dickersin 1 2
TRANSCRIPT

2Selective
Embryology
A knowledge of the early stages of human embryogen-esis is helpful in classifying neoplasms according tocells of origin. The least differentiated tumors obviouslyare the most difficult to categorize, and the key to di-agnosis often lies in finding cells that have sufficient cy-toplasmic or surface differentiation to allow a compar-ison with known cytologic structures in the normalhuman embryo. The brief text, diagrams, and electronmicrographs that follow serve to illustrate some of thebasic embryonic features that are applicable in estab-lishing the histogenesis of various neoplasms.
Embryogenesis from Fertilization Through Three Weeks
Figures 2.1A and B depict embryogenesis from fertil-ization to implantation. The fertilized ovum (zygote)undergoes sequential cleavages, forming a ball of 12–16cells (morula) by the third day. Further cellular divisionand accumulation of extracellular fluid during thefourth through seventh days result in a blastula (blas-tocyst), which consists of an eccentric cavity, an outercell mass (trophoblast), and an inner cell mass (em-bryoblast, Figure 2.1C). Implantation of the blastocystinto the endometrium occurs between the fifth and sev-enth days; by the eighth day, a bilaminar germ disc, con-sisting of epiblast (pre-ectoderm) and hypoblast (pre-endoderm), has formed (Figure 2.1D). During the sec-ond week of development, the amniotic cavity and yolksac are created, and the trophoblast differentiates intotwo layers: cytotrophoblast and syncytiotrophoblast. Bythe 16th day, the primitive streak has formed in the cau-dal end of the embryonic disc, and cells of the epiblastin this region migrate ventrally, laterally, and cephaladto form first a loose primary mesenchyme and then amore dense third germ layer, the mesoderm (Figure2.2A). The notochord (axial mesoderm) is formed fromcephalic migration of some of the cells of the primarymesenchyme, cephalad to the primitive streak, at ap-proximately the 18th day. By the 20th day, the meso-dermal cells have aggregated into three discrete masses:paraxial, intermediate, and lateral (Figure 2.2B). Theparaxial mesoderm becomes more distinct somiteswithin a day, starting at the cranial end and progress-ing caudally; the intermediate mesoderm develops intonephrotomes cranially and the nephrogenic cord cau-dally; the lateral mesoderm divides into somatic andsplanchnic layers, which become the mesothelial liningsof the coelomic cavities.
7

8 DIAGNOSTIC ELECTRON MICROSCOPY: A TEXT/ATLAS
Embryogenesis
Sperm
Fertilization
Oocyte
Fertilized Oocyte (Zygote)30–40 hours
Morula3–4 days
Blastula (Blastocyst)4–7 days
inner cell mass(embryoblast)
outer cell mass(trophoblast)
hypoblast
yolk sac
epiblastamniotic sac
Bilaminar Germ Disc8–14 days
A
B
C
D
Figure 2.1. Diagram of embryogenesis, from fertilization through the 14th day.

Differentiation of the Paraxial Mesoderm
The cells of the somites have epithelial features, andin the fourth week, each somite develops a central cav-ity. The somatic cells continue to proliferate, becomemore loosely arranged, and take on a more irregularshape (secondary mesenchyme). Cells from the medialand ventral aspects of the somites migrate toward thenotochord, resulting in the formation of the sclero-
tomes and ultimate vertebral column (Figure 2.2C).The mesenchymal cells comprising the sclerotomeshave the potential to differentiate into osteoblasts,chondroblasts, and fibroblasts. The dorsal aspects ofthe somites become the dermatomes (the future con-nective tissue of the back and some of the muscles ofthe limbs). The remaining internal regions of thesomites form the myotomes (the anlage of the musclesof the back).
SELECTIVE EMBRYOLOGY 9
Dorsal ViewTransverse View
(caudally) Sagittal View
primitive streak
primitive streak
primitive streak
somites
somites
neural tube
dermatome
myotome
sclerotome
nephrotome
yolksac
coelom
cranio-caudalflexion cranio-caudal
flexion
foreguthindgut
(B) 20th day
(C) 22nd day
neuralgroove
(A) 16th day
neural groove intermediate mesoderm
lateral mesoderm;somatic andsplanchnic
notochord paraxial mesoderm
(pre-somite)
ectoderm
mesoderm
endoderm
endoderm
mesodermectoderm
amniotic sac
yolksac
coelom
Trilaminar Germ Disc
Figure 2.2. Embryogenesis during the third week of ges-tation. A, Appearance of the third germ layer, the meso-derm. B, Formation of discrete mesodermal masses.
C, Migration and differentiation of cells of the mesoder-mal masses.
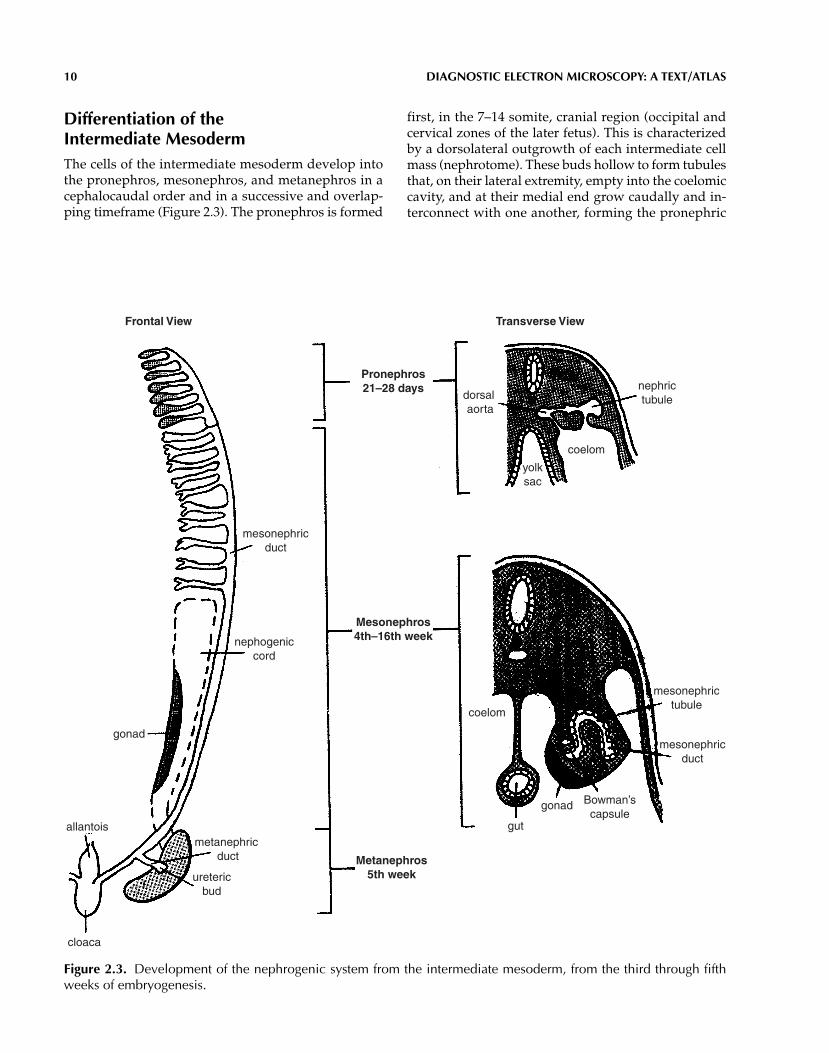
Differentiation of the Intermediate MesodermThe cells of the intermediate mesoderm develop intothe pronephros, mesonephros, and metanephros in acephalocaudal order and in a successive and overlap-ping timeframe (Figure 2.3). The pronephros is formed
first, in the 7–14 somite, cranial region (occipital andcervical zones of the later fetus). This is characterizedby a dorsolateral outgrowth of each intermediate cellmass (nephrotome). These buds hollow to form tubulesthat, on their lateral extremity, empty into the coelomiccavity, and at their medial end grow caudally and in-terconnect with one another, forming the pronephric
10 DIAGNOSTIC ELECTRON MICROSCOPY: A TEXT/ATLAS
Metanephros5th week
Mesonephros4th–16th week
Pronephros21–28 days nephric
tubuledorsal aorta
coelom
coelom
yolksac
mesonephrictubule
mesonephricduct
gonad
gut
Bowman’scapsule
mesonephricduct
gonad
nephogeniccord
metanephric duct
uretericbud
cloaca
allantois
Frontal View Transverse View
Figure 2.3. Development of the nephrogenic system from the intermediate mesoderm, from the third through fifthweeks of embryogenesis.

duct. Sprouts of the aorta grow simultaneously into themedial end of the pronephric buds to form primitiveglomeruli. The pronephros reaches its peak of devel-opment in the fourth week of embryonic life and then,except for its duct, involutes. The pronephric duct re-mains and serves as the duct of the mesonephros, emp-tying into the cloaca.
The mesonephros begins to develop in the fourthand fifth weeks from the nephrogenic cord at the levelsof 14–26 somites (lower cervical, thoracic, and upperlumbar regions).
Vesicles and then tubules develop in the nephrogeniccord. The dorsolateral end of each tubule joins thepronephric duct (now the mesonephric duct), and theopposite end forms a glomerulus with a branch of the aorta. Progressive cephalocaudal degeneration ofthe mesonephros occurs until the end of the 16th weekof life, when only the mesonephric (Wolffian) duct per-sists in the male (vas deferens).
The metanephros begins to develop in the fifth week,in the lower lumbar region, from two primordia: an out-growth (metanephric diverticulum, or ureteric bud) ofthe mesonephric duct, and the nephrogenic cord (meta-nephric blastema) (Figure 2.3). The nephrogenic tissueaggregates into small nodules at the tips of ingrowingcollecting tubules, which are the terminal extensions ofthe outgrowing and dividing urogenital sinus (from theanterior part of the cloaca). The nephrogenic nodules be-come vesicles and then elongate into tubules, connectingwith the collecting tubules on one end, and forming Bow-man’s capsules on the opposite end.
Differentiation of the Lateral Mesoderm
The cells of the lateral (coelomic) mesoderm separateinto two layers around a central, intraembryoniccoelom by the 19th day (Figures 2.2B and C). The outersomatic layer differentiates into the parietal mesothe-lium of the coelomic cavities and the connective tissueand skeletal muscle of the ventral body wall and por-tions of the limbs. The inner splanchnic layer of lateralmesoderm develops into the visceral mesothelium ofthe coelomic cavities, connective tissue and smoothmuscle of the gastrointestinal tract, paramesonephric(Müllerian) ducts, genital ridges, adrenal cortex, andmyocardium.
Comparison of Embryonic MesodermalDifferentiation with EmbryonalRhabdomyosarcoma, Wilms’ Tumor, and Mesothelioma
It is during the third to eighth weeks of embryogenesisthat differentiation is especially interesting to study,particularly in respect to correlating the morphology ofearly derivatives (Figure 2.4) of the third germ layer, themesoderm (Figures 2.5 and 2.6), with the structure ofcertain mesodermally derived neoplasms. For example,myotomes (Figures 2.7 through 2.12) are morphologi-cally recapitulated in embryonal rhabdomyosarcomas,both being composed of small round cells and early
SELECTIVE EMBRYOLOGY 11
Primary mesenchyme
Lateral mesoderm(coelomic)
Intermediate mesoderm(nephrogenic)
Paraxial mesoderm
Somite
Secondary mesenchyme
(sclerotome) (dermatome) (myotome)
Urogenital system
(nephrotome andnephrogenic cord)
(gonad)
osteoblastchondroblast
lipoblastfibroblast
rhabdomyoblast pronephrosmesonephrosmetanephros
(blastema)
mesotheliumgut leiomyoblastcardiac myoblast
adrenal cortical cell
endotheliumhematopoietic cells
lymphocytes
Figure 2.4. Diagram of the differentiation of primary mesenchyme.

strap like cells. Likewise, a well-recognized similarityexists between metanephric blastema (Figures 2.13through 2.16) and the undifferentiated component ofWilms’ tumors. In a similar vein, intraembryonic coe-limic lining cells correlate with epithelioid mesothe-liomas, and, more speculatively, the subsurface cells
(Figures 2.17 and 2.18) may be represented in the cellsthat comprise so-called fibrous mesotheliomas. (Illus-trations of these and related primitive neoplasms arepresented in the chapters on neoplasms).
12 DIAGNOSTIC ELECTRON MICROSCOPY: A TEXT/ATLAS
Figure 2.5. Primary mesenchyme from a 39-day-old human embryo. The cells vary in shape, are loosely arranged inan electron-lucent matrix, and have only focal contact with one another. (� 2100)
(Text continues on page 26)

SELECTIVE EMBRYOLOGY 13
Figure 2.6. Higher magnification of primary mesenchymal cells reveals a high nuclear-cytoplasmic ratio, euchro-matic nuclei, prominent nucleoli, and few cytoplasmic organelles. (� 6480)

14 DIAGNOSTIC ELECTRON MICROSCOPY: A TEXT/ATLAS
Figure 2.7. Human myotome, central dense area. Thecells are polygonal and densely arranged. Their nucleiand nucleoli are similar to those of primary mesenchyme,but their cytoplasm and organelles are somewhat more
abundant. (� 5510) (Permission for reprinting granted byHemisphere Publishing Co., Dickersin GR: Embryonic ul-trastructure as a guide in the diagnosis of tumors. Ultra-struct Pathol 11:609–652, 1987.)

SELECTIVE EMBRYOLOGY 15
Figure 2.8. Higher power of cells of a myotome. A mod-erate number of organelles is visible, including Golgi ap-paratuses (G), mitochondria (M), and rough endoplasmicreticulum (RER). Small junctions (J) are present between
the cells. (� 21,870) (Permission for reprinting grantedby Hemisphere Publishing Co., Dickersin GR: Embryonicultrastructure as a guide in the diagnosis of tumors. Ul-trastruct Pathol 11:609–652, 1987.)

16 DIAGNOSTIC ELECTRON MICROSCOPY: A TEXT/ATLAS
Figure 2.9. Myotome. Among the primitive-appearingcells are two cells (*) showing early myoblastic differen-tiation. This is characterized by a pale cytoplasm and sev-eral electron-dense areas (see Figure 2.10 for higher
power). (� 7020) (Permission for reprinting granted byHemisphere Publishing Co., Dickersin GR: Embryonic ul-trastructure as a guide in the diagnosis of tumors. Ultra-struct Pathol 11:609–652, 1987.)

SELECTIVE EMBRYOLOGY 17
Figure 2.10. Myotome. Higher magnification of two ofthe pale cells in Figure 2.9 reveals most of the cytoplasmto have an open or clear background, consistent with
glycogen, plus many irregularly arranged thin (actin) fil-aments. The dense areas (arrows) consist of thick (myosin)and thin filaments. (� 14,850)

18 DIAGNOSTIC ELECTRON MICROSCOPY: A TEXT/ATLAS
Figure 2.11. Myotome. This elongated cell is foundamong many primitive polygonal cells. It shows the ear-liest sign of skeletal muscle differentiation; that is, paral-lel thick filaments and closely associated rows and clusters of ribosomes (bracketed areas). (� 95,000) (Per-
mission for reprinting granted by Hemisphere PublishingCo., Dickersin GR: Embryonic ultrastructure as a guidein the diagnosis of tumors. Ultrastruct Pathol 11:609–652,1987.)

SELECTIVE EMBRYOLOGY 19
Figure 2.12. Myotome. Another recognizable myogeniccell is found among many undifferentiated ones makingup the myotome. Parallel thick filaments and early Z-bands (Z) are forming sarcomeres. The electron-denseregion at the left side of the field represents early sar-
comeres cut transversely. (� 27,800) (Permission forreprinting granted by Hemisphere Publishing Co., Dick-ersin GR: Embryonic ultrastructure as a guide in the di-agnosis of tumors. Ultrastruct Pathol 11:609–652, 1987.)

20 DIAGNOSTIC ELECTRON MICROSCOPY: A TEXT/ATLAS
Figure 2.13. Metanephric blastema. These cells, whichare derived from the intermediate mesodermal mass, gen-erally are similar to those of the myotome (see Figures 2.7and 2.8). They have a high nuclear-cytoplasmic ratio, eu-chromatic nuclei, prominent nucleoli, and scant cyto-
plasm. (� 3600) (Permission for reprinting granted byHemisphere Publishing Co., Dickersin GR: Embryonic ul-trastructure as a guide in the diagnosis of tumors. Ultra-struct Pathol 11:609–652, 1987.)

SELECTIVE EMBRYOLOGY 21
Figure 2.14. Metanephric blastema. Some of the cells inthis field (center) have grouped together in what is con-sistent with a pretubule. (� 3740) (Permission for reprint-
ing granted by Hemisphere Publishing Co., Dickersin GR:Embryonic ultrastructure as a guide in the diagnosis of tu-mors. Ultrastruct Pathol 11:609–652, 1987.)

22 DIAGNOSTIC ELECTRON MICROSCOPY: A TEXT/ATLAS
Figure 2.15. Metanephric blastema. A more discrete tubule has formed in this region, but only junctional complexesand no open lumen are demonstrable at higher power. (� 3270)

SELECTIVE EMBRYOLOGY 23
Figure 2.16. Metanephric blastema and early tubule. A definite lumen is present in this tubular form, and focal basallamina separating the tubule from the undifferentiated blastema is visible at higher magnification. (� 4750)
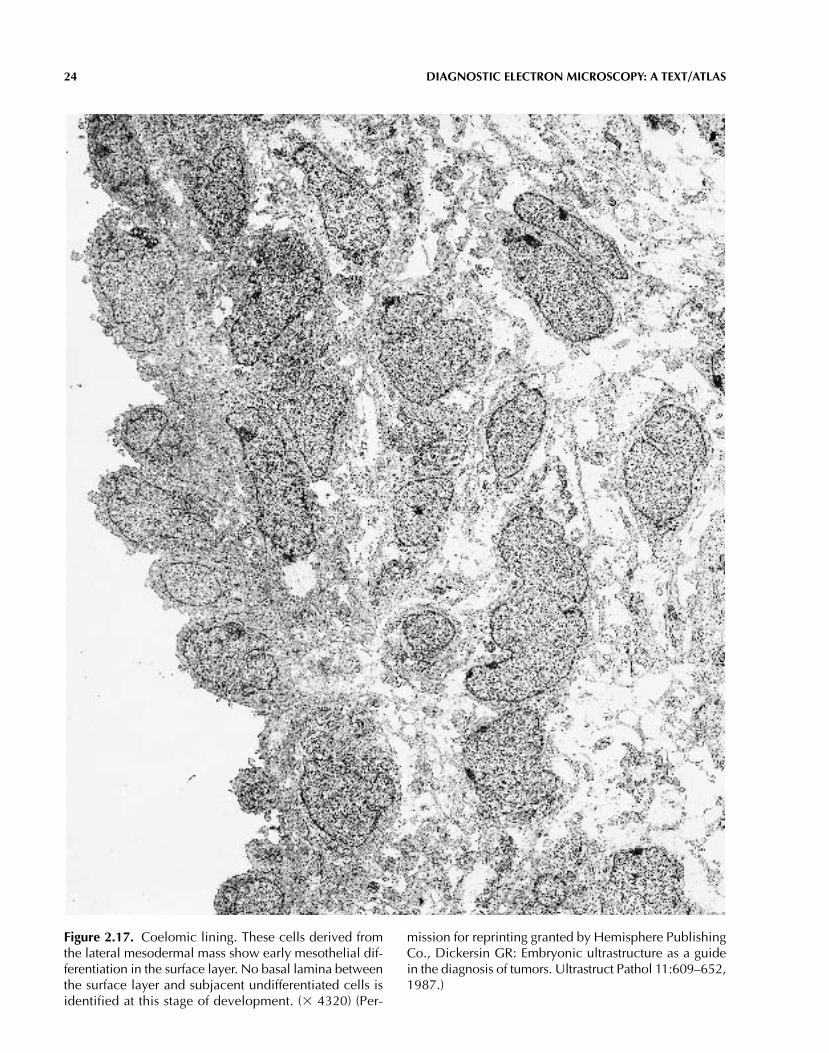
24 DIAGNOSTIC ELECTRON MICROSCOPY: A TEXT/ATLAS
Figure 2.17. Coelomic lining. These cells derived fromthe lateral mesodermal mass show early mesothelial dif-ferentiation in the surface layer. No basal lamina betweenthe surface layer and subjacent undifferentiated cells isidentified at this stage of development. (� 4320) (Per-
mission for reprinting granted by Hemisphere PublishingCo., Dickersin GR: Embryonic ultrastructure as a guidein the diagnosis of tumors. Ultrastruct Pathol 11:609–652,1987.)
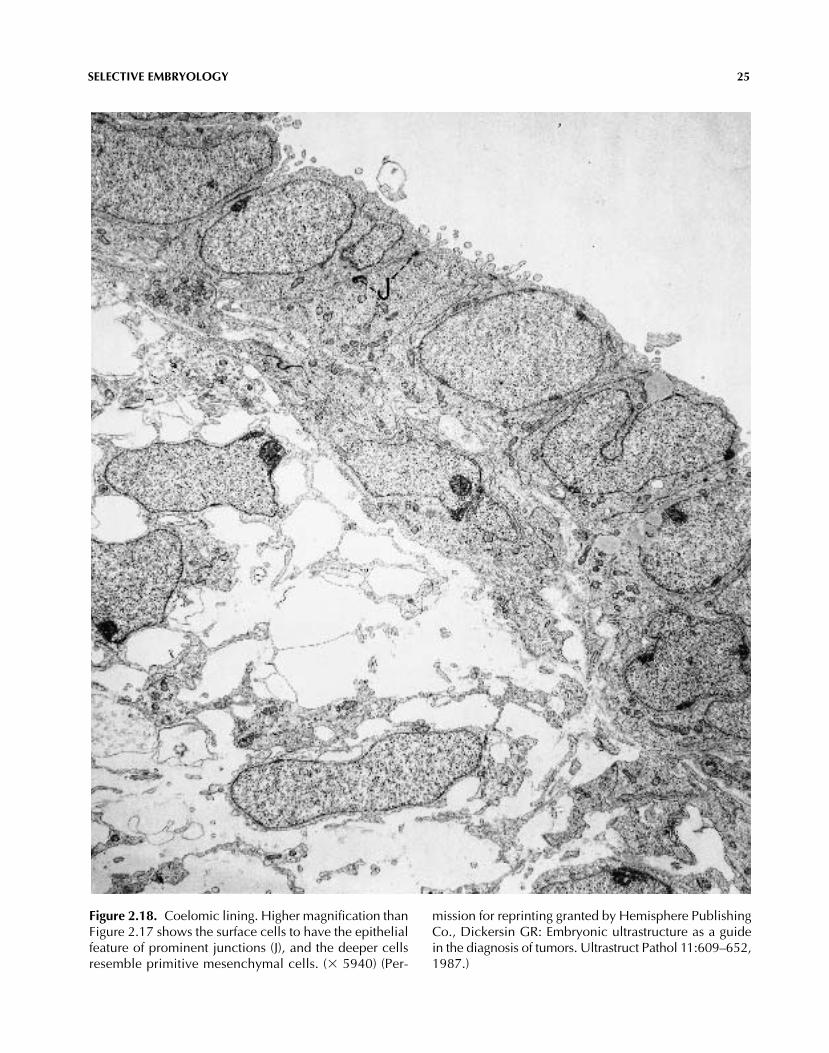
SELECTIVE EMBRYOLOGY 25
Figure 2.18. Coelomic lining. Higher magnification thanFigure 2.17 shows the surface cells to have the epithelialfeature of prominent junctions (J), and the deeper cellsresemble primitive mesenchymal cells. (� 5940) (Per-
mission for reprinting granted by Hemisphere PublishingCo., Dickersin GR: Embryonic ultrastructure as a guidein the diagnosis of tumors. Ultrastruct Pathol 11:609–652,1987.)

(Text continued from page 12)
REFERENCESBriselli M, Mark EJ, Dickersin GR: Solitary fibrous tu-mors of the pleura: Eight new cases and review of 360cases in the literature. Cancer 47:2678–2689, 1981.
Corliss CE: Patten’s Human Embryology. Elements of Clin-ical Development. McGraw-Hill, New York, 1976.
Dickersin GR: The contributions of electron microscopyin the diagnosis and histogenesis of controversial neo-plasms. Clin Lab Med 4:123–164, 1984.
Dickersin GR: Embryonic ultrastructure as a guide inthe diagnosis of tumors. Ultrastruct Pathol 11:609–652,1987.
Fischman DA: An electron microscopic study of myo-fibril formation in embryonic chick skeletal muscle. JCell Biol 32:557–575, 1967.
Friedman J, Bird ES: Electron-microscope investigationof experimental rhabdomyosarcoma. J Pathol 97:375–382,1969.
Hay ED: The fine structure of differentiating muscle inthe salamander tail. Zellforsch Mikrosk Anat 59:6–34,1963.
Jacob F: Expression of embryonic characters by malig-nant cells. Ciba Found Symp 96:4–27, 1983.
Lawrence WD, Whitaker D, Sugimura H, et al: An ul-trastructural study of the developing urogenital tract inearly human fetuses. Am J Obstet Gynecol 167:185–193,1992.
Nishimura H, ed: Atlas of Human Prenatal Histology.Igaku-Shoin, Tokyo, 1983.
Pierce GB: Neoplasms, differentiation and mutations.Am J Pathol 77:103–113, 1974.
Pierce GB: Teratocarcinoma: Model for a developmen-tal concept of cancer. Curr Top Dev Biol 2:223–246, 1967.
Sadler TW: Langman’s Medical Embryology, 7th ed.Williams and Wilkins, Baltimore, 1995.
Schmidt D, Dickersin GR, Vawter GF, et al: Wilms’ tu-mor: Review of ultrastructure and histogenesis. Patho-biol Annu 12:281–300, 1982.
26 DIAGNOSTIC ELECTRON MICROSCOPY: A TEXT/ATLAS