developmental regulation of the glyoxylate cycle in the human pathogen penicillium marneffei
TRANSCRIPT

Developmental regulation of the glyoxylate cycle in thehuman pathogen Penicillium marneffei
David Cánovas and Alex Andrianopoulos*Department of Genetics, University of Melbourne, Vic.3010, Australia.
Summary
Penicillium marneffei is a thermally dimorphic oppor-tunistic human pathogen with a saprophytic filamen-tous hyphal form at 25°C and a pathogenic unicellularyeast form at 37°C. During infection. P. marneffeiyeast cells exist intracellularly in macrophages. Tocope with nutrient deprivation during the infectionprocess, a number of pathogens employ the glyoxy-late cycle to utilize fatty acids as carbon sources. Thegenes which constitute this pathway have been impli-cated in pathogenesis. To investigate acetate and fattyacid utilization, the acuD gene encoding a key glyoxy-late cycle enzyme (isocitrate lyase) was cloned. TheacuD gene is regulated by both carbon source andtemperature in P. marneffei, being strongly induced at37°C even in the presence of a repressing carbonsource such as glucose. When introduced into thenon-pathogenic monomorphic fungus Aspergillusnidulans, the P. marneffei acuD promoter onlyresponds to carbon source. Similarly, when theA. nidulans acuD promoter is introduced into P. marn-effei it only responds to carbon source suggestingthat P. marneffei possesses both cis elements andtrans-acting factors to control acuD by temperature.The Zn(II)2Cys6 DNA binding motif transcriptionalactivator FacB was cloned and is responsible forcarbon source-, but not temperature-, dependentinduction of acuD. The expression of acuD at 37°C isinduced by AbaA, a key regulator of morphogenesis inP. marneffei, but deletion of abaA does not completelyeliminate temperature-dependent induction, suggest-ing that acuD and the glyoxylate cycle are regulatedby a complex network of factors in P. marneffei whichmay contribute to its pathogenicity.
Introduction
Genes encoding enzymes of the glyoxylate cycle havebeen implicated in both fungal and bacterial pathogenesis
(McKinney et al., 2000; Lorenz and Fink, 2001; Munoz-Elias and McKinney, 2005). The glyoxylate cycle involvestwo critical steps catalysed by the enzymes isocitratelyase and malate synthase, which bypass the two decar-boxylation steps of the TCA cycle. Isocitrate lyase hydroly-ses isocitrate (C6) to succinate (C4) and glyoxylate (C2)and subsequent condensation of glyoxylate and acetyl-CoA (C2) by malate synthase produces malate (C4), aTCA cycle intermediate. Malate is further oxidized tooxaloacetate (C4) and then converted into citrate (C6) bythe condensation of another molecule of acetyl-CoA. C4intermediates are required for anabolic processes. Forexample, oxaloacetate is employed during gluconeogen-esis to generate glucose, which is necessary for the incor-poration of carbon into the cellular macromolecules.Malate, oxaloacetate, citrate and isocitrate are intermedi-ates in the glyoxylate and TCA cycles, so the glyoxylatecycle allows two carbon compounds to replenish the TCAcycle. Therefore, the glyoxylate cycle is required forgrowth on gluconeogenic carbon sources, such asacetate, and it is usually activated under conditions ofnutrient deprivation. Such conditions are believed to occurinside macrophages and pose particular challenges tointracellular pathogens (Lorenz and Fink, 2002).
In Candida albicans, the isocitrate lyase gene icl1 isupregulated during growth inside macrophages. Strainsdeficient in this gene are less virulent than wild-typestrains in a mouse pathogenicity model (Lorenz and Fink,2001). Recently it has been reported that the glyoxylatecycle genes in C. albicans are repressed by the physi-ological concentrations of glucose found in thebloodstream. However, these genes are induced uponphagocytosis by macrophages or neutrophils, emphasiz-ing the importance of carbon metabolism during patho-genesis (Barelle et al., 2006). Plant pathogens have alsobeen shown to require the glyoxylate cycle for pathoge-nicity (Idnurm and Howlett, 2002; Wang et al., 2003;Solomon et al., 2004). Despite the importance of the gly-oxylate cycle in pathogenesis, the molecular mechanismswhich regulate the isocitrate lyase encoding genes inpathogens are unknown.
In the non-pathogenic fungus Aspergillus nidulans, theisocitrate lyase encoding acuD gene is expressed duringgrowth on gluconeogenic compounds such as acetate orfatty acids. Acetate induction of acuD is FacB-dependent(Todd et al., 1997). FacB is a transcriptional activator with
Accepted 12 October, 2006. *For correspondence. E-mail [email protected]; Tel. (+61) 383 445 164; Fax (+61) 383 445 139.
Molecular Microbiology (2006) 62(6), 1725–1738 doi:10.1111/j.1365-2958.2006.05477.xFirst published online 13 November 2006
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd

a zinc binuclear cluster (Zn(II)2Cys6) DNA bindingdomain. FacB binds two dissimilar DNA sequences foundin the 5′ regions of acetate-regulated genes such as acuD(Todd et al., 1998). Besides acetate regulation, acuD isalso regulated by fatty acids via FarA (long-chain fattyacids) and ScfA and FarB (short-chain fatty acids). Allthese fatty acid transcriptional regulators also contain aZn(II)2Cys6 DNA binding domain and work independentlyof FacB (Hynes et al., 2006). In Saccharomyces cerevi-siae the glyoxylate cycle genes are activated by the tran-scriptional activators Cat8 and Sip4, which also contain aZn(II)2Cys6 DNA binding domain, in response to glucoselimitation. Cat8 is the main activator of the expression ofthe glyoxylate genes and is also required for the expres-sion of SIP4, which plays a minor role in the upregulationof the glyoxylate genes (Schuller, 2003). In addition tothe acetate and fatty acid induction of the glyoxylatecycle genes, expression is also subject to glucose-mediated repression. In A. nidulans, this is achieved inpart by the action of the carbon catabolite transcriptionalrepressor CreA (Bowyer et al., 1994; De Lucas et al.,1994a).
Penicillium marneffei is an opportunistic fungal patho-gen of humans. It is the only dimorphic species within thePenicillium and related genera. At 25°C, P. marneffeigrows as multicellular hyphal cells, which resemble otherPenicillium species while at 37°C it grows as a unicellularyeast, oval in shape, and divides by fission. Yeast cellsare the pathogenic form. In addition, at 25°C in responseto specific environmental cues it can undergo asexualdevelopment (conidiation) to produce conidia (spores),which are presumed to be the infective form (Andrianopo-ulos, 2002). In the monomorphic fungus A. nidulans,asexual development is controlled by a cascade of tran-scriptional regulators, encompassing brlA, which encodesa C2H2 zinc finger protein (Adams et al., 1988), abaA,which encodes an ATTS/TEA protein (Andrianopoulosand Timberlake, 1991; Burglin, 1991) and wetA, whichencodes a spore-specific protein (Marshall and Timber-lake, 1991). BrlA is activated by conidiation inducing con-ditions, and then in turn activates abaA expression. brlAmutants strains fail to produce most of the conidiophorecell types and are only able to differentiate conidiophorestalks (Clutterbuck, 1969). AbaA activates the expressionof wetA and also feedback regulates the expression ofitself and brlA (for review see Adams et al., 1998). abaAmutant strains produce aberrant conidiophores lackingconidia, instead producing reiterated phialide-like cellswhich bud by acropetal division (Sewall et al., 1990). InP. marneffei, asexual development is similarly regulatedbut AbaA is also required to couple nuclear and cellulardivision during dimorphic switching (Borneman et al.,2000; A.R. Borneman, M.J. Hynes and A. Andrianopoulos,unpublished results). The expression of abaA is upregu-
lated 10-fold during dimorphic switching and 30-foldduring conidiation (Borneman et al., 2000).
In the present work, we show that the P. marneffei acuDgene is required for growth on gluconeogenic carbonsources such as acetate and fatty acids, is stronglyinduced by acetate and is dependent on the FacB tran-scriptional activator for acetate induction. More impor-tantly, P. marneffei acuD is also independently regulatedby the dimorphic switching developmental program andpart of this control is through the AbaA transcriptionalactivator. The developmental regulation of P. marneffeiacuD has both cis- and trans-acting elements which arenot present in the A. nidulans acuD gene or in A. nidulans,showing a unique evolutionary path for acetate and fattyacid regulation in this dimorphic pathogen.
Results
The P. marneffei acuD homologue
P. marneffei acuD was originally identified in a differentialdisplay expression screen as a gene highly expressed inyeast cells (C.R. Cooper Jr, Youngston University, pers.comm.; accession number AF373018). The open readingframe spans 1860 bp and consists of five exons and fourintrons (Fig. 1A). The predicted protein is 540 aa long andhighly similar to other fungal isocitrate lyases. It shows85% identity to the Aspergillus fumigatus predicted isoci-trate lyase (Q6T267), 82% to AcuD of A. nidulans(P28298), 81% to Icl1 of Coccidioides immitis (Q96TP5),and 64% to Icl1 of C. albicans (Q9P8Q7). Although pre-dicted to be localized to glyoxysomes (peroxisomes)these isocitrate lyases, included P. marneffei AcuD, lackobvious peroxisomal targeting sequences including thecarboxyl terminal PTS1 sequence.
Disruption of acuD function
An acuD deletion strain of P. marneffei was generated bytransformation of SPM4 (niaD1 pyrG1) with the deletionconstruct in pDAP4, in which the acuD open readingframe was replaced with the A. nidulans pyrG selectablemarker (Fig. 1A). Transformants were selected for uracilprototrophy and then screened for the absence of growthon minimal media containing acetate as the sole carbonsource. From 22 pyrG+ transformants, 10 were unable togrow on acetate but showed wild-type growth on glucoseas a carbon source. Examination of the deletion arrange-ment in these transformants by Southern blot hybridiza-tion showed that in one transformant the acuD gene wasdeleted and there were no ectopic integration events ofthe deletion construct. The remaining transformants eitherhad the wild-type copy of the gene and/or ectopic integra-tions of the deletion construct (data not shown).
1726 D. Cánovas and A. Andrianopoulos
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 62, 1725–1738

Additional growth tests revealed that the deletionmutant was also unable to grow on butyrate (4C) andTween 80 (18C) as sole carbon sources at 25°C (Fig. 1B).When the DacuD strain was grown on minimal mediacontaining both glucose and acetate at 25°C, growth waspartially impaired (data not shown). At 37°C, the deletionmutant failed to grow on SA (YNB medium containingacetate as the sole carbon source) or SD (YNB mediumcontaining glucose as the sole carbon source) (Fig. 2B).The addition of uracil to the culture medium restoredgrowth of the acuD deletion strain on SD but not on SA at37°C (Fig. 3). Addition of uracil also restored growth of theacuD deletion strain on minimal medium containingglucose and acetate at 25°C (data not shown). Onehypothesis for these observations is that conditions whichinduce expression from the acuD promoter lead to inter-ference with expression of the pyrG selectable marker inthe acuD deletion construct and therefore a failure tocomplement the uracil auxotrophy. To test this hypothesis,an DacuD::pyrG– strain was isolated by selection on5′-fluoroorotic acid. The resultant strain was transformedwith pALX223 (pyrG+) to isolate transformants with ran-domly integrated copies of pyrG in the genome which
would be unlinked to the acuD promoter and not besubject to transcriptional interference from the expressionarising from this promoter. The resultant transformants[DacuD::pyrG, pyrG+ (pALX223)] were able to grow on SD(containing glucose) but not on SA (containing acetate) at37°C (Fig. 2B), showing that a uracil auxotrophy occurringin the DacuD::pyrG+ was responsible for the absence ofgrowth on glucose at 37°C. Introduction of the acuD+ gene(pDAP70) complemented the growth defect of the dele-tion mutant on acetate at 25°C and 37°C (Fig. 2).
A number of additional experiments point to amechanism. First, the acuD deletion mutant also showedpoorer growth on medium containing glucose and acetatethan on glucose alone at 25°C and this was remediated byuracil. Second, a number of transformants containingectopic copies of the acuD deletion construct failed togrow on medium containing acetate as the sole carbonsource but lacking uracil despite having an intact acuDlocus. Coupled with the experiment where additionalectopic copies of pyrG complemented the uracil auxotro-phy, these observations are consistent with the idea thatthese strains are conditional uracil (pyrG–) auxotrophswhere the restrictive condition is dependent on acuD
250 bp
pyrG
Glucose Acetate Butyrate Tween 80
A
B
acuD
Carbon
Free
Xb
aI
Xb
aI
Xb
aI
Eco
RV
Eco
RV
Sal
I Sal
I
Hin
dIII
Hin
dIII
Wild-type
acuD::pyrG
Fig. 1. Construction of a DacuD deletion mutant.A. Diagram showing the gene structure of P. marneffei acuD with representative restriction sites shown. The coding region is indicated byboxes representing the exons interrupted by four introns. An acuD deletion construct was made by replacing an XbaI-EcoRV fragmentcontaining the whole coding region with the selectable marker pyrG.B. Growth of the wild-type and DacuD strains on minimal media containing different carbon sources. Plates were incubated for 10 days at25°C.
Glyoxylate cycle in Penicillium marneffei 1727
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 62, 1725–1738

promoter activity, which in turn interferes with pyrGexpression. One possible mechanism given the invertedorientation of the acuD and pyrG promoters in the deletionconstruct is that acuD promoter activity produces a mRNAcomplementary to the pyrG transcript and that thiscomplementary mRNA acts in trans by RNAi or antisensemechanisms to interfere with the pyrG selectable marker(Agrawal et al., 2003).
The acuD deletion construct removes, in addition to theentire coding region, the first 543 bp of acuD promoter(relative to the ATG) which encompasses four of the fivepredicted FacB binding sites and the transcription initia-tion site (Figs 1 and 6A). Therefore, acuD promotersequences upstream of -543 are sufficient for bothacetate induction and cell type (temperature) induction.Exactly which sequences are responsible for this regula-tion remains to be determined.
acuD expression is induced by acetate and temperature
Expression of acuD is regulated by carbon source inA. nidulans and other fungi like S. cerevisiae (Todd et al.,
1997; Schuller, 2003). To analyse acuD expression inP. marneffei, we performed Northern blot analysis usingRNA isolated from cells grown on SYBD (glucose) orSYBA (acetate) at both 25°C and 37°C (Fig. 3). As forA. nidulans acuD at 25°C, expression of P. marneffeiacuD is dependent on the carbon source, being inducedon acetate while having no detectable expression onglucose at 25°C. At 37°C acuD was expressed in thepresence of either glucose or acetate and the level ofexpression was substantially higher than at 25°C. Theaddition of acetate to the culture medium at 37°C slightlyincreased the expression of acuD over the levels onglucose. This clearly showed a different pattern of acuDregulation between A. nidulans and P. marneffei. Northernblot analysis also revealed a second but minor transcript(Fig. 3).
Comparison of acuD regulation in A. nidulans andP. marneffei
To analyse whether the differing regulation of acuD inA. nidulans and P. marneffei is regulator dependent(trans effect) or promoter dependent (cis effect), weplaced the red fluorescent protein (monomeric) mrfpreporter gene under the control of a 1.4 kb DNA frag-ment containing the acuD promoter from each organism,denoted PmacuD(p) and AnacuD(p). These constructswere used for the transformation of A. nidulans (targetedto the pyrG locus for PmacuD(p)::mrfp and to the pyrGor acuD locus for AnacuD(p)::mrfp) and P. marneffei (tar-geted to the areA locus). Of the pyrG+ transformants,two to three independently isolated strains were grownon glass slides covered with a thin layer of SD (contain-
Wild-type
acuD,vector
acuD::pyrG
acuD,pDAP70(acuD+)
Wild-type
acuD, vector
acuD::pyrG
acuD, pDAP70(acuD+)
Glucose + UGlucose
Acetate
Glucose
A
B
Acetate + U
Acetate
Fig. 2. Complementation of DacuD deletion strain. The pyrGselectable marker was eliminated from DacuD::pyrG+ by growth onmedia containing 5-FOA. The resulting DacuD pyrG– strain wastransformed with a plasmid containing the wild-type acuD (pDAP70)or the empty vector.A. The strains were grown on ANM medium containing differentcarbon sources for 10 days at 25°C.B. The strains were grown on YNB media containing differentcarbon sources in the presence or absence of uracil for 6 days at37°C. Please note that DacuD::pyrG+ did not grow on glucose inthe absence of uracil, but DacuD vector (pyrG+) did grow.
25ºC 37ºC
acuD
rRNA
GA G A
Fig. 3. acuD expression is induced by acetate and temperature.RNA was isolated from wild-type strain grown on SYB containingglucose or acetate as the carbon source at either 25°C or 37°C.RNA was hybridized with a probe specific for acuD. The majorlarger transcript is shown with a heavy arrow while the minorsmaller transcript is marked with a dashed arrow. rRNA is shownas loading controls.
1728 D. Cánovas and A. Andrianopoulos
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 62, 1725–1738
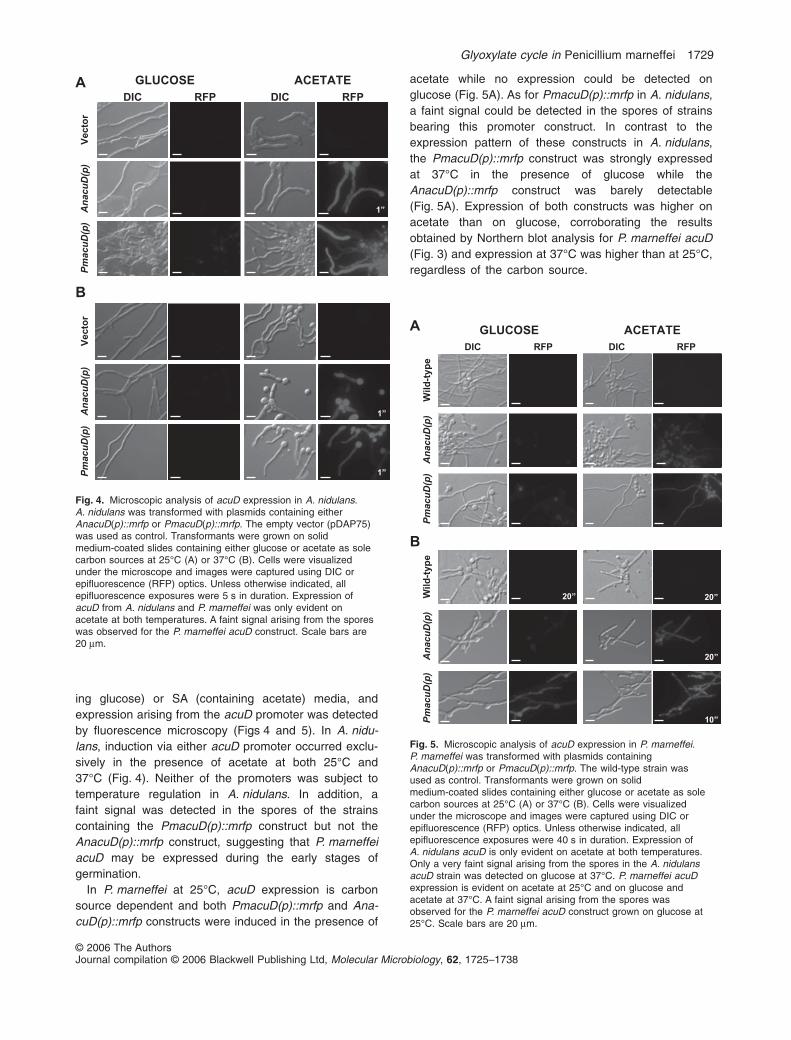
ing glucose) or SA (containing acetate) media, andexpression arising from the acuD promoter was detectedby fluorescence microscopy (Figs 4 and 5). In A. nidu-lans, induction via either acuD promoter occurred exclu-sively in the presence of acetate at both 25°C and37°C (Fig. 4). Neither of the promoters was subject totemperature regulation in A. nidulans. In addition, afaint signal was detected in the spores of the strainscontaining the PmacuD(p)::mrfp construct but not theAnacuD(p)::mrfp construct, suggesting that P. marneffeiacuD may be expressed during the early stages ofgermination.
In P. marneffei at 25°C, acuD expression is carbonsource dependent and both PmacuD(p)::mrfp and Ana-cuD(p)::mrfp constructs were induced in the presence of
acetate while no expression could be detected onglucose (Fig. 5A). As for PmacuD(p)::mrfp in A. nidulans,a faint signal could be detected in the spores of strainsbearing this promoter construct. In contrast to theexpression pattern of these constructs in A. nidulans,the PmacuD(p)::mrfp construct was strongly expressedat 37°C in the presence of glucose while theAnacuD(p)::mrfp construct was barely detectable(Fig. 5A). Expression of both constructs was higher onacetate than on glucose, corroborating the resultsobtained by Northern blot analysis for P. marneffei acuD(Fig. 3) and expression at 37°C was higher than at 25°C,regardless of the carbon source.
An
acu
D(p
)P
mac
uD
(p)
Vecto
r
DIC RFP DIC RFP
GLUCOSE ACETATEV
ecto
rA
nac
uD
(p)
Pm
acu
D(p
)A
B
1”
1”
1”
Fig. 4. Microscopic analysis of acuD expression in A. nidulans.A. nidulans was transformed with plasmids containing eitherAnacuD(p)::mrfp or PmacuD(p)::mrfp. The empty vector (pDAP75)was used as control. Transformants were grown on solidmedium-coated slides containing either glucose or acetate as solecarbon sources at 25°C (A) or 37°C (B). Cells were visualizedunder the microscope and images were captured using DIC orepifluorescence (RFP) optics. Unless otherwise indicated, allepifluorescence exposures were 5 s in duration. Expression ofacuD from A. nidulans and P. marneffei was only evident onacetate at both temperatures. A faint signal arising from the sporeswas observed for the P. marneffei acuD construct. Scale bars are20 mm.
GLUCOSE ACETATEA
B
An
acu
D(p
)P
mac
uD
(p)
Wild
-typ
eA
nac
uD
(p)
Pm
acu
D(p
)
DIC RFP
10”
20”
20”
DIC RFP
20”Wild
-typ
e
Fig. 5. Microscopic analysis of acuD expression in P. marneffei.P. marneffei was transformed with plasmids containingAnacuD(p)::mrfp or PmacuD(p)::mrfp. The wild-type strain wasused as control. Transformants were grown on solidmedium-coated slides containing either glucose or acetate as solecarbon sources at 25°C (A) or 37°C (B). Cells were visualizedunder the microscope and images were captured using DIC orepifluorescence (RFP) optics. Unless otherwise indicated, allepifluorescence exposures were 40 s in duration. Expression ofA. nidulans acuD is only evident on acetate at both temperatures.Only a very faint signal arising from the spores in the A. nidulansacuD strain was detected on glucose at 37°C. P. marneffei acuDexpression is evident on acetate at 25°C and on glucose andacetate at 37°C. A faint signal arising from the spores wasobserved for the P. marneffei acuD construct grown on glucose at25°C. Scale bars are 20 mm.
Glyoxylate cycle in Penicillium marneffei 1729
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 62, 1725–1738

Comparison of the acuD promoter of P. marneffei andthat in the two closest relatives A. nidulans andA. fumigatus
From the expression studies, it was hypothesized thatthere are two levels of regulation of acuD in P. marneffeiwhich are likely to operate independently. We looked forpredicted transcription factor binding sites in the promoterof P. marneffei acuD and then compared these sites withthose found in the acuD promoters of A. nidulans andA. fumigatus (Fig. 6 and Table 1). The acuD promoter ofA. nidulans has eight potential FacB binding sites, ofwhich three sites are bound by FacB in vitro (Todd et al.,1998). In P. marneffei, there are five potential FacBbinding sites, three of which are overlapping andcontiguous. Within the three overlapping FacB bindingsites there is also a predicted binding site for the glucose-dependent repressor CreA (Kulmburg et al., 1993)(Fig. 6). In addition, the A. nidulans acuD promoter hasfive CreA binding sites, one BrlA binding site (Chang andTimberlake, 1993), three AbaA binding sites (Andrianopo-ulos and Timberlake, 1994) and three CCGAGG motifs(Hynes et al., 2006) while the P. marneffei acuD promoterhas two CreA, two AbaA binding sites and one CCGAGGmotif but no BrlA binding sites. The acuD promoter of
A. fumigatus was more similar to the A. nidulans thanP. marneffei containing three CreA binding sites, fourAbaA binding sites, nine FacB binding sites, threeCCGAGG motifs and no BrlA binding sites (Table 1). Inaddition, promoter analysis using MATCH software (http://www.gene-regulation.com/pub/programs.html#match)with a cut-off set to minimize false positives revealed twoCCAAT boxes inverted with respect to each other, sepa-rated by 100 bp and surrounding the three overlappingFacB binding sites and two overlapping CCAAT displace-ment protein (CDP) binding sites in the P. marneffei acuDpromoter but not in the Aspergillus acuD promoters. Inhigher eukaryotes, CDP is believed to act as a repressorof gene transcription (Barberis et al., 1987). However, nohomologues of CDP have been found in fungi to date(Gillingham et al., 2002).
acuD expression is regulated by the acetate-dependentregulator FacB in P. marneffei
FacB is a transcriptional activator involved in theacetate-dependent induction of acuD in A. nidulans(Katz and Hynes, 1989). To test whether acuD expres-sion in P. marneffei was dependent on FacB, Northern
Fig. 6. Diagram of the regulatory sites inacuD promoters. Schematic representation ofthe acuD promoters from A. nidulans (An),A. fumigatus (Af) and P. marneffei (Pm)showing the predicted and confirmed bindingsites for the transcriptional regulators AbaA(CATTCY), BrlA (MRAGGGR), FacB(TCSN8-10SGA and GCMN8-10KGC) and CreA(SYGGGG and GCGGAG). The CCGAGGmotives required for fatty acid regulation, theCCAAT boxes and the CDP binding sites arealso indicated. The presence of twooverlapping FacB binding sites in A. fumigatusand three overlapping FacB binding sites inP. marneffei is indicated in the sequencesabove and below the promoter representationrespectively. The predicted CreA binding siteoverlapping with the FacB binding site isboxed. The XbaI site used to make theP. marneffei acuD deletion construct isindicated.
100 bps
AbaA sites
BrlA sites
GCCTCCTCCGAATTGCGGAGGA
GCCTCGAGAGATGCTCGA
FacB sites (predicted)
FacB sites (confirmed)
An
CCGAGG motif
CreA sites
Pm
XbaI
CCAAT box
CDP sites
Af
Table 1. Summary of predicted transcriptionalregulator binding sites in acuD promoters.A. nidulans A. fumigatus P. marneffei
AbaA (CATTCY) 4 4 2BrlA (MRAGGGR) 1 0 0FacB (TCSN8-10SGA and GCMN8-10KGC) 8a 9 5CreA (SYGGRG) 5 3 2CCGAGG motif 3 3 1
a. Three of which are bound by FacB in vitro (Todd et al., 1998).
1730 D. Cánovas and A. Andrianopoulos
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 62, 1725–1738
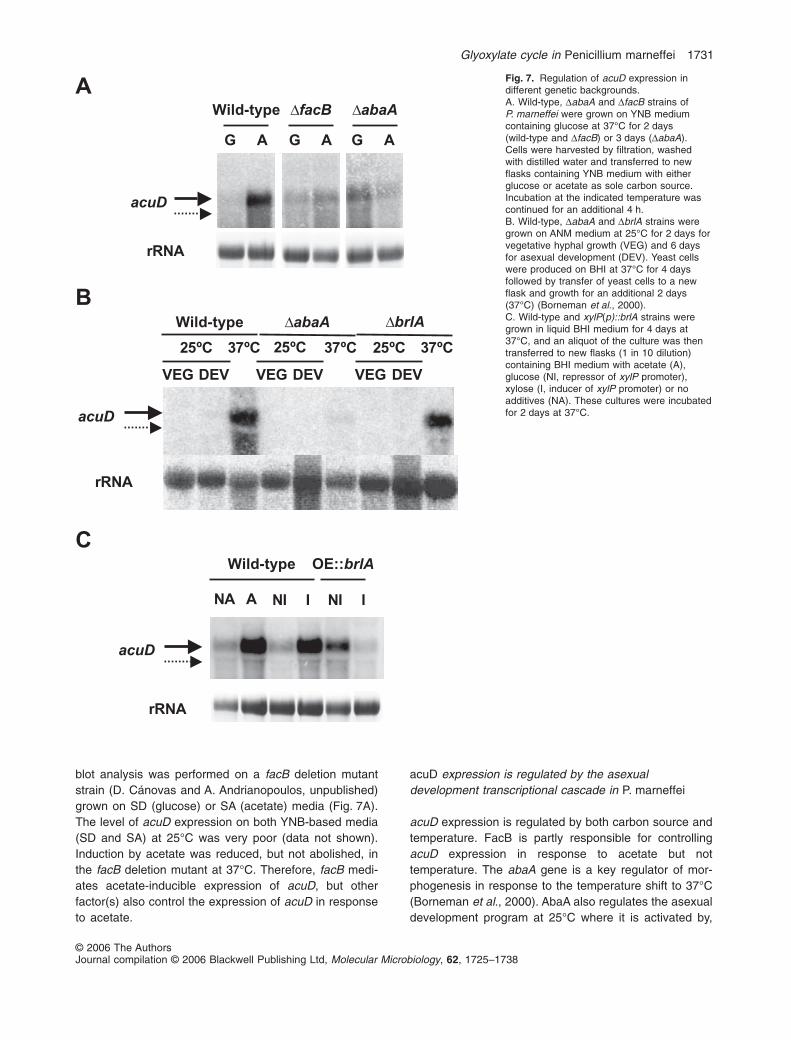
blot analysis was performed on a facB deletion mutantstrain (D. Cánovas and A. Andrianopoulos, unpublished)grown on SD (glucose) or SA (acetate) media (Fig. 7A).The level of acuD expression on both YNB-based media(SD and SA) at 25°C was very poor (data not shown).Induction by acetate was reduced, but not abolished, inthe facB deletion mutant at 37°C. Therefore, facB medi-ates acetate-inducible expression of acuD, but otherfactor(s) also control the expression of acuD in responseto acetate.
acuD expression is regulated by the asexualdevelopment transcriptional cascade in P. marneffei
acuD expression is regulated by both carbon source andtemperature. FacB is partly responsible for controllingacuD expression in response to acetate but nottemperature. The abaA gene is a key regulator of mor-phogenesis in response to the temperature shift to 37°C(Borneman et al., 2000). AbaA also regulates the asexualdevelopment program at 25°C where it is activated by,
Fig. 7. Regulation of acuD expression indifferent genetic backgrounds.A. Wild-type, DabaA and DfacB strains ofP. marneffei were grown on YNB mediumcontaining glucose at 37°C for 2 days(wild-type and DfacB) or 3 days (DabaA).Cells were harvested by filtration, washedwith distilled water and transferred to newflasks containing YNB medium with eitherglucose or acetate as sole carbon source.Incubation at the indicated temperature wascontinued for an additional 4 h.B. Wild-type, DabaA and DbrlA strains weregrown on ANM medium at 25°C for 2 days forvegetative hyphal growth (VEG) and 6 daysfor asexual development (DEV). Yeast cellswere produced on BHI at 37°C for 4 daysfollowed by transfer of yeast cells to a newflask and growth for an additional 2 days(37°C) (Borneman et al., 2000).C. Wild-type and xylP(p)::brlA strains weregrown in liquid BHI medium for 4 days at37°C, and an aliquot of the culture was thentransferred to new flasks (1 in 10 dilution)containing BHI medium with acetate (A),glucose (NI, repressor of xylP promoter),xylose (I, inducer of xylP promoter) or noadditives (NA). These cultures were incubatedfor 2 days at 37°C.
A
B
C
acuD
Wild-type
INIINI
OE::brlA
A
rRNA
NA
Wild-type abaA brlA
25ºC 37ºC
DEV
37ºC 37ºC25ºC
DEV
25ºC
DEV
acuD
rRNA
GEVGEV VEG
acuD
rRNA
G A
abaAfacB
AGG A
Wild-type
Glyoxylate cycle in Penicillium marneffei 1731
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 62, 1725–1738
and feedback activates, the brlA regulatory gene. Giventhat there are two predicted AbaA binding sites in thepromoter of P. marneffei acuD (Fig. 6A), the role of AbaAin acuD expression was examined. Wild-type, DabaA andDbrlA mutant strains of P. marneffei were grown under thestandard growth conditions which promote hyphal growthand asexual development [A. nidulans medium (ANM) at25°C] or dimorphic switching [brain–heart infusion (BHI)at 37°C], and acuD expression was analysed by Northernblot (Fig. 7B). The expression of acuD at 37°C wasseverely reduced in the abaA deletion mutant but unaf-fected in the brlA deletion mutant. At 25°C during asexualdevelopment, acuD was expressed at low levels in thewild-type strain but absent in both the DabaA or DbrlAmutants. Given that AbaA strongly induces the expressionof acuD at 37°C and directs the expression of genesduring conidiation at 25°C, it is not clear why acuDexpression is low during conidiation. One possibility is thatacuD is repressed during asexual development, possiblyvia brlA, thereby overriding AbaA activation. To test thishypothesis, a strain of P. marneffei bearing A. nidulansbrlA under the control of the inducible xylP promoter wasgrown at 37°C on BHI under repressing (glucose) andinducing (xylose) conditions. Northern blot analysisrevealed that overexpression of brlA dramatically reducedthe expression of acuD (Fig. 7C). There are no BrlAbinding sites in the promoter of P. marneffei acuD(Fig. 6A), suggesting that BrlA-mediated repression islikely to be indirect.
AbaA is required for controlling acuD expression in atemperature-dependent manner. To examine whetherAbaA is also involved in the acetate-dependent inductionof acuD, the abaA mutant strain was grown in SD and SA.Northern blot analysis showed that the level of acuDexpression on acetate was significantly lower in theDabaA mutant compared with the wild type suggestingthat acetate induction is partially mediated by AbaA at37°C (Fig. 7A). In addition, the level of acuD expressionon glucose was higher than the wild type.
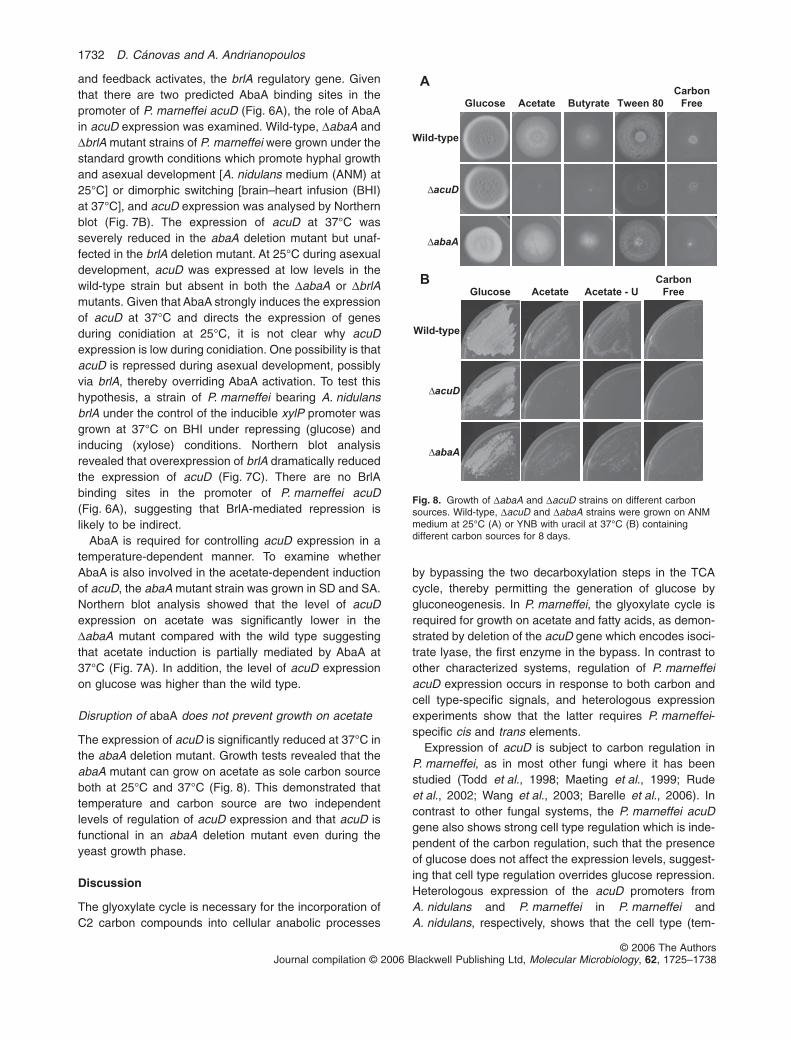
Disruption of abaA does not prevent growth on acetate
The expression of acuD is significantly reduced at 37°C inthe abaA deletion mutant. Growth tests revealed that theabaA mutant can grow on acetate as sole carbon sourceboth at 25°C and 37°C (Fig. 8). This demonstrated thattemperature and carbon source are two independentlevels of regulation of acuD expression and that acuD isfunctional in an abaA deletion mutant even during theyeast growth phase.
Discussion
The glyoxylate cycle is necessary for the incorporation ofC2 carbon compounds into cellular anabolic processes
by bypassing the two decarboxylation steps in the TCAcycle, thereby permitting the generation of glucose bygluconeogenesis. In P. marneffei, the glyoxylate cycle isrequired for growth on acetate and fatty acids, as demon-strated by deletion of the acuD gene which encodes isoci-trate lyase, the first enzyme in the bypass. In contrast toother characterized systems, regulation of P. marneffeiacuD expression occurs in response to both carbon andcell type-specific signals, and heterologous expressionexperiments show that the latter requires P. marneffei-specific cis and trans elements.
Expression of acuD is subject to carbon regulation inP. marneffei, as in most other fungi where it has beenstudied (Todd et al., 1998; Maeting et al., 1999; Rudeet al., 2002; Wang et al., 2003; Barelle et al., 2006). Incontrast to other fungal systems, the P. marneffei acuDgene also shows strong cell type regulation which is inde-pendent of the carbon regulation, such that the presenceof glucose does not affect the expression levels, suggest-ing that cell type regulation overrides glucose repression.Heterologous expression of the acuD promoters fromA. nidulans and P. marneffei in P. marneffei andA. nidulans, respectively, shows that the cell type (tem-
Glucose Acetate Butyrate Tween 80
Carbon
Free
Glucose Acetate
Carbon
FreeAcetate - U
A
B
Wild-type
acuD
abaA
Wild-type
acuD
abaA
Fig. 8. Growth of DabaA and DacuD strains on different carbonsources. Wild-type, DacuD and DabaA strains were grown on ANMmedium at 25°C (A) or YNB with uracil at 37°C (B) containingdifferent carbon sources for 8 days.
1732 D. Cánovas and A. Andrianopoulos
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 62, 1725–1738

perature) regulation is restricted to P. marneffei harbour-ing its native acuD promoter. This suggests the existenceof unique elements in the promoter of P. marneffei acuD,which are controlled by specific regulator(s) present inP. marneffei but not in A. nidulans. This regulatory systemcould also control the expression of other genes includingthose involved in pathogenesis. Comparison of the pro-moter sequence in both organisms revealed a number ofputative binding sites for transcriptional regulatorscommon to both organisms. Besides positional andnumber differences for the various transcription factorbinding sites, the other difference is the presence of twoCCAAT boxes which are binding sites for a wide domain,multimeric transcriptional activator complex found ineukaryotes (Brakhage et al., 1999). This CCAAT bindingcomplex has been shown to contribute to setting highbasal levels of expression as well as in facilitating activa-tion, and in one case repression, by other transcriptionalregulators (Aramayo and Timberlake, 1993; Brakhageet al., 1999; Zhang et al., 1999; Schuller, 2003; McNabband Pinto, 2005). The fact that this complex has beenimplicated in carbon metabolism, nitrogen metabolism,development and virulence in fungi suggests that theCCAAT binding factor may contribute to the expression ofgluconeogenic genes (i.e. acuD) that may be upregulatedduring infection in P. marneffei.
acuD is independently regulated by acetate and tem-perature in P. marneffei and a combination of both induc-ing conditions resulted in the highest level of expression,suggesting additive relationship. A minor transcript wasalso evident but it is currently unclear whether this is aresult of differential splicing or differential transcriptionalinitiation and/or termination. Both A. nidulans acuD andS. cerevisiae ICL1 genes are strictly regulated by carbonsource but not temperature (De Lucas et al., 1994b; Ordizet al., 1995). At the transcriptional level, A. nidulans acuDand S. cerevisiae ICL1 expression is upregulated in thepresence of acetate and repressed by glucose (Fernan-dez et al., 1993; Bowyer et al., 1994; De Lucas et al.,1994a). Glucose repression in S. cerevisiae is mediatedby Mig1, a C2H2 zinc finger regulator; however, glucoserepression of ICL1 is indirect even though the promotercontains a Mig1 binding site. Glucose derepression ismediated by Snf1, which inactivates Mig1 and this, in turn,allows the expression of CAT8, which activates theexpression of SIP4. Both Cat8 and Sip4 act in combina-tion to induce the expression of gluconeogenic genes,including ICL1, by binding to a carbon source responsiveelement, YCCRTTNRNCCG (Schuller, 2003). InA. nidulans, glucose repression is mediated by CreA,which binds to the consensus sequence SYGGRG (Kul-mburg et al., 1993; Cubero and Scazzocchio, 1994), pre-venting the transcription of target genes (Mathieu andFelenbok, 1994). However, in P. marneffei, acuD is also
expressed in the presence of glucose at 37°C, thereforetemperature induction overrides any putative glucoserepression control of acuD expression. It is noteworthythat two potential CreA binding sites are present in theacuD promoter, one of which is located directly on theDNA binding region containing three overlapping FacBsites (Fig. 6).
In A. nidulans acetate induction of acuD expression isdependent on the transcriptional regulator FacB andthree FacB binding sites have been identified, consistingof two dissimilar DNA sequences (De Lucas et al.,1994a; Todd et al., 1998). Examination of the P. marnef-fei acuD promoter sequence and comparison with theA. nidulans and A. fumigatus promoters revealed anumber of conserved binding sites for FacB. There arefive additional potential sites in A. nidulans, nine in A. fu-migatus and five in P. marneffei. The position of the siteswith respect to the ATG are not conserved in the threespecies. Interestingly, the two pathogenic speciescontain overlapping FacB binding sites (two in A. fumi-gatus and three in P. marneffei); however, the biologicalsignificance of this is unclear. In the P. marneffei facBdeletion strain, acetate induction of acuD is substantiallyreduced at 37°C. Given that acuD expression at 37°C isnot completely abolished, this shows that FacB is notrequired for temperature-dependent induction. Further-more, the higher levels of acuD mRNA at 37°C onacetate compared with glucose indicates that this iseither the result of glucose derepression leading to highbasal levels of acuD expression or there is an additionalweak acetate induction mechanism involving an uniden-tified transcriptional activator.
The P. marneffei abaA gene controls both differentiationof the sporogenic phialide cell type during asexual devel-opment at 25°C and the coupling of nuclear division withcell division during yeast cell morphogenesis at 37°C(Borneman et al., 2000). The downstream structuralgenes which are regulated by AbaA are poorlyunderstood. There are two potential AbaA binding sites inthe P. marneffei acuD promoter and four potential bindingsites in the promoters of the A. nidulans and A. fumigatusacuD genes. Deletion of the P. marneffei abaA gene leadsto an almost complete loss of acuD expression at 37°Cshowing that this is the prime morphological regulator ofacuD. In addition, loss of abaA also leads to a defect inacetate induction of acuD at 37°C suggesting that eitherAbaA activates the facB gene which in turn activates acuDexpression or that FacB and AbaA are co-dependent foracetate induction of acuD at 37°C. Studies in othersystems have shown that pathogens reprogram theirmetabolic patterns when growing inside the host (McKin-ney et al., 2000; Lorenz and Fink, 2001) and this is alsolikely to be true for P. marneffei, possibly via the develop-mental regulator AbaA. Interestingly, there are three
Glyoxylate cycle in Penicillium marneffei 1733
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 62, 1725–1738

binding sites for the AbaA homologue Tec1 (Andrianopo-ulos and Timberlake, 1994; Madhani and Fink, 1997) inthe ICL1 promoter of C. albicans but none in the genefrom the non-pathogenic S. cerevisiae. Tec1 is involved infilamentation and pathogenesis in C. albicans (Schweizeret al., 2000) but only in pseudohyphal growth inS. cerevisiae (Lengeler et al., 2000). Whether this is ageneral mechanism occurring in other dimorphic fungiremains to be discovered. A screen for phase-specificexpressed sequence tags in the thermally dimorphicfungal pathogen Paracoccidioides braziliensis also iden-tified an acuD orthologue and it was shown to be differ-entially expressed during the transition from yeast tomycelium (Goldman et al., 2003). The fact that AbaA regu-lates a gene involved in carbon metabolism at 37°C, andimplicated in pathogenicity in some other fungal patho-gens, implies that it may be important for the regulation ofother pathogenic determinants.
The acuD gene is expressed at low levels duringasexual development and this expression is absent inthe abaA deletion mutant. During asexual developmentbut not yeast cell morphogenesis, the abaA gene is acti-vated by the product of the brlA gene (A.R. Bornemanet al., unpublished). Consistent with these observations,loss of brlA has no effect on acuD expression at 37°Cbut does lead to a loss of expression during asexualdevelopment at 25°C. The high level of AbaA-dependentacuD expression in yeast cells is in stark contrast to thelow levels during asexual development. One possibleexplanation for these results is that BrlA is acting asboth an activator and repressor of gene transcription. Insupport of this, ectopic overexpression of BrlA at 37°Cresults in a dramatic downregulation of acuD expression.Such a mechanism may be one way in which the activityof regulatory factors such as AbaA, which participates intwo distinct developmental programs, are cell typelimited in their targets. It is unknown whether BrlA-mediated repression occurs by direct binding to the pro-moter (no BrlA binding site was found in P. marneffeiacuD promoter), by activating the transcription of someother repressing regulator(s) or by some other unknownmechanism. Alternatively, these results may beexplained by the loss of the cell types in which acuD isexpressed.
The second step in the glyoxylate bypass is catalysedby malate synthase and it would be expected that thegene encoding this enzyme would be co-ordinately regu-lated with acuD, The P. marneffei acuE gene encodesmalate synthase and this gene is regulated in an iden-tical manner to acuD responding to both temperatureand acetate induction, showing AbaA-dependent expres-sion and showing reduced expression when brlA isoverexpressed (Fig. S1). Therefore, the described regu-latory pattern is not unique to acuD, as the acuE gene
exhibits identical regulation, showing that the wholeglyoxylate bypass is co-ordinately regulated by thismechanism.
In A. nidulans a suppressive subtraction hybridizationscreen for genes expressed in conidia identified a numberof genes including acuD and it was shown that expressionof acuD is significantly reduced in strains carrying muta-tions in the brlA and abaA genes (Osherov et al., 2002).The acuD gene was also previously identified in a screenfor genes expressed during asexual development (termedCAN genes for conidiation in A. nidulans) (Boylan et al.,1987; Stringer, 1994). The role of acuD expression duringasexual development is not clear but it can be hypoth-esized that the glyoxylate cycle is used to derive energy indeveloping conidiophores which grow away from thegrowth substrate and thus may be deprived of nutrients. InP. marneffei, the acuD deletion strain does not show anysigns of delayed germination at either 25°C or 37°C sug-gesting that it is not required for conidia to exit fromdormancy and initiate growth (data not shown), furtherimplicating its role in conidiogenesis rather thangermination.
The pathology of P. marneffei is poorly understood. It isassumed that infection occurs via inhalation of conidia,which are subsequently phagocytosed by macrophages(Vanittanakom et al., 2006). Macrophages are consideredan environment poor in nutrients. Consequently, genesinvolved in the glyoxylate cycle in other fungi, such asC. albicans, are upregulated when the fungal cells areinside the macrophages (Lorenz and Fink, 2001; Barelleet al., 2006). In C. albicans, concentrations of glucoseas low as those found in the bloodstream are enoughto downregulate glyoxylate cycle genes. Therefore,C. albicans reprograms its carbon metabolism from anactivated glyoxylate cycle during early stages of infectioninside the macrophage to the activation of glycolysisduring the progression of systemic disease (Barelle et al.,2006). Although P. marneffei primarily produces pulmo-nary infections, it can also be disseminated to otherorgans, so that yeast cells are found both in intracellularand extracellular environments within the host (Vanittana-kom et al., 2006). As the acuD gene is tightly regulated byboth temperature and carbon source, it would beexpected that in contrast to C. albicans, the glyoxylatecycle remains upregulated during the entire P. marneffeiinfection cycle. These regulatory differences may be aconsequence of the commensal versus opportunistic lifestyles of these two pathogens.
Induction of the glyoxylate cycle in pathogenic microor-ganisms is presumed to be a response to nutrient depri-vation inside the macrophages (Lorenz and Fink, 2002);however, the regulatory mechanisms controlling theexpression of the glyoxylate cycle genes in pathogenshad not been addressed previously. This study shows that
1734 D. Cánovas and A. Andrianopoulos
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 62, 1725–1738

the genes belonging to the glyoxylate bypass are highlyexpressed during the pathogenic yeast form inP. marneffei. The expression of these genes is tightly con-trolled by the acetate-responsive regulator FacB and thedevelopmental regulator AbaA (Fig. 9). These resultsprovide evidence for a link between carbon metabolism,development and pathogenicity in the dimorphic fungalpathogen P. marneffei.
Experimental procedures
Strains, media and growth conditions
Fungal strains used in this study are listed in Table 2.A. nidulans strains were regularly grown on Aspergillusdefined medium (ANM) (Cove, 1966). P. marneffei strainswere grown on either ANM or yeast synthetic medium(YNB, Yeast Nitrogen Base) (Ausubel et al., 1989) contain-ing the appropriate carbon and nitrogen sources, or thecomplex BHI (Oxoid) or SYB (2% yeast extract, 1% pro-
tease peptone) medium. (NH4)2SO4 at a final concentrationof 10 mM was used as the nitrogen source in the definedmedia. Glucose (1–2%), acetate (50 mM), butyrate (20 mM)or Tween 80 (0.2%) were used as carbon sources in themedia. Auxotrophic supplements were added as required.Escherichia coli TOP10 (Invitrogen Life Technologies) andJM110 dam– were used for the propagation of plasmid DNA.
Molecular techniques
Genomic DNA was isolated as previously described (Borne-man et al., 2001). RNA was prepared by using the FastRNAred kit (BIO101). RNA samples were separated on 1.2%agarose formaldehyde denaturing gels, blotted onto Amer-sham Hybond N + membranes and analysed by Northernblot hybridization (Sambrook et al., 1989). The acuD andacuE probes for Northern blotting were generatedby PCR using the primers pm-icl1upper1 (5′-GTGGTACACTCAAGATTGAGTATC-3′) and pm-icl1lower1 (5′-GTTCTAGAATAACGAACATTAGAG-3′), and pmacuEnort43(5′-GAACTGTCAACAACGAACATC-3′) and pmacuEnort592
Fig. 9. Model for acuD gene regulation inP. marneffei. P. marneffei can undergo twodifferent developmental programs asexualdevelopment (conidiation) at 25°C anddimorphic switching at 37°C. Duringvegetative hyphal growth at 25°C, the FacBgene controls induction of acuD expression inresponse to acetate. The AbaA transcriptionalactivator is induced during both asexualdevelopment at 25°C and dimorphic switchingat 37°C (Borneman et al., 2000). At 25°Cduring asexual development, AbaA activatesconidiation-specific genes and acuDexpression. At 37°C during yeast cellmorphogenesis, AbaA activates yeast-specificgenes and directly activates acuD in responseto temperature and acetate or indirectlyactivates acuD by activating facB expression.
AbaA
acuD
FacB
Acetate
Yeast
genes
Conidiation
genes
25°C 37°C
Yeast
Growth
Asexual
Development
Hyphal
Growth
AbaA
acuDacuD
Table 2. Fungal strains used in this work.
Strain Genotype Reference
P. marneffeiFRR2161 Wild type ATCC 18224SPM4 niaD1 pyrG1 Borneman et al. (2001)
niaD1 pyrG1 areA H. Smith and A. Andrianopoulos (unpubl. results)niaD1 pyrG1DacuD::AnpyrG+ This studyniaD1 pyrG1DacuDniaD1 pyrG1DacuD::pyrG–; PmacuD AnpyrG+ (pDAP70) This studyniaD1 pyrG1DacuD::pyrG–AnpyrG+ (pALX223) This studyniaD1 pyrG1 AnacuD(p)::mrfp AnpyrG+ (pDAP76) This studyniaD1 pyrG1 PmacuD(p)::mrfp AnpyrG+ (pDAP77) This studyniaD1 pyrG1DfacB::AnpyrG+ D. Cánovas and A. Andrianopoulos (unpublished)niaD1 pyrG1DabaA::AnpyrG+ Borneman et al. (2000)niaD1 pyrG1DbrlA::AnpyrG+ A.R. Borneman et al. (unpublished)niaD1 pyrG1 xylP(p)::AnbrlA A.R. Borneman et al. (unpublished)
A. nidulansTN02A3 nkuA::argB pyrG89 pyroA4 Dr Berl Oakley
nkuA::argB pyrG89 pyroA4 AnpyrG+ (pDAP75) This studynkuA::argB pyrG89 pyroA4 AnacuD(p)::mrfp AnpyrG+ (pDAP76) This studynkuA::argB pyrG89 pyroA4 PmacuD(p)::mrfp AnpyrG+ (pDAP77) This study
Glyoxylate cycle in Penicillium marneffei 1735
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 62, 1725–1738

(5′-GACTGTGAAATGCTTCTCCTC-3′) respectively. DNA-mediated transformation of P. marneffei and A. nidulans hasbeen previously described (Borneman et al., 2001; Nayaket al., 2005).
Cloning and plasmid construction
The acuD (ICL1) DNA sequence of P. marneffei is depositedin the GenBank database (accession number AF373018).acuD was amplified from genomic DNA by using primersicl1for1 (5′-GATCGTAGAGACTAGATGACC-3′) and icl1rev1(5′-ACCTGTATCGATTGACAATTC-3′) and cloned intopGEM®-T Easy vector (Promega). The DacuD deletion con-struct plasmid (pDAP4) was generated by replacing an XbaI-EcoRV fragment spanning the entire acuD coding region withthe A. nidulans pyrG selectable marker (Fig. 1B).
PmacuD(p)::mrfp and AnacuD(p)::mrfp were constructedby assembling two PCR products. Both contain 1.4 kb of thepromoter region immediately adjacent to the ATG of eachgene. The promoter of P. marneffei acuD was amplified usingthe primers acudfor5′xho1 (5′-ACTCGAGCACCCCATACCAACAATC-3′) and acudrev5′mrfp (5′-GTCCTCGGAGGAGGCCATTGTGAATTTT-3′). The A. nidulans acuDpromoter was amplified by using primers AnacuDpup(5′-ATCTTCTTCCCATGCAACGG-3′) and AnacuDplowRFP(5′-GTCCTCGGAGGAGGCCATGATGGCAGTATTCAGC-3′).The mrfp gene was amplified from plasmid pMT-mRFP1(Toews et al., 2004) using the primers mRFPforw (5′-CATATGGCCTCCTCCGAGGACG-3′) and mRFPrev (5′-GGCGCCGGTGGAGTGGCG-3′). The PCR fragmentcontaining the P. marneffei promoter fused to the mrfpreporter gene was cloned into pGEM®-T Easy to produce thePmacuD(p)::mrfp fusion vector (pDAP28). pDAP75 was gen-erated by inserting the 3′ region of the P. marneffei areA geneinto pALX223 to allow targeting into the areA locus ofP. marneffei (H. Smith and A. Andrianopoulos, unpubl.results). pDAP28 was then digested with NotI and the Pma-cuD(p)::mrfp fragment cloned into pDAP75 to give the finalplasmid pDAP77 (pyrG+, areA*, PmacuD(p)::mrfp). The PCRfragment containing the A. nidulans acuD promoter fused tothe mrfp reporter gene was cloned into pALX223 digestedwith EcoRV (pDAP71). areA* was then cloned into the EcoRIsite giving the final plasmid pDAP76 (pyrG+, areA*,AnacuD(p)::mrfp).
For complementation of the DacuD deletion mutant, theacuD gene was generated by PCR using primers icl1for1(5′-GATCGTAGAGACTAGATGACC-3′) and icl1rev1 (5′-ACCTGTATCGATTGACAATTC-3′) and cloned into pALX223digested with EcoRV to yield pDAP70.
Microscopy
P. marneffei and A. nidulans strains were grown on slidescovered with a thin layer of solid medium, inoculated withconidia from the appropriate strains and incubated at theindicated temperature, except for P. marneffei cultures at37°C where conidia were allowed to germinate on liquidmedium and then visualized under the microscope. All strainswere grown on yeast nitrogen base containing 10 mM(NH4)2SO4 as a nitrogen source and either 2% glucose (SD)
or 50 mM sodium acetate (SA) as a sole carbon source at theindicated temperature. Slides were visualized on a ReichartJung Polyvar II microscope using differential interferencecontrast (DIC) or epifluorescence optics for RFP. Imageswere captured using a SPOT CCD camera (Diagnostic Instru-ments) and processed in Adobe Photoshop™ 7.0.
Acknowledgements
D.C. thanks all members of the Andrianopoulos, Hynes andDavis groups for helpful discussions. Dr Richard Todd, DrReinhard Fischer and Dr Berl Oakley kindly provided a partialclone of the P. marneffei facB gene, the pMT-mRFP1 plasmidand the A. nidulans TNO2A3 strain respectively. D.C. wassupported by a Marie Curie OIF. This work was supported bygrants from the Australian Research Council, National Healthand Medical Research Council and Howard Hughes MedicalInstitute.
References
Adams, T.H., Boylan, M.T., and Timberlake, W.E. (1988) brlAis necessary and sufficient to direct conidiophore develop-ment in Aspergillus nidulans. Cell 54: 353–362.
Adams, T.H., Wieser, J.K., and Yu, J.H. (1998) Asexualsporulation in Aspergillus nidulans. Microbiol Mol Biol Rev62: 35–54.
Agrawal, N., Dasaradhi, P.V., Mohmmed, A., Malhotra, P.,Bhatnagar, R.K., and Mukherjee, S.K. (2003) RNA interfer-ence: biology, mechanism, and applications. Microbiol MolBiol Rev 67: 657–685.
Andrianopoulos, A. (2002) Control of morphogenesis in thehuman fungal pathogen Penicillium marneffei. Int J MedMicrobiol 292: 331–347.
Andrianopoulos, A., and Timberlake, W.E. (1991) ATTS, anew and conserved DNA binding domain. Plant Cell 3:747–748.
Andrianopoulos, A., and Timberlake, W.E. (1994) TheAspergillus nidulans abaA gene encodes a transcriptionalactivator that acts as a genetic switch to controldevelopment. Mol Cell Biol 14: 2503–2515.
Aramayo, R., and Timberlake, W.E. (1993) The Aspergillusnidulans yA gene is regulated by abaA. EMBO J 12: 2039–2048.
Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D.,Seidman, J.G., Smith, J.R., and Struhl, K. (1989) CurrentProtocols in Molecular Biology. New York, NY: GreeneAssociates/Wiley Interscience.
Barberis, A., Superti-Furga, G., and Busslinger, M. (1987)Mutually exclusive interaction of the CCAAT-binding factorand of a displacement protein with overlapping sequencesof a histone gene promoter. Cell 50: 347–359.
Barelle, C.J., Priest, C.L., MacCallum, D.M., Gow, N.A.R.,Odds, F.C., and Brown, A.J.P. (2006) Niche-specific regu-lation of central metabolic pathways in a fungal pathogen.Cell Microbiol 8: 961–971.
Borneman, A.R., Hynes, M.J., and Andrianopoulos, A. (2000)The abaA homologue of Penicillium marneffei participatesin two developmental programmes: conidiation and dimor-phic growth. Mol Microbiol 38: 1034–1047.
1736 D. Cánovas and A. Andrianopoulos
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 62, 1725–1738

Borneman, A.R., Hynes, M.J., and Andrianopoulos, A. (2001)An STE12 homolog from the asexual, dimorphic fungusPenicillium marneffei complements the defect in sexualdevelopment of an Aspergillus nidulans steA mutant.Genetics 157: 1003–1014.
Bowyer, P., De Lucas, J.R., and Turner, G. (1994) Regulationof the expression of the isocitrate lyase gene (acuD) ofAspergillus nidulans. Mol Gen Genet 242: 484–489.
Boylan, M.T., Mirabito, P.M., Willett, C.E., Zimmerman, C.R.,and Timberlake, W.E. (1987) Isolation and physical char-acterization of three essential conidiation genes fromAspergillus nidulans. Mol Cell Biol 7: 3113–3118.
Brakhage, A.A., Andrianopoulos, A., Kato, M., Steidl, S.,Davis, M.A., Tsukagoshi, N., and Hynes, M.J. (1999) HAP-like CCAAT-binding complexes in filamentous fungi: impli-cations for biotechnology. Fungal Genet Biol 27: 243–252.
Burglin, T.R. (1991) The TEA domain: a novel, highly con-served DNA-binding motif. Cell 66: 11–12.
Chang, Y.C., and Timberlake, W.E. (1993) Identification ofAspergillus brlA response elements (BREs) by geneticselection in yeast. Genetics 133: 29–38.
Clutterbuck, A.J. (1969) A mutational analysis of conidialdevelopment in Aspergillus nidulans. Genetics 63: 317–327.
Cove, D.J. (1966) The induction and repression of nitratereductase in the fungus Aspergillus nidulans. BiochimBiophys Acta 113: 51–56.
Cubero, B., and Scazzocchio, C. (1994) Two different, adja-cent and divergent zinc finger binding sites are necessaryfor CREA-mediated carbon catabolite repression in theproline gene cluster of Aspergillus nidulans. EMBO J 13:407–415.
De Lucas, J.R., Gregory, S., and Turner, G. (1994a) Analysisof the regulation of the Aspergillus nidulans acuD gene,encoding isocitrate lyase, by construction of a hybridpromoter. Mol Gen Genet 243: 654–659.
De Lucas, J.R., Valenciano, S., Laborda, F., and Turner, G.(1994b) Glucose-induced inactivation of isocitrate lyase inAspergillus nidulans. Arch Microbiol 162: 409–413.
Fernandez, E., Fernandez, M., Moreno, F., and Rodicio, R.(1993) Transcriptional regulation of the isocitrate lyaseencoding gene in Saccharomyces cerevisiae. FEBS Lett333: 238–242.
Gillingham, A.K., Pfeifer, A.C., and Munro, S. (2002) CASP,the alternatively spliced product of the gene encoding theCCAAT-displacement protein transcription factor, is a Golgimembrane protein related to giantin. Mol Biol Cell 13:3761–3774.
Goldman, G.H., dos Reis Marques, E., Duarte Ribeiro, D.C.,de Souza Bernardes, L.A., Quiapin, A.C., Vitorelli, P.M.,et al. (2003) Expressed sequence tag analysis of thehuman pathogen Paracoccidioides brasiliensis yeastphase: identification of putative homologues of Candidaalbicans virulence and pathogenicity genes. Eukaryot Cell2: 34–48.
Hynes, M.J., Murray, S.L., Duncan, A., Khew, G.S., andDavis, M.A. (2006) Regulatory genes controlling fatty acidcatabolism and peroxisomal functions in the filamentousfungus, Aspergillus nidulans. Eukaryot Cell 5: 794–805.
Idnurm, A., and Howlett, B.J. (2002) Isocitrate lyase is essen-
tial for pathogenicity of the fungus Leptosphaeria maculansto canola (Brassica napus). Eukaryot Cell 1: 719–724.
Katz, M.E., and Hynes, M.J. (1989) Isolation and analysis ofthe acetate regulatory gene, facB, from Aspergillusnidulans. Mol Cell Biol 9: 5696–5701.
Kulmburg, P., Mathieu, M., Dowzer, C., Kelly, J., and Felen-bok, B. (1993) Specific binding sites in the alcR and alcApromoters of the ethanol regulon for the CREA repressormediating carbon catabolite repression in Aspergillusnidulans. Mol Microbiol 7: 847–857.
Lengeler, K.B., Davidson, R.C., D’Souza, C., Harashima, T.,Shen, W.C., Wang, P., et al. (2000) Signal transductioncascades regulating fungal development and virulence.Microbiol Mol Biol Rev 64: 746–785.
Lorenz, M.C., and Fink, G.R. (2001) The glyoxylate cycle isrequired for fungal virulence. Nature 412: 83–86.
Lorenz, M.C., and Fink, G.R. (2002) Life and death in amacrophage: role of the glyoxylate cycle in virulence.Eukaryot Cell 1: 657–662.
McKinney, J.D., Honer zu Bentrup, K., Munoz-Elias, E.J.,Miczak, A., Chen, B., Chan, W.T., et al. (2000) Persistenceof Mycobacterium tuberculosis in macrophages and micerequires the glyoxylate shunt enzyme isocitrate lyase.Nature 406: 735–738.
McNabb, D.S., and Pinto, I. (2005) Assembly of the Hap2p/Hap3p/Hap4p/Hap5p-DNA complex in Saccharomycescerevisiae. Eukaryot Cell 4: 1829–1839.
Madhani, H.D., and Fink, G.R. (1997) Combinatorial controlrequired for the specificity of yeast MAPK signaling.Science 275: 1314–1317.
Maeting, I., Schmidt, G., Sahm, H., Revuelta, J.L., Stierhof,Y.D., and Stahmann, K.P. (1999) Isocitrate lyase ofAshbya gossypii – transcriptional regulation and peroxiso-mal localization. FEBS Lett 444: 15–21.
Marshall, M.A., and Timberlake, W.E. (1991) Aspergillusnidulans wetA activates spore-specific gene expression.Mol Cell Biol 11: 55–62.
Mathieu, M., and Felenbok, B. (1994) The Aspergillus nidu-lans CREA protein mediates glucose repression of theethanol regulon at various levels through competition withthe ALCR-specific transactivator. EMBO J 13: 4022–4027.
Munoz-Elias, E.J., and McKinney, J.D. (2005) Mycobacte-rium tuberculosis isocitrate lyases 1 and 2 are jointlyrequired for in vivo growth and virulence. Nat Med 11:638–644.
Nayak, T., Szewczyk, E., Oakley, C.E., Osmani, A., Ukil, L.,Murray, S.L., et al. (2005) A versatile and efficient genetargeting system for Aspergillus nidulans. Genetics 172:1557–1566.
Ordiz, I., Herrero, P., Rodicio, R., and Moreno, F. (1995)Glucose-induced inactivation of isocitrate lyase in Saccha-romyces cerevisiae is mediated by an internal decapeptidesequence. FEBS Lett 367: 219–222.
Osherov, N., Mathew, J., Romans, A., and May, G.S. (2002)Identification of conidial-enriched transcripts in Aspergillusnidulans using suppression subtractive hybridization.Fungal Genet Biol 37: 197–204.
Rude, T.H., Toffaletti, D.L., Cox, G.M., and Perfect, J.R.(2002) Relationship of the glyoxylate pathway to the patho-genesis of Cryptococcus neoformans. Infect Immun 70:5684–5694.
Glyoxylate cycle in Penicillium marneffei 1737
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 62, 1725–1738

Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Molecu-lar Cloning: A Laboratory Manual. Cold Spring Harbour,NY: Cold Spring Harbour Laboratory Press.
Schuller, H.J. (2003) Transcriptional control of nonfermenta-tive metabolism in the yeast Saccharomyces cerevisiae.Curr Genet 43: 139–160.
Schweizer, A., Rupp, S., Taylor, B.N., Rollinghoff, M., andSchroppel, K. (2000) The TEA/ATTS transcription factorCaTec1p regulates hyphal development and virulence inCandida albicans. Mol Microbiol 38: 435–445.
Sewall, T.C., Mims, C.W., and Timberlake, W.E. (1990) abaAcontrols phialide differentiation in Aspergillus nidulans.Plant Cell 2: 731–739.
Solomon, P.S., Lee, R.C., Wilson, T.J., and Oliver, R.P.(2004) Pathogenicity of Stagonospora nodorum requiresmalate synthase. Mol Microbiol 53: 1065–1073.
Stringer, M.A. (1994) Analysis of two Aspergillus nidulanssporulation genes encoding members of a new class ofproteins, the fungal hydrophobins. PhD Thesis. Universityof Georgia.
Todd, R.B., Kelly, J.M., Davis, M.A., and Hynes, M.J. (1997)Molecular characterization of mutants of the acetate regu-latory gene facB of Aspergillus nidulans. Fungal Genet Biol22: 92–102.
Todd, R.B., Andrianopoulos, A., Davis, M.A., and Hynes, M.J.(1998) FacB, the Aspergillus nidulans activator of acetateutilization genes, binds dissimilar DNA sequences. EMBOJ 17: 2042–2054.
Toews, M.W., Warmbold, J., Konzack, S., Rischitor, P., Veith,D., Vienken, K., et al. (2004) Establishment of mRFP1 as afluorescent marker in Aspergillus nidulans and constructionof expression vectors for high-throughput protein taggingusing recombination in vitro (GATEWAY). Curr Genet 45:383–389.
Vanittanakom, N., Cooper, C.R., Jr, Fisher, M.C., and Sirisan-thana, T. (2006) Penicillium marneffei infection and recentadvances in the epidemiology and molecular biologyaspects. Clin Microbiol Rev 19: 95–110.
Wang, Z.Y., Thornton, C.R., Kershaw, M.J., Debao, L., andTalbot, N.J. (2003) The glyoxylate cycle is required fortemporal regulation of virulence by the plant pathogenicfungus Magnaporthe grisea. Mol Microbiol 47: 1601–1612.
Zhang, S., Varma, A., and Williamson, P.R. (1999) The yeastCryptococcus neoformans uses ‘mammalian’ enhancersites in the regulation of the virulence gene, CNLAC1.Gene 227: 231–240.
Supplementary material
The following supplementary material is available for thisarticle online:Fig. S1. Regulation of the malate synthase gene (acuE)expression in different genetic backgrounds.
This material is available as part of the online article fromhttp://www.blackwell-synergy.com
1738 D. Cánovas and A. Andrianopoulos
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 62, 1725–1738