen-associated molecular patterns, Pathogen-assoziierte molekulare
Strukturen PRR pattern recognition receptors, Mustererkennungsrezeptoren PBS Phosphatgepufferte Kochsalzlösung pDC plasmacytoid dendritic cell, plasmazytoide dendritische Zelle PE Phycoerythrin Poly I:C polyinosinic-polycytidylic acid (als TLR3-Ligand verwendet) R-848 Resiquimod (als TLR7-Ligand verwendet) RIG-I retinoic-acid-inducible gene I RNA Ribonukleinsäure Rpm rounds per minute, Umdrehungen pro minute RPMI Roswell Park Memorial Institut RT Raumtemperatur SA Streptavidin SDS sodium dodecyl sulfate, Natriumdodecylsulfat SEM standard error of the mean, Standardfehler TMB Tetramethylbenzidin TRIF TIR-domain containing adaptor protein inducing IFN TIR Toll/IL-1R homology domain TLR toll-like receptor(s), Toll-ähnliche Rezeptor(en) TNF Tumornekrosefaktor(en) WT Wildtyp VV Vaccinia-Viren

INHALTSVERZEICHNIS
IV
Inhaltsverzeichnis
1 Einleitung 1
1.1 Parapoxvirus ovis 1
1.2 Dendritische Zellen 6
1.2.1 Subpopulationen von dendritischen Zellen 6
1.2.2 Erkennung von Viren durch dendritische Zellen über
Mustererkennungsrezeptoren 8
1.2.3 Die antivirale Effektorantwort von dendritischen Zellen 11
1.3 Zielstellung 13
2 Material 14
2.1 Tiere und Pathogene 14
2.1.1 Mäuse 14
2.1.2 Parapoxvirus ovis 14
2.2 Zelllinien 14
2.3 Wachstumsfaktoren 15
2.4 Antikörper 15
2.4.1 Fangantikörper für Zytokin-Sandwich-ELISA 15
2.4.2 Detektionsantikörper für Zytokin-Sandwich-ELISA 15
2.4.3 Sekundärantikörper für Zytokin-Sandwich-ELISA 15
2.4.4 Antikörper für durchflusszytometrische Analysen 16
2.5 Proteinstandards 16
2.6 Geräte und Materialien 16
2.7 Software 18
2.8 Chemikalien, Lösungen, Puffer und Nährmedien 18
2.8.1 Allgemeine Chemikalien 18
2.8.2 Puffer und Lösungen für Zellisolation 20
2.8.3 Puffer und Lösungen für Durchflusszytometrie 20
2.8.4 Puffer und Lösungen für ELISA 20
2.8.5 Medien und Lösungen für Zellkultur 21
2.8.6 Kits 22
2.8.7 DNA-Standards 22
2.8.8 Puffer und Lösungen für Isolation viraler DNA 22

INHALTSVERZEICHNIS
V
2.9.9 Puffer und Lösungen für Agarose-Gelelektrophorese 22
3 Methoden 24
3.1 Zellbiologische Methoden 24
3.1.1. Kultivierung von P388-Zellen für die Etablierung der Lebend-/
Tot-Färbung mit Ethidiummonoazidbromid 24
3.1.2. Generierung von dendritischen Zellen aus Knochenmark 24
3.1.3. Stimulation von dendritischen Zellen 25
3.2 Durchflusszytometrische Analysen 25
3.3 Quantifizierung von Zytokinen über Sandwich-ELISA 27
3.4 Generierung und Stimulation von dendritischen Zellen für elektronen-
mikroskopische Aufnahmen 28
3.5 Molekularbiologische Methoden 28
3.5.1. Isolation viraler DNA 28
3.5.2. Agarose-Gelelektrophorese 29
3.6 Statistische Analysen 30
4 Ergebnisse 31
4.1 Etablierung einer Lebend-/Tot-Färbung mit Ethidiummonoazidbromid 31
4.2 Charakterisierung dendritischer Zellen, generiert mit fms-like tyrosine
kinase-3-ligand aus murinen Knochenmarkszellen 32
4.3 Charakterisierung der Aktivierung von dendritischen Zellen nach
Stimulation mit inaktiviertem Parapoxvirus ovis 35
4.3.1 Aufregulation von MHC-I/-II und kostimulatorischen Molekülen 35
4.3.2 Produktion antiviraler (IFN-α) und proinflammatorischer
(IL-12/23p40 und TNF-α) Zytokine 37
4.3.3 Vergleich zwischen DCs, generiert mit humanem und murinem
Flt3L 38
4.3.4 Untersuchungen zur Aufnahme von iPPVO durch DCs 40
4.3.5 Kinetische Analyse der iPPVO-induzierten DC-Aktivierung 42
4.4 Beteiligung der Toll-ähnlichen Rezeptoren 9 und 2 an der Stimulation
von dendritischen Zellen durch inaktiviertes Parapoxvirus ovis 45
4.4.1 Rolle von TLR9 bei der iPPVO-induzierten DC-Aktivierung 45
4.4.2 Rolle von TLR2 bei der iPPVO-induzierten DC-Aktivierung 48

INHALTSVERZEICHNIS
VI
5 Diskussion 51
5.1 Charakterisierung der immunstimulatorischen Eigenschaften von
inaktiviertem Parapoxvirus ovis auf dendritische Zellen 51
5.1.1 Vorbereitende Untersuchungen zu DCs mit Flt3L und EMA 51
5.1.2 iPPVO induziert die Expression kostimulatorischer Moleküle
eher auf cDCs als auf pDCs 52
5.1.3 Der Einfluss von IFN-α auf die durch iPPVO- ausgelöste
DC Aktivierung 53
5.1.4 Rolle von IFN-β für die DC-Effektorantwort nach Kontakt mit
iPPVO 54
5.1.5 Die iPPVO-induzierte DC-Aktivierung führt zur
Freisetzung von IL-12/23p40 und TNF-α 54
5.1.6 Unterschiedliche Kinetik der durch iPPVO induzierten
DC-Aktivierungsmarker 55
5.2 Inaktiviertes Parapoxvirus ovis wird endozytotisch durch dendritische
Zellen aufgenommen 57
5.3 Beteiligung von Toll-ähnlichen Rezeptoren an der durch inaktiviertes
Parapoxvirus ovis induzierten Aktivierung dendritischer Zellen 58
5.3.1 Die DC-Aktivierung durch iPPVO ist TLR9-unabhängig 58
5.3.2 Die DC-Aktivierung nach iPPVO-Stimulation ist TLR2-
unabhängig 61
5.3.3 Weiterführende Untersuchungen an iPPVO mit neuen
Anwendungsbereichen 63
6 Ausblick 64
7 Zusammenfassung 65
8 Literaturangabe 66

VII
Eidesstattliche Erklärung
Ich erkläre an Eides statt, dass ich die vorliegende Arbeit selbstständig und ohne fremde Hilfe
angefertigt habe. Ich habe nur die angegebene Literatur bzw. angeführten Quellen benutzt.
Leipzig,

EINLEITUNG
1
1 Einleitung
1.1 Parapoxvirus ovis
Parapoxvirus ovis (PPVO) aus der Familie der Poxvirdae (Subfamilie Orthopoxvirus) ist ein
Vertreter der Gattung Parapoxvirus. Es verursacht eine weltweit vorkommende Infektion bei
Schafen und Ziegen, bekannt als Ecthyma contagiosum, pustuläre Dermatitis oder
Lippengrind. Betroffene Tiere zeigen Schleimhautläsionen, Pusteln und Abschürfungen
(Abb. 1). Die Infektion ist auf den Menschen übertragbar nach direktem Kontakt mit
infizierten Tieren. Jedoch bleibt die Erkrankung beim Menschen lokal begrenzt und
auftretende Hautpusteln verheilen schnell (1).
Abb. 1: Ausprägungen des durch PPVO verursachten Ecthyma contagiosum bei Tier und
Mensch. (A) Die bei Schafen und Ziegen vorkommende Erkrankung zeichnet sich besonders durch
verletzte Schleimhäute an Maul und Euter der Tiere aus. (B) Lokal auftretende Symptome beim
Menschen nach direktem Kontakt mit infizierten Tieren, auch bekannt als Melkersknoten, verheilen
schnell und ohne Komplikationen (2).
PPVO ist ein epitheliotrophes, doppelsträngiges DNA-Virus, dessen lineares Genom eine
Größe von 140 kb aufweist (Abb. 2). Dabei bilden 3 kb an jedem Ende des Genoms
invertierte terminale Schlaufen (3). Die virale DNA zeichnet sich durch einen sehr hohen
G/C-Gehalt von 63 % im Gegensatz zu anderen Pockenviren aus (4). Das PPVO-Genom
enthält viele immunmodulatorische Gene, die für Proteine wie den vaskulären endothelialen
Wachstumsfaktor VEGF-e, ein Ortholog für Interleukin (IL)-10, und einen inhibitorschen
Faktor des Granulozyten-Makrophagen koloniestimulierenden Faktors (GM-CSF), kodieren.
Sie ermöglichen eine schnelle Replikation des Virus im Wirt, bevor dessen Immunsystem
effektiv die Infektion bekämpfen kann (5). Der in der vorliegenden Arbeit verwendete Stamm
D1701 wurde ursprünglich aus einem Schaf isoliert und in Zellkultur mit der bovinen
Nierenzellinie BKKL3A adaptiert (6). Er zeichnet sich durch eine stark reduzierte Virulenz
EINLEITUNG
2
aus, d.h. es kam in Schafen zu stark verminderten Symptomen bei subkutaner Injektion des
Virus.
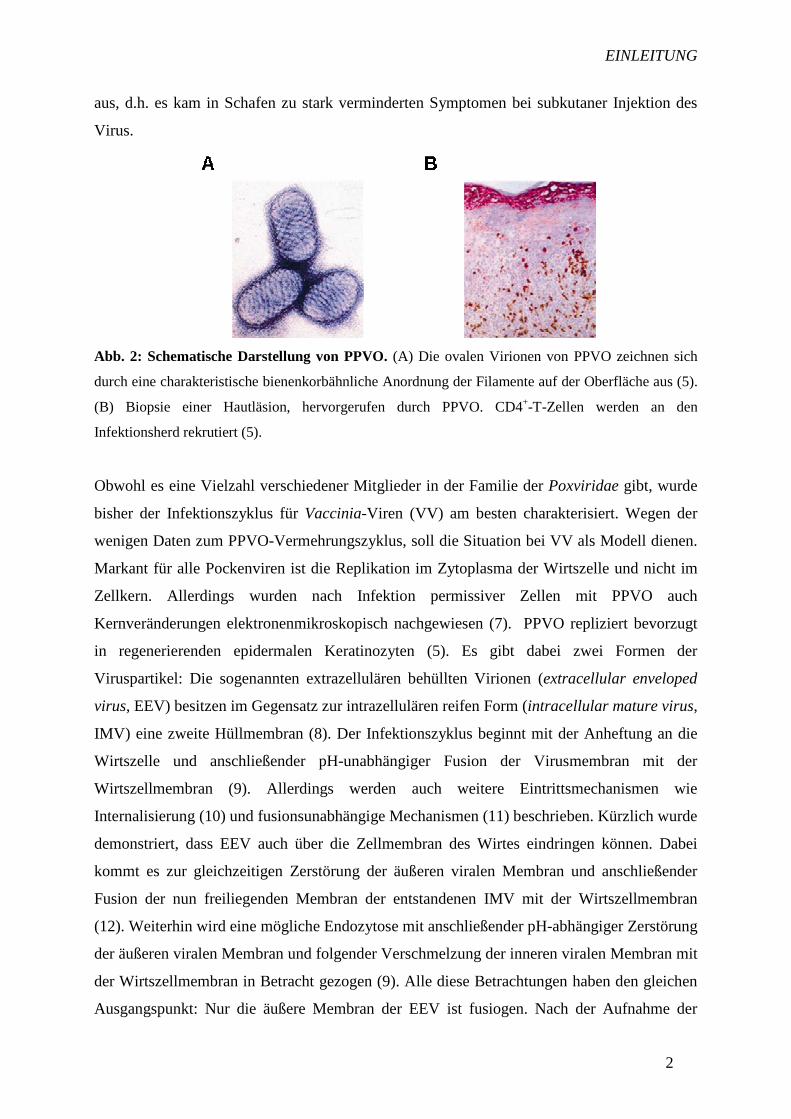
Abb. 2: Schematische Darstellung von PPVO. (A) Die ovalen Virionen von PPVO zeichnen sich
durch eine charakteristische bienenkorbähnliche Anordnung der Filamente auf der Oberfläche aus (5).
(B) Biopsie einer Hautläsion, hervorgerufen durch PPVO. CD4+-T-Zellen werden an den
Infektionsherd rekrutiert (5).
Obwohl es eine Vielzahl verschiedener Mitglieder in der Familie der Poxviridae gibt, wurde
bisher der Infektionszyklus für Vaccinia-Viren (VV) am besten charakterisiert. Wegen der
wenigen Daten zum PPVO-Vermehrungszyklus, soll die Situation bei VV als Modell dienen.
Markant für alle Pockenviren ist die Replikation im Zytoplasma der Wirtszelle und nicht im
Zellkern. Allerdings wurden nach Infektion permissiver Zellen mit PPVO auch
Kernveränderungen elektronenmikroskopisch nachgewiesen (7). PPVO repliziert bevorzugt
in regenerierenden epidermalen Keratinozyten (5). Es gibt dabei zwei Formen der
Viruspartikel: Die sogenannten extrazellulären behüllten Virionen (extracellular enveloped
virus, EEV) besitzen im Gegensatz zur intrazellulären reifen Form (intracellular mature virus,
IMV) eine zweite Hüllmembran (8). Der Infektionszyklus beginnt mit der Anheftung an die
Wirtszelle und anschließender pH-unabhängiger Fusion der Virusmembran mit der
Wirtszellmembran (9). Allerdings werden auch weitere Eintrittsmechanismen wie
Internalisierung (10) und fusionsunabhängige Mechanismen (11) beschrieben. Kürzlich wurde
demonstriert, dass EEV auch über die Zellmembran des Wirtes eindringen können. Dabei
kommt es zur gleichzeitigen Zerstörung der äußeren viralen Membran und anschließender
Fusion der nun freiliegenden Membran der entstandenen IMV mit der Wirtszellmembran
(12). Weiterhin wird eine mögliche Endozytose mit anschließender pH-abhängiger Zerstörung
der äußeren viralen Membran und folgender Verschmelzung der inneren viralen Membran mit
der Wirtszellmembran in Betracht gezogen (9). Alle diese Betrachtungen haben den gleichen
Ausgangspunkt: Nur die äußere Membran der EEV ist fusiogen. Nach der Aufnahme der

EINLEITUNG
3
Virionen beginnt im nun freigesetzten, noch intakten Core die Transkription früher Gene
(Abb. 3) (13). Im Anschluß folgt die Freisetzung des Virusgenoms in das Cytoplasma der
Wirtszelle. Die DNA-, RNA- und Proteinsynthese werden in der Wirtszelle schon im
Anfangsstadium des Infektionszyklus abgeschaltet („shut off“). Die Bereiche der viralen
Transkription und DNA-Replikation im Cytoplasma, auch Virusfabriken genannt (14),
werden von Membranen des endosomalen Retikulums umschlossen (15). Nach der späten
Phase der viralen Transkription findet die Neusynthese von Virionen in granulären Bereichen
des Cytoplasmas statt (Abb. 3). Nur ein Teil der bisher entstandenden intrazellulären
Viruspartikel wird mit einer weiteren Membranhülle aus dem trans-Golginetzwerk versehen
und über Mikrotubuli und Aktinfilamente mittels Exozytose nach Verschmelzung der äußeren
viralen Membran mit der Wirtszellmembran aus der Zelle entlassen (16).

EINLEITUNG
4
Abb. 3: Vereinfachte schematische Darstellung der extrazellulären Form (EEV) von
Pockenviren und des Infektionszyklus. (A) Es ist der schematische Aufbau eines EEV-Virions mit
der inneren und äußeren Hüllmembran und der viralen dsDNA dargestellt. Die Funktion der
Lateralkörper ist bislang ungeklärt (abgeleitet nach 17). (B) Die Viruspartikel werden über
verschiedene Wege in die Wirtszelle aufgenommen. Nach der Verschmelzung beider Membranen wird
der Inhalt des Virions in die Zelle entlassen. Dadurch kommt es zur DNA-Replikation und
Genexpression. Die gebildeten Strukturproteine und die virale DNA werden anschließend als reife
Partikel (IMV) verpackt, welche schließlich mit einem Golgivesikel fusionieren. Die EEV entstehen
durch eine zusätzliche Membranhülle aus dem trans-Golginetzwerk und verlassen die Wirtszelle. Ein
weiterer Replikationszyklus kann in neu infizierten Zellen beginnen (abgeleitet nach 18).
Es existieren verschiedene Möglichkeiten zur chemischen Inaktivierung von Viren. Bei dem
Einsatz von Formaldehyd, welches mit chemischen Gruppen von Proteinen reagiert, kann es
immer noch zu einer Virusreplikation in Wirtszellen durch infektiöse virale Nukleinsäuren
kommen. Weiterhin werden die Oberflächenproteine durch Formaldehyd verändert, was zu
einer Reduktion der Antigenizität der viralen Antigene führt (19). Daher eignet sich die

EINLEITUNG
5
Verwendung von binären Ethyliminen, welche die antigenen Eigenschaften von viralen
Oberflächenproteinen kaum verändern. Die Inaktivierungsreaktion über binäre Ethylimine ist
spezifisch für Nukleinsäuren (20). Das gleiche gilt für den Einsatz von β-Propiolakton als
Inaktivierungsreagens (21) (Abb. 4). Daher wurde PPVO zum einen mit binären Ethyliminen
(Pfizer Animal Health) aber auch β-Propiolakton (Prof. M. Büttner) chemisch inaktiviert
(Abb. 7). Dabei kommt es zu einer Methylierung der viralen DNA, wodurch sich das
inaktivierte Parapoxvirus ovis (iPPVO) in der Wirtszelle nicht mehr replizieren kann.
Interessanterweise wirkt iPPVO immunstimulierend. Schon seit 1990 werden diese
immunstimulatorischen Eigenschaften des iPPVO gezielt in der Veterinärmedizin zur
prophylaktischen und therapeutischen Unterstützung von Infektionen und stressbedingten
Erkrankungen bei Pferden, Schweinen, Rindern und kleineren Haustieren wie Hunde oder
Katzen eingesetzt. Das veterinärmedizinische Präparat Zylexis®, früher als Baypamune®
bekannt, enthält iPPVO (D1701) als Wirkstoff (Abb. 4).
Abb. 4: Agenzien zur chemischen Inaktivierung von Viren, eingesetzt im Präparat Zylexis:
binäre Ethylimine und β-Propiolakton. (A) Gezeigt ist die chemische Herstellung von binärem
Ethylimin. Die Zugabe von 2-Aminoethylhydrogensulfat zu einer kochenden Natriumhydroxidlösung
führt zur Bildung von Aziridin. Säure-induzierte Polymerisation von Aziridinen führt zur Entstehung
von binärem Ethylimin neben weiteren oligomeren Ethyliminen (22). (B) Dargestellt ist die chemische
Strukturformel von β-Propiolakton (1,3-Propiolakton). (C) Inaktiviertes Parapoxvirus ovis (iPPVO)
vom Stamm D1701 zeigt die gleiche Morphologie wie PPVO (2).
Die Mechanismen der immunstimulatorischen Aktivität von iPPVO auf Zellen des
angeborenen Immunsystems wie Makrophagen, neurophile Granulozyten und natürliche
Killerzellen wurden in der Literatur bereits beschrieben (23). iPPVO aktiviert

EINLEITUNG
6
antigenpräsentierende Zellen wie Monozyten über Signale durch CD14 und einen Toll-
ähnlichen Rezeptor (23). Dies führt zu einer Aufregulation von proinflammatorischen und
Th1-assoziierten Zytokinen wie Interferon γ (IFN-γ), Tumornekrosefaktor α (TNF-α),
Interleukin (IL)-6, IL-8, IL-12 und IL-18, im weiteren Verlauf schließlich auch zu
antiinflammatorischen und Th2-assoziierten Zytokinen wie IL-4, IL-10 und der IL-1-
Rezeptorantagonist. IL-12 und IL-18 sind dabei in die Sekretion von IFN-γ durch T-Zellen
und/oder NK-Zellen involviert (23). Des Weiteren konnten antivirale Effekte von iPPVO bei
einer Infektion mit Herpes-Simplex-Virus I im Meerschweinchenmodell nachgewiesen
werden (24). Diese immunstimulatorischen Eigenschaften von iPPVO auf Zellen des
angeborenen Immunsystems könnten für neue antivirale Behandlungsstrategien genutzt
werden. Dafür ist eine vollständige Aufklärung des bisher nur eingeschränkt verstandenden
Wirkunsgmechanismus erforderlich. Dendritische Zellen (DCs), wichtige
antigenpräsentierende Zellen des angeborenen Immunsystems, müssen noch im Hinblick auf
iPPVO charakterisiert werden. Denn für die weitere Anwendung von Zylexis® ist es
notwendig, die immunstimulierenden Eigenschaften auf alle Zellen des Immunsystems genau
zu charakterisieren.
1.2 Dendritische Zellen
1.2.1 Subpopulationen von dendritischen Zellen
DCs stellen eine heterogene Population von Immunzellen des angeborenen Immunsystems
dar, welche Antigene aufnehmen, prozessieren und sie anschließend Zellen des adaptiven
Immunsystems wie z.B. T-Zellen präsentieren. Somit sind sie ein wichtiges Bindeglied
zwischen der angeborenen und adaptiven Immunantwort. DCs entstehen sowohl aus
myeloiden als auch lymphoiden Vorläuferzellen im Knochenmark und wandern als unreife
DCs über das Blut in periphere Gewebe ein. Hier werden sie durch Pathogene während einer
Infektion aktiviert. Die reifen dendritischen Zellen migrieren in die lymphatischen Gewebe,
wo sie die prozessierten pathogenassoziierten Peptide mit Hilfe von
Haupthistokompatibilitätskomplexen (MHC) auf ihrer Oberfläche anderen Zellen präsentieren
(25).
Die Einteilung der DCs kann morphologisch, funktionell und über die Expression von
Zelloberflächenmolekülen in verschiedene Subpopulationen erfolgen (26). Sowohl im
EINLEITUNG
7
humanen als auch im murinen Modell sind sogenannte konventionelle (cDCs) und
plasmazytoide DCs (pDCs) beschrieben. Sie entwickeln sich beide aus hämatopoetischen
Stammzellen, allerdings bleiben die exakte Verwandtschaft und gegenseitige
Wechselwirkungen zwischen den beiden Populationen bis zu diesem Zeitpunkt ungeklärt
(26). cDCs haben eine sternförmige dendritische Zellform und üben die bisher beschriebenen
Funktionen von DCs aus, wie die Aufnahme, Prozessierung und Präsentation von Antigenen
(Abb. 5) (27). Sie werden durchflusszytometrisch als CD11c+, CD11b+ und B220-
charakterisiert (28). Hingegen zeichnen sich pDCs durch eine plasmazellähnliche
Morphologie (Abb. 5) und durch folgendes Oberflächenmolekül-Expressionsmuster aus:
CD11cniedrig, CD11b- und B220+ (29). Sie werden auch als natürliche Interferon-
produzierende Zellen bezeichnet, da sie große Mengen an IFN-α nach viraler Stimulation
freisetzen. Nach der Aktivierung differenzieren pDCs zu DCs mit antigenpräsentierender
Funktion (Abb. 5).
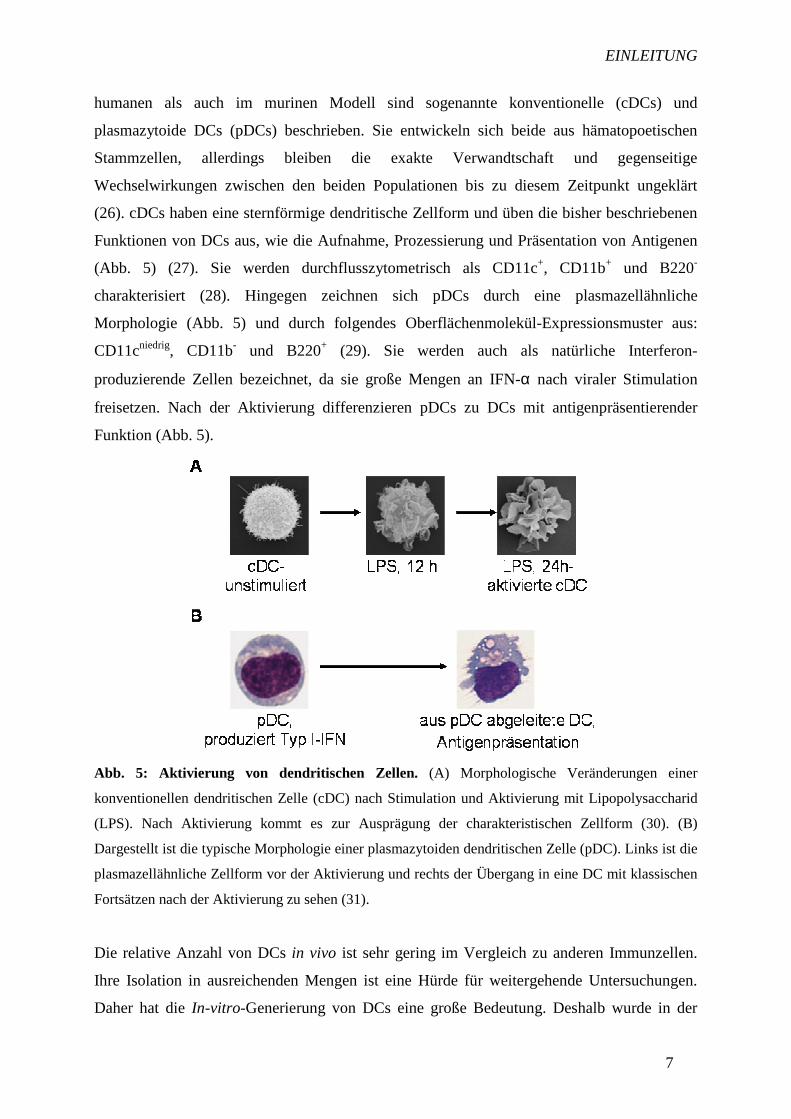
Abb. 5: Aktivierung von dendritischen Zellen. (A) Morphologische Veränderungen einer
konventionellen dendritischen Zelle (cDC) nach Stimulation und Aktivierung mit Lipopolysaccharid
(LPS). Nach Aktivierung kommt es zur Ausprägung der charakteristischen Zellform (30). (B)
Dargestellt ist die typische Morphologie einer plasmazytoiden dendritischen Zelle (pDC). Links ist die
plasmazellähnliche Zellform vor der Aktivierung und rechts der Übergang in eine DC mit klassischen
Fortsätzen nach der Aktivierung zu sehen (31).
Die relative Anzahl von DCs in vivo ist sehr gering im Vergleich zu anderen Immunzellen.
Ihre Isolation in ausreichenden Mengen ist eine Hürde für weitergehende Untersuchungen.
Daher hat die In-vitro-Generierung von DCs eine große Bedeutung. Deshalb wurde in der

EINLEITUNG
8
vorliegenden Arbeit mit DCs gearbeitet, welche aus murinen Knochenmarkszellen unter der
Einwirkung von fms-like tyrosine kinase-3-ligand (Flt3L) differenziert wurden. Dieser
hämatopoetische Wachstumsfaktor ist der einzige bisher bekannte Faktor zur Generierung
von pDCs (32, 33). Diese murinen Knochenmarks-DCs, generiert mit Flt3L, ähneln murinen
steady-state Milz-pDCs und -cDCs in vivo hinsichtlich der Expression von
Oberflächenmolekülen, Transkriptionsfaktoren, TLR, Chemokinrezeptoren, und TLR-
vermittelten Chemokin- und Zytokinantworten (29, 34, 35, 36). Daher sind diese
Knochenmarks-DCs als In-vitro-Modellsystem für die Untersuchungen mit iPPVO geeignet.
1.2.2 Erkennung von Viren durch dendritische Zellen über Mustererkennungs-
rezeptoren
Zellen des angeborenen Immunsystems haben die Aufgabe, Pathogene zu erkennen, zu deren
Eliminierung beizutragen und die adaptive Immunantwort zu initiieren. Für die Erkennung
von Pathogenen ist die Expression von Mustererkennungsrezeptoren (PRR) insbesondere auf
Makrophagen und DCs notwendig. Eine Klasse dieser PRR sind die transmembranen Toll-
ähnlichen Rezeptoren (TLR) (37). Die intrazelluläre Domäne enthält die sogenannte Toll/IL-
1-Rezeptor-homologe Domäne (TIR). Die extrazelluläre Domäne der TLRs beinhaltet
Leucin-reiche Wiederholungssequenzen (38). Für humanen TLR3 konnte anhand der
Kristallstruktur der Ektodomäne ein großes Hufeisen-förmiges Solenoid, gebildet von 23
Leucin-reichen Wiederholungen und stabilisiert durch Asparagine des 24. Leucin-Motivs,
nachgewiesen werden (39). Jeder der bisher 12 entdeckten TLR bei Säugern erkennt über
diese Sequenzen spezifisch pathogenassoziierte Liganden, wie Lipopeptid (TLR2), dsRNA
(TLR3), Lipopolysaccharid (TLR4) oder CpG-DNA (TLR9) (Tab. 1) (40). TLR3, TLR7 und
TLR9 zeichnen sich durch eine endosomale Lokalisation aus, alle anderen TLR befinden sich
auf der Zelloberfläche (37).
EINLEITUNG
9
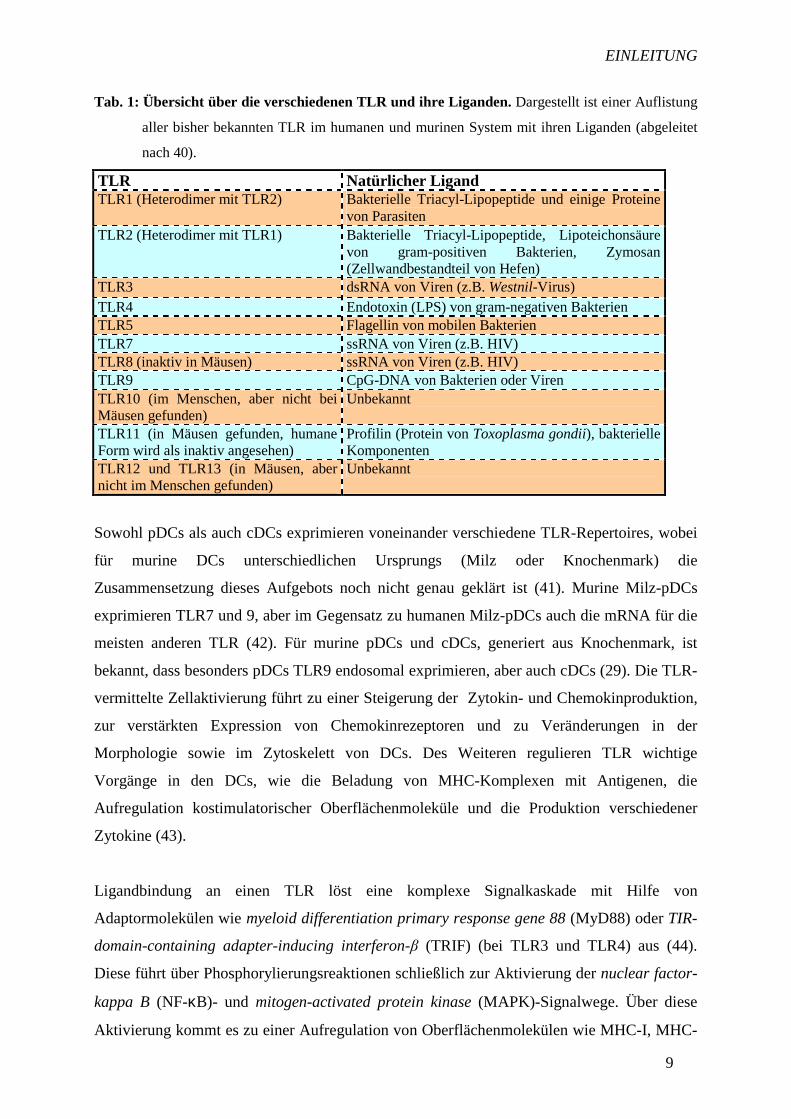
Tab. 1: Übersicht über die verschiedenen TLR und ihre Liganden. Dargestellt ist einer Auflistung
aller bisher bekannten TLR im humanen und murinen System mit ihren Liganden (abgeleitet
nach 40).
TLR Natürlicher Ligand TLR1 (Heterodimer mit TLR2) Bakterielle Triacyl-Lipopeptide und einige Proteine
von Parasiten TLR2 (Heterodimer mit TLR1) Bakterielle Triacyl-Lipopeptide, Lipoteichonsäure
von gram-positiven Bakterien, Zymosan (Zellwandbestandteil von Hefen)
TLR3 dsRNA von Viren (z.B. Westnil-Virus) TLR4 Endotoxin (LPS) von gram-negativen Bakterien TLR5 Flagellin von mobilen Bakterien TLR7 ssRNA von Viren (z.B. HIV) TLR8 (inaktiv in Mäusen) ssRNA von Viren (z.B. HIV) TLR9 CpG-DNA von Bakterien oder Viren TLR10 (im Menschen, aber nicht bei Mäusen gefunden)
Unbekannt
TLR11 (in Mäusen gefunden, humane Form wird als inaktiv angesehen)
Profilin (Protein von Toxoplasma gondii), bakterielle Komponenten
TLR12 und TLR13 (in Mäusen, aber nicht im Menschen gefunden)
Unbekannt
Sowohl pDCs als auch cDCs exprimieren voneinander verschiedene TLR-Repertoires, wobei
für murine DCs unterschiedlichen Ursprungs (Milz oder Knochenmark) die
Zusammensetzung dieses Aufgebots noch nicht genau geklärt ist (41). Murine Milz-pDCs
exprimieren TLR7 und 9, aber im Gegensatz zu humanen Milz-pDCs auch die mRNA für die
meisten anderen TLR (42). Für murine pDCs und cDCs, generiert aus Knochenmark, ist
bekannt, dass besonders pDCs TLR9 endosomal exprimieren, aber auch cDCs (29). Die TLR-
vermittelte Zellaktivierung führt zu einer Steigerung der Zytokin- und Chemokinproduktion,
zur verstärkten Expression von Chemokinrezeptoren und zu Veränderungen in der
Morphologie sowie im Zytoskelett von DCs. Des Weiteren regulieren TLR wichtige
Vorgänge in den DCs, wie die Beladung von MHC-Komplexen mit Antigenen, die
Aufregulation kostimulatorischer Oberflächenmoleküle und die Produktion verschiedener
Zytokine (43).
Ligandbindung an einen TLR löst eine komplexe Signalkaskade mit Hilfe von
Adaptormolekülen wie myeloid differentiation primary response gene 88 (MyD88) oder TIR-
domain-containing adapter-inducing interferon-β (TRIF) (bei TLR3 und TLR4) aus (44).
Diese führt über Phosphorylierungsreaktionen schließlich zur Aktivierung der nuclear factor-
kappa B (NF-κB)- und mitogen-activated protein kinase (MAPK)-Signalwege. Über diese
Aktivierung kommt es zu einer Aufregulation von Oberflächenmolekülen wie MHC-I, MHC-
EINLEITUNG
10
II, CD86, CD80 und CD40 und zur Produktion von proinflammatorischen Zytokinen (z.B. IL-
12/23p40 und TNF-α). Über die interferon regulatoy factors (IRF) 7 (MyD88-abhängig) und
IRF3 (TRIF-abhängig) wird die Produktion von Typ I-Interferonen (IFN) wie IFN-α induziert
(45, 46, 47). Eine vereinfachte Darstellung der Effektorantworten auf diese TLR-
Signalkaskade ist in Abb. 6 zu sehen.
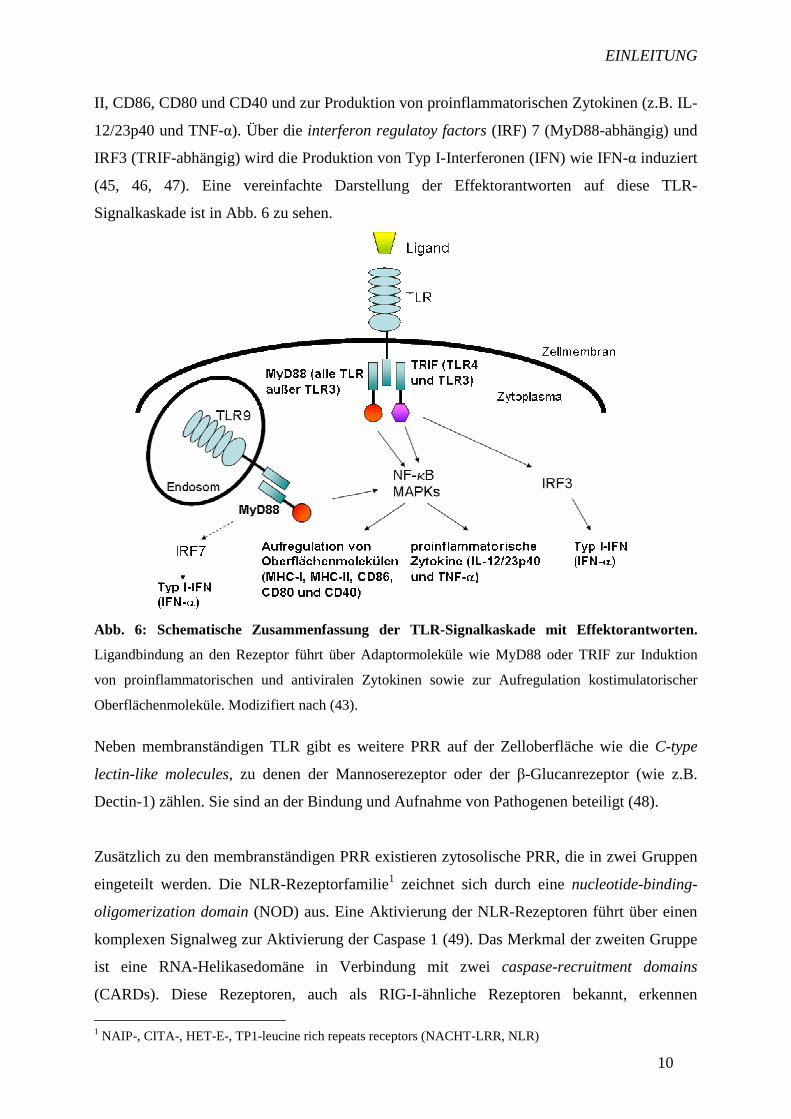
Abb. 6: Schematische Zusammenfassung der TLR-Signalkaskade mit Effektorantworten.
Ligandbindung an den Rezeptor führt über Adaptormoleküle wie MyD88 oder TRIF zur Induktion
von proinflammatorischen und antiviralen Zytokinen sowie zur Aufregulation kostimulatorischer
Oberflächenmoleküle. Modizifiert nach (43).
Neben membranständigen TLR gibt es weitere PRR auf der Zelloberfläche wie die C-type
lectin-like molecules, zu denen der Mannoserezeptor oder der β-Glucanrezeptor (wie z.B.
Dectin-1) zählen. Sie sind an der Bindung und Aufnahme von Pathogenen beteiligt (48).
Zusätzlich zu den membranständigen PRR existieren zytosolische PRR, die in zwei Gruppen
eingeteilt werden. Die NLR-Rezeptorfamilie1 zeichnet sich durch eine nucleotide-binding-
oligomerization domain (NOD) aus. Eine Aktivierung der NLR-Rezeptoren führt über einen
komplexen Signalweg zur Aktivierung der Caspase 1 (49). Das Merkmal der zweiten Gruppe
ist eine RNA-Helikasedomäne in Verbindung mit zwei caspase-recruitment domains
(CARDs). Diese Rezeptoren, auch als RIG-I-ähnliche Rezeptoren bekannt, erkennen
1 NAIP-, CITA-, HET-E-, TP1-leucine rich repeats receptors (NACHT-LRR, NLR)

EINLEITUNG
11
Zwischenprodukte der viralen Replikation, wie dsRNA. dsRNA kann direkt die Proteinkinase
R (PKR) aktivieren oder indirekt über Helikasen, kodiert vom retinoic-acid-inducible gene I
(RIG-I) oder melanoma-differentiation-associated gene 5 (MDA-5), erkannt werden. Diese
zytosolischen PRR ermöglichen auch cDCs, unabhängig von TLR7 und TLR9, Interferone
nach viraler Infektion zu produzieren (50, 51). Des Weiteren wurde kürzlich ein TLR9-
unabhängiger zytosolischer DNA-Rezeptor, DNA-dependent activator of IFN-regulatory
factors (DAI), beschrieben (52). Es existieren zunehmend Hinweise auf mögliche
Kooperationen der zytosolischen PRR mit den TLR. Die Entdeckung neuer Rezeptoren führt
zu einer immer größeren Vielfalt von Kandidatenrezeptoren, die für eine Erkennung von
Viren und anderen Pathogenen in Frage kommen.
1.2.3 Die antivirale Effektorantwort von dendritischen Zellen
Bei viralen Infektionen kommt es zur Aktivierung einer großen Anzahl von Immunantworten,
um die Replikation von Viren und die Ausbreitung der Infektion zu verhindern. Ein wichtiger
antiviraler Mechanismus ist das Typ I-Interferon-System. Hierzu zählen je nach Spezies bis
zu 14 IFN-α-Subtypen, 1 IFN-β und die noch relativ unbekannten IFN-ε, IFN-κ, IFN-ω, IFN-
δ, IFN-τ und IFN-ζ (53). Typ I-IFN verstärken die Expression der eIF2α-Kinase und 2’5’-
Oligoadenylatsynthetase, um die virale Replikation in der Wirtszelle zu verhindern. Weiterhin
stimulieren sie NK-Zellen, verstärken die DC-Aktivierung und unterstützen die Induktion der
adaptiven Immunantwort (43). Besonders pDCs sind in der Lage, sehr hohe Konzentrationen
von Typ I-IFN über TLR7 und TLR9 freizusetzen (43). pDCs antworteten mit einer starken
IFN-α-Sekretion auf die Stimulation mit dem behüllten DNA-Virus Herpex-Simplex-Virus I
(54). Virale Partikel des nicht-behüllten RNA-Virus der Maul- und Klauenseuche, welches
selbst nicht immunstimulatorisch wirkt, konnten im Komplex mit Immunglobulinen,
übertragen durch den Fcγ-Rezeptor II, die IFN-α-Produktion in pDCs auslösen (55). Bisher
konnte weiterhin gezeigt werden, dass die Typ I-Interferon-Produktion in pDCs unabhängig
vom RIG-I/MDA5-Signalweg geschieht (43). Allerdings gibt es auch zunehmend Hinweise
auf MyD88-unabhängige Mechanismen der Typ I-IFN-Produktion in pDCs. So ist sie in
pDCs, generiert aus Knochenmarkszellen, nach Applikation von Vaccinia-Viren (VV) nur
partiell TLR2-abhängig, ebenso scheint die IFN-β-Induktion MyD88-unabhängig zu sein
(56). Andere Zelltypen wie cDCs hingegen produzieren ebenfalls IFN-α als Reaktion auf
Virusstimulation oder transfizierte DNA/RNA, aber diese Produktion verläuft ausschließlich
über TLR7- und TLR9-unabhängige Signalwege, die sowohl die Proteinkinase R als auch die

EINLEITUNG
12
Helikasen RIG-1 und MDA-5, TLR3 und andere, zum Teil noch nicht identifizierte,
zytosolische Rezeptoren einschließen (50, 51). Auch der kürzlich beschriebene zytosolische
DNA-Rezeptor DAI könnte eine Rolle bei der Erkennung Viren-assoziierter Liganden durch
sowohl pDCs als auch cDCs spielen (52).
Neben Typ I-IFN produzieren sowohl cDCs als auch pDCs große Mengen an
proinflammatorischen Zytokinen wie den Tumornekrosefaktor-α (TNF-α) und Zytokine der
IL-12-Familie bei viralen Infektionen (23). TNF-α beeinflusst Wachstum, Differenzierung
und Funktion einer Vielzahl von Immunzellen wie DCs und Makrophagen. Zusätzlich
reguliert TNF-α Diffusionsvorgänge am Infektionsherd und führt zu einer gesteigerten
Freisetzung von Antikörpern, Komplement und Akutphaseproteinen (57). IL-12 und IL-23
besitzen eine heterodimere Struktur. Beide zeichnen sich durch die IL-12/23p40-Untereinheit
aus, IL-12 setzt sich weiterhin aus der p35 und IL-23 aus der p19-Untereinheit zusammen (58,
59, 60). Man kann von einer IL-12-Produktion ausgehen, wenn p40 im Überschuss vorliegt
(60). Deshalb kam in der vorliegenden Arbeit eine Detektion von IL-12 über die p40-
Untereinheit zum Einsatz. IL-12 stimuliert die Produktion von IFN-γ durch T-Zellen, NK-
Zellen und Makrophagen (58, 59). Weiterhin ist es essentiell für die Induktion und Erhaltung
einer Th1-vermittelten zellulären Immunantwort, welche für die Abwehr intrazellulärer
Erreger von Bedeutung ist. Es wurde dokumentiert, dass IL-23, aber nicht IL-12, die murinen
T-Gedächtniszellen zur Produktion des proinflammatorischen Zytokins IL-17 aktivieren
können (60).
Die Aktivierung von DCs als Folge von viralen Infektionen z.B. über TLR führt zu einer
Aufregulation von Oberflächenmolekülen wie MHC-I und MHC-II sowie den
kostimulatorischen Molekülen aus der B7-Familie CD86 und CD80 und außerdem CD40.
Über die Moleküle MHC-I und –II können zytosolische und endosomale Antigene präsentiert
werden (61). T-Zellen können mit Hilfe des T-Zellrezeptors mit den MHC-Peptid-Komplexen
auf der Oberfläche der antigenpräsentierenden Zelle interagieren. Zur Aktivierung der T-Zelle
sind jedoch kostimulatorische Moleküle auf antigenpräsentierenden Zellen wie CD86 und
CD80 notwendig, welche mit CD28 auf der T-Zelle wechselwirken (62). Verstärkt
exprimiertes CD40 auf DCs als Folge der DC-Aktivierung bewirkt mit dem CD40-Liganden
auf der T-Zelle eine Reorganisation des Zytoskeletts der T-Zelle und damit den Beginn der T-
Zellproliferation (63). Allerdings konnte in In-vitro-Experimenten auch eine Aktivierung von
T-Zellen ohne CD40 nachgewiesen werden (64).

EINLEITUNG
13
1.3 Zielstellung
Das Ziel dieser Arbeit besteht in der erstmaligen Charakterisierung der Aktivierung von DCs
durch iPPVO in vitro. Auf dem Hintergrund des veterinärmedizinischen Einsatzes von iPPVO
ist es erforderlich, neben den bisher untersuchten Zellen wie Makrophagen, NK-Zellen und
neutrophilen Granulozyten die für die antivirale und T-Zell-stimulierende Immunantwort
besonders wichtigen DCs zu charakterisieren. Diese sind die besten antigenpräsentierenden
Zellen für T-Zellen und somit auch für die adaptive Immunantwort von essentieller
Bedeutung. In der vorliegenden Arbeit sollten sowohl die Aufregulation der
Oberflächenmoleküle MHC-I, MHC-II sowie CD86, CD80 und CD40, als auch die
Produktion von IFN-α, IL-12/23p40 und TNF-α durch pDCs und cDCs nach Aktivierung mit
iPPVO untersucht werden.
Für iPPVO ist der Eintrittsmechanismus in Wirtszellen noch völlig ungeklärt. In dieser Arbeit
galt es zu analysieren, ob iPPVO-Virionen von DCs erkannt und aufgenommen werden. Als
erstes Experiment zu diesem Punkt wird daher die Aufnahme von PPVO und iPPVO durch
dendritische Zellen mit Hilfe von elektronenmikroskopischen Aufnahmen beschrieben.
Als weiterführende Zielstellung sollten TLR, die an der Erkennung der stimulatorische(n)
virale(n) Komponente(n) durch DCs beteiligt sind, identifiziert werden. Als mögliche
Kandidaten werden TLR9 und TLR2 untersucht. TLR9 erkennt dsDNA und ist somit ein
aussichtsreicher Rezeptorkandidat für ein dsDNA-Virus wie iPPVO (43). Weiterhin wird
TLR2 untersucht, da dieser bei der Aktivierung von DCs durch andere Pockenviren wie
Vaccinia-Viren (VV) eine zentrale Rolle spielt (56).

MATERIAL
14
2 Material
2.1 Tiere und Pathogene
2.1.1 Mäuse
C57BL/6J Wildtyp-Mäuse, C57BL/6J TLR9def (TLR9-/-) und C57BL/6J TLR2def
(TLR2-/-) Mäuse wurden in der Tieranlage an der Technischen Universität München unter
spezifisch pathogenfreien Bedingungen gezüchtet. C57BL/6J Wildtyp-Mäuse wurden am
Institut für Immunologie, Veterinärmedizinische Fakultät der Universität Leipzig, und am
Max-Planck-Institut für Evolutionäre Anthropologie Leipzig im Einklang mit den vom
Tierschutzausschuss des Regierungspräsidiums Leipzig festgelegten Richtlinien gehalten. Für
das Tierversuchsvorhaben lag eine Genehmigung des Regierungspräsidiums Leipzig mit dem
Az 24-9162.11-T17/07 vor. Für alle Experimente kamen weibliche Mäuse im Alter von 8 bis
16 Wochen zum Einsatz. Futter und Wasser wurden ad libitum verabreicht.
2.1.2 Parapoxvirus ovis
Chemisch inaktiviertes Parapoxvirus ovis (iPPVO) wurde freundlicherweise von Dr. Rüdiger
Raue (Pfizer Ltd., Sandwich, GB) und Prof. Mathias Büttner (Institut für Immunologie,
Wissenschaftszentrum für Virusinfektion bei Tieren, Tübingen) zur Verfügung gestellt.
2.2 Zelllinien
Murine P388-Zellen für die Etablierung einer Lebend-/Tot-Färbung mit
Ethidiummonoazidbromid (EMA) wurden freundlich durch Bernd Walloschke, Veterinär-
Physiologisch-Chemisches Institut der Veterinärmedizinischen Fakultät, Universität Leipzig,
zur Verfügung gestellt. Alle Zellen wurden in flüssigem Stickstoff gelagert und regelmäßig
auf Mycoplasma spp. getestet.
MATERIAL
15
2.3 Wachstumsfaktoren
Humaner fms-like tyrosine kinase-3-ligand (Flt3L) stellte freundlicherweise Amgen, Seattle,
USA zur Verfügung. Muriner Flt3L wurde von Tim Sparwasser (Institut für Medizinische
Mikrobiologie, Immunologie und Hygiene, Technische Universität München), bereitgestellt.
2.4 Antikörper
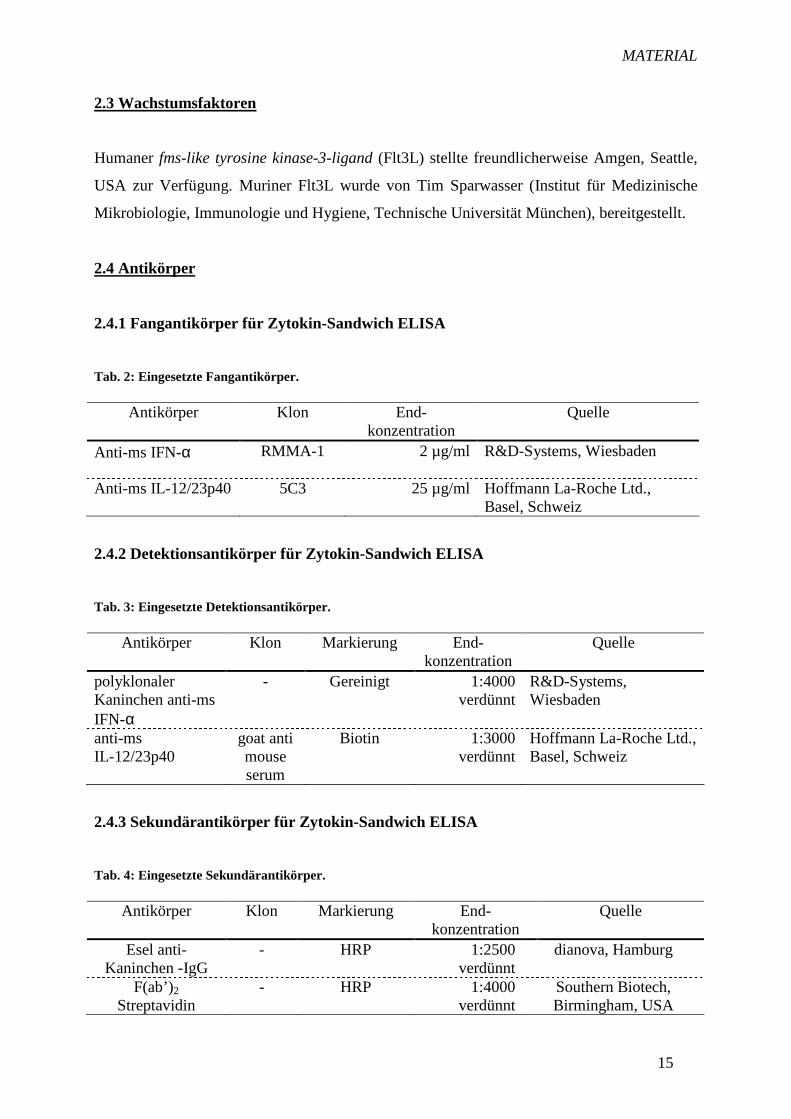
2.4.1 Fangantikörper für Zytokin-Sandwich ELISA
Tab. 2: Eingesetzte Fangantikörper.
Antikörper Klon End-konzentration
Quelle
Anti-ms IFN-α RMMA-1
2 µg/ml R&D-Systems, Wiesbaden
Anti-ms IL-12/23p40
5C3 25 µg/ml Hoffmann La-Roche Ltd., Basel, Schweiz
2.4.2 Detektionsantikörper für Zytokin-Sandwich ELISA
Tab. 3: Eingesetzte Detektionsantikörper.
Antikörper Klon Markierung End-konzentration
Quelle
polyklonaler Kaninchen anti-ms IFN-α
- Gereinigt 1:4000 verdünnt
R&D-Systems, Wiesbaden
anti-ms IL-12/23p40
goat anti mouse serum
Biotin 1:3000 verdünnt
Hoffmann La-Roche Ltd., Basel, Schweiz
2.4.3 Sekundärantikörper für Zytokin-Sandwich ELISA
Tab. 4: Eingesetzte Sekundärantikörper.
Antikörper Klon Markierung End-konzentration
Quelle
Esel anti- Kaninchen -IgG
-
HRP
1:2500 verdünnt
dianova, Hamburg
F(ab’)2 Streptavidin
- HRP 1:4000 verdünnt
Southern Biotech, Birmingham, USA
MATERIAL
16
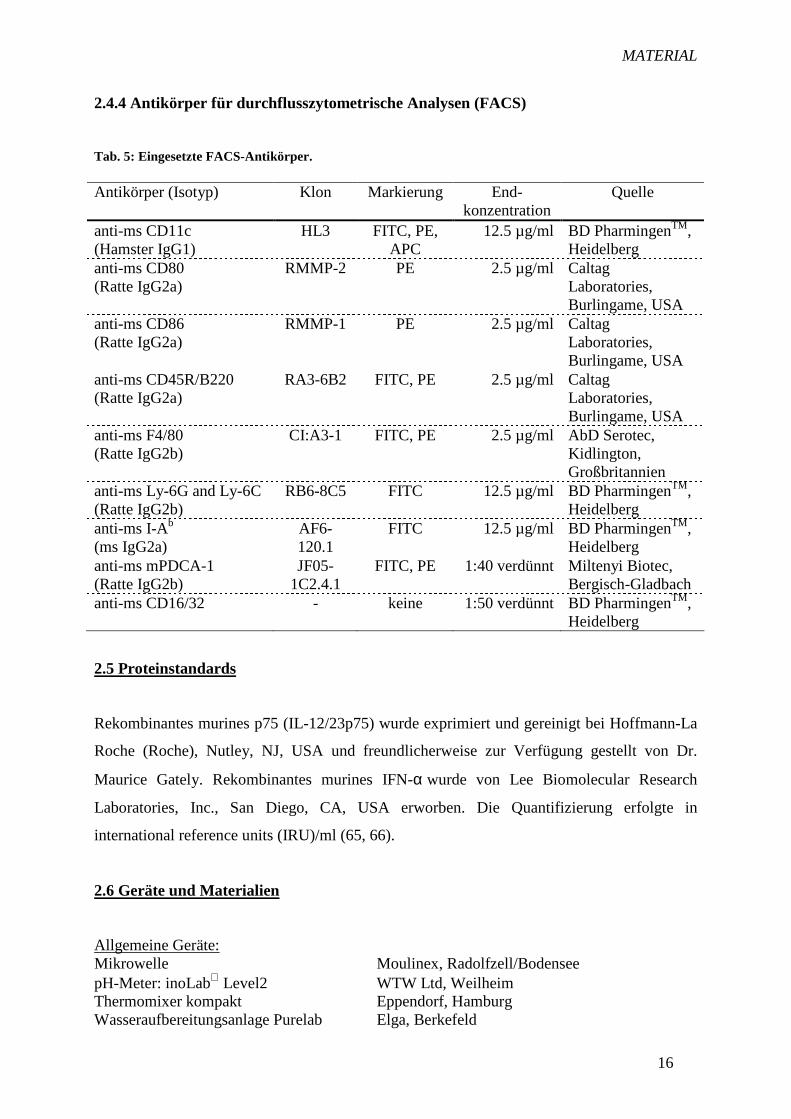
2.4.4 Antikörper für durchflusszytometrische Analysen (FACS)
Tab. 5: Eingesetzte FACS-Antikörper. Antikörper (Isotyp) Klon Markierung End-
konzentration Quelle
anti-ms CD11c (Hamster IgG1)
HL3 FITC, PE, APC
12.5 µg/ml BD PharmingenTM, Heidelberg
anti-ms CD80 (Ratte IgG2a)
RMMP-2 PE 2.5 µg/ml Caltag Laboratories, Burlingame, USA
anti-ms CD86 (Ratte IgG2a)
RMMP-1 PE 2.5 µg/ml Caltag Laboratories, Burlingame, USA
anti-ms CD45R/B220 (Ratte IgG2a)
RA3-6B2 FITC, PE 2.5 µg/ml Caltag Laboratories, Burlingame, USA
anti-ms F4/80 (Ratte IgG2b)
CI:A3-1 FITC, PE 2.5 µg/ml AbD Serotec, Kidlington, Großbritannien
anti-ms Ly-6G and Ly-6C (Ratte IgG2b)
RB6-8C5 FITC 12.5 µg/ml BD PharmingenTM, Heidelberg
anti-ms I-Ab (ms IgG2a)
AF6-120.1
FITC 12.5 µg/ml BD PharmingenTM, Heidelberg
anti-ms mPDCA-1 (Ratte IgG2b)
JF05-1C2.4.1
FITC, PE 1:40 verdünnt Miltenyi Biotec, Bergisch-Gladbach
anti-ms CD16/32 - keine 1:50 verdünnt BD PharmingenTM, Heidelberg
2.5 Proteinstandards
Rekombinantes murines p75 (IL-12/23p75) wurde exprimiert und gereinigt bei Hoffmann-La
Roche (Roche), Nutley, NJ, USA und freundlicherweise zur Verfügung gestellt von Dr.
Maurice Gately. Rekombinantes murines IFN-α wurde von Lee Biomolecular Research
Laboratories, Inc., San Diego, CA, USA erworben. Die Quantifizierung erfolgte in
international reference units (IRU)/ml (65, 66).
2.6 Geräte und Materialien
Allgemeine Geräte: Mikrowelle Moulinex, Radolfzell/Bodensee pH-Meter: inoLab Level2 WTW Ltd, Weilheim Thermomixer kompakt Eppendorf, Hamburg Wasseraufbereitungsanlage Purelab Elga, Berkefeld

MATERIAL
17
Wasserbad: Isotemp 205 Scientific Support, Hayward, CA, USA Allgemeine Materialien: 0,7 ml Reaktionsgefäße (aus PP) Becton Dickinson, Franklin Lakes, NJ, USA 1,5 ml Reaktionsgefäße (aus PP) Becton Dickinson, Franklin Lakes, NJ, USA 2,0 ml Reaktionsgefäße (aus PP) Becton Dickinson, Franklin Lakes, NJ, USA 15 ml Zentrifugenröhrchen (steril, aus PP) Becton Dickinson, Franklin Lakes, NJ, USA 50 ml Zentrifugenröhrchen (steril, aus PP) Becton Dickinson, Franklin Lakes, NJ, USA Injektionsnadeln (27 G) Neolis®, Terumo ®, Leuven, Belgien Pipetten und Pipettenspitzen Eppendorf, Hamburg Reaktionsgefäß Phase Lock Gen I heavy Eppendorf, Hamburg Spritzen Becton Dickinson, Franklin Lakes, NJ, USA Sterilfilter(0,2 µm; 0,5 µm) Sartorius AG, Göttingen Durchflusszytometrie: FACS Calibur Becton Dickinson, Heidelberg 96er Spitzbodenplatten NUNC, Wiesbaden Durchlichtmikroskop: Axiovert 25 Carl-Zeiss, Jena Axioskop 2 plus Carl-Zeiss, Jena ELISA: ELISA-Reader: Spectra-max 340 Molecular Devices, München ELISA-Waschgerät (Ultrawash PLUS) DYNEX Technologies Inc., Chantilly, VA, USA 96er Immunoplatten Maxisorb NUNC, Wiesbaden Gelelektrophoresezubehör: DNA-Sub Cell™ Elektrophoresekammer Bio-Rad Laboratories, Hercules, CA, USA Stromversorgungsgerät 200/2.0 Bio-Rad Laboratories, Hercules, CA, USA UV-Transilluminator MWG-Biotech, Ebersberg Waagen: Mettler PM 4000 Mettler Toledo GmbH, Giessen Mettler AB184-S-A Mettler Toledo GmbH, Giessen Zellkultur: Benchtop Inkubator GFL 3031 Gesellschaft für Labortechnik mbH, Burgwedel CO2-Inkubator für Zellkultur Heraeus, Osterode Neubauer-Zählkammer Fuchs-Rosenthal, Bad Blankenburg Sterilbank: HERA-safe Heraeus, Osterode Zellkulturflaschen TPP®, Trasadingen, Schweiz Zellkulturplatten TPP®, Trasadingen, Schweiz Zellschaber, steril Costar®, Cambridge, MA, USA Zentrifugen: Megafuge 2.0 R Heraeus, Osterode Multifuge 3 S-R Heraeus, Osterode Tischzentrifuge TOMY Tech., Framont, USA Zentrifuge 5417C Eppendorf, Hamburg Zentrifuge 5417R Eppendorf, Hamburg

MATERIAL
18
2.7 Software
BD CellQuest™ pro Becton Dickinson, Heidelberg; Analyse und Auswertung durchflusszytometrischer Daten
Graph Pad Prism™ 4 GraphPad Sofware, San Diego, CA, USA;
Statistische Auswertung und graphische Darstellung von Daten
Reference Manager Professional 10/11 Thomson Research Soft, Carlsbad, CA, USA; Verwaltung von Referenzen und Literatur Softmax Pro 3.1.2 /5.0.0 Molecular Devices, Sunnyvale, CA, USA
Analyse und Auswertung von ELISA-Daten
Microsoft® Office 2002/2003 /XP Microsoft, Redmond, WA, USA Präsentation von Daten, Anfertigung schriftlicher Dokumente, Anfertigung von Abbildungen, Bearbeitung von Daten
2.8 Chemikalien, Lösungen, Puffer und Nährmedien
2.8.1 Allgemeine Chemikalien
Agarose Invitrogen, Karlsruhe
β-Merkaptoethanol Sigma-Aldrich, Taufkirchen
Borsäure Sigma-Aldrich, Taufkirchen
Bromphenolblau Sigma-Aldrich, Taufkirchen
BSA Sigma-Aldrich, Taufkirchen
CpG-ODN 22162 TIB MOLBIOL Syntheselabor GmbH, Berlin
CaCl2 Carl Roth GmbH, Karlsruhe
Chloroform Carl Roth GmbH, Karlsruhe
DNA-Probenpuffer ICN Biomedicals, Aurora, Ohio, USA
DMSO Sigma-Aldrich, Taufkirchen
EDTA (Natriumsalz) Merck, Darmstadt
Ethanol Carl Roth GmbH, Karlsruhe
Ethidiumbromid Sigma-Aldrich, Taufkirchen
2 CpG-ODN 2216 mit der Sequenz 5’-GsGsGGGACGATCGTCsGsGsGsGsGsG-3’ (s = Phosphothioat)

MATERIAL
19
Ethidiummonoazidbromid Invitrogen, Karlsruhe
FKS PAA Laboratories GmbH, Cölbe
Formaldehyd (37 Vol %) Carl Roth GmbH, Karlsruhe
Gelatine (Schwein) Merck, Darmstadt
Glycerol Sigma-Aldrich, Taufkirchen
Glukose Sigma-Aldrich, Taufkirchen
L-Glutamin (200 mM) Sigma-Aldrich, Taufkirchen
HEPES-Lösung, pH 7,4 PAA Laboratories, Cölbe
Isoamylalkohol Carl Roth GmbH, Karlsruhe
KCl Merck, Darmstadt
KH2PO4 Fluka-Chemie Ag, Buchs
K2HPO4 Carl Roth GmbH, Karlsruhe
LPS (von Salmonella Abortus equi, S-Form) Alexis, Grünberg
MgCl2 Promega, Mannheim
NaCl Carl Roth GmbH, Karlsruhe
Na2CO3 Carl Roth GmbH, Karlsruhe
NaHCO3 Sigma-Aldrich, Taufkirchen
NaOH Carl Roth GmbH, Karlsruhe
Na2HPO4 Carl Roth GmbH, Karlsruhe
Natriumacetat Carl Roth GmbH, Karlsruhe
Natriumpyruvat Sigma-Aldrich, Taufkirchen
Naphtylethylendiamid-Dihydrochlorid Sigma-Aldrich, Taufkirchen
Penicillin/Streptomycin Sigma-Aldrich, Taufkirchen
Phenol Sigma-Aldrich, Taufkirchen
Polyinosinische–Polycytidylische Säure,
Kalium-Salz, (poly I:C) Sigma-Aldrich, Taufkirchen
R-848 (Resiquimod) Alexis, Grünberg
SDS Sigma-Aldrich, Taufkirchen
Sukrose Sigma-Aldrich, Taufkirchen
Sulphanilamid Sigma-Aldrich, Taufkirchen
Tris Base AppliChem GmbH, Darmstadt
Trypanblau Sigma-Aldrich, Taufkirchen
Trypton Carl Roth GmbH, Karlsruhe
TWEEN 20 Sigma-Aldrich, Taufkirchen

MATERIAL
20
2.8.2 Puffer und Lösungen für Zellisolation
1x PBS: 0.80 % (w/v) NaCl 0.02 % (w/v) KCl 0.23 % (w/v) Na2HPO4 x 2 H2O 0.02 % (w/v) KH2PO4 pH = 7.4 PBS / FKS:
10 % (v/v) FKS in 10 % 10x PBS pH = 7.4; steril filtriert
Gey´s-Lösung (für die Lyse von Erythrozyten): Lsg.A: 0.65 M NH4Cl
24.80 mM KCl 4.20 mM Na2HPO4
0.81 mM KH2PO4 25.23 mM Glukose 0.14 mM Phenolrot Lsg.B: 0.42 % (w/v) MgCl2 x 6 H2O 0.07 % (w/v) MgSO4
Lsg.C: 2.25 % (w/v) NaHCO3
Arbeitslösung: 20 % (v/v) Lsg. A, 5 % (v/v) Lsg. B and 5 % (v/v) Lsg. C
2.8.3 Puffer und Lösungen für Durchflusszytometrie
Ethidiummonoazidbromid: 1.0 % (w/v) EMA in 97 % (v/v) Ethanol
FACS-Puffer (Wasch-und Verdünnungspuffer):
3.0 % (v/v) FKS 0.1 % (w/v) NaN3 in 1x PBS
Fixierungspuffer: 1.48 % (v/v) Formaldehyd in 1x PBS
2.8.4 Puffer und Lösungen für ELISA
10x PBS: 0.35 % (w/v) KH2PO4 1.58 % (w/v) Na2HPO4 (wasserfrei) 8.50 % (w/v) NaCl Carbonatpuffer: 1.73 % (w/v) NaHCO3 0.86 % (w/v) Na2CO3 pH = 9.5

MATERIAL
21
Blockpuffer: 0.1 % (w/v) Gelatine (Schwein) 0.5 % (w/v) BSA in 1x PBS Serumdiluent: 0.10 % (w/v) Gelatine (Schwein) 0.05 % (v/v) Tween 20 in 1x PBS Waschpuffer:
0.05 % (v/v) Tween 20 in 1x PBS Substratpuffer: TMB-Peroxidase-Substrat KPL, Medac, Wedel, Germany Arbeitslösung: Lsg. A und Lsg. B: 1:1 2.8.5 Medien und Lösungen für Zellkultur
Soweit nicht anders vermerkt, wurden alle Zellkulturreagenzien von PAA-Laboratorien,
Cölbe, erworben.
Kulturmedium zur Differenzierung einer Mischkultur von plasmazytoiden und konventionellen BMDC (Flt3L-Kulturmedium): 10 % (v/v) FKS 1.0 % (v/v) Penicillin/Streptomycin (104 U/ml, 104 µg/ml)
1.0 % (v/v) L-Glutamin (200 mM) 0.2 % (v/v) Vitamine 0.2 % (v/v) essentielle und nicht-essentielle
Aminosäuren 300 ng/ml huFlt3L
50 µM β-Merkaptoethanol
(0.175 %ig) in RPMI1640
Kulturmedium zur Kultivierung von P388-Zellen (P388-Medium): 10 % (v/v) FKS 1.0 % (v/v) Penicillin/Streptomycin
(104 U/ml, 104 µg/ml) 1.0 % (v/v) L-Glutamin (200 mM) in RPMI1640 Cryomedium für Zelllinien:
10 % (v/v) DMSO in FKS

MATERIAL
22
2.8.6 Kits
Venor GeM Mycoplasma detection kit Minerva Biolabs GmbH, Berlin
Murine TNF-α ELISA Development kit Pepro Tech Ec Ltd., London, GB
2.8.7 DNA-Standards
10 kb DNA Ladder Invitrogen, Karlsruhe
2.8.8 Puffer und Lösungen für die Isolierung viraler DNA
Alle Lösungen wurden in DNase-freiem aqua bidest. (2x autoklaviert) hergestellt.
Ethanol: 70.0 % (v/v) Ethanol unvergällt IAC-Lösung:
24 x Volumenanteile Chloroform 1 x Volumenanteil Isoamylalkohol
Natriumacetatlösung (pH 8.4): 30.0 % (w/v) Natriumacetat Proteinase K (1mg/ml in 1,5 mM CaCL2) SDS-Lösung: 20.0 % (w/v) SDS TEN-Puffer: 0.35 % (w/v) KH2PO44 1.58 % (w/v) Na2HPO4 (wasserfrei) 8.50 % (w/v) NaCl TE-Puffer pH 8.0: 0.35 % (w/v) KH2PO44 1.58 % (w/v) Na2HPO4 (wasserfrei) 8.50 % (w/v) NaCl 2.8.9 Puffer und Lösungen für die Agarose-Gelelektrophorese
Agarose: 0.80 % (w/v) Agarose in TBE-Puffer Ethidiumbromid-Lösung: 5 µg/ml in TBE-Puffer

MATERIAL
23
Ladepuffer (6x): 0.25 % (w/v) Bromphenolblau 40.0 % (w/v) Sukrose
in aqua bidest. TBE-Puffer: 10.8 % (w/v) Tris HCl 5.5 % (w/v) Borsäure 0.72 % (w/v) EDTA

METHODEN
24
3 Methoden
3.1 Zellbiologische Methoden
3.1.1 Kultivierung von P388-Zellen für die Etablierung der Lebend-/Tot-Färbung mit
Eithidiummonoazidbromid
Zur Etablierung einer Lebend-/Tot-Färbung mit Eithidiummonoazidbromid (EMA) wurden
wegen besserer Verfügbarkeit Zellen der murinen Zellinie P388 verwendet. EMA wird nur
bei bereits perforiert vorliegender Zellmembran in die Zelle aufgenommen. Über
Photocrosslinking bindet EMA kovalent an die DNA und kann daraufhin mittels
durchflusszytometrischer Analyse nachgewiesen werden.
Die Inkubation der P388-Zellen erfolgte in P388-Medium bei 37 °C und 5 % CO2 mit einer
Dichte von 1.5x106 Zellen/ml für 5 Tage. Anschließend mussten die adhärenten Zellen mit
einem Zellschaber von der Oberfläche der Zellkulturflaschen gelöst und bei 1200 rpm für 8
min bei RT zentrifugiert werden. Der Überstand wurde verworfen, das Zellpellet in P388-
Medium resuspendiert und die Zellen mit Trypanblau in der Neubauerzählkammer
quantifiziert. Danach wurden 2x106 Zellen in 1 ml P388-Medium gegeben. Die Inkubation
der Zellen für die EMA-Positivkontrolle erfolgte für 10 min bei 60 °C mit anschließender
Überprüfung der Zellvitalität mit der Trypanblaufärbung. Zu den Proben wurden 10 µl der
EMA-Arbeitslösung gegeben und für 10 min auf Eis und anschließend 15 min unter
Lichteinfluss inkubiert. Nach der folgenden Zentrifugation bei 300 g für 8 min bei 4 °C fand
eine Resuspension des Zellpellets in 30 µl FACS-Puffer und eine Verteilung der
Zellsuspension auf jeweils 5 µl/FACS-Ansatz statt. Die P388-Zellen wurden dabei
durchflusszytometrisch neben der Vitalität auf das Makrophagen-spezifische
Oberflächenmolekül F4/80 (FITC) und CD86 (PE) untersucht. Als Negativkontrolle wurden
Zellen ohne Inkubation mit EMA verwendet.
3.1.2 Generierung von dendritischen Zellen aus dem Knochenmark
Knochenmarkszellen differenzieren zu einer Mischkultur aus pDCs und cDCs unter der
Einwirkung von Flt3L. Daher wurden die Oberschenkelknochen von C57BL/6J WT-,
TLR2def- und TLR9def-Mäusen entnommen und das Knochenmark mit einer PBS-Lösung mit

METHODEN
25
zusätzlich 5 % FKS mit Hilfe einer Spritze mit Nadel von 27 Gauge Durchmesser ausgespült.
Die Zellen wurden anschließend in Flt3L-Kulturmedium mit einer Zelldichte von 1.5x106
Zellen/ml für 7 Tage bei 37 °C in einer befeuchteten Atmosphäre mit 5 % CO2 inkubiert. Die
nicht-adhärenten Zellen wurden aus den Zellkulturflaschen entnommen und bei 300 g für 8
min bei RT zentrifugiert, in Medium resuspendiert und mit einer Konzentration von 5x105
Zellen/ml in 24er Flachbodenplatten (0.9 ml/Kavität) eingesät.
3.1.3 Stimulation von dendritischen Zellen
Alle Stimulationslösungen wurden in Flt3L-Kulturmedium auf Eis verdünnt und in Volumen
von 100 µl zugegeben. iPPVO wurde mit MOI = 5 eingesetzt. Als Kontrollstimuli wurden
synthetische oder natürliche Liganden der einzelnen TLR verwendet, wie in Tab. 6
angegeben. Die Stimulation der Zellen erfolgte, wenn nicht anders angegeben für 48 h bei 37
°C und 5 % CO2.
Tab. 6: Zellstimuli und ihre Konzentrationen. Als Zellstimuli kamen iPPVO und spezifische,
natürliche oder synthetische TLR-Liganden zum Einsatz.
Stimulus Ausgangs-
konzentration
Endkonzentration
iPPVO 1.78E8 VP/ml 2.5E6 VP/ml
CpG-ODN 2216 (TLR9) 100 µM 1 µM
R-848 (TLR7) 1 mg/ml 1 µg/ml
LPS (TLR4) 1 mg/ml 5 µg/ml
Pam3CSK4 (TLR2) 1 mg/ml 2 µg/ml
Poly-IC (TLR3) 1 mg/ml 100 µg/ml
3.2 Durchflusszytometrische Analysen
Die Durchflusszytometrie ist eine geeignete Methode, um die Expression von
Oberflächenmolekülen auf den Zellen zu untersuchen. Dabei können murine pDCs und cDCs
aufgrund der Expression der Oberflächenmoleküle CD11c (Integrin) und B220
(Tyrosinphosphatase) unterschieden und separat analysiert werden. Tote Zellen wurden durch
eine Lebend-/Tot-Färbung mit EMA (Abschnitt 3.1) von der Analyse ausgeschlossen. Für
jede gefärbte Probe wurde eine Isotypkontrolle angesetzt, um unspezifische Bindungen der

METHODEN
26
Antikörper an die Zellen zu erfassen. Weiterhin können IgG-Antikörper nicht nur mit dem
gewünschten Oberflächenantigen reagieren, sondern auch mit Fcγ-Rezeptoren, so dass falsch
positive Resultate erscheinen. Daher kam ein spezieller Fcγ-Block zum Einsatz.
Alle Inkubationen zur FACS-Färbung wurden bei 4-8 °C durchgeführt. Alle
durchflusszytometrischen Analysen erfolgten am FACScalibur Durchflusszytometer.
Nach der Stimulation der Zellen wurden diese mit dem Überstand in 1,5 ml Reaktionsgefäße
überführt und bei 300 g und 4 °C für 8 min zentrifugiert. Die Resuspension des Zellpellets
erfolgte in 0,99 ml PBS. Anschließend wurden die Proben mit 10 µl EMA-Arbeitslösung mit
folgendem mehrfachen Invertieren der Proben versetzt. Als Negativkontrolle dienten dabei
Zellen, die mit Medium stimuliert und in 1 ml PBS resuspendiert wurden ohne Zugabe von
EMA-Arbeitslösung. Die Herstellung einer Positivkontrolle erfolgte unter Hitzeinkubation
von mit Medium stimulierten Zellen für 10 min bei 60 °C und anschließender Überprüfung
der Vitalität mit Trypanblau-Färbung. Diese Positivkontrollee wurde ebenfalls mit EMA-
Arbeitslösung versetzt, für 10 min im Inkubator bei 37 °C und 5 % CO2 und danach für 15
min mit geöffnetem Verschluss unter Lichteinfluss in der Sterilbank inkubiert. Nach erneuter
Zentrifugation der Proben bei 300 g, 8 min, 4 °C wurde der Überstand verworfen und die
Zellen in Fcγ-Block (Anti-FcγR-AK (1:50); CD16/32) nach den Empfehlungen des
Herstellers resuspendiert, um eine unspezifische Bindung der AK an den Fcγ-Rezeptor zu
verhindern. Pro FACS-Färbung wurden 5 µl Suspension in 96-Kavitätenplatten pipettiert. Zu
den 5 µl Zellsuspension erfolgte die Zugabe von 5 µl der bereits hergestellten Färbelösung
(Tab. 7). Anschließend wurden die Platten kurz anzentrifugiert und 20 min bei 4 °C vor Licht
geschützt inkubiert. Nach dreimaligem Waschen mit 100 µl/Kavität FACS-Puffer mit
dazwischen liegender Zentrifugation für 2 min bei 300 g und 4 °C und Verwerfen des
Überstandes wurden 100 µl/Kavität FACS-Fixierungspuffer hinzugefügt und die Platten
verschlossen und bei 4 °C bis zur durchflusszytometrischen Analyse im Dunkeln aufbewahrt.
Zur Messung der Vierfarben-Immunfluoreszenz (FITC, PE, EMA und APC) musste auf
Grund überlappender Emissionsbereiche die Detektoren für die 4 Fluoreszenzen
gegeneinander kompensiert werden. Zur Einstellung der Kompensation dienten einzelne
Proben von Zellen, die jeweils nur mit einem Antikörper gefärbt wurden.
METHODEN
27
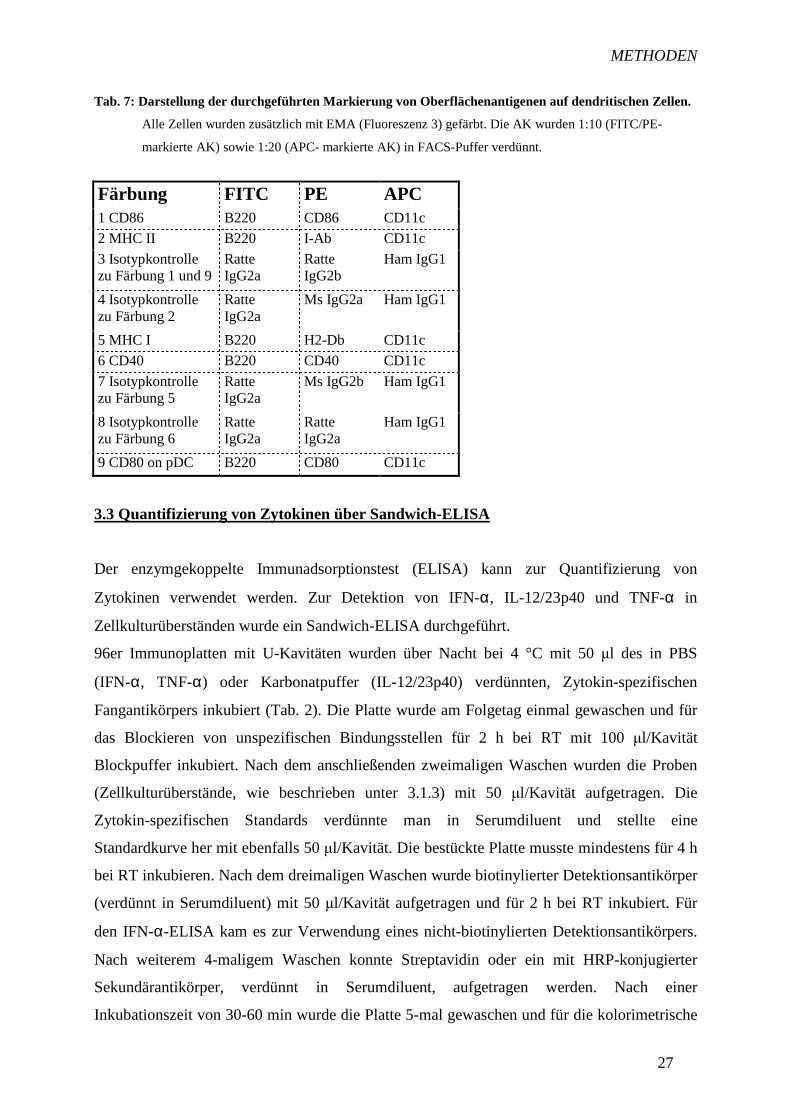
Tab. 7: Darstellung der durchgeführten Markierung von Oberflächenantigenen auf dendritischen Zellen.
Alle Zellen wurden zusätzlich mit EMA (Fluoreszenz 3) gefärbt. Die AK wurden 1:10 (FITC/PE-
markierte AK) sowie 1:20 (APC- markierte AK) in FACS-Puffer verdünnt.
Färbung FITC PE APC 1 CD86 B220 CD86 CD11c
2 MHC II B220 I-Ab CD11c
3 Isotypkontrolle zu Färbung 1 und 9
Ratte IgG2a
Ratte IgG2b
Ham IgG1
4 Isotypkontrolle zu Färbung 2
Ratte IgG2a
Ms IgG2a Ham IgG1
5 MHC I B220 H2-Db CD11c
6 CD40 B220 CD40 CD11c
7 Isotypkontrolle zu Färbung 5
Ratte IgG2a
Ms IgG2b Ham IgG1
8 Isotypkontrolle zu Färbung 6
Ratte IgG2a
Ratte IgG2a
Ham IgG1
9 CD80 on pDC B220 CD80 CD11c
3.3 Quantifizierung von Zytokinen über Sandwich-ELISA
Der enzymgekoppelte Immunadsorptionstest (ELISA) kann zur Quantifizierung von
Zytokinen verwendet werden. Zur Detektion von IFN-α, IL-12/23p40 und TNF-α in
Zellkulturüberständen wurde ein Sandwich-ELISA durchgeführt.
96er Immunoplatten mit U-Kavitäten wurden über Nacht bei 4 °C mit 50 µl des in PBS
(IFN-α, TNF-α) oder Karbonatpuffer (IL-12/23p40) verdünnten, Zytokin-spezifischen
Fangantikörpers inkubiert (Tab. 2). Die Platte wurde am Folgetag einmal gewaschen und für
das Blockieren von unspezifischen Bindungsstellen für 2 h bei RT mit 100 µl/Kavität
Blockpuffer inkubiert. Nach dem anschließenden zweimaligen Waschen wurden die Proben
(Zellkulturüberstände, wie beschrieben unter 3.1.3) mit 50 µl/Kavität aufgetragen. Die
Zytokin-spezifischen Standards verdünnte man in Serumdiluent und stellte eine
Standardkurve her mit ebenfalls 50 µl/Kavität. Die bestückte Platte musste mindestens für 4 h
bei RT inkubieren. Nach dem dreimaligen Waschen wurde biotinylierter Detektionsantikörper
(verdünnt in Serumdiluent) mit 50 µl/Kavität aufgetragen und für 2 h bei RT inkubiert. Für
den IFN-α-ELISA kam es zur Verwendung eines nicht-biotinylierten Detektionsantikörpers.
Nach weiterem 4-maligem Waschen konnte Streptavidin oder ein mit HRP-konjugierter
Sekundärantikörper, verdünnt in Serumdiluent, aufgetragen werden. Nach einer
Inkubationszeit von 30-60 min wurde die Platte 5-mal gewaschen und für die kolorimetrische

METHODEN
28
Bestimmung der Zytokinkonzentrationen 100 µl des Substrates TMB (Microwell Peroxidase
System) eingesetzt. Das Messen der ELISA-Platten erfolgte anschließend am Spectra-max
340 ELISA reader bei 650 nm und 480 nm.
3.4 Generierung und Stimulation von dendritischen Zellen für elektronen-
mikroskopische Aufnahmen
Für die elektronenmikroskopischen Aufnahmen wurde eine Mischkultur aus pDCs und cDCs
mit huFlt3L aus murinem Knochenmark generiert, wie in Abschnitt 3.1.2 beschrieben.
Anschließend erfolgte analog Abschnitt 3.1.3 die Stimulation der Zellen für 6, 12, 24 und 48 h
mit iPPVO und Kontrollstimulus CpG-ODN. Dabei wurden zum einen Stimulationsansätze
von 1x106 Zellen in 2 ml mit 200 µl Stimulus-Lösung für die elektronenmikroskopischen
Aufnahmen durchgeführt. Zum zweiten erfolgte die Stimulation für die FACS-Analyse und
Durchführung des Sandwich-ELISA mit 5x105 Zellen analog 3.1.3.
Die Zellüberstände der für die Elektronenmikroskopie (EM) bestimmten Zellen wurden
verworfen und das Zellpellet dreimal mit PBS gewaschen. Anschließend erfolgte das
vorsichtige Überschichten des Pellets mit 100 µl eines speziellen Fixierungspuffers
(Bereitstellung über Prof. Gabriele Köhler, Medizinische Fakultät, Westfälische Universität
Münster). Die Lagerung bis zum Transport der EM-Proben erfolgte bei 4 °C. Sie wurden
innerhalb eines Tages bei 4 °C nach Münster versandt.
Die Zellen für die parallele Kontrollanalyse zur Aktivierung der DCs durch iPPVO und CpG-
ODN wurden analog 3.1.3 behandelt. Dabei fand, wie in 3.2 beschrieben, eine
Charakterisierung der Aufregulation kostimulatorischer Oberflächenmoleküle (MHC-I, MHC-
II, CD86, CD80, CD40) statt. Des Weiteren wurde die Produktion von IFN-α, IL-12/23p40
und TNF-α mit Sandwich-ELISA untersucht (Abschnitt 3.3).
3.5 Molekularbiologische Methoden
3.5.1 Isolation viraler DNA
Für die Charakterisierung der viralen, methylierten DNA von iPPVO wurde eine DNA-
Extraktion von iPPVO vorgenommen.

METHODEN
29
Zu 300 µl gereinigter iPPVO-Lösung (Prof. M. Büttner, Institut für Immunologie,
Wissenschaftszentrum für Virusinfektionen bei Tieren, Tübingen) wurden in 1,5 ml
Reaktionsgefäße 50 µl Proteinase K, 22,5 µl TEN-Puffer und 22,5 µl SDS-Lösung gegeben
und für 2 h bei 56 °C inkubiert. Anschließend wurde ein PLG-h-tube 30 s bei 12 000 g
zentrifugiert und mit 500 µl Phenol versetzt. Die Virus-Lösung wurde hinzugefügt, das Tube
mehrmals invertiert und für 20 min auf Eis gestellt. Die anschließende Zentrifugation erfolgte
bei 12 000 g für 6 min bei RT. Dieser Schritt wurde mit dem Überstand und weiteren 500 µl
Phenol in einem weiteren PLG-h-Tube wiederholt. Die wässrige Phase wurde in ein 500 µl
ICA-Lösung enthaltendes PLG-h-Tube überführt, das Tube mehrmals invertiert und
anschließend bei 12 000 g für 6 min bei RT zentrifugiert. Nach der Wiederholung dieses
Schritts mit der wässrigen Phase und weiteren 500 µl ICA-Lösung in einem neuen PLG-h-
Tube wurde die wässrige Phase in ein steriles 1,5 ml Tube überführt und das Volumen dabei
bestimmt. Anschließend wurden 1/10 des bestimmten Volumens an Natriumacetat-Lösung
und das doppelte Volumen an eiskaltem absolutem Ethanol gegeben und das Gemisch für 10
min bei -70 °C und für 2 h bei -20 °C inkubiert. Nach dem Ausfällen der viralen DNA wurde
bei 4 °C mit 17 600 g für 30 min zentrifugiert und das Pellet zweimal mit 70 %igem Ethanol
gewaschen. Nach dem Lufttrocknen des Pellets und der Resuspension der vDNA in 10-20 µl
TE-Puffer wurde die DNA-Konzentration bei OD260 und OD280 gemessen. Eine OD260 von 1
entsprach 50 µg dsDNA. Die Reinheit der isolierten DNA wurde über den Quotienten von
260/280 nm überprüft.
3.5.2 Agarose-Gelelektrophorese
Zur Analyse der isolierten DNA wurde eine Agarose-Gelelektrophorese durchgeführt. DNA
ist negativ geladen und wandert daher im elektrischen Feld der Gelkammer zur Anode. Dabei
erfolgt eine Trennung nach der Molekülmasse.
Für die Agarose-Gelelektrophorese wurden 0,8 g Agarose in 100 ml TBE-Puffer gelöst und in
der Mikrowelle zum Sieden gebracht. Nach dem Abkühlen auf unter 60 °C wurde das Gel in
einer Gelkammer mit eingesetztem Kamm gegossen. Nach dem Erkalten des Gels folgten das
Einsetzen in die Elektrophoresekammer sowie das Entfernen des Gelkamms. Nach dem
Überschichten des Gels mit TBE-Laufpuffer wurden 1 µl Probe mit 9 µl DNA-Probenpuffer
gemischt und in die Geltaschen aufgetragen. Als Kontrolle diente ein 10 kb-DNA-Standard.
Die Kammer wurde geschlossen und eine Spannung von 110 V angelegt. Die Laufzeit betrug
1 h. Anschließend musste das Gel aus der Kammer entfernt und 15 min von einem

METHODEN
30
Ethidiumbromidbad umspült werden. Die entstandenen Banden wurden im UV-Licht von
einer CCD-Kamera detektiert und dokumentiert.
3.6 Statistische Analysen
Alle Bestimmungen der Mittelwerte mit zugehöriger Standardabweichung wurden unter der
Verwendung von GraphPad PRISM TMSoftware (GraphPad Software Inc., San Diego, CA,
USA) ermittelt. Statistische Signifikanz wurde unter Anwendung des Kolmogorov-Smirnof-
Tests auf Basis eines P-Wertes von kleiner als 0,05 definiert.

ERGEBNISSE
31
4 Ergebnisse
4.1 Etablierung einer Lebend-/Tot-Färbung mit Ethidiummonoazid
Um DCs hinsichtlich ihrer Aktivierung durch iPPVO zu charakterisieren, werden sie u.a.
durchflusszytometrisch analysiert. Dabei zum Einsatz kommende Antikörper binden häufig
unspezifisch an nicht-vitale Zellen. Bei durchflusszytometrischen Analysen muss dieser
Aspekt berücksichtigt werden, um nur die spezifische Bindung der Antikörper an vitale Zellen
zu erfassen. Eine Abgrenzung der vitalen von der nicht-vitalen Zellpopulation mit Hilfe des
Vorwärts-/Seitwärtsstreulichts ist zu ungenau, da in dieser Darstellungsweise die
Populationen nur anhand ihrer Größe und Granularität eingeteilt werden. Zellen mit
geringerer Größe und daher außerhalb der gesetzten Region (Abb. 7A) liegend, erachtet man
als nicht vital. Es ist aber nicht ausgeschlossen, dass apoptotische und nekrotische Zellen mit
einer gleichen oder ähnlichen Zellgröße wie vitale Zellen noch innerhalb dieser Region liegen
und damit die Analyse beeinflussen. Daher musste eine Lebend-/Tot-Färbung eingeführt
werden, die auch für fixierte Zellen bei durchflusszytometrischen Analysen eingesetzt werden
konnte und eine klare Trennung zwischen vital und nicht-vital ermöglichte.
Ethidiummonoazidbromid (EMA) dringt in nicht-vitale Zellen mit perforierter Membran ein
und bindet kovalent über Photocrosslinking unter Lichteinfluss an die DNA. Die Membranen
vitaler Zellen sind intakt und lassen EMA nicht ins Zellinnere eindringen und an die DNA
binden. Für die ersten Untersuchungen mit EMA wurden wegen der besseren Verfügbarkeit
Zellen der murinen Makrophagen-Zellinie P388 verwendet. Als EMA-Positivkontrolle kamen
ausschließlich nicht-vitale Zellen zum Einsatz, die durch eine Hitzeinkubation bei 60 °C
gewonnen und mit der Trypanblaufärbung überprüft wurden. Wie in Abb. 7B zu sehen ist,
wurden alle Zellen der EMA-Postivkontrolle angefärbt. Die EMA-Negativkontrolle enthielt
kein EMA. Mit Hilfe dieser Kontrollen konnte die Grenze (R2) zwischen vitalen und nicht-
vitalen Zellen festgelegt werden. Für die EMA-Analyse war eine Kompensation der
Fluoreszenzen von EMA und Phycoerythrin (PE) gegeneinander aufgrund der
Emissionsspektrenüberlappung notwendig. In den Untersuchungen von mit iPPVO-
stimulierten DCs, auf welche die so etablierte Lebend-/Tot-Färbung übertragen wurde, kamen
verschiedene markierte Antikörper zum Einsatz. Neben CD11c-APC zur DC-Identifizierung
(pDCs und cDCs sind CD11c+) und B220-FITC zur Differenzierung zwischen cDCs (B220-)
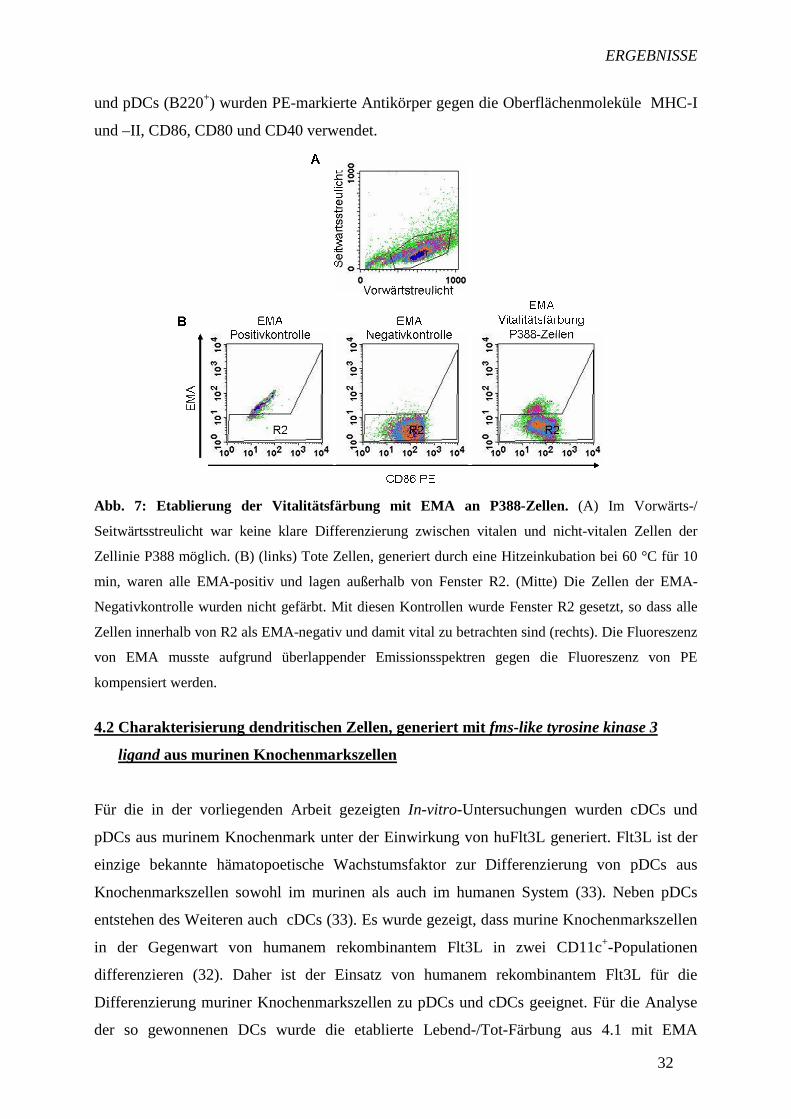
ERGEBNISSE
32
und pDCs (B220+) wurden PE-markierte Antikörper gegen die Oberflächenmoleküle MHC-I
und –II, CD86, CD80 und CD40 verwendet.
Abb. 7: Etablierung der Vitalitätsfärbung mit EMA an P388-Zellen. (A) Im Vorwärts-/
Seitwärtsstreulicht war keine klare Differenzierung zwischen vitalen und nicht-vitalen Zellen der
Zellinie P388 möglich. (B) (links) Tote Zellen, generiert durch eine Hitzeinkubation bei 60 °C für 10
min, waren alle EMA-positiv und lagen außerhalb von Fenster R2. (Mitte) Die Zellen der EMA-
Negativkontrolle wurden nicht gefärbt. Mit diesen Kontrollen wurde Fenster R2 gesetzt, so dass alle
Zellen innerhalb von R2 als EMA-negativ und damit vital zu betrachten sind (rechts). Die Fluoreszenz
von EMA musste aufgrund überlappender Emissionsspektren gegen die Fluoreszenz von PE
kompensiert werden.
4.2 Charakterisierung dendritischen Zellen, generiert mit fms-like tyrosine kinase 3
ligand aus murinen Knochenmarkszellen
Für die in der vorliegenden Arbeit gezeigten In-vitro-Untersuchungen wurden cDCs und
pDCs aus murinem Knochenmark unter der Einwirkung von huFlt3L generiert. Flt3L ist der
einzige bekannte hämatopoetische Wachstumsfaktor zur Differenzierung von pDCs aus
Knochenmarkszellen sowohl im murinen als auch im humanen System (33). Neben pDCs
entstehen des Weiteren auch cDCs (33). Es wurde gezeigt, dass murine Knochenmarkszellen
in der Gegenwart von humanem rekombinantem Flt3L in zwei CD11c+-Populationen
differenzieren (32). Daher ist der Einsatz von humanem rekombinantem Flt3L für die
Differenzierung muriner Knochenmarkszellen zu pDCs und cDCs geeignet. Für die Analyse
der so gewonnenen DCs wurde die etablierte Lebend-/Tot-Färbung aus 4.1 mit EMA

ERGEBNISSE
33
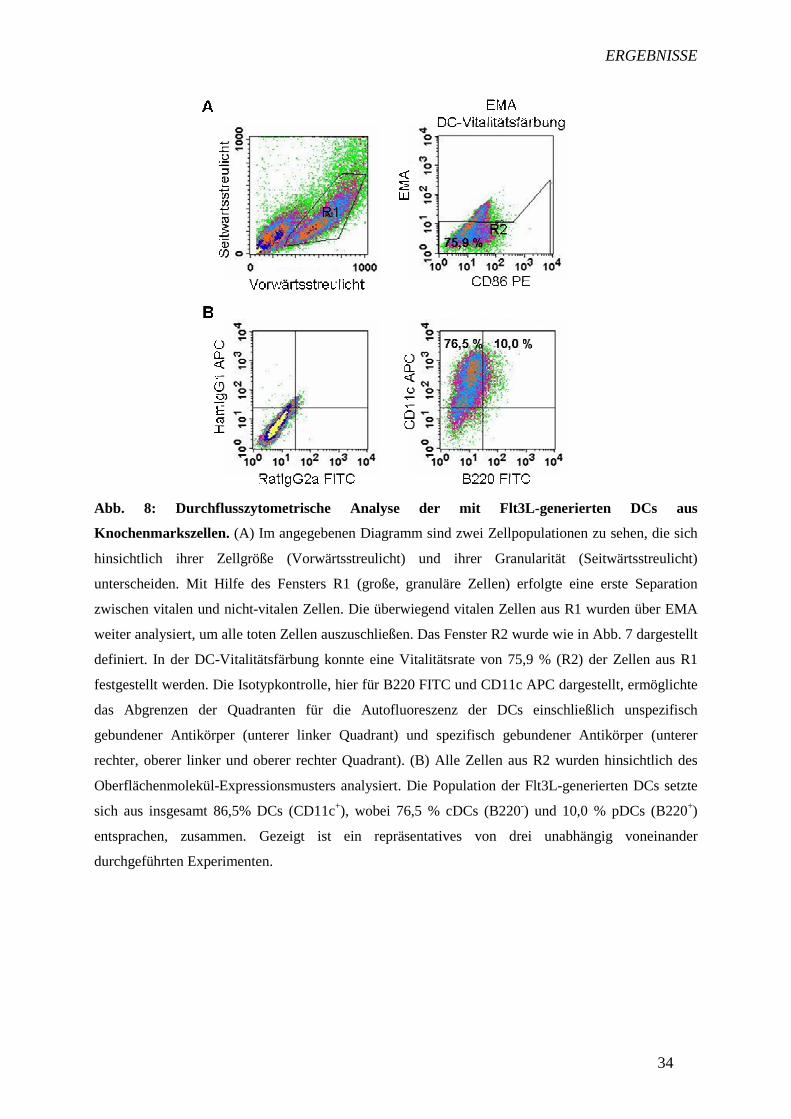
übernommen. Wie in Abb. 8A deutlich zu erkennen ist, konnten 75,9 % aller Zellen aus R1
als vital betrachtet werden.
Murine DCs unterscheidet man anhand verschiedener exprimierter Oberflächenmoleküle, die
in der Durchflusszytometrie zu einer Trennung von Zellpopulationen in der Analyse dienen.
cDCs und pDCs sind CD11c+, hingegen sind pDCs zusätzlich B220+ (28, 29). Dadurch ist
eine Unterscheidung und getrennte Analyse von cDCs und pDCs mittels Fluorochrom-
gekoppelter Antikörper gegen diese Oberflächenmarker in der Durchflusszytometrie möglich.
Von den 75,9 % vitalen Zellen aus R1 waren 86,5 % CD11c-positive Zellen. Von diesen
CD11c+-DCs waren 10 % B220+ und somit pDCs, 76,5 % hingegen B220– und daher cDCs
(Abb. 8B).
Für die durchflusszytometrische Analyse der Flt3L-DCs aus murinem Knochenmark waren
zahlreiche Kontrollen notwendig. Wie in Abb. 8B dargestellt ist, wurde für jeden verwendeten
Antikörper (anti-B220-FITC, anti-CD11c-APC und die PE-markieren AK gegen MHC-I,
MHC-II, CD86, CD80 und CD40) eine Isotypkontrolle mitgeführt, um gegebenenfalls
unspezifische Bindungen dieser AK ausschließen zu können. Bei einer Isotypkontrolle
handelt es sich um markierte Immunglobuline (Ig) der gleichen Klasse wie das zur Analyse
eingesetzte Ig, allerdings mit einer Spezifität für ein von dem zu analysierenden Antigen weit
entfernten Modellantigen. Die Isotypkontrolle ermöglichte daher das Abgrenzen der
Quadranten für die Autofluoreszenz der DCs einschließlich unspezifisch gebundener
Antikörper (unterer linker Quadrant) und spezifisch gebundener Antikörper (unterer rechter,
oberer linker und oberer rechter Quadrant). Alle Zellen sollten bei der
durchflusszytometrischen Analyse einer Isotypkontrolle im linken unteren Quadranten liegen
(aufgrund ihrer Autofluoreszenz).
ERGEBNISSE
34
Abb. 8: Durchflusszytometrische Analyse der mit Flt3L-generierten DCs aus
Knochenmarkszellen. (A) Im angegebenen Diagramm sind zwei Zellpopulationen zu sehen, die sich
hinsichtlich ihrer Zellgröße (Vorwärtsstreulicht) und ihrer Granularität (Seitwärtsstreulicht)
unterscheiden. Mit Hilfe des Fensters R1 (große, granuläre Zellen) erfolgte eine erste Separation
zwischen vitalen und nicht-vitalen Zellen. Die überwiegend vitalen Zellen aus R1 wurden über EMA
weiter analysiert, um alle toten Zellen auszuschließen. Das Fenster R2 wurde wie in Abb. 7 dargestellt
definiert. In der DC-Vitalitätsfärbung konnte eine Vitalitätsrate von 75,9 % (R2) der Zellen aus R1
festgestellt werden. Die Isotypkontrolle, hier für B220 FITC und CD11c APC dargestellt, ermöglichte
das Abgrenzen der Quadranten für die Autofluoreszenz der DCs einschließlich unspezifisch
gebundener Antikörper (unterer linker Quadrant) und spezifisch gebundener Antikörper (unterer
rechter, oberer linker und oberer rechter Quadrant). (B) Alle Zellen aus R2 wurden hinsichtlich des
Oberflächenmolekül-Expressionsmusters analysiert. Die Population der Flt3L-generierten DCs setzte
sich aus insgesamt 86,5% DCs (CD11c+), wobei 76,5 % cDCs (B220-) und 10,0 % pDCs (B220+)
entsprachen, zusammen. Gezeigt ist ein repräsentatives von drei unabhängig voneinander
durchgeführten Experimenten.

ERGEBNISSE
35
4.3 Charakterisierung der Aktivierung von dendritischen Zellen nach Stimulation mit
inaktiviertem Parapoxvirus ovis
4.3.1 Aufregulation von MHC-I/-II und kostimulatorischen Molekülen
Um die Aktivierung von DCs durch iPPVO untersuchen zu können, eignet sich die Methode
der Durchflusszytometrie: Man kann lebende Zellen hinsichtlich der auf der Zelloberfläche
exprimierten Moleküle analysieren und dabei erstens zwischen einzelnen Zellpopulationen
unterscheiden und zweitens deren Aktivierung anhand veränderter Expressionsmuster von
Oberflächenmolekülen nachvollziehen. Über den Einsatz verschiedener TLR-Liganden als
Kontrollstimuli für DCs ist es möglich, mehrere Experimente zu vergleichen und die
Aktivierung von WT-DCs im Kontrast zu TLRdef-DCs zu analysieren (Abschnitt 4.4).
Die Expression von kostimulatorischen Molekülen auf antigenpräsentierenden Zellen wie
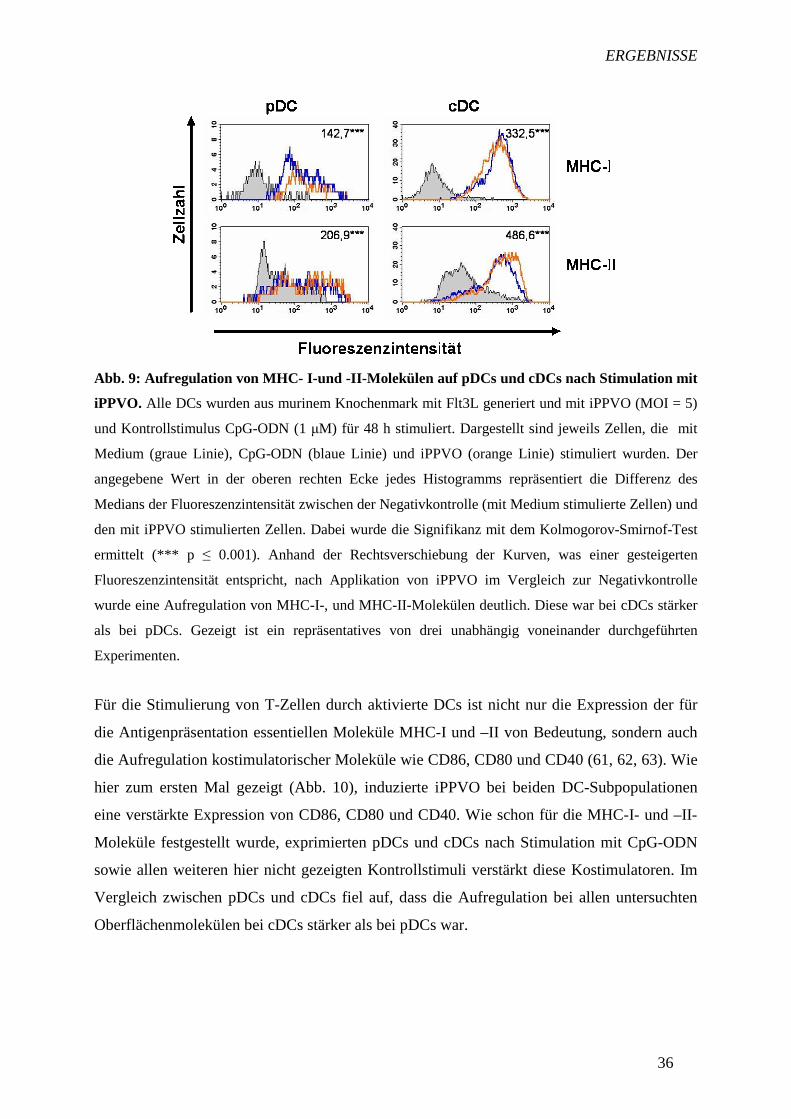
DCs nach deren Aktivierung durch Pathogene korreliert mit ihrer Fähigkeit, zur Aktivierung
von T-Zellen beizutragen. DCs reagierten auf eine Inkubation mit iPPVO mit einer
signifikanten Aufregulation von MHC-I- und -II-Molekülen (Abb. 9). Im Vergleich zwischen
pDCs und cDCs war ersichtlich, dass die Aufregulation deutlich stärker auf cDCs erfolgte als
auf pDCs. Kontrollstimulus CpG-ODN ist ein spezifischer Ligand für den endosomal
vorkommenden TLR9, welcher dsDNA erkennt. Da es sich bei iPPVO um ein dsDNA-Virus
handelt, ist CpG-ODN ein geeigneter Kontrollstimulus. In Abb. 9 ist deutlich zu erkennen,
dass CpG-ODN ebenfalls eine verstärkte Expression von MHC-I und –II-Molekülen in DCs
induzierte. Ein gleiches Ergebnis konnte für die hier nicht dargestellten Kontrollstimuli LPS
(TLR4), Poly-IC (TLR3), R-848 (TLR7) und Pam3CSK4 (TLR2) beobachtet werden. Dieses
Ergebnis demonstriert, dass die mit huFlt3L-generierten DCs TLR2, 3, 4, 7 und 9
exprimierten und über diese stimulierbar waren.
ERGEBNISSE
36
Abb. 9: Aufregulation von MHC- I-und -II-Molekülen auf pD Cs und cDCs nach Stimulation mit
iPPVO. Alle DCs wurden aus murinem Knochenmark mit Flt3L generiert und mit iPPVO (MOI = 5)
und Kontrollstimulus CpG-ODN (1 µM) für 48 h stimuliert. Dargestellt sind jeweils Zellen, die mit
Medium (graue Linie), CpG-ODN (blaue Linie) und iPPVO (orange Linie) stimuliert wurden. Der
angegebene Wert in der oberen rechten Ecke jedes Histogramms repräsentiert die Differenz des
Medians der Fluoreszenzintensität zwischen der Negativkontrolle (mit Medium stimulierte Zellen) und
den mit iPPVO stimulierten Zellen. Dabei wurde die Signifikanz mit dem Kolmogorov-Smirnof-Test
ermittelt (*** p ≤ 0.001). Anhand der Rechtsverschiebung der Kurven, was einer gesteigerten
Fluoreszenzintensität entspricht, nach Applikation von iPPVO im Vergleich zur Negativkontrolle
wurde eine Aufregulation von MHC-I-, und MHC-II-Molekülen deutlich. Diese war bei cDCs stärker
als bei pDCs. Gezeigt ist ein repräsentatives von drei unabhängig voneinander durchgeführten
Experimenten.
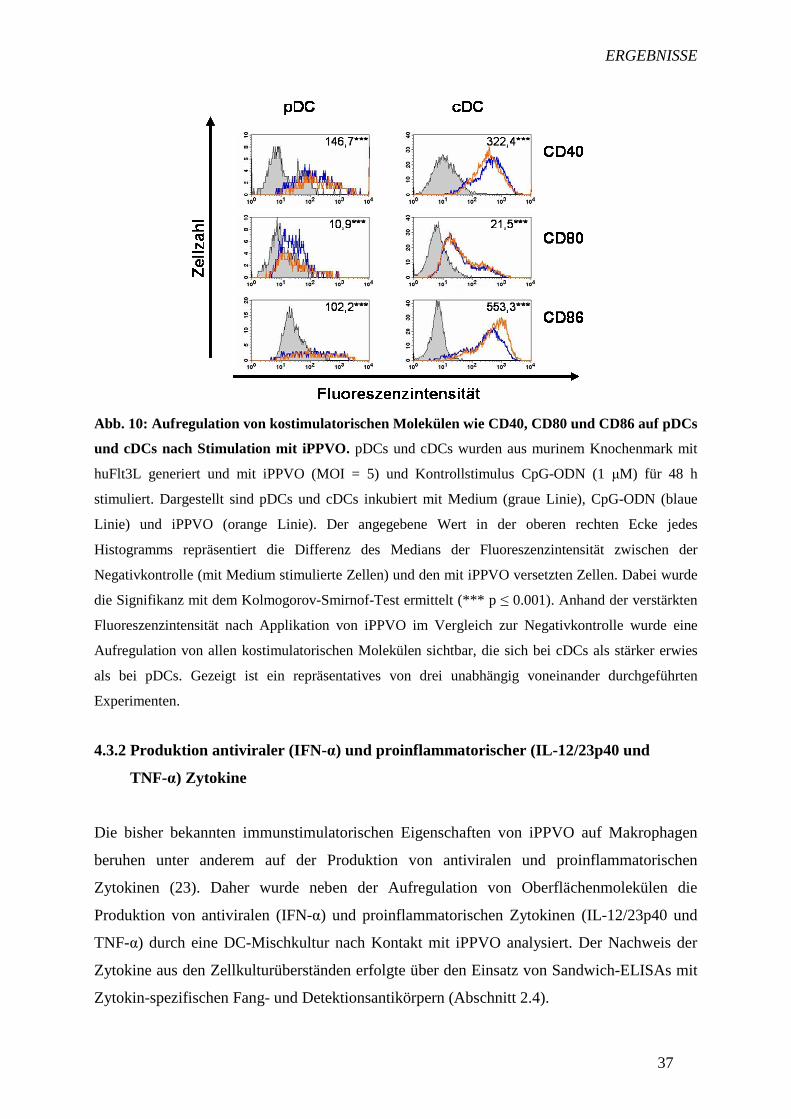
Für die Stimulierung von T-Zellen durch aktivierte DCs ist nicht nur die Expression der für
die Antigenpräsentation essentiellen Moleküle MHC-I und –II von Bedeutung, sondern auch
die Aufregulation kostimulatorischer Moleküle wie CD86, CD80 und CD40 (61, 62, 63). Wie
hier zum ersten Mal gezeigt (Abb. 10), induzierte iPPVO bei beiden DC-Subpopulationen
eine verstärkte Expression von CD86, CD80 und CD40. Wie schon für die MHC-I- und –II-
Moleküle festgestellt wurde, exprimierten pDCs und cDCs nach Stimulation mit CpG-ODN
sowie allen weiteren hier nicht gezeigten Kontrollstimuli verstärkt diese Kostimulatoren. Im
Vergleich zwischen pDCs und cDCs fiel auf, dass die Aufregulation bei allen untersuchten
Oberflächenmolekülen bei cDCs stärker als bei pDCs war.
ERGEBNISSE
37
Abb. 10: Aufregulation von kostimulatorischen Molekülen wie CD40, CD80 und CD86 auf pDCs
und cDCs nach Stimulation mit iPPVO. pDCs und cDCs wurden aus murinem Knochenmark mit
huFlt3L generiert und mit iPPVO (MOI = 5) und Kontrollstimulus CpG-ODN (1 µM) für 48 h
stimuliert. Dargestellt sind pDCs und cDCs inkubiert mit Medium (graue Linie), CpG-ODN (blaue
Linie) und iPPVO (orange Linie). Der angegebene Wert in der oberen rechten Ecke jedes
Histogramms repräsentiert die Differenz des Medians der Fluoreszenzintensität zwischen der
Negativkontrolle (mit Medium stimulierte Zellen) und den mit iPPVO versetzten Zellen. Dabei wurde
die Signifikanz mit dem Kolmogorov-Smirnof-Test ermittelt (*** p ≤ 0.001). Anhand der verstärkten
Fluoreszenzintensität nach Applikation von iPPVO im Vergleich zur Negativkontrolle wurde eine
Aufregulation von allen kostimulatorischen Molekülen sichtbar, die sich bei cDCs als stärker erwies
als bei pDCs. Gezeigt ist ein repräsentatives von drei unabhängig voneinander durchgeführten
Experimenten.
4.3.2 Produktion antiviraler (IFN-α) und proinflammatorischer (IL-12/23p40 und
TNF-α) Zytokine
Die bisher bekannten immunstimulatorischen Eigenschaften von iPPVO auf Makrophagen
beruhen unter anderem auf der Produktion von antiviralen und proinflammatorischen
Zytokinen (23). Daher wurde neben der Aufregulation von Oberflächenmolekülen die
Produktion von antiviralen (IFN-α) und proinflammatorischen Zytokinen (IL-12/23p40 und
TNF-α) durch eine DC-Mischkultur nach Kontakt mit iPPVO analysiert. Der Nachweis der
Zytokine aus den Zellkulturüberständen erfolgte über den Einsatz von Sandwich-ELISAs mit
Zytokin-spezifischen Fang- und Detektionsantikörpern (Abschnitt 2.4).
ERGEBNISSE
38
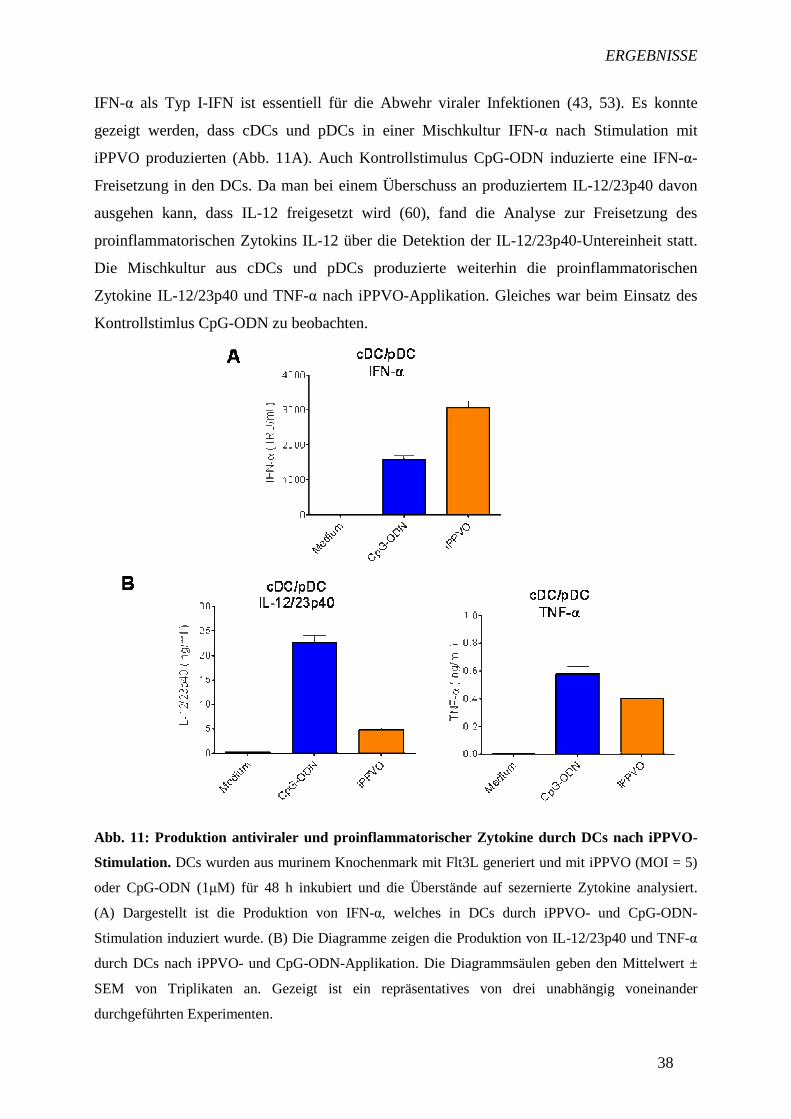
IFN-α als Typ I-IFN ist essentiell für die Abwehr viraler Infektionen (43, 53). Es konnte
gezeigt werden, dass cDCs und pDCs in einer Mischkultur IFN-α nach Stimulation mit
iPPVO produzierten (Abb. 11A). Auch Kontrollstimulus CpG-ODN induzierte eine IFN-α-
Freisetzung in den DCs. Da man bei einem Überschuss an produziertem IL-12/23p40 davon
ausgehen kann, dass IL-12 freigesetzt wird (60), fand die Analyse zur Freisetzung des
proinflammatorischen Zytokins IL-12 über die Detektion der IL-12/23p40-Untereinheit statt.
Die Mischkultur aus cDCs und pDCs produzierte weiterhin die proinflammatorischen
Zytokine IL-12/23p40 und TNF-α nach iPPVO-Applikation. Gleiches war beim Einsatz des
Kontrollstimlus CpG-ODN zu beobachten.
Abb. 11: Produktion antiviraler und proinflammatorischer Zytoki ne durch DCs nach iPPVO-
Stimulation. DCs wurden aus murinem Knochenmark mit Flt3L generiert und mit iPPVO (MOI = 5)
oder CpG-ODN (1µM) für 48 h inkubiert und die Überstände auf sezernierte Zytokine analysiert.
(A) Dargestellt ist die Produktion von IFN-α, welches in DCs durch iPPVO- und CpG-ODN-
Stimulation induziert wurde. (B) Die Diagramme zeigen die Produktion von IL-12/23p40 und TNF-α
durch DCs nach iPPVO- und CpG-ODN-Applikation. Die Diagrammsäulen geben den Mittelwert ±
SEM von Triplikaten an. Gezeigt ist ein repräsentatives von drei unabhängig voneinander
durchgeführten Experimenten.

ERGEBNISSE
39
4.3.3 Vergleich zwischen DCs, generiert mit humanem und murinem Flt3L
Im murinen System ist der Einsatz von humanen hämatopoetischen Faktoren mit Risiken
verbunden. So können Spezies-spezifische Nebenreaktionen auftreten wie z.B. eine
unterschiedliche Wirkung von huFlt3L auf murine pDCs und cDCs im Vergleich zu muFlt3L.
Des Weiteren kann die Quelle des hämatopoetischen Faktors einen Einfluss auf seine
Wirksamkeit haben. HuFlt3L wurde rekombinant hergestellt und konnte daher genau
quantifiziert werden. MuFlt3L wurde aus dem Überstand einer transfizierten Zelllinie
gewonnen und anschließend gereinigt. Eine genaue Konzentration des Wachstumsfaktors ist
unbekannt, er wird in einem Volumen von 10 µl/ml eingesetzt.
Um die Wirkung von rekombinantem huFlt3L und dem Überstand, der muFlt3L enthielt, auf
murine Knochenmarkszellen zu untersuchen, wurde ein Vergleichsexperiment durchgeführt.
Dafür fand die Inkubation für einen Teil der murinen Knochenmarkszellen für 7 Tage mit
humanem, für einen anderen Teil mit murinem Flt3L statt. Diese so gewonnenen Zellen
konnten hinsichtlich ihrer charakteristischen DC-Oberflächenmoleküle (CD11c, B220) sowie
auf veränderte Oberflächenmolekül-Expressionsmuster und die Produktion von IL-12/23p40
nach Stimulation mit iPPVO sowie den Kontrollstimuli CpG-ODN und LPS analysiert
werden. In Abb. 12 ist dargestellt, dass sich muFlt3L-DCs nach Stimulation genauso
verhielten wie huFlt3L-DCs. Sowohl MHC-I-Moleküle als auch das kostimulatorische
Molekül CD86 wurden im vergleichbaren Verhältnis nach Stimulation aufreguliert, wie es bei
huFlt3L-DCs zu beobachten war (Abschnitt 4.3.1). IL-12/23p40 wurde durch die Mischkultur
aus pDCs und cDCs, generiert mit muFlt3L, nach CpG-ODN- und LPS-Stimulation
freigesetzt. MuFlt3L-DCs produzierten dabei geringfügig weniger IL-12/23p40 als huFlt3L-
DCs. Die Konzentrationsunterschiede betrugen weniger als 1 ng/ml und waren auf
experimentelle Schwankungen zurückzuführen. Das Verhältnis zwischen pDCs und cDCs war
gleich nach Einsatz von muFlt3L oder huFlt3L (10-17 % pDCs und 75-85 % cDCs).
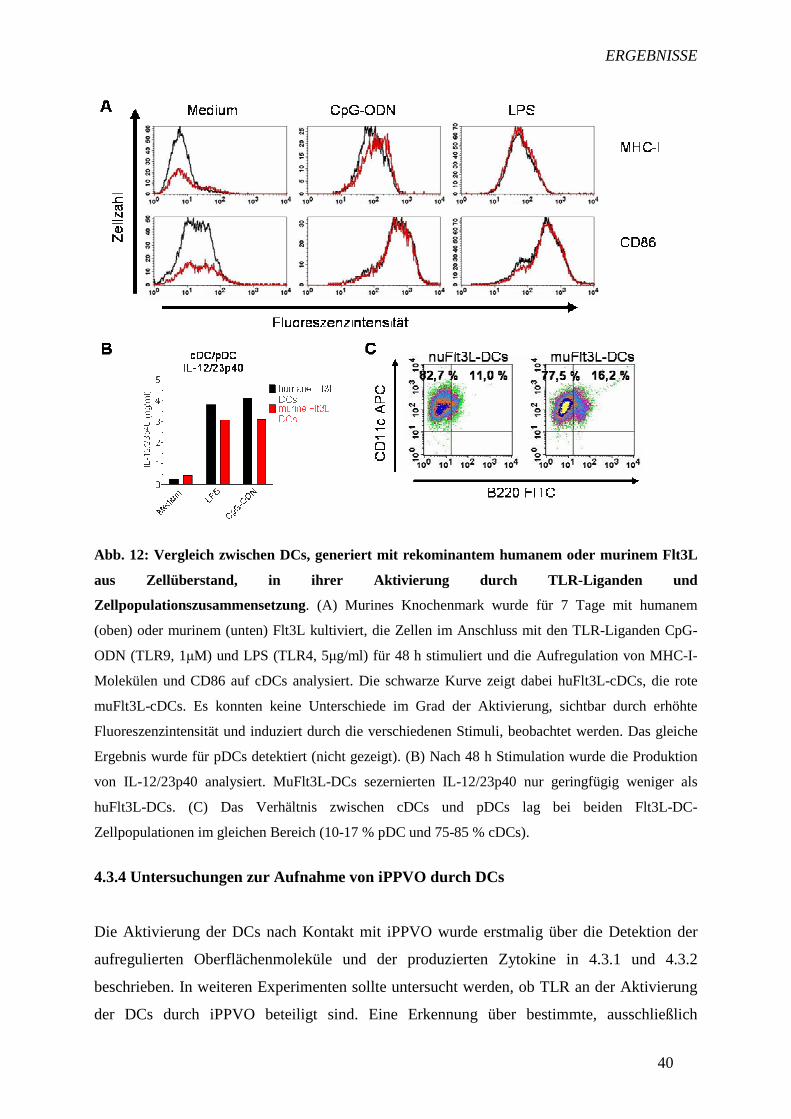
ERGEBNISSE
40
Abb. 12: Vergleich zwischen DCs, generiert mit rekominantem humanem oder murinem Flt3L
aus Zellüberstand, in ihrer Aktivierung durch TLR-Ligande n und
Zellpopulationszusammensetzung. (A) Murines Knochenmark wurde für 7 Tage mit humanem
(oben) oder murinem (unten) Flt3L kultiviert, die Zellen im Anschluss mit den TLR-Liganden CpG-
ODN (TLR9, 1µM) und LPS (TLR4, 5µg/ml) für 48 h stimuliert und die Aufregulation von MHC-I-
Molekülen und CD86 auf cDCs analysiert. Die schwarze Kurve zeigt dabei huFlt3L-cDCs, die rote
muFlt3L-cDCs. Es konnten keine Unterschiede im Grad der Aktivierung, sichtbar durch erhöhte
Fluoreszenzintensität und induziert durch die verschiedenen Stimuli, beobachtet werden. Das gleiche
Ergebnis wurde für pDCs detektiert (nicht gezeigt). (B) Nach 48 h Stimulation wurde die Produktion
von IL-12/23p40 analysiert. MuFlt3L-DCs sezernierten IL-12/23p40 nur geringfügig weniger als
huFlt3L-DCs. (C) Das Verhältnis zwischen cDCs und pDCs lag bei beiden Flt3L-DC-
Zellpopulationen im gleichen Bereich (10-17 % pDC und 75-85 % cDCs).
4.3.4 Untersuchungen zur Aufnahme von iPPVO durch DCs
Die Aktivierung der DCs nach Kontakt mit iPPVO wurde erstmalig über die Detektion der
aufregulierten Oberflächenmoleküle und der produzierten Zytokine in 4.3.1 und 4.3.2
beschrieben. In weiteren Experimenten sollte untersucht werden, ob TLR an der Aktivierung
der DCs durch iPPVO beteiligt sind. Eine Erkennung über bestimmte, ausschließlich

ERGEBNISSE
41
endosomal vorkommende TLR wie TLR9 setzt eine endozytotische Aufnahme der Virionen
in DCs voraus. In der Literatur werden verschiedene Möglichkeiten der Aufnahme von
Pockenviren in die Wirtszelle anhand von zahlreichen Untersuchungen mit VV diskutiert, wie
beispielsweise über Endozytose (9, 10, 11, 12). In dieser Arbeit galt es zu untersuchen, ob
iPPVO von DCs aufgenommen wird und welche der propagierten Aufnahmemechanismen für
nähere Untersuchungen in Frage kommen. Als einleitendes Experiment zu dieser Zielstellung
dienten elektronenmikroskopische Aufnahmen von mit iPPVO versetzten DCs.
Die ersten elektronenmikroskopischen Aufnahmen zeigten die Aufnahme von iPPVO durch
DCs (Abb. 13). Im Vergleich zu Aufnahmen mit Kontroll-DCs (mit Medium stimulierte DCs)
waren nach Stimulation mit iPPVO ovale Partikel mit einer Größe von 200 nm zu sehen.
Diese Partikel sind das von der DC aufgenommene iPPVO (Abb. 13A). Nach 6 h Stimulation
befanden sich zahlreiche Einschlüsse mit Virionen im DC-Zellkörper, besonders in den
Zellrandbereichen (Abb. 13B, D). Im Zellinneren liegende lysomale bzw. endosomale
Gebilde schienen Abbauprodukte verdauter Virionen zu enthalten. Dabei wies die
ultrastrukturelle Morphologie der Einschlusskörper darauf hin, dass es sich bei ihnen um
Lysosomen handelt (Prof. Köhler, mündliche Mitteilung). Weiterhin ließ sich aus den
gewonnenen Aufnahmen vermuten, dass dendritische Ausprägungen der DCs die Virionen
umschließen. Die eindeutige lysosomale Lokalisation von iPPVO (Abb. 13B, D) deutete
daher auf Phagozytose als Aufnahmemechanismus hin, was im Vergleich zum bisherigen
Wissen ein wichtiges neues Ergebnis in der Forschung über die immunstimulierenden
Mechanismen von iPPVO darstellt. Die Zahl der extrazellulären Virionen im zellfreien
Überstand nach 6 h Stimulation war auch im Fall der Infektion hoch permissiver Zellen sehr
gering. Diese Beobachtungen können zum einen mit einer schnellen Aufnahme der Virionen
durch DCs oder durch die zahlreichen Waschschritte während der Präparation des Zellpellets
erklärt werden. Das Wollknäuel-ähnliche (ball of wool morphology) Aussehen der
Coreproteinstruktur der Viren war gut zu erkennen (Abb. 13D). Elektronenmikroskopische
Aufnahmen von DCs nach Inkubation für 12, 24 und 48 h mit iPPVO waren aufgrund der
bereits einsetzenden Virusdegradation nicht für die Analyse der Zellaufnahme von iPPVO
verwertbar. Eine Parallelanalyse der DCs mittels Durchflusszytometrie sowie die Detektion
der produzierten Zytokine als Folge der iPPVO-Applikation ergab das gleiche Bild wie in
4.3.1 - 4.3.3 beschrieben.
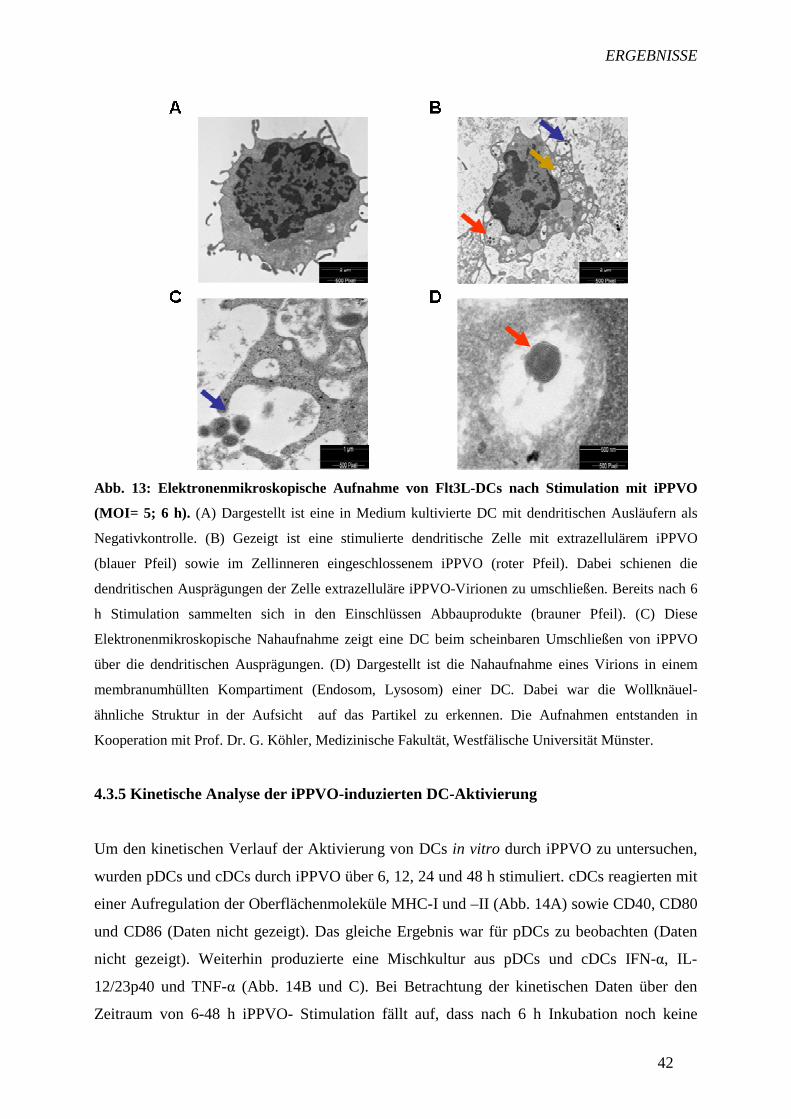
ERGEBNISSE
42
Abb. 13: Elektronenmikroskopische Aufnahme von Flt3L-DCs nach Stimulation mit iPPVO
(MOI= 5; 6 h). (A) Dargestellt ist eine in Medium kultivierte DC mit dendritischen Ausläufern als
Negativkontrolle. (B) Gezeigt ist eine stimulierte dendritische Zelle mit extrazellulärem iPPVO
(blauer Pfeil) sowie im Zellinneren eingeschlossenem iPPVO (roter Pfeil). Dabei schienen die
dendritischen Ausprägungen der Zelle extrazelluläre iPPVO-Virionen zu umschließen. Bereits nach 6
h Stimulation sammelten sich in den Einschlüssen Abbauprodukte (brauner Pfeil). (C) Diese
Elektronenmikroskopische Nahaufnahme zeigt eine DC beim scheinbaren Umschließen von iPPVO
über die dendritischen Ausprägungen. (D) Dargestellt ist die Nahaufnahme eines Virions in einem
membranumhüllten Kompartiment (Endosom, Lysosom) einer DC. Dabei war die Wollknäuel-
ähnliche Struktur in der Aufsicht auf das Partikel zu erkennen. Die Aufnahmen entstanden in
Kooperation mit Prof. Dr. G. Köhler, Medizinische Fakultät, Westfälische Universität Münster.
4.3.5 Kinetische Analyse der iPPVO-induzierten DC-Aktivierung
Um den kinetischen Verlauf der Aktivierung von DCs in vitro durch iPPVO zu untersuchen,
wurden pDCs und cDCs durch iPPVO über 6, 12, 24 und 48 h stimuliert. cDCs reagierten mit
einer Aufregulation der Oberflächenmoleküle MHC-I und –II (Abb. 14A) sowie CD40, CD80
und CD86 (Daten nicht gezeigt). Das gleiche Ergebnis war für pDCs zu beobachten (Daten
nicht gezeigt). Weiterhin produzierte eine Mischkultur aus pDCs und cDCs IFN-α, IL-
12/23p40 und TNF-α (Abb. 14B und C). Bei Betrachtung der kinetischen Daten über den
Zeitraum von 6-48 h iPPVO- Stimulation fällt auf, dass nach 6 h Inkubation noch keine

ERGEBNISSE
43
eindeutige Aktivierung der cDCs (Abb. 14A) und der pDCs (Daten nicht gezeigt) zu
beobachten war, sowohl in Bezug auf die Aufregulation von MHC-I und –II- Molekülen als
auch bei der Zytokinproduktion. Die gesteigerte Expression kostimulatorischer
Oberflächenmoleküle wurde nach etwa 12 h sichtbar, hingegen konnte die Produktion von
IFN-α und IL-12/23p40 erst nach 24 h nachgewiesen werden. Lediglich sezerniertes TNF-α
war schon nach 6 h Stimulation detektierbar, wobei die Menge linear mit steigender
Stimulationszeit zunahm (Abb. 14C). Die analysierten Aktivierungsmarker der DCs waren
somit erst nach einer Inkubation über 12-24 h mit iPPVO erhöht im Vergleich zur
Negativkontrolle (für 48 h mit Medium kultivierte DCs). Die DCs der Negativkontrolle
zeigten im Verlauf der zeitlichen Stimulation von 6-48 h keine Änderung in der
Oberflächenmolekülexpression (Daten nicht gezeigt) und Zytokinproduktion (Abb. 14B). Für
Kontrollstimulus CpG-ODN wurden ausschließlich nach 48 h Stimulation die
Aktivierungsmarker der DCs bestimmt. Wie schon in 4.3.1 und 4.3.2 beschrieben, induzierte
CpG-ODN nach 48 h Inkubation eine Aufregulation von MHC-I, MHC-II und CD86, CD80
sowie CD40 in cDCs und pDCs (Daten nicht gezeigt). Die Produktion von IFN-α, IL-
12/23p40 und TNF-α wurde ebenfalls über CpG-ODN in der Mischkultur aus cDCs und
pDCs aktiviert.
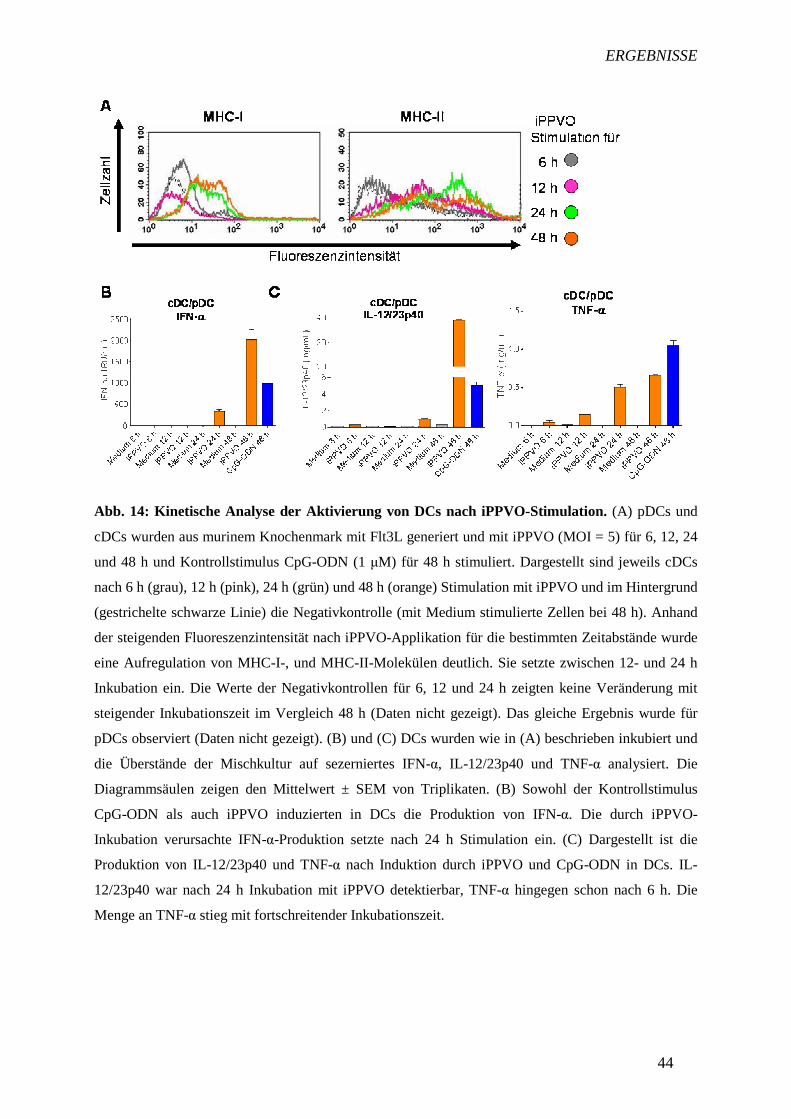
ERGEBNISSE
44
Abb. 14: Kinetische Analyse der Aktivierung von DCs nach iPPVO-Stimulation. (A) pDCs und
cDCs wurden aus murinem Knochenmark mit Flt3L generiert und mit iPPVO (MOI = 5) für 6, 12, 24
und 48 h und Kontrollstimulus CpG-ODN (1 µM) für 48 h stimuliert. Dargestellt sind jeweils cDCs
nach 6 h (grau), 12 h (pink), 24 h (grün) und 48 h (orange) Stimulation mit iPPVO und im Hintergrund
(gestrichelte schwarze Linie) die Negativkontrolle (mit Medium stimulierte Zellen bei 48 h). Anhand
der steigenden Fluoreszenzintensität nach iPPVO-Applikation für die bestimmten Zeitabstände wurde
eine Aufregulation von MHC-I-, und MHC-II-Molekülen deutlich. Sie setzte zwischen 12- und 24 h
Inkubation ein. Die Werte der Negativkontrollen für 6, 12 und 24 h zeigten keine Veränderung mit
steigender Inkubationszeit im Vergleich 48 h (Daten nicht gezeigt). Das gleiche Ergebnis wurde für
pDCs observiert (Daten nicht gezeigt). (B) und (C) DCs wurden wie in (A) beschrieben inkubiert und
die Überstände der Mischkultur auf sezerniertes IFN-α, IL-12/23p40 und TNF-α analysiert. Die
Diagrammsäulen zeigen den Mittelwert ± SEM von Triplikaten. (B) Sowohl der Kontrollstimulus
CpG-ODN als auch iPPVO induzierten in DCs die Produktion von IFN-α. Die durch iPPVO-
Inkubation verursachte IFN-α-Produktion setzte nach 24 h Stimulation ein. (C) Dargestellt ist die
Produktion von IL-12/23p40 und TNF-α nach Induktion durch iPPVO und CpG-ODN in DCs. IL-
12/23p40 war nach 24 h Inkubation mit iPPVO detektierbar, TNF-α hingegen schon nach 6 h. Die
Menge an TNF-α stieg mit fortschreitender Inkubationszeit.

ERGEBNISSE
45
4.4 Beteiligung der Toll-like Rezeptoren 9 und 2 an der Stimulation von dendritischen
Zellen durch inaktiviertes Parapoxvirus ovis
4.4.1 Rolle von TLR9 bei der iPPVO-induzierten DC-Aktivierung
Unmethylierte 2’-Desoxyribo-CpG-DNA-Motive, wie in bakterieller und viraler DNA
vorkommend, sind Liganden von TLR9 (67). Allerdings mehren sich auch zunehmend
Anzeichen für eine Erkennung von methylierter DNA durch TLR9 (Prof. Alber, mündliche
Mitteilung). iPPVO ist ein dsDNA-Virus mit einem hohen G/C-Gehalt (4). Daher schien
TLR9 ein möglicher Rezeptorkandidat für die DC-Aktivierung durch iPPVO zu sein. Im
Gegensatz zu anderen TLR, welche hauptsächlich auf der Zelloberfläche exprimiert werden,
befindet sich TLR9 ausschließlich in endosomalen Kompartimenten (43). Über die
elektronenmikroskopischen Aufnahmen von mit iPPVO-stimulierten DCs konnte eine
endozytotische Aufnahme der Virionen in Lysosomen zweifelsfrei demonstriert werden (Abb.
13 B, D)). Diese endozytotische Aufnahme mit anschließender Virusdegradation führt zur
Freisetzung viraler dsDNA im Endosom (43), wo sie anschließend über TLR9 erkannt und die
intrazelluläre Signaltransduktion MyD88-abhängig eingeleitet werden kann. Zur
Untersuchung bezüglich der Rolle des Rezeptorkandidaten TLR9 in der iPPVO-induzierten
DC-Aktivierung wurden DCs aus Knochenmark, das von TLR9-/-- und WT-Mäusen stammte,
auf eine TLR9-Abhängigkeit der Expression kostimulatorischer Oberflächenmoleküle sowie
die Produktion von IFN-α, IL-12/23p40 und TNF-α nach iPPVO-Applikation analysiert.
Aus den durchflusszytometrischen Daten für die Aufregulation von MHC-I-Molekülen und
CD86 ist deutlich zu erkennen, dass die Aktivierung von cDCs durch iPPVO TLR9-
unabhängig erfolgte (Abb. 15). Hingegen induzierte CpG-ODN (TLR9-Ligand) nur bei WT-
cDCs, nicht aber bei TLR9-/--cDCs, eine verstärkte Expression von MHC-I-Molekülen und
CD86 (Abb. 14A). Das gleiche Ergebnis konnte für pDCs und die kostimulatorischen
Moleküle MHC-II, CD80 und CD40 gefunden werden (Daten nicht gezeigt). Der TLR9-
unabhängige Kontrollstimulus LPS aktivierte sowohl WT-cDCs als auch TLR9-/--cDCs mit
gleicher Stärke. Die TLR9-Defizienz der DCs beeinflusste nicht die Effizienz der DC-
Aktivierung über iPPVO. Das pDC/cDC-Verhältnis innerhalb der TLR9-/--DC-Population
entsprach der WT-DC-Population mit 10-17 % pDC und 75-85 % cDCs (Daten nicht gezeigt).
Isolierte iPPVO-DNA, deren Reinheit über Agarose-Gelelektrophorese überprüft wurde
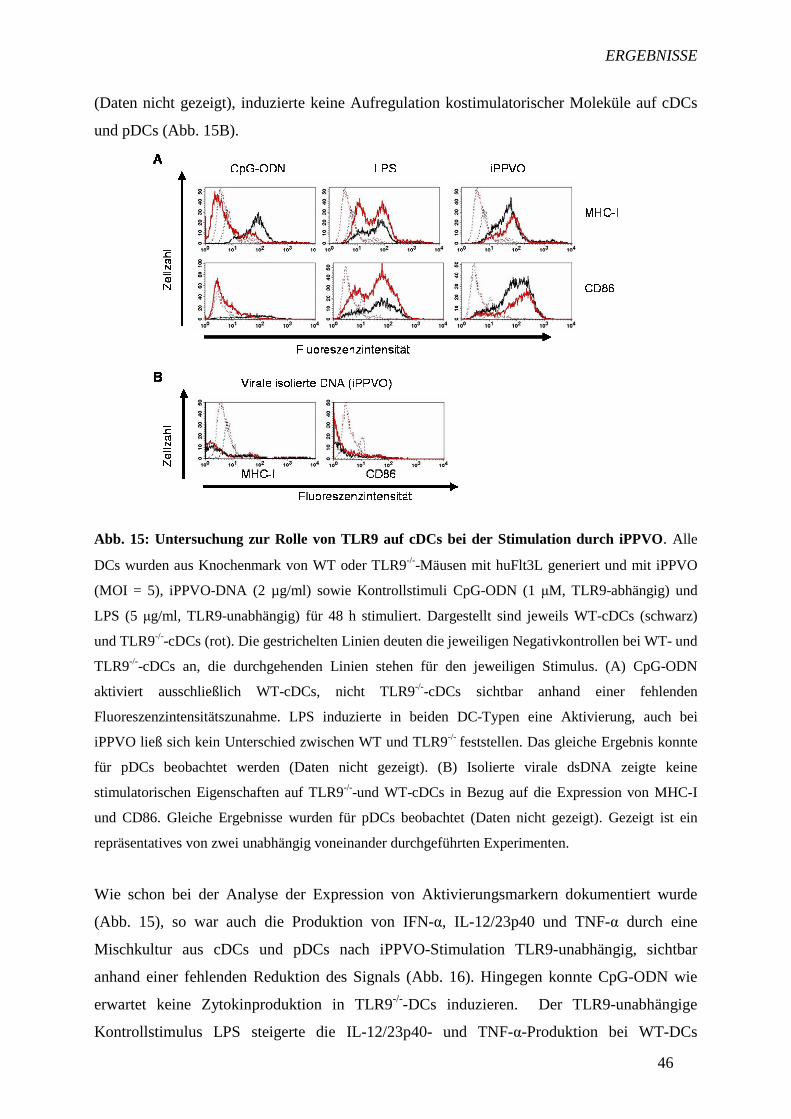
ERGEBNISSE
46
(Daten nicht gezeigt), induzierte keine Aufregulation kostimulatorischer Moleküle auf cDCs
und pDCs (Abb. 15B).
Abb. 15: Untersuchung zur Rolle von TLR9 auf cDCs bei der Stimulation durch iPPVO . Alle
DCs wurden aus Knochenmark von WT oder TLR9-/--Mäusen mit huFlt3L generiert und mit iPPVO
(MOI = 5), iPPVO-DNA (2 µg/ml) sowie Kontrollstimuli CpG-ODN (1 µM, TLR9-abhängig) und
LPS (5 µg/ml, TLR9-unabhängig) für 48 h stimuliert. Dargestellt sind jeweils WT-cDCs (schwarz)
und TLR9-/--cDCs (rot). Die gestrichelten Linien deuten die jeweiligen Negativkontrollen bei WT- und
TLR9-/--cDCs an, die durchgehenden Linien stehen für den jeweiligen Stimulus. (A) CpG-ODN
aktiviert ausschließlich WT-cDCs, nicht TLR9-/--cDCs sichtbar anhand einer fehlenden
Fluoreszenzintensitätszunahme. LPS induzierte in beiden DC-Typen eine Aktivierung, auch bei
iPPVO ließ sich kein Unterschied zwischen WT und TLR9-/- feststellen. Das gleiche Ergebnis konnte
für pDCs beobachtet werden (Daten nicht gezeigt). (B) Isolierte virale dsDNA zeigte keine
stimulatorischen Eigenschaften auf TLR9-/--und WT-cDCs in Bezug auf die Expression von MHC-I
und CD86. Gleiche Ergebnisse wurden für pDCs beobachtet (Daten nicht gezeigt). Gezeigt ist ein
repräsentatives von zwei unabhängig voneinander durchgeführten Experimenten.
Wie schon bei der Analyse der Expression von Aktivierungsmarkern dokumentiert wurde
(Abb. 15), so war auch die Produktion von IFN-α, IL-12/23p40 und TNF-α durch eine
Mischkultur aus cDCs und pDCs nach iPPVO-Stimulation TLR9-unabhängig, sichtbar
anhand einer fehlenden Reduktion des Signals (Abb. 16). Hingegen konnte CpG-ODN wie
erwartet keine Zytokinproduktion in TLR9-/--DCs induzieren. Der TLR9-unabhängige
Kontrollstimulus LPS steigerte die IL-12/23p40- und TNF-α-Produktion bei WT-DCs
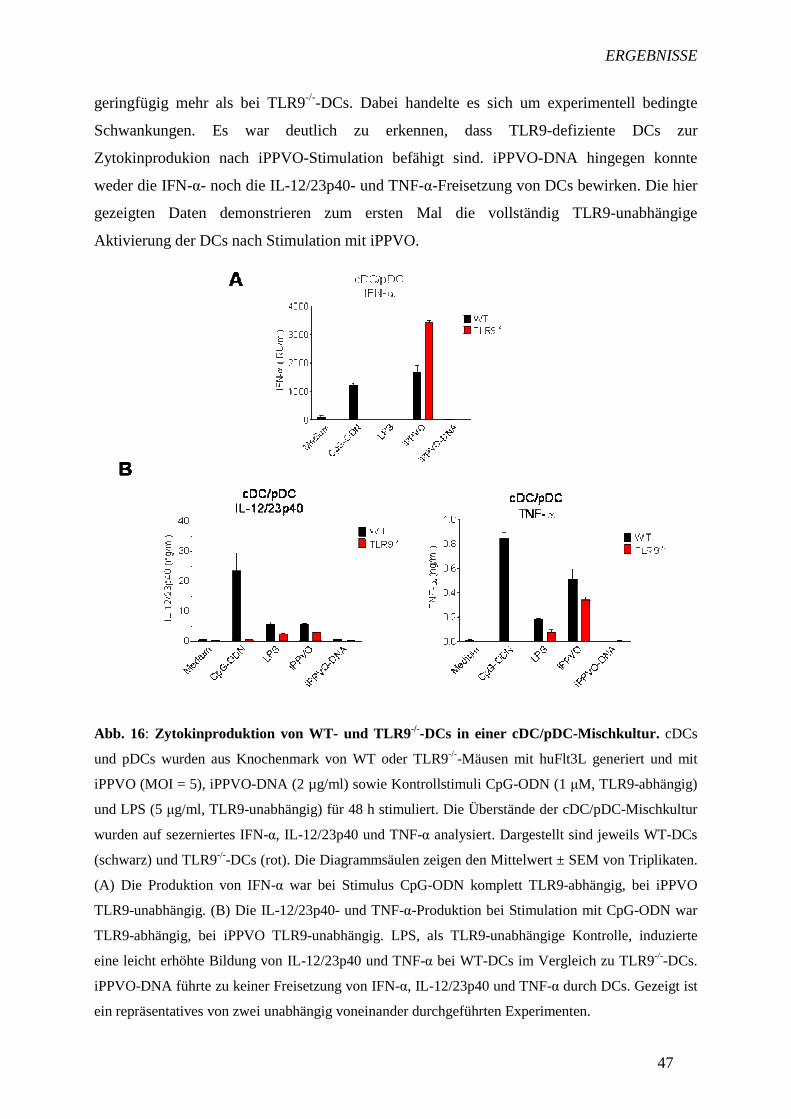
ERGEBNISSE
47
geringfügig mehr als bei TLR9-/--DCs. Dabei handelte es sich um experimentell bedingte
Schwankungen. Es war deutlich zu erkennen, dass TLR9-defiziente DCs zur
Zytokinprodukion nach iPPVO-Stimulation befähigt sind. iPPVO-DNA hingegen konnte
weder die IFN-α- noch die IL-12/23p40- und TNF-α-Freisetzung von DCs bewirken. Die hier
gezeigten Daten demonstrieren zum ersten Mal die vollständig TLR9-unabhängige
Aktivierung der DCs nach Stimulation mit iPPVO.
Abb. 16: Zytokinproduktion von WT- und TLR9 -/--DCs in einer cDC/pDC-Mischkultur. cDCs
und pDCs wurden aus Knochenmark von WT oder TLR9-/--Mäusen mit huFlt3L generiert und mit
iPPVO (MOI = 5), iPPVO-DNA (2 µg/ml) sowie Kontrollstimuli CpG-ODN (1 µM, TLR9-abhängig)
und LPS (5 µg/ml, TLR9-unabhängig) für 48 h stimuliert. Die Überstände der cDC/pDC-Mischkultur
wurden auf sezerniertes IFN-α, IL-12/23p40 und TNF-α analysiert. Dargestellt sind jeweils WT-DCs
(schwarz) und TLR9-/--DCs (rot). Die Diagrammsäulen zeigen den Mittelwert ± SEM von Triplikaten.
(A) Die Produktion von IFN-α war bei Stimulus CpG-ODN komplett TLR9-abhängig, bei iPPVO
TLR9-unabhängig. (B) Die IL-12/23p40- und TNF-α-Produktion bei Stimulation mit CpG-ODN war
TLR9-abhängig, bei iPPVO TLR9-unabhängig. LPS, als TLR9-unabhängige Kontrolle, induzierte
eine leicht erhöhte Bildung von IL-12/23p40 und TNF-α bei WT-DCs im Vergleich zu TLR9-/--DCs.
iPPVO-DNA führte zu keiner Freisetzung von IFN-α, IL-12/23p40 und TNF-α durch DCs. Gezeigt ist
ein repräsentatives von zwei unabhängig voneinander durchgeführten Experimenten.

ERGEBNISSE
48
4.4.2 Rolle von TLR2 bei der iPPVO-induzierten DC-Aktivierung
Nach dem Ausschluss von TLR9 als möglichen Rezeptor der iPPVO-induzierten DC-
Aktivierung wurde in weiterführenden Analysen eine partielle MyD88-Abhängigkeit der
Aufregulation der Oberflächenmolekülexpression sowie der IL-12/23p40- und TNF-α-
Produktion durch DCs nach iPPVO-Stimulation festgestellt (Siegemund et al., Manuskript in
Vorbereitung). Dieses Ergebnis deutet darauf hin, dass TLR an der DC-Aktivierung durch
iPPVO involviert waren und weitere Mitglieder dieser PRR-Familie der TLR in Erwägung
gezogen werden mussten. Erst kürzlich wurde in der Literatur der Zusammenhang zwischen
TLR2 und der Aktivierung von Zellen des angeborenen Immunsystems nach Infektion mit
Vaccinia-Viren (VV) dokumentiert (56). Dabei produzierten DCs (generiert aus murinem
Knochenmark mit IL-4 und GM-CSF) proinflammatorische Zytokine TLR2- und MyD88-
abhängig, IFN-β hingegen TLR2-unabhängig. VV gehören zur Familie der Pockenviren und
sind daher verwandt mit dem in Bezug auf DCs zu untersuchenden iPPVO. Schlussfolgernd
stellte sich die Frage, ob TLR2 auch für die Aktivierung von DCs durch iPPVO eine
entscheidende Rolle spielt. Zur Untersuchung bezüglich der Rolle von TLR2 in der iPPVO-
induzierten DC-Aktivierung wurden DCs aus Knochenmark, das von TLR2-/-- und WT-
Mäusen stammte, auf eine TLR2-Abhängigkeit der Expression der Oberflächenmoleküle
MHC-I, MHC-II, CD86, CD80 und CD40 sowie die Produktion von IFN-α, IL-12/23p40 und
TNF-α nach iPPVO-Applikation untersucht. Als Kontrollstimulus kam Pam3CSK4 als
synthetischer TLR2-Ligand eine entscheidende Funktion zu, denn nur WT-DCs werden durch
diesen Liganden über TLR2 aktiviert, nicht aber TLR2-/--DCs. LPS diente als TLR2-
unabhängiger Kontrollstimulus.
Im Gegensatz zu VV (56) wurden die immunstimulatorischen Effekte von iPPVO auf DCs
nicht über TLR2 geleitet (Abb. 17 und 18). Die Aufregulation kostimulatorischer
Oberflächenmoleküle erfolgte bei WT- und TLR2-/--cDCs in gleicher Stärke, es konnten keine
Unterschiede im Expressionsmuster aktivierter WT-cDCs und TLR2-/--cDCs festgestellt
werden. Wie zu erwarten war, induzierte Pam3CSK4 nur bei WT-cDCs eine verstärkte
Expression von MHC-I-Molekülen und CD86. Bei TLR2-/--cDCs war dieses
Aktivierungssignal nicht mehr vorhanden (Abb. 17). Das gleiche Ergebnis konnte für die
nicht gezeigten pDCs und Oberflächenmoleküle MHC-II, CD80 und CD40 observiert werden.
Der TLR2-unabhängige Kontrollstimulus LPS aktivierte sowohl WT-cDCs als auch TLR2-/--
cDCs mit gleicher Stärke. Somit beeinflusste die TLR2-Defizienz nicht die Effizienz der DC-
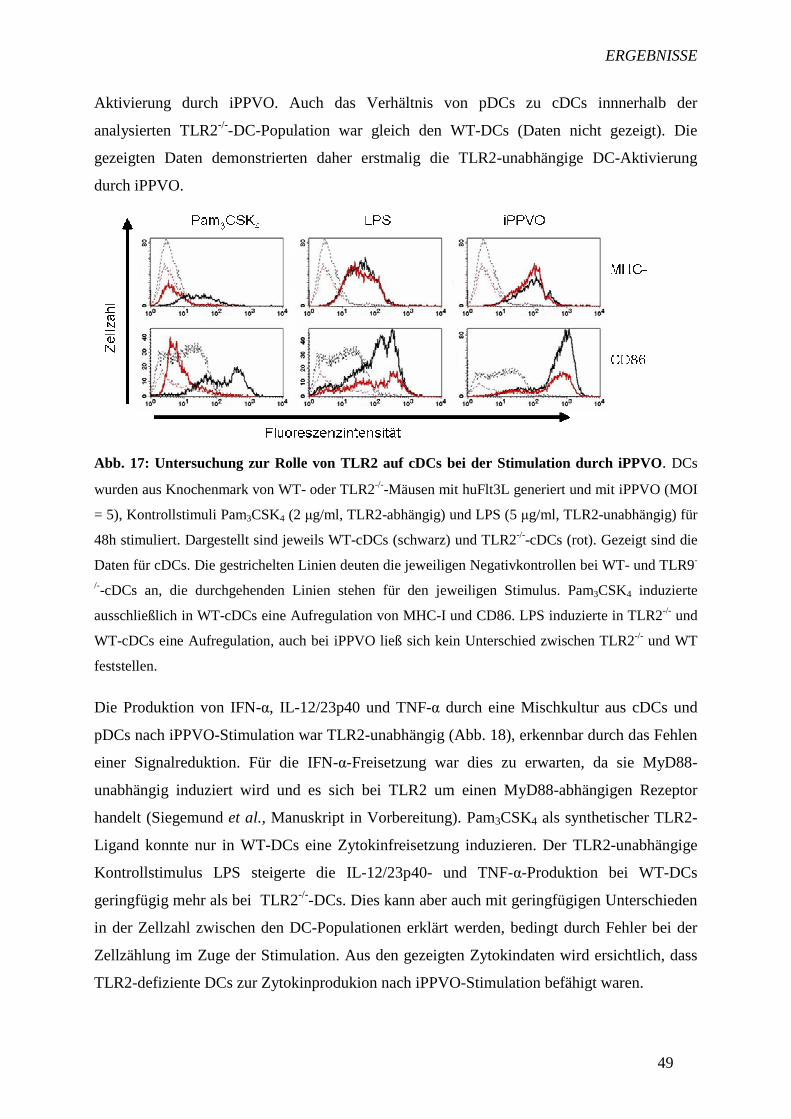
ERGEBNISSE
49
Aktivierung durch iPPVO. Auch das Verhältnis von pDCs zu cDCs innnerhalb der
analysierten TLR2-/--DC-Population war gleich den WT-DCs (Daten nicht gezeigt). Die
gezeigten Daten demonstrierten daher erstmalig die TLR2-unabhängige DC-Aktivierung
durch iPPVO.
Abb. 17: Untersuchung zur Rolle von TLR2 auf cDCs bei der Stimulation durch iPPVO. DCs
wurden aus Knochenmark von WT- oder TLR2-/--Mäusen mit huFlt3L generiert und mit iPPVO (MOI
= 5), Kontrollstimuli Pam3CSK4 (2 µg/ml, TLR2-abhängig) und LPS (5 µg/ml, TLR2-unabhängig) für
48h stimuliert. Dargestellt sind jeweils WT-cDCs (schwarz) und TLR2-/--cDCs (rot). Gezeigt sind die
Daten für cDCs. Die gestrichelten Linien deuten die jeweiligen Negativkontrollen bei WT- und TLR9-
/--cDCs an, die durchgehenden Linien stehen für den jeweiligen Stimulus. Pam3CSK4 induzierte
ausschließlich in WT-cDCs eine Aufregulation von MHC-I und CD86. LPS induzierte in TLR2-/- und
WT-cDCs eine Aufregulation, auch bei iPPVO ließ sich kein Unterschied zwischen TLR2-/- und WT
feststellen.
Die Produktion von IFN-α, IL-12/23p40 und TNF-α durch eine Mischkultur aus cDCs und
pDCs nach iPPVO-Stimulation war TLR2-unabhängig (Abb. 18), erkennbar durch das Fehlen
einer Signalreduktion. Für die IFN-α-Freisetzung war dies zu erwarten, da sie MyD88-
unabhängig induziert wird und es sich bei TLR2 um einen MyD88-abhängigen Rezeptor
handelt (Siegemund et al., Manuskript in Vorbereitung). Pam3CSK4 als synthetischer TLR2-
Ligand konnte nur in WT-DCs eine Zytokinfreisetzung induzieren. Der TLR2-unabhängige
Kontrollstimulus LPS steigerte die IL-12/23p40- und TNF-α-Produktion bei WT-DCs
geringfügig mehr als bei TLR2-/--DCs. Dies kann aber auch mit geringfügigen Unterschieden
in der Zellzahl zwischen den DC-Populationen erklärt werden, bedingt durch Fehler bei der
Zellzählung im Zuge der Stimulation. Aus den gezeigten Zytokindaten wird ersichtlich, dass
TLR2-defiziente DCs zur Zytokinprodukion nach iPPVO-Stimulation befähigt waren.
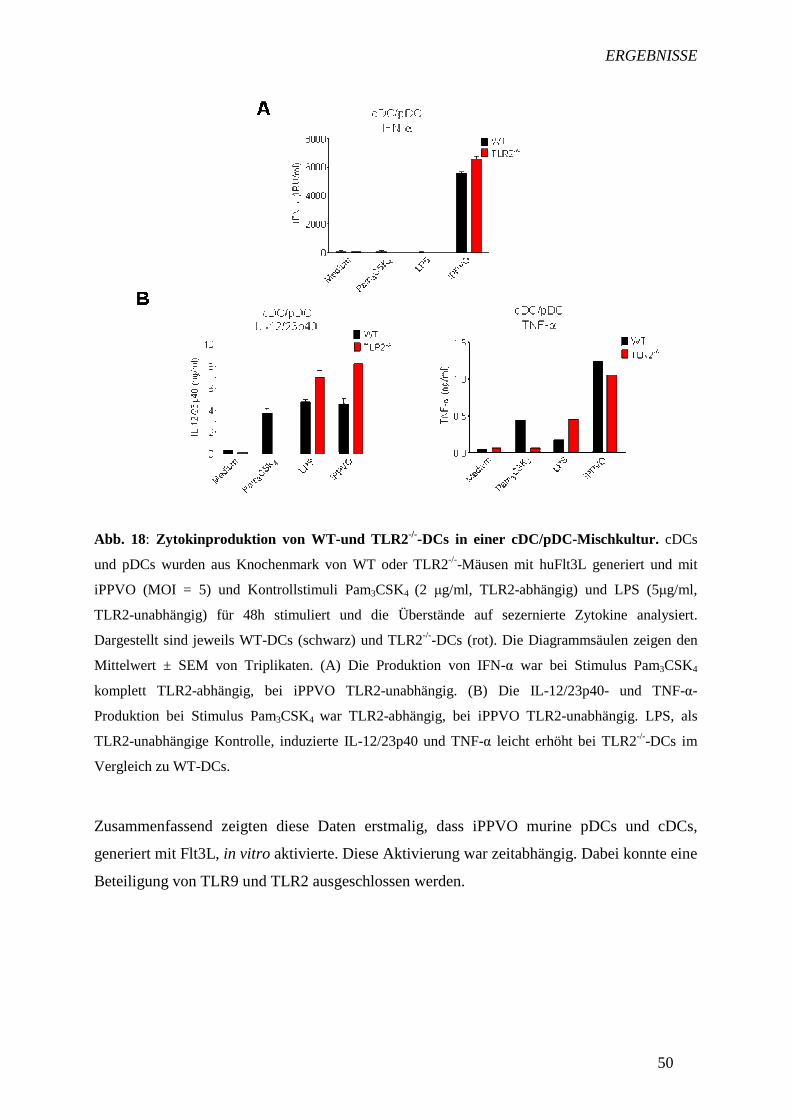
ERGEBNISSE
50
Abb. 18: Zytokinproduktion von WT-und TLR2 -/--DCs in einer cDC/pDC-Mischkultur. cDCs
und pDCs wurden aus Knochenmark von WT oder TLR2-/--Mäusen mit huFlt3L generiert und mit
iPPVO (MOI = 5) und Kontrollstimuli Pam3CSK4 (2 µg/ml, TLR2-abhängig) und LPS (5µg/ml,
TLR2-unabhängig) für 48h stimuliert und die Überstände auf sezernierte Zytokine analysiert.
Dargestellt sind jeweils WT-DCs (schwarz) und TLR2-/--DCs (rot). Die Diagrammsäulen zeigen den
Mittelwert ± SEM von Triplikaten. (A) Die Produktion von IFN-α war bei Stimulus Pam3CSK4
komplett TLR2-abhängig, bei iPPVO TLR2-unabhängig. (B) Die IL-12/23p40- und TNF-α-
Produktion bei Stimulus Pam3CSK4 war TLR2-abhängig, bei iPPVO TLR2-unabhängig. LPS, als
TLR2-unabhängige Kontrolle, induzierte IL-12/23p40 und TNF-α leicht erhöht bei TLR2-/--DCs im
Vergleich zu WT-DCs.
Zusammenfassend zeigten diese Daten erstmalig, dass iPPVO murine pDCs und cDCs,
generiert mit Flt3L, in vitro aktivierte. Diese Aktivierung war zeitabhängig. Dabei konnte eine
Beteiligung von TLR9 und TLR2 ausgeschlossen werden.

DISKUSSION
51
5 Diskussion
5.1 Charakterisierung der immunstimulatorischen Eigenschaften von inaktiviertem
Parapoxvirus ovis auf dendritische Zellen
5.1.1 Vorbereitende Untersuchungen zu DCs mit Flt3L und EMA
Dendritische Zellen (DCs) sind Teil des angeborenen Immunsystems und bilden daher eine
erste wichtige Immunbarriere gegen pathogene Erreger (25). In der vorliegenden Arbeit
sollten erstmalig die immunstimulatorischen Eigenschaften von iPPVO auf DCs
charakterisiert werden. Für die Generierung einer Mischpopulation aus pDCs und cDCs
wurden murine Knochenmarkszellen in Gegenwart von huFlt3L kultiviert. Die Existenz
dieser Subpopulationen ist sowohl für humane als auch murine DCs beschrieben (26). Daher
eignet sich das murine Modell für Untersuchungen mit iPPVO, auch wenn sich muDCs und
huDCs in der Expression einiger TLR unterscheiden (41). Über durchflusszytometrische
Analysen war es möglich, die cDCs getrennt von den pDCs zu analysieren. Für die
Untersuchungen von cDCs und pDCs wurde eine Lebend-/Tot-Färbung mit
Ethidiummonoazidbromid (EMA) für fixierte Zellen im Zuge der durchflusszytometrischen
Analyse etabliert. Es konnte gezeigt werden, dass sich über die EMA-Färbung effektiv vitale
von nicht-vitalen Zellen unterscheiden lassen.
Die Generierung von pDCs und cDCs in vitro mit Hilfe von rekombinantem humanem Flt3L
erwies sich als effiziente Methode, DCs in ausreichendem Maße zu gewinnen (Abb. 8). Im
Vergleich zwischen DCs, die mit humanem, rekombinanten Flt3L generiert wurden, und DCs,
die mit murinem, aus Überständen einer transfizierten Zellinie gewonnenem Flt3L generiert
wurden, zeigten sich keine Unterschiede in der Populationszusammensetzung. Auch fanden
sich keine Unterschiede in der Aktivierbarkeit der DCs (Abb. 12). Mit diesem Experiment
konnten mögliche abweichende Ergebnisse mit Zellen generiert unter Einsatz des
rekombinanten humanen bzw. aus Zellkultur gewonnenem murinem Flt3L ausgeschlossen
werden. HuFlt3L kann daher im murinen System, wie auch bereits beschrieben (32, 33), zur
Generierung von DCs eingesetzt werden. Erst kürzlich wurde die Generierung von pDCs und
cDCs mit Hilfe von rekombinantem M-CSF unabhängig von Flt3L im Zuge der
Makrophagenkultivierung beschrieben (68). Allerdings waren die Ausbeuten an pDCs und

DISKUSSION
52
cDCs innerhalb der Makrophagenkultur deutlich geringer als nach Kultivierung von
Knochenmark mit Flt3L (68).
5.1.2 iPPVO induziert die Expression kostimulatorischer Moleküle eher auf cDCs als
auf pDCs
Unter dem Einsatz von Flt3L-generierten DCs aus murinem Knochenmark und der etablierten
EMA-Lebend-/Tot-Färbung sollte erstmalig die immunstimulatorische Wirkung von iPPVO
auf diese wichtigen antigenpräsentierenden Zellen des angeborenen Immunsystems untersucht
werden. Bisher wurden ausschließlich Makrophagen, neurophile Granulozyten und natürliche
Killerzellen in Bezug auf iPPVO charakterisiert (23). Im natürlichen Wirt des PPVO-
Stammes D1701 wurde eine massive Ansammlung von DCs um die durch PPVO
verursachten Läsionen herum beschrieben. Daher konnte angenommen werden, dass auch
iPPVO DCs chemotaktisch anzieht und sie aktiviert (69). Selbst wenn eine Beteiligung von
DCs an der Entstehung der immunstimulatorischen Effekte durch iPPVO vermutet wurde,
fanden bis jetzt keine detaillierten Untersuchungen zu dieser Fragestellung statt. Aus den in
der vorliegenden Diplomarbeit gezeigten Daten wird deutlich, dass iPPVO mit
immunstimulatorischen Effekten auf DCs in vitro wirkt (Herden et al., 12. Symposium
„Infektion und Immunabwehr“ Burg Rothenfels, 7.- 9. März 2008). Diese Effekte beinhalten
eine verstärkte Expression der Oberflächenmoleküle MHC-I, MHC-II, CD86, CD80 und
CD40 auf cDCs und pDCs. Um ihre Rolle als antigenpräsentierende und T-Zellen
aktivierende Zellen erfüllen zu können, müssen DCs selbst aktiviert werden. Diese DC-
Aktivierung kann über die Wirkung von Zytokinen oder von Pathogen-assoziierten Stimuli
induziert werden. Nur aktivierte DCs exprimieren verstärkt kostimulatorische Moleküle wie
CD86, CD80 und CD40 (Abb. 10) und sind dann in der Lage, T-Zellen zu aktivierten (25).
Somit deuten die gezeigten Daten darauf hin, dass iPPVO in vitro über die Stimulation von
DCs eine T-Zellaktivierung hervorrufen kann. Es muss noch gezeigt werden, ob diese
Aufregulation der Oberflächenmoleküle auf DCs direkt durch iPPVO oder indirekt durch im
Zuge der iPPVO-induzierten DC-Stimulation produzierte Zytokine wie IFN-α, IL-12/23p40
und TNF-α verursacht wird (Abb. 11). Die Aufregulation ist verstärkt bei cDCs zu
beobachten im Vergleich zu pDCs (Abb. 9 und 10). Dieses Ergebnis deckt sich mit bisherigen
Untersuchungen, die auf eine weniger effektive Aktivierung von T-Zellen durch pDCs als
durch cDCs hindeuten (29). Damit scheinen cDCs die potenteren Aktivatoren von T-Zellen
im Zuge einer iPPVO-Applikation zu sein.

DISKUSSION
53
Des Weiteren stellt sich die Frage, ob iPPVO direkt die Aktivierung beider Subpopulationen,
cDCs und pDCs, bewirkt oder ob pDCs zur Aktivierung von cDCs beitragen und umgekehrt.
Für die Infektion von DCs, generiert aus murinem Knochenmark, mit gamma-Herpesvirus 68
wurde eine pDC-abhängige cDC-Aktivierung über eine verstärkte Expression von CD86,
CD80 und CD40 auf cDCs nachgewiesen (70). Mögliche Priming-Effekte dieser Art
zwischen pDCs und cDCs nach Stimulation mit iPPVO könnten durch
Separationsexperimente überprüft werden. Dabei wird aus der Mischkultur von pDCs und
cDCs über eine in einem magnetischen Feld befindliche Säule mit Hilfe von Antikörper-
gekoppelten Magnetkügelchen eine Zellpopulation depletiert, so dass eine reine cDC- bzw.
pDC-Kultur entsteht. Erste Untersuchungen mit dieser Methode weisen auf Priming-Effekte
zwischen pDCs und cDCs bei der iPPVO-induzierten Aktivierung hin (Siegemund et al.,
Manuskript in Vorbereitung).
5.1.3 Der Einfluss von IFN-α auf die durch iPPVO-ausgelöste DC-Aktivierung
Die Mischkultur von cDCs und pDCs produzierte große Mengen an IFN-α nach iPPVO-
Stimulation (Abb. 11). Über dieses antivirale Zytokin werden verschiedene
Effektormechanismen aktiviert, um die Replikation der Viren in ihren Wirtszellen zu
verhindern, z.B. die verstärkte Expression der eIF2α-Kinase und der 2’5’-
Oligoadenylatsynthetase (43). In seiner Funktion als Immunmodulator hilft die durch iPPVO-
induzierte IFN-α-Produktion durch DCs dem mit Zylexis® behandelten Tier, bestehende
virale Infektionen zu überwinden. Erste Daten einer intrazellulären IFN-α-Markierung deuten
darauf hin, dass pDCs verstärkt IFN-α nach Kontakt mit iPPVO synthetisieren (Siegemund et
al., Manuskript in Vorbereitung). Neben pDCs produzieren überraschenderweise auch cDCs
hohe Mengen an IFN-α nach iPPVO-Stimulation. Es bleibt jedoch offen, ob sowohl pDCs als
auch cDCs IFN-α unabhängig voneinander produzieren oder ob die IFN-α-Produktion der
einen Subpopulation zur IFN-α-Induktion in der anderen Subpopulation führt (Abb. 19).
Autokrine und parakrine Effekte zwischen pDCs und cDCs können durch
Separationsexperimente analysiert werden (5.1.2). Erste Resultate deuten allerdings auf keine
vorhandenen Priming-Effekte zwischen pDCs und cDCs hinsichtlich der IFN-α-Produktion
hin (Siegemund et al., Mauskript in Vorbereitung). In Bezug auf autokrine oder parakrine
Rückkopplungsschleifen des nach iPPVO-Stimulation gebildeten IFN-α hat sich für mit
modifizierten VV (Typ Ankara)-stimulierte DCs gezeigt, dass Rückkopplungsschleifen über
den IFN-α/β-Rezeptor existieren. Für die Produktion von IFN-α nach VV-Inkubation war die

DISKUSSION
54
Expression des IFN-α/β-Rezeptors essentiell, nicht aber die Produktion von IFN-β (71). Daher
könnte eine Rückkopplungsschleife über IFN-α bestehen und dieser Aspekt auch auf das mit
VV verwandte iPPVO zutreffen.
Eine Freisetzung von IFN-α durch DCs kann neben direkten antiviralen Effektorfunktionen,
wie in 1.2.3 beschrieben, auch indirekte Auswirkungen auf die Aktivierung der DCs haben.
Die verstärkte Expression von kostimulatorischen Oberflächenmolekülen im Zuge einer
iPPVO-induzierten DC-Aktivierung kann nicht nur direkt über die Signalweiterleitung eines
TLR, sondern auch indirekt über das produzierte IFN-α hervorgerufen werden (72, 73) (Abb.
19). Typ I-IFN wie IFN-α können die DC-Reifung über die Aufregulation von
kostimulatorischen Oberflächenmolekülen wie CD86, CD80 und CD40 einleiten (73). Daher
sind noch Experimente mit IFN-α/β-Rezeptor-defizienten DCs notwendig um festzustellen,
ob iPPVO die Aufregulation über den direkten (TLR) oder indirekten (IFN-α) Weg induziert.
5.1.4 Rolle von IFN-β für die DC-Effektorantwort nach Kontakt mit iPPVO
Neben IFN-α ist auch IFN-β für die antivirale Effektorantwort von Zellen des angeborenen
Immunsystems von entscheidender Bedeutung. IFN-β kann über den IFN-α/β-Rezeptor IFN-α
induzieren (53). Bei Untersuchungen mit durch VV stimulierten DCs war IFN-β entscheidend
für die TLR2-/MyD88-abhängige Produktion proinflammatorischer Zytokine. Weiterhin
konnte IFN-β indirekt zur Aktivierung adaptiver Immunantworten nach VV-Infektion
beitragen. Entgegen den Erwartungen wurde kein IFN-α durch DCs, stimuliert mit VV,
produziert (56). In weiteren Versuchen muss geklärt werden, ob DCs und gegebenenfalls
welche der beiden Subpopulationen nach iPPVO-Stimulation ebenfalls IFN-β freisetzen
sowie welche Effekte IFN-β auf die Effektorantwort von DCs auf iPPVO hat.
5.1.5 Die iPPVO-induzierte DC-Aktivierung führt zur Freisetzung von IL-12/23p40 und
TNF-α
Die Freisetzung von antiviralen und proinflammatorischen Zytokinen durch Zellen des
angeborenenen Immunsystems ist entscheidend für den Verlauf von viralen Infektionen
(Punkt 1.2.3) In Bezug auf iPPVO kommt es in vitro zur Freisetzung von IL-12/23p40 und
TNF-α durch aktivierte DCs. Weitere Untersuchungen sind notwendig, um zu klären, ob
cDCs und pDCs oder nur eine der beiden DC-Subpopulationen IL-12/23p40 und TNF-α nach

DISKUSSION
55
Inkubation mit iPPVO freisetzen. Erste Analysen mit intrazellulärer Färbung von IL-12/23p40
zeigen cDCs als dessen Hauptproduzenten (Siegemund et al., Manuskript in Vorbereitung).
Des Weiteren kann die Produktion von proinflammatorischen Zytokinen auch indirekt über
Typ I-IFN induziert werden (Abb. 19). Bei der durch VV verursachten DC-Aktivierung
wurde eine Abhängigkeit der IL-6-Freisetzung durch DCs von Typ I-IFN wie IFN-α und IFN-
β nachgewiesen (56). Um zu klären, ob die Produktion von IL-12/23p40 und TNF-α über
IFN-α induziert wird, könnten Untersuchungen in vitro an IFN-α/β-Rezeptor-defizienten DCs
durchführt werden.
5.1.6 Unterschiedliche Kinetik der durch iPPVO induzierten DC-Aktivierungsmarker
Die Aktivierung der DCs durch iPPVO unterliegt einem kinetischen Verlauf. Erst nach einer
Zeitspanne von 12-24 h Stimulation war eine verstärkte Expression der Oberflächenmoleküle
MHC-I, MHC-II, CD86, CD80 und CD40 auf cDCs und pDCs sowie die Produktion von IL-
12/23p40 und IFN-α durch die DC-Mischpopulation detektierbar (Abb. 14). Freigesetztes
TNF-α hingegen wurde schon nach 6 h Stimulation nachgewiesen. Diese schnelle Produktion
von TNF-α im Vergleich zu IFN-α und IL-12/23p40 könnte zum einen an einer schnelleren
Synthese von TNF-α durch iPPVO-aktivierte DCs liegen, die keiner autokrinen
Aktivierungssignale durch andere Mediatoren wie IFN-α bedarf. In Untersuchungen zu
immunmodulatorischen Eigenschaften von Viren konnte gezeigt werden, dass Viren
Mechanismen besitzen, um eine TNF-α-Produktion durch Immunzellen des Wirts zu
unterbinden (74, 75). So verhinderten VV in epidermalen DCs aktiv eine TNF-α-Produktion
(74). Humaner Cytomegalovirus konnte über eine Inhibition der NF-κB-Aktivierung die
Synthese von TNF-α in humanen Fibroblastenzellen unterbinden (75). Diese viralen
Mechanismen zum Verhindern einer TNF-α-Produktion in infizierten Wirtszellen deutet auf
eine frühe Rolle von TNF-α in der Virusabwehr hin. Tatsächlich wurde für humane
Cytomegalo-Viren weiterhin beschrieben, dass die TNF-α-Produktion schon in der initialen
Aktivierungsphase der Wirtszelle durch die Virusanheftung und den Viruseintritt aktiviert
wird (76). Zusätzlich könnte bei der iPPVO-induzierten DC-Aktivierung schon vorhandenes
TNF-α notwendig für das Einsetzen der IL-12/23p40- und IFN-α-Produktion sein. Im Zuge
der Infektion von Zellen der humanen epithelialen Lungenzellinie A549 durch Influenza-A-
Viren konnte gezeigt werden, dass TNF-α zu einer verstärkten Expression der Gene von RIG-
I, TLR3, MyD88, TRIF und IRF7 führt (77). Diese Resultate zeigen einen direkten Einfluss
von TNF-α auf die Induktion der Produktion von Signalkomponenten sowohl vom TLR-
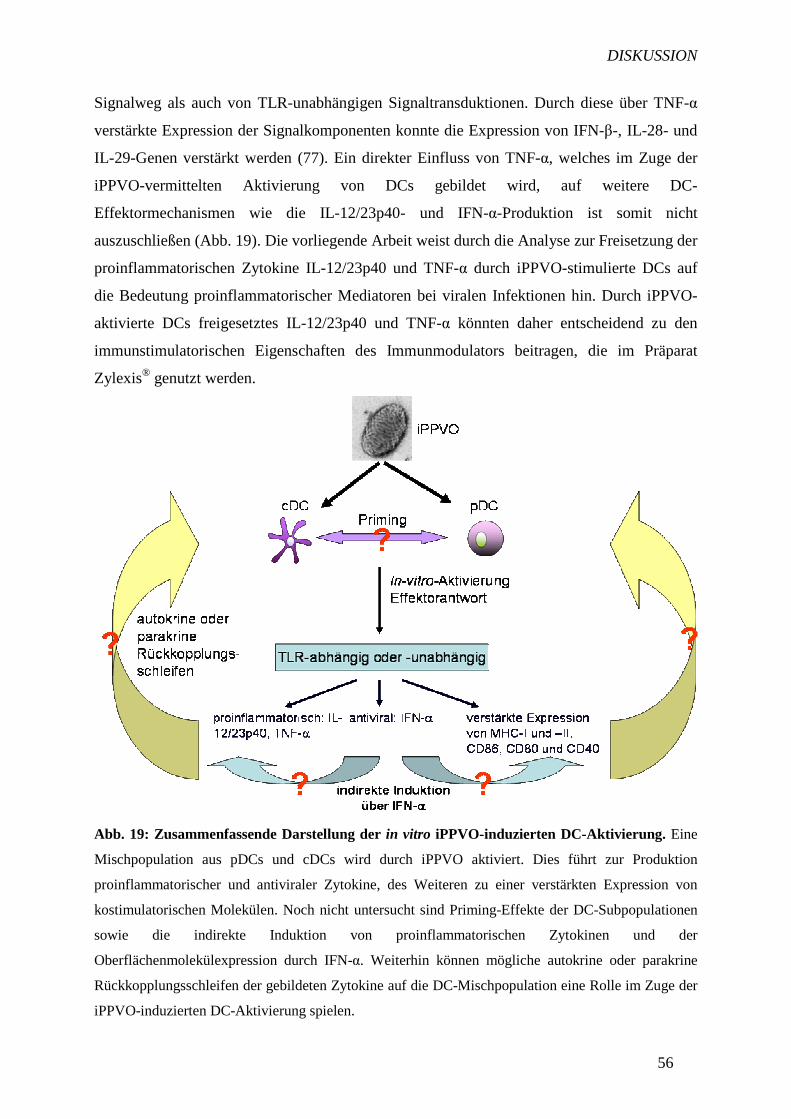
DISKUSSION
56
Signalweg als auch von TLR-unabhängigen Signaltransduktionen. Durch diese über TNF-α
verstärkte Expression der Signalkomponenten konnte die Expression von IFN-β-, IL-28- und
IL-29-Genen verstärkt werden (77). Ein direkter Einfluss von TNF-α, welches im Zuge der
iPPVO-vermittelten Aktivierung von DCs gebildet wird, auf weitere DC-
Effektormechanismen wie die IL-12/23p40- und IFN-α-Produktion ist somit nicht
auszuschließen (Abb. 19). Die vorliegende Arbeit weist durch die Analyse zur Freisetzung der
proinflammatorischen Zytokine IL-12/23p40 und TNF-α durch iPPVO-stimulierte DCs auf
die Bedeutung proinflammatorischer Mediatoren bei viralen Infektionen hin. Durch iPPVO-
aktivierte DCs freigesetztes IL-12/23p40 und TNF-α könnten daher entscheidend zu den
immunstimulatorischen Eigenschaften des Immunmodulators beitragen, die im Präparat
Zylexis® genutzt werden.
Abb. 19: Zusammenfassende Darstellung der in vitro iPPVO-induzierten DC-Aktivierung. Eine
Mischpopulation aus pDCs und cDCs wird durch iPPVO aktiviert. Dies führt zur Produktion
proinflammatorischer und antiviraler Zytokine, des Weiteren zu einer verstärkten Expression von
kostimulatorischen Molekülen. Noch nicht untersucht sind Priming-Effekte der DC-Subpopulationen
sowie die indirekte Induktion von proinflammatorischen Zytokinen und der
Oberflächenmolekülexpression durch IFN-α. Weiterhin können mögliche autokrine oder parakrine
Rückkopplungsschleifen der gebildeten Zytokine auf die DC-Mischpopulation eine Rolle im Zuge der
iPPVO-induzierten DC-Aktivierung spielen.

DISKUSSION
57
5.2 iPPVO wird endozytotisch durch dendritische Zellen aufgenommen
Über elektronenmikroskopische Aufnahmen konnte die endozytotische Aufnahme von
iPPVO-Virionen durch DCs dokumentiert werden (Abb. 13). Die endozytotische Aufnahme
von festen Partikeln wie Virionen wird dabei als Phagozytose bezeichnet. Routinemäßig
werden in der Elektronenmikroskopie (EM) die PPVO-Partikel im „negative staining“-
Verfahren mit Phosphorwolframsäure kontrastiert. Damit können die ovalen Virionen mit
den, im Gegensatz zu den Orthopocken, charakteristisch regelmäßig angeordneten
Coreproteinfilamenten sichtbar und unterscheidbar gemacht werden (78). Die in der
vorliegenden Arbeit gezeigten EM-Aufnahmen lassen diese charakteristische
Oberflächenstruktur von iPPVO erkennen (Abb. 13D).
Die Mehrheit von Virionen nach lytischer Virusvermehrung und Virusernte in permessiven
Zellen sind intrazelluläre, reife Virionen (IMV) (Prof. Büttner, schriftliche Mitteilung). Durch
die Adaption von PPVO-D1701 an die bovine Nierenzellinie BKKL3A mit anschließender
Isolation und Inaktivierung kann man davon ausgehen, dass das vorliegende Präparat
vorwiegend aus IMV besteht. Daher sind in elektronenmikroskopischen Aufnahmen kaum
EEV von iPPVO zu finden, zumal durch die Präparationstechnik die äußere Hülle der EEV
leicht zerstört werden kann (Prof. Büttner, schriftliche Mitteilung). Deshalb sind für die
Aufnahme von iPPVO durch DCs nur in der Literatur diskutierte Aufnahmemechanismen für
IMV möglich. Bisher führte man Analysen zum Infektionszyklus und damit zur Aufnahme
der Virionen in Wirtszellen anhand vom Modellvirus der Pockenviren, den VV, durch. Dabei
werden verschiedene Aufnahmemechanismen der Virionen nach der Anheftung an die
Wirtszelle für IMV diskutiert. Dazu zählen bespielsweise die Internalisierung (10) und
fusionsunabhängige Mechanismen (11), da nur die äußere Membran der EEV als fusiogen
gilt. Diese Mechanismen können auch für die Aufnahme der iPPVO-Virionen in DCs in Frage
kommen. Unter der Berücksichtigung, dass es sich bei murinen Flt3L-DCs mit hoher
Wahrscheinlichkeit nicht um permissive Zielzellen von PPVO handelt, könnte selbst für
iPPVO nach Kontakt mit permissiven epidermalen Keratinocyten von natürlichen Wirten
(Ziegen und Schafe) ein anderer Aufnahmemechanismus als bei DCs vorliegen.
Weiterführende Untersuchungen müssen klären, ob es sich bei der Aufnahme von iPPVO um
eine Rezeptor-vermittelte Endozytose handelt. Durch die chemische Inaktivierung von PPVO
mit binären Ethyliminen oder β-Propiolakton kann man davon ausgehen, dass die viralen

DISKUSSION
58
Hüllproteine nicht verändert sind (20, 21). Ein Viruseintritt in die Wirtszelle wird generell
von der Interaktion von viralen Proteinen mit spezifischen Zelloberflächenproteinen oder
Polysacchariden begleitet (79). Rezeptoren können die Anheftung der viralen Partikel an die
Zellmembran verstärken, Signalkaskaden aktivieren und metastabile virale Fusionsproteine
aktivieren. Es gibt Hinweise, dass virale Proteine von VV an
Zelloberflächenglucosaminglycane binden und damit der Viruseintritt in die Zelle erleichtert
wird (80). Weiterhin muss die Rolle von Lipid-Flößen (lipid rafts) bei der Aufnahme von
iPPVO analysiert werden. Für Pockenviren wurde bisher gezeigt, dass eine Depletion von
Cholesterin in der Membran die Aufnahme der Viruspartikel verhindert (81).
Für die Untersuchungen zur Rezeptor-vermittelten Endozytose empfiehlt sich der Einsatz von
Immunelektronenmikroskopie, eventuell gekoppelt mit Fluoreszenzresonanzenergietransfer,
und/oder Konfokalmikroskopie (82). Um hochauflösende Studien über strukturelle
Rearrangements von Viren oder subviralen Partikeln während bzw. nach der Zellaufnahme
oder Rezeptorbindung durchzuführen, könnten Röntgenanalysen (83) oder sogenannte
Einzelpartikel-Rekonstruktionen mittels Cryo-Elektronenmikroskopie durchgeführt werden
(84).
5.3 Beteiligung von Toll-ähnlichen Rezeptoren an der durch inaktiviertes Parapoxvirus
ovis induzierten Aktivierung dendritischer Zellen
5.3.1 Die DC-Aktivierung durch iPPVO ist TLR9-unabhängig
Für die vollständige Aufklärung des Wirkungsmechanismus von iPPVO auf DCs ist es
unerlässlich, den (die) Rezeptor(en) für die virale Komponente(n) auf den DCs zu
identifizieren. Bisher wurde die Rolle von TLR besonders im Zusammenhang mit Infektionen
mit Bakterien und Pilzen untersucht. Zunehmend kommt aber auch die Bedeutung dieser
Rezeptoren bei der Abwehr viraler Infektionen zum Vorschein (54). TLR9 erkennt
unmethylierte 2’-Desoxyribo-CpG-DNA-Motive, welche sowohl in bakterieller als auch
viraler DNA vorkommen (85). Allerdings mehren sich die Hinweise, dass auch methylierte
Motive erkannt werden (Prof. Alber, mündliche Mitteilung). Zudem sind synthetische CpG-
Oligonukleotide bekannt für die Aktivierung der TLR9-abhängigen IFN-α-Produktion in DCs
(86). Damit bot sich TLR9 als möglicher Kandidat für die Aktivierung von DCs durch
iPPVO-dsDNA an.

DISKUSSION
59
TLR9 kommt ausschließlich in Endosomen vor (43). Nach Behandlung von pDCs mit
Inhibitoren der Lysosomenfunktion wurden die TLR9-abhängigen Signale nach HSV-I-
Inkubation komplett unterbunden (43). Dies deutet darauf hin, dass virale Nukleinsäuren in
das endosomale Kompartiment über Endozytose gelangen, wo ein endosomaler Rezeptor wie
TLR9 die freigesetzten Nukleinsäuren erkennt und intrazelluläre Signaltransduktion aktiviert.
Bei den analysierten elektronenmikroskopischen Aufnahmen konnten Anzeichen einer
endozytotischen Aufnahme von iPPVO-Virionen detektiert werden, die sich noch in weiteren
Experimenten bestätigen müssen. Daher ist die Möglichkeit, dass die virale dsDNA ins
Endosom für die Erkennung durch TLR9 gelangt, gegeben.
Analysen von TLR9-defizienten Mäusen haben gezeigt, dass TLR9 für die Produktion von
IFN-α durch DCs als antivirale Antwort auf DNA-Viren wie Mäuse-Cytomegalo-Virus
(MCMV), Herpes-Simplex-Virus-I (HSV-I) und HSV-II notwendig ist (67, 86). Inaktiviertes
HSV-2 oder gereinigte genomische HSV-2 DNA konnten auch die IFN-α-Produktion TLR9-
abhängig aktivieren (67). Die in dieser Arbeit vorliegenden Daten zeigen jedoch zum ersten
Mal, dass die Aktivierung von DCs durch iPPVO in vitro TLR9-unabhängig erfolgt (Abb. 15
und 16). Dieser Befund weist darauf hin, dass die Erkennungsmechanismen auf DCs für Viren
innerhalb einer Virusfamilie trotz Verwandtschaft unterschiedlich sein können.
Isolierte iPPVO-DNA induzierte keine Aktivierungssignale in DCs über TLR9 (Abb. 15B und
16). Dieses Ergebnis könnte durch eine fehlende Aufnahme eines mit 140 kbp so großen
DNA-Moleküls durch DCs entstanden sein. Jedoch konnte bei Untersuchungen mit
genomischer HSV-2-DNA mit einer Größe von 154 kbp eine deutliche IFN-α-Produktion
durch pDCs detektiert werden ohne den Einsatz einer Transfektion (67). Trotz ihrer
Verwandtschaft können sich die DNAs verschiedener DNA-Viren hinsichtlich ihrer Größe
und Basenzusammensetzung sehr unterscheiden. Daher kann die DNA eines DNA-Virus wie
HSV-2 stimulatorisch, virale DNA von iPPVO nicht stimulatorisch auf DCs wirken. Darüber
hinaus handelt es sich bei iPPVO-DNA um methylierte DNA. Bisher jedoch wurde die
Erkennung von unmethylierten 2’-Desoxyribo-CpG-DNA-Motiven durch TLR9 beschrieben
(85), auch wenn sich Hinweise vermehren auf die Erkennung von methylierter DNA (Prof.
Alber, mündliche Mitteilung). Es ist damit nicht ausgeschlossen, dass TLR9 für die
Aktivierung von DCs durch nicht-inaktiviertes PPVO von Bedeutung sein könnte, auch wenn
für inaktiviertes PPVO mit methylierter DNA dieses Resultat, wie in dieser Arbeit gezeigt,
nicht zutrifft. Weiterhin besteht die Möglichkeit, dass es sich bei der viralen dsDNA von

DISKUSSION
60
iPPVO nicht um die immunstimulatorische Komponente handelt, die zur DC-Aktivierung
führt. Zur Bestätigung dieser These bedarf es neben den hier gezeigten Daten zu TLR9 noch
fortführender Experimente, da neben TLR9 für die DNA-Erkennung auch zytosolische, TLR-
unabhängige Rezeptoren in Frage kommen wie der kürzlich beschriebene DNA-abhängige
Aktivator von IFN-regulatorischen Faktoren (DAI) (52). DAI bindet dsDNA, was zur
Assoziation mit IRF3 mit folgender IFN-Freisetzung führt. Damit könnte DAI als
zytosolischer DNA-Sensor eine entscheidende Rolle in der Aktivierung angeborener
Immunantworten spielen (52).
Da die Produktion von IFN-α TLR9-unabhängig nach iPPVO-Stimulation induziert wird,
stellt sich die Frage, über welche Mechanismen die Bildung von IFN-α erfolgt. pDCs gelten
als die einzigen Zellen mit der Fähigkeit, große Mengen IFN-α ohne vorherige Produktion
von IFN-β über die Aktivierung von TLR7 und TLR9 freizusetzen. Bisher konnte weiterhin
gezeigt werden, dass die Typ I-Interferon-Produktion in pDCs unabhängig vom RIG-
I/MDA5-Signalweg geschieht (43). Es ist jedoch nicht auszuschließen, dass pDCs große
Mengen an IFN-α über TLR- und RIG-I/MDA5-unabhängige Signalwege, wie z.B. mit DAI-
assoziierte Transduktionen, freisetzen können. Daher ist auch ein noch unbekannter TLR9-
und RIG-I/MDA5-unabhängiger Mechanismus zur IFN-α-Produktion in pDCs durch iPPVO
möglich. Ein solcher neuer Mechanismus wurde für pDCs bei Infektion mit dem
modifizierten VV-Ankara gezeigt. Die IFN-α-Freisetzung der pDCs geschieht nach
Stimulation mit VV-Ankara hauptsächlich TLR-unabhängig (71). Überdies wurde
dokumentiert, dass neben pDCs auch andere Zelltypen wie cDCs und Makrophagen IFN-α als
Antwort auf HSV-I-Applikation produzieren. Diese Produktion von IFN-α erfolgte durch
pDCs und cDCs allerdings vollständig TLR9-unabhängig. Herausragend ist, dass dieses
TLR9-unabhängig gebildete IFN-α den Hauptteil des gesamten, von Knochenmarkszellen
gebildeten IFN-α nach HSV-I-Stimulation ausmachte (54).
Bei den Untersuchungen mit HSV-I zur IFN-α-Produktion spielte die Herkunft der pDCs eine
entscheidende Rolle (45). So konnten mit Flt3L generierte pDCs aus Knochenmark IFN-α
auch TLR9-unabhängig herstellen, während die IFN-α-Produktion isolierter pDCs aus der
Milz sich als komplett TLR9-abhängig herausstellte (45). Die hier gezeigten Experimente
wurden mit Flt3L-generierten DCs durchgeführt und ergaben eine TLR9-unabhängige IFN-α-
Produktion. Es ist aber nicht ausgeschlossen, dass die IFN-α-Produktion von DCs anderer
Herkunft nach iPPVO-Stimulation TLR9-abhängig verläuft. Weitere Studien in vivo sind

DISKUSSION
61
notwendig, um den Mechanismus der iPPVO-induzierten IFN-α-Produktion in pDCs und
cDCs aufzuklären.
5.3.2 Die DC-Aktivierung nach iPPVO-Stimulation ist TLR2-unabhängig
Vergleichende Untersuchungen mit MyD88-defizienten DCs ergaben eine partielle MyD88-
Abhängigkeit der IL-12/23p40- und TNF-α-Produktion von iPPVO-aktivierten DCs
(Siegemund et al., Manuskript in Vorbereitung). Demnach mussten ein oder mehrere TLR an
der Induktion von IL-12/23p40 und TNF-α in DCs durch iPPVO beteiligt sein. Nach dem
Ausschluss von TLR9 als möglicher Rezeptor für die DC-Aktivierung durch iPPVO-DNA
wurde TLR2 näher betrachtet. Dies geschah auf der Grundlage, dass TLR2 zusammen mit
TLR9 bei HSV in vitro für die Produktion von IL-6 und IL-12 durch DCs eine wichtige Rolle
spielt (88). Weiterhin ist TLR2 von Bedeutung bei der Aktivierung von DCs durch VV (56).
In derselben Studie wurden proinflammatorische Zytokine TLR2- und MyD88-abhängig,
IFN-β hingegen TLR2-unabhängig von DCs, die aus murinem Knochenmark mit GM-CSF
und IL-4 generiert wurden, produziert. UV-behandeltes VV führte zur gleichen
Immunantwort wie unbehandeltes Virus. Daher schlussfolgerte man, dass der virale Ligand
für TLR2 ein Hüll- oder Coreprotein statt eines neu synthetisierten viralen Produktes im Zuge
der Replikation im Zytoplasma sein muss (56). Die Identifizierung eines solchen Liganden
würde die Herstellung effektiver und gleichzeitig immunstimulatorischer Vakzine
ermöglichen. iPPVO, durch Methylierung der DNA nicht mehr zur Replikation befähigt,
synthetisiert keine viralen Produkte im Zuge einer zytoplasmatischen Vermehrung. Hüll- oder
Coreproteine könnten jedoch als TLR2-Liganden in Frage kommen (Abb. 3A). Eine
endozytotische Aufnahme der iPPVO-Virionen ist dabei für die Erkennung durch TLR2 auf
DCs nicht erforderlich, da TLR2 auf der Plasmamembran exprimiert wird (40). Anhand der
bisherigen Untersuchungen konnte gezeigt werden, dass die Aktivierung von DCs durch
iPPVO in vitro neben TLR9- auch TLR2-unabhängig erfolgt (Abb. 17 und 18). Dieses
Ergebnis demonstriert, dass trotz der bestehenden Verwandtschaft zwischen iPPVO und VV
die Aktivierung der DCs durch diese Viren über verschiedene Mechanismen verläuft.
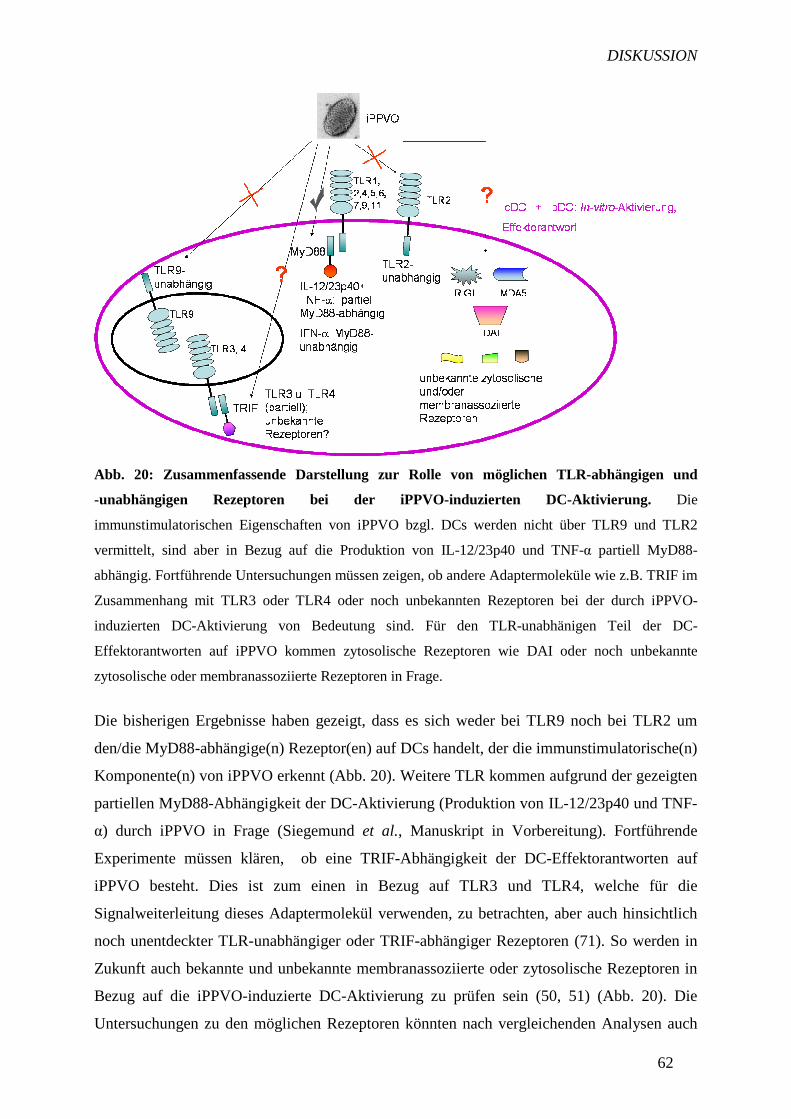
DISKUSSION
62
Abb. 20: Zusammenfassende Darstellung zur Rolle von möglichen TLR-abhängigen und
-unabhängigen Rezeptoren bei der iPPVO-induzierten DC-Aktivierung. Die
immunstimulatorischen Eigenschaften von iPPVO bzgl. DCs werden nicht über TLR9 und TLR2
vermittelt, sind aber in Bezug auf die Produktion von IL-12/23p40 und TNF-α partiell MyD88-
abhängig. Fortführende Untersuchungen müssen zeigen, ob andere Adaptermoleküle wie z.B. TRIF im
Zusammenhang mit TLR3 oder TLR4 oder noch unbekannten Rezeptoren bei der durch iPPVO-
induzierten DC-Aktivierung von Bedeutung sind. Für den TLR-unabhänigen Teil der DC-
Effektorantworten auf iPPVO kommen zytosolische Rezeptoren wie DAI oder noch unbekannte
zytosolische oder membranassoziierte Rezeptoren in Frage.
Die bisherigen Ergebnisse haben gezeigt, dass es sich weder bei TLR9 noch bei TLR2 um
den/die MyD88-abhängige(n) Rezeptor(en) auf DCs handelt, der die immunstimulatorische(n)
Komponente(n) von iPPVO erkennt (Abb. 20). Weitere TLR kommen aufgrund der gezeigten
partiellen MyD88-Abhängigkeit der DC-Aktivierung (Produktion von IL-12/23p40 und TNF-
α) durch iPPVO in Frage (Siegemund et al., Manuskript in Vorbereitung). Fortführende
Experimente müssen klären, ob eine TRIF-Abhängigkeit der DC-Effektorantworten auf
iPPVO besteht. Dies ist zum einen in Bezug auf TLR3 und TLR4, welche für die
Signalweiterleitung dieses Adaptermolekül verwenden, zu betrachten, aber auch hinsichtlich
noch unentdeckter TLR-unabhängiger oder TRIF-abhängiger Rezeptoren (71). So werden in
Zukunft auch bekannte und unbekannte membranassoziierte oder zytosolische Rezeptoren in
Bezug auf die iPPVO-induzierte DC-Aktivierung zu prüfen sein (50, 51) (Abb. 20). Die
Untersuchungen zu den möglichen Rezeptoren könnten nach vergleichenden Analysen auch

DISKUSSION
63
auf lebendes, nicht inaktiviertes PPVO übertragen werden und weitere Informationen über die
Pathogenese von PPVO-Infektionen liefern.
5.3.3 Weiterführende Untersuchungen an iPPVO mit neuen Anwendungsbereichen
Es scheint, dass Untersuchungsergebnisse von anderen dsDNA-Viren, selbst aus der gleichen
Familie, nicht auf alle Vertreter übertragbar sind. Eine Pauschalisierung, beispielsweise auch
bezüglich des Infektionszyklus von VV, auf alle Vertreter der Pockenviren, sollte daher
überdacht werden. Die dokumentierte partielle MyD88-Abhängigkeit der Aktivierung von
DCs durch iPPVO (Siegemund et al., Manuskript in Vorbereitung) beweist, dass ein
Immunmodulator wie iPPVO sowohl TLR-abhängige als auch TLR-unabhängige
immunologische Erkennungswege aktivieren kann und somit eine Verstärkung der
Wirtsimmunantwort erreicht. Dieses Erkenntnis könnte ein Ausgangspunkt für die
Entwicklung effektiver neuer Impfvektoren sein, die neben dem zu exprimierenden
Impfantigenen immunstimulatorische Elemente beinhalten, die eine effektivere Vakzinierung
und damit besseren Schutz vor Infektionen vermitteln als herkömmliche Impfstoffe. Für
PPVO wäre ein Einsatz als Vektor für mikrobielle Antigene mit gleichzeitigen
immunstimulatorischen Eigenschaften denkbar.
Virale Infektionen sind auch heute noch schwer therapierbar, da nur wenige antivirale
Medikamente zur Verfügung stehen. Überdies beeinflussen virale Infektionen den klinischen
Verlauf von gleichzeitigen Infektionen durch andere Pathogene. Die immunstimulatorischen
Eigenschaften des iPPVO werden daher gezielt in der Veterinärmedizin im Präparat Zylexis®,
früher als Baypamune® bekannt, zur prophylaktischen und therapeutischen Unterstützung von
Infektionen und stressbedingten Erkrankungen bei zahlreichen Nutztieren eingesetzt. Zylexis®
zeichnet sich durch geringe Nebenwirkungen aus. Während rekombinant hergestellte
Zytokine durch ihre geringe Halbwertszeit in hohen Dosen verabreicht werden müssen und
ihre Anwendung daher oft Nebenwirkungen mit sich bringen, führt iPPVO zu einer
regulierten Zytokinantwort. Diese beinhaltet gleich mehrere Zytokine parallel und unterstützt
damit die antivirale Immunantwort gegen eine bereits bestehende Infektion (19). Daher ist der
immuntherapeutische Ansatz mit iPPVO infolge zusätzlicher immunstimulatorischer Effekte
für die Heilung bestehender Infektionen sehr vielversprechend. Dies hätte große Vorteile
gegenüber bereits existierenden Immuntherapien mit vielen Nebenwirkungen (19).

AUSBLICK
64
6 Ausblick
Für alle in 5.3.4 beschriebenen Anwendungen ist die genaue Charakterisierung der
molekularen Aktivierungsmechanismen von iPPVO auf Zellen des angeborenen und
adaptiven Immunsystems notwendig, um mögliche Komplikationen und Nebenwirkungen zu
vermeiden. Die vorliegende Arbeit leistet zu dieser Charakterisierung einen entscheidenden
Beitrag. Sie befasst sich erstmals eingehend mit der Darstellung der immunstimulatorischen
Eigenschaften von iPPVO auf DCs in vitro. Die durchgeführten Untersuchungen mit iPPVO
an DCs ermöglichen neue Einblicke in die Wirkungsweise dieses Immunmodulators aus dem
Präparat Zylexis®. Die Analysen unterstreichen durch die beschriebenen DC-
Effektorantworten in vitro dessen Potential, zur Überwindung von viralen und stressbedingten
Erkrankungen bei domestisch gehaltenen Tieren wie Hunde, Katzen oder Pferde beizutragen.
Diese Charakterisierung muss noch fortgeführt und erweitert werden, um auch Aussagen über
die Wirkung von iPPVO auf Immunzellen in vivo treffen zu können. Dies beinhaltet weitere
Analysen des zellulären Interaktionspartners für die stimulatorische virale Komponente. Für
Untersuchungen zur Identifikation der jeweiligen Quelle (cDC, pDC oder beide) der
produzierten Zytokine sowie zu den durch diese möglicherweise induzierten autokrinen bzw.
parakrinen Rückkopplungsschleifen eignen sich Depletionsexperimente in Kombination mit
ELISA zur Detektion der produzierten Zytokine. Mit cryo-elektronenmikroskopischen
Aufnahmen, Konfokalmikroskopie oder Immunfluorenzenz-EM kann der
Aufnahmemechanismus von iPPVO in DCs charakterisiert und verglichen werden mit bisher
diskutierten Aufnahmewegen beim Modellvirus der Pockenviren VV. Analysen wie die in der
vorliegenden Arbeit dienen einem besseren Verständnis der Aktivierung des angeborenen
Immunsystems durch Viren und den damit assoziierten molekularen
Erkennungsmechanismen. Sie führen letztendlich zu neuen effizienteren Strategien für die
Immunisierung und zur Entwicklung antiviraler Medikamente.

ZUSAMMENFASSUNG
65
7 Zusammenfassung
Der bisherige Forschungsstand zur Charakterisierung der molekularen Mechanismen, die sich
hinter den immunstimulatorischen Eigenschaften von iPPVO verbergen, beruhte
ausschließlich auf der Analyse von Makrophagen, NK-Zellen und neutrophilen Granuloyzten
(18). Zur vollständigen Untersuchung der Wirkung des veterinärmedizinischen
Immunmodulators iPPVO auf das Immunsystem müssen jedoch DCs als wichtige
antigenpräsentierende Zellen und Produzenten einer Vielzahl immunmodulatorischer
Zytokine in diese Betrachtungen mit einbezogen werden. Die vorliegende Arbeit beschreibt
umfassend die In-vitro-Aktivierung von Flt3L-generierten DCs (murines Modell) aus
Knochenmark nach Stimulation mit iPPVO und deren vielfältige Effektorantworten. In Bezug
auf die weitere Aktivierung von Zellen des adaptiven Immunsystems scheinen cDCs die
potenteren DCs, was sich anhand verstärkt exprimierter Oberflächenmoleküle nach
Stimulation mit iPPVO im Vergleich zu pDCs zeigt. Des Weiteren konnte eine Freisetzung
von antiviralem IFN-α sowie der proinflammatorischen Mediatoren IL-12/23p40 und TNF-α
nachgewiesen werden. Erste elektronenmikroskopische Aufnahmen von durch iPPVO
aktivierten DCs dokumentieren die endozytotische Aufnahme von iPPVO.
Für die Charakterisierung der immunstimulatorischen Eigenschaften von iPPVO ist es
weiterhin unerlässlich, den/die Rezeptor(en) auf den DCs und die dazugehörige(n) virale(n)
Komponente(n) zu analysieren. Durch eine vorher bestimmte partielle MyD88-Abhängigkeit
der iPPVO-induzierten DC-Aktivierung in Bezug auf IL-12/23p40 und TNF-α rückten die
Toll-ähnlichen Rezeptoren als membranständige Klasse der PRRs in die nähere Auswahl
(Siegemund et al., Manuskript in Vorbereitung). Insbesondere die in Bezug auf virale
Infektionen schon mehrfach beschriebenen TLR9 und TLR2 kamen in Betracht (54, 56).
Jedoch erwies sich keiner der beiden als der MyD88-abhängige Rezeptor für die
immunstimulatorische Komponente von iPPVO, über den DCs aktiviert und zur gesteigerten
Expression von Oberflächenmolekülen wie MHC-I, MHC-II, CD86, CD80 und CD40 und
zur Freisetzung von IL-12/23p40 und TNF-α stimuliert werden. Diese Untersuchungen
zeigen, dass ein Pathogen mehrere verschiedene Rezeptoren auf und in Immunzellen
aktivieren kann und dass Pockenviren, trotz der Verwandtschaft innerhalb dieser
Virusfamilie, über viele verschiedene Erkennungsmechanismen Zellen des Immunsystems
aktivieren. Die vorliegende Arbeit leistet daher einen entscheidenden Beitrag zur Aufklärung
der immunstimulatorischen Wirkungsmechanismen von iPPVO auf dendritische Zellen.

LITERATURANGABE
66
8 Literaturangabe
(1) Haig, D.M., Mercer, A. A. 1998. Orf. Vet. Res. 29: 311-326. (2) Büttner, M., Rhiza, H.-J. 2002. Parapoxviruses: From Lesion to the Viral
Genome. J. Vet. Med. B 49:7-16.
(3) Mercer, A.A., Fraser, K., Barns, G., Robinson, A.J. 1987. The structure and cloning of orf virus DNA. Virology 157:1-12.
(4) Wittek, R., Kuenzle, C.C., Wyler, R.1979. High C+G content in parapoxvirus
DNA. J. Gen. Virol. 43:231-234.
(5) Haig, D.M., McInnes, C.J. 2002. Immunity and counter-immunity during infection with the parapoxvirus orf virus. Virus research 88:3-16
(6) Mayr, A., Herlyn, M., Mahnel, H.1981. Control of contagious ecthyma in
sheep by means of a new parenteral cell culture derived live vaccine. J. Med. Vet. B 28:535-549
(7) Popischil, A., Bachmann, P.A. 1980. Nuclear changes in cells infected with
parapoxviruses stomatitis papulosa and orf: an in vivo and in vitro ultrastructural study. J. Gen Virol 47(1): 113-121
(8) Moss, B. 2005. Poxvirus entry and membrane fusion. Virology 244:48-54.
(9) Vanderplasschen, A., Hollinshead, M., Smith, G.L. 1998. Intracellular and
extracellular vaccinia virions enter cells by different mechanisms. J. Gen. Virol. 79:877-887.
(10) Payne, L.G., Norrby, E. 1978. Adsorption and penetration of enveloped and
naked vaccinia virus particles. J. Virol. 27:19-27.
(11) Locker, J.K., Huehn, A., Schleich, S., Rutter, G., Hohenberg, H., Wepf, R., Griffiths, G. 2002. Entry of two infectious forms of vaccinia virus at the plasma membrane is signalling-dependent for the IMV but not the EEV. Mol. Biol. Cell 11:2497-2511.
(12) Law, M., Carter, G.C., Roberts, K.L., Hollinshead, M., Smith, G.L. 2006.
Ligand-induced and non-fusogenic dissolution of a viral membrane. Proc. Natl. Acad. Sci. USA 103:5989-5994.
(13) Armstrong, J.A., Metz, D.H., Young, M.R. 1973. The mode of entry of
vaccinia virus into L cells. J. Gen. Virol. 21(3):533-537.
(14) Beaud, G., Beaud, R. 1997. Preferential virosomal location of underphosphorylated H5R protein synthesized in vaccinia virus-infected cells. J. Gen. Virol. 78:3297-3302.

LITERATURANGABE
67
(15) Tolonen, N., Doglio, L., Schleich, S., Locker, J.K. 2001. Vaccinia virus DNA replication occurs in endoplasmatic reticulum-enclosed cytoplasmatic mini-nuclei. Mol. Biol. Cell 12:2031-2046.
(16) Smith, L.J., Vanderplasschen, A., Law, M. 2002. The formation and function
of extracellular enveloped vaccinia virus. J. Gen. Virol. 83:2915-2931.
(17) Modrow, S., Falke, D., Truyen, U. 2003. Molekulare Virologie. Heidelberg, Spektrum Akamdemischer Verlag, Berlin, Oxford.
(18) Cyrklaff, M., Linaroudis, A., Boicu, M., Chlanda, P., Baumeister, W.,
Griffiths, G., Locker, J.K. 2007. Whole cell cryo-electron tomography reveals distinct disassembly intermediates of vaccinia virus. PLoS ONE 2(5): e 420.
(19) Brown, F. 1995. Vaccination today and tomorrow. Biosci. Rep. 15(6): 493-
502.
(20) Bahnemann, H.G. 1990. Inactivation of viral antigens for vaccine preparation with particular reference to the application of binary ethylamine. Vaccine 8(4): 299-303.
(21) Troll, W., Berkowitz, E.M. 1969. Modification of dNA and poly rG by
carcinogenic agents assayed by physical and enzymatic methods. In: Bergman E.D., Pullmann, B. 1969. Physico-chemical mechanisms of carcinogenesis. Jerusalem Symp Quant Chem Biochem, Isreal Acad of Sci Human, Jerusalem I: 168-174
(22) Kaisermann, F., Wyss, K., Kempf, C. 2001. Virus inactivation and protein
modifications by ethyleneimines. Antiviral Research 52: 33-41
(23) Friebe, A., Siegling, A., Friederichs, S., Volk, H.D., Weber, o. 2004. Immunomodulatory effects of inactivated parapoxvirus ovis (Orf virus) on human peripheral immune cells: induction of cytokine secretion in monocytes and Th1-like cells. J. Virol. 78(17): 9400-9411.
(24) Weber, O., Siegling, A., Friebe, A., Limmer, A., Schlapp, T., Knolle, P.,
Mercer, A., Schaller, H., Volk, H.D. 2003. Inactivated parapoxvirus ovis (Orf virus) has antiviral activity against hepatitis B virus and herpes simplex virus. J. Gen. Virol. 84:1843-1852.
(25) Steinman, R.M., Inaba, K. 1999. Myeloid dendritic cells. J. Leukoc. Biol.
66:205-208.
(26) Shortman, K., Liu, Y.J. 2002. Mouse and human dendritic cell subypes. Nat. Rev. Immunol. 2:151-161.
(27) Shortman, K., Naik., S.H. 2007. Steady-state and inflammatory dendritic-cell
development. Nat. Rev. Immunol. 7:105-114.
(28) Wick, M.J. 2007. Monocyte and dendritic cell recruitment and activation during oral Salmonella infection. Immunol Lett. 112(2): 68-74

LITERATURANGABE
68
(29) Brawand, P.,Fitzpatrick, D.R., Greenfield, B.W., Brasel, K., Maliszewski, C.R.
und De Smedt, T. 2002. Murine Plasmacytoid Pre-Dendritic cells generated from Flt3 Ligand-Supplemented Bone Marrow Cultures Are Immature APCs. J. Immunol. 169:6711-6719.
(30) Verdijk, P., van Veelen, P.A., de Ru, A.H., Hensbergen, P.J., Mizuno, K.,
Koerten, H.K., Koning, F., Tensen, C.P., Mommaas, A.M. 2004. Morphological changes during dendritic cell maturation correlate with cofilin activation and translocation to the cell membrane. Eur. J. Immunol. 34:156-164.
(31) Soumelis, V., Liu, J-Y. 2006. From plasmacytoid to dendritic cell:
Morphological and functional switches during plasmacytoid pre-dendritic cell differentiation. Eur. J. Immunol. 36:2286-2292.
(32) Gilliet, M., Boosntra, A., Paturel, C. 2002. The development of murine
plasmacytoid dendritic cell precursors is differentially regulated by FLT3-ligand and granulocyte/macrophage colony-stimulating factor. J. Exp. Med. 195:953-958
(33) Brasel, K., DeSmedt, T., Smith, J.L., Maliszewski, C.R. 2000. Generation of
murine dendritic cells from flt3-ligand-supplemented bone marrow cultures. Blood 96:3029-3039.
(34) Cella, M., Jarrossay, D., Facchetti, F., Alebardi, O., Nakajima, H.,
Lanzavecchia, A., Colonna, M. 1999. Plasmacytoid monocytes migrate to inflamed lymph nodes and produce large amounds of type I interferon. Nat Med 5: 919-923.
(35) Asselin-Paturel, C., Boonstra, A., Dalod, M. 2001. Mouse type I IFN-
producing cells are immature APCs with plasmacytoid morphology. Nat Immunol 2: 1144-1150.
(36) Naik, S.H., Proietto, A.I., Wilson, N.S. 2005. Cutting edge: generation of
splenic CD8+ and CD8- dendritic cell equivilants in Fms-like tyrosine kinase 3 ligand bone marrow cultures. J Immunol 174: 6592-6597.
(37) Medzhitov, R. 2001. Toll-like receptors and innate immunity. Nat. Rev.
Immunol. 1:135-145
(38) Takeda, K., Kaisho, T., Akira, S. 2003. Toll-like receptors. Annu.Rev.Immunol. 21:335-376.
(39) Choe, J., Kelker, M.S., Wilson, I.A. 2005. Crystal structure of human Toll-like
Receptor 3 (TLR3) Ectodomain. Science 309: 581-585.
(40) Wickelgren, I. 2007. Targeting the Tolls. Science 312:184-186.
(41) Reis e Sousa, C. 2004. Toll-like receptors and dendritic cells: from whom the bug tolls. Seminars in Immunology 16: 27-34.

LITERATURANGABE
69
(42) Edwards, A.D., Diebold, S.S., Slack, E.M., Tomizawa, H., Hemmi H., Kaisho,
T. 2003. Toll-like receptor expression in murine DC subsets: lack of TLR7 expression by CD8α+ DC correlates with unresponsiveness to imidazoquines. Eur J Immunol 33: 827-833.
(43) Kawai, T., Akira, S. 2006. Innate immunity recognition of viral infection. Nat.
Immunol. 7:131-137.
(44) Honda, K., Yanai, H., Mizutani, T., Negishi, H., Shimada, N., Suzuki, N., Ohba, Y., Takaoka, A., Yeh, W., Taniguchi, T. 2001. Role of a transductional-transcriptional processor complex involving MyD88 and IRF-7 in Toll-like receptor signalling. Proc. Natl. Acad. Sci. USA 101:15416-15421.
(45) Hoshino, K., Sugiyama, T., Matsumoto, M., Tanaka, T., Saito, M., Hemmi, H.,
Ohara, O., Akira, S., Kaisho, T. 2006. IκB kinase α is critical for interferon-α production induced by Toll-like receptors 7 and 9. Nature 440:949-953.
(46) Oganesyan, G., Saha, S.K., Guo, B., He, J.Q., Shahangian, A., Zarnegar, B.,
Perry, A., Cheng, G. 2006. Critical role of TRAF3 in the Toll-like receptor-dependent and –independent antiviral response. Nature 439:208-211.
(47) Honda, K., Yanai, H., Negishi, H., Asagiri, M., Sato, M., Mizutani, T.,
Shimada, N., Ohba, V., Takaoka, A., Yoshida, N. 2005. IRF-7 is the master regulator of type-I-interferon-dependent immune responses. Nature 434:772-777.
(48) McGreal, E.P., Miller, J.L., Gordon, S. 2005. Ligand recognition by antigen-
presenting cell C-type lectin receptors. Curr. Opin. Immunol. 17: 18-24.
(49) Creagh, E.M., O’Neill, L.A. 2006. TLRs, NLRs and RLRs: a trinity of pathogen sensors that co-operate in inate immunity. Trends Immunol. 27: 352-357.
(50) Rothenfusser, S., Goutany, N., DiPerna, G., Gong, M., Monks, B.G.,
Schoenemeyer, A., Yamamoto, M., Akira, S, Fitzgerald, K.A. 2005. The RNA-helicase Lgp2 inhibits TLR-independent sensing of viral replication by retinoic acid-inducible gene-I. J. Immunol. 175:5260-5268.
(51) Yonyama, M., Kikuchi, M., Natsukawa, T., Shinobu, N., Imaizumi, T.,
Miyagishi, M., Taira, K., Akira, S., Fujita, T. 2004. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat. Immunol. 5:730-737
(52) Takaoka,A., Wang, Z.C., Choi, M.K., Yanai, H., Negishi, H., Ban, T., Lu, Y.,
Miyagishi, M., Tatsuhiko, K.,Honda, K., Ohba, Y., Taniguchi, T. 2007. DAI (DM-1/ZBP1) is a cytosolic DNA sensor an activator of innate immune responses. Nature 448:501-506.
(53) Pestka, S., Krause, C.D., Walter, M.R. 2004. Interferons, interferon-like
cytokines, and their receptors. Immunol. Rev. 202:8-32.

LITERATURANGABE
70
(54) Hochrein, H., Schlatter, B., O’Keeffe, M., Wagner, C., Schmitz, F.,
Schiemann, M., Bauer, S., Suter, M., Wahner, H. 2004. Herpes simplex virus type-1 induces IFN-α production via Toll-like receptor 9-dependent and -independent pathways. PNAS 101(31):11416-11421
(55) Guzylack-Piriou, L., Bergamin, F., Gerber, M., McCullough, K.C.,
Summerfield, A. 2006. Plasmacytoid dendritic cell activation by foot-and-mouth disease virus requires immune complexes. Eur. J. Immunol. 36:1674-1683.
(56) Zhu, J., Martinez, J., Huang, X., Yang Y. 2007. Innate immunity against
vaccinia virus is mediated by TLR2 and requires TLR-independent production of IFN-β. BLOOD 109(2):619-625
(57) Pfeffer, K., Matsuyama, T., Kündig, T.M., Wakeham, A., Kishihara, K.,
Shahinian, A., Wiegmann, K., Ohashi, P.S., Krönke, M., Mak, T.W. 1993. Mice deficient for the 55kd tumor necrosis factor receptor are resistant to endotoxic shock, yet succumb to L. monocytogenes infection. Cell 73:457-467.
(58) Magram, J. 1996. IL-12 deficient mice are defective in IFNγ production and
type 1 cytokine responses. Immunity 4:471-481.
(59) Mattner, F. 1996. Genetically resistant mice lacking interleukin-12 are susceptible to infection with Leishmania major and mount a polarized Th2-cell response. Eur. J. Immunol. 26:1553-1559.
(60) Brombacher, F., Kastelein, R.A., Alber, G. 2003. Novel IL-12 family members
shed light on the orchestration of Th1 responses. TRENDS in Immunology 24(4):207-212.
(61) Gowans, J.L. 1996. The Lymphocyte - a disgraceful gap in medical knowledge.
Immunol. Today 17:288-291.
(62) Gunzer, M., Weishaupt, C., Hillmer, A., Basolglu, Y., Friedl, P., Dittmar, K., Kolanus, W. 2004. A spectrum of biophysical interaction modes between T-cells and different antigen-presenting cells during priming in 3-D collagen and in vivo. BlOOD 104:2801-2809
(63) Rothoeft, T., Balkow, S., Krummen, M., Beissert, S., Varga, G., Loser, K.,
Oberanscheidt, P., van den Boom, F., Grabbe, S. 2006. Structure and duration of contact between dendritic cells and T cells are controlled by T cell activation state. Eur. J. Immunol. 36:3105-3117
(64) Whitmire, J.K., Flavell, R.A., Grewal, I.S., Larsen, C.P., Pearson, T.C.,
Ahmend, R. 1999. CD40-CD40 ligand costimulation is required for generating antiviral CD4 T cell responses but is dispensable for CD8 T cell responses. J. Immunol. 163:3194-3201.
(65) Brennan, G.L., Kronenberg, L.H. 1983. Automated bioassay of interferons in
micro-test plates. Biotechniques 1:78-83.

LITERATURANGABE
71
(66) Finter, N.B. 1969. Dye Uptake Methods for Assessing Viral Cytopathogenicity
and their Application to Interferon Assays. J. Gen. Virol. 5:419-427.
(67) Lund, J., Sato, A., Akira, S., Medzhitov, Iwasaki, A. 2003. Toll-like receptor 9 mediated recognition of herpes simplex virus-2 by plasmacytoid dendritic cells. J. Exp. Med. 198:513-520.
(68) Francke, B., Suter, M., Hochrein, H., O’Keeffe, M. 2008. M-CSF: a novel
plasmacytoid and conventional dendritic cell poietin. Blood 111:150-159.
(69) Lear, A., Hutchinson, G., Reid, H.W., Norval, M., Haig, D.M. 1996. Phenotypic characterisation of the dendritic cells accumulating in ovine dermis following primary and secondary Orf virus infection. Eur. J. Dermatol. 6:135-140.
(70) Weslow-Schmidt, J.L., Jewell, N.A., Mertz, S.E., Simas, J.P., Durbin, J.E.,
Flano, E. 2007. Type I interferon inhibition and dendritic cell activation during gammaherpesvirus respiratory infection. J. Virol. 81(18): 9778-9789.
(71) Waibler, Z., Anzaghe, M., Ludwig, H., Akira, S., Weiss, S., Sutter, G.,
Kalinke, U. 2007. Modified VacciniaVirus Ankara induces Toll-like receptor-independent Type I Interferon responses. J. Virol. 81(22):12102-12110.
(72) Honda, K., Sakaguchi, S., Nakajima, C. 2003. Selektive contribution of IFN-
alpha/beta signaling to the maturation of dendritic cells induced by double-stranded RNA or viral infection. Proc. Natl. Acad. Sci. USA 100: 10872-10877
(73) Hoebe, K., Janssen, E.M., Kim, S.O. 2003. Upregulation of costimulatory
molecules induced by lipopolysaccharide and double-stranded RNA occurs by Trif-dependent and –independent pathways. Nat. Immunol. 4:1223-1229
(74) Deng, L., Dai, P., Ding, W., Granstein, R., Shumann, S. 2006. Vaccinia virus
infection attenuates innate immune responses and antigen presentation by epidermal dendritic cells. J. Virol. 80 (20): 9977-9987.
(75) Jarvis, M.A., Borton, J.A., Keech, A.M., Wong, J., Britt, W.J., Magun, B.E.,
Nelson, J.A. 2006. Human cytomegalovirus attenuates interleukin-1β and tumor necrosis factor alpha proinflammatory signalling by inhibition of NF-κB activation. J. Virol. 80(11): 5588-5598.
(76) Kowalik, T.F., Wing, B., Haskill, J.S., Azizkhan, J.C., Baldwin, A.S., Huang,
E.S. 1993. Multiple mechanisms are implicated in the regulation of NF-kappa B activity during human cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 90: 1107-111.
(77) Matikainen, S., Siren, J., Tissari, J., Veckman, V., Pirhonen, J., Severa, M.,
Sun, Q., Lin, R., Meri, S., Uze, G., Hiscott, J., Julkunen, I. 2006. Tumor necrosis factor alpha enhances Influenza A virus-induced expression of antiviral cytokines by activating RIG-I Gene expression. J. Virol. 80 (7): 3515-3522.

LITERATURANGABE
72
(78) Nagington, J., Horne, R.W. 1962. Morphological studies of Orf and Vaccinia
viruses. Virology 16:248-260.
(79) Sieczkarski, S.B., Whittaker, G.R. 2005. Viral entry. Curr. Top. Microbiol. Immunol. 285:1-23.
(80) Hsiao, J.C., Chung, C.S., Chang, W. 1999. Vaccinia Virus envelope D8L
protein binds to cell surface chondroitin sulphate and mediates the adsorption on intracellular mature virions to cells. J. Virol. 73:8750-8761.
(81) Chung, C.S., Huang, C.Y., Chang, W. 2005. Vaccinia virus penetration
requires cholesterol and results in specific viral envelope proteins associated with lipid rafts. J. Virol. 79:1623-1634.
(82) Townsley, A.C., Weisberg, A.S., Wagenaar, T.R., Moss, B. 2006. Vaccinia
virus entry into cells via a low-pH-dependent endosomal pathway. J. Virol. 80:8899-8908
(83) Kwong, P.D., Wyatt, R., Robinson, J., Sweet, R.W., Sodrowski, J. 1998.
Strucutre of an HIV gp120 envelope glycoprotein in complex with CD4 receptor and a neutralizing human antibody. Nature 393:648-659.
(84) Steward, P.L., Dermody, T.S., Nemerow, G.R. 2003. Structural basis of
nonenveloped virus cell entry. Adv. Protein Chem. 64:455-491.
(85) Hemmi, H. 2002. A Toll-like receptor recognizes bacterial DNA. Nature 408:740-745
(86) Hemmi, H., Kaisho, T., Takeda, K., Akira, S. 2003. The roles of Toll-like
receptor 9, MyD88, and DNA-dependent protein kinases catalytic subunit in the effect of two distinct CpG-DNAs on dendritic cell subsets. J. Immunol. 170:3059-3064.
(87) Krug, A. 2004. TLR9-dependent recognition of MCMV by IPC and DC
generates coordinated cytokine responses that activate antiviral NK cell function. Immunity 21:107-119.
(88) Sato, A., Lineham, M. M., Iawsaki, A. 2006. Dual recognition of herpes
simplex viruses by TLR2 and TLR9 in dendritic cells. PNAS 103(46): 17343-17348.

Danksagung
Ich möchte mich bei Prof. Gottfried Alber für die Bereitstellung des spannenden Themas und
aller zu den Experimenten benötigten Arbeitsmaterialien bedanken. Es war sehr schön, einen
Betreuer zu haben, der sich so viel Zeit für konstruktive Gespräche und Hilfestellung bei der
Bearbeitung wissenschaftlicher Fragen für einen Studenten nimmt.
Frau Dr. Sabine Siegemund danke ich für die viele Zeit und Mühe, die sie in meine
Einarbeitung gesteckt und für die Beantwortung meiner vielen Fragen investiert hat. Vor ihrer
Geduld und stetigen Hilfsbereitschaft habe ich den größten Respekt.
Ich danke weiterhin den gesamten Arbeitsgruppen von Prof. Gottfried Alber, Prof. Manfred
Blessing und PD Dr. Reinhard Straubinger für die freundliche Integration in das Team und die
stetige Unterstützung bei allen Fragen und Nöten. So macht wissenschaftliches Arbeiten
einfach viel Freude.
Ich danke Dr. Rüdiger Raue und Pfizer Ltd. und Prof. Mathias Büttner für die Bereitstellung
von iPPVO und die vielen hilfreichen Tipps sowie Dipl.-Biol. Andrea Hartl und Dr. Tim
Sparwasser für die Zulieferung von Knochenmark aus TLR9def- und TLR2def-Mäusen.
Weiterhin gilt mein Dank Frau Prof. Gabriele Köhler für die elektronenmikroskopischen
Aufnahmen.
Frau Prof. Annette Beck-Sickinger danke ich für die Betreuung dieser Arbeit als
Zweitgutachterin und für ihr stetiges Engagement für die Studenten während des gesamten
Biochemiestudiums.
Mein besonderer Dank gilt Philipp, für die unendliche Geduld und stetige Unterstützung im
gesamten Zeitraum der Diplomarbeit. Jeden Tag ist es mein ganzes Glück, mit ihm das Leben
zu teilen. Weiterhin danke ich meinen Eltern, die ich wie Felsen in der Brandung stets hinter
mir weiß, die mir mit Rat und Tat zur Seite stehen und mir mein Studium überhaupt erst
ermöglichten.